RWU-4152 Staff': VMPR Problem: 02 Code: 2.42 … · PX. Khosla.Palampur ... (Canham et o2.,1999)....
53
RWU-4152 Staff': VMPR Problem: 02 Code: 2.42 Reprints: N e. _ t- o__ . _= O >7. t::: , , _ _5. _= M m N, ..=_ 0 _ - _,_ .._- ,._,_ _ .-- "_ e- _ _ .. .- k3 -;_ _.o ==- _< _.__
Transcript of RWU-4152 Staff': VMPR Problem: 02 Code: 2.42 … · PX. Khosla.Palampur ... (Canham et o2.,1999)....

RWU-4152
Staff': VMPR Problem: 02
Code: 2.42 Reprints: N
e. _ t-
o__ . _=
O >7. t::: , ,
_ _5. _=M m N, ..=_
0 _ - _,_ .._-,._,_ _ .--"_ e- _ _ ...- k3
-;_ _.o
==- _<_.__

ii IUFRO Research Series
publishing material in this series are available from the publisher: Books andReference Works, CABI Publishing, CAB International, Wallingford, Oxon
OXIO 8DE, UK; and the IUFRO Secretariat, c/o Federal Forest Research Centre,
-:.:-_ Seckendorff-Gudent-Weg 8, A-1131, Vienna, Austria.
IUFRO Executive Board:
]. Budey, Oxford, UK M.N.Salleh,Kuala Lumpur, MalaysiaD.EDykstra,Portland, Oregon,USA K. Sassa, U}!Kyoto,JapanR. glenaRosello, Madrid,Spain R. Schlaepfer,Lausanne, SwitzerlandD.W.Flinn, Heidelberg.Victoria,Austra_a H. Schmutzenhofex,Vienna, AustriaR. GuevaraMoncada,Turrialba,CostaRlca L.Sennerby-Forsse,Uppsala,SwedenJ.Hong, Beijing, China R. Seppaelae.Helsinki.FinlandD.EKarnosky,Houghton, Michigan,USA B. Solber_ As, NorwayPX. Khosla.Palampur,Hlmachal Pradesl_ V.V.Strakhov,Moscow.RussianFederation
India E.Teissierdu Cros,Avignon. FranceN.g. Koch,Hoexsholm, Denmark ICVancura,Praha, Czech RepublicEJ.Krugex.Sunnyslde,South Africa K.yon Gadow,Goetttngen,GermanyD.ICLee,Seoul, South Korea . J.L.Whltmore, Washington, DC,USAJ.A. PradoDonoso, Santiago, Chile C. W'mget,Ottawa,Ontario,CanadaEN. Sail.Dakar,Senegal J.A. Yotmgquist,Madison, W'k_consimUSA
IUFRO Research Series
Titles available:
1. PorestDynamics in Heavily PollutedRegionsEditedbyJ.L.Innes andJ.Oleksyn
2. ForestHistory: Intonational StudiesonSodoeconomlcandForestEcosystem ChangeEditedbyM. AgnoletUand S.Anderson
3. MethodsandApproachesin ForestHistoryEditedbyM. Agnoletti and S.Anderson
4. Air Pollutionand the ForestsofDeveloplR@andRapidlyIndustrtali_g CountriesEditedbyJ.L.Innes and AJ-LHaron
5. Forestsln Sust_nable Mountmn Ik'vdopment:aState ofKnowledoeReportfor 2000Editedby M.Price and N. Butt
6. ForestsandLandscape_:LinkingF,colooT3,Sust_nabRlty andAestheticsEditedby S.R.J.Sheppardand H.W.Harshaw
7. Criteriaand Indicatorsfor SustainableForestManaoementEditedby J.Raison,A. Brownand D.Fltnn
8. The Impactof CarbonDioxideandOtherGreenhouseGaseson ForestEco_stemsEditedby D.EKarnosky.R. Ceulemans.G.E.Scarascia-Mugno_a and J.L.Innes

_r
/
_ The Impact of Carbon Dioxide and Other......... Greenhouse Gases on Forest Ecosystems
Report No. 3 of the IUFRO TaskForceonEnvironmentalChange
_ted
David E Karnosky__. Michigan Technological University, Houghton, Michigan, USA
Reinhart CeulemansUniversity of Antwerp, W_drijk,Belgium
Giuseppe E. Scarascia-MugnozzaUniversity of Tuscia, Fiterbo, Italy
and
John L. lnnesUniversity of British Columbia, Vancouver, Canada
CABIPublishingin association with
The International Union of Forestry Research Organizations (IUFRO)

CABI Publishing is a division of CAB International
CABIPublishing CABI Publishing_ " _ CAB International 10 E40th Street
............. _ Wallingford Suite 3203Oxen OXIO 8DE New York, NY 10016UK USA
Tel: �Œ�(0)1491832111 Tel:+I 212 481 7018
Fax:+44 (0)1491833508 Fax:+1 212 686 7993
gmail: cabi_cabi.org Email: cabi-nao_cabi.orgWeb site: www.cabi.org
© CAB International 2001. All rights reserved. No part of this publication may bereproduced in any form or by any means, electronically, mechanically, byphotocopying, recording or otherwise, without the prior permission of thecopyright owners.
A catalogue record for this book is available from the British Library, London, UK.
Library of Congress Cataloging-in-Publication DataThe impact of carbon dioxide and other greenhouse gases on forest ecosystems:report no. 3 of the 1UFROTask Force on Environmental Change / edited byDavid E Karnosky ... [et al.].
p. cm. - (IUFROresearch series ; 8)Includes bibliographical references (p.).ISBN O-85199-551-9 (all paper)
1. Forest plants-Effect of atmospheric carbon dioxide on. 2. Forestplants-Effect of greenhouse gases on. 3. Trees-Effect of atmospheric carbon di-oxide on. 4. Trees-Effect of greenhouse gases on. 5. Atmospheric carbondioxide-Environmental aspects. 6. Greenhouse gases-Environmental aspects. 7._Forest ecology. I. Karnosky, David, 1949- ]I. IUFROTask Force on EnvironmentalChange. IH. Series.QK753.C3 I48 2001577.3'276-dc21 2001018103
Published in association with:
The International Union of Forestry Research Organ|TaUons (IUFRO)c/o Federal Forest Research Centre
Seckendorff-Gudent-Weg 8A-1131 ViennaAustria
ISBN 0 85199 551 9
Typeset in 10/12pt Photina by Columns Design Ltd, Reading.Printed and bound in the UKby Biddies Ltd, Guildford and King's Lynn.

• Contents
Contributors vii " -
Preface ix
l The Impact of CO2 and Other Gree_aouse Gases on ForestEcosystems: an Introduction 1G.E. Scarascia-Mugnozza, D.E Karnosky, R. Ceulemans and].L. Innes
2 Effects of Greenhouse Gases on the Gas Exchange ofForest Trees 17D. Eamus and R. Ceulemans
3 The Impacts of Greenhouse Gases on Antioxidants andFoliar Defence Compounds 5 7G.K. Podila, A.1L Paolacciand NL Badiani
4 Above-ground Growth Responses of Forest Trees to Elevated
Atmospheric CO2 Concentrations 12 7B.E. Medlyn, A. P_y, C.V.M. Barton and M. Forstreuter
5 Influence of CO2 on the Growth and Function of Roots andRoot Systems 14:7NLE. Kubiske and D.L. Godbold
6 Impacts of Greenhouse Gases on the Phenology ofForest Trees 19 3
M.E. ]ach, R. Ceulemans and M.B. Murray

vi Contents
7 Effects of Elevated CO2 on Nutrient Cycling in Forests 23 7D.W. Johnson, R.]. Norby and B.A. Hungate
8 Impacts of Interacting Greenhouse Gases on Forest Ecosystems 253:: .; ........ D.F. Karnosky, E. Oksanen, R.E. Dickson and ].G. Isebrands
9 Modelling the Impacts of Greenhouse Gases on Forests 269F.Magnani and G. Matteucci
10 FACESystems for Studying the Impacts of Greenhouse Gases onForest Ecosystems 297D.F.Karnosky, B. Gielen, R. Ceulemans, W.H. $chlesinger,R.]. Norby, E. Oksanen, R. Matyssek and G.R. Hendrey
11 Knowledge Gaps in the Study of the Impacts of Elevated
Atmospheric CO2 and Other Greenhouse Gases onForest Ecosystems 325D.F.Karnosky, G.E. Scarascia-Mugnozza, R. Ceulemans and1. Innes
Index 3 41

Influence of CO2on the Growth and 5Function of Rootsand Root Systems
M.E. KUBISKE1. AND D.L. GODBOLD 2
1Mississippi State University, Department of Forestry, Mississippi State,USA; 2School of Agricultural and Forest Sciences, University of Wales,
• Bangor, UK]
*Present address: USDA Forest Service, Forestery Sciences Laboratory, 5985 Highway K,Rhinelander, W154501, USA.
0 CAB International 2001. The Impact of Carbon Dioxide and Other GreenhouseGases on Forest Ecosystems (eds D.F. Kamosky, R. Ceulemans, G. Scarascia-Mugnozzaand J.L. Innes) 147

148 M.E. Kubiskeand D.L. Godbold
5.1 Introduction
Root systems comprise up to half of total tree biomass; below-ground net primaryproduction may exceed 50% of total net primary production (NPP) in forestsand may be as high as 75% of total NPP (Vogt, 1991; Nadelhoffer and Raich,1992 ). As growth and metabolism of roots are entirely dependent upon photo-synthate suppled by leaves, alterations of the photosynthetic environment hasthe potential to affect root growth, structure and function profoundly. Tree rootsystems must remain in functional equilibrium with above-ground organs tooptimize the efficient use of scarce resources, i.e. water and nutrients, forgrowth and reproduction (Cannel and Dewar, 1994; Luo et al., 1994). Rootsare also important storage organs for plants, and function as capacitors forannual plant carbon (C) and nitrogen (N) budgets that not only influence treegrowth, but also play important roles in seedling survival and forest communitydynamics (Canham et o2., 1999). Because trees are the dominant organisms inany forest ecosystem, tree roots are an important source of organic carbon insoils that provides the primary energy for food webs and microbial processingof nutrients (Kfrner and Arnone, 1992; Diaz et al., 1993; Zak et al., 1993¢j2000a; Hungate et al., 1997; Klironomos eta/., 1997). Tree root systems serveas a platform for fungal and bacterial symbioses that are important for treenutrient and water relations (Koide, 1985; Allen, 1991; Lamhamedi et al.,
1992), ecosystem nutrient cycles (George et al., 1995; Guo et al., 1996;]entschke eta/., 2000b) and community dynamics (Miller, 1983; Janos, 1985).
In this chapter, we will _ briefly some important roles of roots at organ-ismal, community and ecosystem levels that have particular relevance to atmos-
pheric CO2 enrichment. Then, we will focus attention on relatively recent researchinvolving the physiological mechanisms of root responses to an increase in carbo-hydrate production and what might be expected from root systems as the atmos-

Influenceof CO2on RootGrowthand Function 149
pheric CO2 concentration continues to increase. Wewill then review experimentalevidence that either supports or refutes those expectations,including the produc-tion and turnover of fine roots, the role of roots in the deposition of organic Cinto
mils, root-system architecture, root absorbing capacity and symbiotic relationships.
5.1.1 Growth of roots and root systems
The additional C fixed under a CO2-euriched atmosphere is well known toincrease total tree biomass production, including root mass (Poorter, 1993;Stulen and den Hertog, 1993; Wullschleger et al., 1997; Curtis and Wang,1998). Beyond the absolute increase in root biomass, carbon-balance theory pre-dicts that trees should allocate a greater proportion of fixed C to roots to main-rain a functional balance between leaves and roots (Bloom et al., 1985). Whilemany authors have considered static measures of root dry mass as 'below-ground allocation', others have recognil.ed the limited importance of staric mea-sures of the root : shoot ratio (Stulen and den Hcrtog, 1993; Norby, 1994; Norbyet al., 1999). It is clear that standing root biomass is a very poor indicator of totalroot production due to the continuous production and abscission oftine roots,which may account for up to one-third of the total C allocated to roots (Persson,1979; Aber eta/., 1985; Nadelhoffer and Raich, 1992; Hendrick and Pregitzer,
1993). Carbon losses due to root respiration and exudation must also beaccounted for, and the proportion of a tree's Cbudget demanded by root sym-bionts may also be quite substantial. Thus, from the perspective of phloem lrans-port, below-ground allocation must also include all Closses from root systems,such as tlne root mortality, exudation, respiration and symbiotic development.None the less, static estimates of root : shoot biomass provide a snapshot in timeof the functional, assimilatory balance between roots and shoots.
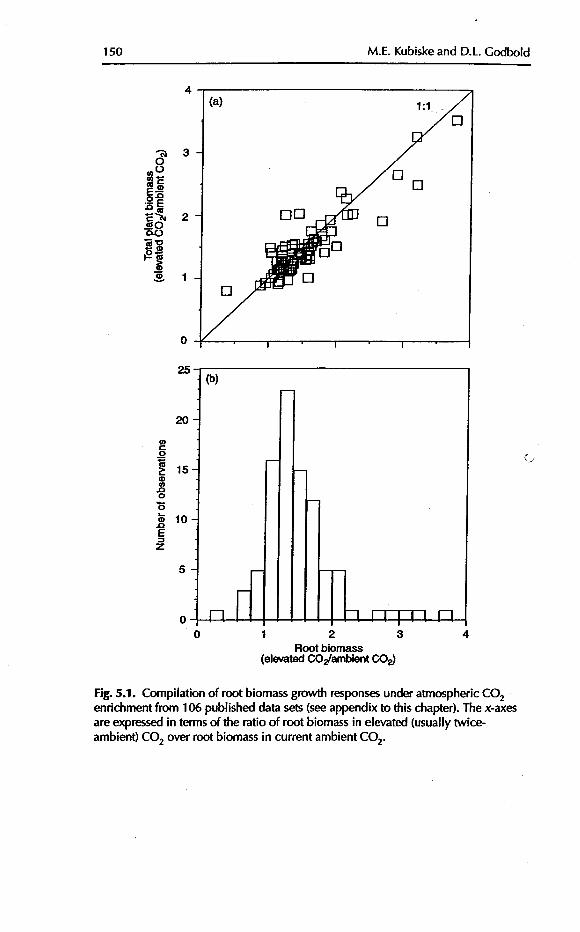
Several authors have found patterns for CO2-induced changes inroot : shoot dry mass partitioning that differed among herbaceous, crop andwoody plants (Farrar and Williams, 1991; Stulen and den Hertog, 1993). Areview of 106 experimental data sets on trees published since 1994 indicates
that root systems respond to elevated CO2 in the same proportion as the wholetree (Fig. 5.1a), and that increase is usually by a factor of 1.2-1.4 (Fig. 5.1b).These findings are entirely consistent with other reviews of the literature(Poortcr, 1993; Wullschleger eta/., 1997; Curtis and Wang, 1998; Norby et al.,1999). In trees, the 32-37% increase in dry mass under twice-ambient CO2(Poorter, 1993) was found to be allocated about equally to all organs(Wullschleger eta/., 1997). Curtis and Wang (1998) provided statistical confir-marion of these conclusions: their meta-analysis of over 500 studies showednostatistically supportable change in root : shoot dry mass partitioning by woody
species under elevated CO2. In the light of carbon-balance theory (Bloom et al.,1985), this is rather surprising; however, the functional need for a proportion:
ally larger root system under elevated CO2 may be questionable.
150 M.E. Kubiske and D.L. Godbold
4
: 0"-_ 3 (a)
_ O o
--_E
0 , , i
25(b)
2O
C
o :j
,,Q0
_ 10
Ez
5
0 1 2 3 4Root biornass
(elevatedCOL/ambientC02)
Fig. 5.1. Compilation of root biomass growth responsesunder atmospheric CO2enrichment from 106 published data sets(seeappendix to this chapter). The x-axesare expressedin terms of the ratio of root biomass in elevated (usually twice-ambient) CO2 over root biomass in current ambient CO2.

