Seed Dispersal Effectiveness of Rhesus...
18
21 Seed Dispersal Effectiveness of Rhesus Macaques In Chapter 2, I examine the main question of my dissertation – the role of a common species, the rhesus macaque, as seed dispersers in the forests of northern India. Due to their large group sizes, home ranges and daily path lengths, occasional high percentage of fruit in the diet, and ecological resilience, I predicted that rhesus macaques may potentially be effective seed dispersers in disturbed forests. To test my prediction I examined seed dispersal effectiveness in rhesus macaques using the SDE (Seed Disperser Effectiveness, Schupp, 1993; modified to Seed Dispersal Effectiveness, Schupp et al., 2010) framework which comprises qualitative and quantitative aspects. More specifically, I studied (i) the number of fruit species dispersed by rhesus macaques, (ii) macaque seed handling methods, (iii) their rates of seed predation and (iv) germination and establishment rates of dispersed seeds.
Transcript of Seed Dispersal Effectiveness of Rhesus...

21
Seed Dispersal Effectiveness of Rhesus Macaques
In Chapter 2, I examine the main question of my dissertation – the role of a common
species, the rhesus macaque, as seed dispersers in the forests of northern India. Due
to their large group sizes, home ranges and daily path lengths, occasional high
percentage of fruit in the diet, and ecological resilience, I predicted that rhesus
macaques may potentially be effective seed dispersers in disturbed forests. To test my
prediction I examined seed dispersal effectiveness in rhesus macaques using the SDE
(Seed Disperser Effectiveness, Schupp, 1993; modified to Seed Dispersal Effectiveness,
Schupp et al., 2010) framework which comprises qualitative and quantitative aspects.
More specifically, I studied (i) the number of fruit species dispersed by rhesus
macaques, (ii) macaque seed handling methods, (iii) their rates of seed predation and
(iv) germination and establishment rates of dispersed seeds.

22
Chapter 2
Seed Dispersal Effectiveness of Rhesus Macaques*
Abstract
Frugivorous primates are important seed dispersers and their absence from forest
patches is predicted to be detrimental to tropical forest regeneration and recruitment.
With the reduction of primate populations globally, ecologically-resilient common
primate species, characterized by dietary flexibility and the ability to thrive in a
variety of habitats, assume new importance as seed dispersers. The most widely
distributed non-human primate, the rhesus macaque Macaca mulatta, has been
intensively studied but little is known about its role in maintaining ecosystem
structure and functions. Due to their frugivorous diet, large group sizes, large home
ranges and tolerance to disturbance, rhesus macaques may be effective seed
dispersers. To test this hypothesis, I studied seed dispersal by rhesus macaques at the
Buxa Tiger Reserve, India, through a combination of behavioural observations and
germination experiments. Rhesus macaques dispersed 84 % of the 49 species they fed
on either through spitting or defecation. Nearly 96 % of the handled seeds were
undamaged and 61% of the species for which germination tests were performed had
enhanced germination. Almost 50 % of the monitored seeds among those deposited
in situ germinated and 22 % established seedlings, suggesting that rhesus macaques
are important seed dispersers in tropical forests. Due to their widespread distribution
and large populations, rhesus macaques are perceived as common and are categorized
as Least Concern on the IUCN Red List, effectively excluding them from any
conservation plans. Based on the results of my study, I argue that rhesus macaques
fulfill critical ecological functions in their habitat and that this parameter must be
taken into consideration when they are reviewed for conservation priorities.
*A version of this Chapter has been published: Sengupta, A., McConkey, K.R. & Radhakrishna, S. (2014)
Seed dispersal by rhesus macaques Macaca mulatta in northern India. American Journal of Primatology,
76, 1175-1184.

23
2.1 Introduction
As seed dispersers, frugivorous primates are ecologically critical species (Garber &
Lambert, 1998; Chapman & Russo, 2007) and their absence from forest patches is
predicted to lead to lowered rates of tropical forest regeneration and recruitment,
thereby altering plant community structure (Anzures-Dadda et al., 2011). Land use
changes involving forest transformations have led to the loss of large numbers of
primate populations globally (Anzures-Dadda et al., 2011). In such a scenario, the
importance of ecologically-resilient primate species, characterized by dietary
flexibility and ability to thrive in a variety of habitats, increases manifold (Albert et al.,
2014). It has, in fact, been suggested that in many parts of tropical and subtropical
Asia, where forests are fragmented or native mammal fauna has gone extinct,
disturbance-tolerant macaques like the rhesus macaque Macaca mulatta, the bonnet
macaque M. radiata and the long-tailed macaque M. fascicularis may be the only seed
dispersers, especially for the large fruit/seed bearing species (Lucas & Corlett, 1998).
Hence extirpation or even a decline in the number of these macaques would
negatively impact seed dispersal in such areas (Lucas & Corlett, 1998).
Rhesus macaques have the largest geographical distribution among non-
human primates with their range spanning Afghanistan, Bangladesh, Bhutan, India,
Nepal, Pakistan, China, Burma, Laos, Thailand, and Vietnam (Fooden, 2000; Brandon-
Jones et al., 2004;). Highly adaptable, often commensal and classified as ‘Least
Concern’ on the IUCN Red List (IUCN, 2015), M. mulatta has been intensively studied
both in the field as well as laboratory (Richard et al., 1989). Yet little is known about
its role in maintaining ecosystem structure and functions. Although highly flexible in
their feeding ecology, fruits may constitute as much as 70 % (Fooden, 2000) of rhesus
macaque diet. Being semi-terrestrial, they can also include fruits of species at different
strata of the forest (Albert et al., 2013). Their large group sizes (mean: 40, Richter et al.,
2013) mean that a large number of individuals can feed on a patch at the same time
thereby dispersing considerable amounts of seeds at a time (sensu Albert et al., 2013).
Their large home (mean: 593.3 ha, Makwana, 1978) and daily ranges (mean: 1803 m,
Makwana, 1978) imply that individuals can carry seeds to large distances from the
parent trees (Albert et al., 2013). Finally, because of their ability to thrive in a range of
habitats (including fragmented and disturbed habitats) these macaques are
ecologically highly resilient (Albert et al., 2014). These characteristics suggest that
rhesus macaques can be highly effective seed dispersers in disturbed areas. However,
available information on the effectiveness of rhesus macaques as seed dispersal agents

