Role of cell walls in the bioaccessibility of lipids in ...ucanr.edu/datastoreFiles/608-33.pdf ·...
10
Role of cell walls in the bioaccessibility of lipids in almond seeds 1–3 Peter R Ellis, Cyril WC Kendall, Yilong Ren, Charlotte Parker, John F Pacy, Keith W Waldron, and David JA Jenkins ABSTRACT Background: Certain nutrients and phytochemicals in almonds may confer protection against cardiovascular disease, but little is known about factors that influence their bioavailability. A crucial and rel- evant aspect is the amount of these dietary components available for absorption in the intestine, which is a concept referred to as bioac- cessibility. Objective: We investigated the role played by cell walls in influ- encing the bioaccessibility of intracellular lipid from almond seeds. Design: Quantitative analyses of nonstarch polysaccharides (NSPs) and phenolic compounds of cell walls were performed by gas-liquid chromatography and HPLC, respectively. In a series of experiments, the effects of mechanical disruption, chewing, and digestion on almond seed microstructure and intracellular lipid release were de- termined. In the digestibility study, fecal samples were collected from healthy subjects who had consumed diets with or without almonds. Almond seeds and fecal samples were examined by mi- croscopy to identify cell walls and intracellular lipid. Results: Cell walls were found to be rich in NSPs, particularly arabinose-rich polysaccharides, with a high concentration of phe- nolic compounds detected in the seed coat cell wall. During disrup- tion of almond tissue by mechanical methods or chewing, only the first layer of cells at the fractured surface was ruptured and able to release lipid. In fecal samples collected from subjects consuming the almond diet, we observed intact cotyledonary cells, in which the cell walls encapsulated intracellular lipid. This lipid appeared suscepti- ble to colonic fermentation once the cotyledonary cell walls were breached by bacterial degradation. Conclusion: The cell walls of almond seeds reduce lipid bioaccessi- bility by hindering the release of lipid available for digestion. Am J Clin Nutr 2004;80:604 –13. KEY WORDS Almond seeds, plant cell wall, nonstarch poly- saccharides, dietary fiber, phenolics, lipid digestion, bioavailability, microstructure, microscopy, chewing, fecal fat INTRODUCTION Studies show that diets rich in tree nuts, including almonds, have beneficial effects on risk factors for coronary artery disease (CAD) (1–7). Various nutrients and phytochemicals found in almonds and other tree nuts, including unsaturated fatty acids, plant sterols, -tocopherol, dietary fiber, and phenolic compo- nents, could be responsible for modifying CAD risk factors (2, 3, 7). However, a major obstacle in evaluating the role of individual food components in modifying disease risk is the paucity of information on factors that influence their bioavailability (8). One critical aspect of this information is the proportion of nutri- ents “released” from a complex food matrix and, therefore, po- tentially available for absorption in the gastrointestinal (GI) tract, a concept that was recently referred to as bioaccessibility (8). To study factors that affect bioaccessibility it is crucial to have a basic understanding of the physical and chemical changes in the food matrix during the digestive process (9 –13). In plant food tissues, the physicochemical structure and prop- erties of cell walls (CWs) in the GI lumen are critical factors involved in influencing bioaccessibility (14, 15). Thus, for ex- ample, intact CWs of starch-rich leguminous seeds can act as a physical barrier to the action of -amylase, thereby hindering the rate and extent of starch digestion (16, 17). In almond seeds, however, lipid is the main storage component, which comprises 50% of the total weight of the seed and is located as intracel- lular oil bodies of diameter size ranging from 1 to 5 m (18). An important question is, therefore, do the CWs of almond tissue act as a physical barrier to the action of lipase after ingestion? Such an effect is likely to reduce the rate and extent of lipolysis in the proximal gut. Furthermore, if undigested lipid from al- mond tissue is transported to more distal sites of the GI tract, this could lead to increased excretion of fecal energy, which has implications for the regulation of body weight. An early, but somewhat limited, study showed that human subjects who had consumed a peanut-rich diet for 6 d excreted a significant amount of fecal lipid (19). More recent studies indicated that significant amounts of lipid were excreted in subjects on diets rich in either almonds (20) or pecans (21). However, none of these studies included an investigation of the factors that are responsible for this apparent “malabsorption,” but a decrease in lipid digestion caused by an intact CW barrier is one possible mechanism. The physiologic effects of the almond seed CW and how intracellular lipid and other nutrients are released will depend on the physical state of the almond tissue in the gut lumen. Thus, a critical factor will be whether the CWs are disrupted during processing and chewing and in transit along the GI tract (14, 15). 1 From the Biopolymers Group, Department of Life Sciences, King’s College London, University of London (PRE, YR, JFP); the Department of Nutritional Sciences, Faculty of Medicine, University of Toronto (CWCK and DJAJ); and the BBSRC Institute of Food Research, Norwich, United Kingdom (CP and KWW). 2 Supported by the Almond Board of California and the BBSRC. 3 Address reprint requests to PR Ellis, Biopolymers Group, Department of Life Sciences, King’s College London, Franklin-Wilkins Building, 150 Stam- ford Street, London SE1 9NN, United Kingdom. E-mail: [email protected]. Received July 7, 2003. Accepted for publication March 9, 2004. 604 Am J Clin Nutr 2004;80:604 –13. Printed in USA. © 2004 American Society for Clinical Nutrition at Univ of California-Davis Carlson Hlth Sci Lib on May 3, 2011 www.ajcn.org Downloaded from
-
Upload
truongngoc -
Category
Documents
-
view
216 -
download
1
Transcript of Role of cell walls in the bioaccessibility of lipids in ...ucanr.edu/datastoreFiles/608-33.pdf ·...

Role of cell walls in the bioaccessibility of lipids in almond seeds1–3
Peter R Ellis, Cyril WC Kendall, Yilong Ren, Charlotte Parker, John F Pacy, Keith W Waldron, and David JA Jenkins
ABSTRACTBackground: Certain nutrients and phytochemicals in almonds mayconfer protection against cardiovascular disease, but little is knownabout factors that influence their bioavailability. A crucial and rel-evant aspect is the amount of these dietary components available forabsorption in the intestine, which is a concept referred to as bioac-cessibility.Objective: We investigated the role played by cell walls in influ-encing the bioaccessibility of intracellular lipid from almond seeds.Design: Quantitative analyses of nonstarch polysaccharides (NSPs)and phenolic compounds of cell walls were performed by gas-liquidchromatography and HPLC, respectively. In a series of experiments,the effects of mechanical disruption, chewing, and digestion onalmond seed microstructure and intracellular lipid release were de-termined. In the digestibility study, fecal samples were collectedfrom healthy subjects who had consumed diets with or withoutalmonds. Almond seeds and fecal samples were examined by mi-croscopy to identify cell walls and intracellular lipid.Results: Cell walls were found to be rich in NSPs, particularlyarabinose-rich polysaccharides, with a high concentration of phe-nolic compounds detected in the seed coat cell wall. During disrup-tion of almond tissue by mechanical methods or chewing, only thefirst layer of cells at the fractured surface was ruptured and able torelease lipid. In fecal samples collected from subjects consuming thealmond diet, we observed intact cotyledonary cells, in which the cellwalls encapsulated intracellular lipid. This lipid appeared suscepti-ble to colonic fermentation once the cotyledonary cell walls werebreached by bacterial degradation.Conclusion: The cell walls of almond seeds reduce lipid bioaccessi-bility by hindering the release of lipid available for digestion. Am JClin Nutr 2004;80:604–13.
