







Running Title: Rickettsial plasmids · 22 Minnesota, 1980 Folwell Ave., St. Paul, MN 55108. Phone:...
48
1 1 Title: Low Copy Number Plasmids are Widely Dispersed in Rickettsia Species Associated with 2 Blood-feeding Arthropods and may have Multiple Origins 3 4 Running Title: Rickettsial plasmids 5 6 Gerald D. Baldridge* 4 , Nicole Y. Burkhardt 4 , Marcelo B. Labruna 1 , Richard C. Pacheco 1 , 7 Christopher D. Paddock 2 , Philip. C. Williamson 3 , Peggy. M. Billingsley 3 , Roderick F. 8 Felsheim 4 , Timothy J. Kurtti 4 , and Ulrike G. Munderloh 4 9 Department of Entomology, University of Minnesota, St. Paul, Minnesota 55108 10 11 1 Departamento de Medicina Veterinaria Preventiva e Saude Animal, Faculdade de Medicina 12 Veterinaria e Zootecnia, Universidade de Sao Paulo, Sao Paulo, Brazil 13 14 2 Infectious Diseases Pathology Branch, Division of Viral and Rickettsial Disease, Centers for 15 Disease Control and Prevention, Atlanta, Georgia 16 17 3 Department of Forensic and Investigative Genetics, University of North Texas Health Science 18 Center, Fort Worth, Texas 19 20 * 4 Corresponding author. Mailing address: Department of Entomology, University of 21 Minnesota, 1980 Folwell Ave., St. Paul, MN 55108. Phone: (612) 624-3688. 22 E-mail: [email protected] 23 Copyright © 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. Appl. Environ. Microbiol. doi:10.1128/AEM.02988-09 AEM Accepts, published online ahead of print on 22 January 2010 on May 12, 2020 by guest http://aem.asm.org/ Downloaded from
Transcript of Running Title: Rickettsial plasmids · 22 Minnesota, 1980 Folwell Ave., St. Paul, MN 55108. Phone:...

1
1
Title: Low Copy Number Plasmids are Widely Dispersed in Rickettsia Species Associated with 2
Blood-feeding Arthropods and may have Multiple Origins 3
4
Running Title: Rickettsial plasmids 5
6
Gerald D. Baldridge*4, Nicole Y. Burkhardt
4, Marcelo B. Labruna
1, Richard C. Pacheco
1, 7
Christopher D. Paddock2, Philip. C. Williamson
3, Peggy. M. Billingsley
3 , Roderick F. 8
Felsheim4, Timothy J. Kurtti
4, and Ulrike G. Munderloh
4 9
Department of Entomology, University of Minnesota, St. Paul, Minnesota 55108 10
11
1Departamento de Medicina Veterinaria Preventiva e Saude Animal, Faculdade de Medicina 12
Veterinaria e Zootecnia, Universidade de Sao Paulo, Sao Paulo, Brazil 13
14
2Infectious Diseases Pathology Branch, Division of Viral and Rickettsial Disease, Centers for 15
Disease Control and Prevention, Atlanta, Georgia 16
17
3Department of Forensic and Investigative Genetics, University of North Texas Health Science 18
Center, Fort Worth, Texas 19
20
*4Corresponding author. Mailing address: Department of Entomology, University of 21
Minnesota, 1980 Folwell Ave., St. Paul, MN 55108. Phone: (612) 624-3688. 22
E-mail: [email protected] 23
Copyright © 2010, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.02988-09 AEM Accepts, published online ahead of print on 22 January 2010
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

2
ABSTRACT 24
25 Plasmids are mobile genetic elements of bacteria that can impart important adaptive traits such as 26
increased virulence or antibiotic resistance. We report existence of plasmids in Rickettsia 27
(Rickettsiales; Rickettsiaceae) species including R. akari, R. amblyommii, R. bellii, R. 28
rhipicephali and REIS, the rickettsial endosymbiont of Ixodes scapularis. All of the rickettsiae 29
were isolated from humans or North and South American ticks. R. parkeri isolates from both 30
continents did not possess plasmids. We have now demonstrated plasmids in nearly all Rickettsia 31
species that we have surveyed from three continents and that represent three of the four major 32
proposed phylogenetic groups associated with blood-feeding arthropods. Gel-based evidence 33
consistent with existence of multiple plasmids in some species was confirmed by cloning 34
plasmids with very different sequences from each of two R. amblyommii isolates. Phylogenetic 35
analysis of rickettsial ParA plasmid partitioning proteins indicated multiple parA gene origins 36
and plasmid incompatibility groups consistent with possible multiple plasmid origins. 37
Phylogenetic analysis of potentially host-adaptive rickettsial small heat shock proteins showed 38
that hsp2 genes were plasmid-specific and that hsp1 genes found only on plasmids of R. 39
amblyommii, R. felis, R. monacensis and R. peacockii were probably acquired independently of 40
the hsp2 genes. Plasmid copy numbers in seven Rickettsia species ranged from 2.4 to 9.2 per 41
chromosomal equivalent, as determined by real-time quantitative PCR. Plasmids may be of 42
significance in rickettsial evolution and epidemiology by conferring genetic plasticity and host 43
adaptive traits via horizontal gene transfer that counteracts the reductive genome evolution 44
typical of obligate intracellular bacteria. 45
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

3
INTRODUCTION 46
The α-proteobacteria of the genus Rickettsia (Rickettsiales: Rickettsiaceae) have 47
undergone the reductive genome evolution typical of obligate intracellular bacteria, resulting in 48
A/T-rich genomes (1.1 x 106 to 1.5 x 10
6 bp) with a high content of pseudogenes undergoing 49
elimination (3, 10, 20, 26). Initial sequencing of rickettsial genomes focused on the important 50
arthropod borne pathogens Rickettsia prowazekii, Rickettsia conorii, and Rickettsia typhi, and 51
appeared to confirm the prevailing belief that plasmids were absent and transposons were rare 52
among Rickettsia spp. (2, 28, 39, 44). As mobile genetic elements in bacteria, plasmids and 53
transposons drive horizontal gene transfer (HGT) and acquisition of virulence determinants and 54
environmental adaptive traits (30, 43, 60, 70). Subsequent sequencing of the Rickettsia felis 55
genome revealed the surprising presence of abundant transposase paralogs and the 63 kbp pRF 56
plasmid with 68 open reading frames (ORFs) encoding predicted proteins, as well as a 39 kbp 57
deletion form, pRFδ (45). Although pRF was suggested to be conjugative, it was initially thought 58
to be unique among the rickettsiae, a reasonable inference given that plasmids are uncommon 59
among the reduced genomes of obligate intracellular bacteria and were previously unknown in 60
the Rickettsiales (3, 4, 13). However, a phylogenetic analysis implied an origin for pRF in 61
ancestral rickettsiae and the possible existence of other rickettsial plasmids (28), which was soon 62
confirmed by the cloning of the 23.5 kbp pRM from Rickettsia monacensis (6). Some of the 23 63
ORFS on pRM had close pRF homologs and both plasmids carried transposon genes and the 64
molecular footprints of transposition events associated with HGT from other bacterial taxa. 65
The discoveries of pRF and pRM made obsolete the long-held dogma that plasmids were 66
not present in members of the genus Rickettsia and implied a source of unexpected genetic 67
diversity in the reduced rickettsial genomes, particularly if potentially conjugative plasmids 68
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

4
carrying transposon genes proved to be common among members of the genus. That hypothesis 69
gained credence when pulsed field gel electrophoresis (PFGE) and Southern blot surveys using 70
plasmid gene-specific probes demonstrated plasmids in Rickettsia helvetica, Rickettsia 71
hoogstraalii and Rickettsia massiliae and possible multiple plasmids in Rickettsia amblyommii 72
isolates (7). The same study demonstrated loss of a plasmid in the non-pathogenic Rickettsia 73
peacockii during long-term serial passage in cultured cells and absence of a plasmid in Rickettsia 74
montanensis M5/6, an isolate with a long laboratory passage history. Genome sequencing of R. 75
massiliae and Rickettsia africae revealed the 15.3 kbp pRMA and 12.4 kbp pRAF sequences 76
with 12 and 11 ORFS, respectively, that were more similar to those of pRF than pRM (11, 24). 77
The absence of plasmids in R. montanensis and important Rickettsia pathogens 78
maintained as laboratory isolates has left unresolved the question of the true extent of plasmid 79
distribution among Rickettsia spp. Until recently, the genus was thought to consist of closely 80
related species known chiefly as typhus and spotted fever pathogens transmitted by lice, fleas, 81
mites and ticks (31). It is now apparent that many, and possibly most, Rickettsia spp. inhabit a 82
diverse range of arthropods that do not feed on blood, as well as leeches, helminthes, crustaceans 83
and protozoans, suggesting an ancient and complex evolutionary history (54). A multi-gene 84
phylogenetic analysis of the Rickettsiales resulted in a “molecular clock”, which indicated the 85
order arose from a presumably free living ancestor and then adapted to intracellular growth 86
during the appearance of metazoan phyla in the Cambrian explosion (76). A transition to a 87
primary association with arthropods followed during the Ordovician and Silurian periods. The 88
genus Rickettsia arose approximately 150 million years ago and evolved into several clades, 89
including the early diverging hydra and torix lineages associated with leeches and protozoans. A 90
rapid radiation occurred about 50 million years ago in the arthropod associated lineages (76). 91
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

5
Whole genome sequencing has led to a revision of phylogenetic relationships among 92
Rickettsia spp. associated with blood feeding arthropods (10, 26, 28). A newly defined ancestral 93
group (AG) contains the earliest diverging species, Rickettsia bellii and Rickettsia canadensis, 94
while R. prowazekii and R. typhi transmitted by lice and fleas, respectively, constitute the typhus 95
group (TG). A proposed transitional group (TRG), consisting of the mite borne Rickettsia akari, 96
the flea borne R. felis, and the tick borne Rickettsia australis, bridges the genotypic and 97
phenotypic differences between the TG and the much larger spotted fever group (SFG) 98
consisting of tick borne rickettsiae (28). However, some presumptive SFG rickettsiae remain 99
poorly characterized and are of uncertain phylogenetic status, while accumulation of genomic 100
data from rickettsiae found in the diverse range of invertebrate hosts may have profound impacts 101
on the currently understood phylogeny of rickettsiae associated with blood feeding arthropods. 102
For example, it appears that the above AG and TRG species have many close relatives in insects 103
(76). Despite the recent phylogenomic advances, the genetic and host adaptive mechanisms 104
underlying the evolution of arthropod transmitted pathogens of vertebrates from ancestral 105
Rickettsia spp., including any possible role of plasmids, remain poorly understood. 106
In this report, we have taken advantage of recent isolations of rickettsiae from North and 107
South America to conclusively demonstrate that low copy number plasmids are indeed common 108
in low passage isolates of AG, TRG and SFG rickettsiae. The only exceptions were multiple 109
isolates from ticks and human eschar biopsies of R. parkeri, newly recognized as a mildly 110
pathogenic SFG rickettsia (49, 50, 52), and the previously characterized R. montanensis (7). We 111
confirmed that some Rickettsia isolates harbor more than one plasmid by cloning and sequencing 112
multiple plasmids from Rickettsia amblyommii isolates AaR/SC and Ac/Pa, and obtained PCR 113
and gel-based evidence that supported genome sequence evidence for existence of multiple 114
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