24
is fragmentary (Dudgeon & Corlett, 1994; Lucas & Corlett, 1998) or limited to some
aspects (Tsuji et al., 2013).
Since its conception in 1993, Schupp’s SDE (initially Seed Disperser
Effectiveness; modified to Seed Dispersal Effectiveness - Schupp et al., 2010)
framework for assessing seed dispersal activities has been seminal in this field of
research. Defined as the ‘number of new adults produced by the dispersal activities of
a disperser, SDE considers the quantitative aspect of seed dispersal (measured as the
product of number of visits made to a tree and the number of seeds dispersed per
visit) as well as the qualitative (measured as the product of the probability of seed
survival in a viable condition post-handling and the probability of the survival and
germination of the dispersed seed and the subsequent production of an adult tree)
(Schupp, 1993; Schupp et al., 2010). The most common variables examined with regard
to assessment of SDE are seed handling behaviour, dispersal distance and handling
effects on germination (Schupp et al., 2010). In terms of seed handling mechanisms,
primate species are categorised as “seed swallowers”, “seed spitters” or “seed
destroyers” (Corlett & Lucas, 1990). While spat out seeds are usually deposited singly
on the forest floor, swallowed (and defecated) seeds mostly have a clumped pattern
of deposition (Chapman, 1995). Suitable dispersal distance is a key component for
effective dispersal; not only should seeds be deposited in an area suitable for
germination, an effective disperser should also carry seeds away from the parent tree
to escape mortality due to distance and density dependent predators, pathogens and
herbivores targeting the adult (Janzen, 1970; Connell, 1971; Howe & Smallwood,
1982). Handled seeds should also be in viable condition for a disperser to be called
effective (Schupp et al., 2010). Mechanical and/or chemical scarification of seed coat in
the gut, separation of seeds from pulp and the presence of fecal matter surrounding
the seeds can all have positive implications for the germination of swallowed and
defecated seeds (Traveset & Verdú, 2002).
The aim of this Chapter was to examine the role of rhesus macaques as seed
dispersers taking into account the aforementioned aspects of SDE. More specifically,
I addressed the following questions: (i) How many plant species are dispersed by
rhesus macaques? (ii) How are the various seeds handled? (iii) How far from the
parent tree are seeds deposited? (iv) What is the effect of seed handling on
germination? and (v) What percentage of seeds deposited on the forest floor achieve
establishment?

25
2.2 Methods
2.2.1 Seed handling mechanisms and dispersal distance
I followed the macaques of Troop C from their waking site to their sleeping tree for 10
days each month (12 hours a day; 5 days each in the 2nd and 3rd weeks of each month)
and recorded feeding activity using a 30-minute interval scan sampling method
(Altmann, 1974). In each sample, I scanned the troop for 15 minutes and noted the first
item (plant species and part thereof- fruit, leaf, shoot, flower; insects; fungi) consumed
by each individual. When macaques fed upon fruits, I employed 30 minute-long focal
sampling (Altmann, 1974) on any randomly chosen adult individual to make detailed
observations on the part of the fruit consumed (whole fruit, pulp, seed) and the way
seeds were handled. I checked fruits to determine the state of ripeness (ripe/unripe)
and classified seed handling mechanisms as follows: swallowed (when the entire fruit
was ingested, digested and the seeds egested intact), spat out (when the fruit was taken
into the mouth, mostly stored in cheek pouches, cleaned of the pulp and the seeds
expectorated), dropped (when portions of fruits, e.g. epicarp, were fed upon and then
discarded) and destroyed (when seeds were consistently crunched by macaques or if
the fruits of those species were consumed in an unripe state – Kunz & Linsenmair,
2008; Albert et al., 2013).
I studied remnants of fruits/seeds beneath the feeding tree to confirm the exact
part fed upon and recorded if the macaques spat out any seed during movement
between trees. Often, a number of macaques fed upon fruits of the same tree and spat
out the seeds. As it was difficult to distinguish seeds handled by different individuals
in such conditions, I divided the number of spat out seeds found under a feeding tree
immediately after a feeding event by the total number of macaques on the tree to
obtain data on dispersal through seed-spitting. I opportunistically collected fresh fecal
material from focal individuals and examined it to check the number and status
(intact/crunched) of seeds within. I measured spat out seeds and those found in fecal
samples using calipers and assigned them to the following size categories according
to their length: small (≤ 5 mm in length), medium (> 5 mm but < 10 mm) and large (≥ 10
mm) (Chapman et al., 2010; Albert et al., 2013). Species whose seeds were observed to
be spat out just once or were found only in one fecal sample (N = 2) were not
considered for the analysis.
I recorded the locations of all fecal samples and feeding trees with a hand held
GPS unit. Additionally, every month, for a period of two days, I revisited the feeding
trees and marked conspecifics within 50 m of their canopies along four 10 m wide

