Research Article Sliding Motility, Biofilm Formation, and ...
12
Research Article Sliding Motility, Biofilm Formation, and Glycopeptidolipid Production in Mycobacterium colombiense Strains Milena Maya-Hoyos, 1 John Leguizamón, 1 Leonardo Mariño-Ramírez, 2,3 and Carlos Y. Soto 1 1 Chemistry Department, Faculty of Sciences, Universidad Nacional de Colombia, Carrera 30, No. 45-03, Ciudad Universitaria, Bogot´ a, Colombia 2 Computational Biology Branch, NCBI, NLM, NIH, Bethesda, MD 20894-6075, USA 3 Pan American Bioinformatics Institute, Santa Marta, Magdalena, Colombia Correspondence should be addressed to Carlos Y. Soto; [email protected] Received 15 December 2014; Revised 15 January 2015; Accepted 16 January 2015 Academic Editor: Mehdi Mirsaeidi Copyright © 2015 Milena Maya-Hoyos et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Mycobacterium colombiense is a novel member of the Mycobacterium avium complex, which produces respiratory and disseminated infections in immunosuppressed patients. Currently, the morphological and genetic bases underlying the phenotypic features of M. colombiense strains remain unknown. In the present study, we demonstrated that M. colombiense strains displaying smooth morphology show increased biofilm formation on hydrophobic surfaces and sliding on motility plates. in-layer chromatography experiments showed that M. colombiense strains displaying smooth colonies produce large amounts of glycolipids with a chromatographic behaviour similar to that of the glycopeptidolipids (GPLs) of M. avium. Conversely, we observed a natural rough variant of M. colombiense (57B strain) lacking pigmentation and exhibiting impaired sliding, biofilm formation, and GPL production. Bioinformatics analyses revealed a gene cluster that is likely involved in GPL biosynthesis in M. colombiense CECT 3035. RT-qPCR experiments showed that motile culture conditions activate the transcription of genes possibly involved in key enzymatic activities of GPL biosynthesis. 1. Introduction e Mycobacterium avium complex (MAC) is widely dis- tributed in soil and water [1], and this complex has been frequently identified as an infectious agent in animals and humans [2, 3]. e MAC comprises the species M. avium, M. intracellulare [3], M. colombiense [4], M. chimaera [5], M. marseillense, M. timonense, M. boucherdurhonense [6], M. vulneris [7], M. arosiense [8], 4 subspecies of M. avium [9], and “MAC-other” species [10]. e opportunistic infections generated from mycobacteria in HIV-infected and immuno- suppressed patients have recently highlighted the clinical relevance of MAC [11, 12]. M. colombiense was originally isolated from HIV-positive patients in Bogot´ a, Colombia [4]. is species is responsible for lymphadenopathy in immunocompetent children in Spain and France [13, 14] and has recently been associated with pulmonary infections that complicate cases of cystic fibrosis [15] and disseminated coinfections with cytomegalovirus [16]. Urease-positive tests and the mycolic acid pattern by thin- layer chromatography (TLC) demonstrate the phenotypic characteristics that distinguish M. colombiense from other MAC species [4]. We recently used TLC to show that the mycolate profile of M. colombiense is characterised by the presence of mycolates I (-mycolate), IV (ketomycolate), and VI (carboxymycolate) and two additional spots with chromatographic behaviours similar to those of mycolates III (hydroxyl-mycolate) and VI [17]. A unique 16S rDNA and the internal transcribed spacer (ITS), MAC-X, facilitated the classification of M. colombiense as a novel sequevar [4]. We also identified a 450-bp exclusive genomic region suitable for M. colombiense identification through PCR [17]. e physiological and molecular bases for MAC virulence have not been entirely established. However, the virulence of MAC strains has been associated with variations in colony morphology [18, 19], genetic markers, and glycolipid compo- sition [20]. MAC strains display three different morphologies: smooth transparent, smooth opaque, and rough [18, 19], with Hindawi Publishing Corporation BioMed Research International Volume 2015, Article ID 419549, 11 pages http://dx.doi.org/10.1155/2015/419549
Transcript of Research Article Sliding Motility, Biofilm Formation, and ...

Research ArticleSliding Motility Biofilm Formation and GlycopeptidolipidProduction in Mycobacterium colombiense Strains
Milena Maya-Hoyos1 John Leguizamoacuten1 Leonardo Marintildeo-Ramiacuterez23 and Carlos Y Soto1
1Chemistry Department Faculty of Sciences Universidad Nacional de Colombia Carrera 30 No 45-03Ciudad Universitaria Bogota Colombia2Computational Biology Branch NCBI NLM NIH Bethesda MD 20894-6075 USA3Pan American Bioinformatics Institute Santa Marta Magdalena Colombia
Correspondence should be addressed to Carlos Y Soto cysotoounaleduco
Received 15 December 2014 Revised 15 January 2015 Accepted 16 January 2015
Academic Editor Mehdi Mirsaeidi
Copyright copy 2015 Milena Maya-Hoyos et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Mycobacterium colombiense is a novel member of theMycobacterium avium complex which produces respiratory and disseminatedinfections in immunosuppressed patients Currently the morphological and genetic bases underlying the phenotypic features ofM colombiense strains remain unknown In the present study we demonstrated that M colombiense strains displaying smoothmorphology show increased biofilm formation on hydrophobic surfaces and sliding onmotility platesThin-layer chromatographyexperiments showed that M colombiense strains displaying smooth colonies produce large amounts of glycolipids with achromatographic behaviour similar to that of the glycopeptidolipids (GPLs) of M avium Conversely we observed a naturalrough variant of M colombiense (57B strain) lacking pigmentation and exhibiting impaired sliding biofilm formation and GPLproduction Bioinformatics analyses revealed a gene cluster that is likely involved in GPL biosynthesis in M colombiense CECT3035 RT-qPCR experiments showed that motile culture conditions activate the transcription of genes possibly involved in keyenzymatic activities of GPL biosynthesis
1 Introduction
The Mycobacterium avium complex (MAC) is widely dis-tributed in soil and water [1] and this complex has beenfrequently identified as an infectious agent in animals andhumans [2 3] The MAC comprises the species M aviumM intracellulare [3] M colombiense [4] M chimaera [5]MmarseillenseM timonenseM boucherdurhonense [6]Mvulneris [7] M arosiense [8] 4 subspecies of M avium [9]and ldquoMAC-otherrdquo species [10] The opportunistic infectionsgenerated from mycobacteria in HIV-infected and immuno-suppressed patients have recently highlighted the clinicalrelevance of MAC [11 12] M colombiense was originallyisolated from HIV-positive patients in Bogota Colombia[4] This species is responsible for lymphadenopathy inimmunocompetent children in Spain and France [13 14]and has recently been associated with pulmonary infectionsthat complicate cases of cystic fibrosis [15] and disseminatedcoinfections with cytomegalovirus [16]
Urease-positive tests and themycolic acid pattern by thin-layer chromatography (TLC) demonstrate the phenotypiccharacteristics that distinguish M colombiense from otherMAC species [4] We recently used TLC to show that themycolate profile of M colombiense is characterised by thepresence of mycolates I (120572-mycolate) IV (ketomycolate)and VI (carboxymycolate) and two additional spots withchromatographic behaviours similar to those of mycolates III(hydroxyl-mycolate) and VI [17] A unique 16S rDNA andthe internal transcribed spacer (ITS) MAC-X facilitated theclassification of M colombiense as a novel sequevar [4] Wealso identified a 450-bp exclusive genomic region suitable forM colombiense identification through PCR [17]
Thephysiological andmolecular bases forMACvirulencehave not been entirely established However the virulence ofMAC strains has been associated with variations in colonymorphology [18 19] genetic markers and glycolipid compo-sition [20]MAC strains display three differentmorphologiessmooth transparent smooth opaque and rough [18 19] with
Hindawi Publishing CorporationBioMed Research InternationalVolume 2015 Article ID 419549 11 pageshttpdxdoiorg1011552015419549
2 BioMed Research International
the smooth variants being the most virulent morphology[18 19] In addition MAC strains spread on solid hydrophilicsurfaces through sliding motility mechanisms that are inde-pendent of extracellular structures [21 22] Bacterial motilityplays a significant role in the colonisation of environmentalsurfaces and cells [21] which in turn has been correlatedin vitro with the capacity to form biofilms on hydrophobicsurfaces [23] In M avium strains motility and biofilmformation have been correlated with colony morphology andthe presence of glycopeptidolipids (GPLs) in the cell envelope[24 25] Specifically smooth transparent M avium variantsshow higher motility on hydrophilic surfaces and increasedGPL production conversely rough variants show diminishedmotility and impaired GPL production [22]
GPLs are glycolipids attached to the outermost portionof some nontuberculous mycobacteria including M aviumM smegmatisM abscessus andM fortuitum [25] This typeof glycolipid comprises a mixture of 3-hydroxy or 3-methoxyC26ndashC34
fatty acids amidated to a tripeptide-amino-alcohol(D-phenylalanine-D-allo-threonine-D-alanine-L-alaninol)[24 25] The lipopeptide core is subsequently glycosylatedthrough 6-deoxytalose (linked to the allo-threonine) and120572-L-rhamnose (linked to L-alaninol) and the resultingoligosaccharide residues are methylated to form the non-serovar-specificGPLs (nsGPLs) present in all GPL-producingmycobacteria However MAC strains also synthesise polarGPLs in which some oligosaccharides residues are attachedto the 6-deoxytalose producing serovar-specific GPLs(ssGPLs) [24 25] In M avium the consecutive actionsof a peptide synthetase (PstAPstB) a fatty acid synthasesystem (FAS) and polyketide synthetase (Pks) polyketidesynthetase associated protein (PapA) glycosyl and methyltransferases (GtfAGtfBRtfA and MtfAMtfBMtfCMtfD)and glycolipid transporters (TmtP and Gap) are responsiblefor the biosynthesis and subsequent translocation of GPLs tothe cell wall surface [24ndash26]
Currently the existence of GPLs inM colombiense strainsis completely unknown In the present study we showed thatM colombiense contains glycolipids with chromatographicbehaviours similar to GPLs In addition this novel speciesforms biofilms on the hydrophobic surfaces of polystyreneand motility is increased in strains displaying smoothcolony morphology Moreover we examined the genes likelyinvolved in GPL biosynthesis in the CECT 3035 strain
2 Material and Methods
21 Bacterial Strains Culture Conditions and Genomic DNAIsolation The M colombiense clinical isolates 19B 57Bthe M colombiense genome sequence strain CECT 3035M avium 104 [27] and M smegmatis mc2155 [28] wereused in this study (Table 1) Planktonic mycobacteria werecultured at 37∘C with agitation (76 rpm) in Middlebrook7H9 media supplemented with ADC (05 (wv) bovineserum albumin 02 (wv) dextrose 0085 (wv) NaCl and00003 (wv) beef catalase) and 005 (vv) glycerol untilan OD
600of 05 was obtained (planktonic conditions) For
the cell motility assay mycobacteria were cultured inmotilitymedium containing 7H9 supplemented with ADC and 035
agarose Pseudomonas aeruginosa ATCC27853 [29] culturedinmotilitymediumwas used as a positive control in the drop-collapsing test
For DNA extraction the mycobacteria were grown in7H9-ADC broth to an OD
600of 05 centrifuged and resus-
pended in TE buffer (10mM Tris-HCl and 1mM EDTA pH8) Subsequently the bacilli were inactivated through incu-bation at 80∘C for 20 minutes Genomic DNA was extractedusing lysozyme SDSproteinase K and CTABNaCl for celldisruption and chloroform isoamyl alcohol (24 1 vv) forgetting rid of proteins [30 31] The DNA pellets were treatedwithDNAse-free RNAse resuspended in 01X TE followed byquantification using a NanoDrop 2000c Spectrophotometer(ThermoScientificMAUSA)TheDNAqualitywas assessedusing agarose gel electrophoresis and spectrophotometry(OD260
OD280
)
22 Cell Motility Assay and Congo Red Staining For the cellmotility assay plates containing 7H9 media supplementedwithADCand035agarosewere incubated at room temper-ature overnight prior to cell inoculation [21] Subsequently3 120583L of mycobacterial culture at an OD
600of 06 (sim27 times
105 CFU) was carefully spotted onto the centre of the plates tominimise the spread of cells from the inoculation point Theplates were dried under sterile conditions for 30min sealedwith Parafilm and incubated at 37∘C for 4-5 weeks underhumidified conditions The motility rate was recorded asthe colony growth halo The motility assays were performedin triplicate from three independent experiments using Mavium 104 as a control Differences among the experimentaldata were compared using Studentrsquos t-test and 119875 lt 005 wasconsidered significantly different
For the characterisation of colony morphology platescontaining 7H10-OADC (ADC + 005 oleic acid) mediasupplemented with 100 120583gmL Congo Red (Sigma-AldrichMO USA) [32] were inoculated as in the cell motility assayand the resulting cultures were incubated for 4-5 weeksat 37∘C under humidified conditions The morphology wasdefined as rough or smooth opaque or transparent andyellow or beige
23 Biofilm FormationAssay Biofilm formationwas assessedas previously described [22] with some modificationsBriefly the wells of polystyrene microtiter plates (TPPSwitzerland) were filled with a mix of 75120583L of planktonicmycobacterial culture (OD
600= 06) and 125 120583L of 7H9-
ADC broth The plates were incubated at 37∘C for 6 weeksSubsequently the plates were washed three times with 1XPBS (014M NaCl 5mM KCl 10mM Na
2HPO4 and 2mM
KH2PO4) 200120583L of 01 crystal violet solution was added
and the cells were incubated at 37∘C for an additional30min The residual crystal violet solution was removedand the wells were washed three times with 1X PBS bufferA total of 200120583L of absolute ethanol was added to eachwell and the plates were incubated for 1 hour at roomtemperature Colouration was quantified at 570 nm using aiMark Microplate Absorbance Reader 168-1135 (BioRad PAUSA) The biofilm assays were performed in triplicate fromthree independent experiments using M avium 104 as a
BioMed Research International 3
Table 1 Bacterial strains and primers used in this study
Strain and primers Description SourceM colombiense (CECT 3035) Sequence genome strain Clinical isolateM colombiense 19B and 57B Clinical isolates Clinical isolateM smegmatismc2155 Reference strain [28]M avium 104 Reference strain [27]P aeruginosa ATCC27853 Reference strain [29]Primers Sequence (51015840-31015840)pstA dir ACAGGGCACGAGGAATTCTA This studypstA rev TAGTCCTCGGAGGCTTCGTA This studygftA dir ATGTGTGCTGGCCAGTTATG This studygftA rev GGAAGAACGACGTCCAGAAG This studyrtfA dir GACTTTTGGAGCGACGAGTT This studyrtfA rev GCCAAATCCTGGTAAAGCTG This studymtfB dir GGACACCGAGCACTACGAG This studymtfB rev TCATACAGATCGCCATCCAG This studymtfC dir ACAAGGCGGATAAAGGGATT This studymtfC rev CTCATACAGATCGCCATCCA This studymtfD dir TACCTGCTCGACACCTTCG This studymtfD rev TCGACCTGCTCGAGTGTCT This studytmtpC dir TTCATTCGGGATACCAGGAG This studytmtpC rev TTGATCCTGACCCGAAGTTT This studytmtpA dir CTCTCGGCTTTGACGACAC This studytmtpA rev ATGGCCGACATCAGCTACTT This studytmtpB dir GAGTGCCCTTGAGTGATTCC This studytmtpB rev CCTCCAAGAATGACGATTCC This study16 sRNA dir GAGATAGGCGTTCCCTTGTG This study16 sRNA rev CTGGACATAAGGGGCATGAT This study
positive control Differences among experimental data werecompared using Studentrsquos t-test and119875 lt 005was consideredsignificantly different
24 Drop-Collapsing Test Mycobacteria grown on motilitymedium were scraped and resuspended in 5mL 1X PBSgently vortexed and centrifuged at 8000 rpm for 1 h Sub-sequently 20120583L of supernatant was carefully spotted ontothe 154mm wells of polystyrene plates (TPP Switzerland)previously coated with 100120583L of mineral oil and incubatedfor 24 h at room temperature [33 34] The aqueous dropswere visually examined incubated for 1min at room tem-perature and subsequently photographed The tests wereperformed in triplicate from three independent experimentsusing deionised water and 1 SDSM smegmatismc2155 andP aeruginosa as controls
25 Lipid Extraction and TLC Analysis Crudemycobacteriallipid extracts were obtained as previously described [35] withsome modifications Briefly mycobacteria cultured underplanktonic and motility conditions were extracted usingchloroform-methanol (1 2 vv) through vigorous stirringfor 48 h at room temperature followed by chloroform-methanol (2 1 vv) extraction under the same experimentalconditions Pooled and dried organic extracts were parti-tioned using a chloroform-methanol-water mixture (8 4 2vvv) The organic phase was separated and evaporated todryness and the free glycolipids extracts were weighed ForGPL detection 1mg of the lipid extracts was dissolved in
chloroform-methanol (2 1 vv) at 20mgmL and analysedon 20 times 20 Silica Gel 60 TLC plates (Merck NJ USA)using chloroform-methanol-water (65 25 4 vvv) [35]Thecarbohydrate-containing compounds were visualised afterspraying the plates with 1 anthrone (Sigma-Aldrich MOUSA) in sulphuric acid followed by heating at 120∘C CrudeM avium 104 lipid extract was used as a control andthe glycolipid profile was obtained from three independentexperiments performed in duplicate
26 Bioinformatics Analysis The genes implicated in the bio-synthesis of GPLs in theM avium 104 strain (httpwwwnc-binlmnihgovnuccoreCP0004791) were used as model forthe bioinformatics analysisThe gene sequences for pstApstB(peptide synthetases) pks10 (polyketide synthetase) gtfAgtfB and rtfA (glycosyl transferases) mtfA mtfB mtfC andmtfD (methyl transferases) tmtpC tmtpA and tmtpB (gly-colipid transporters) and papA2 (acyl transferase) (Table 1)were identified in M colombiense CECT 3035 using the Ge-nomic BLAST tool (httpwwwncbinlmnihgovsutilsge-nom tablecgi) The orthologous sequences identified in theM colombiense CECT 3035 genome [36] were subsequentlyused as targets for primer design in the RT-qPCR experi-ments
27 RNA Isolation and RT-qPCR Analysis RNA ofM colom-biense 19B CECT 3035 and 57B cultured on both motilitymedium and planktonic conditions was isolated for RT-qPCR experimentsMycobacteria grown onmotilitymedium
4 BioMed Research International
(a) (b)
(c) (d)
Figure 1 Colonymorphology ofMycobacterium colombiense strainsMycobacterial strains were grown on 7H10-OADC plates supplementedwith Congo Red and incubated at 37∘C for 4-5 weeks (a)M colombiense 19B (b) CECT 3035 (c) 57B and (d)M avium 104
were scraped and resuspended in diethylpyrocarbonate-(DEPC-) treated water Motile and planktonic mycobacteriawere centrifuged washed with DEPC-treated water andresuspended in TRIzol (Invitrogen USA) The cells werelysed using a Mini-Bead Beater 16 (BioSpec OK USA) andglass beads (01mm) Total RNA was isolated as previouslydescribed [37] and quantified using aNanoDrop 2000c Spec-trophotometer (Thermo Scientific MA USA) After DNaseI treatment the RNA quality was evaluated through agarosegel electrophoresis and spectrophotometry (OD
260OD280
)cDNA libraries were constructed from 1 120583g of RNA using theRevertAid First cDNA Synthesis Kit (Fermentas Lithuania)For reverse transcription the samples were incubated at 37∘Cfor 30min 42∘C for 5 h 72∘C for 1min and 4∘C for 10min
The primers used for the RT-qPCR analyses are listed inTable 1The RT-qPCR reactions were set up in triplicate usingthe Express SYBR GreenER Universal qPCR SuperMix Kit(Invitrogen NY USA) and a CFX-96 thermocycler (BioradPA USA) under conditions including a denaturation stepfor 5min at 95∘C followed by 39 cycles of 95∘C for 10 sec
58∘C for 10 sec and 72∘C for 15 sec Primers or cDNA wereomitted in the negative controls The gene transcriptionwas normalised to the mean value of 16S rRNA (rrs) geneexpressionThe transcription profile was obtained from threeindependent experiments performed in duplicate For eachexperiment the differences among experimental data werecompared using Studentrsquos t-test and119875 lt 005was consideredsignificantly different
3 Results
31M colombiense IsMotile and Forms Biofilms onHydropho-bic Polystyrene Surfaces The M colombiense strains usedin this study including the genome sequence strain CECT3035 presented smooth opaque and brilliant colonies on7H10-OADC plates supplemented with Congo Red exceptstrain 57B which displayed a rough and wrinkled phenotype(Figure 1) Similar results were obtained using medium lack-ing oleic acid (7H9-ADC-035 agarose data not shown)Other M colombiense strains not included in the present
BioMed Research International 5
7
17
27
37
47
57
0 10 20 30 40 50 60 70
Dia
met
er (m
m)
Day
M colombiense 19BM avium
M colombiense CECT 3035
M colombiense 57B
Figure 2 Motility rates for Mycobacterium colombiense strainsMycobacterial strains were cultivated on 7H9-ADC-035 agaroseand incubated at 37∘C for 4-5 weeks The colony growth haloswere measured daily to determine the motility rate Each reportedvalue represents the mean of the colony growth halos from threeindependent experiments The presented data have statisticallysignificant differences compared with the values obtained from Mavium 104 (119875 lt 005)
study (6B 7B 9B and 16B) also displayed a smooth colonymorphology (data not shown) Both smooth and rough bac-terial phenotypes were stable after repetitive culture on solidmedia for the duration of the study (3 years) The smoothM colombiense strains developed yellow pigmentation withage specifically at the stationary phase in all culture mediahowever the rough variant (57B) remained beige in colourunder the same experimental conditions (Figure 1)
Because alterations in the phenotypic characteristicsassociated with colony morphology have been observedin other MAC species motility and biofilm formationwere compared between smooth and rough M colombiensestrains We observed that the M colombiense 19B andCECT3035 strains (smooth colony morphology) showedincreased spreading (4094 plusmn 005 120583mday and 2972 plusmn005 120583mday resp) on hydrophilic agarose comparedwith thenatural rough variant 57B which showed impaired spreading(Figure 2)
Crystal violet staining was used to quantify the degree ofM colombiense biofilm formation onpolystyreneThehighestOD570
values for all M colombiense strains were obtainedusing an inoculum of sim27 times 105 CFUmL In general thestrains displaying smooth morphology showed increasedbiofilm formation on hydrophobic surfaces while therough variant 57B showed reduced adhesion to polystyrene(Figure 3)
As biosurfactants could potentially influence cell motilitythe ldquodrop-collapsingrdquo method was used to detect biosurfac-tant secretion in M colombiense strains [22] As shown inFigure 4 drops of 1 SDS and P aeruginosa aqueous extract(positive controls) spread over the oily surface after the sam-ples were incubated indicating the presence of biosurfactantsubstances In contrast drops of distilled waterM smegmatis(negative control) and M colombiense aqueous extracts didnot collapse on the oily surface but rather appeared as firm
0
005
01
015
02
025
OD570
Mavium
104
Mcolom
biense
CECT
3035
Mcolom
biense19
B
Mcolom
biense57
B
Figure 3 Capacity of Mycobacterium colombiense strains to formbiofilms The OD
570nm represents the proportion of mycobacteria
added to the microtiter wells that reacted with the crystal violetsolutionThebars represent the SD calculated from two independentexperiments each performed in triplicate
drops suggesting the absence of biosurfactants in the culturesupernatants
32 M colombiense Displaying a Smooth Colony Morphol-ogy Contains Glycolipids with Thin-Layer ChromatographicBehaviour Similar to That of the GPLs of M avium TLCanalysis of noncovalently attached lipids extracted frommycobacteria cultured under planktonic and motile condi-tions showed that M colombiense strains contain glycolipidswith chromatographic behaviour similar to that of theGPLsofM avium [23] Regarding planktonic cells (Figure 5(a)) thestrains with smooth colony morphology (19B and CECT3035) showed multiple lipid spots migrating in the region ofcontrol GPLs (M avium 104) however the spots observed inthe CECT 3035 lipid extract were more intense than those inthe 19B strain The natural rough variant (57B) showed onlyone spot that migrated in the same GPLs region
Under motile conditions (Figure 5(b)) the smoothcolony morphology strains 19B and CECT 3035 showed aGPL profile similar to that obtained for cells cultured underplanktonic conditions nevertheless the counterpart spotswere more intense than those observed in planktonic cellsparticularly for the 19B strain Nevertheless the unique spotobserved for motile 57B cells (rough variant) were moreintense than that observed for planktonic 57B cells Inter-estingly all motile M colombiense cells exhibited reducedproduction of polar glycolipids such as phosphatidyl-inositolmannosides (PIMs) and phosphatidylglycerol (PG) com-pared with planktonic cells (Figure 5(b))
33 In Silico Identification of GPL Biosynthesis Genes inM colombiense CECT 3035 The genes involved in GPLbiosynthesis in M colombiense are currently unknown
6 BioMed Research International
(a) (b) (c) (d)
(e) (f) (g) (h)
Figure 4 Drop-collapsing test (a) Water (no surfactant) (b) 1 SDS (surfactant) (c) P aeruginosa (d) M smegmatis mc2155 (e) Mcolombiense 19B (f) M colombiense CECT 3035 (g) M colombiense 57B and (h) M avium 104 Drop spreading over the oily surface aftersamples incubation indicates the presence of biosurfactant substances in the aqueous sampleThis assay was conducted in three independentexperiments each performed in duplicate
Based on the GPL biosynthetic pathway reported for Mavium 104 [25] 30 open reading frames (ORF) encodingpeptide synthetase fatty acid synthases (FAS) polyketidesynthases (PKs) acyltransferase (PapA) glycosyltransferasescarbohydrate synthetases methyl and acyl transferases gly-colipid transporters potential biosynthesis regulators andproteins with unknown functions [25 26] were searchedwithin the recently reported nucleotide sequence of Mcolombiense CECT 3035 [36] This in silico analysis (seeTable S1 of the Supplementary Material available online athttpdxdoiorg1011552015419549) showed that all enzy-matic activities searched were encoded by different ORFsin M colombiense CECT 3035 with an identity rangingbetween 77 and 96 The ORFs in the genome of the Mcolombiense counterpart were located in the contigs 0000100002A 00003A and 00007 of the genome sequence of theCECT 3035 strain (Figure 6) Among the 30 genes of the GPLsynthetic pathway inM avium 104 7 genes (mtfA gtfB gtfDdghA fadD23 pe and gap-like) did not have orthologoussequences inM colombiense CECT 3035 (Table S1)
34 Differential Transcription of the GPL Biosynthesis Genesin M colombiense Displaying Smooth and Rough ColonyMorphologies RNAwas isolated fromM colombienseCECT3035 19B and 57B strains cultured under planktonic andmotile conditions to quantify the transcription levels ofthe pstA gtfA rtfA mtfB mtfC mtfD tmtpC tmtpA andtmtpB genes predicted to encode key enzymes of the GPLbiosynthetic pathway (Table S1) For relative quantificationM colombiense CECT 3035 was grown to the exponentialphase under planktonic conditions and used as the referencepattern for two reasons (1) CECT3035 is theM colombiensegenome sequence strain [36] and consistent with the TLCanalysis (2) planktonic cells displayed the lowest GPL pro-duction
With regard to planktonic cells the rate ofM colombiense19B gene transcription was consistently higher than thatfor the M colombiense 57B genes Conversely the roughvariant (57B) displayed a dramatic decrease in the rate oftranscription for the 9 selected genes particularly thoseencoding peptide synthetase pstA glycosyltransferase gtfA
BioMed Research International 7
1 2 3 41 2 3 4
GPLs
PG
PIMs
(a) (b)
Figure 5 TLC profile of the GPLs in Mycobacterium colombiense strains Crude lipid extracts of (1) M avium 104 (2) M colombiense19B (3) M colombiense CECT 3035 and (4) M colombiense 57B Cells cultivated (a) under planktonic and (b) motile conditions weredeveloped using chloroform-methanol-water (65 25 4 vvv) GPLs glycopeptidolipids PIMs phosphatidyl-inositol mannosides and PGphosphatidylglycerol
the methyltransferases mtfB and mtfC and the lipid trans-porters tmtpA tmtpB and tmtpC (Figure 7(a)) InterestinglyrtfA (rhamnosyltransferase) in the 19B strain (smooth colonymorphology) was the only gene among the strains culturedunder planktonic conditions that showed an increased tran-scription rate (221-fold higher) compared with the controlcells (planktonicM colombiense CECT 3035 at the exponen-tial phase of growth)
Under motile growth conditions M colombiense 19Bshowed the highest transcription rate for all selected genescompared with the CECT 3035 and 57B strains (Figure 7(b))In contrast the transcription rates for all genes in the57B strain (rough variant) except tmtpA and tmtpC (lipidtransporters) were reduced In motile CECT 3035 cellswhereas rtfA mtfB (methyl transfers) and tmtpA showedincreased transcription the pstA gftA mtfC mtfD tmtpBand tmtpC genes exhibited reduced transcription comparedwith control cells
4 Discussion
In the present study TLC experiments were able to show thatM colombiense contains glycolipids with chromatographicbehaviour similar to the GPLs of M avium 104 which havebeen previously identified and completely characterised [23]in addition the colony morphology was associated withthe GPL profile of M colombiense strains Thus the TLCanalysis revealed that (1) strains displaying smooth colonymorphology (19B and CECT3035) produce a higher amountof variable GPLs compared with the rough natural variant(57B) strain and (2) the GPL production was augmentedin M colombiense cells cultivated under the motile growthconditions As expected the TLC analyses did not provide
detailed information about GPL structure however struc-tural determinationwas not the objective of the present study
Themedia composition influences the glycolipid contentof the mycobacterial cell wall The MAC strains growing onsolid medium exhibit a significant rate of transition fromsmooth transparent to smooth opaque and from smooth torough morphologies [18 19 38] We compared planktonicand motile M colombiense cells grown in the same culturemedium (7H9-ADC or 7H9-ADC-agarose) to avoid differ-ences in lipid content produced through alterations in themedia composition The results showed that the low GPLcontent observed in the rough 57B variant is less frequentamong M colombiense strains therefore M colombienseshows a preference for augmented GPL production andsmooth colony morphology Interestingly planktonic Mcolombiense cells displayed increased PIM and PG pro-duction and diminished GPL content compared with cellscultivated under motile conditions suggesting a potentialcompensatory mechanism in cell wall lipid synthesis thatcompensates for the low GPL content
The abundance of GPLs in the outermost portion ofthe cell envelope could be associated with the motility ofM colombiense strains on hydrophilic surfaces Recht andothers [22] proposed that GPLs are linked through thehydrophilic head to the cellular capsule and the hydrophobicfatty acid chain is exposed to the bacterial surface therebyreducing interactions with hydrophilic agarose surfaces andfacilitating the spread of cells on solid medium [21 22]Thus the augmented GPL production of motile 19B andCECT 3035 cells compared with that of the rough variant57B is consistent with the sliding motility on the hydrophilicagarose surface observed for these strains It has also beensuggested that bacterial sliding motility might be favoured
8 BioMed Research International
33kb 2kb9kb51kb
Contig 00001
600kb
87kb 245kb
488kb 36kb320kb
Contig 00002A
Contig 00003A
584kb 452kb
mmpL10
326kb
Contig 00007
fadE
15
atf
rmlB
ecf
rv1174
rmlA
mmpL
4a
mmpL
4b
mmpS4
mbtH
gap rv0926
sap
pks papA3
gtfA
mtfB
mtfC
mtfD
rtfA
pstA
pstB
(a)
ecf
fadE
5
rv1174
atf
rmlB
rmlA
mmpL
4a
mmpL
4b
mmpS
4
mbtH
gap
237kb
rv0926
7kb90kb
530kb
300kb
738kb
mmpL10
sap
Saccharides biosynthesis transfer and modification
Fatty acyl chain biosynthesis modification and transfer
Members of the mmpSL family
Peptide synthesis Unknown
Possible regulatory genes
GPL export to the cell surface
pks pe fadD23 gap-likepapA3
mtfB
gtfA
mtfC
mtfD
dhgA
rtfA
pstA
pstB
mtfA
gtfB
gtfD
(b)
Figure 6Model showing the gene cluster proposed for the GPL biosynthesis ofMycobacterium colombienseCECT 3035 (a)The location andidentity of theM colombiense genes were established using the previously described biosynthetic pathway for GPL biosynthesis annotated intheM avium 104 genomic sequence (b)
