REVIEW Voices of the past: a review of Paleozoic and ...
33
REVIEW Voices of the past: a review of Paleozoic and Mesozoic animal sounds Phil Senter* Department of Natural Sciences, Fayetteville State University, 1200 Murchison Road, Fayetteville, NC 28301, USA (Received 10 March 2009; final version received 6 May 2009) Here, I present a review and synthesis of fossil and neontological evidence to find major trends in the pre-Cenozoic evolution of animal acoustic behaviour. Anatomical, ecological and phylogenetic data support the following scenario. Stridulating insects, including crickets, performed the first terrestrial twilight choruses during the Triassic. The twilight chorus was joined by water boatmen in the Lower Jurassic, anurans in the Upper Jurassic, geckoes and birds in the Lower Cretaceous, and cicadas and crocodilians in the Upper Cretaceous. Parallel evolution of defensive stridulation took place multiple times within Malacostraca, Arachnida and Coleoptera. Parallel evolution of defensive and courtship-related sound production took place in Actinopterygii, possibly as early as the Devonian. Defensive vocalisations by tetrapods probably did not appear until their predators acquired tympanic ears in the Permian. Tympanic ears appeared independently in Diadectomorpha, Seymouriamorpha, Parareptilia, Diapsida and derived Synapsida. Crocodilians and birds acquired vocal organs independently, and there is no anatomical evidence for vocal ability in bird-line archosaurs basal to the avian clade Ornithothoraces. Acoustic displays by non-avian dinosaurs were therefore probably non-vocal. Other aspects of the evolution of acoustic behaviour in these and other lineages are also discussed. Keywords: acoustic behaviour; vocalisation; hearing; Crustacea; Insecta; Orthoptera; Coleoptera; Hemiptera; Crocodylia; Aves; Mammalia Introduction The fossil record does not include audio recordings. As a result, few researchers lose sleep over such questions as whether Triceratops heard crickets chirping in the evening, whether Mesozoic treetops resounded with birdsong or frogsong, or whether the immense corpses of sauropods were surrounded by the buzzing of bottle flies. Questions such as these seem silly at first but are nevertheless worthwhile to ask for three reasons: (1) inferable acoustic details must be included if the paleontologist is to accomplish one of paleontology’s main goals: the reconstruction of the ancient world in as much detail as possible, (2) acoustic signals are of great importance to many animals, so the reconstruction of the lifestyles of ancient organisms must take acoustic signals into consideration, and (3) many studies have addressed whether or not extinct animals could hear (e.g. Reisz 1981; Wu 1994; Clack and Allin 2004), so it is reasonable to ask what they heard. Bioacoustics, the study of animal sound production and reception, is a rich field with much to offer the paleontologist for application to the study of sound production and reception in fossil animals. Such application could aptly be called paleobioacoustics. Thus far, most paleobioacoustical research has been concen- trated on the study of the evolution of hearing in tetrapod vertebrates (e.g. Reisz 1981; Allin 1986; Wu 1994; Clack and Allin 2004; Vater et al. 2004) and the evolution of sound production in the insect orders Orthoptera and Hemiptera (e.g. Sweet 1996; Rust et al. 1999; Be ´thoux and Nel 2002; Gorochov and Rasnitsyn 2002). However, many other paleobioacoustical issues can be addressed with available data from fossil and neontological evidence. For example, the first appearances of fossils of sound- producing taxa can be used to constrain the times of origin of their characteristic sounds. Also, knowledge of directional selection on extant animal sounds can be used to infer characteristics of the sounds produced by their ancestors. In addition, because the functions of animal sounds depend on their reception by the intended recipients, information on the evolution of hearing can be used to constrain the times of origin of certain animal sounds. For example, one can reasonably infer that certain courtship sounds, territorial sounds and other sounds directed at conspecifics were absent in taxa that lacked appropriate sensory structures. Similarly, one can also reasonably infer that certain anti-predator sounds were absent before the appearance of predators with appropriate sensory structures. Much information pertinent to the ISSN 0891-2963 print/ISSN 1029-2381 online q 2008 Taylor & Francis DOI: 10.1080/08912960903033327 http://www.informaworld.com *Email: [email protected] Historical Biology Vol. 20, No. 4, December 2008, 255–287 Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009
Transcript of REVIEW Voices of the past: a review of Paleozoic and ...

REVIEW
Voices of the past: a review of Paleozoic and Mesozoic animal sounds
Phil Senter*
Department of Natural Sciences, Fayetteville State University, 1200 Murchison Road, Fayetteville, NC 28301, USA
(Received 10 March 2009; final version received 6 May 2009)
Here, I present a review and synthesis of fossil and neontological evidence to find major trends in the pre-Cenozoic evolutionof animal acoustic behaviour. Anatomical, ecological and phylogenetic data support the following scenario. Stridulatinginsects, including crickets, performed the first terrestrial twilight choruses during the Triassic. The twilight chorus wasjoined by water boatmen in the Lower Jurassic, anurans in the Upper Jurassic, geckoes and birds in the Lower Cretaceous,and cicadas and crocodilians in the Upper Cretaceous. Parallel evolution of defensive stridulation took place multiple timeswithin Malacostraca, Arachnida and Coleoptera. Parallel evolution of defensive and courtship-related sound productiontook place in Actinopterygii, possibly as early as the Devonian. Defensive vocalisations by tetrapods probably did notappear until their predators acquired tympanic ears in the Permian. Tympanic ears appeared independently inDiadectomorpha, Seymouriamorpha, Parareptilia, Diapsida and derived Synapsida. Crocodilians and birds acquired vocalorgans independently, and there is no anatomical evidence for vocal ability in bird-line archosaurs basal to the avian cladeOrnithothoraces. Acoustic displays by non-avian dinosaurs were therefore probably non-vocal. Other aspects of theevolution of acoustic behaviour in these and other lineages are also discussed.
Keywords: acoustic behaviour; vocalisation; hearing; Crustacea; Insecta; Orthoptera; Coleoptera; Hemiptera; Crocodylia;Aves; Mammalia
Introduction
The fossil record does not include audio recordings. As a
result, few researchers lose sleep over such questions as
whether Triceratops heard crickets chirping in the evening,
whether Mesozoic treetops resounded with birdsong or
frogsong, or whether the immense corpses of sauropods
were surrounded by the buzzing of bottle flies. Questions
such as these seem silly at first but are nevertheless
worthwhile to ask for three reasons: (1) inferable acoustic
details must be included if the paleontologist is to
accomplish one of paleontology’s main goals: the
reconstruction of the ancient world in as much detail as
possible, (2) acoustic signals are of great importance to
many animals, so the reconstruction of the lifestyles of
ancient organisms must take acoustic signals into
consideration, and (3) many studies have addressed
whether or not extinct animals could hear (e.g. Reisz
1981; Wu 1994; Clack and Allin 2004), so it is reasonable
to ask what they heard.
Bioacoustics, the study of animal sound production
and reception, is a rich field with much to offer the
paleontologist for application to the study of sound
production and reception in fossil animals. Such
application could aptly be called paleobioacoustics. Thus
far, most paleobioacoustical research has been concen-
trated on the study of the evolution of hearing in tetrapod
vertebrates (e.g. Reisz 1981; Allin 1986; Wu 1994; Clack
and Allin 2004; Vater et al. 2004) and the evolution of
sound production in the insect orders Orthoptera and
Hemiptera (e.g. Sweet 1996; Rust et al. 1999; Bethoux and
Nel 2002; Gorochov and Rasnitsyn 2002). However, many
other paleobioacoustical issues can be addressed with
available data from fossil and neontological evidence. For
example, the first appearances of fossils of sound-
producing taxa can be used to constrain the times of
origin of their characteristic sounds. Also, knowledge of
directional selection on extant animal sounds can be used
to infer characteristics of the sounds produced by their
ancestors. In addition, because the functions of animal
sounds depend on their reception by the intended
recipients, information on the evolution of hearing can
be used to constrain the times of origin of certain animal
sounds. For example, one can reasonably infer that certain
courtship sounds, territorial sounds and other sounds
directed at conspecifics were absent in taxa that lacked
appropriate sensory structures. Similarly, one can also
reasonably infer that certain anti-predator sounds were
absent before the appearance of predators with appropriate
sensory structures. Much information pertinent to the
ISSN 0891-2963 print/ISSN 1029-2381 online
q 2008 Taylor & Francis
DOI: 10.1080/08912960903033327
http://www.informaworld.com
*Email: [email protected]
Historical Biology
Vol. 20, No. 4, December 2008, 255–287
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009
reconstruction of ancient acoustic behaviour has been
published, but before now no attempt has been made to
pool available information to illuminate broad trends in
such behaviour across large geologic time spans. Here,
I present a review of the available literature so as to
perform such a synthesis for the Paleozoic and Mesozoic
Eras.
Major themes in the evolution of aerial soundproduction and reception
Sound reception
Most invertebrates are aquatic, and several lineages have
independently evolved sense organs that perceive water
displacement (Budelmann 1992a,b; Coffin et al. 2004).
However, it is difficult to say whether most possess a true
sense of hearing, both because the definition of ‘hearing’
varies among researchers and because in aquatic environ-
ments the distinctions between sound, vibration and water
flow are blurred (Budelmann 1992b). In any case, among
extant invertebrates acoustic communication is unknown
outside Arthropoda, and there is no reason to believe that the
case was different among extinct invertebrates.
Airborne sound reception is typically accomplishedwith
tympanic ears. In such ears airborne sounds cause vibrations
in a thin membrane, the tympanum (Figure 1), internal to
which is an air-filled chamber; mechanoreceptors that are
linked to the tympanum detect its movement in response to
sounds (Wever 1978; Yager 1999; Kardong 2006). Among
insects, tympanic ears have appeared independently in
Cicadidae (cicadas), Corixidae (water boatmen), Tachinidae
(tachinid flies), Sarcophagidae (flesh flies), Neuroptera
(lacewings), Mantodea (mantises), Cicindelidae
(tiger beetles), Scarabaeidae (scarab beetles), and several
times within Orthoptera (crickets and grasshoppers) and
Lepidoptera (moths and butterflies) (Yager 1999; Flook et al.
2000; Robert and Hoy 2000; Cokl et al. 2006) (Figure 2).
Tympanic ears in cicadas, water boatmen and crickets are
associated with intraspecific acoustic communication
(Bailey 1991; Gerhardt and Huber 2002; Cokl et al. 2006).
Those of tachinid and flesh flies are used to find their cricket
hosts when the latter stridulate (Yager 1999; Robert andHoy
2000). The tympanic ears of lepidopterans, lacewings,
mantises, beetles and grasshoppers are tuned to frequencies
produced by echolocating bats (Mammalia: Chiroptera), the
sounds of which stimulate evasive behaviours in these
insects (Bailey 1991; Yager 1999; Flook et al. 2000). Such
ears are an evolutionary response to predation by bats
(Bailey 1991; Flook et al. 2000) and, like bats, were
therefore absent before the Cenozoic. Courtship sounds of
grasshoppers and lepidopterans are secondary and appeared
after the advent of tympanic hearing in those taxa (Yager
1999), and were therefore absent before the Cenozoic.
In addition to receptors attuned to frequencies used in
intraspecific communication, some cricket ears also have a
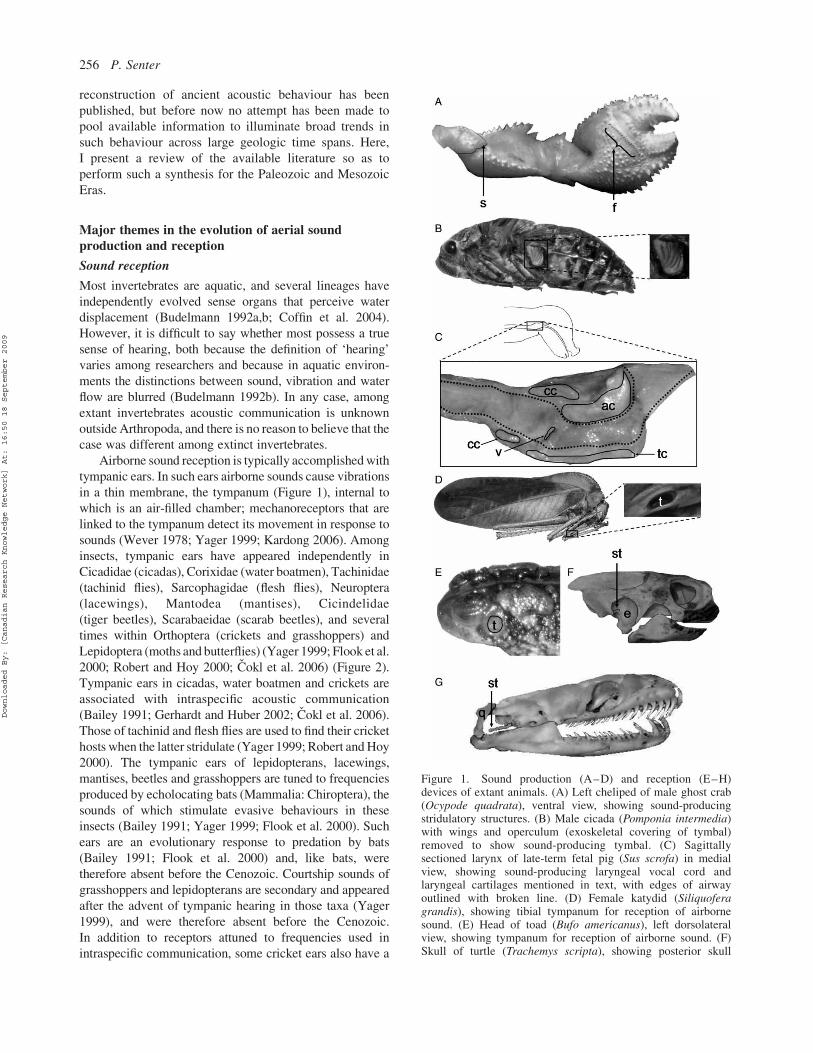
Figure 1. Sound production (A–D) and reception (E–H)devices of extant animals. (A) Left cheliped of male ghost crab(Ocypode quadrata), ventral view, showing sound-producingstridulatory structures. (B) Male cicada (Pomponia intermedia)with wings and operculum (exoskeletal covering of tymbal)removed to show sound-producing tymbal. (C) Sagittallysectioned larynx of late-term fetal pig (Sus scrofa) in medialview, showing sound-producing laryngeal vocal cord andlaryngeal cartilages mentioned in text, with edges of airwayoutlined with broken line. (D) Female katydid (Siliquoferagrandis), showing tibial tympanum for reception of airbornesound. (E) Head of toad (Bufo americanus), left dorsolateralview, showing tympanum for reception of airborne sound. (F)Skull of turtle (Trachemys scripta), showing posterior skull
P. Senter256
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

small number of acoustic receptors for much higher
frequencies produced by echolocating bats (Imaizumi and
Pollack 1999) (Figure 3), a secondary innovation that
presumably appeared after the Cenozoic appearance of bats.
In tetrapod vertebrates with tympanic ears, a bony
connection transmits vibrations from the tympanum to the
inner ear, where resulting fluid vibrations stimulate
mechanoreceptors (Wever 1978; Kardong 2006). This
mechanism appeared independently in Anura (frogs and
toads), Diapsida (lizards, crocodilians, birds and kin)
and Synapsida (mammals and kin) (Clack and Allin 2004),
and apparently also in the extinct tetrapod groups
Seymouriamorpha (Ivakhnenko 1987), Diadectomorpha
(Berman et al. 1992) and Parareptilia (Laurin and Reisz
1995) (Figure 4). Whether the tympanic ears of turtles
(Testudines) appeared independently depends on whether
or not turtles arose from within Diapsida or Parareptilia, an
issue that is disputed (e.g. Rieppel and deBraga 1996; Lee
1997a,b). Osteological evidence, reviewed later in this
embayment and stapes; the embayment houses an air-filled spaceinternal to the tympanum, which is attached to the rim of theembayment, and the stapes conducts auditory vibrations from thetympanum to the inner ear. (G) Skull of snake (Boidae,undetermined species), showing stapes and loosely attachedquadrate bone; the quadrate acts as a tympanum, and the stapes(attached to the quadrate via a cartilaginous extension that is notshown here) conducts auditory vibrations from the quadrate tothe inner ear. ac, arytenoid cartilage; cc, cricoid cartilage; e,embayment; f, file; q, quadrate; s, scraper; st, stapes; t,tympanum; tc, thyroid cartilage; v, vocal cord.
R
Figure 2. Phylogeny of insect orders, after Grimaldi and Engel (2005) showing distribution of sound production and tympanic ears(Dumortier 1963b; Aiken 1985; Yager 1999; Virant-Doberlet and Cokl 2004; Drosopoulos and Claridge 2006). Large symbols indicatewide taxonomic distribution within an order, and small symbols indicate limited taxonomic distribution. Filled circles and squaresindicate occurrence within a family or families with known pre-Cenozoic fossil records. A, expulsion of air through spiracles; B, loudflight buzzing; P, percussion of body parts against substrate; S, stridulation; T, tymbals; t, tympanic ears; W, wing fluttering.
Historical Biology 257
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009
paper, indicates the presence of tympanic ears in all these
groups before the Cenozoic, and in most cases before the
Mesozoic. As with many insect taxa, it is possible that the
original function of tympanic hearing in the various
tetrapod taxa was predator avoidance. The sounds made by
the approach of a large animal often stimulate evasive or
cryptic behaviour in extant reptiles (Greene 1988), birds
and mammals (personal observation). In predatory taxa
hearing may have originally facilitated prey localisation.
Many extant predatory tetrapods rely heavily on chemical
and visual means of prey localisation (Ewer 1973;
Duellman and Trueb 1994; Pough et al. 1998), but some
use hearing to locate prey (Ewer 1973).
Sound production
Methods of animal sound production fall into five main
categories: stridulation, vocalisation, percussion, forced
airflow and the tymbal mechanism. Stridulation, the rubbing
together of body parts, is the most common means of sound
production in arthropods. Usually one of the stridulatory
parts (the file) is ridged, while the other (the scraper or
plectrum) is not (Figure 1). Stridulation has evolved
convergently in several arthropod groups (Figure 2) and is
especially prevalent among groups with highly sclerotised
(hardened) exoskeletons such as Malacostraca (shrimps,
crabs and kin), Arachnida (spiders, scorpions and kin),
Orthoptera (crickets and grasshoppers), Hemiptera (true
bugs) and Coleoptera (beetles) (e.g. Dumortier 1963b).
Unfortunately for paleontology, arthropod fossils are rarely
preserved in enough detail to determine whether or not
stridulatory structures are present. An exception is the taxon
Orthoptera, in which the stridulatory structures are modified
wing veins and can easily be discerned on fossil wings (e.g.
Bethoux and Nel 2002). Stridulation can be effective even if
the intended recipient lacks an auditory apparatus.
Arthropods often detect the stridulation of conspecifics via
substrate-borne vibrations (Crowson 1981; Gogala 1985,
2006; Barth 2002). Stridulation by prey upon seizure by a
vertebrate or arthropod predator stimulates the predator’s
tactile receptors – and auditory receptors, if present – and
often stimulates the predator to release the prey (Masters
1979; Crowson 1981). In many cases the predator’s release
of the stridulating prey may be a hardwired response to an
honest warning, for stridulating prey items often follow
stridulation with painful stimuli (e.g. stinging) or emission
of noxious chemicals, or are externally tough and therefore
difficult to eat (Masters 1979).
Many tetrapod vertebrates (Tetrapoda) produce sounds
by vocalisation, the vibration of mucosal folds called vocal
cords that extend into the lumen of the respiratory tract
(Figure 1). Vocal cords occur in Anura (frogs and toads),
Ambystoma þ Dicamptodon (mole salamanders), Mam-
malia (mammals), Gekkonidae þ Eublepharidae (geck-
oes), Pituophis melanoleucus (the bull snake), Crocodylia
(crocodilians), and Aves (birds) (Reese 1914; Maslin
1950; Kelemen 1963; Gans and Maderson 1973; King
1989; Young et al. 1995). Morphology, topology and
phylogenetic distribution indicate that vocal cords arose
independently in each of these groups (Figure 5). In all but
Aves the vocal cords are in the larynx (Reese 1914; Maslin
1950; Kelemen 1963; Gans and Maderson 1973; Young
et al. 1995); in birds they are in a unique organ called the
syrinx (King 1989).
Percussion is the hitting of a body part against a
substrate. Examples include nuptial abdomen tapping by
stoneflies (Plecoptera) and booklice (Psocoptera) and the
aquatic head-slap displays of crocodilians (Pearman 1928;
Garrick et al. 1978; Thorbjarnarson 1989; Stewart and
Sandberg 2006). These are discussed in more detail below.
Some insects produce defensive sounds by forcing air
through spiracles (Roth and Hartman 1967; Aiken 1985).
However, sound production by forceful airflow is more
characteristic of the vertebrate taxon Amniota, many
members of which hiss by forced ventilation. Hissing as a
threat device, often directed at potential predators, is
widespread among extant amniotes, including lizards,
snakes, turtles, crocodilians, basal birds and basal
mammals (Greene 1988; Davies 2002; Kear 2005;
Nowak 2005). It may therefore be a behavioural
symplesiomorphy for Amniota. If that is correct, then the
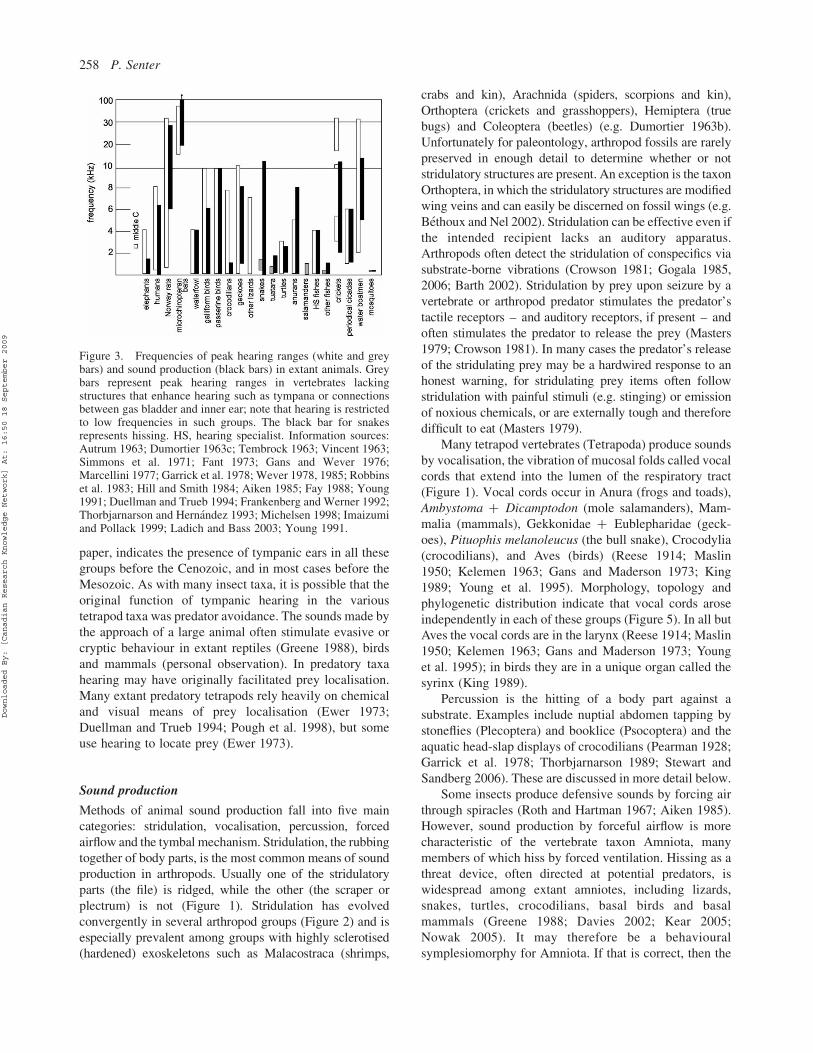
Figure 3. Frequencies of peak hearing ranges (white and greybars) and sound production (black bars) in extant animals. Greybars represent peak hearing ranges in vertebrates lackingstructures that enhance hearing such as tympana or connectionsbetween gas bladder and inner ear; note that hearing is restrictedto low frequencies in such groups. The black bar for snakesrepresents hissing. HS, hearing specialist. Information sources:Autrum 1963; Dumortier 1963c; Tembrock 1963; Vincent 1963;Simmons et al. 1971; Fant 1973; Gans and Wever 1976;Marcellini 1977; Garrick et al. 1978; Wever 1978, 1985; Robbinset al. 1983; Hill and Smith 1984; Aiken 1985; Fay 1988; Young1991; Duellman and Trueb 1994; Frankenberg and Werner 1992;Thorbjarnarson and Hernandez 1993; Michelsen 1998; Imaizumiand Pollack 1999; Ladich and Bass 2003; Young 1991.
P. Senter258
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

hiss was present by the Pennsylvanian Period (Ruta et al.
2003), at which time no vertebrate predator had yet
acquired tympanic ears (Clack and Allin 2004).
A Pennsylvanian hiss therefore had no value as an
acoustic display unless it exhibited the extraordinary
intensity necessary to be heard with atympanic ears.
If hissing is a behavioural symplesiomorphy of Amniota,
its sound may therefore have originally been the passive
result of a visual, inflationary display, as is the case with
some extant reptiles (Kinney et al. 1998). A hiss can be
produced during inspiration (Kinney et al. 1998; Young
1998) or expiration (Gans and Maderson 1973; Young
1998). Either way, forced inspiration occurs, and this
increases apparent body size in an amniote with movable
ribs (e.g. Kinney et al. 1998). In turtles and birds, hissing is
not accompanied by bodily expansion because their
ribcages do not allow it. However, hissing in the
earliest turtles and birds already had value as an acoustic
display because tympanic ears were present in a variety of
tetrapod predators by the time turtles and birds appeared
(reviewed below).
Tymbals, which are unique to Hemiptera (true bugs),
are series of external folds on the first abdominal segment.
Tymbals are buckled by internal muscles to produce
substrate-borne vibrations (Leston and Pringle 1963;
Gogala 1984; Claridge 1985). In cicadas (Cicadidae),
discussed more fully below, tymbals are coupled with air
bladders to produce airborne sounds.
Figure 4. Phylogeny of vertebrates discussed in this paper, showing independent origins of tympanic ears (T). Extant taxa are indicatedwith common names in parentheses. See text for information sources.
Historical Biology 259
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

