The Role of Endotoxin during Typhoid Fever and Tularemia in Man ...

The Role of Endotoxin dunrng
Typhoid Fever and Tularemia in Man
IV. THE INTEGRITY OF THE ENDOTOXIN
TOLERANCEMECHANISMSDURING INFECTION
SHEIELDONE. GREisMAN, RiciARD B. HoRmIc, HENRYN. WAGNER,JR.,WILLIAM E. WOODWARD,and THEODOREE. WOODWARD
From the Departments of Medicine and Physiology, University of MarylandSchool of Medicine, and the Department of Medicine, The Johns HopkinsSchool of Medicine, Baltimore, Maryland 21201
A B S T R A C T Volunteers infected with Salmonella ty-phosa develop a remarkable hyperreactivity to the py-rogenic and subjective toxic activities of homologous(S. typhosa) and heterologous (Pseudomonas) endo-toxins. The present studies quantitate this augmentedreactivity and demonstrate by three differing approachesthat significant tolerance to these endotoxins can bereadily induced within the framework of the hyperreac-tive state. Thus, (a) tolerance induced before illnessby repeated daily intravenous injections of the endotoxinsremained demonstrable during overt illness, (b) dailyintravenous injections of the endotoxins begun duringovert illness evoked progressively increasing tolerance,and (c) continuous intravenous infusions of S. typhosaendotoxin during illness rapidly induced a pyrogenic re-fractory state. Despite unequivocal activation of the en-dotoxin tolerance mechanisms by any of the abovemethods, the febrile and toxic course of typhoid feverproceeded unabated. Similarly, in other volunteers withPasteurella tularensis infection, continuous intravenousinfusions of S. typhosa endotoxin evoked initial hyper-reactive febrile and subjective toxic responses followedby rapid appearance of a pyrogenic refractory state with-out modification of the underlying clinical illness. Theseobservations suggest that circulating endotoxin plays nomajor role in pathogenesis of the sustained fever andtoxemia during typhoid fever and tularemia in man.
This work was presented in part before the eightieth An-nual Meeting of the Association of American Physicians,May 1967.
Received for publication 31 May 1968 and in revised form16 December 1968.
The mechanisms responsible for the systemic hyper-reactivity to endotoxin during typhoid fever and tulare-mia were further investigated. Low grade endotoxemia,nonspecific effects of tissue injury, impaired ability of thereticuloendothelial system to clear circulating endotoxin,and production of cytophilic antibodies capable of sensi-tizing leukocytes to endotoxin did not appear responsi-ble. Inflammatory reactions to intradermal S. typhosaendotoxin increased significantly during typhoid fever.However, since no such dermal hyperreactivity devel-oped to Pseudomonas endotoxin during typhoid fevernor to S. typhosa endotoxin during tularemia, the sys-temic hyperreactivity to bacterial endotoxins duringtyphoid fever and tularemia could not presently be as-cribed to enhanced levels of acquired hypersensitivity.
INTRODUCTIONThe importance of endotoxemia in the production ofclinical illness by Gram-negative bacteria is unknown.It has long been recognized, however, that healthy manrapidly acquires tolerance to the pyrogenic and toxicactivities of intravenous injections of bacterial endo-toxins administered at 24-hr intervals (1). Based uponthese observations in healthy man, the inference hasbeen drawn that circulating endotoxin can play no majorrole in the pathogenesis of fever during sustained Gram-negative bacterial infections (2, 3). While this propo-sition appears reasonable, it rests upon two unprovenassumptions: (a) that the mechanisms responsible fortolerance to daily intravenous injections of endotoxinwill continue to operate effectively during the febrile
The Journal of Clinical Investigation Volume 48 1969 613

phase of infection, and (b) that tolerance to sustainedendotoxemia can also develop during the febrile phase.Recent studies in volunteers with induced typhoid feverand tularemia demonstrated the appearance of a re-markable hyperreactivity to single intravenous injectionsof endotoxins, both homologous and heterologous, duringovert illness. This hyperreactivity occurred regardlessof whether a high degree of tolerance was preinduced byrepeated daily intravenous injections of endotoxin be-fore infection, or whether only a single control intra-venous base line test was performed. It was emphasizedthat it was uncertain as to whether this hyperreactivityto endotoxin involved the inhibition of those mecha-nisms that participate in tolerance, or whether the toler-ance mechanisms remained functional and offered somepartial protection (4). The effect of sustained endo-toxin infusions during illness was not determined. Thepresent studies were designed to evaluate more criticallythe role of endotoxemia in the pathogenesis of the febrileand toxic course of typhoid fever and tularemia in manby quantitating the integrity of the endotoxin tolerancemechanisms during the febrile stage of illness, employingboth intermittent and sustained endotoxin administration.In addition, further studies were conducted on themechanisms underlying the remarkable hyperreactivityto endotoxin seen during these infections.
METHODSVolunteers for these studies were male inmates of the Mary-land House of Correction, Jessup, Md., whose ages rangedfrom 20 to 52 yr. Each volunteer was fully apprised of allaspects of the investigation and the attendant risk. They werefree to cease participation at any time. Complete medical sur-veys were performed to verify the fitness of each participant.The preparation and administration of the inocula of Sal-monella typhosa and Pasteurella tularensis and the clinicalcourse and treatment of the resulting illnesses have been out-lined previously (5, 6). All infected volunteers tested withendotoxin during the present studies were unvaccinatedcontrols who were given the infectious agents for the pri-mary purpose of evaluating the efficacy of existing vaccines.Critical appraisal of the clinical course of induced illnesswas carried out by daily evaluation and recording, by thesame internist, of all symptoms and signs of disease. Inaddition, levels of bacteremia were quantitated serially. Com-parisons of these detailed records, designed primarily toprovide a sensitive assessment of disease modification byvaccines, also permitted reliable assessment of disease modi-fication in the present volunteers treated with purified endo-toxins.
Three preparations of bacterial endotoxins were employed:a highly purified endotoxin from S. typhosa (0-282),' pre-viously described (7), Pseudomonas endotoxin2 preparedby tryptic digestion (8), and Escherichia coli endotoxin(0127B8)8 prepared by Boivin-type extraction (9). All
'Kindly supplied by Dr. Maurice Landy, National Insti-tutes of Health.
2 Kindly supplied as Piromen by Dr. T. A. Garrett, Ba~xterLaboratories, Inc., Morton Grove, Ill.
8 Supplied by Difco Laboratories, Detroit, Mich.
endotoxin preparations were suspended in sterile, pyrogen-free physiologic saline and stored in rubber-stoppered vaccinebottles at 40C. The E. coli endotoxin was heated at 100'Cfor 10 min before use. The preparations were bacteriologi-cally sterile, and the same bottle of diluted endotoxin wasalways employed throughout any given study.
Pyrogen assay in man. Volunteers were hospitalized, andall responses to endotoxin were assayed with the subj ectsconfined to bed and covered with a light blanket. Syringeswere either of the sterile, pyrogen-free, disposable type' orwere heated overnight in a dry air oven at 200'C to eliminateextraneous pyrogens. Each test was begun between 8 and9 a.m. Flexible thermocouples were inserted 6 inches intothe rectum and temperatures recorded by a telethermometer.'Temperatures were monitored immediately before and everyhalf-hour after the intravenous injection of endotoxin. Theresults were plotted on standard graph paper, and feverresponses quantitated by two methods: (a) the area underthe 7 hr fever curve, with the initial temperature as thebase line, was calculated by summing the number of enclosedsmall squares. This value, termed the fever index, representsa measure of the height and duration of the febrile response(10). The coordinates were plotted so that a fever index of100 reflected a 1VF rise in rectal temperature for a 1 hrduration; (b) the maximum rate of temperature rise wascalculated by dividing each temperature increment (0F)after endotoxin administration by the time (hours) requiredto attain this increment and selecting the highest quotient.The derived value reflects the steepest slope of the tempera-ture rise from the origin (Fig. 1 A, slope a). This value,termed the maximum pyrogenic response rate, was consist-ently obtainable within the initial 3 hr. In addition to thefebrile responses, subjective toxic reactions after endotoxinadministration were graded as follows: 1+ = mild head-ache, anorexia; 2+ =moderate headache, anorexia, chills;3+ = severe headache, shaking chills, myalgia; 4+ =ex-tremely severe headache, shaking chills, myalgias, vomiting.
Labeling and clearance assay of Pseudomonas-51Cr endo-toxin. Pseudomonas endotoxin was labeled with 5'CrCl3 orwith Na2'CrO4,' as described by Braude, Carey, Sutherland,and Zalesky (11). For use in man, the endotoxin was recon-stituted to 25 ,Ag/ml in pyrogen-free physiologic saline. Afterproof of bacteriologic sterility, 1-ml aliquots containing amaximum radioactivity of 150 juc were employed for eachstudy. A 20 gauge needle was inserted into the antecubitalvein of the arm opposite to that used for intravenous injec-tion of the radioactively labeled endotoxin. A control bloodsample was obtained and the needle left in situ for collectionof serial timed blood samples after the endotoxin injection.Approximately 5-ml blood samples were collected at eachtime period, placed into plastic test tubes, and then weighedon a precision balance. The mean weight of 10 similar emptytubes was subtracted to give the net weight of each bloodsample. Weighing error was under 5%. The samples werecounted in a 3 inch scintillation crystal, and sufficient countsaccumulated to produce less than 5% error. All counts wereconverted to net counts per minute per 5 g of blood. Theinitial blank sample count was also corrected to 5 g ofblood and this value subtracted from each postendotoxinsample. The counts per minute per 5 g of blood were plottedagainst time on 2-cycle semilog paper and the time for
4 Plastipak, supplied by Becton-Dickinson & Co., Ruther-ford, N. J.
5 Yellow Springs Instrument Company, Inc., YellowSprings, Ohio.
6 Supplied by E. R. Squibb & Sons, New Brunswick, N. J.
614 Greisman, Hornick, Wagner, Woodward, and Woodward
clearance of 50% of the endotoxin (ti) extrapolated em-ploying the 3 min value as the reference point.
Endotoxin clearance studies in rabbits were performed asabove except that the timed serial blood samples wereobtained by cardiac puncture with a 22 gauge needle afterinjection of the endotoxin into a marginal ear vein.
Since bacterial endotoxins are complex molecules, thetoxic moiety of which has not yet been precisely defined,only those labeled preparations which exhibited blood clear-ance rates that could be related to their pyrogenic activity inrabbits were employed for studies in man. Thus, endotoxinpreparations were considered acceptable only when rabbitassay demonstrated: (a) significant acceleration of bloodclearance after pyrogenic tolerance was established by sixdaily intravenous injections of the unlabeled endotoxin; (b)significant retardation of blood clearance after reticulo-endothelial "blockade" with 3 ml/kg of thorotrast; (c) noretardation of blood clearance after reticuloendothelial "block-ade" with a colloid that failed to increase febrile responsive-ness to the endotoxin (i.e., heat-aggregated human serumalbumin); (d) blood clearance rates appreciably slower thanthat of aggregated 'CrCls.7 Two batches of 'Cr-labeled
7This latter criterion was found essential when labelingwith 'CrCls because of spontaneous aggregation occurringin the pH 7 phosphate buffer; i.e., significant amounts of the5'CrC13 became nondialyzable during control labeling studiesin which endotoxin was omitted from the procedure. Whentested in four healthy volunteers, 25-,ug aliquots of suchaggregates of 'CrCls were cleared from the blood at veryrapid rates (tj = 2.9 ±0.25 (SE) min), and these rates werenot significantly accelerated after induction of endotoxintolerance (tj = 2.7 ±0.29 min). This spontaneous aggregationof 'CrC1l was only eliminated by keeping the reaction pHbetween 3 and 4 with acetate buffer, although this procedure
5
4
U. 3
10.z 2
I
4
Z3
cr
I.-29
A( )-NO. SUBJECTS
VWnrzntnMPRR: 1.9
NOVvvelN bxLT d-U :1.0
/I~~
7/A4.