0001
001
01
1
10
Rela
tive g
ene t
rans
crip
tion
M colombiense 57BM colombiense 19B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(a)
01
1
10
100
Rela
tive g
ene t
rans
crip
tion
M colombiense CECT 3035
M colombiense 19B
M colombiense 57B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(b)
Figure 7 In vitro transcription of Mycobacterium colombiense genes involved in GPL biosynthesis Relative quantification expressed as theratio of gene transcription in (a) M colombiense cells grown under planktonic conditionstranscription ofM colombiense CECT 3035 cellsgrown under planktonic conditions and (b)M colombiense cells grown on the motility mediumtranscription ofM colombiense CECT 3035grown under planktonic conditions The presented data have statistically significant differences compared with the values obtained fromMcolombiense CECT 3035 cells (119875 lt 005)
BioMed Research International 9
through the secretion of surfactant substances from cells[21 22] The results of the drop-collapsing test showed thatbiosurfactants were likely not secreted from M colombiensecells suggesting that the motility of M colombiense couldbe favoured through hydrophobic molecules such as GPLsbound to the outermost portion of cells however we cannotcompletely rule out the possibility that some of the surfactantsubstances secreted from M colombiense remained adheredto the mycobacterial cell surface thereby influencing cellmotility
GPLs on the outermost portions of the M colombiensecell envelope generate a more hydrophobic cell surfacethat facilitates initial interactions with hydrophobic surfacesand increase biofilm formation on polystyrene wells Inaddition the increased GPL production in 19B and CECT3035 cells is consistent with the augmented biofilm forma-tion observed for M colombiense strains displaying smoothcolony morphology Moreover increased biofilm formationon hydrophobic surfaces and enhanced sliding over motilityplates for 19B cells compared with the CECT 3035 strainwhich also displays smooth colony morphology likely sug-gest possible differences in GPL structure betweenM colom-biense strains Differences inGPL production between strainscould influence the M colombiense ability to form biofilmson polyvinylchloride (PVC) pipes which would facilitatethe dissemination of these bacteria in natural environmentssuch as in-hospital spaces thereby increasing the chance ofinfection in immunosuppressed patients [23 39 40]
Based on the GPL biosynthetic pathway previouslydescribed for the M avium 104 strain [25] we are currentlyconstructing a potential gene cluster for GPL biosynthesis inM colombiense which will be finished when the nucleotidesequence of the CECT 3035 is completely assembled RT-qPCR experiments showed that the planktonic and motilecells of the rough 57B strain showed decreased transcriptionof the 9 selected genes leading to the low production ofGPLs in this rough variant comparedwith theM colombiensestrains displaying smooth colony morphology Interestinglyplanktonic 19B cells exhibited lower expression of most of theselected genes compared with planktonic CECT 3035 cellsThis behaviour results in the reduced production of GPLs inplanktonic 19B cells compared with the CECT 3035 strainconsistent with the results of the TLC experiments
It has been previously shown that an rtfA mutation inthe M avium serovar-2-specific strain resulted in the lossof serovar-specific GPLs thereby diminishing the variabilityof these glycolipids in the cell envelope [41 42] Thus itis possible that the enzymatic activity of the RtfA proteincould be relevant for the increased glycosylation andor aug-mented production of ssGPLs in strains displaying smoothcolony morphology compared with the rough variant 57BIn strains displaying smooth colony morphology the genesencoding rhamnosyl andmethyl transferases (rtfA andmtfB)are overtranscribed suggesting the increased production ofnsGPLs precursors for ssGPL biosynthesis [24ndash26] therebyincreasing the production of GPLs The diminished tran-scription of pstA gftA mtfC mtfD tmtpBand tmtpC inmotile CECT 3035 compared with control cells (planktonicCECT 3035) is intriguing however this behaviour also
suggests the relevance of rhamnosyl transferases particularlyRtfA in GPL biosynthesis in M colombiense Neverthelessthe actual role for RtfA in M colombiense strains should beconfirmed using rtfA-null mutants
We did not identify 7 of the 30 known GPL cluster genesof M avium 104 in the genome sequence of M colombienseCECT 3035 This interesting observation suggests that theGPLs between these two closely related MAC species couldhave different structural characteristics However furtherexperiments using mass spectrometry and NMR are neces-sary to evaluate potential differences in the GPL structureamongM colombiense strains
5 Conclusions
In conclusion the results of the present study show that Mcolombiense strains displaying smooth morphology exhibitincreased biofilm formation on hydrophobic polystyrenesurfaces and enhanced sliding over motility plates Bioin-formatics analyses indicate that the gene cluster establishedfor GPL formation modification and translocation is intactstrongly suggesting that GPLs are putatively present in Mcolombiense In addition motile culture conditions activatethe transcription of the genes implicated in the key enzymaticactivities of GPL biosynthesis
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
Thisworkwas supported through funding from the ldquoDivisionde Investigacion Bogota (DIB)rdquo-Vicerrectorıa de Investi-gacion Universidad Nacional de Colombia (Grants 1606014837 and 14337) Milena Maya-Hoyos received a fellowshipfrom the Academic Direction Universidad Nacional deColombia and the ldquoJovenes Investigadores e InnovadoresrdquoProgram Colciencias Colombia This research was alsosupported in part through the Intramural Research Programof the National Institutes of Health National Library ofMedicine and National Center for Biotechnology Informa-tion (NCBI)
References
[1] S K Field D Fisher and R L Cowie ldquoMycobacterium aviumcomplex pulmonary disease in patient without HIV infectionrdquoChest vol 126 no 2 pp 566ndash581 2004
[2] M J M Vaerewijck G Huys J C Palomino J Swings and FPortaels ldquoMycobacteria in drinking water distribution systemsecology and significance for human healthrdquo FEMSMicrobiologyReviews vol 29 no 5 pp 911ndash934 2005
[3] E Hagiwara S Komatsu R Nishihira T Shinohara T Babaand T Ogura ldquoClinical characteristics and prevalence of pneu-mothorax in patients with pulmonary Mycobacterium aviumcomplex diseaserdquo Journal of Infection and Chemotherapy vol19 no 4 pp 588ndash592 2013
10 BioMed Research International
[4] M I Murcia E Tortoli M C Menendez E Palenque andM J Garcia ldquoMycobacterium colombiense sp nov a novelmember of theMycobacterium avium complex and descriptionof MAC-X as a new ITS genetic variantrdquo International Journalof Systematic and Evolutionary Microbiology vol 56 no 9 pp2049ndash2054 2006
[5] E Tortoli L Rindi M J Garcia et al ldquoProposal to elevatethe genetic variant MAC-A included in the Mycobacteriumavium complex to species rank as Mycobacterium chimaerasp novrdquo International Journal of Systematic and EvolutionaryMicrobiology vol 54 no 4 pp 1277ndash1285 2004
[6] I B Salah C Cayrou D Raoult and M Drancourt ldquoMycobac-terium marseillense sp nov Mycobacterium timonense sp novand Mycobacterium bouchedurhonense sp novmembers ofthe Mycobacterium avium complexrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 11 pp2803ndash2808 2009
[7] J van Ingen M J Boeree K Kosters et al ldquoProposal toelevate Mycobacterium avium complex ITS sequevar MAC-Qto Mycobacterium vulneris sp novrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 9 pp2277ndash2282 2009
[8] D Bang T Herlin M Stegger et al ldquoMycobacterium arosiensesp nov a slowly growing scotochromogenic species causingosteomyelitis in an immunocompromised childrdquo InternationalJournal of Systematic and Evolutionary Microbiology vol 58part 10 pp 2398ndash2402 2008
[9] C Cayrou C Turenne M A Behr and M Drancourt ldquoGeno-typing of Mycobacterium avium complex organisms usingmultispacer sequence typingrdquoMicrobiology vol 156 part 3 pp687ndash694 2010
[10] S C Smole F McAleese J Ngampasutadol C F von Reynand R D Arbeit ldquoClinical and epidemiological correlates ofgenotypes within theMycobacterium avium complex defined byrestriction and sequence analysis of hsp65rdquo Journal of ClinicalMicrobiology vol 40 no 9 pp 3374ndash3380 2002
[11] AMuwonge C Kankya T B Johansen et al ldquoNon-tuberculousmycobacteria isolated from slaughter pigs in Mubende districtUgandardquo BMC Veterinary Research vol 8 article 52 2012
[12] J O Falkinham III ldquoSurrounded by mycobacteria nontuber-culous mycobacteria in the human environmentrdquo Journal ofApplied Microbiology vol 107 no 2 pp 356ndash367 2009
[13] K Vuorenmaa I B Salah V Barlogis H Chambost and MDrancourt ldquoMycobacterium colombiense and pseudotubercu-lous lymphadenopathyrdquo Emerging Infectious Diseases vol 15no 4 pp 618ndash620 2009
[14] O Esparcia F Navarro M Quer and P Coll ldquoLymphadenopa-thy caused by Mycobacterium colombienserdquo Journal of ClinicalMicrobiology vol 46 no 5 pp 1885ndash1887 2008
[15] S Cohen-Bacrie M David N Stremler J-C Dubus J-MRolain and M Drancourt ldquoMycobacterium chimaera pul-monary infection complicating cystic fibrosis a case reportrdquoJournal of Medical Case Reports vol 5 no 1 article 473 2011
[16] S Poulin C Corbeil M Nguyen et al ldquoFatal Mycobac-terium colombiensecytomegalovirus coinfection associatedwith acquired immunodeficiency due to autoantibodies againstinterferon gamma a case reportrdquo BMC Infectious Diseases vol13 no 1 article 24 2013
[17] J Leguizamon J Hernandez M-I Murcia and C-Y SotoldquoIdentification of potential biomarkers to distinguishMycobac-terium colombiense from other mycobacterial speciesrdquoMolecu-lar and Cellular Probes vol 27 no 1 pp 46ndash52 2013
[18] D Chatterjee and K H Khoo ldquoThe surface glycopeptidolipidsof mycobacteria structures and biological propertiesrdquo Cellularand Molecular Life Sciences vol 58 no 14 pp 2018ndash2042 2001
[19] J B Torrelles D Ellis T Osborne et al ldquoCharacterizationof virulence colony morphotype and the glycopeptidolipid ofMycobacterium avium strain 104rdquo Tuberculosis vol 82 no 6pp 293ndash300 2002
[20] M F Thorel and H L David ldquoSpecific surface antigens of SmTvariants ofMycobacterium aviumrdquo Infection and Immunity vol43 no 1 pp 438ndash439 1984
[21] A Martınez S Torello and R Kolter ldquoSliding motility inmycobacteriardquo Journal of Bacteriology vol 181 no 23 pp 7331ndash7338 1999
[22] J Recht AMartınez S Torello and R Kolter ldquoGenetic analysisof sliding motility in Mycobacterium smegmatisrdquo Journal ofBacteriology vol 182 no 15 pp 4348ndash4351 2000
[23] G Carter M Wu D C Drummond and L E BermudezldquoCharacterization of biofilm formation by clinical isolates ofMycobacterium aviumrdquo Journal of Medical Microbiology vol 52no 9 pp 747ndash752 2003
[24] J S Schorey and L Sweet ldquoThe mycobacterial glycopepti-dolipids structure function and their role in pathogenesisrdquoGlycobiology vol 18 no 11 pp 832ndash841 2008
[25] L Pang X Tian W Pan and J Xie ldquoStructure and function ofmycobacterium glycopeptidolipids from comparative genomicsperspectiverdquo Journal of Cellular Biochemistry vol 114 no 8 pp1705ndash1713 2013
[26] R Mukherjee and D Chatterji ldquoGlycopeptidolipids immuno-modulators in greasy mycobacterial cell enveloperdquo IUBMB Lifevol 64 no 3 pp 215ndash225 2012
[27] K L Horan R Freeman K Weigel et al ldquoIsolation ofthe genome sequence strain Mycobacterium avium 104 frommultiple patients over a 17-year periodrdquo Journal of ClinicalMicrobiology vol 44 no 3 pp 783ndash789 2006
[28] S B Snapper R E Melton S Mustafa T Kieser and W RJacobs Jr ldquoIsolation and characterization of efficient plasmidtransformation mutants of Mycobacterium smegmatisrdquo Molec-ular Microbiology vol 4 no 11 pp 1911ndash1919 1990
[29] M Radji R A Agustama B Elya and C R TjampakasarildquoAntimicrobial activity of green tea extract against isolatesof methicillin-resistant Staphylococcus aureus and multi-drugresistant Pseudomonas aeruginosardquo Asian Pacific Journal ofTropical Biomedicine vol 3 no 8 pp 663ndash667 2013
[30] P Helden T Victor R Warren and E Helden ldquoIsolationof DNA from Mycobacterium tuberculosisrdquo in Mycobacteriumtuberculosis Protocols Humana Press Totowa NJ USA 2001
[31] J C van Kessel L J Marinelli and G F Hatfull ldquoRecom-bineering mycobacteria and their phagesrdquo Nature ReviewsMicrobiology vol 6 no 11 pp 851ndash857 2008
[32] N M Parrish C G Ko J D Dick P B Jones and J LE Ellingson ldquoGrowth Congo Red agar colony morphotypesand antibiotic susceptibility testing of Mycobacterium aviumsubspecies paratuberculosisrdquo Clin Med Res vol 2 no 2 pp107ndash114 2004
[33] T Tugrul and E Cansunar ldquoDetecting surfactant-producingmicroorganisms by the drop-collapse testrdquo World Journal ofMicrobiology and Biotechnology vol 21 no 6-7 pp 851ndash8532005
[34] D K Jain D L Collins-Thompson H Lee and J T TrevorsldquoA drop-collapsing test for screening surfactant-producingmicroorganismsrdquo Journal ofMicrobiologicalMethods vol 13 no4 pp 271ndash279 1991
BioMed Research International 11
[35] M Munoz M-A Laneelle M Luquin et al ldquoOccurrence ofan antigenic triacyl trehalose in clinical isolates and referencestrains of Mycobacterium tuberculosisrdquo FEMS MicrobiologyLetters vol 157 no 2 pp 251ndash259 1997
[36] M Gonzalez-Perez M I Murcia D Landsman I K Jordanand L Marino-Ramırez ldquoGenome sequence of the Mycobac-terium colombiense type strain CECT 3035rdquo Journal of Bacte-riology vol 193 no 20 pp 5866ndash5867 2011
[37] J C Betts P T Lukey L C Robb R A McAdam andK Duncan ldquoEvaluation of a nutrient starvation model ofMycobacterium tuberculosis persistence by gene and proteinexpression profilingrdquoMolecular Microbiology vol 43 no 3 pp717ndash731 2002
[38] W W Barrow and P J Brennan ldquoIsolation in high frequencyof rough variants of Mycobacterium intracellulare lacking C-mycoside glycopeptidolipid antigensrdquo Journal of Bacteriologyvol 150 no 1 pp 381ndash384 1982
[39] W Costerton R Veeh M Shirtliff M Pasmore C Post andG Ehrlich ldquoThe application of biofilm science to the studyand control of chronic bacterial infectionsrdquo Journal of ClinicalInvestigation vol 112 no 10 pp 1466ndash1477 2003
[40] J Recht and R Kolter ldquoGlycopeptidolipid acetylation affectssliding motility and biofilm formation inMycobacterium smeg-matisrdquo Journal of Bacteriology vol 183 no 19 pp 5718ndash57242001
[41] T M Eckstein F S Silbaq D Chatterjee N J Kelly P JBrennan and J T Belisle ldquoIdentification and recombinantexpression of a Mycobacterium avium rhamnosyltransferasegene (rtfA) involved in glycopeptidolipid biosynthesisrdquo Journalof Bacteriology vol 180 no 21 pp 5567ndash5573 1998
[42] J N Maslow V R Irani S-H Lee T M Eckstein J MInamine and J T Belisle ldquoBiosynthetic specificity of therhamnosyltransferase gene of Mycobacterium avium serovar 2as determined by allelic exchange mutagenesisrdquo Microbiologyvol 149 no 11 pp 3193ndash3202 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 BioMed Research International
the smooth variants being the most virulent morphology[18 19] In addition MAC strains spread on solid hydrophilicsurfaces through sliding motility mechanisms that are inde-pendent of extracellular structures [21 22] Bacterial motilityplays a significant role in the colonisation of environmentalsurfaces and cells [21] which in turn has been correlatedin vitro with the capacity to form biofilms on hydrophobicsurfaces [23] In M avium strains motility and biofilmformation have been correlated with colony morphology andthe presence of glycopeptidolipids (GPLs) in the cell envelope[24 25] Specifically smooth transparent M avium variantsshow higher motility on hydrophilic surfaces and increasedGPL production conversely rough variants show diminishedmotility and impaired GPL production [22]
GPLs are glycolipids attached to the outermost portionof some nontuberculous mycobacteria including M aviumM smegmatisM abscessus andM fortuitum [25] This typeof glycolipid comprises a mixture of 3-hydroxy or 3-methoxyC26ndashC34
fatty acids amidated to a tripeptide-amino-alcohol(D-phenylalanine-D-allo-threonine-D-alanine-L-alaninol)[24 25] The lipopeptide core is subsequently glycosylatedthrough 6-deoxytalose (linked to the allo-threonine) and120572-L-rhamnose (linked to L-alaninol) and the resultingoligosaccharide residues are methylated to form the non-serovar-specificGPLs (nsGPLs) present in all GPL-producingmycobacteria However MAC strains also synthesise polarGPLs in which some oligosaccharides residues are attachedto the 6-deoxytalose producing serovar-specific GPLs(ssGPLs) [24 25] In M avium the consecutive actionsof a peptide synthetase (PstAPstB) a fatty acid synthasesystem (FAS) and polyketide synthetase (Pks) polyketidesynthetase associated protein (PapA) glycosyl and methyltransferases (GtfAGtfBRtfA and MtfAMtfBMtfCMtfD)and glycolipid transporters (TmtP and Gap) are responsiblefor the biosynthesis and subsequent translocation of GPLs tothe cell wall surface [24ndash26]
Currently the existence of GPLs inM colombiense strainsis completely unknown In the present study we showed thatM colombiense contains glycolipids with chromatographicbehaviours similar to GPLs In addition this novel speciesforms biofilms on the hydrophobic surfaces of polystyreneand motility is increased in strains displaying smoothcolony morphology Moreover we examined the genes likelyinvolved in GPL biosynthesis in the CECT 3035 strain
2 Material and Methods
21 Bacterial Strains Culture Conditions and Genomic DNAIsolation The M colombiense clinical isolates 19B 57Bthe M colombiense genome sequence strain CECT 3035M avium 104 [27] and M smegmatis mc2155 [28] wereused in this study (Table 1) Planktonic mycobacteria werecultured at 37∘C with agitation (76 rpm) in Middlebrook7H9 media supplemented with ADC (05 (wv) bovineserum albumin 02 (wv) dextrose 0085 (wv) NaCl and00003 (wv) beef catalase) and 005 (vv) glycerol untilan OD
600of 05 was obtained (planktonic conditions) For
the cell motility assay mycobacteria were cultured inmotilitymedium containing 7H9 supplemented with ADC and 035
agarose Pseudomonas aeruginosa ATCC27853 [29] culturedinmotilitymediumwas used as a positive control in the drop-collapsing test
For DNA extraction the mycobacteria were grown in7H9-ADC broth to an OD
600of 05 centrifuged and resus-
pended in TE buffer (10mM Tris-HCl and 1mM EDTA pH8) Subsequently the bacilli were inactivated through incu-bation at 80∘C for 20 minutes Genomic DNA was extractedusing lysozyme SDSproteinase K and CTABNaCl for celldisruption and chloroform isoamyl alcohol (24 1 vv) forgetting rid of proteins [30 31] The DNA pellets were treatedwithDNAse-free RNAse resuspended in 01X TE followed byquantification using a NanoDrop 2000c Spectrophotometer(ThermoScientificMAUSA)TheDNAqualitywas assessedusing agarose gel electrophoresis and spectrophotometry(OD260
OD280
)
22 Cell Motility Assay and Congo Red Staining For the cellmotility assay plates containing 7H9 media supplementedwithADCand035agarosewere incubated at room temper-ature overnight prior to cell inoculation [21] Subsequently3 120583L of mycobacterial culture at an OD
600of 06 (sim27 times
105 CFU) was carefully spotted onto the centre of the plates tominimise the spread of cells from the inoculation point Theplates were dried under sterile conditions for 30min sealedwith Parafilm and incubated at 37∘C for 4-5 weeks underhumidified conditions The motility rate was recorded asthe colony growth halo The motility assays were performedin triplicate from three independent experiments using Mavium 104 as a control Differences among the experimentaldata were compared using Studentrsquos t-test and 119875 lt 005 wasconsidered significantly different
For the characterisation of colony morphology platescontaining 7H10-OADC (ADC + 005 oleic acid) mediasupplemented with 100 120583gmL Congo Red (Sigma-AldrichMO USA) [32] were inoculated as in the cell motility assayand the resulting cultures were incubated for 4-5 weeksat 37∘C under humidified conditions The morphology wasdefined as rough or smooth opaque or transparent andyellow or beige
23 Biofilm FormationAssay Biofilm formationwas assessedas previously described [22] with some modificationsBriefly the wells of polystyrene microtiter plates (TPPSwitzerland) were filled with a mix of 75120583L of planktonicmycobacterial culture (OD
600= 06) and 125 120583L of 7H9-
ADC broth The plates were incubated at 37∘C for 6 weeksSubsequently the plates were washed three times with 1XPBS (014M NaCl 5mM KCl 10mM Na
2HPO4 and 2mM
KH2PO4) 200120583L of 01 crystal violet solution was added
and the cells were incubated at 37∘C for an additional30min The residual crystal violet solution was removedand the wells were washed three times with 1X PBS bufferA total of 200120583L of absolute ethanol was added to eachwell and the plates were incubated for 1 hour at roomtemperature Colouration was quantified at 570 nm using aiMark Microplate Absorbance Reader 168-1135 (BioRad PAUSA) The biofilm assays were performed in triplicate fromthree independent experiments using M avium 104 as a
BioMed Research International 3
Table 1 Bacterial strains and primers used in this study
Strain and primers Description SourceM colombiense (CECT 3035) Sequence genome strain Clinical isolateM colombiense 19B and 57B Clinical isolates Clinical isolateM smegmatismc2155 Reference strain [28]M avium 104 Reference strain [27]P aeruginosa ATCC27853 Reference strain [29]Primers Sequence (51015840-31015840)pstA dir ACAGGGCACGAGGAATTCTA This studypstA rev TAGTCCTCGGAGGCTTCGTA This studygftA dir ATGTGTGCTGGCCAGTTATG This studygftA rev GGAAGAACGACGTCCAGAAG This studyrtfA dir GACTTTTGGAGCGACGAGTT This studyrtfA rev GCCAAATCCTGGTAAAGCTG This studymtfB dir GGACACCGAGCACTACGAG This studymtfB rev TCATACAGATCGCCATCCAG This studymtfC dir ACAAGGCGGATAAAGGGATT This studymtfC rev CTCATACAGATCGCCATCCA This studymtfD dir TACCTGCTCGACACCTTCG This studymtfD rev TCGACCTGCTCGAGTGTCT This studytmtpC dir TTCATTCGGGATACCAGGAG This studytmtpC rev TTGATCCTGACCCGAAGTTT This studytmtpA dir CTCTCGGCTTTGACGACAC This studytmtpA rev ATGGCCGACATCAGCTACTT This studytmtpB dir GAGTGCCCTTGAGTGATTCC This studytmtpB rev CCTCCAAGAATGACGATTCC This study16 sRNA dir GAGATAGGCGTTCCCTTGTG This study16 sRNA rev CTGGACATAAGGGGCATGAT This study
positive control Differences among experimental data werecompared using Studentrsquos t-test and119875 lt 005was consideredsignificantly different
24 Drop-Collapsing Test Mycobacteria grown on motilitymedium were scraped and resuspended in 5mL 1X PBSgently vortexed and centrifuged at 8000 rpm for 1 h Sub-sequently 20120583L of supernatant was carefully spotted ontothe 154mm wells of polystyrene plates (TPP Switzerland)previously coated with 100120583L of mineral oil and incubatedfor 24 h at room temperature [33 34] The aqueous dropswere visually examined incubated for 1min at room tem-perature and subsequently photographed The tests wereperformed in triplicate from three independent experimentsusing deionised water and 1 SDSM smegmatismc2155 andP aeruginosa as controls
25 Lipid Extraction and TLC Analysis Crudemycobacteriallipid extracts were obtained as previously described [35] withsome modifications Briefly mycobacteria cultured underplanktonic and motility conditions were extracted usingchloroform-methanol (1 2 vv) through vigorous stirringfor 48 h at room temperature followed by chloroform-methanol (2 1 vv) extraction under the same experimentalconditions Pooled and dried organic extracts were parti-tioned using a chloroform-methanol-water mixture (8 4 2vvv) The organic phase was separated and evaporated todryness and the free glycolipids extracts were weighed ForGPL detection 1mg of the lipid extracts was dissolved in
chloroform-methanol (2 1 vv) at 20mgmL and analysedon 20 times 20 Silica Gel 60 TLC plates (Merck NJ USA)using chloroform-methanol-water (65 25 4 vvv) [35]Thecarbohydrate-containing compounds were visualised afterspraying the plates with 1 anthrone (Sigma-Aldrich MOUSA) in sulphuric acid followed by heating at 120∘C CrudeM avium 104 lipid extract was used as a control andthe glycolipid profile was obtained from three independentexperiments performed in duplicate
26 Bioinformatics Analysis The genes implicated in the bio-synthesis of GPLs in theM avium 104 strain (httpwwwnc-binlmnihgovnuccoreCP0004791) were used as model forthe bioinformatics analysisThe gene sequences for pstApstB(peptide synthetases) pks10 (polyketide synthetase) gtfAgtfB and rtfA (glycosyl transferases) mtfA mtfB mtfC andmtfD (methyl transferases) tmtpC tmtpA and tmtpB (gly-colipid transporters) and papA2 (acyl transferase) (Table 1)were identified in M colombiense CECT 3035 using the Ge-nomic BLAST tool (httpwwwncbinlmnihgovsutilsge-nom tablecgi) The orthologous sequences identified in theM colombiense CECT 3035 genome [36] were subsequentlyused as targets for primer design in the RT-qPCR experi-ments
27 RNA Isolation and RT-qPCR Analysis RNA ofM colom-biense 19B CECT 3035 and 57B cultured on both motilitymedium and planktonic conditions was isolated for RT-qPCR experimentsMycobacteria grown onmotilitymedium
4 BioMed Research International
(a) (b)
(c) (d)
Figure 1 Colonymorphology ofMycobacterium colombiense strainsMycobacterial strains were grown on 7H10-OADC plates supplementedwith Congo Red and incubated at 37∘C for 4-5 weeks (a)M colombiense 19B (b) CECT 3035 (c) 57B and (d)M avium 104
were scraped and resuspended in diethylpyrocarbonate-(DEPC-) treated water Motile and planktonic mycobacteriawere centrifuged washed with DEPC-treated water andresuspended in TRIzol (Invitrogen USA) The cells werelysed using a Mini-Bead Beater 16 (BioSpec OK USA) andglass beads (01mm) Total RNA was isolated as previouslydescribed [37] and quantified using aNanoDrop 2000c Spec-trophotometer (Thermo Scientific MA USA) After DNaseI treatment the RNA quality was evaluated through agarosegel electrophoresis and spectrophotometry (OD
260OD280
)cDNA libraries were constructed from 1 120583g of RNA using theRevertAid First cDNA Synthesis Kit (Fermentas Lithuania)For reverse transcription the samples were incubated at 37∘Cfor 30min 42∘C for 5 h 72∘C for 1min and 4∘C for 10min
The primers used for the RT-qPCR analyses are listed inTable 1The RT-qPCR reactions were set up in triplicate usingthe Express SYBR GreenER Universal qPCR SuperMix Kit(Invitrogen NY USA) and a CFX-96 thermocycler (BioradPA USA) under conditions including a denaturation stepfor 5min at 95∘C followed by 39 cycles of 95∘C for 10 sec
58∘C for 10 sec and 72∘C for 15 sec Primers or cDNA wereomitted in the negative controls The gene transcriptionwas normalised to the mean value of 16S rRNA (rrs) geneexpressionThe transcription profile was obtained from threeindependent experiments performed in duplicate For eachexperiment the differences among experimental data werecompared using Studentrsquos t-test and119875 lt 005was consideredsignificantly different
3 Results
31M colombiense IsMotile and Forms Biofilms onHydropho-bic Polystyrene Surfaces The M colombiense strains usedin this study including the genome sequence strain CECT3035 presented smooth opaque and brilliant colonies on7H10-OADC plates supplemented with Congo Red exceptstrain 57B which displayed a rough and wrinkled phenotype(Figure 1) Similar results were obtained using medium lack-ing oleic acid (7H9-ADC-035 agarose data not shown)Other M colombiense strains not included in the present
BioMed Research International 5
7
17
27
37
47
57
0 10 20 30 40 50 60 70
Dia
met
er (m
m)
Day
M colombiense 19BM avium
M colombiense CECT 3035
M colombiense 57B
Figure 2 Motility rates for Mycobacterium colombiense strainsMycobacterial strains were cultivated on 7H9-ADC-035 agaroseand incubated at 37∘C for 4-5 weeks The colony growth haloswere measured daily to determine the motility rate Each reportedvalue represents the mean of the colony growth halos from threeindependent experiments The presented data have statisticallysignificant differences compared with the values obtained from Mavium 104 (119875 lt 005)
study (6B 7B 9B and 16B) also displayed a smooth colonymorphology (data not shown) Both smooth and rough bac-terial phenotypes were stable after repetitive culture on solidmedia for the duration of the study (3 years) The smoothM colombiense strains developed yellow pigmentation withage specifically at the stationary phase in all culture mediahowever the rough variant (57B) remained beige in colourunder the same experimental conditions (Figure 1)
Because alterations in the phenotypic characteristicsassociated with colony morphology have been observedin other MAC species motility and biofilm formationwere compared between smooth and rough M colombiensestrains We observed that the M colombiense 19B andCECT3035 strains (smooth colony morphology) showedincreased spreading (4094 plusmn 005 120583mday and 2972 plusmn005 120583mday resp) on hydrophilic agarose comparedwith thenatural rough variant 57B which showed impaired spreading(Figure 2)
Crystal violet staining was used to quantify the degree ofM colombiense biofilm formation onpolystyreneThehighestOD570
values for all M colombiense strains were obtainedusing an inoculum of sim27 times 105 CFUmL In general thestrains displaying smooth morphology showed increasedbiofilm formation on hydrophobic surfaces while therough variant 57B showed reduced adhesion to polystyrene(Figure 3)
As biosurfactants could potentially influence cell motilitythe ldquodrop-collapsingrdquo method was used to detect biosurfac-tant secretion in M colombiense strains [22] As shown inFigure 4 drops of 1 SDS and P aeruginosa aqueous extract(positive controls) spread over the oily surface after the sam-ples were incubated indicating the presence of biosurfactantsubstances In contrast drops of distilled waterM smegmatis(negative control) and M colombiense aqueous extracts didnot collapse on the oily surface but rather appeared as firm
0
005
01
015
02
025
OD570
Mavium
104
Mcolom
biense
CECT
3035
Mcolom
biense19
B
Mcolom
biense57
B
Figure 3 Capacity of Mycobacterium colombiense strains to formbiofilms The OD
570nm represents the proportion of mycobacteria
added to the microtiter wells that reacted with the crystal violetsolutionThebars represent the SD calculated from two independentexperiments each performed in triplicate
drops suggesting the absence of biosurfactants in the culturesupernatants
32 M colombiense Displaying a Smooth Colony Morphol-ogy Contains Glycolipids with Thin-Layer ChromatographicBehaviour Similar to That of the GPLs of M avium TLCanalysis of noncovalently attached lipids extracted frommycobacteria cultured under planktonic and motile condi-tions showed that M colombiense strains contain glycolipidswith chromatographic behaviour similar to that of theGPLsofM avium [23] Regarding planktonic cells (Figure 5(a)) thestrains with smooth colony morphology (19B and CECT3035) showed multiple lipid spots migrating in the region ofcontrol GPLs (M avium 104) however the spots observed inthe CECT 3035 lipid extract were more intense than those inthe 19B strain The natural rough variant (57B) showed onlyone spot that migrated in the same GPLs region
Under motile conditions (Figure 5(b)) the smoothcolony morphology strains 19B and CECT 3035 showed aGPL profile similar to that obtained for cells cultured underplanktonic conditions nevertheless the counterpart spotswere more intense than those observed in planktonic cellsparticularly for the 19B strain Nevertheless the unique spotobserved for motile 57B cells (rough variant) were moreintense than that observed for planktonic 57B cells Inter-estingly all motile M colombiense cells exhibited reducedproduction of polar glycolipids such as phosphatidyl-inositolmannosides (PIMs) and phosphatidylglycerol (PG) com-pared with planktonic cells (Figure 5(b))
33 In Silico Identification of GPL Biosynthesis Genes inM colombiense CECT 3035 The genes involved in GPLbiosynthesis in M colombiense are currently unknown
6 BioMed Research International
(a) (b) (c) (d)
(e) (f) (g) (h)
Figure 4 Drop-collapsing test (a) Water (no surfactant) (b) 1 SDS (surfactant) (c) P aeruginosa (d) M smegmatis mc2155 (e) Mcolombiense 19B (f) M colombiense CECT 3035 (g) M colombiense 57B and (h) M avium 104 Drop spreading over the oily surface aftersamples incubation indicates the presence of biosurfactant substances in the aqueous sampleThis assay was conducted in three independentexperiments each performed in duplicate