Influenceof CO2 on Root Growth andFunction 151
5.1.2 Functioning of roots and rootsystems
5.1.2. I Functional balance
As the assimilatory organs of trees, the allomefa'ic relationship between fine rootsand leaves is of particular interest. Photosynthetic C assimilation by leaves ishighly dependent on the supply of N from fine roots (Field and Mooney, 1986;Evans, 1989 ). !.ikewise, the uptake of N is dependent upon the availability of Csubstrates to roots (Oaks, 1993 ) and the availability of N in the soil is dependentupon the deposition of root-derived organic Cin the soil (Zaket aL, 1993,200Oa;Mikan et al., 2000). There is considerable empirical evidence that these factors
interact strongly to affect tree root responses to elevated CO2 (Norby _ aL, 1986;Mousseau and Enoch, 1989; Stulen and den Hcrtog, 1993; Rogers eta/., 1994;Pre_tzer et aL, 1995, 2000; Kubiske et o2., 1998). In a series of five experiments
investigating interactions between soil N availability and atmospheric CO2 con-centratiou, Pregitzer, Zak, Curtis and their co-workers found no significantchanges in the root : shoot ratio under elevated CD2,consistent with the analysispresented above (Zaketa/., 1993, 200Oa; Curtis eta/., 1994; Pregitzer et _, 1995;
Mikan et o2.,2DOO;Kubiske eta/., 2001). However, a compilation of results from
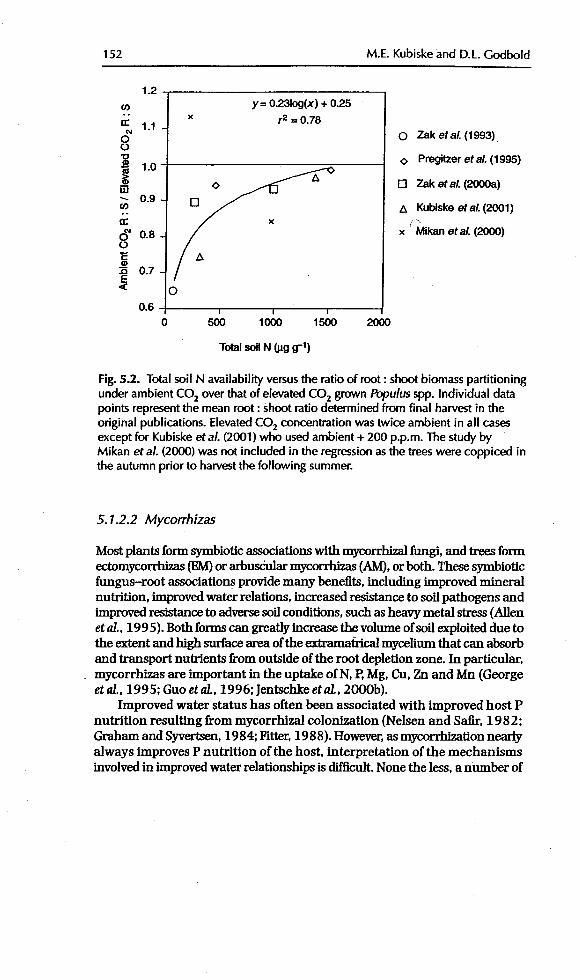
those experiments suggests that soil N availability and atmospheric CO2 con-centration interact to affect root : shoot biomass partitioning (Fig. 5.2). The
logarithmic relationship demonstrates that elevated CD2 may be expected to alterroot : shoot biomass partitioning in situations where soil resources are severelylimiting to growth (Stulen and den Hertog, 1993; Cm'tisand Wang, 1998). Onestudy didnot fitthis pattern: l_flkan et aL (2000) coppiced Populus tremuloides trees
following one growing season of exposure to elevated CO2 and harvested the
experiment the following snmmer. In that study, there were no siLmificant CO2effects on biomass partitioning among leaves, stems, coarse roots and fine roots,but coppiced trees in high N soil allocated 35% more biomass to stems and 15%less biomass to coarse roots compared to those in low N soil.
We still have much to learn about the balance between CO2 and N avail-ability vis-a-vis leaf and root physiology. For example, in the functional balance
hypothesis, allocation depends on both the environmental availability ofresources and the intrinsic assimilatory capacity or efficiency of roots and leaves
(Cannel] and Dewar, 1994). However, this model does not consider the abilityof leaves to ufiliT.eN, nor the ability of roots to utilize C;the former being knownto change with CO2 enrichment (see Chapter 9). Luo eta/. (1994) proposed analternative model that predicts a relative increase in the root fraction whenphotosynthesis increases more than total plant growth. While both of these
models call for better integration among studies on root and shoot growth and
function, predictions involving the long-term response ofroots to elevated CO2continue to be difficult.

Influenceof CO2on RootGrowthandFunction 151
5.1.2 Functioning of roots and rootsystems
5.1.2.1 Functional balance
As the asstm|latory organs of trees, the allometric relationship between fine rootsand leaves is of particular interest. Photosynthetic C assimilation by leaves ishighly dependent on the supply of N from fine roots (Field and Mooney, 1986;Evans, 1989). Likewise, the uptake of N is dependent upon the availability of CsubsWates to roots (Oaks, 1993 ) and the availability of N in the soil is dependentupon the deposition of root-derived organic C in the soil (Zak et aL, 1993, 2000a;Mikan et al., 2000). There is considerable empirical evidence that these factors
interact strongly to affect tree root responses to elevated CO2 (Norby eta/., 1986;Mousseau and Enoch, 1989; Stulen and den Hertog, 1993; Rogers eta/., 1994;Pregitser eta/., 1995, 2000; Kubisl¢ eta/., 1998). In a series offive experiments
investigating interactions between soft N availability and atmospheric CO2 con-centration, Pregitzer, 7.ak, Curtis and their co-workers found no significant
changes in the root : shoot ratio under elevated CO2, consistent with the analysispresented above (Zaket aL, 1993, 2000a; Curtis et a_L,1994; Pregitzer eta/., 1995;
Mikan et aL, 20()0; Kubiske eta/., 2001). However, a compilation of results fromthose experiments suggests that soft N availability and atmospheric CO2 con-centration interact to affect root : shoot biomass partitioning (Fig. 5.2). The
logarithmic relationship demonstrates that elevated CO2 may be expected to alterroot : shoot biomass partitioning in situations where soil resources are severelylimiting to growth (Stulen and den Hertog, 1993; Curtis and Wang, 1998 ). Onestudy did not fit this pattern: kfikan et aL (2000) coppiced Populus tremuloides trees
following one growing season of exposure to elevated CO2 and harvested theexperiment the following snmmer. In that study, there were no significant CO2effects on biomass partitioning among leaves, stems, coarse roots and fine roots,but coppiced trees in high N soil allocated 35% more biomass to stems and 15%less biomass to coarse roots compared to those in low N soft.
We slill have much to learn about the balance between CO2 and N avail-ability vis-d-vts leaf and root physiology. Forexample, in the functional balancehypothesis, allocation depends on both the environmental availability ofresources and the intrinsic assimilatory capacity or efficie_acyof roots and leaves(Cannell and Dewar, 1994). However, this model does not consider the abilityof leaves to utilize hi, nor the ability of roots to otilize C;the former being known
to change with COz enrichment (see Chapter 9). Luo et aJ. (1994) proposed analternative model that predicts a relative increase in the root fraction whenphotosynthesis increases more than total plant growth. While both of thesemodels call for better integration among studies on root and shoot growth and
function, predictions involving the long-term response of roots to elevated CO2continue to be difficult.
152 M.E. Kubiske and D.L. Godbold
1.2
y= 0.23log(x) + 0.25•- x r 2 = 0.78n- 1.1(_ O Zak eta/. (1993)0
O Pregitzereta/. (1995)
o [] Zak et al. (2000a)
_ 0.9.. A Kubiskeeta/. (2001)
(_ 0.8 x Mikan eta/. (2000)0Ca)
0.7E< 0
0.6 I I !
0 500 1000 1500 2000
Total soil N (v.gg-l)
Fig. 5.2. Total soil N availability versusthe ratio of root : shoot biomass partitioningunder ambient COz over that of elevated COz grown Populus spp. Individual datapoints representthe mean root : shoot ratio determined from final harvest in the
original publications. Elevated CO2 concentration was twice ambient in all casesexcept for Kubiskeetal. (2001) who used ambient + 200 p.p.m. The study byMikan et al. (2000) was not included in the regressionas the trees were coppiced inthe autumn prior to harvestthe following summer.
5.1.2.2 Mycorrhizas
Most plants form symbiotic associations with mycorrhizal fungi, and trees formectomycorrhizas (EM) or arbuscular mycorrhizas (AM), or both. These symbiotic
fungus-root associations provide many benefits, including improved mineralnutrition, improved water relations, increased resistance to soil pathogens and
improved resistance to adverse soft conditions, such as heavy metal stress (Alleneta/., 1995). Both forms can greatly increase the volume of soil exploited due to
the extent and high surface area of the extrama/rical mycelium that can absorb
and transport nutrients from outside of the root depletion zone. In particular,mycorrhizas are important in the uptake of N, P, Mg, Cu, Zn and Mn (George
eta/., 1995; Guo et aL, 1996; ]entschke et aL, 200Oh).
Improved water status has often been associated with improved host Pnutrition resulting from mycorrhizal colonization (Nelsen and Safir, 1982;
Graham and Syvertsen, 1984; Fitter, 1988). However, as mycorrhization nearlyalways improves P nutrition of the host, interpretation of the mechanisms
involved in improved water relationships is difficult. None the less, a number of

InfluenceofCO2onRootGrowthandFunction 153
authors have shown that host water status is improvedindependent of host Pnutrition (Aug6et al., 1986). Mechanismsofincreased host drought toleranceinclude increased roothydraulicconductivity (Koide,1985) and hyphal watertransport(Hardie,1985; FaberetaL 1991; Ruiz-LozanoetaL, 1995), alterationof stomata regulation (Allen etaL, 1982; Barea and Azcon-Aguilar, 1982) andosmotic adjustment (Aug6eta/., 1986).
Mycorrhizas may be an important component influencing plant-plantinteractions and thus community composition (Janos, 1985). At a plant com-munity level,mycorrhizalinfectionmaymediatecompetitionbetweenconiferoustree species by reducing competition between individuals and improvingcommunity P uptake (Perryet al., 1989). Hyphal linkages may also inducefacih'tationofone plant byanother, by providinga pathway for the transferof Cand nutrients (Simardet al., 1997; Perry,1999; Simard, 1999). This may beparticularly important in unconnected nurseplants. Transferofnutrients andcarbohydrates between host plants connected by hyphal linkages has beendemonstrated (Readeta/., 1985; Francis et al., 1986). However,the existenceof that phenomenon in arbuscular mycorrhizashas been challenged recently(Fitteret a/., 1999 ). Itis unclear to what extent these transfers influence treegrowth, and they maybe mostimportantin the maintenance of the fungal asso-ciations themselves (Fitteret al., 1998).
5.2 Root growth in elevated CO 2
Rootgrowth, and thereforethe growth responseto elevatedCO2,consistsofrootlength extension (primary growth) that permits exploration ofbroader regionsofsoil, branching that permits more thorough exploitation ofsoil patches, andsecondary growth that allowsroots to enlarge for more effectivetransport andstorage. The functional architecture of the root system is, in large part, a bal-ance between extension and branching (Fitter et al., 1991 ). As with all plantfunctions, root growth is under both genetic and environmental control.Genetic factors include the sequence in which lateral root primordia are formed,the species' propensity for development of adventitious roots, and root apicalbifurcations (Charlton, 1991). Edaphic factors such as soil texture, structureand heterogeneity are known to exert considerable influence on root architec-ture (Theodorouand Bowen, 1993 ).Forexample, the abundance ofNO3-in thesoil may act as a signal rather than a nutrient for apical meristem activity, withbiochemical similarities to the auxin response (Zhang et aL, 19 99). Thoseauthors proposed that the mode of action was mediated by the C/Nratio in theroot tissues, so that if the internal N concentration became too great, lateralroot elongation was systemically inhibited rather than locally stimulated. Asgreater Cassimilation under elevated CO2 suppliesmore substrate for growthprocesses and increases tissue C/Nratios, any occurrences of NO 3- inducedlateral root inhibition may becomeincreasingly rare. In any case, soil conditions

154 ' M.E. Kubiskeand D.L. Godbold
in general are likely to exert considerable influence upon the response of root
systems to atmospheric CO2 concentrations.
5.2.1 Physiologyof primary growth
Because of the importance of root extension and branching to the overall effi-ciency of root systems, primary growth of roots has been studied much moreextensively than secondary growth, particularly with respect to atmospheric CO2enrichment. Primary growth of roots involves a sequence of processes includ-ing cell division, elongation and differentiation. Cell divisions generally occurwithin the first several millimetres from the root meristem, and elongationoccurs within 1 or 2 cm of the tip. Elongation is closely coupled with differenti-ation, which includes the development of lateral root primordia, to the extentthat lateral root primordia are often formed within the zone of active cell elon-gation (Klepper, 19 8 7; Charlton, 19 91). Consequently, meristematic activityinfluences both elongation and the potential for branching.
5.2.1.1 Cell division
Root primary meristems are well-organized assemblages of rapidly dividing cellsthat vary by taxon in their complexi_ Roots elongate when meristematic initialsin the root tip divide and produce meristematic derivatives that further divide5-7 times (Luxovfi and Ciamporovfi, 1992). Division of meristematic initialsoccurs with surprising regularity, such that workers have accurately predictedthe morphology and structure of roots based on cell divisions in the apicalmeristem. Perturbations in the rooting environment, however, can induce reor-ganization of meristematic activity.
The cell cycle, that is, the rate at which cells progress from mitosis, through
the stages of pre-DNA synthesis (G1), DNA synthesis (S), post-DNA synthesis(G2) and back to mitosis again, is sensitive to sucrose concentration (Soni et al.,1995). Cell division is one stage of the cell cycle. The transition stages from Gx
to S and from G2 to mitosis are carbohydrate-dependent (Van't Hof, 1985);when cells are starved of carbohydrate, cycle arrest occurs at either the G1or G2phases. ATP is also required for cell division (Barlow, 19 8 7). Oxidative phos-phorylation, as driven by respiration of imported sucrose, is the primary sourceof ATP in roots. Lambers (1987) estimated that the construction of i g of rootdry mass requires 21.54 mrnol of ATP.
These aspects of cell division beg the question: ff the cell cycle and cytoki-nesis can be arrested by sucrose (and ATP) starvation, does the availability ofsucrose influence the rate of the cell cycle and the rate of cell division? The cell
cycle in maize roots was arrested by sucrose starvation, but on reintroductionof exogenous sucrose, the cycle rate was correlated with sucrose concentration(Evans eta/., 1994). The cell cycle in root tips of Dactyls glomerata was only

Influenceof COz on RootGrowthandFunction 155
slightly accelerated by shoot exposure to elevated CO2 but, more importantly,the proportion of rapidly dividing cells increased by factors of 1.3-1.9 under ele-
vated CO2 (Kinsman et aL, 1997). The mechanism by which sucrose supplymight stimulate the cell cycle is unclear, as little work has been done on this.None the less, growth is most certainly closely coupled with carbohydrate sup-ply as a substrate for respiration, cell-wall synthesis and carbon skeletons. Asthe substrate for carbohydrate production, elevated CO2 may be expected tostimulate growth of plant roots in part by increasing the rate at which new cellsare produced. This leads to the next question: does cell elongation, as the com-plement to cell division, also respond to elevated CO27
5.2.1.2 Cell elongation
Taylor and co-workers (Ferris and Taylor, 1994; Taylor et al., 1994, 1995;
Crookshanks et aL, 1998) reasoned that increased growth under elevated CO2must be the result of either increased meristematic cell division, increased rate
of cell expansion or both. They used the Lockhart equation to investigate thebiomechanics of root responses to elevated CO2 at the cellular level.
The process of cell expansion has been expressed simply as:
1 dV- _ (P-X")i
V dt (5.1)
where V is the cell volume; P, turgor pressure; Y, the yield threshold (i.e. turgotpressure at which the cell wall is forced to expand); and _ is a coefficient relat-ing growth rate to the turgor in excess of Y (the cell-wall extensibility)(Lockhart, 1965; Cosgrove, 1985). Expansion occurs when water flows intocells under osmotic tension and the cell wall responds by relaxing turgor.
One possible way in which dV/dt might increase is for the cell-solute concen-tration to increase, which would increase the osmotic gradient for water influx,
thereby increasing the difference (P - 10. Increased cellular non-structural car-bohydrate concentration under elevated CO2 is an attractive hypothesis thatwould account for this mecbani._m. Root tips ofPopulus deltoides x Populus nigrahad a tendency forhigher solute accumulation and turgot pressure under elevated
CO2 (Bosae et aL, 1995). However, the data of Taylor et aL (1995) indicated nochange in vacuolar solute concentration of Arabidopsis thaliana roots, butincreased rates of cell expansion within the growing region of the root tip. In awater-stress experiment, the roots of sugar maple (Acer saccharum), Americanplane lree (Platanus occidentalis) and sweetgum (Liquidambar stgraciflua) showed
no CO2-related change in total solute concentration (Tsehaplin,_ki et al., 1995).In that study, there were CO2-related increases in fructose, glucose and sucroseconcentrations under drought conditions, but those free carbohydratesaccounted for a small proportion of total osmolites.