Ar - I I I I I a
C MEAN MPRR1.6 1.2S.TYPHOSA ENDOTOXIN/ // 0.8
1 1// (~~~~~~~~3)
o /I I I I I I
rIII I,,,,, .,~(22
0 1 2 3 4 5 6 7HOURS
Pseudomonas endotoxin were prepared which met thesecriteria, one employing 'CrCls, the other Na25CrO4.
RESULTS
Quantitation of endotoxin tolerance during ty-phoid fever and tularemiaConcept of maximum pyrogenic response rate. The
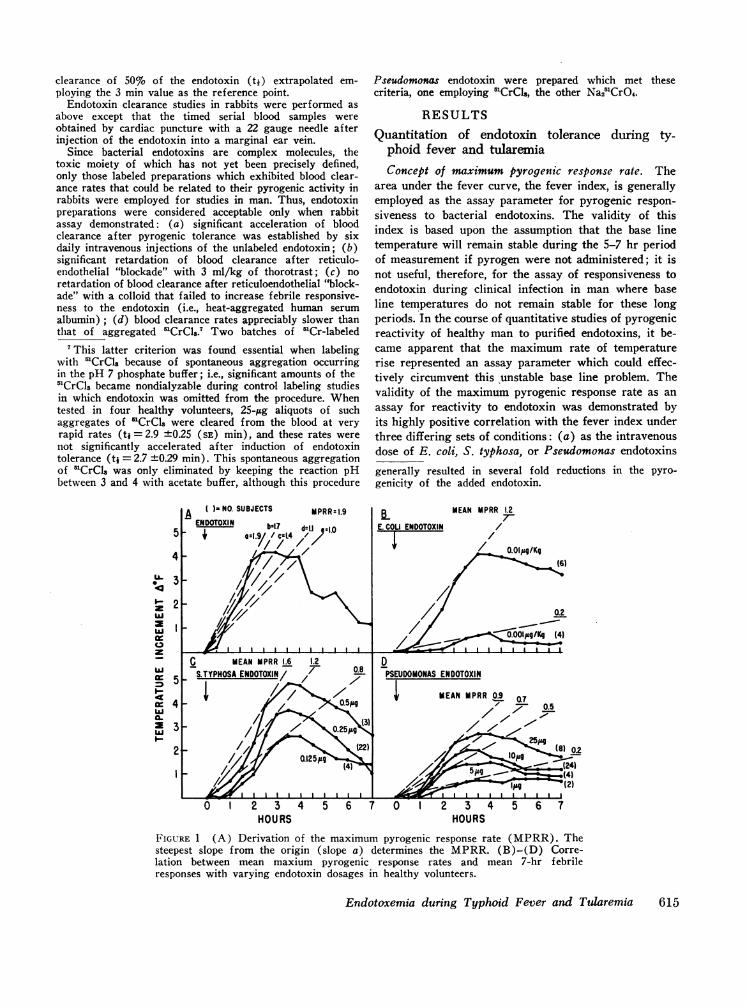
area under the fever curve, the fever index, is generallyemployed as the assay parameter for pyrogenic respon-siveness to bacterial endotoxins. The validity of thisindex is based upon the assumption that the base linetemperature will remain stable during the 5-7 hr periodof measurement if pyrogen were not administered; it isnot useful, therefore, for the assay of responsiveness toendotoxin during clinical infection in man where baseline temperatures do not remain stable for these longperiods. In the course of quantitative studies of pyrogenicreactivity of healthy man to purified endotoxins, it be-came apparent that the maximum rate of temperaturerise represented an assay parameter which could effec-tively circumvent this 'unstable base line problem. Thevalidity of the maximum pyrogenic response rate as anassay for reactivity to endotoxin was demonstrated byits highly positive correlation with the fever index underthree differing sets of conditions: (a) as the intravenousdose of E. coli, S. typhosa, or Pseudomonas endotoxinsgenerally resulted in several fold reductions in the pyro-genicity of the added endotoxin.
BE.COU ENDOTOXIN-F--
MEAN MPRR .27-
/ 0.0lug/Kg
//// 0.2
/ ~~~~~.~0.001,ugIKg (4)/t~~~C__,.}1
DPSEUDOMONASENDOTOXIN
MEAN MPRR 0.9 OLI'll 0.5
7- .
pg(8) 0.2
Itg -ft (2)en_ -- I I I I I L I I _IX_J l.=u--f0 1 2 3 4 5 6 7
HOURSFIGURE 1 (A) Derivation of the maximum pyrogenic response rate (MPRR). Thesteepest slope from the origin (slope a) determines the MPRR. (B)-(D) Corre-lation between mean maxium pyrogenic response rates and mean 7-hr febrileresponses with varying endotoxin dosages in healthy volunteers.
Endotoxemia during Typhoid Fever and Tularemia
I l ikza r I I I I I I I I I Il
615
Lr
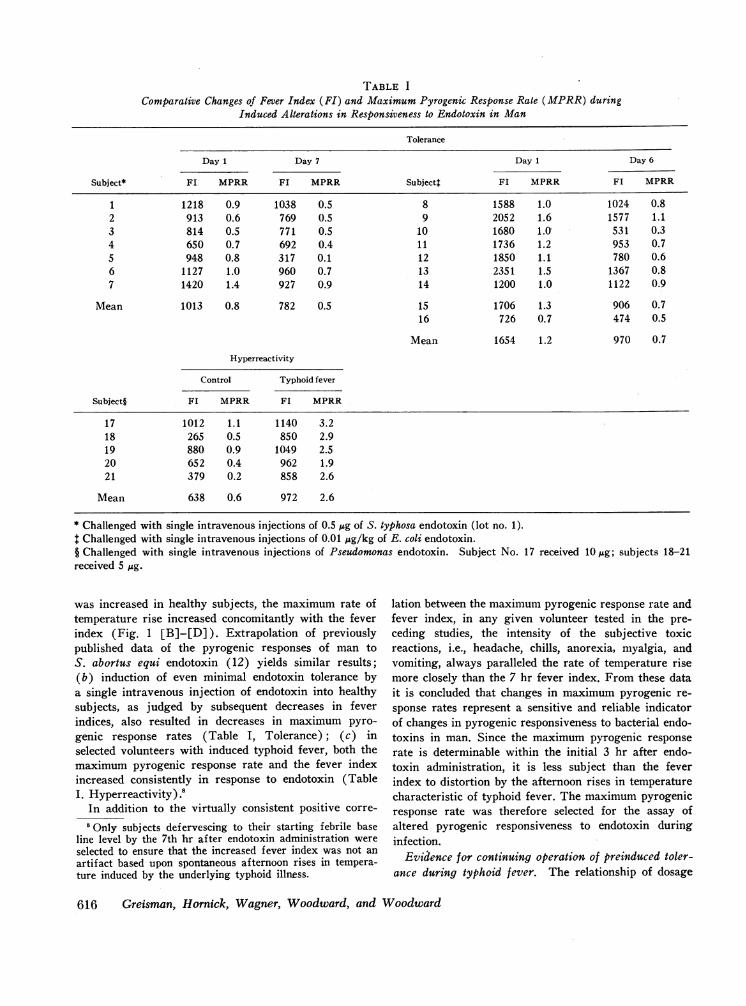
TABLE I
Comparative Changes of Fever Index (FI) and Maximum Pyrogenic Response Rate (MPRR) duringInduced Alterations in Responsiveness to Endotoxin in Man
Tolerance
Day I Day 7 Day I Day 6
Subject* FI MPRR F1 MPRR Subjectt FL MPRR FL MPRR
1 1218 0.9 1038 0.5 8 1588 1.0 1024 0.82 913 0.6 769 0.5 9 2052 1.6 1577 1.13 814 0.5 771 0.5 10 1680 1. 531 0.34 650 0.7 692 0.4 11 1736 1.2 953 0.75 948 0.8 317 0.1 12 1850 1.1 780 0.66 1127 1.0 960 0.7 13 2351 1.5 1367 0.87 1420 1.4 927 0.9 14 1200 1.0 1122 0.9
Mean 1013 0.8 782 0.5 15 1706 1.3 906 0.716 726 0.7 474 0.5
Mean 1654 1.2 970 0.7
Hyperreactivity
Control Typhoid fever
Subject§ FI MPRR FI MPRR
17 1012 1.1 1140 3.218 265 0.5 850 2.919 880 0.9 1049 2.520 652 0.4 962 1.921 379 0.2 858 2.6
Mean 638 0.6 972 2.6
* Challenged with single intravenous injections of 0.5 ,gg of S. typhosa endotoxin (lot no. 1).t Challenged with single intravenous injections of 0.01 Mug/kg of E. coli endotoxin.§ Challenged with single intravenous injections of Pseudomonas endotoxin. Subject No. 17 received lOug; subjects 18-21received 5 Mug.
was increased in healthy subj ects, the maximum rate oftemperature rise increased concomitantly with the feverindex (Fig. 1 [B]-[D]). Extrapolation of previouslypublished data of the pyrogenic responses of man toS. abortus equi endotoxin (12) yields similar results;(b) induction of even minimal endotoxin tolerance bya single intravenous injection of endotoxin into healthysubjects, as judged by subsequent decreases in feverindices, also resulted in decreases in maximum pyro-genic response rates (Table I, Tolerance); (c) inselected volunteers with induced typhoid fever, both themaximum pyrogenic response rate and the fever indexincreased consistently in response to endotoxin (TableI. Hyperreactivity).8
In addition to the virtually consistent positive corre-
8Only subjects defervescing to their starting febrile baseline level by the 7th hr after endotoxin administration wereselected to ensure that the increased fever index was not anartifact based upon spontaneous afternoon rises in tempera-ture induced by the underlying typhoid illness.
lation between the maximum pyrogenic response rate andfever index, in any given volunteer tested in the pre-ceding studies, the intensity of the subjective toxicreactions, i.e., headache, chills, anorexia, myalgia, andvomiting, always paralleled the rate of temperature risemore closely than the 7 hr fever index. From these datait is concluded that changes in maximum pyrogenic re-sponse rates represent a sensitive and reliable indicatorof changes in pyrogenic responsiveness to bacterial endo-toxins in man. Since the maximum pyrogenic responserate is determinable within the initial 3 hr after endo-toxin administration, it is less subject than the feverindex to distortion by the afternoon rises in temperaturecharacteristic of typhoid fever. The maximum pyrogenicresponse rate was therefore selected for the assay ofaltered pyrogenic responsiveness to endotoxin duringinfection.