6
plasmids in REIS, the rickettsial endosymbiont of Ixodes scapularis. Phylogenetic analysis 115
provided strong evidence for multiple plasmid incompatibility groups and possible multiple 116
origins of plasmid-encoded parA genes in the genus Rickettsia. Other than genes encoding 117
plasmid replication initiation and partitioning proteins, the newly sequenced R. amblyommii 118
plasmids resembled the previously sequenced rickettsial plasmids in sharing limited similarities 119
in coding capacity (6, 7, 22). However, we have previously drawn attention to the presence of 120
hsp genes encoding α-crystalline small heat shock proteins as a conserved feature of most 121
rickettsial plasmids that may play a role in host adaptation (7). Phylogenetic analysis indicated 122
that the hsp2 genes were plasmid specific, while the hsp1 genes found on four rickettsial 123
plasmids may have been acquired by a chromosome-to-plasmid transfer event in a TRG-like 124
species. 125
126
MATERIALS AND METHODS 127
Rickettsiae. All rickettsiae were originally isolated in Vero (primate) cells unless stated 128
otherwise (Table 1). Whenever possible, low-passage isolates were used to eliminate the 129
possibility that isolates testing negative for presence of plasmids had done so due to their loss 130
during serial passage in the laboratory (7). Identity of each isolate was confirmed before analysis 131
for presence of plasmids by PCR amplification, DNA sequencing and comparison to Genbank 132
reference sequences of the gltA, ompA and 17-kilodalton antigen genes commonly used to 133
genotype Rickettsia spp. (1, 23). Rickettsiae were cultivated in Vero E6 or tick ISE6 cells as 134
described (39) with the exception of REIS, the rickettsial endosymbiont of Ixodes scapularis, 135
which was cultivated in IRE11 cells. Rickettsiae were released from host cells, separated from 136
cellular debris by filtration and concentrated by centrifugation (7). 137
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

7
Pulsed-field gel electrophoresis (PFGE). Purified rickettsiae were resuspended and 138
embedded in low-melting point agarose, digested with proteinase K in the presence of sodium 139
lauryl sarcosine and 0.5 M EDTA, and subjected to pulsed-field gel electrophoresis (PFGE) as 140
described (6) except that R. bellii isolates were run in 0.9% agarose gels to enhance separation of 141
larger plasmid isomers from chromosomal DNA. 142
Southern blot analyses. Rickettsial DNA was prepared and electrophoresed in PFGE 143
gels, depurinated and transferred onto Zeta Probe GT genomic membrane (Bio-Rad, Hercules, 144
CA) as described (5). The blots were hybridized with digoxigenin–labeled probes prepared by 145
PCR amplification of R. monacensis pRM plasmid genes, washed, and exposed to Kodak X-146
OMAT AR film (6, 7). 147
Determination of plasmid copy numbers. Plasmid copy numbers were determined as 148
ratios of the single copy plasmid hsp2 and chromosomal gltA genes using real-time quantitative 149
PCR (QPCR) and the relative quantification method (36, 37). To construct plasmids for 150
generation of species-specific standard curves, gltA sequences were PCR-amplified from each 151
Rickettsia sp. using 0.25 µM of the primer pair CS877F and CS1273R (54) and PfuTurbo 152
Hotstart DNA polymerase (Stratagene, La Jolla, CA). Cycle parameters with a Stratagene 153
robocycler were: 1 cycle at 95 0C for 2 min; 40 cycles at 95
0C for 30 s, 42
0C for 30 s, 1 min at 154
680C; and a final 10-min cycle at 68
0C. The PCR products were purified on spin-columns 155
(Qiagen, Valencia, CA), treated with Taq enzyme (Promega, Madison, WI) to create 3’-A 156
overhangs and cloned in the pCR4 vector (see below). The hsp2 sequences were amplified using 157
the Hsp2F3/Hsp2R3 primer pair (Table 2; all primers synthesized by Integrated DNA 158
Technologies, Coralville, Iowa) and GoTaq DNA Polymerase (Promega) with the following 159
cycle parameters: 1 cycle at 950C for 2 min; 40 cycles at 95
0C for 30 s, 46
0C for 30 s, 1 min at 160
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

8
720C; and a final 7-min period at 72
0C. Spin column-purified products were ligated into the 161
pCR4 vector with the TOPO TA Cloning kit and transformed into ONESHOT TOP10 competent 162
cells according to the manufacturer’s protocol (Invitrogen, Carslbad, CA). Plasmid DNA from 163
Kanamycin-resistant clones was prepared using the High Pure Plasmid Isolation Kit (Roche, 164
Indianapolis, IN) and sequenced to verify identity of the cloned PCR products (ABI 377 165
automated sequencer at Advanced Genetic Analysis Center, University of Minnesota). 166
For QPCR, serial dilutions of plasmid and rickettsial genomic DNAs were adjusted to 10 167
ng DNA per sample with salmon sperm DNA (Promega) and transferred to 96-well plates for 168
PCR amplification. The hsp2 sequences were amplified with species-specific primers and the 169
gltA sequences with the CS-F/CS-R primer pair (69) or species-specific primers (Table 2), using 170
the Brilliant or Brilliant II SYBR Green QPCR Master Mix, 240 nM primers, and an Mx3005p 171
qPCR cycler according to the manufacturer’s protocol (Stratagene). Cycle parameters were: 1 172
cycle at 950C for 10 min; 40 cycles at 95
0C for 30 s, 54
0C for 1 min, and 72
0C for 30 s. 173
Amplification specificity was confirmed by melting curve analysis. Data acquisition and analysis 174
were carried out with the MxPro software package, Version 4. Plasmid copy numbers per 175
chromosomal equivalent were determined by referencing values obtained from genomic samples 176
to the standard curves and were expressed as the mean ratios of hsp2/gltA amplification products 177
from three separate plates with all samples run in triplicate on each plate. 178
Cloning and sequencing R. amblyommii plasmids. Genomic DNA from R. amblyommii 179
isolates AaR/SC and Ac/Pa was partially digested with HpaI and SwaI (New England Biolabs, 180
Ipswich, MA), whose recognition sites occur at low frequency in rickettsial DNA. The digestion 181
products were ligated into the linear plasmid vector, pJAZZ (Lucigen Corp., Madison, WI) that 182
is optimized for cloning large A/T-rich inserts. Colony lifts of the libraries were hybridized with 183
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

9
probes derived from the R. monacensis pRM6, 8 and 16 plasmid genes (6). Positive clones that 184
contained inserts of approximately 6.5 – 25 kbp were Sanger sequenced by a primer walk 185
strategy using custom primers (Invitrogen, Carlsbad, CA) and BigDye* Terminator v.3.1 Cycle 186
Sequencing (Applied Biosystems, Inc. Foster City, CA using either an ABI PRISM* 310 or 187
3130xl Genetic Analyzer (Applied Biosystems). Sequence analysis, assembly and editing was 188
performed using Sequencher v4.7 (Gene Codes Corporation, Ann Arbor MI). Edited sequence 189
was prepared for submission to GenBank using Sequin Application Version 9.5 (NCBI) and 190
annotated using the NCBI Prokaryotic Genomes Automatic Annotation Pipeline (PGAAP). 191
Phylogenetic analyses of predicted ParA and Hsp proteins. Maximum Parsimony (67) 192
and Neighbor Joining (61) phylogenetic analyses were conducted using ParA, Hsp1 and Hsp2 193
proteins deduced from sequenced PCR products (see below) or plasmids and/or chromosomal 194
sequences of Rickettsia spp. and members of other bacterial taxa available at GenBank. 195
Sequences were manually aligned with Clustal X, edited to remove gaps, and imported into 196
PAUP* 4.0b (72) to construct phylogenetic trees. Node stability of dendrograms was estimated 197
with 1,000,000 random bootstrap replications (21). Additional Hsp2 sequences from R. bellii, 198
and R. hoogstraalii were derived from sequenced GoTaq PCR amplification products obtained 199
using the Hsp2F3/Hsp2R primer pair, and from R. helvetica using the Hsp2F3/Hsp2R3 and 200
Hsp2F/Hsp2R primer pairs (Table 2). Cycle parameters were: 1 cycle at 95 0C for 2 min; 40 201
cycles at 950C for 30 s, 46
0C for 30 s, 1 min at 72
0C; and a final 7-min step at 72
0C, except that 202
R. helvetica Hsp2F/Hsp2R reactions were annealed at 500C. The ParA sequences of REIS were 203
derived from sequenced Accu Taq (Sigma, St. Louis, MO) amplification products obtained using 204
pREIS1, 2, 3, and 4 primer pairs (Table 2). Cycle parameters were: 1 cycle at 94 0C for 3 min; 35 205
cycles at 940C for 30 s, 50
0C for 30 s, 1 min at 72
0C; and a final 5-min step at 72
0C. 206
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

10
Nucleotide sequence accession numbers. The R. amblyommii plasmid and PCR-207
amplified rickettsial hsp2 sequences reported here have been deposited in GenBank 208
(http://www.ncbi.nlm.nih.gov/Genbank) with the following accession numbers: pRAM18 209
(GU322808), pRAM23 (GU322807), R. bellii hsp2 (GU180086), R. helvetica hsp2 (GU180087) 210
and R. hoogstraalii hsp2 (GU180088). 211
212
RESULTS 213
Plasmids occur in R. rhipicephali, REIS and R. akari. In this study, we expanded the range of 214
arthropod-associated Rickettsia spp. analyzed for presence of plasmids using our previously 215
developed PFGE and Southern blot methods (7). In a PFGE lane loaded with an extract of the 216
South American R. rhipicephali HJ#5 isolate cultivated in tick ISE6 cells, plasmid isomers were 217
visible as SYBR Green-stained DNA bands that migrated at approximately 20, 45 and 55 kbp 218
relative to the linear DNA markers (Fig. 1A). The relative migration of the plasmid isomer and 219
DNA markers does not allow precise estimation of plasmid size because the plasmid isomers 220
represent a mixture of supercoiled and open circular forms as well as presumptive replication 221
single-stranded intermediates (6). Host cell mitochondrial DNA bands (7) co-migrated with 222
those in an uninfected ISE6 cell extract at approximately 25 and 35 kbp. In a lane loaded with 223
REIS cultivated in IRE11 cells, plasmid isomer bands migrated at approximately 40, 45 and 55 224
kbp, but only the smaller mitochondrial band was visible. The gel was Southern blotted and 225
hybridized (Fig. 1B) with a probe derived from the R. monacensis pRM16 gene, encoding a 226
DnaA-like replication initiator protein. In the lane with R. monacensis IrR/Munich, the 227
characteristic hybridization pattern of linear, open circular and supercoiled pRM isomers was 228
present with the major bands at approximately 25 and 45 kbp (6, 7). In the R. rhipicephali and 229
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

