Skeletal Muscle Skeletal Muscle Anatomy Figure 12-3a-2: ANATOMY SUMMARY: Skeletal Muscle.
Upload
duongxuyenCategory
view
218download
0
Mammalian Skeletal MusclePHYSIOLOGY & BIOENERGETICSRevision NotesA “Zoology afresh” initiativehttps://www.facebook.com/ZoologyOption/ IAS ZoologyOption Syllabus Types, mechanism of contraction of skeletal muscles, effects of exercise on muscles
IAS Previous Years’ Questions on Muscles
2015 : Explain with diagrams the molecular mechanism contraction of skeletal muscle. Discuss the role of ions and bioenergetics in muscle contraction. Attribute the cause(s) for muscle cramps. [20 marks]
2012 : Draw the diagram to show the arrangements of various types of contractile proteins in skeletal muscles, and explain the bioenergetics of muscle contraction. [20 marks]
2009 : Write short notes on molecular mechanism of skeletal muscle contraction. [15 marks]
RESOURCE NOTE UPDATE VERSION : MUSCLES_001_08082017
TOPICS COVERED IN THIS RESOURCE
Mammalian Skeletal Muscle Tissue : General Introduction & Types Skeletal Muscle Fibre Types (red, white & intermediate) Microscopic Anatomy of a Skeletal Muscle Fiber Filaments and the Sarcomere Muscle Proteins (structural & functional) The Contraction Cycle and Excitation–Contraction Coupling Contraction & Relaxation of Skeletal Muscle Fibers The Sliding Filament Mechanism The Neuromuscular Junction : Innervation (En plaque, en grappe) Muscle Metabolism

Production of ATP in Muscle Fibers, Creatine Phosphate Muscle Fatigue Motor Unit, Muscle Tone, Muscle Twitch, Latent Period, Summation
(complete & incomplete), Tetanus, Rigor mortis. Muscle & Exercise : Isotonic & Isometric Contractions Skeletal Muscle Hypertrophy, atrophy & dystrophy Muscle Diseases : Muscular Dystrophy, Tetanus Neuromuscular Diseases : Motorneuron Disease, Paralysis
Skeletal muscle types, based upon morphologyFusiform, bipinnate, pinnate (or pennate, or unipinnate), digastric, bicipital, cross-banded tendinous muscles, parallel, convergent, sphincter (circular). Not important
Skeletal muscle types based on the ‘biochemical profile’ of their constituent fibres (discuss over phone)Red, white & intermediate muscles (and muscle fibres) – oxidative (aerobic) and non-oxidative (non-anerobic or glycolytic) muscle
Skeletal muscles (or muscle groups) as per their specific functional actionExtensors, flexors, rotators, supinators, adductors, abductors, agonistic, antagonistic & synergistic muscles
Skeletal muscle types based upon their contractile characteristicsPhasic, tonic, fast-twitch & slow-twitch muscles; catch muscles & fibrillary muscles (in invertebrates) – the molluscan catch-mechanism
Be familiar the following terms (discuss over phone)Motor Unit, All or None Principle in relation to a muscle, Sensory muscle fibres (muscle spindles) – the intrafusal & extrafusal muscle fibre terminology
Detailed structure of a skeletal muscle as we proceed from a muscle surface to its interior (towards the muscle fibres)
Structure of a muscle fibre – from microscopic to ultrastructural to – detailed structure of a sarcomere
Transverse-tubular system in a skeletal muscle fibre. The TRIAD. Excitation-contraction coupling – basics of initiation & end of muscular contraction