KEY WORDS Almond seeds, plant cell wall, nonstarch poly-saccharides, dietary fiber, phenolics, lipid digestion, bioavailability,microstructure, microscopy, chewing, fecal fat
INTRODUCTION
Studies show that diets rich in tree nuts, including almonds,have beneficial effects on risk factors for coronary artery disease(CAD) (1–7). Various nutrients and phytochemicals found inalmonds and other tree nuts, including unsaturated fatty acids,plant sterols, �-tocopherol, dietary fiber, and phenolic compo-nents, could be responsible for modifying CAD risk factors (2, 3,7). However, a major obstacle in evaluating the role of individualfood components in modifying disease risk is the paucity ofinformation on factors that influence their bioavailability (8).
One critical aspect of this information is the proportion of nutri-ents “released” from a complex food matrix and, therefore, po-tentially available for absorption in the gastrointestinal (GI) tract,a concept that was recently referred to as bioaccessibility (8). Tostudy factors that affect bioaccessibility it is crucial to have abasic understanding of the physical and chemical changes in thefood matrix during the digestive process (9–13).
In plant food tissues, the physicochemical structure and prop-erties of cell walls (CWs) in the GI lumen are critical factorsinvolved in influencing bioaccessibility (14, 15). Thus, for ex-ample, intact CWs of starch-rich leguminous seeds can act as aphysical barrier to the action of �-amylase, thereby hindering therate and extent of starch digestion (16, 17). In almond seeds,however, lipid is the main storage component, which comprises�50% of the total weight of the seed and is located as intracel-lular oil bodies of diameter size ranging from �1 to 5 �m (18).An important question is, therefore, do the CWs of almond tissueact as a physical barrier to the action of lipase after ingestion?Such an effect is likely to reduce the rate and extent of lipolysisin the proximal gut. Furthermore, if undigested lipid from al-mond tissue is transported to more distal sites of the GI tract, thiscould lead to increased excretion of fecal energy, which hasimplications for the regulation of body weight. An early, butsomewhat limited, study showed that human subjects who hadconsumed a peanut-rich diet for 6 d excreted a significant amountof fecal lipid (19). More recent studies indicated that significantamounts of lipid were excreted in subjects on diets rich in eitheralmonds (20) or pecans (21). However, none of these studiesincluded an investigation of the factors that are responsible forthis apparent “malabsorption,” but a decrease in lipid digestioncaused by an intact CW barrier is one possible mechanism.
The physiologic effects of the almond seed CW and howintracellular lipid and other nutrients are released will depend onthe physical state of the almond tissue in the gut lumen. Thus, acritical factor will be whether the CWs are disrupted duringprocessing and chewing and in transit along the GI tract (14, 15).
1 From the Biopolymers Group, Department of Life Sciences, King’sCollege London, University of London (PRE, YR, JFP); the Department ofNutritional Sciences, Faculty of Medicine, University of Toronto (CWCKand DJAJ); and the BBSRC Institute of Food Research, Norwich, UnitedKingdom (CP and KWW).
2 Supported by the Almond Board of California and the BBSRC.3 Address reprint requests to PR Ellis, Biopolymers Group, Department of
Life Sciences, King’s College London, Franklin-Wilkins Building, 150 Stam-ford Street, London SE1 9NN, United Kingdom. E-mail: [email protected].
Received July 7, 2003.Accepted for publication March 9, 2004.
604 Am J Clin Nutr 2004;80:604–13. Printed in USA. © 2004 American Society for Clinical Nutrition
at Univ of C
alifornia-Davis C
arlson Hlth S
ci Lib on May 3, 2011
ww
w.ajcn.org
Dow
nloaded from

In the present study, we investigated the role played by CWsin influencing lipid release during laboratory processing, chew-ing, and digestion of almond seeds (kernels). Quantitative chem-ical analyses of CWs were performed to provide some under-standing of CW properties in vitro and in vivo. Also, a detailedexamination of almond seed microstructure was carried out, fo-cusing mainly on the topologic relation between the CW and theintracellular lipid. One important objective of this work was toidentify undigested almond tissue in fecal material recoveredfrom human subjects who consumed an almond-rich diet.
SUBJECTS AND METHODS
Test products
Almond seeds or kernels (Amygdalus communis L; variety,Nonpareil) selected for this study were kindly provided by theAlmond Board of California. Roasted almond kernels of thesame variety were also used in this investigation. All the sampleswere stored at a refrigerated temperature of 3–5 °C. The nutrientcontent (percentage by weight of edible portion) of the raw al-monds was as follows: moisture 5.5%, protein 21.2%, lipid55.1%, available carbohydrate (mainly sugars) 5.5%, dietaryfiber 11.8% (derived from the CW), and ash (minerals) 3.1%.Values for moisture and lipid were obtained by proximate anal-ysis (described in Chemical analysis) and for other nutrients fromthe US Department of Agriculture nutrient database (22).
Subjects
Seven human volunteers (5 men and 2 women) with a meanage (� SE) of 37 � 5.9 y (range: 23–64 y) and an average bodymass index of 24.7 � 1.9 kg/m2 (range: 17.4–32.5 kg/m2) par-ticipated in the chewing study. For the digestibility study, 3healthy human subjects (3 men) with mean (� SE) age of 31 �5.4 y (range: 22–40 y) and body mass index of 24.3 � 0.6 kg/m2
participated.
Protocol of human studies
Chewing study
To assess the effects of physical disruption by chewing, amethod mainly based on a technique developed by Granfeldt etal (9) was used. Each volunteer was asked not to eat for 2 h beforethe experiment. The subjects were instructed to rinse theirmouths with drinking water (bottled still) and then chew a 2-gsample of almond seeds (both raw and roasted types chewed onseparate occasions) 30 times for �30 s. In an earlier experiment,this technique was found to provide the conditions required tomasticate the seeds to a physical consistency that allows swal-lowing to take place. The subjects were then asked to expectoratethe contents of their mouths into a petri dish, samples of whichwere then immediately chemically fixed and later stained andexamined by microscopy, as described in the section “Micro-structural analysis.” Chewed samples from each of the 7 volun-teers were examined separately. The subjects had previouslygiven their informed consent to participate in the study, whichwas approved by the Research Ethics Committee of King’s Col-lege London (Reference: 01/02–26).
Digestibility study
Three human subjects consumed increasing amounts of rawalmonds over a 3-d period (100, 150, and 200 g/d, which is
equivalent to �55, 83, and 110 g/d lipid and 11.8, 17.7, and 23.5g/d dietary fiber, respectively). Subjects were requested to limitintakes of other plant foods, to facilitate the location of the al-mond tissue in the fecal samples. On the day immediately afterthe feeding period (ie, day 4), a fecal sample was collected fromeach subject in a plastic bag with use of an under-seat frame andthen weighed. Core biopsies of these fecal samples were imme-diately taken and then placed in a primary fixative within 10 min;the rest of the samples were frozen (�20 °C) and stored for futurechemical analysis. In a similar manner, a fecal sample (negativecontrol) from the same human subjects was also collected after a3-d period, during which no almonds or other nuts were con-sumed. All fecal samples were coded, so that the investigatorswere not aware of which were the control and test samples. Thefixed fecal samples from each of the subjects were examined withuse of a full range of microscopy techniques, and other sampleswere quantitatively analyzed for macronutrients, as described in“Chemical analysis.” The subjects had previously given theirinformed consent to participate in the study, which was approvedby the University of Toronto Research Ethics Board.