Based on the GPL biosynthetic pathway reported for Mavium 104 [25] 30 open reading frames (ORF) encodingpeptide synthetase fatty acid synthases (FAS) polyketidesynthases (PKs) acyltransferase (PapA) glycosyltransferasescarbohydrate synthetases methyl and acyl transferases gly-colipid transporters potential biosynthesis regulators andproteins with unknown functions [25 26] were searchedwithin the recently reported nucleotide sequence of Mcolombiense CECT 3035 [36] This in silico analysis (seeTable S1 of the Supplementary Material available online athttpdxdoiorg1011552015419549) showed that all enzy-matic activities searched were encoded by different ORFsin M colombiense CECT 3035 with an identity rangingbetween 77 and 96 The ORFs in the genome of the Mcolombiense counterpart were located in the contigs 0000100002A 00003A and 00007 of the genome sequence of theCECT 3035 strain (Figure 6) Among the 30 genes of the GPLsynthetic pathway inM avium 104 7 genes (mtfA gtfB gtfDdghA fadD23 pe and gap-like) did not have orthologoussequences inM colombiense CECT 3035 (Table S1)
34 Differential Transcription of the GPL Biosynthesis Genesin M colombiense Displaying Smooth and Rough ColonyMorphologies RNAwas isolated fromM colombienseCECT3035 19B and 57B strains cultured under planktonic andmotile conditions to quantify the transcription levels ofthe pstA gtfA rtfA mtfB mtfC mtfD tmtpC tmtpA andtmtpB genes predicted to encode key enzymes of the GPLbiosynthetic pathway (Table S1) For relative quantificationM colombiense CECT 3035 was grown to the exponentialphase under planktonic conditions and used as the referencepattern for two reasons (1) CECT3035 is theM colombiensegenome sequence strain [36] and consistent with the TLCanalysis (2) planktonic cells displayed the lowest GPL pro-duction
With regard to planktonic cells the rate ofM colombiense19B gene transcription was consistently higher than thatfor the M colombiense 57B genes Conversely the roughvariant (57B) displayed a dramatic decrease in the rate oftranscription for the 9 selected genes particularly thoseencoding peptide synthetase pstA glycosyltransferase gtfA
BioMed Research International 7
1 2 3 41 2 3 4
GPLs
PG
PIMs
(a) (b)
Figure 5 TLC profile of the GPLs in Mycobacterium colombiense strains Crude lipid extracts of (1) M avium 104 (2) M colombiense19B (3) M colombiense CECT 3035 and (4) M colombiense 57B Cells cultivated (a) under planktonic and (b) motile conditions weredeveloped using chloroform-methanol-water (65 25 4 vvv) GPLs glycopeptidolipids PIMs phosphatidyl-inositol mannosides and PGphosphatidylglycerol
the methyltransferases mtfB and mtfC and the lipid trans-porters tmtpA tmtpB and tmtpC (Figure 7(a)) InterestinglyrtfA (rhamnosyltransferase) in the 19B strain (smooth colonymorphology) was the only gene among the strains culturedunder planktonic conditions that showed an increased tran-scription rate (221-fold higher) compared with the controlcells (planktonicM colombiense CECT 3035 at the exponen-tial phase of growth)
Under motile growth conditions M colombiense 19Bshowed the highest transcription rate for all selected genescompared with the CECT 3035 and 57B strains (Figure 7(b))In contrast the transcription rates for all genes in the57B strain (rough variant) except tmtpA and tmtpC (lipidtransporters) were reduced In motile CECT 3035 cellswhereas rtfA mtfB (methyl transfers) and tmtpA showedincreased transcription the pstA gftA mtfC mtfD tmtpBand tmtpC genes exhibited reduced transcription comparedwith control cells
4 Discussion
In the present study TLC experiments were able to show thatM colombiense contains glycolipids with chromatographicbehaviour similar to the GPLs of M avium 104 which havebeen previously identified and completely characterised [23]in addition the colony morphology was associated withthe GPL profile of M colombiense strains Thus the TLCanalysis revealed that (1) strains displaying smooth colonymorphology (19B and CECT3035) produce a higher amountof variable GPLs compared with the rough natural variant(57B) strain and (2) the GPL production was augmentedin M colombiense cells cultivated under the motile growthconditions As expected the TLC analyses did not provide
detailed information about GPL structure however struc-tural determinationwas not the objective of the present study
Themedia composition influences the glycolipid contentof the mycobacterial cell wall The MAC strains growing onsolid medium exhibit a significant rate of transition fromsmooth transparent to smooth opaque and from smooth torough morphologies [18 19 38] We compared planktonicand motile M colombiense cells grown in the same culturemedium (7H9-ADC or 7H9-ADC-agarose) to avoid differ-ences in lipid content produced through alterations in themedia composition The results showed that the low GPLcontent observed in the rough 57B variant is less frequentamong M colombiense strains therefore M colombienseshows a preference for augmented GPL production andsmooth colony morphology Interestingly planktonic Mcolombiense cells displayed increased PIM and PG pro-duction and diminished GPL content compared with cellscultivated under motile conditions suggesting a potentialcompensatory mechanism in cell wall lipid synthesis thatcompensates for the low GPL content
The abundance of GPLs in the outermost portion ofthe cell envelope could be associated with the motility ofM colombiense strains on hydrophilic surfaces Recht andothers [22] proposed that GPLs are linked through thehydrophilic head to the cellular capsule and the hydrophobicfatty acid chain is exposed to the bacterial surface therebyreducing interactions with hydrophilic agarose surfaces andfacilitating the spread of cells on solid medium [21 22]Thus the augmented GPL production of motile 19B andCECT 3035 cells compared with that of the rough variant57B is consistent with the sliding motility on the hydrophilicagarose surface observed for these strains It has also beensuggested that bacterial sliding motility might be favoured
8 BioMed Research International
33kb 2kb9kb51kb
Contig 00001
600kb
87kb 245kb
488kb 36kb320kb
Contig 00002A
Contig 00003A
584kb 452kb
mmpL10
326kb
Contig 00007
fadE
15
atf
rmlB
ecf
rv1174
rmlA
mmpL
4a
mmpL
4b
mmpS4
mbtH
gap rv0926
sap
pks papA3
gtfA
mtfB
mtfC
mtfD
rtfA
pstA
pstB
(a)
ecf
fadE
5
rv1174
atf
rmlB
rmlA
mmpL
4a
mmpL
4b
mmpS
4
mbtH
gap
237kb
rv0926
7kb90kb
530kb
300kb
738kb
mmpL10
sap
Saccharides biosynthesis transfer and modification
Fatty acyl chain biosynthesis modification and transfer
Members of the mmpSL family
Peptide synthesis Unknown
Possible regulatory genes
GPL export to the cell surface
pks pe fadD23 gap-likepapA3
mtfB
gtfA
mtfC
mtfD
dhgA
rtfA
pstA
pstB
mtfA
gtfB
gtfD
(b)
Figure 6Model showing the gene cluster proposed for the GPL biosynthesis ofMycobacterium colombienseCECT 3035 (a)The location andidentity of theM colombiense genes were established using the previously described biosynthetic pathway for GPL biosynthesis annotated intheM avium 104 genomic sequence (b)
0001
001
01
1
10
Rela
tive g
ene t
rans
crip
tion
M colombiense 57BM colombiense 19B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(a)
01
1
10
100
Rela
tive g
ene t
rans
crip
tion
M colombiense CECT 3035
M colombiense 19B
M colombiense 57B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(b)
Figure 7 In vitro transcription of Mycobacterium colombiense genes involved in GPL biosynthesis Relative quantification expressed as theratio of gene transcription in (a) M colombiense cells grown under planktonic conditionstranscription ofM colombiense CECT 3035 cellsgrown under planktonic conditions and (b)M colombiense cells grown on the motility mediumtranscription ofM colombiense CECT 3035grown under planktonic conditions The presented data have statistically significant differences compared with the values obtained fromMcolombiense CECT 3035 cells (119875 lt 005)
BioMed Research International 9
through the secretion of surfactant substances from cells[21 22] The results of the drop-collapsing test showed thatbiosurfactants were likely not secreted from M colombiensecells suggesting that the motility of M colombiense couldbe favoured through hydrophobic molecules such as GPLsbound to the outermost portion of cells however we cannotcompletely rule out the possibility that some of the surfactantsubstances secreted from M colombiense remained adheredto the mycobacterial cell surface thereby influencing cellmotility
GPLs on the outermost portions of the M colombiensecell envelope generate a more hydrophobic cell surfacethat facilitates initial interactions with hydrophobic surfacesand increase biofilm formation on polystyrene wells Inaddition the increased GPL production in 19B and CECT3035 cells is consistent with the augmented biofilm forma-tion observed for M colombiense strains displaying smoothcolony morphology Moreover increased biofilm formationon hydrophobic surfaces and enhanced sliding over motilityplates for 19B cells compared with the CECT 3035 strainwhich also displays smooth colony morphology likely sug-gest possible differences in GPL structure betweenM colom-biense strains Differences inGPL production between strainscould influence the M colombiense ability to form biofilmson polyvinylchloride (PVC) pipes which would facilitatethe dissemination of these bacteria in natural environmentssuch as in-hospital spaces thereby increasing the chance ofinfection in immunosuppressed patients [23 39 40]
Based on the GPL biosynthetic pathway previouslydescribed for the M avium 104 strain [25] we are currentlyconstructing a potential gene cluster for GPL biosynthesis inM colombiense which will be finished when the nucleotidesequence of the CECT 3035 is completely assembled RT-qPCR experiments showed that the planktonic and motilecells of the rough 57B strain showed decreased transcriptionof the 9 selected genes leading to the low production ofGPLs in this rough variant comparedwith theM colombiensestrains displaying smooth colony morphology Interestinglyplanktonic 19B cells exhibited lower expression of most of theselected genes compared with planktonic CECT 3035 cellsThis behaviour results in the reduced production of GPLs inplanktonic 19B cells compared with the CECT 3035 strainconsistent with the results of the TLC experiments
It has been previously shown that an rtfA mutation inthe M avium serovar-2-specific strain resulted in the lossof serovar-specific GPLs thereby diminishing the variabilityof these glycolipids in the cell envelope [41 42] Thus itis possible that the enzymatic activity of the RtfA proteincould be relevant for the increased glycosylation andor aug-mented production of ssGPLs in strains displaying smoothcolony morphology compared with the rough variant 57BIn strains displaying smooth colony morphology the genesencoding rhamnosyl andmethyl transferases (rtfA andmtfB)are overtranscribed suggesting the increased production ofnsGPLs precursors for ssGPL biosynthesis [24ndash26] therebyincreasing the production of GPLs The diminished tran-scription of pstA gftA mtfC mtfD tmtpBand tmtpC inmotile CECT 3035 compared with control cells (planktonicCECT 3035) is intriguing however this behaviour also
suggests the relevance of rhamnosyl transferases particularlyRtfA in GPL biosynthesis in M colombiense Neverthelessthe actual role for RtfA in M colombiense strains should beconfirmed using rtfA-null mutants
We did not identify 7 of the 30 known GPL cluster genesof M avium 104 in the genome sequence of M colombienseCECT 3035 This interesting observation suggests that theGPLs between these two closely related MAC species couldhave different structural characteristics However furtherexperiments using mass spectrometry and NMR are neces-sary to evaluate potential differences in the GPL structureamongM colombiense strains
5 Conclusions
In conclusion the results of the present study show that Mcolombiense strains displaying smooth morphology exhibitincreased biofilm formation on hydrophobic polystyrenesurfaces and enhanced sliding over motility plates Bioin-formatics analyses indicate that the gene cluster establishedfor GPL formation modification and translocation is intactstrongly suggesting that GPLs are putatively present in Mcolombiense In addition motile culture conditions activatethe transcription of the genes implicated in the key enzymaticactivities of GPL biosynthesis
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
Thisworkwas supported through funding from the ldquoDivisionde Investigacion Bogota (DIB)rdquo-Vicerrectorıa de Investi-gacion Universidad Nacional de Colombia (Grants 1606014837 and 14337) Milena Maya-Hoyos received a fellowshipfrom the Academic Direction Universidad Nacional deColombia and the ldquoJovenes Investigadores e InnovadoresrdquoProgram Colciencias Colombia This research was alsosupported in part through the Intramural Research Programof the National Institutes of Health National Library ofMedicine and National Center for Biotechnology Informa-tion (NCBI)
References
[1] S K Field D Fisher and R L Cowie ldquoMycobacterium aviumcomplex pulmonary disease in patient without HIV infectionrdquoChest vol 126 no 2 pp 566ndash581 2004
[2] M J M Vaerewijck G Huys J C Palomino J Swings and FPortaels ldquoMycobacteria in drinking water distribution systemsecology and significance for human healthrdquo FEMSMicrobiologyReviews vol 29 no 5 pp 911ndash934 2005
[3] E Hagiwara S Komatsu R Nishihira T Shinohara T Babaand T Ogura ldquoClinical characteristics and prevalence of pneu-mothorax in patients with pulmonary Mycobacterium aviumcomplex diseaserdquo Journal of Infection and Chemotherapy vol19 no 4 pp 588ndash592 2013
10 BioMed Research International
[4] M I Murcia E Tortoli M C Menendez E Palenque andM J Garcia ldquoMycobacterium colombiense sp nov a novelmember of theMycobacterium avium complex and descriptionof MAC-X as a new ITS genetic variantrdquo International Journalof Systematic and Evolutionary Microbiology vol 56 no 9 pp2049ndash2054 2006
[5] E Tortoli L Rindi M J Garcia et al ldquoProposal to elevatethe genetic variant MAC-A included in the Mycobacteriumavium complex to species rank as Mycobacterium chimaerasp novrdquo International Journal of Systematic and EvolutionaryMicrobiology vol 54 no 4 pp 1277ndash1285 2004
[6] I B Salah C Cayrou D Raoult and M Drancourt ldquoMycobac-terium marseillense sp nov Mycobacterium timonense sp novand Mycobacterium bouchedurhonense sp novmembers ofthe Mycobacterium avium complexrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 11 pp2803ndash2808 2009
[7] J van Ingen M J Boeree K Kosters et al ldquoProposal toelevate Mycobacterium avium complex ITS sequevar MAC-Qto Mycobacterium vulneris sp novrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 9 pp2277ndash2282 2009
[8] D Bang T Herlin M Stegger et al ldquoMycobacterium arosiensesp nov a slowly growing scotochromogenic species causingosteomyelitis in an immunocompromised childrdquo InternationalJournal of Systematic and Evolutionary Microbiology vol 58part 10 pp 2398ndash2402 2008
[9] C Cayrou C Turenne M A Behr and M Drancourt ldquoGeno-typing of Mycobacterium avium complex organisms usingmultispacer sequence typingrdquoMicrobiology vol 156 part 3 pp687ndash694 2010
[10] S C Smole F McAleese J Ngampasutadol C F von Reynand R D Arbeit ldquoClinical and epidemiological correlates ofgenotypes within theMycobacterium avium complex defined byrestriction and sequence analysis of hsp65rdquo Journal of ClinicalMicrobiology vol 40 no 9 pp 3374ndash3380 2002
[11] AMuwonge C Kankya T B Johansen et al ldquoNon-tuberculousmycobacteria isolated from slaughter pigs in Mubende districtUgandardquo BMC Veterinary Research vol 8 article 52 2012
[12] J O Falkinham III ldquoSurrounded by mycobacteria nontuber-culous mycobacteria in the human environmentrdquo Journal ofApplied Microbiology vol 107 no 2 pp 356ndash367 2009
[13] K Vuorenmaa I B Salah V Barlogis H Chambost and MDrancourt ldquoMycobacterium colombiense and pseudotubercu-lous lymphadenopathyrdquo Emerging Infectious Diseases vol 15no 4 pp 618ndash620 2009
[14] O Esparcia F Navarro M Quer and P Coll ldquoLymphadenopa-thy caused by Mycobacterium colombienserdquo Journal of ClinicalMicrobiology vol 46 no 5 pp 1885ndash1887 2008
[15] S Cohen-Bacrie M David N Stremler J-C Dubus J-MRolain and M Drancourt ldquoMycobacterium chimaera pul-monary infection complicating cystic fibrosis a case reportrdquoJournal of Medical Case Reports vol 5 no 1 article 473 2011
[16] S Poulin C Corbeil M Nguyen et al ldquoFatal Mycobac-terium colombiensecytomegalovirus coinfection associatedwith acquired immunodeficiency due to autoantibodies againstinterferon gamma a case reportrdquo BMC Infectious Diseases vol13 no 1 article 24 2013
[17] J Leguizamon J Hernandez M-I Murcia and C-Y SotoldquoIdentification of potential biomarkers to distinguishMycobac-terium colombiense from other mycobacterial speciesrdquoMolecu-lar and Cellular Probes vol 27 no 1 pp 46ndash52 2013
[18] D Chatterjee and K H Khoo ldquoThe surface glycopeptidolipidsof mycobacteria structures and biological propertiesrdquo Cellularand Molecular Life Sciences vol 58 no 14 pp 2018ndash2042 2001
[19] J B Torrelles D Ellis T Osborne et al ldquoCharacterizationof virulence colony morphotype and the glycopeptidolipid ofMycobacterium avium strain 104rdquo Tuberculosis vol 82 no 6pp 293ndash300 2002
[20] M F Thorel and H L David ldquoSpecific surface antigens of SmTvariants ofMycobacterium aviumrdquo Infection and Immunity vol43 no 1 pp 438ndash439 1984
[21] A Martınez S Torello and R Kolter ldquoSliding motility inmycobacteriardquo Journal of Bacteriology vol 181 no 23 pp 7331ndash7338 1999
[22] J Recht AMartınez S Torello and R Kolter ldquoGenetic analysisof sliding motility in Mycobacterium smegmatisrdquo Journal ofBacteriology vol 182 no 15 pp 4348ndash4351 2000
[23] G Carter M Wu D C Drummond and L E BermudezldquoCharacterization of biofilm formation by clinical isolates ofMycobacterium aviumrdquo Journal of Medical Microbiology vol 52no 9 pp 747ndash752 2003
[24] J S Schorey and L Sweet ldquoThe mycobacterial glycopepti-dolipids structure function and their role in pathogenesisrdquoGlycobiology vol 18 no 11 pp 832ndash841 2008
[25] L Pang X Tian W Pan and J Xie ldquoStructure and function ofmycobacterium glycopeptidolipids from comparative genomicsperspectiverdquo Journal of Cellular Biochemistry vol 114 no 8 pp1705ndash1713 2013
[26] R Mukherjee and D Chatterji ldquoGlycopeptidolipids immuno-modulators in greasy mycobacterial cell enveloperdquo IUBMB Lifevol 64 no 3 pp 215ndash225 2012
[27] K L Horan R Freeman K Weigel et al ldquoIsolation ofthe genome sequence strain Mycobacterium avium 104 frommultiple patients over a 17-year periodrdquo Journal of ClinicalMicrobiology vol 44 no 3 pp 783ndash789 2006
[28] S B Snapper R E Melton S Mustafa T Kieser and W RJacobs Jr ldquoIsolation and characterization of efficient plasmidtransformation mutants of Mycobacterium smegmatisrdquo Molec-ular Microbiology vol 4 no 11 pp 1911ndash1919 1990
[29] M Radji R A Agustama B Elya and C R TjampakasarildquoAntimicrobial activity of green tea extract against isolatesof methicillin-resistant Staphylococcus aureus and multi-drugresistant Pseudomonas aeruginosardquo Asian Pacific Journal ofTropical Biomedicine vol 3 no 8 pp 663ndash667 2013
[30] P Helden T Victor R Warren and E Helden ldquoIsolationof DNA from Mycobacterium tuberculosisrdquo in Mycobacteriumtuberculosis Protocols Humana Press Totowa NJ USA 2001
[31] J C van Kessel L J Marinelli and G F Hatfull ldquoRecom-bineering mycobacteria and their phagesrdquo Nature ReviewsMicrobiology vol 6 no 11 pp 851ndash857 2008
[32] N M Parrish C G Ko J D Dick P B Jones and J LE Ellingson ldquoGrowth Congo Red agar colony morphotypesand antibiotic susceptibility testing of Mycobacterium aviumsubspecies paratuberculosisrdquo Clin Med Res vol 2 no 2 pp107ndash114 2004
[33] T Tugrul and E Cansunar ldquoDetecting surfactant-producingmicroorganisms by the drop-collapse testrdquo World Journal ofMicrobiology and Biotechnology vol 21 no 6-7 pp 851ndash8532005
[34] D K Jain D L Collins-Thompson H Lee and J T TrevorsldquoA drop-collapsing test for screening surfactant-producingmicroorganismsrdquo Journal ofMicrobiologicalMethods vol 13 no4 pp 271ndash279 1991
BioMed Research International 11
[35] M Munoz M-A Laneelle M Luquin et al ldquoOccurrence ofan antigenic triacyl trehalose in clinical isolates and referencestrains of Mycobacterium tuberculosisrdquo FEMS MicrobiologyLetters vol 157 no 2 pp 251ndash259 1997
[36] M Gonzalez-Perez M I Murcia D Landsman I K Jordanand L Marino-Ramırez ldquoGenome sequence of the Mycobac-terium colombiense type strain CECT 3035rdquo Journal of Bacte-riology vol 193 no 20 pp 5866ndash5867 2011
[37] J C Betts P T Lukey L C Robb R A McAdam andK Duncan ldquoEvaluation of a nutrient starvation model ofMycobacterium tuberculosis persistence by gene and proteinexpression profilingrdquoMolecular Microbiology vol 43 no 3 pp717ndash731 2002
[38] W W Barrow and P J Brennan ldquoIsolation in high frequencyof rough variants of Mycobacterium intracellulare lacking C-mycoside glycopeptidolipid antigensrdquo Journal of Bacteriologyvol 150 no 1 pp 381ndash384 1982
[39] W Costerton R Veeh M Shirtliff M Pasmore C Post andG Ehrlich ldquoThe application of biofilm science to the studyand control of chronic bacterial infectionsrdquo Journal of ClinicalInvestigation vol 112 no 10 pp 1466ndash1477 2003
[40] J Recht and R Kolter ldquoGlycopeptidolipid acetylation affectssliding motility and biofilm formation inMycobacterium smeg-matisrdquo Journal of Bacteriology vol 183 no 19 pp 5718ndash57242001
[41] T M Eckstein F S Silbaq D Chatterjee N J Kelly P JBrennan and J T Belisle ldquoIdentification and recombinantexpression of a Mycobacterium avium rhamnosyltransferasegene (rtfA) involved in glycopeptidolipid biosynthesisrdquo Journalof Bacteriology vol 180 no 21 pp 5567ndash5573 1998
[42] J N Maslow V R Irani S-H Lee T M Eckstein J MInamine and J T Belisle ldquoBiosynthetic specificity of therhamnosyltransferase gene of Mycobacterium avium serovar 2as determined by allelic exchange mutagenesisrdquo Microbiologyvol 149 no 11 pp 3193ndash3202 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 3
Table 1 Bacterial strains and primers used in this study
Strain and primers Description SourceM colombiense (CECT 3035) Sequence genome strain Clinical isolateM colombiense 19B and 57B Clinical isolates Clinical isolateM smegmatismc2155 Reference strain [28]M avium 104 Reference strain [27]P aeruginosa ATCC27853 Reference strain [29]Primers Sequence (51015840-31015840)pstA dir ACAGGGCACGAGGAATTCTA This studypstA rev TAGTCCTCGGAGGCTTCGTA This studygftA dir ATGTGTGCTGGCCAGTTATG This studygftA rev GGAAGAACGACGTCCAGAAG This studyrtfA dir GACTTTTGGAGCGACGAGTT This studyrtfA rev GCCAAATCCTGGTAAAGCTG This studymtfB dir GGACACCGAGCACTACGAG This studymtfB rev TCATACAGATCGCCATCCAG This studymtfC dir ACAAGGCGGATAAAGGGATT This studymtfC rev CTCATACAGATCGCCATCCA This studymtfD dir TACCTGCTCGACACCTTCG This studymtfD rev TCGACCTGCTCGAGTGTCT This studytmtpC dir TTCATTCGGGATACCAGGAG This studytmtpC rev TTGATCCTGACCCGAAGTTT This studytmtpA dir CTCTCGGCTTTGACGACAC This studytmtpA rev ATGGCCGACATCAGCTACTT This studytmtpB dir GAGTGCCCTTGAGTGATTCC This studytmtpB rev CCTCCAAGAATGACGATTCC This study16 sRNA dir GAGATAGGCGTTCCCTTGTG This study16 sRNA rev CTGGACATAAGGGGCATGAT This study
positive control Differences among experimental data werecompared using Studentrsquos t-test and119875 lt 005was consideredsignificantly different
24 Drop-Collapsing Test Mycobacteria grown on motilitymedium were scraped and resuspended in 5mL 1X PBSgently vortexed and centrifuged at 8000 rpm for 1 h Sub-sequently 20120583L of supernatant was carefully spotted ontothe 154mm wells of polystyrene plates (TPP Switzerland)previously coated with 100120583L of mineral oil and incubatedfor 24 h at room temperature [33 34] The aqueous dropswere visually examined incubated for 1min at room tem-perature and subsequently photographed The tests wereperformed in triplicate from three independent experimentsusing deionised water and 1 SDSM smegmatismc2155 andP aeruginosa as controls
25 Lipid Extraction and TLC Analysis Crudemycobacteriallipid extracts were obtained as previously described [35] withsome modifications Briefly mycobacteria cultured underplanktonic and motility conditions were extracted usingchloroform-methanol (1 2 vv) through vigorous stirringfor 48 h at room temperature followed by chloroform-methanol (2 1 vv) extraction under the same experimentalconditions Pooled and dried organic extracts were parti-tioned using a chloroform-methanol-water mixture (8 4 2vvv) The organic phase was separated and evaporated todryness and the free glycolipids extracts were weighed ForGPL detection 1mg of the lipid extracts was dissolved in
chloroform-methanol (2 1 vv) at 20mgmL and analysedon 20 times 20 Silica Gel 60 TLC plates (Merck NJ USA)using chloroform-methanol-water (65 25 4 vvv) [35]Thecarbohydrate-containing compounds were visualised afterspraying the plates with 1 anthrone (Sigma-Aldrich MOUSA) in sulphuric acid followed by heating at 120∘C CrudeM avium 104 lipid extract was used as a control andthe glycolipid profile was obtained from three independentexperiments performed in duplicate
26 Bioinformatics Analysis The genes implicated in the bio-synthesis of GPLs in theM avium 104 strain (httpwwwnc-binlmnihgovnuccoreCP0004791) were used as model forthe bioinformatics analysisThe gene sequences for pstApstB(peptide synthetases) pks10 (polyketide synthetase) gtfAgtfB and rtfA (glycosyl transferases) mtfA mtfB mtfC andmtfD (methyl transferases) tmtpC tmtpA and tmtpB (gly-colipid transporters) and papA2 (acyl transferase) (Table 1)were identified in M colombiense CECT 3035 using the Ge-nomic BLAST tool (httpwwwncbinlmnihgovsutilsge-nom tablecgi) The orthologous sequences identified in theM colombiense CECT 3035 genome [36] were subsequentlyused as targets for primer design in the RT-qPCR experi-ments
27 RNA Isolation and RT-qPCR Analysis RNA ofM colom-biense 19B CECT 3035 and 57B cultured on both motilitymedium and planktonic conditions was isolated for RT-qPCR experimentsMycobacteria grown onmotilitymedium
4 BioMed Research International
(a) (b)
(c) (d)
Figure 1 Colonymorphology ofMycobacterium colombiense strainsMycobacterial strains were grown on 7H10-OADC plates supplementedwith Congo Red and incubated at 37∘C for 4-5 weeks (a)M colombiense 19B (b) CECT 3035 (c) 57B and (d)M avium 104
were scraped and resuspended in diethylpyrocarbonate-(DEPC-) treated water Motile and planktonic mycobacteriawere centrifuged washed with DEPC-treated water andresuspended in TRIzol (Invitrogen USA) The cells werelysed using a Mini-Bead Beater 16 (BioSpec OK USA) andglass beads (01mm) Total RNA was isolated as previouslydescribed [37] and quantified using aNanoDrop 2000c Spec-trophotometer (Thermo Scientific MA USA) After DNaseI treatment the RNA quality was evaluated through agarosegel electrophoresis and spectrophotometry (OD
260OD280
)cDNA libraries were constructed from 1 120583g of RNA using theRevertAid First cDNA Synthesis Kit (Fermentas Lithuania)For reverse transcription the samples were incubated at 37∘Cfor 30min 42∘C for 5 h 72∘C for 1min and 4∘C for 10min
The primers used for the RT-qPCR analyses are listed inTable 1The RT-qPCR reactions were set up in triplicate usingthe Express SYBR GreenER Universal qPCR SuperMix Kit(Invitrogen NY USA) and a CFX-96 thermocycler (BioradPA USA) under conditions including a denaturation stepfor 5min at 95∘C followed by 39 cycles of 95∘C for 10 sec
58∘C for 10 sec and 72∘C for 15 sec Primers or cDNA wereomitted in the negative controls The gene transcriptionwas normalised to the mean value of 16S rRNA (rrs) geneexpressionThe transcription profile was obtained from threeindependent experiments performed in duplicate For eachexperiment the differences among experimental data werecompared using Studentrsquos t-test and119875 lt 005was consideredsignificantly different
3 Results
31M colombiense IsMotile and Forms Biofilms onHydropho-bic Polystyrene Surfaces The M colombiense strains usedin this study including the genome sequence strain CECT3035 presented smooth opaque and brilliant colonies on7H10-OADC plates supplemented with Congo Red exceptstrain 57B which displayed a rough and wrinkled phenotype(Figure 1) Similar results were obtained using medium lack-ing oleic acid (7H9-ADC-035 agarose data not shown)Other M colombiense strains not included in the present
BioMed Research International 5
7
17
27
37
47
57
0 10 20 30 40 50 60 70
Dia
met
er (m
m)
Day
M colombiense 19BM avium
M colombiense CECT 3035
M colombiense 57B
Figure 2 Motility rates for Mycobacterium colombiense strainsMycobacterial strains were cultivated on 7H9-ADC-035 agaroseand incubated at 37∘C for 4-5 weeks The colony growth haloswere measured daily to determine the motility rate Each reportedvalue represents the mean of the colony growth halos from threeindependent experiments The presented data have statisticallysignificant differences compared with the values obtained from Mavium 104 (119875 lt 005)
study (6B 7B 9B and 16B) also displayed a smooth colonymorphology (data not shown) Both smooth and rough bac-terial phenotypes were stable after repetitive culture on solidmedia for the duration of the study (3 years) The smoothM colombiense strains developed yellow pigmentation withage specifically at the stationary phase in all culture mediahowever the rough variant (57B) remained beige in colourunder the same experimental conditions (Figure 1)
Because alterations in the phenotypic characteristicsassociated with colony morphology have been observedin other MAC species motility and biofilm formationwere compared between smooth and rough M colombiensestrains We observed that the M colombiense 19B andCECT3035 strains (smooth colony morphology) showedincreased spreading (4094 plusmn 005 120583mday and 2972 plusmn005 120583mday resp) on hydrophilic agarose comparedwith thenatural rough variant 57B which showed impaired spreading(Figure 2)
Crystal violet staining was used to quantify the degree ofM colombiense biofilm formation onpolystyreneThehighestOD570
values for all M colombiense strains were obtainedusing an inoculum of sim27 times 105 CFUmL In general thestrains displaying smooth morphology showed increasedbiofilm formation on hydrophobic surfaces while therough variant 57B showed reduced adhesion to polystyrene(Figure 3)
As biosurfactants could potentially influence cell motilitythe ldquodrop-collapsingrdquo method was used to detect biosurfac-tant secretion in M colombiense strains [22] As shown inFigure 4 drops of 1 SDS and P aeruginosa aqueous extract(positive controls) spread over the oily surface after the sam-ples were incubated indicating the presence of biosurfactantsubstances In contrast drops of distilled waterM smegmatis(negative control) and M colombiense aqueous extracts didnot collapse on the oily surface but rather appeared as firm
0
005
01
015
02
025
OD570
Mavium
104
Mcolom
biense
CECT
3035
Mcolom
biense19
B
Mcolom
biense57
B
Figure 3 Capacity of Mycobacterium colombiense strains to formbiofilms The OD
570nm represents the proportion of mycobacteria
added to the microtiter wells that reacted with the crystal violetsolutionThebars represent the SD calculated from two independentexperiments each performed in triplicate
drops suggesting the absence of biosurfactants in the culturesupernatants
32 M colombiense Displaying a Smooth Colony Morphol-ogy Contains Glycolipids with Thin-Layer ChromatographicBehaviour Similar to That of the GPLs of M avium TLCanalysis of noncovalently attached lipids extracted frommycobacteria cultured under planktonic and motile condi-tions showed that M colombiense strains contain glycolipidswith chromatographic behaviour similar to that of theGPLsofM avium [23] Regarding planktonic cells (Figure 5(a)) thestrains with smooth colony morphology (19B and CECT3035) showed multiple lipid spots migrating in the region ofcontrol GPLs (M avium 104) however the spots observed inthe CECT 3035 lipid extract were more intense than those inthe 19B strain The natural rough variant (57B) showed onlyone spot that migrated in the same GPLs region
Under motile conditions (Figure 5(b)) the smoothcolony morphology strains 19B and CECT 3035 showed aGPL profile similar to that obtained for cells cultured underplanktonic conditions nevertheless the counterpart spotswere more intense than those observed in planktonic cellsparticularly for the 19B strain Nevertheless the unique spotobserved for motile 57B cells (rough variant) were moreintense than that observed for planktonic 57B cells Inter-estingly all motile M colombiense cells exhibited reducedproduction of polar glycolipids such as phosphatidyl-inositolmannosides (PIMs) and phosphatidylglycerol (PG) com-pared with planktonic cells (Figure 5(b))