Structural & functional integrity of Skeletal Muscles is always dependent upon the motor nerve innervating them, and, in fact, motor nerve fibres and the skeletal muscle fibres it is innervating always work TOGETHER as an integrated entity called a ‘motor unit’.
Motor nerve fibres end up on the surface of skeletal muscle fibres as ‘motor end-plates’ … if the nerve fibre ends in a single ‘plate-like’ thickening, such ending is referred to as “en plaque’ type of innervation, and if it end after repeated fibre-end divisions into a buch on small end-plates on the sarcolemma, then the ending is referred to as “en grappe’ type of innervation.
A motor nerve fibre PLUS all the skeletal muscle fibres innervated by it, or its terminal branches, is called a “MOTOR UNIT”. Characteristically, all the members skeletal muscle fibres are all alike – that is, either all are RED type, or all are WHITE type … they can never belong to BOTH red & white types. Whenever a stimulus activates a motor unit, all its component muscle fibres contract TOGTHER, or none contracts, if the stimulus is sub-threshold – this is referred to as the “ALL or NONE PRINCIPLE” in regard to a skeletal muscle.Muscle contraction is initiated by a signal sent by the CNS via a motor neuron. Impulse from motor nerve stimulates a muscle fiber at neuro muscular junctions. Neurotransmitter releases here which generates an action potential in sarcolemma. These causes release of Ca++ into sarcoplasm. These Ca++ binds with troponin, thereby remove masking of active site. Myosin head binds to exposed active site on actin to form a cross bridge, utilizing energy from ATP hydrolysis. This pulls the acin filament towards the centre of ‘A’ band. ‘Z’ lines also pulled inward thereby causing a shortening of sarcomere i.e. contraction. ‘I’ band get reduced, whereas the ‘A’ band retain the length. This entire electophysiological process is called as “excitation-contraction” coupling. During relaxation, the cross bridges between the actin and myosin break. Ca++ is pumped back to sarcoplasmic cisternae. Actin filament slide out of ‘A’ band and length of ‘I’ band increases. This returns the muscle to its original resting state.
STRUCTURE OF SKELETAL MUSCLE
Skeletal muscles vary considerably in size, shape, and arrangement of fibers. They range from extremely tiny strands such as the stapedium muscle of the middle ear to large masses such as the muscles of the thigh. Some skeletal muscles are broad in shape and some narrow. In some muscles the fibers are parallel to the long axis of the muscle; in some they converge to a narrow attachment; and in some they are oblique.

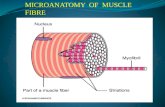
Each skeletal muscle fiber is a single cylindrical muscle cell. An individual skeletal muscle may be made up of hundreds, or even thousands, of muscle fibers bundled together and wrapped in a connective tissue covering. Each muscle is surrounded by a connective tissue sheath called the epimysium. Fascia, connective tissue outside the epimysium, surrounds and separates the muscles. Portions of the epimysium project inward to divide the muscle into compartments. Each compartment contains a bundle of muscle fibers. Each bundle of muscle fiber is called a fasciculus and is surrounded by a layer of connective tissue called the perimysium. Within the fasciculus, each individual muscle cell, called a muscle fiber, is surrounded by connective tissue called the endomysium.
Commonly, the epimysium, perimysium, and endomysium extend beyond the fleshy part of the muscle, the belly or gaster, to form a thick ropelike tendon or a broad, flat sheet-like aponeurosis. The tendon and aponeurosis form indirect

attachments from muscles to the periosteum of bones or to the connective tissue of other muscles. Typically a muscle spans a joint and is attached to bones by tendons at both ends. One of the bones remains relatively fixed or stable while the other end moves as a result of muscle contraction.

Skeletal Muscle
Skeletal muscle, attached to bones, is responsible for skeletal movements. The peripheral portion of the central nervous system (CNS) controls the skeletal muscles. Thus, these muscles are under conscious, or voluntary, control. The basic unit is the muscle fiber with many nuclei. These muscle fibers are striated (having transverse streaks) and each acts independently of neighboring muscle fibers.
Smooth Muscle
Smooth muscle, found in the walls of the hollow internal organs such as blood vessels, the gastrointestinal tract, bladder, and uterus, is under control of the autonomic nervous system. Smooth muscle cannot be controlled consciously and thus acts involuntarily. The non-striated (smooth) muscle cell is spindle-shaped and has one central nucleus. Smooth muscle contracts slowly and rhythmically.
Cardiac Muscle
Cardiac muscle, found in the walls of the heart, is also under control of the autonomic nervous system. The cardiac muscle cell has one central nucleus, like smooth muscle, but it also is striated, like skeletal muscle. The cardiac muscle cell is rectangular in shape. The contraction of cardiac muscle is involuntary, strong, and rhythmical. Intercalated discs are a characteristic of cardiac muscle.

