
Self-Organisation in Reaction-Diffusion...
18
Self-Organisation in Reaction-Diffusion Systems Peter Rashkov University of Exeter Biosciences 22.1.2015 “ . . . Later in the book we discuss in considerable detail various possible mechanisms for generating spatial patterns, including reaction-diffusion systems. I firmly believe that the process here is mechanistic and not genetic.” — J. D. Murray, Mathematical Biology
Transcript of Self-Organisation in Reaction-Diffusion...

Self-Organisation in Reaction-Diffusion Systems
Peter Rashkov
University of Exeter Biosciences
22.1.2015
“ . . . Later in the book we discuss in considerable detail variouspossible mechanisms for generating spatial patterns, includingreaction-diffusion systems. I firmly believe that the process here ismechanistic and not genetic.”
— J. D. Murray, Mathematical Biology

Overview
Morphogenesis
Hair follicle spacing in miceAnalytic propertiesLimiting formTuring space
Outlook & Literature

Mathematical models of morphogenesis
I chemical gradients influence fate of surrounding tissue
I cell differentiation
I tissue and organ formation – epidermal appendages (hairs,feathers)
I mathematical model for the chemical basis of morphogenesis– Turing (1952)
I activator-inhibitor model – Gierer and Meinhardt (1972)
I various models for generation of spatial patterns (spatialheterogeneity)
1 / 16

Turing’s model
I reaction kinetic system
(u, v)t = F (u, v), u, v ∈ R, F : R2 → R2
I steady state (u, v) : F (u, v) = 0
I asymptotically stable to spatially homogeneous perturbations
I reaction-diffusion system
(u, v)t = D∇2(u, v) + F (u, v)
I unstable to spatially heterogeneous perturbations
I D 6= I – diffusion rates for u, v must be different
2 / 16

Gierer and Meinhardt’s model
ut − du∆u =up
vq+ σ1(x)− µuu in [0,T )× Ω
vt − dv∆v =ur
v s+ σ2(x)− µvv in [0,T )× Ω
(1)
I Ω is a bounded smooth domain in Rn, and T ∈ (0,∞]
I initial data u(0, ·), v(0, ·) ∈ L∞(Ω)
I homogeneous Neumann boundary conditions
n · ∇u = n · ∇v = 0 in ∂Ω
I p = r = 2, q = 1, s = 0, Turing space (µu, µv )
I vast literature on existence and uniqueness of solutions fordifferent p, q, r , s, σi
3 / 16

Gierer and Meinhardt’s model
(Loading movie...)
4 / 16

Model for hair follicle localisation in mice
wild-type mouse vs. DKK-transgenetic mutant mouse
5 / 16Sick, Reinker, Timmer & Schlake, Science, 314, 2006.

Model for hair follicle spacing in mice
[Sick et al., 2006]
I WNT signalling pathway in hair follicle localisation
I DKK has inhibitory effect on WNT
I GM kinetics with saturation, Turing instability
I identical production term for activator u & inhibitor v
ut − du∇2u =ρu
v + γ· u2
1 + κu2− µuu in [0,T )× Ω
vt − dv∇2v =ρv
v + γ· u2
1 + κu2− µvv in [0,T )× Ω
(2)
6 / 16

Reaction-kinetic system
I GM type without source terms
I normalise ρu = ρv = 1, µu := µu
ρu, µv := µv
ρv
ut = u2
(v+γ)(1+κu2) − µuu
vt = u2
(v+γ)(1+κu2) − µvv
ut = 0
vt = 0
v
uO
µv ≤ µ2u
µv > µ2u
vt = 0
E
E ′
−γ
7 / 16

Analytical properties
I solution (u, v) of (2) has global existence in L∞ on (0,∞]× Ω forstrictly positive initial data in L∞
I (0, 0) is the unique homogeneous steady state for µv < µ2u
(corresponds to values used by Sick et al. (2006))
I it can be shown analytically that u = 0, v = 0 is local attractor forthe model system (2)
I Turing instability is never possible at (0, 0) no matter what du, dvare!
8 / 16

