Role of Capsid Structure and Membrane Protein Processing in Determining the Size and Copy Number of...
13
J. Mol. Biol. (1996) 260, 9–21 Role of Capsid Structure and Membrane Protein Processing in Determining the Size and Copy Number of Peptides Displayed on the Major Coat Protein of Filamentous Bacteriophage Pratap Malik, Tamsin D. Terry, Lalitha R. Gowda, Amagoia Langara Sergei A. Petukhov, Martyn F. Symmons, Liam C. Welsh Donald A. Marvin and Richard N. Perham* Filamentous bacteriophage virions can be engineered to display small Cambridge Centre for foreign peptides in the N-terminal regions of all 2700 copies of the major Molecular Recognition coat protein (pVIII), but larger peptides can be accommodated only in Department of Biochemistry hybrid virions, in which modified and wild-type coat protein subunits are University of Cambridge interspersed. The copy number of peptides accepted in hybrid virions is Tennis Court Road generally believed to be related to peptide size: the larger the insert, the Cambridge CB2 1QW, UK lower the number of modified coat protein subunits in the assembled virion. However, we show here that some large peptides can be displayed at a much higher copy number than smaller ones and that some relatively small peptides are poorly displayed, if at all, in hybrid virions. X-ray diffraction studies of a recombinant virion together with model building experiments with peptide and protein epitopes of known structure demonstrated that it is feasible to accommodate much larger structures, without perturbation of the capsid protein packing, than it has proved possible to generate in vivo . We show further that the insertion of certain peptides greatly slowed or even prevented the processing of the pVIII pro-coat by leader peptidase at the inner membrane of the Escherichia coli cell. A good correlation was found between the effect of the insert on the rate of the processing of the pro-coat, an essential step in virus assembly, and the number of the mature but modified proteins in the subsequently assembled hybrid virion. These results have important implications for the design of peptide display systems based on filamentous bacteriophage. 7 1996 Academic Press Limited Keywords: filamentous bacteriophage; epitope display; membrane protein translocation; leader peptidase; peptide libraries *Corresponding author Introduction The virion of the filamentous bacteriophage fd (f1 and M13 are very similar strains) is a flexible rod about 1 mm long and 6 nm in diameter. It comprises a tubular sheath of approximately 2700 copies of the major coat protein subunit (pVIII) surrounding a single-stranded, circular DNA core. There are a few copies (three to five) of each of four minor coat proteins at the ends of the virion: pVII and pIX at the end where assembly is initiated, pIII and pVI at the end where the process is terminated (for reviews, see Rasched & Oberer, 1986; Model & Russel, 1988; Russel, 1991). The major coat protein contains 50 amino acid residues and is largely a-helical; in the virion the pVIII subunits form a shingled, helical array, with the long axes of the a-helices making a small angle with the axis of the virus particle (Marvin, 1990; Glucksman et al ., 1992; Marvin et al ., 1994). The negatively charged N- terminal regions are exposed on the outer surface of the virion, whereas the C-terminal segments line Present addresses: L. R. Gowda, Department of Biochemistry & Nutrition, Central Food Technological Research Institute, Mysore 570013, Karnataka, India; A. Langara, Department of Virology, John Innes Centre, Colney Lane, Norwich NR4 7UH, UK; S. Petukhov, Institute of Cell & Molecular Biology, Darwin Building, Mayfield Road, University of Edinburgh, Edinburgh EH9 2JR, UK. 0022–2836/96/260009–13 $18.00/0 7 1996 Academic Press Limited
-
Upload
pratap-malik -
Category
Documents
-
view
213 -
download
0
Transcript of Role of Capsid Structure and Membrane Protein Processing in Determining the Size and Copy Number of...

J. Mol. Biol. (1996) 260, 9–21
Role of Capsid Structure and Membrane ProteinProcessing in Determining the Size and CopyNumber of Peptides Displayed on the Major CoatProtein of Filamentous Bacteriophage
Pratap Malik, Tamsin D. Terry, Lalitha R. Gowda, Amagoia LangaraSergei A. Petukhov, Martyn F. Symmons, Liam C. WelshDonald A. Marvin and Richard N. Perham*
Filamentous bacteriophage virions can be engineered to display smallCambridge Centre forforeign peptides in the N-terminal regions of all 2700 copies of the majorMolecular Recognitioncoat protein (pVIII), but larger peptides can be accommodated only inDepartment of Biochemistryhybrid virions, in which modified and wild-type coat protein subunits areUniversity of Cambridgeinterspersed. The copy number of peptides accepted in hybrid virions isTennis Court Roadgenerally believed to be related to peptide size: the larger the insert, theCambridge CB2 1QW, UKlower the number of modified coat protein subunits in the assembledvirion. However, we show here that some large peptides can be displayedat a much higher copy number than smaller ones and that some relativelysmall peptides are poorly displayed, if at all, in hybrid virions. X-raydiffraction studies of a recombinant virion together with model buildingexperiments with peptide and protein epitopes of known structuredemonstrated that it is feasible to accommodate much larger structures,without perturbation of the capsid protein packing, than it has provedpossible to generate in vivo. We show further that the insertion of certainpeptides greatly slowed or even prevented the processing of the pVIIIpro-coat by leader peptidase at the inner membrane of the Escherichia colicell. A good correlation was found between the effect of the insert on therate of the processing of the pro-coat, an essential step in virus assembly,and the number of the mature but modified proteins in the subsequentlyassembled hybrid virion. These results have important implications for thedesign of peptide display systems based on filamentous bacteriophage.
7 1996 Academic Press Limited
Keywords: filamentous bacteriophage; epitope display; membraneprotein translocation; leader peptidase; peptide libraries*Corresponding author
Introduction
The virion of the filamentous bacteriophage fd (f1and M13 are very similar strains) is a flexible rodabout 1 mm long and 6 nm in diameter. It comprisesa tubular sheath of approximately 2700 copies ofthe major coat protein subunit (pVIII) surrounding
a single-stranded, circular DNA core. There are afew copies (three to five) of each of four minor coatproteins at the ends of the virion: pVII and pIX atthe end where assembly is initiated, pIII and pVIat the end where the process is terminated (forreviews, see Rasched & Oberer, 1986; Model &Russel, 1988; Russel, 1991). The major coat proteincontains 50 amino acid residues and is largelya-helical; in the virion the pVIII subunits form ashingled, helical array, with the long axes of thea-helices making a small angle with the axis of thevirus particle (Marvin, 1990; Glucksman et al., 1992;Marvin et al., 1994). The negatively charged N-terminal regions are exposed on the outer surfaceof the virion, whereas the C-terminal segments line
Present addresses: L. R. Gowda, Department ofBiochemistry & Nutrition, Central Food TechnologicalResearch Institute, Mysore 570013, Karnataka, India; A.Langara, Department of Virology, John Innes Centre,Colney Lane, Norwich NR4 7UH, UK; S. Petukhov,Institute of Cell & Molecular Biology, DarwinBuilding, Mayfield Road, University of Edinburgh,Edinburgh EH9 2JR, UK.
0022–2836/96/260009–13 $18.00/0 7 1996 Academic Press Limited

Peptide Display on Filamentous Bacteriophage10
the cylindrical hole generated by the protein sheath,in which their positively charged side-chainsneutralize the negatively charged phosphodiesterlinks of the DNA (Symmons et al., 1995).
In recent years there has been much interest inmodifying the bacteriophage genome to allowdisplay of foreign peptides and proteins on thesurface of the virion as fusion coat proteins,particularly in the form of peptide libraries that canbe screened in various ways to identify novelbiologically active peptides or to test large numbersof mutants for loss or acquisition of biologicalactivity (reviewed by Smith & Scott, 1993; Hoess,1993; Cortese et al., 1994). Two proteins have beenemployed for this purpose: the minor coat protein,pIII, which provides for display of one copy (or atmost, five copies) of the polypeptide insert pervirion; and the major coat protein, pVIII, which canoffer multiple display. Peptides inserted near the Nterminus of pVIII are prominently exposed on thesurface of the virion and have been shown to elicita powerful and highly specific immune response(Greenwood et al., 1991; Willis et al., 1993; Meolaet al., 1995). Indeed, if the displayed peptide is aknown continuous epitope, it can also mimic in ahighly effective way the immunological propertiesof the protein from which it is drawn (Veroneseet al., 1994).