Evidence for continuing operation of preinduced toler-ance during typhoid fever. The relationship of dosage
616 Greisman, Hornick, Wagner, Woodward, and Woodward
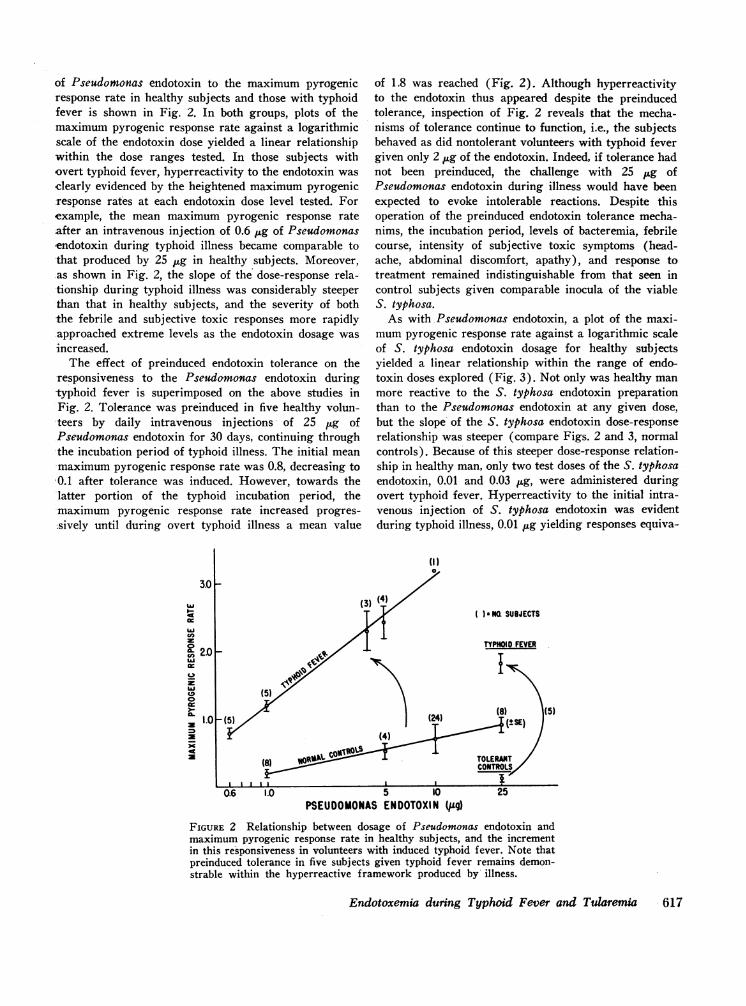
of Pseudomonas endotoxin to the maximum pyrogenicresponse rate in healthy subjects and those with typhoidfever is shown in Fig. 2. In both groups, plots of themaximum pyrogenic response rate against a logarithmicscale of the endotoxin dose yielded a linear relationshipwithin the dose ranges tested. In those subjects withovert typhoid fever, hyperreactivity to the endotoxin wasclearly evidenced by the heightened maximum pyrogenicresponse rates at each endotoxin dose level tested. Forexample, the mean maximum pyrogenic response rateafter an intravenous injection of 0.6 1ug of Pseudomonasendotoxin during typhoid illness became comparable tothat produced by 25 jug in healthy subjects. Moreover,as shown in Fig. 2, the slope of the dose-response rela--tionship during typhoid illness was considerably steeper-than that in healthy subjects, and the severity of boththe febrile and subjective toxic responses more rapidlyapproached extreme levels as the endotoxin dosage wasincreased.
The effect of preinduced endotoxin tolerance on theresponsiveness to the Pseudomonas endotoxin during-typhoid fever is superimposed on the above studies inFig. 2. Tolerance was preinduced in five healthy volun-
-teers by daily intravenous injections of 25 jug ofPseudomonas endotoxin for 30 days, continuing through
-the incubation period of typhoid illness. The initial mean-maximum pyrogenic response rate was 0.8, decreasing to0.1 after tolerance was induced. However, towards thelatter portion of the typhoid incubation period, themaximum pyrogenic response rate increased progres-sively until during overt typhoid illness a mean value
of 1.8 was reached (Fig. 2). Although hyperreactivityto the endotoxin thus appeared despite the preinducedtolerance, inspection of Fig. 2 reveals that the mecha-nisms of tolerance continue to function, i.e., the subjectsbehaved as did nontolerant volunteers with typhoid fevergiven only 2 jg of the endotoxin. Indeed, if tolerance hadnot been preinduced, the challenge with 25 jAg ofPseudomonas endotoxin during illness would have beenexpected to evoke intolerable reactions. Despite thisoperation of the preinduced endotoxin tolerance mecha-nims, the incubation period, levels of bacteremia, febrilecourse, intensity of subjective toxic symptoms (head-ache, abdominal discomfort, apathy), and response totreatment remained indistinguishable from that seen incontrol subjects given comparable inocula of the viableS. typhosa.
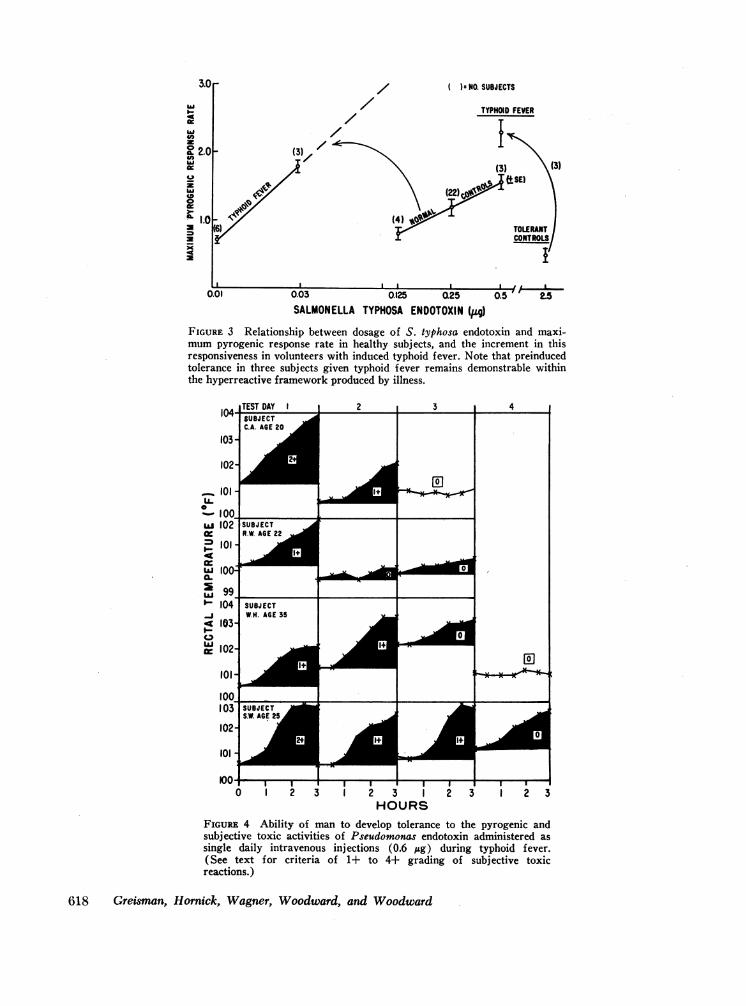
As with Pseudomonas endotoxin, a plot of the maxi-mumpyrogenic response rate against a logarithmic scaleof S. typhosa endotoxin dosage for healthy subjectsyielded a linear relationship within the range of endo-toxin doses explored (Fig. 3). Not only was healthy manmore reactive to the S. typhosa endotoxin preparationthan to the Pseudomonas endotoxin at any given dose,but the slope of the S. typhosa endotoxin dose-responserelationship was steeper (compare Figs. 2 and 3, normalcontrols). Because of this steeper dose-response relation-ship in healthy man, only two test doses of the S. typhosaendotoxin, 0.01 and 0.03 jug, were administered duringovert typhoid fever. Hyperreactivity to the initial intra-venous injection of S. typhosa endotoxin was evidentduring typhoid illness, 0.01 jug yielding responses equiva-
3.0 F
LL
z
ox
2.0
z
xU
CO400.
1h.0w
ccC>
( )a NQ SUBJECTS
TYPHOID FEVER
0.6 1.0 25
FIGURE 2 Relationship between dosage of Pseudomonas endotoxin andmaximum pyrogenic response rate in healthy subjects, and the incrementin this responsiveness in volunteers with induced typhoid fever. Note thatpreinduced tolerance in five subjects given typhoid fever remains demon-strable within the hyperreactive framework produced by illness.
Endotoxemia during Typhoid Fever and Tularemia 617
5 10PSEUDOMONASENDOTOXIN (jig)
_ LtSE)
X t4Ia
0
**1.0 (4)
N6) ~~~~~~~~~~~~~~~~TOLERAT1
0.01 0.03 0.125 Q25 0.5 2.5SALMONELLA TYPHOSA ENDOTOXIN(po)
FIGURE 3 Relationship between dosage of S. typhosa endotoxin and maxi-mumpyrogenic response rate in healthy subjects, and the increment in thisresponsiveness in volunteers with induced typhoid fever. Note that preinducedtolerance in three subjects given typhoid fever remains demonstrable withinthe hyperreactive framework produced by illness.
14TEST DAY I 1 2 3 410-SUBJECT ,
3- C.A. AGE
102 SE
us 102 SUBJECT
_J 02W.".AGE35L t
0 1 2 3 1 2 3 1 2 3 1 2 3HOURS
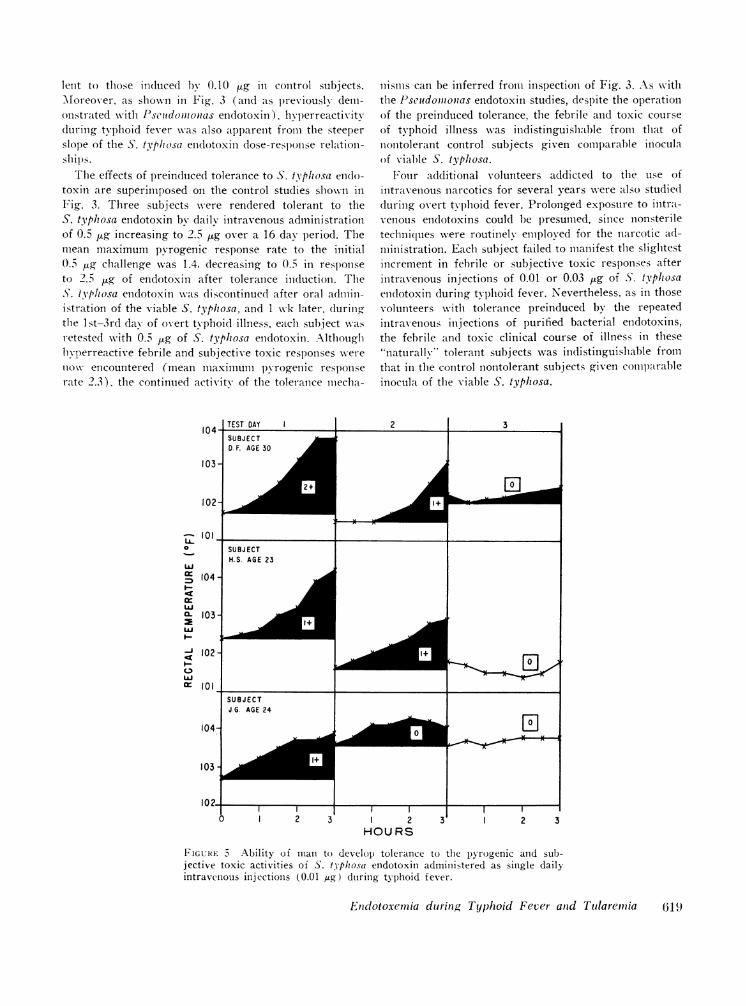
FIGURE 4 Ability of man to develop tolerance to the pyrogenic andsubjective toxic activities of Pseudomonas endotoxin administered assingle daily intravenous injections (0.6 Mg) during typhoid fever.(See text for criteria of 1+ to 4+ grading of subjective toxicreactions.)