33 In Silico Identification of GPL Biosynthesis Genes inM colombiense CECT 3035 The genes involved in GPLbiosynthesis in M colombiense are currently unknown
6 BioMed Research International
(a) (b) (c) (d)
(e) (f) (g) (h)
Figure 4 Drop-collapsing test (a) Water (no surfactant) (b) 1 SDS (surfactant) (c) P aeruginosa (d) M smegmatis mc2155 (e) Mcolombiense 19B (f) M colombiense CECT 3035 (g) M colombiense 57B and (h) M avium 104 Drop spreading over the oily surface aftersamples incubation indicates the presence of biosurfactant substances in the aqueous sampleThis assay was conducted in three independentexperiments each performed in duplicate
Based on the GPL biosynthetic pathway reported for Mavium 104 [25] 30 open reading frames (ORF) encodingpeptide synthetase fatty acid synthases (FAS) polyketidesynthases (PKs) acyltransferase (PapA) glycosyltransferasescarbohydrate synthetases methyl and acyl transferases gly-colipid transporters potential biosynthesis regulators andproteins with unknown functions [25 26] were searchedwithin the recently reported nucleotide sequence of Mcolombiense CECT 3035 [36] This in silico analysis (seeTable S1 of the Supplementary Material available online athttpdxdoiorg1011552015419549) showed that all enzy-matic activities searched were encoded by different ORFsin M colombiense CECT 3035 with an identity rangingbetween 77 and 96 The ORFs in the genome of the Mcolombiense counterpart were located in the contigs 0000100002A 00003A and 00007 of the genome sequence of theCECT 3035 strain (Figure 6) Among the 30 genes of the GPLsynthetic pathway inM avium 104 7 genes (mtfA gtfB gtfDdghA fadD23 pe and gap-like) did not have orthologoussequences inM colombiense CECT 3035 (Table S1)
34 Differential Transcription of the GPL Biosynthesis Genesin M colombiense Displaying Smooth and Rough ColonyMorphologies RNAwas isolated fromM colombienseCECT3035 19B and 57B strains cultured under planktonic andmotile conditions to quantify the transcription levels ofthe pstA gtfA rtfA mtfB mtfC mtfD tmtpC tmtpA andtmtpB genes predicted to encode key enzymes of the GPLbiosynthetic pathway (Table S1) For relative quantificationM colombiense CECT 3035 was grown to the exponentialphase under planktonic conditions and used as the referencepattern for two reasons (1) CECT3035 is theM colombiensegenome sequence strain [36] and consistent with the TLCanalysis (2) planktonic cells displayed the lowest GPL pro-duction
With regard to planktonic cells the rate ofM colombiense19B gene transcription was consistently higher than thatfor the M colombiense 57B genes Conversely the roughvariant (57B) displayed a dramatic decrease in the rate oftranscription for the 9 selected genes particularly thoseencoding peptide synthetase pstA glycosyltransferase gtfA
BioMed Research International 7
1 2 3 41 2 3 4
GPLs
PG
PIMs
(a) (b)
Figure 5 TLC profile of the GPLs in Mycobacterium colombiense strains Crude lipid extracts of (1) M avium 104 (2) M colombiense19B (3) M colombiense CECT 3035 and (4) M colombiense 57B Cells cultivated (a) under planktonic and (b) motile conditions weredeveloped using chloroform-methanol-water (65 25 4 vvv) GPLs glycopeptidolipids PIMs phosphatidyl-inositol mannosides and PGphosphatidylglycerol
the methyltransferases mtfB and mtfC and the lipid trans-porters tmtpA tmtpB and tmtpC (Figure 7(a)) InterestinglyrtfA (rhamnosyltransferase) in the 19B strain (smooth colonymorphology) was the only gene among the strains culturedunder planktonic conditions that showed an increased tran-scription rate (221-fold higher) compared with the controlcells (planktonicM colombiense CECT 3035 at the exponen-tial phase of growth)
Under motile growth conditions M colombiense 19Bshowed the highest transcription rate for all selected genescompared with the CECT 3035 and 57B strains (Figure 7(b))In contrast the transcription rates for all genes in the57B strain (rough variant) except tmtpA and tmtpC (lipidtransporters) were reduced In motile CECT 3035 cellswhereas rtfA mtfB (methyl transfers) and tmtpA showedincreased transcription the pstA gftA mtfC mtfD tmtpBand tmtpC genes exhibited reduced transcription comparedwith control cells
4 Discussion
In the present study TLC experiments were able to show thatM colombiense contains glycolipids with chromatographicbehaviour similar to the GPLs of M avium 104 which havebeen previously identified and completely characterised [23]in addition the colony morphology was associated withthe GPL profile of M colombiense strains Thus the TLCanalysis revealed that (1) strains displaying smooth colonymorphology (19B and CECT3035) produce a higher amountof variable GPLs compared with the rough natural variant(57B) strain and (2) the GPL production was augmentedin M colombiense cells cultivated under the motile growthconditions As expected the TLC analyses did not provide
detailed information about GPL structure however struc-tural determinationwas not the objective of the present study
Themedia composition influences the glycolipid contentof the mycobacterial cell wall The MAC strains growing onsolid medium exhibit a significant rate of transition fromsmooth transparent to smooth opaque and from smooth torough morphologies [18 19 38] We compared planktonicand motile M colombiense cells grown in the same culturemedium (7H9-ADC or 7H9-ADC-agarose) to avoid differ-ences in lipid content produced through alterations in themedia composition The results showed that the low GPLcontent observed in the rough 57B variant is less frequentamong M colombiense strains therefore M colombienseshows a preference for augmented GPL production andsmooth colony morphology Interestingly planktonic Mcolombiense cells displayed increased PIM and PG pro-duction and diminished GPL content compared with cellscultivated under motile conditions suggesting a potentialcompensatory mechanism in cell wall lipid synthesis thatcompensates for the low GPL content
The abundance of GPLs in the outermost portion ofthe cell envelope could be associated with the motility ofM colombiense strains on hydrophilic surfaces Recht andothers [22] proposed that GPLs are linked through thehydrophilic head to the cellular capsule and the hydrophobicfatty acid chain is exposed to the bacterial surface therebyreducing interactions with hydrophilic agarose surfaces andfacilitating the spread of cells on solid medium [21 22]Thus the augmented GPL production of motile 19B andCECT 3035 cells compared with that of the rough variant57B is consistent with the sliding motility on the hydrophilicagarose surface observed for these strains It has also beensuggested that bacterial sliding motility might be favoured
8 BioMed Research International
33kb 2kb9kb51kb
Contig 00001
600kb
87kb 245kb
488kb 36kb320kb
Contig 00002A
Contig 00003A
584kb 452kb
mmpL10
326kb
Contig 00007
fadE
15
atf
rmlB
ecf
rv1174
rmlA
mmpL
4a
mmpL
4b
mmpS4
mbtH
gap rv0926
sap
pks papA3
gtfA
mtfB
mtfC
mtfD
rtfA
pstA
pstB
(a)
ecf
fadE
5
rv1174
atf
rmlB
rmlA
mmpL
4a
mmpL
4b
mmpS
4
mbtH
gap
237kb
rv0926
7kb90kb
530kb
300kb
738kb
mmpL10
sap
Saccharides biosynthesis transfer and modification
Fatty acyl chain biosynthesis modification and transfer
Members of the mmpSL family
Peptide synthesis Unknown
Possible regulatory genes
GPL export to the cell surface
pks pe fadD23 gap-likepapA3
mtfB
gtfA
mtfC
mtfD
dhgA
rtfA
pstA
pstB
mtfA
gtfB
gtfD
(b)
Figure 6Model showing the gene cluster proposed for the GPL biosynthesis ofMycobacterium colombienseCECT 3035 (a)The location andidentity of theM colombiense genes were established using the previously described biosynthetic pathway for GPL biosynthesis annotated intheM avium 104 genomic sequence (b)
0001
001
01
1
10
Rela
tive g
ene t
rans
crip
tion
M colombiense 57BM colombiense 19B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(a)
01
1
10
100
Rela
tive g
ene t
rans
crip
tion
M colombiense CECT 3035
M colombiense 19B
M colombiense 57B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(b)
Figure 7 In vitro transcription of Mycobacterium colombiense genes involved in GPL biosynthesis Relative quantification expressed as theratio of gene transcription in (a) M colombiense cells grown under planktonic conditionstranscription ofM colombiense CECT 3035 cellsgrown under planktonic conditions and (b)M colombiense cells grown on the motility mediumtranscription ofM colombiense CECT 3035grown under planktonic conditions The presented data have statistically significant differences compared with the values obtained fromMcolombiense CECT 3035 cells (119875 lt 005)
BioMed Research International 9
through the secretion of surfactant substances from cells[21 22] The results of the drop-collapsing test showed thatbiosurfactants were likely not secreted from M colombiensecells suggesting that the motility of M colombiense couldbe favoured through hydrophobic molecules such as GPLsbound to the outermost portion of cells however we cannotcompletely rule out the possibility that some of the surfactantsubstances secreted from M colombiense remained adheredto the mycobacterial cell surface thereby influencing cellmotility
GPLs on the outermost portions of the M colombiensecell envelope generate a more hydrophobic cell surfacethat facilitates initial interactions with hydrophobic surfacesand increase biofilm formation on polystyrene wells Inaddition the increased GPL production in 19B and CECT3035 cells is consistent with the augmented biofilm forma-tion observed for M colombiense strains displaying smoothcolony morphology Moreover increased biofilm formationon hydrophobic surfaces and enhanced sliding over motilityplates for 19B cells compared with the CECT 3035 strainwhich also displays smooth colony morphology likely sug-gest possible differences in GPL structure betweenM colom-biense strains Differences inGPL production between strainscould influence the M colombiense ability to form biofilmson polyvinylchloride (PVC) pipes which would facilitatethe dissemination of these bacteria in natural environmentssuch as in-hospital spaces thereby increasing the chance ofinfection in immunosuppressed patients [23 39 40]
Based on the GPL biosynthetic pathway previouslydescribed for the M avium 104 strain [25] we are currentlyconstructing a potential gene cluster for GPL biosynthesis inM colombiense which will be finished when the nucleotidesequence of the CECT 3035 is completely assembled RT-qPCR experiments showed that the planktonic and motilecells of the rough 57B strain showed decreased transcriptionof the 9 selected genes leading to the low production ofGPLs in this rough variant comparedwith theM colombiensestrains displaying smooth colony morphology Interestinglyplanktonic 19B cells exhibited lower expression of most of theselected genes compared with planktonic CECT 3035 cellsThis behaviour results in the reduced production of GPLs inplanktonic 19B cells compared with the CECT 3035 strainconsistent with the results of the TLC experiments
It has been previously shown that an rtfA mutation inthe M avium serovar-2-specific strain resulted in the lossof serovar-specific GPLs thereby diminishing the variabilityof these glycolipids in the cell envelope [41 42] Thus itis possible that the enzymatic activity of the RtfA proteincould be relevant for the increased glycosylation andor aug-mented production of ssGPLs in strains displaying smoothcolony morphology compared with the rough variant 57BIn strains displaying smooth colony morphology the genesencoding rhamnosyl andmethyl transferases (rtfA andmtfB)are overtranscribed suggesting the increased production ofnsGPLs precursors for ssGPL biosynthesis [24ndash26] therebyincreasing the production of GPLs The diminished tran-scription of pstA gftA mtfC mtfD tmtpBand tmtpC inmotile CECT 3035 compared with control cells (planktonicCECT 3035) is intriguing however this behaviour also
suggests the relevance of rhamnosyl transferases particularlyRtfA in GPL biosynthesis in M colombiense Neverthelessthe actual role for RtfA in M colombiense strains should beconfirmed using rtfA-null mutants
We did not identify 7 of the 30 known GPL cluster genesof M avium 104 in the genome sequence of M colombienseCECT 3035 This interesting observation suggests that theGPLs between these two closely related MAC species couldhave different structural characteristics However furtherexperiments using mass spectrometry and NMR are neces-sary to evaluate potential differences in the GPL structureamongM colombiense strains
5 Conclusions
In conclusion the results of the present study show that Mcolombiense strains displaying smooth morphology exhibitincreased biofilm formation on hydrophobic polystyrenesurfaces and enhanced sliding over motility plates Bioin-formatics analyses indicate that the gene cluster establishedfor GPL formation modification and translocation is intactstrongly suggesting that GPLs are putatively present in Mcolombiense In addition motile culture conditions activatethe transcription of the genes implicated in the key enzymaticactivities of GPL biosynthesis
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
Thisworkwas supported through funding from the ldquoDivisionde Investigacion Bogota (DIB)rdquo-Vicerrectorıa de Investi-gacion Universidad Nacional de Colombia (Grants 1606014837 and 14337) Milena Maya-Hoyos received a fellowshipfrom the Academic Direction Universidad Nacional deColombia and the ldquoJovenes Investigadores e InnovadoresrdquoProgram Colciencias Colombia This research was alsosupported in part through the Intramural Research Programof the National Institutes of Health National Library ofMedicine and National Center for Biotechnology Informa-tion (NCBI)
References
[1] S K Field D Fisher and R L Cowie ldquoMycobacterium aviumcomplex pulmonary disease in patient without HIV infectionrdquoChest vol 126 no 2 pp 566ndash581 2004
[2] M J M Vaerewijck G Huys J C Palomino J Swings and FPortaels ldquoMycobacteria in drinking water distribution systemsecology and significance for human healthrdquo FEMSMicrobiologyReviews vol 29 no 5 pp 911ndash934 2005
[3] E Hagiwara S Komatsu R Nishihira T Shinohara T Babaand T Ogura ldquoClinical characteristics and prevalence of pneu-mothorax in patients with pulmonary Mycobacterium aviumcomplex diseaserdquo Journal of Infection and Chemotherapy vol19 no 4 pp 588ndash592 2013
10 BioMed Research International
[4] M I Murcia E Tortoli M C Menendez E Palenque andM J Garcia ldquoMycobacterium colombiense sp nov a novelmember of theMycobacterium avium complex and descriptionof MAC-X as a new ITS genetic variantrdquo International Journalof Systematic and Evolutionary Microbiology vol 56 no 9 pp2049ndash2054 2006
[5] E Tortoli L Rindi M J Garcia et al ldquoProposal to elevatethe genetic variant MAC-A included in the Mycobacteriumavium complex to species rank as Mycobacterium chimaerasp novrdquo International Journal of Systematic and EvolutionaryMicrobiology vol 54 no 4 pp 1277ndash1285 2004
[6] I B Salah C Cayrou D Raoult and M Drancourt ldquoMycobac-terium marseillense sp nov Mycobacterium timonense sp novand Mycobacterium bouchedurhonense sp novmembers ofthe Mycobacterium avium complexrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 11 pp2803ndash2808 2009
[7] J van Ingen M J Boeree K Kosters et al ldquoProposal toelevate Mycobacterium avium complex ITS sequevar MAC-Qto Mycobacterium vulneris sp novrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 9 pp2277ndash2282 2009
[8] D Bang T Herlin M Stegger et al ldquoMycobacterium arosiensesp nov a slowly growing scotochromogenic species causingosteomyelitis in an immunocompromised childrdquo InternationalJournal of Systematic and Evolutionary Microbiology vol 58part 10 pp 2398ndash2402 2008
[9] C Cayrou C Turenne M A Behr and M Drancourt ldquoGeno-typing of Mycobacterium avium complex organisms usingmultispacer sequence typingrdquoMicrobiology vol 156 part 3 pp687ndash694 2010
[10] S C Smole F McAleese J Ngampasutadol C F von Reynand R D Arbeit ldquoClinical and epidemiological correlates ofgenotypes within theMycobacterium avium complex defined byrestriction and sequence analysis of hsp65rdquo Journal of ClinicalMicrobiology vol 40 no 9 pp 3374ndash3380 2002
[11] AMuwonge C Kankya T B Johansen et al ldquoNon-tuberculousmycobacteria isolated from slaughter pigs in Mubende districtUgandardquo BMC Veterinary Research vol 8 article 52 2012
[12] J O Falkinham III ldquoSurrounded by mycobacteria nontuber-culous mycobacteria in the human environmentrdquo Journal ofApplied Microbiology vol 107 no 2 pp 356ndash367 2009
[13] K Vuorenmaa I B Salah V Barlogis H Chambost and MDrancourt ldquoMycobacterium colombiense and pseudotubercu-lous lymphadenopathyrdquo Emerging Infectious Diseases vol 15no 4 pp 618ndash620 2009
[14] O Esparcia F Navarro M Quer and P Coll ldquoLymphadenopa-thy caused by Mycobacterium colombienserdquo Journal of ClinicalMicrobiology vol 46 no 5 pp 1885ndash1887 2008
[15] S Cohen-Bacrie M David N Stremler J-C Dubus J-MRolain and M Drancourt ldquoMycobacterium chimaera pul-monary infection complicating cystic fibrosis a case reportrdquoJournal of Medical Case Reports vol 5 no 1 article 473 2011
[16] S Poulin C Corbeil M Nguyen et al ldquoFatal Mycobac-terium colombiensecytomegalovirus coinfection associatedwith acquired immunodeficiency due to autoantibodies againstinterferon gamma a case reportrdquo BMC Infectious Diseases vol13 no 1 article 24 2013
[17] J Leguizamon J Hernandez M-I Murcia and C-Y SotoldquoIdentification of potential biomarkers to distinguishMycobac-terium colombiense from other mycobacterial speciesrdquoMolecu-lar and Cellular Probes vol 27 no 1 pp 46ndash52 2013
[18] D Chatterjee and K H Khoo ldquoThe surface glycopeptidolipidsof mycobacteria structures and biological propertiesrdquo Cellularand Molecular Life Sciences vol 58 no 14 pp 2018ndash2042 2001
[19] J B Torrelles D Ellis T Osborne et al ldquoCharacterizationof virulence colony morphotype and the glycopeptidolipid ofMycobacterium avium strain 104rdquo Tuberculosis vol 82 no 6pp 293ndash300 2002
[20] M F Thorel and H L David ldquoSpecific surface antigens of SmTvariants ofMycobacterium aviumrdquo Infection and Immunity vol43 no 1 pp 438ndash439 1984
[21] A Martınez S Torello and R Kolter ldquoSliding motility inmycobacteriardquo Journal of Bacteriology vol 181 no 23 pp 7331ndash7338 1999
[22] J Recht AMartınez S Torello and R Kolter ldquoGenetic analysisof sliding motility in Mycobacterium smegmatisrdquo Journal ofBacteriology vol 182 no 15 pp 4348ndash4351 2000
[23] G Carter M Wu D C Drummond and L E BermudezldquoCharacterization of biofilm formation by clinical isolates ofMycobacterium aviumrdquo Journal of Medical Microbiology vol 52no 9 pp 747ndash752 2003
[24] J S Schorey and L Sweet ldquoThe mycobacterial glycopepti-dolipids structure function and their role in pathogenesisrdquoGlycobiology vol 18 no 11 pp 832ndash841 2008
[25] L Pang X Tian W Pan and J Xie ldquoStructure and function ofmycobacterium glycopeptidolipids from comparative genomicsperspectiverdquo Journal of Cellular Biochemistry vol 114 no 8 pp1705ndash1713 2013
[26] R Mukherjee and D Chatterji ldquoGlycopeptidolipids immuno-modulators in greasy mycobacterial cell enveloperdquo IUBMB Lifevol 64 no 3 pp 215ndash225 2012
[27] K L Horan R Freeman K Weigel et al ldquoIsolation ofthe genome sequence strain Mycobacterium avium 104 frommultiple patients over a 17-year periodrdquo Journal of ClinicalMicrobiology vol 44 no 3 pp 783ndash789 2006
[28] S B Snapper R E Melton S Mustafa T Kieser and W RJacobs Jr ldquoIsolation and characterization of efficient plasmidtransformation mutants of Mycobacterium smegmatisrdquo Molec-ular Microbiology vol 4 no 11 pp 1911ndash1919 1990
[29] M Radji R A Agustama B Elya and C R TjampakasarildquoAntimicrobial activity of green tea extract against isolatesof methicillin-resistant Staphylococcus aureus and multi-drugresistant Pseudomonas aeruginosardquo Asian Pacific Journal ofTropical Biomedicine vol 3 no 8 pp 663ndash667 2013
[30] P Helden T Victor R Warren and E Helden ldquoIsolationof DNA from Mycobacterium tuberculosisrdquo in Mycobacteriumtuberculosis Protocols Humana Press Totowa NJ USA 2001
[31] J C van Kessel L J Marinelli and G F Hatfull ldquoRecom-bineering mycobacteria and their phagesrdquo Nature ReviewsMicrobiology vol 6 no 11 pp 851ndash857 2008
[32] N M Parrish C G Ko J D Dick P B Jones and J LE Ellingson ldquoGrowth Congo Red agar colony morphotypesand antibiotic susceptibility testing of Mycobacterium aviumsubspecies paratuberculosisrdquo Clin Med Res vol 2 no 2 pp107ndash114 2004
[33] T Tugrul and E Cansunar ldquoDetecting surfactant-producingmicroorganisms by the drop-collapse testrdquo World Journal ofMicrobiology and Biotechnology vol 21 no 6-7 pp 851ndash8532005
[34] D K Jain D L Collins-Thompson H Lee and J T TrevorsldquoA drop-collapsing test for screening surfactant-producingmicroorganismsrdquo Journal ofMicrobiologicalMethods vol 13 no4 pp 271ndash279 1991
BioMed Research International 11
[35] M Munoz M-A Laneelle M Luquin et al ldquoOccurrence ofan antigenic triacyl trehalose in clinical isolates and referencestrains of Mycobacterium tuberculosisrdquo FEMS MicrobiologyLetters vol 157 no 2 pp 251ndash259 1997
[36] M Gonzalez-Perez M I Murcia D Landsman I K Jordanand L Marino-Ramırez ldquoGenome sequence of the Mycobac-terium colombiense type strain CECT 3035rdquo Journal of Bacte-riology vol 193 no 20 pp 5866ndash5867 2011
[37] J C Betts P T Lukey L C Robb R A McAdam andK Duncan ldquoEvaluation of a nutrient starvation model ofMycobacterium tuberculosis persistence by gene and proteinexpression profilingrdquoMolecular Microbiology vol 43 no 3 pp717ndash731 2002
[38] W W Barrow and P J Brennan ldquoIsolation in high frequencyof rough variants of Mycobacterium intracellulare lacking C-mycoside glycopeptidolipid antigensrdquo Journal of Bacteriologyvol 150 no 1 pp 381ndash384 1982
[39] W Costerton R Veeh M Shirtliff M Pasmore C Post andG Ehrlich ldquoThe application of biofilm science to the studyand control of chronic bacterial infectionsrdquo Journal of ClinicalInvestigation vol 112 no 10 pp 1466ndash1477 2003
[40] J Recht and R Kolter ldquoGlycopeptidolipid acetylation affectssliding motility and biofilm formation inMycobacterium smeg-matisrdquo Journal of Bacteriology vol 183 no 19 pp 5718ndash57242001
[41] T M Eckstein F S Silbaq D Chatterjee N J Kelly P JBrennan and J T Belisle ldquoIdentification and recombinantexpression of a Mycobacterium avium rhamnosyltransferasegene (rtfA) involved in glycopeptidolipid biosynthesisrdquo Journalof Bacteriology vol 180 no 21 pp 5567ndash5573 1998
[42] J N Maslow V R Irani S-H Lee T M Eckstein J MInamine and J T Belisle ldquoBiosynthetic specificity of therhamnosyltransferase gene of Mycobacterium avium serovar 2as determined by allelic exchange mutagenesisrdquo Microbiologyvol 149 no 11 pp 3193ndash3202 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 BioMed Research International
(a) (b)
(c) (d)
Figure 1 Colonymorphology ofMycobacterium colombiense strainsMycobacterial strains were grown on 7H10-OADC plates supplementedwith Congo Red and incubated at 37∘C for 4-5 weeks (a)M colombiense 19B (b) CECT 3035 (c) 57B and (d)M avium 104
were scraped and resuspended in diethylpyrocarbonate-(DEPC-) treated water Motile and planktonic mycobacteriawere centrifuged washed with DEPC-treated water andresuspended in TRIzol (Invitrogen USA) The cells werelysed using a Mini-Bead Beater 16 (BioSpec OK USA) andglass beads (01mm) Total RNA was isolated as previouslydescribed [37] and quantified using aNanoDrop 2000c Spec-trophotometer (Thermo Scientific MA USA) After DNaseI treatment the RNA quality was evaluated through agarosegel electrophoresis and spectrophotometry (OD
260OD280
)cDNA libraries were constructed from 1 120583g of RNA using theRevertAid First cDNA Synthesis Kit (Fermentas Lithuania)For reverse transcription the samples were incubated at 37∘Cfor 30min 42∘C for 5 h 72∘C for 1min and 4∘C for 10min
The primers used for the RT-qPCR analyses are listed inTable 1The RT-qPCR reactions were set up in triplicate usingthe Express SYBR GreenER Universal qPCR SuperMix Kit(Invitrogen NY USA) and a CFX-96 thermocycler (BioradPA USA) under conditions including a denaturation stepfor 5min at 95∘C followed by 39 cycles of 95∘C for 10 sec
58∘C for 10 sec and 72∘C for 15 sec Primers or cDNA wereomitted in the negative controls The gene transcriptionwas normalised to the mean value of 16S rRNA (rrs) geneexpressionThe transcription profile was obtained from threeindependent experiments performed in duplicate For eachexperiment the differences among experimental data werecompared using Studentrsquos t-test and119875 lt 005was consideredsignificantly different
3 Results
31M colombiense IsMotile and Forms Biofilms onHydropho-bic Polystyrene Surfaces The M colombiense strains usedin this study including the genome sequence strain CECT3035 presented smooth opaque and brilliant colonies on7H10-OADC plates supplemented with Congo Red exceptstrain 57B which displayed a rough and wrinkled phenotype(Figure 1) Similar results were obtained using medium lack-ing oleic acid (7H9-ADC-035 agarose data not shown)Other M colombiense strains not included in the present
BioMed Research International 5
7
17
27
37
47
57
0 10 20 30 40 50 60 70
Dia
met
er (m
m)
Day
M colombiense 19BM avium
M colombiense CECT 3035
M colombiense 57B
Figure 2 Motility rates for Mycobacterium colombiense strainsMycobacterial strains were cultivated on 7H9-ADC-035 agaroseand incubated at 37∘C for 4-5 weeks The colony growth haloswere measured daily to determine the motility rate Each reportedvalue represents the mean of the colony growth halos from threeindependent experiments The presented data have statisticallysignificant differences compared with the values obtained from Mavium 104 (119875 lt 005)
study (6B 7B 9B and 16B) also displayed a smooth colonymorphology (data not shown) Both smooth and rough bac-terial phenotypes were stable after repetitive culture on solidmedia for the duration of the study (3 years) The smoothM colombiense strains developed yellow pigmentation withage specifically at the stationary phase in all culture mediahowever the rough variant (57B) remained beige in colourunder the same experimental conditions (Figure 1)
Because alterations in the phenotypic characteristicsassociated with colony morphology have been observedin other MAC species motility and biofilm formationwere compared between smooth and rough M colombiensestrains We observed that the M colombiense 19B andCECT3035 strains (smooth colony morphology) showedincreased spreading (4094 plusmn 005 120583mday and 2972 plusmn005 120583mday resp) on hydrophilic agarose comparedwith thenatural rough variant 57B which showed impaired spreading(Figure 2)
Crystal violet staining was used to quantify the degree ofM colombiense biofilm formation onpolystyreneThehighestOD570
values for all M colombiense strains were obtainedusing an inoculum of sim27 times 105 CFUmL In general thestrains displaying smooth morphology showed increasedbiofilm formation on hydrophobic surfaces while therough variant 57B showed reduced adhesion to polystyrene(Figure 3)
As biosurfactants could potentially influence cell motilitythe ldquodrop-collapsingrdquo method was used to detect biosurfac-tant secretion in M colombiense strains [22] As shown inFigure 4 drops of 1 SDS and P aeruginosa aqueous extract(positive controls) spread over the oily surface after the sam-ples were incubated indicating the presence of biosurfactantsubstances In contrast drops of distilled waterM smegmatis(negative control) and M colombiense aqueous extracts didnot collapse on the oily surface but rather appeared as firm
0
005
01
015
02
025
OD570
Mavium
104
Mcolom
biense
CECT
3035
Mcolom
biense19
B
Mcolom
biense57
B
Figure 3 Capacity of Mycobacterium colombiense strains to formbiofilms The OD
570nm represents the proportion of mycobacteria
added to the microtiter wells that reacted with the crystal violetsolutionThebars represent the SD calculated from two independentexperiments each performed in triplicate
drops suggesting the absence of biosurfactants in the culturesupernatants
32 M colombiense Displaying a Smooth Colony Morphol-ogy Contains Glycolipids with Thin-Layer ChromatographicBehaviour Similar to That of the GPLs of M avium TLCanalysis of noncovalently attached lipids extracted frommycobacteria cultured under planktonic and motile condi-tions showed that M colombiense strains contain glycolipidswith chromatographic behaviour similar to that of theGPLsofM avium [23] Regarding planktonic cells (Figure 5(a)) thestrains with smooth colony morphology (19B and CECT3035) showed multiple lipid spots migrating in the region ofcontrol GPLs (M avium 104) however the spots observed inthe CECT 3035 lipid extract were more intense than those inthe 19B strain The natural rough variant (57B) showed onlyone spot that migrated in the same GPLs region
Under motile conditions (Figure 5(b)) the smoothcolony morphology strains 19B and CECT 3035 showed aGPL profile similar to that obtained for cells cultured underplanktonic conditions nevertheless the counterpart spotswere more intense than those observed in planktonic cellsparticularly for the 19B strain Nevertheless the unique spotobserved for motile 57B cells (rough variant) were moreintense than that observed for planktonic 57B cells Inter-estingly all motile M colombiense cells exhibited reducedproduction of polar glycolipids such as phosphatidyl-inositolmannosides (PIMs) and phosphatidylglycerol (PG) com-pared with planktonic cells (Figure 5(b))
33 In Silico Identification of GPL Biosynthesis Genes inM colombiense CECT 3035 The genes involved in GPLbiosynthesis in M colombiense are currently unknown
6 BioMed Research International
(a) (b) (c) (d)
(e) (f) (g) (h)
Figure 4 Drop-collapsing test (a) Water (no surfactant) (b) 1 SDS (surfactant) (c) P aeruginosa (d) M smegmatis mc2155 (e) Mcolombiense 19B (f) M colombiense CECT 3035 (g) M colombiense 57B and (h) M avium 104 Drop spreading over the oily surface aftersamples incubation indicates the presence of biosurfactant substances in the aqueous sampleThis assay was conducted in three independentexperiments each performed in duplicate
Based on the GPL biosynthetic pathway reported for Mavium 104 [25] 30 open reading frames (ORF) encodingpeptide synthetase fatty acid synthases (FAS) polyketidesynthases (PKs) acyltransferase (PapA) glycosyltransferasescarbohydrate synthetases methyl and acyl transferases gly-colipid transporters potential biosynthesis regulators andproteins with unknown functions [25 26] were searchedwithin the recently reported nucleotide sequence of Mcolombiense CECT 3035 [36] This in silico analysis (seeTable S1 of the Supplementary Material available online athttpdxdoiorg1011552015419549) showed that all enzy-matic activities searched were encoded by different ORFsin M colombiense CECT 3035 with an identity rangingbetween 77 and 96 The ORFs in the genome of the Mcolombiense counterpart were located in the contigs 0000100002A 00003A and 00007 of the genome sequence of theCECT 3035 strain (Figure 6) Among the 30 genes of the GPLsynthetic pathway inM avium 104 7 genes (mtfA gtfB gtfDdghA fadD23 pe and gap-like) did not have orthologoussequences inM colombiense CECT 3035 (Table S1)
34 Differential Transcription of the GPL Biosynthesis Genesin M colombiense Displaying Smooth and Rough ColonyMorphologies RNAwas isolated fromM colombienseCECT3035 19B and 57B strains cultured under planktonic andmotile conditions to quantify the transcription levels ofthe pstA gtfA rtfA mtfB mtfC mtfD tmtpC tmtpA andtmtpB genes predicted to encode key enzymes of the GPLbiosynthetic pathway (Table S1) For relative quantificationM colombiense CECT 3035 was grown to the exponentialphase under planktonic conditions and used as the referencepattern for two reasons (1) CECT3035 is theM colombiensegenome sequence strain [36] and consistent with the TLCanalysis (2) planktonic cells displayed the lowest GPL pro-duction
With regard to planktonic cells the rate ofM colombiense19B gene transcription was consistently higher than thatfor the M colombiense 57B genes Conversely the roughvariant (57B) displayed a dramatic decrease in the rate oftranscription for the 9 selected genes particularly thoseencoding peptide synthetase pstA glycosyltransferase gtfA
BioMed Research International 7
1 2 3 41 2 3 4
GPLs
PG
PIMs
(a) (b)
Figure 5 TLC profile of the GPLs in Mycobacterium colombiense strains Crude lipid extracts of (1) M avium 104 (2) M colombiense19B (3) M colombiense CECT 3035 and (4) M colombiense 57B Cells cultivated (a) under planktonic and (b) motile conditions weredeveloped using chloroform-methanol-water (65 25 4 vvv) GPLs glycopeptidolipids PIMs phosphatidyl-inositol mannosides and PGphosphatidylglycerol