Temporal patterns
Across taxa, certain temporal patterns in sound production
are nearly universal. Animals tend to be noisiest at dawn,
dusk and night, with very little sound production during
the day (Schneider 1967; Marcellini 1977; Garrick et al.
1978; Welty and Baptista 1988; Thorbjarnarson and
Hernandez 1993; Zelick et al. 1999; McCauley and Cato
2000; Gerhardt and Huber 2002). A circannual pattern is
also present across taxa, with more sound production
during summers than during winters (Garrick et al. 1978;
Aiken 1985; Thorbjarnarson and Hernandez 1993; Morton
1996; McCauley and Cato 2000; Gerhardt and Huber
2002). There is no reason to believe that such temporal
patterns were absent before the Cenozoic.
Parallel evolution
Certain body plans are more conducive than others to the
evolution of certain sound-producing or -receiving organs,
and parallel evolution of such structures is common in taxa
with appropriate body plans. For example, posterior and
internal to the tetrapod cheek is a bone called the stapes
(Figure 1) that primitively served as a brace between cheek
and braincase (Clack 1992). Due to its opportune location
the stapes has been coupled with an external tympanum for
transmission of airborne sound to the inner ear in several
tetrapod lineages independently (Clack and Allin 2004)
(Figure 4). Another example of acoustic parallelism in
tetrapods is the parallel evolution of sound-producing
vocal cords within the larynx (Maslin 1950; Kelemen
1963; Gans and Maderson 1973) (Figure 5).
The plesiomorphic presence of laryngeal muscles that
can be evolutionarily co-opted for vocal modulation
makes the larynx a particularly appropriate organ to
modify for vocalisation. The appearance of elytra in
Coleoptera (beetles) and the sclerotisation of the beetle
exoskeleton have also encouraged repeated parallel
evolution of stridulatory structures in Coleoptera, in
which many stridulatory structures are modifications of the
elytra-closing mechanism (Dumortier 1963b). Other
examples of acoustic parallelism include the repeated
independent appearance of trichobothria (sensory hairs
that respond to airborne sounds) within Arachnida (spiders
and kin) and Hemiptera (true bugs), tibial tympana in
Orthoptera (crickets and kin), use of wings for stridulation
in Orthoptera and use of chelipeds for stridulation in
Malacostraca (shrimp and kin) (Dumortier 1963b;
Gwynne 1995; Sweet 1996; Barth 2002; Bethoux and
Nel 2002; Desutter-Grandcolas 2003).
Caveats
Sound-producing structures often fossilise poorly or not at
all. Because of this the inference that a given animal sound
was present during a given geological period is rarely
certain. If a sound producing structure is present in all
extant members of a given crown clade, then it is most
parsimonious to infer that the structure was present in the
common ancestor of the clade. However, confidence in
such inferences cannot always be absolute, because in
some cases the prevalence of acoustic parallelism renders
hypotheses of homology of acoustic structures and
behaviour uncertain even among closely related species.
Even when such an inference is correct, it places only a
tentative bound on the time of origin of the structure,
because the crown clade may have appeared earlier than its
earliest known fossils and also because it may be
impossible to determine whether the structure was present
in fossil members of the lineage that are basal to the crown
clade. Another compounding factor is the fact that some
taxa with extant members are not defined as crown clades.
The earliest members of such taxa therefore do not
necessarily possess the features characteristic of the crown
clade. For these reasons the paleobioacoustical inferences
presented here must be treated as hypotheses to test, rather
than firm conclusions. Nevertheless, available evidence
Figure 5. Phylogenetic distribution of vocal cords in the larynx(L) and syrinx (S) of extant jawed vertebrates. Note the largenumber of intervening taxa without vocal cords, which indicatesmultiple independent origins of vocal cords.
P. Senter260
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

allows enough inferences to be made to make paleobioa-
coustical studies worthwhile.
Pre-Cenozoic paleobioacoustics of non-insect
invertebrates
Incidental sounds
Extant squid make a popping sound that appears to be due
to fluttering of mantle lips during expulsion of water from
the siphon (Iversen et al. 1963). Although cephalopods are
the only invertebrates outside Arthropoda that possess
organs for unambiguous underwater sound reception
(modified statocysts) (Coffin et al. 2004), there is no
evidence that their locomotor sounds are involved in
communication. Cephalopods appeared during the Upper
Cambrian and stem-group squid (Teuthidea) during the
Upper Triassic (Doyle 1993; King 1993). Squid popping
sounds may therefore have existed as early as the Upper
Triassic, and such sounds may have been present as early
as the Cambrian if other cephalopods made them.
However, no extant teuthid family has a known pre-
Cenozoic fossil record (Doyle 1993), so if popping sounds
are restricted to crown-group squid such sounds may have
been absent from the Mesozoic.
Saltwater mussels (Mollusca: Bivalvia: Mytilidae)
attach themselves to the substrate with protein secretions
from the foot called byssal threads. When a mussel uses its
foot for locomotion, the stretching and breaking of the
byssal threads make loud snapping sounds (Fish and
Mowbray 1970). Summation of the snaps produced by the
various colony members produces a continuous crackling
sound (Fish and Mowbray 1970). The family Mytilidae is
known from as early as the Lower Triassic (Skelton and
Benton 1993). Members of several other bivalve families
use byssal threads as anchors, but only in Mytilidae are the
threads used in locomotion (Ruppert and Barnes 1994), so
we should not expect that the snapping sounds were
present before the appearance of mytilids.
Colonies of sea urchins (Echinodermata: Echinoidea)
make sustained crackling sounds that have been likened to
the sound of something frying (Fish and Mowbray 1970).
Individuals contribute to the sound by movement of spines
and possibly by action of the calcareous feeding apparatus
called Aristotle’s lantern (Fish and Mowbray 1970). Sea
urchins are known from as early as the Upper Ordovician;
their diversity exploded during the Jurassic Period
(Kier 1987), at which time their frying sounds presumably
became more prevalent.
Extant barnacles (Arthropoda: Cirripedia) make crack-
ling sounds during feeding as their appendages scrape
against their calcareous shells (Budelmann 1992a,b). These
sounds are often continuous due to high population density
(Dumortier 1963b). Convergent acquisition of calcareous
shells, and presumably crackling sounds, by various
lineages of barnacles occurred in the Triassic Period and
continued through the Mesozoic (Foster and Buckeridge
1987). Paleozoic barnacles lacked calcareous shells (Wills
1963; Schram 1975; Collins and Rudkin 1981) and
therefore probably did not produce the crackling sounds.
The appendages of non-sessile arthropods sometimes
produce low-amplitude clicking or ticking sounds during
locomotion when the appendages contact the substrate or
other body parts. Arthropods are known from early in the
Cambrian Period (Briggs et al. 1993), so their incidental
locomotor sounds may have been present that early.
Crustaceans (Arthropoda: Crustacea)
Many members of the crustacean taxon Malacostraca
(shrimp, lobsters and crabs) stridulate when seized
(Budelmann 1992a). The taxonomically erratic distri-
bution of malacostracan stridulatory structures, along with
their great variety and lack of transitional forms, suggests
multiple parallel origins of stridulation within the group
(Dumortier 1963b) (Table 1). While most malacostracans
stridulate with hard exoskeletal parts, spiny lobsters
(Palinuridae) stridulate by rubbing a soft plectrum at the
base of the antenna over a file beneath the eye; the
mechanism of the resulting rasping sound resembles that
of the bow and string of a stringed instrument (Patek
2001). The oldest known malacostracans are from the
Upper Devonian (Briggs et al. 1993). The bauplan that
allowed repeated evolution of stridulation was therefore
present that early, and it is possible that anti-predator
Table 1. Malacostracan crustacean families that have stridulating members and are known from before the Cenozoic, and theirstridulatory organs.
Taxon Earliest record Location of scraper Location of file
Calappidae (box crabs) Lower Cretaceous Cheliped CephalothoraxDiogenidae (left-handed hermit crabs) Upper Cretaceous Left cheliped Right chelipedLysiosquillidae (mantis shrimp) Upper Cretaceous Uropods Lower surface of telsonPalinuridae (spiny lobsters) Lower Jurassic Base of antenna Antennular plate below eyePenaeidae (prawns) Lower Triassic First abdominal segment Posterior CephalothoraxSquillidae (mantis shrimp) Upper Cretaceous Uropods Lower surface of telsonXanthidae (mud crabs) Lower Cretaceous Cheliped Walking legs
References: Dumortier 1963b; Briggs et al. 1993; Patek 2001.
Historical Biology 261
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

stridulation evolved multiple times in Paleozoic and
Mesozoic malacostracan lineages. Stridulatory structures
have been identified on the flanks of members of the
Cretaceous palinurid lobster genus Linuparus (Feldmann
et al. 2007).
A non-stridulatory malacostracan sound is the buzzing
of clawed lobsters (Nephropidae) when seized, accom-
plished by vibrating the carapace (Henniger and Watson
2005). The family is known from as early as the Middle
Jurassic (Briggs et al. 1993).
Arachnids (Arthropoda: Arachnida)
Stridulation in response to disturbance is present in a wide
variety of arachnids, including spiders (Araneae),
scorpions (Scorpiones), whip scorpions (Amblypygi),
windscorpions (Solifugae) and harvestmen (Opiliones)
(Dumortier 1963a,b; Uetz and Stratton 1981; Cloudsley-
Thompson and Constantinou 1984). While the sounds
produced by arachnid stridulation are often of low
intensity and cannot be heard further away than a few
centimetres, in some cases spider and scorpion stridulation
produces a loud buzz or hiss (Uetz and Stratton 1981;
McCormick and Polis 1990). Only six of the many extant
arachnid families with known stridulating members
(Dumortier 1963b; Uetz and Stratton 1981) have known
pre-Cenozoic fossil records, and all six are spider families
(Selden 1993; Penney et al. 2003) (Table 2). However, the
known pre-Cenozoic fossil record of Arachnida is so
spotty that many extant lineages probably existed long
before their known fossil records reveal (Penney et al.
2003). Also, the wide variety of arachnid stridulatory
structures (Dumortier 1963b; Uetz and Stratton 1981;
Hjelle 1990) indicates that parallel evolution of stridula-
tion is rampant in Arachnida, which in turn suggests high
likelihood that many extinct arachnids also stridulated in
response to disturbance. A patch of spines found on a limb
fragment of a Middle Devonian arachnid of unknown
affinity from New York may be a stridulatory structure
(Shear et al. 1987).
The earliest known arachnids are from the Silurian
(Jeram et al. 1990). The earliest known harvestman is from
the Devonian (Dunlop et al. 2003), and the earliest known
solifuge and amblypygid are from the Pennsylvanian
(Selden 1993; Dunlop 1994). The earliest terrestrial
scorpions are from the Devonian (Selden and Dunlop
1998). Basal spiders are known from as early as the Lower
Devonian, with representatives of extant clades present by
the Pennsylvanian (Mesothelae), Middle Triassic (Myga-
lomorphae) and Middle Jurassic (Araneomorphae) (Pen-
ney et al. 2003). Stridulation by or directed toward those
taxa may have been present by those times.
In addition to defensive stridulation, members of
several extant spider families stridulate during courtship or
interspecific aggression (Uetz and Stratton 1981), and the
same is conceivably true of members of extinct spider
families. Some spiders make courtship sounds by means
other than stridulation. Giant crab spiders (Sparassidae)
vibrate their appendages like tuning forks, and crab spiders
(Thomisidae) drum body parts against the substrate
(Uetz and Stratton 1981). Both families were present by
the Cretaceous Period (Grimaldi et al. 2002a,b).
Trichobothria, sensory hairs sensitive to air move-
ments, have evolved convergently in several extant
arachnid taxa (Sissom 1990; Barth 2002). The trichobo-
thria of spiders can detect low-frequency sounds of
10–950Hz, with peak sensitivity from 50 to 120Hz, and
spiders use this ability to locate flying flies and other prey
(Barth 2002). Extant scorpions use trichobothria to detect
air currents caused by prey movements (McCormick and
Polis 1990), but it is uncertain whether their trichobothria
are used for sound reception. Either way, Paleozoic
scorpions lacked trichobothria (Sissom 1990) and there-
fore did not locate prey by means of airborne sound.
Millipedes and centipedes (Arthropoda: Myriapoda)
Some myriapods stridulate in response to disturbance, and
in some cases detached appendages make creaking sounds
by an unknown mechanism, apparently to distract a
predator while the myriapod escapes (Dumortier 1963a,b).
Centipedes (Chilopoda) are known from as early as the
Upper Silurian (Jeram et al. 1990) and millipedes
(Diplopoda) from as early as the Lower Devonian
(Shear 1997). Defensive stridulation by myriapods,
directed at arachnid or centipede predators, may therefore
Table 2. Stridulating spider families that are known from before the Cenozoic, and their known stridulatory organs.
Taxon Earliest record Location of scraper Location of file
Araneidae (orb weavers) Upper Cretaceous Appendage AbdomenDipluridae (funnel-web tarantulas) Lower Cretaceous Pedipalp CheliceraLinyphiidae (sheet-web weavers) Lower Cretaceous Pedipalp Chelicera
Leg II Leg ISegestriidae (tube-dwelling spiders) Upper Cretaceous – –Theridiidae (comb-footed spiders) Cretaceous Abdomen ProsomaUloboridae (hackled orb weavers) Lower Cretaceous Pedipalp Chelicera
References: Uetz and Stratton 1981; Grimaldi et al. 2002a,b; Penney et al. 2003.
-----------------------------------------------------------
P. Senter262
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

have existed this early. However, airborne predator-
distracting sounds, such as the creaking of detached
appendages, would have been ineffective against deaf
predators and so were probably absent before terrestrial
tetrapods acquired the ability to hear airborne sounds in
the Permian Period (see below).
The earliest terrestrial animal communities, from the
Silurian Period, included centipedes and arachnids
(Jeram et al. 1990), both of which are predatory.
The earliest known millipedes and basal insects (Insecta)
appeared shortly thereafter, in the Lower Devonian
(Jeram et al. 1990; Shear 1997). Anti-predator stridulation
is known in extant members of all four taxa (Dumortier
1963a,b; Uetz and Stratton 1981) and may therefore have
existed in these early communities. Anti-predator stridula-
tion is most likely to have evolved first in taxa with
especially tough exoskeletons or in taxa capable of
producing unpleasant stimuli such as stings, bites, or
noxious emissions, so that it functioned as an honestwarning
to predators, as in extant arthropods (Masters 1979).
Directions for further research
To my knowledge, stridulatory structures have not been
described in fossil arthropods outside Insecta with the
exception of the Devonian arachnid and the Cretaceous
lobster mentioned above. A search for stridulatory
structures among fossil malacostracans, trilobites, eur-
ypterids and perhaps other pre-Cenozoic arthropods would
be worthwhile. It would also be informative to compare
sound-producing structures across Malacostraca and
Arachnida to determine whether any such structures are
homologous between families or genera.
Pre-Cenozoic insect paleobioacoustics
Stoneflies (Plecoptera)
Intraspecific acoustic communication is characteristic of
the extant stonefly clade Arctoperlaria (Stewart and
Sandberg 2006). After emergence, arctoperlarian stone-
flies perform male-female vibratory duets at aggregation
sites while the males search for the females (Stewart and
Sandberg 2006). The duets consist of vibrations with
species-specific rhythms, produced by audible tapping of
the abdomen on the substrate (Stewart and Sandberg
2006). Tapping duets are known in all extant arctoperlar-
ian families and appear to be a behavioural symplesio-
morphy of the clade (Stewart and Sandberg 2006). If this is
correct then the resulting sounds were present at the time
of origin of the arctoperlarian crown clade, the earliest
known fossils of which are from the Middle Triassic
(Grimaldi and Engel 2005).
Members of some extant stonefly families augment
the courtship tapping with scratching of the abdomen on
the substrate (producing a raspy sound), or rubbing
of specialised abdominal parts on the substrate (producing
a squeak) (Stewart and Sandberg 2006). However, none
of these families has a pre-Cenozoic fossil record
(Sinitshenkova 2002).
Crickets and kin (Orthoptera)
Males of several extant orthopteran taxa stridulate during
courtship by rubbing together specialised parts of the
wings, producing sounds with species-specific rhythms.
Such stridulation often stimulates chorusing and aggres-
sion from nearby males, and the approach of one male too
close to another may elicit a different temporal pattern of
stridulation called a disturbance song (Dumortier 1963a).
Stridulatory wing venation patterns are absent in Paleozoic
orthopterans, the earliest known of which are from the
Pennsylvanian (Bethoux and Nel 2002; Gorochov and
Rasnitsyn 2002). Stridulatory mechanisms on the wings
were present in the extinct, Upper Triassic orthopteran
family Mesoedischiidae and in members of Ensifera
(crickets) from the Upper Triassic and later (Bethoux and
Nel 2002; Gorochov and Rasnitsyn 2002). The earliest
known members of the extant stridulating ensiferan taxa
Gryllidae (true crickets) and Gryllotalpidae (mole crick-
ets) are from the Lower Cretaceous (Ross and Jarzem-
bowski 1993). Stridulation is also characteristic of
‘Prophalangopsidae’ (hump-winged and sagebrush crick-
ets), a taxon that is probably paraphyletic with respect to
Tettigoniidae (katydids) (Desutter-Grandcolas 2003).
‘Prophalangopsids’ are known from as early as the
Lower Jurassic, while katydids, grasshoppers (Acridoi-
dea), and other extant orthopteran families with stridulat-
ing members have no known pre-Cenozoic fossil record
(Ross and Jarzembowski 1993; Rust et al. 1999; Gorochov
and Rasnitsyn 2002).
Extant stridulating crickets simultaneously perceive
both the substrate-borne vibrations (via mechanoreceptors
in the legs) and the airborne sounds (via tibial tympana;
Figure 1) generated by the singing of conspecifics
(Kuhne et al. 1985). Morphological phylogenetic analyses
of extant ensiferans suggest that stridulatory structures and
tibial tympana were absent in the common ancestor of
extant orthopterans and have evolved in parallel a number
of times within the ensiferan crown group (Gwynne 1995;
Desutter-Grandcolas 2003). However, stridulatory wing
venation is present in all members of a long phylogenetic
series of Upper Triassic crickets that are basal to crown
Orthoptera (Bethoux and Nel 2002), which indicates that
stridulatory behaviour is plesiomorphic for Upper Triassic
and later crickets. This suggests that either stridulation was
lost once in the crown clade’s ancestor and then regained
multiple times by its descendants, or that it was present in
the crown clade’s ancestor and was lost multiple times
within the crown clade. The latter interpretation is
supported by a recent molecular phylogenetic analysis,
Historical Biology 263
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