618 Greisman, Hornick, Wagner, Woodward, and Woodward
leint to those induced by 0.10 Mg in control subjects.Moreov,-er, as shown in Fig. 3 (and s l)rexiousl detlm-onstrated with Pscindoi onuas endotoxin), hv-perreactiv itvduring typhoid fever was also apparent from the steeperslope of the S. typlhosa en(lotoxin dose-response relation-ships.
The effects of preinduced tolerance to S. typliosa enido-toxin are superimposed on the control studies shown inFig. 3. Three subjects were rendered tolerant to theS. typhosa endotoxin by daily intravenous administrationof 0.5 ,ug increasing to 2.5 Mg over a 16 day period. Themean maximum pyrogenic response rate to the initial0.5 Mg challenge was 1.4, decreasing to 0.5 in responseto 2.5; Mg of endotoxin after tolerance induction. TheS. tvplhosa endotoxin w,%-as discontinued after oral admiiin-istration of the viable S. typhosa, and 1 w-k l]ater, duriillthe I st-3rd day of overt typhoid illness, each subject wasretested with 0.5 Mlg of S. tyvphosa endotoxin. Althoughhyperreactiye febrile and subjective toxic responses werenow encountered (mean maximumlpyrogenic response
rate 2.3), the continued activity of the tolerance mecha-
104 -TEST DAY
SUBJECTD. F. AGE 30
103-
102-
LL-
LUJ
ar-wC-
I-C-,LUI
I0ISUBJECTH.S. AGE 23
104
103-
1023 11
nisms can be inferred from inspection of Fig. 3. As withthe Pseuidomin onas endotoxin studies, despite the operationof the preinduced tolerance, the febrile and toxic courseof typhoid illness was indistinguishable from that ofnontolerant control subjects given comI)aral)le inoculaof viable S. typhosa.
Four additional volunteers addicted to the use ofintravenous narcotics for several years w--ere also studiedduring overt typhoid fever. Prolonged exposure to intra-venous endotoxins could be presumed, since nonsteriletechniques were routinely employed for the narcotic ad-ministration. Each subject failed to manifest the slightestincrement in febrile or subjective toxic responses afterintravenous injections of 0.01 or 0.03 Mg of .S. typhosaendotoxin during typhoid fever. Nevertheless, as in thosevolunteers with tolerance preinduced by the repeatedintravenous injections of purified bacterial endotoxins,the febrile and toxic clinical course of illness in these'naturally' tolerant subjects was indistinguishable fromthat in the control nontolerant subjects given comparableinocula of the viable S. typhosa.
2 1
LIa
3 2HOURS
FIGURE 5 Ability of man to develop tolerance to the p)yrogenic and sub-jective toxic activities of S. tpliosa endotoxin administered as single dailyintravenous injections (0.01 fug) during typhoid fever.
Endotoxemia during Typhoid Fever and Tularemia 619
b**--
0)
3
2 30 2 3
14+)
XxS.TYPHOSA ENDOTOXIN(6 x 106JA/min)
2X 5X lox STOPt t t t
8 9 10 11 12 13 14
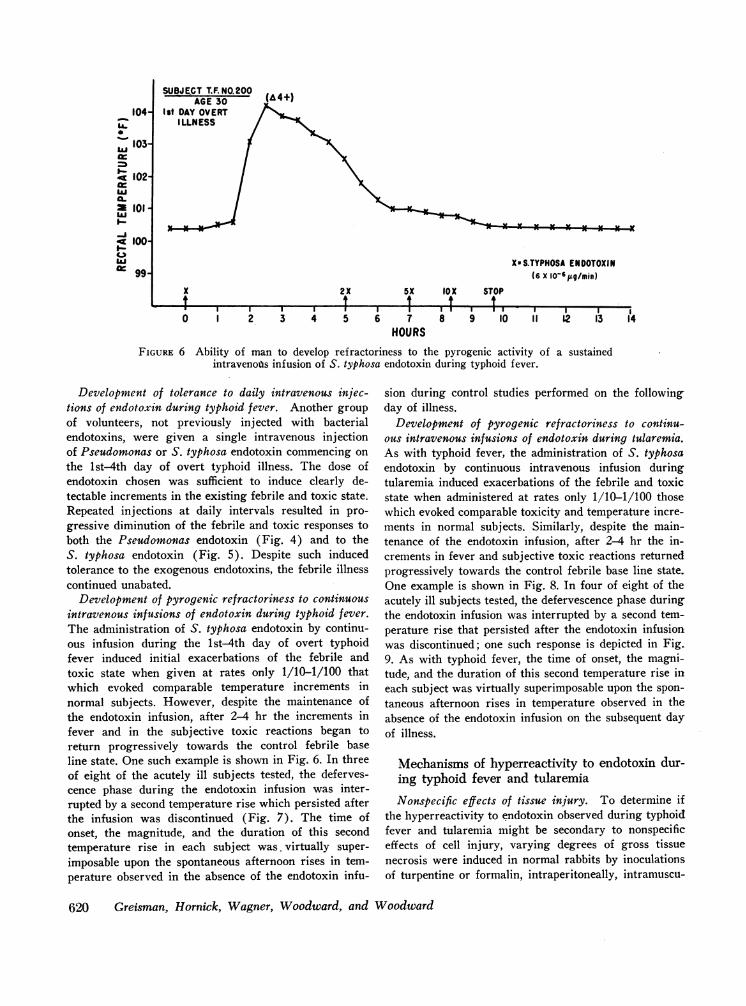
FIGURE 6 Ability of man to develop refractoriness to the pyrogenic activity of a sustainedintravenous infusion of S. typhosa endotoxin during typhoid fever.
Development of tolerance to daily intravenous injec-tions of endotoxin during typhoid fever. Another group
of volunteers, not previously injected with bacterialendotoxins, were given a single intravenous injectionof Pseudomonas or S. typhosa endotoxin commencing on
the lst-4th day of overt typhoid illness. The dose ofendotoxin chosen was sufficient to induce clearly de-tectable increments in the existing febrile and toxic state.Repeated injections at daily intervals resulted in pro-
gressive diminution of the febrile and toxic responses toboth the Pseudomonas endotoxin (Fig. 4) and to theS. typhosa endotoxin (Fig. 5). Despite such inducedtolerance to the exogenous endotoxins, the febrile illnesscontinued unabated.
Development of pyrogenic refractoriness to continuousintravenous infusions of endotoxin during typhoid fever.The administration of S. typhosa endotoxin by continu-ous infusion during the lst-4th day of overt typhoidfever induced initial exacerbations of the febrile andtoxic state when given at rates only 1/10-1/100 thatwhich evoked comparable temperature increments innormal subjects. However, despite the maintenance ofthe endotoxin infusion, after 2-4 hr the increments infever and in the subjective toxic reactions began toreturn progressively towards the control febrile baseline state. One such example is shown in Fig. 6. In threeof eight of the acutely ill subjects tested, the deferves-cence phase during the endotoxin infusion was inter-rupted by a second temperature rise which persisted afterthe infusion was discontinued (Fig. 7). The time ofonset, the magnitude, and the duration of this secondtemperature rise in each subject was. virtually super-
imposable upon the spontaneous afternoon rises in tem-perature observed in the absence of the endotoxin infu-
sion during control studies performed on the followingday of illness.
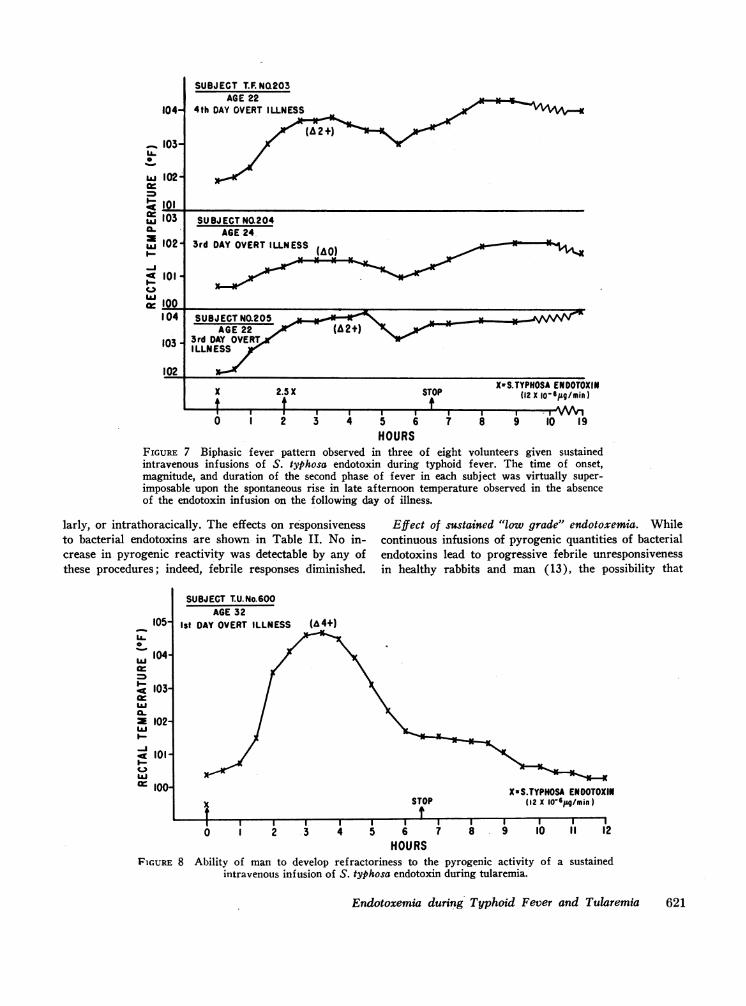
Development of pyrogenic refractoriness to continu-ous intravenous infusions of endotoxin during tularemia.As with typhoid fever, the administration of S. typhosaendotoxin by continuous intravenous infusion duringtularemia induced exacerbations of the febrile and toxicstate when administered at rates only 1/10-1/100 thosewhich evoked comparable toxicity and temperature incre-ments in normal subjects. Similarly, despite the main-tenance of the endotoxin infusion, after 2-4 hr the in-crements in fever and subjective toxic reactions returnedprogressively towards the control febrile base line state.One example is shown in Fig. 8. In four of eight of theacutely ill subjects tested, the defervescence phase duringthe endotoxin infusion was interrupted by a second tem-perature rise that persisted after the endotoxin infusionwas discontinued; one such response is depicted in Fig.9. As with typhoid fever, the time of onset, the magni-tude, and the duration of this second temperature rise ineach subject was virtually superimposable upon the spon-
taneous afternoon rises in temperature observed in theabsence of the endotoxin infusion on the subsequent dayof illness.
Mechanisms of hyperreactivity to endotoxin dur-ing typhoid fever and tularemia
Nonspecific effects of tissue injury. To determine ifthe hyperreactivity to endotoxin observed during typhoidfever and tularemia might be secondary to nonspecificeffects of cell injury, varying degrees of gross tissuenecrosis were induced in normal rabbits by inoculationsof turpentine or formalin, intraperitoneally, intramuscu-
620 Greisman, Hornick, Wagner, Woodward, and Woodward
SUBJECT T.F.NO.200AGE 30
I t DAY OVERTILLNESS
104-LU0
a-3.