the methyltransferases mtfB and mtfC and the lipid trans-porters tmtpA tmtpB and tmtpC (Figure 7(a)) InterestinglyrtfA (rhamnosyltransferase) in the 19B strain (smooth colonymorphology) was the only gene among the strains culturedunder planktonic conditions that showed an increased tran-scription rate (221-fold higher) compared with the controlcells (planktonicM colombiense CECT 3035 at the exponen-tial phase of growth)
Under motile growth conditions M colombiense 19Bshowed the highest transcription rate for all selected genescompared with the CECT 3035 and 57B strains (Figure 7(b))In contrast the transcription rates for all genes in the57B strain (rough variant) except tmtpA and tmtpC (lipidtransporters) were reduced In motile CECT 3035 cellswhereas rtfA mtfB (methyl transfers) and tmtpA showedincreased transcription the pstA gftA mtfC mtfD tmtpBand tmtpC genes exhibited reduced transcription comparedwith control cells
4 Discussion
In the present study TLC experiments were able to show thatM colombiense contains glycolipids with chromatographicbehaviour similar to the GPLs of M avium 104 which havebeen previously identified and completely characterised [23]in addition the colony morphology was associated withthe GPL profile of M colombiense strains Thus the TLCanalysis revealed that (1) strains displaying smooth colonymorphology (19B and CECT3035) produce a higher amountof variable GPLs compared with the rough natural variant(57B) strain and (2) the GPL production was augmentedin M colombiense cells cultivated under the motile growthconditions As expected the TLC analyses did not provide
detailed information about GPL structure however struc-tural determinationwas not the objective of the present study
Themedia composition influences the glycolipid contentof the mycobacterial cell wall The MAC strains growing onsolid medium exhibit a significant rate of transition fromsmooth transparent to smooth opaque and from smooth torough morphologies [18 19 38] We compared planktonicand motile M colombiense cells grown in the same culturemedium (7H9-ADC or 7H9-ADC-agarose) to avoid differ-ences in lipid content produced through alterations in themedia composition The results showed that the low GPLcontent observed in the rough 57B variant is less frequentamong M colombiense strains therefore M colombienseshows a preference for augmented GPL production andsmooth colony morphology Interestingly planktonic Mcolombiense cells displayed increased PIM and PG pro-duction and diminished GPL content compared with cellscultivated under motile conditions suggesting a potentialcompensatory mechanism in cell wall lipid synthesis thatcompensates for the low GPL content
The abundance of GPLs in the outermost portion ofthe cell envelope could be associated with the motility ofM colombiense strains on hydrophilic surfaces Recht andothers [22] proposed that GPLs are linked through thehydrophilic head to the cellular capsule and the hydrophobicfatty acid chain is exposed to the bacterial surface therebyreducing interactions with hydrophilic agarose surfaces andfacilitating the spread of cells on solid medium [21 22]Thus the augmented GPL production of motile 19B andCECT 3035 cells compared with that of the rough variant57B is consistent with the sliding motility on the hydrophilicagarose surface observed for these strains It has also beensuggested that bacterial sliding motility might be favoured
8 BioMed Research International
33kb 2kb9kb51kb
Contig 00001
600kb
87kb 245kb
488kb 36kb320kb
Contig 00002A
Contig 00003A
584kb 452kb
mmpL10
326kb
Contig 00007
fadE
15
atf
rmlB
ecf
rv1174
rmlA
mmpL
4a
mmpL
4b
mmpS4
mbtH
gap rv0926
sap
pks papA3
gtfA
mtfB
mtfC
mtfD
rtfA
pstA
pstB
(a)
ecf
fadE
5
rv1174
atf
rmlB
rmlA
mmpL
4a
mmpL
4b
mmpS
4
mbtH
gap
237kb
rv0926
7kb90kb
530kb
300kb
738kb
mmpL10
sap
Saccharides biosynthesis transfer and modification
Fatty acyl chain biosynthesis modification and transfer
Members of the mmpSL family
Peptide synthesis Unknown
Possible regulatory genes
GPL export to the cell surface
pks pe fadD23 gap-likepapA3
mtfB
gtfA
mtfC
mtfD
dhgA
rtfA
pstA
pstB
mtfA
gtfB
gtfD
(b)
Figure 6Model showing the gene cluster proposed for the GPL biosynthesis ofMycobacterium colombienseCECT 3035 (a)The location andidentity of theM colombiense genes were established using the previously described biosynthetic pathway for GPL biosynthesis annotated intheM avium 104 genomic sequence (b)
0001
001
01
1
10
Rela
tive g
ene t
rans
crip
tion
M colombiense 57BM colombiense 19B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(a)
01
1
10
100
Rela
tive g
ene t
rans
crip
tion
M colombiense CECT 3035
M colombiense 19B
M colombiense 57B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(b)
Figure 7 In vitro transcription of Mycobacterium colombiense genes involved in GPL biosynthesis Relative quantification expressed as theratio of gene transcription in (a) M colombiense cells grown under planktonic conditionstranscription ofM colombiense CECT 3035 cellsgrown under planktonic conditions and (b)M colombiense cells grown on the motility mediumtranscription ofM colombiense CECT 3035grown under planktonic conditions The presented data have statistically significant differences compared with the values obtained fromMcolombiense CECT 3035 cells (119875 lt 005)
BioMed Research International 9
through the secretion of surfactant substances from cells[21 22] The results of the drop-collapsing test showed thatbiosurfactants were likely not secreted from M colombiensecells suggesting that the motility of M colombiense couldbe favoured through hydrophobic molecules such as GPLsbound to the outermost portion of cells however we cannotcompletely rule out the possibility that some of the surfactantsubstances secreted from M colombiense remained adheredto the mycobacterial cell surface thereby influencing cellmotility
GPLs on the outermost portions of the M colombiensecell envelope generate a more hydrophobic cell surfacethat facilitates initial interactions with hydrophobic surfacesand increase biofilm formation on polystyrene wells Inaddition the increased GPL production in 19B and CECT3035 cells is consistent with the augmented biofilm forma-tion observed for M colombiense strains displaying smoothcolony morphology Moreover increased biofilm formationon hydrophobic surfaces and enhanced sliding over motilityplates for 19B cells compared with the CECT 3035 strainwhich also displays smooth colony morphology likely sug-gest possible differences in GPL structure betweenM colom-biense strains Differences inGPL production between strainscould influence the M colombiense ability to form biofilmson polyvinylchloride (PVC) pipes which would facilitatethe dissemination of these bacteria in natural environmentssuch as in-hospital spaces thereby increasing the chance ofinfection in immunosuppressed patients [23 39 40]
Based on the GPL biosynthetic pathway previouslydescribed for the M avium 104 strain [25] we are currentlyconstructing a potential gene cluster for GPL biosynthesis inM colombiense which will be finished when the nucleotidesequence of the CECT 3035 is completely assembled RT-qPCR experiments showed that the planktonic and motilecells of the rough 57B strain showed decreased transcriptionof the 9 selected genes leading to the low production ofGPLs in this rough variant comparedwith theM colombiensestrains displaying smooth colony morphology Interestinglyplanktonic 19B cells exhibited lower expression of most of theselected genes compared with planktonic CECT 3035 cellsThis behaviour results in the reduced production of GPLs inplanktonic 19B cells compared with the CECT 3035 strainconsistent with the results of the TLC experiments
It has been previously shown that an rtfA mutation inthe M avium serovar-2-specific strain resulted in the lossof serovar-specific GPLs thereby diminishing the variabilityof these glycolipids in the cell envelope [41 42] Thus itis possible that the enzymatic activity of the RtfA proteincould be relevant for the increased glycosylation andor aug-mented production of ssGPLs in strains displaying smoothcolony morphology compared with the rough variant 57BIn strains displaying smooth colony morphology the genesencoding rhamnosyl andmethyl transferases (rtfA andmtfB)are overtranscribed suggesting the increased production ofnsGPLs precursors for ssGPL biosynthesis [24ndash26] therebyincreasing the production of GPLs The diminished tran-scription of pstA gftA mtfC mtfD tmtpBand tmtpC inmotile CECT 3035 compared with control cells (planktonicCECT 3035) is intriguing however this behaviour also
suggests the relevance of rhamnosyl transferases particularlyRtfA in GPL biosynthesis in M colombiense Neverthelessthe actual role for RtfA in M colombiense strains should beconfirmed using rtfA-null mutants
We did not identify 7 of the 30 known GPL cluster genesof M avium 104 in the genome sequence of M colombienseCECT 3035 This interesting observation suggests that theGPLs between these two closely related MAC species couldhave different structural characteristics However furtherexperiments using mass spectrometry and NMR are neces-sary to evaluate potential differences in the GPL structureamongM colombiense strains
5 Conclusions
In conclusion the results of the present study show that Mcolombiense strains displaying smooth morphology exhibitincreased biofilm formation on hydrophobic polystyrenesurfaces and enhanced sliding over motility plates Bioin-formatics analyses indicate that the gene cluster establishedfor GPL formation modification and translocation is intactstrongly suggesting that GPLs are putatively present in Mcolombiense In addition motile culture conditions activatethe transcription of the genes implicated in the key enzymaticactivities of GPL biosynthesis
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
Thisworkwas supported through funding from the ldquoDivisionde Investigacion Bogota (DIB)rdquo-Vicerrectorıa de Investi-gacion Universidad Nacional de Colombia (Grants 1606014837 and 14337) Milena Maya-Hoyos received a fellowshipfrom the Academic Direction Universidad Nacional deColombia and the ldquoJovenes Investigadores e InnovadoresrdquoProgram Colciencias Colombia This research was alsosupported in part through the Intramural Research Programof the National Institutes of Health National Library ofMedicine and National Center for Biotechnology Informa-tion (NCBI)
References
[1] S K Field D Fisher and R L Cowie ldquoMycobacterium aviumcomplex pulmonary disease in patient without HIV infectionrdquoChest vol 126 no 2 pp 566ndash581 2004
[2] M J M Vaerewijck G Huys J C Palomino J Swings and FPortaels ldquoMycobacteria in drinking water distribution systemsecology and significance for human healthrdquo FEMSMicrobiologyReviews vol 29 no 5 pp 911ndash934 2005
[3] E Hagiwara S Komatsu R Nishihira T Shinohara T Babaand T Ogura ldquoClinical characteristics and prevalence of pneu-mothorax in patients with pulmonary Mycobacterium aviumcomplex diseaserdquo Journal of Infection and Chemotherapy vol19 no 4 pp 588ndash592 2013
10 BioMed Research International
[4] M I Murcia E Tortoli M C Menendez E Palenque andM J Garcia ldquoMycobacterium colombiense sp nov a novelmember of theMycobacterium avium complex and descriptionof MAC-X as a new ITS genetic variantrdquo International Journalof Systematic and Evolutionary Microbiology vol 56 no 9 pp2049ndash2054 2006
[5] E Tortoli L Rindi M J Garcia et al ldquoProposal to elevatethe genetic variant MAC-A included in the Mycobacteriumavium complex to species rank as Mycobacterium chimaerasp novrdquo International Journal of Systematic and EvolutionaryMicrobiology vol 54 no 4 pp 1277ndash1285 2004
[6] I B Salah C Cayrou D Raoult and M Drancourt ldquoMycobac-terium marseillense sp nov Mycobacterium timonense sp novand Mycobacterium bouchedurhonense sp novmembers ofthe Mycobacterium avium complexrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 11 pp2803ndash2808 2009
[7] J van Ingen M J Boeree K Kosters et al ldquoProposal toelevate Mycobacterium avium complex ITS sequevar MAC-Qto Mycobacterium vulneris sp novrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 9 pp2277ndash2282 2009
[8] D Bang T Herlin M Stegger et al ldquoMycobacterium arosiensesp nov a slowly growing scotochromogenic species causingosteomyelitis in an immunocompromised childrdquo InternationalJournal of Systematic and Evolutionary Microbiology vol 58part 10 pp 2398ndash2402 2008
[9] C Cayrou C Turenne M A Behr and M Drancourt ldquoGeno-typing of Mycobacterium avium complex organisms usingmultispacer sequence typingrdquoMicrobiology vol 156 part 3 pp687ndash694 2010
[10] S C Smole F McAleese J Ngampasutadol C F von Reynand R D Arbeit ldquoClinical and epidemiological correlates ofgenotypes within theMycobacterium avium complex defined byrestriction and sequence analysis of hsp65rdquo Journal of ClinicalMicrobiology vol 40 no 9 pp 3374ndash3380 2002
[11] AMuwonge C Kankya T B Johansen et al ldquoNon-tuberculousmycobacteria isolated from slaughter pigs in Mubende districtUgandardquo BMC Veterinary Research vol 8 article 52 2012
[12] J O Falkinham III ldquoSurrounded by mycobacteria nontuber-culous mycobacteria in the human environmentrdquo Journal ofApplied Microbiology vol 107 no 2 pp 356ndash367 2009
[13] K Vuorenmaa I B Salah V Barlogis H Chambost and MDrancourt ldquoMycobacterium colombiense and pseudotubercu-lous lymphadenopathyrdquo Emerging Infectious Diseases vol 15no 4 pp 618ndash620 2009
[14] O Esparcia F Navarro M Quer and P Coll ldquoLymphadenopa-thy caused by Mycobacterium colombienserdquo Journal of ClinicalMicrobiology vol 46 no 5 pp 1885ndash1887 2008
[15] S Cohen-Bacrie M David N Stremler J-C Dubus J-MRolain and M Drancourt ldquoMycobacterium chimaera pul-monary infection complicating cystic fibrosis a case reportrdquoJournal of Medical Case Reports vol 5 no 1 article 473 2011
[16] S Poulin C Corbeil M Nguyen et al ldquoFatal Mycobac-terium colombiensecytomegalovirus coinfection associatedwith acquired immunodeficiency due to autoantibodies againstinterferon gamma a case reportrdquo BMC Infectious Diseases vol13 no 1 article 24 2013
[17] J Leguizamon J Hernandez M-I Murcia and C-Y SotoldquoIdentification of potential biomarkers to distinguishMycobac-terium colombiense from other mycobacterial speciesrdquoMolecu-lar and Cellular Probes vol 27 no 1 pp 46ndash52 2013
[18] D Chatterjee and K H Khoo ldquoThe surface glycopeptidolipidsof mycobacteria structures and biological propertiesrdquo Cellularand Molecular Life Sciences vol 58 no 14 pp 2018ndash2042 2001
[19] J B Torrelles D Ellis T Osborne et al ldquoCharacterizationof virulence colony morphotype and the glycopeptidolipid ofMycobacterium avium strain 104rdquo Tuberculosis vol 82 no 6pp 293ndash300 2002
[20] M F Thorel and H L David ldquoSpecific surface antigens of SmTvariants ofMycobacterium aviumrdquo Infection and Immunity vol43 no 1 pp 438ndash439 1984
[21] A Martınez S Torello and R Kolter ldquoSliding motility inmycobacteriardquo Journal of Bacteriology vol 181 no 23 pp 7331ndash7338 1999
[22] J Recht AMartınez S Torello and R Kolter ldquoGenetic analysisof sliding motility in Mycobacterium smegmatisrdquo Journal ofBacteriology vol 182 no 15 pp 4348ndash4351 2000
[23] G Carter M Wu D C Drummond and L E BermudezldquoCharacterization of biofilm formation by clinical isolates ofMycobacterium aviumrdquo Journal of Medical Microbiology vol 52no 9 pp 747ndash752 2003
[24] J S Schorey and L Sweet ldquoThe mycobacterial glycopepti-dolipids structure function and their role in pathogenesisrdquoGlycobiology vol 18 no 11 pp 832ndash841 2008
[25] L Pang X Tian W Pan and J Xie ldquoStructure and function ofmycobacterium glycopeptidolipids from comparative genomicsperspectiverdquo Journal of Cellular Biochemistry vol 114 no 8 pp1705ndash1713 2013
[26] R Mukherjee and D Chatterji ldquoGlycopeptidolipids immuno-modulators in greasy mycobacterial cell enveloperdquo IUBMB Lifevol 64 no 3 pp 215ndash225 2012
[27] K L Horan R Freeman K Weigel et al ldquoIsolation ofthe genome sequence strain Mycobacterium avium 104 frommultiple patients over a 17-year periodrdquo Journal of ClinicalMicrobiology vol 44 no 3 pp 783ndash789 2006
[28] S B Snapper R E Melton S Mustafa T Kieser and W RJacobs Jr ldquoIsolation and characterization of efficient plasmidtransformation mutants of Mycobacterium smegmatisrdquo Molec-ular Microbiology vol 4 no 11 pp 1911ndash1919 1990
[29] M Radji R A Agustama B Elya and C R TjampakasarildquoAntimicrobial activity of green tea extract against isolatesof methicillin-resistant Staphylococcus aureus and multi-drugresistant Pseudomonas aeruginosardquo Asian Pacific Journal ofTropical Biomedicine vol 3 no 8 pp 663ndash667 2013
[30] P Helden T Victor R Warren and E Helden ldquoIsolationof DNA from Mycobacterium tuberculosisrdquo in Mycobacteriumtuberculosis Protocols Humana Press Totowa NJ USA 2001
[31] J C van Kessel L J Marinelli and G F Hatfull ldquoRecom-bineering mycobacteria and their phagesrdquo Nature ReviewsMicrobiology vol 6 no 11 pp 851ndash857 2008
[32] N M Parrish C G Ko J D Dick P B Jones and J LE Ellingson ldquoGrowth Congo Red agar colony morphotypesand antibiotic susceptibility testing of Mycobacterium aviumsubspecies paratuberculosisrdquo Clin Med Res vol 2 no 2 pp107ndash114 2004
[33] T Tugrul and E Cansunar ldquoDetecting surfactant-producingmicroorganisms by the drop-collapse testrdquo World Journal ofMicrobiology and Biotechnology vol 21 no 6-7 pp 851ndash8532005
[34] D K Jain D L Collins-Thompson H Lee and J T TrevorsldquoA drop-collapsing test for screening surfactant-producingmicroorganismsrdquo Journal ofMicrobiologicalMethods vol 13 no4 pp 271ndash279 1991
BioMed Research International 11
[35] M Munoz M-A Laneelle M Luquin et al ldquoOccurrence ofan antigenic triacyl trehalose in clinical isolates and referencestrains of Mycobacterium tuberculosisrdquo FEMS MicrobiologyLetters vol 157 no 2 pp 251ndash259 1997
[36] M Gonzalez-Perez M I Murcia D Landsman I K Jordanand L Marino-Ramırez ldquoGenome sequence of the Mycobac-terium colombiense type strain CECT 3035rdquo Journal of Bacte-riology vol 193 no 20 pp 5866ndash5867 2011
[37] J C Betts P T Lukey L C Robb R A McAdam andK Duncan ldquoEvaluation of a nutrient starvation model ofMycobacterium tuberculosis persistence by gene and proteinexpression profilingrdquoMolecular Microbiology vol 43 no 3 pp717ndash731 2002
[38] W W Barrow and P J Brennan ldquoIsolation in high frequencyof rough variants of Mycobacterium intracellulare lacking C-mycoside glycopeptidolipid antigensrdquo Journal of Bacteriologyvol 150 no 1 pp 381ndash384 1982
[39] W Costerton R Veeh M Shirtliff M Pasmore C Post andG Ehrlich ldquoThe application of biofilm science to the studyand control of chronic bacterial infectionsrdquo Journal of ClinicalInvestigation vol 112 no 10 pp 1466ndash1477 2003
[40] J Recht and R Kolter ldquoGlycopeptidolipid acetylation affectssliding motility and biofilm formation inMycobacterium smeg-matisrdquo Journal of Bacteriology vol 183 no 19 pp 5718ndash57242001
[41] T M Eckstein F S Silbaq D Chatterjee N J Kelly P JBrennan and J T Belisle ldquoIdentification and recombinantexpression of a Mycobacterium avium rhamnosyltransferasegene (rtfA) involved in glycopeptidolipid biosynthesisrdquo Journalof Bacteriology vol 180 no 21 pp 5567ndash5573 1998
[42] J N Maslow V R Irani S-H Lee T M Eckstein J MInamine and J T Belisle ldquoBiosynthetic specificity of therhamnosyltransferase gene of Mycobacterium avium serovar 2as determined by allelic exchange mutagenesisrdquo Microbiologyvol 149 no 11 pp 3193ndash3202 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 5
7
17
27
37
47
57
0 10 20 30 40 50 60 70
Dia
met
er (m
m)
Day
M colombiense 19BM avium
M colombiense CECT 3035
M colombiense 57B
Figure 2 Motility rates for Mycobacterium colombiense strainsMycobacterial strains were cultivated on 7H9-ADC-035 agaroseand incubated at 37∘C for 4-5 weeks The colony growth haloswere measured daily to determine the motility rate Each reportedvalue represents the mean of the colony growth halos from threeindependent experiments The presented data have statisticallysignificant differences compared with the values obtained from Mavium 104 (119875 lt 005)
study (6B 7B 9B and 16B) also displayed a smooth colonymorphology (data not shown) Both smooth and rough bac-terial phenotypes were stable after repetitive culture on solidmedia for the duration of the study (3 years) The smoothM colombiense strains developed yellow pigmentation withage specifically at the stationary phase in all culture mediahowever the rough variant (57B) remained beige in colourunder the same experimental conditions (Figure 1)
Because alterations in the phenotypic characteristicsassociated with colony morphology have been observedin other MAC species motility and biofilm formationwere compared between smooth and rough M colombiensestrains We observed that the M colombiense 19B andCECT3035 strains (smooth colony morphology) showedincreased spreading (4094 plusmn 005 120583mday and 2972 plusmn005 120583mday resp) on hydrophilic agarose comparedwith thenatural rough variant 57B which showed impaired spreading(Figure 2)
Crystal violet staining was used to quantify the degree ofM colombiense biofilm formation onpolystyreneThehighestOD570
values for all M colombiense strains were obtainedusing an inoculum of sim27 times 105 CFUmL In general thestrains displaying smooth morphology showed increasedbiofilm formation on hydrophobic surfaces while therough variant 57B showed reduced adhesion to polystyrene(Figure 3)
As biosurfactants could potentially influence cell motilitythe ldquodrop-collapsingrdquo method was used to detect biosurfac-tant secretion in M colombiense strains [22] As shown inFigure 4 drops of 1 SDS and P aeruginosa aqueous extract(positive controls) spread over the oily surface after the sam-ples were incubated indicating the presence of biosurfactantsubstances In contrast drops of distilled waterM smegmatis(negative control) and M colombiense aqueous extracts didnot collapse on the oily surface but rather appeared as firm
0
005
01
015
02
025
OD570
Mavium
104
Mcolom
biense
CECT
3035
Mcolom
biense19
B
Mcolom
biense57
B
Figure 3 Capacity of Mycobacterium colombiense strains to formbiofilms The OD
570nm represents the proportion of mycobacteria
added to the microtiter wells that reacted with the crystal violetsolutionThebars represent the SD calculated from two independentexperiments each performed in triplicate
drops suggesting the absence of biosurfactants in the culturesupernatants
32 M colombiense Displaying a Smooth Colony Morphol-ogy Contains Glycolipids with Thin-Layer ChromatographicBehaviour Similar to That of the GPLs of M avium TLCanalysis of noncovalently attached lipids extracted frommycobacteria cultured under planktonic and motile condi-tions showed that M colombiense strains contain glycolipidswith chromatographic behaviour similar to that of theGPLsofM avium [23] Regarding planktonic cells (Figure 5(a)) thestrains with smooth colony morphology (19B and CECT3035) showed multiple lipid spots migrating in the region ofcontrol GPLs (M avium 104) however the spots observed inthe CECT 3035 lipid extract were more intense than those inthe 19B strain The natural rough variant (57B) showed onlyone spot that migrated in the same GPLs region
Under motile conditions (Figure 5(b)) the smoothcolony morphology strains 19B and CECT 3035 showed aGPL profile similar to that obtained for cells cultured underplanktonic conditions nevertheless the counterpart spotswere more intense than those observed in planktonic cellsparticularly for the 19B strain Nevertheless the unique spotobserved for motile 57B cells (rough variant) were moreintense than that observed for planktonic 57B cells Inter-estingly all motile M colombiense cells exhibited reducedproduction of polar glycolipids such as phosphatidyl-inositolmannosides (PIMs) and phosphatidylglycerol (PG) com-pared with planktonic cells (Figure 5(b))
33 In Silico Identification of GPL Biosynthesis Genes inM colombiense CECT 3035 The genes involved in GPLbiosynthesis in M colombiense are currently unknown
6 BioMed Research International
(a) (b) (c) (d)
(e) (f) (g) (h)
Figure 4 Drop-collapsing test (a) Water (no surfactant) (b) 1 SDS (surfactant) (c) P aeruginosa (d) M smegmatis mc2155 (e) Mcolombiense 19B (f) M colombiense CECT 3035 (g) M colombiense 57B and (h) M avium 104 Drop spreading over the oily surface aftersamples incubation indicates the presence of biosurfactant substances in the aqueous sampleThis assay was conducted in three independentexperiments each performed in duplicate
Based on the GPL biosynthetic pathway reported for Mavium 104 [25] 30 open reading frames (ORF) encodingpeptide synthetase fatty acid synthases (FAS) polyketidesynthases (PKs) acyltransferase (PapA) glycosyltransferasescarbohydrate synthetases methyl and acyl transferases gly-colipid transporters potential biosynthesis regulators andproteins with unknown functions [25 26] were searchedwithin the recently reported nucleotide sequence of Mcolombiense CECT 3035 [36] This in silico analysis (seeTable S1 of the Supplementary Material available online athttpdxdoiorg1011552015419549) showed that all enzy-matic activities searched were encoded by different ORFsin M colombiense CECT 3035 with an identity rangingbetween 77 and 96 The ORFs in the genome of the Mcolombiense counterpart were located in the contigs 0000100002A 00003A and 00007 of the genome sequence of theCECT 3035 strain (Figure 6) Among the 30 genes of the GPLsynthetic pathway inM avium 104 7 genes (mtfA gtfB gtfDdghA fadD23 pe and gap-like) did not have orthologoussequences inM colombiense CECT 3035 (Table S1)
34 Differential Transcription of the GPL Biosynthesis Genesin M colombiense Displaying Smooth and Rough ColonyMorphologies RNAwas isolated fromM colombienseCECT3035 19B and 57B strains cultured under planktonic andmotile conditions to quantify the transcription levels ofthe pstA gtfA rtfA mtfB mtfC mtfD tmtpC tmtpA andtmtpB genes predicted to encode key enzymes of the GPLbiosynthetic pathway (Table S1) For relative quantificationM colombiense CECT 3035 was grown to the exponentialphase under planktonic conditions and used as the referencepattern for two reasons (1) CECT3035 is theM colombiensegenome sequence strain [36] and consistent with the TLCanalysis (2) planktonic cells displayed the lowest GPL pro-duction
With regard to planktonic cells the rate ofM colombiense19B gene transcription was consistently higher than thatfor the M colombiense 57B genes Conversely the roughvariant (57B) displayed a dramatic decrease in the rate oftranscription for the 9 selected genes particularly thoseencoding peptide synthetase pstA glycosyltransferase gtfA
BioMed Research International 7
1 2 3 41 2 3 4
GPLs
PG
PIMs
(a) (b)
Figure 5 TLC profile of the GPLs in Mycobacterium colombiense strains Crude lipid extracts of (1) M avium 104 (2) M colombiense19B (3) M colombiense CECT 3035 and (4) M colombiense 57B Cells cultivated (a) under planktonic and (b) motile conditions weredeveloped using chloroform-methanol-water (65 25 4 vvv) GPLs glycopeptidolipids PIMs phosphatidyl-inositol mannosides and PGphosphatidylglycerol
the methyltransferases mtfB and mtfC and the lipid trans-porters tmtpA tmtpB and tmtpC (Figure 7(a)) InterestinglyrtfA (rhamnosyltransferase) in the 19B strain (smooth colonymorphology) was the only gene among the strains culturedunder planktonic conditions that showed an increased tran-scription rate (221-fold higher) compared with the controlcells (planktonicM colombiense CECT 3035 at the exponen-tial phase of growth)
Under motile growth conditions M colombiense 19Bshowed the highest transcription rate for all selected genescompared with the CECT 3035 and 57B strains (Figure 7(b))In contrast the transcription rates for all genes in the57B strain (rough variant) except tmtpA and tmtpC (lipidtransporters) were reduced In motile CECT 3035 cellswhereas rtfA mtfB (methyl transfers) and tmtpA showedincreased transcription the pstA gftA mtfC mtfD tmtpBand tmtpC genes exhibited reduced transcription comparedwith control cells
4 Discussion
In the present study TLC experiments were able to show thatM colombiense contains glycolipids with chromatographicbehaviour similar to the GPLs of M avium 104 which havebeen previously identified and completely characterised [23]in addition the colony morphology was associated withthe GPL profile of M colombiense strains Thus the TLCanalysis revealed that (1) strains displaying smooth colonymorphology (19B and CECT3035) produce a higher amountof variable GPLs compared with the rough natural variant(57B) strain and (2) the GPL production was augmentedin M colombiense cells cultivated under the motile growthconditions As expected the TLC analyses did not provide
detailed information about GPL structure however struc-tural determinationwas not the objective of the present study
Themedia composition influences the glycolipid contentof the mycobacterial cell wall The MAC strains growing onsolid medium exhibit a significant rate of transition fromsmooth transparent to smooth opaque and from smooth torough morphologies [18 19 38] We compared planktonicand motile M colombiense cells grown in the same culturemedium (7H9-ADC or 7H9-ADC-agarose) to avoid differ-ences in lipid content produced through alterations in themedia composition The results showed that the low GPLcontent observed in the rough 57B variant is less frequentamong M colombiense strains therefore M colombienseshows a preference for augmented GPL production andsmooth colony morphology Interestingly planktonic Mcolombiense cells displayed increased PIM and PG pro-duction and diminished GPL content compared with cellscultivated under motile conditions suggesting a potentialcompensatory mechanism in cell wall lipid synthesis thatcompensates for the low GPL content
The abundance of GPLs in the outermost portion ofthe cell envelope could be associated with the motility ofM colombiense strains on hydrophilic surfaces Recht andothers [22] proposed that GPLs are linked through thehydrophilic head to the cellular capsule and the hydrophobicfatty acid chain is exposed to the bacterial surface therebyreducing interactions with hydrophilic agarose surfaces andfacilitating the spread of cells on solid medium [21 22]Thus the augmented GPL production of motile 19B andCECT 3035 cells compared with that of the rough variant57B is consistent with the sliding motility on the hydrophilicagarose surface observed for these strains It has also beensuggested that bacterial sliding motility might be favoured
8 BioMed Research International
33kb 2kb9kb51kb
Contig 00001
600kb
87kb 245kb
488kb 36kb320kb
Contig 00002A
Contig 00003A
584kb 452kb
mmpL10
326kb
Contig 00007
fadE
15
atf
rmlB
ecf
rv1174
rmlA
mmpL
4a
mmpL
4b
mmpS4
mbtH
gap rv0926
sap
pks papA3
gtfA
mtfB
mtfC
mtfD
rtfA
pstA
pstB
(a)
ecf
fadE
5
rv1174
atf
rmlB
rmlA
mmpL
4a
mmpL
4b
mmpS
4
mbtH
gap
237kb
rv0926
7kb90kb
530kb
300kb
738kb
mmpL10
sap
Saccharides biosynthesis transfer and modification
Fatty acyl chain biosynthesis modification and transfer
Members of the mmpSL family
Peptide synthesis Unknown
Possible regulatory genes
GPL export to the cell surface
pks pe fadD23 gap-likepapA3
mtfB
gtfA
mtfC
mtfD
dhgA
rtfA
pstA
pstB
mtfA
gtfB
gtfD
(b)
Figure 6Model showing the gene cluster proposed for the GPL biosynthesis ofMycobacterium colombienseCECT 3035 (a)The location andidentity of theM colombiense genes were established using the previously described biosynthetic pathway for GPL biosynthesis annotated intheM avium 104 genomic sequence (b)
0001
001
01
1
10
Rela
tive g
ene t
rans
crip
tion
M colombiense 57BM colombiense 19B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(a)
01
1
10
100
Rela
tive g
ene t
rans
crip
tion
M colombiense CECT 3035
M colombiense 19B
M colombiense 57B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(b)
Figure 7 In vitro transcription of Mycobacterium colombiense genes involved in GPL biosynthesis Relative quantification expressed as theratio of gene transcription in (a) M colombiense cells grown under planktonic conditionstranscription ofM colombiense CECT 3035 cellsgrown under planktonic conditions and (b)M colombiense cells grown on the motility mediumtranscription ofM colombiense CECT 3035grown under planktonic conditions The presented data have statistically significant differences compared with the values obtained fromMcolombiense CECT 3035 cells (119875 lt 005)
BioMed Research International 9