However, unlike the display of peptides on pIII,where there appears to be little restriction on thesize of the insert, only small peptides, up to five(Il’ichev et al., 1989) or about six to eight(Greenwood et al., 1991; Iannolo et al., 1995) aminoacid residues in length, can normally be incorpor-ated directly in pVIII to make a recombinant phagevirion in which all 2700 copies of pVIII aremodified. Larger peptides must be displayed inhybrid virions, in which modified coat proteinsdisplaying the foreign peptide are interspersedwith wild-type coat protein subunits (Greenwoodet al., 1991; Kang et al., 1991; Felici et al., 1991). Thecopy number of peptides accepted in hybrid virionsappears to be related inversely, at least in part, topeptide size. Thus, whereas some peptides of about10 to 15 amino acid residues may be represented inup to 30 to 40% (800 to 1000 copies) of the coatprotein subunits in a hybrid virion (Greenwoodet al., 1991; Veronese et al., 1994), only about ten
copies of bovine pancreatic trypsin inhibitor(Markland et al., 1991), a few copies of antibody Fabfragments (Kang et al., 1991) or a zinc finger domain(Bianchi et al., 1995) and less than one copy oftrypsin (Corey et al., 1993) are incorporated pervirion. The reasons for this are unclear.
We show here that there is no direct correlationbetween peptide size and the copy number ofpeptide display in hybrid virions. X-ray fibrediffraction analysis indicates that foreign peptidesare incorporated into the N-terminal region of pVIIIwithout any detectable effect on the structure of theviral capsid. Model building suggests that thepeptide inserts are able to adopt folded confor-mations that carry them significantly clear of theviral surface. Structural constraints are thereforeunlikely to limit the size of the peptide insert inpVIII. In contrast, we found that the inclusion ofcertain peptides in the N-terminal region of pVIIIcan greatly slow the processing of the pro-coat format the inner membrane of the Escherichia coli cell. Agood correlation was noted between the effect ofthe insert on the rate of the processing of thepro-coat, an essential step in virus assembly (Kuhnet al., 1986; Kuhn & Troschel, 1992; Russel, 1991),and the number of the mature but modifiedproteins in the subsequently assembled hybridvirion. These results have important implicationsfor the design of peptide display systems based onfilamentous bacteriophage and the composition ofpeptide libraries.
Results
Display of different peptide inserts on pVIII
A selection of widely differing peptides, whichare typical of continuous epitopes, was tested forthe frequency of display of the modified coatprotein in hybrid phage virions. Oligonucleotidesequences encoding these peptides were clonedinto plasmid pKfdH (Greenwood et al., 1991) orpfd8SHU (Malik & Perham, 1996). This placed thepeptide insert between amino acid residues 3 and4 of pVIII (V and N, in the fdH or pKfdH system,and G and N or D, in the pfd8SHU system). The
Table 1. Effects of peptide inserts in pVIII pro-coatInsert N-terminal sequencea Insert sizeb Per cent displayc Relative processing rate
fdH AEVNDP . . . + + + +A AEVGPGRAFNDP . . . 6 35 + + +B AEGALWGFFPVLDDP . . . 9 5–10 +C AEVIHIGPGRAFYTTNDP . . . 12 15 + +D AEVPKLRGDLQVLAQNDP . . . 12 40 + + + +E AEVIRIQRGPGRAFVTINDP . . . 14 5 + +F AEGPAGFAILKSNNKTFNYDDP . . . 16 25 + + +G AEGKQIIMMWQEVGKAMYADDP . . . 16 10 ND
ND, not determined.a The N-terminal sequence of the mature coat protein is shown, with the sequence of the peptide insert underlined.b The number of amino acid residues in the peptide insert.c The percentage of modified coat proteins incorporated into the hybrid virion.

Peptide Display on Filamentous Bacteriophage 11
Figure 1. Analysis of hybrid virions by SDS-PAGE. Theamino acid sequences of the peptide inserts are listed inTables 1 (Figure 1a) and 2 (Figure 1b). a, Lanes 1 to 7,peptides A to G, respectively. b, Lane 1, wild-type;lane 2, peptide N; lane 3, peptide NH; lane 4, peptide H;lane 5, BD; lane 6, as lane 5 but heavily overloaded.
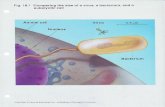
Figure 2. Model of bacteriophage fd displaying the34-residue structured region of the peripheral subunit-binding domain of the B. stearothermophilus pyruvatedehydrogenase complex as an insert between residues 3and 4 of pVIII. Inset, structure of the 44-residue bindingdomain, as determined by NMR spectroscopy. Thestructure is colour-coded red to violet from N toC terminus, respectively. The N-terminal sequenceVIAMPS is shown in red; the N-terminal helix is inorange; and the C-terminal sequence AGGA is in violet.The structure was generated using the programMOLSCRIPT (Kraulis, 1991). a, Side-view of a section ofthe bacteriophage virion, with fivefold rings of pVIIImolecules displaying the insert, depicted in red andgreen in alternate rings. b, As in a, but viewed incross-section and showing the central hole in the capsidin which the viral DNA is located. Structures a and bwere generated using the program GRASP (Nichollset al., 1991).
peptides ranged in size from six to 16 amino acidresidues (Table 1). Hybrid virions were purifiedfrom E. coli TG1 recO cells as described (Greenwoodet al., 1991), the wild-type coat protein beingderived from bacteriophage fd and the modifiedcoat protein from the plasmid.
The modified coat protein was detected bysubjecting the virions to SDS-PAGE (Figure 1a) andthe extent of incorporation was estimated from therelative yields of the two N-terminal sequences(modified protein and wild-type) obtained fromdirect analyses of the purified virions. There wasconsiderable variation in the level of display, from5 to 40% (Figure 1a and Table 1). For example,peptide F (16 residues), which was displayed atrelatively high level (25%), may be compared with,say, peptide B (nine residues) for which little (ca.5%) incorporation of the modified coat proteincould be detected. It is clear that the incorporationof the modified coat protein is not limited simplyby the size of the peptide displayed.
Display of a folded protein domain on thesurface of the virion
The peptides tested above are likely to be tooshort to assume a stable folded structure. Thesmallest known folded protein domain not stabil-ized by disulphide bridges or liganded metal ionsis the peripheral subunit-binding domain (BD) ofthe dihydrolipoyl acyltransferase (E2) polypeptidechain of 2-oxo acid dehydrogenase multienzymecomplexes (Perham, 1991). This domain from thepyruvate dehydrogenase complex of Bacillusstearothermophilus contains only 34 amino acidresidues, but adopts a stable folded structure(Figure 2, inset) consisting of two parallel a-helices
linked by a helical turn and loop region builtaround a small but well-defined hydrophobic core(Kalia et al., 1993). The amino acid sequence isshown in Table 2, flanked by short sequences of six(VIAMPS) and four (AGGA) amino acid residuesthat lie on the N and C-terminal sides, respectively.These are part of longer flanking sequences, moreextended and flexible, that act as linker regions forthe domain in the intact E2 chain (Kalia et al., 1993).
A plasmid, pKBD, expressing this domain wasused to construct hybrid virions, but no modifiedcoat protein could be detected in the bacteriophage

Peptide Display on Filamentous Bacteriophage12
Table 2. Amino acid sequences of BD and related peptidesInsert Plasmid N-terminal sequence
Insert pdf9BDNH VQGTGKNGRVLKEDIDAFLAGGAVNDP . . . .BD pKBD AEVVIAMPSVRKYAREKGVDIRL
VQGTGKNGRVLKEDIDAFLAGGAVNDP . . .N pfd8BDN AEGVIAMPSNDP . . .H pfd8BDH AEGSVRKYAREKGNDP . . .NH pfd8BDNH AEGVIAMPSVRKYAREKGNDP . . .