Laboratory processing of almond seeds
In the initial experiments, laboratory methods that simulate thechewing process were used to provide physically disrupted al-mond seed. Microscopic examination of this material provideduseful information about the effects of trituration on the almondtissue under controlled conditions of physical processing. Tosimulate a cutting action during chewing, the whole almondseeds (raw and roasted types) were mechanically processed withuse of a food processor (type 643; Moulinex, Alencon, France)for 30 s with the cutting blades set at the highest speed setting. Tosimulate a pulverizing action during chewing, a hand-grindingprocess of the almond samples was performed with use of amortar and pestle, grinding continuously for 10 min.
To facilitate characterization of almond seeds macerated bythe 2 above-mentioned methods it was decided to separate theparticulate material into 4 fractions according to particle sizedistribution. This separation would facilitate the characterizationof the almond tissue. Also, it was anticipated that more lipidwould be released from the smaller particles, given the greatersurface area-to-volume ratio of these particles, relative to thelarger ones. These 4 fractions were separated by a mechanicalsieving process (23) according to the following particle sizeranges: between 3.35 and 2.00 mm, 1.99 and 1.00 mm, 0.99 and0.5 mm, and �0.5 mm.
Microstructural analysis
The laboratory-processed, chewed, and fecal samples wereexamined by light, scanning, and transmission electron micros-copy to assess the morphology of the CWs and the morphologyand distribution of intracellular lipids released from any rupturedcells.
Laboratory-processed and chewed samples were rapidly fixed(5-min duration) in 1% (wt:vol) osmium tetroxide and then trans-ferred to 2.5% (by vol) glutaraldehyde in 0.1 M sodium cacody-late buffer (pH � 7.2) and left overnight. Fecal samples werefixed in 2.5% (wt:vol) paraformaldehyde and 0.5% (by vol)glutaraldehyde. Subsequently, all the samples were washedtwice, each time for 30 min, in 0.1 M sodium cacodylate orphosphate buffer and then postfixed in 1% (wt:vol) osmium
CELL WALLS AND BIOACCESSIBILITY OF LIPIDS 605
at Univ of C
alifornia-Davis C
arlson Hlth S
ci Lib on May 3, 2011
ww
w.ajcn.org
Dow
nloaded from

tetroxide for 2 h. Samples were dehydrated in graded acetone orethanol serial dilutions (ie, 50%, 70%, and 90%, by vol, acetoneor ethanol and distilled water) for 30 min for each solution andthen finally in 100% acetone or ethanol for 30 min (3 times).
For the transmission electron microscopy (TEM) and lightmicroscopy, the almond samples were infiltrated with Spurrresin and embedded in molds and polymerized at 60 °C. Sections(1 �m for light microscopy) were cut on a Reichert Ultracutultramicrotome (Leica Microsystems Ltd, Milton Keynes,United Kingdom), mounted on glass slides, and stained in 1%(wt:vol) Toluidine Blue, Sudan black B, or periodic acid-Schiffreagent. Sections of seed testa (skin) were also prepared andstained for lignin with use of phloroglucinol-HCl. Ultrathin sec-tions (�70 nm) were cut for TEM and viewed in a JEOL 100CXMk.II transmission electron microscope (JEOL Ltd, WelwynGarden City, United Kingdom).
For examination by scanning electron microscopy (SEM) thesamples were critically point dried in a Polaron E3000 CP Drier(Quorum Technologies, Newhaven, United Kingdom), mountedon stubs, and sputter coated with gold in a Polaron E5100 sputtercoating unit, and viewed in a JEOL 25SM and a Philips 501scanning electron microscope (FEI Company, Cambridge,United Kingdom).
Chemical analysis
Cell wall components from cotyledon and seed coat ofalmond seeds
Cell wall material (CWM) was prepared from almond seeds(raw and roasted types) with use of methods adapted from pre-vious work (24). Almond seeds were weighed (�25 g of eachtype) and blended in 1.5% (wt:vol) sodium dodecyl sulfate (SDS)with use of a Ystral homogenizer (Ystral GmbH, Dottingen,Germany) for 5 min. A few drops of octanol were added to reducefoaming. The homogenate was filtered through a 75-�m nylonmesh. The residue was ball-milled at 0 °C in 0.5% (wt:vol) SDSat 60 rpm for 12 h. Microscopy was carried out to ensure that theCWM was free of any intracellular material. The CW residue wassuspended in 200 mL 0.5% (wt:vol) SDS after filtering through75-�m nylon mesh and homogenized for 5 min. This procedurewas repeated twice. Further homogenization was carried out withresidue resuspended in 200 mL deionized water. After filteringthrough 75-�m nylon mesh the residue was suspended in 100 mLethanol and homogenized. This procedure was repeated twice.The residue was extracted with hot-ethanol (80 °C) and filteredthrough a 3-mm Whatman filter (Whatman plc, Brentford, Mid-dlesex, United Kingdom) overlaid with a nylon mesh (75 �m).After several washes with acetone the CWM was allowed to dryovernight.
Analysis of CW sugar composition was carried out by acidhydrolysis of polysaccharides followed by reduction, acetyla-tion, and quantitation of alditol acetates by gas-liquid chroma-tography, using methods adapted from Blakeney et al (25).Uronic acids were measured by the method of Blumenkrantz andAsboe-Hansen (26).
Ester-linked phenolic components of CWM were analyzed bythe method of Waldron et al (27). CWM (�30 mg) prepared fromalmond seed was extracted with 0.1 M NaOH for 1 h at roomtemperature under N2. The suspension was filtered through aWhatman (GFA) filter paper overlaid with nylon mesh (75 �m).The residue was retained for further extraction. trans-Cinnamic
acid was added (200 �L, 1.67 mg/50 mL methanol) to the filtrateas an internal standard. The filtrate was then acidified with con-centrated HCl and extracted 3 times with ethyl acetate. Theextracts were combined and rotary evaporated to dryness thenreconstituted in 200 �L methanol and analyzed by HPLC. Pre-viously retained residue was sequentially extracted further in 0.1M NaOH (24 h), 1 M NaOH (24 h), and 2 M NaOH (24 h) withuse of the same procedure. Extracted phenolic compounds wereanalyzed by HPLC (27).
Macronutrient analysis of raw almond seeds and humanfecal samples collected during digestibility study
Total moisture (by oven drying) and lipid (Soxhlet; light pe-troleum, bp 40–60 °C, diethyl ether) contents of the originalalmond seeds (on flour samples prepared with use of the Mou-linex food processor, to pass through a sieve size 500 �m) weredetermined with use of standard methods of the Association ofOfficial Analytic Chemists (AOAC) (28).
Frozen fecal samples were partially thawed, homogenized ina blender (Osterizer model; Sunbeam Corporation, Mississauga,Canada), freeze dried (freeze dry–shell freeze system; LabconcoCorporation, Kansas City, KS), and then analyzed for moisturecontent and total lipids with use of methods of AOAC (28). Thetotal moisture content of feces at the time of collection wasdetermined by an AOAC method (28) and performed on samplesthat were partially thawed and homogenized only.
Statistical analysis
Paired Student’s t tests (2-tailed) were performed on the quan-titative digestibility data (ie, fecal weight and fecal moisture andlipid contents). Statistical differences between the control andtest (almond-containing) samples were accepted at P � 0.05.