11
REIS lanes, four- and three-band hybridization patterns, respectively, were present at positions 230
of the plasmid bands in Fig. 1A, consistent with their identity as plasmid isomers. A replicate gel 231
was hybridized (Fig. 1C) with a probe derived from the pRM6 gene, encoding hsp2, which 232
produced the expected three-band hybridization pattern in the R. monacensis lane. In the R. 233
rhipicephali lane, the pRM6 probe hybridized in a two-band versus the four-band pattern of the 234
pRM16 probe, suggesting plasmid isomers with different gene complements. In the REIS lane, 235
the pRM6 probe replicated the hybridization pattern of the pRM16 probe but a fourth higher 236
molecular weight band was also present. A second replicate gel was hybridized (Fig. 1D) with a 237
probe derived from the pRM8 gene, encoding a helicase RecD/TraA protein. It hybridized as 238
expected in the R. monacensis lane but to only two plasmid isomer bands in the REIS lane and 239
did not produce a signal in the R. rhipicephali lane. Analysis of R. akari Bronx revealed a 240
plasmid that migrated as a predominant isomer at approximately 47 kbp (Fig. 2A). It hybridized 241
to the pRM16 probe (Fig. 2B) but not to the pRM6 and pRM8 probes (Figs. 2C and 2D). 242
In summary, the results demonstrated that the mite-associated TRG member, R. akari, 243
and three SFG Rickettsia spp. isolated from ticks collected on three continents have plasmids. R. 244
rhipicephali and REIS appear to have more than one plasmid. Four putative plasmid sequences 245
from REIS have been obtained as a consequence of the I. scapularis genome sequencing project 246
and deposited in GenBank as the circular pREIS1 and pREIS2, the incompletely circularized 247
pREIS3, and the plasmid scaffold NZ_GG688316 (referred to here as pREIS4). We confirmed 248
the REIS identity of the parA genes on those plasmids by PCR amplification of DNA extracts 249
from our REIS isolate using primers flanking the parA genes on the GenBank-deposited pREIS 250
sequences. In all four cases, PCR products of the predicted lengths (815 to 1075 bp) were 251
obtained and their sequences were identical to those of the cognate pREIS1, 2, 3, and 4 252
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

12
sequences (data not shown), consistent with a REIS origin of the plasmids rather than from other 253
bacteria that may have been present in tissues used to prepare I. scapularis DNA. 254
Plasmids occur in North and South American R. amblyommii but not in R. parkeri. 255
Analysis of the North American SFG member, R. amblyommii Darkwater, revealed plasmid 256
isomers that migrated at approximately 47 and 62 kbp (Fig. 2A). In contrast, plasmids were 257
absent in lanes containing R. parkeri Ft. Story, Portsmouth and Tates Hell (Fig. 2A) and in three 258
additional North American isolates, High Bluff, Grand Bay and Oktibbeha (data not shown). The 259
pRM16 probe hybridized to the R. amblyommii plasmids but did not hybridize to R. parkeri 260
DNA (Fig. 2B). A replicate gel hybridized with the pRM6 probe (Fig. 2C) reproduced the R. 261
amblyommii two-band hybridization pattern of the pRM16 probe and identified a third band that 262
probably corresponded to a plasmid that was poorly resolved from the 25 kbp mitochondrial 263
DNA band in Fig. 1A. The pRM6 probe did not hybridize to DNA of R. parkeri. A second 264
replicate gel hybridized with the pRM8 probe (Fig. 2D) reproduced the R. amblyommii pRM6 265
hybridization pattern, but did not yield a signal in the R. parkeri lanes. 266
Analysis of the South American R. amblyommii An13 and Ac37 isolates revealed 267
plasmid isomers that migrated at approximately 17 and 50 kbp versus 17, 33, and 55 kbp, 268
respectively (Fig. 3A). The South American R. parkeri isolates At#24 and At#5 did not have 269
plasmid DNA bands. The pRM16 probe hybridized to plasmid isomers of R. amblyommii An13 270
in a three-band pattern that included an additional 33 kbp band not revealed by SYBR Green-271
staining, but recognized only the 17 and 33 kbp plasmid isomers of R. amblyommii Ac37 (Fig. 272
3B). The probe did not hybridize to R. parkeri DNA. Replicate gels hybridized with the pRM6 273
probe (Fig. 3C) and the pRM8 probe (Fig. 3D) produced similar hybridization patterns except 274
that the 17 kbp plasmid isomer of R. amblyommii An13 was not recognized. The results were 275
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

13
consistent with presence of plasmids carrying pRM6, 8 and 16 homologs in the R. amblyommii 276
isolates and their absence in the R. parkeri isolates. 277
In summary, the results demonstrated presence of plasmid in North and South American 278
R. amblyommii isolates that carried pRM6, 8 and 16 homologs. In contrast, eight R. parkeri 279
isolates from both continents did not carry plasmids and none of the probes hybridized to their 280
sheared chromosomal or low molecular weight DNA, consistent with absence of plasmid gene 281
homologs on the rickettsial chromosomes. The R. parkeri isolates were the only low passage 282
rickettsia isolates that we have analyzed that did not possess a plasmid. Previously analyzed high 283
passage R. montanensis also lacked a plasmid, while low passage R. peacockii possessed a 284
plasmid that was lost during serial passage (7). 285
Plasmids occur in low passage R. bellii, but not in a high passage isolate. Analysis of 286
South American isolates of R. bellii, a member of the AG rickettsiae, revealed prominent 287
plasmids that migrated at approximately 55 – 65 kbp in lanes containing isolates ovale, Ad25, 288
An4 and Mogi, and at 75 and 90 kbp in the isolate HJ#7 lane (Fig. 4A). The upper mitochondrial 289
DNA band migrated as a doublet in the 0.9% agarose gels. Plasmid bands were absent in the lane 290
loaded with R. bellii 369-C, which was originally isolated in 1966 from Dermacentor variabilis 291
ticks collected in Arkansas, USA. Replicate gels hybridized with the pRM16 probe (Fig. 4B) 292
and the pRM6 probe (Fig. 4C) revealed plasmids in two- or three-band patterns seen in Fig. 4A 293
and less abundant smaller isomers. However, none of the patterns were identical and the plasmid 294
of the ovale isolate did not hybridize with the pRM6 probe. The pRM8 probe hybridized weakly, 295
or not all (isolate An4), to the plasmid bands but strongly to sheared chromosomal DNA, 296
including that of isolate 369-C (Fig. 4D). Because R. bellii occupies a basal position in rickettsial 297
phylogenetic trees, we hybridized a replicate gel blot with a probe derived from the pRM12 298
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

14
gene, encoding a proline/betaine transporter. It was previously shown to recognize the plasmid of 299
R. hoogstraalii isolated from a North American Argasid tick, but not the plasmids of Rickettsia 300
spp. from other Ixodid ticks or R. felis from cat fleas (7). The pRM12 probe hybridized to 301
plasmids of three of the five R. bellii isolates and to chromosomal DNA of all five (Fig. 4E). In 302
summary, the results demonstrated absence of a plasmid in the high passage 369-C isolate in 303
contrast to the universal presence of relatively large plasmids in the low passage R. bellii 304
isolates. The plasmids varied in size and those of the An4 and ovale isolates were less conserved 305
than the others relative to pRM of R. monacensis. 306
To further assess conservation of rickettsial plasmid gene complement, we hybridized 307
PFGE Southern blots of rickettsiae reported in this study with a probe derived from the R. 308
monacensis pRM23 gene encoding a transposon resolvase similar to that of Burkholderia 309
thailandensis and probably derived by HGT (6). The probe hybridized to plasmid isomers of R. 310
akari Bronx, R. rhipicephali HJ#5 and REIS (Fig. 5B) but not to plasmids of R. amblyommii or 311
R. bellii (data not shown). Results of Southern blot analyses shown in Figs. 1 – 5 are summarized 312
in Table 3. 313
Plasmid Copy Numbers. Rickettsial plasmid copy numbers were estimated by QPCR as 314
the relative ratios of plasmid-encoded hsp2 gene homologs and the single-copy gltA genes found 315
on rickettsial chromosomes. Plasmid copy numbers among seven Rickettsia spp. fell within a 316
four-fold range from 2.4 in R. helvetica to 9.2 in R. peacockii (Table 4). Means and standard 317
deviations of standard curve DNA and rickettsial sample DNA hlp2 reaction efficiencies were 318
96.8 +/- 2.6 and 94.1 +/- 2.9, respectively, while the cognate values for gltA reactions were 99.5 319
+/- 1.5 and 95.9 +/- 2.9, respectively. In all cases, reaction products had well-defined single peak 320
melting curves. 321
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

15
R. amblyommii AaR/SC and Ac/Pa isolates each have multiple distinct plasmids. In 322
this (Fig. 1) and a previous study (7), PFGE, Southern blot, PCR and I. scapularis genome 323
sequence data suggested existence of two or more distinct plasmid species in R. amblyommii, R. 324
rhipicephali and REIS. We verified that hypothesis by isolation, cloning and sequencing 325
plasmids from genomic libraries of R. amblyommii AaR/SC and Ac/Pa. Screening of the libraries 326
by hybridization with a cocktail of probes derived from pRM genes yielded positive clones at a 327
frequency of less than 1%. Eight AaR/SC clones and seven Ac/Pa clones were selected for 328
further study. Southern blot analysis revealed plasmid DNA inserts of 6.5 to 24 kbp (data not 329
shown). Four R. amblyommii AaR/SC clones have been sequenced to confirm existence of at 330
least two plasmids in that isolate. The entire 18,497 bp pRAM18 was contained in a single Swa1 331
clone (verified by PCR amplification using end terminal primers), while the 22,781 bp pRAM23 332
sequence was obtained from a 21.9 kbp Hpa1 clone and a 0.9 kbp PCR product amplified with 333
primers complementary to end sequences of the Hpa1 clone. An as yet incompletely circularized 334
sequence represents a provisional third plasmid (pRAM30) estimated to be 30 kbp in length. The 335
completed pRAM18 and pRAM23 sequences encode homologs of the DnaA-like replication 336
initiator and ParA plasmid partitioning proteins encoded by the pRM16 and pRM18 genes. 337
Homologs of the pRM6 and pRM7 genes encoding Hsp1 and Hsp2 proteins were present on 338
pRAM23 but not on pRAM18, which was nearly identical to a slightly larger Ac/Pa plasmid 339
contained in a single clone. Incompletely sequenced Ac/Pa clones indicated presence of at least 340
one more plasmid in that isolate. Other notable aspects of the AaR/SC plasmids included the 341
presence on pRAM23 of a homolog of the R. bellii and Orientia tsustsugamushi phrB genes that 342
encode DNA UV-damage repair enzymes as well as two sca12 gene homologs encoding outer 343
membrane proteins with potential roles in host cell interactions. A predicted gene on pRAM18 344
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