The N-terminal sequence of the mature coat protein is shown, with the sequenceof the peptide insert underlined. The additional V at the C terminus of the BDsequence insert in pKBD is a consequence of the cloning procedure used.
preparation when it was subjected to SDS-PAGE(Figure 1b, lane 5). It appears that for some reasonthis protein domain cannot be displayed in readilydetectable numbers on the surface of the filamen-tous bacteriophage virion. However, an additionalprotein (with an electrophoretic mobility corre-sponding to a molecular mass of ca. 10 kDa) wasdetected when the gels were substantially over-loaded (Figure 1b, lane 6). This band reactedpositively in a Western blot with rabbit antiserumdirected against the peripheral subunit-bindingdomain (data not shown). Larger amounts of thesame protein could be detected after SDS-PAGE ofwhole E. coli TG1 recO cells harbouring the plasmidpKBD (but not the parent plasmid pKK223-3),which had been induced by the addition ofisopropyl-b,D-thiogalactopyranoside (IPTG). Afterelectrotransfer to PVDF membrane, its N-terminalsequence was found to be MKKSLVVLKAS-, whichis the same as that of the signal peptide in thepro-coat form of pVIII (Kuhn et al., 1986). Thisstrongly suggests that a very small amount of theenlarged pro-coat was incorporated into thebacteriophage virion, similar to the levels of otherlarge proteins or domains displayed on pVIIIelsewhere (Markland et al., 1991; Kang et al., 1991;Corey et al., 1991; Bianchi et al., 1995) and the factthat it is displayed as a pro-coat suggests that theremay be a defect in the membrane processingprocess (see below).
X-ray fibre diffraction analysis of recombinantbacteriophage displaying a peptide epitope
The amino acid sequence GPGRAF can bedisplayed at high frequency on hybrid bacterio-phage virions (Figure 1, see above) and, using therelated bacteriophage fdH (Greenwood et al., 1991),it proved possible to make recombinant bacterio-phage virions in which all 2700 copies of pVIIIembody the epitope between residues 3 and 4 (datanot shown). There was no significant differencebetween the X-ray fibre diffraction patterns of thewild-type and recombinant virions (Figure 3),indicating that incorporation of the GPGRAFepitope caused no detectable change in the subunitstructure or helical parameters that define thecapsid. The lack of any significant change in theoverall distribution of diffraction intensity may atfirst seem surprising, since the protein subunit has
been increased in size by about 10%, but theadditional material is near the outer diameter of thevirion, where its position is likely to be poorlydefined, as found for the Pf1 strain of filamentousbacteriophage (Gonzalez et al., 1995). Furthermore,calculations of the X-ray scattering from perfectlyrigid models of wild-type virions, and thosedisplaying the GPGRAF epitope, show that thefibre diffraction patterns are relatively insensitive tochanges in the structure at the outer diameter of thevirion. The intensity changes seen in thesecalculations for rigid inserts are small, and it seemslikely that such changes will usually be below thenoise level of typical experimental fibre diffractiondata.
Model-building of folded structures on thesurface of the virion
To ascertain whether the capsid structureimposes any physical restriction on the display oflarger proteins or protein domains, we built twoexperimental models. In the first, using thecoordinates for the structures of the B. stearother-mophilus BD domain (Kalia et al., 1993) and theprotein capsid of bacteriophage fd (Marvin et al.,1994), we attempted to build the domain as aninsert into the N-terminal region of pVIII betweenresidues 3 and 4. The first four or five residues ofthe wild-type coat protein are conformationallyflexible in the virion, as judged by NMRspectroscopy (Colnago et al., 1987), which makes itunlikely that they contribute to any importantprotein-protein interaction in the virion.
As shown in Figure 2, the 34 amino acid residuesof the structured region of the BD domain in theirnative folded conformation can be incorporatedbetween residues 3 and 4 while maintaining theprotein-protein interactions beyond residue 5 thatdefine the helical viral capsid. Moreover, there isample room on the surface of the virion for all pVIIIsubunits to display the domain, consistent with aconsideration of peptide spacing of the surfacelattice of the closely related filamentous bacterio-phage M13 (Glucksman et al., 1992). Likewise, thereshould also be ample room to display a 26-residuezinc finger domain on every copy of pVIII, yet veryfew copies of the BD domain (Figure 1b) or the zincfinger domain (Bianchi et al., 1995) are incorporatedinto hybrid virions.

Peptide Display on Filamentous Bacteriophage 13
The amino acid sequence GPGRAF is derivedfrom the principal neutralizing determinant in thethird hypervariable region of the surface glyco-protein gp120 of HIV-1 (Javaherian et al., 1989). Thestructure of this epitope when bound to the Fabfragment of a monoclonal antibody directed againstit has been solved by means of X-ray crystallogra-phy (Ghiara et al., 1994). Using these coordinates,we attempted to build this much larger foldedstructure (molecular mass ca. 100 kDa) on thesurface of the recombinant GPGRAF virion, againwithout disturbing the protein-protein interactionsthat define the capsid. The Fab-epitope complexcould readily be assembled, as shown in Figure 4,although each complex would prevent the inter-action of another Fab molecule with the sixneighbouring epitopes. A Monte Carlo simulationof the surface packing indicates that, at saturation,up to 24% of the pVIII molecules in a hybrid virionwould be capable of displaying a comparably largeprotein, although it is unlikely that this level ofincorporation could be achieved in vivo, sinceperfect random packing could not be ensuredduring the assembly process. We conclude thatthere is no major impediment imposed by thestructure of the virus on the size of peptide/proteinthat might be displayed in hybrid virions. Given therestrictions that are found experimentally, it is clearthat other factors must be operating.
Effect of peptide inserts on membraneinsertion and processing of pVIII
The first event involving pVIII in the filamentousbacteriophage assembly process is the insertion ofthe pro-coat into the E. coli cell inner membrane andits processing by leader peptidase (Russel, 1991;Kuhn & Troschel, 1992); a defect at this stage couldobviously lower incorporation of a modified coatprotein into a hybrid virion. The rate of processingof pro-coats containing various peptide inserts inthe N-terminal region of the mature coat protein(Table 1) was therefore examined by means of apulse-chase technique (Russel & Model, 1981;Rowitch & Perham, 1987). Samples of E. colicells transformed with the relevant plasmid andpulse-labelled with [35S]methionine were submittedto SDS-PAGE at various time intervals to monitorthe conversion of pro-coat to mature pVIII.
In the cells transformed with pKfdH, processingof the pro-coat takes place quickly (Figure 5a).There were no radioactive bands in the samepositions in the gel for cells that had not beeninduced with IPTG, nor for cells transformed withthe parent plasmid pKK223-3 (data not shown). Thepeptide inserts were found to have widely differingeffects on the processing of the pro-coat expressedfrom the relevant plasmid, as judged from the rateof disappearance of pro-coat and the appearance ofthe mature coat protein (Figure 5a, A to F). In A, theloss of the pro-coat is hard to see against thebackground of the endogenous proteins, butmature coat protein appears very quickly, as
Figure 3. Diffraction patterns of bacteriophage fd anda recombinant virion displaying the GPGRAF epitope. a,Wide-angle fibre diffraction pattern of the Y21M mutantof fd. b, Wide-angle fibre diffracion pattern of arecombinant fdY21M displaying the GPGRAF epitope.The Y21M mutant was used to avoid layer-line fanning(Marvin et al., 1994); apart from the amino acidreplacement at position 21, the structure of the mutant isindistinguishable from that of wild-type fd. Thediffraction patterns were first mapped from detector toreciprocal space and quadrant averaged, and thebackground scattering was then subtracted. The diffrac-tion patterns shown consist of an averaged reciprocalspace quadrant mirrored about both the equator andthe meridian. The outermost edge of a is at 0.33 A−1;and that of b is at 0.32 A−1. The tilt of the fibre is differentin the two diffraction patterns shown. c, Integratedlayer-line amplitude data extracted from the diffractionpatterns shown in a and b. Curves correspond to onequadrant of the fibre pattern, with the meridian verticaland the equator horizontal. Each curve represents onelayer-line. The layer-line index, l, is shown to the left ofeach layer-line. Horizontal scale divisions alongR = 2 sin u/l are at intervals of 0.025 A−1 from R = 0 atthe left-hand side of each curve. The data from thetwo patterns have been scaled together. Continuouslines, amplitudes for fdY21M; broken lines, amplitudesfor GPGRAF, fdY21M. The low-resolution data onlayer-lines 0 and 1 from fdY21M are from the diffracionpattern of a wet fibre; the remaining fdY21M data arefrom the dry fibre pattern shown in a. This causes asmall break in the data on layer-lines 0 and 1. All theGPGRAF fdY21M data are from the wet fibre patternshown in b. The correlation coefficient betweenthe fdY21M and GPGRAF fdY21M amplitudes (exclud-ing the low resolution data on layer-lines 0 and 1) is0.96.