the results of which differ greatly from those of
morphological phylogenetic analyses (Jost and Shaw
2006). Most of the non-stridulatory extant families are
fossorial (Gwynne 1995), and there may be a connection
between the acquisition of such a lifestyle and the loss of
stridulatory behaviour in crickets.
Gryllacrididae (locust crickets and leaf-rolling crick-
ets) are non-stridulating crickets without tympana, some of
which are fossorial. Males and females duet by drumming
the tarsus or abdomen on the substrate (Hale and Rentz
2001). This family is known from as early as the Upper
Cretaceous (Ross and Jarzembowski 1993).
Among extant orthopterans of various lineages,
females generally prefer male calls that are longer and
occur at higher rates (Gerhardt and Huber 2002).
If selective pressure for these traits has existed long
enough, then within each orthopteran lineage male calls
were originally of shorter duration and occurred at lower
rates than those of their extant descendants.
Rust et al. (1999) used comparison with extant
orthopterans to calculate the dominant frequency of the
stridulatory sounds produced by the wings of a Cenozoic
fossil katydid. The same may be possible with Mesozoic
orthopterans but has not yet been attempted.
Stridulatory wing structures are also present in members
of the extinct, Middle and Upper Triassic taxon Titanoptera
(¼Mesotitanida). This taxon is closely related to Orthoptera
and consists of large (wingspans up to 40 cm) insects from
Eurasia and Australia (Gorochov and Rasnitsyn 2002;
Grimaldi and Engel 2005).
Roaches (Blattaria)
Roachlike insects from as early as the Carboniferous
Period are often placed in Blattaria, but as a group
Paleozoic ‘roachoids’ are paraphyletic with respect to
Isoptera (termites), Mantodea (mantises), and Blattaria
(true roaches) (Grimaldi and Engel 2005). The earliest
known members of Blattaria sensu stricto are from the
Lower Cretaceous (Grimaldi and Engel 2005). Many
extant roaches make courtship or disturbance sounds by a
variety of means, but most such noisy roaches are
members of Blaberidae (Roth and Hartman 1967), which
is unknown before the Cenozoic (Grimaldi and Engel
2005). Members of other roach families are generally
silent, although one member of Blattellidae, a family
known from the Lower Cretaceous (Grimaldi and Engel
2005), makes a clicking sound before taking flight, and
another blattellid is known to squeak when caught
(Roth and Hartman 1967).
Termites (Isoptera)
Termite soldiers tap their heads on the substrate with a
rhythm that is recognised by conspecifics, in response
to disturbance, most likely as an alarm signal (Kirchner
et al. 1994). This behaviour is taxonomically widespread
within Isoptera and may therefore be a behavioural
symplesiomorphy for the group (Virant-Doberlet and Cokl
2004). Early termites are known from most continents in
the Lower Cretaceous, including Laurasian members of
the extant head-tapping family Termopsidae (Grimaldi
and Engel 2005).
Booklice (Psocoptera)
A widespread behaviour among booklice is the tapping of
the abdomen on the substrate by females to call males; it
makes a ticking or creaking sound (Pearman 1928).
The earliest known booklice are from the Upper Jurassic;
fossils from as early as the Lower Permian have been
attributed to Psocoptera but more likely belong to some
related taxon (Grimaldi and Engel 2005). Extant psocop-
teran families with tapping species and known pre-Cenozoic
fossil records include Psocidae (Upper Jurassic) and
Trogiidae (Upper Cretaceous) (Rasnitsyn 2002).
True bugs (Hemiptera)
Extant hemipterans comprise five major clades: Sternor-
rhyncha (aphids, scale insects and kin), Fulguromorpha
(planthoppers), Coleorrhyncha (peloridiid bugs), Prosor-
rhyncha (water bugs, assassin bugs, stink bugs and kin)
and Clypeomorpha (cicadas, leafhoppers, froghoppers and
kin). Members of the latter four clades possess tymbals,
which are buckled by internal muscles in species-specific
rhythms to produce vibrations called ‘songs’ during
courtship and, in some cases, during copulation
(Leston and Pringle 1963; Gogala 1984; Claridge 1985;
Sweet 1996). Tymbals are absent in Sternorrhyncha, and
their presence in the other four clades suggests that
the presence of tymbals is a synapomorphy that unites the
other four clades into a taxon (hereafter called the
‘tymbaled superclade’ below) that excludes Sternor-
rhyncha (Sweet 1996). That interpretation of hemipteran
phylogeny is supported by molecular systematics
(Campbell et al. 1995; von Dohlen and Moran 1995).
In most members of the tymbaled superclade the
tymbals are simple and do not produce airborne sound but
instead cause vibrations in the substrate, usually a plant
stem (Leston and Pringle 1963; Claridge 1985; Gogala
1984, 1985, 2006; Sweet 1996). Both sexes sing, lack
tympana and perceive the songs of conspecifics via
mechanoreceptors in the legs (Leston and Pringle 1963;
Claridge 1985; Gogala 1985, 2006; Sweet 1996). This
appears to be the plesiomorphic condition for the tymbaled
superclade (Sweet 1996). The earliest known members of
the tymbaled superclade, which is a crown clade, are from
the Lower Permian (Shcherbakov and Popov 2002), and
P. Senter264
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

the use of tymbals for substrate-borne courtship songs
presumably originated then.
Within the tymbaled superclade are several taxa in
which courtship is modified such that it produces airborne
sounds. The most familiar example is probably the treetop
singing of members of Cicadidae (cicadas), the earliest
known fossils of which are from the Upper Cretaceous
(Turonian) of New Jersey (Grimaldi et al. 2002b). In cicadas
the females have lost the tymbals, so singing is done only by
the males, in which the tymbals are elaborate in comparison
with those of other hemipterans (Leston and Pringle 1963;
Claridge 1985). Air sacs internal to the tymbals amplify the
songs so that they are audible from several meters away and
attract members of both sexes to a congregational area for
reproduction (Dumortier 1963a; Claridge 1985). Cicadas
hear each other’s songs with tympanic ears that are ventral
to the tymbals on the abdomen (Leston and Pringle 1963;
Claridge 1985). After attracting a female, a male cicada
changes the rhythm of his song; in some species he also adds
audible wing-flicking, and in some species the female
responds with wing-flicking of her own (Boulard 2006).
Some cicada species also have stridulatory structures, the
behavioural significance of which is unknown (Boulard
2006). Another enigmatic acoustic behaviour by cicadas is
the drumming of the wings on the substrate in some species,
especially in those that have lost tymbals altogether
(Boulard 2006).
In many members of Prosorrhyncha, the earliest
known members of which are from the Upper Triassic
(Shcherbakov and Popov 2002), tymbal songs are
augmented by simultaneous, audible stridulation, which
adds higher frequencies to the songs (Gogala 1984, 1985,
2006; Sweet 1996). Often, after the female has been
attracted by the male’s calling song, he switches to a
courtship song (Dumortier 1963a). The variety of
stridulatory structures in Prosorrhyncha (Table 3)
indicates that augmentation of tymbal songs with
stridulation arose multiple times in parallel within
Prosorrhyncha (Leston and Pringle 1963). Trichobothria
are present in many prosorrhynchans, fulguromorphs, and
coleorrhynchans, but not in clypeomorphs (Sweet 1996).
They appear to be used as acoustic receptors for low-
frequency, low-amplitude, audible song and differences
between the trichobothria of various taxa suggest that they
appeared in parallel multiple times in the tymbaled
superclade (Sweet 1996).
In the prosorrhynchan taxon Nepomorpha a switch has
occurred from the ancestral hemipteran plant-dwelling
habitat to an aquatic one (Shcherbakov and Popov 2002).
The new habitat required that plant stems be traded for the
watery environment as a medium to carry vibrational
signals to conspecifics. Accordingly, many nepomorphs
stridulate underwater. Among nepomorph families with
known pre-Cenozoic fossil records, stridulation is known
in Corixidae (water boatmen), Naucoridae (creeping water
bugs), Nepidae (waterscorpions), and Notonectidae (back-
swimmers) (Dumortier 1963b). In Corixidae and Noto-
nectidae stridulation by males is known to attract females
(Dumortier 1963a; Cokl et al. 2006). The variety of
nepomorph stridulatory structures (Table 3) suggests that
stridulation arose multiple times independently in
Nepomorpha. Because evolution of stridulation is rampant
in Nepomorpha, it may have been present in extinct
nepomorphs as early as the Upper Triassic, from which
time period the earliest nepomorphs are known (Shcher-
bakov and Popov 2002). Non-stridulatory courtship
sounds (buzzing or chirping) are made by members of
Belostomatidae (giant water bugs) by expulsion of air
Table 3. Prosorrhynchan families that are known from before the Cenozoic with members that are known to stridulate, and theirstridulatory organs.
Taxon Earliest record Location of scraper Location of file
Alydidae (broad-headed bugs) Upper Jurassic Femur HemelytraAradidae (flat bugs) Lower Cretaceous Hind femur Abdomen
Hind tibia AbdomenCoreidae (leaf-footed bugs) Upper Jurassic Pronotum Fore wingCorixidae (water boatmen) Lower Jurassic Fore tarsus Opposite fore femur
Fore femur HeadCydnidae (burrowing bugs) Lower Jurassic Abdomen Hind wingLygaeidae (seed bugs) Lower Cretaceous Wing Abdomen
Foreleg ProsternumMiridae (plant bugs) Upper Jurassic Hind femur EmboliumNabidae (damsel bugs) Upper Jurassic – –Naucoridae (creeping water bugs) Upper Triassic – –Nepidae (waterscorpions) Upper Jurassic Coxa FemurNotonectidae (backswimmers) Upper Triassic Fore femur Head or fore coxa
Tibia RostrumReduviidae (assassin bugs) Lower Cretaceous Rostrum ProsternumThaumastellidae (no common name) Lower Cretaceous Abdomen Wing
References: Dumortier 1963b; Leston and Pringle 1963; Gogala 1984, 1985, 2006; Aiken 1985; Ross and Jarzembowski 1993; Shcherbakov and Popov 2002.
----------------------
------------------------
-----------------------
-----------------------
--------------------------------------------------------
----------------------------------------------------
Historical Biology 265
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

through spiracles (Aiken 1985); belostomatids are known
from as early as the Upper Triassic (Ross and
Jarzembowski 1993).
In one nepomorph family, Corixidae (water boatmen) –
known from as early as the Lower Jurassic (Ross and
Jarzembowski 1993) – perception of conspecific stridula-
tion is truly auditory. Water boatmen receive such
vibrations with tympana that are surrounded by air bubbles,
whereas other nepomorphs use atympanic mechanorecep-
tors for reception of water-borne vibrations (Cokl et al.
2006). Unlike the low-amplitude calls of most nepomorphs,
which are audible to humans only if the bug is close to the
listener’s ear, the underwater songs of corixids are quite
loud; resonation by the same air bubbles that enable
tympanic hearing in corixids amplifies the songs so that they
are audible from beyond the shoreline (Aiken 1985).
Courtship sounds are not the only sounds made by
hemipterans. Many clypeomorphs cry audibly when
caught and during rough treatment (Dumortier 1963a;
Leston and Pringle 1963; Claridge 1985). Many prosor-
rhynchans stridulate audibly when caught, including
members of Cydnidae (burrowing bugs), Lygaeidae
(seed bugs) and Reduviidae (assassin bugs) (Dumortier
1963a; Leston and Pringle 1963; Gogala 2006). In some
reduviid species the female stridulates upon being
mounted, deterring the male, if she is unwilling to mate
(Lazzari et al. 2006). Members of the prosorrhynchan
family Coreidae (leaf-footed bugs), the earliest known
members of which are from the Upper Jurassic (Ross and
Jarzembowski 1993), make sounds by unknown means
(Dumortier 1963b); the biological significance of these
sounds is unclear, and they are probably by-products of
locomotion (Gogala 1985). Among members of Sternor-
rhyncha, some colonies of aphids (Aphididae) make
scraping sounds when disturbed (Leston and Pringle
1963). These stridulatory sounds are made by rubbing the
hind tibiae on rough spots on the abdomen (Dumortier
1963a). The earliest known aphids are from the Lower
Cretaceous (Ross and Jarzembowski 1993).
Stridulatory structures are known in some fossil
hemipterans. Members of the stem-hemipteran family
Dysmorphoptilidae possessed a stridulatory apparatus
with a scraper on the hind knee and the file on the
underside of the fore wing; the family is known from the
Upper Permian to the Upper Jurassic (Shcherbakov and
Popov 2002). A similar stridulatory apparatus is present in
Triassic members of the extinct prosorrhynchan family
Ipsviciidae (Shcherbakov and Popov 2002).
Beetles (Coleoptera)
Beetles of many extant families produce sounds, often by
stridulation (Alexander and Moore 1963; Dumortier
1963b). Adult and larval beetles often stridulate in response
to disturbance, although some adult males stridulate
to attract females or repel rival males (Dumortier 1963a;
Rudinsky and Ryker 1976; Aiken 1985). The broad
taxonomic span of families with stridulating members and
the wide variety of stridulatory structures shows that
repeated parallel evolution of stridulation is rampant in
Coleoptera (Table 4). Beetle stridulationmay therefore have
been present as early as the Lower Permian, from which the
earliest beetle fossils are known (Ponomarenko 2002).
Various beetles are also known to make non-
stridulatory sounds. Female members of Anobiidae
(death-watch beetles), a family known from as early as
the Lower Cretaceous (Ross and Jarzembowski 1993),
attract males by drumming the head on the substrate
(Dumortier 1963a). In some members of Buprestidae
(metallic wood-boring beetles), a family known from as
early as the Middle Jurassic (Ross and Jarzembowski
1993), members of both sexes drum their abdomens on the
substrate and answer each other’s drumming in like
manner (Crowson 1981). Drumming of body parts against
the substrate is also known in Cerambycidae (long-horned
beetles) (Crowson 1981), the earliest known members of
which are from the Lower Cretaceous (Ross and
Jarzembowski 1993). Some cerambycids also make
purring sounds by vibrating the body within a crevice in
bark (Dumortier 1963b). Larvae of some members of
Dytiscidae (predaceous diving beetles), a family known
from as early as the Upper Jurassic (Ponomarenko 2002),
squeak by expelling air through spiracles when disturbed
(Aiken 1985). Larvae of some members of Hydrophilidae
(water scavenger beetles), a taxon known from as early as
the Upper Triassic (Ponomarenko 2002), hiss by an
unknown mechanism upon disturbance (Aiken 1985).
Members of Elateridae (click beetles), produce a loud
click by snapping a spine on the prosternum into a notch
on the mesosternum, which suddenly flexes and bounces
the beetle and is used to right the beetle when upside-down
or to make it difficult for predators to hold (Grimaldi and
Engel 2005); the earliest known elaterids are from the
Upper Triassic (Ponomarenko 2002). The wings of
members of Scarabaeidae (scarab beetles), a family
known from as early as the Lower Jurassic (Ponomarenko
2002), often buzz loudly during flight. The exoskeletons of
these beetles are particularly tough, so it is possible that
the loud scarabaeid flight buzz is an honest warning of
culinary difficulty to potential predators.
Lacewings and kin (Neuropterida)
The supraordinal insect taxon Neuropterida includes the
order Raphidioptera (snakeflies) and the sister orders
Megaloptera (dobsonflies and alderflies) and Neuroptera
(lacewings). Courtship in all extant families of Raphi-
dioptera and Megaloptera involves abdominal tremulation,
sensed via substrate- (stem- or leaf-) borne vibrations with
species-specific rhythms; in Corydalidae (dobsonflies)
P. Senter266
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

only the males tremulate, but in Sialidae (alderflies) and
Raphidioptera the sexes perform tremulatory duets
(Henry 2006). While tremulation does not produce
airborne sounds, it is often accompanied by audible wing
fluttering in Raphidioptera or by audible drumming of the
abdomen on the substrate in Sialidae, and is replaced by
audible wing fluttering in some species of Corydalidae
(Henry 2006).
Among lacewings, wing fluttering during courtship is
known in Coniopterygidae (dustywings) and Berothidae
(beaded lacewings) (Henry 2006), known from as early as
the Upper Jurassic and the Lower Cretaceous respectively
(Grimaldi and Engel 2005). Tremulation, wing fluttering
and abdominal drumming during courtship are all known
in Chrysopidae (green lacewings) (Henry 2006), a family
known from as early as the Lower Cretaceous (Grimaldi
and Engel 2005).
Among lacewings, tremulation is absent in all but three
derived families, the relationships between which suggest
an independent origin of tremulation in each (Henry 2006).
The presence of tremulation in Raphidioptera and
Megaloptera but not basal Neuroptera makes it unclear
whether tremulation appeared independently in the three
neuropterid orders or was present in the common
neuropterid ancestor, lost in ancestral lacewings, and
then regained in derived lacewings. If tremulation was
present in the neuropterid ancestor, then it (and possibly
the accompanying sounds of drumming and/or wing
fluttering) existed as early as the Permian Period, from
which the earliest neuropterid fossils are known
(Grimaldi and Engel 2005). If tremulation evolved
separately in the three orders, then the times of origin of
its separate appearances are less clear. Snakeflies are
known from the Lower Jurassic, but the earliest known
member of an extant family is from the Tertiary (Grimaldi
and Engel 2005). Megalopterans are known from as early
as the Upper Triassic, but extant megalopteran families are
unknown before the Middle Jurassic (Grimaldi and Engel
2005). Estimated times of origin of tremulation and
accompanying sounds in Raphidioptera and Megaloptera
therefore depend on whether tremulation was present in
each order’s ancestor or only in members of extant
tremulating families. It is plausible that the neuropterid
bauplan encouraged repeated evolution of tremulation,
just as the beetle bauplan encouraged repeated evolution of
stridulation. It is therefore possible that these neuropterid
Table 4. Beetle families that are known from before the Cenozoic in which adults are known to stridulate, and their stridulatory organs,where known.
Taxon Earliest record Location of scraper Location of file
Anobiidae (death-watch beetles) Lower Cretaceous Prosternum GulaBruchidae (seed beetles) Upper Jurassic – –Buprestidae (metallic wood-boring beetles) Upper Jurassic – –Carabidae (ground beetles) Lower Jurassic Elytra Abdomen
Femur ElytraCerambycidae (long-horned beetles) Lower Cretaceous Pronotum Mesonotum
Elytra HindfemurMetasternum Coax
Chrysomelidae (leaf beetles) Upper Jurassic Prosternum GulaPronotum MesonotumElytra Abdomen
Curculionidae (weevils) Lower Cretaceous Elytra AbdomenAbdomen Elytra
Dytiscidae (predaceous diving beetles) Upper Jurassic Wing AbdomenEndomychidae (handsome fungus beetles) Upper Cretaceous Pronotum HeadHeteroceridae (variegated mud-loving beetles) Lower Cretaceous Femur AbdomenHydrophilidae (water scavenger beetles) Upper Triassic Elytra AbdomenLucanidae (stag beetles) Upper Jurassic Elytra HindfemurNemonychidae (no common name) Upper Jurassic – –Nitidulidae (sap beetles) Middle Jurassic Pronotum HeadPassalidae (bessbugs) Lower Cretaceous Abdomen WingScarabaeidae (scarab beetles) Lower Jurassic Elytra Abdomen
Elytra HindfemurCoax MetasternumAbdomen ElytraAbdomen WingFemur Elytra
Scolytidae (bark beetles and ambrosia beetles) Lower Cretaceous Prosternum GulaAbdomen Elytra
Silphidae (carrion beetles) Lower Jurassic Elytra Abdomen
References: Dumortier 1963b; Ross and Jarzembowski 1993; Ponomarenko 2002; Wessel 2006.
------------------------------------------------------
--------------------------------------------------
----------------------------------------
-----------------------------------------------------------------------------
-----------------------
---------------------------------------------------------------------------------------------------------
----------------------------------------------
-----------------------------------------------
-----------------------
Historical Biology 267
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

behaviours and their accompanying sounds appeared
multiple times before and during the Cenozoic.
Wasps, bees and ants (Hymenoptera)
Wasps (Hymenoptera) are known from as early as the
Upper Triassic (Grimaldi and Engel 2005). The wasp clade
Aculeata, known from as early as the Upper Jurassic, is
characterised by modification of the ovipositor into a sting
(Grimaldi and Engel 2005). Aculeates tend to buzz audibly
in flight, and it is possible that, as with the stridulatory
sounds of certain other insects, the flight fuzz deters
potential predators by warning them of the painful
consequences of predatory attempts.
Aculeate flight sounds also serve social functions. Some
male braconid wasps (Braconidae) also use their wings to
make courtship sounds when near females (Sotavalta 1963);
braconids are known from as early as the Lower Cretaceous
(Ross and Jarzembowski 1993). Members of Vespidae,
which includes hornets and kin, are known to buzz with
their wings during flight and during copulation (O’Neill
2001); vespids are known from as early as the Lower
Cretaceous (Ross and Jarzembowski 1993). In some
members of Sphecidae (mud daubers and kin), a family
known from as early as the Lower Cretaceous (Ross and
Jarzembowski 1993), males use buzzing as an acoustic
threat when defending the nest against marauders or rival
males (O’Neill 2001); males of some sphecid species also
locate virgin females by the loud buzz made by the females
as they dig themselves out of the ground. The buzzing of
bees (Apoidea) cannot be heard by fellow bees during flight,
but in some bees, once females have alighted, males use the
flight buzz to court them (Sotavalta 1963). The earliest
known bee is from the Lower Cretaceous (Poinar and
Danforth 2006). Reports of pre-Cretaceous bee body and
trace fossils exist, but the former have been reidentified as
indeterminate insects and the latter are more likely beetle
than bee traces (Engel 2001).
A variety of sounds are known in extant bees,
particularly those of the closely related tribes Apini
(honeybees and kin), Bombini (bumblebees and kin) and
Meliponini (stingless bees). Sounds that have been
described as tooting, quacking, hissing and piping serve
various social functions in Apini and Bombini (Hrncir et al.
2006), but neither taxon is known from before the Cenozoic
(Engel 2001). Stingless bees are known from the Upper
Cretaceous (Engel 2001). Stingless bees share with Apini
and Bombini the tendency for foragers to use thoracic
pulsation to make sounds that stimulate nestmates to fly out
– sometimes after first buzzing in response – and collect
food (Hrncir et al. 2006). If thoracic pulsation is a
behavioural symplesiomorphy for stingless bees, honeybees
and bumblebees, then it was present at least as early as the
Upper Cretaceous. Bees lack tympanic ears and sense the
sounds of conspecifics via substrate-borne vibrations and via
mechanoreceptors in the antennae that detect air movement
caused by the vibrations of the wings of conspecifics
engaging in thoracic pulsation (Hrncir et al. 2006).
Ants (Formicidae) are also members of Aculeata.
The earliest known ants are from the Cretaceous Period
and include members of four extant subfamilies:
Ponerinae (trap-jaw ants and kin), Dolichoderinae (cone
ants and kin), Formicinae (carpenter ants, wood ants and
kin), and Myrmicinae (harvester ants, fire ants and kin)
(Moreau et al. 2006). The earliest known fossil records of
the former two subfamilies are from the Lower
Cretaceous, and those of the latter two are from the
Upper Cretaceous (Moreau et al. 2006). Some dolicho-
derine and formicine ant workers tap their mandibles and
abdomens on the substrate to produce alarm sounds in
response to nest disturbance (Holldobler and Wilson
1990). Stridulation is used by ponerine and myrmicine
ants – especially in soil-dwelling species – for group
cohesion, warning of danger, as a call for help by wounded
or imperiled individuals, and as a signal by females to
males that copulation is complete (Dumortier 1963a;
Holldobler and Wilson 1990). Stridulation occurs by
rubbing the last segment of the petiole against the striated
dorsal edge of the first segment of the gaster in Ponerinae
and Myrmicinae (Dumortier 1963b); in some myrmicines
this is coupled with the rubbing of a second, ventral
stridulatory apparatus at the joint between the same two
segments (Dumortier 1963b). In small ant species
stridulation is inaudible to humans further away than a
few centimetres, but in large members of Ponerinae and
Myrmicinae it can be audible further away (Dumortier
1963b,c). Ants themselves are deaf to airborne sounds and
detect conspecific stridulation via substrate-borne
vibrations (Holldobler and Wilson 1990).
Stridulation between edges of abdominal segments is
also known in some members of Mutillidae (velvet-ants)
(Dumortier 1963b; O’Neill 2001), an aculeate family
known from as early as the Upper Cretaceous (Ross and
Jarzembowski 1993). In some species the male makes a
honking sound by vibrations of the wings and thorax as he
approaches the female, and both members of the pair
alternately stridulate during copulation (O’Neill 2001).
Flies, gnats and mosquitoes (Diptera)
Dipterans are known from as early as the Middle Triassic
(Blagoderov et al. 2002). Audible wing vibration by the
male during mounting is widespread in Diptera (Kanmiya
2006). In addition, many dipterans use flight sounds as a
means of intraspecific communication. Male mosquitoes
(Culicidae) have a hearing apparatus in the antennae,
which is used to find the female by her flight hum and to
keep together in all-male swarms (Sotavalta 1963); the
earliest known mosquitoes are from the Cretaceous
(Grimaldi et al. 2002a,b). Male hover flies (Syrphidae)
P. Senter268
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