16
encoded a chimeric protein containing both SpoT and leucine-rich repeat domains, thus 345
combining potential stringent response and protein interaction functions. 346
The pRAM18 gene complement, but not that of pRAM23, was highly similar to that of 347
pRMA from R. massiliae, and included a homolog of the pRMA-p05 site specific recombinase 348
gene. In a PFGE/Southern blot analysis, a probe derived from the pRAM18 p05 homolog 349
hybridized to a subset of the R. amblyommii AaR/SC plasmid isomers that were not recognized 350
by the pRM6 probe (data not shown), further confirming existence of multiple plasmid species in 351
a single rickettsia isolate. 352
Phylogenetic analysis of rickettsial ParA proteins. Maximum parsimony analysis of 154 353
amino acids (145 informative) of the rickettsial ParA proteins showed that those encoded on 354
rickettsial chromosomes clustered tightly within the tree (Fig. 6, at left center) as a group that 355
diverged from a node on a branch bearing a ParA from Ehrlichia chaffeensis, also a member of 356
the Rickettsiaceae. Of all the rickettsial plasmid-encoded type Ib ParA proteins, only that 357
encoded by pREIS4 from the rickettsial endosymbiont of I. scapularis was present on the same 358
branch with the chromosomal ParA proteins, diverging from the most basal node on the branch. 359
The rickettsial plasmid ParA proteins were highly diverse relative to the chromosomal ParA 360
proteins, as illustrated by comparison with the tree positions of plasmid-encoded ParA proteins 361
from other families of bacteria. The majority of the rickettsial plasmid ParA sequences clustered 362
in three groups with good bootstrap support. The first group (at right) was at a distal position on 363
a branch with a basal node that led off to pRAF-encoded ParA of R. africae and a medial node 364
that led off to plasmid-encoded ParA sequences from Yersinia and Salmonella spp. The distal 365
group consisted of highly similar ParA homologs encoded by pRAM23 of R. amblyommii and 366
pRMA of R. massiliae, as well as a less similar homolog encoded on a contig reported from an 367
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

17
incomplete genome sequence assembly of the aquatic eukaryote Trichoplax adhaerens (68), the 368
simplest known multi-cellular animal. We detected many homologs of Rickettsia genes on the 369
Trichoplax contigs and predict that previously observed bacterial endosymbionts in Trichoplax 370
(62) will prove to be rickettsiae. The second rickettsial plasmid ParA group (bottom right) 371
consisted of homologs encoded by pRPR of R. peacockii and pRF of R. felis that clustered at the 372
distal ends of a branch containing an adjacent node that led to a ParA encoded by a plasmid of 373
Pseudomonas syringae. A more basal node on the branch led to ParA proteins from from 374
pREIS1 and plasmids of Yersinia and Pseudomonas spp., while the most basal node led to the 375
ParA encoded by pRM of R. monacensis. The third group (upper left) consisted of ParA proteins 376
encoded by pRFδ of R. felis, pREIS2 and pREIS3. The ParA proteins encoded by pRAM18 and 377
the provisional pRAM30 diverged from nodes without bootstrap support on a branch (top right) 378
that also contained a ParA encoded by the Borrelia burgdorferi cp32 plasmid. Neighbor joining 379
analysis supported the maximum parsimony results (not shown). 380
Phylogenetic analysis of rickettsial small heat shock proteins. Maximum parsimony 381
analysis of the rickettsial plasmid-encoded Hsp2 proteins (113 amino acids; 100 informative) 382
showed that they clustered tightly as a single group (Fig. 6B, circled at bottom right). The most 383
basal group member was encoded by the R. felis chromosomal RF1004 gene. Sequence analysis 384
indicated it was a chimera that may have originally been a plasmid gene that was integrated into 385
the chromosome by a recombination event adjacent to RF1005, which suffered a 5’end-terminal 386
deletion of its ORF (data not shown). The chromosome-encoded Hsp proteins of three facultative 387
intracellular pathogens from other bacterial taxa and of Wolbachia pipientis (Rickettsiales) 388
occupied intermediate positions in the tree. The Rickettsia chromosome-encoded Hsp1 proteins 389
clustered tightly as a group that branched away from the Legionella pneumophila node. The “R. 390
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

18
rickettsii complex” in that group included the SFG members R. rickettsii, R. africae, R. conorii, 391
R. massiliae, R. peacockii and R. sibirica. The plasmid-encoded Hsp1 proteins of R. felis, R. 392
monacensis, R. peacockii and R. amblyommii pRAM23 (pRAM18 has no hsp genes) formed a 393
second group (circled at top) that clustered as a side branch to the chromosomal Hsp1 cluster. 394
Neighbor joining analysis produced the same tree topology (not shown). 395
396
DISCUSSION 397
With this and previous PFGE/Southern blot analyses (6, 7), we have demonstrated 398
presence of plasmids in low passage isolates of AG, TRG and SFG Rickettsia spp. obtained from 399
arthropods or clinical samples collected from humans in North and South America and Europe. 400
Similar isolates of R. africae from Africa have plasmids (22) and it now seems likely that 401
plasmids occur in rickettsiae associated with blood-feeding arthropods throughout the world. 402
However, plasmids were not present in R. parkeri, newly recognized as a mild SFG pathogen 403
(49, 50, 52), or in the GenBank-deposited genome sequences of major pathogens in the SFG (R. 404
conorii and R. rickettsii) and the TG (R. prowazekii and R. typhi). Plasmids were detected in five 405
low-passage isolates of R. bellii, but not in R. bellii 369-C (Fig. 4) or in R. montanensis M5/6 (7), 406
both having undergone serial passage since their isolations in the 1960s (9, 56 and E. J. Bell, 407
unpublished). In this context, it would be reasonable to re-evaluate the major rickettsial 408
pathogens (i.e. R. conorii, R. prowazekii, R. rickettsii and R. typhi) as low-passage isolates for 409
presence of plasmids using PFGE/Southern blot assays. 410
We obtained the first estimates of rickettsial plasmid copy numbers. Single copy hsp2 411
genes occur on the pRF, pRM, pRAM23 and pRPR sequenced plasmids. The unsequenced 412
plasmids of R. bellii, R. helvetica, and R. hoogstraali have hsp2 homologs (Fig. 4C and ref. 7) 413
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

19
but their existence as single copies was an assumption. Rickettsial chromosomes typically carry 414
single copy gltA genes but whether this is true for the unsequenced R. amblyommii, R. helvetica 415
and R. hoogstraalii awaits confirmation. With the exception of R. felis, hsp2 homologs are not 416
present in the known rickettsial chromosome sequences. Given those caveats, plasmid copy 417
numbers in seven Rickettsia spp. representing the AG, TRG and SFG averaged 4.4 per 418
chromosomal equivalent (Table 4). The rickettsiae thus have low copy number plasmids (i.e. less 419
than 10 per chromosome) consistent with their possession of par genes, which are essential for 420
the maintenance and stable inheritance of such plasmids (25, 27, 73). Plasmids are absent in 421
members of the other genera of the Rickettsiales and are rare in other obligate intracellular 422
bacteria associated with arthropods. Among such bacteria, the plasmids of the Buchnera 423
endosymbionts of aphids (Hemiptera: Aphididae) are the best known. Ratios of the leuABCD and 424
trpEG genes encoded on separate plasmids to single copy chromosomal genes among Buchnera 425
spp. associated with three aphid hosts ranged from 0.6 to 23.5 (35, 58, 75). However, copy 426
number interpretations of those values may be complicated by trpEG gene amplification and 427
fluctuations in Buchnera chromosome ploidy during host lifetimes (75). 428
Apparent homologs of the R. monacensis pRM16 and pRM6 genes encoding plasmid 429
maintenance and probable host-adaptive functions, respectively, were well conserved among the 430
plasmids of 21 Rickettsia isolates (Table 3 and ref. 7), but there were interesting divergences in 431
plasmid gene conservation versus host association and phylogeny. Among SFG members, the 432
plasmid gene complement of R. hoogstraalii, isolated from a North American Argasid tick, was 433
much better conserved relative to that of R. monacensis than was that of R. helvetica, both of 434
which were isolated from the same European Ixodid tick. Apparent homologs of the pRM23 435
gene encoding a transposon resolvase were present on plasmids of the mite borne R. akari of the 436
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

20
TRG as well as R. rhipicephali and REIS, SFG members that were isolated from South and 437
North American ticks of different genera (Fig. 5). In contrast, pRM23 homologs were not present 438
on plasmids of other SFG rickettsiae or those of R. felis (TRG) and R. bellii (AG). The plasmid 439
gene complements were neither wholly consistent with rickettsial phylogeny and host 440
associations nor with descent from a single ancestral plasmid, suggesting possible multiple 441
origins and/or the influence of HGT. 442
We obtained evidence for multiple origins of the plasmids through phylogenetic analysis. 443
Plasmids carry conserved partitioning genes (par) that are usually organized in an auto-regulated 444
operon and are required for plasmid segregation at cell division (25, 73). The encoded ParA 445
proteins are Walker-type ATPases whose ATP-bound forms interact with a nucleoprotein 446
complex consisting of ParB protein dimers bound to sequence repeats at the parS centromere to 447
mediate intracellular location, movement, segregation and incompatibility of plasmids (14, 25). 448
The ParB protein sequences are highly conserved but the more variable ParA sequences allow 449
phylogenetic analysis of plasmid lineages (27). Maximum parsimony analysis showed that most 450
rickettsial plasmid-encoded ParA proteins fell into three groups that clustered with ParA proteins 451
encoded on plasmids from other bacterial genera rather than with the highly conserved ParA 452
sequences encoded on rickettsial chromosomes (Fig. 6). In conjunction with the evidence for 453
multiple plasmids in single Rickettsia spp. discussed below, the results provided strong evidence 454
for the presence of plasmids from multiple incompatibility groups in the genus Rickettsia. 455
Similar phylogenetic analyses of plasmid maintenance proteins encoded by repABC operons of 456
the repABC plasmids have demonstrated presence of multiple plasmid incompatibility groups 457
within several genera of α-proteobacteria, including as many as nine in the Roseobacter clade 458
that can stably coexist in the same cell, while six occur in each of two Rhizobium spp. (17, 55). 459
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