26
transects radiating east, west, north and south from the tree bases. Upon identification
of a particular seed species within a fecal sample, I measured straight line distances
between the location of the fecal sample and all marked conspecifics of that tree
species. I considered dispersal distance of fecal seeds as the linear distance between
the defecated seed and the nearest (based on shortest straight line distance) tree of the
same species (McConkey, 2000) and dispersal distances of spat out seeds as the
straight line distance between the actual feeding tree and the seed deposition site
(Albert et al., 2013). Dispersal distances were estimated only for those species which
were found in at least 30 fecal samples or 30 clumps of spat out seeds. For these
analyses, I only used seeds which I had directly observed being spat out or being
egested via fecal matter.
2.2.2. Seed collection and germination experiments
In order to test the effect of rhesus macaque seed handling mechanisms on seed
germination, I collected spat out and defecated seeds from different fecal samples/spat
out clumps and different locations and sowed them along with seeds (cleaned of the
pulp) collected from intact fruits (control seeds) in a shaded nursery under similar
conditions. For Ficus species and Anthocephalus chinensis, groups of 50 seeds were
placed on moist tissue papers on petri dishes (McConkey, 2000). I monitored sown
seeds every day and watered them at an interval of 4 days during the wet months
(April to October) and at an interval of 2 days during the dry months (November to
March). I sowed a total of 1020 treated and 980 control seeds and recorded the percent
of seeds that germinated (emergence of radicle). The germination percent of control
and treated seeds and their latency periods were compared using Chi-square and
Mann-Whitney U test respectively (Zar, 2010).
I conducted in situ germination tests for spat-out seeds of 8 species (Artocarpus
chaplasha, Chisocheton paniculatus, Careya arborea, Elaeocarpus varuna, Populus gamblei,
Premna benghalensis, Cissus elongata, Syzygium formosa) and defecated seeds of 3 species
(Elaeocarpus varuna, Populus gamblei, Premna benghalensis). The choice of these species
was determined by (i) the ease of recognizing their seeds on the forest floor, and (ii)
the timing of the fruiting period that allowed us to monitor them for at least 9 months.
I inserted small pegs with identifying tapes on the ground next to the seeds and
monitored a total of 603 seeds (280 fecal, 323 spat out) to note seed germination and
seedling establishment (defined as the stage when the emerged seedling is no longer
attached to the seed and its root is firmly in the soil, Balcomb & Chapman, 2003).
2.3 Results

27
2.3.1 Seed handling and dispersal
I collected a total of 2865 scans (total observation time = 716.25 h) and 600 focal animal
samples (total observation time = 300 h) on macaque behaviour. Individuals spent 58
% of their active time on feeding. Fruits comprised 79 % of the diet (percent of diet
scans), and the macaques fed on the fruits of 49 species (seed species henceforth)
(Table 2.1). The most common seed handling mechanism was spitting; significantly
more seed species were spat out (53.1 %) than were swallowed (20.4 %), dropped (4.1
%) or destroyed (12.2 %) (χ2 = 75.3, df = 4, p < 0.001) (Fig. 2.1, Table 2.1). Seeds of four
species were both spat out and defecated (Fig. 2.1, Table 2.1), and seeds of five species
were destroyed: Acacia auriculiformis, Acacia pennata, Delonix regia, Michelia champaka,
Quercus lancaefolia and an unidentified liana species (Liana species 1).
Figure 2.1 Seed fate of species fed upon by rhesus macaques (N = 49 species)
Eighty-seven percent of fecal samples (N = 311) contained seeds and comprised
nine small-seeded (71150 seeds) and two medium-sized species (235 seeds). The mean
number of small seeds/fecal sample was 317.63 ± SD 46.95 (N = 224, range: 148-778) and
the mean number of medium seeds/fecal sample was 5.10 ± SD 10 (N = 46, range: 1-
63); the number of fecal samples containing small seeds was significantly more than
those with medium seeds (χ2 = 116.03, df = 1, p < 0.001). Seeds of all sizes were spat
out but the number of observations involving large seeds (n = 678; 20 species, 11450
seeds) was significantly greater than those involving medium (n = 68; 2 species, 2448
seeds) or small seeds (n=135; 4 species, 10403 seeds) (χ2 = 762.13, df = 2, p < 0.001). The
mean number of spat out seeds/ individual was 8.56 ± SD 1.4 for small seeds (range: 6
- 13), 6 ± SD 2.24 for medium seeds (range: 3 - 8), and 3.4 ± SD 1.3 for large seeds (range:
0
10
20
30
40
50
60
Per
ceta
ng
e o
f se
ed
sp
ecie
s
Seed Fate

28
2 - 6). None of the fecal seeds had any visible damage, while 82.5 % of the spat out
seeds were found to be intact.
Table 2.1 Seed fate of species fed upon by rhesus macaques (Troop C)
Species Family Spat out Swallowed Destroyed
Acacia auriculiformis Fabaceae x
Acacia pennata Fabaceae x
Amoora rohituka Meliaceae x
Anogeissus latifolia Combretaceae x
Anthocephalus chinensis Rubiaceae x
Antidesma diandrum Euphorbiaceae x
Artocarpus chaplasha Moraceae x
Baccauria sapida Euphorbiaceae x
Beilschmedia
gammieana
Lauraceae x
Bridelia retusa Euphorbiaceae x
Careya arborea Lecythidaceae x
Casearia spp. Flacourtiaceae x x
Chisocheton paniculatus Meliaceae x
Cissus elongata Vitaceae x x
Crataeva unilocularis Capparaceae x
Delonix regia Fabaceae x
Elaeocarpus aristatus Elaeocarpaceae x
Elaeocarpus floribundus Elaeocarpaceae x
Elaeocarpus varuna Elaeocarpaceae x x
Emblica officinalis Phyllanthaceae x
Eurya acuminata Theaceae x
Ficus benghalensis Moraceae x
Ficus benjamina Moraceae x
Ficus racemosa Moraceae x
Ficus spp. Moraceae x
Gmelina arborea Verbeneceae x
Leea spp. Leeaceae x
Mangifera indica Anacardiaceae x
Mangifera sylvatica Anacardiaceae x
Michelia champaka Magnoliaceae x
Populus gamblei Salicaceae x x