I-A4_ 100-I-9C.,)LUa: 99-
Xt8 I I I I I0 1 2 3 4 5 6 7
HOURS
SUBJECT T.F. NQ203AGE 22
4th DAY OVERT ILLNESS
(A2+
SUBJ ECTNO.204AGE 24
3rd DAY OVERTILLNESS (AO) o
(AO
SUBJECTNO.205 S %AO~3rd DAY OVERTI LLN ESS
X 2.5 X
6 iSTOP
fXwS.TYPKOSAENDOTOXIN
(12 x lo-6j9g/min)
4 5 6 7 8 9 10 19HOURS
FIGURE 7 Biphasic fever pattern observed in three of eight volunteers given sustainedintravenous infusions of S. typhosa endotoxin during typhoid fever. The time of onset,magnitude, and duration of the second phase of fever in each subject was virtually super-imposable upon the spontaneous rise in late afternoon temperature observed in the absenceof the endotoxin infusion on the following day of illness.
larly, or intrathoracically. The effects on responsivenessto bacterial endotoxins are shown in Table II. No in-crease in pyrogenic reactivity was detectable by any ofthese procedures; indeed, febrile responses diminished.
105-U-0
S 104-
o 103-
ILX 102-LuI.-
-J41 101-C 1)1= 100-
SUBJECT T.U.No.600AGE 32
Ist DAY OVERT ILLNESS
Effect of sustained "low grade" endotoxemia. Whilecontinuous infusions of pyrogenic quantities of bacterialendotoxins lead to progressive febrile unresponsivenessin healthy rabbits and man (13), the possibility that
(A4+)
STOPXsS.TYPHOSA ENDOTOXIN
(12 X 10-6,g/min)
Endotoxemia during Typhoid Fever and Tularemia 621
104-
_ 103-LLU-w 102
c 101= 103a-X 102I-
C 101
wcr 100
104
103
102
I~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~i I ~I I I I I I I I I I
0 1 2 3 4 5 6 7 8 9 10 11 12HOURS
FIGURE 8 Ability of man to develop refractoriness to the pyrogenic activity of a sustainedintravenous infusion of S. typhosa endotoxin during tularemia.
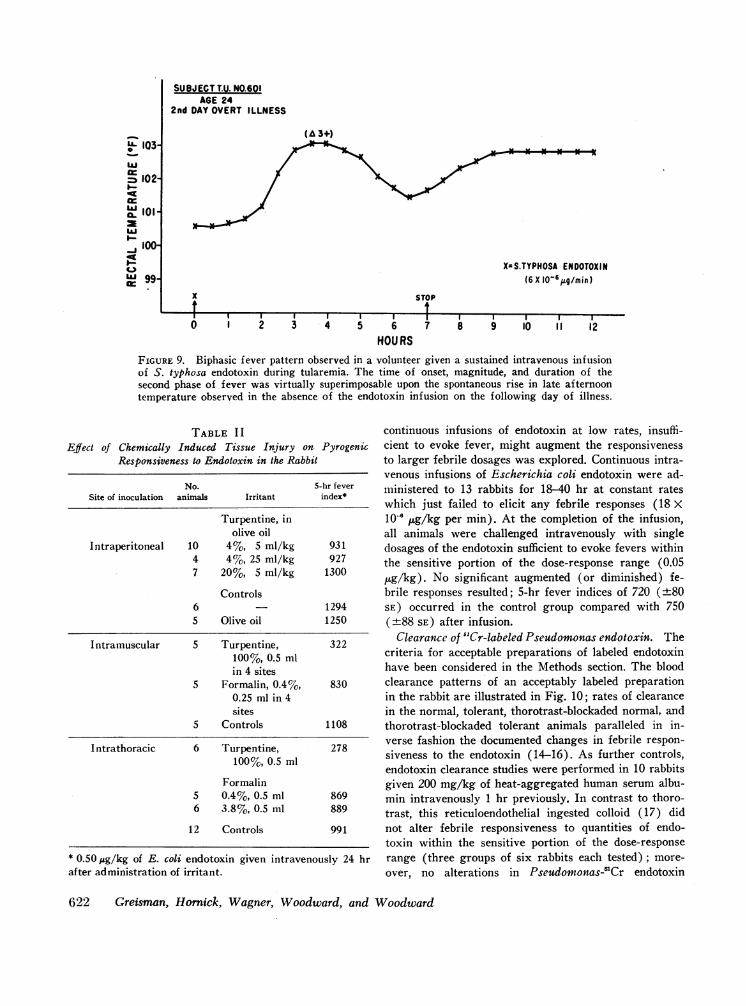
SUBJ ECTT.U. NO.601AGE 24
2nd DAY OVERT ILLNESS
X=S.TYPHOSA ENDOTOXIN(6X 10O6 g/min)
x
tSTOP
II I II I I. I I I I.
1 2 3 4 5 6
HOURS
I I17 8 9 10 11 12
FIGURE 9. Biphasic fever pattern observed in a volunteer given a sustained intravenous infusionof S. typhosa endotoxin during tularemia. The time of onset, magnitude, and duration of thesecond phase of fever was virtually superimposable upon the spontaneous rise in late afternoontemperature observed in the absence of the endotoxin infusion on the following day of illness.
TABLE I IEffect of Chemically Induced Tissue Injury on Pyrogenic
Responsiveness to Endotoxin in the Rabbit
No. 5-hr feverSite of inoculation animals Irritant index*
Turpentine, inolive oil
Intraperitoneal 10 4%, 5 ml/kg 9314 4%, 25 ml/kg 9277 20%, 5 ml/kg 1300
Controls6 12945 Olive oil 1250
Intramuscular 5 Turpentine, 322100%, 0.5 mlin 4 sites
5 Formalin, 0.4%, 8300.25 ml in 4sites
5 Controls 1108
I ntrathoracic 6 Turpentine, 278100%, 0.5 ml
Formalin5 0.4%, 0.5 ml 8696 3.8%, 0.5 ml 889
12 Controls 991
* 0.50/jg/kg of E. coli endotoxin given intravenously 24 hrafter administration of irritant.
continuous infusions of endotoxin at low rates, insuffi-cient to evoke fever, might augment the responsivenessto larger febrile dosages was explored. Continuous intra-venous infusions of Escherichia coli endotoxin were ad-ministered to 13 rabbits for 18-40 hr at constant rateswhich just failed to elicit any febrile responses (18 X10' ug/kg per min). At the completion of the infusion,all animals were challenged intravenously with singledosages of the endotoxin sufficient to evoke fevers withinthe sensitive portion of the dose-response range (0.05,ug/kg). No significant augmented (or diminished) fe-brile responses resulted; 5-hr fever indices of 720 (±80SE) occurred in the control group compared with 750(±+88 SE) after infusion.
Clearance of "Cr-labeled Pseudomonas endotoxin. Thecriteria for acceptable preparations of labeled endotoxinhave been considered in the Methods section. The bloodclearance patterns of an acceptably labeled preparationin the rabbit are illustrated in Fig. 10; rates of clearancein the normal, tolerant, thorotrast-blockaded normal, andthorotrast-blockaded tolerant animals paralleled in in-verse fashion the documented changes in febrile respon-siveness to the endotoxin (14-16). As further controls,endotoxin clearance studies were performed in 10 rabbitsgiven 200 mg/kg of heat-aggregated human serum albu-min intravenously 1 hr previously. In contrast to thoro-trast, this reticuloendothelial ingested colloid (17) didnot alter febrile responsiveness to quantities of endo-toxin within the sensitive portion of the dose-responserange (three groups of six rabbits each tested) ; more-
over, no alterations in Pseudomonas-'Cr endotoxin
622 Greisman, Hornick, Wagner, Woodward, and Woodward
L03wLU
= 102'
X, 101
> 100
C-)LU 99.
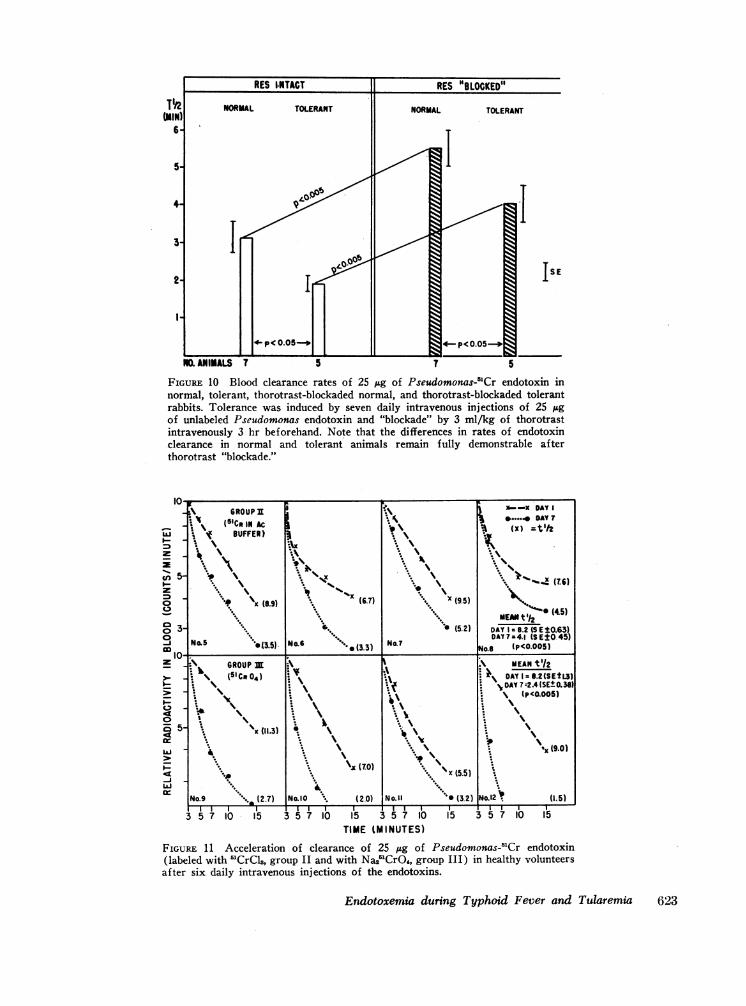
FIGURE 10 Blood clearance rates of 25 jtg of Pseudomonas-'Cr endotoxin innormal, tolerant, thorotrast-blockaded normal, and thorotrast-blockaded tolerantrabbits. Tolerance was induced by seven daily intravenous injections of 25 sgof unlabeled Pseudomonas endotoxin and "blockade" by 3 ml/kg of thorotrastintravenously 3 hr beforehand. Note that the differences in rates of endotoxinclearance in normal and tolerant animals remain fully demonstrable afterthorotrast "blockade."
(20) INO.II15 3 5 7 10
TIME (MINUTES)
FIGURE 11 Acceleration of clearance of 25 lug of Pseudomonas-'Cr endotoxin(labeled with 5'CrCl8, group II and with Na21CrO4, group III) in healthy volunteersafter six daily intravenous injections of the endotoxins.
