
Secondary production and consumer energetics The consumer energy budget Determinants of energy flow...
45
Secondary production and consumer energetics • The consumer energy budget • Determinants of energy flow • Ecological efficiencies • Definition of secondary production • Measurement of secondary production • Predicting secondary production – For individual populations – For guilds of consumers – For the entire community of consumers
-
Upload
georgiana-dalton -
Category
Documents
-
view
237 -
download
0
Transcript of Secondary production and consumer energetics The consumer energy budget Determinants of energy flow...

Secondary production and consumer energetics
• The consumer energy budget• Determinants of energy flow• Ecological efficiencies• Definition of secondary production• Measurement of secondary production• Predicting secondary production
– For individual populations– For guilds of consumers– For the entire community of consumers


Ingestion (I)
→

Ingestion (I)
→Assimilation (A)
I = A + E
→Egestion (E)

Ingestion (I)
→ Assimilation (A)
I = A + EA = R + P (+ U)
→Egestion (E)
Respiration (R)
→
Growth (G), or Production (P)
(Excretion (U))

Ingestion (I)=loss to preypopulation→ Assimilation (A)
=energy available to consumer
I = A + EA = R + P (+U)
→Egestion (E)=input to detritus
Respiration (R)=loss of useful energy
→
Growth (G), or Production (P)=energy available to predators
(Excretion (U))

What affects rates of energy flow?

Temperature affects energetic rates(Q10 ~2)
Peters 1983

Body size affects energetic rates(~M-0.25)
Peters 1983
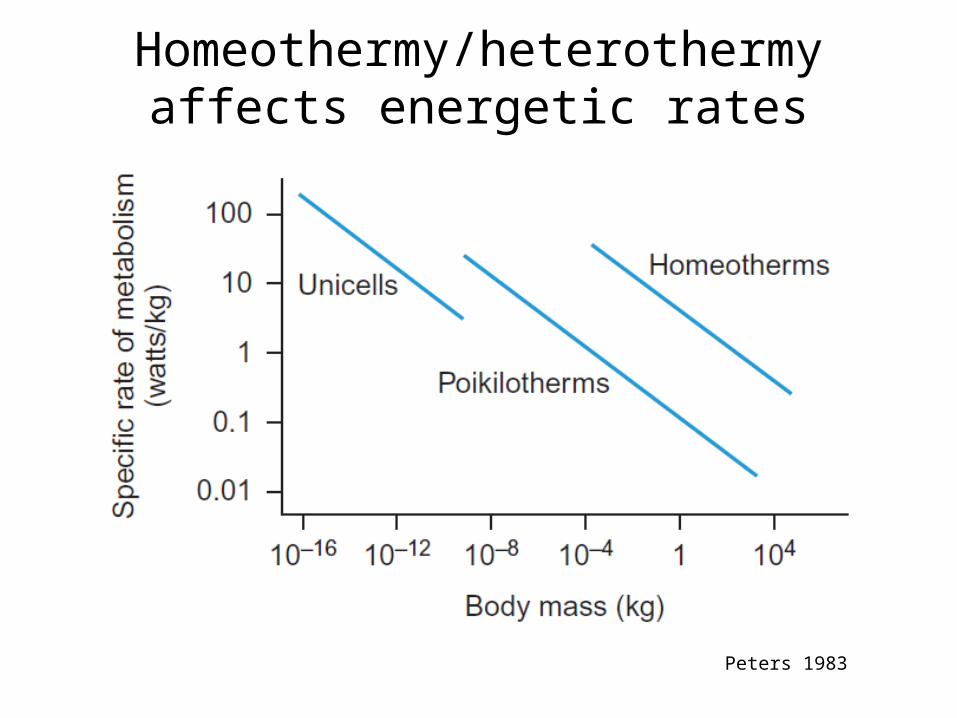
Homeothermy/heterothermy affects energetic rates
Peters 1983

Metabolic rates are evolutionarily flexible
Data on flatworms from Gourbault 1972

Ecological efficiencies
A/I = assimilation efficiency
P/A = net growth efficiency
P/I = gross growth efficiency

Typical values of ecological efficiencies
Assimilation efficiency (%)
Net growth efficiency (%)
Gross growth efficiency (%)
Plants 1 – 2 30 – 75 0.5 – 1
Bacteria - 5 – 60 -
Heterotherms 10 – 90 10 – 60 5 – 30
Homeotherms 40 – 90 1 – 5 1 - 4

What affects ecological efficiencies (partitioning of energy)?