STRUCTURE O0F MYO0FIBRIL & SARCOMERES
Each myofibril consist of alternate dark and light band.Dark band - contain myosim protien and is called A-band or Anisotroic band. It appears dark, as it REFRACTS twice as much light as does the ‘A’ band. Or, it REFLECTS only half as much as light as is done by the ‘I’ band.Light band - Contains actin protein and is called I band or Isotropic band.I band is bisected by a thin ‘Z’ line (Zwischenscheibe, a German word, meaning a CROSS disk). Actin filament (thin filament) are firmly attached to the ‘Z’ lines.Myosin filament (thick filament) are positioned in the ‘A’ band, are also held together spatially in the middle of ‘I’ band by thin fibrous membrane called ‘M’ line. The portion between two successive ‘Z’ lines is considered as the smallest integrated structural & functional unit of muscle contraction - and is called a sarcomere.

Excitation-Contraction coupling (to be discussed personally)
Major Proteins in Skeletal Muscle Sarcomere (molecular architecture)Structural proteins (architectural) > Actin & MyosinRegulatory proteins of muscular contraction > Troponin & Tropomyosin

Mechanism of Muscle contraction : Sliding filament theorySarcomere Lattice Geometry : Spatial cooperation of Actins & Myosins
As shown in the diagram above, actin & myosin molecules within a sarcomere special a characteristic ‘spatial’ relationship. Actin myofilament emerge from the Z discs in a ‘hexagonal configuration, whereas as, in contrast, myosin myofilaments are so arranged in a ‘triangular’ pattern. Both these molecular patterns are so

configured that the centre of each hexagon (of actins) is occupied by an angle of the triangle (myosin).
The contraction of muscle fiber takes place by the sliding of the smaller & thinner actins (thin filament) on thicker & bigger myosins (thick filament). Just before the process of contraction ensues by the sliding of these two respective thin & thick filaments, the CROSS-BRIDGES of the myosin molecule get hooked on to the receptor sites on the actin filaments … a process that keeps repeating in a cyclic, conformational pattern as long as the Ca2+ concentration within the sarcomere stays risen to a level of 10-4 consequent to ‘excitation-contraction’ coupling … facilitated by the transverse-tubular system of a skeletal muscle fibre through a special triple-gated opening called ‘TRIAD’ at the level of Z-lines between two apposing sarcomere on adjacent myofibrils.
All sort of skeletal muscles & their fibres (red, white or intermediate types) require instant energy for contraction, as do smooth & cardiac muscles. The primary energy-rich compound for mammalian muscles is ATP, in combination with CP (Creatine Phosphate) – the energy rich bond of the latter is instantaneously transferred to ADP to regenerate ATP, whenever the muscles are contracting faster than the rate of oxidative rephosporylation of ADP to ATP.
ATP & ADP
ATP is manufactured as a result of several cell processes including fermentation, respiration and photosynthesis. Most commonly the cells use ADP as a precursor molecule and then add a phosphorus to it. In eukaryotes this can occur either in the soluble portion of the cytoplasm (cytosol) or in special energy-producing structures called mitochondria. Charging ADP to form ATP in the mitochondria is called chemiosmotic phosphorylation. This process occurs in specially constructed chambers located in the mitochondrion’s inner membranes. ATP-synthase converts ADP into ATP, a process called charging Adenosine triphosphate (ATP) is an organic molecule which stores energy used to carry out life processes. ATP is made of an adenine nucleoside, ribose sugar, and three phosphate groups. The high energy bonds between phosphate groups are broken when hydrolyzed, thus releasing energy in the system. Either one or two phosphate groups can break off, releasing Gibb's free energy, which can then be used to drive other reactions.