Endotoxemia during Typhoid Fever and Tularemia 623
TIME (MINUTES)
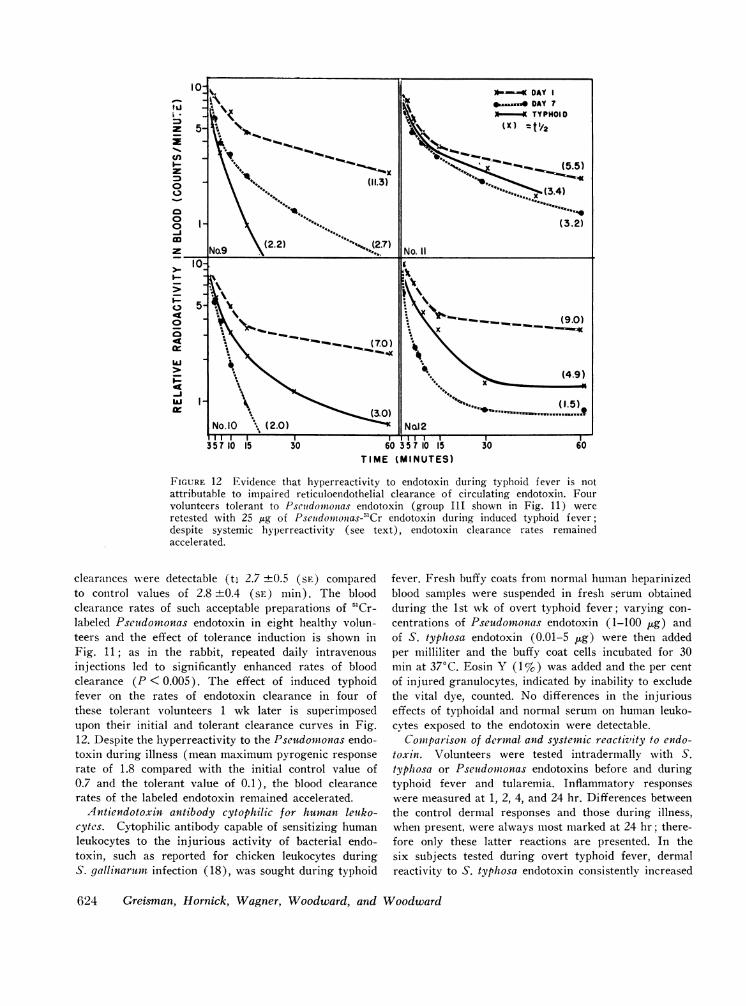
FIGURE 12 Evidence that hyperreactivity to endotoxin during typhoid f ever is notattributable to impaired reticuloendothelial clearance of circulating endotoxin. Fourvolunteers tolerant to Pseutdomtonias endotoxin (group III shown in Fig. 11) wereretested with 25 ,ug of Pseitdoiiionzas-51Cr endotoxin during induced typhoid fever;despite systemic hyperreactivity (see text), endotoxin clearance rates remainedaccelerated.
clearances were detectable (t, 2.7 ±0.5 (SE) comparedto control values of 2.8 ±0.4 (SE) min). The bloodclearance rates of such acceptable preparations of 51Cr-labeled Pseuidomonas endotoxin in eight healthy volun-teers and the effect of tolerance induction is shown inFig. 11; as in the rabbit, repeated daily intravenousinjections led to significantly enhanced rates of bloodclearance (P < 0.005). The effect of induced typhoidfever on the rates of endotoxin clearance in four ofthese tolerant volunteers 1 wk later is superimposedupon their initial and tolerant clearance curves in Fig.12. Despite the hyperreactivity to the Pseudomonas endo-toxin during illness (mean maximum pyrogenic responserate of 1.8 compared with the initial control value of0.7 and the tolerant value of 0.1), the blood clearancerates of the labeled endotoxin remained accelerated.
Antiendotoxin antibody cytophilic for human leuko-cytes. Cytophilic antibody capable of sensitizing humanleukocytes to the injurious activity of bacterial endo-toxin, such as reported for chicken leukocytes duringS. gallinarum infection (18), was sought during typhoid
fever. Fresh buffy coats from normal human heparinizedblood samples were suspended in fresh serum obtainedduring the 1st wk of overt typhoid fever; varying con-centrations of Pseudomonas endotoxin (1-100 fig) andof S. typhosa endotoxin (0.01-5 jig) were then addedper milliliter and the buffy coat cells incubated for 30min at 370C. Eosin Y (1%) was added and the per centof injured granulocytes, indicated by inability to excludethe vital dye, counted. No differences in the injuriouseffects of typhoidal and normal serum on human leuko-cytes exposed to the endotoxin were detectable.
Coniparison of dermal and systemic reactivity to endo-toxin. Volunteers were tested intradermally with S.typhosa or Pseudonionas endotoxins before and duringtyphoid fever and tularemia. Inflammatory responseswere measured at 1, 2, 4, and 24 hr. Differences betweenthe control dermal responses and those during illness,when present, were always most marked at 24 hr; there-fore only these latter reactions are presented. In thesix subjects tested during overt typhoid fever, dermalreactivity to S. typhosa endotoxin consistently increased
624 Greisnman, Hornick, Wagner, Woodward, and Woodward
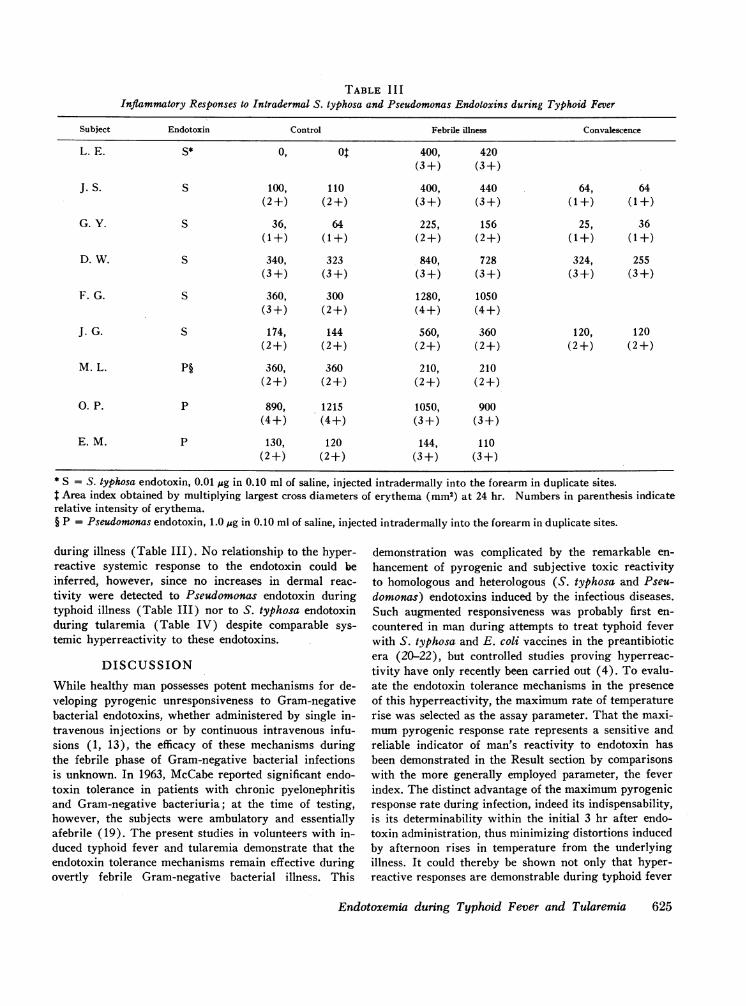
TABLE I I IInflammatory Responses to Intradermal S. typhosa and Pseudomonas Endotoxins during Typhoid Fever
Subject Endotoxin Control Febrile illness Convalescence
L. E. S* 0, 01 400, 420(3 +) (3 +)
J. S. S 100, 110 400, 440 64, 64(2+) (2+) (3+) (3+) (1+) (1+)
G. Y. S 36, 64 225, 156 25, 36(1+) (1+) (2+) (2+) (1+) (1+)
D. W. S 340, 323 840, 728 324, 255(3 +) (3+) (3+) (3 +) (3 +) (3 +)
F. G. S 360, 300 1280, 1050(3+) (2+) (4+) (4+)
J. G. S 174, 144 560, 360 120, 120(2+) (2+) (2+) (2+) (2+) (2+)
M. L. P§ 360, 360 210, 210(2+) (2+) (2+) (2+)
0. P. P 890, 1215 1050, 900(4+) (4+) (3+) (3+)
E. M. P 130, 120 144, 110(2+) (2+) (3+) (3+)
* S = S. typhosa endotoxin, 0.01 fig in 0.10 ml of saline, injected intradermally into the forearm in duplicate sites.t Area index obtained by multiplying largest cross diameters of erythema (mm2) at 24 hr. Numbers in parenthesis indicaterelative intensity of erythema.§ P = Pseudomonas endotoxin, 1.0 pig in 0.10 ml of saline, injected intradermally into the forearm in duplicate sites.
during illness (Table III). No relationship to the hyper-reactive systemic response to the endotoxin could beinferred, however, since no increases in dermal reac-tivity were detected to Pseudomonas endotoxin duringtyphoid illness (Table III) nor to S. typhosa endotoxinduring tularemia (Table IV) despite comparable sys-temic hyperreactivity to these endotoxins.
DISCUSSION
While healthy man possesses potent mechanisms for de-veloping pyrogenic unresponsiveness to Gram-negativebacterial endotoxins, whether administered by single in-travenous injections or by continuous intravenous infu-sions (1, 13), the efficacy of these mechanisms duringthe febrile phase of Gram-negative bacterial infectionsis unknown. In 1963, McCabe reported significant endo-toxin tolerance in patients with chronic pyelonephritisand Gram-negative bacteriuria; at the time of testing,however, the subjects were ambulatory and essentiallyafebrile (19). The present studies in volunteers with in-duced typhoid fever and tularemia demonstrate that theendotoxin tolerance mechanisms remain effective duringovertly febrile Gram-negative bacterial illness. This
demonstration was complicated by the remarkable en-hancement of pyrogenic and subjective toxic reactivityto homologous and heterologous (S. typhosa and Pseu-domonas) endotoxins induced by the infectious diseases.Such augmented responsiveness was probably first en-countered in man during attempts to treat typhoid feverwith S. typhosa and E. coli vaccines in the preantibioticera (20-22), but controlled studies proving hyperreac-tivity have only recently been carried out (4). To evalu-ate the endotoxin tolerance mechanisms in the presenceof this hyperreactivity, the maximum rate of temperaturerise was selected as the assay parameter. That the maxi-mumpyrogenic response rate represents a sensitive andreliable indicator of man's reactivity to endotoxin hasbeen demonstrated in the Result section by comparisonswith the more generally employed parameter, the feverindex. The distinct advantage of the maximum pyrogenicresponse rate during infection, indeed its indispensability,is its determinability within the initial 3 hr after endo-toxin administration, thus minimizing distortions inducedby afternoon rises in temperature from the underlyingillness. It could thereby be shown not only that hyper-reactive responses are demonstrable during typhoid fever
Endotoxemia during Typhoid Fever and Tularemia 625
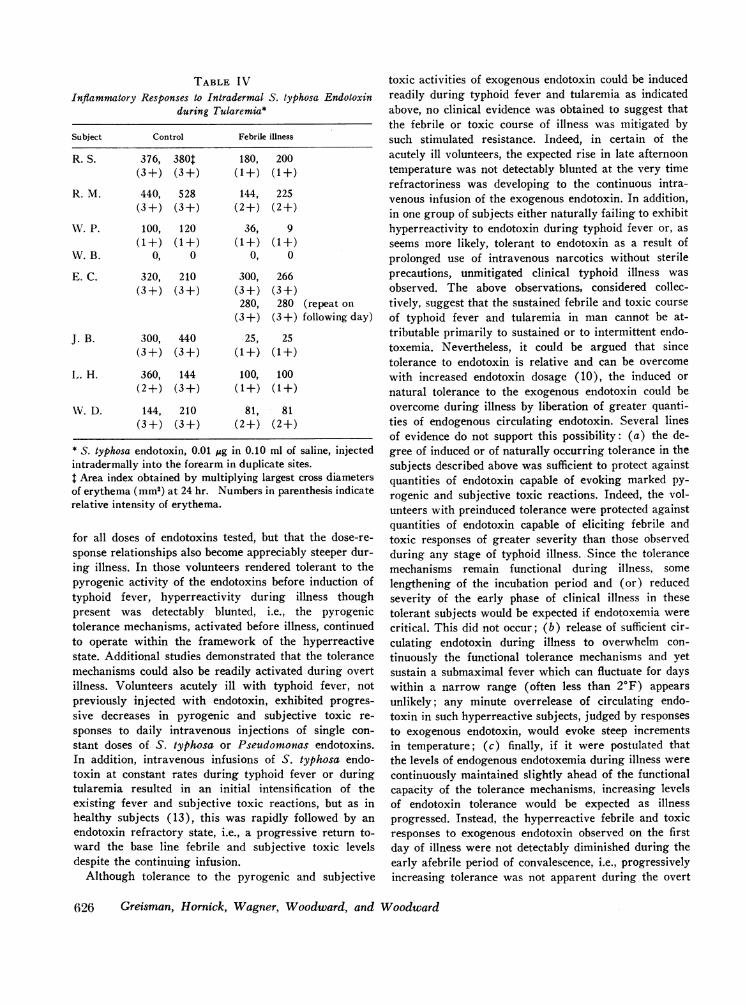
TABLE IVInflammatory Responses to Intradermal S. typhosa Endotoxin
during Tularemui*
Subject Control Febrile illness
R. S. 376, 380T 180, 200(3+) (3+) (1+) (1+)
R. M. 440, 528 144, 225(3+) (3+) (2+) (2+)
WV. P. 100, 120 36, 9(1+) (1+) (1+) (1+)
W.B. 0, 0 0, 0
E. C. 320, 210 300, 266(3 +) (3 +) (3 +) (3+)
280, 280 (repeat on(3+) (3+) following day)
J. B. 300, 440 25, 25(3+) (3+) (1+) (1+)
L,. H. 360, 144 100, 100(2+) (3+) (1+) (1+)
WV. D. 144, 210 81, 81(3+) (3+) (2+) (2+)
* S. typhosa endotoxin, 0.01 jug in 0.10 ml of saline, injectedintradermally into the forearm in duplicate sites.$ Area index obtained by multiplying largest cross diametersof erythema (mm2) at 24 hr. Numbers in parenthesis indicaterelative intensity of erythema.