156 M.E. Kubiskeand D.L. Goclbold
Taiz (1984) argued that solute concentration does not normally limit cellexpansion, rather, growth may be controlled by cell-wall properties that define_b,and are under the influence of plant growth substances. During periods ofactive cell expansion, cell-wall extensibility generally increases due to auxin-mediated, wall-loosening enzymes (Taiz, 1984; Cosgrove, 1985). Four species
of grassland herbs had significant increases in cell-wall exteusibility under ele-
vated CO2 (Ferris and Taylor, 1994). One enzyme that has been implicatedin cell-wall loosening is xyloglucan endotransglycosylase (Fry et al., 1992).Taylor et al: (1994) measured increased activity ofxyloglucan endotransglyco-sylase in Phaseolus vulgaris leaves under elevated CO2, although activity of thisenzyme has not been studied in roots. However, the transcription of ribosomalproteins involved with cell growth and development in maize root tips was reg-dated by sucrose, glucose and fructose concentrations (Chevalier et aL, 1996).Thus, stimulation of cell elongation rate may be the direct result of photosyn-
thate supply to root meristems. Unfortunately, there is no evidence to supportthese relationships in the roots of woody species.
5.2.1.3 Cell differentiation and root branching
How elevated atmospheric CO2 might influence root architecture (root lengthand branching) is a difficult matter to resolve. Root-system branching is com-prised of any combination of several processes, including root bifurcation at theapical meristem, the differentiation of endodermal cells into lateral root pri-mordia, and the 'meristemi_,.ation' of quiescent endodermal cells that formadventitious roots even following secondary growth (Charlton, 1991).
Bifurcation is a common occurrence in Pinus roots when cell division of the
apical meristem is interrupted. When growth resumes, divisions occur aroundtwo foal rather than one (Badow, 1987). At this time, there is no reason to expectthat increased carbohydrate supply to root meristems would induce increasedrates of bifurcations. One possible exception, however, is that bifurcations are
relatively common in ectomycorrhizas. If elevated CO2 increases carbohydratesupply for mycorrhizas, then the incidence of root bifurcation may also increasewith the rate of infection. This speculation is consistent with findings of an
increased number of mycorrhizal tips under elevated CO2, as discussed below.Lateral root primordia generally develop within 1-2 cm of the root tip,
within the region of active cell dongation, from pericycle parenchyma that aresituated near to protoxylem strands (Klepper, 198 7; Charlton, 1991). Thisjuxtaposition ofprotoxylem and pericycle parenchyma occurs at the xylempoles (with 3-6 xylem poles being common in woody species) (Klepper, 1987;Luxovfi and Ciamporovfi, 1992). Consequently, lateral roots form in longitudi-nal ranks, although not all lateral primordia develop into lateral roots (Klepper,1987; Barlow and Adam, 1988; Charlton, 1991). Since the initiation of lateral
root primordia occurs with striking regularity within ranks, some authors have

InfluenceofCO 2 on RootGrowthandFunction 157
suggested that it is regulated as part of the general pattern of cell division anddifferentiation in the root tip (Charlton, 1991).
In light of this possibility,it is attractive to speculate that the accelerationof root cell division and elongation rates discussed above might somehow berelated to an increased rate ofdifferentiation into lateral primordia. The spac-ing oflateral root primordia oftomato (Lycopersiconesculentum)roots in culturemedia was unaffected by modification of exogenous sucrose concentration,despite significant alteration ofroot extension rate and an increase in the lengthofthe zone bearing unemerged primordia (Barlowand Adam, 1988). Charlton(1991 )elaborated on this by stating that the relative positions of lateral root pri-mordia initiation in angiosperms are not fixed,even within the same root. Nonethe less, another study indicated that the initiation of root primordia was stim-ulated by the addition of exogenous sucrose (Evans et d., 1994). Extrapolation
of these findings to roots with increased sucrose concentrations due to CO2enrichment leads to the conclusion that CO2enrichment stimulates the forma-tion and emergence of lateral root primordia.
5.2.2 Rootsystemgrowthanddevelopmentunderfield conditions
The data ofTaylor et al. (1995) for Populuseuramericanacv.Primo clearly indi-cated that root cells within 7 mm of the root tip were expanding faster underelevated compared to ambient CO2, but that there was no difference in the sizeof mature cells.Similarresnlts were reportedfor Fraxinusexcelsior(Crookshankseta/., 1998). Therefore.increased root growth under elevated CO2appears tobe a function of increased rates of production and expansion ofcells near theroot apex, rather than increased cell size. Thisis an importantpoint with directbearing on C and N construction costs of new roots. Increased cell numberstranslates to increased quantities of N-rich constituents (cell nuclei, nucleicacids, ribosomes, etc.) as well as C-rich cell walls, with the functional propor-tion of Cand N in roots remaining relatively constant. If true, then increased
production of new roots with CO2 enrichment would require proportionalinputs of construction resources. Other factors remaining constant, the cost ofproducing each gram of new primary root under elevated CO2should notchange appreciably.
In terms ofCcosts, the very small termina'lroots are most efficientat nutri-ent uptake, most expensive to maintain, but rather inexpensive to construct(Yanai, 1994; Pregitzer eta/., 1997). Moreover,small-diameter rootswith highspecificroot length (m g-I) havemore absorptive length and tend to be shorterlived than low specific root length roots of the same mass (Eissenstat, 1992).Thus, it is more favourable for trees to invest Cin the construction of new butshort-lived roots with high specificroot length in previously unexplored softpatches, rather than in the more expensive maintenance of roots in softdeple-tion zones.This has implications for modifications ofroot architecture with CO2

158 M.E. Kubiskeand D.L. Godbold
enrichment. As the availability of C for root construction increases, the best
economic alternative would be to invest in the area of highest return: the pro-duction ofnew, fine roots. An increase in Cavailability may preferentially favour
the production of efficient, nutrient-absorbing terminal roots, thereby improv-ing the nutrient-absorbing capacity of the root system. However, if otherresources are limiting for rapid construction of new roots, additional C may beinvested in maintenance of existing roots, or mycorrhizas with lower N costs.
5.2.2.1 Root length extension and branching
Total standing root mass generally increases with elevated CO2 in proportion tototal tree growth rate (Fig. S. 1; Farrar and Williams, 1991; Stulen and den
Hertog, 1993; Wullschleger etal., 1997; Curtis and Wang, 1998; Norby et al.,1999). Specific root lengths generally either decrease, owing to accumulations
of starch or actual increases in root diameter, or stay the same with CO2 enrich-ment (Table 5.1). The reported decreases in specific root length are largely dueto accumulation of non-structural carbohydrates (Larigauderie et aL, 1994;Crookshanks et al., 1998). Regardless of specific root length responses, trees
consistently respond to elevated (302 with increases in total length of root sys-tems (Table 5.1; K/_rner and Arnone, 1992; Prior eta/., 1997).
Reviewing several studies on herbaceous plants, Stulen and den Hertog
(1993) concluded that elevated CO2 increased the number of growing fine-roottips, but that axial extension was not altered. For trees, authors have reportedvarious combinations of longer individual roots and greater proliferation of pri-mary laterals that contribute to greater root system length (Table 5.1). Populusroots repeatedly exhibited significant quadratic effects in root length responses
to elevated CO2, indicating increased rates of Free root length production(Pregit_er et al., 1995; Kubiske et al., 1998). Consistent with the conclusions ofStulen and den Hertog (1993), significant increases in total root system lengthwere concurrent with increased numbers of fine roots in species of Populus,Fraxinus, Quercus and Pinus (Zak et al., 1993; Crookshank._ et al., 1998). Thetotal length of visible Populus deltoides × Populus nigra roots was significantly
greater under elevated CA)2, which was attributed solely to an increase in thenumber of roots, with no change in the extension rate of individual root tips(Bosac et al., 1995). A common interpretation of such findings has been thatroot systems will explore the soil volume more effectively and therefore absorp-tion capability will increase. Not all studies have these results, however. The rootsystem of Pinus taeda had significant increases in the length of second-order lat-erals and in the ratio of second-order : first-order root length, but no change inthe number of lateral roots per centimetre of parent root (Larigauderie et aL,
1994). In another study involving P. taeda, the fine root biomass as a proportion
of lateral root biomass decreased with elevated CO2 (Gebauer et al., 1996).
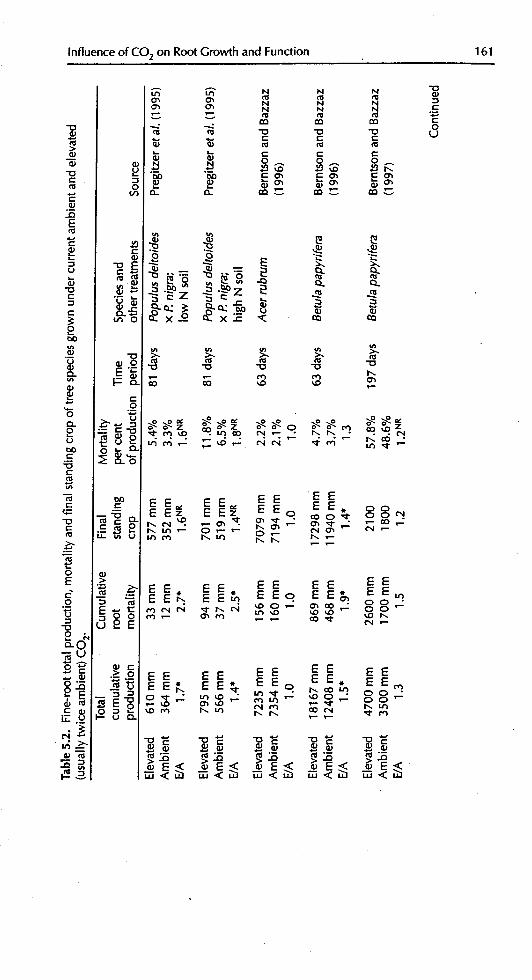
Influence of CO2 on Root Growth and Function 159
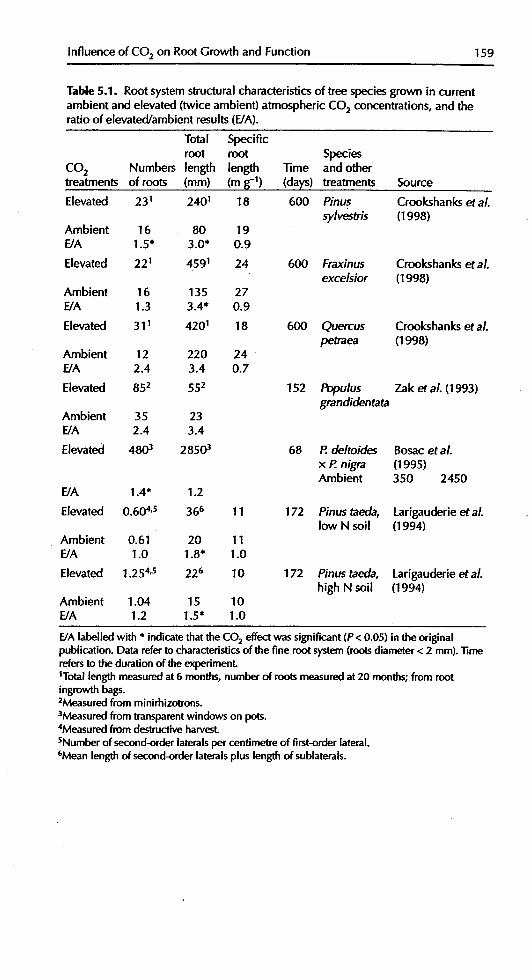
Table 5.1. Root systemstructural characteristics of tree speciesgrown in currentambient and elevated (twice ambient) atmospheric CO2 concentrations, and theratio of elevated/ambient results(E/A).
Total Specificroot root Species
CO 2 Numbers length length .time and othertreatments of roots (ram) (m_ -1) (days) treatments Source
Elevated 231 2401 18 600 Pinus Crookshanks et aL
sylvestris (1998)Ambient 16 80 19E/A 1.5" 3.0* 0.9
Elevated 221 4591 24 600 Fraxinus Crookshanks etal.excelsior (1998)
Ambient 16 135 27E/A 1.3 3.4* 0.9
Elevated 311 4201 18 600 Quercus Crookshanks et al.petraea (1998)
Ambient 12 220 24E/A 2.4 3.4 0.7
Elevated 852 552 152 Populus Zak et al. (1993)grandidentata
Ambient 35 23E/A 2.4 3.4
Elevated 4803 28503 68 P.deltoides Bosac et aL
x P.nigra (1995)Ambient 350 2450
E/A 1.4* 1.2
Elevated 0.604,s 366 11 172 Pinus taecla, Larigauderie et al. . .low N soil (1994)
Ambient 0.61 20 11E/A 1.0 1.8" 1.0
Elevated 1.254,5 226 10 172 Pinus taecla, Larigauderie et aLhigh N soil (1994)
Ambient 1.04 15 10E/A 1.2 1.5" 1.0
E/Alabelledwith * indicatethattheCO2 effectwassignificant(P< 0.05) intheoriginalpublication.Datarefertocharacteristicsofthefine rootsystem(rootsdiameter< 2 mm)..timerefersto thedurationof theexperiment.1Totallengthmeasuredat 6 months,numberof rootsmeasuredat20 months;from rootingrowthbags.2Measuredfrom minirhizotrons.
3Measuredfromtransparentwindowson pots.4Measuredfromdestructiveharvest.SNumherof second-orderlateralspercentimetreof first-orderlateral.6Meanlengthofsecond-orderlateralspluslengthof sublaterals.

160 M.E. Kubiskeand D.L. Godbold
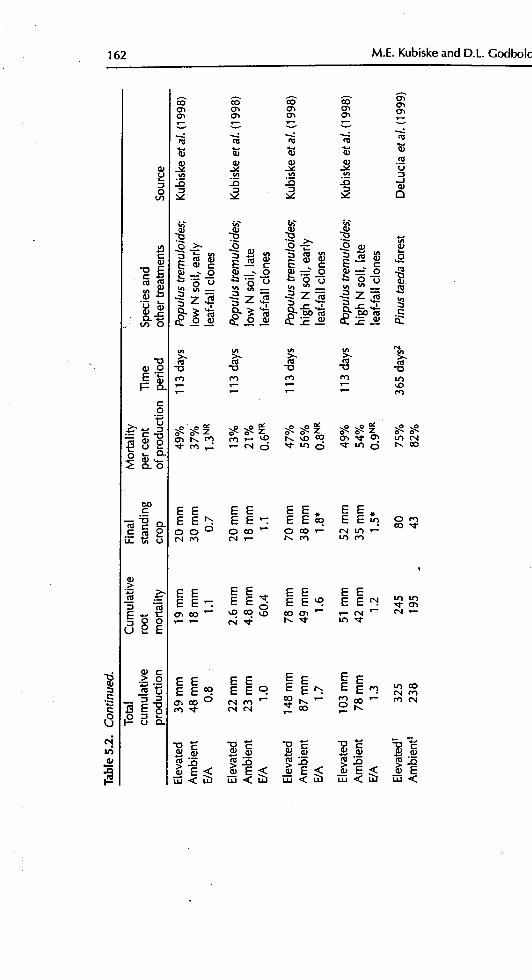
5.2.2.2 Production and turnover of fine roots
If tree root systems do, in fact, respond to elevated CO2by the greater productionof fine roots, then fine-root turnover might also be expected to change. The pro-duction and turnover of fine tree roots is a critical issue with implications for
ecosystem nutrient cycling and C sequestration, but few studies have addressedit (Pregitzer eta/., 2000). Of those that have, nearly all have reported substantialincreases in fine-root length production and mortality with CO2 enrichment
(Table 5.2). About half of the studies reported a significant change in the demo-graphic relationship between fine-root production and mortality, but the resultsare greatly varied. Pregitzer eta/. (1995) found significantly reduced fine-rootlifespans ofP. deltoides x P. nigra under elevated CO2, but Kubiske et aL (1998)found no such responses in P. tremuloides. The rates of production and mortalityofPmusponden_sa mycorrl_za tips was significantly greaterwith CO2 enrichment,but there was no change in lifespan (Rygiewicz et aL, 1997). Elevated CO2 had noeffect on root systems ofAcer rubrum, but Betulapap!trifera had a non-si_r_ificantincrease in fine root mortality by a factor of 1.3 (Berntson and Ba_ 1996). In
contrast, Tingey eta/. (1997) reported that P.ponderosa had increased root life-spans by about 30 days with elevated CO2. They went on to suggest that rootlffespan is a function of the amount of carbohydrate supplied to fine roots at the
time of primary growth, thus greater carbohydrate supply under CO2enrichmentshould lead to longer fine-root lifespans. However, this is further complicated byinteractions with ectomycorrhizas. Elevated CO2 may alter ectomycorrhizal
species composition (see below) and field observations of Fagus s!tlvatica haveshown that different ectomycorrhiza species vary in lifespan (Wittig, 1999).
Changes in fine-root turnover and lifespan do not necessarily reflect a CO2-induced increase in C allocation to roots if the increases in production are
consistent with the normal, allometric dynamics of C allocation versus plant
size. The demographic relationship between fine-root production and mortal-
ity is logarithmic, so that a larger proportion of fine-root mortality may beexpected as the population of fine roots increases (Berntson and Bazzaz, 1996).Apparent shifts in below-ground allocation under elevated CO2may simply rep-resent normal, size-dependent changes in C allocation as trees grow larger
(Tissue et al., 1997). Thus, the extent to which increased fine-root mortality isa function of direct treatment effects on fine-root lifespan or larger plant size is
not clearly defined. These results have important implications for ecosystem-level soil processes, as discussed elsewhere in this volume. Although the extentto which fine roots may retranslocate N and Cprior to abscission is _mknown,
increased mortality under CO2 enrichmen_ is directly responsible for enhanc-ing soil C deposition.
Influenceof CO2on RootGrowthandFunction 161
162 M.E. Kubiskeand D.L Godbold
E E _" E E'- E E_o = = _ o_• " ° ° CO0 0 0 0 0 oO _'" 0 aO "_" r,,I L_ _--