29
Polyalthia simiarum Annonaceae x
Premna benghalensis Verbeneceae x x
Quercus lancaefolia Fagaceae x
Spondias mangifera Anacardiaceae x
Syzygium cumini Myrtaceae x
Syzygium formosa Myrtaceae x
Talauma hodgsonii Magnoliaceae x
Zanthoxylum budrunga Rutaceae x
Ziziphus mauritiana Rhamnaceae x
Liana species 1* x
Liana species 2* x
Liana species 3* x
Liana species 4* x
Liana species 5* x
Liana species 6* x
Liana species 7* x * These species were identified by their vernacular (Bengali/Nepali) names
2.3.2 Dispersal distance
Dispersal distances of fecal seeds (mean = 116.5 ± SD 78.60 m, range: 29.3-774.0 m, n =
370) were significantly greater than those of spat out seeds (mean = 19.2 ± SD 7.85 m,
range: 4.9-69.0 m, n = 174; t = 22, df = 542, p < 0.001; Table 2.2). About 39 % of spat out
seeds were found within the canopy radii of the feeding trees; none of the fecal seeds
were found within the canopy radii.
Table 2.2 Dispersal distances in study area
Treatment Species (N = no. of seeds,
each collected from different
dispersal events )
Mean distance
(m)
Maximum
distance (m)
Fecal Anthocephalus chinensis (34) 64.1 84.4
Eurya acuminata (31) 69.7 90.9
Ficus benghalensis (31) 73.2 144.3
Ficus benjamina (32) 76.4 96.8
Ficus spp.(32) 73.7 103
Liana 7 (37) 91.1 126.3
Leea spp. (32) 98.6 103.6
Premna benghalensis (36) 126.6 158.7
Populus gamblei (34) 132.1 163.2
Cissus elongata (37) 160.6 226.1

30
Elaeocarpus varuna (34) 299 774
Spat Spondias mangifera (38) 15.6 22.6
Polyalthia simiarum (35) 16.6 25.7
Liana 2 (32) 16.8 23.7
Chisocheton paniculatus (35) 18.7 28.7
Syzygium formosa (33) 27.4 43.7
Artocarpus chaplasha (37) 41.9 58.1
Careya arborea (31) 42.3 56.2
Populus gamblei (32) 51.6 79.7
2.3.3. Germination trials
I obtained germination data for 68.2 % of the 2000 seeds sowed; 637 seeds were either
damaged by insects or desiccated. Of the 18 species that germinated, 12 species
showed significant differences either in the germination percent or in the latency
period of the treated seeds (Table 2.3). For Populus gamblei, although none of the
control or the spat-out seeds germinated, all the fecal seeds germinated. Premna
benghalensis and Talauma hodgsonii were two other species where only the treated seeds
germinated. For Anthocephalus chinensis, the germination percent of fecal seeds was
significantly lower than control seeds. However, the mean latency period of fecal
seeds was significantly lower than control seeds.
Table 2.3 Germination percent and latency period of treated and control seeds
Species Treatment Germination percent Mean Latency period
(days)
Control (N) Treated (N) Control Treated
Anthocephalus
chinensis
Fecal 40 (50) 20* (50) 150 50***
Artocarpus chaplasha Spat 83.3 (30) 80 (30) 33.4 34.4
Careya arborea Spat 100 (35) 100 (35) 30 17.75
Chisocheton paniculatus Spat 100 (37) 85.7 (37) 54.9 66.5
Cissus elongata Fecal 69 (34) 61 (34) 65 48
Elaeocarpus varuna Spat 100 (30) 87 (30) 48 40
Fecal 100 (30) 92.5 (30) 34 39
Ficus benghalensis Fecal 90 (50) 95 (50) 40 13***
Ficus benjamina Fecal 90 (50) 83 (50) 43 20.5**
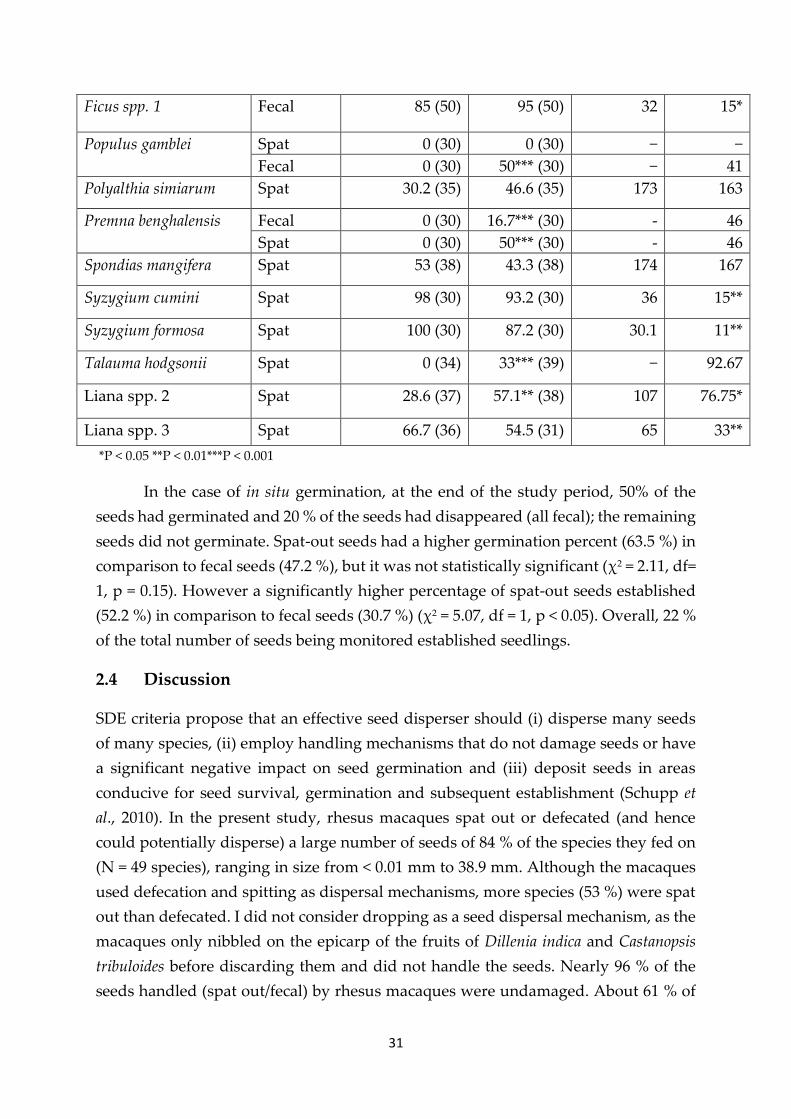
31
Ficus spp. 1 Fecal 85 (50) 95 (50) 32 15*
Populus gamblei Spat 0 (30) 0 (30) − −
Fecal 0 (30) 50*** (30) − 41
Polyalthia simiarum Spat 30.2 (35) 46.6 (35) 173 163
Premna benghalensis Fecal 0 (30) 16.7*** (30) - 46
Spat 0 (30) 50*** (30) - 46
Spondias mangifera Spat 53 (38) 43.3 (38) 174 167
Syzygium cumini Spat 98 (30) 93.2 (30) 36 15**
Syzygium formosa Spat 100 (30) 87.2 (30) 30.1 11**
Talauma hodgsonii Spat 0 (34) 33*** (39) − 92.67
Liana spp. 2 Spat 28.6 (37) 57.1** (38) 107 76.75*
Liana spp. 3 Spat 66.7 (36) 54.5 (31) 65 33**
*P < 0.05 **P < 0.01***P < 0.001
In the case of in situ germination, at the end of the study period, 50% of the
seeds had germinated and 20 % of the seeds had disappeared (all fecal); the remaining
seeds did not germinate. Spat-out seeds had a higher germination percent (63.5 %) in
comparison to fecal seeds (47.2 %), but it was not statistically significant (χ2 = 2.11, df=
1, p = 0.15). However a significantly higher percentage of spat-out seeds established
(52.2 %) in comparison to fecal seeds (30.7 %) (χ2 = 5.07, df = 1, p < 0.05). Overall, 22 %
of the total number of seeds being monitored established seedlings.
2.4 Discussion
SDE criteria propose that an effective seed disperser should (i) disperse many seeds
of many species, (ii) employ handling mechanisms that do not damage seeds or have
a significant negative impact on seed germination and (iii) deposit seeds in areas
conducive for seed survival, germination and subsequent establishment (Schupp et
al., 2010). In the present study, rhesus macaques spat out or defecated (and hence
could potentially disperse) a large number of seeds of 84 % of the species they fed on
(N = 49 species), ranging in size from < 0.01 mm to 38.9 mm. Although the macaques
used defecation and spitting as dispersal mechanisms, more species (53 %) were spat
out than defecated. I did not consider dropping as a seed dispersal mechanism, as the
macaques only nibbled on the epicarp of the fruits of Dillenia indica and Castanopsis
tribuloides before discarding them and did not handle the seeds. Nearly 96 % of the
seeds handled (spat out/fecal) by rhesus macaques were undamaged. About 61 % of