21
Similar to the rickettsial ParA phylogeny, different repABC replicons within the same bacterial 460
strain tend to belong to different phylogenetic clades with lineages that are not congruent with 461
species trees, suggesting that incompatibility groups arise as a consequence of divergent 462
evolution that may be interrupted by HGT events between plasmid lineages (16). 463
Differential hybridization patterns of the pRM6 and pRM16 probes to plasmid isomers of 464
both R. rhipicephali and REIS (Fig. 1) and several R. amblyommii isolates (Fig. 2, and ref. 7) 465
were reminiscent of the simultaneous presence of the 63-kbp pRF and the 39-kbp pRFδ deletion 466
form in R. felis (45). Although the pREIS1, 2, 3 and 4 plasmid sequence scaffolds from the I. 467
scapularis genome sequence project have varying degrees of similarity, they possess different 468
parA genes and none clearly represents a deletion form of another. We obtained PCR evidence 469
for their legitimate identity as REIS plasmids and obtained physical confirmation that multiple 470
plasmids exist in single rickettsiae isolates by cloning and sequencing pRAM18 and pRAM23 471
from R. amblyommii AaR/SC. Their sequence similarities were confined to genes that encoded 472
DnaA-like replication initiators and Par proteins and they were therefore distinct plasmids rather 473
than a major plasmid accompanied by a deletion form. We are sequencing a third plasmid 474
(pRAM30) from the AaR/SC isolate and two from the R. amblyommii Ac/Pa isolate. Because 475
those isolates represent uncloned bacterial populations it is not yet clear whether individual cells 476
contain only single or multiple plasmid species. 477
The presence of multiple plasmid species in single R. amblyommii isolates is intriguing 478
given the biology of their primary hosts. Amblyomma ticks are widely distributed from tropical 479
to temperate climates and are known for their aggressive propensity to feed on a wide range of 480
hosts that are parasitized by other ticks and blood-feeding arthropods (18, 29). Those attributes 481
are well suited to facilitating HGT within the “intracellular arena” of bacterial genetic exchange 482
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

22
in potentially co-infected arthropods (12, 13). A surprisingly diverse range of bacteria occur in 483
ticks (40) and co-infections of single ticks with different obligate intracellular microbes have 484
been demonstrated in several genera, including Amblyomma (19), and as many as three 485
Rickettsia spp. have been found in single ticks (15). The wide distribution and host biology of 486
Amblyomma ticks, and the presence of multiple plasmids that may be of different lineages in 487
single R. amblyommii strains, is consistent with the possibility that Amblyomma ticks have been 488
an active locus of HGT into and within the genus Rickettsia. 489
The high content of transposon-related sequences characteristic of all sequenced 490
rickettsial plasmids suggests that they may be HGT “hotspots” within rickettsial genomes, 491
perhaps as a consequence of their exposed positions as cytoplasmic episomes relative to the 492
packaged chromosomes associated with the bacterial cell walls. As mobile genetic elements, 493
plasmids are crucial drivers of HGT that enhance bacterial diversity and often provide the host 494
bacterium with functions such as drug resistance or environmental adaptive capacity that play 495
roles in pathogenicity (30, 43, 60, 70, 74). Plasmids may play those roles in rickettsiae by acting 496
as a mechanism for gain of new genes in the otherwise reductive Rickettsia genomes. The 497
rickettsial α-crystalline hsp genes that are prime candidates for provision of host adaptive 498
functions (7) provide a likely example. Phylogenetic analysis indicated that the hsp2 genes of 499
the Rickettsia were plasmid-specific, with the exception of a chromosomal homolog in R. felis, 500
and were probably acquired independently of the hsp1 genes (Fig. 6B). The plasmid-encoded 501
Hsp1 cluster consisted of proteins from R. felis (TRG) and three SFG spp. (R. monacensis, R. 502
peacockii and R. amblyommii) but branched from a deep node within the chromosomal Hsp1 503
group that lies between R. bellii (AG) and the TRG and TG rickettsiae. That result was consistent 504
with a potential evolutionary origin of the plasmid hsp1 genes in a chromosome-to-plasmid 505
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

23
transfer event in a TRG-like Rickettsia species. This possibility is supported by phylogenetic 506
analyses that showed the R. monacensis OmpA and OmpB proteins have much greater similarity 507
to homologs from TRG rickettsiae than to those from SFG rickettsiae (38), consistent with a 508
much closer affinity between R. monacensis and the TRG members than has previously been 509
realized. Additional support derives from presence of a 12 kbp plasmid-like sequence in the R. 510
typhi chromosome (28, 39) and our observation that the R. felis RF1004 gene may have 511
undergone a plasmid to chromosome transfer. 512
We have now demonstrated that plasmids occur in nearly all arthropod-borne AG, TRG 513
and SFG rickettsiae that we have surveyed for their presence. The pRF plasmid of R. felis was 514
the first to be discovered in the genus Rickettsia and was suggested to be conjugative on the basis 515
of encoding conjugative transfer genes and the presence of pili on the surfaces of R. felis cells 516
(45). It is now known that conjugative genes are widespread in the genus and that they are 517
horizontally transmitted (77). The presence of wide-spread and potentially mobile plasmids in 518
Rickettsia spp. has evolutionary and epidemiologic implications. The true impact of those 519
implications requires further investigation of the full extent of the distribution of rickettsial 520
plasmids within the many members of the genus not found in arthropod vectors and whether the 521
plasmids are currently mobile. 522
523
ACKNOWLEDGEMENT 524
This research was supported by NIH grant RO1 AI49424 to U.G.M. 525
526
REFERENCES 527
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

24
1. Anderson, B. E. and T. Tzianabos. 1989. Comparative sequence analysis of a genus-528
common rickettsial antigen gene. J. Bacteriol. 171:5199-5201.2 529
2. Andersson, S. G., A. Zomorodipour, J. O. Andersson, T. Sicheritz-Ponten, and U. C. Alsmark. 530
R. M. Podowski, A. K. Naslund, A. Eriksson, H. H. Winkler and C. G. Kurland. 1998. The 531
genome sequence of Rickettsia prowazekii and the origin of mitochondria. Nature. 396:133-532
140. 533
3. Andersson, S. G. E. and C. G. Kurland. 1998. Reductive evolution of resident genomes. 534
Trends Microbiol. 6:263-268. 535
4. Andersson, J. O. and S. G. E. Andersson. 2001. Pseudogenes, junk DNA and the 536
dynamics of rickettsia genomes. Mol. Biol. Evol. 18:829-839. 537
5. Baldridge, G. D., N. Burkhardt, M. J. Herron, T. J. Kurtti, and U. G. Munderloh. 2005. 538
Analysis of fluorescent protein expression in transformants of Rickettsia monacensis, an obligate 539
intracellular tick symbiont. Appl. Environ. Microbiol. 71:2095-2105. 540
6. Baldridge, G. D., N. Y. Burkhardt, R. F. Felsheim, T. J. Kurtti, and U. G. Munderloh. 541
2007. Transposon insertion reveals pRM, a plasmid of Rickettsia monacensis Appl. Environ. 542
Microbiol. 73:4984-4995. 543
7. Baldridge, G. D., N. Y. Burkhardt, R. F. Felsheim, T. J. Kurtti, and U. G. Munderloh. 544
2008. Plasmids of the pRM/pRF family occur in diverse Rickettsia species. Appl. Environ. 545
Microbiol. 74:645-652. 546
8. Beati, L., O. Peter, W. Burgdorfer, A. Aeschlimann, and D. Raoult. 1993. Confirmation 547
that Rickettsia helvetica spp. nov. is a distinct species of the spotted fever group of rickettsiae. 548
Int. J. Syst. Bacteriol. 43:521-526. 549
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

25
9. Bell, E. J., G. M. Kohls, H. G. Stoenner, and D. B. Lackman. 1963. Nonpathogenic 550
rickettsias related to the spotted fever group isolated from ticks, Dermacentor variabilis and 551
Dermacentor andersoni from eastern Montana. J. Immunol. 90:770-781. 552
10. Blanc, G., H. Ogata, C. Robert, S. Audic, K. Suhre, G. Vestris, J. - M. Claverie, and D. 553
Raoult. 2007. Reductive genome evolution from the mother of Rickettsia. PLoS Genet. 3(1):e14. 554
11. Blanc, G., H. Ogata, C. Robert, S. Audic, J. - M. Claverie, and D. Raoult. 2007. Lateral 555
gene transfer between obligate intracellular bacteria: Evidence from the Rickettsia massiliae 556
genome. Genome Research. 17:1657-1664. 557
12. Bordenstein, S. R., and J. J. Wernegreen. 2004. Bacteriophage flux in endosymbionts 558
(Wolbachia): Infection frequency, lateral transfer, and recombination rates. Mol. Biol. Evol. 559
21:1981-1991. 560
13. Bordenstein, S. R., and W.S. Reznikoff. 2005. Mobile DNA in obligate intracellular 561
bacteria. Nature Rev. 3:688-699. 562
14. Bouet, J. -Y., K. Nordstrom, and D. Lane. 2007. Plasmid partition and incompatibility – 563
the focus shifts. Mol. Microbiol. 65:1405-1414. 564
15. Carmichael, J. R., and P. A. Fuerst. 2009. Molecular detection of Rickettsia bellii, 565
Rickettsia montanensis, and Rickettsia rickettsii in a Dermacentor variabilis tick from nature. 566
Vector Borne Zoonotic Dis. Epub ahead of print. 567
16. Castillo-Ramirez, S., J. F. Vazquez-Castellanos, V. Gonzalez, and M. A. Cevallos. 2009. 568
Horizontal gene transfer and diverse functional constrains within a common replication-569
partitioning system in Alphaproteobacteria: the repABC operon. BMC Genomics. 10:536. 570
17. Cevallos, M. A., R. Cervantes-Rivera, and R. M. Gutierrez-Rios. 2008. The repABC 571
plasmid family. Plasmid. 60:19-37. 572
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

26
18. Childs, J.E., and C.D. Paddock. 2003. The ascendancy of Amblyomma americanuum as a 573
vector of pathogens affecting humans in the United States. Annu. Rev. Entomol. 48:307-337. 574
19. Clay, K., O. Klyachko, N. Grindle, D. Civitello, D. Oleske, and C. Fuqua. 2008. 575
Microbial communities and interactions in the lone star tick, Amblyomma americanum. Mol. 576
Ecol. 17:4371-4381. 577
20. Darby, A. C., N. -H. Cho, H. -H. Fuxelius, J. Westberg, and S. G. E. Andersson. 2007. 578
Intracellular pathogens go extreme: genome evolution in the Rickettsiales. Trends Genet. 579
23:511-520. 580
21. Felsenstein, J. 1985. Confidence limits on phylogenies: an approach using the bootstrap. 581
Evolution. 39:783-791. 582
22. Felsheim, R. F., T. J. Kurtti, and U. G. Munderloh. 2009. Genome sequence of the 583
endosymbiont Rickettsia peacockii and comparison with virulent Rickettsia rickettsii: 584
Identification of virulence factors. PLoS One. 4(12): e8361. 585
23. Fournier, P. -E., J. S. Dumler, G. Greub, J. Zhang, Y. Wu, and D. Raoult. 2003. Gene 586
sequence-based criteria for identification of new Rickettsia isolates and description of Rickettsia 587
heilongjiangensis spp. nov. J. Clin. Microbiol. 41:5456-5465. 588
24. Fournier, P. -E., K. E. Karkouri, Q. Leroy, C. Robert, B. Giumelli, P. Renesto, C. 589
Socolovschi, P. Parola, S. Audic, and D. Raoult. 2009. Analysis of the Rickettsia africae 590
genome reveals that virulence acquisition in Rickettsia species may be explained by genome 591
reduction. BMC Genomics. 10:166. doi: 10.1186/1471-2164-10-166. 592
25. Funnell, B. 2005. Partition-mediated plasmid pairing. Plasmid 53:119-125. 593
26. Fuxelius, H. -H., A. Darby, C. -K. Min, N. -M. Cho, and S. G. E. Andersson. 2007. The 594
genomic and metabolic diversity of Rickettsia. Res. Microbiol. 158:745-753. 595
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