RESULTS
Release of lipids by laboratory processing and chewing
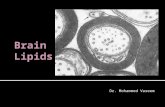
Many of the cotyledonary cells of the almond seed tissueremained physically intact after mechanical trituration. Thus, themicrographs seen in Figure 1, A and B, were prepared fromalmond seed, which was physically disrupted by hand-grinding(pulverizing), and show structurally intact areas but not the frac-tured surfaces. Carbohydrates were located with use of Schiff’sreagent in the CW and intracellular inclusions (Figure 1A). InFigure 1B, the intracellular lipid bodies, stained with Sudanblack, appear to fill much of the intracellular space. In all of thephysically disrupted particles of almond tissue, the CW on thefractured surface (ie, the first layer of cells) were ruptured, so thatmuch of the lipid was released from this location, as indicated inFigure 1, C and D. In these sections Toluidine Blue was used forstaining the CW and intracellular components, including lipid,which is quantitatively the major nutrient of the seed. The lighterblue stain surrounding the seed particle section indicates lipidthat was released from the ruptured cells during processing (Fig-ure 1, C and D).
The SEM micrograph of almond samples disrupted by thecutting method revealed a surface mainly covered in lipid drop-lets (Figure 1E). These droplets were significantly larger in di-ameter size (�10–40 �m) than the intact intracellular lipid bod-ies of �1–5 �m (TEM micrograph in Figure 2), indicating thatthese bodies had coalesced on the fractured surfaces of the cut
606 ELLIS ET AL
at Univ of C
alifornia-Davis C
arlson Hlth S
ci Lib on May 3, 2011
ww
w.ajcn.org
Dow
nloaded from
tissue. The presence of lipid on the particle surfaces (Figure 1E)indicates that the CW of the first layer was ruptured, which wasseen more clearly when most of the lipid was removed by gentlewashing with petroleum ether for 3–4 min (Figure 1F).
Examining the almond tissue over a range of particle sizesprovided no evidence of cell separation, which is consistent withthe view that cotyledonary cells are ruptured at the fracturedsurface, releasing intracellular lipids. The only explanation ofthis is that the cell-cell adhesion of the almond tissue is verystrong, so that the cells can only be disrupted by breakage ofthe CW.
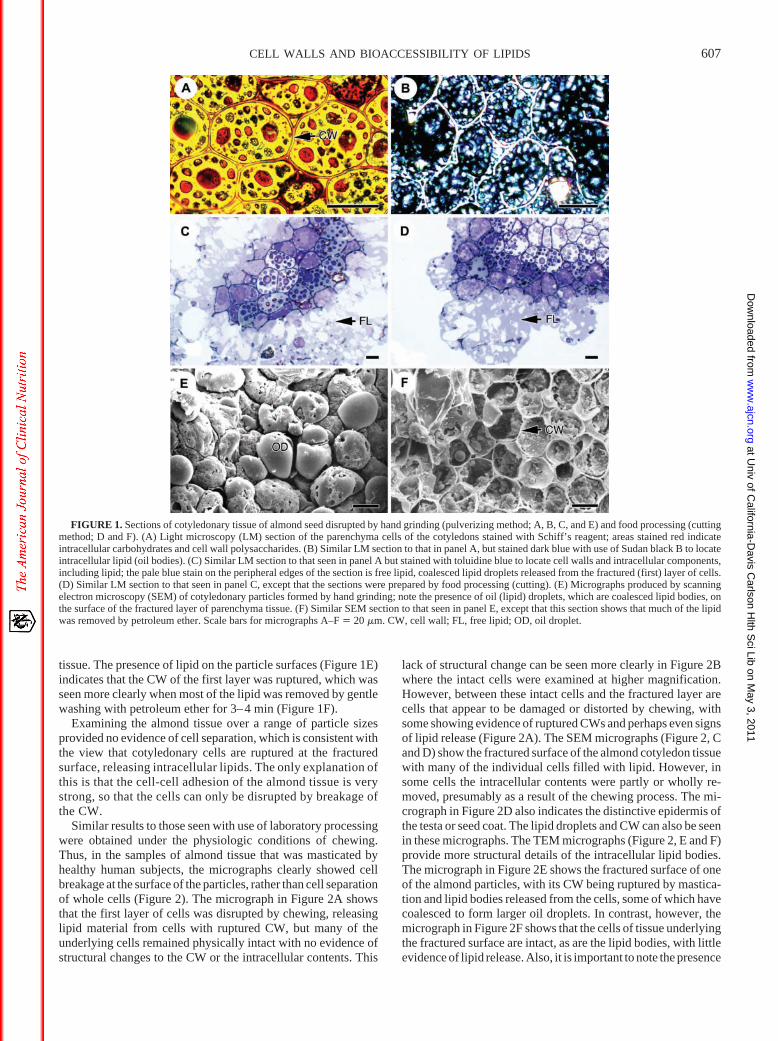
Similar results to those seen with use of laboratory processingwere obtained under the physiologic conditions of chewing.Thus, in the samples of almond tissue that was masticated byhealthy human subjects, the micrographs clearly showed cellbreakage at the surface of the particles, rather than cell separationof whole cells (Figure 2). The micrograph in Figure 2A showsthat the first layer of cells was disrupted by chewing, releasinglipid material from cells with ruptured CW, but many of theunderlying cells remained physically intact with no evidence ofstructural changes to the CW or the intracellular contents. This
lack of structural change can be seen more clearly in Figure 2Bwhere the intact cells were examined at higher magnification.However, between these intact cells and the fractured layer arecells that appear to be damaged or distorted by chewing, withsome showing evidence of ruptured CWs and perhaps even signsof lipid release (Figure 2A). The SEM micrographs (Figure 2, Cand D) show the fractured surface of the almond cotyledon tissuewith many of the individual cells filled with lipid. However, insome cells the intracellular contents were partly or wholly re-moved, presumably as a result of the chewing process. The mi-crograph in Figure 2D also indicates the distinctive epidermis ofthe testa or seed coat. The lipid droplets and CW can also be seenin these micrographs. The TEM micrographs (Figure 2, E and F)provide more structural details of the intracellular lipid bodies.The micrograph in Figure 2E shows the fractured surface of oneof the almond particles, with its CW being ruptured by mastica-tion and lipid bodies released from the cells, some of which havecoalesced to form larger oil droplets. In contrast, however, themicrograph in Figure 2F shows that the cells of tissue underlyingthe fractured surface are intact, as are the lipid bodies, with littleevidence of lipid release. Also, it is important to note the presence
FIGURE 1. Sections of cotyledonary tissue of almond seed disrupted by hand grinding (pulverizing method; A, B, C, and E) and food processing (cuttingmethod; D and F). (A) Light microscopy (LM) section of the parenchyma cells of the cotyledons stained with Schiff’s reagent; areas stained red indicateintracellular carbohydrates and cell wall polysaccharides. (B) Similar LM section to that in panel A, but stained dark blue with use of Sudan black B to locateintracellular lipid (oil bodies). (C) Similar LM section to that seen in panel A but stained with toluidine blue to locate cell walls and intracellular components,including lipid; the pale blue stain on the peripheral edges of the section is free lipid, coalesced lipid droplets released from the fractured (first) layer of cells.(D) Similar LM section to that seen in panel C, except that the sections were prepared by food processing (cutting). (E) Micrographs produced by scanningelectron microscopy (SEM) of cotyledonary particles formed by hand grinding; note the presence of oil (lipid) droplets, which are coalesced lipid bodies, onthe surface of the fractured layer of parenchyma tissue. (F) Similar SEM section to that seen in panel E, except that this section shows that much of the lipidwas removed by petroleum ether. Scale bars for micrographs A–F � 20 �m. CW, cell wall; FL, free lipid; OD, oil droplet.
CELL WALLS AND BIOACCESSIBILITY OF LIPIDS 607
at Univ of C
alifornia-Davis C
arlson Hlth S
ci Lib on May 3, 2011
ww
w.ajcn.org
Dow
nloaded from
of a thickened middle lamella at the cell junctions, indicative ofa high concentration of pectic material (Table 1), which is likelyto play an important role in maintaining cell-cell adhesion.