Peptide Display on Filamentous Bacteriophage14
Figure 4. Model of bacteriophage fd with animmunoglobulin Fab fragment interacting with a pepideepitope displayed as an insert between residues 3 and 4of pVIII. The peptide epitope (GPGRAF) is depicted inred, the Fab fragment in green, and GPGRAF epitopesnot interacting with the Fab fragment in blue. A sectionof the virion about 100 A long is shown. a, Side view;b, the same structure rotated by 90° around the virionaxis; c, an end-on view. The structures were generatedusing the program GRASP (Nicholls et al., 1991).
(judged on a four-point scale) are listed in Table 1.It is clear that size of peptide insert (i.e. number ofamino acid residues) is not alone in determininghow much the rate of pro-coat processing is slowed.On the other hand, the correlation between theeffect of a peptide insert on pro-coat processing(Figure 5a) and the frequency of its display on ahybrid virion (Figure 1a) is strong (Table 1): theslower the processing, the lower the incorporationof the modified coat protein in the assembledvirion.
These experiments were repeated with BD andrelated peptides. Introduction of the full 44 aminoacid residues (Table 2) prevented the processing ofthe much larger pro-coat over the time-scale of theexperiment (Figure 5b). This is in keeping with theinability to display the domain at a substantial copynumber on a hybrid virion, and the detection ofvery small amounts of unprocessed pro-coat in thebacteriophage capsid (Figure 1b). This was notsimply a problem of the sequence specificity ofleader peptidase, since the insertion of a six-residuepeptide with the same amino acid sequence(VIAMPS) as the N-terminal region of BD (plasmidpfd8BDN, Table 2) permitted efficient processing ofthe modified pro-coat (Figure 5b). The samesix-residue sequence could be displayed at highfrequency in a hybrid phage virion (Figure 1b). Incontrast, insertion of the ten-residue sequenceSVRKYAREKG (plasmid pfd8BDH, Table 2),embodying the first a-helix in the BD-domain(Figure 2), abolished detectable pro-coat processing(Figure 5b), as did the insertion of the 15-resi-due sequence VIAMPSVRKYAREKG (plasmidpfd8BDNH, Table 2) comprising the N-terminalregion plus first a-helix of BD (Figure 5b). Neitherof these latter sequences yielded a hybrid bacterio-phage virion exhibiting a detectable amount of themodified coat protein on a normal silver-stained gel(Figure 1b).
Membrane protein insertion and leaderpeptidase cleavage
The lack of processing of the pro-coat of apVIII-fusion protein could be due to a failure toinsert into the E. coli cell membrane or of theinserted protein to undergo subsequent cleavage bythe periplasmic leader peptidase. To determine theintracellular location of the protein, cultures ofE. coli cells transformed with plasmids pKBD orpfd8BDH were induced with IPTG and then, twohours after induction, the cells were fractionated bythe NaOH technique of Russel & Model (1982). Thetwo fractions, membrane and cytoplasmic, weresubjected to SDS-PAGE and the products examinedby means of Western blotting with antisera directedagainst the virus coat protein or BD (Figure 6). Thebulk (at least 90%) of the pro-coat in each instancewas found associated with the membrane fraction(Figure 6), suggesting that both pro-coat proteinswere capable of membrane insertion.
indicated by lane 2, and rises to its final level earlyin the series of time-points. Hence we deem it tohave processed rapidly. Likewise, in B, thedisappearance of the pro-coat is hard to judge, butthe appearance of the mature coat protein can beseen to be very slow. In C to F, the relativeelectrophoretic mobilities of the pro-coat andmature coat make it easier to assess the rate ofprocessing, though it can be regarded only assemi-quantitative. The relative rates of processing

Peptide Display on Filamentous Bacteriophage 15
The membrane insertion of the pro-coat wastested by analysing its susceptibility to proteinasedigestion. Protoplasts of E. coli cells transformedwith pKBD or pKfdH were exposed to proteinaseK at 0°C. The treated protoplasts were subjected toSDS-PAGE and the products examined by means ofWestern blotting with antisera directed against thevirus coat protein or BD (Figure 7). The pVIII-BDpro-coat was very sensitive to proteinase K,strongly suggesting that it had been correctlyinserted into the inner cell membrane, thereby
making it accessible to the proteinase. A smallamount of the pVIII-BD pro-coat remained undi-gested, presumably because it was in the cytoplasmand thus protected from the proteinase. Asexpected, it was totally digested if the cells wereexposed to the detergent, 3-[(3-cholamidopropyl)-dimethylammonio]-1-propane sulphonate (Chaps),to solubilize the cell membrane before the pro-teinase K was added (Figure 7, lanes 2 and 4).Identical results were obtained for the fdH pro-coat,which is efficiently translocated and processed to
(a) (b)
Figure 5. Processing of modified pro-coat proteins in E. coli. Cultures of E. coli TG1 recO cells transformed withexpression plasmids for the modified pro-coats were induced with IPTG and, after 15 to 20 minutes, pulse-labelledfor ten seconds with L-[35S]methionine. An excess of unlabelled methionine was then added and samples werewithdrawn at measured times and analysed by SDS-PAGE. In each panel, lane 1 represents cells uninduced with IPTG;lanes 2 to 7 are samples taken from induced cells 10, 20, 30, 45, 60 and 180 seconds after the addition of the unlabelledmethionine. a, fdH, processing of fdH pro-coat with no insert; A to F, processing of pro-coats with peptides A to F,respectively, as inserts. b, Processing of pro-coats with peptides BD, N, H and NH, respectively, as inserts. The positionsof the pro-coat, p, and mature coat protein, c, are indicated. In some instances the pro-coat runs in the same positionas a highly expressed host protein. The pro-coat containing peptide B is expressed constitutively. The position of thewild-type fd coat protein is marked (fd).

Peptide Display on Filamentous Bacteriophage16
Figure 6. Cellular location of modified pro-coatproteins. Cultures of E. coli TG1 recO cells transformedwith expression plasmids encoding the modified pro-coats were induced with IPTG and, two hours afterinduction, 1 ml of cell culture was added to 1 ml of cold0.2 M NaOH. Samples were prepared as described inMaterials and Methods and subjected to SDS-PAGE.Immunodetection with rabbit antiserum directed againstwild-type coat protein. b, Immunodetection with rabbitantiserum directed against BD. L, whole-cell lysate; S,alkali-soluble (cytoplasmic) fraction; and M, membranefraction. Lanes 1 to 3, pfd8BDH; lanes 4 to 6, pKBD. Theposition of processed wild-type pVIII is indicated by thearrow.