32
dispersed species either had enhanced germination percentages or decreased latency
periods and rhesus seed handling was critical for three species. For the remaining 7
species, the effect of rhesus seed handling on seed germination and latency period was
neutral. Hence the results of my study establish that the rhesus macaque fulfils two of
three criteria to be assessed an effective disperser.
Long-distance dispersal has been documented to be critical to plant dynamics,
being a principal driver of population spread, persistence of subpopulation,
recolonization and gene flow (Schupp et al., 2010). Primate seed dispersal studies
employ various techniques to measure dispersal distance. Stevenson (2000), for
example, considers dispersal distance as the distance between the exact parent tree
and the point where the seed is deposited, on the condition that the focal animal fed
at one tree of a particular species only once during the sampling time and seeds of that
species were defecated/spat out for the first time in that sample 5 or more hours after
the beginning of the observations. Dispersal distances have also been calculated based
on the daily range of animals and their gut passage rates (Chapman & Russo, 2007).
Owing to the large size of our study group, and lack of information regarding rhesus
macaque gut passage rates, I did not use either of these methods in our study. Due to
similar methodological constraints, some studies (eg. Albert et al., 2013) do not report
dispersal distances of fecal seeds at all.
In general, dispersal distance of > 100 m from the parent tree is defined as long-
distance dispersal (Cain et al., 2000). However, shorter dispersal distances may also
permit seeds to escape distance and density dependent effects under parent crowns.
Appropriate dispersal distances are often species and habitat-specific (Lambert &
Chapman, 2005) and have been reported to vary between 2 and < 20 m from the crown
of the parent tree (Webb & Willson, 1985; Schupp, 1988; Ahumada et al., 2004; Hubbell
et al., 2001; Uriarte et al., 2005). In fact, it has been observed that the chance of survival
of seeds dispersed to even 5 m away from the crown is higher than that of those which
are deposited beneath it (Chapman, 1995). In this study, over 90% of all seeds (spat
out and fecal) were deposited beyond the canopy radii of feeding trees and the
maximum distance attained by dispersed seeds was 69 m for spat out seeds and 774
m for fecal seeds. Also, the assumption that the nearest adult is the parent tree
underestimates dispersal distances as the actual parent trees maybe much further
away (Nathan & Muller-Landau, 2000). Hence dispersal distances reported in this
chapter may actually be greater than what my calculations indicate.
Many studies highlight the importance of long dispersal distances away from
parental trees and negative density dependent effects (Wills et al., 1997; Cain et al.,