In all the microscopic sections examined, no differences inmicrostructure were observed between raw and roasted almondseed samples, whether in an intact or a physically disrupted state.
Digestibility of almond seeds in healthy human subjects
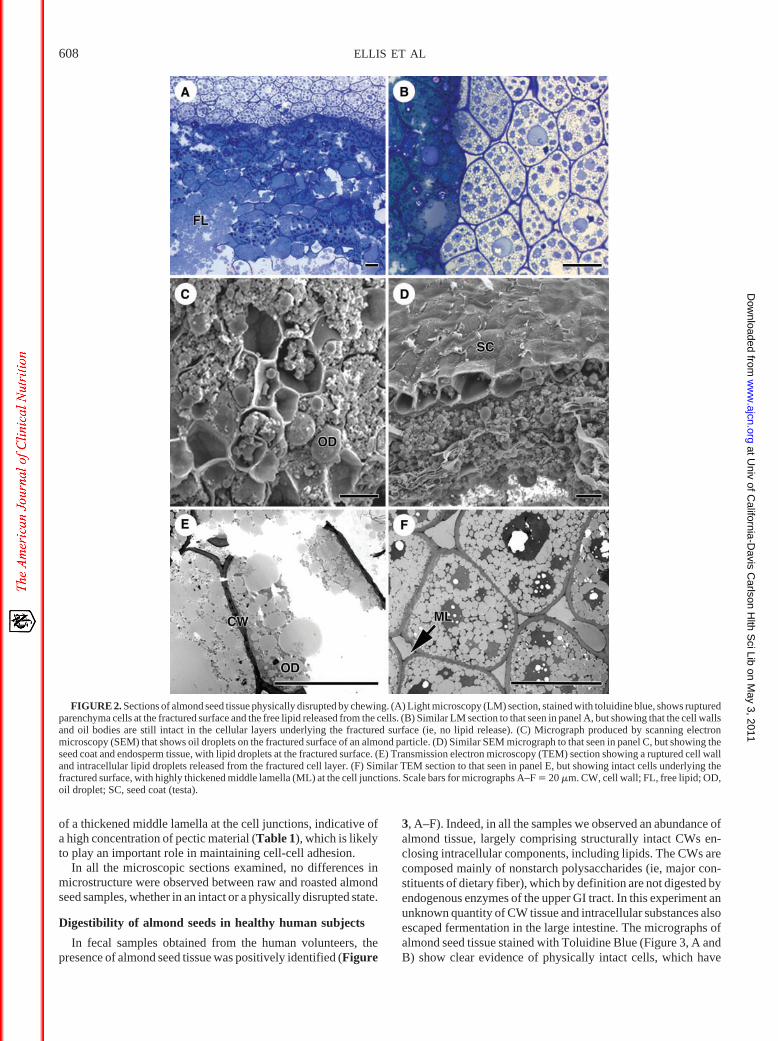
In fecal samples obtained from the human volunteers, thepresence of almond seed tissue was positively identified (Figure
3, A–F). Indeed, in all the samples we observed an abundance ofalmond tissue, largely comprising structurally intact CWs en-closing intracellular components, including lipids. The CWs arecomposed mainly of nonstarch polysaccharides (ie, major con-stituents of dietary fiber), which by definition are not digested byendogenous enzymes of the upper GI tract. In this experiment anunknown quantity of CW tissue and intracellular substances alsoescaped fermentation in the large intestine. The micrographs ofalmond seed tissue stained with Toluidine Blue (Figure 3, A andB) show clear evidence of physically intact cells, which have
FIGURE 2. Sections of almond seed tissue physically disrupted by chewing. (A) Light microscopy (LM) section, stained with toluidine blue, shows rupturedparenchyma cells at the fractured surface and the free lipid released from the cells. (B) Similar LM section to that seen in panel A, but showing that the cell wallsand oil bodies are still intact in the cellular layers underlying the fractured surface (ie, no lipid release). (C) Micrograph produced by scanning electronmicroscopy (SEM) that shows oil droplets on the fractured surface of an almond particle. (D) Similar SEM micrograph to that seen in panel C, but showing theseed coat and endosperm tissue, with lipid droplets at the fractured surface. (E) Transmission electron microscopy (TEM) section showing a ruptured cell walland intracellular lipid droplets released from the fractured cell layer. (F) Similar TEM section to that seen in panel E, but showing intact cells underlying thefractured surface, with highly thickened middle lamella (ML) at the cell junctions. Scale bars for micrographs A–F � 20 �m. CW, cell wall; FL, free lipid; OD,oil droplet; SC, seed coat (testa).
608 ELLIS ET AL
at Univ of C
alifornia-Davis C
arlson Hlth S
ci Lib on May 3, 2011
ww
w.ajcn.org
Dow
nloaded from

survived degradation by digestive secretions and microbial en-zymes. The stain also picked up the presence of intestinal bac-teria, which appear to be using the CWs and also the intracellularnutrients, including lipid. Interestingly, the slightly darker blueand more uniform staining on the periphery of the almond sectionreveals cotyledonary cells that are less packed together. Indeed,there is evidence of cell separation, caused possibly by bacterialdegradation of the middle lamella, which is likely to consistmainly of pectic material [described in Results section underNonstarch polysaccharides (dietary fiber) and phenolic compo-nents of almond seed cell wall material]. Further evidence of lossof the middle lamella and cell separation can be seen in the TEMmicrographs (Figure 3, E and F).
More detailed pictures of microbial action on specific struc-tures of the almond seed can be seen in the SEM micrographs. InFigure 3C, the relatively smooth surface of the seed coat or testa,which contains pigments and phenolic compounds, shows littleevidence of bacterial degradation. Thus, no obvious structuralchanges were observed compared with structural changes seen inmicrographs of predigested testa (Figure 2D). Also, there waslittle evidence of bacteria growing on the surface, because thenumber of bacteria observed in this location appeared to be verylow. In contrast, large numbers of bacteria were detected on theCW surfaces of cotyledonary tissue. It appears that these bacteriacould well be active, given that they co-locate with partiallydigested (fermented) CW polysaccharides (Figure 3D). In thiscase, microbial degradation created numerous holes in the CW,thus allowing microflora to enter contiguous cells and then useother potential substrates, notably the intracellular lipid. TheTEM micrographs (Figure 3, E and F) can provide some insightinto the behavior of these bacteria, in that in cells in which theCWs were largely but not completely intact, much of the intra-cellular contents appeared to be used by bacteria. In some cells,all the cellular contents had virtually disappeared with only theCWs surrounding the bacteria.
Concentrations of fecal lipids in human subjects oncontrol and almond-rich diets
All results in this section are presented as mean (� SE) values.In the digestibility study, the total wet weight of fecal samplescollected from human subjects on the control and almond-rich(test) diets was 70 � 20 and 196 � 126 g, respectively (NS). Themoisture content of fecal samples from subjects on the controland almond diets was 75.2 � 3.1 and 72.7 � 2.4%, respectively(NS). Analysis of the fecal samples showed a marked increase inlipid concentration in subjects on the almond-rich diet. Thus, thetotal lipid concentrations of fecal samples (wet weight basis)from subjects on the control and almond-rich diets were 3.5 �
0.9% and 9.9 � 1.1%, respectively (P � 0.015). On the basis ofthe total fecal weight values, the actual weight of lipid in thecontrol feces was 2.8 � 1.5 g, and in the test feces it was 21.4 �14.4 g.
