
Retroviruses205.ppt - RCI
36
Genetic recombination during reverse transcription • All recombination occurs between coencapsidated genomes at the time of reverse transcription • The copy choice model postulates a mechanism during (-) strand DNA synthesis
Transcript of Retroviruses205.ppt - RCI

Genetic recombination during reverse transcription
• All recombination occurs between coencapsidated genomes at the time of reverse transcription
• The copy choice model postulates a mechanism during (-) strand DNA synthesis

Strand displacement assimilation model proposes that recombination occurs during (+) strand DNA synthesis

Domain and subunit relationships of RTs from different retroviruses
•Integrase is encoded at the 3’ end of the pol gene
•Mature protein is made by protease mediated processing of the Gag-Pol precursor

Retroviral DNA Integration
6 bp target site is duplicated on either side of proviral DNA
Viral DNA is shortened by 2 bp at each end

• Integrase (IN) is the enzyme that catalyzes integration of the reverse transcribed viral DNA into the host genome
• Viral DNA is shortened by 2 bp from each end and a short (4-6 bp) duplication of host DNA flanks the provirus at either end.
• Proviral ends of all retroviruses comprise the same dinucleotide: 5’-TG----CA-3’
• This dinucleotide is found in an inverted repeat characteristic of the virus
• The inverted repeat, conserved terminal dinucleotide sequence and flanking direct repeats of host DNA are characteristic of features of bacterial insertion sequences
• These similarities suggest common mechanisms of retroviral DNA integration and DNA transposition
Retroviral DNA Integration

Endonucleolytic nicking and removal of 2 nt and formation of a new 3’ recessed end
Joining of 3’ ends to phosphates at the target site
Gapped intermediate Gaps are repaired
The two ends of viral DNA are recognized, nicked and then joined covalently to host DNA in random locations at staggered nicks also introduced by Integrase
Steps in retroviral DNA integration

• Most retrotransposons are distinguished from retroviruses by lack of an extracellular phase
• They have no env gene, thus virus like particles formed in vivo are noninfectious

Retrovirus assembly• Association of Gag molecules with the plasma membrane and with the RNA molecules initiates assembly at the inner surface of the plasma membrane
• A minor fraction of Gag translation products carry the retroviral enzymes, PR, RT and IN at their C-termini
• Assembly continues by incorporation of additional molecules of Gag
• Fusion of the membrane around the budding particle releases the immature noninfectious particle
• Cleavage of Gag and Gag-Pol polyproteins by the viral protease produces infectious particles

Retrovirus assembly

Phylogenetic relationships among retroviruses

The Origin of HIV
“HIV-1 probably originated from SIVcpz in chimpanzees less than 100 years ago. HIV-2, a virus still largely confined in Western Africa, probably originated from SIVsm in
Mangabes”.

http://www.cdc.gov/ncidod/EID/index.htm

Similarities and differences between Lentiviruses• Similarities
– Nucleic acid homology, organization– Transmission (sex, blood, milk)– Macrophage often a target cell– Lentivirus never cleared – Slow pathogenesis / Long time to disease – Vaccines hard– Diseases : no cancers; not endogenous; immune system
dysregulation; immune complexes; neurological• Differences
– Many silent infections vs. high penetrance of AIDS – CD4 lymphocytes in addition to macrophages
• Examples of lentiviruses infecting a different host, under different conditions, and producing a new disease.

Oncogenic retroviruses• Cancer is a genetic disease- oncogenesis consists of the processes that result in growth of cells in which mutations have accumulated
• Viruses are a contributing factor in about 20% of all human cancers
• Growth properties and morphologies of cultured cells could be changed upon infection with certain viruses- cells become transformed
• Cells become immortal in an early step in oncogenesis-they continue to grow and divide even though the body has sufficient numbers of these cells
• They lose contact inhibition and the need to adhere to a surface
• They look different, more rounded

• Transformed cells grow to high densities, they grow on top of untransformed cells forming clumps or foci
• They lose the need to adhere to a surface, can grow in agar
• They are more rounded and look different from normal cells
Avian cells transformed with two strains of the Rous sarcoma virus:

The phases of a eukaryotic cell cycle:
Errors in the signaling pathways that regulate cell cycle progression can lead to cancer
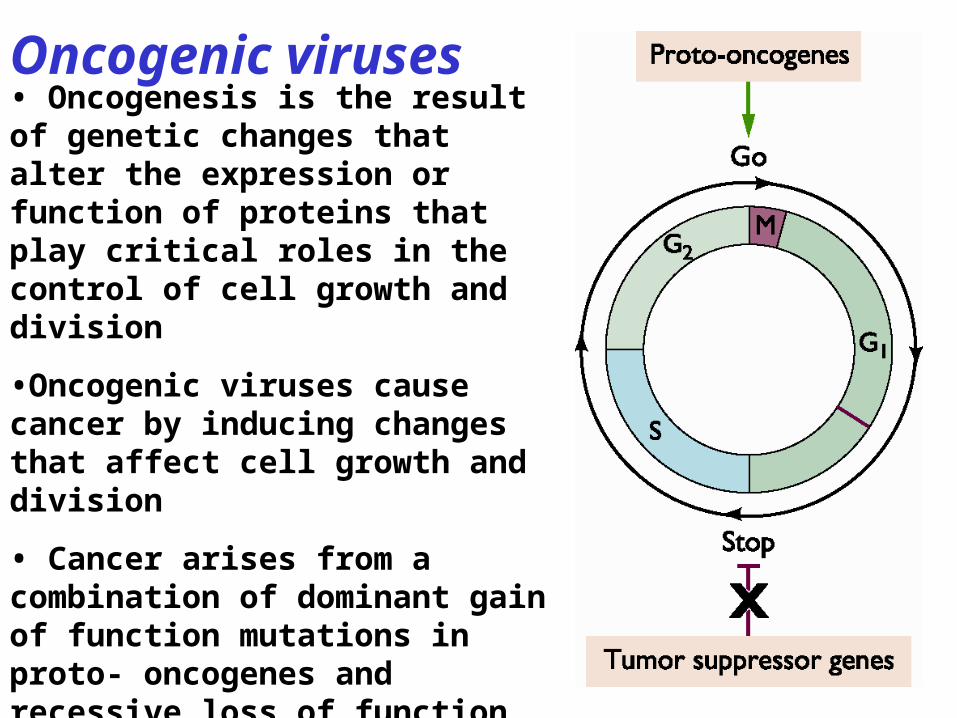
• Oncogenesis is the result of genetic changes that alter the expression or function of proteins that play critical roles in the control of cell growth and division
•Oncogenic viruses cause cancer by inducing changes that affect cell growth and division
• Cancer arises from a combination of dominant gain of function mutations in proto- oncogenes and recessive loss of function mutations in tumor suppressor genes
Oncogenic viruses

• Oncogenic viruses were discovered by Ellerman and Bang in 1908 who showed that avian leukemia can be transmitted by filtered extracts of leukemic cells
• In 1911 Peyton Rous showed that solid tumors can be produced in chicken using cell free extracts

Oncogenic retroviruses are classified into two groups:1) Transducing oncogenic retroviruses:
- highly carcinogenic, cause malignancies in 100% of the infected animals in a matter of days
- cause cancer because their genomes contain transduced cellular genes that become oncogenes
-virally transduced versions of cellular genes are called v-oncogenes, their cellular counterparts are called c-oncogenes or proto-oncogenes
2) Nontransducing oncogenic retroviruses:
-less carcinogenic
-do not encode cell derived oncogenes
-activate transcription of proto-oncogenes by integration of the provirus close to these genes in the host genome


Properties of viral transforming genes• Results with Rous sarcoma virus showed that transformation and viral replication are distinct processes
• With the exception of RSV, these viruses are all replication defective
• Defective transducing viruses can be propagated in mixed infections with replication-competent helper viruses
• In many transducing retroviruses, the viral and cellular protein coding sequences are fused
• In most cases, the captured oncogenes have undergone additional changes that contribute to their transforming potential

Possible mechanisms of oncogene capture by retroviruses


• The discovery that the transforming gene of RSV was a transduced cellular gene led to identification of cellular proto-oncogenes and the pathways in which they function
• Mutations introduced into these genes during or following capture into retroviral genomes lead to constitutive activation of signaling
• Viral transformation can be the result of either constitutive activation of cytoplasmic signal transduction cascades or disruption of nuclear pathways that negatively regulate cell cycle progression
Mechanisms of transformation by oncogenes

Family Flaviviridae

Family Flaviviridae
IRES
Hepatitis C virus

3’ ends of flavivirus RNAs are not polyadenylated

• HCV first identified in 1989
• Blood-borne infection is often subclinical, despite persistent and progressive inflammation and fibrosis of the liver, resulting in liver cirrhosis, hepatic failure and hepatocellular carcinoma
• It is estimated that HCV has infected more than 170 million people globally -- nearly five times more than HIV-infected individuals
• HCV first identified in 1989
• Blood-borne infection is often subclinical, despite persistent and progressive inflammation and fibrosis of the liver, resulting in liver cirrhosis, hepatic failure and hepatocellular carcinoma
• It is estimated that HCV has infected more than 170 million people globally -- nearly five times more than HIV-infected individuals
HCV Infection

HCV Infection
• HCV establishes a chronic infection in ~85% of cases
• HCV infection has become the most common cause of liver cancer and the primary reason for liver transplants among adults in western countries
• Currently no broadly effective anti-HCV therapies are available

Difficulties in treatment
• Persistence of virus
• Genetic diversity during replication in the host
• Development of drug-resistant viral mutants
• Lack of reproducible infectious culture systems and small-animal models for HCV replication and pathogenesis

HCV Genome and Lifecycle
• Enveloped single-stranded RNA Hepacivirus in the Flaviviridae family
• Closely related human viruses include GB virusC (GBV-C), hepatitis H virus, yellow fever virus and dengue virus

Cap-dependent vs.cap-independent translation
initiation
Cap-Dependent Cap-Independent
40S40S40S
4B
40S40S

Where internal ribosome entry sites (IRESs) are found
• Picornavirus– Poliovirus– Hepatitis A– Apthovirus– Rhinovirus– Encephalomyocarditis
virus
• Flavivirus– Yellow Fever Virus– GBV-B– HCV
• Pestivirus– BVDV– CSFV
• Eukaryotic mRNA– Fibroblast growth factor 2– Insulin-like growth factor 2– Platelet derived growth
factor 2– c-myc– BiP– Ornithine decarboxylase
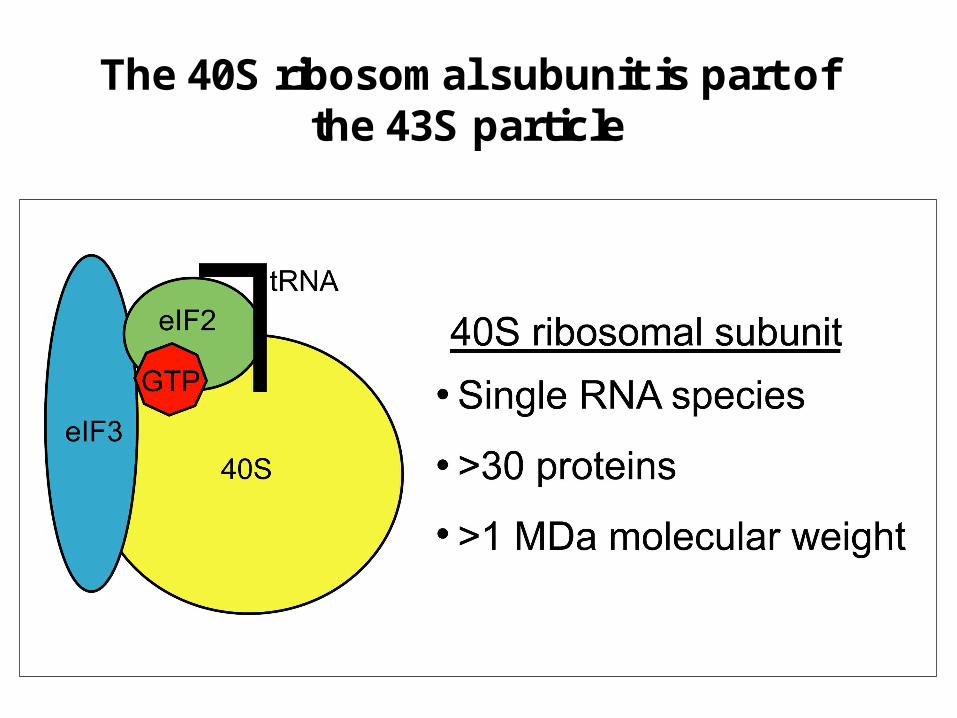
The 40S ribosomal subunit is part of the 43S particle

HCV 5’ untranslated
region
• One of the most conserved regions of genome
• Extensive secondary structure

Complex of the 40S subunit with the hepatitis C virus IRES
• HCV IRES binds directly to the 40S subunit
• Binding of HCV IRES induces a conformational change in the 40S subunit of the ribosome


