Analytical properties
I solution (u, v) of (2) has global existence in L∞ on (0,∞]× Ω forstrictly positive initial data in L∞
I (0, 0) is the unique homogeneous steady state for µv < µ2u
(corresponds to values used by Sick et al. (2006))
I it can be shown analytically that u = 0, v = 0 is local attractor forthe model system (2)
I Turing instability is never possible at (0, 0) no matter what du, dvare!
9 / 16

Analytical properties
I solution (u, v) of (2) has global existence in L∞ on (0,∞]× Ω forstrictly positive initial data in L∞
I (0, 0) is the unique homogeneous steady state for µv < µ2u
(corresponds to values used by Sick et al. (2006))
I it can be shown analytically that u = 0, v = 0 is local attractor forthe model system (2)
I Turing instability is never possible at (0, 0) no matter what du, dvare!
10 / 16

Analytical properties
I solution (u, v) of (2) has global existence in L∞ on (0,∞]× Ω forstrictly positive initial data in L∞
I (0, 0) is the unique homogeneous steady state for µv < µ2u
(corresponds to values used by Sick et al. (2006))
I it can be shown analytically that u = 0, v = 0 is local attractor forthe model system (2)
I Turing instability is never possible at (0, 0) no matter what du, dvare!
11 / 16

Limiting form of model system
ut − du∇2u =u2
v(1 + κu2)− µuu in [0,T )× Ω
vt − dv∇2v =u2
v(1 + κu2)− µvv in [0,T )× Ω
(3)
I γ = 0, du = 1, dv = d – approximation when γ ≈ 0
I solution (u, v) of (3) has global existence in L∞ on(0,∞]× Ω for strictly positive initial data in L∞
I a-priori bounds on stationary solutions in t
I find Turing space
12 / 16
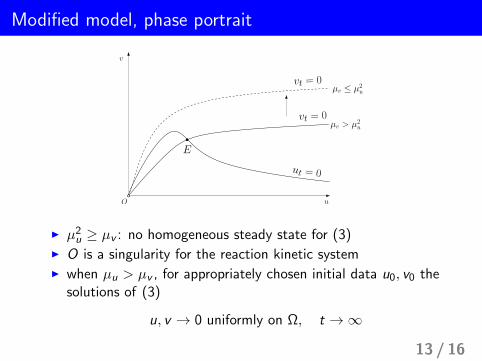
Modified model, phase portrait
ut = 0
vt = 0
v
uO
µv ≤ µ2u
µv > µ2u
vt = 0
E
I µ2u ≥ µv : no homogeneous steady state for (3)I O is a singularity for the reaction kinetic systemI when µu > µv , for appropriately chosen initial data u0, v0 the
solutions of (3)
u, v → 0 uniformly on Ω, t →∞
13 / 16

Turing space for fixed Ω, d , κ
µv
µu0
µ v=µ2 uI
I
V
C
IVh1(µu, µv) = 0
II
IIIh 2(µ
u, µ
v)=0
subregion V – Turing instability, subregion C – collapsing solutionssubregion IV – far-from-equilibrium stationary solutions
14 / 16

Summary
I “modified” GM – uniform boundedness, global existence
I conditions for pattern formation in Sick et al. (2006) not relevant
I patterns observed are not due to Turing bifurcation, butconvergence to a far-from-equilibrium solution
I sensitivity to initial conditions because of collapsing solutions
u
0 20 40 60 80 100
0
20
40
60
80
100
0
0.5
1
1.5
2
2.5
v
0 20 40 60 80 100
0
20
40
60
80
100
0
0.5
1
1.5
2
Figure: Asymmetric pattern converging to some non-trivial solutionbranch stemming from some (unknown) steady state
15 / 16

Literature
I P. Rashkov.Remarks on pattern formation in a model for hair follicle spacing.(under review)
I P. Rashkov.Regular and discontinuous solutions in a reaction-diffusion model forhair follicle spacing.Biomath J. 3(2): 1411111, 2014.
I S. Sick, S. Reinker, J. Timmer, and T. Schlake.WNT and DKK determine hair follicle spacing through areaction-diffusion mechanism.
Science 314(5804): 1447–1450, 2006.
Thanks to : S. Dahlke, B. Schmitt (Marburg), J. Sherratt(Heriott-Watt), A. Marciniak-Czochra (Heidelberg)
16 / 16


