through the secretion of surfactant substances from cells[21 22] The results of the drop-collapsing test showed thatbiosurfactants were likely not secreted from M colombiensecells suggesting that the motility of M colombiense couldbe favoured through hydrophobic molecules such as GPLsbound to the outermost portion of cells however we cannotcompletely rule out the possibility that some of the surfactantsubstances secreted from M colombiense remained adheredto the mycobacterial cell surface thereby influencing cellmotility
GPLs on the outermost portions of the M colombiensecell envelope generate a more hydrophobic cell surfacethat facilitates initial interactions with hydrophobic surfacesand increase biofilm formation on polystyrene wells Inaddition the increased GPL production in 19B and CECT3035 cells is consistent with the augmented biofilm forma-tion observed for M colombiense strains displaying smoothcolony morphology Moreover increased biofilm formationon hydrophobic surfaces and enhanced sliding over motilityplates for 19B cells compared with the CECT 3035 strainwhich also displays smooth colony morphology likely sug-gest possible differences in GPL structure betweenM colom-biense strains Differences inGPL production between strainscould influence the M colombiense ability to form biofilmson polyvinylchloride (PVC) pipes which would facilitatethe dissemination of these bacteria in natural environmentssuch as in-hospital spaces thereby increasing the chance ofinfection in immunosuppressed patients [23 39 40]
Based on the GPL biosynthetic pathway previouslydescribed for the M avium 104 strain [25] we are currentlyconstructing a potential gene cluster for GPL biosynthesis inM colombiense which will be finished when the nucleotidesequence of the CECT 3035 is completely assembled RT-qPCR experiments showed that the planktonic and motilecells of the rough 57B strain showed decreased transcriptionof the 9 selected genes leading to the low production ofGPLs in this rough variant comparedwith theM colombiensestrains displaying smooth colony morphology Interestinglyplanktonic 19B cells exhibited lower expression of most of theselected genes compared with planktonic CECT 3035 cellsThis behaviour results in the reduced production of GPLs inplanktonic 19B cells compared with the CECT 3035 strainconsistent with the results of the TLC experiments
It has been previously shown that an rtfA mutation inthe M avium serovar-2-specific strain resulted in the lossof serovar-specific GPLs thereby diminishing the variabilityof these glycolipids in the cell envelope [41 42] Thus itis possible that the enzymatic activity of the RtfA proteincould be relevant for the increased glycosylation andor aug-mented production of ssGPLs in strains displaying smoothcolony morphology compared with the rough variant 57BIn strains displaying smooth colony morphology the genesencoding rhamnosyl andmethyl transferases (rtfA andmtfB)are overtranscribed suggesting the increased production ofnsGPLs precursors for ssGPL biosynthesis [24ndash26] therebyincreasing the production of GPLs The diminished tran-scription of pstA gftA mtfC mtfD tmtpBand tmtpC inmotile CECT 3035 compared with control cells (planktonicCECT 3035) is intriguing however this behaviour also
suggests the relevance of rhamnosyl transferases particularlyRtfA in GPL biosynthesis in M colombiense Neverthelessthe actual role for RtfA in M colombiense strains should beconfirmed using rtfA-null mutants
We did not identify 7 of the 30 known GPL cluster genesof M avium 104 in the genome sequence of M colombienseCECT 3035 This interesting observation suggests that theGPLs between these two closely related MAC species couldhave different structural characteristics However furtherexperiments using mass spectrometry and NMR are neces-sary to evaluate potential differences in the GPL structureamongM colombiense strains
5 Conclusions
In conclusion the results of the present study show that Mcolombiense strains displaying smooth morphology exhibitincreased biofilm formation on hydrophobic polystyrenesurfaces and enhanced sliding over motility plates Bioin-formatics analyses indicate that the gene cluster establishedfor GPL formation modification and translocation is intactstrongly suggesting that GPLs are putatively present in Mcolombiense In addition motile culture conditions activatethe transcription of the genes implicated in the key enzymaticactivities of GPL biosynthesis
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
Thisworkwas supported through funding from the ldquoDivisionde Investigacion Bogota (DIB)rdquo-Vicerrectorıa de Investi-gacion Universidad Nacional de Colombia (Grants 1606014837 and 14337) Milena Maya-Hoyos received a fellowshipfrom the Academic Direction Universidad Nacional deColombia and the ldquoJovenes Investigadores e InnovadoresrdquoProgram Colciencias Colombia This research was alsosupported in part through the Intramural Research Programof the National Institutes of Health National Library ofMedicine and National Center for Biotechnology Informa-tion (NCBI)
References
[1] S K Field D Fisher and R L Cowie ldquoMycobacterium aviumcomplex pulmonary disease in patient without HIV infectionrdquoChest vol 126 no 2 pp 566ndash581 2004
[2] M J M Vaerewijck G Huys J C Palomino J Swings and FPortaels ldquoMycobacteria in drinking water distribution systemsecology and significance for human healthrdquo FEMSMicrobiologyReviews vol 29 no 5 pp 911ndash934 2005
[3] E Hagiwara S Komatsu R Nishihira T Shinohara T Babaand T Ogura ldquoClinical characteristics and prevalence of pneu-mothorax in patients with pulmonary Mycobacterium aviumcomplex diseaserdquo Journal of Infection and Chemotherapy vol19 no 4 pp 588ndash592 2013
10 BioMed Research International
[4] M I Murcia E Tortoli M C Menendez E Palenque andM J Garcia ldquoMycobacterium colombiense sp nov a novelmember of theMycobacterium avium complex and descriptionof MAC-X as a new ITS genetic variantrdquo International Journalof Systematic and Evolutionary Microbiology vol 56 no 9 pp2049ndash2054 2006
[5] E Tortoli L Rindi M J Garcia et al ldquoProposal to elevatethe genetic variant MAC-A included in the Mycobacteriumavium complex to species rank as Mycobacterium chimaerasp novrdquo International Journal of Systematic and EvolutionaryMicrobiology vol 54 no 4 pp 1277ndash1285 2004
[6] I B Salah C Cayrou D Raoult and M Drancourt ldquoMycobac-terium marseillense sp nov Mycobacterium timonense sp novand Mycobacterium bouchedurhonense sp novmembers ofthe Mycobacterium avium complexrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 11 pp2803ndash2808 2009
[7] J van Ingen M J Boeree K Kosters et al ldquoProposal toelevate Mycobacterium avium complex ITS sequevar MAC-Qto Mycobacterium vulneris sp novrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 9 pp2277ndash2282 2009
[8] D Bang T Herlin M Stegger et al ldquoMycobacterium arosiensesp nov a slowly growing scotochromogenic species causingosteomyelitis in an immunocompromised childrdquo InternationalJournal of Systematic and Evolutionary Microbiology vol 58part 10 pp 2398ndash2402 2008
[9] C Cayrou C Turenne M A Behr and M Drancourt ldquoGeno-typing of Mycobacterium avium complex organisms usingmultispacer sequence typingrdquoMicrobiology vol 156 part 3 pp687ndash694 2010
[10] S C Smole F McAleese J Ngampasutadol C F von Reynand R D Arbeit ldquoClinical and epidemiological correlates ofgenotypes within theMycobacterium avium complex defined byrestriction and sequence analysis of hsp65rdquo Journal of ClinicalMicrobiology vol 40 no 9 pp 3374ndash3380 2002
[11] AMuwonge C Kankya T B Johansen et al ldquoNon-tuberculousmycobacteria isolated from slaughter pigs in Mubende districtUgandardquo BMC Veterinary Research vol 8 article 52 2012
[12] J O Falkinham III ldquoSurrounded by mycobacteria nontuber-culous mycobacteria in the human environmentrdquo Journal ofApplied Microbiology vol 107 no 2 pp 356ndash367 2009
[13] K Vuorenmaa I B Salah V Barlogis H Chambost and MDrancourt ldquoMycobacterium colombiense and pseudotubercu-lous lymphadenopathyrdquo Emerging Infectious Diseases vol 15no 4 pp 618ndash620 2009
[14] O Esparcia F Navarro M Quer and P Coll ldquoLymphadenopa-thy caused by Mycobacterium colombienserdquo Journal of ClinicalMicrobiology vol 46 no 5 pp 1885ndash1887 2008
[15] S Cohen-Bacrie M David N Stremler J-C Dubus J-MRolain and M Drancourt ldquoMycobacterium chimaera pul-monary infection complicating cystic fibrosis a case reportrdquoJournal of Medical Case Reports vol 5 no 1 article 473 2011
[16] S Poulin C Corbeil M Nguyen et al ldquoFatal Mycobac-terium colombiensecytomegalovirus coinfection associatedwith acquired immunodeficiency due to autoantibodies againstinterferon gamma a case reportrdquo BMC Infectious Diseases vol13 no 1 article 24 2013
[17] J Leguizamon J Hernandez M-I Murcia and C-Y SotoldquoIdentification of potential biomarkers to distinguishMycobac-terium colombiense from other mycobacterial speciesrdquoMolecu-lar and Cellular Probes vol 27 no 1 pp 46ndash52 2013
[18] D Chatterjee and K H Khoo ldquoThe surface glycopeptidolipidsof mycobacteria structures and biological propertiesrdquo Cellularand Molecular Life Sciences vol 58 no 14 pp 2018ndash2042 2001
[19] J B Torrelles D Ellis T Osborne et al ldquoCharacterizationof virulence colony morphotype and the glycopeptidolipid ofMycobacterium avium strain 104rdquo Tuberculosis vol 82 no 6pp 293ndash300 2002
[20] M F Thorel and H L David ldquoSpecific surface antigens of SmTvariants ofMycobacterium aviumrdquo Infection and Immunity vol43 no 1 pp 438ndash439 1984
[21] A Martınez S Torello and R Kolter ldquoSliding motility inmycobacteriardquo Journal of Bacteriology vol 181 no 23 pp 7331ndash7338 1999
[22] J Recht AMartınez S Torello and R Kolter ldquoGenetic analysisof sliding motility in Mycobacterium smegmatisrdquo Journal ofBacteriology vol 182 no 15 pp 4348ndash4351 2000
[23] G Carter M Wu D C Drummond and L E BermudezldquoCharacterization of biofilm formation by clinical isolates ofMycobacterium aviumrdquo Journal of Medical Microbiology vol 52no 9 pp 747ndash752 2003
[24] J S Schorey and L Sweet ldquoThe mycobacterial glycopepti-dolipids structure function and their role in pathogenesisrdquoGlycobiology vol 18 no 11 pp 832ndash841 2008
[25] L Pang X Tian W Pan and J Xie ldquoStructure and function ofmycobacterium glycopeptidolipids from comparative genomicsperspectiverdquo Journal of Cellular Biochemistry vol 114 no 8 pp1705ndash1713 2013
[26] R Mukherjee and D Chatterji ldquoGlycopeptidolipids immuno-modulators in greasy mycobacterial cell enveloperdquo IUBMB Lifevol 64 no 3 pp 215ndash225 2012
[27] K L Horan R Freeman K Weigel et al ldquoIsolation ofthe genome sequence strain Mycobacterium avium 104 frommultiple patients over a 17-year periodrdquo Journal of ClinicalMicrobiology vol 44 no 3 pp 783ndash789 2006
[28] S B Snapper R E Melton S Mustafa T Kieser and W RJacobs Jr ldquoIsolation and characterization of efficient plasmidtransformation mutants of Mycobacterium smegmatisrdquo Molec-ular Microbiology vol 4 no 11 pp 1911ndash1919 1990
[29] M Radji R A Agustama B Elya and C R TjampakasarildquoAntimicrobial activity of green tea extract against isolatesof methicillin-resistant Staphylococcus aureus and multi-drugresistant Pseudomonas aeruginosardquo Asian Pacific Journal ofTropical Biomedicine vol 3 no 8 pp 663ndash667 2013
[30] P Helden T Victor R Warren and E Helden ldquoIsolationof DNA from Mycobacterium tuberculosisrdquo in Mycobacteriumtuberculosis Protocols Humana Press Totowa NJ USA 2001
[31] J C van Kessel L J Marinelli and G F Hatfull ldquoRecom-bineering mycobacteria and their phagesrdquo Nature ReviewsMicrobiology vol 6 no 11 pp 851ndash857 2008
[32] N M Parrish C G Ko J D Dick P B Jones and J LE Ellingson ldquoGrowth Congo Red agar colony morphotypesand antibiotic susceptibility testing of Mycobacterium aviumsubspecies paratuberculosisrdquo Clin Med Res vol 2 no 2 pp107ndash114 2004
[33] T Tugrul and E Cansunar ldquoDetecting surfactant-producingmicroorganisms by the drop-collapse testrdquo World Journal ofMicrobiology and Biotechnology vol 21 no 6-7 pp 851ndash8532005
[34] D K Jain D L Collins-Thompson H Lee and J T TrevorsldquoA drop-collapsing test for screening surfactant-producingmicroorganismsrdquo Journal ofMicrobiologicalMethods vol 13 no4 pp 271ndash279 1991
BioMed Research International 11
[35] M Munoz M-A Laneelle M Luquin et al ldquoOccurrence ofan antigenic triacyl trehalose in clinical isolates and referencestrains of Mycobacterium tuberculosisrdquo FEMS MicrobiologyLetters vol 157 no 2 pp 251ndash259 1997
[36] M Gonzalez-Perez M I Murcia D Landsman I K Jordanand L Marino-Ramırez ldquoGenome sequence of the Mycobac-terium colombiense type strain CECT 3035rdquo Journal of Bacte-riology vol 193 no 20 pp 5866ndash5867 2011
[37] J C Betts P T Lukey L C Robb R A McAdam andK Duncan ldquoEvaluation of a nutrient starvation model ofMycobacterium tuberculosis persistence by gene and proteinexpression profilingrdquoMolecular Microbiology vol 43 no 3 pp717ndash731 2002
[38] W W Barrow and P J Brennan ldquoIsolation in high frequencyof rough variants of Mycobacterium intracellulare lacking C-mycoside glycopeptidolipid antigensrdquo Journal of Bacteriologyvol 150 no 1 pp 381ndash384 1982
[39] W Costerton R Veeh M Shirtliff M Pasmore C Post andG Ehrlich ldquoThe application of biofilm science to the studyand control of chronic bacterial infectionsrdquo Journal of ClinicalInvestigation vol 112 no 10 pp 1466ndash1477 2003
[40] J Recht and R Kolter ldquoGlycopeptidolipid acetylation affectssliding motility and biofilm formation inMycobacterium smeg-matisrdquo Journal of Bacteriology vol 183 no 19 pp 5718ndash57242001
[41] T M Eckstein F S Silbaq D Chatterjee N J Kelly P JBrennan and J T Belisle ldquoIdentification and recombinantexpression of a Mycobacterium avium rhamnosyltransferasegene (rtfA) involved in glycopeptidolipid biosynthesisrdquo Journalof Bacteriology vol 180 no 21 pp 5567ndash5573 1998
[42] J N Maslow V R Irani S-H Lee T M Eckstein J MInamine and J T Belisle ldquoBiosynthetic specificity of therhamnosyltransferase gene of Mycobacterium avium serovar 2as determined by allelic exchange mutagenesisrdquo Microbiologyvol 149 no 11 pp 3193ndash3202 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 BioMed Research International
(a) (b) (c) (d)
(e) (f) (g) (h)
Figure 4 Drop-collapsing test (a) Water (no surfactant) (b) 1 SDS (surfactant) (c) P aeruginosa (d) M smegmatis mc2155 (e) Mcolombiense 19B (f) M colombiense CECT 3035 (g) M colombiense 57B and (h) M avium 104 Drop spreading over the oily surface aftersamples incubation indicates the presence of biosurfactant substances in the aqueous sampleThis assay was conducted in three independentexperiments each performed in duplicate
Based on the GPL biosynthetic pathway reported for Mavium 104 [25] 30 open reading frames (ORF) encodingpeptide synthetase fatty acid synthases (FAS) polyketidesynthases (PKs) acyltransferase (PapA) glycosyltransferasescarbohydrate synthetases methyl and acyl transferases gly-colipid transporters potential biosynthesis regulators andproteins with unknown functions [25 26] were searchedwithin the recently reported nucleotide sequence of Mcolombiense CECT 3035 [36] This in silico analysis (seeTable S1 of the Supplementary Material available online athttpdxdoiorg1011552015419549) showed that all enzy-matic activities searched were encoded by different ORFsin M colombiense CECT 3035 with an identity rangingbetween 77 and 96 The ORFs in the genome of the Mcolombiense counterpart were located in the contigs 0000100002A 00003A and 00007 of the genome sequence of theCECT 3035 strain (Figure 6) Among the 30 genes of the GPLsynthetic pathway inM avium 104 7 genes (mtfA gtfB gtfDdghA fadD23 pe and gap-like) did not have orthologoussequences inM colombiense CECT 3035 (Table S1)
34 Differential Transcription of the GPL Biosynthesis Genesin M colombiense Displaying Smooth and Rough ColonyMorphologies RNAwas isolated fromM colombienseCECT3035 19B and 57B strains cultured under planktonic andmotile conditions to quantify the transcription levels ofthe pstA gtfA rtfA mtfB mtfC mtfD tmtpC tmtpA andtmtpB genes predicted to encode key enzymes of the GPLbiosynthetic pathway (Table S1) For relative quantificationM colombiense CECT 3035 was grown to the exponentialphase under planktonic conditions and used as the referencepattern for two reasons (1) CECT3035 is theM colombiensegenome sequence strain [36] and consistent with the TLCanalysis (2) planktonic cells displayed the lowest GPL pro-duction
With regard to planktonic cells the rate ofM colombiense19B gene transcription was consistently higher than thatfor the M colombiense 57B genes Conversely the roughvariant (57B) displayed a dramatic decrease in the rate oftranscription for the 9 selected genes particularly thoseencoding peptide synthetase pstA glycosyltransferase gtfA
BioMed Research International 7
1 2 3 41 2 3 4
GPLs
PG
PIMs
(a) (b)
Figure 5 TLC profile of the GPLs in Mycobacterium colombiense strains Crude lipid extracts of (1) M avium 104 (2) M colombiense19B (3) M colombiense CECT 3035 and (4) M colombiense 57B Cells cultivated (a) under planktonic and (b) motile conditions weredeveloped using chloroform-methanol-water (65 25 4 vvv) GPLs glycopeptidolipids PIMs phosphatidyl-inositol mannosides and PGphosphatidylglycerol
the methyltransferases mtfB and mtfC and the lipid trans-porters tmtpA tmtpB and tmtpC (Figure 7(a)) InterestinglyrtfA (rhamnosyltransferase) in the 19B strain (smooth colonymorphology) was the only gene among the strains culturedunder planktonic conditions that showed an increased tran-scription rate (221-fold higher) compared with the controlcells (planktonicM colombiense CECT 3035 at the exponen-tial phase of growth)
Under motile growth conditions M colombiense 19Bshowed the highest transcription rate for all selected genescompared with the CECT 3035 and 57B strains (Figure 7(b))In contrast the transcription rates for all genes in the57B strain (rough variant) except tmtpA and tmtpC (lipidtransporters) were reduced In motile CECT 3035 cellswhereas rtfA mtfB (methyl transfers) and tmtpA showedincreased transcription the pstA gftA mtfC mtfD tmtpBand tmtpC genes exhibited reduced transcription comparedwith control cells
4 Discussion
In the present study TLC experiments were able to show thatM colombiense contains glycolipids with chromatographicbehaviour similar to the GPLs of M avium 104 which havebeen previously identified and completely characterised [23]in addition the colony morphology was associated withthe GPL profile of M colombiense strains Thus the TLCanalysis revealed that (1) strains displaying smooth colonymorphology (19B and CECT3035) produce a higher amountof variable GPLs compared with the rough natural variant(57B) strain and (2) the GPL production was augmentedin M colombiense cells cultivated under the motile growthconditions As expected the TLC analyses did not provide
detailed information about GPL structure however struc-tural determinationwas not the objective of the present study
Themedia composition influences the glycolipid contentof the mycobacterial cell wall The MAC strains growing onsolid medium exhibit a significant rate of transition fromsmooth transparent to smooth opaque and from smooth torough morphologies [18 19 38] We compared planktonicand motile M colombiense cells grown in the same culturemedium (7H9-ADC or 7H9-ADC-agarose) to avoid differ-ences in lipid content produced through alterations in themedia composition The results showed that the low GPLcontent observed in the rough 57B variant is less frequentamong M colombiense strains therefore M colombienseshows a preference for augmented GPL production andsmooth colony morphology Interestingly planktonic Mcolombiense cells displayed increased PIM and PG pro-duction and diminished GPL content compared with cellscultivated under motile conditions suggesting a potentialcompensatory mechanism in cell wall lipid synthesis thatcompensates for the low GPL content
The abundance of GPLs in the outermost portion ofthe cell envelope could be associated with the motility ofM colombiense strains on hydrophilic surfaces Recht andothers [22] proposed that GPLs are linked through thehydrophilic head to the cellular capsule and the hydrophobicfatty acid chain is exposed to the bacterial surface therebyreducing interactions with hydrophilic agarose surfaces andfacilitating the spread of cells on solid medium [21 22]Thus the augmented GPL production of motile 19B andCECT 3035 cells compared with that of the rough variant57B is consistent with the sliding motility on the hydrophilicagarose surface observed for these strains It has also beensuggested that bacterial sliding motility might be favoured
8 BioMed Research International
33kb 2kb9kb51kb
Contig 00001
600kb
87kb 245kb
488kb 36kb320kb
Contig 00002A
Contig 00003A
584kb 452kb
mmpL10
326kb
Contig 00007
fadE
15
atf
rmlB
ecf
rv1174
rmlA
mmpL
4a
mmpL
4b
mmpS4
mbtH
gap rv0926
sap
pks papA3
gtfA
mtfB
mtfC
mtfD
rtfA
pstA
pstB
(a)
ecf
fadE
5
rv1174
atf
rmlB
rmlA
mmpL
4a
mmpL
4b
mmpS
4
mbtH
gap
237kb
rv0926
7kb90kb
530kb
300kb
738kb
mmpL10
sap
Saccharides biosynthesis transfer and modification
Fatty acyl chain biosynthesis modification and transfer
Members of the mmpSL family
Peptide synthesis Unknown
Possible regulatory genes
GPL export to the cell surface
pks pe fadD23 gap-likepapA3
mtfB
gtfA
mtfC
mtfD
dhgA
rtfA
pstA
pstB
mtfA
gtfB
gtfD
(b)
Figure 6Model showing the gene cluster proposed for the GPL biosynthesis ofMycobacterium colombienseCECT 3035 (a)The location andidentity of theM colombiense genes were established using the previously described biosynthetic pathway for GPL biosynthesis annotated intheM avium 104 genomic sequence (b)
0001
001
01
1
10
Rela
tive g
ene t
rans
crip
tion
M colombiense 57BM colombiense 19B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(a)
01
1
10
100
Rela
tive g
ene t
rans
crip
tion
M colombiense CECT 3035
M colombiense 19B
M colombiense 57B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(b)
Figure 7 In vitro transcription of Mycobacterium colombiense genes involved in GPL biosynthesis Relative quantification expressed as theratio of gene transcription in (a) M colombiense cells grown under planktonic conditionstranscription ofM colombiense CECT 3035 cellsgrown under planktonic conditions and (b)M colombiense cells grown on the motility mediumtranscription ofM colombiense CECT 3035grown under planktonic conditions The presented data have statistically significant differences compared with the values obtained fromMcolombiense CECT 3035 cells (119875 lt 005)
BioMed Research International 9
through the secretion of surfactant substances from cells[21 22] The results of the drop-collapsing test showed thatbiosurfactants were likely not secreted from M colombiensecells suggesting that the motility of M colombiense couldbe favoured through hydrophobic molecules such as GPLsbound to the outermost portion of cells however we cannotcompletely rule out the possibility that some of the surfactantsubstances secreted from M colombiense remained adheredto the mycobacterial cell surface thereby influencing cellmotility
GPLs on the outermost portions of the M colombiensecell envelope generate a more hydrophobic cell surfacethat facilitates initial interactions with hydrophobic surfacesand increase biofilm formation on polystyrene wells Inaddition the increased GPL production in 19B and CECT3035 cells is consistent with the augmented biofilm forma-tion observed for M colombiense strains displaying smoothcolony morphology Moreover increased biofilm formationon hydrophobic surfaces and enhanced sliding over motilityplates for 19B cells compared with the CECT 3035 strainwhich also displays smooth colony morphology likely sug-gest possible differences in GPL structure betweenM colom-biense strains Differences inGPL production between strainscould influence the M colombiense ability to form biofilmson polyvinylchloride (PVC) pipes which would facilitatethe dissemination of these bacteria in natural environmentssuch as in-hospital spaces thereby increasing the chance ofinfection in immunosuppressed patients [23 39 40]
Based on the GPL biosynthetic pathway previouslydescribed for the M avium 104 strain [25] we are currentlyconstructing a potential gene cluster for GPL biosynthesis inM colombiense which will be finished when the nucleotidesequence of the CECT 3035 is completely assembled RT-qPCR experiments showed that the planktonic and motilecells of the rough 57B strain showed decreased transcriptionof the 9 selected genes leading to the low production ofGPLs in this rough variant comparedwith theM colombiensestrains displaying smooth colony morphology Interestinglyplanktonic 19B cells exhibited lower expression of most of theselected genes compared with planktonic CECT 3035 cellsThis behaviour results in the reduced production of GPLs inplanktonic 19B cells compared with the CECT 3035 strainconsistent with the results of the TLC experiments
It has been previously shown that an rtfA mutation inthe M avium serovar-2-specific strain resulted in the lossof serovar-specific GPLs thereby diminishing the variabilityof these glycolipids in the cell envelope [41 42] Thus itis possible that the enzymatic activity of the RtfA proteincould be relevant for the increased glycosylation andor aug-mented production of ssGPLs in strains displaying smoothcolony morphology compared with the rough variant 57BIn strains displaying smooth colony morphology the genesencoding rhamnosyl andmethyl transferases (rtfA andmtfB)are overtranscribed suggesting the increased production ofnsGPLs precursors for ssGPL biosynthesis [24ndash26] therebyincreasing the production of GPLs The diminished tran-scription of pstA gftA mtfC mtfD tmtpBand tmtpC inmotile CECT 3035 compared with control cells (planktonicCECT 3035) is intriguing however this behaviour also
suggests the relevance of rhamnosyl transferases particularlyRtfA in GPL biosynthesis in M colombiense Neverthelessthe actual role for RtfA in M colombiense strains should beconfirmed using rtfA-null mutants
We did not identify 7 of the 30 known GPL cluster genesof M avium 104 in the genome sequence of M colombienseCECT 3035 This interesting observation suggests that theGPLs between these two closely related MAC species couldhave different structural characteristics However furtherexperiments using mass spectrometry and NMR are neces-sary to evaluate potential differences in the GPL structureamongM colombiense strains
5 Conclusions
In conclusion the results of the present study show that Mcolombiense strains displaying smooth morphology exhibitincreased biofilm formation on hydrophobic polystyrenesurfaces and enhanced sliding over motility plates Bioin-formatics analyses indicate that the gene cluster establishedfor GPL formation modification and translocation is intactstrongly suggesting that GPLs are putatively present in Mcolombiense In addition motile culture conditions activatethe transcription of the genes implicated in the key enzymaticactivities of GPL biosynthesis
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
Thisworkwas supported through funding from the ldquoDivisionde Investigacion Bogota (DIB)rdquo-Vicerrectorıa de Investi-gacion Universidad Nacional de Colombia (Grants 1606014837 and 14337) Milena Maya-Hoyos received a fellowshipfrom the Academic Direction Universidad Nacional deColombia and the ldquoJovenes Investigadores e InnovadoresrdquoProgram Colciencias Colombia This research was alsosupported in part through the Intramural Research Programof the National Institutes of Health National Library ofMedicine and National Center for Biotechnology Informa-tion (NCBI)
References
[1] S K Field D Fisher and R L Cowie ldquoMycobacterium aviumcomplex pulmonary disease in patient without HIV infectionrdquoChest vol 126 no 2 pp 566ndash581 2004
[2] M J M Vaerewijck G Huys J C Palomino J Swings and FPortaels ldquoMycobacteria in drinking water distribution systemsecology and significance for human healthrdquo FEMSMicrobiologyReviews vol 29 no 5 pp 911ndash934 2005
[3] E Hagiwara S Komatsu R Nishihira T Shinohara T Babaand T Ogura ldquoClinical characteristics and prevalence of pneu-mothorax in patients with pulmonary Mycobacterium aviumcomplex diseaserdquo Journal of Infection and Chemotherapy vol19 no 4 pp 588ndash592 2013
10 BioMed Research International
[4] M I Murcia E Tortoli M C Menendez E Palenque andM J Garcia ldquoMycobacterium colombiense sp nov a novelmember of theMycobacterium avium complex and descriptionof MAC-X as a new ITS genetic variantrdquo International Journalof Systematic and Evolutionary Microbiology vol 56 no 9 pp2049ndash2054 2006
[5] E Tortoli L Rindi M J Garcia et al ldquoProposal to elevatethe genetic variant MAC-A included in the Mycobacteriumavium complex to species rank as Mycobacterium chimaerasp novrdquo International Journal of Systematic and EvolutionaryMicrobiology vol 54 no 4 pp 1277ndash1285 2004
[6] I B Salah C Cayrou D Raoult and M Drancourt ldquoMycobac-terium marseillense sp nov Mycobacterium timonense sp novand Mycobacterium bouchedurhonense sp novmembers ofthe Mycobacterium avium complexrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 11 pp2803ndash2808 2009
[7] J van Ingen M J Boeree K Kosters et al ldquoProposal toelevate Mycobacterium avium complex ITS sequevar MAC-Qto Mycobacterium vulneris sp novrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 9 pp2277ndash2282 2009
[8] D Bang T Herlin M Stegger et al ldquoMycobacterium arosiensesp nov a slowly growing scotochromogenic species causingosteomyelitis in an immunocompromised childrdquo InternationalJournal of Systematic and Evolutionary Microbiology vol 58part 10 pp 2398ndash2402 2008
[9] C Cayrou C Turenne M A Behr and M Drancourt ldquoGeno-typing of Mycobacterium avium complex organisms usingmultispacer sequence typingrdquoMicrobiology vol 156 part 3 pp687ndash694 2010
[10] S C Smole F McAleese J Ngampasutadol C F von Reynand R D Arbeit ldquoClinical and epidemiological correlates ofgenotypes within theMycobacterium avium complex defined byrestriction and sequence analysis of hsp65rdquo Journal of ClinicalMicrobiology vol 40 no 9 pp 3374ndash3380 2002
[11] AMuwonge C Kankya T B Johansen et al ldquoNon-tuberculousmycobacteria isolated from slaughter pigs in Mubende districtUgandardquo BMC Veterinary Research vol 8 article 52 2012
[12] J O Falkinham III ldquoSurrounded by mycobacteria nontuber-culous mycobacteria in the human environmentrdquo Journal ofApplied Microbiology vol 107 no 2 pp 356ndash367 2009
[13] K Vuorenmaa I B Salah V Barlogis H Chambost and MDrancourt ldquoMycobacterium colombiense and pseudotubercu-lous lymphadenopathyrdquo Emerging Infectious Diseases vol 15no 4 pp 618ndash620 2009
[14] O Esparcia F Navarro M Quer and P Coll ldquoLymphadenopa-thy caused by Mycobacterium colombienserdquo Journal of ClinicalMicrobiology vol 46 no 5 pp 1885ndash1887 2008
[15] S Cohen-Bacrie M David N Stremler J-C Dubus J-MRolain and M Drancourt ldquoMycobacterium chimaera pul-monary infection complicating cystic fibrosis a case reportrdquoJournal of Medical Case Reports vol 5 no 1 article 473 2011
[16] S Poulin C Corbeil M Nguyen et al ldquoFatal Mycobac-terium colombiensecytomegalovirus coinfection associatedwith acquired immunodeficiency due to autoantibodies againstinterferon gamma a case reportrdquo BMC Infectious Diseases vol13 no 1 article 24 2013
[17] J Leguizamon J Hernandez M-I Murcia and C-Y SotoldquoIdentification of potential biomarkers to distinguishMycobac-terium colombiense from other mycobacterial speciesrdquoMolecu-lar and Cellular Probes vol 27 no 1 pp 46ndash52 2013
[18] D Chatterjee and K H Khoo ldquoThe surface glycopeptidolipidsof mycobacteria structures and biological propertiesrdquo Cellularand Molecular Life Sciences vol 58 no 14 pp 2018ndash2042 2001
[19] J B Torrelles D Ellis T Osborne et al ldquoCharacterizationof virulence colony morphotype and the glycopeptidolipid ofMycobacterium avium strain 104rdquo Tuberculosis vol 82 no 6pp 293ndash300 2002
[20] M F Thorel and H L David ldquoSpecific surface antigens of SmTvariants ofMycobacterium aviumrdquo Infection and Immunity vol43 no 1 pp 438ndash439 1984
[21] A Martınez S Torello and R Kolter ldquoSliding motility inmycobacteriardquo Journal of Bacteriology vol 181 no 23 pp 7331ndash7338 1999
[22] J Recht AMartınez S Torello and R Kolter ldquoGenetic analysisof sliding motility in Mycobacterium smegmatisrdquo Journal ofBacteriology vol 182 no 15 pp 4348ndash4351 2000
[23] G Carter M Wu D C Drummond and L E BermudezldquoCharacterization of biofilm formation by clinical isolates ofMycobacterium aviumrdquo Journal of Medical Microbiology vol 52no 9 pp 747ndash752 2003