for all doses of endotoxins tested, but that the dose-re-sponse relationships also become appreciably steeper dur-ing illness. In those volunteers rendered tolerant to thepyrogenic activity of the endotoxins before induction oftyphoid fever, hyperreactivity during illness thoughpresent was detectably blunted, i.e., the pyrogenictolerance mechanisms, activated before illness, continuedto operate within the framework of the hyperreactivestate. Additional studies demonstrated that the tolerancemechanisms could also be readily activated during overtillness. Volunteers acutely ill with typhoid fever, notpreviously injected with endotoxin, exhibited progres-sive decreases in pyrogenic and subjective toxic re-sponses to daily intravenous injections of single con-stant doses of S. typhosa or Pseudomonas endotoxins.In addition, intravenous infusions of S. typhosa endo-toxin at constant rates during typhoid fever or duringtularemia resulted in an initial intensification of theexisting fever and subjective toxic reactions, but as inhealthy subjects (13), this was rapidly followed by anendotoxin refractory state, i.e., a progressive return to-ward the base line febrile and subjective toxic levelsdespite the continuing infusion.
Although tolerance to the pyrogenic and subjective
toxic activities of exogenous endotoxin could be inducedreadily during typhoid fever and tularemia as indicatedabove, no clinical evidence was obtained to suggest thatthe febrile or toxic course of illness was mitigated bysuch stimulated resistance. Indeed, in certain of theacutely ill volunteers, the expected rise in late afternoontemperature was not detectably blunted at the very timerefractoriness was developing to the continuous intra-venous infusion of the exogenous endotoxin. In addition,in one group of subjects either naturally failing to exhibithyperreactivity to endotoxin during typhoid fever or, asseems more likely, tolerant to endotoxin as a result ofprolonged use of intravenous narcotics without sterileprecautions, unmitigated clinical typhoid illness wasobserved. The above observations, considered collec-tively, suggest that the sustained febrile and toxic courseof typhoid fever and tularemia in man cannot be at-tributable primarily to sustained or to intermittent endo-toxemia. Nevertheless, it could be argued that sincetolerance to endotoxin is relative and can be overcomewith increased endotoxin dosage (10), the induced ornatural tolerance to the exogenous endotoxin could beovercome during illness by liberation of greater quanti-ties of endogenous circulating endotoxin. Several linesof evidence do not support this possibility: (a) the de-gree of induced or of naturally occurring tolerance in thesubjects described above was sufficient to protect againstquantities of endotoxin capable of evoking marked py-rogenic and subjective toxic reactions. Indeed, the vol-unteers with preinduced tolerance were protected againstquantities of endotoxin capable of eliciting febrile andtoxic responses of greater severity than those observedduring any stage of typhoid illness. Since the tolerancemechanisms remain functional during illness, somelengthening of the incubation period and (or) reducedseverity of the early phase of clinical illness in thesetolerant subjects would be expected if endotoxemia werecritical. This did not occur; (b) release of sufficient cir-culating endotoxin during illness to overwhelm con-tinuously the functional tolerance mechanisms and yetsustain a submaximal fever which can fluctuate for dayswithin a narrow range (often less than 2°F) appearsunlikely; any minute overrelease of circulating endo-toxin in such hyperreactive subjects, judged by responsesto exogenous endotoxin, would evoke steep incrementsin temperature; (c) finally, if it were postulated thatthe levels of endogenous endotoxemia during illness werecontinuously maintained slightly ahead of the functionalcapacity of the tolerance mechanisms, increasing levelsof endotoxin tolerance would be expected as illnessprogressed. Instead, the hyperreactive febrile and toxicresponses to exogenous endotoxin observed on the firstday of illness were not detectably diminished during theearly afebrile period of convalescence, i.e., progressivelyincreasing tolerance was not apparent during the overt
626 Greisman, Hornick, Wagner, Woodward, and Woodward

phase of illness (4). Additional support for the conceptthat endotoxemia is not critical for the sustained feverduring Gram-negative bacterial infection is provided bythe well controlled studies of Bennett employing Esch-erichia coli-induced peritonitis in rabbits (23).
It is emphasized that the conclusion presently drawnis that endotoxemia plays no major role in pathogenesisof the sustained fever and toxemia of typhoid fever andtularemia. The above conclusion requires careful quali-fication in three respects: (a) it does not preclude thepresence of endotoxemia during infection; indeed, sero-logic evidence exists for endotoxemia during the initial7-10 days of typhoid fever (24). Rather, it is the re-sponse of man to endotoxemia, i.e., the rapid ability todevelop tolerance, and the lack of clinical protection af-forded by high levels of induced tolerance, that formsthe basis for the inference that such endotoxemia cannotprimarily account for the sustained fever and toxemiaduring typhoid fever; (b) the possibility is not precludedthat abrupt increments in circulating endotoxin levelsduring typhoid fever or tularemia mediate acute exacer-bations of fever and toxemia and shock in the hyperreac-tive host. Rather, it is postulated from the present datathat these exacerbations would be superimposed uponthe more basic mechanisms responsible for the sustainedfever and toxemia, and that their frequency and intensitywould be limited by the development of systemic toler-ance; (c) the present conclusion does not preclude animportant role of endotoxin acting directly at the sitesof bacterial lodgement to enhance the local inflammatoryresponse and the release of endogenous pyrogen fromthe surrounding leukocyte and macrophage populations.Indeed, hyperreactive inflammatory responses were ob-served after local (i.e., intradermal) injections of ho-mologous endotoxin during typhoid fever. Such abilityof S. typhosa endotoxin to potentiate local tissue in-jury during typhoid fever actually may contribute sig-nificantly to the pathogenesis of sustained fever andtoxemia of illness.9
In light of the above considerations, the significanceof the pyrogenic tolerance to endotoxin observed duringlate convalescence from typhoid fever and tularemia(26, 27), and the vascular hyperreactivity to catechola-mines observed during typhoid fever (28), previouslyregarded as evidence for a major role of circulating en-dotoxin in pathogenesis of these illnesses, requires re-evaluation. These findings are simply compatible withthe presence of endotoxemia; they in no way define itsrelative importance in the production of illness. The
9 This latter possibility is not invalidated by the unmitigatedclinical illness in volunteers rendered tolerant to the systemiceffects of endotoxin. Systemic tolerance is not synonymouswith local tolerance. At least one site in man, the skin,exhibits enhanced inflammatory responses to the endotoxinemployed to induce systemic tolerance (25).
present studies, on the other hand, now permit the in-terpretation that sufficient endotoxemia occurs duringillness to produce or contribute to tolerance and vascu-lar hyperreactivity without being primarily responsiblefor the sustained clinical disease. Additional support forthis interpretation is presently available. The pyrogenictolerance after recovery from typhoid fever and tulare-mia was evident by 50-70% reductions in mean feverindices (26, 27). Comparable degrees of tolerance re-cently have been induced in normal volunteers by in-travenous injections of subfebrile doses of Boivin-ex-tracted endotoxins, i.e., induction of significant pyro-genic tolerance to endotoxin in man does not requireclinically demonstrable symptoms.10 Exposure of man tosuch small but nevertheless tolerance-evoking quantitiesof endotoxin incidental to the prolonged typhoid andtularemia illness would not be unexpected. As for thevascular hyperreactivity to catecholamines observedduring typhoid fever, this persisted for several weeksinto the afebrile convalescent phase, was not demonstra-ble during the other comparably febrile illness inducedby the endotoxin-containing Gram-negative microbePasteurella tularensis, and could not be reproduced innormal man by intravenous injections of toxic quan-tities of purified bacterial endotoxins. It was thereforeemphasized that mechanisms other than endotoxemia,such as release of serotonin from the inflammed in-testinal mucosa, might primarily account for thisphenomenon (28). The recent demonstration that avariety of inflammatory diseases of the intestine evokevascular hyperreactivity to catecholamines in man (29)renders this latter explanation more plausible.
The mechanisms underlying the hyperreactivity toendotoxin during typhoid fever and tularemia were fur-ther explored in the present studies. It has already beendemonstrated that the phenomenon is not a nonspecificreaction to fever per se; the hyperreactivity was detec-table during the afebrile incubation period and continuedinto the afebrile convalescent phase; moreover, volun-teers febrile with the virus of Sandfly fever exhibitedno detectable hyperreactivity (4). Nonspecific effects oftissue injury also do not appear responsible. The presentfindings demonstrate that rabbits with varying degreesof gross tissue necrosis induced by chemical irritants(turpentine and formalin) administered intraperitoneally,intramuscularly, or intrathoracically did not exhibit hy-perreactive febrile responses to endotoxin; indeed, re-duced responses were observed, confirming the findingsof Raskova (30). Removal of natural protective circu-lating factors or enhancement of cellular reactivity bysubfebrile levels of endotoxemia also appears an unlikelymechanism of hyperreactivity, since infusions of endo-toxin for up to 40 hr at rates that just failed to elicit
10 Unpublished observations.
Endotoxemia during Typhoid Fever and Tularemia 627

fever did not render rabbits more responsive to subse-quent challenge with pyrogenic doses. Nor could evidencebe obtained that the hyperreactive febrile responses dur-ing typhoid fever were mediated by inability of the retic-uloendothelial system to clear the blood of the injectedendotoxin labeled with 5"Cr; indeed, endotoxin bloodclearance rates, accelerated as a result of induction ofendotoxin tolerance before typhoid fever, remained ac-celerated during infection. Such findings further sup-port the thesis that the mechanisms responsible for en-dotoxin tolerance continue to operate during infection.Moreover, the present endotoxin blood clearance datain man parallel those in mice infected with Mycobac-terium tuberculosis (BCG strain) wherein hyperreac-tivity to E. coli endotoxin developed during infection,but clearance of the radioactively labeled endotoxin wasaccelerated (31 ).