Influenceof COz on RootGrowthandFunction 163

164 M.E. Kubiskeand D.L. Godbold
5.2.2.3 Root exudation and respiration
In addition to fine-root turnover, rhizodeposition of C includes water-solubleexudates such as sugars and organic acids, and insoluble materials such assloughed tissues and mucilage. These exudates can account for up to 20% offixed C (Merbach et al., 1999) and so constitute an important, but often over-
looked, component of below-ground plant C budgets. The soluble materials arereadily utiliT.e_lby soft microbes, so the materials are important for soft ecology(Martens, 1990). Several studies have examined rates of C rhizodeposition
under CO2 enrichment, usually by measuring bulk transfer of labelled Cinto thesoil solution. Ineson et al. (1996) grew Betula pendula seedlings in soft that wasisotopically labelled with a C4 513C signature. After 186 days under elevated
CO2, the C3 B. pendula seedlings added 1.5 g C to the soil, which represented25% of live-root C, compared to 0.6 g or 17% of live-root C added to the soft
under current ambient CO2. Similar findings were reported by Rouhier et al.(1994), who concluded that C allocation below ground may increase with nodetectable change in root : shoot dry mass partitioning. In contrast, Norby eta/. (1987) found that soluble exudes per gram fine-root mass ofPinus echinata
grown in pots decreased with CA)2 enrichment, but that total soluble exudatesincreased as a function of greater fine-root production. More recently, elevated
CO2 did not affect the allometry of either biomass or 14Callocation by Populustremuloidesrooted in open-bottom mesocosms (Mlkan et al., 2000). However, aswith fine root turnover, a clearer understanding of the aUometric relationshipsbetween root system size and activity must precede a definitive answer to
whether elevated CO2 stimulates a shift in below-ground allocation. Moreover,clearer definition of exudate constituents is needed because of the various roles
they play in microbial and geochemical processing of C and nutrients.
Poorter et al. (1992) reviewed root respiration under elevated CO2 andfound no significant response. However, their study included no data from
woody species. Since then, a number of investigators have attempted to mea-
sure respiration of fine roots from trees grown in elevated CO2. Total soft respi-ration was enhanced by 19--44% with CO2 enrichment in studies on P.tremuloides (l_rakanet al., 2000; Pregitzer eta/., 2000). While it is not possible toseparate the contributions of roots and microbes to total soil respiration by mea-
suring CO2 efflux at the soft surface, the evolved C is ultimately of plant originand the increase in respiration represents an increase in below-ground C flux(Mikan et al., 2000). Moreover, soft respiration increases were closely linearlyrelated to fine-root (r 2 = 0.87), coarse-root (r2 = 0.96) and total root
(r 2 = 0.96) biomass, indicating that the bulk of soil CO2 efflux is influenceddirectly by root biomass (Pregit_cr et al., 2000).

Influenceof COz onRootGrowthandFunction 165
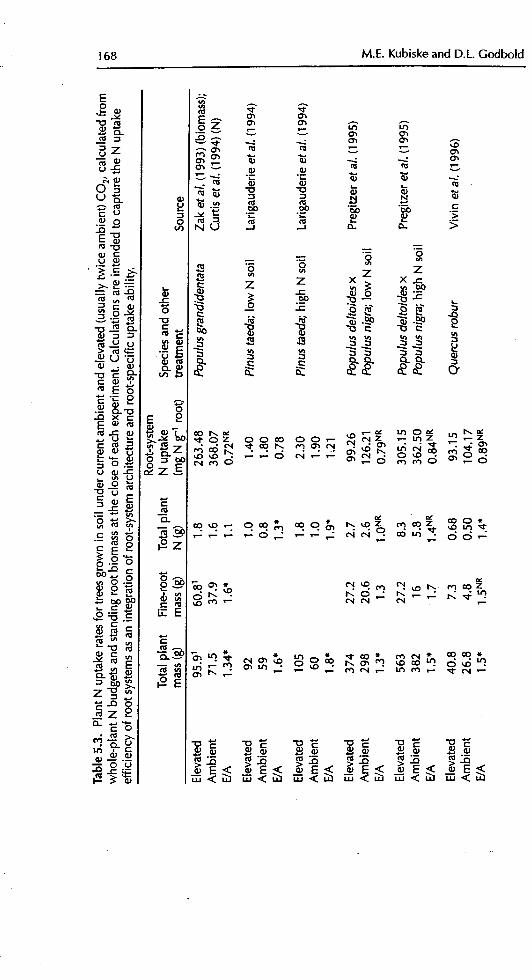
5.3 Root function under elevated CO 2
Root-system function is a complex integration of root-system architecture andthe absorptive capability of individual roots. The functional response of roots to
elevated CO2 may be more closely related to root-system architecture and mor-phology than to changes in absorptive physiology (Stulen and den Hertog,1993; Larigauderie et al., 1994; Rogers et al., 1994; Bosac et al., 1995;BassiriRad et M., 1996a; Norby eta/., 1999). Authors have reported increased
N uptake at elevated CO2 that they attribute to changes in root architecture thatenable plants to explore volumes of soft more thoroughly (Larigauderie et aL,1994; BassiriRad et aI., 1996a). None the less, experiments on the primaryfunction of roots, i.e. uptake physiology, have produced some interesting results.
5.3.1 Nutrient relations
5.3.1.1 Root-specific N uptake
Carbohydrates are important effectors of NO_ uptake, such that the addition ofglucose to excised roots can increase nitrate reductase activity (Oaks, 1993).
The metabolic cost of protein synthesis with NO_3as the N source was found tobe about 35% more expensive (in glucose equivalents) compared to protein syn-
thesis with NH_ as the N source (Zerihun et aL, 1997). Rideout and Raper(1994) reported that shoot-to-root carbohydrate flux was highly correlated with
root NO] uptake. In light of these reports, here we ask the question, doeselevated CO2 facilitate N uptake metabolism?
There is mixed evidence that N uptake physiology may be enhanced by
elevated CO2 in conifers (Larigauderie ¢tat, 1994; BassiriRad et at, 1996a), butwoody angiosperms have exhibited no such response (Vivin et al., 1996;
BassiriRad et at, 1997b; Rothstein et at, 2000). Rlevated CO2 resulted in greaterN uptake per root dry mass of Pinus taedagrown in high-N soft, but less N uptakerate in low-N soft (Iarigauderie et at, 1994). In another study, P. taedaunder ele-
vated CO2 had increased maYimum rates for NO_auptake (i/max)by 65 and 58%in fertilized and unfertilized treatments, respectively, which coincided withincreased non-structural carbohydrate concentration of the roots (BassiriRad
et at, 1996a). Uptake kinetics were not altered for NH_ uptake in that study, norwhen NH_ was the sole source ofN (BassiriRad et al., I996b).
These studies suggest that increased carbohydrate availability to roots
under elevated CO2 may increase the maximal capacity of roots for NO_uptake, but not necessarily the steady-state uptake ofN (BassiriRad et aL,1996a). ff increased carbohydrate availability increases the capacity of roots
to reduce NO_ and/or synthesize proteins, then the maximum reaction veloc-ity (Vmax) would be expected to increase with an increase in enzyme availabil-ity (e.g. nitrite reductase, nitrate reductase, glutamine synthetase), whereas

166 M.E. Kubiskeand D.L. Godbold
enzymatic affinity (Kin) would not be expected to change. Furthermore, the bio-mechanics of C source-sink relations tells us that sink strength drives C trans-
port and allocation, and not source production (Dickson, 1989, 1991). Thus,ffuitrate reduction and assimilation by roots is limited by internal carbohy-
drate supply, COz enrielament may help ameliorate such limitations; however,increased carbohydrate production is not likely to drive an increase in root-spe-cific N uptake.
Another important consideration is the proportion of NO_3assimilated inroots versus the proportion transported and assimilated in shoots. It is meta-bolically less expensive for plants to assimilate NO_3in shoots for several reasons.First, it is less expensive to passively transport NO_3in xylem sap from roots toshoots, than to actively transport sucrose in phloem from shoots to roots.Second, reductant from the photosynthetic light reactions, rather than from
respiration of glucose, may be utilized directly to reduce NO_3in leaves (Zerihtmeta/., 1997). However, photosynthetic down-regulation with CO2 enrichmentis well documented, and increased carboxylation rates with CO2 enrichmentmay demand a larger proportion of reductant from the light reactions. Thus, ifany particular species depends heavily upon NO_ reduction in shoots, then
NO_3uptake kinetics of excised roots could also be subject to negative feedbackinhibition as NO_ accumulates in root tissues (d. Rothstein et at, 2000).
The relative importance of these caveats is unclear, but certainly not all stud-
ies report a CO2-related change in root-specific N uptake (for example, BassiriRadet at, 1997a; DeLucia et aL, 1997). Elevated CO2 had no effect on specific root
15NO_3uptake ofQuercus robur despite a significant increase in newly fixed z3Callocated to fine roots (Vivin et at, 1996). The authors noted that it was unclearwhether the lack of N uptake stimulation resulted from soil N limitation or root
physiology. Similar results were found by Rothstein et at (2000) for P.tremuloides.But, in solution culture, which would seemingly preclude any rooting mediumN limitation, Larrea tridentata secdFmgs had inhibited NO_3uptake rate under ele-
vated CO2, and decreased total plant N (BassiriRad et at, 1997b), as did Pinustaeda and P.ponderosa (Friend et at, 1998).
5.3.1.2 Root-system N uptake
Root uptake kinetics are more complicated than simple relationships with car-bohydrate status; they depend also upon the relative availability of nutrients insoft, which may also change under a C02-enriched atmosphere. None the less,root symbionts are highly dependent on C from roots to process N and othernutrients. The roots of nitrogen fixers such as legumes and Alnus spp. may
respond quite differently to elevated (302 in terms of N budget compared to otherspecies. Root nodule mass and N fixation were stimulated by elevated CO2 inAlnas glutinosa (Vogel et at, 1997), but NO_- uptake of non-nodulated Prosopisglandulosa was unresponsive to elevated CO2 (BassiriRad eta/., 199 7b). Alnusglutinosa plants under elevated CO2 had 40% greater N content, 50% greater

Influenceof CO2on RootGrowthand Function 167
leafN content and 16% greater maximum Rubisco activity, all of which wereattributed to increased fixed C fuelling nodule nitrogenase activity (Vogel and
Curtis, 1995 ).The B1sN from elevated C02-grown tissues was similar to that fromtissues grown in ambient CO2, suggesting that trees derived similar proportionsof their total N budget from root symbionts (Vogel et al., 199 7). However,the authors suggested that the increase in N fixation lagged well behind rootinoculation, which would discount 092 stimulation of non-symbiotic N uptake.Similar results were reported by Thomas et al. (1991) for a tropical woodylegume. Uselman et al. (1999) found no stimulation of dissolved organic N fromsymbiotic Robinia pseudoacacia seedlings under elevated C02; thus, whilesymbiotic N fixation may be stimulated under elevated CO2, it was utilizedalmost exclusively by the host plant, providing little benefit to surrounding
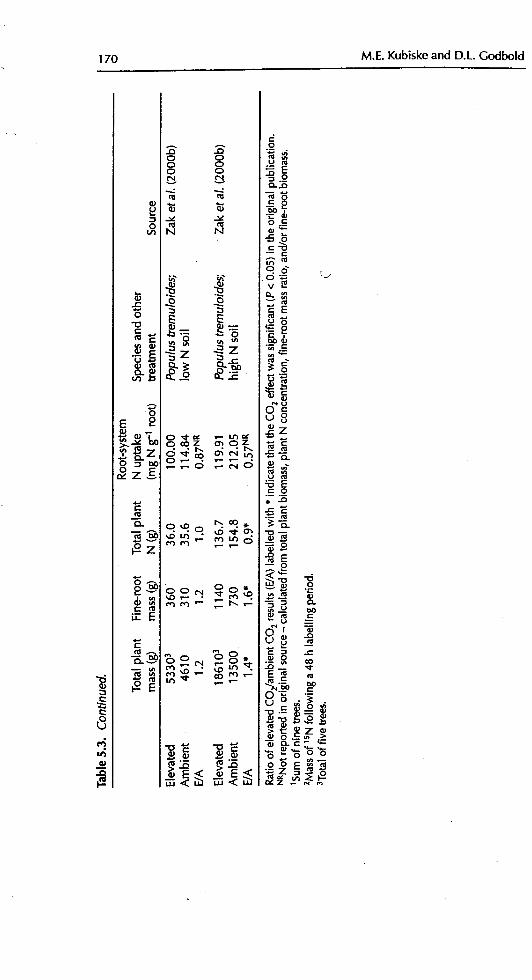
vegetation.To assess the N uptake efficiency of non-N-fix'ing tree species, we calculated
root-system N uptake efficiency for a number of studies as the total plant N con-tent per unit fine-root mass (Table 5.3). In most studies, total plant N uptakewas outpaced by biomass accumulation, such that total plant N content perfine-root blomass decreased. Similarly, in studies where root-specific NO_ uptake
increased under elevated CO2, it was almost always outpaced by increased bio-mass accumulation, resulting in a smaller total plant N concentration
(BassiriRad eta/., 1997a; Friend et al., 1998 ). As the proportion of fine-rootturnover may increase with CO2 enrichment (Table 5.2), fine-root mortalitymay carry with it a larger proportion of the total plant N budget. A crude cal-culation was made using the data of Pregitzer et aL (1995). Assuming that fine-root biomass mortality occurs at the same proportion as fine-root lengthmortality, then elevated CO2 resulted in a twofold increase in the percentage oftree N budget lost in fine-root turnover (Table 5.4). However, the losses were stillconsiderably less than 1% of the total N budget. Another assumption in thesecalculations was that of no N retranslocation prior to senescence. This is prob-
ably not a valid assumption, the violation of which would only serve to decreasethe proportions shown in Table 5.3.
In any case, these results are in direct contradiction with the idea thatchanges in root-system architecture and physiology lead to more efficient Nuptake by increased branching. One possible explanation may be that increasedbranching does not necessarily confer an increase in root-system uptake effi-ciency. For example, Fitter eta/. (1991) calculated the 'exploitation efficiency'(ratio of total root-system depletion zone volume to total root system volume)of idealized root systems. The architectural analysis included only fine-root sys-tems, and not large, woody structural roots. The most efficient root systemswere described as having long terminal root tips and internodes, with branchingonly along the primary axis. This provided for the least overlap of depletionzones, while ma_mi_ing total depletion zone volume. In the Fitter et al. (1991)model, higher-order branches tended to develop within the depletion zone of theparent, so that higher-order roots simply reduced overall exploitation efficiencyof the system.
168 M.E. Kubiskeand D.L. Godbold

Influence of CO 2 on Root Growth and Function 169
"o
o_ o_ _"t.-- v-- 0"3 _
_ CO _ 0 0-_ --_ _-_ _ _ 0 0
_= r= .__ o_ o_ _ _._
-_ =_ _
o oZ z
0 _. ,_ m0
s _. _ _" _ z _z
• . • • ,.. ooo _ ,.o _oo6 _6
_ o_ _ _ _ _ _6o _o oo _
_,--,_- _,-_ _ _o{,-: _,-" _,-" ,-"
• . • ° . ° ° • v-- •r,, u'_ ,_ _ _ _.. u_ _- • c_ C_l _-- i,_ u_ v--
....
170 M.E. Kubiskeand D.L. Godbold

Influenceof CO2on RootGrowthandFunction 171
Whether or not increased branching increases absorption efficiency of rootsystems, Table 5.3 suggests that root systems may function somewhat belowtheir capability while maintaining or even increasing the absorption capacityof individual roots. This essentially amounts to fail-safe production of fine rootsthat should confer greater resilience in tree root systems, making trees moreresistant to what might otherwise be catastrophic losses of roots or periodicshortages of soil resources. Continuing with the data of Pregitzer et al. (1995)
as an example, Populus trees under elevated CO2 could have maintained thesame N absorption efficiency as those under ambient O92 with only 21.4 g of fineroots in low N soil, and 22.9 g of fine roots in high N soil (compare with Table 5.3).
Another aspect that has received very little attention is the competitive abil-ity of root systems. Under low soil resource availability, the root systems of
Liquiclambar st!traciflua and Pinus taeda were more responsive to elevated CO2when grown as monocultures than when in combination (Jifon eta/., 1995).However, when soft resources were high, the reverse was true. It is difficult toplace this isolated study in context since virtually all other below-ground stud-ies involve single trees or monoeultures. None the less, it illus_ates that speciesinteractions have important ramifications for below-ground growth responses
of trees under elevated CO2. This is clearly an area that calls for more research.
5.4 Formation and function of mycorrhizas
Mycorrhizas are vital components of soil ecosystems, differing from non-symbiotic roots in both structure and function. Unfortunately, studies thatinvestigate root-system growth and function rarely mention the presence,
Table 5.4. Estimated losses of tree nitrogen associated with the senescence offine rootsofpopulustrees.
Total PlantN lostdueFine-root fine-root Fine-root to rootmortalityharvest Percentagebiornass N (%of totalbiomass mortality production (ml__-1) (m_) plantN)
LowN soil
ElevatedCO2 27.2 5.7 28.7 8.2 12.7 0.47AmbientCO2 20.6 3.4 21.3 8.9 6.2 0.24
HighN soilElevatedCO2 27.2 13.4 30.8 14.5 52.8 0.64AmbientCO2 16.0 7.1 17.1 16.0 18.2 0.31
Fine-rootproductionandmortalitydatafromminirhizotronsreportedinPregitzeretal. (1995)wereused.Oneassumptionwasthattheproportionoffine-rootmortalitytofinalstandingcropwasthesameforfine-rootlengthandbiomass.A secondassumptionwasthatnoN wasretranslocatedfromfinerootspriortosenescence.Moreaccurateestimateswouldhavetoaccountfordifferentordersof finerootsintermsofspecificrootlengthandN concentration.