27
27. Gerdes, K., J. Moller-Jensen, and R. Bugge Jensen. 2000. Plasmid and chromosome 596
partitioning: surprises from phylogeny. Mol. Microbiol. 37:455-466. 597
28. Gillespie, J. J., M. S. Beier, M. S. Rahman, N. C. Ammerman, J. M. Shallom, A. 598
Purkayastha, B. S. Sobral, and A. F. Azad. 2007. Plasmids and rickettsial evolution: Insight 599
from Rickettsia felis. PLoS One. 3:1-17. 600
29. Goddard, J. and A. S. Varela-Stokes. 2009. Role of the lone star tick, Amblyomma 601
americanum (L.), in human and animal diseases. Veterin. Parasitol. 160:1-12. 602
30. Hochut, B., U. Dobrindt, and J. Hacker. 2006. The contribution of pathogenicity islands to 603
the evolution of bacterial pathogens, pp83-108. In H. S. Siefert and V. J. Dirita (ed.) Evolution of 604
Microbial Pathogens. ASM Press, Washington, D.C. 605
31. Jones, K.E., N.G. Patel, M.A. Levy, A. Storeygard, D. Balk, J.L. Gittleman, and P. 606
Daszak. 2008. Global trends in emerging infectious diseases. Nature. 451:990-993. 607
32. Labruna, M. B., T. Whitworth, D. H. Bouyer, J. McBride, L. Marcelo, A. Camargo, E. 608
P. Camargo, V. Popov, and D. H. Walker. 2004. Rickettsia bellii and Rickettsia amblyommii in 609
Amblyomma ticks from the state of Rondonia, Western Amazon, Brazil. J. Med. Entomol. 610
41:1073-1081. 611
33. Labruna, M. B., R. C. Pacheco, S. Nava, P. E. Brandao, L. J. Richtzenhain, and A. A. 612
Guglielmone. 2007. Infection by Rickettsia bellii and Candidatus “Rickettsia amblyommii” in 613
Amblyomma neumanni ticks from Argentina. Microbial Ecol. 54:126-133. 614
34. Labruna, M. B., R. C. Pacheco, L. J. Richtzenhain, and M. P. J. Szabo. 2007. Isolation of 615
Rickettsia rhipicephali and Rickettsia bellii from Haemaphysalis juxtakochi ticks in the state of 616
Sao Paulo, Brazil. Appl. Environ. Microbiol. 73:869-873. 617
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

28
35. Lai, C. – Y., P. Baumann, and N. Moran. 1996. The endosymbiont (Buchnera sp.) of the 618
aphid Diuraphis noxia contains plasmids consisting of trpEG and tandem repeats of trpEG 619
pseudogenes. Appl. Environ. Microbiol. 62:332-339. 620
36. Lee, C., J. Kim, S. G. Shin, and S. Hwang. 2006. Absolute and relative QPCR 621
quantification of plasmid copy number in Escherichia coli. J. Biotechnol. 123:273-280. 622
37. Livak, K. J., and T. D. Schmittgen. 2001. Analysis of relative gene expression data using 623
real-time quantitative PCR and the 2-∆∆C
T method. Methods. 25:402-408. 624
38. Mattila, J. T., N. Y. Burkhardt, H. J. Hutcheson, U. G. Munderloh and T. J. Kurtti. 625
2007. Isolation of cell lines and a rickettsial endosymbiont from the soft tick Carios capensis 626
(Neumann) (Acari: Argasidae: Ornithodorinae). J. Med. Ent. 44:1091-1101. 627
39. McLeod, M. P., X. Qin, S. E. Karpathy, J. Gioia, S.K. Highlander, G. E. Fox, T. Z. 628
McNeill, H. Jiang, D. Muzny, L. S. Jacob, A. C. Hawes, E. Sodergren, R. Gill, J. Hume, M. 629
Morgan, G. Fan, A. G. Amin, R. A. Gibbs, C. Hong, X. Yu, D. H. Walker, and G. M. 630
Weinstock. 2004. Complete genome sequence of Rickettsia typhi and comparison with 631
sequences of other rickettsiae. J. Bacteriol. 186:5842-5855. 632
40. Moreno, C.X., F. Moy, T.J. Daniels, H.P. Godfrey, and F.C. Cabello. 2006. Molecular 633
analysis of microbial communities identified in different developmental stages of Ixodes 634
scapularis ticks from Westchester and Dutchess counties, New York. Environ. Microbiol. 8:761-635
772. 636
41. Munderloh, U. G., S. D. Jauron, V. Fingerle, L. Leitritz, S. F. Hayes, J. M. Hautman, C. 637
M. Nelson, B. W. Huberty, T. J. Kurtti, G. G. Ahlstrand, B. Greig, M. A. Mellencamp, and 638
J. L. Goodman. 1999. Invasion and intracellular development of the human granulocytic 639
ehrlichiosis agent in tick cell culture. J. Clin. Microbiol. 37:2518-2524. 640
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

29
42. Noda, H., U. G. Munderloh, and T. J. Kurtti. 1997. Endosymbionts of ticks and their 641
relationship to Wolbachia and tick-borne pathogens of man and animals. Appl. Environ. 642
Microbiol. 63:3926-3932. 643
43. Ochmann, H., J. G. Lawrence, and E. A. Grolsman. 2000. Lateral gene transfer and the 644
nature of bacterial innovation. Nature. 405:299-304. 645
44. Ogata, H., S. Audic, P. Renesto-Audiffren, P. – E. Fournier, V. Barbe, D. Samson, V. 646
Roux, P. Cossart, J. Weissenbach, J. – M. Claverie, and D. Raoult. 2001. Mechanisms of 647
evolution in Rickettsia conorii and R. prowazekii. Science. 293:2093-2098. 648
45. Ogata, H., P. Renesto, S. Audic, C. Robert, G. Blanc, P. - E. Fournier, H. Parinello, J. - 649
M. Claverie, and D. Raoult. 2005. The genome sequence of Rickettsia felis identifies the first 650
putative conjugative plasmid in an obligate intracellular parasite. PLoS Biol. 3(8):e248. 651
46. Pacheco, R. C., J. M. Venzal. L. J. Richtzenhain, and M. B. Labruna. 2006. Rickettsia 652
parkeri in Uruguay. Emerg. Infect. Dis. 12:1804-1805. 653
47. Pacheco, R., S. Rosa, L. Richtzenhain, M. P. J. Szabó, and M. B. Labruna. 2008. 654
Isolation of Rickettsia bellii from Amblyomma ovale and Amblyomma incisum ticks from 655
southern Brazil. Rev. MVZ Córdoba 13:1273-1279. 656
48. Pacheco, R. C., M. C. Horta, A. Pinter, J. Moraes-Filho, T. F. Martins, M. S. Nardi, S. 657
S. Souza, C. E. Souza, M. P. Szabó, L. J. Richtzenhain, and M. B. Labruna. 2009. Pesquisa 658
de Rickettsia spp em carrapatos Amblyomma cajennense e Amblyomma dubitatum no Estado de 659
São Paulo. Rev. Soc. Bras. Med. Trop. 42:351-353. 660
49. Paddock, C. D. 2005. Rickettsia parkeri as a paradigm for multiple causes of tick-borne 661
spotted fever in the Western Hemisphere. Ann. N.Y. Acad. Sci. 1063:315-326. 662
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

30
50. Paddock, C. D., J. W. Sumner, J. A. Comer, S. R. Zaki, C. S. Goldsmith, J. G. Goddard, 663
S. L. F. McLellan, C. L. Tamminga, and C. A. Ohl. 2004. Rickettsia parkeri: A newly 664
recognized cause of spotted fever rickettsiosis in the United States. Clin. Infec. Dis. 38:805-811. 665
51. Paddock, P. D., T. Koss, M. E. Eremeeva, G. A. Dasch, S. R. Zaki, and J. W. Sumner. 666
2006. Isolation of Rickettsia akari from eschars of patients with rickettsialpox. Am. J. Trop. 667
Med. Hyg. 75:732-738. 668
52. Paddock, C. D., R. W. Finley, C. S. Wright, H. N. Robinson, B. J. Schrodt, C. C. Lane, 669
O. Ekenna, M. A. Blass, C. L. Tamminga, C. A. Ohl, S. L. F. McLellan, J. Goddard, R. C. 670
Holman, J. J. Openshaw, J. W. Sumner, S. R. Zaki, and M. Eremeeva. 2008. Rickettsia 671
parkeri rickettsioisis and its clinical distinction from Rocky Mountain spotted fever. Clin. Infect. 672
Dis. 47:1188-1196. 673
53. Paddock, C. D., P. – E. Fournier, J. W. Sumner, J. Goddard, Y. Elshenawy, M. G. 674
Metcalfe, and A. Varela-Stokes. In press. Isolation of Rickettsia parkeri and identification of a 675
novel spotted fever group Rickettsia sp. from Gulf Coast ticks (Amblyomma maculatum) in the 676
United States. Appl. Environ. Microbiol. 677
54. Perlman, S. J., M. S. Hunter and E. Zchori-Fein. 2006. The emerging diversity of 678
Rickettsia. Proc. R. Soc. B. 273:2097-2106. 679
55. Petersen, J., H. Brinkmann, and S. Pradella. 2009. Diversity and evolution of repABC 680
type plasmids in Rhodobacterales. Environ. Microbiol. 11:2627-2638. 681
56. Philip, R. N., E. A. Casper, R. L. Anacker, J. Cory, S. F. Hayes, W. Burgdorfer, and C. 682
E. Yunker. 1983. Rickettsia bellii sp. nov.: a tick-borne Rickettsia, widely distributed in the 683
United States, that is distinct from the spotted fever and typhus biogroups. Internat. J. Syst. 684
Bacteriol. 33:94-106. 685
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