court females by flying with a high-pitched hum over a
female on a flower (Sotavalta 1963); the earliest known
hover flies are from the Upper Cretaceous (Blagoderov
et al. 2002).
Other dipteran families with audible flight sounds and
known pre-Cenozoic fossils include Bombyliidae (bee
flies), Tabanidae (horse flies), and bottle flies (Calliphor-
idae). The oldest known members of Bombyliidae and
Tabanidae are from the Lower Cretaceous, and the oldest
known member of Calliphoridae is from the Upper
Cretaceous (Blagoderov et al. 2002). In some dipterans,
morphology and flight sounds mimic those of stinging
hymenopterans, apparently to deter predators (Chapman
1975). Such mimicry is unlikely to have arisen before the
appearance of stinging hymenopterans, which occurred in
the Upper Jurassic (Grimaldi and Engel 2005).
Directions for further research
Restudy of fossil insects – especially those that are well
preserved in amber – to search for potentially overlooked
bioacoustical structures might be worthwhile. A search for
stridulatory structures, highly sclerotised ventral surfaces
(for percussion), and other sound-producing structures
could prove informative. Phylogenetic studies to deter-
mine homologies between sound-producing parts could be
used to constrain phylogenetic levels at which homologous
sound production occurred within given insect taxa, and
fossils could then be used to constrain the times of origin
of such sounds. For this, a set of reliable phylogenies
incorporating these fossil taxa would be necessary to
determine historical homology as opposed to convergence.
It would also be interesting to examine sound-producing
and auditory parts of extant insects for correlations
between morphology and dominant frequencies, and to
apply the results to fossils to determine more precisely the
nature of the sounds produced and heard by fossil insects
(e.g. Rust et al. 1999).
Pre-Cenozoic vertebrate paleobioacoustics
Non-bony fishes
In vertebrates the inner ear exhibits a number of separate
neuromast organs, some of which are used for 3D
orientation and some of which are used for sound reception
(Ladich and Popper 2004). Sound reception has not been
demonstrated in the basal taxa Myxini (hagfishes) and
Petromyzontida (lampreys) but is present in Gnathosto-
mata (jawed vertebrates) (Ladich and Popper 2004). It may
have appeared independently in cartilaginous fishes
(Gnathostomata: Chondrichthyes) and bony fishes
(Gnathostomata: Osteichthyes) because the former use a
neuromast organ called the macula neglecta for hearing,
whereas in the latter the macula neglecta is reduced
or absent and other parts of the inner ear are used for
hearing (Popper et al. 1992; Ladich and Popper 2004).
Cartilaginous fishes are generally silent (Hueter et al.
2004). An exception is the cownose ray (Rhinoptera
bonasus), which emits clicks in response to a human touch
(Fish and Mowbray 1970). The cownose ray is a member
of the family Myliobatidae, the earliest known members of
which are from the latest Cretaceous (Campanian-
Maastrichtian) (Cappetta 1987).
Hearing has been demonstrated in Elasmobranchii
(sharks and rays) but has not been studied in Holocephali
(ratfishes). Elasmobranchs are known from as early as the
Lower Devonian (Cappetta et al. 1993), so it is possible
that anti-predator sounds directed at them existed by the
late Paleozoic.
Lobe-finned fishes (Osteichthyes: Sarcopterygii)
Extant lungfishes (Sarcopterygii: Dipnoi) are generally
silent, although the Australian lungfish (Neoceratodus
forsteri) is known to breathe air noisily and to do it more
frequently during the breeding season, sometimes in
concert in the evening (Kemp 1986). The sound is more
likely incidental than of use in intraspecific communi-
cation, because lungfishes lack specialisations for airborne
sound reception. Incidental air-breathing sounds that
resemble whistling and clucking are also known in extant
salamanders (Maslin 1950), so the phenomenon is not
limited to sarcopterygians outside Tetrapoda. Sarcopter-
ygians are known from as early as the Upper Silurian
(Zhu et al. 2009), so the incidental sounds of air their
breathing may have existed that early.
Ray-finned bony fishes (Osteichthyes: Actinopterygii)
Many actinopterygian taxa produce grunts, clicks, squeaks,
whistles and other sounds for intraspecific communication
and when disturbed or apprehended (Fish and Mowbray
1970; Myrberg 1981; Ladich and Bass 2003). Such sounds
are typically produced by muscular vibration of the gas
bladder, but in some species sound production is
accomplished by tendon snapping, pectoral girdle vibration,
stridulation of pharyngeal teeth, or other methods (Ladich
and Bass 2003). Actinopterygian acoustic behaviour has
been studied mainly in teleosts (Teleostei). Not all teleosts
are known to make sounds, and the variety of their sound
production mechanisms indicates that acoustic communi-
cation evolved independently in several lineages (Schneider
1967; Ladich and Bass 2003). In addition to teleosts,
acoustic communication is known in basal ray-finned
groups such as sturgeons (Acipenseridae) and bichirs
(Polypteridae) (Johnston and Phillips 2003; Ladich and
Bass 2003). Although the known fossil records of extant
ray-finned fish families that communicate acoustically go
back no further than the Upper Jurassic (Table 5), those
Historical Biology 269
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

families phylogenetically bracket taxa from as early as the
Upper Devonian (Carroll 1988; Grande and Bemis 1996).
The sounds of actinopterygian communication may there-
fore have existed through the late Paleozoic and Mesozoic.
Basal ray-finned fishes must have been present by the
Upper Silurian, because Sarcopterygii, the sister taxon to
Actinpterygii, was present then (Zhu et al. 2009). The lungs
of extant basal actinopterygians may be homologous with
those of lungfishes (Kardong 2006), in which case the
sound of noisy air-breathing by ray-finned fishes may have
existed alongside that of their lobe-finned counterparts as
early as the Upper Silurian.
Underwater hearing is present in actinopterygians in
general. A number of teleost taxa, known as hearing
specialists, independently evolved connections between
the gas bladder and the inner ear (Ladich and Bass 2003).
This increases the upper frequency range through which
underwater sounds can be heard (Yan et al. 2000; Ladich
and Bass 2003). Accordingly, sounds made by hearing
specialist fishes are of higher frequency than those of other
fishes (Figure 3) (Ladich and Bass 2003). Hearing
specialist taxa include Otophysi (carp, catfishes and kin),
Anabantoidea (gouramis and kin), Mormyridae (mormyr-
ids) and Holocentridae (squirrelfishes) (Ladich and Bass
2003). Hearing specialist fishes typically inhabit quiet,
shallow water, which enhances high-frequency sound
transmission (Ladich and Bass 2003). It stands to reason,
then, that high-frequency acoustic behaviour of fossil
actinopterygians would have been more prevalent in quiet,
shallow water than in noisy or deep water. The earliest
known members of extant hearing specialist fish taxa are
from the Upper Cretaceous (Table 5).
Basal tetrapods (Tetrapoda)
In basal tetrapods a bone called the stapes served as a brace
between the palate and braincase (Clack 1992). The stapes of
most extant frogs (Anura) and amniotes (Amniota) has been
reoriented so that it couples the inner ear to a tympanum,
enabling detection of airborne sounds. The sound-sensitive
region of the inner ear is homologous in Anura and Amniota
(Fritzsch 1992) but its coupling to a tympanum evolved
separately in the ancestors of extant frogs, diapsids
(Amniota: Diapsida) and mammals (Amniota: Mammalia)
(Bolt and Lombard 1985; Carroll 1991; Clack and Allin
2004) (Figure 4). The earliest tetrapod skulls exhibit
posterior notches that are often called otic (ear) notches
because they superficially resemble the posterior skull
notches that house tympana in extant frogs and amniotes.
However, early tetrapod stapedial and otic morphology are
inconsistent with tympanic hearing (Clack et al. 2003).
The bony architecture of the notches indicates that they
housed spiracles, not tympana (Brazeau and Ahlberg 2006).
Even so, early tetrapod inner ears probably detected low-
frequency water- and substrate-borne sounds, as do the
atympanic ears of extant fishes, salamanders and caecilians
(Smotherman and Narins 2004; McCormick 1999; Ladich
and Bass 2003).
The early tetrapod Ichthyostega exhibits otic modifi-
cations analogous to those of hearing-specialist fishes.
Uniquely among basal tetrapods, its skull appears to have
had an air-filled otic chamber and a mobile stapes with a
morphology and position conducive to sound conduction
from the air-filled chamber to the inner ear (Clack et al.
2003). The otic chamber apparently functioned as a
resonator for high-frequency sounds, as does the gas bladder
of hearing specialist fishes, which raises the possibility of
high-frequency sonic communication in Ichthyostega.
Among members of Lissamphibia (the smallest clade
including extant amphibians and all taxa phylogenetically
bracketed by them), tympanic ears are present inmost extant
frogs (Anura) but are absent in basal frogs (Ascaphidae and
Leiopelmatidae), caecilians (Gymnophiona) and salaman-
ders (Caudata) (Bogert 1958; Smotherman and Narins
2004). This suggests that the lissamphibian ancestor lacked
tympanic ears and that within Lissamphibia tympanic ears
are a synapomorphy of derived frogs. This interpretation is
compatible with all three of the competing hypotheses of
lissamphibian origins. According to one hypothesis,
Lissamphibia arose from within the otherwise Paleozoic
Table 5. Families of sound-producing fishes that are known from before the Cenozoic, and details about their sounds.
Family Earliest record Description of sound Function of sound Mechanism, if known
Acipenseridae (sturgeons) Upper Cretaceous Squeak, chirp, knock, groan – –Albulidae (bonefishes) Upper Cretaceous Click, scratch, knock R –Ariidae (HS) (marine catfishes) Upper Cretaceous Grunt, creak, yelp R EElopidae (tarpons) Upper Jurassic Thump R SGadidae (cods) Upper Cretaceous Grunt A, C, G, R INHolocentridae (HS) (squirrelfishes) Upper Cretaceous Staccato, grunt A, R, G EMyliobatidae (eagle rays) Upper Cretaceous Click R IOphidiidae (cusk-eels) Upper Cretaceous – – –Polypteridae (bichirs) Upper Cretaceous Thump, moan A, R I
A, aggression; C, courtship; E, contraction of extrinsic swim bladder muscles; G, group coordination; HS, hearing specialist; I, sound produced by unknown internal mechanism;IN, contraction of intrinsic swim bladder muscles; R, response to predator, human touch, or startling; S, sound made by swimming. References: Myrberg 1981; Cappetta 1987;Ladich and Tadler 1988; Arratia 1993; Meunier and Gayet 1993; Nolf and Stringer 1993; Patterson 1993; Schwarzhans 1993; Stewart 1993; Johnston and Phillips 2003; Ladichand Bass 2003; Wilson et al. 2003.
P. Senter270
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

clade Lepospondyli (Laurin and Reisz 1995, 1997, 1999;
Laurin 1998; Vallin and Laurin 2004). According to the
second hypothesis, Lissamphibia arose from within the
otherwise mostly Paleozoic and Triassic clade Temnospon-
dyli (Milner 1988; Trueb and Cloutier 1991; Ruta et al.
2003). According to the third hypothesis, frogs and
salamanders arose from within tymnospondyli but caeci-
lians arose from within Lepospondyli (Carroll 2007;
Anderson et al. 2008). Lepospondyls show no osteological
evidence of tympanic ears (Clack and Allin 2004). Many
temnospondyls exhibit posterior cranial notches that have
been interpreted as having housed tympana (Clack and Allin
2004). However, recent studies show that the morphology of
the temporal area and the massive size of the stapes in
temnospondyls are incompatible with the presence of a
tympanum; the notchesmore likely housed spiracles (Laurin
1998; Laurin and Soler-Gijon 2006). The common ancestor
of extant amphibians therefore lacked tympanic ears,
regardless of which phylogenetic hypothesis is correct.
Many large temnospondyls had large heads like those of
extant crocodilians (Romer 1947), which acoustically
advertise presence by slapping their heads on the water
(Vliet 1989; Thorbjarnarson 1991; Thorbjarnarson and
Hernandez 1993). It is therefore tempting to imagine large-
headed temnospondyls employing the same communication
device. Had they done so, their underwater hearing would
have detected the sound and their lateral line systems
(Romer 1947) would have detected the location of its source
(Coombs and Braun 2003), even without tympana.
However, the extreme proximity between shoulder girdle
and skull in temnospondyls and other non-amniote tetrapods
made the neck extremely short (Romer 1947), so it is
unlikely that they could have raised their heads enough to
have performed crocodilian-style head-slaps.
This does not necessarily mean that extinct temnos-
pondyls and other basal tetrapods were silent. It is possible
that they performed other activities involving audible
water displacement. Also, several extant salamander
species – which lack tympanic ears – produce airborne
anti-predator sounds in response to capture (Maslin 1950).
Such defensive sound production may have evolved
convergently in extinct basal tetrapods after the appear-
ance of tympanic ears in their predators. Many extant
lissamphibians that make such sounds produce noxious
chemicals that discourage or even kill predators (Duell-
man and Trueb 1994). Others, such as amphiumas
(Amphiumidae), can deliver a painful bite. Lissamphibian
anti-predator sounds may therefore be honest warnings to
predators (Duellman and Trueb 1994), as is the case with
stridulating insects (Masters 1979). Noxious chemical
production in extinct temnospondyls cannot be evaluated,
but cranial and dental morphology of many temnospondyls
indicates the ability to produce a nasty bite, so they may
have made honest warning sounds. While the earliest
known predators with tympanic ears are Lower Permian
seymouriamorphs (Ivakhnenko 1987) (Figure 6), basal
tetrapod anti-predator sounds directed at fishes may have
existed before then.
Mechanisms of defensive sound production in extant
salamanders are listed in Table 6. Among extant salamander
families, Cryptobranchidae (giant salamanders) and
Amphiumidae (amphiumas) have a pre-Cenozoic fossil
record. The former are known from the Middle Jurassic of
China (Gao and Shubin 2003) and the latter from the Upper
Cretaceous of North America (Milner 1993). It is therefore
possible that their characteristic defensive sounds existed
before the Cenozoic.
Arthropod stridulation in response to seizure by
tetrapod predators began at the earliest in the Mississip-
pian, at which time the earliest insectivorous tetrapods
Figure 6. Estimated times of origin of sounds and tympanic earsin various animal lineages. See text for information sources.RPE, rampant parallel evolution of.
Historical Biology 271
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

appeared (Hotton et al. 1997). Mississippian tetrapods
lacked tympanic ears (see below) and therefore would
have been deterred by the substrate-transmitted mechan-
ical vibrations produced by stridulation, rather than by the
resulting sounds. Predator deterrence by airborne stridu-
latory sounds would have been effective only after the
appearance of tympanic ears in predatory tetrapods in the
Permian (see below).
Frogs (Lissamphibia: Anura)
In lunged fishes, including lungfishes, the glottis (the
opening in the pharynx that leads to the lung) is ringed by a
muscular sphincter that closes the glottis (Negus 1949).
To this arrangement tetrapods have added the larynx
(Figure 1), an organ made of cartilage bodies (arytenoid
cartilages cranially and cricoid cartilages caudally) that
function as attachment sites formuscles that open the glottis
(Negus 1949;Kelemen 1963). In anurans paired vocal cords
extend into the lumen from the arytenoid part of the larynx
and are vibrated to produce vocalisation during forced
expulsion of air from the lungs (Kelemen 1963), or during
inspiration in Bombina (Duellman and Trueb 1994).
Members of Pipidae (clawed frogs) are exceptions; they
lack vocal cords but produce a clicking noise by means of
the sudden popping movement of a pair of articulating
cartilage disks attached to bony rods inside the larynx
(Duellman and Trueb 1994). In most frogs, males possess a
gular vocal sac that resonates and radiates the sound
(Duellman and Trueb 1994).
Members of most families of anurans make anti-
predator distress calls when disturbed (Bogert 1958). This
is true even of basal taxa such as Leiopelma in which
tympana, vocal sacs and courtship vocalisations are absent
(Bogert 1958). Mesozoic anurans may therefore have
made anti-predator sounds before they acquired tympanic
ears. Basal crown-group frogs are known from the Upper
Jurassic of Argentina (Henrici 1998), so anuran anti-
predator sounds may have existed that early.
Tympanic ears and chorusing behaviour are present in
the anuran clades Discoglossidae (midwife toads),
Mesobatrachia (spadefoot toads, clawed frogs and their
relatives) and Neobatrachia (typical frogs, tree frogs, true
toads and their relatives), and therefore may be
symplesiomorphies of higher anurans. Discoglossoids
and mesobatrachians are known from Laurasia as early as
the Upper Jurassic (Henrici 1998) and from Gondwana as
early as the Early Cretaceous (Baez et al. 2000). Anuran
chorusing therefore likely arose on those landmasses by
those times. The only known Mesozoic members of
Neobatrachia are South American members of Leptodac-
tylidae from the Upper Cretaceous (Sanchiz 1998).
Because no members of arboreal anuran clades are
known from the Mesozoic, we can safely infer that
Mesozoic anuran choruses took place at ground level.
During choruses, male anurans call to attract mates
(advertisement calls) and also emit special aggressive calls
in response to the loud vocalisations of nearby male
neighbours (Schwartz 2001). Aggressive calls may differ
qualitatively or quantitatively from advertisement calls
(Schwartz 2001). Females of many anuran species prefer
males whose calls are louder, delivered at higher rates, and
delivered for longer periods; in fact, male anurans often
increase these parameters in their own calls in response to
the calls of rivals (Schwartz 2001). If such directional
selection by female choice was present through anuran
geological history, then Mesozoic anuran calls exhibited
lower amplitude, rate and duration than extant anuran calls.
During choruses, male frogs tend to vocalise during the
silences between calls of nearby conspecifics – or, for
species with multi-note calls, if the songs overlap, notes are
sung by one individual between the notes sung by another
(Schwartz 2001). Such vocal alternation enables males to
detect each other’s calls and enables females to discriminate
between individual males’ calls (Schwartz 2001). If vocal
alternation during anuran chorusing evolved by means of
directional selection, we can safely infer that it was absent
or poor in the earliest anuran choruses.
Table 6. Mechanisms of defensive sound production by extant salamanders (Maslin 1950).
Family Description of sound Mechanism (if known)
Ambystomatidae (mole salamanders) Grunt Laryngeal vocal cordsClick, kissing sound Forcefully opening mouth (sound produced by
breakage of moist seal around mouth)Amphiumidae (amphiumas) Whistle, peep Forcing air through gill slits
Cryptobranchidae (giant salamanders)Cry similar to that of ahuman infant
–
Dicamptodontidae (mole salamanders) Bark Laryngeal vocal cordsPlethodontidae (lungless salamanders) Squeak –Proteidae (mudpuppies) Hiss –
Salamandridae (newts and kin) Peep, squeak, kissing soundForcefully opening mouth (sound produced bybreakage of moist seal around mouth)
Sirenidae (sirens) Croak, hiss –
------------------------------------------------------
P. Senter272
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

Extinct basal tetrapods with tympanic ears
Skull and the stapes morphology preclude tympana in
most basal tetrapods, including basal amniotes (Reisz
1981; Carroll 1991; Laurin 1998; Clack and Allin 2004;
Laurin and Soler-Gijon 2006), but there were exceptions.
A rodlike stapes oriented toward a strongly embayed
posterior skull notch suggests a tympanic ear in the Lower
and Upper Permian taxon Seymouriamorpha (Ivakhnenko
1987), which is phylogenetically intermediate between
Temnospondyli and Amniota (Laurin 1998; Laurin and
Reisz 1999; Ruta et al. 2003) (Figure 4). A posterior skull
embayment that suggests a tympanum is also present in the
Upper Carboniferous and Lower Permian taxon Diadecto-
morpha (Berman et al. 1992). Diadectomorpha is
phylogenetically positioned immediately outside Amniota
(Laurin and Reisz 1995; Ruta et al. 2003) (Figure 4).
A reduced stapes and a posterior skull embayment that
suggest a tympanum are present in most members of
Parareptilia, a clade of dumpy-looking basal amniotes
from the Permian and Triassic (Carroll and Lindsay 1985;
Laurin and Reisz 1995; Tsuji 2006; Muller and Tsuji
2007). The embayment is absent in Mesosauridae, the
most basal parareptilian group, and its presence is a
synapomorphy of higher parareptiles (Laurin and Reisz
1995; Tsuji 2006). Within Parareptilia the otic embayment
is particularly well developed in the Upper Permian and
Triassic clade Procolophonoidea and a Middle Permian
clade informally called ‘nycteroleters’ (Carroll and
Lindsay 1985; Muller and Tsuji 2007). The orbits are
enlarged in both groups, suggesting crepuscular or
nocturnal habits, hence occupation of a visually compro-
mised niche, to which the enhanced auditory apparatus
may be functionally related (Muller and Tsuji 2007).
Taxa without tympanic ears fill the phylogenetic gaps
between Seymouriamorpha, Diadectomorpha and Pararep-
tilia, and also between these taxa and other amniote taxa
with tympanic ears (Figure 4). Such ears therefore appeared
independently in each group. The presence of tympanic ears
in Seymouriamorpha, Diadectomorpha and Parareptilia
made possible the use of airborne sounds in intraspecific
communication. However, there is no certainty that it
occurred. These groups may have used their tympanic ears
for other functions instead of (or in addition to) intraspecific
communication. Seymouriamorphs were carnivorous
(Hotton et al. 1997) and may therefore have used hearing
to locate prey. A prey-locating function is unlikely in the
largely herbivorous clades Diadectomorpha and Pararepti-
lia. All three taxa may have used hearing for predator
avoidance, as do extant reptiles (Greene 1988).
Lower Permian seymouriamorphs are the earliest
known animals with tympanic ears that preyed on
vertebrates. The earliest airborne, anti-predator sounds
made by vertebrates may therefore have been directed at
Lower Permian seymouriamorphs.
Turtles (Reptilia: Testudines)
Turtles and tortoises are usually silent but do occasionally
make sounds by unknown means. During copulation or
while chasing the female, males often make sounds that
have been described as grunts, groans, moans, bellows and
roars (Gans and Maderson 1973; Auffenberg 1977). Such
mating sounds are little studied but are known from such a
wide taxonomic spectrum of turtle families (Campbell and
Evans 1972; Gans and Maderson 1973) that all extant
turtles are phylogenetically bracketed by taxa known to
produce mating sounds (Shaffer et al. 1997). This suggests
that turtle mating sounds were present in the common
ancestor of crown group turtles, which appeared in the
Lower Jurassic (Rougier et al. 1995). Osteological
correlates of hearing in extant turtles are also present in
the Upper Triassic and Lower Jurassic family Australo-
chelidae, the sister taxon to the turtle crown clade
(Gaffney and Kitching 1990). The situation is ambiguous
in the Upper Triassic taxon Proganochelys, the most basal
known fossil turtle, because of poor preservation in the
relevant region of the skull (Gaffney 1990).
The morphology of the tympanic region of the turtle
skull strongly resembles that of some parareptiles, and the
same pair of bones (squamosal and quadratojugal) supports
the tympanum in both groups (Carroll and Lindsay 1985;
Laurin and Reisz 1995; Rougier et al. 1995; Tsuji 2006).
If turtles arose from within Parareptilia (Laurin and Reisz
1995; Lee 1997a,b), then the tympanic ears of turtles are
probably homologous with those of parareptiles. This
introduces the possibility that parareptiles made turtle-like
courtship and mating sounds, in which case such sounds
may have existed as early as the Permian.
While some phylogenetic analyses of morphological
data place turtles within Parareptilia (Laurin and Reisz
1995; Lee 1997a,b), others place them just outside Diapsida
(Gauthier et al. 1988) or within crown Diapsida (Rieppel
and deBraga 1996; deBraga and Rieppel 1997). Curiously,
molecular analyses usually place them within or closely
related to the diapsid taxon Archosauria (Mannen et al.
1997; Platz and Conlon 1997; Hedges and Poling 1999;
Zardoya and Meyer 2001), for which there is no
documented morphological support, although one recent
molecular study places turtles in a phylogenetic position
compatible with parareptile status (Frost et al. 2006). If
turtles are diapsids then their tympanic ears are homologous
with those of other diapsids and not with those of
parareptiles.
Lizards and kin (Diapsida: Lepidosauromorpha)
Within the amniote taxon Diapsida are two major clades:
Lepidosauromorpha (lizards, tuataras and extinct kin) and
Archosauromorpha (crocodilians, birds and extinct kin,
including dinosaurs and pterosaurs) (Benton 1985)
(Figure 4). Extinct diapsids basal to the clade
Historical Biology 273
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