SUBSTRATE-LEVEL & OXIDATIVE PHOSPHORYLATION
Substrate-level phosphorylation is a type of metabolic reaction that results in the formation of adenosine triphosphate (ATP) or guanosine triphosphate (GTP) by the direct transfer and donation of a phosphoryl (PO3) group to adenosine diphosphate (ADP) or guanosine diphosphate (GDP) from a phosphorylated reactive intermediate. Note that the phosphate group does not have to come directly from the substrate. By convention, the phosphoryl group that is transferred is referred to as a phosphate group. An alternative way to create ATP is through oxidative phosphorylation, which takes place during the process of cellular respiration, in addition to the substrate-level phosphorylation that occurs during glycolysis and the Krebs cycle. During oxidative phosphorylation, NADH is oxidized to NAD+, yielding 2.5 ATPs, and FADH2 yields 1.5 ATPs when it is oxidized. Oxidative phosphorylation uses an electrochemical or chemiosmotic gradient of protons (H+) across the inner mitochondrial membrane to generate ATP from ADP, which is a key difference from substrate-level phosphorylation. Unlike oxidative phosphorylation, oxidation and phosphorylation are not coupled in the process of substrate-level phosphorylation, although both types of phosphorylation result in the formation of ATP and reactive intermediates are most often gained in course of oxidation processes in catabolism. However, usually most of the ATP is generated by oxidative phosphorylation in aerobic or anaerobic respiration. Substrate-level phosphorylation serves as fast source of ATP independent of external electron acceptors and respiration. This is the case for example in human erythrocytes, which have no mitochondria, and in the muscle during oxygen depression. CREATINE PHOSPHATE
Creatine phosphate (CP) is stored in muscles as a reservoir of high-potential phosphoryl groups. CP can readily transfer its phosphoryl group to ADP (catalyzed by creatine kinase). Creatine phosphate + ADP + H+<---> ATP + CreatineThe abundance of CP and its high phosphoryl transfer potential relative to that of ATP make it a highly effective ~ P buffer. Depletion of the high energy phosphate bonds is the primary cause of fatigue within the muscles, and also results in excessive lactic acid accumulation consequent to operation of glycolytic (anaerobic) energy-yielding pathway. Creatine is catabolized to creatinine.

The pentose phosphate pathway (also called the phosphogluconate pathway and the hexose monophosphate shunt) is a metabolic pathway parallel to glycolysis. It generates NADPH and pentoses (5-carbon sugars) as well as ribose 5-phosphate, the last one a precursor for the synthesis of nucleotides.
The pentose phosphate pathway (also called the phosphogluconate pathway and the hexose monophosphate shunt) is a metabolic pathway parallel to glycolysis. It generates NADPH and pentoses (5-carbon sugars) as well as ribose 5-phosphate, the last one a precursor for the synthesis of nucleotides. While it does involve oxidation of glucose, its primary role is anabolic rather than catabolic.
RIGOR MORTIS
Rigor Mortis is the stiffening of the body after death following loss of ATP from the body's muscles. ATP is required to make the muscles work, and without ATP muscles tend to become stiff & inflexible. Rigor begins throughout the body at the same time, but the body's smaller muscles - such as those in the face, neck, arms and shoulders - are affected first, and then the subsequent muscles throughout the body … the larger ones are affected later, as ATP depletion in big muscles isn’t instantaneous . Rigor normally appears within the body around two hours after death, with the facial and upper neck & shoulder muscles (upper scapular region)