33
2000; Wright, 2002; Swamy & Terborgh, 2010) and macaques have been dismissed as
effective seed dispersers because they spit out a large number of seeds which are
usually not carried far from the parent tree (Balasubramanian & Bole, 1993). However
long dispersal distances of spat out seeds have been reported for macaques (265 m
by Macaca leonina- Albert et al., 2013; upto 1 km by M. fuscata- Yumoto et al., 1998) and
spitting out might confer certain benefits to seeds. Spat out seeds, for example, are
completely cleaned of their flesh and may therefore be less susceptible to fungal
attack as compared to seeds with pulp attached to them (Lambert, 2001). For the same
reason, spat out seeds would also be less attractive to seed predators, whereas
defecated seeds may attract seed predators due to the presence of dung (Andresen,
2002). Data on Prunus javanica showed that seeds dispersed by spitting by M. leonina
were deposited closer to the parent tree than seeds dispersed by hornbills and
gibbons, but accounted for 67 % of the established seedlings at the end of the year
(McConkey & Brockelman, 2011). All the monitored seeds which disappeared in the
present study were fecal seeds. Of course, these may have been removed by
secondary dispersers (Andresen, 2002) and the post-deposition seed fate of rhesus
macaque-dispersed seeds clearly requires further study.
In this study, nearly 50 % of the monitored seeds of those deposited in situ
germinated and 22 % established seedlings, thereby implying that rhesus macaques
may potentially be responsible for the emergence of a considerable number of
seedlings in tropical forests. SDE though is measured as the number of adult trees
produced by the activities of a disperser (Schupp et al., 2010) and post-deposition
seed fate estimation should essentially take into account seed survival, germination,
survival and growth of seedling to adult stage (Schupp & Fuentes, 1995). Thus, it is
imperative to carry out long-term studies to monitor establishment of adult trees
from seeds dispersed by rhesus macaques before a valid conclusion can be drawn
regarding the effectiveness of rhesus macaques as seed dispersers.
An effective frugivore should disperse the seeds of many plant species
(Andresen, 2002) but what number of species would qualify as ‘many’ would depend
on the plant community composition and fruit availability. The composition of seeds
dispersed by macaques may not be rigid but rather a function of vegetation
associated with different habitats (Tsuji et al., 2013), and further research is required
to investigate how dietary diversity and hence seed dispersal activities of rhesus
macaques may vary with resource availability (or phenological diversity) across
seasons and in different habitats. Also, mutualistic interactions like seed dispersal
involve plant species interacting with multiple frugivores, thereby forming networks

34
(Donatti et al., 2011) and certain plant-frugivore interactions maybe more specialized
and stronger than others within an assemblage (Schleuning et al., 2011). Therefore, a
complete understanding of the importance of the rhesus macaque as a seed disperser
would entail investigating the various dispersal interactions that it is a part of and
also if it links many modules within such networks. Such studies would also help
determine how vulnerable different plant species are to changes in rhesus macaque
abundance (sensu Schupp et al., 2010).
Ecologically-resilient and disturbance-tolerant primates are typically perceived
to be commonly present and therefore tend to categorised as Least Concern on the
IUCN RedList, which effectively excludes them from any conservation plans.
However, although such primate species are highly adaptable, they are also
negatively impacted by habitat degradation, poaching and indiscriminate use in
biomedical research and commercial trade (Malik, 1992; Molur et al., 2003; Eudey,
2008; Radhakrishna & Sinha, 2011; Albert et al., 2014). At least two Least Concern
species, the bonnet macaque Macaca radiata and the long-tailed macaque M.
fascicularis, have suffered major declines in population size over the past couple of
decades (Singh & Rao, 2004; Eudey, 2008; Kumara et al., 2010; Kumar et al., 2011; Singh
et al., 2011; Sengupta & Radhakrishna, 2013). A widespread geographic distribution,
and perceptions regarding its large population numbers, makes the rhesus macaque
particularly vulnerable to commodification and even a brief history of rhesus
macaque-human interactions underline an unregulated use of the species by humans
(Malik, 1992; Ahuja, 2013). More information about the ecological role of the species,
leading to a greater appreciation of its functional significance, may improve the
conservation value of the species. The results of this Chapter imply that rhesus
macaques potentially fulfil crucial ecological roles as seed dispersers, particularly in
disturbed forest areas, and I strongly urge that this parameter be considered to re-
evaluate their conservation importance in the ecosystem.
2.5 References
Ahuja, N. (2013) Macaques and Biomedicine: Notes on Decolonization, Polio, and
Changing Representations of Indian Rhesus in the United States, 1930-1960. The
Macaque Connection: Cooperation and conflict between humans and macaques (ed. by
S. Radhakrishna, M.A. Huffman & A. Sinha), pp 71-92, Springer, New York.

35
Ahumada, J.A., Hubbell, S.P., Condit, R. & Foster, R.B. (2004) Long term tree survival
in a Neotropical forest: the influence of local biotic neighbourhood. Forest
diversity and dynamism: findings from a network of large‐scale tropical forest
plots (ed. by E. Losos, E.G. Leigh Jr. & R. Condit), pp. 408-432, University of
Chicago Press, Chicago, IL.
Albert, A., Hambuckers, A., Culot, L., Savini, T. & Huynen, M-C. (2013) Frugivory and
seed dispersal by northern pigtaled macaques (Macaca leonina) in Thailand.
International Journal of Primatology, 34, 170-193.
Albert, A., McConkey, K., Savini, T. & Huynen, M‐C. (2014) The value of disturbance‐
tolerant cercopithecine monkeys as seed dispersers in degraded habitats.
Biological Conservation, 170, 300–310.
Altmann, J. (1974) Observational study of behavior: sampling methods. Behaviour, 49,
227-267.
Andresen, E. (2002). Primary seed dispersal by red howler monkeys and the effect of
defecation patterns on the fate of dispersed seeds. Biotropica, 34, 261–272.
Anzures‐Dadda, A., Andresen, E., Martinez, M. & Manson, R.H. (2011) Absence of
howlers (Alouatta palliata) influences tree seedling densities in tropical rain forest
fragments in southern Mexico. International Journal of Primatology, 32, 634–651.
Balasubramanian, P. & Bole, P.V. (1993) Seed dispersal by mammals at Point Calimere
Wildlife Sanctuary, Tamil Nadu. Journal of Bombay Natural History Society, 90, 33–
44.
Balcomb, S.R. & Chapman, C.A. (2003) Bridging the seed dispersal gap: consequences
of seed deposition for seedling recruitment in primate-tree interactions. Ecological
Monographs, 73, 625–642.
Brandon-Jones, D., Eudey, A.A., Geissmann, T., Groves, C.P., Melnick, D.J., Morales,
J.C., Shekelle, M. & Stewart, C.B. (2004) Asian primate classification. International
Journal of Primatology, 25, 97–164.
Cain, M.L., Milligan, B.G. & Strand, A.E. (2000) Long‐distance seed dispersal in plant
populations. American Journal of Botany, 87, 1217–1227.
Chapman, C.A. (1995) Primate seed dispersal: coevolution and conservation
implications. Evolutionary Anthropology, 4, 74–82.
Chapman, C.A. & Russo, S.E. (2007) Primate seed dispersal: linking behavioral
ecology with forest community structure. Primates in perspective (ed. by C.J.
Campbell, A.F. Fuentes, K.C. MacKinnon, M. Panger & S. Bearder), pp. 510–525.
Oxford University Press, Oxford, UK.
Chapman, H.M., Goldson, S.L. & Beck, J. (2010) Post-dispersal removal and
germination of seed dispersed by Cercopithecus nictitans in a West African
montane forest. Folia Primatologia, 81, 41–50.
Connell, J.H. (1971) On the role of natural enemies in preventing competitive exclusion
in some marine animals and rain forest trees. Dynamics of Populations (ed. By P.J.
Dem Boer & G.R. Gradwell), Centre for Agricultural Publishing and
Documentation, Wageningen.

36
Corlett, R.T. & Lucas, P.W. (1990) Alternative seed-handling strategies in primates:
Seed-spitting by long-tailed macaques (Macaca fascicularis). Oecologica, 82, 166–
171.
Donatti, C.I., Guimaraes, P.R., Galetti, M., Pizo, M.A., Marquitti, F.M.D. & Dirzo, R.
(2011) Analysis of a hyper-diverse seed dispersal network: modularity and
underlying mechanisms. Ecology Letters, 14, 773–78.
Dudgeon, D. & Corlett, R.T. (1994) Hills and streams: an ecology of Hong Kong. Hong
Kong University Press, Hong Kong.
Eudey, A.A. (2008) The crab-eating macaque (Macaca fascicularis): Widespread and
rapidly declining. Primate Conservation, 23, 129–132.
Fooden, J. (2000) Systematic review of the rhesus macaque, Macaca mulatta
(Zimmermann, 1780). Fieldiana Zoology New Series, 96, 1–180.
Garber, P.A. & Lambert, J.E. (1998) Introduction to primate seed dispersal. Primates
as seed dispersers: ecological processes and directions for future research.
American Journal of Primatology, 45, 3–8.
Howe, H.F. & Smallwood, J. (1982) Ecology of seed dispersal. Annual Review of Ecology
and Systematics, 13, 201-228.
Hubbell, S.P., Ahumuda, J.A., Condit, R. & Foster, R.B. (2001) Local neighbourhood
effects on long‐term survival of individual trees in a Neotropical forest. Ecological
Research, 16, 859–875.
IUCN (2015) IUCN red list of threatened species. Version. 2015.2. Available online at:
www.iucnredlist.org. Downloaded on 22 March 2015.
Janzen, D.H. (1970) Herbivores and the number of trees species in tropical forests.
American Naturalist, 104, 501-527.
Kumar, R., Radhakrishna, S. & Sinha, A. (2011) Of Least Concern? Range extension by
rhesus macaques (Macaca mulatta) threatens long-term survival of bonnet
macaques (M. radiata) in Peninsular India. International Journal of Primatology, 32,
945–959.
Kumara, H.N., Kumar, S. & Singh, M. (2010) Of how much concern are the ‘Least
Concern’ species? Distribution and conservation status of bonnet macaques,
rhesus macaques and Hanuman langurs in Karnataka, India. Primates, 51, 37–42.
Kunz, B.A. & Linsenmair, K.E. (2008) The role of olive baboons as seed dispersers in
the savannah‐forest mosaic of West Africa. Journal of Tropical Ecology, 24, 235–
246.
Lambert, J.E. (2001) Red-tailed guenons (Cercopithecus ascanius) and Strychnos mitis:
evidence for plant benefits beyond seed dispersal. International Journal of
Primatology, 22, 189–201.
Lambert, J.E. & Chapman, C.A. (2005). The fate of primate‐dispersed seeds: deposition
pattern, dispersal distance and implications for conservation. Seed fate—
predation, dispersal and seedling establishment (ed. by P.M. Forget, J.E. Lambert, P.E.
Hulme & S.B. Vander Wall), pp. 137-150, CAB International, Cambridge, UK.
Lucas, P.W. & Corlett, R.T. (1998) Seed dispersal by long-tailed macaques. American
Journal of Primatology, 45, 29–44.