172 M.E. Kubiskeand D.L. Godbold
abundance or influence of mycorrhizas. Conversely, studies that report on theabundance and roles ofmycorrhizas rarely discuss them in terms of free-rootproduction and turnover, and whole root-system function. There is a need forbetter experimental integration among these different aspects of tree root sys-tems so that their biologically integrated responses to global change conditionscan be assessed adequately. Until then, we will be forced to deal with mycor-rhizas as a separate issue from other root-system responses.
5.4.1 Carbon requirements
At both the community and individual levels, the benefits of mycorrhizas canbe traced back to the hyphal net of extramatrical mycelium in the soil. Theextramatrical mycelium provides both the connections between plants and a
large surface area for the uptake of water and nutrients. Differences in the abilityof ectomycorrhizas to improve nutrient and water relations of the host tree havebeen related to the extent and structure of the mycelium (Read, 1992).However, production of the extramatrical mycelinm may put a high C demandon the host plant, as the fungi require plant C for both structure and function.For example, ectomycorrhizal fungi exude large amounts of malate and oxalatethat liberate phosphoric acid from soft colloids for plant uptake (Lapeyrie et al.,
1987; Marschner, 1994). F_mgal Cdemand isparticularly obvious in experimentsinvolving high nutrient availability, where the advantage of mycorrhizal-medi-ated nutrient acquisition is not expressed. In such cases, the drain on the Cresourcx_ of the host plant may result in lower above-ground growth (Kfihr andArveby, 1986; Nyland and Wallander, 1989; Peng eta/., 1993). For example,Pinus sylvestris inoculated with a range of ectomycorrhizal fungi under lownutrient conditions had a negative correlation between plant growth and fun-gal biomass (Colpaert eta/., 1992). These costs must be balanced against thebenefits of a particular species. Costs and benefits are not always directly relatedto the mass of extramatrical mycelinm, especially in the case of mineral nutri-
ent uptake. None the less, the amount of extramatrical mycelium produced isan important factor (Marschner, 1994; Jentschke et al., 20OOa). Since theamount of extramatrical mycelium produced by an ectomycorrhizal fungus isspecies-specific, different species may have different costs to the tree.
The species composition of ectomycorrhizal fungi colonizing a root systemchanges over the life of a tree, and even within a particular growing season.Over a period of 3 years, Wu et al. (1993) observed changes in the type ofectomycorrhizas colonizing the roots of Pinus resinosa seedlings. By far the
largest changes in species composition occur during the lifespan of a tree andthese compositional changes are accompanied by increased diversity. Forexample, in stands ofPinus strobus, fruiting bodies of 5, 37 and 78 mycorrhizalspecies were found in 15-, 40- and 175-215-year-old stands, respectively

Influenceof CO2onRootGrowthandFunction 173
(Miller, 1983). This also illustrates that as trees or stands age, they are i_f_t_by a succession of different fungal species of mycorrhizas. The mechanismsfacilitating the succession are not certain, but there appear to be differences inthe Gcost and mode of infection of the different species. Early-stage fungi havea lower carbohydrate demand for mycelium growth than late-stage fungi(Deacon and Fleming, 1992). While there are many other differences between
early- and late-stage fungi in their ability to infect roots, such as the ability toinfect from spores, there is a strong case for the involvement of carbohydratesupply from the host tree in determining the mycorrhizal succession. It has beenreported that the degree of infection with late-stage fungi is correlated to rootsugar concentrations (Marx et td., 1977). For seedlings under normal condi-tions, the cost of late-stage fungi may be too high, even though these fungi mayconfer a number of benefits.
5.4.2 Mycorrhizal colonization
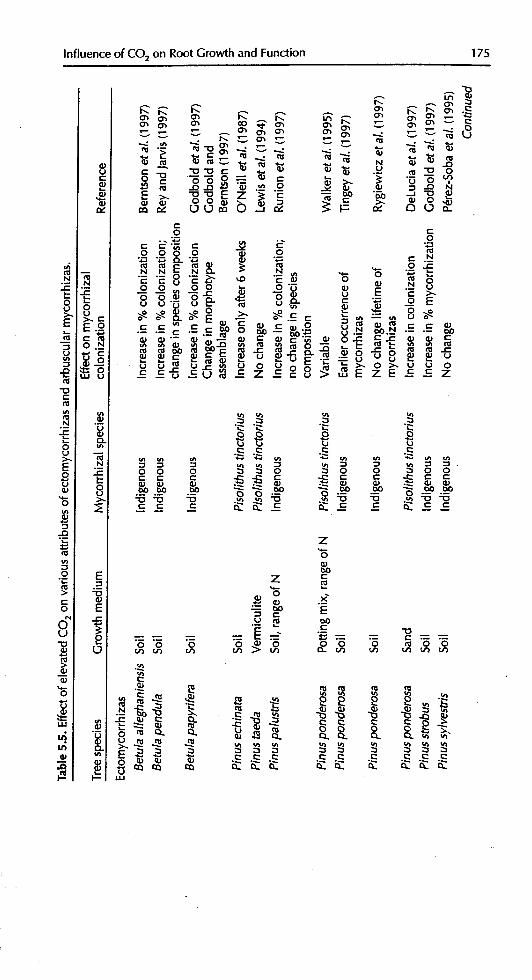
A number of studies have addressed the effects of elevated CO2 on the degree ofmycorrhization of tree roots (Table 5.5). These studies have used both indige-nous fungi using soil as an inoculum, and inoculation with a specific fungus, inmost cases Pisolithus tinctorius. In the majority of cases, elevated atmospheric
CO2 increased the degree of colonization by ectomycorrhizas on their host trees.This has been shown for indigenous ectomycorrhizas on Betula alleghaniensis(Berntson et el., 1997), B. papyrifera, P.strobus (Godbold et el., 1997), P.sylvestris(Rouhier and Read, 1998) and Pinuspalustris (Runion et al., 1997), and forPisolithus tinctorius on Pinus eclffnata (O'Neill eta/., 1987), P. ponderosa (Walkeret al., 1995; DeLucia et el., 1997), P. sylvestris (Ineichen et aI., 1995) and
Ouercus alba (O'Neill et el., 1987). However, no effect of elevated CO2 on col-onization was found for indigenous fungi on P. s#lvestris (P6rez-Soba et al.,1995; Markkola et al., 1996) and Tsuga canadensis (Godbold et el., 1997) orPisolithus tinctorius on P. taeda (Lewis et el., 1994) and P.ponderosa (Walker et el.,
1995). There is no apparent reason for these differences, either between speciesor the culture methods used. However, the few studies that have compared a
• number of species (O'Neill et eL, 1987; Godbold eta/., 1997) have reported agreater increase in colonization in deciduous angiosperms (O.a/be, B, papyrifera)compared to conifers (P.echinata, P.strobus, Z canadensis).
In O. a/be colonized with indigenous ectomycorrhizal fungi, the total pro-portion of mycorrhizal tips increased, but the percentage colonization by the
easily identifiable species Cenococcum grardforme decreased with elevated CO2relative to all mycorrhizal species (O'Neill et el., 1987). Cenococcum graniformemay confer increased drought resistance on its host (Read et al., 1977). Theectomycorrhizal species Paxillus involutus and Suillus bovinus both respondedwith greater fungal biomass when the host, P. sylvestris was _ to elevated
CO2 (Rouhicr and Read, 1998). Similarly, in B. papyrifera and P..strobus, the total

174 M.E. Kubiskeand D.L.Godbold
fraction of eetomycorrhizal root tips increased, and the assemblage of morpho-
types coloni_ing the roots changed (Godbold et aL, 1997). In B. papyrifera, themorphotype that showed the single largest increase had an extremely thickmantle and a large number of emanating hyphae and rhizomorphs. Theincrease in this morphotype was at the expense ofa morphotype with a thinnermantle, few emanating hyphae and no rhizomorphs (Godbold and Berntson,
1997). However, nnlike the study by O'Neill et aL (1987), there was no changein the frequency of a Cenococcuramorphotype. In P. strobus, no single morpho-type increased in frequency to the extent observed in B. pap!trifera; however, amorphotype was found on P. strobus under elevated COs that did not occurunder ambient 092. The changes in the ectomycorrhizal species assemblage inthe almost completely ectomycorrhizal B. papyrifera suggest that there may havebeen competition between morphotypes for available non-mycorrhizal root tips,and that those competitive interactions were modified by COs enrichment.Similarly, Rey and ]arvis (1997) found a change in ectomycorrhizal speciescomposition for/3, pendu/a. Under elevated COs, there was an increase in the fre-quency of Leccinum sp., which formed the domin ant fungal species, togetherwith P. involutus and Laccatia sp., whereas the most frequent species under ambi-
ent COs were Laccaria sp., Hebdoma sacchariolens and Thelephora terrestris. Theauthors explained this as accelerated tree ontogeny and a shift towards latersuccessional mycorrhizal species. Similar results and conclusions were reportedfor P.ponderosa (Tingey et aL, 1997).
Changes in the degree of ectomycorrhizal colonization may be driven bychanges in root carbohydrates for both ectomycorrhizal (Marx et al., 1977) andarbuscular mycorrhizal fungi (Douds and Schenk, 1990). Marx et aL (1977)reported that the degree of colonization with late-stage fungi was correlatedwith sugar concentrations in the roots. Late-stage fungi may have a higherdemand for carbohydrates than early-stage ectomycorrhizal fungi (Deacon andFleming, 1992). Independently, Godbold et a/. (1997) and Rey and Jarvis (1997)suggested that the increased total allocation of carbon below-ground by plants
in elevated 092 environments may be a controlling factor in the response of dif-ferent ectomycorrhizas to elevated COs, possibly favouring a shift in mycorrhizalassemblages toward late-stage fungi. However, species of fungi may also simplyvary in their ability to utilize higher carbohydrate availability for faster growth.It is not yet clear if this mechanism truly exists. Lewis et al. (1994) foundincreased soluble sugar concentrations in Pinus spp. roots under elevated CO2without a change in the rate of mycorrhizal colonization. Saikkonen et aL
(1999) showed a change in mycorrhizal morphotype composition after defoli-ation of 10-15-year-old P. sylvestr/s. There was a decrease in a tuberculate mor-
photype with a thick mantle and rhizomorphs, and a decrease in smoothmorphotypes.
In contrast to ectomycorrhizas, where increases in the degree of coloniza-
tion under elevated CO2 are common, responses of arbuscular mycorrhizalinfection to elevated CO2 are more variable. Using indigenous mycorrhizas on
Influenceof CO 2 on RootGrowth and Function 175
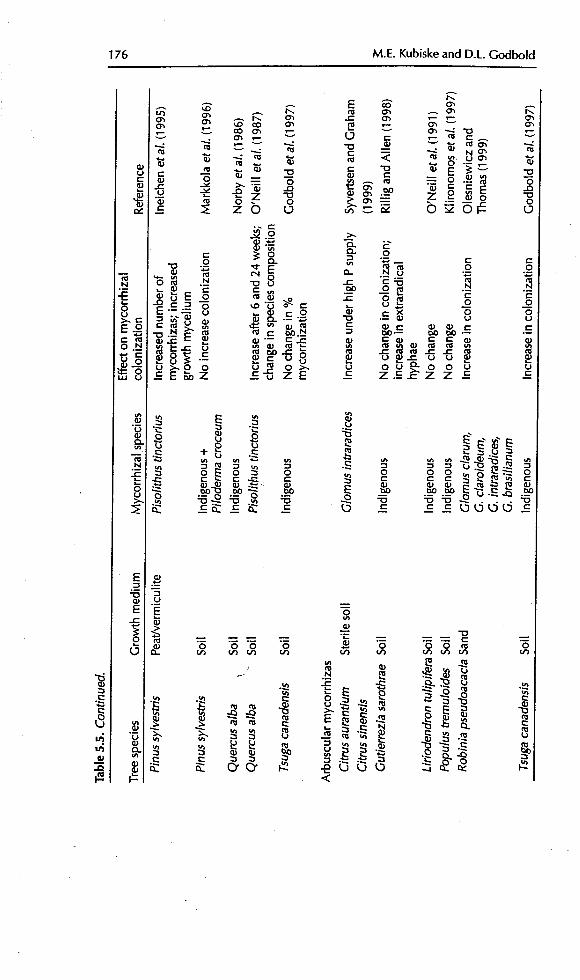
176 M.E. Kubiskeand D.L. Godbold

Influence of CO2on RootGrowth and Function 177
Liriodendron tulipifera under elevated CO2, no effect on the degree of infectionwas found (O'Neill et al., 1991). Similarly, in P. tremuloides (Klironomos et al.,1997) and Gutierrezia sarothrae (Rillig and Allen, 1998), no change was found.However, inoculation into sterilized soft or sand with a range of Glomus speciesresulted in increased rates of colonization (Olesniewicz and Thomas, 1999;8yvertsen and Graham, 1999). Godbold et al. (1997) also found an increaseddegree of colonization by indigenous fungi in non-sterile soft on T.canadensis.
In a number of cases for both ectomycorrhizas (Ineichen et al., 1995;Godbold et d., 1997) and arbuscular mycorrhizas (Rillig and Allen, 1998 ) the
density of extramatrical or extraradical hyphae under elevated CO2 hasincreased. This has also been shown in non-woody arbuscular mycorrhizalplants (Sanders eta/., 1998).
5.4.3 Mycorrhizal function
Although little is known about the nutrient acquisition by individual species ofectomycorrhizal fungi, it is becoming increasingly clear that there are differ-
ences in physiological performance (Bending and Read, 1995) and hyphal pro-duction (Rousseau et aI., 1994). Central to the benefits of mycorrhizas innutrient acquisition is the formation of an extensive extramatrical mycelium.Large differences have been shown in the utilization of both inorganic andorganic N forms. Finlay et al. (1988) showed differences in the assimilation of
lSNH 4 when fed to the extramatrical mycelittm of Rhizopogon roseolus, S. bovi-nus, P.involutus and P. tinctorius in symbiosis with P. sylvestris. In all fungi exceptP. involutus, a high proportion of assimilated ]SN was found inaspartate-asparagine. Bending and Read (1995) reported differences in Ndepletion from organic matter colonized by the extramatrical mycelium of S.bovinus and Thelephora terrestris. A higher depletion was found with S. bovinus.Additionally, mycorrhization alone has been shown, with a number of ectomy-corrhizas, to improve tree utilization of organic N sources (Abuzinadah andRead, 1986). Similar differences have also been shown between ectomycor-rhizal species in their ability to facilitate P uptake (Jentschke et a/., 2000a) andto improve heavy-metal tolerance (Godbold et a/., 1998). At present there are
no clear indications that elevated CO2 changes either P uptake (Walker eta/.,1995; Lewis and Strain, 1996; DeLucia eta/., 1997) or tolerance to toxic met-als, specifically aluminium (Al) (8chier and McOuattie, 1998). DeLucia et aL(1997) reported an increase in specific absorption rate and specific utiliTationrate of P in P.ponderosa mycorrhiml with P. tinctorius. They suggested that thecontribution of mycorrhizas to total P uptake may have been greater under ele-
vated (202, possibly related to an increase in oxalate exudation, which is believedto be involved in P release from soils (Strom, 1997).
Exudation of organic acids, particularly oxalate, is also believed to be an
important effector of Al tolerance (Delhaize eta/., 1993; Zheng eta/., 1998).