Assimilation efficiency
depends on food quality
Valiela 1984

Bacterial growth efficiency depends on food quality
Del Giorgio andCole 1998

Bacterial growth efficiency depends on temperature
Rivkin and Legendre 2001

Introduction to secondary production
• “All non-photosynthetic production (growth), regardless of its fate”
• NOT the same as biomass accumulation
• NOT just the production of herbivores
• Much better studied than other parts of the consumer energy budget– Easier to measure– Historically considered more important

Secondary production is aquatic and empirical
• 167 papers published on subject in 2005• 52% marine or estuarine, 35% freshwater, 3% terrestrial• 55% microbial, 39% invertebrate, 7% vertebrate• Very little theoretical work• Are generalizations about secondary production
really generalizations about aquatic ecosystems?

How do we estimate secondary production?
• Tracer methods
• Demographic methods
• Turnover methods
• Empirical methods

How do we estimate secondary production?
Organism Method Data requirements Limitations
Bacteria tracers (radioactive nucleotides or amino acids)
uptake of label subject to large errors because of (i) critical assumptions about fate and use of label and non-radioactive analogues, which may be hard to test; (ii) uncertain conversion factors to get from uptake of label to carbon production
Fungi ergosterol synthesis (from radioactive acetate)
uptake of label into ergosterol method still under development; potential problems similar to those for bacterial production
animals with recognizable cohorts
increment summation, mortality summation, Allen curve
density and body size of animals at frequent intervals over the life of the cohort
data intensive
animals without recognizable cohorts
growth increment summation, instantaneous growth
density, body size, and growth rates of animals in various size classes throughout the year
data intensive; growth rates often measured in the lab and extrapolated to the field
egg ratio density and development time of eggs, body mass of animals at death
suitable only in the special case in which the body mass at death is known
size-frequency (“Hynes method”)
density and body size of animals in various size classes throughout the year
data intensive
any organism empirical models population biomass; perhaps body size, temperature, habitat type
subject to large error; may be data intensive

Controls on/prediction of secondary production
• Individual populations
• Guilds of consumers
• Entire communities

Predicting secondary production:(1) individual populations
Marine benthic invertebrates
Log10(P) = 0.18 + 0.97 log10(B) - 0.22 log10(W) + 0.04 (T) – 0.014 (T*log10depth)
R2 = 0.86, N = 125
Tumbiolo and Downing 1994