Nonstarch polysaccharides (dietary fiber) and phenoliccomponents of almond seed cell wall material
The concentrations of monosaccharide residues of the hydro-lyzed CWM of seed cotyledon and testa were determined withuse of gas-liquid chromatography. The predominant sugarsfound in the raw and roasted almond cotyledon were arabinose,uronic acids, glucose, xylose, galactose, rhamnose, fucose, andmannose, and in the skin they were uronic acid, arabinose, glu-cose, xylose, galactose, rhamnose, mannose, and fucose (con-centrations shown in Table 1). The arabinose, rhamnose, anduronic acid contents represent more than 60% of the total sugarcontent, indicating the presence of one or more arabinose-richpolysaccharides, including pectic material. The presence of thelatter is consistent with the relatively thick middle lamella at CWjunctions, as seen in Figures 2F and 3F. The relatively highconcentration of glucose found in the CWM suggests the pres-ence of either cellulose or perhaps even a mixed linkage�-glucan. Another possibility is that the glucose could also formthe cellulosic backbone of a xyloglucan, because xylose andgalactose residues were also identified in the CW preparations.The ratio of glucose, xylose, and galactose (3.19:1.00:0.65) issimilar to ratios reported for the xyloglucan group of CW storagepolysaccharides (29, 30). The sugar concentrations found in thehydrolyzed CWM extracted from the roasted and raw almondsamples were similar.
Several phenolic compounds, mainly protocatechuic acid,p-hydroxybenzoic acid, and vanillic acid, were identified in theCWM of almond skin, but not in the cotyledonary tissue in whichonly trace amounts were recorded. The mean concentrations(duplicate values) of the main phenolic components protocate-chuic acid, p-hydroxybenzoic acid, and vanillic acid were 0.64,0.46, and 0.10 �g/mg, respectively. A positive stain with use ofphloroglucinol-HCl was observed for lignin in the same skinsamples.
DISCUSSION
This paper provides an explanation of why in one study sig-nificant amounts of lipid were excreted in feces obtained fromhealthy human subjects who consumed an almond-rich diet (20).Microstructural examination of almond tissue showed that theCWs remained intact, except for the fractured surface and someof the underlying cells, even after laboratory processing and
TABLE 1Concentration of sugar (monosaccharide) residues (dry weight) in the hydrolyzed cell wall material of raw and roasted almond seeds and almond seed testa(skin)1
Rhamnose Fucose Arabinose Xylose Mannose Galactose Glucose Uronic acids Total
�g/mgRaw almond 23.6 10.2 259.2 46.2 7.0 30.1 147.6 201.4 725.3Roasted almond 31.6 13.9 335.1 64.0 9.4 40.2 174.6 205.7 874.5Raw almond skin 22.8 8.0 143.1 25.0 10.3 26.0 157.7 199.7 592.6Roasted almond skin 19.5 4.3 137.7 23.7 9.3 25.0 159.8 215.5 594.8
1 Values for the total and each sugar concentration are the means of 4 replicates; the CV of the replicates was �3%.
CELL WALLS AND BIOACCESSIBILITY OF LIPIDS 609
at Univ of C
alifornia-Davis C
arlson Hlth S
ci Lib on May 3, 2011
ww
w.ajcn.org
Dow
nloaded from
digestion in the GI tract. Thus, in the digestibility study, intactalmond seed cotyledon and testa (seed coat) were identified infecal samples obtained from subjects on the almond-rich diet.Moreover, intracellular lipid bodies that were observed in theoriginal almonds were also positively identified in the same fecalsamples. These results substantiate our original hypothesis thatan increase in fecal fat excretion in subjects on a diet containing
almonds is attributed to intact CWs hindering the digestion ofintracellular lipid.
The lipid fraction of almonds constitutes the largest proportionof available energy in the seed. In people consuming a dietcontaining almonds, any fat that escapes digestion in the smallintestine could represent an appreciable loss of energy. However,a proportion of this energy is likely to be salvaged in the large
FIGURE 3. Sections of almond tissues found in feces collected from healthy subjects. Panels A and B are light microscopy (LM) micrographs, which werestained with toluidine blue. (A) Shows intact almond tissue containing intracellular lipid and other nutrients and almond tissue surrounded by fecal bacteria.(B) Similar LM section to that seen in panel A, except showing the almond tissue at higher magnification and some evidence of cell separation of the outsidelayers of the tissue. In some parts of the tissue, it appears that the cell walls are ruptured, releasing lipid, and bacteria are located inside the cells. (C) Scanningelectron microscopy (SEM) micrograph showing the epidermis of the seed coat, largely undamaged by endogenous secretions of the gut and by bacterialfermentation; few bacteria appear to be growing on the epidermal surface. (D) Similar SEM section to that seen in panel C, but showing the surface of theparenchyma cells, with bacterial fermentation appearing to erode the cell walls of fractured and contiguous cells. Numerous bacteria are seen to be growingon the cell wall surface. (E) TEM section showing fecal bacteria that have digested the cotyledonary cell walls and gained access to the nutrients inside the cell.(F) Similar TEM section to that seen in panel E, except that in one cotyledonary cell it appears that the bacteria have used all the intracellular nutrients, includingthe lipids. It also appears that bacterial enzymes have eroded the middle lamella, which has contributed to cell separation. Scale bars for micrographs A–F �20 �m. BT, bacteria; CW, cell wall; SC, seed coat (testa).
610 ELLIS ET AL
at Univ of C
alifornia-Davis C
arlson Hlth S
ci Lib on May 3, 2011
ww
w.ajcn.org
Dow
nloaded from

intestine, where lipid that is more accessible after cell rupturecould be used by resident microflora. Nevertheless, in the presentstudy we observed a marked increase in fecal fat excretion inhealthy subjects eating almonds relative to the control diet. Thisresult is consistent with data from Zemaitis and Sabaté (20) andHaddard and Sabaté (21), who reported increases in fecal fatexcretion in healthy subjects who consumed diets enriched withalmonds or pecans. Because almonds and other tree nuts areconsidered to be high-fat foods and concern was expressed thathigh-fat diets could cause weight gain (31, 32), these results areof particular interest to health professionals concerned with obe-sity management. The importance of the health implications isunderlined by the results of a recent study in which overweightand obese subjects, consuming a formula-based low-energy diet,showed greater weight loss on a diet enriched with almondsrather than complex carbohydrates (33).
After physical disruption by in vitro methods or by chewing,the cotyledonary cells of the almond seeds were seen to rupture,rather than separate, as seen in many of the micrographs. Theprocess by which almond tissue fractures, like all plant tissues,depends on CW strength and cell-cell adhesion (15). Thus, infruits and vegetables that are crisp or crunchy the process oftissue fracture occurs by cell rupture. However, during cookingand ripening of these plant foods a process of cell separationcan occur, which leads to tissue softening, and is attributedmainly to the weakening of pectic substances involved incell-cell adhesion (15, 34).
In the present study, the rupturing of CWs was manifest pre-dominantly in the outer layer of almond seed particles, reflectingthe fracture planes that were created during disruption. Thisrupturing resulted in the exposure and release of intracellularlipid bodies, which coalesced to form larger oil droplets (sizerange, �10–40 �m). We presume that lipid from ruptured cellsis much more accessible for emulsification and subsequent di-gestion by pancreatic lipase in the small intestine (35). The po-tential bioaccessibility of this lipid was demonstrated by the easewith which lipid could be solubilized and removed when thefractured surface of the almond tissue was gently washed withpetroleum ether. This ease of removal of lipid in the fracturedlayer contrasted with the retention of intracellular lipid in intactcotyledonary cells after solvent washing. Moreover, in the di-gestibility study, the fractured surfaces of almond tissue exam-ined in human feces were relatively free of coalesced lipid drop-lets. This finding suggests that lipid from ruptured cells wasbioaccessible and, therefore, digested and absorbed, perhapspartly facilitated by the action of microbial enzymes. In contrast,however, there was no evidence that intracellular lipid retainedby the CW barrier was degraded by lipases. In future studies, itwill be necessary to quantify the extent and rate of release oflipids from the almond tissue and other lipid-bearing plant foods.The kinetics of lipid digestion and absorption is considered to beof importance in influencing postprandial lipemia, which canitself influence the atherosclerotic process of CAD (36, 37).