Lepidosauromorpha þ Archosauromorpha lacked tympanic
ears (Reisz 1981; Clack and Allin 2004). Extant lepidosaur-
omorphs and archosauromorphs possess tympanic ears,
except for some in which tympanic ears have been lost:
snakes (Serpentes), amphisbaenians (Amphisbaenia), Cali-
fornia legless lizards (Anniella), chameleons (Chamaeleoni-
dae) and tuataras (Sphenodon) (Wever 1978). Most such
groups are subterranean and therefore have no need for
airbornehearing (Wever 1978).Whilemost extant snakes are
not subterranean, their ancestors probablywere subterranean
or aquatic (Apesteguıa and Zaher 2006; Palci and Caldwell
2007), so their lack of tympanic ears is a vestige of ancestry.
Basal lepidosauromorphs and archosauromorphs exhi-
bit osteological evidence of tympanic ears (posteriorly
embayed quadrate, thin stapes) (Gregory 1945; Robinson
1962; Kuhn-Schnyder and Peyer 1973; Carroll 1975;
Gow 1975; Evans 1980; Dilkes 1998), which suggests
inheritance from a common ancestor. A recently published
inference that the two lineages acquired tympanic ears
separately is based on the assumption that the basal diapsid
Youngina – which lacks evidence for tympanic ears – is a
basal lepidosauromorph (Clack and Allin 2004). However,
recent phylogenetic analyses place Youngina outside the
lepidosauromorph-archosauromorph clade (Laurin 1991;
Rieppel and deBraga 1996; Lee 1997a,b; Modesto and
Reisz 2002; Senter 2004). Enlarged orbits in basal
lepidosauromorphs and archosauromorphs (Robinson
1962; Kuhn-Schnyder and Peyer 1973; Carroll 1975;
Gow 1975; Evans 1980, Evans 1991; Modesto and Reisz
2002) suggest crepuscular or nocturnal habits, an
interpretation supported by the fact that a reconstructed
ancestral archosaur visual pigment seems made for low
light levels (Chang et al. 2002). As with parareptiles
(Muller and Tsuji 2007), the enhancement of hearing in
these animals may be functionally related to ancestral
occupation of a visually compromised niche.
The earliest known lepidosauromorphs and archosaur-
omorphs are from the Upper Permian (Carroll 1975; Evans
and King 1993). Whether or not the two groups acquired
airborne hearing independently, it was present in both by
then. Anti-predator sounds directed at diapsids may
therefore have appeared in the Upper Permian. The small,
peglike teeth of many basal diapsids, including basal
lepidosauromorphs and basal archosauromorphs, suggest
a diet of invertebrates (Colbert 1970; Carroll 1975;
Gow 1975; Evans 1980). Anti-predator sounds directed at
Permian diapsids may therefore have included stridulation
by arachnids and beetles.
The ability to hear airborne sounds allows acoustic
communication, but among extant lizard families with
Mesozoic records only geckoes (Gekkonidae and Euble-
pharidae) vocalise. Using laryngeal vocal cords, geckoes
make sounds that are described as squeaks, growls and
barks, which carry only for a few meters in some cases and
as far as 20m in others (Gans and Maderson 1973;
Marcellini 1977). Single chirps are used for threat calls
(when lunging at a predator or conspecific rival) and
distress calls (upon being attacked or grasped), both in
nocturnal and diurnal geckoes (Marcellini 1977). Tem-
porally patterned multiple chirps are used by nocturnal
geckoes, usually males, in intraspecific social situations
(Marcellini 1977). Multiple chirps are produced at the
beginning of the nocturnal activity period, and may be
uttered in response to other individuals’ calls, in response
to the sight of rival males, and during courtship (Marcellini
1977). Multiple chirp calls vary between species and
appear to be related to spacing and territory (Marcellini
1977). The earliest known geckoes are from the Aptian-
Albian (Lower Cretaceous) of Mongolia (Alifanov 1989),
and it is possible that gecko calls were present then.
The lepidosauromorph taxon Sphenodontia is today
represented by only one genus: Sphenodon (tuataras).
Tuataras croak loudly when disturbed (Gans and Wever
1976). Sphenodontians are known from as early as the
Upper Triassic (Fraser 1982). Sphenodon lacks tympana
and has lost the embayment in the quadrate bone that is
associated with the tympanum in lizards. The embayment
is present in basal sphenodontians (Evans 1980; Fraser
1982; Wu 1994) but was lost in the lineage leading to
Sphenodon by the Lower Jurassic (Reynoso and Clark
1998). The loss of tympanic ears indicates that the use of
airborne sounds for intraspecific communication was not
important in the lineage.
Without tympanic ears extant tuataras have trouble
hearing airborne sounds over 1 kHz but can hear their own
croaks, which are of appropriate frequency (Gans and
Wever 1976) (Figure 3). Tuataras are not known to vocalise
for intraspecific communication, but the fact that they
can hear their own voices suggests that they have the
equipmentwithwhich to do so, evenwithout tympanic ears.
This fact might tempt the researcher to conclude that basal,
terrestrial tetrapods without tympanic ears may have used
low-frequency vocalisations for intraspecific communi-
cation. However, such temptation should be resisted.
The loss of the tympanum in extant reptiles results not only
in a limitation of auditory sensitivity to low-frequency
sounds (Figure 3), but also results in an inability to hear
such sounds except at very high amplitudes (Wever 1978).
Snakes are exceptions to this rule, because they can hear
low-frequency sounds at low amplitudes (Wever 1978).
However, the example of the snake’s ear is inapplicable to
basal tetrapods, for two reasons. First, the quadrate bone of
a snake’s skull (Figure 1) is so loose that it vibrates
freely and has essentially become a bony tympanum
(Wever 1978), whereas no posterior skull bonewas loose in
basal tetrapods. Second, the stapes of a snake is slender and
contacts the quadrate as if contacting a tympanum (Wever
1978) (Figure 1), whereas the basal tetrapod stapes was a
robust, solid brace between the bones of the cheek or palate
and the braincase (Carroll 1986; Clack 1992).
P. Senter274
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

Mesozoic marine reptiles
Stapedial and posterior skull morphology indicate
retention of tympanic ears in Nothosauria, a marine
archosauromorph clade from the Triassic Period
(Carroll and Gaskill 1985). This indicates an ability to
hear airborne sounds in these animals, despite their aquatic
habits. In Plesiosauria, a Mesozoic marine taxon closely
related to Nothosauria, a thin stapes suggests evolution
from an ancestor with tympanic ears (Hetherington 2008).
However, posterior skull morphology indicates that the
tympanum was lost in Plesiosauria (Carpenter 1997;
Storrs 1997; Hetherington 2008).
A tympanic notch is absent in the Mesozoic marine
taxon Ichthyosauria (Maisch 1997), which may or may not
be a member of Diapsida (Maisch 1997; Motani et al.
1998). The ichthyosaur stapes is robust and serves as a
brace between the cheek and braincase (Hetherington
2008), as in basal amniotes (Clack and Allin 2004). This
suggests that ichthyosaur ancestors never passed through a
stage with tympanic ears. The lack of tympanic ears in
plesiosaurs and ichthyosaurs does not necessarily mean
that they were deaf to underwater sounds but does indicate
a lack of importance of airborne sounds to them.
Mosasauridae, a marine family of Upper Cretaceous
lizards, retained tympanic ears, but they were modified for
underwater sound reception. The tympanum was often
ossified, increasing resistance to rupture in deep water
(Russell 1967). The middle ear cavity was encased in bone
and lacked acoustic isolation from the rest of the skull,
allowing the skull to conduct sound to the middle ear
(Hetherington 2008). Underwater hearing was therefore
apparently important to mosasaurs. It is impossible to
know, however, whether mosasaur hearing was of
importance in prey localisation, avoidance of dangerous
animals emitting honest sonic warnings, intraspecific
communication, or a combination of factors.
Crocodilians and kin (Archosauromorpha: Archosauria:Crurotarsi)
Basal diapsids appear to have been insectivorous, but the
serrated, ziphodont teeth of the archosauromorph clade
Archosauriformes indicate a switch to a diet of vertebrates
(Senter 2003). Archosauriformes exploded in diversity in
the Triassic Period but at least one of its basal members
was present in the Upper Permian (Hughes 1963). Anti-
predator sounds from vertebrates, directed at archosauri-
forms, may therefore have been present that early.
Members of Archosauria, the archosauriform crown
clade, are known from as early as the Middle Triassic
(Sereno 1991). Archosauria includes two major lineages,
Crurotarsi and Ornithodira (Sereno 1991) (Figure 4).
Crurotarsi includes crocodilians (Crocodylia) and several
extinct, mostly Triassic taxa (Sereno 1991). Ornithodira
includes dinosaurs (Dinosauria) – including birds (Aves)
– and pterosaurs (Pterosauria) and a few other extinct
forms (Sereno 1991). The vocal organ in extant members
of Crurotarsi (crocodilians) is the larynx, whereas in extant
members of Ornithodira (birds) it is the syrinx. Because
the larynx and syrinx are not homologous, it is most
parsimonious to infer that vocalisation arose indepen-
dently in the two lineages, in which case their common
ancestor lacked vocal ability.
Crocodilians vocalise by vibrating three mucosal
folds – absent in the avian larynx (McLelland 1989a,b) –
that project into the laryngeal lumen under part of the
arytenoid cartilage (Reese 1914). Unlike the avian syrinx
(see below), the crocodilian vocal organ leaves no trace of
its presence on fossil skeletons, so it is impossible to know
whether vocalisation was present further down the
crurotarsian tree than Crocodylia. We therefore cannot
know whether the Triassic and Jurassic Periods were filled
with the roaring of basal crurotarsians.
On the other hand, we can infer that the homologous
acoustic behaviours of gharials (Gavialidae), crocodiles
(Crocodylidae), alligators and caimans (Alligatoridae)
were present in their common ancestor (Senter 2008),
which appeared in the Upper Cretaceous (Brochu 2003).
Juvenile crocodilians of all three groups squeak or grunt
when disturbed, sometimes even from within the egg if the
nest is disturbed, and the sound attracts adults
(Modha 1967; Whitaker and Basu 1982; Cintra 1989;
Whitaker and Whitaker 1989). Adult males of all three
groups engage in head-slapping, in which the head is
raised and then slammed downward onto the water’s
surface to create a loud splash that is often followed by a
hiss in gharials or a roar in alligators and crocodiles
(Garrick et al. 1978; Whitaker and Basu 1982;
Thorbjarnarson 1991; Thorbjarnarson and Hernandez
1993). The head-slap is used in displays of presence and
dominance and in courtship (Garrick et al. 1978;
Thorbjarnarson 1989; Whitaker and Whitaker 1989).
Courtship in gharials, alligators and crocodiles also
involves geysering (shooting jets of water upward from
the nostrils, which are held just below the surface) and
blowing bubbles in the water with the mouth (Garrick et al.
1978; Whitaker and Basu 1982; Thorbjarnarson 1989;
Whitaker and Whitaker 1989; Thorbjarnarson and
Hernandez 1993). Narial geysering is also used as a
territorial display (Modha 1967; Garrick et al. 1978).
Territorial fights between crocodilian males often involve
loud splashing as bodies are hurled against each other
(Modha 1967; Singh and Rao 1990).
The above are all water-based behaviours that would
obviously have been absent in the terrestrial precursors of
Crocodylia. Aquatic or semi-aquatic habits are character-
istic of a large clade, Neosuchia, which includes
Crocodylia and several extinct Mesozoic taxa
(Clark 1994). If the aquatic acoustic behaviours of
crocodilians were present further down neosuchian
Historical Biology 275
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

phylogeny, they may have been present as early as the
Upper Jurassic, from which the earliest non-marine
neosuchians are known (Carroll 1988).
Most phylogenetic analyses place the marine taxon
Thalattosuchia within Neosuchia (e.g. Clark 1994; Ortega
et al. 2000; Sereno et al. 2001; Company et al. 2005).
Thalattosuchians, known from as early as the Lower
Jurassic (Carroll 1988), had narrow snouts (Clark 1994).
Thalattosuchian head-slapping, if it occurred, would
therefore have been weak for the sake of protecting the
narrow snout, as in the extant gharial (Whitaker and Basu
1982). Snouts were wider in most other extinct
neosuchians (Clark 1994), allowing strong head-slaps.
Head-slaps, if present, would have been particularly
impressive in the gigantic taxa Sarcosuchus and Deino-
suchus, in each of which head length approached 2m
(Sereno et al. 2001; Schwimmer 2002). Sarcosuchus is a
neosuchian from the Lower Cretaceous (Aptian-Albian) of
western Africa (Sereno et al. 2001), and Deinosuchus is an
alligatoroid from the Upper Cretaceous (Campanian) of
North America (Schwimmer 2002). Deinosuchus is more
certain than Sarcosuchus to have employed head-slapping
behaviour because Deinosuchus lies within Crocodylia
(Sereno et al. 2001), whereas Sarcosuchus does not (Sereno
et al. 2001). Also, Sarcosuchus is phylogenetically
bracketed by narrow-snouted forms (Sereno et al. 2001),
which increases the likelihood that head-slapping beha-
viour, if present that far back in neosuchian phylogeny, was
lost or weakened in the clade that includes Sarcosuchus.
Members of Alligatoridae (alligators and caimans)
employ both head-slapping and a vocal acoustic advertise-
ment display called bellowing (Garrick 1975; Garrick et al.
1978;Gorzula andSeijas 1989).Maleand femalealligatorids
bellow to advertise presence. Bellowing is contagious and its
sound is sexually dimorphic (Garrick 1975; Garrick et al.
1978). Alligatoridae is unknown before the Cenozoic
(Brochu 2003), but other members of Alligatoroidea, the
larger taxon that includes Alligatoridae but excludes
CrocodylidaeandGavialidae,werepresent inNorthAmerica
as early as the Campanian (Upper Cretaceous) (Brochu
1999). If basal alligatoroids bellowed, then the sound may
have been present in the Upper Cretaceous.
Birds and kin (Archosauria: Ornithodira)
The syrinx of birds (Ornithodira: Aves) is a series of
cartilage rings at the junction of the trachea and primary
bronchi, with membranous folds that protrude into the
lumen and can be vibrated to produce sound (Brackenbury
1989; King 1989). Vocal production by the syrinx depends
on the presence of a clavicular air sac (Brackenbury 1989),
a soft structure that invades and pneumatises the bones of
the avian pectoral girdle and limb, marking its presence
with a characteristic opening on the humerus (McLelland
1989a,b). Such an opening is present in members of the
avian clade Ornithothoraces – which includes modern
birds and the Lower and Upper Cretaceous taxon
Enantiornithes (Chiappe and Walker 2002) – but not in
birds basal to Ornithothoraces (Chiappe et al. 1999; Zhou
and Zhang 2003).
The clavicular air sac does not develop individually
but instead represents a fusion of outgrowths from two pre-
existing air sacs and a pair of outgrowths from the lungs.
The four outgrowths do not meet in the midline, in the
vicinity of the syrinx, until late in embryology (Locy and
Larsell 1916). The status of the clavicular air sac as an
outgrowth of pre-existing structures, two of which are
other air sacs, suggests that it is a later evolutionary
acquisition than the original set of ornithodiran air sacs.
This is consistent with the interpretation that it arrived
relatively late in ornithodiran history. Evidence exists that
non-avian ornithodirans, including basal birds, possessed
other air sacs homologous with those of birds (Britt et al.
1998; O’Connor 2006; Wedel 2006), but not the clavicular
air sac. Outside Ornithothoraces, humeral pneumatisation
is present in Pterosauria (O’Connor 2006) but not in the
many intervening dinosaurian taxa (Britt 1997). Humeral
pneumatisation in pterosaurs is therefore not homologous
with that of birds and so is not evidence for the presence of
a clavicular air sac homologous with that of birds.
Pneumatisation of the furcula is present in the allosauroid
theropod Aerosteon riocoloradensis, suggesting the pre-
sence of a clavicular air sac (Sereno et al. 2008). However,
such a sac is not likely homologous with that of
ornithothoracine birds because its osteological indicator
(furcular pneumatisation) is absent in phylogenetically
intermediate taxa, including other allosauroids (Chure and
Madsen 1996), basal birds (Chiappe et al. 1999; Zhou and
Zhang 2003; Mayr et al. 2007) and non-avian coelur-
osaurian theropods (Norell et al. 1997; Makovicky and
Currie 1998; Clark et al. 1999; Xu and Norell 2004).
Without evidence for a clavicular air sac homologous with
that of birds, we should not presume that basal birds and
non-avian ornithodirans possessed a functioning syrinx.
The lack of evidence of a syrinx in ornithodirans
outside Ornithothoraces will, no doubt, disappoint fans of
roaring movie dinosaurs. However, lack of ability to
vocalise does not necessarily mean that such animals were
silent altogether. Many extant reptiles communicate with
each other and with potential predators by non-vocal
acoustic means such as hissing, clapping jaws together,
grinding mandibles against upper jaws, rubbing scales
together, or use of environmental materials (e.g. splashing
against water) (Campbell and Evans 1972; Gans and
Maderson 1973; Garrick et al. 1978; Thorbjarnarson and
Hernandez 1993). Birds also use non-vocal acoustic means
of communication such as hissing, bill-clapping, stamping
and wing beating (Welty and Baptista 1988; Kear 2005;
Nelson 2005). Non-avian theropods with feathered wings
may have beaten their wings in acoustic displays as extant
P. Senter276
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

birds often do. Sauropod dinosaurs of the family
Diplodocidae, known from the Upper Jurassic of North
America and Africa (Upchurch et al. 2004), possessed
whiplike tail tips that could have produced loud, whiplike
cracking sounds for intraspecific communication or in
response to predators (Myhrvold and Currie 1997).
The morphology of the nasal passage in Lambeosaur-
inae, a Laurasian clade of Upper Cretaceous ornithischians
(Horner et al. 2004), suggests a role in sonic resonation
(Weishampel 1981). However, resonation need not be for
vocal sounds. Some extant snakes lack vocal cords but
possess resonating chambers that emphasise the low
frequencies of the hiss (Young 1991). The variety of visual
display structures in pterosaurs and dinosaurs (Chapman
et al. 1997; Molnar 2005; Unwin 2006) shows that visual
communication was important to these animals. They may
therefore have relied largely on visual means of
communication. Extant precedent for such reliance is
found among lizards, in which non-chemical communi-
cation is primarily visual (Pough et al. 1998), despite their
excellent hearing (Wever 1978).
Members of a few extant bird orders are known from
before the Cenozoic. Members of Anseriformes (water-
fowl) are known from the latest Cretaceous (Campanian-
Maastrichtian) of Asia, North America and Antarctica
(Hope 2002; Clarke et al. 2005). Several acoustic
behaviours are taxonomically widespread within Anser-
iformes and may be behavioural symplesiomorphies of the
group: near-constant contact calls of low amplitude among
groups, pairs or parents and offspring; a loud duet called a
triumph display by a pair after an agonistic encounter; a
special greeting call; a special post-copulatory call; a
special alarm call; calling during takeoff and throughout
flight; defensive hissing and among juveniles, special
distress calls, begging calls and calls expressing sleepiness
(Kear 2005). Vocal pitch usually differs between the sexes
and is higher in juveniles than adults (Kear 2005).
Members of Galliformes (game birds) are known from
the latest Cretaceous (Campanian-Maastrichtian) of North
America (Hope 2002). Several acoustic behaviours are
taxonomically widespread within Galliformes and may be
behavioural symplesiomorphies of the group: frequent
utterance of contact calls of low pitch and amplitude
among aggregations and pairs; a loud, far-carrying, often
multi-syllable advertisement call by the male that is often
answered by the female to form a vocally dimorphic duet
that may be answered by neighbouring males or pairs; a
special alarm call; and a loud, rapid, multi-syllable call
that differs from the usual alarm call, upon takeoff when
flushed (Jones et al. 1995; Madge and McGowan 2002).
Anseriformes and Galliformes together form the clade
Galloanserae, which is the sister clade to all other extant
birds except ratites and tinamous (van Tuinen et al. 2000).
The frequent occurrence in both Anseriformes and
Galliformes of duetting and near-constant, low-amplitude
contact calls suggests that they may be behavioural
symplesiomorphies for Galloanserae and therefore were
present in the Cretaceous.
Members of Gaviiformes (loons) are known from the
latest Cretaceous (Maastrichtian) of South America (Hope
2002). Loons emit trains of clucking sounds during flight
and a variety of calls in other situations (Oberholser 1974).
Far-reaching, drawn-out advertisement calls known as
wails or yodels are produced at dawn and dusk during the
breeding season (Oberholser 1974). In most loons takeoff
involves much noise because a loon has to run along the
water’s surface for a long distance to become airborne, and
landing may involve a powerful splash (Oberholser 1974).
Members of the extant family Phalacrocoracidae
(cormorants, order Pelecaniformes) are known from the
latest Cretaceous (Maastrichtian) of Asia and North
America (Hope 2002). Several acoustic characteristics and
behaviours found in cormorants are widespread in
Pelecaniformes and may be behavioural symplesiomor-
phies for the group. These include general silence away
from the nesting colony except for loud communal fishing
calls; high-pitched begging calls from chicks; sexual
dimorphism in voice; emission of hisses and special
disturbance calls during nest-usurpation attempts; copula-
tory vocalisations and the ability to recognise the voices of
offspring and neighbours (Nelson 2005). Other acoustic
characteristics that are typical of cormorants but not other
pelecaniformes include squeaky complaint calls from
chicks when disturbed; loss of the female voice after five
to six weeks; growling or loud barks by males in nest
defense; and a display by the male when advertising for a
female in which he throws his head back and makes
neighing or gargling sounds and follows this with ‘Goww,
gow, goww,’ if the female approaches (Nelson 2005).
Some fragmentary Upper Cretaceous (Maastrichtian)
bird specimens have been referred to Charadriiformes
(shorebirds) and Psittaciformes (parrots), but these
assignments are doubtful (Hope 2002). We therefore
cannot confidently infer that typical shorebird or parrot
calls were present before the Cenozoic.
Among extant birds, the vocabularies of basal groups –
ratites, tinamous, waterfowl, and galliforms – tend to be
differentiated at least into loud advertisement calls, soft
contact calls, special alarm calls, and (in Ratitae and
Anseriformes) the use of hissing as a threat (Jones et al.
1995; Davies 2002; Madge and McGowan 2002; Kear
2005). Such differentiation also occurs in more derived
groups (e.g. Nelson 2005) and therefore may be
symplesiomorphic for Neornithes, the clade that includes
all birds phylogenetically bracketed by the extant bird
orders. The earliest known neornithean birds are from the
Upper Cretaceous (Campanian) (Hope 2002), at which
time the differentiation was probably present.
Extant bird vocalisations vary in pitch according to body
size, with smaller birds emitting vocalisations of higher
Historical Biology 277
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