178 M.E. Kubiskeand D.L.Godbold
There have been few direct determinations of increased rhizodeposition under
elevated atmospheric CO2 (see above). Elevated atmospheric CO2 could increaseAl tolerance of plants directly by increasing the production of organic root exu-
dates, or indirectly through mycorrhizal effects. However, elevated CO2 did notincrease the Al tolerance ofPinus rigida mycorrhizal with P. tinctorius (Schierand McOuattie, 1998) or of non-mycorrhizal B. papyrifera (Li et aL, 1998) orPice.a rubens (Shipley eta/., 1992).
5.5 Conclusionsand researchneeds
Elevated CO2 results in greater production of photosynthate, which enhancesthe carbohydrate supply to root systems. Root-system growth increases,
although the allometric relationship between roots and shoots is generally notaffected except under the most nutrient-limiting conditions. The increase intotal root-system length is usually a function of increased branching, eitherfrom an increase in mycorrhizal tips, an increase in the production of lateral-root primordia at the apical meristem, or both. This larger population of fineroots is generally accompanied by increased fine-root turnover and total soft Cinput, although it is not clear ff this is a direct effect of C02 enrichment, or sire-
:-' ply a normal, allometric result of larger root systems. An increase in the num-ber of fine-root tips may dilute the total root-system sucrose supply, so thatindividual root elongation may not be stimulated by an increase in sucrose con-centration, although the possibility for increased growth rate of primary rootsis mechanistically feasible. Consmlction costs of producing new fine roots is notlikely to be altered, although a more complex root system with more growingroot tips is more expensive to maintain than a less complex system.
While a great deal of effort has been placed on understanding tree growth
under atmospheric CO2 enrichment, and there are many reports on the pro-duction of below-ground biomass with elevated CO2, reports detailing the phys-iological and morphological responses of root systems are relatively scarce. Evenless well understood is the functional balance between shoots and roots as envi-
ronmental conditions change, particularly with regard to variation between,and interactions among, tree species. K6rner and Arnone (1992) pointed outthe problems associated with attempting to scale up from physiological basefines without regard for mechanisms that operate at the ecosystem level. Forexample, studies on root-system architecture and physiology suggest that rootsmay become more effective in terms of soil exploration and nutrient uptake.However, thi.qis not consistent with tree N budgets, which show decreased total
plant N per unit fine-root biomass. It is nnknown ffincreased mycorrhizationmight lead to increased uptake efficiency as extramatrical mycelium extendswell beyond the reach of the root system proper. In any case, the increased pro-duction of both fine roots and mycorrhizas suggest an added buffer against rootlosses to herbivory or mechanical stresses.

Influenceof CO2on RootGrowth and Function 179
Better understanding of the allometric relationships within root systems isneeded in order to determine whether changes in fine-root production andturnover represent real changes in the growth and function of root systems. Therole of mycorrhizas clearly deserves more attention, particularly integrated withassessments of nutrient acquisition and soil Cinputs. Here the importance of under-
standing differences in the physiology of mycorrhizal species is vital. However, atpresent even basic culture methods to produce trees mycorrhizal with most late-stage fungi are missing. Ourknowledge of the physiology of ectomycorrhizal fungiis based almost entirely on early-stage fungi, which are of lesser ecological signifi-cance for mature forestecosystems. Finally, in our review of the litecature, we foundno data concerning the development and storage capacity of structural roots. Thisis probably because most studies to date have only dealt with seedling or sapling-sized trees that possess very few structural roots. With the development of FACEtechnology, perhaps more emphasis will be placed on understanding potentialchanges in root secondary growth and storage, which represent considerablebelow-ground C sinlc_,and their roles in the perennial growth of trees.
Acknowledgements
We thank those individuals who were willing to share details of their unpub-
lished data, answer our inquiries, and provide additional information as werequested it. We also thank Alex Friend and Mark Coleman, who reviewed ear-lier versions of the manuscript. Approved for publication as article No. F0142of the Forest and Wildlife Research Center, Mississippi State University.
References
Aber, J.D., MeliUo,].M.,Nadelhoffer,K.].,McClaugherty,C.A. and Pastor,J.(1985) Fineroot turnoverin forestecosystemsin relation toquantity and formofnitrogen avail-ability:a comparisonof two methods. Oecolo_a66, 317-321.
Abnzinadah, R.A.and Read,D.J.(1986) Theroleofproteinsin the nitrogen nutrition ofectomycorrhizal plants. L Utilizationof peptidesand proteins by ectomycorrhizalfun_. New Phytoloffist103, 481-493.
Allen, E.B., Allen, M.E, Helm, D.J.,Trappe, J.M.,Molina, R. and Rincon, E. (1995)Patterns andregulation ofmyeorrhizal plant and fungal diversity.Plantand Soft170,47-62.
Allen, M.F.(1991 ) TheEcoloffyofMycorrhizae.Cambridge University Press, Cambridge,pp. 9-22.
Allen, M.E, Moore, T.S.Jr and Chrlstensen, M. (1982) Phytohormone changes inBoutelouagrac///sinfected by vesicular-arbuscular myeorrhizae. IT.Altered levelsofgibbereUin-likesubstances and abscisicacid in the host plant. CanadianJournalofBotany 60, 468-471.

180 M.E. Kubiske and D.L. Godbold
: Aug6, R.M., Schekel, K.A. and Wample, R.L. (1986) Greater leaf conductance of well-: watered VA mycorrhizal rose plants is not related to phosphorus nutrition. New
Phytologist 103, 107-116.Barea, J.M. and Azcon-Agul]ar, C. (1982) Production of plant growth-regulating sub-
stances by the vesicular--arbuscular mycorrhizal fungus Glomus mosseae. AppliedEnvironmental Microbiology 43, 810-813.
Barlow, RW. (1987) Cellular packets, cell division and morphogenesis in the primary rootmeristem of Zea mays L. New Phytologist 105, 27-56.
Barlow, RW. and Adam, J.S. (1988) The position and growth of lateral roots on culturedroot axes of tomato, Lycopersicon esculentum (Solanaceae). Plant Systematics andEvolution 158, 141-154.
BassiriRad, H., Thomas, R.B., Reynolds, J.F. and Strain, B.R. (1996a) Differential
responses of root uptake kinetics of NH_ and NO_ to enriched atmospheric CO2con-centration in field-grown loblol]y pine. Plant, Cell and Environment 19, 367-371.
i_i BassiriRad, H., Griffin,K.L, Str-din, B.R. and Reynolds, J.E (1996b) Effects of CO2enrich-
ment on growth and root lSNH_ uptake rate of loblolly pine and ponderosa pineseedlings. Tre_Physiology 16,957-962.
BassiriRad, H., GritS, K.L., Reynolds, J.E and Strain, B.K. (1997a) Changes in root NH_
and NO_ absorption rates of loblolly pine and ponderosa pine in response to CO2enrichment. Plant and Soil 190, 1-9.
BassiriRad, H., Reynolds, J.E,V'w_i'nia,R.A. and Brunelle, M.H. (1997b) Growth and root
NO_ and PO_ uptake capacity of three desert species in response to atmosphericCO2 enrichment. Australian Journal of Plant Physiology 24, 353-358.
Bending, G.D. and Read, D.J. (1995) The structure and function of the vegetativemycelium of ectomycorrhizal plants. V. Foraging behaviour and translocation ofnutrients from exploited litter. New Phytologist 130, 401--409.
Berntson, G.M., and Bazzaz. EA. (1996) The allometry of root production and loss inseedlings of Acer rubrum (Aeeraceae) and B. papyrifem (Betulaceae): implications for
root dynamics in elevated CO2"American Journal of Botany 83, 608-616.Berntson, G.M. and Bazzaz, EA. (1997) Elevated CO2 and the magnitude and seasonal
dynamics of root production and loss in Betula papyrifera. Plant and Soil 190,211-216.
Berntson, G.M. Wayne, RM. and Bazzaz, EA. (1997) Below-ground architectural and
mycorrhizal responses to elevated CO2 in Betula alleghaniensis populations.Functional Ecology 11,684--695.
Bloom, A.J., Chapin, ES. and Mooney, H.A. (1985) Resource limitation in plants- aneconomic analogy. Annual Review of Ecology and Systematics 16, 363-392.
Bosac, C., Gardner, S.D.L., Taylor, G. and Wilkins, D. (1995) Elevated CO2 and hybridpoplar: a detailed investigation of root and shoot growth and physiology of Popu/useuramericana 'Primo'. Forest Ecology and Management 74, 103-116.
Canham, C.D., Kobe, R.K., Larry, E.E and Chazdon, R.L. (1999) Interspecific and
intraspodfic variation in tree seexlling survival: effects of allocation to roots versuscarbohydrate reserves. Oecologia 121, 1-11.
Cannel], M.G.R. and Dewar, R.C. (1994) Carbon allocation in trees: a review ofconeeptsfor modeling. Advances in Ecological Research 25, 59-104.
Charlton, W.A. (1991 ) Lateral root initiation. In: Waisel, Y., Eshel A. and Katkafi, U. (eds)Plant Roots, the Hidden Half. Marcel Dekker, New York, pp. 103-128.
Chevalier, C., LeOuerrec, E and Raymond, P.(1996) Sugar levels regulate the expression

Influence of CO 2 on Root Growth and Function 181
of ribosomal protein genes encoding protein $28 and ubiquitin-fnsed protein $27ain maize primary root tips. Plant Science 117, 95-105.
Colpaert, J.V., van Assche, J.A. and Luijtens, K. (1992) The growth of the extramatricalmycelinm of ectomycorrhiral fungi and the growth response of Pinus sylvestris L.NewPhyWlogist 120, 127-135.
Cosgrove, D.J. (1985) Cell wall yield properties of growing tissue. Plant Physiology 78,347-356.
Crookshanks, M., Taylor, G. and Broadmeadow, M. (1998) Elevated CO2 and tree rootgrowth: contrasting responses in Fraxlnus excelsior, Quercus petraea and Pinussylvestris. New Phytologist 138,241-250.
Curtis, P.S.and Wang, X. (1998) A recta-analysis of elevated t202 effects on woody plantmass, form, and physiology. Oecologia 113,299-313.
Cm:tis, ES., Zak, D.R., Pregitzer, K.S. and Teed, ]_.A.(1994) Above- and belowground
response of Populus grandidentata to elevated atmospheric CO2 and soil N availabil-i : :::ii:[_ i! [.i: _ :i:i ity. Plant and Soft 165, 45-51.
.... . :: • Deacon, J.W. and Fleming, L.V. (1992) Interactions ofectomycorrhizal fungi. In: Alien,M.]. (ed.) Mycorrhizal Functioning. Chapman & Hall, London, pp. 249-300.
Delhatze, E., Ryan, P.R. and Randall, EJ. (1993) Aluminium tolerance In wheat (_Y/ticumaest2v//m L) H.Alnminillm-sl_hnulated e_r_-'retionofinalh2 acid from root apices. PlantPhysiology 103,695-702.
DeLueia, ILH., Callaway, R.M., Thomas, g.M. and Schlesinger, W.H. (1997) Mechanisms
of phosphorns acquisition for Ponderosa pine seedlings under high CO2 and tem-perature. Annals of Botany 79, 111-120.
DeLucia, E.FL,Hamilton, J.G.,Naldu, S.L., Thomas, R_., Andrew_, J.A., Fin_, A., Lavine,M., Matamala, R., Mohan, J.E., Hendrey, G.R. and Schlesinger, W.H. (1999) Net pri-
mary production of a forest ecosystem with experimental CO2enrichment. Science284, 1177-1179.
Diaz, 8., Grime, J.P.,Harris, J.and McPherson, g. (1993) Evidence of a feedback mecha-nism limiting plant responses to elevated carbon dioxide. Nature 364, 616-617.
Dickson, R.E. (1989) Carbon and nitrogen allocation in trees. Annales des SciencesForestiires 46 (Suppl.), 631--647.
Dickson, R.tl. (1991) Assimilate distribution and storage. In: Raghavendra, A.S. (ed.)Physiology of Trees.John W'dey & Sons, New York, pp. 51-85.
Douds, D.D. and Schenk, N.C. (1990) Relationship of colonization and sporulation by VAmycorrhizal fungi to plant nutrient and carbohydrate contents. New Phytologist116, 621-627.
Eissenstat, D.M. (1992) Costs and benefits of constructing roots of small diameter, lournalof Plant Nutrition 15,763-782.
Entry, J.A., l_nnton, G.B., Prior, S_., Mitchell, R.J. and Rogers, H.H. (1998) Influence of
CO2enrichment and nitrogen fertilization on tissue chemistry and carbon alloca-tion in longleaf pine seedlings. Plant and SoI1200, 3-11.
Evans, J.R. (1989) Photosynthesis and nitrogen relationships in leaves of C3 plants.Oecologia 78, 9-19.
Evans, J., Francis, D. and Farrar, I.E (1994) Regulation of the cell cycle and the growthof maize roots in culture by exogenous sucrose. Journal of Experimental Botany 45(special issue), 36 [abstract].
Faber, B.A., Zasoski, R.J., Munns, D.N. and Shackel, K. (1991) A method for measuringhyphal nutrient and water uptake in mycorrhizal plants. Canadian Journal of Botany69, 87-94.

182 M.E. Kubiskeand D.L. Godbold
Fat-rat,J.F.and W'dllams, M.L (1991) The effectsof Increasedatmospheric carbon diox-ide and temperature on carbon partitioning, so--ink relations and recitation.Plant, Cell and Enviromn_mt14, 819-830.
Ferris, R. and Taylor, G. (1994) Increased root growth in elevated CO2: a biophysicalanalysis of root cell elongation. Journal of Experimental Botany 45, 1603-I 612.
Field, C. and Mooney, H.A. (1986) The photosynthesis-nitrogen relationship in wild
plants. In: Givnish I'./. (ed.) On the Economy of Plant Form and Function. CambridgeUniversity Press, Cambridge, pp. 25-55.
Finlay, R.D., Ek, H., Odham, G. and S6clerstr/_m, B. (1988) Mycelial uptake, transloca-
tion and assimilation of nitrogen from 15N labelled ammonium by Pinus sylvestrisplants infected with four different ectomycorrhizal fungi. New Phytologist 110,59-66.
Fitter, A.H. (1988) Water relations of red clover (Trifolium pratense L.) as affected by VA
: : .:iii i! _. .... i:i;i mycorrhizal infection and phosphorus supply before and during drought. Journal of. :: Experimental Botany 39, 595-603.
Fitter, A.H., StricHand, T.R., Harry, M.L. and Wilson, G.W.(1991) Architectural analy-sis of plant root systems 1. Architectural correlates of exploitation efficiency. NewPhytologist 118, 375-382.
Fitter, A.H., Graves, J.D., Watldn_, N.K., Robinson, D. and Scrimgeour, C. (1998) Carbontransfer between plants and its control in networks of arbuscular mycorrhizas.Functional Ecology 12, 406-412.
Fitter, A.H., Hodge, A., Daniell, T.J. and Robinson, D. (1999) Resource sharing inplant-fungus communities: did the carbon move for you? Trends in Ecology andEvolution 14, 70-71.
Francis, R., Finlay, R.D. and Read, D.J. ( 1986) Vesicular-arbuscular mycorrhiza in nat-ural vegetation systems. IV. Transfer of nutrients in inter- and intra-specific combi-nations of host plants. New Phytologist 102, 103-111.
Friend, A.L., Johnson, D.W., Yturiaga, J.V..and Modley, J. (1998 ) Impacts of elevated (302on N uptake of lobloUy and ponderosa pine. Soil Science Society of America AnnualMeeting, Madison, Wisconsin, p. 298.
Fry, S.C., Smith, R.C., Renwick, K.E, Martin, D.J. and Hodge, S.K. (1992 ) Xyloglucanendotransglycosylase, a new wall loosening enzyme activity from plants.Biochemistry Journal 282, 821-828.
Gebauer, R.LE., Reynolds, J.E and Strain, B.R. (1996) Allometric relations and growthin Pinus taeda: the effect of elevated CO2 and changing N availability. New Phytologist134, 85-93.
George, K, Marschner, H. and Jakobsen, I. (1995) Role of arbuscular mycorrhizal fungiin uptake of phosphorus and nitrogen from soft. Critiad Reviews in Biotechnology 15,257-270.
Godbold, D.L. and Berntson, G.M. (1997) Elevated atmospheric CO2 concentrationchanges eetomycorrhizal morphotype assemblages in Betula papyrifera. TreePhysiology 17, 347-350.
Godbold, D.L., Berntsan, G.M. and Baz_az, EA. (1997) Growth and mycorrhizal colo-
nization of three North American tree species under elevated atmospheric CO2.NewPhytologist 13 7, 433-440.
Godbold, D.L.,Jentschke, G., Winter, S. and Marschner, P. (1998) Ectomycorrhisas andamelioration of metal stress in forest trees. Chemosphere 36, 757-762.
Graham, J.H. and Syvertsen, J.E ( 1984) Influence of vesicular-arbuscular mycorrhiza