Figure 7. Mapping the topology of the pro-coat formof pVIII-BD fusion protein in the cell membrane. Culturesof E. coli TG1 recO cells transformed with the plasmidspKBD and pKfdH were grown at 37°C and induced withIPTG. Two hours after induction, cells were harvestedand treated with lysozyme to release the protoplasts,which were then exposed to proteinase K in the presenceor absence of detergent Chaps before being subjected toSDS-PAGE. a, Immunodetecion of BD-fusion proteinwith rabbit antiserum directed against wild-type coatprotein; b, immunodetection of BD-fusion protein withrabbit antiserum directed against BD; c, immunodetec-tion of fdH protein with rabbit antiserum directed againstwild-type coat protein. Lane 1, cells not exposed tolysozyme or proteinase K; lane 2, protoplasts exposed toproteinase K; lane 3, protoplasts not exposed toproteinase K; lane 4, protoplasts exposed to detergentand proteinase K.
sustain virus assembly (Figure 5a). In this instance,all the protein appeared as mature coat protein andwas totally accessible to proteinase K withouttreatment of the protoplasts with detergent beforeexposure to the enzyme (Figure 7).
Discussion
The experiments described above make it clearthat the display of peptides on pVIII molecules inhybrid virions is not governed simply by peptidesize (Figure 1 and Table 1). Peptide A (six residues)was displayed at high frequency, whereas it provedimpossible to incorporate readily detectable levelsof the ten-residue sequence SVRKYAREKG or the44-residue BD. In contrast, peptide F (16 residues)was displayed at a substantially higher copynumber than peptide B (nine residues). X-ray fibrediffraction analysis of the recombinant virion(Figure 3) demonstrated that the hexapeptideGPGRAF can be accommodated in all 2700 pVIIImolecules without disturbing the protein-proteininteractions in the capsid. Similarly, no changewas detected in the helical symmetry of recombi-nant virions displaying five-residue peptides(Kishchenko et al., 1994), suggesting that this resultis likely to be general. Moreover, model-building
experiments demonstrated that it is possible toconstruct the 34-residue folded region of the BDdomain as part of the N-terminal region of each ofthe 2700 pVIII molecules in a recombinant virion(Figure 2), despite the inability to incorporate BDinto a hybrid virion except at very low levelsof what appeared to be the pro-coat version(Figure 1b). Even more significantly, it provedpossible to build a model of a virion exhibiting thecomplex formed between the GPGRAF epitope andan Fab fragment directed against it (Figure 4).Although the complex obscured the six neighbour-ing epitopes, it can safely be inferred that it shouldbe possible to display a large protein (molecularmass ca. 100 kDa) at a reasonable copy number (upto 24% at saturation) in a hybrid virion. We inferthat there is no structural impediment to thedisplay of large peptides and proteins on thesurface of the filamentous bacteriophage virion, atmuch higher levels than has proved possiblein vivo.
Based on the X-ray fibre diffraction analysis of arecombinant bacteriophage M13 displaying a

Peptide Display on Filamentous Bacteriophage 17
five-residue peptide, Kishchenko et al. (1994)proposed that the peptide is accommodated on thesurface of the virion in an extended conformationin a shallow groove about 17 to 20 A long betweenneighbouring a-helical pVIII subunits, andsuggested further that it is the length of the groovethat limits the length of peptide displayed onrecombinant virions to about six to eight aminoacid residues, akin to the groove between a-helicesthat accommodates peptide epitopes on the surfaceof the major histocompatibility complexes (Brownet al., 1993). However, the differences betweensuccessive diffraction patterns for the wild-type(Glucksman et al., 1992; Kishchenko et al., 1994)and recombinant bacteriophage M13 (Kishchenkoet al., 1994) used for the above analysis aresufficiently small as to be near the noise level,suggesting that such differences should be inter-preted with caution. Moreover, we doubt thatresidence in a groove can be obligatory, given thehigh frequency of display achieved for peptides aslong as 16 residues (e.g. peptide F, Table 1) onhybrid virions.
An extended conformation for a displayedpeptide lying in a groove is also not easilyreconciled with the properties of the recombinantvirion displaying the GPGRAF sequence. Thissequence adopts a double-turn conformation whenbound to the Fab fragment of an HIV-1 neutralizingantibody (Ghiara et al., 1994) and essentially thesame conformation has been determined by meansof NMR spectroscopy for the hexapeptide dis-played in the pVIII molecule contained in anSDS-micelle (unpublished results). The propensityof the GPGRAF sequence to adopt its naturaldouble-turn conformation would account for thefact that the recombinant virion reacts strongly withnatural antisera from HIV-infected patients in anELISA assay (unpublished results). For this tohappen, the full IgG molecule must make contactwith a significant number of the displayed peptideepitopes in a recognisable conformation, consistentwith our model-building studies based on the Fabfragment (Figure 4).
A better correlation (Table 1, Figure 5a and b)was found to exist between the copy number ofmodified coat proteins in a hybrid virion and theability of the relevant pro-coat to undergomembrane insertion and processing, the essentialfirst step in virion assembly. For the BD domainand the ten-residue sequence SVRKYAREKGrepresenting its first a-helical region, the pro-coatswere successfully inserted into the E. coli cell innermembrane (Figures 6 and 7). In these instances atleast, the lesion appears to lie in a poor subsequentprocessing of the modified pro-coat by leaderpeptidase. Insertion of the hexapeptide VIAMPS inthe pro-coat still permitted rapid processing,whereas insertion of the 15-residue peptideVIAMPSVRKYAREKG or the whole BD domain,both of which begin with this same N-terminalsequence (Table 2), strongly inhibited processing(Figure 5b). The lesion is thus not simply due to a
change in protein sequence in the immediatevicinity of the leader peptidase cleavage site.
Many studies of leader peptidase have high-lighted the importance of small amino acid residuesat positions −1 and −3, and a helix-breaking aminoacid (proline in bacteriophage fd pro-coat) atposition −6 from the cleavage site (Shen et al., 1991;Izard & Kendall, 1994). Rather less is known aboutwhat sequences are accepted on the C-terminalside, although a Glu to Leu mutation at position +2in the M13 pro-coat permits membrane insertionbut inhibits subsequent leader peptidase cleavage(Russel & Model, 1981). This region of the pro-coatis fairly tolerant to change, as indicated by thesuccessful membrane insertion and processing of somany different pro-coat molecules incorporatingpeptides for bacteriophage display (Greenwoodet al., 1991; Felici et al., 1991; Iannolo et al., 1995).Other mutations altering the predominantly nega-tively charged character of this region of pro-coathave been reported to sustain membrane insertionand processing, and to retain the sec-independenceof the process, although an accumulation ofpositively charged residues does slow the mem-brane translocation (Kuhn et al., 1990; Cao et al.,1995). Peters et al. (1994) have shown that severalpositively charged amino acids in a region adjacentto the leader peptidase cleavage site inhibit properinsertion of the minor coat protein, pIII, into theE. coli inner membrane, thereby blocking theassembly and extrusion of phage particles. Theydemonstrated that these defects in membraneinsertion can be corrected by prlA suppressors,taking this as strong evidence for the sec-dependentinsertion of pIII. However, pVIII translocation issec-independent and such inhibition is unlikely.Moreover, the pro-coat of pVIII with the peptideinsert KILKVIRKNIV, containing four positivelycharged amino acid residues, is efficiently pro-cessed and incorporated into a hybrid virion(unpublished results). Factors other than the netcharge of a peptide insert must be influencing theprocessing of the modified pVIII pro-coat.
There is some evidence that the conformation ofthe cleavage region may be important in renderingit susceptible to leader peptidase. For example, theactive site of leader peptidase may be in closeproximity to the periplasmic surface of the E. colicell inner membrane (Jain et al., 1994). Secondarystructure motifs, especially a b-turn in the leaderpeptide but perhaps also in the amino acidsequence that immediately follows the cleavagesite, may have an important part to play (Jain et al.,1994; Barkocy-Gallagher et al., 1994). The gener-ation of an unfavourable secondary structure by thepeptide insert in some instances, e.g. a propensityto form an a-helix by the sequence SVRKYAREKG,or simply the bulk of the insert in other instances,may therefore be responsible for lowering thesusceptibility to leader peptidase.