frequency (Welty and Baptista 1988). Avian body size is
also correlated with the range of frequencies detectable by
the inner ear, with smaller birds capable of detecting higher
frequencies (Gleich et al. 2004). There is no reason to expect
that the same was not true of Mesozoic birds.
Whether or not Mesozoic birds sang is at least partly a
semantic issue. According to the technical definition, an
avian advertisement call is considered a ‘song’ if it is
acoustically complex, multi-syllabic, and exhibits shorter
pauses between notes than the pauses between groups of
notes (Welty and Baptista 1988). In practice, however,
avian advertisement vocalisations are called ‘songs’ if they
issue from the mouths of songbirds (Passeriformes), doves
(Columbiformes) or tinamous (Tinamiformes), whether or
not they are complex or multi-syllabic. This is probably
because the advertisement vocalisations of these taxa have a
quality that is musical to human ears. The less pleasant (to
humans) advertisement vocalisations of other bird taxa are
considered ‘calls’ rather than ‘songs,’ despite their similar
function and the fact that in some taxa (e.g. galliforms and
loons) they fit the technical definition of ‘song.’ In any case,
Mesozoic bird sounds probably lacked musical quality
because songbirds, doves and tinamous lack Mesozoic
records (Unwin 1993; Boles 1995), and musical vocalisa-
tions are not the norm in other birds.
Mammals and kin (Amniota: Synapsida)
The amniote clade Synapsida includes mammals (Mam-
malia) and their extinct relatives. Basal synapsids
(‘pelycosaurs’) lacked tympanic ears (Carroll 1986). The
bones that support the tympanum in mammals and connect
it with the stapes are homologous to the posterior bones of
the mandible and jaw joint in other synapsids (Allin 1986;
Luo and Crompton 1994). In the synapsids most closely
related to mammals these bones became smaller and less
solidly attached to the anterior mandible (Allin 1986;
Crompton and Hylander 1986). In basal mammals – and
possibly inHadrocodium, the genus most closely related to
mammals (Luo et al. 2001) – these bones detached from
the mandible and became incorporated into the middle ear
(Wang et al. 2001). This specialisation was present by the
Middle Jurassic at the latest (Kielan-Jaworowska et al.
2004), and by the Lower Jurassic if it was present in
Hadrocodium (Luo et al. 2001). It indicates definitive
evidence for tympanic hearing, hence the possibility of
acoustic communication, in the earliest mammals. The
earliest mammals and their predecessors were probably
nocturnal (Crompton et al. 1978), so their evolution of
airborne hearing may have been functionally related to
their occupation of a visually compromised niche.
The possibility of tympanic hearing in synapsids
intermediate between ‘pelycosaurs’ and mammals has
received attention. There are strong osteological indicators
that these animals lacked a tympanum posterior to the
quadrate such as diapsids possess (Luo and Crompton
1994). Some researchers have suggested that a mandibular
tympanum was present in derived Permian and Triassic
synapsids with a posterior mandibular cleft bordered by
bones homologous to those that contact the mammalian
tympanum (e.g. Allin 1986). However, three lines of
evidence support the absence of a tympanum outside a
derived synapsid clade that appeared in the Upper Triassic.
That clade, hereafter called the DS (derived synapsid) clade
for concision, includesmammals and their extinct outgroups
Morganucodontidae and Tritylodontidae (Figure 4). First,
the synapsid homologue of the cochlea retained its
plesiomorphic, globular shape in other synapsids (Allin
1986) but became elongated, indicating increased auditory
acuity, in the DS clade (Vater et al. 2004). Second,
osteological evidence suggests that the posterior mandibular
bones were muscle attachment sites in other synapsids but
not in the DS clade, and the attachment of muscles would
have impeded transmission of vibrations from a tympanum
(Kemp 2005). Third, the attachment of the posterior
mandibular bones was solid, impeding transmission of
vibrations from a tympanum, in other synapsids but was
loose, facilitating transmission of vibrations, in the DS clade
(Crompton and Hylander 1986).
The same three lines of evidence also suggest an
increase in auditory acuity at the base of the DS clade.
However, they do not demonstrate the presence of
tympana. New osteological evidence from a particularly
well-preserved skull of Chiniquodon, a non-mammalian
member of the DS clade, shows that the posterior
mandibular bones were acoustically isolated from the
anterior mandible and were free to vibrate as a unit about
their collective longitudinal axis in response to low-
frequency airborne sound (Kemp 2007). However, the
arrangement of the rest of the skull bones precludes the
presence of an air-filled tympanic cavity, without which a
tympanum is pointless (Kemp 2007). Basal members of
the DS clade could therefore hear airborne sounds but only
at low frequencies (Kemp 2007).
Extant mammals of all three clades – Monotremata
(egg-laying mammals: echidna and platypus), Metatheria
(marsupials) and Eutheria (placental mammals) – vocalise
using vocal cords in the larynx. Their common ancestor,
which appeared in the Middle Jurassic (Kielan-Jawor-
owska et al. 2004), therefore presumably also could
vocalise with laryngeal vocal cords. It is uncertain how
much, if any, further back in synapsid phylogeny vocal
cords were present. Venomous hindlimb spurs are
plesiomorphic for the clade that includes Mammalia and
its close outgroup Docodonta (Hurum et al. 2006), which
phylogenetically bracket Lower Jurassic taxa (Kielan-
Jaworowska et al. 2004). If proto-mammals employed
acoustic ‘honest warnings’ to predators, as do many
insects and amphibians (Masters 1979; Duellman and
Trueb 1994), vocal cords may have been present as far
P. Senter278
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

back as the common ancestor of Docodonta and
Mammalia in the Lower Jurassic.
In monotremes and the extinct basal mammalian clade
Multituberculata, the cochlea is more elongated than in
more basal synapsids, indicating that a wider range of
frequencies can be heard; even so, monotreme auditory
acuity is poor, with a peak at 5 kHz and low ability to hear
low-amplitude sounds (Vater et al. 2004). Therefore, in
Mesozoic mammals of this grade, any vocalisations used in
intraspecific communication likely exhibited low pitch.
In contrast, the cochleae of therian (marsupial and placental)
mammals are extremely elongated and coiled, allowing
hearing through wide frequency ranges even at low
amplitudes (Vater et al. 2004). The frequencies of peak
sensitivity for therian ears vary inversely according to body
size, with the largest therian ears sensitive to infrasound and
the smallest sensitive to ultrasound (Long 1994) (Figure 1).
Therefore, in Mesozoic therians, vocalisations used in
intraspecific communication likely were of highest pitch in
small species and of lower pitch in larger species.
The monotreme and marsupial vocal apparatus is poor,
due to fusion of the thyroid cartilage (a mammalian
apomorphy) of the larynx with the cricoid cartilage and the
possession of primitive laryngeal musculature (Negus 1949;
Kelemen 1963). Perhaps because of this, monotreme and
marsupial vocal sounds are generally simpler than those of
placental mammals, and their vocabularies are smaller.
Cooing, purring and snuffing sounds of unclear significance
are known in echidnas (Tachyglossidae) (Hutchins 2004).
Platypuses (Ornithorhynchus) growl when disturbed, and
the female emits pre-copulatory squeaks that are answered
by the male; juveniles make squeaking contact calls to each
other during play and are also known to hiss (Tembrock
1963; Griffiths 1978). Marsupial vocabularies often include
courtship vocalisations, aggressive vocalisations, and
hissing as a defensive sound, although in some taxa
defensive sounds are vocal (Nowak 2005). More extensive
vocabularies exist in the marsupial genus Macropus (true
kangaroos), but this is probably related to eusociality, a trait
characteristic of Macropus and uncharacteristic of marsu-
pials in general (Menkhorst and Knight 2004). It is therefore
doubtful that complex vocabularies existed among Meso-
zoic marsupials.
Stem-group marsupials are known from as early as the
Lower Cretaceous (Aptian-Albian) (Kielan-Jaworowska
et al. 2004), but no member of the marsupial crown group
is known from before the Cenozoic (Rougier et al. 1998).
None of the vocalisations characteristic of extant
marsupial families can therefore be inferred to have
existed before the Cenozoic. However, the fact that
vocabularies are differentiated into different vocalisations
for different situations in both platypuses and marsupials
suggests that such differentiation is an ancient trait. Vocal
differentiation may therefore have been present in the
common mammalian ancestor in the Middle Jurassic.
Also, the possibility of behavioural homology of defensive
hissing in marsupials and reptiles suggests that hissing was
a typical defensive behaviour through the synapsid
ancestry of mammals.
In placental mammals the thyroid cartilage has detached
from the cricoid cartilage (Figure 1) and the laryngeal
musculature is elaborated, so the eutherian larynx facilitates
more elaborate vocalisation than is possible in monotremes
and marsupials (Kelemen 1963). Perhaps because of this the
vocabularies of placental mammals tend to be larger than
those of monotremes and marsupials and can convey
information about a wider variety of situations and
emotional states (Estes 1991). The earliest known placental
mammals are from the Lower Cretaceous (Barremian)
(Ji et al. 2002), and complex vocabularies may have existed
among placentals that early.
One extant placental order, Soricomorpha (shrews,
tenrecs, hedgehogs and kin), is known from the Mesozoic
(Kielan-Jaworowska et al. 2004). Echolocation, clicking
sounds, defensive hissing and aggressive squeaking are
common and taxonomically widespread in extant sorico-
morphs (Nowak and Paradiso 1983). If these are ancestral
acoustic behaviours for Soricomorpha, they were present
before the Cenozoic. Soricomorphs are known from as
early as the Coniacian (Upper Cretaceous) of Asia and the
Campanian (Upper Cretaceous) of North America
(Kielan-Jaworowska et al. 2004).
Other extant placental orders are unknown before the
Cenozoic, but members of some extant placental supraor-
dinal taxa are present. Basal members of Archonta, which
includes the orders Dermoptera (colugos), Chiroptera (bats)
and Primates (primates), are known from the latest
Cretaceous (Maastrichtian) of India (Kielan-Jaworowska
et al. 2004). Basal members of the supraordinal taxon
Ungulatomorpha (hoofed mammals) are known from as
early as the Upper Cretaceous (Turonian) of Asia and the
latest Cretaceous (Campanian-Maastrichtian) of Europe and
South America (Kielan-Jaworowska et al. 2004). Sounds
made by extant archontans and ungulatomorphs are too
varied across taxa (Hill and Smith 1984; Estes 1991) to infer
ancestral acoustic behaviour for either group. Basal
members of Ferae, which includes the order Carnivora
and a few extinct orders, are known from the latest
Cretaceous (Campanian-Maastrichtian) of North America
(Kielan-Jaworowska et al. 2004). Sounds made by extant
carnivorans vary, but three types of sounds are common to
so many members of so many carnivoran families that they
may be plesiomorphic for Carnivora or perhaps even Ferae:
low-pitched growling to communicate threat, high-pitched
vocalisations in response to physical pain, and high-pitched
distress and contact calls from juveniles (Estes 1991).
Drumming with the hind feet on the substrate is a
common method to communicate alarm or the presence of
danger among mammals with hindlimbs that are specialised
for saltatory locomotion (hopping). This method has
Historical Biology 279
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

appeared convergently in Macropodidae (kangaroos and
their saltatory relatives), Macroscelididae (elephant
shrews), Leporidae (rabbits), Dipodidae (jerboas) and
Dipodomys (kangaroo rats) (Nowak and Paradiso 1983;
Menkhorst and Knight 2004). It is probably no coincidence
that this form of communication is prevalent among
mammals of this morphotype and that in all cases it
transmits the same type of information. Theoretically, a
novel reflexive behaviour in response to a given stimulus
could most easily evolve by modification of a preexisting
reflexive behaviour involving the same set of muscles in
response to the same stimulus. An oscillatory hindlimb
motor reflex (drumming) in response to danger might
therefore most easily evolve inmammals that already have a
preexisting oscillatory hindlimb motor reflex (hopping) in
response to danger. Because of its prevalence among
saltatory mammals, it is possible that this method of
acoustic communication of alarm/danger was present in
Zalambdalestes, a saltatory placental mammal from the
Upper Cretaceous (Campanian) of Mongolia (Kielan-
Jaworowska 1978).
Directions for further research
Rigorous analysis is needed to determine whether the
potential acoustic behavioural homologies proposed here
within Aves and Mammalia meet tests of homology. It
would also be informative to compare aspects of
vocalisation across anuran, avian and mammalian
phylogeny to determine whether or not it is possible to
infer ancestral characteristics of vocal sounds in each
group. It would also be informative to study inner ear
endocasts of more fossil archosaurian and synapsid taxa to
determine hearing ranges, as has been done for a few taxa
(Gleich et al. 2004; Vater et al. 2004). In addition, it would
be interesting to study sound-producing structures across
the phylogeny of ray-finned fishes to determine whether
homologies in acoustic anatomy can be assessed.
The big picture
The evidence and inferences presented above suggest that
the questions posed in this paper’s opening paragraph can
be answered. Triceratops and indeed even the earliest
dinosaurs heard crickets chirping in the evening.
The Mesozoic treetops did not resound with frogsong,
although Upper Jurassic and Cretaceous bodies of water
did. Cretaceous treetops were filled with the sounds of
honking, cackling and wailing birds in what must have
been an awful cacophony that would not become sweet
and musical until the advent of songbirds, doves and
tinamous in the Cenozoic. The immense corpses of
sauropods were surrounded with the buzzing of legions of
bottle flies, but not until the Upper Cretaceous.
In addition to such tidbits, a larger paleobioacoustical
scenario emerges from the pooled information presented
here (Figure 6). Incidental animal sounds were probably
present in the oceans by the beginning of the Paleozoic
Era, but those sounds were not heard (except perhaps by
cephalopods) until much later. The first terrestrial
arthropod communities arose in the Silurian Period, and
defensive stridulation in terrestrial arthropods may have
appeared soon thereafter. In the Silurian Period cartilagi-
nous and bony fishes appeared, and each independently
acquired the ability to hear low-frequency water-borne
sounds. Parallel evolution of defensive, aggressive and
courtship sounds is rampant among ray-finned fishes and
may have begun in the Silurian or Devonian. Malacos-
tracan crustaceans also appeared in the Devonian, and
several lineages evolved defensive stridulation in parallel,
possibly in response to predation by fishes with underwater
hearing. Bony fishes and early tetrapods of the Silurian and
Devonian Periods engaged in noisy air-breathing without
fear of being overheard by predators, because no
vertebrate had yet acquired tympanic ears.
Tetrapods exploded in diversity in the Carboniferous
Period. Amniotes appeared in the Pennsylvanian and
employeddefensivedisplays that incorporatedbody inflation
as a visual signal. The resulting hissing sound was first heard
by seymouriamorph predators in the Lower Permian and by
archosauriform predators in the Upper Permian. Arthropod
defensive stridulation was heard by diapsid predators for the
first time in the Upper Permian, although the mechanical
vibrations produced by stridulation may have deterred other
tetrapod predators as early as the Mississippian.
An explosion of terrestrial sound appeared in the
Triassic Period. Low-frequency twilight choruses may
already have existed among ray-finned fishes, but the
earliest terrestrial twilight choruses appeared in the Triassic
with the advent of crickets and their mesoedischiid and
titanopteran relatives. The drumming of stoneflies appeared
in theMiddle Triassic. The hemipteran taxon Prosorrhyncha
appeared in the Upper Triassic and radiated into several
lineages that evolved defensive and courtship stridulation in
parallel. Turtles first appeared in the Upper Triassic; their
courtship sounds may have appeared soon thereafter, or
those sounds may be evolutionary continuations of court-
ship sounds that were present in parareptiles from perhaps as
early as the Permian.
Water boatmen joined the twilight chorus in the Lower
Jurassic, and anurans joined it in the Upper Jurassic.
Venomous mammals began making defensive vocalisa-
tions in the Lower Jurassic. Tympanic ears appeared in
mammals by the Middle Jurassic and may have been used
to locate (chorusing?) insect prey. The tapping of booklice,
the buzzing of aculeate wasps, and tail-cracking sounds by
diplodocid sauropods appeared in the Upper Jurassic.
In the Lower Cretaceous, geckoes and birds joined the
twilight chorus. The tapping of death-watch beetles and
P. Senter280
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

termite soldiers, and the buzzing of horseflies, bottle flies
and mosquitoes appeared. Mammalian vocalisations
increased in pitch as therians gained the ability to hear
high-frequency sounds. In the Upper Cretaceous, cicadas
and head-slapping crocodilians joined the twilight chorus,
along with birds from a few extant orders. The buzzing of
bottle flies and the echolocating chirps of soricomorph
mammals also appeared, possibly along with the growling
of angry basal members of Ferae. High-frequency fish
sounds appeared with the advent of hearing-specialist
fishes.
This scenario is unlikely to be the last word on
Paleozoic andMesozoic animal sounds. Future discoveries
will augment, hone and perhaps even falsify hypotheses
put forth here. It is my hope that this synthesis will
stimulate more research on the voices of the past.
Acknowledgements
Of those who provided help with this project, M. Holmandeserves a particularly huge amount of thanks. She was in chargeof inter-library loan at Lamar State College-Orange in Orange,Texas while I worked there on the first phase of this project, andshe was run ragged during the massive literature search requiredfor this study. B. Karpinecz helped with the typing of thebibliography. Several ichthyologists provided help with findingreferences on acoustic behaviour in fishes: A. Popper,R. Rountree, C. Johnston, A. Bass, K. Huebert and J. McManus.R. Fortey and P. Selden helped me track down references onstridulatory structures in Paleozoic arthropods. Conversationswith R. Senter and M. McClure sparked useful ideas. Of thespecimens in Figure 1, M. McClure of Lamar State College-Orange provided access to the crab, toad and pig; L. Wilmore ofLamar State College-Orange provided access to the snake skull;and T. Rowe and C. Jass of the Texas Memorial Museumprovided access to the turtle skull. M. Laurin provided usefulinput that improved this paper.
References
Aiken RB. 1985. Sound production by aquatic insects. Biol Rev.65:163–211.
Alexander RD, Moore TE. 1963. The evolutionary differentiation ofstridulatory signals in beetles (Insecta: Coleoptera). Anim Behav.11:111–115.
Alifanov VR. 1989. The oldest gecko (Lacertilia, Gekkonidae) from theLower Cretaceous of Mongolia. Paleontol J. 23:128–131.
Allin EF. 1986. The auditory apparatus of advanced mammal-like reptilesand early mammals. In: Hotton N III, MacLean PD, Roth JJ, RothEC, editors. The ecology and biology of mammal-like reptiles.Washington, DC: Smithsonian Institution Press. p. 283–294.
Anderson JS, Reisz RR, Scott D, Frobisch ND, Sumida SS. 2008. A stembatrachians from the early Permian of Texas and the origin of frogsand salamanders. Nature. 453:515–518.
Apesteguıa S, Zaher H. 2006. A Cretaceous terrestrial snake with robusthindlimbs and a sacrum. Nature. 440:1037–1040.
Arratia G. 1993. The Jurassic and the early history of teleosts. In: ArratiaG, Viohl G, editors. Mesozoic fishes. Systematics and paleoecology.Munich: Verlag Dr. Friedrich Pfiel. p. 243–259.
Auffenberg W. 1977. Display behavior in tortoises. Am Zool.17:241–250.
Autrum H. 1963. Anatomy and physiology of sound receptorsin invertebrates. In: Busnel R-G, editor. Acoustic behaviour ofanimals. Amsterdam: Elsevier Publishing Company. p. 412–433.
Baez AM, Trueb L, Calvo JO. 2000. The earliest known pipoid frog fromSouth America: a new genus from the middle Cretaceous ofArgentina. J Vert Paleontol. 20:490–500.
Bailey WJ. 1991. Acoustic behaviour of insects, an evolutionaryperspective. London: Chapman and Hall.
Barth FG. 2002. A Spider’sWorld. Senses and behavior. Berlin: Springer.Benton MJ. 1985. Classification and phylogeny of the diapsid reptiles.
Zool J Linn Soc. 84:97–164.Berman DS, Sumida SS, Lombard ER. 1992. Reinterpretation of the
temporal and occipital regions in Diadectes and the relationships ofdiadectomorphs. J Paleontol. 66:481–499.
Bethoux O, Nel A. 2002. Venation pattern and revision of Orthopterasensu nov. and sister groups. Zootaxa. 96:1–88.
Blagoderov VA, Lukashevich ED, Mostovski MB. 2002. Order DipteraLinne, 1758. The true flies. In: Rasnitsyn AP, Quicke DLJ, editors.History of insects. Dordrecht: Kluwer Academic Publishers.p. 227–240.
Bogert CM. 1958. Influence of sound on the behavior of amphibians andreptiles. In: Layton WE, Tavolga WN, editors. Animal sounds andcommunication. Washington, DC: American Institute of BiologicalSciences. p. 137–320.
Boles WE. 1995. The world’s oldest songbird. Nature. 374:21–22.Bolt JR, Lombard RE. 1985. Evolution of the amphibian tympanic ear
and the origin of frogs. Biol J Linn Soc. 24:83–99.Boulard M. 2006. Acoustic signals, diversity and behaviour of cicadas
(Cicadidae, Hemiptera). In: Drosopoulos S, Claridge MF, editors.Insect sounds and communication. Boca Raton (FL): Taylor andFrancis. p. 331–349.
Brackenbury JH. 1989. Functions of the syrinx and the control of soundproduction. In: King AS, McLelland J, editors. Form and function inbirds. Vol. 4. San Diego (CA): Academic Press. p. 193–220.
Brazeau MD, Ahlberg PE. 2006. Tetrapod-like middle ear architecture ina Devonian fish. Nature. 439:318–321.
Briggs DEG, Weedon MJ, Whyte MA. 1993. Arthropoda (Crustaceaexcluding Ostracoda). In: Benton MJ, editor. The fossil record 2.London: Chapman and Hall. p. 321–342.
Britt BB. 1997. Postcranial pneumaticity. In: Currie PJ, Padian K, editors.Encyclopedia of Dinosaurs. San Diego (CA): Academic Press.p. 590–593.
Britt BB, Makovicky PJ, Gauthier J, Bonde N. 1998. Postcranialpneumatization in Archaeopteryx. Nature. 395:374–376.
Brochu CA. 1999. Phylogenetics, taxonomy, and historical biogeographyof Alligatoroidea. Soc Vert Paleontol Mem. 6:9–100.
Brochu CA. 2003. Phylogenetic approaches toward crocodylian history.Annu Rev Earth Plan Sci. 31:357–397.
Budelmann BU. 1992a. Hearing in Crustacea. In: Webster DB, Fay RR,Popper AN, editors. The evolutionary biology of hearing. New York:Springer-Verlag. p. 131–139.
Budelmann BU. 1992b. Hearing in nonarthropod invertebrates. In:Webster DB, Fay RR, Popper AN, editors. The evolutionary biologyof hearing. New York: Springer-Verlag. p. 141–155.
Campbell HW, Evans WE. 1972. Observations on the vocal behavior ofchelonians. Herpetologica. 28:277–280.
Campbell BC, Steffen-Campbell JD, Sorensen JT, Gill RJ. 1995.Paraphyly of Homoptera and Auchenorrhyncha inferred from 18SrDNA nucleotide sequences. Syst Entomol. 20:175–194.
Cappetta H. 1987. Handbook of Palaeoichthyology. Chondrichthyes II.Mesozoic and Cenozoic Elasmobranchii. Stuttgart: Gustav FischerVerlag.
Cappetta H, Duffin C, Zidek J. 1993. Chondrichthyes. In: Benton MJ,editor. The fossil record 2. London: Chapman and Hall. p. 593–609.
Carpenter K. 1997. Comparative cranial anatomy of two North AmericanCretaceous plesiosaurs. In: Callaway JM, Nicholls EL, editors.Ancient marine reptiles. San Diego (CA): Academic Press.p. 191–216.
Carroll RL. 1975. Permo-Triassic “lizards” from the Karroo. PalaeontolAfr. 18:71–87.
Carroll RL. 1986. The skeletal anatomy and some aspects of thephysiology of primitive reptiles. In: Hotton N III, MacLean PD, RothJJ, Roth EC, editors. The ecology and biology of mammal-likereptiles. Washington, DC: Smithsonian Institution Press. p. 25–45.
Carroll RL. 1988. Vertebrate paleontology and evolution. New York:W.H. Freeman and Company.
Historical Biology 281
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