37
Makwana, S.C. (1978) Field ecology and behaviour of the rhesus macaque (Macaca
mulatta): I. Group composition, home range, roosting sites, and foraging routes
in the Asarori Forest. Primates, 19, 483-492.
Malik, I. (1992). Consequences of export and trapping of monkeys. Primate Report, 34,
5-11.
McConkey, K.R. (2000) Primary seed shadow generated by gibbons in the rain forest
of Barito Ulu, Central Borneo. American Journal of Primatology, 52, 13–29.
McConkey, K.R. & Brockelman, W.Y. (2011) Non-redundancy in the dispersal
network of a generalist tropical forest tree. Ecology, 92, 1492–1502.
Molur, S., Brandon-Jones, D., Dittus, W., Eudey, A., Kumar, A., Singh, M., Feeroz,
M.M., Chalise, M., Priya, P. & Walker, S. (2003) Status of South Asian Primates:
Conservation Assessment and Management Plan (C.A.M.P.) Workshop Report.
Zoo Outreach Organisation/ CBSG-South Asia, Coimbatore, India.
Nathan, R. & Muller‐Landau, H.C. (2000) Spatial patterns of seed dispersal, their
determinants and consequences for recruitment. Trends in Ecology and Evolution,
15, 278–285.
Radhakrishna, S. & Sinha, A. (2011). Less than wild? Commensal primates and
wildlife conservation. Journal of Biosciences, 36, 1–5.
Richard, A., Goldstein, S. & Dewar, R. (1989) Weed macaques: the evolutionary
implications of macaque feeding ecology. International Journal of Primatology, 10,
569-594.
Richter, C., Taufiq, A., Hodges, K., Ostner, J. & Schülke, O. (2013) Ecology of an
endemic primate species (Macaca siberu) on Siberut Island, Indonesia.
SpringerPlus 2, 137, http://www.springerplus.com/content/2/1/137.
Schleuning, M., Blüthgen, N., Flörchinger, M., Braun, J., Schaefer, H.M. & Böhning-
Gaese, K. (2011) Specialization and interaction strength in a tropical plant–
frugivore network differ among forest strata. Ecology, 92, 26–36.
Schupp, E.W. (1988) Factors affecting post-dispersal seed survival in a tropical forest.
Oecologia, 76, 525–530.
Schupp, E.W. (1993) Quantity, quality and the effectiveness of seed dispersal.
Vegetatio, 107/108, 15-29.
Schupp, E.W. & Fuentes, M. (1995) Spatial patterns of seed dispersal and the
unification of plant population ecology. Ecoscience, 2, 267–275.
Schupp, E.W., Jordano, P. & Gômez, J.M. (2010) Seed dispersal effectiveness revisited:
a conceptual review. New Phytologist, 188, 333–353.
Sengupta, A. & Radhakrishna, S. (2013) Of concern yet? Distribution and conservation
status of the bonnet macaque (Macaca radiata) in Goa, India. Primate Conservation,
27, 109–114.
Singh, M. & Rao, N.R. (2004). Population dynamics and conservation of commensal
bonnet macaques. International Journal of Primatology, 25, 847–859.
Singh, M., Erinjery, J.J., Kavana, T.S., Roy, K. & Singh, M. (2011) Drastic population
decline and conservation prospects of roadside dark-bellied bonnet macaques
(Macaca radiata radiata) of southern India. Primates, 52, 149–154.

38
Stevenson, P.R. (2000) Seed dispersal by woolly monkeys (Lagothrix lagothricha) at
Tinigua National Park, Colombia: dispersal distance, germination rates, and
dispersal quantity. American Journal of Primatology, 50, 275–289.
Swamy, V. & Terborgh, J.W. (2010) Distance‐responsive natural enemies strongly
influence seedling establishment patterns of multiple species in an Amazonian
rain forest. Journal of Ecology, 98, 1096–1107.
Traveset, A. & Verdú, M. (2002) A meta‐analysis of the effect of gut treatment on seed
germination. Seed dispersal and frugivory: ecology, evolution and conservation (ed. by
D.J. Levey, W.R. Silva & M. Galetti), pp. 339-350, CAB International, New York,
NY.
Tsuji, Y., Minh, N.V., Kitamura, S., Van, N.H. & Hamada, Y. (2013) Seed dispersal by
rhesus macaques (Macaca mulatta) in Son Tra Nature Reserve, central Vietnam:
A preliminary report. Vietnamese Journal of Primatology, 2, 65-73.
Uriarte, M., Hubbell, S.P., Condit, R. & Canham, C.D. (2005) Neighbourhood effects
on sapling growth and survival in a neotropical forest and the ecological
equivalence hypothesis. Biological interactions in the tropics: their role in the
maintenance of species diversity (ed. by D. Burslem, M. Pinard & S. Hartley), pp.
89-106, Cambridge University Press, Cambridge, UK.
Webb, S.L. & Willson, M.F. (1985) Spatial heterogeneity in post-dispersal predation on
Prunus and Uvularia seeds. Oecologia, 67, 150–153.
Wills, C., Condit, R., Foster, R.B. & Hubbell, S.P. (1997). Strong density and diversity‐
related effects help to maintain tree species diversity in a Neotropical forest.
Proceedings of the National Academy of Sciences USA, 94, 1252–1257.
Wright, S.J. (2002) Plant diversity in tropical forests: a review of mechanisms of
species coexistence. Oecologia, 130, 1–14.
Yumoto, T., Noma, N. & Maruhashi, T. (1998) Cheek-pouch Dispersal of Seeds by
Japanese Monkeys (Macaca fuscata yakui) on Yakushima Island, Japan. Primates,
39, 325-338.
Zar, J.H. (2010) Biostatistical Analysis, 4th Edition, Pearson Prentice-Hall, Upper Saddle
River, NJ.