Any failure to supply the modified coat proteinat a rate sufficient to compete effectively with thewild-type coat protein in the assembly process at

Peptide Display on Filamentous Bacteriophage18
the inner cell membrane would inevitably lower itsincorporation into a hybrid virion. Of course, theremay be additional, and perhaps insuperable,impediments later in the assembly pathway. Forexample, the proposed channel formed of pIVmolecules through which the virion emerges fromthe infected E. coli cell may be of insufficientdiameter to allow egress of a greatly enlargedcapsid (Kazmierczak et al., 1994). Similarly, asproposed by Endemann & Model (1995), it may beimpossible to generate a recombinant viriondisplaying peptides of more than about six to eightamino acid residues because the modified pVIIImolecule cannot interact properly with pVII toinitiate the viral assembly process. The constructionof hybrid virions overcomes this potential problemby the provision of the wild-type pVIII to initiateassembly. Nonetheless, if there is a defect inpro-coat processing, no recombinant or hybridvirion can be generated unless this separateproblem can be surmounted. These findings have
important implications for the design of filamen-tous bacteriophage peptide display systems, for thecomposition of peptide libraries, and for themechanism of leader peptidase that merit furtherattention.
Materials and Methods
Materials
Bacteriological media were from Difco. E. coli strainTG1 recO (K12,D(lac-proAB), supE, thi, hsdD5, recO::Tn5Kanr/F'traD36, proA+B+, lacIq, lacZDM15) used in allexperiments was a gift from Dr P. Oliver (Department ofGenetics, University of Cambridge). The expressionvector pKK223-3 and all enzymes, unless otherwisestated, were from Pharmacia LKB Biotechnology. Rabbitantisera against wild-type bacteriophage fd (Greenwoodet al., 1991) and B. stearothermophilus BD (Hipps &Perham, 1992) have been described.
Recombinant DNA techniques
Recombinant DNA techniques were carried out asdescribed (Ausubel et al., 1995). Polymerase chainreactions (PCR) were performed with Pfu DNApolymerase (Strategene). DNA fragments were purifiedusing the WizardTM PCR (Promega) system. Thefidelity of amplified PCR fragments were established byDNA sequencing using the T7 SequencingTM kit(Pharmacia) after sub-cloning into the vector. Oligonucle-otides were synthesized on a Pharmacia Gene AssemblerPlusTM.
Construction of plasmids
Plasmid pKMN encoding gVIIIp molecules displayingthe insert represented as peptide C has been described(Veronese et al., 1994). The following olignucleotideinserts were cloned into the HpaI site of plasmid pKfdH(Greenwood et al., 1991), peptides A, D and E, or intoSacII + StuI-digested pfd8SHU (Malik & Perham, 1996),peptides B and H:
For peptide F, a SacII + HindIII-digested PCR fragmentfrom the reaction using oligonucleotide OLP24Sfor.(5' - GCCTCTTTCGCCGCGGAGGGTCCGGCTGGTTTC -GCTATCCTGAAATCCAACAACAA AACCTTCAAC-TACGACGATCCTGCAAAAGCGGCCTTTGACTCCCT-GC-3') and OLfdH3rev. (5'-CGTTAGTAAATGAAGCTT-CTGTATGAGGTT-3') and plasmid pfd8SHU as templatewas cloned into SacII + HindIII-digested pfd8SHU toobtain pfd8P24S.
To create pKBD, a 155 bp DNA fragment encoding theperipheral subunit-binding domain from B. stearother-mophilus was produced by PCR using oligonucleotidesOLBDPETfor. (5'-GTACGTGCATCACGTGATCGATGC-CGTCCGTGCG-3') and OLBDPETrev. (5'-GAATGCCG-TAGTTAACTGCGCCGCCGGCAAGGAAAGCGTC - 3')and pET11D2D (Hipps & Perham, 1992; and unpub-lished results) as the template. The PCR-amplified DNAwas digested with PmlI (New England Biolabs) andHpaI and cloned into the HpaI site of pKfdH to yieldpKBD.
To create pfd8BDN, the oligonucleotides OLBDNfor.(5'-GCTGTCTTTCGCCGCGGAGGGTGTCATCGCCAT -

Peptide Display on Filamentous Bacteriophage 19
GCCTTCGAACGATCCTGCAAAAGCGGCCTTTGAC-TCC-3') and OLfdH3rev. were used as primers, withpKBD as template. The DNA fragment from thePCR-amplification was digested with SacII and HindIIIand cloned into SacII + HindIII-digested pfd8SHU.
To construct pfd8BDNH, two PCR reactions wereperformed with pfd8BDN as the template and oligo-nucleotides OLfdRIfor. (5'-CGTTTTAGGTTGGTGAATT-CGTAGTGGCAT-3') and OLBDNHrev. (5'-ATCGTTGC-CTTTTTCGCGCGCATACTTGCGCACCGAAGGCTGG -CGATGACACCCTCCGC-3') as primers, and withpfd8BDH as template and oligonucleotides OLBDNHfor.(5' - GGAGGGTGTCATCGCCATGCCTTCGGTGCGCA -AGTATGCGCGCGAAAAAGGCAACGAT-3') and OLf-dH3rev. as primers. The complete fragment was thengenerated by a PCR reaction using these smallerfragments and the oligonucleotides OLfdR1for. andOLfdH3rev. This product was digested with EcoRI andHindIII and cloned into EcoRI + HindIII-digestedpfd8SHU.
Production and purification of hybrid virions
A single plaque of bacteriophage fd wasintroduced into 3 ml of a 1:100 dilution of anovernight culture in 2 × TY medium (Ausubel et al.,1995) of E. coli cells transformed with the relevantplasmid. This culture was grown at 37°C untilturbid and then used to inoculate 100 ml of freshmedium. When the culture had grown to an A600 of0.9 to 1.0, IPTG was added to a final concentrationof 1 mM. After 30 minutes, the cells were collectedby centrifugation and resuspended in 100 ml offresh 2 × TY medium containing 50 mg/ml ampi-cillin and 1 mM IPTG and grown overnight. Thebacteriophage virions were purified from theculture supernatant (Rowitch et al., 1988) andanalysed by means of SDS-PAGE as described(Greenwood et al., 1991).
X-ray fibre diffraction
Magnetically aligned fibres of bacteriophage fd wereprepared by drying the virions for 30 hours at 92%relative humidity in a 9 T magnetic field, essentially asdescribed (Nave et al., 1981). Diffraction patterns werecollected on Station 7.2 of the synchrotron at the CLRCDaresbury Laboratory, using a wavelength of 1.488 Aand diffraction data were calibrated and processed usingthe programs from the CCP13 suite (Symmons et al., 1995;and R. Denny, personal communication).
Model building
Molecular models of recombinant fd virions displayingthe GPGRAF and BD sequences were built from themodel of the diad symmetry form of the fd virion(Marvin et al., 1994; entry 1IFJ in the Brookhaven ProteinData Bank) using the graphics program O (Jones et al.,1991). The structure of the GPGRAF sequence was thatdetermined for the GPGRAF-Fab complex by X-raycrystallography (Ghiara et al., 1994; BPDB entry 1ACY);the structure of the BD was that determined fromtwo-dimensional solution NMR data (Kalia et al., 1993;BPDB entry 2PDD). The fd pVIII is essentially all a-helixexcept for the residues on the immediate N and
C-terminal sides of the insert; in all cases these residuesare in stereochemically permissible conformations. Thestereochemistry of the models was checked using theprogram PROCHECK (Laskowski et al., 1993) and the3D-1D profiles reported by Luthy et al. (1992).