Carroll RL. 1991. The origin of reptiles. In: Schultze H-P, Trueb L,editors. Origins of the higher groups of tetrapods. Ithaca (NY):Cornell University Press. p. 331–353.
Carroll RL. 2007. The Palaeozoic ancestry of salamanders, frogs, andcaecilians. Zool J Linn Soc. 150:1–140.
Carroll RL, Gaskill P. 1985. The nothosaur Pachypleurosaurus and theorigin of plesiosaurs. Phil Trans R Soc Lond B. 309:343–393.
Carroll RL, Lindsay W. 1985. Cranial anatomy of the primitive reptileProcolophon. Can J Earth Sci. 22:1571–1587.
Chang BSW, Jonsson K, Kazmi MA, Donoghue MJ, Sakmar TP. 2002.Recreating a functional ancestral archosaur pigment. Mol Biol Evol.19:1483–1489.
Chapman RF. 1975. The insects. Structure and function. New York:American Elsevier Publishing Company. p. 83–93.
Chapman RE, Weishampel DB, Hunt G, Rasskin-Gutman D. 1997.Sexual dimorphism in dinosaurs. In: Wolberg DL, Stump E,Rosenberg GD, editors. Dinofest international: proceedings of asymposium held at Arizona State University. Philadelphia: Academyof Natural Sciences. p. 83–93.
Chiappe LM, Walker CA. 2002. Skeletal morphology and systematics ofthe Cretaceous Euenantiornithes (Ornithothoraces: Enantiornithes).In: Chiappe LM, Witmer LM, editors. Mesozoic birds. Above theheads of Dinosaurs. Berkeley (CA): University of California Press.p. 240–267.
Chiappe LM, Ji S, Ji Q, Norell MA. 1999. Anatomy and systematics ofthe Confuciusornithidae (Theropoda: Aves) from the late Mesozoicof northeastern China. Bull Am Mus Nat Hist. 242:1–89.
Chure DJ, Madsen JH. 1996. On the presence of furculae in some non-maniraptoran theropods. J Vert Paleontol. 16:573–577.
Cintra R. 1989. Maternal care and daily patterns of behavior in a family ofcaimans, Caiman yacare in the Brazilian Pantanal. Copeia.1989:320–322.
Clack JA. 1992. The stapes of Acanthostega gunnari and the role of thestapes in early tetrapods. In: Webster DB, Fay RR, Popper AN,editors. The evolutionary biology of hearing. New York: Springer-Verlag. p. 405–420.
Clack JA, Allin E. 2004. The evolution of single- and multiple-ossicleears in fishes and tetrapods. In: Manley GA, Popper AN, Fay RR,editors. Evolution of the vertebrate auditory system. New York:Springer-Verlag. p. 128–163.
Clack JA, Ahlberg PE, Finney SM, Dominguez Alonso P, Robinson J,Ketcham RA. 2003. A uniquely specialized ear in a very earlytetrapod. Nature. 425:65–69.
Claridge MF. 1985. Acoustic signals in the Homoptera: behavior,taxonomy, and evolution. Annu Rev Entomol. 30:297–317.
Clark JM. 1994. Patterns of evolution in Mesozoic Crocodyliformes. In:Fraser NC, Sues H-D, editors. In the shadow of the Dinosaurs.Cambridge: Cambridge University Press. p. 84–97.
Clark JM, Norell MA, Chiappe LM. 1999. An oviraptorid skeleton fromthe Late Cretaceous of Ukhaa Tolgod, Mongolia, preserved in anavianlike brooding position over an oviraptorid nest. Am Mus Nov.3265:1–36.
Clarke JA, Tambussi CP, Noriega JI, Erickson GM, Ketcham RA. 2005.Definitive fossil evidence for the extant avian radiation in theCretaceous. Nature. 433:305–308.
Cloudsley-Thompson JL, Constantinou C. 1984. Stridulatory apparatus ofSolifugae (Solpugida). J Arid Env. 7:365–369.
Coffin A, Kelley M, Manley GA, Popper AN. 2004. Evolution of sensoryhair cells. In: Manley GA, Popper AN, Fay RR, editors. Evolution ofthe vertebrate auditory system. New York: Springer-Verlag.p. 55–94.
Cokl A, Virant-Doberlet M, Zorovic M. 2006. Sense organs involved inthe vibratory communication of bugs. In: Drosopoulos S, ClaridgeMF, editors. Insect sounds and communication. Boca Raton (FL):Taylor and Francis. p. 71–80.
Colbert EH. 1970. The Triassic gliding reptile Icarosaurus. Bull AmMusNat Hist. 143:87–142.
Collins D, Rudkin DM. 1981. Priscansermarinus barnetti, a probablelepadomorph barnacle from the Middle Cambrian Burgess Shale ofBritish Columbia. J Paleontol. 53:1006–1015.
Company J, Suberbiola XP, Ruiz-Omenaca JI. 2005. A new species ofDoratodon (Crocodyliformes: Ziphosuchia) from the Late Cretac-eous of Spain. J Vert Paleontol. 25:343–353.
Coombs S, Braun CB. 2003. Information processing by the lateral linesystem. In: Collin SP, Marshall NJ, editors. Sensory processing inaquatic environments. New York: Springer. p. 122–138.
Crompton AW, Hylander WL. 1986. Changes in mandibular functionfollowing the acquisition of a dentary-squamosal jaw articulation. In:Hotton N III, MacLean PD, Roth JJ, Roth EC, editors. The ecologyand biology of mammal-like reptiles. Washington, DC: SmithsonianInstitution Press. p. 263–282.
Crompton AW, Tyler CR, Jagger JJ. 1978. Evolution of homeothermy inmammals. Nature. 272:333–336.
Crowson RA. 1981. The biology of Coleoptera. London: Academic Press.Davies SJJF. 2002. Ratites and Tinamous. Oxford: Oxford University
Press.deBraga M, Rieppel O. 1997. Reptile phylogeny and the interrelation-
ships of turtles. Zool J Linn Soc. 120:281–354.Desutter-Grandcolas L. 2003. Phylogeny and the evolution of acoustic
communication in extant Ensifera (Insecta, Orthoptera). Zool Scr.32:525–561.
Dilkes DW. 1998. The Early Triassic rhynchosaur Mesosuchus browniand the interrelationships of basal archosauromorph reptiles. PhilTrans R Soc Lond B. 353:501–541.
Doyle P. 1993. Cephalopoda (Coleoidea). In: Benton MJ, editor. Thefossil record 2. London: Chapman and Hall. p. 229–236.
Drosopoulos S, Claridge MF. 2006. Insect Sounds and Communication.Boca Raton (FL): Taylor and Francis.
Duellman WE, Trueb L. 1994. Biology of amphibians. Baltimore (MD):Johns Hopkins University Press. p. 229–236.
Dumortier B. 1963a. Ethological and physiological study of soundemissions in Arthropoda. In: Busnel R-G, editor. Acoustic behaviourof animals. Amsterdam: Elsevier Publishing Company. p. 583–654.
Dumortier B. 1963b. Morphology of sound emission apparatus inArthropoda. In: Busnel R-G, editor. Acoustic behaviour of animal.Amsterdam: Elsevier Publishing Company. p. 278–345.
Dumortier B. 1963c. The physical characteristics of sound emissions inArthropoda. In: Busnel R-G, editor. Acoustic behaviour of animal.Amsterdam: Elsevier Publishing Company. p. 346–373.
Dunlop JA. 1994. An Upper Carboniferous amblypygid from theWrithlington Geological Nature Reserve. Proc Geol Assoc.105:245–250.
Dunlop JA, Anderson LI, Kerp H, Hass H. 2003. Preserved organs ofDevonian harvestmen. Nature. 425:916.
Engel MS. 2001. A monograph on the Baltic amber bees and evolution ofthe Apoidea (Hymenoptera). Bull Am Mus Nat Hist. 259:1–192.
Estes RD. 1991. The behavior guide to African Mammals. Berkeley(CA): University of California Press.
Evans SE. 1980. The skull of a new eosuchian reptile from the LowerJurassic of South Wales. Zool J Linn Soc. 70:203–264.
Evans SE. 1991. A new lizard-like reptile (Diapsida: Lepidosauromor-pha) from the Middle Jurassic of England. Zool J Linn Soc.103:391–412.
Evans SE, King MS. 1993. A new specimen of Protorosaurus (Reptilia:Diapsida) from the Marl Slate (late Permian) of Britain. ProcYorkshire Geol Soc. 49:229–234.
Ewer RF. 1973. The Carnivores. Ithaca (NY): Cornell University Press.494 pp.
Fant G. 1973. Speech sounds and features. Cambridge, MA: MIT Press.Fay RR. 1988. Hearing in vertebrates: a psychophysics databook.
Winnetka (IL): Hill-Fay Associates.Feldmann RM, Schweitzer CE, Redman CM, Morris NJ, Ward DJ. 2007.
New Late Cretaceous lobsters from the Kyzylkum Desert ofUzbekistan. J Paleontol. 81:701–713.
Fish MP, Mowbray WH. 1970. Sounds of Western North Atlantic Fishes.Baltimore (MD): Johns Hopkins Press.
Flook PK, Klee S, Rowell CHF. 2000. Molecular phylogenetic analysis ofthe Pneumoroidea (Orthoptera: Caelifera): molecular data resolvemorphological character conflicts in the basal Acridomorpha. MolPhyl Evol. 15:45–354.
Foster BA, Buckeridge JS. 1987. Barnacle palaeontology. In: SouthwardAJ, editor. Barnacle biology. Rotterdam: A.A. Balkema. p. 43–61.
Frankenberg E,Werner YL. 1992. Vocal communication in the Reptilia –facts and questions. Acta Zool Lill. 41:45–62.
Fraser NC. 1982. A new rhynchocephalian from the British Upper Trias.Palaeontology. 25:709–725.
P. Senter282
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

Fritzsch B. 1992. The water-to-land transition: evolution of the tetrapodbasal papilla, middle ear, and auditory nuclei. In: Webster DB,Fay RR, Popper AN, editors. The evolutionary biology of hearing.New York: Springer-Verlag. p. 351–375.
Frost DR, Grant T, Faivovich J, Bain RH, Haas A, Haddad CFB, de SaRO, Channing A, Wilkinson M, Donnellan SC, et al. 2006. Theamphibian tree of life. Bull Am Mus Nat Hist. 297:1–370.
Gaffney ES. 1990. The comparative osteology of the Triassic turtleProganochelys. Bull Am Mus Nat Hist. 194:1–263.
Gaffney ES, Kitching JW. 1990. The most ancient African turtle. Nature.369:55–58.
Gans C, Maderson PFA. 1973. Sound producing mechanisms in recentreptiles: review and comment. Am Zool. 13:1195–1203.
Gans C, Wever EG. 1976. Ear and hearing in Sphenodon punctatus. ProcNatl Acad Sci USA. 73:4244–4246.
Gao K, Shubin NH. 2003. Earliest known crown-group salamanders.Nature. 422:424–428.
Garrick LD. 1975. Structure and pattern of the roars of Chinese alligators(Alligator sinensis Fauvel). Herpetologica. 31:26–31.
Garrick LD, Lang JW, Herzog HA Jr. 1978. Social signals of adultAmerican alligators. Bull Am Mus Nat Hist. 160:157–192.
Gauthier J, Kluge AG, Rowe T. 1988. The early evolution of the Amniota.In: Benton MJ, editor. The phylogeny and classification of theTetrapods. Amphibians, reptiles, birds. Vol. 1. Oxford: ClarendonPress. p. 103–155.
Gerhardt HC, Huber F. 2002. Acoustic communication in insects andAnurans. Common problems and diverse solutions. Chicago (IL):University of Chicago Press.
Gleich O, Fischer FP, Kopl C, Manley GA. 2004. Hearing organevolution and specialization in archosaurs. In: Manley GA, PopperAN, Fay RR, editors. Evolution of the vertebrate auditory system.New York: Springer-Verlag. p. 224–255.
Gogala M. 1984. Vibration producing structures and songs of terrestrialHeteroptera as systematic characters. Biol Vestn. 32:19–36.
Gogala M. 1985. Vibrational songs of land bugs and their production. In:Kalmring K, Elsner N, editors. Acoustic and vibrational communi-cation in insects. Berlin: Verlag Paul Parey. p. 143–150.
Gogala M. 2006. Vibratory signals produced by Heteroptera –Pentatomorpha and Cimicomorpha. In: Drosopoulos S, ClaridgeMF, editors. Insect sounds and communication. Boca Raton (FL):Taylor and Francis. p. 275–295.
Gorochov AV, Rasnitsyn AP. 2002. Superorder Gryllidea Laicharting,1781. In: Rasnitsyn AP, Quicke DLJ, editors. History of insects.Dordrecht: Kluwer Academic Publishers. p. 293–303.
Gorzula S, Seijas AE. 1989. The common caiman. In: CrocodileSpecialist Group, editor. Crocodiles. Their ecology, management,and conservation. Gland: International Union for Conservation ofNature and Natural Resources. p. 44–61.
Gow CE. 1975. The morphology and relationships of Youngina capensisBroom and Prolacerta broomi Parrington. Palaeontol Afr.18:89–131.
Grande L, Bemis WE. 1996. Interrelationships of Acipenseriformes, withcomments on “Chondrostei”. In: Stiassny MJL, Parenti LR, JohnsonGD, editors. Interrelationships of fishes. San Diego (CA): AcademicPress. p. 85–116.
Greene HW. 1988. Antipredator mechanisms in reptiles. In: Gans C,Huey RB, editors. Biology of the Reptilia. Ecology B. Vol. 16. NewYork: Alan R. Liss. p. 1–152.
Gregory JT. 1945. Osteology and relationships of Trilophosaurus. UnivTexas Publ. 4401:273–359.
Griffiths M. 1978. The biology of the Monotremes. New York: AcademicPress.
Grimaldi DA, Engel MS. 2005. Evolution of the insects. Cambridge:Cambridge University Press.
Grimaldi DA, Engel MS, Nascimbene PC. 2002a. FossiliferousCretaceous amber from Myanmar (Burma): its rediscovery, bioticdiversity, and paleontological significance. Am Mus Nov.3361:1–71.
Grimaldi D, Shedrinsky A, Wampler TP. 2002b. A remarkable deposit offossiliferous amber from the Upper Cretaceous (Turonian) of NewJersey. In: Grimaldi D, editor. Studies on fossils in Amber, withparticular reference to the Cretaceous of New Jersey. Leyden:Backhuys Publishers. p. 1–76.
Gwynne DT. 1995. Phylogeny of the Ensifera (Orthoptera): a hypothesissupporting multiple origins of acoustical signalling, complexspermatophores and maternal care in crickets, katydid, and weta.J Orthop Res. 4:203–218.
Hale RJ, Rentz DFC. 2001. Gryllacrididae: an overview of the worldfauna with emphasis on Australian examples. In: Field LH, editor.The biology of wetas, king crickets, and their allies. Oxford: CABInternational. p. 95–110.
Hedges SB, Poling LL. 1999. A molecular phylogeny of reptiles. Science.283:998–1011.
Henniger HP, Watson WH III. 2005. Mechanisms underlying theproduction of carapace vibrations and associated waterborne soundsin the American lobster, Homarus americanus. J Exp Biol.208:3421–3429.
Henrici AC. 1998. A new pipoid anuran from the Late Jurassic MorrisonFormation at Dinosaur National Monument, Utah. J Vert Paleontol.18:321–332.
Henry CS. 2006. Acoustic communication in neuropterid insects. In:Drosopoulos S, Claridge MF, editors. Insect soundsand communication. Boca Raton (FL): Taylor and Francis.p. 153–166.
Hetherington T. 2008. Comparative anatomy and function of hearing inaquatic amphibians, reptiles, and birds. In: Thewissen JGM,Nummela S, editors. Sensory evolution on the threshold. Adaptationsin secondarily aquatic vertebrates. Berkeley (CA): University ofCalifornia Press. p. 183–209.
Hill JE, Smith JD. 1984. Bats: a natural history. Austin (TX): Universityof Texas Press.
Hjelle JT. 1990. Anatomy and morphology. In: Polis GA, editor. Thebiology of scorpions. Stanford (CA): Stanford University Press.p. 9–63.
Holldobler B, Wilson EO. 1990. The ants. Cambridge (MA): HarvardUniversity Press.
Hope S. 2002. The Mesozoic radiation of Neornithes. In: Chiappe LM,Witmer LM, editors. Mesozoic birds. Above the heads of Dinosaurs.Berkeley (CA): University of California Press. p. 339–388.
Horner JH, Weishampel DB, Forster CA. 2004. Hadrosauridae. In:Weishampel DB, Osmolska H, Dodson P, editors. The Dinosauria.2nd ed. Berkeley (CA): University of California Press. p. 438–463.
Hotton N III, Olson EC, Beerbower R. 1997. Amniote origins and thediscovery of herbivory. In: Sumida SS, Martin KLM, editors.Amniote origins: completing the transition to land. San Diego (CA):Academic Press. p. 207–264.
Hrncir M, Barth FG, Tautz J. 2006. Vibratory and airborne-sound signalsin bee communication (Hymenoptera). In: Drosopoulos S, ClaridgeMF, editors. Insect sounds and communication. Boca Raton (FL):Taylor and Francis. p. 421–436.
Hueter RE, Mann DA, Maruska KP, Sisneros JA, Demski LS. 2004.Sensory biology of elasmobranchs. In: Carrier JC, Musick JA,Heithaus MR, editors. Biology of sharks and their relatives. BocaRaton (FL): CRC Press. p. 325–368.
Hughes B. 1963. The earliest archosaurian reptiles. S Afr J Sci.59:221–241.
Hurum JH, Luo Z, Kielan-Jaworowska Z. 2006. Were mammalsoriginally venomous? Acta Palaeontol Pol. 51:1–11.
Hutchins M. 2004. Grzimek’s animal life encyclopedia. Mammals 1.Vol. 12. 2nd ed. Detroit (MI): Thomson Gale.
Imaizumi K, Pollack GS. 1999. Neural coding of sound frequency bycricket auditory receptors. J Neurosci. 19:1508–1516.
Ivakhnenko MF. 1987. Permian parareptiles of the U.S.S.R. Ak NaukSSSR. 223:1–159.
Iversen RTS, Perkins PJ, Dionne RD. 1963. An indication of underwatersound production by squid. Nature. 199:250–251.
Jeram AJ, Selden PA, Edwards D. 1990. Land animals in the Silurian:arachnids and myriapods from Shropshire, England. Science.250:658–661.
Ji Q, Luo Z, Yuan C, White JR, Zhang J, Georgi JA. 2002. The earliestknown eutherian mammal. Nature. 416:816–822.
Johnston CE, Phillips CT. 2003. Sound production in sturgeonScaphirhynchus albus and S. platyrhynchus (Acipenseridae). EnvBiol Fishes. 68:59–64.
Jones DN, Dekker RWRJ, Roselaar CS. 1995. The megapodes. Oxford:Oxford University Press.
Historical Biology 283
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

Jost MC, Shaw KL. 2006. Phylogeny of Ensifera (Hexapoda: Orthoptera)using three ribosomal loci, with implications for the evolution ofacoustic communication. Mol Phyl Evol. 38:510–530.
Kanmiya K. 2006. Communication by vibratory signals in Diptera. In:Drosopoulos S, Claridge MF, editors. Insect sounds and communi-cation. Boca Raton (FL): Taylor and Francis. p. 381–396.
Kardong KV. 2006. Vertebrates. Comparative anatomy, function,evolution. 4th ed. Boston (MA): McGraw-Hill.
Kear J. 2005. Ducks, geese and swans. Oxford: Oxford University Press.Kelemen G. 1963. Comparative anatomy and performance of the vocal
organ in vertebrates. In: Busnel R-G, editor. Acoust behaviour ofanimals. Amsterdam: Elsevier Publishing Company. p. 489–519.
Kemp A. 1986. The biology of the Australian lungfish, Neoceratodusforsteri (Kreft 1870). J Morphol Suppl. 1:81–198.
Kemp TS. 2005. The origin and evolution of mammals. Oxford: OxfordUniversity Press.
Kemp TS. 2007. Acoustic transformer function of the postdentary bonesand quadrate of a nonmammalian cynodont. J Vert Paleontol.27:431–441.
Kielan-Jaworowska Z. 1978. Evolution of the therian mammals in theLate Cretaceous of Asia. Part II. Postcranial skeleton inZalambdalestidae. Palaeontol Pol. 38:5–41.
Kielan-Jaworowska Z, Cifelli RL, Luo Z. 2004. Mammals from the age ofDinosaurs. New York: Columbia University Press.
Kier PM. 1987. Class Echinoidea. In: Boardman RS, Cheetham AH,Rowell AJ, editors. Fossil invertebrates. Cambridge (MA): Black-well Science. p. 596–611.
King AS. 1989. Functional anatomy of the syrinx. In: King AS,McLelland J, editors. Form and function in birds. Vol. 4. San Diego(CA): Academic Press. p. 105–192.
King AH. 1993. Cephalopoda (Nautiloidea). In: Benton MJ, editor. Thefossil record 2. London: Chapman and Hall. p. 169–188.
Kinney C, Abisahin G, Young BA. 1998. Hissing in rattlesnakes:redundant signaling or inflationary epiphenomenon? J Exp Zool.280:107–113.
Kirchner WH, Broeker I, Tautz J. 1994. Vibrational alarm communi-cation in the damp-wood termite Zootermopsis nevadensis. PhysiolEntomol. 19:187–190.
Kuhn-Schnyder E, Peyer B. 1973. Die Triasfauna der Tessiner KalkalpenXXIII. Tanystropheus longobardicus (Bassani). Schw Palaontol Abh.95:1–155.
Kuhne R, Silver S, Lewis B. 1985. Processing of vibratory signals in thecentral nervous system of the cricket. In: Kalmring K, Elsner N,editors. Acoustic and vibrational communication in insects. Berlin:Verlag Paul Parey. p. 183–192.
Ladich F, Bass AH. 2003. Underwater sound generation and acousticreception in fishes with some notes on frogs. In: Collin SP, MarshallNJ, editors. Sensory processing aquatic environments. New York:Springer. p. 173–193.
Ladich F, Popper AN. 2004. Parallel evolution in fish hearing organs. In:Manley GA, Popper AN, Fay RR, editors. Evolution of the vertebrateauditory system. New York: Springer-Verlag. p. 95–127.
Ladich F, Tadler A. 1988. Sound production in Polypterus (Osteichthyes:Polypteridae). Copeia. 1988:1076–1077.
Laurin M. 1991. The osteology of a Lower Permian eosuchian from Texasand a review of diapsid phylogeny. Zool J Linn Soc. 101:59–95.
Laurin M. 1998. The importance of global parsimony and historical biasin understanding tetrapod evolution. Part I. Systematics, middle earevolution and jaw suspension. Ann Sci Nat. 1:1–42.
Laurin M, Reisz RR. 1995. A reevaluation of early amniote phylogeny.Zool J Linn Soc. 113:165–223.
Laurin M, Reisz RR. 1997. A new perspective on tetrapod phylogeny. In:Sumida SS, Martin KLM, editors. Amniote origins: completing thetransition to land. San Diego (CA): Academic Press. p. 9–59.
Laurin M, Reisz RR. 1999. A new study of Solenodonsaurus janenschi,and a reconsideration of amniote origins and stegocephalianevolution. Can J Earth Sci. 36:1239–1255.
Laurin M, Soler-Gijon R. 2006. The oldest known stegocephalian(Sarcopterygii: Temnospondyli) from Spain. J Vert Paleontol.26:284–299.
Lazzari CR, Manrique G, Schilman PE. 2006. Vibratory communicationin Triatominae (Heteroptera). In: Drosopoulos S, Claridge MF,
editors. Insect sounds and communication. Boca Raton (FL): Taylorand Francis. p. 297–304.
Lee MSY. 1997a. Pareiasaur phylogeny and the origin of turtles. Zool JLinn Soc. 120:197–280.
Lee MSY. 1997b. Reptile relationships turn turtle . . . . Nature.389:245–246.
Leston D, Pringle JWS. 1963. Acoustical behaviour of Hemiptera. In:Busnel R-G, editor. Acoustic behaviour of animals. Amsterdam:Elsevier Publishing Company. p. 391–411.
Locy WA, Larsell O. 1916. The embryology of the bird’s lung, based onobservations of the domestic fowl, part II. Am J Anat. 20:1–44.
Long GR. 1994. Psychobioacoustics. In: Fay RR, Popper AN, editors.Comparative Hearing: Mammals. New York: Springer. p. 18–56.
Luo Z, Crompton AW. 1994. Transformation of the quadrate (incus)through the transition from non-mammalian cynodonts to mammals.J Vert Paleontol. 14:341–374.
Luo Z, Crompton AW, Sun A. 2001. A new mammaliaform from theEarly Jurassic and evolution of mammalian characteristics. Science.292:1535–1540.
Madge S, McGowan P. 2002. Pheasants, partridges, and grouse. Princeton(NJ): Princeton University Press.
Maisch MW. 1997. A case against a diapsid origin of the Ichthyosauria.N Jb Geol Palaontol Abh. 205:111–127.
Makovicky PJ, Currie PJ. 1998. The presence of a furcula intyrannosaurid theropods, and its phylogenetic and functionalimplications. J Vert Paleontol. 18:143–149.
Mannen H, Tsoi SC, Krushkal JS, Li W, Li SS. 1997. The cDNA cloningand molecular evolution of reptile and pigeon lactate dehydrogenaseisozymes. Mol Biol Evol. 14:1081–1087.
Marcellini D. 1977. Acoustic and visual display behavior of gekkonidlizards. Am Zool. 17:251–260.
Maslin PT. 1950. The production of sound in caudate Amphibia. UnivCol Stud Biol Ser. 1:20–45.
Masters WM. 1979. Insect disturbance stridulation: its defensive role.Behav Ecol Sociobiol. 5:187–200.
Mayr GB, Pohl B, Hartmann S, Peters DS. 2007. The tenth skeletalspecimen of Archaeopteryx. Zool J Linn Soc. 149:97–116.
McCauley RD, Cato DH. 2000. Patterns of fish calling in a nearshoreenvironment in the Great Barrier Reef. Phil Trans R Soc Lond B.355:1289–1293.
McCormick CA. 1999. Anatomy of the central auditory pathways of fishand amphibians. In: Fay RR, Popper AN, editors. Comparativehearing: fish and amphibians. New York: Springer-Verlag.p. 155–217.
McCormick SJ, Polis GA. 1990. Prey, predators, and parasites. In: PolisGA, editor. The biology of scorpions. Stanford (CA): StanfordUniversity Press. p. 294–320.
McLelland J. 1989a. Anatomy of the lungs and air sacs. In: King AS,McLelland J, editors. Form and function in birds. Vol. 4. San Diego(CA): Academic Press. p. 221–279.
McLelland J. 1989b. Larynx and trachea. In: King AS, McLelland J,editors. Form and function in birds. Vol. 4. San Diego (CA):Academic Press. p. 69–103.
Menkhorst P, Knight F. 2004. A field guide to the mammals of Australia.South Melbourne: Oxford University Press.
Meunier FJ, Gayet M. 1993. A new polypteriform from the LateCretaceous and the middle Paleocene of South America. In: ArratiaG, Viohl G, editors. Mesozoic fishes. Systematics and paleoecology.Munich: Verlag Dr. Friedrich Pfiel. p. 95–103.
Michelsen A. 1998. Biophysics of sound localization in insects. In: HoyRR, Popper AN, Fay RR, editors. Comparative hearing: insects. NewYork: Springer. p. 18–62.
Milner AR. 1988. The relationships and origin of living amphibians.In: Benton MJ, editor. The phylogeny and classification ofTetrapods. Amphibians, reptiles, birds. Vol. 1. Oxford: ClarendonPress. p. 59–102.
Milner AR. 1993. Amphibian-grade Tetrapoda. In: Benton MJ, editor.The fossil record 2. London: Chapman and Hall. p. 665–679.
Modesto SP, Reisz RR. 2002. An enigmatic new diapsid reptile from theUpper Permian of eastern Europe. J Vert Paleontol. 22:851–855.
Modha ML. 1967. The ecology of the Nile crocodile (Crocodylusniloticus Laurenti) on Central Island, Lake Rudolf. East Afr Wildl J.5:74–95.
P. Senter284
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