being the first to be visibly suffer from its effects. Once the state of Rigor is initiated, it normally takes from eight to twelve hours during which period the body becomes completely stiff all over. This ‘fixed’ state lasts for up to another eighteen hours. Contrary to common perception, the process of Rigor Mortis actually does reverse, and the body returns to a flaccid state; the muscles losing their tightness in reverse of how they gained it, i.e.: the larger muscles that hardened the last lose their stiffness first and return to their pre-Rigor condition. Rigor Mortis is a good means of indicating time of death, as it is normally visible within the first thirty-six to forty-eight hours after death.
EFFECT OF EXERCISE ON SKELETAL MUSCLE
The TONE (state of visible contracture) of skeletal muscles is a sign of their health. The better the tone, the better their intrinsic health. Flaccid & loose muscles indicate bad health. Exercise tones up (firms up) skeletal muscles. Further, regular exercise makes them bigger (hypertrophy). However, even in hypertrophy, new muscle cells are never formed, only the pre-existing one gain in size. Lack of exercise or muscle use (as when a limb is in a plaster cast following a fracture) results is decreased muscle tone & size (atrophy). When certain muscle diseases make the muscle become weaker and smaller – it is called as muscle ‘dystrophy’.
PLEASE NOTE THAT AFTER BIRTH NEW(ER) NERVE CELLS & SKELETAL MUSCLE CELLS ARE NEVER FORMED – SUCH A PHENOMENON IS CALLED AS POST-MITOTIC FIXATION. Regaining muscle function following an attack of paralysis (stroke), doesn’t at all imply the formation of any new(er) neurons, but quite simply, the activation of some pre-existing, dormant neurons that were brought out of their latency following stimulation provided by ‘physiotherapy’ exercises. There are millions of such dormant/latent neurons within our CNS at any given point of time.
Muscle Twitch : is an individual contractile response of a skeletal muscle to a single threshold stimulus. The time taken by a ‘muscle twitch’ to complete following the application of a stimulus is referred to as ‘twitch time’. Twitch time comprises (1) Latent Period. Following the application of a stimulus to a nerve supplying a muscle, the muscle doesn’t contract immediately. This time gap, during which a muscle is actually preparing for contraction, is called as latent

period. This is followed first by a ‘contraction period’ and then by a ‘relaxation period’.
SUMMATION OF STIMULI & TETANY in SKELETAL MUSCLES
At times, especially in laboratory experimentation, successive stimuli to a muscle follow the initial stimulus so quickly that after the very first stimulus the muscle didn’t have sufficient time to relax, before the next stimulus made it contract again. The muscle responds by showing into a degree of partial contracture (incopmplete tetany) in response to the experiment (incomplete summation of stimuli). Sometimes, the external stimuli may come in so close together that the muscle has no time to relax at all and it goes into a state of permanent contracture - (copmplete tetany) in response to the experiment (complete summation of stimuli). Such tetanized muscle take an inordinately long time in relaxing fully, and do so only after sufficient ATP has been available to them 0for the purpose, Something very similar happens in the disease Tetanus, where the bodily muscles are almost constantly stimulated by a chemotoxin from the infecting populations of the bacterium Clostridium tetani. In a tetanized muscle, the contracted protein ACTOMYOSIN takes a fairly long time in getting resolved to actin & myosins.
TREPPE (or the STAIRCASE PHENOMENON) … if a muscle is subject to a volley of separate but repeated stimuli quickly together … its contractile response (amplitude of contraction) increases in response to each successive stimulus … yielding an almost staircase sort of pattern – first contraction having the least amplitude, the 2nd slightly higher, the 3rd higher still and so on, till a plateau is reached. Such a phenomenon is called as TREPPE.
MUSCLE SPINDLE (intrafusal & extrafusal) muscle fibres) … certain stretch-sensitive, sensory muscle fibres often lie enclosed in a separate sheathed bag (spindle). All the normal, motor muscle fibres outside this ‘spindle’ are called as ‘extrafusal’ muscle fibres, whereas these specialized, sensory muscle fibres inside the spindle are referred to as ‘intrafusal’ fibres.
Denervation : surgical/experimental loss of the normal nerve supply to a muscle. Reinnervation : surgical/experimental restoration of a muscle’s original nerve. Cross-innervation : surgical/experimental exchange of the nerves of two muscles.