[24] J S Schorey and L Sweet ldquoThe mycobacterial glycopepti-dolipids structure function and their role in pathogenesisrdquoGlycobiology vol 18 no 11 pp 832ndash841 2008
[25] L Pang X Tian W Pan and J Xie ldquoStructure and function ofmycobacterium glycopeptidolipids from comparative genomicsperspectiverdquo Journal of Cellular Biochemistry vol 114 no 8 pp1705ndash1713 2013
[26] R Mukherjee and D Chatterji ldquoGlycopeptidolipids immuno-modulators in greasy mycobacterial cell enveloperdquo IUBMB Lifevol 64 no 3 pp 215ndash225 2012
[27] K L Horan R Freeman K Weigel et al ldquoIsolation ofthe genome sequence strain Mycobacterium avium 104 frommultiple patients over a 17-year periodrdquo Journal of ClinicalMicrobiology vol 44 no 3 pp 783ndash789 2006
[28] S B Snapper R E Melton S Mustafa T Kieser and W RJacobs Jr ldquoIsolation and characterization of efficient plasmidtransformation mutants of Mycobacterium smegmatisrdquo Molec-ular Microbiology vol 4 no 11 pp 1911ndash1919 1990
[29] M Radji R A Agustama B Elya and C R TjampakasarildquoAntimicrobial activity of green tea extract against isolatesof methicillin-resistant Staphylococcus aureus and multi-drugresistant Pseudomonas aeruginosardquo Asian Pacific Journal ofTropical Biomedicine vol 3 no 8 pp 663ndash667 2013
[30] P Helden T Victor R Warren and E Helden ldquoIsolationof DNA from Mycobacterium tuberculosisrdquo in Mycobacteriumtuberculosis Protocols Humana Press Totowa NJ USA 2001
[31] J C van Kessel L J Marinelli and G F Hatfull ldquoRecom-bineering mycobacteria and their phagesrdquo Nature ReviewsMicrobiology vol 6 no 11 pp 851ndash857 2008
[32] N M Parrish C G Ko J D Dick P B Jones and J LE Ellingson ldquoGrowth Congo Red agar colony morphotypesand antibiotic susceptibility testing of Mycobacterium aviumsubspecies paratuberculosisrdquo Clin Med Res vol 2 no 2 pp107ndash114 2004
[33] T Tugrul and E Cansunar ldquoDetecting surfactant-producingmicroorganisms by the drop-collapse testrdquo World Journal ofMicrobiology and Biotechnology vol 21 no 6-7 pp 851ndash8532005
[34] D K Jain D L Collins-Thompson H Lee and J T TrevorsldquoA drop-collapsing test for screening surfactant-producingmicroorganismsrdquo Journal ofMicrobiologicalMethods vol 13 no4 pp 271ndash279 1991
BioMed Research International 11
[35] M Munoz M-A Laneelle M Luquin et al ldquoOccurrence ofan antigenic triacyl trehalose in clinical isolates and referencestrains of Mycobacterium tuberculosisrdquo FEMS MicrobiologyLetters vol 157 no 2 pp 251ndash259 1997
[36] M Gonzalez-Perez M I Murcia D Landsman I K Jordanand L Marino-Ramırez ldquoGenome sequence of the Mycobac-terium colombiense type strain CECT 3035rdquo Journal of Bacte-riology vol 193 no 20 pp 5866ndash5867 2011
[37] J C Betts P T Lukey L C Robb R A McAdam andK Duncan ldquoEvaluation of a nutrient starvation model ofMycobacterium tuberculosis persistence by gene and proteinexpression profilingrdquoMolecular Microbiology vol 43 no 3 pp717ndash731 2002
[38] W W Barrow and P J Brennan ldquoIsolation in high frequencyof rough variants of Mycobacterium intracellulare lacking C-mycoside glycopeptidolipid antigensrdquo Journal of Bacteriologyvol 150 no 1 pp 381ndash384 1982
[39] W Costerton R Veeh M Shirtliff M Pasmore C Post andG Ehrlich ldquoThe application of biofilm science to the studyand control of chronic bacterial infectionsrdquo Journal of ClinicalInvestigation vol 112 no 10 pp 1466ndash1477 2003
[40] J Recht and R Kolter ldquoGlycopeptidolipid acetylation affectssliding motility and biofilm formation inMycobacterium smeg-matisrdquo Journal of Bacteriology vol 183 no 19 pp 5718ndash57242001
[41] T M Eckstein F S Silbaq D Chatterjee N J Kelly P JBrennan and J T Belisle ldquoIdentification and recombinantexpression of a Mycobacterium avium rhamnosyltransferasegene (rtfA) involved in glycopeptidolipid biosynthesisrdquo Journalof Bacteriology vol 180 no 21 pp 5567ndash5573 1998
[42] J N Maslow V R Irani S-H Lee T M Eckstein J MInamine and J T Belisle ldquoBiosynthetic specificity of therhamnosyltransferase gene of Mycobacterium avium serovar 2as determined by allelic exchange mutagenesisrdquo Microbiologyvol 149 no 11 pp 3193ndash3202 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
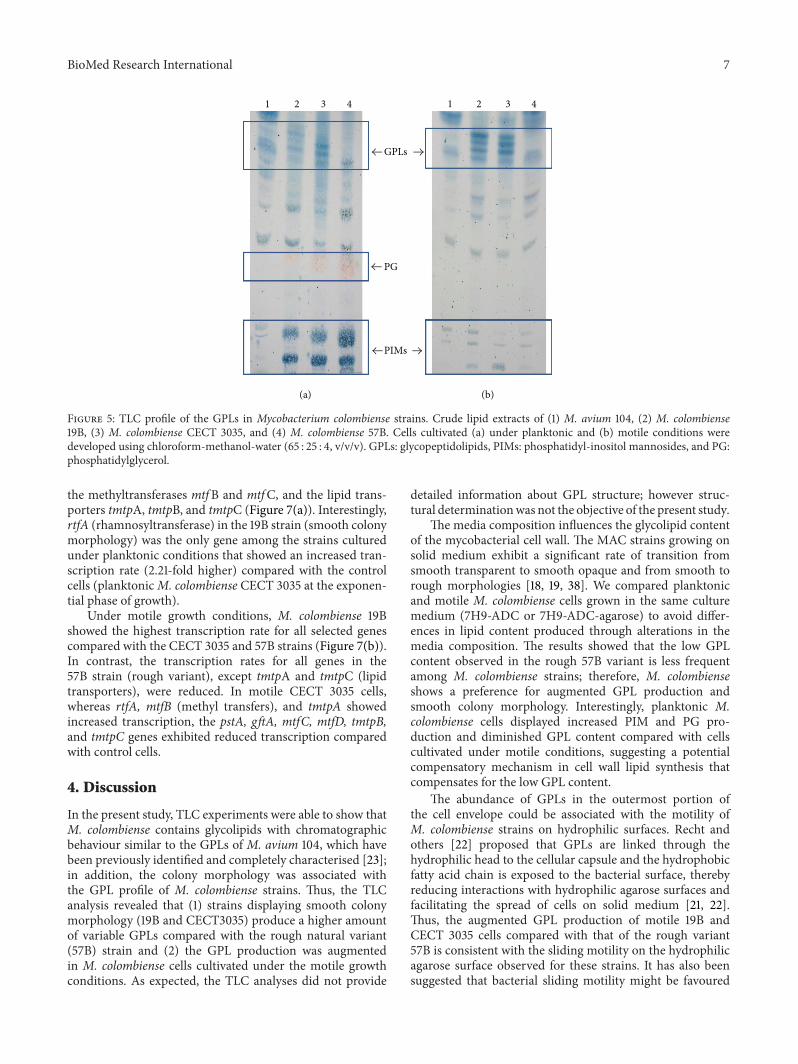
BioMed Research International 7
1 2 3 41 2 3 4
GPLs
PG
PIMs
(a) (b)
Figure 5 TLC profile of the GPLs in Mycobacterium colombiense strains Crude lipid extracts of (1) M avium 104 (2) M colombiense19B (3) M colombiense CECT 3035 and (4) M colombiense 57B Cells cultivated (a) under planktonic and (b) motile conditions weredeveloped using chloroform-methanol-water (65 25 4 vvv) GPLs glycopeptidolipids PIMs phosphatidyl-inositol mannosides and PGphosphatidylglycerol
the methyltransferases mtfB and mtfC and the lipid trans-porters tmtpA tmtpB and tmtpC (Figure 7(a)) InterestinglyrtfA (rhamnosyltransferase) in the 19B strain (smooth colonymorphology) was the only gene among the strains culturedunder planktonic conditions that showed an increased tran-scription rate (221-fold higher) compared with the controlcells (planktonicM colombiense CECT 3035 at the exponen-tial phase of growth)
Under motile growth conditions M colombiense 19Bshowed the highest transcription rate for all selected genescompared with the CECT 3035 and 57B strains (Figure 7(b))In contrast the transcription rates for all genes in the57B strain (rough variant) except tmtpA and tmtpC (lipidtransporters) were reduced In motile CECT 3035 cellswhereas rtfA mtfB (methyl transfers) and tmtpA showedincreased transcription the pstA gftA mtfC mtfD tmtpBand tmtpC genes exhibited reduced transcription comparedwith control cells
4 Discussion
In the present study TLC experiments were able to show thatM colombiense contains glycolipids with chromatographicbehaviour similar to the GPLs of M avium 104 which havebeen previously identified and completely characterised [23]in addition the colony morphology was associated withthe GPL profile of M colombiense strains Thus the TLCanalysis revealed that (1) strains displaying smooth colonymorphology (19B and CECT3035) produce a higher amountof variable GPLs compared with the rough natural variant(57B) strain and (2) the GPL production was augmentedin M colombiense cells cultivated under the motile growthconditions As expected the TLC analyses did not provide
detailed information about GPL structure however struc-tural determinationwas not the objective of the present study
Themedia composition influences the glycolipid contentof the mycobacterial cell wall The MAC strains growing onsolid medium exhibit a significant rate of transition fromsmooth transparent to smooth opaque and from smooth torough morphologies [18 19 38] We compared planktonicand motile M colombiense cells grown in the same culturemedium (7H9-ADC or 7H9-ADC-agarose) to avoid differ-ences in lipid content produced through alterations in themedia composition The results showed that the low GPLcontent observed in the rough 57B variant is less frequentamong M colombiense strains therefore M colombienseshows a preference for augmented GPL production andsmooth colony morphology Interestingly planktonic Mcolombiense cells displayed increased PIM and PG pro-duction and diminished GPL content compared with cellscultivated under motile conditions suggesting a potentialcompensatory mechanism in cell wall lipid synthesis thatcompensates for the low GPL content
The abundance of GPLs in the outermost portion ofthe cell envelope could be associated with the motility ofM colombiense strains on hydrophilic surfaces Recht andothers [22] proposed that GPLs are linked through thehydrophilic head to the cellular capsule and the hydrophobicfatty acid chain is exposed to the bacterial surface therebyreducing interactions with hydrophilic agarose surfaces andfacilitating the spread of cells on solid medium [21 22]Thus the augmented GPL production of motile 19B andCECT 3035 cells compared with that of the rough variant57B is consistent with the sliding motility on the hydrophilicagarose surface observed for these strains It has also beensuggested that bacterial sliding motility might be favoured
8 BioMed Research International
33kb 2kb9kb51kb
Contig 00001
600kb
87kb 245kb
488kb 36kb320kb
Contig 00002A
Contig 00003A
584kb 452kb
mmpL10
326kb
Contig 00007
fadE
15
atf
rmlB
ecf
rv1174
rmlA
mmpL
4a
mmpL
4b
mmpS4
mbtH
gap rv0926
sap
pks papA3
gtfA
mtfB
mtfC
mtfD
rtfA
pstA
pstB
(a)
ecf
fadE
5
rv1174
atf
rmlB
rmlA
mmpL
4a
mmpL
4b
mmpS
4
mbtH
gap
237kb
rv0926
7kb90kb
530kb
300kb
738kb
mmpL10
sap
Saccharides biosynthesis transfer and modification
Fatty acyl chain biosynthesis modification and transfer
Members of the mmpSL family
Peptide synthesis Unknown
Possible regulatory genes
GPL export to the cell surface
pks pe fadD23 gap-likepapA3
mtfB
gtfA
mtfC
mtfD
dhgA
rtfA
pstA
pstB
mtfA
gtfB
gtfD
(b)
Figure 6Model showing the gene cluster proposed for the GPL biosynthesis ofMycobacterium colombienseCECT 3035 (a)The location andidentity of theM colombiense genes were established using the previously described biosynthetic pathway for GPL biosynthesis annotated intheM avium 104 genomic sequence (b)
0001
001
01
1
10
Rela
tive g
ene t
rans
crip
tion
M colombiense 57BM colombiense 19B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(a)
01
1
10
100
Rela
tive g
ene t
rans
crip
tion
M colombiense CECT 3035
M colombiense 19B
M colombiense 57B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(b)
Figure 7 In vitro transcription of Mycobacterium colombiense genes involved in GPL biosynthesis Relative quantification expressed as theratio of gene transcription in (a) M colombiense cells grown under planktonic conditionstranscription ofM colombiense CECT 3035 cellsgrown under planktonic conditions and (b)M colombiense cells grown on the motility mediumtranscription ofM colombiense CECT 3035grown under planktonic conditions The presented data have statistically significant differences compared with the values obtained fromMcolombiense CECT 3035 cells (119875 lt 005)
BioMed Research International 9
through the secretion of surfactant substances from cells[21 22] The results of the drop-collapsing test showed thatbiosurfactants were likely not secreted from M colombiensecells suggesting that the motility of M colombiense couldbe favoured through hydrophobic molecules such as GPLsbound to the outermost portion of cells however we cannotcompletely rule out the possibility that some of the surfactantsubstances secreted from M colombiense remained adheredto the mycobacterial cell surface thereby influencing cellmotility
GPLs on the outermost portions of the M colombiensecell envelope generate a more hydrophobic cell surfacethat facilitates initial interactions with hydrophobic surfacesand increase biofilm formation on polystyrene wells Inaddition the increased GPL production in 19B and CECT3035 cells is consistent with the augmented biofilm forma-tion observed for M colombiense strains displaying smoothcolony morphology Moreover increased biofilm formationon hydrophobic surfaces and enhanced sliding over motilityplates for 19B cells compared with the CECT 3035 strainwhich also displays smooth colony morphology likely sug-gest possible differences in GPL structure betweenM colom-biense strains Differences inGPL production between strainscould influence the M colombiense ability to form biofilmson polyvinylchloride (PVC) pipes which would facilitatethe dissemination of these bacteria in natural environmentssuch as in-hospital spaces thereby increasing the chance ofinfection in immunosuppressed patients [23 39 40]
Based on the GPL biosynthetic pathway previouslydescribed for the M avium 104 strain [25] we are currentlyconstructing a potential gene cluster for GPL biosynthesis inM colombiense which will be finished when the nucleotidesequence of the CECT 3035 is completely assembled RT-qPCR experiments showed that the planktonic and motilecells of the rough 57B strain showed decreased transcriptionof the 9 selected genes leading to the low production ofGPLs in this rough variant comparedwith theM colombiensestrains displaying smooth colony morphology Interestinglyplanktonic 19B cells exhibited lower expression of most of theselected genes compared with planktonic CECT 3035 cellsThis behaviour results in the reduced production of GPLs inplanktonic 19B cells compared with the CECT 3035 strainconsistent with the results of the TLC experiments
It has been previously shown that an rtfA mutation inthe M avium serovar-2-specific strain resulted in the lossof serovar-specific GPLs thereby diminishing the variabilityof these glycolipids in the cell envelope [41 42] Thus itis possible that the enzymatic activity of the RtfA proteincould be relevant for the increased glycosylation andor aug-mented production of ssGPLs in strains displaying smoothcolony morphology compared with the rough variant 57BIn strains displaying smooth colony morphology the genesencoding rhamnosyl andmethyl transferases (rtfA andmtfB)are overtranscribed suggesting the increased production ofnsGPLs precursors for ssGPL biosynthesis [24ndash26] therebyincreasing the production of GPLs The diminished tran-scription of pstA gftA mtfC mtfD tmtpBand tmtpC inmotile CECT 3035 compared with control cells (planktonicCECT 3035) is intriguing however this behaviour also
suggests the relevance of rhamnosyl transferases particularlyRtfA in GPL biosynthesis in M colombiense Neverthelessthe actual role for RtfA in M colombiense strains should beconfirmed using rtfA-null mutants
We did not identify 7 of the 30 known GPL cluster genesof M avium 104 in the genome sequence of M colombienseCECT 3035 This interesting observation suggests that theGPLs between these two closely related MAC species couldhave different structural characteristics However furtherexperiments using mass spectrometry and NMR are neces-sary to evaluate potential differences in the GPL structureamongM colombiense strains
5 Conclusions
In conclusion the results of the present study show that Mcolombiense strains displaying smooth morphology exhibitincreased biofilm formation on hydrophobic polystyrenesurfaces and enhanced sliding over motility plates Bioin-formatics analyses indicate that the gene cluster establishedfor GPL formation modification and translocation is intactstrongly suggesting that GPLs are putatively present in Mcolombiense In addition motile culture conditions activatethe transcription of the genes implicated in the key enzymaticactivities of GPL biosynthesis
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
Thisworkwas supported through funding from the ldquoDivisionde Investigacion Bogota (DIB)rdquo-Vicerrectorıa de Investi-gacion Universidad Nacional de Colombia (Grants 1606014837 and 14337) Milena Maya-Hoyos received a fellowshipfrom the Academic Direction Universidad Nacional deColombia and the ldquoJovenes Investigadores e InnovadoresrdquoProgram Colciencias Colombia This research was alsosupported in part through the Intramural Research Programof the National Institutes of Health National Library ofMedicine and National Center for Biotechnology Informa-tion (NCBI)
References
[1] S K Field D Fisher and R L Cowie ldquoMycobacterium aviumcomplex pulmonary disease in patient without HIV infectionrdquoChest vol 126 no 2 pp 566ndash581 2004
[2] M J M Vaerewijck G Huys J C Palomino J Swings and FPortaels ldquoMycobacteria in drinking water distribution systemsecology and significance for human healthrdquo FEMSMicrobiologyReviews vol 29 no 5 pp 911ndash934 2005
[3] E Hagiwara S Komatsu R Nishihira T Shinohara T Babaand T Ogura ldquoClinical characteristics and prevalence of pneu-mothorax in patients with pulmonary Mycobacterium aviumcomplex diseaserdquo Journal of Infection and Chemotherapy vol19 no 4 pp 588ndash592 2013
10 BioMed Research International
[4] M I Murcia E Tortoli M C Menendez E Palenque andM J Garcia ldquoMycobacterium colombiense sp nov a novelmember of theMycobacterium avium complex and descriptionof MAC-X as a new ITS genetic variantrdquo International Journalof Systematic and Evolutionary Microbiology vol 56 no 9 pp2049ndash2054 2006
[5] E Tortoli L Rindi M J Garcia et al ldquoProposal to elevatethe genetic variant MAC-A included in the Mycobacteriumavium complex to species rank as Mycobacterium chimaerasp novrdquo International Journal of Systematic and EvolutionaryMicrobiology vol 54 no 4 pp 1277ndash1285 2004
[6] I B Salah C Cayrou D Raoult and M Drancourt ldquoMycobac-terium marseillense sp nov Mycobacterium timonense sp novand Mycobacterium bouchedurhonense sp novmembers ofthe Mycobacterium avium complexrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 11 pp2803ndash2808 2009
[7] J van Ingen M J Boeree K Kosters et al ldquoProposal toelevate Mycobacterium avium complex ITS sequevar MAC-Qto Mycobacterium vulneris sp novrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 9 pp2277ndash2282 2009
[8] D Bang T Herlin M Stegger et al ldquoMycobacterium arosiensesp nov a slowly growing scotochromogenic species causingosteomyelitis in an immunocompromised childrdquo InternationalJournal of Systematic and Evolutionary Microbiology vol 58part 10 pp 2398ndash2402 2008
[9] C Cayrou C Turenne M A Behr and M Drancourt ldquoGeno-typing of Mycobacterium avium complex organisms usingmultispacer sequence typingrdquoMicrobiology vol 156 part 3 pp687ndash694 2010
[10] S C Smole F McAleese J Ngampasutadol C F von Reynand R D Arbeit ldquoClinical and epidemiological correlates ofgenotypes within theMycobacterium avium complex defined byrestriction and sequence analysis of hsp65rdquo Journal of ClinicalMicrobiology vol 40 no 9 pp 3374ndash3380 2002
[11] AMuwonge C Kankya T B Johansen et al ldquoNon-tuberculousmycobacteria isolated from slaughter pigs in Mubende districtUgandardquo BMC Veterinary Research vol 8 article 52 2012
[12] J O Falkinham III ldquoSurrounded by mycobacteria nontuber-culous mycobacteria in the human environmentrdquo Journal ofApplied Microbiology vol 107 no 2 pp 356ndash367 2009
[13] K Vuorenmaa I B Salah V Barlogis H Chambost and MDrancourt ldquoMycobacterium colombiense and pseudotubercu-lous lymphadenopathyrdquo Emerging Infectious Diseases vol 15no 4 pp 618ndash620 2009
[14] O Esparcia F Navarro M Quer and P Coll ldquoLymphadenopa-thy caused by Mycobacterium colombienserdquo Journal of ClinicalMicrobiology vol 46 no 5 pp 1885ndash1887 2008
[15] S Cohen-Bacrie M David N Stremler J-C Dubus J-MRolain and M Drancourt ldquoMycobacterium chimaera pul-monary infection complicating cystic fibrosis a case reportrdquoJournal of Medical Case Reports vol 5 no 1 article 473 2011
[16] S Poulin C Corbeil M Nguyen et al ldquoFatal Mycobac-terium colombiensecytomegalovirus coinfection associatedwith acquired immunodeficiency due to autoantibodies againstinterferon gamma a case reportrdquo BMC Infectious Diseases vol13 no 1 article 24 2013
[17] J Leguizamon J Hernandez M-I Murcia and C-Y SotoldquoIdentification of potential biomarkers to distinguishMycobac-terium colombiense from other mycobacterial speciesrdquoMolecu-lar and Cellular Probes vol 27 no 1 pp 46ndash52 2013
[18] D Chatterjee and K H Khoo ldquoThe surface glycopeptidolipidsof mycobacteria structures and biological propertiesrdquo Cellularand Molecular Life Sciences vol 58 no 14 pp 2018ndash2042 2001
[19] J B Torrelles D Ellis T Osborne et al ldquoCharacterizationof virulence colony morphotype and the glycopeptidolipid ofMycobacterium avium strain 104rdquo Tuberculosis vol 82 no 6pp 293ndash300 2002
[20] M F Thorel and H L David ldquoSpecific surface antigens of SmTvariants ofMycobacterium aviumrdquo Infection and Immunity vol43 no 1 pp 438ndash439 1984
[21] A Martınez S Torello and R Kolter ldquoSliding motility inmycobacteriardquo Journal of Bacteriology vol 181 no 23 pp 7331ndash7338 1999
[22] J Recht AMartınez S Torello and R Kolter ldquoGenetic analysisof sliding motility in Mycobacterium smegmatisrdquo Journal ofBacteriology vol 182 no 15 pp 4348ndash4351 2000
[23] G Carter M Wu D C Drummond and L E BermudezldquoCharacterization of biofilm formation by clinical isolates ofMycobacterium aviumrdquo Journal of Medical Microbiology vol 52no 9 pp 747ndash752 2003
[24] J S Schorey and L Sweet ldquoThe mycobacterial glycopepti-dolipids structure function and their role in pathogenesisrdquoGlycobiology vol 18 no 11 pp 832ndash841 2008
[25] L Pang X Tian W Pan and J Xie ldquoStructure and function ofmycobacterium glycopeptidolipids from comparative genomicsperspectiverdquo Journal of Cellular Biochemistry vol 114 no 8 pp1705ndash1713 2013
[26] R Mukherjee and D Chatterji ldquoGlycopeptidolipids immuno-modulators in greasy mycobacterial cell enveloperdquo IUBMB Lifevol 64 no 3 pp 215ndash225 2012
[27] K L Horan R Freeman K Weigel et al ldquoIsolation ofthe genome sequence strain Mycobacterium avium 104 frommultiple patients over a 17-year periodrdquo Journal of ClinicalMicrobiology vol 44 no 3 pp 783ndash789 2006
[28] S B Snapper R E Melton S Mustafa T Kieser and W RJacobs Jr ldquoIsolation and characterization of efficient plasmidtransformation mutants of Mycobacterium smegmatisrdquo Molec-ular Microbiology vol 4 no 11 pp 1911ndash1919 1990
[29] M Radji R A Agustama B Elya and C R TjampakasarildquoAntimicrobial activity of green tea extract against isolatesof methicillin-resistant Staphylococcus aureus and multi-drugresistant Pseudomonas aeruginosardquo Asian Pacific Journal ofTropical Biomedicine vol 3 no 8 pp 663ndash667 2013
[30] P Helden T Victor R Warren and E Helden ldquoIsolationof DNA from Mycobacterium tuberculosisrdquo in Mycobacteriumtuberculosis Protocols Humana Press Totowa NJ USA 2001
[31] J C van Kessel L J Marinelli and G F Hatfull ldquoRecom-bineering mycobacteria and their phagesrdquo Nature ReviewsMicrobiology vol 6 no 11 pp 851ndash857 2008
[32] N M Parrish C G Ko J D Dick P B Jones and J LE Ellingson ldquoGrowth Congo Red agar colony morphotypesand antibiotic susceptibility testing of Mycobacterium aviumsubspecies paratuberculosisrdquo Clin Med Res vol 2 no 2 pp107ndash114 2004
[33] T Tugrul and E Cansunar ldquoDetecting surfactant-producingmicroorganisms by the drop-collapse testrdquo World Journal ofMicrobiology and Biotechnology vol 21 no 6-7 pp 851ndash8532005
[34] D K Jain D L Collins-Thompson H Lee and J T TrevorsldquoA drop-collapsing test for screening surfactant-producingmicroorganismsrdquo Journal ofMicrobiologicalMethods vol 13 no4 pp 271ndash279 1991
BioMed Research International 11
[35] M Munoz M-A Laneelle M Luquin et al ldquoOccurrence ofan antigenic triacyl trehalose in clinical isolates and referencestrains of Mycobacterium tuberculosisrdquo FEMS MicrobiologyLetters vol 157 no 2 pp 251ndash259 1997
[36] M Gonzalez-Perez M I Murcia D Landsman I K Jordanand L Marino-Ramırez ldquoGenome sequence of the Mycobac-terium colombiense type strain CECT 3035rdquo Journal of Bacte-riology vol 193 no 20 pp 5866ndash5867 2011
[37] J C Betts P T Lukey L C Robb R A McAdam andK Duncan ldquoEvaluation of a nutrient starvation model ofMycobacterium tuberculosis persistence by gene and proteinexpression profilingrdquoMolecular Microbiology vol 43 no 3 pp717ndash731 2002
[38] W W Barrow and P J Brennan ldquoIsolation in high frequencyof rough variants of Mycobacterium intracellulare lacking C-mycoside glycopeptidolipid antigensrdquo Journal of Bacteriologyvol 150 no 1 pp 381ndash384 1982
[39] W Costerton R Veeh M Shirtliff M Pasmore C Post andG Ehrlich ldquoThe application of biofilm science to the studyand control of chronic bacterial infectionsrdquo Journal of ClinicalInvestigation vol 112 no 10 pp 1466ndash1477 2003
[40] J Recht and R Kolter ldquoGlycopeptidolipid acetylation affectssliding motility and biofilm formation inMycobacterium smeg-matisrdquo Journal of Bacteriology vol 183 no 19 pp 5718ndash57242001
[41] T M Eckstein F S Silbaq D Chatterjee N J Kelly P JBrennan and J T Belisle ldquoIdentification and recombinantexpression of a Mycobacterium avium rhamnosyltransferasegene (rtfA) involved in glycopeptidolipid biosynthesisrdquo Journalof Bacteriology vol 180 no 21 pp 5567ndash5573 1998
[42] J N Maslow V R Irani S-H Lee T M Eckstein J MInamine and J T Belisle ldquoBiosynthetic specificity of therhamnosyltransferase gene of Mycobacterium avium serovar 2as determined by allelic exchange mutagenesisrdquo Microbiologyvol 149 no 11 pp 3193ndash3202 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 BioMed Research International
33kb 2kb9kb51kb
Contig 00001
600kb
87kb 245kb
488kb 36kb320kb
Contig 00002A
Contig 00003A
584kb 452kb
mmpL10
326kb
Contig 00007
fadE
15
atf
rmlB
ecf
rv1174
rmlA
mmpL
4a
mmpL
4b
mmpS4
mbtH
gap rv0926
sap
pks papA3
gtfA
mtfB
mtfC
mtfD
rtfA
pstA
pstB
(a)
ecf
fadE
5
rv1174
atf
rmlB
rmlA
mmpL
4a
mmpL
4b
mmpS
4
mbtH
gap
237kb
rv0926
7kb90kb
530kb
300kb
738kb
mmpL10
sap
Saccharides biosynthesis transfer and modification
Fatty acyl chain biosynthesis modification and transfer
Members of the mmpSL family
Peptide synthesis Unknown
Possible regulatory genes
GPL export to the cell surface
pks pe fadD23 gap-likepapA3
mtfB
gtfA
mtfC
mtfD
dhgA
rtfA
pstA
pstB
mtfA
gtfB
gtfD
(b)
Figure 6Model showing the gene cluster proposed for the GPL biosynthesis ofMycobacterium colombienseCECT 3035 (a)The location andidentity of theM colombiense genes were established using the previously described biosynthetic pathway for GPL biosynthesis annotated intheM avium 104 genomic sequence (b)
0001
001
01
1
10
Rela
tive g
ene t
rans
crip
tion
M colombiense 57BM colombiense 19B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(a)
01
1
10
100
Rela
tive g
ene t
rans
crip
tion
M colombiense CECT 3035
M colombiense 19B
M colombiense 57B
pstA
gtfA
rtfA
mtfB
mtfC
mtfD
tmtpA
tmtpB
tmtpC
(b)
Figure 7 In vitro transcription of Mycobacterium colombiense genes involved in GPL biosynthesis Relative quantification expressed as theratio of gene transcription in (a) M colombiense cells grown under planktonic conditionstranscription ofM colombiense CECT 3035 cellsgrown under planktonic conditions and (b)M colombiense cells grown on the motility mediumtranscription ofM colombiense CECT 3035grown under planktonic conditions The presented data have statistically significant differences compared with the values obtained fromMcolombiense CECT 3035 cells (119875 lt 005)
BioMed Research International 9
through the secretion of surfactant substances from cells[21 22] The results of the drop-collapsing test showed thatbiosurfactants were likely not secreted from M colombiensecells suggesting that the motility of M colombiense couldbe favoured through hydrophobic molecules such as GPLsbound to the outermost portion of cells however we cannotcompletely rule out the possibility that some of the surfactantsubstances secreted from M colombiense remained adheredto the mycobacterial cell surface thereby influencing cellmotility
GPLs on the outermost portions of the M colombiensecell envelope generate a more hydrophobic cell surfacethat facilitates initial interactions with hydrophobic surfacesand increase biofilm formation on polystyrene wells Inaddition the increased GPL production in 19B and CECT3035 cells is consistent with the augmented biofilm forma-tion observed for M colombiense strains displaying smoothcolony morphology Moreover increased biofilm formationon hydrophobic surfaces and enhanced sliding over motilityplates for 19B cells compared with the CECT 3035 strainwhich also displays smooth colony morphology likely sug-gest possible differences in GPL structure betweenM colom-biense strains Differences inGPL production between strainscould influence the M colombiense ability to form biofilmson polyvinylchloride (PVC) pipes which would facilitatethe dissemination of these bacteria in natural environmentssuch as in-hospital spaces thereby increasing the chance ofinfection in immunosuppressed patients [23 39 40]
Based on the GPL biosynthetic pathway previouslydescribed for the M avium 104 strain [25] we are currentlyconstructing a potential gene cluster for GPL biosynthesis inM colombiense which will be finished when the nucleotidesequence of the CECT 3035 is completely assembled RT-qPCR experiments showed that the planktonic and motilecells of the rough 57B strain showed decreased transcriptionof the 9 selected genes leading to the low production ofGPLs in this rough variant comparedwith theM colombiensestrains displaying smooth colony morphology Interestinglyplanktonic 19B cells exhibited lower expression of most of theselected genes compared with planktonic CECT 3035 cellsThis behaviour results in the reduced production of GPLs inplanktonic 19B cells compared with the CECT 3035 strainconsistent with the results of the TLC experiments
It has been previously shown that an rtfA mutation inthe M avium serovar-2-specific strain resulted in the lossof serovar-specific GPLs thereby diminishing the variabilityof these glycolipids in the cell envelope [41 42] Thus itis possible that the enzymatic activity of the RtfA proteincould be relevant for the increased glycosylation andor aug-mented production of ssGPLs in strains displaying smoothcolony morphology compared with the rough variant 57BIn strains displaying smooth colony morphology the genesencoding rhamnosyl andmethyl transferases (rtfA andmtfB)are overtranscribed suggesting the increased production ofnsGPLs precursors for ssGPL biosynthesis [24ndash26] therebyincreasing the production of GPLs The diminished tran-scription of pstA gftA mtfC mtfD tmtpBand tmtpC inmotile CECT 3035 compared with control cells (planktonicCECT 3035) is intriguing however this behaviour also
suggests the relevance of rhamnosyl transferases particularlyRtfA in GPL biosynthesis in M colombiense Neverthelessthe actual role for RtfA in M colombiense strains should beconfirmed using rtfA-null mutants
We did not identify 7 of the 30 known GPL cluster genesof M avium 104 in the genome sequence of M colombienseCECT 3035 This interesting observation suggests that theGPLs between these two closely related MAC species couldhave different structural characteristics However furtherexperiments using mass spectrometry and NMR are neces-sary to evaluate potential differences in the GPL structureamongM colombiense strains
5 Conclusions
In conclusion the results of the present study show that Mcolombiense strains displaying smooth morphology exhibitincreased biofilm formation on hydrophobic polystyrenesurfaces and enhanced sliding over motility plates Bioin-formatics analyses indicate that the gene cluster establishedfor GPL formation modification and translocation is intactstrongly suggesting that GPLs are putatively present in Mcolombiense In addition motile culture conditions activatethe transcription of the genes implicated in the key enzymaticactivities of GPL biosynthesis
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
Thisworkwas supported through funding from the ldquoDivisionde Investigacion Bogota (DIB)rdquo-Vicerrectorıa de Investi-gacion Universidad Nacional de Colombia (Grants 1606014837 and 14337) Milena Maya-Hoyos received a fellowshipfrom the Academic Direction Universidad Nacional deColombia and the ldquoJovenes Investigadores e InnovadoresrdquoProgram Colciencias Colombia This research was alsosupported in part through the Intramural Research Programof the National Institutes of Health National Library ofMedicine and National Center for Biotechnology Informa-tion (NCBI)
References
[1] S K Field D Fisher and R L Cowie ldquoMycobacterium aviumcomplex pulmonary disease in patient without HIV infectionrdquoChest vol 126 no 2 pp 566ndash581 2004
[2] M J M Vaerewijck G Huys J C Palomino J Swings and FPortaels ldquoMycobacteria in drinking water distribution systemsecology and significance for human healthrdquo FEMSMicrobiologyReviews vol 29 no 5 pp 911ndash934 2005
[3] E Hagiwara S Komatsu R Nishihira T Shinohara T Babaand T Ogura ldquoClinical characteristics and prevalence of pneu-mothorax in patients with pulmonary Mycobacterium aviumcomplex diseaserdquo Journal of Infection and Chemotherapy vol19 no 4 pp 588ndash592 2013
10 BioMed Research International
[4] M I Murcia E Tortoli M C Menendez E Palenque andM J Garcia ldquoMycobacterium colombiense sp nov a novelmember of theMycobacterium avium complex and descriptionof MAC-X as a new ITS genetic variantrdquo International Journalof Systematic and Evolutionary Microbiology vol 56 no 9 pp2049ndash2054 2006
[5] E Tortoli L Rindi M J Garcia et al ldquoProposal to elevatethe genetic variant MAC-A included in the Mycobacteriumavium complex to species rank as Mycobacterium chimaerasp novrdquo International Journal of Systematic and EvolutionaryMicrobiology vol 54 no 4 pp 1277ndash1285 2004