Molnar RE. 2005. Sexual selection and sexual dimorphism in theropods.In: Carpenter K, editor. The Carnivorous Dinosaurs. Bloomington(IN): Indiana University Press. p. 284–312.
Moreau CS, Bell CD, Vila R, Archibald B, Pierce NE. 2006. Phylogenyof the ants: diversification in the age of angiosperms. Science.312:101–104.
Morton ES. 1996. A comparison of vocal behavior among tropical andtemperate passerine birds. In: Kroodsma DE, Miller EH, editors.Ecology and evolution of acoustic communication in birds. Ithaca(NY): Comstock Publishing Associates. p. 258–268.
Motani R, Mioura N, Ando T. 1998. Ichthyosaurian relationshipsilluminated by new primitive skeletons from Japan. Nature.393:255–257.
Myhrvold NP, Currie PJ. 1997. Supersonic sauropods? Tail dynamics inthe diplodocids. Paleobiology. 23:393–409.
Myrberg AA Jr. 1981. Sound communication and interception infishes. In: Tavolga WN, Popper AN, Fay RR, editors. Hearing andsound communication in fishes. New York: Springer-Verlag.p. 395–425.
Muller J, Tsuji LA. 2007. Impedance-matching hearing in Paleozoicreptiles: evidence of advanced sensory perception at an early stage ofamniote evolution. PLoS ONE. 9:1–7.
Negus VE. 1949. The comparative anatomy and physiology of theLarynx. New York: Hafner.
Nelson JB. 2005. Pelicans, cormorants, and their relatives. Oxford:Oxford University Press.
Nolf D, Stringer GL. 1993. Cretaceous fish Otoliths – a synthesis of theNorth American record. In: Arratia G, Viohl G, editors. Mesozoicfishes. Systematics and Paleoecology. Munich: Verlag Dr. FriedrichPfiel. p. 433–459.
Norell MA, Makovicky PJ, Clark JM. 1997. A Velociraptor wishbone.Nature. 389:447.
Nowak RM. 2005. Walker’s marsupials of the world. Baltimore (MI):Johns Hopkins University Press.
Nowak RM, Paradiso JL. 1983. Walker’s mammals of the world. 4th ed.Baltimore (MD): Johns Hopkins University Press.
O’Connor PM. 2006. Postcranial pneumaticity: an evaluation of soft-tissue inferences on the postcranial skeleton and the reconstruction ofpulmonary anatomy in archosaurs. J Morphol. 267:1199–1226.
O’Neill KM. 2001. Solitary Wasps: behavior and natural history. Ithaca(NY): Cornell University Press.
Oberholser HC. 1974. The bird life of Texas. Austin (TX): University ofTexas at Austin Press.
Ortega F, Gasparini Z, Buscalioni AD, Calvo JO. 2000. A new species ofAraripesuchus (Crocodylomorpha, Mesoeucrocodylia) from theLower Cretaceous of Patagonia. J Vert Paleontol. 20:57–76.
Palci A, Caldwell MW. 2007. Vestigial forelimbs and axial elongation ina 95 million-year-old non-snake squamate. J Vert Paleontol. 27:1–7.
Patek SN. 2001. Spiny lobsters stick and slip to make sound. Nature.411:153–154.
Patterson C. 1993. Osteichthyes: Teleostei. In: Benton MJ, editor. Thefossil record 2. London: Chapman and Hall. p. 621–656.
Pearman JV. 1928. On sound production in the Psocoptera and on apresumed stridulatory organ. Entomol Monthly Mag. 64:179–186.
Penney D, Wheater CP, Selden PA. 2003. Resistance of spiders toCretaceous-Tertiary extinction events. Evolution. 57:2599–2607.
Platz JE, Conlon JM. 1997. . . . and back again. Nature. 389:246.Poinar GO Jr, Danforth BN. 2006. A fossil bee from Early Cretaceous
Burmese amber. Science. 314:614.Ponomarenko AG. 2002. Superorder Scarabaeidea Laicharting, 1781.
Order Coleoptera Linne, 1758. The beetles. In: Rasnitsyn AP,Quicke DLJ, editors. History of insects. Dordrecht: KluwerAcademic Publishers. p. 164–176.
Popper AN, Platt C, Edds PL. 1992. Evolution of the vertebrate inner ear:an overview of ideas. In: Webster DB, Fay RR, Popper AN, editors.The evolutionary biology of hearing. New York: Springer-Verlag.p. 49–57.
Pough FH, Andrews RM, Cadle JE, Crump ML, Savitzky AH, Wells KD.1998. Herpetology. Upper Saddle River (NJ): Prentice Hall.
Rasnitsyn AP. 2002. Superorder Psocidea Leach, 1814. In: Rasnitsyn AP,Quicke DLJ, editors. History of insects. Dordrecht: KluwerAcademic Publishers. p. 125–131.
Reese AM. 1914. The alligator and its allies. New York: G.P. Putnam’sSons.
Reisz R. 1981. A diapsid reptile from the Pennsylvanian of Kansas. UnivKans Mus Nat Hist Sp Publ. 7:1–74.
Reynoso VH, Clark JM. 1998. A dwarf sphenodontian from the JurassicLa Boca Formation of Tamaulipas, Mexico. J Vert Paleontol.18:333–339.
Rieppel O, deBraga M. 1996. Turtles as diapsid reptiles. Nature.384:453–455.
Robbins CS, Bruun B, Zim HS. 1983. Birds of North America. A guide tofield identification. New York: Golden Press.
Robert D, Hoy RR. 2000. The evolutionary innovation of tympanalhearing in Diptera. In: Hoy RR, Popper AN, Fay RR, editors.Comparative hearing: insects. New York: Springer. p. 197–227.
Robinson PL. 1962. Gliding lizards from the Upper Keuper of GreatBritain. Proc Geol Soc Lond. 1601:137–146.
Romer AS. 1947. Review of the Labyrinthodontia. Bull Mus Comp Zool.99:1–368.
Ross AJ, Jarzembowski EA. 1993. Arthropoda (Hexapoda; Insecta). In:Benton MJ, editor. The fossil record 2. London: Chapman and Hall.p. 363–426.
Roth LM, Hartman HB. 1967. Sound production and its evolutionarysignificance in the Blattaria. Ann Enomol Soc Am. 60:740–752.
Rougier GW, de la Fuente MS, Arcucci AB. 1995. Late Triassic turtlesfrom South America. Science. 268:855–858.
Rougier GW, Wible JR, Novacek MJ. 1998. Implications ofDeltatheridium specimens for early marsupial history. Nature.396:459–463.
Rudinsky JA, Ryker LC. 1976. Sound production in Scolytidae: rivalryand premating stridulation of male douglas-fir beetle. J Ins Physiol.22:997–1003.
Ruppert EE, Barnes RD. 1994. Invertebrate zoology. 6th ed. Fort Worth:Saunders College Publishing.
Russell DA. 1967. Systematics and morphology of American mosasaurs.Peabody Mus Nat Hist Bull. 23:1–240.
Rust J, Stumpner A, Gottwald J. 1999. Singing and hearing in a Tertiarybushcricket. Nature. 399:650.
Ruta M, Coates MI, Quicke DLJ. 2003. Early tetrapod relationshipsrevisited. Biol Rev. 78:251–345.
Sanchiz B. 1998. Encyclopedia of paleoherpetology, part 4, Salientia.Munich: Verlag Dr. Friedrich Pfeil.
Schneider H. 1967. Morphology and physiology of sound-producingmechanisms in teleost fishes. In: Tavolga WN, editor. Marine bio-acoustics. Vol. 2. Oxford: Pergamon Press. p. 135–158.
Schram FR. 1975. A Pennsylvanian lepadomorph barnacle from theMazon Creek area, Illinois. J Paleontol. 49:928–930.
Schwartz JJ. 2001. Call monitoring and interactive playback systems inthe study of acoustic interactions among male anurans. In: Ryan MJ,editor. Anuran communication. Washington, DC: SmithsonianInstitution Press. p. 183–204.
Schwarzhans W. 1993. Otoliths from the Maastrichtian of Bavaria andtheir evolutionary significance. In: Arratia G, Viohl G, editors.Mesozoic fishes. Systematics and paleoecology. Munich: VerlagDr. Friedrich Pfiel. p. 417–431.
Schwimmer DR. 2002. King of the Crocodylians. Bloomington (IN):Indiana University Press.
Selden PA. 1993. Arthropoda (Aglaspidida, Pycnogonida and Chelicer-ata). In: Benton MJ, editor. The fossil record 2. London: Chapmanand Hall. p. 297–320.
Selden PA, Dunlop JA. 1998. Fossil taxa and relationships of chelicerates.In: Edgecombe GD, editor. Arthropod fossils and phylogeny. NewYork: Columbia University Press. p. 303–331.
Senter P. 2003. New information on cranial and dental features of theTriassic archosauriform reptile Euparkeria capensis. Palaeontology.46:613–621.
Senter P. 2004. Phylogeny of Drepanosauridae (Reptilia: Diapsida). JSyst Palaeont. 2:257–268.
Senter P. 2008. Homology between and antiquity of stereotypedcommunicatory behaviors of crocodilians. J Herpetol. 42:354–360.
Sereno PC. 1991. Basal archosaurs: phylogenetic relationships andfunctional implications. Soc Vert Paleont Mem. 2:1–53.
Historical Biology 285
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

Sereno PC, Larsson HCE, Sidor CA, Gado B. 2001. The giantcrocodyliform Sarcosuchus from the Cretaceous of Africa. Science.294:1516–1519.
Sereno PC, Martinez RN, Wilson JA, Varricchio DJ, Alcober OA,Larsson HCE. 2008. Evidence for avian intrathoracic air sacs in anew predatory dinosaur from Argentina. PLoS One. 3(9):1–20.
Shaffer HB, Meylan P, McKnight ML. 1997. Tests of turtle phylogeny:molecular, morphological, and paleontological approaches. SystBiol. 46:235–268.
Shcherbakov DE, Popov YA. 2002. Superorder Cimicidea Laicharting,1781. Order Hemiptera Linne, 1758. The bugs, cicadas, plantlice,scale insects, etc. In: Rasnitsyn AP, Quicke DLJ, editors. History ofinsects. Dordrecht: Kluwer Academic Publishers. p. 143–157.
Shear WA. 1997. The fossil record and evolution of the Myriapoda. In:Fortey RA, Thomas RH, editors. Arthropod relationships. London:Chapman and Hall. p. 211–219.
Shear WA, Selden PA, Rolfe WDI, Bonamo PM, Grierson JD. 1987. Newterrestrial arachnids from the Devonian of Gilboa, New York(Arachnida, Trigonotarbida). Am Mus Nov. 2901:1–74.
Simmons JA, Wever EG, Strother WF, Pylka JM, Long GR. 1971.Acoustic behavior of three sympatric species of 17-yr cicadas. JAcoust Soc Am. 49:93.
Singh LAK, Rao RJ. 1990. Territorial behaviour of male gharial (Gavialisgangeticus) in the National Chambal Sanctuary, India. J Bombay NatHist Soc. 87:149–151.
Sinitshenkova ND. 2002. Order Perlida Latreille, 1810. The stoneflies(¼Plecoptera Burmeister, 1839). In: Rasnitsyn AP, Quicke DLJ,editors. History of insects. Dordrecht: Kluwer Academic Publishers.p. 281–287.
Sissom WD. 1990. Systematics, biogeography, and paleontology. In:Polis GA, editor. The biology of scorpions. Stanford (CA): StanfordUniversity Press. p. 64–160.
Skelton PW, Benton MJ. 1993. Mollusca: Rostroconchia, Scaphopoda,and Bivalvia. In: Benton MJ, editor. The fossil record 2. London:Chapman and Hall. p. 237–263.
Smotherman M, Narins P. 2004. Evolution of the amphibian ear. In:Manley GA, Popper AN, Fay RR, editors. Evolution of the vertebrateauditory system. New York: Springer-Verlag. p. 164–193.
Sotavalta O. 1963. The flight-sounds of insects. In: Busnel R-G, editor.Acoustic behaviour of animals. Amsterdam: Elsevier PublishingCompany. p. 374–390.
Stewart JD. 1993. Cretaceous acanthomorphs from North America. In:Arratia G, Viohl G, editors. Mesozoic fishes. Systematics andpaleoecology. Munich: Verlag Dr. Friedrich Pfiel. p. 383–394.
Stewart KW, Sandberg JB. 2006. Vibratory communication and matesearching in stonefiles. In: Drosopoulos S, Claridge MF, editors.Insect Sounds and Communication. Boca Raton (FL): Taylor andFrancis. p. 179–186.
Storrs GW. 1997. Morphological and taxonomic clarification of the genusPlesiosaurus. In: Callaway JM, Nicholls EL, editors. Ancient marinereptiles. San Diego (CA): Academic Press. p. 145–190.
Sweet MH. 1996. Comparative external morphology of the pregenitalabdomen of the Hemiptera. In: Schaefer CW, editor. Studies onHemipteran Phylogeny. Lanham, MD: Entomological Society ofAmerica. p. 119–158.
Tembrock G. 1963. Acoustic behaviour of mammals. In: Busnel R-G,editor. Acoustic behaviour of animals. Amsterdam: ElsevierPublishing Company. p. 751–786.
Thorbjarnarson JB. 1989. Ecology of the American crocodile, Crocodylusacutus. In: Crocodile Specialist Group, editor. Crocodiles. Theirecology, management, and conservation. Gland: International Unionfor Conservation of Nature and Natural Resources. p. 228–258.
Thorbjarnarson JB. 1991. Crocodylus acutus (American crocodile).Social behavior. Herpetol Rev. 22:130.
Thorbjarnarson JB, Hernandez G. 1993. Reproductive ecology of theOrinoco crocodile (Crocodylus intermedius) in Venezuela. II.Reproductive and social behavior. J Herpetol. 27:371–379.
Trueb L, Cloutier R. 1991. A phylogenetic investigation of the inter- andintrarelationships of the Lissamphibia (Amphibia: Temnospondyli).In: Schultze H-P, Trueb L, editors. Origins of the higher groups ofTetrapods. Controversy and consensus. Ithaca (NY): ComstockPublishing Associates. p. 223–313.
Tsuji LA. 2006. Cranial anatomy and phylogenetic affinities of thePermian parareptile Macroleter poezicus. J Vert Paleontol.26:849–865.
Uetz GW, Stratton GE. 1981. Acoustic communication and reproductiveisolation in spiders. In: Witt PN, Rover JS, editors. Spidercommunication. Mechanisms and ecological significance. Princeton(NJ): Princeton University Press. p. 123–159.
Unwin DM. 1993. Aves. In: Benton MJ, editor. The fossil record 2.London: Chapman and Hall. p. 717–737.
Unwin DM. 2006. The Pterosaurs: from deep time. New York: Pi Press.Upchurch P, Barrett PM, Dodson P. 2004. Sauropoda. In: Weishampel
DB, Dodson P, Osmolska H, editors. The Dinosauria. 2nd ed.Berkeley (CA): University of California Press. p. 259–324.
Vallin G, Laurin M. 2004. Cranial morphology and affinities ofMicrobrachis, and a reappraisal of the phylogeny and lifestyle of thefirst amphibians. J Vert Paleontol. 24:56–72.
van Tuinen M, Sibley CG, Hedges SB. 2000. The early history of modernbirds inferred from DNA sequences of nuclear and mitochondrialribosomal genes. Mol Biol Evol. 17:451–457.
Vater M, Meng J, Fox RC. 2004. Hearing organ evolution andspecialization: early and later mammals. In: Manley GA, Popper AN,Fay RR, editors. Evolution of the vertebrate auditory system. NewYork: Springer-Verlag. p. 256–288.
Vincent F. 1963. Acoustic signals for auto-information of echolocation.In: Busnel R-G, editor. Acoustic behaviour of animals. Amsterdam:Elsevier Publishing Company. p. 183–227.
Virant-Doberlet M, Cokl A. 2004. Vibratory communication in insects.Neotrop Entomol. 33:121–134.
Vliet KA. 1989. Social displays of the American alligator (Alligatormississippiensis). Am Zool. 29:1019–1031.
von Dohlen CD, Moran NA. 1995. Molecular phylogeny of theHomoptera: a paraphyletic taxon. J Mol Evol. 41:211–223.
Wang Y, Hu Y, Li C. 2001. An ossified Meckel’s cartilage in twoCretaceous mammals and the origin of the mammalian middle ear.Science. 294:357–361.
Wedel MJ. 2006. Origin of postcranial skeletal pneumaticity in dinosaurs.Integr Zool. 2006:80–85.
Weishampel DB. 1981. Acoustic analysis of potential vocalization inlambeosaurine dinosaurs (Reptilia: Ornithischia). Paleobiology.7:252–261.
Welty JC, Baptista L. 1988. The life of birds. 4th ed. Fort Worth (TX):Harcourt Brace Jovanovich.
Wessel A. 2006. Stridulation in the Coleoptera – an overview. In:Drosopoulos S, Claridge MF, editors. Insect soundsand communication. Boca Raton (FL): Taylor and Francis.p. 397–403.
Wever EG. 1978. The reptile ear. Princeton (NJ): Princeton UniversityPress.
Wever EG. 1985. The amphibian ear. Princeton (NJ): PrincetonUniversity Press.
Whitaker R, Basu D. 1982. The gharial (Gavialis gangeticus): a review. JBomb Nat Hist Soc. 79:531–548.
Whitaker R, Whitaker Z. 1989. Ecology of the mugger crocodile. In:Crocodile Specialist Group, editor. Crocodiles. Their ecology,management, and conservation. Gland: International Union forConservation of Nature and Natural Resources. p. 276–296.
Wills LJ. 1963. Cyprilepas holmi Wills 1962, a peduncular cirrepedefrom the Upper Silurian of Oesel, Esthonia. Palaeontology.6:161–165.
Wilson B, Batty RS, Dill LM. 2003. Pacific and Atlantic herring produceburst pulse sounds. Proc R Soc Lond B Suppl. 271:S95–S97.
Wu X. 1994. Late Triassic-Early Jurassic sphenodontians from China andthe phylogeny of the Sphenodontia. In: Fraser NC, Sues H-D, editors.In the shadow of the Dinosaurs. Early Mesozoic Tetrapods.Cambridge: Cambridge University Press. p. 38–69.
Xu X, Norell MA. 2004. A new troodontid dinosaur from China withavian-like sleeping posture. Nature. 431:838–841.
Yager DD. 1999. Structure, evolution, and development of insect auditorystructures. Microsc Res Techn. 47:380–400.
Yan HY, Fine ML, Horn NS, Colon WE. 2000. Variability in the role ofthe gas bladder in fish audition. J Comp Physiol A. 186:435–445.
Young BA. 1991. Morphological basis of “growling” in the king cobra,Ophiophagus hannah. J Exp Zool. 260:275–287.
P. Senter286
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009

Young BA. 1998. How to hiss loudly: on narial flaring and acousticamplification in Russell’s viper (Daboia russellii). Hamadryad.23:103–110.
Young BA, Sheft S, Yost W. 1995. Sound production in Pituophismelanoleucus (Serpentes: Colubridae) with the first description of avocal cord in snakes. J Exp Zool. 273:472–481.
Zardoya R, Meyer A. 2001. On the origin of and phylogeneticrelationships among living amphibians. Proc Natl Ac Sci USA.98:7380–7383.
Zelick R, Mann DA, Popper AN. 1999. Acoustic communication in fishesand frogs. In: Fay RR, Popper AN, editors. Comparative hearing: fishand Amphibians. New York: Springer-Verlag. p. 363–411.
Zhou Z, Zhang F. 2003. Anatomy of the primitive bird Sapeornischaoyangensis from the Early Cretaceous of Liaoning, China. Can JEarth Sci. 40:731–747.
Zhu M, Zhao W, Jia L, Lu J, Qiao T, Qu Q. 2009. The oldest articulatedosteicthyan reveals mosaic gnathostome characters. Nature.458:469–474.
Historical Biology 287
Downloaded By: [Canadian Research Knowledge Network] At: 16:50 18 September 2009