Pulse-chase analysis
E. coli cells transformed with the relevant plasmid weregrown at 37°C in M9 minimal medium (Ausubel et al.,1995) containing 50 mg/ml ampicillin, 0.4% (w/v)glucose, 20 mg/ml thiamin and all amino acids (finalconcentration 1 mM) except methionine. Overnightcultures were diluted 1:30 in 3 ml of fresh medium,shaken for two hours at 37°C and induced with IPTG(2 mM final concentration). After 15 to 20 minutes, asample (200 ml) of the cells was labelled with 60 mCi ofL-[35S]methionine (Amersham International) for tenseconds and the chase was then initiated by the additionof unlabelled methionine to a final concentration of1.4 mg/ml (Russel & Model, 1981; Rowitch & Perham,1987). At various time intervals, samples were precipi-tated in ice-cold 5% (w/v) trichloroacetic acid andexamined by means of SDS-PAGE (Greenwood et al.,1991). Gels were dried and autoradiographed usingHyperfilm b-max (Amersham International).
Cell fractionation
E. coli cells harbouring the relevant plasmid weregrown and induced with IPTG as described above. Aftertwo hours, a sample (1 ml) of the culture was added to1 ml of 0.2 M NaOH on ice. The membrane-associatedproteins were separated from the cytoplasmic fraction bycentrifugation for 30 minutes at 40,000 g and 4°C (Russel& Model, 1982).
Preparation of E. coli protoplasts and proteinaseK treatment
Two hours after induction with IPTG, a sample (1 ml)of an E. coli cell culture was centrifuged and the cellswere resuspended in buffer (0.5 M sucrose, 30 mMTris-HCl (pH 8.0), containing 20 mg/ml lysozyme and1 mM EDTA) on ice for 20 minutes. The protoplasts weredivided into three samples: one sample was retained asa control; the other two samples were treated withproteinase K (75 mg /ml), one in the presence and one inthe absence of 0.2% Chaps detergent. All three sampleswere incubated at 0°C for 20 minutes, then at 25°C forfive minutes, at which time the protease inhibitor,phenylmethylsulphonyl fluoride (1 mM final concen-tration), was added and the cells were collected bycentrifugation.
Other techniques
After polyacrylamide gel electrophoresis, proteinswere transferred onto a nitrocellulose membrane andanalysed by immunoblotting (Greenwood et al., 1991).Selected protein bands were electroblotted on to a PVDFmembrane (Millipore) and their N-terminal sequencesdetermined with an Applied Biosystems 477A proteinsequencer (Packman et al., 1988). The N-terminalsequences of coat proteins in hybrid virions weredetermined similarly after applying the virions to PVDFmembrane.

Peptide Display on Filamentous Bacteriophage20
AcknowledgementsWe thank Dr L. C. Packman, Dr C. Hill and Mr M.
Weldon for protein sequence analysis and oligonucle-otide synthesis, Mr M. Grant for preparing the cloneexpressing peptide G, Mr A. Gleeson for help withmagnetic alignment at Daresbury, and Mr C. Fuller forskilled technical assistance. We are grateful to TheWellcome Trust, the Biotechnology and BiologicalSciences Research Council and The Leverhulme Trust forfinancial support and to Dr E. A. Marseglia forencouragement of L.C.W. L.R.G. was supported by agrant from The British Council, T.D.T. was the holder ofa Wellcome Prize Studentship and L.C.W. holds aResearch Studentship from the Engineering and PhysicalSciences Research Council.
ReferencesAusubel, F. M., Brent, R., Kingston, R. E., Moore, D. D.,
Seidman, J. G., Smith, J. A. & Struhl, K. (1995).Current Protocols in Molecular Biology, John Wiley &Sons Inc., New York.
Barkocy-Gallagher, G. A., Cannon, J. G. & Bassford, P. J.,Jr (1994). b-Turn formation in the processing regionis important for efficient maturation of Escherichiacoli maltose-binding protein by signal peptidase Iin vivo. J. Biol. Chem. 269, 13609–13613.
Bianchi, E., Folgori, A., Wallace, A., Nicotra, M., Acali, S.,Phalipon, A., Barbato, G., Bazzo, R., Cortese, R.,Felici, F. & Pessi, A. (1995). A conformationallyhomogeneous combinatorial peptide library. J. Mol.Biol. 247, 154–160.
Brown, J. H., Jardetzky, T. S., Gorga, J. C., Stern, L. J.,Urban, R. G., Strominger, J. L. & Wiley, D. C. (1993).Three-dimensional structure of the human class IIhistocompatibility antigen HLA-DRl. Nature, 364,33–39.
Cao, G., Kuhn, A. & Dalbey, R. E. (1995). Thetranslocation of negatively charged residues acrossthe membrane is driven by the electrochemicalpotential: evidence for an electrophoresis-like mem-brane transfer mechanism. EMBO J. 14, 866–875.
Colnago, L. A., Valentine, K. G. & Opella, S. J. (1987).Dynamics of fd coat protein in the bacteriophage.Biochemistry, 26, 847–854.
Corey, D. R., Shiau, A. K., Yang, Q., Janowski, B. A. &Craik, C. S. (1993). Trypsin display on the surface ofbacteriophage. Gene, 128, 129–134.
Cortese, R., Felici, F., Galfre, G., Luzzago, A., Monaci, P.& Nicosia, A. (1994). Epitope discovery usingpeptide libraries displayed on phage. Trends Biotech-nol. 12, 262–267.
Endemann, H. & Model, P. (1995). Location offilamentous phage minor coat proteins in phage andin infected cells. J. Mol. Biol. 250, 496–506.
Felici, F., Castagnoli, L., Musacchio, A., Jappelli, R. &Cesareni, G. (1991). Selection of antibody ligandsfrom a large library of oligopeptides expressed on amultivalent exposition vector. J. Mol. Biol. 222,301–310.
Ghiara, J. B., Stura, E. A., Stanfield, R. L., Profy, A. T. &Wilson, I. A. (1994). Crystal structure of the principalneutralization site of HIV-1. Science, 264, 82–85.
Glucksman, M. J., Bhattacharjee, S. & Makowski, L.(1992). Three-dimensional structure of a cloningvector. X-ray diffraction studies of filamentousbacteriophage M13 at 7 A resolution. J. Mol. Biol. 226,455–470.
Gonzalez, A., Nave, C. & Marvin, D. A. (1995). Pf1filamentous bacteriophage: refinement of a molecu-lar model by simulated annealing using 3.3 Aresolution X-ray fibre diffraction data. Acta Crystal-log. sect. D, 51, 792–804.
Greenwood, J., Willis, A. E. & Perham, R. N. (1991).Multiple display of foreign peptides on a filamen-tous bacteriophage. J. Mol. Biol. 220, 821–827.
Hipps, D. A. & Perham, R. N. (1992). Expression inEscherichia coli of a sub-gene encoding the lipoyl andperipheral subunit-binding domains of the dihy-drolipoamide acetyltransferase component of thepyruvate dehydrogenase complex of Bacillusstearothermophilus. Biochem. J. 283, 665–671.
Hoess, R. H. (1993). Phage display of peptides andprotein domains. Curr. Opin. Struct. Biol. 3, 572–579.
Iannolo, G., Minenkova, O., Petruzzelli, R. & Cesareni, G.(1995). Modifying filamentous phage capsid: limitsin the size of the major capsid protein. J. Mol. Biol.248, 835–844.
Il’ichev, A. A., Minenkova, O. O., Tat’kov, S. I.,Karpyshev, N. N., Eroshkin, A. M., Petrenko, V. A.& Sandakhchiev, L. S. (1989) Production of a viablevariant of phage M13 with incorporated foreignpeptide in the major envelope protein. Dokl. Akad.Nauk. SSSR, 307, 481–483.
Izard, J. W. & Kendall, D. A. (1994). Signal peptides:exquisitely designed transport promoters. Mol.Microbiol. 13, 765–773.
Jain, R. G., Rusch, S. L. & Kendall, D. A. (1994). Signalpeptide cleavage regions. Functional limits on lengthand topological implications. J. Biol. Chem. 269,16305–16310.
Javaherian, K., Langlois, A. J., McDanal, C., Ross, K. L.,Eckler, L. I., Jellis, C. L., Profy, A. T., Rusche, J. R.,Bolognesi, D. P., Putney, S. D. & Matthews, T. J.(1989). Principal neutralizing domain of the humanimmunodeficiency virus type 1 envelope protein.Proc. Natl Acad. Sci. USA, 86, 6768–6772.