The encapsulation of intracellular lipids and other nutrients byCWs is not only likely to prevent their digestion in the upper GItract, but also, from the results of our digestibility study in hu-mans, it seems that a significant proportion of CWs and cellcontents survived the fermentation process in the large intestine.However, the fecal samples provided indirect, but likely, evi-dence of bacterial fermentation of the almond tissue, includingintracellular contents. This evidence is based on the presence of
bacteria on the surface of CWs that appear to be actively growingand severe erosion of the walls, exposing the inside of the cells(micrograph in Figure 3D). Furthermore, the micrographs pro-duced by TEM clearly show the presence of bacteria inside thecells, apparently “using” the intracellular contents.
These results raise interesting questions about the types ofbacteria that use substrates from the almond seed, the extent towhich these substrates are fermented, and the types of productsthat arise from this process (eg, short-chain fatty acids). Becausethe cells are particularly rich in lipid, it would be surprising ifbacteria did not exploit this carbon source for metabolic pur-poses. Moreover, it was known for some time that polyunsatu-rated fatty acids can influence the growth and mucus adhesion ofintestinal microflora and probiotic supplements (38–40). Al-though the triacylglycerols of almond lipids contain some poly-unsaturated fatty acids, mainly linoleic acid, the predominantfatty acid is oleic acid, a monounsaturated variety comprising�65% of the total oil fraction (41). As far as we are aware theeffects of almond seed lipids on bacterial growth and propertieshave not been investigated.
Evidence of microbial degradation of the almond seed CW isconsistent with the plethora of data showing that CW constitu-ents (ie, nonstarch polysaccharides) are fermented to a variabledegree in the large intestine (42). This evidence raises a furtherinteresting question about which polysaccharides are most sus-ceptible to fermentation. The partial susceptibility of pectic sub-stances to degradation by the gut microflora is borne out by theerosion of the middle lamella seen in the fecal samples, which canpartly explain the cell separation seen in the outer layers of theparticles of almond tissue. However, the effects of endogenoussecretions of the gut and physical mixing as a result of peristalsisare also likely to play a role in any cell separation process. OtherCW polysaccharides could also be susceptible to microbial deg-radation. However, further studies are required to establish thestructures and relative proportions of polysaccharides in the al-mond seed CW, which can then be more closely linked to theirsusceptibility to microbial fermentation.
The degradation of the CW can also be significantly influ-enced by the presence of noncarbohydrate compounds, such asphenolic components (43), which are known also to have strongantioxidant properties (44, 45). In the current study only traceamounts of phenolic compounds were found in the cotyledonarytissue. This finding indicates that the considerable quantity ofarabinose-containing polysaccharides lack significant phenolicsubstitution. However, significant concentrations of protocate-chuic acid, p-hydroxybenzoic acid, and vanillic acid were foundin the CW of the seed coat (skin). Also, (Klason) lignin waspositively identified in the same skin samples. Some of thesecompounds are likely to play an important structural role and,therefore, modify the mechanical properties of the CW (15, 34),which could, in turn, confer greater resistance to bacterial fer-mentation and lipid release. Some evidence of this resistancecomes from the microstructural examination of the seed coat,which appears to be largely intact in the fecal samples providedby the human subjects. Furthermore, the number of bacteriaobserved on the testa surface in the fecal samples appeared to berelatively low, certainly compared with that found on the surfaceof cotyledonary CWs. At this stage we are not able to quantifythese observations.
In conclusion, we have successfully identified almond seedtissue in fecal material collected from healthy subjects on an
CELL WALLS AND BIOACCESSIBILITY OF LIPIDS 611
at Univ of C
alifornia-Davis C
arlson Hlth S
ci Lib on May 3, 2011
ww
w.ajcn.org
Dow
nloaded from

almond-rich diet. The main structures of almond tissue werefound to be preserved after laboratory processing or even afterchewing and digestion. In particular, the CWs were intact andhindered the release of intracellular lipid. Thus, an increase infecal fat excretion after consumption of an almond-rich diet isprimarily attributed to impairment of lipid bioaccessibility, as aresult of an intact CW barrier.
We thank Karen Lapsley (Almond Board of California) for providing thealmond seeds and useful discussion of the results and manuscript. We thankour colleagues Mary Parker, Anthony Brain, and Battista Calvieri for assis-tance with microscopy analysis; Alan Howard for helping to produce themicrographs; Yu-Min Lee, George Koumbridis, and Augustine Marchie fortechnical assistance with digestibility study; and Kenneth Bruce for usefulinsights of bacterial fermentation.
All the authors participated in the experimental design, the collection andinterpretation of data, and the writing of the manuscript. Specifically, PREand CWCK were involved in the design and data interpretation of the studyoverall and took the main responsibility for writing the manuscript. YRperformed the laboratory processing and chewing experiments. DJAJ andCWCK were responsible for the digestibility part of the study. CP and KWWwere responsible for the cell wall analysis. JFP was responsible for devel-opment of techniques and preparation of samples for microscopy analysis.None of the authors had any financial or personal conflict of interest.
REFERENCES1. Kris-Etherton PM, Zhao G, Binkoski AE, Coval SM, Etherton TD. The
effect of nuts on coronary heart disease risk. Nutr Rev 2001;59:103–11.2. Hu FB, Stampfer MJ. Nut consumption and risk of coronary heart dis-
ease: a review of epidemiologic evidence. Curr Athersclerosis Reports1999;1:205–10.
3. Spiller GA, Jenkins DA, Bosello O, et al. Nuts and plasma lipids: analmond-based diet lowers LDL-C while preserving HDL-C. J Am CollNutr 1998;17:285–90.
4. Almario RU, Vonghavaravat V, Wong R, et al. Effects of walnut con-sumption on plasma fatty acids and lipoproteins in combined hyperlip-idemia. Am J Clin Nutr 2001;74:72–9.
5. Sabaté J, Haddad E, Tanzman JS, Jambazian P, Rajaram S. Serum lipidresponse to the graduated enrichment of a Step 1 diet with almonds: arandomized feeding trial. Am J Clin Nutr 2003;77:1379–84.
6. Hyson D, Schneeman PA, Davis PA. Almonds and almond oil havesimilar effects on plasma lipids and LDL oxidation in healthy men andwomen. J Nutr 2002;132:703–7.
7. Jenkins DJA, Kendall CWC, Marchie A, et al. Dose response of almondson coronary heart disease risk factors: blood lipids, oxidised low-densitylipoproteins, lipoprotein(a), homocysteine, and pulmonary nitric oxide.Circulation 2002;106:1327–32.
8. Stahl W, van den Berg H, Arthur J, et al. Bioavailability and metabolism.Mol Aspects Med 2002;23:39–100.
9. Granfeldt Y, Björck I, Drews A, Tovar J. An in-vitro procedure based onchewing to predict metabolic response to starch in cereal and legumeproducts. Eur J Clin Nutr 1992;46:649–60.
10. Ellis PR, Rayment P, Wang Q. Physico-chemical perspective of plantpolysaccharides in relation to glucose absorption, insulin secretion andthe entero-insular axis. Proc Nutr Soc 1996;55:881–98.