Predicting secondary production:(1) individual populations
Marine benthic invertebrates
Log10(P) = 0.18 + 0.97 log10(B) - 0.22 log10(W) + 0.04 (T) – 0.014 (T*log10depth)
R2 = 0.86, N = 125
Tumbiolo and Downing 1994
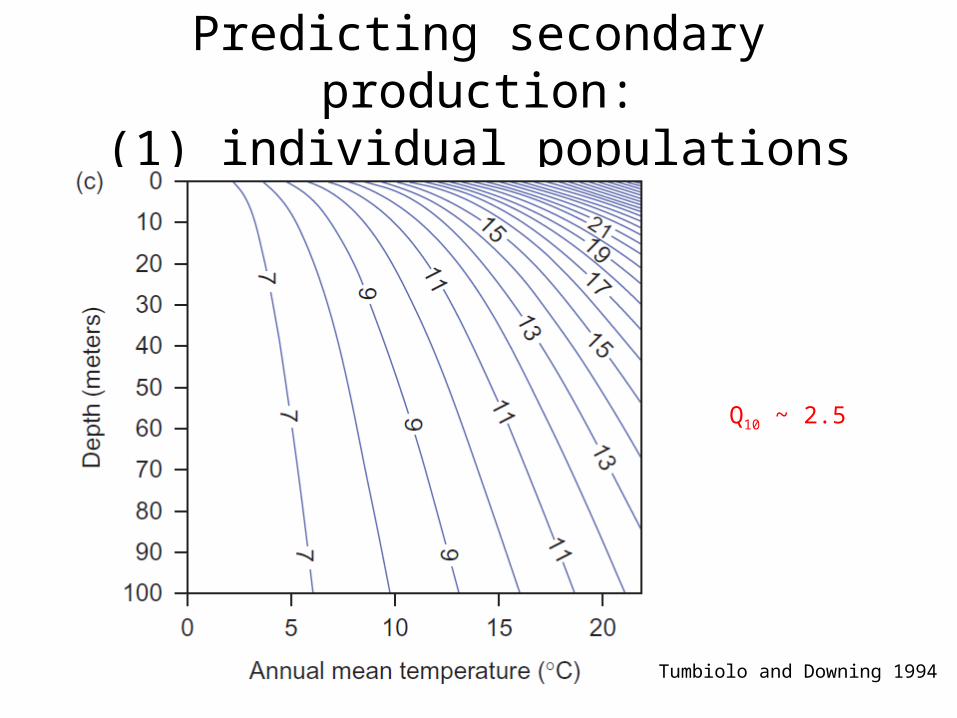
Predicting secondary production:(1) individual populations
Tumbiolo and Downing 1994
Q10 ~ 2.5

Predicting secondary production:(1) individual populations
Tumbiolo and Downing 1994

Predicting secondary production of individual populations
• Feasible if you know mean annual biomass, body size, and temperature
• Very imprecise
• If you’re going to measure mean annual biomass, why not just estimate production directly?

Predicting secondary production: (2) guilds
(aquatic bacterial production as a function of phytoplankton production – Cole et al. 1988)

Predicting secondary production: (2) guilds
(aquatic invertebrate production in experimentally manipulated streams (Wallace et al. 1999)
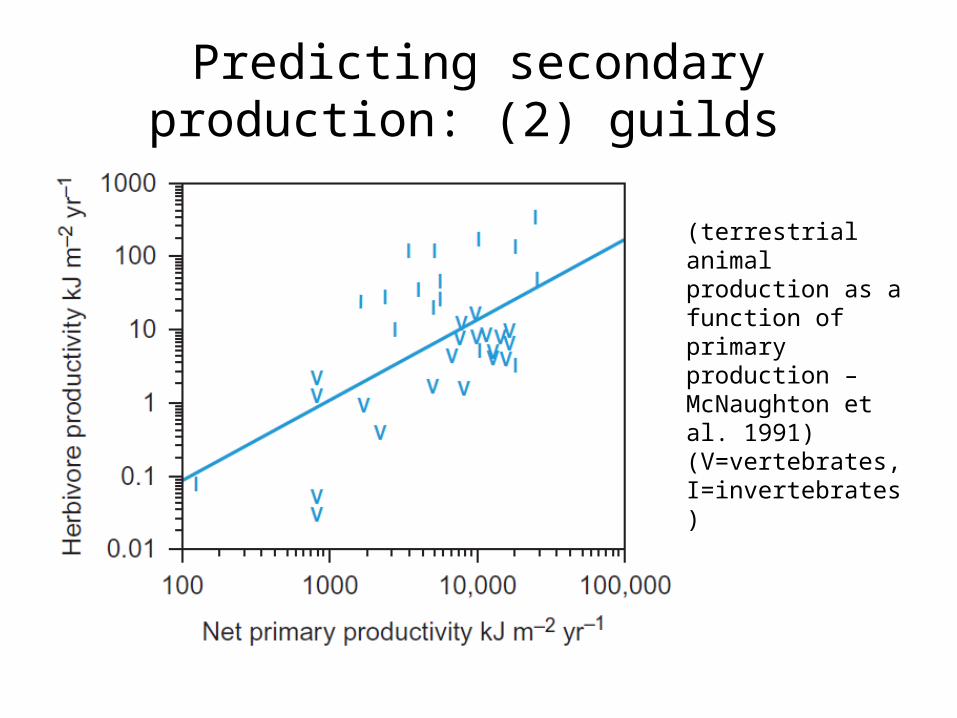
Predicting secondary production: (2) guilds
(terrestrial animal production as a function of primary production – McNaughton et al. 1991) (V=vertebrates, I=invertebrates)