Jones, T. A., Zou, J. Y., Cowan, S. W. & Kjeldgaard, M.(1991). Improved methods for building proteinmodels in electron density maps and the location oferrors in these models. Acta Crystallog. sect. A, 47,110–119.
Kalia, Y. N., Brocklehurst, S. M., Hipps, D. S., Appella, E.,Sakaguchi, K. & Perham, R. N. (1993). The highresolution structure of the peripheral subunit-bind-ing domain of dihydrolipoamide acetyltransferasefrom the pyruvate dehydrogenase multienzymecomplex of Bacillus stearothermophilus. J. Mol. Biol.230, 323–341.
Kang, A. S., Barbas, C. F., Janda, K. D., Benkovic, S. J. &Lerner, R. A. (1991). Linkage of recognition andreplication functions by assembling combinatorialantibody Fab libraries along phage surfaces. Proc.Natl Acad. Sci. USA, 88, 4363–4366.
Kazmierczak, B., Mielke, D. L., Russel, M. & Model, P.(1994). Filamentous phage pIV forms a multimer thatmediates phage export across the bacterial cellenvelope. J. Mol. Biol. 238, 187–198.
Kishchenko, G., Batliwala, H. & Makowski, L. (1994).Structure of a foreign peptide displayed on thesurface of bacteriophage M13. J. Mol. Biol. 241,208–213.
Kraulis, P. J. (1991). MOLSCRIPT—a program to produceboth detailed and schematic plots of proteinstructures. J. Appl. Crystallog. 24, 946–950.
Kuhn, A. & Troschel, D. (1992). Distinct steps in theinsertion pathway of bacteriophage coat proteins. In

Peptide Display on Filamentous Bacteriophage 21
Membrane Biogenesis and Protein Targeting (Neupert,W. & Lill, R., eds), pp. 33–47, Elsevier, Amsterdam.
Kuhn, A., Wickner, W. & Kreil, G. (1986). Thecytoplasmic carboxy terminus of M13 procoat isrequired for the membrane insertion of its centraldomain. Nature, 322, 335–339.
Kuhn, A., Zhu, H.-Y. & Dalbey, R. E. (1990). Efficienttranslocation of positively charged residues of M13procoat protein across the membrane excludeselelctrophoresis as the primary force for membraneinsertion. EMBO J. 9, 2385–2389.
Laskowski, R. A., MacArthur, M. W., Moss, D. S. &Thornton, J. M. (1993). PROCHECK—a program tocheck the stereochemical quality of protein struc-tures. J. Appl. Crystallog. 26, 283–291.
Luthy, R., Bowie, J. U. & Eisenberg, D. (1992). Assessmentof protein models with 3-dimensional profiles.Nature, 356, 83–85.
Malik, P. & Perham, R. N. (1996). New vectors for peptidedisplay on the surface of filamentous bacteriophage.Gene, 171, 49–51.
Markland, W., Roberts, B. L., Saxena, M. J., Guterman,S. K. & Ladner, R. C. (1991). Design, construction andfunction of a multicopy display vector using fusionsto the major coat protein of bacteriophage M13. Gene,109, 13–19.
Marvin, D. A. (1990). Model building studies of Inovirus:genetic variations on a geometric theme. Int. J. Biol.Macromol. 12, 125–138.
Marvin, D. A., Hale, R. D., Nave, C. & Helmer Citterich,M. (1994). Molecular models and structural compari-sons of native and mutant class I filamentousbacteriophages. J. Mol. Biol. 235, 260–286.
Meola, A., Delmastro, P., Monaci, P., Luzzago, A.,Nicosia, A., Felici, F., Cortese, R. & Galfe’, G. (1995).Derivation of vaccines from mimotopes. Immuno-logical properties of HBsAg mimotopes displayed onfilamentous phage. J. Immunol. 154, 3162–3172.
Model, P. & Russel, M. (1988). Filamentous bacterio-phage. In The Bacteriophages (Calendar, R., ed.), vol. 2,pp. 375–456, Plenum Press, New York.
Nave, C., Brown, R. S., Fowler, A. G., Ladner, J. E.,Marvin, D. A., Provencher, S. W., Tsugita, A.,Armstrong, J. & Perham, R. N. (1981). Pf1 filament-ous bacterial virus. X-ray fibre diffraction analysis of2 heavy-atom derivatives. J. Mol. Biol. 149, 675–707.
Nicholls, A., Sharp, K. A. & Honig, B. H. (1991). Proteinfolding and association–insights from the interfacialand thermodynamic properties of hydrocarbons.Proteins: Struct. Funct. Genet. 11, 281–296.
Packman, L. C., Borges, A. & Perham, R. N. (1988). Aminoacid sequence analysis of the lipoyl and peripheralsubunit-binding domains in the lipoate acetyltrans-ferase component of the pyruvate dehydrogenasecomplex from Bacillus stearothermophilus. Biochem. J.252, 79–86.
Perham, R. N. (1991). Domains, motifs and linkers in2-oxo acid dehydrogenase multienzyme complexes:a paradigm in the design of a multifunctionalprotein. Biochemistry, 30, 8501–8512.
Peters, E. A., Schatz, P. J., Johnson, S. S. & Dower, W. J.(1994). Membrane insertion defects caused bypositive charges in the early mature region of protenpIII of filamentous phage fd can be corrected by prlAsuppressors. J. Bacteriol. 176, 4296–4305.
Rasched, I. & Oberer, E. (1986). Ff coliphages: structuraland functional relationships. Microbiol. Rev. 50,401–427.
Rowitch, D. H. & Perham, R. N. (1987). Cloning andexpression of the filamentous bacteriophage Pf1major coat protein gene in Escherichia coli. Membraneprotein processing and virus assembly. J. Mol. Biol.195, 873–884.
Rowitch, D. H., Hunter, G. J. & Perham, R. N. (1988).Variable electrostatic interaction between DNA andcoat protein in filamentous bacteriophage assembly.J. Mol. Biol. 204, 663–674.
Russel, M. (1991). Filamentous phage assembly. Mol.Microbiol. 5, 1607–1613.
Russel, M. & Model, P. (1981). A mutation downstreamfrom the signal peptidase cleavage site affectscleavage but not membrane insertion of phagecoat protein. Proc. Natl Acad. Sci. USA, 78, 1717–1721.
Russel, M. & Model, P. (1982). Filamentous phagepre-coat is an integral membrane protein: analysis bya new method of membrane preparation. Cell, 28,177–184.
Shen, L. M., Lee, J.-I., Cheng, S., Jutte, H., Kuhn, A. &Dalbey, R. E. (1991). Use of site-directed mutagenesisto define the limits of sequence variation toleratedfor processing of the M13 procoat protein by theEscherichia coli leader peptidase. Biochemistry, 30,11775–11781.
Smith, G. P. & Scott, J. K. (1993). Libraries of peptides andproteins displayed on filamentous phage. MethodsEnzymol. 217, 228–257.
Symmons, M. F., Welsh, L. C., Nave, C., Marvin, D. A. &Perham, R. N. (1995). Matching electrostatic chargebetween DNA and coat protein in filamentousbacteriophage. Fibre diffraction of charge-deletionmutants. J. Mol. Biol. 245, 86–91.
Veronese, F. di M., Willis, A. E., Boyer-Thompson, C.,Appella, E. & Perham, R. N. (1994). Structuralmimicry and enhanced immunogenicity of peptideepitopes displayed on filamentous bacteriophage.The V3 loop of HIV-1 gp120. J. Mol. Biol. 243,167–172.
Willis, A. E., Perham, R. N. & Wraith, D. (1993).Immunological properties of foreign peptides inmultiple display on a filamentous bacteriophage.Gene, 128, 79–83.
Edited by J. Karn
(Received 2 February 1996; received in revised form 3 April 1996; accepted 12 April 1996)