
Reversibilite de la dedifferenciation observee dan les s ... · reposant sur une grille de type...
14
/. Embryol. exp. Morph. Vol. 35, 2, pp. 227-240, 1976 227 Printed in Great Britain Reversibilite de la dedifferenciation observee dans les cultures cellulaires de foie embryonnaire d'oiseau ParE. HOUSSAINT 1 Laboratoire d'Embryologie, Universite de Nantes, France SUMMARY Reversibility of the morphological and functional dedifferentiation occurring in cultures of avion embryonic liver cells Primary cell cultures are established from 8-day quail embryo livers. During the first three days the culture is made up of areas of epithelial-like cells and scattered fibroblasts. The cytoplasm of the epithelial cells shows a high glycogen content as detected by the PAS reaction controlled with salivary amylase digestion. During the following days an im- portant increase in the number of fibroblastic cells is observed. After 6-7 days of cultivation, the epithelial cells have disappeared and the culture is entirely fibroblastic. PAS technique does not show any trace of glycogen in these cultures which have been prolonged up to 45 days. Six- to 45-day primary cultures entirely made up of fibroblasts were associated with hepatic or pulmonary mesenchyme in organotypic culture for 3-4 days. In some cases the explant was first cultivated in vitro for 2 days and then grafted into a 5-day-old chick embryo on the chorioallantoic membrane for 6 days. In the secondary cultures hepatocytes showing an epithelial arrangement and a high glycogen content were observed. It appears from this observation that some of the primary culture fibroblasts are in fact dedifferentiated parenchymal cells. Such a dedifferentiation is a reversible phenomenon since the cells retain the ability to express their initial determination if they are placed in convenient environmental conditions. The role of the specific tissular arrangement inthe stability of the differentiated state is discussed. INTRODUCTION De nombreux auteurs ont tente d'etablir des lignees cellulaires a partir de cultures de tissu hepatique d'oiseaux et de mammiferes. Les cellules du foie de mammifere peuvent etre cultivees pendant de longues periodes et des cultures clonales ont meme pu etre obtenues a partir de foie de rat nouveau ne (Coon, 1969; Lambiotte, Susor & Cahn, 1972; Chessebeuf et ah 1974); de foie de souris (Evans et ah 1952; Waymouth, Chen &Wood, 1971); de foie humain foetal ou adulte (Chang, 1954) et de foie de veau (Pieck &Kuyper, 1961). Le but des recherches effectuees dans ce domaine etait d'etablir des cultures massives de cellules hepatiques metaboliquement actives. Le plus souvent les 1 Adresse de Vauteur: Laboratoire d'Embryologie, Universite de Nantes, U.E.R. des Sciences de la Nature, 38 Boulevard Michelet, B.P. 1044-44037 Nantes Cedex, France. 15-2
Transcript of Reversibilite de la dedifferenciation observee dan les s ... · reposant sur une grille de type...

/ . Embryol. exp. Morph. Vol. 35, 2, pp. 227-240, 1976 2 2 7
Printed in Great Britain
Reversibilite de la dedifferenciationobservee dans les cultures cellulaires de foie
embryonnaire d'oiseau
ParE. HOUSSAINT1
Laboratoire d'Embryologie, Universite de Nantes, France
SUMMARY
Reversibility of the morphological and functional dedifferentiation occurringin cultures of avion embryonic liver cells
Primary cell cultures are established from 8-day quail embryo livers. During the first threedays the culture is made up of areas of epithelial-like cells and scattered fibroblasts. Thecytoplasm of the epithelial cells shows a high glycogen content as detected by the PASreaction controlled with salivary amylase digestion. During the following days an im-portant increase in the number of fibroblastic cells is observed. After 6-7 days of cultivation,the epithelial cells have disappeared and the culture is entirely fibroblastic. PAS technique doesnot show any trace of glycogen in these cultures which have been prolonged up to 45 days.
Six- to 45-day primary cultures entirely made up of fibroblasts were associated with hepaticor pulmonary mesenchyme in organotypic culture for 3-4 days. In some cases the explantwas first cultivated in vitro for 2 days and then grafted into a 5-day-old chick embryo on thechorioallantoic membrane for 6 days. In the secondary cultures hepatocytes showing anepithelial arrangement and a high glycogen content were observed.
It appears from this observation that some of the primary culture fibroblasts are in factdedifferentiated parenchymal cells. Such a dedifferentiation is a reversible phenomenon sincethe cells retain the ability to express their initial determination if they are placed in convenientenvironmental conditions. The role of the specific tissular arrangement in the stability of thedifferentiated state is discussed.
INTRODUCTION
De nombreux auteurs ont tente d'etablir des lignees cellulaires a partir decultures de tissu hepatique d'oiseaux et de mammiferes. Les cellules du foie demammifere peuvent etre cultivees pendant de longues periodes et des culturesclonales ont meme pu etre obtenues a partir de foie de rat nouveau ne (Coon,1969; Lambiotte, Susor & Cahn, 1972; Chessebeuf et ah 1974); de foie de souris(Evans et ah 1952; Waymouth, Chen & Wood, 1971); de foie humain foetal ouadulte (Chang, 1954) et de foie de veau (Pieck & Kuyper, 1961).