31
57. Pinter, A., and M. B. Labruna. 2006. Isolation of Rickettsia rickettsii and Rickettsia bellii 686
in cell culture from the tick Amblyomma aureolatum in Brazil. Ann. N. Y. Acad. Sci. 1078:523-687
529. 688
58. Plague, G. R., C. Dale, and N. A. Moran. 2003. Low and homogenous copy number of 689
plasmid-borne symbiont genes affecting host nutrition in Buchnera aphidicola of the aphid 690
Uroleucon ambrosiae. 691
59. Pornwiroon, W., S. S. Pourciau, L. D. Foil, and K. R. Macaluso. 2006. Rickettsia felis 692
from colonized cat fleas: Isolation and culture in a tick-derived cell line. Appl. Environ. 693
Microbiol.72:5589-5595. 694
60. Rashkin, D. M., R. Seshadri, S. U. Pukatzki, and J. J. Mekalanos. 2006. Bacterial 695
genomics and pathogen evolution. Cell. 124:703-714. 696
61. Saitou, N., and M. Nei. 1987. The neighbor-joining method: a new method for 697
reconstructing phylogenetic trees. Mol. Biol. Evol. 4:406-425. 698
62. Schierwater, B. 2005. My favorite animal, Trichoplax adhaerens. BioEssays 27:1294-1302. 699
63. Sekeyova, Z., P.E. Fournier, J. Rehacek, and D. Raoult. 2000. Characterization of a new 700
Spotted Fever Group Rickettsia detected in Ixodes ricinus (Acari: Ixodidae) collected in Slovakia. 701
J. Med. Entomol. 37:707-713. 702
64. Silveira, I., R. C. Pacheco, M. P. Szabo, H. G. Ramos, and M. B. Labruna. 2007. Rickettsia 703
parkeri in Brazil. Emerg. Infec. Dis. 13:1111-1113. 704
65. Simser, J. A., A. T. Palmer, U. G. Munderloh and T. J. Kurtti. 2001. Isolation of a 705
spotted fever group rickettsia, Rickettsia peacockii, in a Rocky Mountain Wood Tick, 706
Dermacentor andersoni, cell line. Appl. Environ, Microbiol. 67:546-552 707
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

32
66. Simser, J. A., A. T. Palmer, V. Fingerle, B. Wilske, T. J. Kurtti, and U. G. Munderloh. 708
2002. Rickettsia monacensis spp. nov., a spotted fever group rickettsia, from ticks (Ixodes 709
ricinus) collected in a European city park. Appl. Environ. Microbiol. 68:4559-4566. 710
67. Sober, E. 1988. Reconstructing the past: parsimony, evolution and inference. MIT Press, 711
Cambridge, Mass. 712
68. Srivastava, M., E. Begovic, J. Chapman, N. H. Putman, U. Hellsten, T. Kawashima, A. 713
Kuo, T. Mitros, A. Salamov, M. L. Carpenter, A. Y. Signorovitch, M. A. Moreno, K. 714
Kamm, J. Grimwood, J. Schmutz, H. Shapiro, I. V. Grigoriev, L. W. Buss, B. Schierwater, 715
S. L. Dellaporta, and D. S. Rokhsar. 2008. The Trichoplax genome and the nature of 716
placozoans. Nature. 454:955-960. 717
69. Stenos, J., S. R. Graves, and N. B. Unsworth. 2005. A highly sensitive and specific real-718
time PCR assay for the detection of spotted fever and typhus group rickettsiae. Am. J. Trop. 719
Med. Hyg. 73:1083-1085. 720
70. Stine, O. C., and J. P. Nataro. 2006. The evolution of bacterial toxins. In: Evolution of 721
Microbial Pathogens. H. S. Siefert and V. J. Dirita, Eds. Pp. 167-188. ASM Press. Washington 722
D.C. 723
71. Stothard, D. 1995. Ph.D. thesis. The evolutionary history of the genus Rickettsia as inferred 724
from 16S and 23S ribosomal RNA genes and the 17 kilodalton cell surface antigen gene. The 725
Ohio State University. Columbus, OH. 726
72. Swofford, D. L. 2001. PAUP*. Phylogenetic analysis using parsimony (*and other 727
methods), version 4.0b8 (PPC). 728
73. Thomas, C. M. 2000. Paradigms of plasmid organization. Mol. Microbiol. 37:585-491. 729
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

33
74. Thomas, C. M. 2000. The Horizontal Gene Pool. Harwood Academic Publishers, 730
Amsterdam, The Netherlands. 731
75. Thao, M. L., L. Baumann, P. Baumann, and N. A. Moran. 1998. Endosymbionts 732
(Buchnera) from the aphids Schizaphis graminum and Diuraphis noxia have different copy 733
numbers of the plasmid containing the leucine biosynthetic genes. Current Microbiol. 36:238-734
240. 735
76. Weinert, L. A., J. H. Werren, A. Aebi, G. N. Stone, and F. M. Jiggins. 2009. Evolution 736
and diversity of Rickettsia bacteria. BMC Biology. 7:6. doi:10.1186/1741-7007-7-6. 737
77. Weinert, L. A., J. J. Welch, and F. M. Higgins. 2009. Conjugation genes are common 738
throughout the genus Rickettsia and are transmitted horizontally. Proc. R. Soc. B 276:3619-3627. 739
78. Weller, S. J., G. D. Baldridge, U. G. Munderloh, H. Noda, J. Simser, and T. J. Kurtti. 740
1998. Phylogenetic placement of rickettsiae from the ticks Amblyomma americanum and Ixodes 741
scapularis. J. Clin. Microbiol. 36:1305-1317. 742
79. Whitman, T. J., A. L. Richards, C. D. Paddock, C. L. Tamminga, P. J. Sniezek, J. Jiang, 743
D. K. Byers, and J. W. Sanders. 2007. Rickettsia parkeri infection after tick bite, Virginia. 744
Emerg. Infec. Dis. 13:334-336. 745
746
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

34
FIGURE LEGENDS 747
748
Fig. 1. Presence of plasmids in R. rhipicephali from S. America and REIS from N. America. (A) 749
PFGE of DNA from REIS isolate Camp Ripley, R. rhipicephali isolate HJ#5 and R. monacensis 750
isolate IrR/Munich. Relative migration positions of rickettsial chromosomal (C), ISE6 host cell 751
mitochondrial (M), and prominent rickettsial plasmid (P) DNAs are indicated at right. Migration 752
positions of 25, 50 and 100 kbp linear DNA markers are indicated at left. Panels B, C and D: 753
Southern blots of replicate gels hybridized with pRM16, pRM6 and pRM8 gene probes, 754
respectively. Band identities (right) and migration positions of marker DNAs (left). 755
756
Fig. 2. Presence of plasmids in R. akari and R. amblyommii isolates and absence of plasmids in 757
R. parkeri isolates from N. America. (A) PFGE of DNA from R. akari isolate Bronx, R. parkeri 758
isolates Ft. Story, Portsmouth and Tates Hell and R. amblyommii isolate Darkwater. Panels B, C, 759
and D: Southern blots of replicate gels hybridized with pRM16, pRM6 and pRM8 gene probes. 760
Band identities (right) and migration positions of marker DNAs (left) as in Fig. 1. 761
762
Fig. 3. Presence and absence of plasmids in South American isolates of R. amblyommii and R. 763
parkeri, respectively. (A) PFGE of DNA from R. amblyommii isolates An13 and Ac37 and R. 764
parkeri isolates At#24 and At#5. Panels B, C and D: Southern blots of replicate gels hybridized 765
with pRM16, pRM6 and pRM8 gene probes, respectively. Band identities (right) and migration 766
positions of marker DNAs (left) are as in Fig. 1. 767
768
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

35
Fig. 4. Presence of plasmids in S. American isolates of R. bellii. (A) PFGE of DNA from R. 769
bellii isolates ovale, Ad25, An4, Mogi and HJ#7 (S. America) and RML 369-C (N. America). 770
(B) Southern blot of same gel hybridized with the pRM16 gene probe. Panels C, D and E: 771
Southern blots of replicate gels hybridized with pRM6, pRM8 and pRM12 gene probes, 772
respectively. Band identities (right) and migration positions of marker DNAs (left) as in Fig. 1. 773
774
Fig. 5. A probe derived from the R. monacensis pRM23 gene encoding a homolog of a 775
Burkholderia thailandensis transposon resolvase gene hybridizes to plasmids of three Rickettsia 776
spp. (A) PFGE of DNA from R. monacensis IrR/Munich, R. akari Bronx, REIS and R. 777
rhipicephali HJ#5. (B) Southern blot of same gel hybridized with the pRM23 probe. Band 778
identities (right) and migration positions of marker DNAs (left) as in Fig. 1. 779
780
Fig. 6. Maximum parsimony phylogenetic tree (unrooted) of rickettsial ParA proteins. Scale bars 781
indicate number amino acid changes within branch lengths. Bootstrap scores indicated by 782
numerals. GenBank Accession numbers: plasmid-encoded ParA sequences were pRAM18 783
(GU322808), pRAM23 (GU322807), pRAF (YP_002845772), pREIS1, 2, 3 and 4 (EER20867, 784
EER20809, EER20748, and NZ_GG688316 – translated nucleotides 10818 to 11573), pRF 785
(YP_247439), pRM (YP_001967398), pRMA (YP_001497196), pRPR (YP_002921997), cp32 786
(AAL60459), pPSR1 (NP_940697), pWes-1 (YP_002332254), pYptb32953 (YP_068544), 787
pMATVIM7 (YP_001427366), 153 kb plasmid (YP_001393406). Trichoplax adhaerens contig: 788
(EDV19036). Chromosome-encoded ParA sequences were: E. chaffeensis (ABD44691), R. 789
africae (YP_002844799), R. bellii (YP_538520), R. canadensis (YP_001491792), R. felis 790
(YP_246133), I. scapularis endosymbiont (ZP_04698724), R. massiliae (YP_001498948), 791
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

36
R. peacockii (ACR47329), R. typhi (YP_067042). 792
793
Fig. 7. Maximum parsimony phylogenetic tree (unrooted) of rickettsial small heat shock 794
proteins. Scale bars indicate number amino acid changes within branch lengths. Bootstrap scores 795
indicated by numerals. GenBank Accession numbers: plasmid-encoded or *PCR-amplified 796
plasmid-encoded Hsp sequences were pRAM23 (GU322807), pRB* (GU180086), pRF 797
(YP_247467 and YP_170678), pRHE* (GU180087), pRHO* (GU180088), pRIS 798
(NZ_CM000771), pRM (YP_001967388 and YP_001967389), pRPR (YP_002922008 and 799
YP_002922009); chromosome-encoded Hsp sequences were R. africae (ZP_02336191), R. akari 800
(YP_001493212), R. bellii (YP_538007), R. canadensis (YP_001492044), R. conorii 801
(NP_360000), R. felis (YP_247020 and YP_247021), R. massiliae (YP_001499170), R. 802
peacockii (YP_002916350), R. prowazekii (NP_220658), R. rickettsii (YP_001494483), R. 803
siberica (ZP_00142181), R. typhi (YP_067227), B. thailandensis (ZP_02384335), Francisella 804
tularensis (YP_170678), L. pneumophila (YP_096204), T. adhaerens (XP_002118475), and W. 805
pipientis (YP_001975380). 806
807
808
809
810
811
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