Influence of CO2 on Root Growth and Function 183
on the hydraulic conductivity of roots of two citrus rootstocks (Carrizo citrange, souroranges, inoculation with the fungus Glomus intraradices). New Phytologist 97,277-284.
Guo, Y., George, E.and Marschner, H. (1996) Contribution of an arbuscular mycorrhizal
fungus to the uptake of cadmium and nickel in bean and maize plants. Plant and Soil184, 195-205.
Hardie, K. ( 1985) The effect of removal of extraradical hyphae on water uptake by vesic-ular-arbuscular mycorrhizal plants. New Fhytologist 101, 677-684.
Hendrick, R.L and Pregitzer, K.S. (1993) The dynamics of fine root length, biomass, andnitrogen content in two northern hardwood ecosystems. Canadian Journal of ForestResearch 23, 2507-2520.
Hungate, B.A., Holland, E.A., Jackson, R.B., Chapin, ES., Mooney, H.A. and Field, C.B.(1997) The fate of carbon in grasslands under carbon dioxide enrichment. Nature388, 576-579.
Ineichen, K., Wiemken, V.and Wiemk_n, A. (1995) Shoots, roots and ectomycorrhizaformation of pine seedlings at elevated atmospheric carbon dioxide. Plant, Cell andEnvironment 18, 703-707.
Ineson, P., Cotrufo, M.E, Bol, R., Harkness, D.D. and Blum, H. (1996) Ouantillcation ofsoil carbon inputs under elevated CO2: C3 plants in a C4 soil. Plant and Soil 187,345-350.
Janos, D.E (1985) Mycorrhizal fungi: agents or symptoms of _vpical community com-position? In: Molina, R. (ed.) Proceedings of the 6th North American Conference onM!tcorrhizas. Forest Research Laboratory, Corvallis, Oregon, pp. 98-103.
lentschke, G., Brandes, B., Kuhn, A.J., SchrSder, W.H. and Godbold, D.L. (2000a)Phosphorus, nitrogen, potassium and magnesium translocation by the ectomycor-rhizal fungus Paxillus involutus connected to phosphorus-deficient Picea abiesseedlings. New Phytologist (in press).
lentschke, G., Brandes, B., Kuhn, AJ., SchrSder, W.H., Becker, J.S. and Godbold, D.L.(2000b) The mycorrhizal fungus Paxillus involutus transports magnesium toNorway spruce seedlings. Plant and Soil 220, 243-246.
Jffon, J.L., Friend, A.L. and Berrang, P.C. (1995) Species mixture and soil-resource avail-ability affect the root growth response of tree seedlings to elevated atmospheric CO2.Canadian Journal of Forest Research 25, 824--832.
K_ar, M. and Arveby, A.S. (1986) A method for establishing ectomycorrhiza on conifer
seedlings in steady-state conditions of nutrition. Physiologia Plantarum 67,333-339.
Kinsman, E.A., Lewis, C., Davies, M.S., Young, J.g., Francis, D., Vilhar, B. and Ougham,
H.]. (1997) Elevated CO2 stimulates cells to divide in grass meristems: a differentialeffect in two natural populations of Dactglis glomerata. Plant, Cell and Environment20, 1309-1316.
Kiepper, B. (1987) Origin, branching and distribution of root systems. In: Gregory, P.J.,Lake, ].V. and Rose, D.A. (eds) Root Development and Function. Society forExperimental Biology Seminar Series 30. Cambridge University Press, Cambridge,pp. 103-124.
Klironomos, J.N., Rillig, M.C., Allen, M.F., Zak, D.R., Kubiske, M.E. and Pregitser, K.S.
(1997) Soil fungal-arthropod responses to Populus tremuloides grown underenriched atmospheric CO2 under field conditions. Global Change Biology 3,473-478.

184 M.E. Kubiske and D.L. Godbold
Koide, R. (1985 ) The effect of VA mycorrhizal infection and phosphorus status on sun-flower hydraulic and stomatal properties. Journal of Experimental Botany 36,108 7-1098.
K6rner, Ch. and Arnone, J.A. (1992) Responses to elevated carbon dioxide in artificialtropical ecosystems. Science 257, 1672-1675.
Kubiske, M.E., Pregitzer, K.S., Zak, D.R. and Mikan, C.J.(1998) Growth and C allocation
of Popu/us tremuloides genotypes in response to atmospheric CO2 and soil N avail-ability. New Ph!!tologist 140, 251-260.
Kubiske, M.E., Zak, D.R., Pregi_er, K.S., Higgins, P., King, J.S. and Holmes, W. (2001)Growth and allometry ofPopulus tremuloides and Acer saccharum in response to ele-
vated CO2and soil N. Canadian Journal of Forest Research, in preparation.Lambers, H. (1987) Growth, respiration, exudation and symbiotic associations: the fate
of carbon translocated to the roots. In: Gregory, EJ., Lake J.V. and Rose, D.A. (eds)Root Development and Function. Society for Experimental Biology Seminar Series 30.Cambridge University Press, Cambridge, pp. 125-145.
Lamhamedi, M., Bernier, EY. and Fortin, J.A. (1992) Hydraulic conductance and softwater potential at the soil-root interface of Pmus pinaster seedlings inoculated withdifferent dikaryons of Pisoilthus sp. Tree Physiology 1O, 231-244.
Lapeyrie, F., Chilvers, G.A. and Bebm, C.A. (1987) Oxalic acid synthesis by the mycor-rhizal fungus Paxillus involutus (Batsch ex Ft.) Fr. New Phytologist 106, 139-146.
Larigauderie, A., Reynolds, J.E and Strain, B.IL (1994) Root response to CO2enrichmentand nitrogen supply in loblolly pine. Plant and Soil 165, 21-32.
Lewis, J.D.and Strain, B.R. ( 1996) The role of mycorrhizas in the response of P/nus taeda
seedlings to elevated CO2. New Phytologist 133, 431-443.Lewis, J_D.,Thomas, R.B. and Strain, B.R. (1994) Effect of elevated CO2 on mycorrhizal
colonization of loblolly pine (Pinus taeda L.) seedlings. Plant and Soil 165, 81-88.Li, A-, Berntson, G.M., Godbold, D.L. and Bazzaz, F.A. (1998) The dyvamics of root pro-
duction and loss in Betula papyrifera seedlings in response to elevated CO2 and anall]minium pulse. Zeitschrifl fdr Pfl_anzenerndhrang und Bodenkunde 161, 17-21.
Lockhart, J.A. (1965) An analysis of irreversible plant cell elongation. Journal ofTheoretical Biology 8,264--275.
Luo, Y., Field, C.B. and Mooney, H.A. (1994) Predicting responses of photosynthesis and
root fraction to elevated CO2:interactions among carbon, nitrogen, and growth.Plant, Ceil and Environment 17, 1195-1204.
Luxovfi, M. and Ciamporovfi, M. ( 1992 ) Root structure. In: Kolek, J.and Kozinka, V.(eds)Physiology of the Plant Root System. Kluwer Academic Publishers, Dordrecht, TheNetherlands, pp. 31-81.
Markkola, A.M., Ohtonen, A., Ahonen-Jonnarth, U. and Ohtonen, R. (1996) Scots pine
responses to CO2 enrichment - L Ectomycorrhizal fungi and soil fauna.Environmental Pollution 94, 309-316.
Marschner, P. (1994) Einflul_ der Mykorrhizierung auf die Aufnahme von Blei beiFichtenkeimlingen. PhD thesis, University of G6ttingen, G6ttingen, Germany.
Martens, R. (1990) Contn'bution of rhizodeposits to the maintenance and growth of soilmicrobial biomass. Soil Biology and Biochemistry 22,141-147.
Marx, D., Hatch, A.B. and Mendicino, J.E (1977) High fertility decreases sucrose contentand susceptibility of loblolly pine roots to ectomycorrhizal infection by Pisolithustinctorius. Canadian Journal of Botany 55, 1569-1574.
Merbach, W., lVlirus,F_,Knof_G.,Remus, R., Ruppel, S., Russow, IL,Gransee, A. and Schtdze, J.

Influence of CO2 on Root Growth and Function 185
(1999) Release ofcadxm and nitrogen compounds by plant roots and their possible eco-_ importance. ]ota'_ ofPlant Nutrition and soil Scienee 162,373-383.
Mikan, C.J., Zak, D.R., Kubiske, M.E. and Pregitzer, K.S. (2000) Combined effects of
atmospheric C02 and N availability on the belowground carbon and nitrogendynamics of aspen mesocosms. Oecologia 124, 432-445.
Miller, O.K. (1983 ) Ectomycorrhizae in the Agricales and Gasteromycetes. Canadian]ournd of Botany 61,909-916.
Mousseau, M. and Enoch, Z.FI.(1989) Carbon dioxide enrichment reduces shoot growthin sweet chestnut seedlings ( Castanea sativa). Plant, Cell and Environment 12,927-934.
Nadelhoffer, K.J. and Raich, J.W.(1992 ) Fine root production estimates and belowgroundcarbon allocation in forest ecosystems. Ecology 73, 1139-1147.
Nelsen, C.E. and Safir, G.R. (1982) Increased drought tolerance of mycorrhizal onionplants caused by improved phosphorus nutrition. Planta 154, 407-413.
Norby, R.J. (1994) Issues and perspectives for investigating root responses to elevatedatmospheric carbon dioxide. Plant and SOil 165, 9-20.
Norby, R.J., O'Neill, E.G. and Luxmoore, R.J. (1986) Effects of atmospheric CO2 enrich-ment on the growth and mineral nutrition of Quercus a/ba seedlings in nutrient-poorsoil. Plant Physiology 82, 83-89.
Norby, R.J., O'Neill, E.G., Hood, G.W. and Lnx'moore, R.J.(1987) Carbon allocation, rootexudation and mycorrhizal colonization ofPinus echinata seedlings grown under
CO2enrichment. Tree Physiology 3, 203-210.Norby, R.J., Wullschleger, S.D., Gunderson, C.A., Johnson, D.W. and Ceulemans, R.
( 1999 ) Tree responses to rising CO2 in field experiments: impfications for the futureforest. Plant, Celland Environment 22, 683-714.
Nyland, J.E.and Wallander, H. (1989) Effects of ectomycorrhiza on host growth and car-bon balance in a semi-hydroponic cultivation system. New Phytologist 112,
389-398.
Oaks, A. (1993 ) Primary nitrogen assimilation in higher plants and its regulation.Canadian Journal of Botany 72, 739-_750.
Olesniewicz, K.S. and Thomas, R.B. ( 1999) Effects of mycorrhizal colonisation on bio-mass production and nitrogen production of black locust (Robinia peeudoacaeia)seedlings under elevated atmospheric carbon dioxide. New Phytologist 142,133-140.
O'Neill, E.G., Lu_rnoore, R.J. and Norby, R.J. (1987) Increases in mycorrhizal colonisa-
tion and seedling growth in Pinus echinata and O.uercus aim in an enriched CO2atmosphere. Canadian Journal of Forest Research 17, 878-883.
O'Neill, E.G., O'Neill, R.V.and Norby, R.J. (1991 ) Hierarchy theory as a guide to mycor-rhizal research on large-scale problems. Environmental Pollution 73, 271-284.
Peng. S., Eissenstat, D.M., Graham, J.H., Williams, K. and Hodge, N.C. (1993) Growthdepression in mycorrhizal citrus at high-phosphorus supply. Analysis of carboncosts. Plant Physiology 101, 1063-1071.
P6rez-Soba, M., Dueck, T.A., Puppi, G. and Kuiper, EJ.C. (1995) Interactions of elevated
CO2, NH 3 and O3 on mycorrhizal infection, gas exchange and N metabolism insaplings of Scots pine. Plant and Soil 176, 107-116.
Perry, D.A. ( 1999 ) Reply from D.A. Perry. Trends in Ecology and B volution 14, 71 - 72.Perry, D.A., Margolis, H., Choquette, C., Molina, R. and Trappe, J.M. (1989)
Ectomycorrhizal mediation of competition between coniferous tree species. NewPhytologist 112, 501-511.

186 M.E. Kubiske and D.L. Godbold
Persson, H. (1979) Fine-root production, mortality and decomposition in forest ecosys-tems. Vogetatio 41,101-109.
Poorter, H. ( 1993) Interspecific variation in the growth response of plants to an elevated
ambient CO2concentration. Vegetatio 104/105, 77-97.Poorter, H., Gifford, R.M., Kriedeman, I.E. and Chin Wong, S. (1992) A quantitative
analysis of dark respiration and carbon content as factors in the growth response
of plants to elevated CO2. Australian]ournal of Botany 40, 501-513.Pregitzer, K.S., Zak, D.R., Curtis, ES., Kubiske, M.E., Teeri, J.A. and Vogel, C.S. (1995)
Atmospheric CO2, soil nitrogen and turnover of fine roots. New Phytologist 129,579-585.
Pregitzer, K.S., Kubiske, M.K, Yu, C.K. and Hendrick, R.L (1997) Relationships among rootbranch order, carbon, and nilroge_ in four temper_ specie_ Oecolog/a111,302-308.
Pregitger, K.S., Zak, D.R., Maziasz, J., DeForest, J.L., Curtis, E8. and Lussenhop, J. (2000)Interactive effects of atmospheric CO2 and sofl-N availability on fine roots of Populustremuloides. Ecological Applications 10, 18-33.
Prior, S.A., Runlon, G.B., Mitchell, R.J., Rogers, H.H. and Amthor, J.S. (1997) Effects of
atmospheric CO2 on longleaf pine: productivity and allocation as influenced bynitrogen and water. Tree Physiology 17, 397-405.
Read, D.J. (1992) The mycorrhizal myeelinm. In: Allen, M.J. (ed.) MycorrhizalFunctioning. Chapman & Hall, London, pp. 102-133.
Read, D.J., Kianmehr, H. and Malibari, A. (1977) The biology of mycorrhiza inHelianthemum Mill. New Phytologist 78, 305-312.
Read, D.J., Francis, R. and Finlay, R.D. ( 1985) Mycorrhizal mycefia and nutrient cycling
in plant communities. In: Fitter, A.H., Read, D.J. and Usher, M.B. (eds) EcologicalInteractions in SoiL Black-well, Oxford, pp. 193-217.
Rey, A. and Jarvis, P.G. (1997) Growth response of young birch trees (Betula pendula
Roth.) after four and a half years of CO2 exposure. Annals of Botang 80, 809-816.Rideout, J.W. and Raper, C.D. (1994) Diurnal changes in net uptake rate of nitrate are
associated with changes in e_imated export of carbohydrates to roots. International
Journal of Plant Science 155, 173-179.Rillig, M.C. and Allen, M.F. (1998) Arbuscular mycorrhizae ofGutierrezia sarothrae and
elevated carbon dioxide: evidence for shifts in C allocation to and within the myco-
biont. Soil Biology and Biochemistry 30, 2001-2008.Rogers, H.H., Runion, G.B. and Krupa, 8.V. (1994) Plant responses to atmospheric CO2
enrichment with emphasis on roots and the rhizosphere. Environmental Pollution83, 155-189.
Rothstein, D.E., Zak, D.R., Pregitzer, K.S. and Curtis, P.S. (2000) Kinetics of nitrogan
uptake by Populus tremuloides in relation to atmospheric CO2 and soil nitrogen. TreePhysiology 2O, 265-270.
Rouhler, H. and Read, D.J. (1998) Plant and fungal responses to elevated atmosphericcarbon dioxide in mycorrhizal seedlings of Pinus sylvestris. Environmental and
Experimental Botany 40, 237-246.Rouhier, H., Billes, G., ElKohen, A., Mousseau, M. and Bottner, P. (1994) Effect of ele-
vated CO2 on carbon and nitrogen distribution within a tree (Castanea sativaMill.)--soft system. Plant and $o//162, 281-292.
Rousseau, J.V., Sylvia, D.M. and Fox, A.J. (1994) Contribution of ectomycorrhiza to thepotential nutrient-absorbing surface of pine. New Phytologist 128, 639-644.
Ruiz-Lozano, I.M., Azcon, R. and Gomez, M. (1995) Effects of arbuscular--mycorrhizal