Le but des recherches effectuees dans ce domaine etait d'etablir des culturesmassives de cellules hepatiques metaboliquement actives. Le plus souvent les
1 Adresse de Vauteur: Laboratoire d'Embryologie, Universite de Nantes, U.E.R. desSciences de la Nature, 38 Boulevard Michelet, B.P. 1044-44037 Nantes Cedex, France.
15-2

228 E. HOUSSAINT
cultures de foie de mammifere sont constitutes de cellules presentant un aspectepithelial. Cependant, bien que dans certains cas il ait ete possible de decelerdans les cellules certaines proprietes des hepatocytes (production de proteinesseriques - Coon, 1969; Namba, Hirox & Kishimoto, 1968; Kaighn & Prince,1971; Waymouth et al. 1971; Bissell & Tilles, 1971; presence de glycogene-Gerschenson, Andersson & Molson, 1970; existence de la forme B de l'aldolase- Chessebeuf et al. 1974), le plus generalement les caracteristiques biochimiqueset fonctionnelles du tissu hepatique disparaissent rapidement en culture (Evanset al. 1952; Perske, Parks & Walker, 1957; Auerbach & Walker, 1959; Hillis &Bang, 1962; Watanabe, 1966; Fogel, 1968; Rose, Kumegawa & Cattoni, 1968;Alexander & Grisham, 1970; Lambiotte et al. 1972).
L'explantation de foie embryonnaire de poulet n'a donne lieu qu'a descultures de duree limitee ne supportant pas de repiquages successifs (Kuroda &Nagatani, 1965; Angulo, 1968; Laschi & Rizzoli, 1968; Verne & Hebert, 1968;Skea & Nemeth, 1969; Lambiotte, 1970; Verne, Hebert, Richshoffer & Roux,1971). Elles sont constitutes au cours des premiers jours, de plages epithelialeset de cellules d'aspect fibroblastique qui deviennent rapidement predominates.Apres quelques jours, les plages epitheliales disparaissent et les fibroblastes sontalors le seul type cellulaire de la culture.
II apparait done que les conditions de la culture cellulaire sont defavorablesau maintien de l'activite fonctionnelle de la cellule hepatique. Nous noussommes pose la question de savoir si les fibroblastes resultant de l'explantationprolongee du foie d'oiseau derivaient exclusivement des cellules endotheliales etconjonctives du tissu hepatique ou si certains d'entre eux pouvaient provenir decellules parenchymateuses morphologiquement et physiologiquement dedif-ferenciees. Dans ce dernier cas il etait interessant de rechercher si la dedifferen-ciation imposee par les conditions de la culture etait ou non reversible.
Dans des recherches precedentes de notre groupe le role de la composantemesodermique du foie sur la differenciation de l'endoderme en hepatocytes etsur le maintien de l'activite fonctionnelle de ces cellules a ete demontre (LeDouarin, 1964; Le Douarin & Houssaint, 1967; Le Douarin, 1968; Houssaint,1972). De plus il a ete etabli que l'ensemble du mesoderme des lames lateralespeut jouer le meme role que le mesenchyme propre du foie sur la differenciationdes cellules endodermiques en parenchyme hepatique (Le Douarin, Bussonnet& Chaumont, 1968).
Dans le present travail nous avons etudie revolution morphologique et lecontenu en glycogene des cellules hepatiques d'embryons de caille de 8 joursdissociees par action de la trypsine puis transplanted en milieu liquide. Letemps de culture maximum a ete de 45 jours. Lorsque les cultures ont atteintun etat fibroblastique homogene nous les avons associees en culture organo-typique avec du mesenchyme d'origine splanchnique et nous avons pu montrerque des cellules hepatiques fonctionnelles derivant de la culture primaireapparaissent dans les explants.

Reversibilite de la dedifferenciation 229
MATERIEL ET METHODES
(a) Cultures pritnaires de cellules hepatiques
Les cultures cellulaires sont realisees a partir de foies d'embryons de caille de 8jours. Les organes preleves sterilement sont decoupes en minces fragments dansdu liquide de Tyrode puis incubes a 37 °C, pendant 30 min, dans une solution detrypsine (NBC) a 0,25 % dans du liquide de Tyrode sans Ca ni Mg. Au cours decette incubation, la solution est agitee a trois reprises par pipetages repetes.Apres nitration sur gaze, les cellules isolees ainsi que les petits agregats cellu-laires sont laves deux fois dans du milieu de Eagle additionne de 20 % de serumfoetal de veau. Les cellules sont alors mises en culture dans des boites de PetriFalcon de 35 mm de diametre, a raison de 106 cellules par ml de milieu. Onplace le plus souvent une lamelle de verre dans le fond du recipient de culture.Nous avons egalement realise des cultures dans des boites dont le fond estrecouvert d'un film de collagene. Nous utilisons le milieu MEM de Eagle(Wellcome) additionne de 10 % de serum foetal de veau. De la penicilline(100 unites/ml) et de la streptomycine (lOO^g/ml) sont ajoutees au milieu. Lescultures sont incubees en atmosphere humide dans une etuve Nacional acirculation gazeuse (95 % air-5 % CO2) et le milieu de culture est renouvele tousles trois jours. Les cultures sont observees avec un microscope a contraste dephase Wild M40.
(b) Obtention des mesenchymes hepatique et pulmonaire
Mesenchyme hepatique
La technique qui permet d'obtenir du mesenchyme hepatique a l'etat pur aete mise au point par Le Douarin (1964). On peut isoler la partie posterieure del'aire presomptive du mesenchyme hepatique en empechant experimentalementla progression des cellules endodermiques dans ce territoire. Pour cela, onexcise un fragment de l'aire laterale au niveau des somites anterieurs, au-dessousdu niveau de la levre anterieure de l'ombilic intestinal, chez des embryons de6 a 13 somites (Fig. 1 A). La progression caudale des cordons endodermiques estarretee par Foperation. En arriere du territoire excise, du mesenchyme hepatiquese developpe; il est preleve du 6eme au 9eme jour de l'incubation. Le mesenchymehepatique ainsi obtenu est dans tous les cas totalement depourvu d'hepatocytes(Fig. 1B). Ceci a ete eprouve au cours de nombreuses experiences realisees avecle mesenchyme propre du foie isole experimentalement (Le Douarin, 1964;Houssaint & Le Douarin, 1971; Houssaint, 1972) (Fig. 1A, B).
Mesenchyme pulmonaire
On preleve les ebauches pulmonaires sur des embryons de poulet de 4 jours.La separation du mesenchyme et de la bronche est obtenue par action d'unesolution de trypsine a 1 % dans du liquide de Tyrode sans Ca ni Mg selon une

230 E. HOUSSAINT
mh
B
Fig. 1. (A) Isolement du mesenchyme hepatique (mh). On excise un fragment del'aire laterale (ex) au niveau des somites anterieurs./: region ou le foie se differencienormalement. (D'apres Le Douarin, 1964.) (B) Mesenchyme hepatique preleve surun embryon de poulet de 6£ jours. Les travees mesenchymateuses (tm) sont depour-vues d'hepatocytes. Hematoxyline-eosine. x 315.
technique decrite par Dameron (1961). Le mesenchyme est rince dans duTyrode normal additionne de 10 % de serum de poulain pour epuiser la trypsineavant la mise en culture.
(c) Cultures secondaires
Dans une premiere serie experimentale, les cellules hepatiques cultivees pen-dant 6 a 45 jours sont decollees de la lamelle de verre, a laquelle elles adherent, al'aide d'une spatule puis elles sont retransplantees soit sur un milieu semi-solide(Wolff & Haffen, 1952) soit sur un filtre millipore pose sur une grille de typeTrowell. Dans ce dernier cas, le milieu a la meme composition que celui que nousutilisons pour les cultures primaires de cellules hepatiques (milieu MEM deEagle +10 % de serum foetal de veau). L'explant est cultive pendant 3 a 4 joursdans ces conditions puis fixe pour l'examen histologique.
Dans une deuxieme serie experimentale, nous avons realise des associations decultures primaires avec le mesenchyme hepatique ou pulmonaire. La cultureprimaire que Ton a laisse evoluer pendant 6 a 45 jours, est alors placee en cultureorganotypique au contact d'un mesenchyme hepatique ou pulmonaire. Cetteassociation est realisee sur un milieu de Wolff et Haffen ou sur un filtre millipore

Reversibilite de la de differentiation 231
Culture primaire(6 a 45 jours)
mh
Culturesecondaire
(3 a 5 jours)
Greffe coelomiqueou
Greffe chorioallantoidienne
Fig. 2. Realisation de cultures secondaires par association de cultures primaires de6 a 45 jours avec du mesenchyme hepatique. L'association est realisee in vitropendant 3 a 4 jours. Dans certains cas elle est prolongee en greffe in ovo pendant6 jours.
reposant sur une grille de type Trowell. L'explant est cultive pendant 3 a 4 jourspuis fixe par le liquide de Gendre. Dans certaines series experimentales, l'explantest cultive 1 ou 2 jours puis retransplante in ovo pendant 6 jours dans le ccelomed'un embryon de poulet de 3 jours d'incubation ou sur la membrane chorio-allantoidienne (CAM) d'un embryon de 6 jours (Fig. 2).
(d) Detection du glycogene
Les lamelles portant les cultures cellulaires, ainsi que les explants resultant dela combinaison des cultures primaires et du mesenchyme hepatique ou pul-monaire sont fixes par le liquide de Gendre a - 20 °C pendant 10 min, puis 20min a 4 °C. La deshydratation est egalement effectuee a 4 °C. Des resultatsanterieurs ont en effet montre que la fixation par le liquide de Gendre pratiqueea basse temperature et tres rapidement, permet de preserver de tres petitesquantites de glycogene (Houssaint et Le Douarin, 1974). Les tissus sontensuite soumis a la reaction a l'acide periodique Schiff (APS) selon la methodede Hotchkiss-MacManus (1948), en vue de la detection du glycogene, puispost colores par l'hematoxyline de Groat.

232 E. HOUSSAINT

Reversibilite de la dedifferenciation 233
RESULTATS
/. Evolution des cultures primaires d'hepatocytes
Observation au microscope a contraste de phase
Quelques heures apres leur mise en culture, les cellules adherent a la lamelle deverre placee au fond de la boite de Petri. Dans les cultures de 12 h on observe depetites plages epitheliales constitutes de 8 a 10 cellules arrondies pourvues d'ungros nucleole. Entre ces plages sont dispersees quelques volumineuses cellulesisolees d'aspect fibroblastique possedant un noyau hypertrophie. Ces cellulesse multiplient rapidement et apres 48 heures les plages epitheliales sont beaucoupplus larges tandis que les cellules fibroblastiques deviennent beaucoup plusnombreuses (Fig. 3 A).
A partir du 4 erne jour, les cellules de type fibroblastique prennent de plus enplus d'importance par rapport aux cellules epitheliales. Apres 96 h, la cultureforme une couche continue constitute en majorite de fibroblastes. On reconnaitencore a ce stade quelques grandes plages de cellules epitheliales dont le cyto-plasme apparait charge de vacuoles. Au bord des plages epitheliales on observedes cellules d'aspect intermediate a noyau ovale qui semblent s'echapper.Les cellules continuent a se multiplier activement formant meme plusieurscouches superposees dans certaines zones. Apres 6 a 7 jours les cellules epi-theliales ne sont pratiquement plus decelables et la culture revet un aspectuniquement fibroblastique (Fig. 4A) qu'elle gardera par la suite. Les culturesrealisees sur substrat de collagene montrent la meme evolution. Certaines cul-tures ont ete prolongees jusqu'a 45 jours. Des essais de repiquage, par dissocia-tion de la culture primaire sous Faction de la trypsine, se sont soldes par unechec.
L'evolution des cultures de cellules hepatiques d'embryons de caille permetdone d'observer une disparition progressive des cellules epitheliales tandis queles fibroblastes deviennent les elements dominants puis exclusifs de la culture.
FIGURES 3 ET 4
Cultures primaires realisees a partir de foies d'embryon de caille de 8 jours.Fig. 3(A). Culture de 48 h observee au microscope a contraste de phase. Quelquesvolumineux fibroblastes (/) sont observes au bord des plages epitheliales (c.ep). x 540.(B). Culture de 48 h traitee par la reaction a TAPS. Certaines cellules epitheliales(c.ep.) sont riches en glycogene, d'autres en sont deja depourvues. La reaction estnegative au niveau des fibroblastes (/). APS-hematoxyline. x 125.Fig. 4(A). Culture primaire de 6 jours observee au contraste de phase. Toutes lescellules ont un aspect fibroblastique. x 540.(B). Culture primaire de 6 jours traitee par la reaction a TAPS. La reaction estnegative. Le cytoplasme des fibroblastes contient seulement des mucopolysacchari-des qui ne disparaissent pas apres action de l'amylase salivaire. APS-hematoxyline.x315.
234 E. HOUSSAINT

Reversibilite de la dedifferenciation 235
Detection du glycogene dans les cultures d'hepatocytes
La reaction a TAPS appliquee aux cultures de 24 et 48 h est fortement positiveau niveau des plages epitheliales (Fig. 3B) et le controle a l'amylase salivairemontre que le polysaccharide present dans les cellules est du glycogene. Apres3 jours de culture, seules quelques unes des cellules epitheliales constituant lesplages sont APS positives et apres 4 jours on n'observe plus aucune trace deglycogene. Dans les cultures plus agees, le reactif de Schiff permet de raettreen evidence des polysaccharides resistants a Faction de 1'amylase salivaire(Fig. 4B).
//. Realisation de cultures secondaires
Dans une premiere etape, nous laissons evoluer des cultures histiotypiquespendant 6 a 45 jours, puis le film cellulaire detache a l'aide d'une spatule estplace en culture organotypique sur un milieu de Wolff et Haffen ou sur un filtresupporte par une grille de type Trowell. Au bout de 3 jours de culture l'observa-tion histologique des explants, apres fixation par le liquide de Gendre ettraitement par la reaction a TAPS, montre que les cellules degenerent rapide-ment. Elles apparaissent dispersees dans une substance fondamentale riche enpolysaccharides. Aucune structure de type hepatique n'est reconstitute dans cecas et les cellules ne possedent aucune des caracteristiques des cellules parenchy-mateuses. En particulier elles ont un noyau de petite taille et sont totalementdepourvues de glycogene.
Dans un deuxieme temps, nous avons associe des cultures cellulaires primairesde 6 a 45 jours au mesenchyme hepatique ou pulmonaire.
Lorsque la culture primaire est associee au mesenchyme hepatique apres 6 a22 jours de culture, l'examen histologique des explants ou des greffons permetd'observer que des hepatocytes sont presents au sein du mesenchyme hepatique.Us sont caracterises par une structure de type epithelial et un noyau contenant
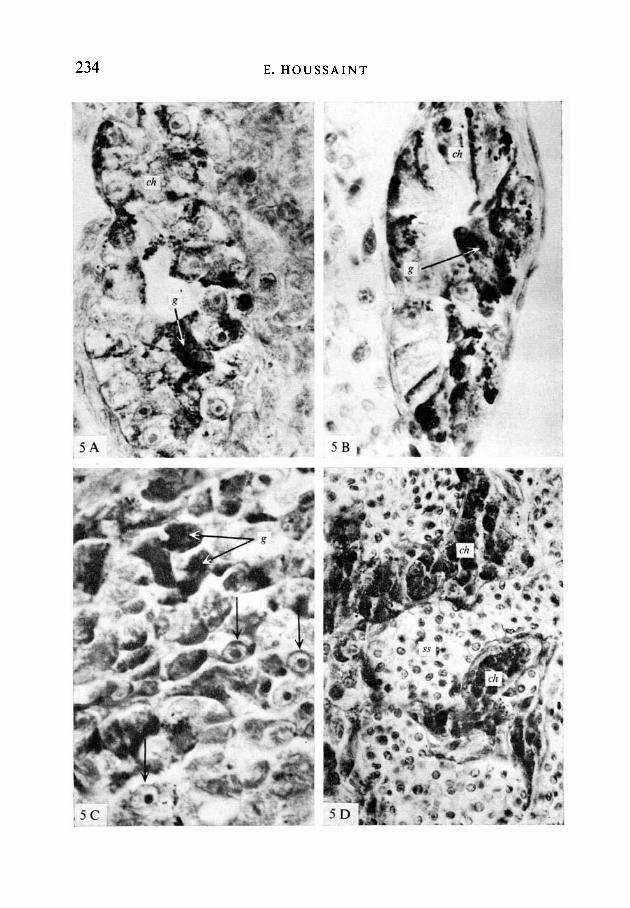
FIGURE 5
Cultures secondaires: association de cultures primaires avec du mesenchymehepatique d'embryon de Poulet. On observe au sein du mesenchyme des celluleshepatiques (ch) riches en glycogene (g).(A). Culture primaire de 6 jours. Duree de la culture secondaire: 3 jours en culturein vitro. APS-hematoxyline. x 900.(B). Culture primaire de 10 jours associee au mesenchyme hepatique pendant3 jours en culture in vitro. APS-hematoxyline. x 1100.(C). Culture primaire de 12 jours associee au mesenchyme hepatique pendant 4 joursen culture in vitro. Les cellules hepatiques possedent le volumineux nucleole carac-teristique des cellules de caille (fleches) et elles sont riches en glycogene (g).APS-hematoxyline. xll50.(D). Culture primaire de 18 jours associee avec du mesenchyme hepatique pendant24 h en culture in vitro puis pendant 6 jours en greffe dans le coelome d'un embryonde poulet de 3 jours. Les cellules hepatiques sont organisees en cordons (ch) separespar des sinusoides sanguins (ss). APS-hematoxyline. x 600.

236 E. HOUSSAINT
Tableau 1. Recuperation de la capacite de synthetiser du glycogene par des culturesprimaires de foies d'embryons de caille de 8 jours associees au mesenchymehepatique ou pulmonaire d'embryons de poulet
Culture secondaire
Culture primaire+
mesenchymehepatique
Culture primaire+
mesenchymepulmonaire
(6 a 10 jours
26 cas positifssur 33
4 cas positifssur 10
Duree de la
11 a 18 jours
12 cas positifssur 38
2 cas positifssur 6
culture primaireA
19 a 22 jours
12 cas positifssur 54
4 cas positifssur 10
22 a 45 jours
21 cas negatifs
5 cas negatifs
un nucleole particulierement volumineux comme c'est le cas dans les cellulesparenchymateuses de foie de caille normal. II est possible en effet de reconnaitreles cellules de caille des cellules de poulet apres une simple coloration du noyaupar l'hematoxyline de Groat (Le Douarin, 1969, 1973). De plus, la reaction aTAPS montre que leur cytoplasme est tres riche en glycogene (Fig. 5 A a D).Lorsque l'association est realisee in vitro, le tissu hepatique forme le plus souventun ou deux ilots compacts au voisinage desquels on observe des cellules quipossedent un noyau de petite taille et qui sont depourvues de glycogene. Lorsquel'association est prolongee en greffe in ovo, le tissu hepatique qui se reconstitue aune structure proche de celle du foie normal. II est alors forme de cordonsd'hepatocytes separes par des sinusoides sanguins (Fig. 5D). Des resultatssimilaires ont ete obtenus dans les associations de mesenchyme pulmonaire avecdes cultures primaires de foie de 6 a 22 jours (Tableau 1).
Lorsque la culture primaire a ete prolongee entre 22 et 45 jours, aucune cellulehepatique n'a ete observee dans les cultures secondaires quel que soit le typed'association realisee (Tableau 1).
CONCLUSION ET DISCUSSION
Lorsque du tissu hepatique differencie d'embryons de caille de 8 jours estcultive in vitro les cellules subissent des transformations morphologiques etphysiologiques profondes. Des plages de cellules d'aspect epithelial subsistentquelques jours puis font place a des cellules fibroblastiques qui deviennentapres 6 jours le seul constituant de la culture. Simultanement, lescellules se modifient du point de vue physiologique et le glycogene decel-able dans les plages epitheliales disparait totalement a partir du 4eme jourde culture.
La perte des fonctions et des caracteristiques structurales des constituants

Reversibilite de la dedifferenciation 237
cellulaires du foie en culture in vitro a ete tres generalement constatee par diversauteurs quelle que soit l'espece utilisee. Sauf dans quelques cas particuliers telsque les cultures de foie humain etablies par Kaighn & Prince (1971) dans les-quelles on a pu mettre en evidence la synthese d'une molecule antigeniquementsemblable a la serum albumine, les cellules d'aspect fibroblastique ne presententpas les proprietes biochimiques caracteristiques des hepatocytes. La 'trans-formation' fibroblastique des cellules de foie en culture pose le probleme d'unededifferenciation morphologique et fonctionnelle possible des elements paren-chymateux. On peut concevoir en effet que dans le cas du foie d'oiseau lesfibroblastes derivent non seulement des cellules mesenchymateuses mais corres-pondent aussi a des hepatocytes apparemment dedifferencies.
Cette conception s'est revelee exacte. En effet, si une culture primaire de foiepurement fibroblastique est associee a du mesenchyme splanchnique d'originehepatique ou pulmonaire, elle fournit des groupes de cellules parenchymateusesriches en glycogene.
L'association interspecifique d'une culture primaire de foie de caille avec dumesenchyme hepatique de poulet permet de se rendre compte que les cellulesparenchymateuses derivent de la culture fibroblastique primaire de caille. Lesproprietes du noyau chez la caille permettent en effet de reconnaitre les cellulesde cette espece de celles du poulet (Le Douarin, 1969, 1973).
Dans les cultures secondaires obtenues par l'association des 'fibroblastes'avec le mesenchyme splanchnique, on constate que les cellules contenant duglycogene sont organisees en un epithelium et sont en contact avec les elementsmesenchymateaux. Ainsi, l'architecture tissulaire caracteristique du foie estreconstitute en meme temps qu'est retablie dans les hepatocytes la capacited'accumuler le glycogene. Le role des contacts specifiques, etablis entre les cellulesd'une ebauche, sur la differenciation et l'activite fonctionnelle des elements qui lacomposent a ete demontre dans d'autres systemes tels que le tissu cartilagineux(Holtzer & Abbott, 1968) et la retine nerveuse (Moscona, 1971). Dans le cas dufoie, il est vraisemblable que l'invasion du tissu mesenchymateux par les cellulesendodermiques s'accompagne de processus de division. Quoiqu'il en soit, cesobservations montrent que la dedifferenciation morphologique apparente descellules parenchymateuses qui s'effectue en culture n'est pas irreversible.L'association des cultures primaires avec le mesenchyme hepatique organisepermet en effet de faire reapparaitre dans ces cellules les caracteres pheno-typiques specifiques du parenchyme hepatique. II est interessant de souligner acet egard que 1'etat de differenciation fonctionnelle des hepatocytes n'est stableque si les cellules sont placees dans des conditions d'environnement favorables.Le mesenchyme splanchnique qui s'est revele indispensable a la differenciationde l'endoderme hepatique (Le Douarin, 1964; Le Douarin & Houssaint, 1967;Le Douarin, 1968; Houssaint, 1972) joue done par la suite un role fondamentaldans le maintien des caracteres de la differenciation. Des experiences anterieuresont montre que ^interposition entre l'endoderme et le mesenchyme hepatique

238 E. HOUSSAINT
d'une seule couche de mesenchyme somitique suffit a empecher la differentiationde Pendoderme en hepatocytes (Fontaine & Le Douarin, 1972). Les resultats quenous rapportons ici montrent que seules les cellules qui enhavissent le mesen-chyme hepatique ou pulmonaire recuperent une morphologie et un fonctionne-ment de type hepatique. 11 semble done que les interactions cellulaires s'exercantentre les deux constituants du foie, aussi bien au cours de l'organogenese que dansle tissu hepatique differencie, ne peuvent s'effectuer que si les cellules etablissententre elles des contacts etroits.
RESUME
Des cultures cellulaires primaires sont etablies a partir de foies d'embryons de caille de 8jours. Au cours des trois premiers jours, la culture est constitute de plages epithelialesseparees par quelques cellules d'aspect fibroblastique. Le cytoplasme des cellules epithelialesest alors riche en glycogene decelable par la reaction histochimique a l'acide periodique Schiff(APS) controlee par le test de Pamylase salivaire. Au cours de revolution de la culture, leselements fibroblastiques deviennent de plus en plus nombreux et apres 6 a 7 jours, les plagesepitheliales ont disparu et la culture est totalement fibroblastique. Aucune trace de glycogenene peut alors y etre decelee par la methode a TAPS. La culture qui garde ensuite cet aspect apu etre prolongee dans certains cas jusqu'a 45 jours.
Des cultures primaires de 6 a 45 jours, ayant acquis un aspect purement fibroblastique, ontete associees a un mesenchyme d'origine splanchnique, hepatique ou pulmonaire. Ces culturessecondaires sont realisees in vitro pendant 3 a 4 jours, soit sur un milieu de Wolff et Hafifensoit sur un filtre millipore pose sur une grille de type Trowell. Dans certains cas, l'associationest maintenue pendant deux jours en culture in vitro puis elle est prolongee en greffe inovo pendant 6 jours. L'observation des cultures secondaires, fixees par le liquide de Gendreet traitees par la reaction a TAPS, montre que des cellules hepatiques ayant une structureepitheliale caracteristique et dont le cytoplasme est riche en glycogene sont presentes dansles explants.
On peut conclure de cette observation que certains fibroblastes de la culture primairesont en fait des cellules parenchymateuses apparemment dedifferenciees. Ces cellules gardentcependant la capacite de reexprimer leur determination initiale quand on les associe a unmesenchyme splanchnique.
Le role des contacts cellulaires dans le maintien de la differentiation est envisage dans ladiscussion.
TRAVAUX CITES
ALEXANDER, R. & GRISHAM, J. (1970). Explant culture of rat liver. I. Method, morphologyand cytogenesis. Lab. Invest. 22, 50-62.
ANGULO, E. (1968). Contribution a l'etude morphologique et histochimique de la culture decellules hepatiques sur un substrat de collagene. Rec. e'd. veter. Ec. Alfort, 144, 1035-1042.
AUERBACH, V. & WALKER, D. (1959). Further studies on the enzymic pattern of a culturedcell line from liver. Biochim. biophys. Acta 31, 268.
BISSELL, D. & TILLES, J. (1971). Morphology and function of cells of human embryonic livermonolayer culture. / . Cell Biol. 50, 222-231.
CHANG, R. (1954). Continuous subcultivation of epithelial-like cells from normal humantissues. Proc. Soc. exp. Biol. Med. 87, 440.
CHESSEBEUF, M., OLSSON, A., BOURNOT, P., DESGRES, J., GUIGUET, M., MAUME, G., MAUME,B., PERISSEL, B. & PADIEU, P. (1974). Long term cell culture of rat liver epithelial cellsretaining some hepatic functions. Biochimie 56, 10, 1365-1379.
COON, H. (1969). Clonal culture of differentiated cells from mammals: rat liver cell culture.Carnegie Institution of Washington, year book, 67, 419-421.

Reversibilite de la dedifferenciation 239DAMERON, F. (1961). Influence de divers mesenchymes sur la differentiation de l'epithelium
pulmonaire de l'embryon de poulet en culture in vitro. J. Embryol. exp. Morph. 9,628-633.
EVANS, V., EARLE, W., WILSSON, E., WALTZ, M. & MADREY, C. (1952). The growth in vitro ofmassive cultures of liver cells. / . natn. Cancer Inst. 12, 1245-1267.
FOGEL, M. (1968). Phenotypic changes in cultured hamster embryonic liver cells as studied byimmunofluorescence. Devi Biol. 17, 85-100.
FONTAINE, J. & LE DOUARIN, N. (1972). Essai d'analyse en culture in vitro de l'effet inhibiteurexerce par le mesenchyme somitique sur le developpement de 1'endoderme hepatique.C. r. Seanc. Soc. Biol. 165, 1972-1976.
GERSCHENSON, L., ANDERSSON, M. & MOLSON, J. (1970). Tyrosine transaminase induction bydexamethasone in a new rat liver cell line. Science N.Y. 170, 859-861.
HILLIS, W. & BANG, F. (1962). The cultivation of human embryonic liver cells. Expl Cell Res.26, 9.
HOLTZER, M. & ABBOTT, J. (1968). Oscillations of the chondrogenic phenotype in vitro. InThe stability of the Differentiated State (ed. H. Ursprung), pp. 1-16. Berlin: Springer.
HOTCHKISS, R. (1948). A microchemical reaction resulting in the staining of polysaccharidestructures in fixed tissue preparations. Archs Biochem. Biophys. 16, 131-141.
HOUSSAINT, E. (1972). Etude des capacites de differentiation de 1'endoderme hepatique dePembryon de Souris. C. r. hebd. Seanc. Acad. Sci., Paris 275, 461-464.
HOUSSAINT, E. & LE DOUARIN, N. (1971). Activites cholinesterasiques dans le foie de poulet.Role de 1'endoderme dans l'apparition d'activites cholinesterasiques dans la composantemesenchymateuse du tissu hepatique. / . Embryol. exp. Morph. 26, 481-495.
HOUSSAINT, E. & LE DOUARIN. N. (1974). Mise en evidence de glycogene et d'une activitede la glycogene synthetase dans l'ebauche hepatique aux stades precoces du developpmentembryonnaire chez le poulet. / . Embryol exp. Morph. 32, 637-650.
KAIGHN, M. & PRINCE, A. (1971). Production of albumin and other serum proteins by clonalcultures of normal human liver. Proc. natn. Acad. Sci. 68, 2396-2400.
KURODA, Y. & NAGATANI, T. (1965). Experimentally induced changes in fructose-l,6-diphos-phatase activity in embryonic chick liver and heart cells in vitro. Devi Biol. 11, 325-351.
LAMBIOTTE, M. (1970). Observations morphologiques et biochimiques sur les celluleshepatiques en culture cellulaire; obtention d'un clone d'hepatocytes de rat. Anneebiologique 9, 349-357.
LAMBIOTTE, M., SUSOR, W. & CAHN, R. (1972). Morphological and biochemical observationson mammalian liver cells in culture. Isolation of a clonal strain from rat liver. Biochimie 54,1179-1187.
LASCHI, R. & RIZZOLI, C. (1968). Etudes ultrastructurales de cultures primaires de foieembryonnaire. Mise au point de techniques et premieres observations. / . Microscopie 7,533-538.
LE DOUARIN, N. (1964). Etude experimentale de l'organogenese du tube digestif et du foiechez l'embryon de Poulet. Bull. biol. Fr. Belg. 98, 544-676.
LE DOUARIN, N. (1968). Synthese du glycogene dans les hepatocytes en voie de differentiation:role des mesenchymes homologues et heterologues. Devi Biol. 17, 101-114.
LE DOUARIN, N. (1969). Particularity du noyau interphasique chez la Caille japonaise(Coturnix coturnix japonica). Utilisation de ces particularity comme 'marquage biologique'dans des recherches sur les interactions tissulaires et les migrations cellulaires au cours de1'ontogenese. Bull. biol. Fr. Belg. 103, 435-452.
LE DOUARIN, N. (1973). A Feulgen-positive nucleolus. Expl Cell Res. 11, 459-468.LE DOUARIN, N., BUSSONNET, C. & CHAUMONT, F. (1968). Etude des capacites de differentia-
tion et du role morphogene de 1'endoderme pharyngien chez l'embryon d'Oiseau. AnnlsEmbryol. Morph. exp. 1, 29-40.
LE DOUARIN, N. & HOUSSAINT, E. (1967). Role du mesoderme dans l'induction de la synthesedu glycogene dans les cellules hepatiques en voie de differentiation. C. r. hebd. Seanc. Acad.Sci., Paris 264, 1872-1874.
MOSCONA, A. (1971). Embryonic and neoplastic cell surface: availability of receptors forCon A and wheat germ agglutinin. Science, N.Y. Ill, 905-906.

240 E. HOUSSAINT
NAMBA, M., HrROX, M. & KISFHMOTO, R. (1968). Properties and identification of rat livercells in long-term cultivation. Acta med. Okayama 22, 209-217.
PERSKE, W., PARKS, R. & WALKER, D. (1957). Metabolic differences between hepatic paren-chymal cells and a cultured cell line from liver. Science, N.Y. 125,1290.
PIECK, A. & KUYPER, C. (1961). Establishment of an epithelial cell strain from calf liver incontinuous culture. Experientia 17, 115.
ROSE, G., KUMEGAWA, M. & CATTONF, M. (1968). The circumfusion system for multipurposeculture chambers. II. The protracted maintenance of differentiation of foetal and newbornmouse liver in vitro. J. Cell Biol. 39, 430-450.
SKEA, B. & NEMETH, A. (1969). Factors influencing premature induction of UDP-glucuronyltransferase activity in cultured chick embryo liver cells. Proc. natn. Acad. Sci. U.S.A. 64,795-802.
VERNE, J. & HEBERT, S. (1968). Comparison of the mode of action of insulin and glucagon onthe hepatocyte glycogen and glucose-6-phosphatase in tissue culture. Folia Histochemica etCytochemica 6, 5-10.
VERNE, J., HEBERT, S., RICHSHOFFER, N. & Roux, F. (1971). Interpretation des processus desynthese et de degradation du glycogene dans les hepatocytes en culture trypsinee. C. r.Seanc. Soc. Biol. 165, 1003-1005.
WATANABE, H. (1966). A fine structural study of liver culture. Expl Cell Res. 42, 685-699.WAYMOUTH, C, CHEN, H. & WOOD, B. (1971). Characteristics of mouse liver parenchymal
cells in chemically defined media. In Vitro 6, 371 A.WOLFF, ET. & HAFFEN, K. (1952). Sur une methode de culture d'organes embryonnaires in
vitro. Texas Rep. Biol. Med. 10, 463-472.
(Regu le 16 juin 1975, revise le 23 octobre 1975)


