Activity of consumer guilds rises roughly linearly with food supply
Ecosystem type Consumer activity RMA slope Source
Lakes Zoobenthos production
0.8 Kajak et al. 1980
Aquatic ecosystems Bacterial production 1.1 Cole et al. 1988
Terrestrial ecosystems
Aboveground production
1.8 McNaughton et al. 1991
Aquatic ecosystems Herbivore ingestion 1.05 Cebrian and Lartigue 2004
All ecosystems Herbivore ingestion 1.1 Cebrian 1999
Marine ecosystems Herbivore ingestion 1.0 Cebrian 2002

Nutrients affect production of guilds
Cross et al. 2006

Predicting secondary production (or ingestion): (2) guilds
(Cebrian and Lartigue 2004)
Aquatic is white (left) or blue (center and right); terrestrial is black (left) or green (center and right)

Terrestrial/aquatic differences
• Herbivores ingest a higher proportion of NPP in aquatic systems (higher nutrient content of NPP)
• Herbivore production possibly much higher in aquatic systems (higher ingestion, higher assimilation efficiency?, less homeothermy so higher net growth efficiency)

Predicting secondary production of guilds
• Predictable (and linear?) from resource supply• Too imprecise to be very useful as a predictor• Maybe strong terrestrial/aquatic differences
arising from nutrient content of primary producers
• Nutrients as well as energy affect guild production

Predicting secondary production: (3) entire communities

Predicting secondary production: (3) entire communities
S = R + L, so R = S – L
(S = net supply of organic matter, L = non-respiratory losses)

Predicting secondary production: (3) entire communities
S = R + L, so R = S – L
εng = P/(P + R), so P = εng(P + R)
(εng = net growth efficiency,S = net supply of organic matter, L = non-respiratory losses)

Predicting secondary production: (3) entire communities
S = R + L, so R = S – L
εng = P/(P + R), so P = εng(P + R)
Therefore, P = εng(P + S – L)

Predicting secondary production: (3) entire communities
S = R + L, so R = S – L
εng = P/(P + R), so P = εng(P + R)
Therefore, P = εng(P + S – L);
Rearranging, P(1- εng) = εng(S – L)

Predicting secondary production: (3) entire communities
S = R + L, so R = S – L
εng = P/(P + R), so P = εng(P + R)
Therefore, P = εng(P + S – L);
Rearranging, P(1- εng) = εng(S – L)
And P = (S – L)εng/(1 – εng)

Predicting secondary production: (3) entire communities
P = (S – L) εng/(1 – εng)
A = (S – L)/(1 – εng)
I = (S – L)/(εa(1 - εng))
εa = assimilation efficiency, εng = net growth efficiency,S = net supply of organic matter, L = non-respiratory losses

Predicting secondary
production: (3) entire communities

Predicting secondary production of entire communities
• Secondary production is large compared to primary production (if NGE=30%, secondary production = 43% of NPP)
• Decomposers see a lot of consumer tissue (not just plant tissue)
• Secondary production is larger in systems dominated by heterotherms than in systems dominated by homeotherms
• Energy available for ingestion and assimilation by consumers is greater than primary production (if NGE=30% and AE = 20%, A=143% of NPP, I = 714% of NPP)

Conclusions
• It’s easier to predict the secondary production of an entire community than a single population
• Consumer activity is tightly linked with other processes that control the movement and fate of organic matter
• When considered at the community level, secondary production (maybe) is controlled by the same factors that control primary production: supply of energy and nutrients, and their retention


















