
Soc., 1988, 7(4):503-524 - Coweeta LTER | Coweeta LTERcoweeta.uga.edu/publications/574.pdf ·...
22
J. N. Am. Benthol. Soc., 1988, 7(4):503-524 © 1988 by The North American Benthological Society Patch dynamics in lotic systems: the stream as a mosaic* CATHERINE M. PRINGLE*, ROBERT J. NAiMAN 2 , GERNOT BRETScroco 3 , JAMES R. KARR*, MARK W. Oswooo 5 , JACKSON R. WEBSTER*, ROBIN L. WELCOMME 7 , AND MICHAEL J. WlNTERBOURN 8 1 Department of Botany, University of California, Berkeley, California 94720 USA 2 Natural Resources Research Institute, University of Minnesota, Duluth, Minnesota 55811 USA (Present address: Centre for Streamside Studies, University of Washington, Seattle, Washington 98195 USA) 3 Biological Station, Austrian Academy of Science, A-3293 Lunz am See, Austria * Smithsonian Tropical Research Institute, Box 2072, Balboa, Panama (Present address: Department of Biology, Virginia Polytechnic Institute and State University, Blacksburg, Virginia 24061 USA) 5 Department of Biology, Fisheries and Wildlife, University of Alaska, Fairbanks, Alaska 99775-1780 USA 6 Department of Biology, Virginia Polytechnic Institute and State University, Blacksburg, Virginia 24061 USA 7 Fishery Resources and Environment Division, Food and Agricultural Organization, Via delle Terms di Caracalla, 00100 Rome, Italy 8 Zoology Department, University of Canterbury, Christchurch 1, New Zealand Abstract. This paper applies concepts of landscape ecology and patch dynamics to lotic systems. We present a framework for the investigation of pattern and process in lotic ecosystems that considers how specific patch characteristics determine biotic and abiotic processes over various scales. Patch characteristics include: size, size distribution within the landscape, juxtaposition, diversity, duration, and mechanisms affecting patch formation. Several topics of current interest in lotic ecology are examined from a patch-dynamics perspective: (1) response of periphyton communities to nutrient patches; (2) effects of patch dynamics on nutrient spiralling; (3) riparian patch dynamics and effects of leaf litter characteristics on lotic food webs; (4) beaver-induced patch dynamics; and (5) patch dynamics of river floodplains. We conclude that a patch-dynamics perspective coupled with a strong experimental approach can enhance the utility and predictive power of unifying concepts in lotic ecology, such as the river continuum hypothesis and nutrient spiralling, through its focus on organismal and process-specific building blocks of lotic systems. The effectiveness of a patch-dynamics approach as a framework for the study of lotic systems lies in the strength of the linkage between reductionist and whole- stream perspectives. Key words: streams, rivers, patch dynamics, landscape ecology, communities, scale, disturbance, periphyton, floodplains. The analysis of spatial patterns within land- scapes has been widely applied in the study of terrestrial ecosystems (e.g., Forman 1983, For- man and Godron 1981,1986, Noss 1983, Picket* and White 1985, Risser et al. 1984, Wiens et al. 1985). Landscapes have been viewed as mosaics of heterogeneous land forms, vegetation pat- * Paper resulting from a Working Group discussion at a symposium on "Community structure and func- tion in temperate and tropical streams" held 24-28 April 1987 at Flathead Lake Biological Station, Uni- versity of Montana, Poison, USA. terns, and land uses (Urban et al. 1987). In turn, patterns and processes within terrestrial eco- systems have been clarified by the study of spa- tial relationships among landscape patches as well as dynamics of the mosaic (Forman 1983). While this approach has not been as widely applied in aquatic systems, it has played a cen- tral role in explaining the distribution, relative abundance, and diversity of attached algae and invertebrates in marine intertidal habitats (e.g., Levin and Paine 1974, Paine and Levin 1981, Sousa 1985). 503
Transcript of Soc., 1988, 7(4):503-524 - Coweeta LTER | Coweeta LTERcoweeta.uga.edu/publications/574.pdf ·...

J. N. Am. Benthol. Soc., 1988, 7(4):503-524© 1988 by The North American Benthological Society
Patch dynamics in lotic systems: the stream as a mosaic*
CATHERINE M. PRINGLE*, ROBERT J. NAiMAN2, GERNOT BRETScroco3,JAMES R. KARR*, MARK W. Oswooo5, JACKSON R. WEBSTER*,
ROBIN L. WELCOMME7, AND MICHAEL J. WlNTERBOURN8
1 Department of Botany, University of California, Berkeley, California 94720 USA2 Natural Resources Research Institute, University of Minnesota,
Duluth, Minnesota 55811 USA(Present address: Centre for Streamside Studies, University of Washington,
Seattle, Washington 98195 USA)3 Biological Station, Austrian Academy of Science, A-3293 Lunz am See, Austria
* Smithsonian Tropical Research Institute, Box 2072, Balboa, Panama(Present address: Department of Biology, Virginia Polytechnic Institute and State University,
Blacksburg, Virginia 24061 USA)5 Department of Biology, Fisheries and Wildlife, University of Alaska,
Fairbanks, Alaska 99775-1780 USA6 Department of Biology, Virginia Polytechnic Institute and State University,
Blacksburg, Virginia 24061 USA7 Fishery Resources and Environment Division, Food and Agricultural Organization,
Via delle Terms di Caracalla, 00100 Rome, Italy8 Zoology Department, University of Canterbury, Christchurch 1, New Zealand
Abstract. This paper applies concepts of landscape ecology and patch dynamics to lotic systems.We present a framework for the investigation of pattern and process in lotic ecosystems that considershow specific patch characteristics determine biotic and abiotic processes over various scales. Patchcharacteristics include: size, size distribution within the landscape, juxtaposition, diversity, duration,and mechanisms affecting patch formation. Several topics of current interest in lotic ecology areexamined from a patch-dynamics perspective: (1) response of periphyton communities to nutrientpatches; (2) effects of patch dynamics on nutrient spiralling; (3) riparian patch dynamics and effectsof leaf litter characteristics on lotic food webs; (4) beaver-induced patch dynamics; and (5) patchdynamics of river floodplains.
We conclude that a patch-dynamics perspective coupled with a strong experimental approachcan enhance the utility and predictive power of unifying concepts in lotic ecology, such as the rivercontinuum hypothesis and nutrient spiralling, through its focus on organismal and process-specificbuilding blocks of lotic systems. The effectiveness of a patch-dynamics approach as a frameworkfor the study of lotic systems lies in the strength of the linkage between reductionist and whole-stream perspectives.
Key words: streams, rivers, patch dynamics, landscape ecology, communities, scale, disturbance,periphyton, floodplains.
The analysis of spatial patterns within land-scapes has been widely applied in the study ofterrestrial ecosystems (e.g., Forman 1983, For-man and Godron 1981,1986, Noss 1983, Picket*and White 1985, Risser et al. 1984, Wiens et al.1985). Landscapes have been viewed as mosaicsof heterogeneous land forms, vegetation pat-
* Paper resulting from a Working Group discussionat a symposium on "Community structure and func-tion in temperate and tropical streams" held 24-28April 1987 at Flathead Lake Biological Station, Uni-versity of Montana, Poison, USA.
terns, and land uses (Urban et al. 1987). In turn,patterns and processes within terrestrial eco-systems have been clarified by the study of spa-tial relationships among landscape patches aswell as dynamics of the mosaic (Forman 1983).While this approach has not been as widelyapplied in aquatic systems, it has played a cen-tral role in explaining the distribution, relativeabundance, and diversity of attached algae andinvertebrates in marine intertidal habitats (e.g.,Levin and Paine 1974, Paine and Levin 1981,Sousa 1985).
503

504 C. M. PRINGLE ET AL. [Volume 7
*-_ - ~ Y ! " . '~I ^ t-. > 7 7 ~
In streams, as in intertidal zones, the distri-bution of both sedentary and mobile organismsis strongly influenced by the movements ofoverlying waters. While the unidirectional flowof water in streams can create spatial and tem-poral heterogeneity (in patterns of current ve-locity, substratum sorting, faunal habitat, etc.),mixing processes also reduce heterogeneity ofother factors (e.g., spatial and temporal differ-ences in nutrient concentrations). Surface waterflow is an abiotic vector that imposes a direc-tional flux of materials on the general back-ground of resource concentration gradients(Forman 1983, Wiens et al. 1985).
Viewing streams as mosaics of patches pro-vides a fresh perspective for lotic studies. Mech-anisms of spatial and temporal complexity on avariety of scales can be investigated in a man-ageable fashion by examining interrelation-ships between basic building blocks of the sys-tem. For instance, unifying concepts in loticecology that have emphasized longitudinallinkages, such as the river continuum concept(Minshall et al. 1985, Vannote et al. 1980) andnutrient spiralling (Newbold et al. 1981, Web-ster 1975), can be evaluated by studying changesin characteristics of patches and interactions be-tween specific patch types along a stream con-tinuum. If there are net imbalances in fluxes orthe spread of disturbances across boundaries inrelation to within-patch processes, a directionalbias to system fluxes can be established (Wienset al. 1985). Thus, a patch dynamics approachcan complement unifying theories that empha-size longitudinal continua by providing a re-ductionist tool for evaluation of such concepts.As stressed by Orians (1980), one of the greatchallenges in ecology is the bridging of theconceptual gap between micro- and macro-ecol-ogy. The study of patch dynamics with its em-phasis on processes and interactions at variousscales is one logical approach to making thetransition.
It has long been recognized that rivers andstreams are heterogeneous environments wherehighly clumped distributions of organisms posesevere sampling problems (e.g., Hynes 1970).Traditionally, aquatic ecologists have designat-ed riffles, pools, runs, etc. as relatively homo-geneous units for study (e.g., Hynes 1970) and,at a finer scale, patchiness has been describedin terms of physical factors such as depth, width,current speed, and substratum particle size (e.g..
Gorman and Karr 1978). However, patchinesswithin streams at scales perceived and/or ex-ploited by stream organisms is a reality that hasoften been ignored. Research in lotic ecologyoften has been constrained by preconceived no-tions of what a "patch" is.
To a considerable extent, studies have stressedcomparisons of conditions and/or communitieswithin and between patches, rather than inter-active processes that control form and functionof patch mosaics. An instructive exception isthe work on thermal effluent communities,which demonstrated the continually changingnature of these systems and the interdepen-dence of temperature, flow, grazing, and pre-dation (Wickstrom and Wiegert 1980). Recentdescriptions of stream riffle ecosystems as opennon-equilibrium systems (Peckarsky 1985) andpatches in transitory equilibrium with otherpatches (Sheldon 1984) indicate an increasingperception of stream patches in dynamic terms.
The strength of a patch perspective to providea better understanding of functional relation-ships is demonstrated by the work of Statzner(1987) and Statzner and Higler (1986). They havedrawn attention to regions of hydraulic stresswhich are important discontinuities in riversystems and influence the distribution of zoo-benthos (see also Statzner et al. [1988] in thisissue). Recent studies of beaver (Johnston andNaiman 1987, Naiman and Melillo 1984, Nai-man et al. 1986) provide an example of howlarge animals can create patches in streams andact as biotic vectors that affect fluxes of energyand materials across patch boundaries. A patchperspective has been employed in lotic systemson successively smaller scales in investigationsof fish (Angermeier and Karr 1984, Power 1984),sediment-fauna-leaf pack associations (Reice1974,1977,1980,1985), behavioral responses ofgrazing invertebrates to patchiness in algal foodabundance (Hart 1981, Hart and Resh 1980,Kohler 1984,1985), and periphyton communityresponse to nutrient patches (Pringle 1985b,1987, Pringle and Bowers 1984).
Our objective is to develop the concept ofpatch dynamics (Pickett and White 1985,Thompson 1978, Wiens et al. 1985) as a usefultool that will enhance predictive understand-ing of lotic systems. We present an operationaldefinition of "patch" and discuss patch charac-teristics as they relate to the function of loticecosystems on differing scales. We then apply

1988] PATCH DYNAMICS IN LOTIC SYSTEMS 505
a patch perspective to various ecological prob-lems in lotic systems, ending with a series ofconclusions and recommendations.
Patch definition: what scales ofvariation are perceived by
the organism?
Appropriate scales in ecological studies de-pend upon the organism(s) of study, the timeunit during which the organism(s) will be af-fected by environmental fluctuations and thequestion(s) being asked. For example, in his ex-amination of environmental factors determin-ing spatial and temporal heterogeneity of phy-toplankton populations, Reynolds (1984)adopted the time unit of "cell generation time",asking the question, "which scales of variationare perceived by the intact cell?" Human per-ceptions often operate on radically differenttemporal and spatial scales from those of otherorganisms we study, contributing to the dis-parity between relevant and tractable scales ofinvestigation. As stated by Allen (1977), "theecologist is likely to measure the environmentat frequencies too long and at grain sizes toolarge to correspond to the short frequency, finegrain actuality Small life forms are so farremoved from human scales of perception thateven the best amplifiers of our senses can't beused directly." Connell (1978) likewise notedthat with long-lived organisms such as trees orcorals, gradual changes in climate over severalhundred years represent the same scale as sea-son does to a phytoplankton community.
Our working definition of a "patch" is an op-erational one: A patch is a spatial unit that isdetermined by the organism(s) and problem(s)in question. This definition incorporates a scalarcomponent since organisms and processes canoperate within several spatial and temporalscales simultaneously. As discussed by Pickettand White (1985), patch implies a spatial pat-tern where patches are spatially related to eachother and to the surrounding, less affected ma-trix. The study of patch dynamics focuses onmechanisms behind patch change and inter-actions between patches.
While the distinction between riffles and poolsmay yield patch units appropriate to some prob-lems in lotic ecology such as factors determin-ing fish distribution and habitat preference, sucha level of spatial heterogeneity may be inap-
propriate for studies addressing fine-grainedheterogeneity such as efficient nutrient cyclingwithin microbial communities. In streams,patches may be determined by many interact-ing factors, including substratum conditions,topography, current patterns, organisms, anddisturbance. Clearly, patch size and patchboundaries perceived by individual organismsvary significantly among organisms and evenfor the same organism at different times. As aboundary, a pond margin is impermeable to afish, permeable and of major importance to abeaver (Castor canadensis), and not really per-ceived by a parasite with life history stageswithin both fishes and birds.
Characteristics of patches
The effects of patch dynamics on populations,communities, and ecosystems have been de-scribed in detail for a variety of terrestrial andmarine habitats (e.g., Pickett and White 1985),often focusing on characteristics of patch typeswithin these different systems (e.g., Sousa 1985).Here we provide a framework for applicationof patch dynamics to problems in lotic ecology(Table 1) by consideration of the following patchcharacteristics and how they affect biotic andabiotic processes over various scales: (1) size andsize distribution; (2) density; (3) juxtaposition;(4) diversity; (5) duration; and (6) mechanismsaffecting formation.
How do these patch characteristics affect loticecosystem function? All are critical determi-nants of both within- and between-patch dy-namics. This importance is dramatically illus-trated by the loss of biotic integrity thataccompanies major structural modifications ofstream channels. The erosive dynamics of waterin meandering streams creates habitat mosaicsof major consequence to the distribution of manyorganisms (Karr and Schlosser 1978). For in-stance, with respect to fish habitat, the struc-tural simplification that results from streamchannelization generally results in a decreasein the variance of patch sizes, patch density, thenumber of juxtapositions between patches, patchdiversity, and even in mechanisms affectingsubsequent patch formation.
Variations in patch characteristics that occurover extremes in spatial and temporal scales caninfluence stream structure and function. For ex-ample, the dynamics of fish populations are in-

506 C. M. PRINGLE ET AL. [Volume 7
TABLE 1. Guidelines for the application of patch dynamics in lotic systems.
I. A first step in applying a patch-dynamics perspective to a specific ecological problem is to define theconceptual patch unit(s) after consideration of the following questions:A. On what scales is variation perceived or exploited by the organism(s) and/or process under study?B. On what scale is environmental heterogeneity (patch-to-patch variation) affected by the organism
or process under study?n. Once the path unit is determined, the following characteristics may be considered with respect to the
ecological problem in question: (a) patch juxtaposition; (b) patch size and density; (c) patch size distri-bution; (d) patch diversity; (e) patch duration; and (f) mechanisms affecting patch formation.
III. Sampling units, sampling time frames, and/or experimental/manipulative approaches should be selectedthat are appropriate to the scale of the problem to be addressed.
IV. To assess system-level effects of patch-specific phenomena, one must stand back and adopt a broad focus,examining key characteristics of patches (listed above) and process-oriented interactions between patches,within the larger context of the stream landscape.
fluenced by spatial variations in habitat patchmosaics ranging in size from localized substra-tum patches to entire catchments. In short streamsegments, the presence of instream cover orhabitat patches such as undercut banks, logs,etc. are important determinants of fish biomass,species diversity, and community composition(e.g., Angermeier and Karr 1984, Fraser and Sise1980, Hunt 1971, Sheldon 1968). Some fish pop-ulations may be composed of both sedentaryand mobile groups depending upon the relativesuitability of local habitats, particularly duringchanging environmental conditions (Fagen1962, Harima and Mundy 1974, Karr and Dud-ley 1981). The physical process of fish migra-tion, itself, presupposes the existence and/orabsence of certain patch types within migratorypaths. On the floodplain for instance, the pres-ence of floating vegetation may prohibit pas-sage of fish species that have high oxygen de-mands because of low oxygen concentrationsunder the mat. On the other hand, fishes indewatered desert streams inhabit refugialpatches under rocks or algal mats to await thenext spate (Stanford and Ward 1986). Long dis-tance migrants in rivers are frequently pre-sented with the need to negotiate rapids and,thus, are dependent on the presence of a suit-able patch structure in the form of resting pools.A serious consequence of human interventionin river systems (e.g., dam building, defores-tation, channelization, etc.) is that the patchstructure necessary for fish migrations may bealtered, impeding upstream and/or down-stream movements. Decreased habitat mosaicson a broad scale may account for declines infish species diversity on a regional level. Theobserved decline in diversity of fishes from the
eastern to western region of the central plainsof North America is a good example (Cross 1970,Matthews 1988—see this issue). The six generalpatch characteristics listed above may also in-fluence the level of system stability and resi-liency in response to disturbance. Disturbancesare less likely to spread over a spatially heter-ogeneous area than through a homogeneous re-gion. If patches are isolated in a biotically orabiotically dissimilar matrix, their response toa particular disturbance will probably differ fromthat when barriers do not exist (Pickett andWhite 1985). For instance, geomorphic hetero-geneity in streams (e.g., size, density, and sizedistribution of patches within the stream land-scape, and juxtaposition and diversity of patches,as defined by topographic characteristics) mayplay a major role in determining effects of dis-turbances, since localized areas of differing geo-morphometry may respond differently to thesame disturbance regime (Pickett and White1985). On a microscopic scale, algal cells livingwithin hollows and indented areas of suspend-ed sediment particles are subject to less abrasionthan cells located on exposed portions of sed-iment grains. Algal populations inhabitingcrevices on sand grains may be considered asmicropatches that serve as sources of new col-onists for sediment particles following thescouring effect of spates. Furthermore, algal andmacrophyte communities living in topograph-ically different substratum patches can experi-ence different current regimes and nutrient dif-fusion rates, thus influencing local algalcommunity composition and production. On alarger scale, subterranean burrows withinstreams may serve as refugia for fish and in-vertebrates during floods and during periods

1988] PATCH DYNAMICS IN LOTIC SYSTEMS 507
when stream channels are dewatered (e.g., Glo-dek 1978).
There is also evidence that a high diversityof resource patches confers community stabil-ity. For example, Tilman (1982) hypothesizedthat diversity of plant communities may be atleast partially explained by resource competi-tion within a spatially heterogeneous environ-ment. If individual plant species tend to max-imize competitive abilities within specific andnarrow ranges of resources (e.g., N:P), spatialvariation in resource supplies may lead to thecoexistence of many species. Evidence that dis-similarity of nutrient regimes between patchesin streams can promote periphyton species di-versity (Pringle 1985b) is consistent with Til-man's hypothesis.
Response of lotic periphytoncommunities to nutrient patches
The turbulent nature of many, if not most,lotic environments tends to homogenize dis-solved and particulate materials as they aretransported downstream, at least in comparisonto more sequestered supplies in lentic and ter-restrial systems. Nonetheless, contiguous tem-poral and spatial patterns in concentrations andsupply rates of resources essential to productionof plant and associated bacterial communitiesmay be identified (Meyer et al. 1988—see thisissue). Little is known about the effects of suchnutrient patches on algal growth and distri-bution in streams. Attached algae may experi-ence variable nutrient levels over wide ex-tremes of temporal scale: (1) seasonal (e.g., inputsof decomposing leaves and other allochthonousmaterials which release nutrients in a high ini-tial pulse [Elwood et al. 1981, Howarth and Fish-er 1976]); (2) hourly (e.g., following storms[Hynes 1970, Wetzel and Likens 1979]); and (3)milliseconds (e.g., when the diffusion regimeexperienced by a sessile cell is altered by themovement of an adjacent motile cell). Effects ofnutrient patches on microbial communities aredifficult to determine because "microbes areclearly scaled very differently in their biologi-cal strategies than are humans .... So large isthe surface area of a cell (relative to its volume),that the effect of nutrient uptake might be anutrient "draft" analogous to the breeze near toan air intake of a large building" (Allen 1977).
Algal response to variations in nutrient sup-
plies that occur on larger scales (e.g., mass load-ing) are comfortably addressed according to ourhuman sense of time and space and are the mostthoroughly documented; however, little isknown about effects of finer levels of temporaland spatial variation in nutrient supplies. Giventhat a plant a few jim in diameter is responsiveto the nutrients contained in the environmentimmediately surrounding it (e.g., Pasciak andGavis 1974) and that algal turnover rates areoften a matter of hours, these finer levels ofvariation may be crucial in determining micro-bial distribution and abundance. It follows thatsuch patch dynamic responses may vitally in-fluence overall food-web complexity and pat-tern.
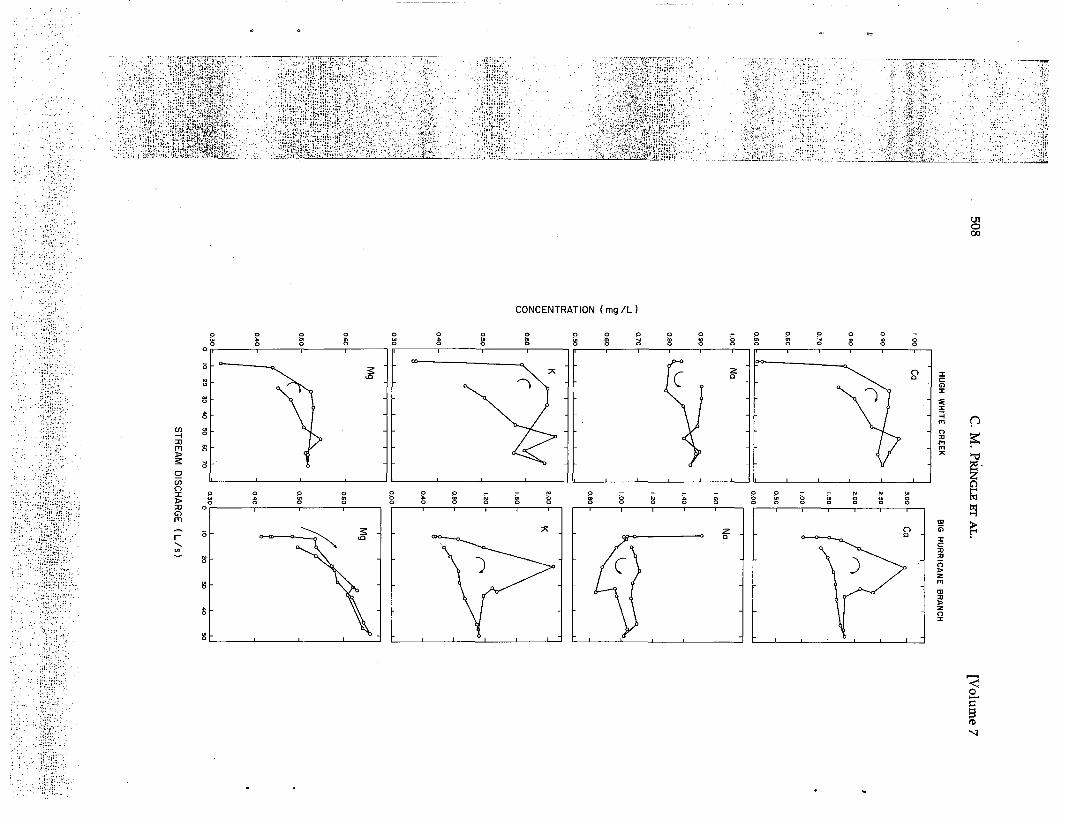
Daily measurement of ambient nutrient levelsis a widely accepted way of characterizing thegeneral nutrient environment experienced byattached microbes in streams (e.g., Peterson etal. 1983, Pringle et al. 1986, Stockner and Short-reed 1978): however, such measurements inte-grate information at a scale that is orders ofmagnitude too large to detect fine grain tem-poral and spatial variation in nutrient patches(e.g., Fig. 1).
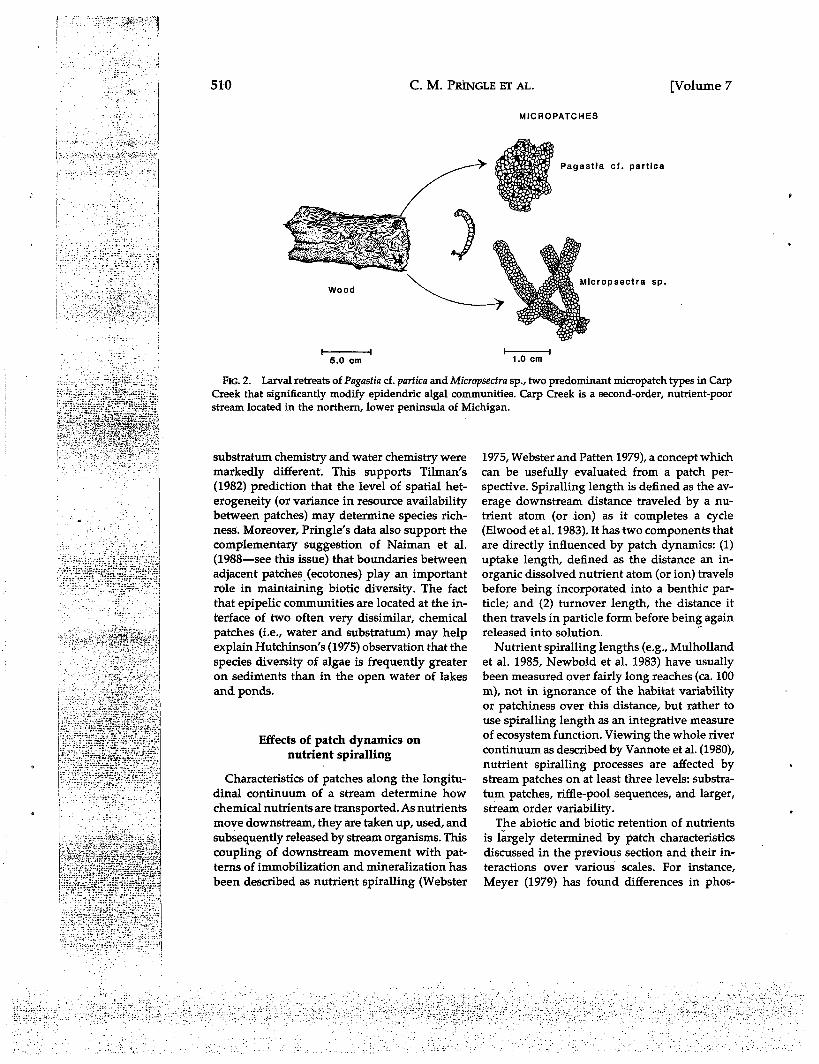
A case study in point concerns the relation-ship between organisms in the periphyton andthe surfaces to which they are attached. Mac-rophytes, decomposing wood, and leaf litter canbe direct sources of nutrients for attached mi-crobes. Retreats constructed by many zooberi-thic species also may constitute natural nu-trient-diffusing surfaces for attached algae,owing to metabolic and excretory activities oflarval inhabitants (Pringle 1985a). For instance,many Chironomidae and Trichoptera larvaeconsolidate sand grains into tube-like retreats.In a nutrient-deficient stream in northern Mich-igan, significantly greater amounts of ammonia-N, ortho-phosphate, and total phosphorus wereextracted from tubes of the chironomid Pagastiacf. partica in comparison with controls of un-consolidated sand (Pringle 1985b). The retreatswere covered by dense mats of diatoms, sup-porting over 10 times the chlorophyll a thanequivalent weights of control sand (Pringle1985b) and over 12 times the diatom biomasscompared with adjacent wood surfaces (Prin-gle 1985a). In this instance, algal communitydynamics apparently respond to nutrientpatches mediated by the chironomid larvae.Such efficient biophysical coupling representsa lower spatial limit within the framework of
3 ~"i
^
CONCENTRATION (mg/L)
00
S §
3m >
o o o o o o~i 1 r
GJ3:S
to Nl U
i
n
Om
sftXI

1988] PATCH" DYNAMICS IN LOTIC SYSTEMS 509
nutrient spiralling (Elwood et al. 1981, Web-ster 1975).
Ecosystem-level nutrient fluxes may occur inpatch mosaics far smaller than those generallyrecognized in contemporary plant ecology. Forexample, denitrificahon with biophysical ag-gregates in aerobic soils can be the major path-way of nitrogen loss from forests (Myrold et al.1982 as cited by Vitousek 1985). Likewise, smalllocalized nutrient patches around individualchironomids and their retreats may cumula-tively influence ecosystem-level properties ofstreams because chironomids are often ex-tremely abundant (e.g., Saether 1968, Wiley1976). In a nutrient-poor northern Michiganstream, larval retreats of the chironomids Mi-cropsectra sp. and Pagastia partica (Fig. 2) typi-cally covered 40-75% of submerged wood sur-faces in midsummer and supported 7 to 12 timesgreater algal biomass than wood unmodified bylarval retreats (Pringle 1985a). When present atmaximum densities, a conservative estimate wasthat retreats of the two taxa supported over 95%of total epidendric periphyton standing crop inthe stream. Population dynamics, distribution,and feeding behavior of chironomids can havea direct effect on all characteristics of patchesdiscussed in the previous section: juxtaposition,density, size and size distribution, diversity, andmechanisms affecting patch formation and du-ration. For example, when chironomid larvaeemerge as winged adults, patterns of nutrientcycling in the stream may be altered owing tothe loss of spatially-fixed nutrient patches pre-viously maintained by larvae. In accordance withthis prediction, Hershey et al. (1988) measuredlower concentrations of chlorophyll a on pupaltubes than on larval tubes of the chironomidOrthodadius rivulorum. One might predict thateffects of localized nutrient patches on stream-wide patterns of algal distribution would bemost pronounced in a stream with low ambient
nutrient concentrations; periphyton would befound in contiguous patches in response to lo-calized nutrient sources. In a nutrient-richstream, however, localized patch effects areameliorated and periphyton may be more uni-formly distributed.
Viewing effects of nutrients derived from dif-ferent sources as patch-specific phenomena is aprerequisite for the careful examination of in-teractions between nutrient sources (patches).Taking this approach, one can quantify effectsof nutrient patches through experimental ma-nipulation. For example, Pringle (1985b, 1987)developed an integrated bioassay to separatepatch-specific effects of substratum and waternutrient supplies on algal growth and com-munity structure. Experiments that enriched thewater and substratum in a fully crossed designshowed that nutrients derived from differentsources interacted to determine periphytongrowth response; periphyton responded tocombined influences of water and substratumenrichment in an additive or synergistic fash-ion, depending on amounts and types of nu-trients added from each source. In another study,algal epiphytes of Phragmites were found to bedependent on silicate released from host stems,when competition for silicate between plank-tonic and epiphytic diatoms resulted in silicatedepletion in the water (Jorgensen 1957). Whensilica concentrations are low, epipelic diatomsobtain supplies of this mineral from the sedi-ment and interstitial water (Happey-Wood andPriddle 1984, Werner 1977). Accordingly, astream can be envisioned as a mosaic of nutrientmicro-patches, which differ dynamically inchemical nature and, thus, may impart variablebut potentially predictable biotic responses be-tween patches.
Indeed, results of integrated bioassays em-ployed by Pringle (1985b) showed that peri-phyton species diversity was highest where
FIG. 1. Data illustrating short-term temporal variation in ambient concentrations of calcium, sodium,potassium, and magnesium as a function of discharge in Hugh White Creek and Big Hurricane Branch duringa storm on 30 June 1981. Both streams are located at Coweeta Hydrologic Laboratory in southwestern NorthCarolina, USA. Hugh White Creek drains an undisturbed hardwood forest watershed and Big HurricaneBranch drains a watershed that was logged in 1977. The arrows show the time sequence of samples over thetotal sampling period of about three hours. Samples from Hugh White Creek were taken every 15 minutes.Samples from Big Hurricane Branch were taken even more frequently when the hydrograph was rising. Thetime between the onset of the storm and peak discharge was 50 minutes in Big Hurricane Branch and 65minutes in Hugh White Creek. Data from J. R. Webster and S. W. Golladay, Virginia Polytechnic Instituteand State University.
!
510
i-vr
C. M. PRINGLE ET AL.
MICROPATCHES
[Volume 7
Pagastla cf. partlca
Mlcropsectra sp.
5.0 cm 1.0 cm
FIG. 2. Larval retreats of Pagastia cf. partica and Micropsectra sp., two predominant micropatch types in CarpCreek that significantly modify epidendric algal communities. Carp Creek is a second-order, nutrient-poorstream located in the northern, lower peninsula of Michigan.
substratum chemistry and water chemistry weremarkedly different. This supports Tilman's(1982) prediction that the level of spatial het-erogeneity (or variance in resource availabilitybetween patches) may determine species rich-ness. Moreover, Pringle's data also support thecomplementary suggestion of Naiman et al.(1988—see this issue) that boundaries betweenadjacent patches (ecotones) play an importantrole in maintaining biotic diversity. The factthat epipelic communities are located at the in-terface of two often very dissimilar, chemicalpatches (i.e., water and substratum) may helpexplain Hutchinson's (1975) observation that thespecies diversity of algae is frequently greateron sediments than in the open water of lakesand ponds.
Effects of patch dynamics onnutrient spiralling
Characteristics of patches along the longitu-dinal continuum of a stream determine howchemical nutrients are transported. As nutrientsmove downstream, they are taken up, used, andsubsequently released by stream organisms. Thiscoupling of downstream movement with pat-terns of immobilization and mineralization hasbeen described as nutrient spiralling (Webster
1975, Webster and Patten 1979), a concept whichcan be usefully evaluated from a patch per-spective. Spiralling length is defined as the av-erage downstream distance traveled by a nu-trient atom (or ion) as it completes a cycle(Elwood et al. 1983). It has two components thatare directly influenced by patch dynamics: (1)uptake length, defined as the distance an in-organic dissolved nutrient atom (or ion) travelsbefore being incorporated into a benthic par-ticle; and (2) turnover length, the distance itthen travels in particle form before being againreleased into solution.
Nutrient spiralling lengths (e.g., Mulhollandet al. 1985, Newbold et al. 1983) have usuallybeen measured over fairly long reaches (ca. 100m), not in ignorance of the habitat variabilityor patchiness over this distance, but rather touse spiralling length as an integrative measureof ecosystem function. Viewing the whole rivercontinuum as described by Vannote et al. (1980),nutrient spiralling processes are affected bystream patches on at least three levels: substra-tum patches, riffle-pool sequences, and larger,stream order variability.
The abiotic and biotic retention of nutrientsis largely determined by patch characteristicsdiscussed in the previous section and their in-teractions over various scales. For instance,Meyer (1979) has found differences in phos-

1988] PATCH DYNAMICS IN LOTIC SYSTEMS 511
phate sorption on silty versus sandy substrata.Most stream segments or reaches are mosaics ofdifferent substratum types (e.g., gravel, sand,silt, and organic accumulations) that may dif-ferentially affect abiotic uptake of nutrients.Therefore, the relative size of different physicalsubstratum patches, their size distribution anddensity within the stream landscape, combinedwith their juxtaposition, diversity, and dura-tion, all interact to affect the overall nutrientretention of the reach and will determine, inpart, the dynamics of nutrient cycling withinthe drainage network.
Nutrient dynamics are often mediated by bothautotrophic and heterotrophic biota (e.g., El-wood et al. 1981, Gregory 1978, Lock and Johns1979) which may be distributed in fairly dis-crete, but non-uniform patches. For example,Pringle (1985a, 1987) found dense patches ofdiatoms on the nutrient-rich retreats of chiron-omids and on artificial substrates enriched withnutrients. Also, Hart (1985) has shown thatgrazing caddisflies can create patchy distribu-tions of attached algae, and thus influencenutrient cycling. On somewhat larger scales,vascular macrophytes have highly patchy distri-butions in streams and can be major componentsof nutrient cycles (e.g., Hill 1979, Meyer 1979,Vincent and Downes 1980). Even when plantdistributions appear to be fairly uniform, theiractivity may have considerable spatial variabil-ity related to current patterns. Several studieshave shown that periphyton production andnutrient uptake is significantly affected by cur-rent velocity (Lock and Johns 1979, Mclntire1966, Whitford and Schumacher 1964). More-over, attached algae possess a variety of char-acteristics and adaptations that maximize theirability to exploit patch-specific variations in nu-trient availability, hence increasing the overallretentiveness of the system and shortening nu-trient spiralling lengths. Such characteristicsinclude cell permeability, cell motility, pres-ence of mucilage and/or enzymes that facilitatenutrient uptake (e.g., alkaline phosphatase),mode of enzyme release, heterotrophic abilities,luxury consumption, and storage. Virtuallynothing is known about how these factors in-teract to determine nutrient cycling processesoperating over a range of scales.
In many streams, especially small shadedheadwater streams, nutrient uptake by hetero-trophic organisms associated with decaying or-
ganic matter may be considerably more impor-tant than autotrophic processes. For example,in their studies of a small Tennessee stream,Newbold et al. (1983) and Mulholland et al.(1985) found that phosphorus spiralling lengthwas inversely correlated with the abundance oflarge particulate detritus on the streambed. Infall, when this material was abundant, spiral-ling length of phosphorus was short. In sum-mer, once most of this material had decayed,phosphorus spiralling length was longer. Asimilar situation probably exists for other nu-trients, especially nitrogen, as many studies havedemonstrated nitrogen immobilization by de-caying leaves (e.g., Kaushik and Hynes 1968,Triska and Buckley 1978) and wood (e.g., Au-men et al. 1985, Melillo et al. 1983). Althoughleaf fall may initially distribute leaves uniform-ly over a streambed, these leaves are quicklyrearranged by the current into a highly clumpeddistribution (Fisher and Likens 1973). Forexample, fine and coarse particulate organicmatter in samples taken from five first- and sec-ond-order streams in North Carolina was sig-nificantly clumped in every case (Table 2).
In addition to immobilizing dissolved nu-trients, decaying organic material may also bea site for denitrification. Several studies (e.g.,Kaushik and Robinson 1976, Swank and Caskey1982) have suggested that denitrification is amajor pathway of nitrogen loss from streams.Swank and Caskey (1982) found that rates ofdenitrification were highly variable over thestreambed and tended to be correlated with sed-iment organic matter and the presence of in-termittent seeps.
At a larger scale, woody debris has been shownto be an important component of many streams(e.g., Triska and Cromack 1980). The distribu-tion of this material is extremely patchy andusually occurs in debris dams. These woody de-bris dams are major sites for nutrient transfor-mations associated with the decomposition ofwood, nutrient uptake, and nitrogen fixation(e.g., Triska et al. 1984). These dams also slowwater velocity, thus shortening nutrient spi-ralling; but, their greatest effect on nutrient spi-ralling probably is that they retard transport oforganic and inorganic particles and their asso-ciated nutrients (e.g., Bilby 1981).
A higher level of patchiness in streams is as-sociated with the alternation of riffles and pools.Because of differences in slope and hence water

512 C. M. PRINGLE ET AL. [Volume 7
• _ ' . _ --^"" * 5 f "J.
TABLE 2. Clumping of fine and coarse particulateorganic matter (FPOM and CPOM) taken from fivestreams at Coweeta Hydrologic Laboratory, MaconCounty, North Carolina. Samples were collected witha 0.071-m2 pot sampler in July 1985. The ratio of thevariance to the mean (s2/x) is an indication of thedispersion of the material. A ratio greater than oneindicates clumping. In all cases the ratio was signif-icantly greater than one (chi-square test, Cox 1985).Data from S. W. Golladay, Virginia Polytechnic In-stitute and State University.
Stream
FPOMSawmill BranchBig Hurricane BranchCarpenter BranchHugh White CreekGrady Branch
CPOMSawmill BranchBig Hurricane BranchCarpenter BranchHugh White CreekGrady Branch
(g/mz)
156.046.1
499.6102.274.1
97.298.0
277.3117.7133.8
n
3060306030
3060306030
sV*
387.730.5
407.947.423.9
63.9208.1179.9105.4132.7
velocity, riffles and pools differ in substratumtype, particulate organic matter accumulation,and plant distribution. These factors all affectnutrients in ways similar to the smaller scalepatterns discussed previously. Meyer (1980)found significant differences between the phos-phorous dynamics of pools and rapids and Nai-man et al. (1986) discussed the differences be-tween nutrient processes in undammed andbeaver-dammed streams.
Patchiness at a comparative spatial scale oc-curs when treefall creates a canopy gap over awoodland stream, increasing light penetrationto the streambed. Primary production in manywoodland streams appears to be limited byavailable sunlight (e.g., Triska et al. 1983). Can-opy gaps may create patches where autotrophicprocesses temporarily dominate nutrient dy-namics (S. W. Golladay, Virginia PolytechnicInstitute, personal communication).
Finally, over the length of an entire river,nutrient spiralling efficiency or length may dif-fer dramatically between reaches. In shaded,headwater streams, heterotrophic processes andparticle retention dominate. In somewhat larg-
er streams, that are open to sunlight but are stillshallow, autotrophic nutrient uptake, release ofdissolved organic compounds, and plant decayare probably the most important processes af-fecting nutrient dynamics. Farther down-stream, in deep turbid rivers, heterotrophic nu-trient processes may again predominate, owingto decomposition of relatively higher organiccarbon loads. Minshall et al. (1983) and Naimanet al. (1987) showed that carbon turnover lengthsincreased several orders of magnitude in down-stream reaches of large rivers. However, neitherstudy was extended to the near lentic condi-tions of very large rivers where flood plain dy-namics may be very important.
When viewed from a landscape perspective,nutrient spiralling is controlled by a shiftingmosaic of patch-specific biotic and abiotic pro-cesses existing on various spatial scales. We haveattempted to discuss typical patterns of patch-iness and how they affect nutrient spiralling.Perhaps more interesting than some of the typ-ical patterns discussed, may be the variabilityin patch-specific nutrient cycling processeswithin and between biomes and ways in whichnatural and anthropogenic disturbances (suchas volcanoes, major storms, clear-cutting, anddam building) affect patterns of patchiness andrelated nutrient transport.
Riparian patch dynamics and effects ofleaf litter characteristics on
lotic food webs
Streams and rivers are like threads traversingthe shifting patchwork of terrestrial landscapes.Some patches are obvious (e.g., fires and agri-cultural or urban areas), whereas others are moresubtle. Impacts of terrestrial patches on streamsare often buffered and/or obscured by dilutionand downstream transport of nutrients and or-ganic materials. The extent to which each ter-restrial patch imposes ecological changes uponits drainage system depends upon both spatial(terrestrial patch size in relation to stream size)and temporal (longevity of the patch) scales.
Few studies have investigated stream riparianzones from a patch-dynamics perspective, andsuch an approach may be beneficial since land-stream interactions are so complex. Riparianvegetation acts as both a source and filter forenergy and materials entering stream ecosys-

1988] PATCH DYNAMICS IN LOTIC SYSTEMS 513
terns. It influences water temperature by shad-ing, although the effect varies seasonally in re-lation to the deciduous versus evergreen natureof the flora. Shading may also limit instreamprimary production; and streamside vegetationis a major source of particulate detritus used byzoobenthos in headwater streams. As discussedabove with respect to nutrient spiralling, bothof these influences, in turn, may affect lotic nu-trient cycling processes and trophic structure.A desert stream with limited riparian vegeta-tion may be predominately autotrophic, where-as a stream of similar size in the deciduous biomemay be dominated by heterotrophic processes,owing to allochthonous leaf fall from the ri-parian canopy (Minshall 1978, Minshall et al.1983). Riparian vegetation also influences chan-nel form by limiting bank and catchment ero-sion. It acts as a moderator of run-off volume(Jackson 1987), decreasing sediment, nutrient,and water transport from adjacent disturbedareas (e.g., Karr and Schlosser 1978, Peterjohnand Correll 1984).
The ability of catchments as a whole to retainnutrients may be addressed from the perspec-tive of patch dynamics (Vitousek 1985). A catch-ment composed of a mosaic of vegetation patchesmay be more retentive than catchments domi-nated by climax or uniformly distributed vege-tation. But how do riparian vegetation patchesinteract to affect the relative importance of al-lochthonous versus autochthonous energysources, nutrient and sediment inputs, dis-charge fluctuations, habitat requirements foradult stages of "aquatic" insects, the quantityand quality of allochthonous food sources avail-able to stream organisms, and consequently thetrophic structure of stream communities? Clear-ly these are complicated questions because ofthe interacting effects of many different factors.We suggest that a patch-dynamics perspective,coupled with a strong experimental approachis particularly useful in identifying and quan-tifying key factors that have heretofore beenobscured by interrelated effects.
How do riparian patch characteristics affectthe lotic food webs in terms of quantity andquality of leaf litter? Diversity, species com-position, and successional stage of riparianvegetation is clearly important. Patches of re-duced vegetational diversity (e.g., monocul-tures such as orchards or intensive silviculture)may not provide quantity or quality of leaf litter
in a time sequence that favors high diversity ofstream organisms. Processing rates of leaf littervary by tree species (Anderson and Sedell 1979).Thus, sustained availability to shredders of mi-crobially conditioned leaf detritus is presum-ably dependent upon a suitable mix or patchi-ness of riparian tree species (Golladay et al. 1983).Experiments by McArthur et al. (1985) indicatethat within stream reaches, resident microflorascan be adapted to use dissolved organic com-pounds derived from particular groups of ri-parian plants. Likewise, Molles (1982) demon-strated how benthic community structure(taxonomic composition and abundance offunctional feeding groups) was influenced bystructural and decompositional characteristicsof the riparian vegetation at different stages offorest succession; his temporal model focusesattention on streams as both patchy and dynam-ic environments.
A variety of poorly understood environmen-tal factors interact at different time scales toproduce a diverse array of patch characteristicsexhibited by riparian vegetation. We have littleperspective, for example, of how life historiesof stream organisms that are dependent on ri-parian leaf litter may be linked to evolutionaryprocesses in riparian vegetation. Plants haveevolved numerous strategies for defending theirtissues from herbivory and microbial attack.Among these defenses are physical barriers (e.g.,tough cuticle, spines, or trichomes) and sec-ondary metabolites that act as feeding deter-rents or are toxic (e.g., Feeny 1976, Harborne1982, Janzen and Rosenthal 1979, Levin 1976,Rhodes and Gates 1976). Insect herbivores have,in turn, often developed the ability to tolerateor detoxify plant defensive chemicals, frequent-ly using the chemical deterrent as a feeding cueor attractant (Ehrlich and Raven 1965, Futuyma1983). This action and reaction has been likenedto a coevolutionary "arms race" but direct evi-dence for long-term stepwise adaptations of in-teracting herbivorous insects and plants is mea-ger (Futuyma 1983). Shredder insects in streamsconsume the same leaf material that has evolveddefenses against terrestrial herbivory and mi-crobial attack (Fig. 3). There is, of course, noselective pressure on plants to evolve defensesagainst these shredders. However, shredderstream insects have presumably adapted tochanges in chemical characteristics of leaf de-tritus in both evolutionary time (responses of

514 C. M. PRINGLE ET AL. [Volume 7
VERTEBRATE HERBIVORYINSECT HERBIVORYMICROBIAL ATTACK
CHEMICAL AND STRUCTURALPLANT DEFENSES
TOXIC COMPOUNDS (E.G. ALKALOIDS)DIGESTION INHIBITORS (E.G. TANNINS)LEAF TOUGHNESS
\
CHANGES INC AND N BASEDCHEMICAL DEFENSES
SENESCENCE (DIFFERENTIALRECLAMATION OF NUTRIENTS ANDCHEMICAL DEFENSES)
CHANGES IN NUTRIENT CONTENT \LEACHING IN STREAM
VARYING NUTRIENT AVAILABILITY \MICROBIAL COLONIZATION
\NUTRITIONAL PROPERTIESOF LEAF DETRITUS
FIG. 3. Schematic diagram illustrating how plant chemical defenses and soil nutrient availability mayaffect the quality of riparian leaf litter available to shredder invertebrates in streams.
riparian plants to terrestrial herbivores and mi-crobes) and ecological time (successionalchanges in species composition of riparianvegetation).
Nutrient availability within soils or ground-waters may also influence riparian patch dy-namics and the nutritional quality of stream leaflitter (Fig. 3). Increased availability of soil nu-trients to riparian vegetation (e.g., through ag-ricultural runoff) should increase the nutrientcontent of detrital inputs into the stream. Ni-trogen (or protein) is of critical nutritional im-portance to both terrestrial herbivores (Mattson1980) and stream detritivores (Cummins andKlug 1979). Leaf nitrogen content declines withsenescence and abscission so that nitrogen con-tent of leaf detritus may be especially criticalto detritivores. Changes in nutrient status mayalso cause shifts in plant tissue concentrationsof carbon-based (e.g., phenolics) and nitrogen-based (e.g., alkaloids) defensive chemicals(Bryant et al. 1983, Chew and Rodman 1979, delMoral 1972).
While patchiness in the quantity of leaf litteravailable to a stream system is likely to be ob-vious (e.g., loss of riparian vegetation duringland development), patchiness in leaf litter
quality is likely to be widespread, complex, andnot at all obvious. Catchment management mustinclude an understanding of how qualitativepatchiness of riparian vegetation affects leaf lit-ter characteristics and hence detrital food websin streams.
A recent study by Irons et al. (1988) providesan example of how changes in the species com-position and nutrient status of riparian vege-tation might affect consumption of leaf litter byshredders. Four species of Alaskan trees com-mon in riparian areas (alder, birch, willow, andpoplar) were fertilized with nitrogen, phos-phorus, nitrogen + phosphorus, or given nofertilizer (control). Senescent leaves were of-fered to a shredder, Hydatophylax variabilis (Tri-choptera:Limnephilidae). Consumption of leafmaterial was compared among nutrient treat-ments (within tree species) and among treespecies (within nutrient treatments). Alder waspreferred over other tree species in all nutrienttreatments. Larvae preferred leaf material treat-ed with nitrogen + phosphorus for all speciesexcept poplar. Poplar leaves fertilized with ni-trogen were highly preferred whereas fertil-ization with nitrogen + phosphorus apparentlyrendered leaf material nearly inedible. Nutrient

1988] PATCH DYNAMICS IN LOTIC SYSTEMS 515
(N, P) and tannin content, and leaf toughnessdiffered among tree species and nutrient treat-ments. Consumption of leaf litter was positivelycorrelated with nitrogen content and negative-ly correlated with tannin content but was un-correlated with leaf toughness. A next step isto assess consequences of these qualitative dif-ferences in leaf detritus in terms related toshredder fitness (growth, mortality, reproduc-tion) for a wide range of shredder organisms.But the work of Irons et al. (1988) suggests thatthe benefits of riparian vegetation as bufferzones (e.g., decreasing nutrient runoff into thestream by means of nutrient uptake by riparianplants) may be offset if stream organisms de-pendent on leaf litter cannot use it efficientlybecause of excessive nutrient content, or inap-propriate ratios, or both (i.e., as derived from"fertilized" riparian trees). Thus, ecosystem-level repercussions of patch-specific phenom-ena in the riparian zone need to be addressed.For instance, how does species composition ofriparian patches interact with the nutrient sta-tus of the vegetation to affect the trophic struc-ture of stream communities? Do these interac-tions vary along tropical-temperate gradients?
Beaver-induced patch dynamics
In North America, beaver (Castor canadensis)have a strong influence on patch dynamics instream ecosystems. Since near extermination atthe beginning of this century, the populationsize of North American beaver has increaseddramatically. Although current beaver popu-lations have reached only 10-15% of the pop-ulation size before the arrival of Europeans,changes in stream patch dynamics over the last80 years have been substantial as beaver recol-onize areas where habitat and food resourceshave not been used for many decades (Naimanand Melillo 1984, Naiman et al. 1986).
Long-term population dynamics, in combi-nation with feeding and dam building activi-ties, act to produce a startling array of patchcharacteristics within the drainage network. InQuebec, Naiman and Melillo (1984) and Nai-man et al. (1986) found that beaver ponds hada' density averaging 10.6/km2 in second- andfourth-order streams, while in northern Min-nesota the number of beaver ponds on the Ka-batogama Peninsula increased from 0.15/km2 in1940 to 1.9/km2 in 1981 (Naiman, unpublished
data). These patches have significantly differentstanding stocks and fluxes of carbon and nitro-gen (Naiman and Melillo 1984, Naiman et al.1986), a different community composition(McDowell and Naiman 1986), and alternativepathways for carbon metabolism (Ford and Nai-man 1988) compared with the original stream.Beaver have once again become a key compo-nent in the structure and function of streamecosystems in the boreal forest. The overall in-fluence of beaver-created patches on the drain-age network needs to be considered in terms ofmechanisms of patch formation and patch size,distribution, density, juxtaposition, diversity,and longevity. In combination, these factors in-teract to influence the overall biogeochemicaleconomy of the drainage network.
Beaver create patches in two primary ways;by dam building and by foraging both in theimpoundment and in the adjacent riparian zone.Dam building alters the hydrologic regime, ac-cumulating material in the stream channel andcreating extensive anaerobic zones as oxygenavailability to the deep sediments is decreased(Naiman et al. 1986). Foraging by beaver is se-lective (Jenkins 1979), changing the vegetativecommunity composition and the nature andamounts of allochthonous inputs to the aquaticenvironment (Conners and Naiman 1984). Bothmechanisms of patch formation have long-termconsequences for the stream depending uponthe hydrologic regime, the response of the ri-parian vegetation to foraging, and the abilityof beaver to maintain activity at the site. Theontogeny of a beaver pond, from its formationto its eventual decay, may range from a year tomany centuries, with the physicochemical char-acteristics shifting during the aging process.When viewed in a landscape perspective, thedistribution of beaver ponds becomes a dynam-ic mosaic of constantly shifting environmentalsituations reflecting pond age and size, degreeof community maturity, substratum, hydrolog-ical characteristics, and resource inputs. Thisshifting mosaic of beaver-created habitat, adominant part of the landscape for tens of thou-sands of years (Hays 1871, Jenkins and Busker1979), overlies the natural development of thedrainage network driven by climate and geol-ogy. The changing pattern of beaver ponds hasboth spatial and temporal components. Sinceall ponds along a reach of stream are not iden-tical, specific ecosystem parameters are not spa-

L <>•""
516 C. M. PRINGLE ET AL. [Volume 7
tially constant. The result is a dynamic habitatwhere the relative importance of an ecosystemparameter varies along the channel. For ex-ample, one pond may have a primary produc-tion rate typical of a third-order stream, anotherthe rate of a ninth-order stream, while the con-necting riffles have rates of the normal second-order stream. This assumes that primary pro-duction, as well as other parameters, are notsignificantly affected by upstream ponds. Fur-ther, since beaver populations are known towax and wane over time, the changes they causewill similarly have temporal variations in den-sity and diversity. These changes may be man-ifested by changes in the internal biogeochem-ical economy of the watershed or, in addition,by changes in the total watershed budget for aparticular parameter. Historically, this shiftingmosaic, both spatially and temporally, was un-doubtedly an important determinant of biogeo-chemical economies for pristine watersheds.This complex situation is analogous to the pres-ent variety of human-induced perturbationssubjected upon stream systems. The concept ofa shifting patch mosaic offers a convenient andessential approach for quantifying patch dy-namics when biotic parameters continually ad-just to abiotic gradients, such as those naturallyoccurring along stream-river continua.
The density of beaver-created patches is a re-sponse to beaver population dynamics, with asufficient lag time to account for the return tonormal stream conditions after the habitat isabandoned. This lag time may range from de-
' cades to centuries depending upon local con-ditions (Rudemann and Schoonmaker 1938). Atpresent, for most of North America, cyclicchanges in patch density are not well knownnor are the long-term consequences well under-stood. Beaver populations in many areas arecontinuing to increase with the virtual absenceof natural predators and an abundance of earlysuccessional forage along stream channels.Consideration of the large-scale consequencesof habitat abandonment on stream stability andlong-term productivity can only be speculativeat present.
Patch dynamics of floodplains
Patch dynamics have yet to be formally ap-plied to explain the functioning of the flood-
plains of the potomon, mainly because such sys-tems have been less studied than other types offluvial systems. Sufficient information does ex-ist, however, to show the potential of the con-cept in explaining the distribution and abun-dance of fish and other biota in the lower reachesof rivers and to suggest future avenues of ex-ploration.
The potomon of most large, unmodified riverscan be thought of as existing in two major states:(1) a low flow state in which aquatic compo-nents of the system are confined to channelsand residual water bodies distributed over arelatively flat alluvial plain; and (2) a high flowstate where aquatic components of the systemfill the valley of the river to a greater or lesserextent. This means that a large proportion ofthe system alternates between a wet and a drystate at least once per year. Between these twostates are periods, usually relatively brief, dur-ing which water levels rise or fall.
Floodplains may be considered to be subjectto temporal events of four orders:
(1) Annual (or twice a year in some equatorialrivers) cycles of flood and low water;
(2) Periods of 10-102 years during which ero-sional and depositional processes producea slow evolution and succession in the var-ious components of the system. Meandersmigrate laterally reworking the sedimentsof the plain. New water bodies are createdon the floodplain, and existing ones agethrough siltation and eventually disappear;
(3) Periods of 102-103 years during which dis-turbance events, in the form of greater thanaverage floods or droughts, locally reformthe river and its floodplain; and
(4) Longer geological periods of >104 yearswhen landform processes may substantiallyalter the nature of the river basin by rivercapture, flow reversals, etc.
To these natural processes should be added ef-fects of dams, deforestation, agriculture, andother human disturbances, which have sub-stantially changed the nature of the world'srivers.
Erosional and depositional processes com-bine over time to impart a distinct texture tothe river and its floodplain (Amoros et al. 1982,Bravard et al. 1986, Salo et al. 1986) that can bedescribed in terms of patches. Individual patchescan be remarkably persistent even though they

1988] PATCH DYNAMICS IN LOTIC SYSTEMS 517
typically undergo a successional process thatmay consist of: (1) a slow transition from onestate to another, as in the silting of individualwater bodies; (2) a rapid change followed by arelatively steady state, as in the silting of theupper end of a channel transforming it into adead arm; or (3) a local catastrophic event, suchas the collapse of an eroded bank or the lateraldisplacement of a channel into a new bed. G.H. Copp (Universite Claude Bernard [Lyon],personal communication) has pointed out thatsome changes in patch structure that appear toevolve slowly, in fact proceed through a seriesof critical threshold events. Thus an anabranch(a stream leaving a river and joining it lowerdown) that supports a certain community oforganisms will switch rapidly to a differentphase that supports a totally different commu-nity as soon as its upper end is closed and itbecomes a dead arm. A similar transition to anisolated water body occurs when the lower endis closed.
Large river systems characteristically exhibitan orderly succession of features that repeatthemselves for considerable distances, oftenthousands of kilometers along the river chan-nel. Depending on slope and silt loadings, thesecan take the form of braided anabranches andislands or meanders which tend to change slow-ly with downstream progression. Some of thesefeatures are complex. In the case of a meanderbend, at least three distinct patches can be iden-tified: (1) the deepwater at the outer edge of themeander; (2) the point bar; and (3) the slackdownstream of the point bar. Several workers(Araujo-Lima et al. 1986, Junk et al. 1983) haveidentified fish community structures associatedwith such habitat patches. The nature of thesefeatures can change downstream as mean par-ticle size decreases. For instance, levees are re-duced and finally disappear. Sandy point barsare gradually replaced by mud banks. Patchstructures in the main channel can be remark-ably subtle. In the lowest reaches of the river,where the bottom sediments appear to be a ho-mogeneous cover of fine particulate substrata,differences in nutrient content can be sufficient-ly great to be detected by illiogphagous fishsuch as Prochilodus which feed on the organiccompounds absorbed onto particles. Differ-ences in concentrations of such compoundslargely explain the patchy distribution of pop-ulations of this fish taxon (Quiros and Delfino
1986). Mud-feeding fish of this type can locallyexhaust nutrient-rich patches (Bakare 1970), andpatch structure is determined by a feed-backmechanism between depositional processes andfish feeding dynamics.
Mats of floating vegetation tend to producestrong vertical gradients in temperature anddissolved oxygen in underlying waters that cansignificantly affect the distribution of fish andother organisms. Many fish species are intol-erant of low dissolved oxygen and will tend tomove to zones of higher flow and better aera-tion. Other species are equipped with auxiliarybreathing organs and may actively seek regionsof low dissolved oxygen. Thus, patches of float-ing vegetation may segregate other biota or pro-cesses. The larger the patch, the greater its ten-dency to act in this way. Floodplain poolsblocked with large masses of vegetation willoften be populated only by species that are ableto penetrate the barrier of the deoxygenatedwater under the mat. Smaller patches are lessstable and are more readily dislodged by cur-rent and carried out into the main stream.
The effect of increasing size of floodplainwater bodies (FAO/UN 1971) is also closelylinked to the capacity of vegetation mats to low-er dissolved oxygen. Small residual water bod-ies on the floodplain are often completely cov-ered with floating vegetation, and underlyingwaters are typically completely devoid of oxy-gen. As a result, the few fish species that pop-ulate them are those which are adapted tobreathe atmospheric oxygen. Somewhat largerwater bodies in the patch become less homo-geneous and areas of open water appear in whichoxygen levels are high enough to support anumber of more resistant species. Further in-creases in the size of water bodies introduce ahierarchy of patches of floating, emergent andsubmersed vegetation, and open water. This di-versity of habitat patches, and the improvedoxygen conditions arising from the effect ofwind, enable a greater variety of species to sur-vive. Further expansion in the size of waterbodies is accompanied by an equivalent in-crease in habitat and species diversity.
Few fluvial ecosystems have escaped the ac-tivities of man. Dahm et al. (1987) point out thathistorical reconstructions have shown repeat-edly that many floodplain landscapes were morecomplex and heterogeneous in space and timethan is now the case. They suggest that recon-

518 C. M. PRINGLE ET AL. [Volume 7
structions are prerequisites for understandingcontemporary patterns. Modifications of flood-plain patch dynamics can be considered as pro-ceeding in three main phases: (1) colonization,(2) utilization, and (3) destruction. Originallymost floodplains appear to have been heavilywooded (at least with gallery forests), even indrier parts of ,the world. This condition stillexists in some equatorial basins where the sub-tle interactions between trees and fish com-munities (Goulding 1980) or the adaptations tocolonize dead wood lying in river channels(Lowe-McConnell 1964) attest to the complex-ities of patch structure in unaltered environ-ments.
Early in the colonization process, human ac-tivities progressively reduced the forest coverand replaced much of the woodland with thesavannah plains that are so typical of Africanand South American rivers today. This processled to drastic changes in the ecosystems whichwere aggravated as the size of woodland patchesdecreased (e.g., Alho et al. 1988). Greater insta-bility was introduced into the physical systemwith an acceleration of erosion-depositioncycles. More long-lasting and stable structuralpatches (trees) were replaced by mosaics offloating, emergent, and submersed vegetation.Increased insolation led to higher temperaturesand to more rapid recycling of organic matter.Phyto- and zooplankton increased in abun-dance and floating grasses became the domi-nant form of primary production. These changeswere probably accompanied by changes in fishpopulation structure that have not been docu-mented. However, more recent observations(e.g., Goulding 1980) indicate the types ofchanges that may have occurred. Decreases inthe size of woodland patches leads to the de-cline in the number of frugivorous species.Likewise, the disappearance of wood fromstreambeds leads to a diminution of species de-pendent on such habitats for refuge.
During the utilization phase, river channelsand floodplains are subject to intensive ruraluse involving agriculture, fisheries, cattle rear-ing, and even human occupation (e.g., Alho etal. 1988). Use of the plain can be highly orga-nized while the flood pattern is maintained.Traditional values of human communities at thislevel of use often show a deep understandingof the role of patch structure of floodplains and
many features are adapted to maximize pro-duction. Examples include plantations of float-ing rice, patches of floating vegetation that areused as devices to concentrate fish for later cap-ture, and floodplain water bodies that are adapt-ed as drains in fish ponds. Further modificationsto the structure of floodplain systems have re-sulted from the introduction of new organisms.The introduction of floating weeds such as Sal-vinia, Azolla, and Eichomia have frequently ledto changes in nutrient cycling, with massiverepercussions throughout the system. On asmaller scale, introduction of a fish such as thegrass carp (Ctenapharyngodon idellus) can elimi-nate patches of submerged vegetation resultingin a disappearance of water bodies, or conver-sion in primary production to phytoplanktonand shifts in the species composition of com-munities of aquatic fauna.
During the destruction phase, the character-istics of the river and its floodplain are so al-tered by channelization, drainage, poldering,and urbanization that few of the original fea-tures of the system remain. The breakdown ofpatch structures essential to the breeding andfeeding of many species results in a simplifi-cation of the fish community and greatly low-ered productivity.
An understanding of various elements of flu-vial ecosystems and the way in which they re-late to one another is crucial to effective man-agement. Such knowledge is needed both at theutilization level, where maintenance of the cor-rect balance of water level and water use canpermit sustained harvesting of a number ofaquatic and terrestrial food organisms, and atthe destruction level, where similar informa-tion is needed to mitigate changes and restorewhat has been lost.
Studies on energy flow and structural dynam-ics of biota between and among terrestrial, ri-parian, and riverine patches, as superimposedon the floodplain surface, are needed. Compar-isons of temporal dynamics of floodplain pro-cesses in desert, forested, and savannah (i.e.,transition between forested and drier biomes)environments may be most instructive. In par-ticular, it is vital to document how floodplain-dependent fishes are influenced by intensityand duration of flood periods. Conservation ofthe few remaining pristine big river floodplainscannot occur without this information (Wei-
' j

1988] PATCH DYNAMICS IN LOTIC SYSTEMS 519
comme 1988—see this issue). A patch dynamicsapproach may offer a new synthesis for suchstudies.
Conclusions and recommendationsIn addition to specific recommendations pre-
sented with the preceding discussion topics, weoffer the following conclusions and recommen-dations in hopes of facilitating the applicationof a patch-dynamics perspective to problems inlotic ecology, and stimulating ideas and ap-proaches for future study:
(1) A patch dynamics approach has the poten-tial to yield information central to the ef-fective management and long-term survivalof lotic systems by increasing our under-standing of the role of various elements instreams and the way in which they relateto one another.
(2) The effectiveness of a patch-dynamics ap-proach as a framework for the study of loticsystems lies in the strength of the linkagebetween reductionist and whole-systemperspectives.
(3) Examination of patch-dynamics can en-hance the utility and predictive power ofunifying concepts in lotic ecology such asthe river continuum hypothesis (Vannoteet al. 1980) and nutrient spiralling (New-bold et al. 1981) through its focus on or-ganism and process-specific building blocksof lotic systems. Examination of patches andprocess-oriented interactions betweenpatches will allow quantitative comparisonsto be made within and between stream sys-tems.
(4) Patch-to-patch variability in lotic systemsshould be viewed as information, rather thanstatistical noise to be overcome with ade-quate sample size. This concept needs to beapplied over extremes in spatial and tem-poral scales, from the response of microbialcommunities to nutrient patches on a scaleof /im and milliseconds, to differences be-tween catchments in different biomes.
(5) With respect to tropical-temperate compar-isons, more data are necessary from a varietyof lotic systems within given geographicalareas; variability within biomes must be ex-amined before cross-biome comparisons canbe made. Broad and often inaccurate gen-
eralizations regarding properties of tropicalecosystems have generated restrictive as-sumptions regarding functional character-istics of tropical streams.
(6) A patch perspective is recommended as anideal framework to test Tilman's (1982) con-sumer resource theories as they apply tolotic systems. The study of patch dynamics.,with a landscape perspective, also providesa useful perspective with which to examinelittle-understood biotic/abiotic interactionsin streams (e.g., Power et al. 1988—see thisissue).
(7) A greater emphasis should be placed uponthose characteristics of a study organism thatallow it to perceive and affect patch-specificphenomena or variation in its environment.
Acknowledgements
We thank Jack A. Stanford and Alan C. Co-vich for organizing the symposium. The writ-ing of the paper was supported in part by Na-tional Science Foundation grant BSR-87-17746to CMP.
Literature Cited
ALHO, C. J. R., T. E. LUCHER, AND H. C. GONCALVES.1988. Environmental degradation in the Panta-nal ecosystem. BioScience 38:164-171.
ALLEN, T. F. H. 1977. Scale in microscopic algal ecol-ogy: a neglected dimension. Phycologia 16:253-257.
AMOROS, C., M. RICHARDOT-COULET, AND G. PATOU.1982. Les ensembles fonctionelles: des entilesecologiques qui traduisent 1'evolution de ITiy-drosysteme en integrant le geomorphologie et1'anthropisation (exemple de Haul Rh6ne fran-cais). Revue Geographic de Lyon 57:49-62.
ANDERSON, N. H., AND J. R. SEDELL. 1979. Detritusprocessing by macroinvertebrates in stream eco-systems. Annual Review of Entomology 24:351-377.
ANGERMEER, P. L., AND J. R. KARR. 1984. Relation-ships between eroding debris and fish habitat ina small stream. Transactions of the AmericanFisheries Society 113:716-726.
ARAUjo-LiMA, A. R. M., L. P. S. PORTUGAL, AND E. G.FERRHRA. 1986. Fish-macrophyte relationshipsin the Anavihanas Archipelago, a black watersystem in the Central Amazon. Journal of FishBiology 29:1-11.
AUMEN, N. G., P. J. BOTTOMLEY, AND S. V. GREGORY.1985. Nitrogen dynamics in stream wood sam-
I

520 C. M. PRINGLE ET AL. [Volume 7
pies incubated with [14C]lignocellulose and po-tassium [15N]nitrate. Applied and EnvironmentalMicrobiology 49:1119-1123.
BAKARE, O. 1970. Bottom deposits as food for inlandfreshwater fish. Pages 65-85 in S. A. Visser (ed-itor). Kaiji Lake studies, Volume 1, Ecology. Iba-dan University Press, Ibadan.
BILBY, R. E. 1981. Role of organic debris dams inregulating the export of dissolved and particulatematter from a forested watershed. Ecology 62:1234-1243.
BRAVARD, J. P., C. AMOROS, AND G. PATOU. 1986. Im-pact of civil engineering works on the succes-sions of communities in a fluvial system. Oikos47:92-111.
BRYANT, J. P., F. S. CHAPIN, AND D. R. KLEIN. 1983.Carbon/nutrient balance of boreal plants in re-lation to vertebrate herbivory. Oikos 40:357-368.
CHEW, F. S., AND J. E. RODMAN. 1979. Plant resourcesfor chemical defense. Pages 271-307 in G. A. Ro-senthal and D. H. Janzen (editors). Herbivores:their interaction with secondary plant metabo-lites. Academic Press, New York.
CONNELL, J. 1978. Diversity in tropical rainforestsand coral reefs. Science 199:1302-1310.
CONNERS, M. E., AND R. J. NAIMAN. 1984. Particulateallochthonous inputs: relationships with streamorder in an undisturbed watershed. CanadianJournal of Fisheries and Aquatic Sciences 41:1473-1484.
CROSS, F. B. 1970. Fishes as indicators of pleistoceneand recent environments in the central plains.Pages 241-257 in Pleistocene and recent environ-ments of the central great plains. University ofKansas, Special Publication No. 3, Lawrence.
CUMMINS, K. W., AND M. J. KLUG. 1979. Feedingecology of stream invertebrates. Annual Reviewof Ecology and Systematics 10:147-172.
DAHM, C. N., E. H. TROTTER, AND J. R. SEDELL. 1987.Role of anaerobic zones and processes in streamecosystem productivity. Pages 157-178 in R. C.Averett and D. M. McKnight (editors). Chemicalquality of water and the hydrologic cycle. LewisPublishers, Inc., Chelsea, Michigan.
DEL MORAL, R. 1972. On the variability of chloro-genic acid concentration. Oecologia 9:289-300.
EHRUCH, P. R., AND P. H. RAVEN. 1965. Butterfliesand plants: a study in coevolution. Evolution 18:586-608.
ELWOOD, J. W., J. D. NEWBOLD, R. V. O'NEILL, AND W.VANWINKLE. 1983. Resource spiralling: an op-erational paradigm for analyzing lotic ecosys-tems. Pages 3-27 in T. D. Fontaine and S. M. Bar-tell (editors). Dynamics of lotic ecosystems. AnnArbor Science, Ann Arbor, Michigan.
ELWOOD, J. W., J. D. NEWBOLD, A. F. TRIMBLE, AND R.W. STARK. 1981. The limiting role of phos-phorus in a woodland stream ecosystem: effect
of P enrichment on leaf decomposition and pri-mary producers. Ecology 62:146-158.
FAGEN, O. F. 1962. Influence of stream stability onhoming behavior of two smallmouth bass pop-ulations. Transactions of the American FisheriesSociety 91:346-349.
FEENY, P. 1976. Plant apparency and chemical de-fense. Pages 1-40 in J. W. Wallace and R. L. Man-sell (editors). Biochemical interactions betweenplants and insects. Plenum Press, New York.
FISHER, S. G., AND G. E. LIKENS. 1973. Energy flowin Bear Brook, New Hampshire: an integrativeapproach to stream ecosystem metabolism. Eco-logical Monographs 43:421-439.
FOOD AND AGRICULTURE ORGANIZATION OF THE UNITEDNATIONS. 1971. Rapport au Gouvernement duDahomey sur 1'evolution de la peche interieur,son etat actuel et ses possibilites, establi sur labase des travaux de R. L. Welcomme, Specialistede la peche. Report Number FAO/PNUD(AT)(2938), Rome.
FORD, T. E., AND R. J. NAIMAN. 1988. Alteration ofcarbon cycling by beaver: methane evasion ratesfrom boreal forest streams and rivers. CanadianJournal of Zoology 66:529-533.
FORMAN, R. T. T. 1983. An ecology of the landscape.BioScience 33:535.
FORMAN, R. T. T., AND M. GODRON. 1981. Patchesand structural components for a landscape ecol-ogy. BioScience 31:733-740.
FORMAN, R. T. T., AND M. GODRON. 1986. Landscapeecology. John Wiley & Sons, New York.
FRASER, D. F., AND T. E. SISE. 1980. Observations onstream minnows in a patchy environment: a testof a theory of habitat distribution. Ecology 61:790-797.
FUTUYMA, D. J. 1983. Evolutionary interactionsamong herbivorous insects and plants. Pages 207-231 in D. J. Futuyma and M. Slatkin (editors).Coevolution. Sinauer Associates, Inc., Sunder-land, Massachusetts.
GLODEK, G. S. 1978. The importance of catfish bur-rows in maintaining fish populations of tropicalfreshwater streams in Western Ecuador. Fieldi-ana Zoology 73:1-8.
GOLLADAY, S. W., J. E. WEBSTER, AND E. F. BENFIELD.1983. Factors affecting food utilization by a leafshredding aquatic insect: leaf species and con-ditioning time. Holarctic Ecology 6:157-162.
GORMAN, O. T., AND J. R. KARR. 1978. Habitat struc-ture and stream fish communities. Ecology 59:507-515.
GOULDING, M. 1980. The fish and the forest: explo-rations in Amazonian natural history. Universityof California Press, Berkeley.
GREGORY, S. V. 1978. Phosphorus dynamics on or-ganic and inorganic substrates in streams. Ver-handlungen der Internationalen Vereinigung fur

1988] PATCH DYNAMICS IN LOTIC SYSTEMS 521
Theoretische und Angewandte Limnologie 20:1340-1346.
HAPPEY-WOOD,C.,ANDJ.PRIDDLE. 1984. The ecologyof epipelic algae of five Welsh lakes with specialreference to volvocalean green flagellates (Chlo-rophyceae). Journal of Phycology 20:109-124.
HARBORNE, J. B. 1982. Introduction to ecological bio-chemistry. Second edition. Academic Press, NewYork.
HARIMA, H., AND P. R. MUNDY. 1974. Diversity in-dices applied to the fish biofacies of a small stream.Transactions of the American Fisheries Society103:457-461.
HART, D. D. 1981. Foraging and resource patchiness:field experiments with a grazing stream insect.Oikos 37:46-52.
HART, D. D. 1985. Causes and consequences of ter-ritoriality in a grazing stream insect. Ecology 66:404-414.
HART, D. D., AND V. H. RESH. 1980. Movement pat-terns and foraging ecology of a stream caddisflylarva. Canadian Journal of Zoology 58:1174-1185.
HAYS, W. J. 1871. Notes on the range of some of theanimals in America at the time of the arrival ofthe white men. American Naturalist 5:25-30.
HERSHEY, A. E., A. L. HILTNER, M. A. HULLAR, M. C.MILLER, J. R. VESTAL, M. A. LOCK, S. RUNDLE, ANDB. J. PETERSON. 1988. Nutrient influences on astream grazer: Orthocladius microcommunities re-spond to nutrient input. Ecology 69:1383-1392.
HILL, B. H. 1979. Uptake and release of nutrients byaquatic macrophytes. Aquatic Botany 7:87-93.
HOWARTH, R. W., AND S. G. FISHER. 1976. Carbon,nitrogen and phosphorus dynamics during leafdecay in nutrient-enriched stream microecosys-tems. Freshwater Biology 6:221-228.
HUNT, R. L. 1971. Responses of a brook trout pop-ulation to habitat development in LawrenceCreek. Wisconsin Department of Natural Re-sources, Madison, Technical Bulletin No. 48.
HUTCHINSON, G. E. 1975. A treatise on limnology.II. John Wiley and Sons, New York.
HYNES,H. B. N. 1970. The ecology of running waters.University of Toronto Press, Toronto.
IRONS, J. G., M. W. OSWOOD, AND J. P. BRYANT. 1988.Consumption of leaf detritus by a stream shred-der: influence of tree species and nutrient status.Hydrobiologia 160:53-61.
JACKSON, R. J. 1987. Hydrology of an acid wetlandbefore and after draining for afforestation, west-ern New Zealand. Pages 465-474 in Forest hy-drology and watershed management (Proceed-ings of the Vancouver symposium, August 1987),I.H.A.S. Publication No. 167.
JANZEN, D. A., AND G. A. ROSENTHAL. 1979. Herbi-vores: their interaction with plant secondary me-tabolites. Academic Press, New York.
JENKINS, S. H. 1979. Seasonal and year-to-year dif-ferences in food selection by beavers. Oecologia44:112-116.
JENKINS, S. H., AND P. E. BUSKER. 1979. Castor cana-densis. Mammalian Species 120:1-9.
JOHNSTON, C. A., AND R. J. NAIMAN. 1987. Boundarydynamics at the aquatic terrestrial interface: theinfluence of beaver and geomorphology. Land-scape Ecology 1:47-57.
JORGENSEN, E. G. 1957. Diatom periodicity and sili-con assimilation. Dansk Botanisk Arkiv 18:1-54.
JUNK, W. J., G. M. SCARES, AND F. M. CARVALHO. 1983,Distribution of fish species in a lake of the Am-azon river floodplain (Lago Camaleao), with spe-cial reference to extreme oxygen conditions.Amazoniana 7:397-431.
KARR, J. R., AND D. R. DUDLEY. 1981. Ecological per-spectives on water quality goals. EnvironmentalManagement 5:55-68.
KARR, J. R., AND I. J. SCHLOSSER. 1978. Water re-sources and the land-water interface. Science 201:229-234.
KAUSHDC, N. K., AND H. B. N. HYNES. 1968. Experi-mental study on the role of autumn shed leavesin aquatic environments. Journal of Ecology 56:229-245.
KAUSHK, N. K., AND J. B. ROBINSON. 1976. Prelimi-nary observations on nitrogen transport duringsummer in a small spring-fed Ontario stream.Hydrobiologia 49:59-63.
KOHLER, S. L. 1984. Search mechanisms of a streamgrazer in patchy environments: the role of foodabundance. Oecologia 62:209-218.
KOHLER, S. L. 1985. Identification of stream driftmechanisms: an experimental and observationalapproach. Ecology 66:1749-1761.
LEVIN, S. A. 1976. The chemical defenses of plantsto pathogens and herbivores. Annual Review ofEcology and Systematics 7:121-159.
LEVIN, S. A., AND R. T. PAINE. 1974. Disturbance,patch formation and community structure. Pro-ceedings of the National Academy of Sciences,USA 71:2744-2747.
LOCK, M. A., AND P. H. JOHNS. 1979. The effect offlow pattern on uptake of phosphorus by riverphytoplankton. Limnology and Oceanography29:376-382.
LowE-McCONNELL, R. H. 1964. The fishes of the Ru-pununi savannah district of British Guiana, SouthAmerica. Part 1. Ecological groupings of fishspecies and effects of the seasonal cycle on thefish. Journal of the Linnean Society of London(Zoology) 45:103-144.
MATTHEWS, W. J. 1988. North American prairiestreams as systems for ecological study. Journalof the North American Benthological Society 7:387-409.
MATTSON, W. J. 1980. Herbivory in relation to plant

522 C. M. PRINGLE ET AL. [Volume 7
nitrogen content. Annual Review of Ecology andSystematics 11:119-161.
MCARTHUR, J. V., G. R. MASZOLF, AND J. E. URBAN.1985. Response of bacteria isolated from a pris-tine prairie stream to concentration and sourceof soluble organic carbon. Applied and Environ-mental Microbiology 49:238-241.
MCDOWELL, D.M., AND R.J.NAIMAN. 1986. Structureand function of a benthic invertebrate streamcommunity as influenced by beaver (Castor can-adensis). Oecologia 68:481-489.
MclNTffiE, C. D. 1966. Some effects of current ve-locity on periphyton communities in laboratorystreams. Hydrobiologia 27:557-570.
MEULLO, J. M., R. J. NAIMAN, J. D. ABER, AND K. N.ESHLEMAN. 1983. The influence of substratequality and stream size on wood decompositiondynamics. Oecologia 58:281-285.
MEYER, J. L. 1979. The role of sediments and bryo-phytes in phosphorus dynamics in a headwaterstream ecosystem. Limnology and Oceanography24:365-376.
MEYER, J. L. 1980. Dynamics of phosphorus and or-ganic matter during leaf decomposition in a for-est stream. Oikos 34:44-53.
MEYER, J. L., W. H. MCDOWELL, T. L. Borr, J. W. ELWOOD,C. IsmzAia, J. M. MELACK, B. L. PECKARSKY, B.PETERSON, AND P. RUBLEB. 1988. Elemental dy-namics in streams. Journal of the North AmericanBenthological Society 7:410-432.
MINSHALL, G. W. 1978. Autotrophy in stream eco-systems. BioScience 28:767-771.
MINSHALL, G. W., K. W. CUMMINS, R. C. PETERSEN, C.E. CUSHING, D. A. BRUNS, J. R. SEDELL, AND R. L.VANNOTE. 1985. Developments in stream eco-system theory. Canadian Journal of Fisheries andAquatic Sciences 42:1045-1055.
MINSHALL, G. W., R. C. PETERSEN, K. W. CUMMINS, T.L. BOTT, J. R. SEDELL, C. E. CUSHING, AND R. L.VANNOTE. 1983. Interbiome comparison ofstream ecosystem dynamics. Ecological Mono-graphs 53:1-25.
MOLLES, M. C. 1982. Trichopteran communities ofstreams associated with aspen and conifer forests:long-term structural change. Ecology 63:1-6.
MULHOLLAND, P. J., J. D. NEWBOLD, J. W. ELWOOD, L.A. FERREN, AND J. R. WEBSTER. 1985. Phosphorusspiralling in a woodland stream: seasonal varia-tions. Ecology 66:1012-1023.
MYROLD, D. D., J. R. Gosz, AND J. M. TIEDJE. 1982.Moisture gradient effects on denitrification rateswithin a non-aggrading aspen site. Bulletin ofthe Ecological Society of America 63:95.
NAIMAN, R. J., H. DECAMPS, J. PASTOR, AND C. A.JOHNSTON. 1988. The potential importance ofboundaries to fluvial ecosystems. Journal of theNorth American Benthological Society 7:289-306.
NAIMAN, R. J., AND J. M. MEULLO. 1984. Nitrogen
budget of a subarctic stream altered by beaver(Castor canadensis). Oecologia 62:150-155.
NAIMAN, R. J., J. M. MEULLO, AND J. F. HOBBIE. 1986.Ecosystem alteration of boreal forest streams bybeaver (Castor canadensis). Ecology 67:1254-1269.
NAIMAN, R. J., J. M. MEULLO, M. A. LOCK, T. E. FORD,AND S. R. REICH. 1987. Longitudinal patterns ofecosystem processes and community structure ina subarctic river continuum. Ecology 68:1139-1156.
NEWBOLD, J. D., J. W. ELWOOD, R. V. O'NEILL, AND A.L. SHELDON. 1983. Phosphorus dynamics in awoodland stream ecosystem: a study of nutrientspiralling. Ecology 64:1249-1265.
NEWBOLD, J. D., J. W. ELWOOD, R. V. O'NEILL, AND W.VAN WINKLE. 1981. Measuring nutrient spiral-ling in streams. Canadian Journal of Fisheriesand Aquatic Sciences 38:860-863.
Noss, R. F. 1983. A regional landscape approach tomaintain diversity. BioScience 33:700-706.
ORIANS, G. H. 1980. Micro and macro in ecologicaltheory. BioScience 30:79.
PAINE, R. T., AND S. A. LEVIN. 1981. Intertidal land-scapes: disturbance and the dynamics of pattern.Ecological Monographs 51:145-178.
PASOAK, W. J., AND J. GAVIS. 1974. Transport limi-tations of nutrient uptake in phytoplankton.Limnology and Oceanography 19:881-888,
PECKARSKY, B. L. 1985. Do predaceous stoneflies andsiltation affect the structure of stream insect com-munities colonizing enclosures? Canadian Jour-nal of Zoology 63:1519-1530.
PETERJOHN,W. T.,ANDD.L.CORRELL. 1984. Nutrientdynamics in an agricultural watershed: obser-vation of a riparian forest. Ecology 65:1466-1475.
PETERSON, B. J., J. E. HOBBIE, T. L. CORLISS, AND K.KRIET. 1983. A continuous flow periphytonbioassay: tests of nutrient limitation in a tundrastream. Limnology and Oceanography 28:583-591.
PICKETT, S. T. A., AND P. S. WHITE. 1985. The ecologyof natural disturbance and patch dynamics. Ac-ademic Press, Orlando, Florida.
POWER, M. E. 1984. Depth distribution of armoredcatfish: predation induced resource avoidance.Ecology 65:523-528.
POWER, M. E., R. J. STOUT, C. E. CUSHING, P. P. HARPER,F. R. HAUER, W. J. MATTHEWS, P. B. MOYLE, B.STATZNER,ANDI.R.WAISDEBADGEN. 1988. Biot-ic and abiotic controls in river and stream com-munities. Journal of the North American Ben-thological Society 7:456-479.
PRINGLE, C. M. 1985a. Effects of chironomid (Insecta:Diptera) tube-building activities on stream dia-tom communities. Journal of Phycology 21:185-194.
PRINGLE, C. M. 1985b. Nutrient heterogeneity andthe maintenance of species diversity: periphytonresponse to substratum and water enrichment in

1988] PATCH DYNAMICS IN LOTIC SYSTEMS 523
a nutrient-poor stream. Ph.D. Dissertation, Uni-versity of Michigan, Ann Arbor.
PRINGLE, C. M. 1987. Effects of water and substratumnutrient supplies on lotic periphyton growth: anintegrated bioassay. Canadian Journal of Fish-eries and Aquatic Sciences 44:619-629.
PRINGLE, C. M., AND J. A. BOWERS. 1984. An in situsubstratum fertilization technique: diatom colo-nization on nutrient-enriched sand substrata. Ca-nadian Journal of Fisheries and Aquatic Sciences41:1247-1251.
PRINGLE, C M., P. PAABY, P. D. VAUX, AND C. GOLDMAN.1986. In situ nutrient assays of periphyton growthin a lowland Costa Rican stream. Hydrobiologia134:207-213.
QUIROS, R., AND R. DELFINO. 1986. Estimation em-pirica de la abundantia de peces en subambientesdel embalse de Salto Grande. Revista de la Aso-ciacion de Ciencias Naturales del Literal 16:199-214.
REICE, S. R. 1974. Environmental patchiness and thebreakdown of leaf litter in a woodland stream.Ecology 55:1271-1282.
REICE, S. R. 1977. The role of animal associations andcurrent velocity in sediment-specific leaf litterdecomposition. Oikos 29:357-365.
REICE, S. R. 1980. The role of substratum in benthicmacroinvertebrate microdistribution and litterdecomposition in a woodland stream. Ecology 61:580-590.
REICE, S. R. 1985. Experimental disturbance and themaintenance of species diversity in a stream com-munity. Oecologia 67:90-97.
REYNOLDS, C. S. 1984. The ecology of freshwaterphytoplankton. Cambridge University Press, NewYork.
RHODES, D. P., AND R. G. GATES. 1976. Toward ageneral theory of plant antiherbivore chemistry.Pages 168-213 in J. W. Wallace and R. L. Mansell(editors). Biochemical interactions between in-sects and plants. Plenum Press, New York.
RISSER, P. G., J. R. KARR, AND R. T. FORMAN. 1984.Landscape ecology: directions and approaches.Illinois Natural History Survey, Champaign,Special Publication No. 2.
RUDEMANN, R., AND W. J. SCHOONMAKER. 1938. Bea-ver dams as geologic agents. Science 88:523-525.
SAETHER, O. A. 1968. Chironomids of the Finse Area,Norway, with special reference to their distri-bution in a glacier brook. Archiv fur Hydrobio-logie 64:426-488.
SALO, J., R. KALUOLA, I. HAKMNEN, Y. MAMNEN, P.NffiMELA, M. PUHAKKA, AND P. D. COLBY. 1986.River dynamics and the diversity of Amazon low-land forest. Nature 322:254-258.
SHELDON, A. L. 1968. Species diversity and longi-tudinal succession in stream fishes. Ecology 49:193-198.
SHELDON, A. L. 1984. Colonization dynamics ofaquatic insects. Pages 401-429 in V. H. Resh andD. M. Rosenberg (editors). The ecology of aquaticinsects. Praeger Publishers, New York.
SOUSA, W. P. 1985. Disturbance and patch dynamicson rocky intertidal shores. Pages 101-124 in S. T.A. Pickett and P. S. White (editors). The ecologyof natural disturbance and patch dynamics. Ac-ademic Press, New York.
STANFORD, J. A., AND J. V. WARD. 1986. Fish of theColorado system. Pages 385-402 in B. R. Daviesand K. F. Walker (editors). The ecology of riversystems. Dr. W. Junk Publishers, Dordrecht.
STATZNER, B. 1987. Characteristics of lotic ecosys-tems and consequences for future research di-rections. Pages 365-390 in E.-D. Schulze and H.Zwolfer (editors). Potentials and limitations ofecosystem analysis. Ecological Studies 61, Spring-er-Verlag, New York.
STATZNER, B., J. A. GORE, AND V. H. RESH. 1988. Hy-draulic stream ecology: observed patterns andpotential applications. Journal of the NorthAmerican Benthological Society 7:307-360.
STATZNER, B., AND B. HIGLER. 1986. Stream hydrau-lics as a major determinant of benthic inverte-brate zonation patterns. Freshwater Biology 16:127-139.
STOCKNER, J. G., AND K. R. S. SHORTREED. 1978. En-hancement of autotrophic production by nu-trient addition in a coastal rainforest stream onVancouver Island. Journal of the Fisheries Re-search Board of Canada 35:28-34.
SWANK, W. T., AND W. H. CASKEY. 1982. Nitrate de-pletion in a second-order mountain stream. Jour-nal of Environmental Quality 11:581-584.
THOMPSON, J. N. 1978. Within patch structure anddynamics in Pastinaca saliva and resource avail-ability to a specialized herbivore. Ecology 59:443-448.
TILMAN, D. 1982. Resource competition and com-munity structure. Monographs in Population Bi-ology No. 17. Princeton University Press, Prince-ton, New Jersey.
TRISKA, F. J., AND BUCKLEY, B. M. 1978. Patterns ofnitrogen uptake and loss in relation to Utter dis-appearance and associated invertebrate biomassin six streams of the Pacific Northwest, USA. Ver-handlungen der Internationalen Vereinigung furTheoretische und Angewandte Limnologie 20:1324-1332.
TRISKA, F. J., AND K. CROMACK. 1980. The role ofwood debris in forests and streams. Pages 171-190 in R. H. Waring (editor). Forests: fresh per-spectives from ecosystem analysis. Oregon StateUniversity Press, Corvallis, Oregon.
TRISKA, F. J., V. C. KENNEDY, R. J. AVANZINO, AND B.N. REILLY. 1983. Effect of simulated canopy cov-er on regulation of nitrate uptake and primary

524 C. M. PRINGLE ET AL. [Volume 7
production by natural periphyton assemblages.Pages 129-159 in T. D. Fontaine and S. M. Bartell(editors). Dynamics of lotic ecosystems. Ann Ar-bor Science, Ann Arbor, Michigan.
TRISKA, F. J., J. R. SEDELL, K. CROMACK, S. V. GREGORY,AND F. McCoRlsoN. 1984. Nitrogen budget fora small coniferous forest stream. EcologicalMonographs 54:119-140.
URBAN, D. L., R. V. O'NEILL, AND H. H. SHUGART.1987. Landscape ecology. BioScience 37:119-127
VANNOTE, R. L., G. W. MINSHALL, K. W. CUMMINS, J.R. SEDELL, AND C. E. GUSHING. 1980. The rivercontinuum concept. Canadian Journal of Fish-eries and Aquatic Sciences 37:130-137.
VINCENT, W. F., AND M. T. DOWNES. 1980. Variationin nutrient removal in a stream by watercress(Nasturtium offidnale R. Br.). Aquatic Botany 9:221-235.
VrrousEK, P. M. 1985. Community turnover and eco-system nutrient dynamics. Pages 325-334 in S. T.A. Pickett and P. S. White (editors). The ecologyof natural disturbance and patch dynamics. Ac-ademic Press, New York.
WEBSTER, J. R. 1975. Analysis of potassium and cal-cium dynamics in stream ecosystems on threesouthern Appalachian watersheds of contrastingvegetation. Ph.D. Dissertation, University ofGeorgia, Athens.
WEBSTER, J. R., AND B. C. PATTEN. 1979. Effects ofwatershed perturbation on stream potassium andcalcium dynamics. Ecological Monographs 49:51-72.
WELCOMME, R. 1988. Concluding remarks I: On thenature of large tropical river, floodplains, andfuture research directions. Journal of the NorthAmerican Benthological Society 7:525-526.
WERNER, D. 1977. Silicate metabolism. Pages 110-149 in D. Werner (editor). The biology of diatoms.Blackwell Scientific Publications, Oxford.
WETZEL, R. G., AND G. E. LIKENS. 1979. Limnologicalanalyses. W. B. Saunders Company, Philadelphia.
WHTTFORD, L. A., AND G. J. SCHUMACHER. 1964. Effectof a current on respiration and mineral uptakein Spirogyra and Oedogonium. Ecology 45:168-170.
WICKSTROM, C. E., AND R. G. WiEGERT. 1980. Re-sponse of thermal algal-bacterial mat to grazingby brine flies. Microbial Ecology 6:303-315.
WIENS, J. A., C. S. CRAWFORD, AND J. R. Gosz. 1985.Boundary dynamics: a conceptual framework forstudying landscape ecosystems. Oikos 45:421-427.
WILEY, M.J. 1976. The ecology of the Chironomidae(Diptera:Chironomidae) of Hunt Creek with spe-cial reference to substrate relations. Masters The-sis, University of Michigan, Ann Arbor.


