Influence of CO2 on Root Growth and Function 187
Glomus species on drought tolerance: physiological and nutritional plant responses.Applied Environmental Microbiology 61,456-460.
Runion, G.B., Mitchell, R.I., Rogers, H.H., Prior, S.A. and Counts, T.K. (1997) Effects of
nitrogen and water limitation and elevated atmospheric CO2on ectomycorrhiza oflongleaf pine. New Phytologist 137, 681-689.
Rygiewic_ ET., Johnson, M.G., Ganio, L.M., Tingey, D.T.and Storm, M.J. (1997) Lifetimeand temporal occurrence of ectomycorrhizae on ponderosa pine (Pinus ponderosa
Laws.) seedlings grown under varied atmospheric CO2 and nitrogen levels. Plant andSoil 189,275-287.
Saikkonen, K., Ahonen-Jounarth, U., Markkola A.M., Helander M., Tuomi, J.,Roitto, M.and Ranta, H. (1999) Defoliation and mycorrhizal symbiosis: a functional balancebetween carbon sources and below-ground s|nk._. Ecology Letters 2, 19-26.
Sanders, I.R., Streitwolf-Engel, R., van der Heijden, M.G.A., Boiler, T. and Wiemken, A.(1998) Increased allocation to external hyphae of arbuscular mycorrhizal fungi
under CO2enrichment. Oecologiii 117, 496--503.Schier, G.A. and McQuattie, C.J. (1998). Effects of carbon dioxide enrichment response
of mycorrhizal pitch pine (Pinus rigida) to aluminium: growth and mineral nutri-tion. Trees: Structure and Function 12,340-346.
Shipley, B., Lechowicz, M., Dumont, S. and Hendershot, W.H. (1992) Interacting effects
of nutrients, pH, ,4,1and elevated CO2on the growth of red spruce (Picea abies Sarg.)seedlings. Water, Air, and Soil Pollution 64, 585-600.
Simard, S.W. (1999 ) Below-ground connections among trees: implications for enhancedsilviculture and restoration. In: Egan, B. (ed.) Helping the Land Heak EcologicalRestoration in British Colunrbia.BC Environmental Network Educational Foundation,
Victoria, BC, Canada, pp. 179-184.Simard, S.W., Perry, D.A., Jones, M.D., Myrold, D.D., Dural, D.M. and Molina, R. (1997)
Net transfer of carbon between tree species with shared ectomycorrhizal fungi.Nature 388,579-582.
Soni, R., Carmichael, J.R, Shah, Z.H. and Murray, J.A.H. (1995) A family of cyclin Dhomologues from plants differently controlled by growth regulators and contain-ing the conserved retinoblastoma protein interaction motif. Plant Cell 7, 85-103.
Strom, L. ( 1997) Root exudation of organic acids: importance to nutrient availability andthe calcifuge and calcicole behavior of plants. Oikos 80, 459-466.
Stulen, I. and den Hertog, J. (1993 ) Root growth and functioning under atmospheric CO2enrichment. Vegetatio 104/105, 99-115.
Syvertsen, J.E and Graham, J.H. (1999) Phosphorus supply and arbnscular mycorrhizasincrease growth and net gas exchange responses of two Otrus spp. grown at ele-
vated COz. Plant and Soil 208,209-219.Taiz, L. (1984) Plant cell expansion: regulation of cell wall mechanical properties. Armual
Review ofPlant Physiology 35, 585-657.Taylor, G., Ranasinghe, S., Bosac, C., Gardner, S.D_L.and Ferris, R. (1994) Elevated CO2
and plant growth: cellular mecbani_ms and responses of whole plants. Journal ofExperimental Botany 45,1761-1774.
Taylor, G., Gardner, S_D.L.,Bosac, C., Flowers, T.J.,Crookshank._, M. and Dolan, L. (1995)
Effects of elevated CO2on cellular mechanisms, growth and development of treeswith particular reference to hybrid poplar. Forestry 68, 379-390.
Theodorou, C. and Bowen, G.D. (1993) Root morphology, growth and uptake of phos-phorus and nitrogen of Pinus radiata families in different soils. Forest EcologyManagement 56, 43-56.

188 M.E Kubiske and D.L. Godbold
Thomas, R.B., Richter, D.D., Ye, H., Heine, El{. and Strain, B.R. (1991) Nitrogen dynam-ics and growth of seedlings of a N-fixing tree (Gliricidia sepium (Jacq.) Walp.) exposedto elevated atmospheric carbon dioxide. Oecologia 88,415--421.
Tingey, D.T., Phillips, D.L., Johnson, M.G., Storm, M.J. and Ball, T.J. (1997) Effects of ele-
vated CO2 and N fertilization on fine root dynamics and fungal growth in seedlingPinus ponderosa. Enviromnental and Experimental Botany 37, 73-83.
Tissue, D.T., Thomas, R.B. and Strain, B.R. (1997) Atmospheric CO2 enrichmentincreases growth and photosynthesis ofPinus taex/a:a 4 year experiment in the field.Plant, Cell and Environment 20, 1123-1134.
Tschaplinski, T.J.,Stewart, D.B. and Norby, R.J. (1995) Interactions between drought and
elevated CO2 on osmotic adjustment and solute concentrations of tree seedlings.New Phytologist 131, 169-177.
Uselman, S.M., Quails, R.G. and Thomas, R.B. (1999) A test of a potential short cut inthe nitrogen cycle: the role of exudation of symbiotically fixed nitrogen from the
roots of a N-fixing tree and the effects of increased atmospheric CO2and tempera-ture. Plant and Sot/210, 21-32.
Van't Hof, 1. (1985) Control points within the cell cycle. In: Bryant, J.A. and Francis, D.
(eds) The Cell Division Cycle in Plants. Society for Experimental Biology SeminarSeries 26. Cambridge University Press, Cambridge, pp. 1-13.
Vivin, P., Martin, E and Guehl, J.-M. (1996) Acquisition and within-plant allocation of
13Cand lsN in CO2-enriched Quercus roburplants. Physiologia P/antarum 98, 89-96.Vogel, C.S. and Curtis, P.S. (1995) Leaf gas exchange and nitrogen dynamics of N2-fix-
ing, field-grown Alnus glutinosa under elevated atmospheric CO2. Global ChangeBiology 1, 55-61.
Vogel, C.S., Curtis, ES. and Thomas, R.B. (1997) Growth and nitrogen accretion of dini-trogen-fixing Alnus glutinosa (L.) Gaertn. under elevated carbon dioxide. PlantEcology 130, 63-70.
Vogt, K. (1991) Carbon budgets of temperate forest ecosystems. Tree Physiology 9,69-86.
Walker, R.E, Geisinger, D.R., Johnson, D.W. and Ball J.T. (1995) Interactive effects of
atmospheric CO2 enrichment and soft N on growth and ectomycorrhizal colonisa-tion of ponderosa pine seedlings. Forest Science 41,491-500.
Wittig, U. (1999 ) Zur saisonal Dynamik yon Ek-tomyeorrhizan der Buche (Fagus sglvaticaL.). PhD thesis, University of G6ttingen, Gfttingen, Germany.
Wu, Y, Gale, M.R., Cattelino, EJ., Richter, D.L. and Br_Thn: J.N. (1993) Temporal dynam-ics of ectomycorrhizal populations and seedling characteristics on red pine (Pinusresinosa). Cana_an Journal of Forest Research 23, 810-815.
Wullschleger, S.D., Norby, R.J. and Gunderson, C.A. (1997) Forest trees and their
response to atmospheric carbon dioxide enrichment: a compilation of results. In:Allen, LH., Kirkham, M.B., Olszyk, D.M. and Whitman, C.E. (eds) Advances in CarbonDioxide Effects Research. American Society of Agronomy Special Publication Number61, Madison, W_.onsin, pp. 79-99.
Yanai, R.D. (1994) A steady-state model of nutrient uptake improved to account fornewly-grown roots. Soil Science Society of America Journal 58, 1562-1571.
Zak, D.R., Pregitzer, K.S., Curtis, P.S., Teeri, J.A., Fogel, R. and Randlett, D.L. (1993)
Elevated atmospheric 092 and feedback between carbon and nitrogen cycles. PlantandSoi1151,105-117.

Influence of CO2 on Root Growth and Function 189
Zak, D.R., Pregitzer, K.S., Curtis, ES. and Holmes, W.E. (20OOa) Atmospheric CO2andthe composition and function of soft microbial communities. Ecological Applications10, 3_ _6.
Zak, DR., Pregitzer, K.S., Curtis, ES., Vogel, C.S., Holmes, W.E. and Lussenhop, J. (200Ob)
Atmospheric CO2, soil N availability, and the allocation of biomass and nitrogen,Ecological Applications 10, 47-50.
Zerihun, A., McKenzie, B.A. and Morton, J.D. (1997) Photosynthate costs associatedwith the utilization of different nitrogen-forms: influence on the carbon balanceofplants and shoot-root biomass partitioning. New PhytologiSt 138, 1-11.
ghang, H., Jennings, A., Barlow, P.W. and Forde, B.G. (1999) Dualpathways for regula-tion of root branching by nitrate. Proceedings of the National Academy of Sciences USA96, 6529-6534.
Zheng, S.I., Ma, J.E and Matsumoto, H. (1998) Continuous secretion of organic acids isrelated to aluminium resistance during relatively long term exposure to aluminiumstress. Physiologia Plantarum 103,209-214.
Appendix:literatureusedin Fig.5.1
Atkinson, C.J., Taylor, J.M., Willdn.%D. and Besford, R.T. (1997) Effects of elevated CO2on chloroplast components, gas exchange and growth of oak and cherry. TreePhysiology 17, 319-325.
BassiriRad, H., Griffin, K.L., Strain, B.R. and Reynolds, J.E (1996) Effects of CO2 enrich-ment on growth and root lSNH_ uptake rate of loblolly pine and ponderosa pineseedlings. TreePhysiology 16, 957-962.
Berntson, G.M., Wayne, P.M. and Bazzaz, EA. (1997) Below-ground architectural and
mycorrhizal responses to elevated CO2 in Betula alleghaniensis populations.Functional Ecology 11,684--695.
Bosac, C., Gardner, S.D.L., Taylor, G. and Wilidns, D. (1995) Elevated CO2 and hybridpoplar: a detailed investigation of root and shoot growth and physiology of Popu/useuramericana 'Primo'. Forest Ecology and Management 74, 103-116.
Crookshanks, M., Taylor, G. and Broadmeadow, M. (1998) Elevated CO2 and tree rootgrowth: contrasting responses in Fraxinus excelsior, Quercus petraea and Pinussylvestris. New Phytologist 138, 241-250.
DeLucia, EAt., Callaway, R.M., Thomas, E.M. and Schlesinger, W.H. ( 1997) Mecbani._ms
of phosphorus acquisition for Ponderosa pine seedlings under high CO2 and tem-perature. Annals of Botany 79, 111-120.
Entry, J.A., Runion, G.B., Prior, S.A., Mitchell, R.J. and Rogers, H.H. ( 1998) Influence of
CO2enrichment and nitrogen fertilization on tissue chemistry and carbon alloca-tion in longleaf pine seedlings. Plant and Soft 200, 3-11.
gpron, D., Liozon, R. and Mousseau, M. ( 1996 ) Effects of elevated CO2concentration onleaf characteristics and photosynthetic capacity of beech (Fagus sylvatica) during
• the growing season. Tree Physiology 16, 425-432.Gebauer, R.L.E., Reynolds, J.E and Strain, B.R. (1996) Allometric relations and growth
in P/nus taeda:the effect of elevated (202 and chan_ng N availability. New Phytologist134, 85-93.
Godbold, G.L., Rerntson, G.M. and Bazzaz, EA. (1997)Growth and mycorrhizal colo-

190 M.E. Kubiskeand D.L. Godbold
nizarion of three North American tree species under elevated atmospheric 092. NewPhytologist 137, 433-440.
Goodfellow, J.,gamus, D. and _ G. (1997) Diurnal and seasonal changes in the impactof CO2 enrichment on assimilation, stomatal conductance and growth in a long-term study of Mangifem lnd/ca in the wet-dry tropics of Australia. Tree Physiology17, 291-299.
Gorissen, A. (1996) Elevated CO2 evokes quantitative and qualitative changes in carbondynamics in a plant/soil system: mechanisms and implications. Plant and Soil 187,289-298.
Guehl, J.M., Picon, C., Anssenac, G. and Gross, P. (1994) Interactive effects of elevated
CO2and soil drought on growth and transpiration efficiency and its determinantsin two European forest tree species. Tree Physiology 14, 707-724.
Heath, J.and Kersriens, G. (1997) Effects of elevated CO2on leaf gas exchange in beechand oak at two levels of nutrient supply: consequences for sensitivity to drought inbeech. Plant, Cell and Environment 20, 57-67.
Hoddinott, J. and Scott, R. (1996) The influence of light quality and carbon dioxide
enrichment on the growth and physiology ofseedlings of three conifer species. [.Growth responses. CanadianJournal of Botany 74, 383-390.
Ineichen, K., Wiem_n, V.and Wiemken, A. (1995) Shoot, roots and ectomycorrhiza for-marion of Scots pine seedlings at elevated atmospheric carbon dioxide. Plant, Celland Environment 18, 703-707.
Ineson, P., Cotnffo, M_., 13ol,R., Harkness, D_D.and Blum, tL (1996) QuantiAcarion of soil
carbon inputs under elevated O32: C3plants in a C4 soil. P/ant and Soil 187, 345-350.K/Srner,Ch. and Arnone, J.A. (1992 ) Responses to elevated carbon dioxide in artificial
tropical ecosystems. Science 257, 1672-1675.
Larigauderie, A., Reynolds, J.E and Strain, B.R. (1994) Root response to COz enrichmentand nitrogen supply in loblolly pine. Plant and Soil 165, 21-32.
Markkola, A.M., Ohtonen, A., Ahonen-Jonnarth, U. and Ohtonen, R. (1996) Scots pine
responses to CO2 enrichment - I. Ectomycorrhizal fungi and soil fauna.EnvironmentaIPollutlon 94, 309-316.
Mikan, C.J., gak, D.R., Kubiske, M.E. and Pregitzer, K.S. (2000) Combined effects of
atmospheric CO2 and N availability on the belowground carbon and nitrogendynamics of aspen mesocosms. Oecologia (in press).
Polley, H.W.,Johnson, H.B., Mayeux, H.8., Tischler, C.R. and Brown, D.A. (1996) Carbondioxide enrichment improves growth, water relations and survival of droughtedhoney mesqnite (Prosop/s glandu/osa) seedlings. TreePhysiology 16, 817-823.
Pregitzer, K.S., Zak, D.R., Curtis, ES., Kubiske, M.E., Teeri, ].A. and Vogel, C.S. (1995)
Atmospheric CO2, soil nitrogen and turnover of fine roots. New Phytologist 129,579-585.
Progitzer, K.S., Zak, D.R., Maziasg, ]., DeForest, ].L., Curtis, ES. and Lussenhop, ]. (2000)
Interactive effects of atmospheric 002 and soil-N availability on fine roots of Popu/ustremuloides. Ecological Applications 1O, 18-33.
Rey, A. and ]arvis, P.G.(1997) Growth response ofyoung birch trees (Betulapendula
Roth.) after four and a half years of CO2 exposure. Annals of Botany 80, 809-816.Rouhier, H., Billes, G., gl Kohen, A., Mousseau, M. and Bottner, P. (1994) Effect of ele-
vated CO2 on carbon and nitrogen distribution within a tree (Castanea sativaMill. )-soil system. Plant and Soil 162, 281-292.
Tissue, D.T.,Thomas, R.B. and Stxain, B.R. (1996 ) Growth and photosynthesis of lobloUy

Influence of CO2 on Root Growth and Function 191
pine (Pinus taeda) after exposure to elevated CO2 for 19 months in the field. TreePhysiology 16, 49-59.
V'nrin, E, Martin, E and Guehl, J.-M. (1996) Acquisition and within-plant allocation of
13Cand 15Nin 032-enriched Ouemus robur plants. Physiologia Plantarura 98, 89-96.Vogel, C.S., Curtis, ES. and Thomas, R_B.(1997) Growth and nitrogen accretion of dini-
trogen-fixing Alnus glutinosa (L.) Gaertn. under elevated carbon dioxide. PlantEcology 130, 63-70.
Volin, J.C. and Reich, P.B.(1996) Interaction of elevated CO2 and 03 on growth, photo-synthesis and respiration of three perennial species grown in low and high nitro-gen. Physiologia P/antarum 97, 674-684.
Watt, A.D., Jones, C.G., Wynn, J. and Woodward, EI. (1999) The fraction of expanding
to expanded leaves determines the biomass response of Populus to elevated CO2.Oecologia 121,193-200.
Walker, R.E, Geisinger, D.R., Johnson, D.W. and Ball, ].T. (1998) Atmospheric CO2enrichment and soil N fertility effects on juvenile ponderosa pine: growth, ectomy-corrhizal development, and xylem water potential. Forest Ecology and Management102, 33-44.
Will, R.E. and Teskey, R.O. (1997) Effect of elevated carbon dioxide concentration and
root restriction on net photosynthesis, water relations and fofiar carbohydrate sta-tns of loblolly pine seedlings. Tree Physiology 17, 655--661.
Zak, D.R., Pregitzer, K.S., Curtis, ES., Vogel, C.S., Holmes, W_ and Lnssenhop, J. (2000)Atmospheric CO2, soil N availability, and the allocation of binmass and nitrogen.EcologicalAppiications 10, 47-50.