11. Hoebler C, Karinthi A, Devaux M-F, et al. Physical and chemical trans-formations of cereal food during oral digestion in human subjects. Br JNutr 1998;80:429–36.
12. van den Berg H. Carotenoids: factors affecting bioavailability. In:Pfannhuaser W, Fenwick GR, Khokhar S, eds. Biologically-active phy-tochemicals in food. Analysis, metabolism, bioavailability and function.Cambridge, United Kingdom: The Royal Society of Chemistry, 2001:407–15.
13. Slaughter SL, Ellis PR, Butterworth PJ. An investigation of the action ofporcine pancreatic �-amylase on native and gelatinised starches. Bio-chim Biophys Acta 2001;1525:29–36.
14. MacDougall AJ, Selvedran RR. Chemistry, architecture, and com-position of dietary fiber from plant cell walls. In: Cho SS, Dreher ML,eds. Handbook of dietary fiber. New York: Marcel Dekker Inc, 2001:281–319.
15. Waldron KW, Smith AC, Parr AJ, Ng A, Parker ML. New approaches tounderstanding and controlling cell separation in relation to fruit andvegetable texture. Trends Food Sci Technol 1997;8:213–21.
16. Tovar J, Defrancisco A, Björck I, Asp NG. Relationship between mi-crostructure and in vitro digestibility of starch in precooked leguminousseed flours. Food Structure 1991;10:19–26.
17. Noah L, Guillon F, Bouchet B, et al. Digestion of carbohydrate fromwhite beans (Phaseolus vulgaris L.) in healthy humans. J Nutr 1998;128:977–85.
18. Ren Y, Waldron KW, Pacy JF, Brain A, Ellis PR. Chemical and histo-chemical characterisation of cell wall polysaccharides in almond seedsin relation to lipid bioavailability. In: Pfannhauser W, Fenwick GR,Khokhar S, eds. Biologically-active phytochemicals in food. Cam-bridge, United Kingdom: The Royal Society of Chemistry, 2001:448 –52.
19. Levine AS, Silvis SE. Absorption of whole peanuts, peanut oil, andpeanut butter. N Engl J Med 1980;304:359–60.
20. Zemaitis J, Sabaté J. Effect of almond consumption on stool weight andstool fat. Fed Am Soc Exp Biol J 2001;15(4):A602 (abstr).
21. Haddard EH, Sabaté J. Effect of pecan consumption on stool fat. Fed AmSoc Exp Biol J 2001;14(4):A294 (abstr).
22. US Department of Agriculture, Agricultural Research Service. USDAnational nutrient database for standard reference, release 16, 2003. In-ternet: http://www.nal.usda.gov/fnic/foodcomp (accessed 30 October2003).
23. Lauer O. Grain size measurements on commercial powders. Ausberg,Germany: Alpine AG Ausberg, 1966.
24. Parker ML, Waldron KW. Texture of Chinese water chestnut: involve-ment of cell-wall phenolics. J Sci Food Agric 1995;68:337–46.
25. Blakeney AB, Harris PJ, Stone BAA. A simple and rapid preparation ofalditol acetates for monosaccharide analysis. Carbohydr Res 1983;113:291–9.
26. Blumenkrantz N, Asboe-Hansen G. New method for quantitative deter-mination of uronic acids. Anal Biochem 1973;54:484–9.
27. Waldron KW, Parr AJ, Ng A, Ralph J. Cell wall esterified phenolicdimers: identification and quantification by reverse phase high perfor-mance liquid chromatography and diode array detection. PhytochemicalAnalysis 1996;7:305–12.
28. Association of Official Analytical Chemists. AOAC Official Methods ofAnalysis. Washington, DC: AOAC, 1995.
29. Wang Q, Ellis PR, Ross-Murphy SB, Reid JSG. A new plant polysac-charide from a traditional Nigerian plant food: Detarium senegalenseGmelin. Carbohydr Res 1996;284:229–39.
30. Reid JSG. Cell wall storage carbohydrates in seeds. Biochemistry of theseed ‘gum’ and ‘hemicelluloses’. Adv Bot Res 1985;11:125–55.
31. Krauss RM, Eckel RH, Howard B, et al. AHA dietary guidelines: revi-sion 2000: a statement for healthcare professionals from the NutritionCommittee of the American Heart Association. Circulation 2000;102:2284–99.
32. Fraser GE. Nut consumption, lipids, and risk of a coronary event. ClinCardiol 1999;22:1–5.
33. Wien MA, Sabaté JM, Iklé DN, Cole SE, Kandeel FR. Almonds vscomplex carbohydrates in a weight reduction program. Int J Obes RelatMetab Disord 2003;27:1365–72.
34. Brett CT, Waldron KW. Physiology and biochemistry of plant cell walls.2nd ed. London: Chapman Hall, 1996.
35. Castro GA. Digestion and absorption. In: Johnson LR, ed. Gastrointes-tinal physiology, 4th ed. St Louis: Mosby Year Book, Inc, 1991:108–30.
36. Sethi S, Gibney MJ, Williams CM. Postprandial lipoprotein metabolism.Nutr Res Rev 1993;6:161–83.
37. Sanders TM, de Grassi T, Miller GJ, Morrissey JH. Influence of fattyacid chain length and cis/trans isomerization on postprandial lipemiaand factor VII in healthy subjects (postprandial lipids and factor VII).Atherosclerosis 2000;149:413–20.
38. Kodicek E, Worden AN. The effect of unsaturated fatty acids on Lac-tobacillus helveticus and other Gram-positive micro-organisms. Bio-chem J 1945;39:78–82.
39. Kabara JJ, Swieczkowski DM, Conley AJ, Truant JP. Fatty acids andderivatives as antimicrobial agents. Antimicrob Agents Chemother1972;2:23–38.
612 ELLIS ET AL
at Univ of C
alifornia-Davis C
arlson Hlth S
ci Lib on May 3, 2011
ww
w.ajcn.org
Dow
nloaded from

40. Kankaanpaa PA, Salminen SJ, Isolauri E, Lee YK. The influence ofpolyunsaturated fatty acids on probiotic growth and adhesion. FEMSMicrobiol Lett 2001;194:149–53.
41. Abdallah A, Ahumada MH, Gradziel TM. Oil content and fatty acidcomposition of almond kernals from different genotypes and Californiaproduction regions. J Am Soc Hort Sci 1998;123:1029–103.
42. Edwards CA, Rowland IR. Bacterial fermentation in the colon and itsmeasurement. In: Schweizer TF, Edwards CA, eds. Dietary fiber: acomponent of food. London: Springer, 1992:119–36.
43. Parker ML, Ng A, Smith AC, Waldron KW. Esterified phenolics of thecell walls of chufa (Cyperus esculentus L.) tubers and their role intexture. J Agric Food Chem 2000;48:6284–91.
44. Bors W, Heller W, Michel C, Stettmaier K. Flavanoids and polyphenols:chemistry and biology. In: Cadenas E, Packer L, eds. Handbook ofantioxidants. New York: Dekker, 1996:409–46.
45. Sang S, Lapsley K, Jeong W-S, Lachance PA, Ho C-T, Rosen RT.Antioxidive phenolic compounds isolated from almond skins (Prunusamygdalus Batsch). J Agric Food Chem 2002;50:2459–63.
CELL WALLS AND BIOACCESSIBILITY OF LIPIDS 613
at Univ of C
alifornia-Davis C
arlson Hlth S
ci Lib on May 3, 2011
ww
w.ajcn.org
Dow
nloaded from