An alternative hypothesis for the systemic hyperreac-tivity to endotoxin during typhoid fever and tularemiawould be that suggested by Abernathy and Spink forthe hyperreactivity to Brucella endotoxin during brucel-losis, i.e., acquired hypersensitivity (32, 33). Importantsupport for this hypothesis was derived from the ap-parent correlation between the intensity of the dermalinflammatory responses elicited by brucella skin testantigens and the intensity of the pyrogenic and toxicsystemic responses to Brucella endotoxin (32). How-ever, the Brucella endotoxin preparation employed byAbernathy and Spink possessed pyrogenic activitiesquite different from those of classical purified prepara-tions."' As already emphasized by Beeson (2). it istherefore uncertain as to whether the observed svstemichyperreactivity during brucellosis reflects acquired hy-persensitivity to the endotoxin per se or to other anti-genic components in this preparation. In the presentstudies, significant increments in dermal inflammatoryresponses to highly purified S. typhosa endotoxin weredetectable during overt typhoid fever; however, unlikebrucellosis, the dermal hyperreactivity could not becorrelated with the systemic hyperreactivitv to the en-dotoxin. Thus, no dermal hyperreactivity was found toPseudomonas endotoxin during typhoid fever nor toS. typhosa endotoxin during tularemia, although com-parably hyperreactive pyrogenic and toxic responses fol-lowed the intravenous administration of these endo-toxins. Moreover, whereas systemic hyperreactivity per-sisted for years in the subjects with brucellosis tested
" The pyrogenic activity of the Briucella endotoxin was notdemonstrable in normal subjects until dosages of 250 ug wereadministered intravenously (compared to less than 5 ,ig forPseudomonas or 0.1 jug for S. typhosa or E. coli endotoxins).Moreover, the latent period before fever onset averagedmore than 3 hr (even in the highly reactive group of subjectswith brucellosis) compared with approximately 1 hr for theclassical endotoxin preparations.
with the Brucella preparation (32, 33), systemic hyper-reactivity waned rapidly during convalescence from ty-phoid fever and tularemia with the purified endotoxinpreparations employed in the present studies (4). Al-ternative attempts to demonstrate increases in cytophilicantibody capable of sensitizing leukocytes to the in-jurious activity of bacterial endotoxin, as has been re-ported in the chicken during S. galliniaruin infection(18), were also unsuccessful. It must be concluded,therefore, that the systemic hyperreactivity to bacterialendotoxins during typhoid fever and tularemia cannotbe ascribed at present to enhanced levels of acquired hy-persensitivity, and further studies to define the responsi-ble mechanisms are currently in progress.
ACKNOWLEDGMENTSThe authors express their deepest appreciation to the volun-teers who contributed so faithfully to these studies and tothe officials of the Maryland House of Correction for theirgenerous cooperation and interest, without which thesestudies could not have been accomplished. Appreciation is alsoexpressed to Mrs. Judy Bucchanan for the excellent technicalassistance with the labeling and counting during the Pseuido-monas-"Cr endotoxin clearance studies.
This investigation was supported by the U. S. Army Medi-cal Research and Development Command, contract DA-49-193-MD-2867, under the auspices of the Commission onEpidemiological Survey of the Armed Forces Epidemiologi-cal Board, by the U. S. Public Health Service ResearchGrant AI-07052, and by Career Development Award 2-K3-He-15,237, from the U. S. Public Health Service.
REFERENCES1. Morgan, H. R. 1948. Resistance to the action of the
endotoxin of enteric bacilli in man. J. C/in. Invest. 27:706.
2. Beeson, P. B. 1954. Discussion of Spink, XV. XV. Thesignificance of endotoxin in brucellosis: experimental andclinical studies. Tranls. Ass. Amner. Physicians Phila-delphia. 67: 292.
3. Atkins, E. 1960. Pathogenesis of fever. PhXsiol. Rev.40: 580.
4. Greisman, S. E., R. B. Hornick, and T. E. WVoodward.1964. The role of endotoxin during typhoid fever andtularemia in man. III. Hyperreactivity to endotoxinduring infection. J. Clin. Inivest. 43: 1747.
5. Hornick, R. B., T. E. Woodward, F. R. McCrumb, M. J.Snyder, A. T. Dawkins, J. T. Bulkeley, F. D. L. Macorra,and F. A. Carozza. 1966. Study of induced typhoid feverin man. I. Evaluation of vaccine effectiveness. Traits. Ass.Amer. Phisicians (Philadelphia). 79: 361.
6. Hornick, R. B., and H. T. Eigelsbach. 1966. Aerogenicimmunization of man with live tularemia vaccine. Bac-teriol. Rev. 30: 532.
7. Webster, M. E., J. F. Sagin, M. Landy, and A. G.Johnson. 1955. Studies on the 0 antigen of Salmonellatyphosa. I. Purification of the antigen. J. Imininnol. 74:455.
8. Nesset, N. M., J. McLallen, P. Z. Anthony, and L. G.Ginger. 1950. Bacterial pyrogens. I. Pyrogenic prepara-
628 Greisman, Hornick, Wagner, Woodward, and Woodward

tion from a Pseudomonas species. J. Amcr. Pharmn. Ass.39: 456.
9. Boivin, A., and L. Mesrobeanu. 1935. Recherches sur lesantigenes somatiques et sur les endotoxines des bacteries.I. Considerations generales et expose des techniquesutilisees. Rev. Imminol. (Paris). 1: 553.
10. Beeson, P. B. 1947. Tolerance to bacterial pyrogens.I. Factors influencing its development. J. Exp. Med.86: 29.
11. Braude, A. I., F. J. Carey, D. Sutherland, and M.Zalesky. 1955. Studies with radioactive endotoxin. I. Theuse of Cr5' to label endotoxin of Eschericlia coli.J. Clin. Invest. 34: 850.
12. Mechanic, R. C., E. Frei III, M. Landy, and W. W.Smith. 1962. Quantitative studies of human leukocyticand febrile response to single and repeated doses ofpurified bacterial endotoxin. J. Clin. Invest. 41: 162.
13. Greisman, S. E., and W. E. Woodward. 1965. Mecha-nisms of endotoxin tolerance. III. The refractory stateduring continuous intravenous infusions of endotoxin.J. Exp. Med. 121: 911.
14. Greisman, S. E., F. A. Carozza, Jr., and J. D. Hills.1963. Mechanisms of endotoxin tolerance. I. Relationshipbetween tolerance and reticuloendothelial system phago-cytic activity in the rabbit. J. Exp. Med. 117: 663.
15. Wolff, S. M., J. H. Mulholland, and S. B. Ward. 1965.Quantitative aspects of the pyrogenic response of rabbitsto endotoxin. J. Lab. Clin. Med. 65: 268.
16. Shulman, J. A., and R. G. Petersdorf. 1965. Relationshipof endogenous pyrogen and serum augmenting factor toendotoxin tolerance. Proc. Soc. Exp. Biol. Med. 119: 218.
17. lio, M., and H. N. Wagner, Jr. 1963. Studies of thereticuloendothelial system (RES). I. Measurements ofthe phagocytic capacity of the RES in man and dog.J. Clin. Invest. 42: 417.
18. Buxton, A., and D. Allan. 1963. Studies on immunityand pathogenesis of Salmonellosis. I. Antigen-antibodyreactions on circulating leucocytes of chickens infectedwith Salmonella gallinarum. Immntinology. 6: 520.
19. McCabe, W. R. 1963. Endotoxin tolerance. II. Its occur-rence in patients with pyelonephritis. .J. Clin. Invest. 42:618.
20. Ditthorn, F., and W. Schultz. 1915. Zur Antigenbe-handlung des Typhus. Med. Klin. 11: 100.
21. Kraus, R., and S. Mazza. 1914. Zur Frage der Vakzine-therapie des Typhus abdominalis. Deut. MVed. Wochenschr.40: 1556.
22. Kraus, R. 1914. Bemurkungen uber schutzimpfungen undeine Bakteriotherapie des Typhus abdominalis. WienerKlin. Wochenschr. 27: 1443.
23. Bennett, I. L., Jr. 1948. Observations on the fever causedby bacterial pyrogens. I. A study of the relationshipbetween the fever caused by bacterial pyrogens and thefever accompanying acute infections. J. Exp. Aled. 88:267.
24. Dennis, E. W., and A. S. Saigh. 1945. Precipitabletyphoid somatic antigen in the serum of typhoid feverpatients. Science. 102: 280.
25. Greisman, S. E., H. N. Wagner, Jr., M. lio, and R. B.Hornick. 1964. Mechanisms of endotoxin tolerance. II.Relationship between endotoxin tolerance and reticulo-endothelial system phagocytic activity in man. J. Exp.Med. 119: 241.
26. Greisman, S. E., R. B. Hornick, F. A. Carozza, Jr., andT. E. Woodward. 1963. The role of endotoxin duringtyphoid fever and tularemia in man. I. Acquisition oftolerance to endotoxin. J. Clin. Invest. 42: 1064.
27. Neva, F. A., and H. R. Morgan. 1950. Tolerance to theaction of endotoxins of enteric bacilli in patients con-valescent from typhoid and paratyphoid fevers. J. Lab.Clin. Med. 35: 911.
28. Greisman, S. E., R. B. Hornick, F. A. Carozza, Jr., andT. E. Woodward. 1964. The role of endotoxin duringtyphoid fever and tularemia in man. II. Altered cardio-vascular responses to catecholamines. J. Clin. Invest. 43:986.
29. Bowen, G. E., and J. B. Kirsner. 1965. Positive epineph-rine skin test for "circulating endotoxin" in inflammatorydisease of the intestine. Amer. J. Clin. Pathol. 44: 642.
30. Raskova, H. 1964. Nonspecific endotoxin-like resistanceinduced by simple chemical compounds. In BacterialEndotoxins; proceedings of a symposium. M. Landy andW. Braun, editors. Institute of Microbiology, Rutgers,The State University, New Brunswick. 1: 546.
31. Howard, J. G., G. Biozzi, B. N. Halpern, S. Stiffel, andD. Mouton. 1959. The effect of Mycobacterihn tuibercu-losis (BCG) infection on the resistance of mice to bac-terial endotoxin and Salmonella enteritidis infection.Brit. J. Exp. Pathol. 40: 281.
32. Abernathy, R. S., and W. W. Spink. 1958. Studies withbrucella endotoxin in humans: the significance of sus-ceptibility to endotoxin in the pathogenesis of brucellosis.J. Clin. Invest. 37: 219.
33. Spink, W. W. 1954. The significance of endotoxin inbrucellosis: experimental and clinical studies. Trans. Ass.Amer. Physicians (Philadelphia). 67: 283.
Endotoxemia during Typhoid Fever and Tularemia 629