Table 1. Origins and passage histories of Rickettsia species evaluated for plasmids and plasmid copy number.
Rickettsia sp. Isolate Source Geographic origin Year of isolation Passage history1
Reference(s)
R. akari Bronx human New York 2002 5 (E6) 51
“R. amblyommii” AaR/SC Amblyomma. americanum South Carolina 1999 6 (ISE6)2
T.J. Kurtti, unpubl.
Ac/Pa Amblyomma cajennense Panama 2007 2 (ISE6)2 U.G. Munderloh, unpubl
Ac37 A. cajennense Brazil 2004 5 (E6) 2 (ISE6) 32
An13 Amblyomma neumanni Argentina 2007 7 (E6), 2(ISE6) 33
Darkwater A. americanum Florida 2006 5 (E6) C.D. Paddock, unpubl.
R. bellii RML 369-C Dermacentor variabilis Arkansas 1966 uncertain, 2 (ISE6) E.J. Bell, unpubl, 53
An4 Amblyomma neumanni Argentina 2007 5 (E6), 4 (ISE6) 33
HJ#7 Haemaphysalis juxtakochi Brazil 2007 8 (E6), 2 (ISE6) 34
Ad25 Amblyomma dubitatum Brazil 2007 3 (E6), 2 (ISE6) 48
Ovale Amblyomma ovale Brazil 2007 3 (E6), 2 (ISE6) 47
Mogi Amblyomma aureolatum Brazil 2006 11 (E6), 2 (ISE6) 57
R. felis LSU Ctenocephalides felis Louisiana 2006 9 (ISE6)2 59
R. helvetica C9P9 Ixodes ricinus Switzerland 1993 uncertain, 14 (ISE6) 8
R. hoogstraalii RCCE3 Carios capensis Georgia 2007 37 (CCE5)2, 12 (ISE18) 38
R. monacensis IrR/Munich I. ricinus Germany 2002 18 - 29 (ISE6)2 65
R. parkeri Portsmouth human Virginia 2002 8 (E6) 50
Ft. Story human Virginia 2006 5 (E6) 52
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

Tate’s Hell Amblyomma maculatum Florida 2005 4 (E6) 53
High Bluff A. maculatum Florida 2007 3 (E6) 53
Grand Bay A. maculatum Mississippi 2007 3 (E6) 53
Oktibbeha A. maculatum Mississippi 2007 6 (E6) 53
AT#5 Amblyomma triste Uruguay 2006 9 (E6), 2 (ISE6) 46
AT#24 A. triste Brazil 2007 8 (E6), 2 (ISE6) 64
R. peacockii DAE100R Dermacentor andersoni Colorado 2001 11 (DAE100)2
, 5 (ISE6) 65
R. rhipicephali HJ#5 H. juxtakochi Brazil 2007 5 (E6), 2 (ISE6) 34
REIS same Ixodes scapularis Minnesota 2008 7 (IRE11)2
T.J. Kurtti, unpubl.
1Number of passages in cultured cell lines: Vero (E6); I. scapularis embryonic (ISE6) and (ISE18); I. ricinus embryonic (IRE11); D. andersoni
embryonic (DAE100). Carios capensis (CCE5). 2Indicates cell line of Rickettsia sp. isolation if other than Vero. R. bellii 369-C was isolated in
embryonated chicken eggs.
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from
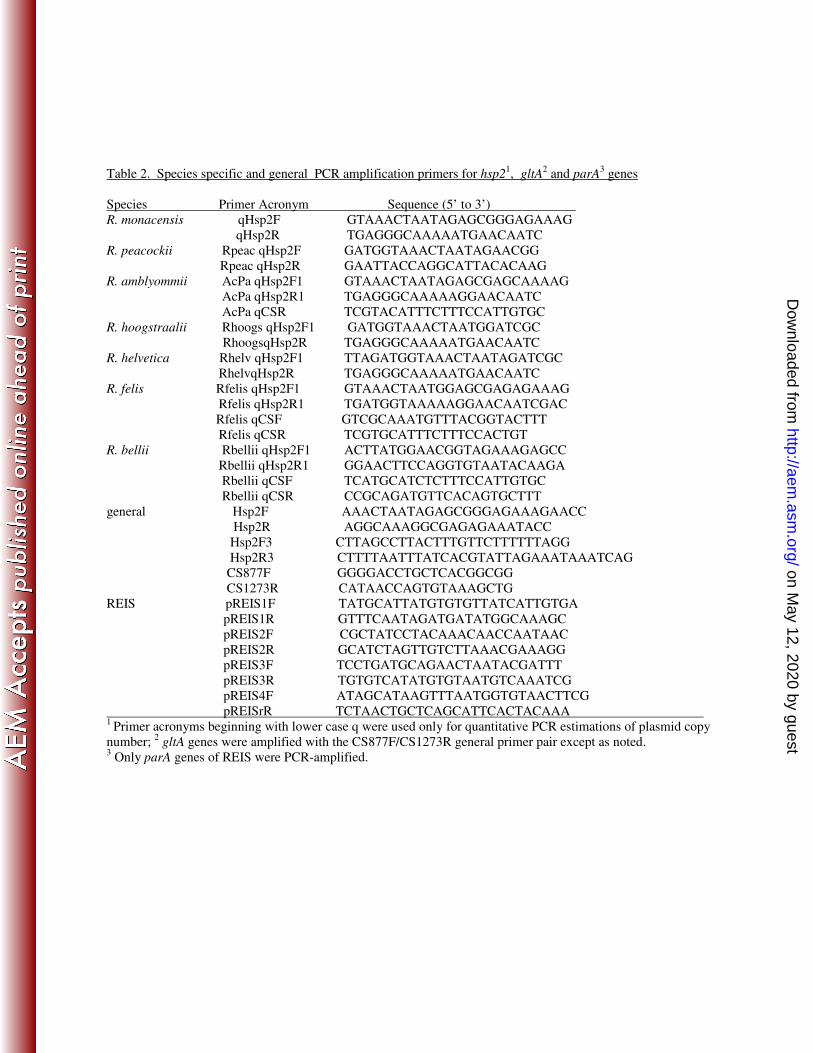
Table 2. Species specific and general PCR amplification primers for hsp21, gltA
2 and parA
3 genes
Species Primer Acronym Sequence (5’ to 3’)
R. monacensis qHsp2F GTAAACTAATAGAGCGGGAGAAAG
qHsp2R TGAGGGCAAAAATGAACAATC
R. peacockii Rpeac qHsp2F GATGGTAAACTAATAGAACGG
Rpeac qHsp2R GAATTACCAGGCATTACACAAG
R. amblyommii AcPa qHsp2F1 GTAAACTAATAGAGCGAGCAAAAG
AcPa qHsp2R1 TGAGGGCAAAAAGGAACAATC
AcPa qCSR TCGTACATTTCTTTCCATTGTGC
R. hoogstraalii Rhoogs qHsp2F1 GATGGTAAACTAATGGATCGC
RhoogsqHsp2R TGAGGGCAAAAATGAACAATC
R. helvetica Rhelv qHsp2F1 TTAGATGGTAAACTAATAGATCGC
RhelvqHsp2R TGAGGGCAAAAATGAACAATC
R. felis Rfelis qHsp2F1 GTAAACTAATGGAGCGAGAGAAAG
Rfelis qHsp2R1 TGATGGTAAAAAGGAACAATCGAC
Rfelis qCSF GTCGCAAATGTTTACGGTACTTT
Rfelis qCSR TCGTGCATTTCTTTCCACTGT
R. bellii Rbellii qHsp2F1 ACTTATGGAACGGTAGAAAGAGCC
Rbellii qHsp2R1 GGAACTTCCAGGTGTAATACAAGA
Rbellii qCSF TCATGCATCTCTTTCCATTGTGC
Rbellii qCSR CCGCAGATGTTCACAGTGCTTT
general Hsp2F AAACTAATAGAGCGGGAGAAAGAACC
Hsp2R AGGCAAAGGCGAGAGAAATACC
Hsp2F3 CTTAGCCTTACTTTGTTCTTTTTTAGG
Hsp2R3 CTTTTAATTTATCACGTATTAGAAATAAATCAG
CS877F GGGGACCTGCTCACGGCGG
CS1273R CATAACCAGTGTAAAGCTG
REIS pREIS1F TATGCATTATGTGTGTTATCATTGTGA
pREIS1R GTTTCAATAGATGATATGGCAAAGC
pREIS2F CGCTATCCTACAAACAACCAATAAC
pREIS2R GCATCTAGTTGTCTTAAACGAAAGG
pREIS3F TCCTGATGCAGAACTAATACGATTT
pREIS3R TGTGTCATATGTGTAATGTCAAATCG
pREIS4F ATAGCATAAGTTTAATGGTGTAACTTCG
pREISrR TCTAACTGCTCAGCATTCACTACAAA 1
Primer acronyms beginning with lower case q were used only for quantitative PCR estimations of plasmid copy
number; 2 gltA genes were amplified with the CS877F/CS1273R general primer pair except as noted.
3 Only parA genes of REIS were PCR-amplified.
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

Table 3. Southern blot detection of pRM gene homologs on plasmids of New World Rickettsia isolates.
pRM R. akari R. amblyommii R. bellii R. rhipicephali REIS
genea Bronx An13 Ac37 Dark
b Ad25 An4 HJ#37 Mogi ovale HJ#5 Camp Ripley
pRM6 - + + + + + + + - + +
pRM8 - + + + + - + +
+ - -
pRM12 - - - - + - + + - -
+
pRM16 + + + + + + + + + + +
pRM23 + - - - - - - - - + +
aThe R. monacensis pRM plasmid genes encode respectively, from top to bottom, a small heat shock
protein, a RecD/TraA protein, a proline betaine membrane transporter, a DnaA-like replication
initiator and a transposon resolvase (6). b isolate Darkwater.
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from

Table 4. Plasmid Copy Numbers in Selected Rickettsia spp.
Species (isolate) copy numbera SD
b
R. helvetica (C9P9) 2.4 0.3
R. monacensis (IrR/Munich) 2.7 0.3
R. amblyommii (Ac/Pa) 3.5 0.1
R. hoogstraalii (RCCE3) 5.5 1.1
R. bellii (An4) 4.3 0.4
R. felis (LSU) 5.9 0.5
R. peacockii ( DAE100R) 9.2 1. 9
aratio of hsp2 to gltA gene number expressed as means of
three experiments of three replicates each. bstandard deviation.
on May 12, 2020 by guest
http://aem.asm
.org/D
ownloaded from


