[6] I B Salah C Cayrou D Raoult and M Drancourt ldquoMycobac-terium marseillense sp nov Mycobacterium timonense sp novand Mycobacterium bouchedurhonense sp novmembers ofthe Mycobacterium avium complexrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 11 pp2803ndash2808 2009
[7] J van Ingen M J Boeree K Kosters et al ldquoProposal toelevate Mycobacterium avium complex ITS sequevar MAC-Qto Mycobacterium vulneris sp novrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 9 pp2277ndash2282 2009
[8] D Bang T Herlin M Stegger et al ldquoMycobacterium arosiensesp nov a slowly growing scotochromogenic species causingosteomyelitis in an immunocompromised childrdquo InternationalJournal of Systematic and Evolutionary Microbiology vol 58part 10 pp 2398ndash2402 2008
[9] C Cayrou C Turenne M A Behr and M Drancourt ldquoGeno-typing of Mycobacterium avium complex organisms usingmultispacer sequence typingrdquoMicrobiology vol 156 part 3 pp687ndash694 2010
[10] S C Smole F McAleese J Ngampasutadol C F von Reynand R D Arbeit ldquoClinical and epidemiological correlates ofgenotypes within theMycobacterium avium complex defined byrestriction and sequence analysis of hsp65rdquo Journal of ClinicalMicrobiology vol 40 no 9 pp 3374ndash3380 2002
[11] AMuwonge C Kankya T B Johansen et al ldquoNon-tuberculousmycobacteria isolated from slaughter pigs in Mubende districtUgandardquo BMC Veterinary Research vol 8 article 52 2012
[12] J O Falkinham III ldquoSurrounded by mycobacteria nontuber-culous mycobacteria in the human environmentrdquo Journal ofApplied Microbiology vol 107 no 2 pp 356ndash367 2009
[13] K Vuorenmaa I B Salah V Barlogis H Chambost and MDrancourt ldquoMycobacterium colombiense and pseudotubercu-lous lymphadenopathyrdquo Emerging Infectious Diseases vol 15no 4 pp 618ndash620 2009
[14] O Esparcia F Navarro M Quer and P Coll ldquoLymphadenopa-thy caused by Mycobacterium colombienserdquo Journal of ClinicalMicrobiology vol 46 no 5 pp 1885ndash1887 2008
[15] S Cohen-Bacrie M David N Stremler J-C Dubus J-MRolain and M Drancourt ldquoMycobacterium chimaera pul-monary infection complicating cystic fibrosis a case reportrdquoJournal of Medical Case Reports vol 5 no 1 article 473 2011
[16] S Poulin C Corbeil M Nguyen et al ldquoFatal Mycobac-terium colombiensecytomegalovirus coinfection associatedwith acquired immunodeficiency due to autoantibodies againstinterferon gamma a case reportrdquo BMC Infectious Diseases vol13 no 1 article 24 2013
[17] J Leguizamon J Hernandez M-I Murcia and C-Y SotoldquoIdentification of potential biomarkers to distinguishMycobac-terium colombiense from other mycobacterial speciesrdquoMolecu-lar and Cellular Probes vol 27 no 1 pp 46ndash52 2013
[18] D Chatterjee and K H Khoo ldquoThe surface glycopeptidolipidsof mycobacteria structures and biological propertiesrdquo Cellularand Molecular Life Sciences vol 58 no 14 pp 2018ndash2042 2001
[19] J B Torrelles D Ellis T Osborne et al ldquoCharacterizationof virulence colony morphotype and the glycopeptidolipid ofMycobacterium avium strain 104rdquo Tuberculosis vol 82 no 6pp 293ndash300 2002
[20] M F Thorel and H L David ldquoSpecific surface antigens of SmTvariants ofMycobacterium aviumrdquo Infection and Immunity vol43 no 1 pp 438ndash439 1984
[21] A Martınez S Torello and R Kolter ldquoSliding motility inmycobacteriardquo Journal of Bacteriology vol 181 no 23 pp 7331ndash7338 1999
[22] J Recht AMartınez S Torello and R Kolter ldquoGenetic analysisof sliding motility in Mycobacterium smegmatisrdquo Journal ofBacteriology vol 182 no 15 pp 4348ndash4351 2000
[23] G Carter M Wu D C Drummond and L E BermudezldquoCharacterization of biofilm formation by clinical isolates ofMycobacterium aviumrdquo Journal of Medical Microbiology vol 52no 9 pp 747ndash752 2003
[24] J S Schorey and L Sweet ldquoThe mycobacterial glycopepti-dolipids structure function and their role in pathogenesisrdquoGlycobiology vol 18 no 11 pp 832ndash841 2008
[25] L Pang X Tian W Pan and J Xie ldquoStructure and function ofmycobacterium glycopeptidolipids from comparative genomicsperspectiverdquo Journal of Cellular Biochemistry vol 114 no 8 pp1705ndash1713 2013
[26] R Mukherjee and D Chatterji ldquoGlycopeptidolipids immuno-modulators in greasy mycobacterial cell enveloperdquo IUBMB Lifevol 64 no 3 pp 215ndash225 2012
[27] K L Horan R Freeman K Weigel et al ldquoIsolation ofthe genome sequence strain Mycobacterium avium 104 frommultiple patients over a 17-year periodrdquo Journal of ClinicalMicrobiology vol 44 no 3 pp 783ndash789 2006
[28] S B Snapper R E Melton S Mustafa T Kieser and W RJacobs Jr ldquoIsolation and characterization of efficient plasmidtransformation mutants of Mycobacterium smegmatisrdquo Molec-ular Microbiology vol 4 no 11 pp 1911ndash1919 1990
[29] M Radji R A Agustama B Elya and C R TjampakasarildquoAntimicrobial activity of green tea extract against isolatesof methicillin-resistant Staphylococcus aureus and multi-drugresistant Pseudomonas aeruginosardquo Asian Pacific Journal ofTropical Biomedicine vol 3 no 8 pp 663ndash667 2013
[30] P Helden T Victor R Warren and E Helden ldquoIsolationof DNA from Mycobacterium tuberculosisrdquo in Mycobacteriumtuberculosis Protocols Humana Press Totowa NJ USA 2001
[31] J C van Kessel L J Marinelli and G F Hatfull ldquoRecom-bineering mycobacteria and their phagesrdquo Nature ReviewsMicrobiology vol 6 no 11 pp 851ndash857 2008
[32] N M Parrish C G Ko J D Dick P B Jones and J LE Ellingson ldquoGrowth Congo Red agar colony morphotypesand antibiotic susceptibility testing of Mycobacterium aviumsubspecies paratuberculosisrdquo Clin Med Res vol 2 no 2 pp107ndash114 2004
[33] T Tugrul and E Cansunar ldquoDetecting surfactant-producingmicroorganisms by the drop-collapse testrdquo World Journal ofMicrobiology and Biotechnology vol 21 no 6-7 pp 851ndash8532005
[34] D K Jain D L Collins-Thompson H Lee and J T TrevorsldquoA drop-collapsing test for screening surfactant-producingmicroorganismsrdquo Journal ofMicrobiologicalMethods vol 13 no4 pp 271ndash279 1991
BioMed Research International 11
[35] M Munoz M-A Laneelle M Luquin et al ldquoOccurrence ofan antigenic triacyl trehalose in clinical isolates and referencestrains of Mycobacterium tuberculosisrdquo FEMS MicrobiologyLetters vol 157 no 2 pp 251ndash259 1997
[36] M Gonzalez-Perez M I Murcia D Landsman I K Jordanand L Marino-Ramırez ldquoGenome sequence of the Mycobac-terium colombiense type strain CECT 3035rdquo Journal of Bacte-riology vol 193 no 20 pp 5866ndash5867 2011
[37] J C Betts P T Lukey L C Robb R A McAdam andK Duncan ldquoEvaluation of a nutrient starvation model ofMycobacterium tuberculosis persistence by gene and proteinexpression profilingrdquoMolecular Microbiology vol 43 no 3 pp717ndash731 2002
[38] W W Barrow and P J Brennan ldquoIsolation in high frequencyof rough variants of Mycobacterium intracellulare lacking C-mycoside glycopeptidolipid antigensrdquo Journal of Bacteriologyvol 150 no 1 pp 381ndash384 1982
[39] W Costerton R Veeh M Shirtliff M Pasmore C Post andG Ehrlich ldquoThe application of biofilm science to the studyand control of chronic bacterial infectionsrdquo Journal of ClinicalInvestigation vol 112 no 10 pp 1466ndash1477 2003
[40] J Recht and R Kolter ldquoGlycopeptidolipid acetylation affectssliding motility and biofilm formation inMycobacterium smeg-matisrdquo Journal of Bacteriology vol 183 no 19 pp 5718ndash57242001
[41] T M Eckstein F S Silbaq D Chatterjee N J Kelly P JBrennan and J T Belisle ldquoIdentification and recombinantexpression of a Mycobacterium avium rhamnosyltransferasegene (rtfA) involved in glycopeptidolipid biosynthesisrdquo Journalof Bacteriology vol 180 no 21 pp 5567ndash5573 1998
[42] J N Maslow V R Irani S-H Lee T M Eckstein J MInamine and J T Belisle ldquoBiosynthetic specificity of therhamnosyltransferase gene of Mycobacterium avium serovar 2as determined by allelic exchange mutagenesisrdquo Microbiologyvol 149 no 11 pp 3193ndash3202 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 9
through the secretion of surfactant substances from cells[21 22] The results of the drop-collapsing test showed thatbiosurfactants were likely not secreted from M colombiensecells suggesting that the motility of M colombiense couldbe favoured through hydrophobic molecules such as GPLsbound to the outermost portion of cells however we cannotcompletely rule out the possibility that some of the surfactantsubstances secreted from M colombiense remained adheredto the mycobacterial cell surface thereby influencing cellmotility
GPLs on the outermost portions of the M colombiensecell envelope generate a more hydrophobic cell surfacethat facilitates initial interactions with hydrophobic surfacesand increase biofilm formation on polystyrene wells Inaddition the increased GPL production in 19B and CECT3035 cells is consistent with the augmented biofilm forma-tion observed for M colombiense strains displaying smoothcolony morphology Moreover increased biofilm formationon hydrophobic surfaces and enhanced sliding over motilityplates for 19B cells compared with the CECT 3035 strainwhich also displays smooth colony morphology likely sug-gest possible differences in GPL structure betweenM colom-biense strains Differences inGPL production between strainscould influence the M colombiense ability to form biofilmson polyvinylchloride (PVC) pipes which would facilitatethe dissemination of these bacteria in natural environmentssuch as in-hospital spaces thereby increasing the chance ofinfection in immunosuppressed patients [23 39 40]
Based on the GPL biosynthetic pathway previouslydescribed for the M avium 104 strain [25] we are currentlyconstructing a potential gene cluster for GPL biosynthesis inM colombiense which will be finished when the nucleotidesequence of the CECT 3035 is completely assembled RT-qPCR experiments showed that the planktonic and motilecells of the rough 57B strain showed decreased transcriptionof the 9 selected genes leading to the low production ofGPLs in this rough variant comparedwith theM colombiensestrains displaying smooth colony morphology Interestinglyplanktonic 19B cells exhibited lower expression of most of theselected genes compared with planktonic CECT 3035 cellsThis behaviour results in the reduced production of GPLs inplanktonic 19B cells compared with the CECT 3035 strainconsistent with the results of the TLC experiments
It has been previously shown that an rtfA mutation inthe M avium serovar-2-specific strain resulted in the lossof serovar-specific GPLs thereby diminishing the variabilityof these glycolipids in the cell envelope [41 42] Thus itis possible that the enzymatic activity of the RtfA proteincould be relevant for the increased glycosylation andor aug-mented production of ssGPLs in strains displaying smoothcolony morphology compared with the rough variant 57BIn strains displaying smooth colony morphology the genesencoding rhamnosyl andmethyl transferases (rtfA andmtfB)are overtranscribed suggesting the increased production ofnsGPLs precursors for ssGPL biosynthesis [24ndash26] therebyincreasing the production of GPLs The diminished tran-scription of pstA gftA mtfC mtfD tmtpBand tmtpC inmotile CECT 3035 compared with control cells (planktonicCECT 3035) is intriguing however this behaviour also
suggests the relevance of rhamnosyl transferases particularlyRtfA in GPL biosynthesis in M colombiense Neverthelessthe actual role for RtfA in M colombiense strains should beconfirmed using rtfA-null mutants
We did not identify 7 of the 30 known GPL cluster genesof M avium 104 in the genome sequence of M colombienseCECT 3035 This interesting observation suggests that theGPLs between these two closely related MAC species couldhave different structural characteristics However furtherexperiments using mass spectrometry and NMR are neces-sary to evaluate potential differences in the GPL structureamongM colombiense strains
5 Conclusions
In conclusion the results of the present study show that Mcolombiense strains displaying smooth morphology exhibitincreased biofilm formation on hydrophobic polystyrenesurfaces and enhanced sliding over motility plates Bioin-formatics analyses indicate that the gene cluster establishedfor GPL formation modification and translocation is intactstrongly suggesting that GPLs are putatively present in Mcolombiense In addition motile culture conditions activatethe transcription of the genes implicated in the key enzymaticactivities of GPL biosynthesis
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
Thisworkwas supported through funding from the ldquoDivisionde Investigacion Bogota (DIB)rdquo-Vicerrectorıa de Investi-gacion Universidad Nacional de Colombia (Grants 1606014837 and 14337) Milena Maya-Hoyos received a fellowshipfrom the Academic Direction Universidad Nacional deColombia and the ldquoJovenes Investigadores e InnovadoresrdquoProgram Colciencias Colombia This research was alsosupported in part through the Intramural Research Programof the National Institutes of Health National Library ofMedicine and National Center for Biotechnology Informa-tion (NCBI)
References
[1] S K Field D Fisher and R L Cowie ldquoMycobacterium aviumcomplex pulmonary disease in patient without HIV infectionrdquoChest vol 126 no 2 pp 566ndash581 2004
[2] M J M Vaerewijck G Huys J C Palomino J Swings and FPortaels ldquoMycobacteria in drinking water distribution systemsecology and significance for human healthrdquo FEMSMicrobiologyReviews vol 29 no 5 pp 911ndash934 2005
[3] E Hagiwara S Komatsu R Nishihira T Shinohara T Babaand T Ogura ldquoClinical characteristics and prevalence of pneu-mothorax in patients with pulmonary Mycobacterium aviumcomplex diseaserdquo Journal of Infection and Chemotherapy vol19 no 4 pp 588ndash592 2013
10 BioMed Research International
[4] M I Murcia E Tortoli M C Menendez E Palenque andM J Garcia ldquoMycobacterium colombiense sp nov a novelmember of theMycobacterium avium complex and descriptionof MAC-X as a new ITS genetic variantrdquo International Journalof Systematic and Evolutionary Microbiology vol 56 no 9 pp2049ndash2054 2006
[5] E Tortoli L Rindi M J Garcia et al ldquoProposal to elevatethe genetic variant MAC-A included in the Mycobacteriumavium complex to species rank as Mycobacterium chimaerasp novrdquo International Journal of Systematic and EvolutionaryMicrobiology vol 54 no 4 pp 1277ndash1285 2004
[6] I B Salah C Cayrou D Raoult and M Drancourt ldquoMycobac-terium marseillense sp nov Mycobacterium timonense sp novand Mycobacterium bouchedurhonense sp novmembers ofthe Mycobacterium avium complexrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 11 pp2803ndash2808 2009
[7] J van Ingen M J Boeree K Kosters et al ldquoProposal toelevate Mycobacterium avium complex ITS sequevar MAC-Qto Mycobacterium vulneris sp novrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 9 pp2277ndash2282 2009
[8] D Bang T Herlin M Stegger et al ldquoMycobacterium arosiensesp nov a slowly growing scotochromogenic species causingosteomyelitis in an immunocompromised childrdquo InternationalJournal of Systematic and Evolutionary Microbiology vol 58part 10 pp 2398ndash2402 2008
[9] C Cayrou C Turenne M A Behr and M Drancourt ldquoGeno-typing of Mycobacterium avium complex organisms usingmultispacer sequence typingrdquoMicrobiology vol 156 part 3 pp687ndash694 2010
[10] S C Smole F McAleese J Ngampasutadol C F von Reynand R D Arbeit ldquoClinical and epidemiological correlates ofgenotypes within theMycobacterium avium complex defined byrestriction and sequence analysis of hsp65rdquo Journal of ClinicalMicrobiology vol 40 no 9 pp 3374ndash3380 2002
[11] AMuwonge C Kankya T B Johansen et al ldquoNon-tuberculousmycobacteria isolated from slaughter pigs in Mubende districtUgandardquo BMC Veterinary Research vol 8 article 52 2012
[12] J O Falkinham III ldquoSurrounded by mycobacteria nontuber-culous mycobacteria in the human environmentrdquo Journal ofApplied Microbiology vol 107 no 2 pp 356ndash367 2009
[13] K Vuorenmaa I B Salah V Barlogis H Chambost and MDrancourt ldquoMycobacterium colombiense and pseudotubercu-lous lymphadenopathyrdquo Emerging Infectious Diseases vol 15no 4 pp 618ndash620 2009
[14] O Esparcia F Navarro M Quer and P Coll ldquoLymphadenopa-thy caused by Mycobacterium colombienserdquo Journal of ClinicalMicrobiology vol 46 no 5 pp 1885ndash1887 2008
[15] S Cohen-Bacrie M David N Stremler J-C Dubus J-MRolain and M Drancourt ldquoMycobacterium chimaera pul-monary infection complicating cystic fibrosis a case reportrdquoJournal of Medical Case Reports vol 5 no 1 article 473 2011
[16] S Poulin C Corbeil M Nguyen et al ldquoFatal Mycobac-terium colombiensecytomegalovirus coinfection associatedwith acquired immunodeficiency due to autoantibodies againstinterferon gamma a case reportrdquo BMC Infectious Diseases vol13 no 1 article 24 2013
[17] J Leguizamon J Hernandez M-I Murcia and C-Y SotoldquoIdentification of potential biomarkers to distinguishMycobac-terium colombiense from other mycobacterial speciesrdquoMolecu-lar and Cellular Probes vol 27 no 1 pp 46ndash52 2013
[18] D Chatterjee and K H Khoo ldquoThe surface glycopeptidolipidsof mycobacteria structures and biological propertiesrdquo Cellularand Molecular Life Sciences vol 58 no 14 pp 2018ndash2042 2001
[19] J B Torrelles D Ellis T Osborne et al ldquoCharacterizationof virulence colony morphotype and the glycopeptidolipid ofMycobacterium avium strain 104rdquo Tuberculosis vol 82 no 6pp 293ndash300 2002
[20] M F Thorel and H L David ldquoSpecific surface antigens of SmTvariants ofMycobacterium aviumrdquo Infection and Immunity vol43 no 1 pp 438ndash439 1984
[21] A Martınez S Torello and R Kolter ldquoSliding motility inmycobacteriardquo Journal of Bacteriology vol 181 no 23 pp 7331ndash7338 1999
[22] J Recht AMartınez S Torello and R Kolter ldquoGenetic analysisof sliding motility in Mycobacterium smegmatisrdquo Journal ofBacteriology vol 182 no 15 pp 4348ndash4351 2000
[23] G Carter M Wu D C Drummond and L E BermudezldquoCharacterization of biofilm formation by clinical isolates ofMycobacterium aviumrdquo Journal of Medical Microbiology vol 52no 9 pp 747ndash752 2003
[24] J S Schorey and L Sweet ldquoThe mycobacterial glycopepti-dolipids structure function and their role in pathogenesisrdquoGlycobiology vol 18 no 11 pp 832ndash841 2008
[25] L Pang X Tian W Pan and J Xie ldquoStructure and function ofmycobacterium glycopeptidolipids from comparative genomicsperspectiverdquo Journal of Cellular Biochemistry vol 114 no 8 pp1705ndash1713 2013
[26] R Mukherjee and D Chatterji ldquoGlycopeptidolipids immuno-modulators in greasy mycobacterial cell enveloperdquo IUBMB Lifevol 64 no 3 pp 215ndash225 2012
[27] K L Horan R Freeman K Weigel et al ldquoIsolation ofthe genome sequence strain Mycobacterium avium 104 frommultiple patients over a 17-year periodrdquo Journal of ClinicalMicrobiology vol 44 no 3 pp 783ndash789 2006
[28] S B Snapper R E Melton S Mustafa T Kieser and W RJacobs Jr ldquoIsolation and characterization of efficient plasmidtransformation mutants of Mycobacterium smegmatisrdquo Molec-ular Microbiology vol 4 no 11 pp 1911ndash1919 1990
[29] M Radji R A Agustama B Elya and C R TjampakasarildquoAntimicrobial activity of green tea extract against isolatesof methicillin-resistant Staphylococcus aureus and multi-drugresistant Pseudomonas aeruginosardquo Asian Pacific Journal ofTropical Biomedicine vol 3 no 8 pp 663ndash667 2013
[30] P Helden T Victor R Warren and E Helden ldquoIsolationof DNA from Mycobacterium tuberculosisrdquo in Mycobacteriumtuberculosis Protocols Humana Press Totowa NJ USA 2001
[31] J C van Kessel L J Marinelli and G F Hatfull ldquoRecom-bineering mycobacteria and their phagesrdquo Nature ReviewsMicrobiology vol 6 no 11 pp 851ndash857 2008
[32] N M Parrish C G Ko J D Dick P B Jones and J LE Ellingson ldquoGrowth Congo Red agar colony morphotypesand antibiotic susceptibility testing of Mycobacterium aviumsubspecies paratuberculosisrdquo Clin Med Res vol 2 no 2 pp107ndash114 2004
[33] T Tugrul and E Cansunar ldquoDetecting surfactant-producingmicroorganisms by the drop-collapse testrdquo World Journal ofMicrobiology and Biotechnology vol 21 no 6-7 pp 851ndash8532005
[34] D K Jain D L Collins-Thompson H Lee and J T TrevorsldquoA drop-collapsing test for screening surfactant-producingmicroorganismsrdquo Journal ofMicrobiologicalMethods vol 13 no4 pp 271ndash279 1991
BioMed Research International 11
[35] M Munoz M-A Laneelle M Luquin et al ldquoOccurrence ofan antigenic triacyl trehalose in clinical isolates and referencestrains of Mycobacterium tuberculosisrdquo FEMS MicrobiologyLetters vol 157 no 2 pp 251ndash259 1997
[36] M Gonzalez-Perez M I Murcia D Landsman I K Jordanand L Marino-Ramırez ldquoGenome sequence of the Mycobac-terium colombiense type strain CECT 3035rdquo Journal of Bacte-riology vol 193 no 20 pp 5866ndash5867 2011
[37] J C Betts P T Lukey L C Robb R A McAdam andK Duncan ldquoEvaluation of a nutrient starvation model ofMycobacterium tuberculosis persistence by gene and proteinexpression profilingrdquoMolecular Microbiology vol 43 no 3 pp717ndash731 2002
[38] W W Barrow and P J Brennan ldquoIsolation in high frequencyof rough variants of Mycobacterium intracellulare lacking C-mycoside glycopeptidolipid antigensrdquo Journal of Bacteriologyvol 150 no 1 pp 381ndash384 1982
[39] W Costerton R Veeh M Shirtliff M Pasmore C Post andG Ehrlich ldquoThe application of biofilm science to the studyand control of chronic bacterial infectionsrdquo Journal of ClinicalInvestigation vol 112 no 10 pp 1466ndash1477 2003
[40] J Recht and R Kolter ldquoGlycopeptidolipid acetylation affectssliding motility and biofilm formation inMycobacterium smeg-matisrdquo Journal of Bacteriology vol 183 no 19 pp 5718ndash57242001
[41] T M Eckstein F S Silbaq D Chatterjee N J Kelly P JBrennan and J T Belisle ldquoIdentification and recombinantexpression of a Mycobacterium avium rhamnosyltransferasegene (rtfA) involved in glycopeptidolipid biosynthesisrdquo Journalof Bacteriology vol 180 no 21 pp 5567ndash5573 1998
[42] J N Maslow V R Irani S-H Lee T M Eckstein J MInamine and J T Belisle ldquoBiosynthetic specificity of therhamnosyltransferase gene of Mycobacterium avium serovar 2as determined by allelic exchange mutagenesisrdquo Microbiologyvol 149 no 11 pp 3193ndash3202 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 BioMed Research International
[4] M I Murcia E Tortoli M C Menendez E Palenque andM J Garcia ldquoMycobacterium colombiense sp nov a novelmember of theMycobacterium avium complex and descriptionof MAC-X as a new ITS genetic variantrdquo International Journalof Systematic and Evolutionary Microbiology vol 56 no 9 pp2049ndash2054 2006
[5] E Tortoli L Rindi M J Garcia et al ldquoProposal to elevatethe genetic variant MAC-A included in the Mycobacteriumavium complex to species rank as Mycobacterium chimaerasp novrdquo International Journal of Systematic and EvolutionaryMicrobiology vol 54 no 4 pp 1277ndash1285 2004
[6] I B Salah C Cayrou D Raoult and M Drancourt ldquoMycobac-terium marseillense sp nov Mycobacterium timonense sp novand Mycobacterium bouchedurhonense sp novmembers ofthe Mycobacterium avium complexrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 11 pp2803ndash2808 2009
[7] J van Ingen M J Boeree K Kosters et al ldquoProposal toelevate Mycobacterium avium complex ITS sequevar MAC-Qto Mycobacterium vulneris sp novrdquo International Journal ofSystematic and Evolutionary Microbiology vol 59 no 9 pp2277ndash2282 2009
[8] D Bang T Herlin M Stegger et al ldquoMycobacterium arosiensesp nov a slowly growing scotochromogenic species causingosteomyelitis in an immunocompromised childrdquo InternationalJournal of Systematic and Evolutionary Microbiology vol 58part 10 pp 2398ndash2402 2008
[9] C Cayrou C Turenne M A Behr and M Drancourt ldquoGeno-typing of Mycobacterium avium complex organisms usingmultispacer sequence typingrdquoMicrobiology vol 156 part 3 pp687ndash694 2010
[10] S C Smole F McAleese J Ngampasutadol C F von Reynand R D Arbeit ldquoClinical and epidemiological correlates ofgenotypes within theMycobacterium avium complex defined byrestriction and sequence analysis of hsp65rdquo Journal of ClinicalMicrobiology vol 40 no 9 pp 3374ndash3380 2002
[11] AMuwonge C Kankya T B Johansen et al ldquoNon-tuberculousmycobacteria isolated from slaughter pigs in Mubende districtUgandardquo BMC Veterinary Research vol 8 article 52 2012
[12] J O Falkinham III ldquoSurrounded by mycobacteria nontuber-culous mycobacteria in the human environmentrdquo Journal ofApplied Microbiology vol 107 no 2 pp 356ndash367 2009
[13] K Vuorenmaa I B Salah V Barlogis H Chambost and MDrancourt ldquoMycobacterium colombiense and pseudotubercu-lous lymphadenopathyrdquo Emerging Infectious Diseases vol 15no 4 pp 618ndash620 2009
[14] O Esparcia F Navarro M Quer and P Coll ldquoLymphadenopa-thy caused by Mycobacterium colombienserdquo Journal of ClinicalMicrobiology vol 46 no 5 pp 1885ndash1887 2008
[15] S Cohen-Bacrie M David N Stremler J-C Dubus J-MRolain and M Drancourt ldquoMycobacterium chimaera pul-monary infection complicating cystic fibrosis a case reportrdquoJournal of Medical Case Reports vol 5 no 1 article 473 2011
[16] S Poulin C Corbeil M Nguyen et al ldquoFatal Mycobac-terium colombiensecytomegalovirus coinfection associatedwith acquired immunodeficiency due to autoantibodies againstinterferon gamma a case reportrdquo BMC Infectious Diseases vol13 no 1 article 24 2013
[17] J Leguizamon J Hernandez M-I Murcia and C-Y SotoldquoIdentification of potential biomarkers to distinguishMycobac-terium colombiense from other mycobacterial speciesrdquoMolecu-lar and Cellular Probes vol 27 no 1 pp 46ndash52 2013
[18] D Chatterjee and K H Khoo ldquoThe surface glycopeptidolipidsof mycobacteria structures and biological propertiesrdquo Cellularand Molecular Life Sciences vol 58 no 14 pp 2018ndash2042 2001
[19] J B Torrelles D Ellis T Osborne et al ldquoCharacterizationof virulence colony morphotype and the glycopeptidolipid ofMycobacterium avium strain 104rdquo Tuberculosis vol 82 no 6pp 293ndash300 2002
[20] M F Thorel and H L David ldquoSpecific surface antigens of SmTvariants ofMycobacterium aviumrdquo Infection and Immunity vol43 no 1 pp 438ndash439 1984
[21] A Martınez S Torello and R Kolter ldquoSliding motility inmycobacteriardquo Journal of Bacteriology vol 181 no 23 pp 7331ndash7338 1999
[22] J Recht AMartınez S Torello and R Kolter ldquoGenetic analysisof sliding motility in Mycobacterium smegmatisrdquo Journal ofBacteriology vol 182 no 15 pp 4348ndash4351 2000
[23] G Carter M Wu D C Drummond and L E BermudezldquoCharacterization of biofilm formation by clinical isolates ofMycobacterium aviumrdquo Journal of Medical Microbiology vol 52no 9 pp 747ndash752 2003
[24] J S Schorey and L Sweet ldquoThe mycobacterial glycopepti-dolipids structure function and their role in pathogenesisrdquoGlycobiology vol 18 no 11 pp 832ndash841 2008
[25] L Pang X Tian W Pan and J Xie ldquoStructure and function ofmycobacterium glycopeptidolipids from comparative genomicsperspectiverdquo Journal of Cellular Biochemistry vol 114 no 8 pp1705ndash1713 2013
[26] R Mukherjee and D Chatterji ldquoGlycopeptidolipids immuno-modulators in greasy mycobacterial cell enveloperdquo IUBMB Lifevol 64 no 3 pp 215ndash225 2012
[27] K L Horan R Freeman K Weigel et al ldquoIsolation ofthe genome sequence strain Mycobacterium avium 104 frommultiple patients over a 17-year periodrdquo Journal of ClinicalMicrobiology vol 44 no 3 pp 783ndash789 2006
[28] S B Snapper R E Melton S Mustafa T Kieser and W RJacobs Jr ldquoIsolation and characterization of efficient plasmidtransformation mutants of Mycobacterium smegmatisrdquo Molec-ular Microbiology vol 4 no 11 pp 1911ndash1919 1990
[29] M Radji R A Agustama B Elya and C R TjampakasarildquoAntimicrobial activity of green tea extract against isolatesof methicillin-resistant Staphylococcus aureus and multi-drugresistant Pseudomonas aeruginosardquo Asian Pacific Journal ofTropical Biomedicine vol 3 no 8 pp 663ndash667 2013
[30] P Helden T Victor R Warren and E Helden ldquoIsolationof DNA from Mycobacterium tuberculosisrdquo in Mycobacteriumtuberculosis Protocols Humana Press Totowa NJ USA 2001
[31] J C van Kessel L J Marinelli and G F Hatfull ldquoRecom-bineering mycobacteria and their phagesrdquo Nature ReviewsMicrobiology vol 6 no 11 pp 851ndash857 2008
[32] N M Parrish C G Ko J D Dick P B Jones and J LE Ellingson ldquoGrowth Congo Red agar colony morphotypesand antibiotic susceptibility testing of Mycobacterium aviumsubspecies paratuberculosisrdquo Clin Med Res vol 2 no 2 pp107ndash114 2004
[33] T Tugrul and E Cansunar ldquoDetecting surfactant-producingmicroorganisms by the drop-collapse testrdquo World Journal ofMicrobiology and Biotechnology vol 21 no 6-7 pp 851ndash8532005
[34] D K Jain D L Collins-Thompson H Lee and J T TrevorsldquoA drop-collapsing test for screening surfactant-producingmicroorganismsrdquo Journal ofMicrobiologicalMethods vol 13 no4 pp 271ndash279 1991
BioMed Research International 11
[35] M Munoz M-A Laneelle M Luquin et al ldquoOccurrence ofan antigenic triacyl trehalose in clinical isolates and referencestrains of Mycobacterium tuberculosisrdquo FEMS MicrobiologyLetters vol 157 no 2 pp 251ndash259 1997
[36] M Gonzalez-Perez M I Murcia D Landsman I K Jordanand L Marino-Ramırez ldquoGenome sequence of the Mycobac-terium colombiense type strain CECT 3035rdquo Journal of Bacte-riology vol 193 no 20 pp 5866ndash5867 2011
[37] J C Betts P T Lukey L C Robb R A McAdam andK Duncan ldquoEvaluation of a nutrient starvation model ofMycobacterium tuberculosis persistence by gene and proteinexpression profilingrdquoMolecular Microbiology vol 43 no 3 pp717ndash731 2002
[38] W W Barrow and P J Brennan ldquoIsolation in high frequencyof rough variants of Mycobacterium intracellulare lacking C-mycoside glycopeptidolipid antigensrdquo Journal of Bacteriologyvol 150 no 1 pp 381ndash384 1982
[39] W Costerton R Veeh M Shirtliff M Pasmore C Post andG Ehrlich ldquoThe application of biofilm science to the studyand control of chronic bacterial infectionsrdquo Journal of ClinicalInvestigation vol 112 no 10 pp 1466ndash1477 2003
[40] J Recht and R Kolter ldquoGlycopeptidolipid acetylation affectssliding motility and biofilm formation inMycobacterium smeg-matisrdquo Journal of Bacteriology vol 183 no 19 pp 5718ndash57242001
[41] T M Eckstein F S Silbaq D Chatterjee N J Kelly P JBrennan and J T Belisle ldquoIdentification and recombinantexpression of a Mycobacterium avium rhamnosyltransferasegene (rtfA) involved in glycopeptidolipid biosynthesisrdquo Journalof Bacteriology vol 180 no 21 pp 5567ndash5573 1998
[42] J N Maslow V R Irani S-H Lee T M Eckstein J MInamine and J T Belisle ldquoBiosynthetic specificity of therhamnosyltransferase gene of Mycobacterium avium serovar 2as determined by allelic exchange mutagenesisrdquo Microbiologyvol 149 no 11 pp 3193ndash3202 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 11
[35] M Munoz M-A Laneelle M Luquin et al ldquoOccurrence ofan antigenic triacyl trehalose in clinical isolates and referencestrains of Mycobacterium tuberculosisrdquo FEMS MicrobiologyLetters vol 157 no 2 pp 251ndash259 1997
[36] M Gonzalez-Perez M I Murcia D Landsman I K Jordanand L Marino-Ramırez ldquoGenome sequence of the Mycobac-terium colombiense type strain CECT 3035rdquo Journal of Bacte-riology vol 193 no 20 pp 5866ndash5867 2011
[37] J C Betts P T Lukey L C Robb R A McAdam andK Duncan ldquoEvaluation of a nutrient starvation model ofMycobacterium tuberculosis persistence by gene and proteinexpression profilingrdquoMolecular Microbiology vol 43 no 3 pp717ndash731 2002
[38] W W Barrow and P J Brennan ldquoIsolation in high frequencyof rough variants of Mycobacterium intracellulare lacking C-mycoside glycopeptidolipid antigensrdquo Journal of Bacteriologyvol 150 no 1 pp 381ndash384 1982
[39] W Costerton R Veeh M Shirtliff M Pasmore C Post andG Ehrlich ldquoThe application of biofilm science to the studyand control of chronic bacterial infectionsrdquo Journal of ClinicalInvestigation vol 112 no 10 pp 1466ndash1477 2003
[40] J Recht and R Kolter ldquoGlycopeptidolipid acetylation affectssliding motility and biofilm formation inMycobacterium smeg-matisrdquo Journal of Bacteriology vol 183 no 19 pp 5718ndash57242001
[41] T M Eckstein F S Silbaq D Chatterjee N J Kelly P JBrennan and J T Belisle ldquoIdentification and recombinantexpression of a Mycobacterium avium rhamnosyltransferasegene (rtfA) involved in glycopeptidolipid biosynthesisrdquo Journalof Bacteriology vol 180 no 21 pp 5567ndash5573 1998
[42] J N Maslow V R Irani S-H Lee T M Eckstein J MInamine and J T Belisle ldquoBiosynthetic specificity of therhamnosyltransferase gene of Mycobacterium avium serovar 2as determined by allelic exchange mutagenesisrdquo Microbiologyvol 149 no 11 pp 3193ndash3202 2003
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom