
Research Article Nonlinear Pulse Vaccination in an...
14
Hindawi Publishing Corporation Abstract and Applied Analysis Volume 2013, Article ID 670263, 13 pages http://dx.doi.org/10.1155/2013/670263 Research Article Nonlinear Pulse Vaccination in an SIR Epidemic Model with Resource Limitation Wenjie Qin, 1 Sanyi Tang, 1 and Robert A. Cheke 2 1 College of Mathematics and Information Science, Shaanxi Normal University, Xi’an 710062, China 2 Natural Resources Institute, University of Greenwich at Medway, Central Avenue, Chatham Maritime, Chatham, Kent ME4 4TB, UK Correspondence should be addressed to Wenjie Qin; [email protected] Received 3 October 2013; Accepted 18 November 2013 Academic Editor: Juan C. Cort´ es Copyright © 2013 Wenjie Qin et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Mathematical models can assist in the design and understanding of vaccination strategies when resources are limited. Here we propose and analyse an SIR epidemic model with a nonlinear pulse vaccination to examine how a limited vaccine resource affects the transmission and control of infectious diseases, in particular emerging infectious diseases. e threshold condition for the stability of the disease free steady state is given. Latin Hypercube Sampling/Partial Rank Correlation Coefficient uncertainty and sensitivity analysis techniques were employed to determine the key factors which are most significantly related to the threshold value. Comparing this threshold value with that without resource limitation, our results indicate that if resources become limited pulse vaccination should be carried out more frequently than when sufficient resources are available to eradicate an infectious disease. Once the threshold value exceeds a critical level, both susceptible and infected populations can oscillate periodically. Furthermore, when the pulse vaccination period is chosen as a bifurcation parameter, the SIR model with nonlinear pulse vaccination reveals complex dynamics including period doubling, chaotic solutions, and coexistence of multiple attractors. e implications of our findings with respect to disease control are discussed. 1. Introduction Epidemiology is the study of the spread of diseases with the objective of tracing factors that are responsible for or contribute to their occurrence and serves as the foundation and logic of interventions made in the interest of public health and preventive medicine. Mathematical models describing the population dynamics of infectious diseases have played an important role in better understanding epidemiological patterns and disease control for a long time. Various epidemic models have been proposed and explored extensively and considerable progress has been achieved in the studies of disease control and prevention (see [1–3] and the references therein). Outbreaks of infectious diseases have not only caused the loss of billions of lives but have oſten also rapidly dam- aged social economic systems, bringing about much human misery. Consequently, the focus of our research has been on how to prevent and cure infectious diseases effectively. It is well known that one of the most important concerns in the analysis of epidemic logical models is the efficacy of vaccination programmes. is subject gained prominence as a result of highly successful application of vaccinations for the worldwide eradication of smallpox and the restriction of diseases such as poliomyelitis, measles, tetanus, diphtheria, pertussis, and tuberculosis. Vaccination is one of the most cost effective of health investments as it can prevent or ameliorate morbidity due to infections. e effectiveness of vaccination has been widely studied and verified for the influenza vaccine [4], the HPV vaccine [5], the chicken pox vaccine [6], and others. In particular, Agur et al. [7] first proposed a pulse vaccina- tion strategy (PVS), which consists of periodical repetitions of impulsive vaccinations of all the age cohorts in a population, which has been confirmed as an important and effective strategy for the elimination of infectious diseases. At each vaccination time a constant fraction of susceptible people is vaccinated. is kind of vaccination is called impulsive since all the vaccine doses are applied within a very short time, with respect to the dynamics of the target disease. PVS allows
-
Upload
duongxuyen -
Category
Documents
-
view
217 -
download
0
Transcript of Research Article Nonlinear Pulse Vaccination in an...

Hindawi Publishing CorporationAbstract and Applied AnalysisVolume 2013 Article ID 670263 13 pageshttpdxdoiorg1011552013670263
Research ArticleNonlinear Pulse Vaccination in an SIR Epidemic Modelwith Resource Limitation
Wenjie Qin1 Sanyi Tang1 and Robert A Cheke2
1 College of Mathematics and Information Science Shaanxi Normal University Xirsquoan 710062 China2Natural Resources Institute University of Greenwich at Medway Central Avenue ChathamMaritime Chatham Kent ME4 4TB UK
Correspondence should be addressed to Wenjie Qin wenjieqinhotmailcom
Received 3 October 2013 Accepted 18 November 2013
Academic Editor Juan C Cortes
Copyright copy 2013 Wenjie Qin et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Mathematical models can assist in the design and understanding of vaccination strategies when resources are limited Here wepropose and analyse an SIR epidemicmodelwith a nonlinear pulse vaccination to examine how a limited vaccine resource affects thetransmission and control of infectious diseases in particular emerging infectious diseasesThe threshold condition for the stabilityof the disease free steady state is given Latin Hypercube SamplingPartial Rank Correlation Coefficient uncertainty and sensitivityanalysis techniques were employed to determine the key factors which are most significantly related to the threshold valueComparing this threshold value with that without resource limitation our results indicate that if resources become limited pulsevaccination should be carried out more frequently than when sufficient resources are available to eradicate an infectious diseaseOnce the threshold value exceeds a critical level both susceptible and infected populations can oscillate periodically Furthermorewhen the pulse vaccination period is chosen as a bifurcation parameter the SIR model with nonlinear pulse vaccination revealscomplex dynamics including period doubling chaotic solutions and coexistence of multiple attractors The implications of ourfindings with respect to disease control are discussed
1 Introduction
Epidemiology is the study of the spread of diseases withthe objective of tracing factors that are responsible for orcontribute to their occurrence and serves as the foundationand logic of interventionsmade in the interest of public healthand preventive medicine Mathematical models describingthe population dynamics of infectious diseases have playedan important role in better understanding epidemiologicalpatterns and disease control for a long time Various epidemicmodels have been proposed and explored extensively andconsiderable progress has been achieved in the studies ofdisease control and prevention (see [1ndash3] and the referencestherein)
Outbreaks of infectious diseases have not only causedthe loss of billions of lives but have often also rapidly dam-aged social economic systems bringing about much humanmisery Consequently the focus of our research has beenon how to prevent and cure infectious diseases effectivelyIt is well known that one of the most important concerns
in the analysis of epidemic logical models is the efficacy ofvaccination programmes This subject gained prominence asa result of highly successful application of vaccinations forthe worldwide eradication of smallpox and the restriction ofdiseases such as poliomyelitis measles tetanus diphtheriapertussis and tuberculosis Vaccination is one of the mostcost effective of health investments as it can prevent orameliorate morbidity due to infections The effectiveness ofvaccination has been widely studied and verified for theinfluenza vaccine [4] the HPV vaccine [5] the chicken poxvaccine [6] and others
In particular Agur et al [7] first proposed a pulse vaccina-tion strategy (PVS)which consists of periodical repetitions ofimpulsive vaccinations of all the age cohorts in a populationwhich has been confirmed as an important and effectivestrategy for the elimination of infectious diseases At eachvaccination time a constant fraction 119901 of susceptible people isvaccinated This kind of vaccination is called impulsive sinceall the vaccine doses are applied within a very short timewith respect to the dynamics of the target disease PVS allows
2 Abstract and Applied Analysis
the eradication of a disease with some practical advantagesas discussed in [7ndash9] Recently epidemic models with pulsevaccination have been the subject of intense theoreticalanalysis For example the dynamical behaviors of variousepidemic models with PVS are studied in [8 10ndash13] Thetheoretical results show that PVS can be distinguished fromthe conventional strategies in leading to disease eradicationat relatively low values of vaccination (see [7])
Traditional epidemic models with PVS of populationdynamics have assumed that the pulse vaccination propor-tion 119901 is constant which implies that medical resourcessuch as drugs vaccines hospital beds and isolation unitsare sufficient for the infectious disease in question Howeverin reality every community or country has an appropriateor limited capacity for treatment especially for emerginginfectious diseases and so understanding resource limitationis critical to effective management
During the last decade the outbreaks of SARS in 2003and avian influenza among humans (H5N1 in 2003 H1N1 in2009 andH7N9 in 2013) emphasized the need to enhance thecapacity to fight emerging infectious diseases which remainsa challenge for public health Several different vaccines havebeen developed specifically targeting avian influenza amonghumans but limited production has exposed weaknesses[14ndash16] A lack of sufficient vaccine at the outset of a newpandemic is to be expected given the crucial lag betweenthe timing of the emergence of a new strain and when avaccine has been developed and is ready for distributionAlthough most vaccination programmes aim to vaccinate asmany susceptible individuals as possible this is not always thecase for emerging infectious diseases due to limited vaccineavailability particularly in developing countries
However resource limitation is difficult to quantify partlybecause it is a dynamic process (see [17]) Recently continu-ous SIR models concerning limited medical resources havebeen developed and investigated (see [18 19]) Chow et al[18] explored the dynamics of an SIR epidemic model tounderstand how limited medical resources and their supplyefficiency affect the transmission of infectious diseases Zhouand Fan [19] studied a multigroup SIR epidemiologicalmodel to explore the effects of limited medical resourcesand group-targeted vaccination strategies on disease controland prevention To the best of our knowledge no workhas been done for the effects of resource limitation on anSIR model with PVS In order to investigate the effect oflimited vaccine availability on the spread of infectious diseasea saturation phenomenon of limited medical resources isconsideredThat is we will study the dynamic behavior of anSIR epidemic model with nonlinear pulse vaccination Suchmathematical models are suitable for simulating processeswith short duration perturbations during their development(see [20])
In this paper the dynamical behavior of an SIR epidemicmodel with nonlinear pulse vaccination is proposed and ana-lyzed The main purpose is to address and understand how alimited vaccine resource affects the transmission and controlof infectious diseases The rest of this paper is organized asfollows In the next section an SIR epidemic model withnonlinear PVS is introduced In Section 3 we investigate the
existence and stability of the disease-free periodic solutionby using the method of differential inequality qualitativeanalysis a discrete dynamical system determined by the stro-boscopic map and comparison theorem Meanwhile LatinHypercube Sampling (LHS)Partial Rank Correlation Coef-ficients (PRCCs) uncertainty and sensitivity analysis tech-niques are employed to investigate the key control parameterswhich are most significantly related to threshold values (see[21ndash23]) Section 4 focuses on finding the sufficient conditionunderwhich the endemic periodic solution exists by using thebifurcation theorem The paper ends with some interestingbiological conclusions and numerical bifurcation analyseswhich complement the theoretical findings
2 Model Formulation
In the classical epidemiological model (see [24ndash26]) the totalpopulation 119873 is composed of three groups of individualssusceptible (119878) infective (119868) and recovered (119877) Let 119878(119905)denote the number ofmembers of a population susceptible tothe disease at time 119905 119868(119905) represents the number of infectivemembers and 119877(119905) denotes the number of members whohave been removed from the population This leads to thefollowing SIR model
d119878 (119905)d119905
= 119887 minus 120573119878 (119905) 119868 (119905) minus 119887119878 (119905)
d119868 (119905)d119905
= 120573119878 (119905) 119868 (119905) minus 120574119868 (119905) minus 119887119868 (119905)
d119877 (119905)d119905
= 120574119868 (119905) minus 119887119877 (119905)
(1)
In model (1) we assume that the three classes of subpop-ulations have the same constant birth and death rates 119887(ie the total population has a constant size) which can benormalized to unity that is119873(119905) = 119878(119905)+ 119868(119905)+119877(119905) = 1 andthe infectious individuals recover at a rate 120574 gt 0 so that 1120574is the mean infectious period Susceptibles become infectedat a rate 120573119868 where 120573 gt 0 is the transmission rate In practicethe equation for d119877(119905)d119905 is redundant because 119877(119905) can beobtained from119873(119905) = 1 A detailed description of model (1)and its dynamics may be found in [9 27 28]
Let 119879 gt 0 be the time between two consecutive pulsevaccinations and let 119901(119905) (0 le 119901(119905) lt 1) be the fraction ofsusceptible subjects who are inoculated with vaccine whichdepends on the number of susceptibles in the population attime 119905 This yields the following model with PVS [8 9]
d119878 (119905)d119905
= 119887 minus 120573119878 (119905) 119868 (119905) minus 119887119878 (119905)
d119868 (119905)d119905
= 120573119878 (119905) 119868 (119905) minus 120574119868 (119905) minus 119887119868 (119905)
119905 = 119899119879 119899 isinN
119878 (119905+) = (1 minus 119901 (119905)) 119878 (119905)
119868 (119905+) = 119868 (119905)
119905 = 119899119879 119899 isinN
(2)
hereN = 0 1 2
Abstract and Applied Analysis 3
Table 1 Parameter definitions values and source for model (4)
Parameter Definition Value Source119887 Rate of natural birth or death rate of the population (yearminus1) 002 [33]120573 Probability of transmission per contact (yearminus1) 1800 [33]120574 Natural recovery rate of the infective individuals (yearminus1) 100 [33]119901max Themaximal vaccination proportion (yearminus1) [0 1) [8 9]120579 The half saturation constant [0 1] Assumption119879 Period of pulse vaccination (year) [0 20] [8 9]
The formula for 119901(119905) plays a key role in investigating theeffect of the limited capacity for treatment on the spread ofinfectious disease To do this we assume without loss of gen-erality that the PVS is implemented in a developing countrybut one in which vaccines are relatively abundant Note thatthere are often major shortages of medical personnel in ruralareas and in certain specialities such as surgery paediatricsand obstetrics [14] Let vaccination pulses occur every 119879 timeunits When a pulse day occurs the medical personnel haveto go out into the rural population and vaccinate as manysusceptibles as they can find on that particular day If thereare very few susceptibles then it will be hard to find them sothe vaccination coverage will be low The more susceptiblesthere are the easier it is to find them so the coverage per pulsewill be greater But there is a limit to how fast the medicalteam can find and handle each susceptible and this causesthe vaccination rate 119901(119905) to saturate that is a saturationphenomenon of limited medical resources By employingthe Holling type II functional response for a predator-preymodel we can define 119901(119905) as follows
119901 (119905) =119901max119878 (119905)
119878 (119905) + 120579 0 le 119901max lt 1 (3)
where 119901max denotes the maximal vaccination proportion and120579 is the half saturation constant Note that the vaccinationcoverage is less than 100 and in practice a vaccine efficacyis also less than 100 which lead to the parameter 119901max lt 1Furthermore generalized formulae such as the Hill function[29] can be employed to characterize the saturation phe-nomenon of the limited medical resources
Taking into account the resource limitation and satura-tion effects for convenience and simplification then model(2) can be rewritten as
d119878 (119905)d119905
= 119887 minus 120573119878 (119905) 119868 (119905) minus 119887119878 (119905)
d119868 (119905)d119905
= 120573119878 (119905) 119868 (119905) minus 120574119868 (119905) minus 119887119868 (119905)
119905 = 119899119879 119899 isinN
119878 (119905+) =
(1 minus 119901max) 1198782
(119905) + 120579119878 (119905)
119878 (119905) + 120579
119868 (119905+) = 119868 (119905)
119905 = 119899119879 119899 isinN
(4)
where the parameters 119887 120573 120574 and 119901max 120579 are defined in mod-els (1) and (3) respectively Note that model (4) is reduced tothe classical SIRmodel with constant pulse vaccination when120579 = 0 which was investigated by Agur et al [7] Stone et al
[8] and Shulgin et al [9]The parameter definitions of model(4) are summarized in Table 1
3 Existence of the Disease-FreePeriodic Solution
We first demonstrate the existence of a disease-free periodicsolution of model (4) in which infectious individuals areentirely absent from the population permanently that is119868(119905) = 0 for a sufficiently long time Under some thresholdconditions we show below that the susceptible populationoscillates with period 119879 in synchronization with the periodicpulse vaccination and the infectious class will die out eventu-allyThus in this section we focus on determining the globalattraction of this disease-free periodic solution To addressthis we first consider the following submodel over the timeinterval 119899119879 lt 119905 le (119899 + 1)119879
d119878 (119905)d119905
= 119887 minus 119887119878 (119905) 119905 = 119899119879
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579 119905 = 119899119879
119878 (0+
) = 1198780
(5)
Integrating the first equation of model (5) between pulsesyields
119878 (119905) = [119878 (119899119879+
) minus 1] eminus119887(119905minus119899119879) + 1 (6)
for 119899119879 lt 119905 le (119899 + 1)119879 It follows from the second equation ofmodel (5) that
119878 ((119899 + 1) 119879+
) =(1 minus 119901max) 119878
2
((119899 + 1) 119879) + 120579119878 ((119899 + 1) 119879)
119878 ((119899 + 1) 119879) + 120579
(7)
Substitution of (6) into (7) gives
119878 ((119899 + 1) 119879+
) = ((1 minus 119901max) [119878 (119899119879+
) minus 1] eminus119887119879 + 12
+ 120579 [119878 (119899119879+
) minus 1] eminus119887119879 + 1)
times ([119878 (119899119879+
) minus 1] eminus119887119879 + 1 + 120579)minus1
(8)
4 Abstract and Applied Analysis
Denote 119878(119899119879+) = 119878119899then the previous equation can be
rewritten as the following difference equation
119878119899+1
=(1 minus 119901max) [(119878119899 minus 1) e
minus119887119879+ 1]2
+ 120579 [(119878119899minus 1) eminus119887119879 + 1]
(119878119899minus 1) eminus119887119879 + 1 + 120579
≐ 119891 (119878119899)
(9)
which is the so-called stroboscopic map of model (5) and itdescribes the relations of the number of susceptibles in thepopulation between any two successive pulse vaccinationsConsequently the existence of the positive steady state ofmodel (9) implies the existence of a positive periodic solutionof model (5) Therefore we first discuss the conditions whichguarantee the existence of a positive steady state of (9) Takingthe derivative of 119891(119878
119899) with respect to 119878
119899yields
1198911015840
(119878119899) = (1 minus 119901max) +
119901max1205792
[(119878119899minus 1) eminus119887119879 + 1 + 120579]2
eminus119887119879
(10)
and it is easy to see that 0 lt 1198911015840(119878119899) lt 1 holds true
For convenience we denote the positive fixed point ofthe stroboscopic map (9) by 119878 which satisfies the followingequation
11986011198782
+ 1198611119878 + 1198621= 0 (11)
where
1198601= eminus119887119879 [1 minus (1 minus 119901max) e
minus119887119879
] gt 0
1198611= (eminus119887119879 minus 1) [2 (1 minus 119901max) e
minus119887119879
minus (120579 + 1)]
1198621= minus [(1 minus 119901max) (1 minus eminus119887119879)
2
+ 120579 (1 minus eminus119887119879)] lt 0
(12)
Obviously (11) has a unique positive root that is
119878 =minus1198611+ radic11986121minus 411986011198621
21198601
(13)
which is stable due to 0 lt 1198911015840(119878) lt 1According to the above discussion on the relations
between a fixed point of the stroboscopic map (9) and theperiodic solution of model (5) we conclude that the sub-model (5) has a unique nontrivial positive periodic solutiondenoted by 119878lowast(119905) and
119878lowast
(119905) = (119878 minus 1) eminus119887(119905minus119899119879) + 1 for 119899119879 lt 119905 le (119899 + 1) 119879 (14)
and it follows from 0 lt 1198911015840(119878119899) lt 1 and [30] that we have the
following lemma
Lemma 1 Model (5) has a positive periodic solution 119878lowast(119905) andfor any solution 119878(119905) of (5) we have |119878(119905) minus 119878lowast(119905)| rarr 0 as119905 rarr +infin
0 2 4 6 8 100
1
2
3
4
5
6
T
T0max
120579 = 04
120579 = 02
120579 = 03
120579 = 01
120579 = 0
R1 0
Figure 1 The effects of resource limitation parameter 120579 and pulsevaccination period 119879 on the threshold value 1198771
0with 119901max = 075
All other parameter values are given in Table 1
By using Lemma 1 we have thus shown that the uniquefixed point 119878 of (9) is globally stable
In the following we will present the sufficient conditionfor the global attraction of the disease-free periodic solution(119878lowast(119905) 0) of model (4)
Theorem 2 The disease-free periodic solution (119878lowast(119905) 0) ofmodel (4) is globally asymptotically stable in the first quadrantprovided that 1198771
0lt 1 where
1198771
0=120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]
119887119879 (120574 + 119887) (15)
here 119878 is defined in (13)
The proof of Theorem 2 is given in Appendix ANote that although the threshold value 1198771
0depends on
all parameters of model (4) the most interesting parametershere are pulse vaccination period 119879 maximum proportion ofpulse vaccination 119901max and the parameter related to resourcelimitation that is 120579 In order to address how those factors (119879and 120579) affect the threshold value 1198771
0 we first note that
lim119879rarr0
1198771
0= lim119879rarr0
120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]119887119879 (120574 + 119887)
= 0 (16)
which indicates the importance of the frequency of vaccina-tion in eradicating an infectious diseaseThen letting 120579 and 119879vary and fixing all other parameters as those in Figure 1 wesee that the threshold value 1198771
0is a monotonically increasing
function of 119879 for fixed 120579 and it is a monotonically increasingfunction of 120579 for fixed period 119879 Moreover there are somecritical values of 119879 and 120579 such that 1198771
0= 1 which indicate
that there exists one maximum allowable period of pulsevaccination 119879120579max such that 119877
1
0lt 1 for all 119879 lt 119879120579max Moreover
the larger 120579 the smaller the value that119879120579max has Unfortunately
Abstract and Applied Analysis 5
we cannot get its analytical expression due to the complexityof 11987710 So we turn to find some approximations for 119879120579max in the
followingIf the PVS is applied frequently enough such that
d119868(119905)d119905 le 0 for all 119905 gt 0 in model (4) then the numberof infectious individuals is a decreasing function of time Itis possible to satisfy this condition if pulsing ensures that119878(119905) le 119878
119888≐ (119887 + 120574)120573 for all 119905 that is pulse vaccination
is applied every time once the 119878(119905) approaches the thresholdvalue 119878
119888(see [7 27] and the references therein)
It follows from the first equation of model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (17)
Consider the following comparison equation
d119909 (119905)d119905
= 119887 minus 119887119909 (119905) 119909 (0) =(1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
(18)
which gives
119909 (119905) = 1 + ((1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
minus 1) eminus119887119905 (19)
Hence
119878 (119905) le 119909 (119905) = 1 + ((1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
minus 1) eminus119887119905 le 119878119888
(20)
and solving the above inequality with respect to 119905 yields
0 le 119905 le1
119887ln(1 +
119901max1198782
119888
(119878119888+ 120579) (1 minus 119878
119888)) (21)
which indicates that
119879120579
max =1
119887ln(1 +
119901max1198782
119888
(119878119888+ 120579) (1 minus 119878
119888)) (22)
Thus if the nonlinear pulse vaccination strategy is appliedperiodically with a period less than 119879120579max then the numberof infectious in the population will decrease forever andeventually die out
We also note that if 120579 = 0 the maximum allowableperiod 119879120579max becomes 1198790max = (1119887) ln(1 + ((119901max119878119888)(1 minus119878119888))) and this is the case studied in [7 9] for a constant
proportion pulse vaccination strategy Obviously 119879120579max is amonotonically decreasing function with respect to 120579 whichgives that 119879120579max lt 119879
0
max This theoretical result indicates that ifwe aim to eradicate an infectious disease then it is necessaryto carry out pulse vaccination programs more frequentlyunder resource limitation than when necessary resourcesare available When various nonlinear factors including119901max 120579 are seriously considered our results show that it isgetting more and more difficult to eliminate the endemicdiseases It also confirms that getting vaccinated is by farthe most effective action that susceptibles can take to protect
themselves and their family from infectious disease Otheroptions available to health practitioners in response to limitedmedical resources include the release of prescriptive healtheducation training vaccinators and promoting the use ofvaccine-delivery patchwith dissolvingmicroneedles (see [14ndash16] for details)
A typical solution of the SIR model with a nonlinearpulse vaccination strategy is shown in Figure 2 where weobserve how the variable 119878(119905) oscillates in a stable cycle(as shown in Figure 2(a)) In contrast the infection 119868(119905)
rapidly decreases to zero (Figure 2(b)) if we set 120579 = 0That is without resource limitation the infectious diseaseis eventually eradicated (Figure 2(b)) However if there isresource limitation that is the parameter 120579 is larger thanzero then both susceptible and infected populations oscillateperiodically as shown in Figures 2(c) and 2(d) with 120579 = 02
Therefore if we have enough vaccine and vaccinators tocombat the infectious disease then it is reasonable that thesusceptible population vaccinated each time is assumed to beproportional to the number of susceptible individuals (here119901max) However this approximation cannot reflect the realcase if we aim to mitigate the emerging infectious diseaseUsually the number of susceptible individuals needing tobe vaccinated may exceed the capacity of local medicalconditions due to shortages of vaccine and doctors especiallyin rural areas in many developing countries where reachingall of the target population may be difficult For instancein 2010 in Lesotho diphtheria tetanus toxoid and pertussis(DTP3) immunization coverage among 1-year-olds was only83 (but an improvement on 78 in 2001 see [31]) and boththe measles and tuberculosis (BCG) vaccination programswere hampered by supplies of vaccine running out in someparts of the country and then allowing vaccinations only forchildren born in hospitals and not for those born elsewherein the communities Thus it is important to design pulsevaccination campaigns carefully For example how frequentlyshould the vaccination strategy be applied And how can thesize of the susceptible population be evaluatedThose are keyissues for evaluating vaccination coverages and the distribu-tion of vaccines Therefore with the help of mathematicalmodels the vaccination strategy can be properly designed
It is well known that in order to eradicate an infectiousdisease the vaccination strategy must be designed such thatthe threshold value 1198771
0is less than 1 which is equivalent to
1198772
0lt 0 where
1198772
0=120573 (119878 minus 1) (1 minus eminus119887119879)
119887+ (120573 minus 120574 minus 119887) 119879 (23)
In the following we employ the formula 11987720to investigate
the important factors which affect the threshold values mostsignificantly by using uncertainty and sensitivity analysis(see [21 22 32]) Sensitivity analysis of the most significantparameters including the birth or death rate (119887) the contactrate (120573) the maximum proportion vaccinated (119901max) and thepulse vaccination period (119879) was performed by evaluatingthe PRCCs for various input parameters against the thresholdcondition 1198772
0with the LHS method [21 22 32] By using
6 Abstract and Applied Analysis
0 10 20 30 400
002
004
006
008
01
S(t)
120579 = 0
(a)
0 10 20 30 40minus800
minus600
minus400
minus200
0
t
Log(I(t))
(b)
0 10 20 30 400
002
004
006
008
01
S(t)
120579 = 02
(c)
0 10 20 30 40minus100
minus80
minus60
minus40
minus20
0
t
Log(I(t))
(d)
Figure 2 The effects of resource limitation on dynamics of model (4) with 119901max = 05 119879 = 2 and all other parameter values are given inTable 1 (a and b) 120579 = 0 (c and d) 120579 = 02
LHS with 3000 samples uncertainty and sensitivity analysesfor all parameters in model (4) were determined A uniformdistribution function was used and tested for significantPRCCs for all parameters with wide ranges such as 119887 sim
119880(0001 01) 120573 sim 119880(1000 2500) 119879 sim 119880(01 20) and 120579 sim119880(0 1) and the baseline values of all parameters are givenin the figure legend of Figure 3 Note that for comparativepurposes we chose the same baseline parameter values asthose used in literatures [8 9 33] Unfortunately we do nothave the realistic ranges of parameter values and we chosethose parameter ranges just for illustrative purposes only andthen provided a scientific basis for disease control
Figure 3(a) shows the PRCC results which illustrate thedependence of 1198772
0on each parameter and PRCC scatter
plots for each parameter are given in Figures 3(b)ndash3(g)respectively We considered absolute values of PRCC valueswhich belong to (04 1) (02 04) and (0 02) as indicatingvery importantmoderate or correlations of little significancebetween input parameters and output variables respectivelyThe positive signs of their PRCCs indicate that if the param-eters are increased the value of 1198772
0increases (vice versa)
The negative sign suggests that if increased the value of 11987720
decreases (vice versa)
Therefore the parameters 119887 120573 120579 and 119879 are responsiblefor increasing the values of 1198772
0 so decreasing all those four
parameters can reduce the threshold value and consequentlyare beneficial for infectious disease control As the parametermaximum proportion of pulse vaccination 119901max changes itis negatively and significantly correlated with large PRCCsThus increasing this parameter will result in significantlyreducing the threshold value The most significant controlparameters are 119887 120573 119901max and 119879 (as shown in Figures 3(b)3(c) 3(e) and 3(g)) It is interesting to note that the period119879and the maximum proportion vaccinated 119901max are stronglycorrelated with the threshold value 1198772
0 which indicates that
carefully designing a pulse vaccination strategy is crucial forinfectious disease control
4 Existence of Endemic Periodic Solution andComplex Dynamics
An important issue is how the infectious disease develops ifthe threshold condition 1198771
0is larger than one In particular
what types of dynamic behavior maymodel (4) have once theperiod of pulse vaccination exceeds certain threshold levelsTherefore in this section we first focus on the existence of an
Abstract and Applied Analysis 7
minus1
minus05
0
05
1
b 120573 120574 120579 T
PRCC
s
pmax
(a)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [072346 0]
b
R2 0
(b)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [05492 0]
120573
R2 0
(c)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000
120574
R2 0
[minus012283 14766e minus 011]
(d)
minus2000 0 2000minus2000
minus1000
0
1000
2000 [minus071273 0]
R2 0
pmax
(e)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [004117 0024135]
R2 0
120579
(f)
minus2000 0 2000minus2000
minus1000
0
1000
2000 [091801 0]
T
R2 0
(g)
Figure 3 PRCC results and PRCC scatter plots of the baseline parameters as those shown in Table 1 Here we fix 119901max = 075 120579 = 02 119879 = 5(a) PRCC results (bndashg) PRCC scatter plots The title of each plot represents the PRCC value with the corresponding P value
endemic periodic solution that is to investigate under whatconditions will the infected population oscillate periodicallywith small amplitude and then we choose the period ofpulse vaccination as a bifurcation parameter to investigate thecomplex patterns of the infectious disease dynamics
Next we analyze the existence of an endemic periodicsolution of model (4) near the disease-free periodic solution(119878lowast(119905) 0) using bifurcation theory [34] and set the impulsive
period 119879 as bifurcation parameter For ease of narrative werewrite the model (4) as
d119878 (119905)d119905
= 119887 minus 120573119878 (119905) 119868 (119905) minus 119887119878 (119905) ≐ 1198651(119878 (119905) 119868 (119905))
d119868 (119905)d119905
= 120573119878 (119905) 119868 (119905) minus 120574119868 (119905) minus 119887119868 (119905) ≐ 1198652(119878 (119905) 119868 (119905))
119905 = 119899119879 119899 isinN
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579≐ Θ1(119878 (119905) 119868 (119905))
119868 (119905+
) = 119868 (119905) ≐ Θ2(119878 (119905) 119868 (119905))
119905 = 119899119879 119899 isinN
(24)According to the bifurcation theorem in [34] we can
obtain the following result
Theorem 3 If there exists a 1198790gt 0 satisfying 1198771
0(1198790) = 1 and
all the parameters in model (24) satisfy
120573 (119878 (1198790) minus 1) eminus1198871198790 (
11988711987901198603
1 minus 1198603eminus1198871198790
minus 1) lt 120573 minus 119903 minus 119887 (25)
where 1198603= 1 minus 119901max + ((120579
2119901max)(120579 + 119878
lowast(1198790))2
) then thereexists a supercritical branch at the point 119879
0
8 Abstract and Applied Analysis
5 10 15 20002
004
006
008
01
T
S(t)
(a)
2 4 6 8 10004
005
006
007
008
T
S(t)
(b)
5 6 7 8 9 10005
0055
006
0065
007
T
S(t)
(c)
7 75 8005
0055
006
0065
007
T
S(t)
(d)
Figure 4 Bifurcation diagrams of model (4) with respect to bifurcation parameter 119879 Here 119901max = 075 and all other parameter values aregiven in Table 1 (a) 120579 = 0 which are reproduced from [9] (b) 120579 = 02 (c and d) a magnified part of Figure 4(b)
The existence of 1198790has been indicated in Figure 1 and the
proof ofTheorem 3 is given in Appendix BTheorem 3 showsthat there exists an endemic periodic solution under someconditions providing that the disease-free periodic solutionbecomes unstable It follows from Theorem 3 that if 119879 gt 119879
0
and is close to 1198790 the endemic periodic solution of model (4)
is stableTo investigate the complex dynamics that model (4) can
have we chose the pulse period 119879 as a bifurcation parameterand fixed all other parameters as those in Figure 4 for twodifferent 120579 values Figure 4(a) is a bifurcation diagram with-out resource limitation that is 120579 = 0 which was obtainedby Shulgin et al [9] Figure 4(b) is a bifurcation diagramwith resource limitation that is 120579 = 02 Comparing thosetwo bifurcation diagrams we conclude that the nonlinearpulse vaccination can produce more complex dynamics thanthose for the model with linear pulse vaccination Once thepulse period 119879 exceeds 119879120579max defined in (22) both susceptibleand infected populations can oscillate periodically with alarge amplitude that corresponds to periodic outbreaks ofepidemics As the pulse period is further increased a series ofcomplex and interesting bifurcation phenomena are observed(see Figures 4(a) and 4(b)) Figures 4(b)ndash4(d) indicate thatthe dynamical behavior of model (4) is very complexincluding period doubling bifurcation chaotic attractors
multistability periodic-adding chaos crisis and periodicwindowsMeanwhile bifurcation analyses (ie Figures 4(b)ndash4(d)) also indicate that the model (4) has several differentattractors which can coexist for a wide range of parametersFor example Figure 5 provides an example of two attractorcoexistence when 119879 = 78 Multiple attractor coexistenceindicates that the infectious outbreak patterns depend onthe initial values which may cause difficulties for infectiousdisease control
5 Discussion and Biological Conclusions
In order to understand the effect of resource limitation inparticular lack of vaccine on the transmission of infectiousdisease we deliberately investigated the dynamical behaviorof an SIR epidemic model which incorporates a nonlinearpulse vaccination strategy To this end we introduced anonlinear form 119901(119905) = (119901max119878(119905))(119878(119905) + 120579) as vaccinationproportionThis resulted in interesting and dramatic changesin the dynamical behavior of solutions and they becamemoreand more complicated which means that it is a very difficulttask to control infectious diseases in particular emerginginfectious diseases under resource limitation We discussedthe control strategy based on the threshold value in moredetail through theoretical analysis numerical studies and
Abstract and Applied Analysis 9
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(a)
100 110 120 130 140 150minus13
minus12
minus11
minus10
minus9
minus8
minus7
t
Log(I(t)
)
(b)
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(c)
100 110 120 130 140 150minus16
minus14
minus12
minus10
minus8
minus6
t
Log(I(t)
)
(d)
Figure 5 Coexistence of two stable attractors of model (4) Here 119901max = 075 120579 = 02 119879 = 78 and all other parameter values are given inTable 1 (a and b) The initial value (119878
0 1198680) = (012 005) (c and d) the initial value (119878
0 1198680) = (01 002)
sensitivity analysis As pointed out byAgur et al [7] the costsrisks and the efficacy of the pulse vaccinations are key factorsin eliminating transmission of the measles virus In orderto depict those factors the nonlinear resource limitationsshould be taken into account [9] and the main purpose ofthe present work is to formulate the disease model to addressthe effects of resource limitations on disease control
The results indicate that vaccination as an approach tocontrolling epidemics must be committed to a long-termstrategy especially when resources arelimited To understandthe interactions amongst 119878(119905) and 119868(119905) which are importantfor successful control of the spread of infectious diseasewe have proposed detailed modeling methods involvingvaccination on 119878(119905) and determining the most significantparameters for the basic reproductive number 1198772
0by using
LHSPRCC uncertainty and sensitivity analysis techniques(see Figure 3) Our results indicate that we should paymore attention to the vaccination period and vaccinationproportion which reduce the threshold value 1198772
0or 11987710to
prevent the outbreak of disease Our results clarify that itis more significant to improve immunization programs (ievaccinations as mentioned in [15 16]) under conditions ofresource limitation
The results also indicate that the dynamic behavior ofmodel (4) may be dramatically affected by small changes in
the value of initial densities of susceptible and infected withresource limitation Bifurcation diagrams shown in Figure 4indicate that there are many forms of complexities in model(4) which are related to chaotic bands with periodic win-dows and attractor crises and Figure 5 reveals that theoccurrence of multiple attractors is a common property ofthe SIR model with resource limitation which can help usto further understand the application of a nonlinear pulsevaccination strategy in an SIR model (for more informationabout bifurcation diagrams of impulsive control strategiessee [35 36]) Some complexities are related to the long-termbehavior of population dynamics characterized either bywell behaving relatively simple or very complicated strangeattractors We also found that the routes to chaos are verycomplicated For instance with resource limitation there areseveral hidden factors that can adversely affect the controlstrategy The increasing number of potential complexitiespredicted by the theory does not seem to make this taskany easier Nevertheless identifying complicated possiblychaotic dynamics in population data has remained a majorchallenge in epidemiology studies In particular the sug-gestion that the dynamics of measles are chaotic has beenmuch debated (eg see [37]) and when seeking to establishwhether measles dynamics in New York City were chaoticor not Sugihara and May [38] omitted data from after 1963when vaccinations were introduced as the immunisations
10 Abstract and Applied Analysis
had altered the intrinsic dynamics However our resultsimply that even with immunisations disease dynamics couldbe chaotic under some circumstances and so a reanalysis ofthe New York data including the vaccination period might beworthwhile
In the present paper we have focused on studyingthe effects of dynamical behaviors on the SIR model withnonlinear pulse vaccination For our simple model we haveassumed that the maximal vaccination proportion 119901max isa constant and the pulse period 119879 can be varied Howeverdue to limited vaccine availability the maximal vaccinationproportion 119901max should depend on the vaccine obtained ineach pulse period Thus to describe the impact of a limitedvaccine stockpile a separate differential equation for theamount of vaccine denoted by 119881(119905) could be included inthe model and then 119901(119905) should be a function of 119881(119905) and119878(119905) Meanwhile it is essential to link the costs of developingand implementing controls to population dynamic modelingof disease epidemics in order to consider other resourcelimitations such as quantities of drugs availability of isolationunits numbers of hospital beds and equipmentThese topicswill be considered in further work in the future
Appendices
A Proof of Theorem 2
Firstly we prove the local stability of a periodic solution(119878lowast(119905) 0) which may be determined by considering the
behavior of small amplitude perturbations of the solutionDefine
119878 (119905) = 119880 (119905) + 119878lowast
(119905) 119868 (119905) = 119881 (119905) (A1)
here119880(119905) 119881(119905) are small perturbations whichmay be writtenas
(119880 (119905)
119881 (119905)) = Φ (119905) (
119880 (0)
119881 (0)) 0 le 119905 lt 119879 (A2)
where the fundamental matrixΦ(119905) satisfies
dΦ (119905)d119905
= (minus119887 minus120573119878
lowast
(119905)
0 120573119878lowast
(119905) minus 120574 minus 119887)Φ (119905) (A3)
and Φ(0) = 119868 is the identity matrix So
Φ (119879) = (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 ) (A4)
where 995333 is not necessarily computed in detail as it is notrequired in the following analysis
The resetting impulsive conditions of model (4) become
(119880 (119899119879
+)
119881 (119899119879+)) = (
1 minus 119901max +1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)(119880 (119899119879)
119881 (119899119879))
≐ 119861 (119899119879) (119880 (119899119879)
119881 (119899119879))
(A5)
Hence according to Floquet theory if the moduli of botheigenvalues of the matrix
119872 = 119861 (119879)Φ (119879)
= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)
times (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 )
(A6)
are less than one then the periodic solution (119878lowast(119905) 0) islocally stable In fact two Floquet expressions multiplied arethus
1205821= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119879))2) eminus119887119879
1205822= eint
119879
0(120573119878lowast(119905)minus120574minus119887)d119905
(A7)
It is obvious that 0 lt 1205821lt 1 Thus the stability of (119878lowast(119905) 0)
is decided by whether 1205822lt 1 or not So we conclude that
the disease-free periodic solution (119878lowast(119905) 0) is locally stable ifint119879
0(120573119878lowast(119905) minus 120574 minus 119887)d119905 lt 0 that is
1
119879int
119879
0
119878lowast
(119905) d119905 =(119878 minus 1) (1 minus eminus119887119879) + 119887119879
119887119879lt120574 + 119887
120573 (A8)
and by simple calculation we have
120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]119887119879 (120574 + 119887)
lt 1 (A9)
which holds true due to 11987710lt 1 Therefore we can draw
a conclusion from the above analysis that if 11987710lt 1 the
disease-free periodic solution (119878lowast(119905) 0) of model (4) is locallyasymptotically stable
Next we will prove the global attraction of the disease-free periodic solution (119878lowast(119905) 0) of model (4) Since 1198771
0lt 1
we can choose 1205761gt 0 sufficiently small such that
1
119879int
119879
0
(119878lowast
(119905) + 1205761) d119905 lt
120574 + 119887
120573 (A10)
It follows from model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) 119905 = 119899119879
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579 119905 = 119899119879
(A11)
Consider the comparison equation
d119885 (119905)d119905
= 119887 minus 119887119885 (119905) 119905 = 119899119879
119885 (119905+
) =(1 minus 119901max) 119885
2
(119905) + 120579119885 (119905)
119885 (119905) + 120579 119905 = 119899119879
(A12)
Abstract and Applied Analysis 11
From Lemma 1 and the comparison theorem on impul-sive differential equations we have 119878(119905) le 119885(119905) and 119885(119905) rarr119878lowast(119905) as 119905 rarr +infin Hence there exists a 119905
1gt 0 such that
119878 (119905) le 119885 (119905) lt 119878lowast
(119905) + 1205761 (A13)
for all 119905 ge 1199051 here 120576
1gt 0 is small enough
From the second equation of model (4) and (A13) we get
d119868 (119905)d119905
le (120573 (119878lowast
(119905) + 1205761) minus 120574 minus 119887) 119868 (119905) (A14)
for 119905 gt 1199051 then we consider the following comparison
equation with pulses
d1199101(119905)
d119905= (120573 (119878
lowast
(119905) + 1205761) minus 120574 minus 119887) 119910
1(119905) 119905 = 119899119879
1199101(119905+
) = 1199101(119905) 119905 = 119899119879
(A15)
Integrating model (A15) between pulses (119899119879 (119899 + 1)119879] wehave
1199101((119899 + 1) 119879) = 119910
1(119899119879) eint
(119899+1)119879
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101(119899119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A16)
Then by using step by step iterations
1199101(119899119879) = 119910
1((119899 minus 1) 119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101((119899 minus 2) 119879) e2 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= sdot sdot sdot
= 1199101(0) e119899 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A17)
where 1199101(0) = 119910
1(0+) gt 0 we obtain lim
119899rarrinfin1199101(119899119879) =
0 according to (A10) On the other hand integrating andsolving the first equation of model (A15) between pulses itgives
1199101(119905) = 119910
1(119899119879) eint
119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905 119905 isin (119899119879 (119899 + 1) 119879]
(A18)
incorporating into the boundedness of eint119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
due to 11987710lt 1 we have lim
119905rarrinfin1199101(119905) = 0
Let (119878(119905) 119868(119905)) be any solution of model (4) with initialvalue (119878
0 1198680) and 119878
0= 119878(0
+) gt 0 119868
0= 119868(0
+) =
1199101(0) gt 0 according to the comparison theorem we have
lim119905rarrinfin
sup 119868(119905) le lim119905rarrinfin
sup1199101(119905) = 0 Incorporating into
the positivity of 119868(119905) we know that lim119905rarrinfin
119868(119905) = 0Consequently there exists a 119905
2gt 1199051such that 0 lt 119868(119905) le 120576
2
for 119905 ge 1199052 and 120576
2is small enough Further we have
119887 minus (1205731205762+ 119887) 119878 (119905) le
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (A19)
for all 119905 gt 1199052 from which we can obtain the following
equation
d1199102(119905)
d119905= 119887 minus (120573120576
2+ 119887) 119910
2(119905) 119905 = 119899119879
1199102(119905+
) =(1 minus 119901max) 119910
2
2(119905) + 120579119910
2(119905)
1199102(119905) + 120579
119905 = 119899119879
(A20)
By employing the same methods as used for the proof ofLemma 1 we get that the model (A20) has a positive periodicsolution 119910lowast
2(119905) which is globally attractive where
119910lowast
2(119905) = (119910
2minus
119887
1205731205762+ 119887) eminus(1205731205762+119887)(119905minus119899119879) + 119887
1205731205762+ 119887
119905 isin (119899119879 (119899 + 1) 119879]
1199102= 1199102(119899119879+
) = 119910lowast
2(0+
) =minus1198612+ radic11986122minus 411986021198622
21198602
1198602= eminus(1205731205762+119887)119879 [ 119887
1205731205762+ 119887
minus (1 minus 119901max) eminus(1205731205762+119887)119879]
1198612= (eminus(1205731205762+119887)119879 minus 119887
1205731205762+ 119887)
times [2 (1 minus 119901max) eminus(1205731205762+119887)119879minus (120579 +
119887
1205731205762+ 119887)]
1198622= minus[(1 minus 119901max) (
119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)2
+ 120579(119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)]
(A21)
It follows from the comparison theorem on impulsive differ-ential equations that 119910
2(119905) le 119878(119905) le 119885(119905) Moreover 119910
2(119905) rarr
119910lowast
2(119905) and 119885(119905) rarr 119878
lowast(119905) as 119905 rarr +infin Consequently there
exists a 1199053for 1205763small enough such that 119905
3ge 1199052and
119910lowast
2(119905) minus 120576
3lt 119878 (119905) lt 119878
lowast
(119905) + 1205763
(A22)
for 119905 ge 1199053 Let 120576
2rarr 0 Then
119878lowast
(119905) minus 1205763lt 119878 (119905) lt 119878
lowast
(119905) + 1205763 (A23)
Therefore 119878(119905) rarr 119878lowast(119905) as 119905 rarr +infin Thus we
have proved the global stability of the disease-free periodicsolution (119878lowast(119905) 0) of model (4) This completes the proof
B Proof of Theorem 3
In order to apply the bifurcation theory of [34] in the maintext we make the following calculations
1198891015840
0= 1 minus eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
(B1)
If 11988910158400= 0 then 119879
0satisfies the following equation
120573 [(119878 minus 1) (1 minus eminus1198871198790) + 1198871198790]
119887 (120574 + 119887) 1198790
= 1 (B2)
12 Abstract and Applied Analysis
which indicates that there exists a1198790such that 1198771
0= 1 and the
disease-free periodic solution 120575 = (119878lowast(119905) 0) loses its stabilityFurther
120597Φ1(1198790 1198830)
120597119878= eminus1198871198790 gt 0
120597Φ2(1198790 1198830)
120597119868= eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
120597Φ1(1198790 1198830)
120597119868= minusint
1198790
0
120573119878lowast
(V) eminus119887(1198790minusV)eintV
0(120573119878lowast(119905)minus120574minus119887)d119905dV
≐ 119860 lt 0
1198861015840
0= 1 minus 119860
3eminus1198871198790
1198871015840
0= 1 minus 119860
3119860
1205972Φ2(1198790 1198830)
120597119868120597119878= 1205731198790eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
1205972Φ2(1198790 1198830)
1205971198682
= minus1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
119878lowast
(120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585minus119887(Vminus120579)
d120579 dV
lt 0
1205972Φ2(1198790 1198830)
120597119868120597119879= (120573119878
lowast
(1198790) minus 120574 minus 119887) eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
120597Φ1(1198790 1198830)
120597119879= minus119887 (119878 minus 1) eminus1198871198790
119861 = 1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887)
times eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
119862 = 1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
eminus119887(119905minus120579)119878lowast (120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585
d120579 dV
+ 21205731198790
1 minus 1198603119860
1 minus 1198603eminus1198871198790
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
(B3)
It follows that 119861119862 lt 0 and we get the condition that is if theparameters satisfy condition 119861 lt 0 then the model (4) has asupercritical branch at 119879
0
In fact
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0 (B4)
Therefore 119861 lt 0 is equivalent to
1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887) lt 0
(B5)
Obviously 119861 lt 0 due to (25) The proof is completed
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thiswork is supported by theNational Natural Science Foun-dation of China (NSFCs 11171199 11371030 11301320) andthe Fundamental Research Funds for the Central Universities(GK201305010) The authors would like to thank the editorand the referees for their careful reading of the original paperand many valuable comments and suggestions that greatlyimproved the presentation of this paper
References
[1] R M Anderson and R M May ldquoPopulation biology of infec-tious diseases part Irdquo Nature vol 280 no 5721 pp 361ndash3671979
[2] E Hanert E Schumacher and E Deleersnijder ldquoFront dynam-ics in fractional-order epidemic modelsrdquo Journal of TheoreticalBiology vol 279 no 1 pp 9ndash16 2011
[3] H W Hethcote ldquoMathematics of infectious diseasesrdquo SIAMReview vol 42 no 4 pp 599ndash653 2000
[4] A E Fiore C B Bridges and N J Cox ldquoSeasonal influenzavaccinesrdquo Current Topics in Microbiology and Immunology vol333 no 1 pp 43ndash82 2009
[5] Y Chang N T Brewer A C Rinas K Schmitt and J S SmithldquoEvaluating the impact of human papillomavirus vaccinesrdquoVaccine vol 27 no 32 pp 4355ndash4362 2009
[6] T J Liesegang ldquoVaricella zoster virus vaccines effective butconcerns lingerrdquo Canadian Journal of Ophthalmology vol 44no 4 pp 379ndash384 2009
[7] Z Agur L Cojocaru G Mazor R M Anderson and Y LDanon ldquoPulse mass measles vaccination across age cohortsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 90 no 24 pp 11698ndash11702 1993
[8] L Stone B Shulgin and Z Agur ldquoTheoretical examinationof the pulse vaccination policy in the SIR epidemic modelrdquoMathematical and ComputerModelling vol 31 no 4-5 pp 207ndash215 2000
[9] B Shulgin L Stone and Z Agur ldquoPulse vaccination strategy inthe SIR epidemic modelrdquo Bulletin of Mathematical Biology vol60 no 6 pp 1123ndash1148 1998
[10] A DrsquoOnofrio ldquoStability properties of pulse vaccination strategyin SEIR epidemicmodelrdquoMathematical Biosciences vol 179 no1 pp 57ndash72 2002
[11] A Franceschetti and A Pugliese ldquoThreshold behaviour of a SIRepidemicmodel with age structure and immigrationrdquo Journal ofMathematical Biology vol 57 no 1 pp 1ndash27 2008
Abstract and Applied Analysis 13
[12] A J Terry ldquoPulse vaccination strategies in a metapopulationSIR modelrdquo Mathematical Biosciences and Engineering vol 7no 2 pp 455ndash477 2010
[13] S J Gao L S Chen J J Nieto and A Torres ldquoAnalysis of adelayed epidemic model with pulse vaccination and saturationincidencerdquo Vaccine vol 24 no 35-36 pp 6037ndash6045 2006
[14] L Matrajt M E Halloran and I M Longini Jr ldquoOptimal vac-cine allocation for the early mitigation of pandemic influenzardquoPLoS Computational Biology vol 9 no 3 Article ID e10029642013
[15] S P Sullivan D G Koutsonanos M D P Martin et alldquoDissolving polymer microneedle patches for influenza vacci-nationrdquo Nature Medicine vol 16 no 8 pp 915ndash920 2010
[16] Science Daily Web ldquoVaccine-delivery patch with dissolvingmicroneedles eliminates ldquosharpsrdquo boosts protectionrdquo httpwwwsciencedailycomreleases201007100718204733htm
[17] M J Reynolds-Hogland J S Hogland and M S MitchellldquoEvaluating intercepts fromdemographicmodels to understandresource limitation and resource thresholdsrdquo Ecological Mod-elling vol 211 no 3-4 pp 424ndash432 2008
[18] L Chow M Fan and Z L Feng ldquoDynamics of a multigroupepidemiological model with group-targeted vaccination strate-giesrdquo Journal of Theoretical Biology vol 291 no 1 pp 56ndash642011
[19] L H Zhou and M Fan ldquoDynamics of an SIR epidemic modelwith limited medical resources revisitedrdquo Nonlinear AnalysisReal World Applications vol 13 no 1 pp 312ndash324 2012
[20] V Lakshmikantham D D Bainov and P S Simeonov Theoryof Impulsive Differential Equations World Scientific Singapore1989
[21] S M Blower and H Dowlatabadi ldquoSensitivity and uncertaintyanalysis of complex-models of disease transmission An HIVmodel as an examplerdquo International Statistical Review vol 62no 2 pp 229ndash243 1994
[22] S Marino I B Hogue C J Ray and D E Kirschner ldquoA meth-odology for performing global uncertainty and sensitivityanalysis in systems biologyrdquo Journal of Theoretical Biology vol254 no 1 pp 178ndash196 2008
[23] S Y Tang J H Liang Y S Tan and R A Cheke ldquoThresholdconditions for integrated pest management models with pesti-cides that have residual effectsrdquo Journal ofMathematical Biologyvol 66 no 1-2 pp 1ndash35 2013
[24] F Brauer and C Castillo-ChavezMathematical Models in Pop-ulation Biology and Epidemiology Springer New York NYUSA 2000
[25] O Diekmann and J A P Heesterbeek Mathematical Epidemi-ology of Infectious Diseases JohnWiley amp Sons Chichester UK2000
[26] J D MurrayMathematical Biology Springer Berlin Germany1989
[27] R Anderson and R May Infectious Diseases of HumansDynamics and Control Oxford University Press Oxford UK1995
[28] M Hethcote ldquoThree basic epidemiological modelsrdquo in AppliedMathematical Ecology L Gross T G Hallam and S A LevinEds pp 119ndash144 Springer Berlin Germany 1989
[29] A V Hill ldquoThe possible effects of the aggregation of themolecules of haemoglobin on its dissociation curvesrdquo TheJournal of Physiology vol 40 no 4 pp 4ndash7 1910
[30] P Cull ldquoLocal and global stability for population modelsrdquoBiological Cybernetics vol 54 no 3 pp 141ndash149 1986
[31] WHOWeb ldquoGlobal health observatory data repositoryrdquo 2001httpappswhointghodatatheme=country
[32] M D McKay R J Beckman and W J Conover ldquoComparisonof three methods for selecting values of input variables in theanalysis of output from a computer coderdquo Technometrics vol21 no 2 pp 239ndash245 1979
[33] W M Schaffer L F Olsen G L Truty S L Fulmer andD J Graser ldquoPeriodic and chaotic dynamics in childhoodepidemicsrdquo in From Chemical to Biological Organization MMarkus S Muller and G Nicolis Eds Springer BerlinGermany 1988
[34] A Lakmeche and O Arino ldquoBifurcation of non trivial periodicsolutions of impulsive differential equations arising chemother-apeutic treatmentrdquo Dynamics of Continuous Discrete andImpulsive Systems B vol 7 no 2 pp 265ndash287 2000
[35] S Y Tang and L S Chen ldquoMultiple attractors in stage-structured population models with birth pulsesrdquo Bulletin ofMathematical Biology vol 65 no 3 pp 479ndash495 2003
[36] S Y Tang Y N Xiao and R A Cheke ldquoMultiple attractorsof host-parasitoid models with integrated pest managementstrategies eradication persistence and outbreakrdquo TheoreticalPopulation Biology vol 73 no 2 pp 181ndash197 2008
[37] L F Olsen andWM Schaffer ldquoChaos versus noisy periodicityalternative hypotheses for childhood epidemicsrdquo Science vol249 no 4968 pp 499ndash504 1990
[38] G Sugihara and R M May ldquoNonlinear forecasting as a way ofdistinguishing chaos from measurement error in time seriesrdquoNature vol 344 no 6268 pp 734ndash741 1990
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical Problems in Engineering
Hindawi Publishing Corporationhttpwwwhindawicom
Differential EquationsInternational Journal of
Volume 2014
Applied MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Probability and StatisticsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical PhysicsAdvances in
Complex AnalysisJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
OptimizationJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CombinatoricsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Operations ResearchAdvances in
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Function Spaces
Abstract and Applied AnalysisHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of Mathematics and Mathematical Sciences
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Algebra
Discrete Dynamics in Nature and Society
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Decision SciencesAdvances in
Discrete MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014 Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Stochastic AnalysisInternational Journal of

2 Abstract and Applied Analysis
the eradication of a disease with some practical advantagesas discussed in [7ndash9] Recently epidemic models with pulsevaccination have been the subject of intense theoreticalanalysis For example the dynamical behaviors of variousepidemic models with PVS are studied in [8 10ndash13] Thetheoretical results show that PVS can be distinguished fromthe conventional strategies in leading to disease eradicationat relatively low values of vaccination (see [7])
Traditional epidemic models with PVS of populationdynamics have assumed that the pulse vaccination propor-tion 119901 is constant which implies that medical resourcessuch as drugs vaccines hospital beds and isolation unitsare sufficient for the infectious disease in question Howeverin reality every community or country has an appropriateor limited capacity for treatment especially for emerginginfectious diseases and so understanding resource limitationis critical to effective management
During the last decade the outbreaks of SARS in 2003and avian influenza among humans (H5N1 in 2003 H1N1 in2009 andH7N9 in 2013) emphasized the need to enhance thecapacity to fight emerging infectious diseases which remainsa challenge for public health Several different vaccines havebeen developed specifically targeting avian influenza amonghumans but limited production has exposed weaknesses[14ndash16] A lack of sufficient vaccine at the outset of a newpandemic is to be expected given the crucial lag betweenthe timing of the emergence of a new strain and when avaccine has been developed and is ready for distributionAlthough most vaccination programmes aim to vaccinate asmany susceptible individuals as possible this is not always thecase for emerging infectious diseases due to limited vaccineavailability particularly in developing countries
However resource limitation is difficult to quantify partlybecause it is a dynamic process (see [17]) Recently continu-ous SIR models concerning limited medical resources havebeen developed and investigated (see [18 19]) Chow et al[18] explored the dynamics of an SIR epidemic model tounderstand how limited medical resources and their supplyefficiency affect the transmission of infectious diseases Zhouand Fan [19] studied a multigroup SIR epidemiologicalmodel to explore the effects of limited medical resourcesand group-targeted vaccination strategies on disease controland prevention To the best of our knowledge no workhas been done for the effects of resource limitation on anSIR model with PVS In order to investigate the effect oflimited vaccine availability on the spread of infectious diseasea saturation phenomenon of limited medical resources isconsideredThat is we will study the dynamic behavior of anSIR epidemic model with nonlinear pulse vaccination Suchmathematical models are suitable for simulating processeswith short duration perturbations during their development(see [20])
In this paper the dynamical behavior of an SIR epidemicmodel with nonlinear pulse vaccination is proposed and ana-lyzed The main purpose is to address and understand how alimited vaccine resource affects the transmission and controlof infectious diseases The rest of this paper is organized asfollows In the next section an SIR epidemic model withnonlinear PVS is introduced In Section 3 we investigate the
existence and stability of the disease-free periodic solutionby using the method of differential inequality qualitativeanalysis a discrete dynamical system determined by the stro-boscopic map and comparison theorem Meanwhile LatinHypercube Sampling (LHS)Partial Rank Correlation Coef-ficients (PRCCs) uncertainty and sensitivity analysis tech-niques are employed to investigate the key control parameterswhich are most significantly related to threshold values (see[21ndash23]) Section 4 focuses on finding the sufficient conditionunderwhich the endemic periodic solution exists by using thebifurcation theorem The paper ends with some interestingbiological conclusions and numerical bifurcation analyseswhich complement the theoretical findings
2 Model Formulation
In the classical epidemiological model (see [24ndash26]) the totalpopulation 119873 is composed of three groups of individualssusceptible (119878) infective (119868) and recovered (119877) Let 119878(119905)denote the number ofmembers of a population susceptible tothe disease at time 119905 119868(119905) represents the number of infectivemembers and 119877(119905) denotes the number of members whohave been removed from the population This leads to thefollowing SIR model
d119878 (119905)d119905
= 119887 minus 120573119878 (119905) 119868 (119905) minus 119887119878 (119905)
d119868 (119905)d119905
= 120573119878 (119905) 119868 (119905) minus 120574119868 (119905) minus 119887119868 (119905)
d119877 (119905)d119905
= 120574119868 (119905) minus 119887119877 (119905)
(1)
In model (1) we assume that the three classes of subpop-ulations have the same constant birth and death rates 119887(ie the total population has a constant size) which can benormalized to unity that is119873(119905) = 119878(119905)+ 119868(119905)+119877(119905) = 1 andthe infectious individuals recover at a rate 120574 gt 0 so that 1120574is the mean infectious period Susceptibles become infectedat a rate 120573119868 where 120573 gt 0 is the transmission rate In practicethe equation for d119877(119905)d119905 is redundant because 119877(119905) can beobtained from119873(119905) = 1 A detailed description of model (1)and its dynamics may be found in [9 27 28]
Let 119879 gt 0 be the time between two consecutive pulsevaccinations and let 119901(119905) (0 le 119901(119905) lt 1) be the fraction ofsusceptible subjects who are inoculated with vaccine whichdepends on the number of susceptibles in the population attime 119905 This yields the following model with PVS [8 9]
d119878 (119905)d119905
= 119887 minus 120573119878 (119905) 119868 (119905) minus 119887119878 (119905)
d119868 (119905)d119905
= 120573119878 (119905) 119868 (119905) minus 120574119868 (119905) minus 119887119868 (119905)
119905 = 119899119879 119899 isinN
119878 (119905+) = (1 minus 119901 (119905)) 119878 (119905)
119868 (119905+) = 119868 (119905)
119905 = 119899119879 119899 isinN
(2)
hereN = 0 1 2
Abstract and Applied Analysis 3
Table 1 Parameter definitions values and source for model (4)
Parameter Definition Value Source119887 Rate of natural birth or death rate of the population (yearminus1) 002 [33]120573 Probability of transmission per contact (yearminus1) 1800 [33]120574 Natural recovery rate of the infective individuals (yearminus1) 100 [33]119901max Themaximal vaccination proportion (yearminus1) [0 1) [8 9]120579 The half saturation constant [0 1] Assumption119879 Period of pulse vaccination (year) [0 20] [8 9]
The formula for 119901(119905) plays a key role in investigating theeffect of the limited capacity for treatment on the spread ofinfectious disease To do this we assume without loss of gen-erality that the PVS is implemented in a developing countrybut one in which vaccines are relatively abundant Note thatthere are often major shortages of medical personnel in ruralareas and in certain specialities such as surgery paediatricsand obstetrics [14] Let vaccination pulses occur every 119879 timeunits When a pulse day occurs the medical personnel haveto go out into the rural population and vaccinate as manysusceptibles as they can find on that particular day If thereare very few susceptibles then it will be hard to find them sothe vaccination coverage will be low The more susceptiblesthere are the easier it is to find them so the coverage per pulsewill be greater But there is a limit to how fast the medicalteam can find and handle each susceptible and this causesthe vaccination rate 119901(119905) to saturate that is a saturationphenomenon of limited medical resources By employingthe Holling type II functional response for a predator-preymodel we can define 119901(119905) as follows
119901 (119905) =119901max119878 (119905)
119878 (119905) + 120579 0 le 119901max lt 1 (3)
where 119901max denotes the maximal vaccination proportion and120579 is the half saturation constant Note that the vaccinationcoverage is less than 100 and in practice a vaccine efficacyis also less than 100 which lead to the parameter 119901max lt 1Furthermore generalized formulae such as the Hill function[29] can be employed to characterize the saturation phe-nomenon of the limited medical resources
Taking into account the resource limitation and satura-tion effects for convenience and simplification then model(2) can be rewritten as
d119878 (119905)d119905
= 119887 minus 120573119878 (119905) 119868 (119905) minus 119887119878 (119905)
d119868 (119905)d119905
= 120573119878 (119905) 119868 (119905) minus 120574119868 (119905) minus 119887119868 (119905)
119905 = 119899119879 119899 isinN
119878 (119905+) =
(1 minus 119901max) 1198782
(119905) + 120579119878 (119905)
119878 (119905) + 120579
119868 (119905+) = 119868 (119905)
119905 = 119899119879 119899 isinN
(4)
where the parameters 119887 120573 120574 and 119901max 120579 are defined in mod-els (1) and (3) respectively Note that model (4) is reduced tothe classical SIRmodel with constant pulse vaccination when120579 = 0 which was investigated by Agur et al [7] Stone et al
[8] and Shulgin et al [9]The parameter definitions of model(4) are summarized in Table 1
3 Existence of the Disease-FreePeriodic Solution
We first demonstrate the existence of a disease-free periodicsolution of model (4) in which infectious individuals areentirely absent from the population permanently that is119868(119905) = 0 for a sufficiently long time Under some thresholdconditions we show below that the susceptible populationoscillates with period 119879 in synchronization with the periodicpulse vaccination and the infectious class will die out eventu-allyThus in this section we focus on determining the globalattraction of this disease-free periodic solution To addressthis we first consider the following submodel over the timeinterval 119899119879 lt 119905 le (119899 + 1)119879
d119878 (119905)d119905
= 119887 minus 119887119878 (119905) 119905 = 119899119879
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579 119905 = 119899119879
119878 (0+
) = 1198780
(5)
Integrating the first equation of model (5) between pulsesyields
119878 (119905) = [119878 (119899119879+
) minus 1] eminus119887(119905minus119899119879) + 1 (6)
for 119899119879 lt 119905 le (119899 + 1)119879 It follows from the second equation ofmodel (5) that
119878 ((119899 + 1) 119879+
) =(1 minus 119901max) 119878
2
((119899 + 1) 119879) + 120579119878 ((119899 + 1) 119879)
119878 ((119899 + 1) 119879) + 120579
(7)
Substitution of (6) into (7) gives
119878 ((119899 + 1) 119879+
) = ((1 minus 119901max) [119878 (119899119879+
) minus 1] eminus119887119879 + 12
+ 120579 [119878 (119899119879+
) minus 1] eminus119887119879 + 1)
times ([119878 (119899119879+
) minus 1] eminus119887119879 + 1 + 120579)minus1
(8)
4 Abstract and Applied Analysis
Denote 119878(119899119879+) = 119878119899then the previous equation can be
rewritten as the following difference equation
119878119899+1
=(1 minus 119901max) [(119878119899 minus 1) e
minus119887119879+ 1]2
+ 120579 [(119878119899minus 1) eminus119887119879 + 1]
(119878119899minus 1) eminus119887119879 + 1 + 120579
≐ 119891 (119878119899)
(9)
which is the so-called stroboscopic map of model (5) and itdescribes the relations of the number of susceptibles in thepopulation between any two successive pulse vaccinationsConsequently the existence of the positive steady state ofmodel (9) implies the existence of a positive periodic solutionof model (5) Therefore we first discuss the conditions whichguarantee the existence of a positive steady state of (9) Takingthe derivative of 119891(119878
119899) with respect to 119878
119899yields
1198911015840
(119878119899) = (1 minus 119901max) +
119901max1205792
[(119878119899minus 1) eminus119887119879 + 1 + 120579]2
eminus119887119879
(10)
and it is easy to see that 0 lt 1198911015840(119878119899) lt 1 holds true
For convenience we denote the positive fixed point ofthe stroboscopic map (9) by 119878 which satisfies the followingequation
11986011198782
+ 1198611119878 + 1198621= 0 (11)
where
1198601= eminus119887119879 [1 minus (1 minus 119901max) e
minus119887119879
] gt 0
1198611= (eminus119887119879 minus 1) [2 (1 minus 119901max) e
minus119887119879
minus (120579 + 1)]
1198621= minus [(1 minus 119901max) (1 minus eminus119887119879)
2
+ 120579 (1 minus eminus119887119879)] lt 0
(12)
Obviously (11) has a unique positive root that is
119878 =minus1198611+ radic11986121minus 411986011198621
21198601
(13)
which is stable due to 0 lt 1198911015840(119878) lt 1According to the above discussion on the relations
between a fixed point of the stroboscopic map (9) and theperiodic solution of model (5) we conclude that the sub-model (5) has a unique nontrivial positive periodic solutiondenoted by 119878lowast(119905) and
119878lowast
(119905) = (119878 minus 1) eminus119887(119905minus119899119879) + 1 for 119899119879 lt 119905 le (119899 + 1) 119879 (14)
and it follows from 0 lt 1198911015840(119878119899) lt 1 and [30] that we have the
following lemma
Lemma 1 Model (5) has a positive periodic solution 119878lowast(119905) andfor any solution 119878(119905) of (5) we have |119878(119905) minus 119878lowast(119905)| rarr 0 as119905 rarr +infin
0 2 4 6 8 100
1
2
3
4
5
6
T
T0max
120579 = 04
120579 = 02
120579 = 03
120579 = 01
120579 = 0
R1 0
Figure 1 The effects of resource limitation parameter 120579 and pulsevaccination period 119879 on the threshold value 1198771
0with 119901max = 075
All other parameter values are given in Table 1
By using Lemma 1 we have thus shown that the uniquefixed point 119878 of (9) is globally stable
In the following we will present the sufficient conditionfor the global attraction of the disease-free periodic solution(119878lowast(119905) 0) of model (4)
Theorem 2 The disease-free periodic solution (119878lowast(119905) 0) ofmodel (4) is globally asymptotically stable in the first quadrantprovided that 1198771
0lt 1 where
1198771
0=120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]
119887119879 (120574 + 119887) (15)
here 119878 is defined in (13)
The proof of Theorem 2 is given in Appendix ANote that although the threshold value 1198771
0depends on
all parameters of model (4) the most interesting parametershere are pulse vaccination period 119879 maximum proportion ofpulse vaccination 119901max and the parameter related to resourcelimitation that is 120579 In order to address how those factors (119879and 120579) affect the threshold value 1198771
0 we first note that
lim119879rarr0
1198771
0= lim119879rarr0
120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]119887119879 (120574 + 119887)
= 0 (16)
which indicates the importance of the frequency of vaccina-tion in eradicating an infectious diseaseThen letting 120579 and 119879vary and fixing all other parameters as those in Figure 1 wesee that the threshold value 1198771
0is a monotonically increasing
function of 119879 for fixed 120579 and it is a monotonically increasingfunction of 120579 for fixed period 119879 Moreover there are somecritical values of 119879 and 120579 such that 1198771
0= 1 which indicate
that there exists one maximum allowable period of pulsevaccination 119879120579max such that 119877
1
0lt 1 for all 119879 lt 119879120579max Moreover
the larger 120579 the smaller the value that119879120579max has Unfortunately
Abstract and Applied Analysis 5
we cannot get its analytical expression due to the complexityof 11987710 So we turn to find some approximations for 119879120579max in the
followingIf the PVS is applied frequently enough such that
d119868(119905)d119905 le 0 for all 119905 gt 0 in model (4) then the numberof infectious individuals is a decreasing function of time Itis possible to satisfy this condition if pulsing ensures that119878(119905) le 119878
119888≐ (119887 + 120574)120573 for all 119905 that is pulse vaccination
is applied every time once the 119878(119905) approaches the thresholdvalue 119878
119888(see [7 27] and the references therein)
It follows from the first equation of model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (17)
Consider the following comparison equation
d119909 (119905)d119905
= 119887 minus 119887119909 (119905) 119909 (0) =(1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
(18)
which gives
119909 (119905) = 1 + ((1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
minus 1) eminus119887119905 (19)
Hence
119878 (119905) le 119909 (119905) = 1 + ((1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
minus 1) eminus119887119905 le 119878119888
(20)
and solving the above inequality with respect to 119905 yields
0 le 119905 le1
119887ln(1 +
119901max1198782
119888
(119878119888+ 120579) (1 minus 119878
119888)) (21)
which indicates that
119879120579
max =1
119887ln(1 +
119901max1198782
119888
(119878119888+ 120579) (1 minus 119878
119888)) (22)
Thus if the nonlinear pulse vaccination strategy is appliedperiodically with a period less than 119879120579max then the numberof infectious in the population will decrease forever andeventually die out
We also note that if 120579 = 0 the maximum allowableperiod 119879120579max becomes 1198790max = (1119887) ln(1 + ((119901max119878119888)(1 minus119878119888))) and this is the case studied in [7 9] for a constant
proportion pulse vaccination strategy Obviously 119879120579max is amonotonically decreasing function with respect to 120579 whichgives that 119879120579max lt 119879
0
max This theoretical result indicates that ifwe aim to eradicate an infectious disease then it is necessaryto carry out pulse vaccination programs more frequentlyunder resource limitation than when necessary resourcesare available When various nonlinear factors including119901max 120579 are seriously considered our results show that it isgetting more and more difficult to eliminate the endemicdiseases It also confirms that getting vaccinated is by farthe most effective action that susceptibles can take to protect
themselves and their family from infectious disease Otheroptions available to health practitioners in response to limitedmedical resources include the release of prescriptive healtheducation training vaccinators and promoting the use ofvaccine-delivery patchwith dissolvingmicroneedles (see [14ndash16] for details)
A typical solution of the SIR model with a nonlinearpulse vaccination strategy is shown in Figure 2 where weobserve how the variable 119878(119905) oscillates in a stable cycle(as shown in Figure 2(a)) In contrast the infection 119868(119905)
rapidly decreases to zero (Figure 2(b)) if we set 120579 = 0That is without resource limitation the infectious diseaseis eventually eradicated (Figure 2(b)) However if there isresource limitation that is the parameter 120579 is larger thanzero then both susceptible and infected populations oscillateperiodically as shown in Figures 2(c) and 2(d) with 120579 = 02
Therefore if we have enough vaccine and vaccinators tocombat the infectious disease then it is reasonable that thesusceptible population vaccinated each time is assumed to beproportional to the number of susceptible individuals (here119901max) However this approximation cannot reflect the realcase if we aim to mitigate the emerging infectious diseaseUsually the number of susceptible individuals needing tobe vaccinated may exceed the capacity of local medicalconditions due to shortages of vaccine and doctors especiallyin rural areas in many developing countries where reachingall of the target population may be difficult For instancein 2010 in Lesotho diphtheria tetanus toxoid and pertussis(DTP3) immunization coverage among 1-year-olds was only83 (but an improvement on 78 in 2001 see [31]) and boththe measles and tuberculosis (BCG) vaccination programswere hampered by supplies of vaccine running out in someparts of the country and then allowing vaccinations only forchildren born in hospitals and not for those born elsewherein the communities Thus it is important to design pulsevaccination campaigns carefully For example how frequentlyshould the vaccination strategy be applied And how can thesize of the susceptible population be evaluatedThose are keyissues for evaluating vaccination coverages and the distribu-tion of vaccines Therefore with the help of mathematicalmodels the vaccination strategy can be properly designed
It is well known that in order to eradicate an infectiousdisease the vaccination strategy must be designed such thatthe threshold value 1198771
0is less than 1 which is equivalent to
1198772
0lt 0 where
1198772
0=120573 (119878 minus 1) (1 minus eminus119887119879)
119887+ (120573 minus 120574 minus 119887) 119879 (23)
In the following we employ the formula 11987720to investigate
the important factors which affect the threshold values mostsignificantly by using uncertainty and sensitivity analysis(see [21 22 32]) Sensitivity analysis of the most significantparameters including the birth or death rate (119887) the contactrate (120573) the maximum proportion vaccinated (119901max) and thepulse vaccination period (119879) was performed by evaluatingthe PRCCs for various input parameters against the thresholdcondition 1198772
0with the LHS method [21 22 32] By using
6 Abstract and Applied Analysis
0 10 20 30 400
002
004
006
008
01
S(t)
120579 = 0
(a)
0 10 20 30 40minus800
minus600
minus400
minus200
0
t
Log(I(t))
(b)
0 10 20 30 400
002
004
006
008
01
S(t)
120579 = 02
(c)
0 10 20 30 40minus100
minus80
minus60
minus40
minus20
0
t
Log(I(t))
(d)
Figure 2 The effects of resource limitation on dynamics of model (4) with 119901max = 05 119879 = 2 and all other parameter values are given inTable 1 (a and b) 120579 = 0 (c and d) 120579 = 02
LHS with 3000 samples uncertainty and sensitivity analysesfor all parameters in model (4) were determined A uniformdistribution function was used and tested for significantPRCCs for all parameters with wide ranges such as 119887 sim
119880(0001 01) 120573 sim 119880(1000 2500) 119879 sim 119880(01 20) and 120579 sim119880(0 1) and the baseline values of all parameters are givenin the figure legend of Figure 3 Note that for comparativepurposes we chose the same baseline parameter values asthose used in literatures [8 9 33] Unfortunately we do nothave the realistic ranges of parameter values and we chosethose parameter ranges just for illustrative purposes only andthen provided a scientific basis for disease control
Figure 3(a) shows the PRCC results which illustrate thedependence of 1198772
0on each parameter and PRCC scatter
plots for each parameter are given in Figures 3(b)ndash3(g)respectively We considered absolute values of PRCC valueswhich belong to (04 1) (02 04) and (0 02) as indicatingvery importantmoderate or correlations of little significancebetween input parameters and output variables respectivelyThe positive signs of their PRCCs indicate that if the param-eters are increased the value of 1198772
0increases (vice versa)
The negative sign suggests that if increased the value of 11987720
decreases (vice versa)
Therefore the parameters 119887 120573 120579 and 119879 are responsiblefor increasing the values of 1198772
0 so decreasing all those four
parameters can reduce the threshold value and consequentlyare beneficial for infectious disease control As the parametermaximum proportion of pulse vaccination 119901max changes itis negatively and significantly correlated with large PRCCsThus increasing this parameter will result in significantlyreducing the threshold value The most significant controlparameters are 119887 120573 119901max and 119879 (as shown in Figures 3(b)3(c) 3(e) and 3(g)) It is interesting to note that the period119879and the maximum proportion vaccinated 119901max are stronglycorrelated with the threshold value 1198772
0 which indicates that
carefully designing a pulse vaccination strategy is crucial forinfectious disease control
4 Existence of Endemic Periodic Solution andComplex Dynamics
An important issue is how the infectious disease develops ifthe threshold condition 1198771
0is larger than one In particular
what types of dynamic behavior maymodel (4) have once theperiod of pulse vaccination exceeds certain threshold levelsTherefore in this section we first focus on the existence of an
Abstract and Applied Analysis 7
minus1
minus05
0
05
1
b 120573 120574 120579 T
PRCC
s
pmax
(a)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [072346 0]
b
R2 0
(b)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [05492 0]
120573
R2 0
(c)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000
120574
R2 0
[minus012283 14766e minus 011]
(d)
minus2000 0 2000minus2000
minus1000
0
1000
2000 [minus071273 0]
R2 0
pmax
(e)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [004117 0024135]
R2 0
120579
(f)
minus2000 0 2000minus2000
minus1000
0
1000
2000 [091801 0]
T
R2 0
(g)
Figure 3 PRCC results and PRCC scatter plots of the baseline parameters as those shown in Table 1 Here we fix 119901max = 075 120579 = 02 119879 = 5(a) PRCC results (bndashg) PRCC scatter plots The title of each plot represents the PRCC value with the corresponding P value
endemic periodic solution that is to investigate under whatconditions will the infected population oscillate periodicallywith small amplitude and then we choose the period ofpulse vaccination as a bifurcation parameter to investigate thecomplex patterns of the infectious disease dynamics
Next we analyze the existence of an endemic periodicsolution of model (4) near the disease-free periodic solution(119878lowast(119905) 0) using bifurcation theory [34] and set the impulsive
period 119879 as bifurcation parameter For ease of narrative werewrite the model (4) as
d119878 (119905)d119905
= 119887 minus 120573119878 (119905) 119868 (119905) minus 119887119878 (119905) ≐ 1198651(119878 (119905) 119868 (119905))
d119868 (119905)d119905
= 120573119878 (119905) 119868 (119905) minus 120574119868 (119905) minus 119887119868 (119905) ≐ 1198652(119878 (119905) 119868 (119905))
119905 = 119899119879 119899 isinN
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579≐ Θ1(119878 (119905) 119868 (119905))
119868 (119905+
) = 119868 (119905) ≐ Θ2(119878 (119905) 119868 (119905))
119905 = 119899119879 119899 isinN
(24)According to the bifurcation theorem in [34] we can
obtain the following result
Theorem 3 If there exists a 1198790gt 0 satisfying 1198771
0(1198790) = 1 and
all the parameters in model (24) satisfy
120573 (119878 (1198790) minus 1) eminus1198871198790 (
11988711987901198603
1 minus 1198603eminus1198871198790
minus 1) lt 120573 minus 119903 minus 119887 (25)
where 1198603= 1 minus 119901max + ((120579
2119901max)(120579 + 119878
lowast(1198790))2
) then thereexists a supercritical branch at the point 119879
0
8 Abstract and Applied Analysis
5 10 15 20002
004
006
008
01
T
S(t)
(a)
2 4 6 8 10004
005
006
007
008
T
S(t)
(b)
5 6 7 8 9 10005
0055
006
0065
007
T
S(t)
(c)
7 75 8005
0055
006
0065
007
T
S(t)
(d)
Figure 4 Bifurcation diagrams of model (4) with respect to bifurcation parameter 119879 Here 119901max = 075 and all other parameter values aregiven in Table 1 (a) 120579 = 0 which are reproduced from [9] (b) 120579 = 02 (c and d) a magnified part of Figure 4(b)
The existence of 1198790has been indicated in Figure 1 and the
proof ofTheorem 3 is given in Appendix BTheorem 3 showsthat there exists an endemic periodic solution under someconditions providing that the disease-free periodic solutionbecomes unstable It follows from Theorem 3 that if 119879 gt 119879
0
and is close to 1198790 the endemic periodic solution of model (4)
is stableTo investigate the complex dynamics that model (4) can
have we chose the pulse period 119879 as a bifurcation parameterand fixed all other parameters as those in Figure 4 for twodifferent 120579 values Figure 4(a) is a bifurcation diagram with-out resource limitation that is 120579 = 0 which was obtainedby Shulgin et al [9] Figure 4(b) is a bifurcation diagramwith resource limitation that is 120579 = 02 Comparing thosetwo bifurcation diagrams we conclude that the nonlinearpulse vaccination can produce more complex dynamics thanthose for the model with linear pulse vaccination Once thepulse period 119879 exceeds 119879120579max defined in (22) both susceptibleand infected populations can oscillate periodically with alarge amplitude that corresponds to periodic outbreaks ofepidemics As the pulse period is further increased a series ofcomplex and interesting bifurcation phenomena are observed(see Figures 4(a) and 4(b)) Figures 4(b)ndash4(d) indicate thatthe dynamical behavior of model (4) is very complexincluding period doubling bifurcation chaotic attractors
multistability periodic-adding chaos crisis and periodicwindowsMeanwhile bifurcation analyses (ie Figures 4(b)ndash4(d)) also indicate that the model (4) has several differentattractors which can coexist for a wide range of parametersFor example Figure 5 provides an example of two attractorcoexistence when 119879 = 78 Multiple attractor coexistenceindicates that the infectious outbreak patterns depend onthe initial values which may cause difficulties for infectiousdisease control
5 Discussion and Biological Conclusions
In order to understand the effect of resource limitation inparticular lack of vaccine on the transmission of infectiousdisease we deliberately investigated the dynamical behaviorof an SIR epidemic model which incorporates a nonlinearpulse vaccination strategy To this end we introduced anonlinear form 119901(119905) = (119901max119878(119905))(119878(119905) + 120579) as vaccinationproportionThis resulted in interesting and dramatic changesin the dynamical behavior of solutions and they becamemoreand more complicated which means that it is a very difficulttask to control infectious diseases in particular emerginginfectious diseases under resource limitation We discussedthe control strategy based on the threshold value in moredetail through theoretical analysis numerical studies and
Abstract and Applied Analysis 9
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(a)
100 110 120 130 140 150minus13
minus12
minus11
minus10
minus9
minus8
minus7
t
Log(I(t)
)
(b)
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(c)
100 110 120 130 140 150minus16
minus14
minus12
minus10
minus8
minus6
t
Log(I(t)
)
(d)
Figure 5 Coexistence of two stable attractors of model (4) Here 119901max = 075 120579 = 02 119879 = 78 and all other parameter values are given inTable 1 (a and b) The initial value (119878
0 1198680) = (012 005) (c and d) the initial value (119878
0 1198680) = (01 002)
sensitivity analysis As pointed out byAgur et al [7] the costsrisks and the efficacy of the pulse vaccinations are key factorsin eliminating transmission of the measles virus In orderto depict those factors the nonlinear resource limitationsshould be taken into account [9] and the main purpose ofthe present work is to formulate the disease model to addressthe effects of resource limitations on disease control
The results indicate that vaccination as an approach tocontrolling epidemics must be committed to a long-termstrategy especially when resources arelimited To understandthe interactions amongst 119878(119905) and 119868(119905) which are importantfor successful control of the spread of infectious diseasewe have proposed detailed modeling methods involvingvaccination on 119878(119905) and determining the most significantparameters for the basic reproductive number 1198772
0by using
LHSPRCC uncertainty and sensitivity analysis techniques(see Figure 3) Our results indicate that we should paymore attention to the vaccination period and vaccinationproportion which reduce the threshold value 1198772
0or 11987710to
prevent the outbreak of disease Our results clarify that itis more significant to improve immunization programs (ievaccinations as mentioned in [15 16]) under conditions ofresource limitation
The results also indicate that the dynamic behavior ofmodel (4) may be dramatically affected by small changes in
the value of initial densities of susceptible and infected withresource limitation Bifurcation diagrams shown in Figure 4indicate that there are many forms of complexities in model(4) which are related to chaotic bands with periodic win-dows and attractor crises and Figure 5 reveals that theoccurrence of multiple attractors is a common property ofthe SIR model with resource limitation which can help usto further understand the application of a nonlinear pulsevaccination strategy in an SIR model (for more informationabout bifurcation diagrams of impulsive control strategiessee [35 36]) Some complexities are related to the long-termbehavior of population dynamics characterized either bywell behaving relatively simple or very complicated strangeattractors We also found that the routes to chaos are verycomplicated For instance with resource limitation there areseveral hidden factors that can adversely affect the controlstrategy The increasing number of potential complexitiespredicted by the theory does not seem to make this taskany easier Nevertheless identifying complicated possiblychaotic dynamics in population data has remained a majorchallenge in epidemiology studies In particular the sug-gestion that the dynamics of measles are chaotic has beenmuch debated (eg see [37]) and when seeking to establishwhether measles dynamics in New York City were chaoticor not Sugihara and May [38] omitted data from after 1963when vaccinations were introduced as the immunisations
10 Abstract and Applied Analysis
had altered the intrinsic dynamics However our resultsimply that even with immunisations disease dynamics couldbe chaotic under some circumstances and so a reanalysis ofthe New York data including the vaccination period might beworthwhile
In the present paper we have focused on studyingthe effects of dynamical behaviors on the SIR model withnonlinear pulse vaccination For our simple model we haveassumed that the maximal vaccination proportion 119901max isa constant and the pulse period 119879 can be varied Howeverdue to limited vaccine availability the maximal vaccinationproportion 119901max should depend on the vaccine obtained ineach pulse period Thus to describe the impact of a limitedvaccine stockpile a separate differential equation for theamount of vaccine denoted by 119881(119905) could be included inthe model and then 119901(119905) should be a function of 119881(119905) and119878(119905) Meanwhile it is essential to link the costs of developingand implementing controls to population dynamic modelingof disease epidemics in order to consider other resourcelimitations such as quantities of drugs availability of isolationunits numbers of hospital beds and equipmentThese topicswill be considered in further work in the future
Appendices
A Proof of Theorem 2
Firstly we prove the local stability of a periodic solution(119878lowast(119905) 0) which may be determined by considering the
behavior of small amplitude perturbations of the solutionDefine
119878 (119905) = 119880 (119905) + 119878lowast
(119905) 119868 (119905) = 119881 (119905) (A1)
here119880(119905) 119881(119905) are small perturbations whichmay be writtenas
(119880 (119905)
119881 (119905)) = Φ (119905) (
119880 (0)
119881 (0)) 0 le 119905 lt 119879 (A2)
where the fundamental matrixΦ(119905) satisfies
dΦ (119905)d119905
= (minus119887 minus120573119878
lowast
(119905)
0 120573119878lowast
(119905) minus 120574 minus 119887)Φ (119905) (A3)
and Φ(0) = 119868 is the identity matrix So
Φ (119879) = (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 ) (A4)
where 995333 is not necessarily computed in detail as it is notrequired in the following analysis
The resetting impulsive conditions of model (4) become
(119880 (119899119879
+)
119881 (119899119879+)) = (
1 minus 119901max +1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)(119880 (119899119879)
119881 (119899119879))
≐ 119861 (119899119879) (119880 (119899119879)
119881 (119899119879))
(A5)
Hence according to Floquet theory if the moduli of botheigenvalues of the matrix
119872 = 119861 (119879)Φ (119879)
= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)
times (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 )
(A6)
are less than one then the periodic solution (119878lowast(119905) 0) islocally stable In fact two Floquet expressions multiplied arethus
1205821= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119879))2) eminus119887119879
1205822= eint
119879
0(120573119878lowast(119905)minus120574minus119887)d119905
(A7)
It is obvious that 0 lt 1205821lt 1 Thus the stability of (119878lowast(119905) 0)
is decided by whether 1205822lt 1 or not So we conclude that
the disease-free periodic solution (119878lowast(119905) 0) is locally stable ifint119879
0(120573119878lowast(119905) minus 120574 minus 119887)d119905 lt 0 that is
1
119879int
119879
0
119878lowast
(119905) d119905 =(119878 minus 1) (1 minus eminus119887119879) + 119887119879
119887119879lt120574 + 119887
120573 (A8)
and by simple calculation we have
120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]119887119879 (120574 + 119887)
lt 1 (A9)
which holds true due to 11987710lt 1 Therefore we can draw
a conclusion from the above analysis that if 11987710lt 1 the
disease-free periodic solution (119878lowast(119905) 0) of model (4) is locallyasymptotically stable
Next we will prove the global attraction of the disease-free periodic solution (119878lowast(119905) 0) of model (4) Since 1198771
0lt 1
we can choose 1205761gt 0 sufficiently small such that
1
119879int
119879
0
(119878lowast
(119905) + 1205761) d119905 lt
120574 + 119887
120573 (A10)
It follows from model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) 119905 = 119899119879
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579 119905 = 119899119879
(A11)
Consider the comparison equation
d119885 (119905)d119905
= 119887 minus 119887119885 (119905) 119905 = 119899119879
119885 (119905+
) =(1 minus 119901max) 119885
2
(119905) + 120579119885 (119905)
119885 (119905) + 120579 119905 = 119899119879
(A12)
Abstract and Applied Analysis 11
From Lemma 1 and the comparison theorem on impul-sive differential equations we have 119878(119905) le 119885(119905) and 119885(119905) rarr119878lowast(119905) as 119905 rarr +infin Hence there exists a 119905
1gt 0 such that
119878 (119905) le 119885 (119905) lt 119878lowast
(119905) + 1205761 (A13)
for all 119905 ge 1199051 here 120576
1gt 0 is small enough
From the second equation of model (4) and (A13) we get
d119868 (119905)d119905
le (120573 (119878lowast
(119905) + 1205761) minus 120574 minus 119887) 119868 (119905) (A14)
for 119905 gt 1199051 then we consider the following comparison
equation with pulses
d1199101(119905)
d119905= (120573 (119878
lowast
(119905) + 1205761) minus 120574 minus 119887) 119910
1(119905) 119905 = 119899119879
1199101(119905+
) = 1199101(119905) 119905 = 119899119879
(A15)
Integrating model (A15) between pulses (119899119879 (119899 + 1)119879] wehave
1199101((119899 + 1) 119879) = 119910
1(119899119879) eint
(119899+1)119879
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101(119899119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A16)
Then by using step by step iterations
1199101(119899119879) = 119910
1((119899 minus 1) 119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101((119899 minus 2) 119879) e2 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= sdot sdot sdot
= 1199101(0) e119899 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A17)
where 1199101(0) = 119910
1(0+) gt 0 we obtain lim
119899rarrinfin1199101(119899119879) =
0 according to (A10) On the other hand integrating andsolving the first equation of model (A15) between pulses itgives
1199101(119905) = 119910
1(119899119879) eint
119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905 119905 isin (119899119879 (119899 + 1) 119879]
(A18)
incorporating into the boundedness of eint119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
due to 11987710lt 1 we have lim
119905rarrinfin1199101(119905) = 0
Let (119878(119905) 119868(119905)) be any solution of model (4) with initialvalue (119878
0 1198680) and 119878
0= 119878(0
+) gt 0 119868
0= 119868(0
+) =
1199101(0) gt 0 according to the comparison theorem we have
lim119905rarrinfin
sup 119868(119905) le lim119905rarrinfin
sup1199101(119905) = 0 Incorporating into
the positivity of 119868(119905) we know that lim119905rarrinfin
119868(119905) = 0Consequently there exists a 119905
2gt 1199051such that 0 lt 119868(119905) le 120576
2
for 119905 ge 1199052 and 120576
2is small enough Further we have
119887 minus (1205731205762+ 119887) 119878 (119905) le
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (A19)
for all 119905 gt 1199052 from which we can obtain the following
equation
d1199102(119905)
d119905= 119887 minus (120573120576
2+ 119887) 119910
2(119905) 119905 = 119899119879
1199102(119905+
) =(1 minus 119901max) 119910
2
2(119905) + 120579119910
2(119905)
1199102(119905) + 120579
119905 = 119899119879
(A20)
By employing the same methods as used for the proof ofLemma 1 we get that the model (A20) has a positive periodicsolution 119910lowast
2(119905) which is globally attractive where
119910lowast
2(119905) = (119910
2minus
119887
1205731205762+ 119887) eminus(1205731205762+119887)(119905minus119899119879) + 119887
1205731205762+ 119887
119905 isin (119899119879 (119899 + 1) 119879]
1199102= 1199102(119899119879+
) = 119910lowast
2(0+
) =minus1198612+ radic11986122minus 411986021198622
21198602
1198602= eminus(1205731205762+119887)119879 [ 119887
1205731205762+ 119887
minus (1 minus 119901max) eminus(1205731205762+119887)119879]
1198612= (eminus(1205731205762+119887)119879 minus 119887
1205731205762+ 119887)
times [2 (1 minus 119901max) eminus(1205731205762+119887)119879minus (120579 +
119887
1205731205762+ 119887)]
1198622= minus[(1 minus 119901max) (
119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)2
+ 120579(119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)]
(A21)
It follows from the comparison theorem on impulsive differ-ential equations that 119910
2(119905) le 119878(119905) le 119885(119905) Moreover 119910
2(119905) rarr
119910lowast
2(119905) and 119885(119905) rarr 119878
lowast(119905) as 119905 rarr +infin Consequently there
exists a 1199053for 1205763small enough such that 119905
3ge 1199052and
119910lowast
2(119905) minus 120576
3lt 119878 (119905) lt 119878
lowast
(119905) + 1205763
(A22)
for 119905 ge 1199053 Let 120576
2rarr 0 Then
119878lowast
(119905) minus 1205763lt 119878 (119905) lt 119878
lowast
(119905) + 1205763 (A23)
Therefore 119878(119905) rarr 119878lowast(119905) as 119905 rarr +infin Thus we
have proved the global stability of the disease-free periodicsolution (119878lowast(119905) 0) of model (4) This completes the proof
B Proof of Theorem 3
In order to apply the bifurcation theory of [34] in the maintext we make the following calculations
1198891015840
0= 1 minus eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
(B1)
If 11988910158400= 0 then 119879
0satisfies the following equation
120573 [(119878 minus 1) (1 minus eminus1198871198790) + 1198871198790]
119887 (120574 + 119887) 1198790
= 1 (B2)
12 Abstract and Applied Analysis
which indicates that there exists a1198790such that 1198771
0= 1 and the
disease-free periodic solution 120575 = (119878lowast(119905) 0) loses its stabilityFurther
120597Φ1(1198790 1198830)
120597119878= eminus1198871198790 gt 0
120597Φ2(1198790 1198830)
120597119868= eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
120597Φ1(1198790 1198830)
120597119868= minusint
1198790
0
120573119878lowast
(V) eminus119887(1198790minusV)eintV
0(120573119878lowast(119905)minus120574minus119887)d119905dV
≐ 119860 lt 0
1198861015840
0= 1 minus 119860
3eminus1198871198790
1198871015840
0= 1 minus 119860
3119860
1205972Φ2(1198790 1198830)
120597119868120597119878= 1205731198790eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
1205972Φ2(1198790 1198830)
1205971198682
= minus1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
119878lowast
(120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585minus119887(Vminus120579)
d120579 dV
lt 0
1205972Φ2(1198790 1198830)
120597119868120597119879= (120573119878
lowast
(1198790) minus 120574 minus 119887) eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
120597Φ1(1198790 1198830)
120597119879= minus119887 (119878 minus 1) eminus1198871198790
119861 = 1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887)
times eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
119862 = 1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
eminus119887(119905minus120579)119878lowast (120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585
d120579 dV
+ 21205731198790
1 minus 1198603119860
1 minus 1198603eminus1198871198790
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
(B3)
It follows that 119861119862 lt 0 and we get the condition that is if theparameters satisfy condition 119861 lt 0 then the model (4) has asupercritical branch at 119879
0
In fact
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0 (B4)
Therefore 119861 lt 0 is equivalent to
1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887) lt 0
(B5)
Obviously 119861 lt 0 due to (25) The proof is completed
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thiswork is supported by theNational Natural Science Foun-dation of China (NSFCs 11171199 11371030 11301320) andthe Fundamental Research Funds for the Central Universities(GK201305010) The authors would like to thank the editorand the referees for their careful reading of the original paperand many valuable comments and suggestions that greatlyimproved the presentation of this paper
References
[1] R M Anderson and R M May ldquoPopulation biology of infec-tious diseases part Irdquo Nature vol 280 no 5721 pp 361ndash3671979
[2] E Hanert E Schumacher and E Deleersnijder ldquoFront dynam-ics in fractional-order epidemic modelsrdquo Journal of TheoreticalBiology vol 279 no 1 pp 9ndash16 2011
[3] H W Hethcote ldquoMathematics of infectious diseasesrdquo SIAMReview vol 42 no 4 pp 599ndash653 2000
[4] A E Fiore C B Bridges and N J Cox ldquoSeasonal influenzavaccinesrdquo Current Topics in Microbiology and Immunology vol333 no 1 pp 43ndash82 2009
[5] Y Chang N T Brewer A C Rinas K Schmitt and J S SmithldquoEvaluating the impact of human papillomavirus vaccinesrdquoVaccine vol 27 no 32 pp 4355ndash4362 2009
[6] T J Liesegang ldquoVaricella zoster virus vaccines effective butconcerns lingerrdquo Canadian Journal of Ophthalmology vol 44no 4 pp 379ndash384 2009
[7] Z Agur L Cojocaru G Mazor R M Anderson and Y LDanon ldquoPulse mass measles vaccination across age cohortsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 90 no 24 pp 11698ndash11702 1993
[8] L Stone B Shulgin and Z Agur ldquoTheoretical examinationof the pulse vaccination policy in the SIR epidemic modelrdquoMathematical and ComputerModelling vol 31 no 4-5 pp 207ndash215 2000
[9] B Shulgin L Stone and Z Agur ldquoPulse vaccination strategy inthe SIR epidemic modelrdquo Bulletin of Mathematical Biology vol60 no 6 pp 1123ndash1148 1998
[10] A DrsquoOnofrio ldquoStability properties of pulse vaccination strategyin SEIR epidemicmodelrdquoMathematical Biosciences vol 179 no1 pp 57ndash72 2002
[11] A Franceschetti and A Pugliese ldquoThreshold behaviour of a SIRepidemicmodel with age structure and immigrationrdquo Journal ofMathematical Biology vol 57 no 1 pp 1ndash27 2008
Abstract and Applied Analysis 13
[12] A J Terry ldquoPulse vaccination strategies in a metapopulationSIR modelrdquo Mathematical Biosciences and Engineering vol 7no 2 pp 455ndash477 2010
[13] S J Gao L S Chen J J Nieto and A Torres ldquoAnalysis of adelayed epidemic model with pulse vaccination and saturationincidencerdquo Vaccine vol 24 no 35-36 pp 6037ndash6045 2006
[14] L Matrajt M E Halloran and I M Longini Jr ldquoOptimal vac-cine allocation for the early mitigation of pandemic influenzardquoPLoS Computational Biology vol 9 no 3 Article ID e10029642013
[15] S P Sullivan D G Koutsonanos M D P Martin et alldquoDissolving polymer microneedle patches for influenza vacci-nationrdquo Nature Medicine vol 16 no 8 pp 915ndash920 2010
[16] Science Daily Web ldquoVaccine-delivery patch with dissolvingmicroneedles eliminates ldquosharpsrdquo boosts protectionrdquo httpwwwsciencedailycomreleases201007100718204733htm
[17] M J Reynolds-Hogland J S Hogland and M S MitchellldquoEvaluating intercepts fromdemographicmodels to understandresource limitation and resource thresholdsrdquo Ecological Mod-elling vol 211 no 3-4 pp 424ndash432 2008
[18] L Chow M Fan and Z L Feng ldquoDynamics of a multigroupepidemiological model with group-targeted vaccination strate-giesrdquo Journal of Theoretical Biology vol 291 no 1 pp 56ndash642011
[19] L H Zhou and M Fan ldquoDynamics of an SIR epidemic modelwith limited medical resources revisitedrdquo Nonlinear AnalysisReal World Applications vol 13 no 1 pp 312ndash324 2012
[20] V Lakshmikantham D D Bainov and P S Simeonov Theoryof Impulsive Differential Equations World Scientific Singapore1989
[21] S M Blower and H Dowlatabadi ldquoSensitivity and uncertaintyanalysis of complex-models of disease transmission An HIVmodel as an examplerdquo International Statistical Review vol 62no 2 pp 229ndash243 1994
[22] S Marino I B Hogue C J Ray and D E Kirschner ldquoA meth-odology for performing global uncertainty and sensitivityanalysis in systems biologyrdquo Journal of Theoretical Biology vol254 no 1 pp 178ndash196 2008
[23] S Y Tang J H Liang Y S Tan and R A Cheke ldquoThresholdconditions for integrated pest management models with pesti-cides that have residual effectsrdquo Journal ofMathematical Biologyvol 66 no 1-2 pp 1ndash35 2013
[24] F Brauer and C Castillo-ChavezMathematical Models in Pop-ulation Biology and Epidemiology Springer New York NYUSA 2000
[25] O Diekmann and J A P Heesterbeek Mathematical Epidemi-ology of Infectious Diseases JohnWiley amp Sons Chichester UK2000
[26] J D MurrayMathematical Biology Springer Berlin Germany1989
[27] R Anderson and R May Infectious Diseases of HumansDynamics and Control Oxford University Press Oxford UK1995
[28] M Hethcote ldquoThree basic epidemiological modelsrdquo in AppliedMathematical Ecology L Gross T G Hallam and S A LevinEds pp 119ndash144 Springer Berlin Germany 1989
[29] A V Hill ldquoThe possible effects of the aggregation of themolecules of haemoglobin on its dissociation curvesrdquo TheJournal of Physiology vol 40 no 4 pp 4ndash7 1910
[30] P Cull ldquoLocal and global stability for population modelsrdquoBiological Cybernetics vol 54 no 3 pp 141ndash149 1986
[31] WHOWeb ldquoGlobal health observatory data repositoryrdquo 2001httpappswhointghodatatheme=country
[32] M D McKay R J Beckman and W J Conover ldquoComparisonof three methods for selecting values of input variables in theanalysis of output from a computer coderdquo Technometrics vol21 no 2 pp 239ndash245 1979
[33] W M Schaffer L F Olsen G L Truty S L Fulmer andD J Graser ldquoPeriodic and chaotic dynamics in childhoodepidemicsrdquo in From Chemical to Biological Organization MMarkus S Muller and G Nicolis Eds Springer BerlinGermany 1988
[34] A Lakmeche and O Arino ldquoBifurcation of non trivial periodicsolutions of impulsive differential equations arising chemother-apeutic treatmentrdquo Dynamics of Continuous Discrete andImpulsive Systems B vol 7 no 2 pp 265ndash287 2000
[35] S Y Tang and L S Chen ldquoMultiple attractors in stage-structured population models with birth pulsesrdquo Bulletin ofMathematical Biology vol 65 no 3 pp 479ndash495 2003
[36] S Y Tang Y N Xiao and R A Cheke ldquoMultiple attractorsof host-parasitoid models with integrated pest managementstrategies eradication persistence and outbreakrdquo TheoreticalPopulation Biology vol 73 no 2 pp 181ndash197 2008
[37] L F Olsen andWM Schaffer ldquoChaos versus noisy periodicityalternative hypotheses for childhood epidemicsrdquo Science vol249 no 4968 pp 499ndash504 1990
[38] G Sugihara and R M May ldquoNonlinear forecasting as a way ofdistinguishing chaos from measurement error in time seriesrdquoNature vol 344 no 6268 pp 734ndash741 1990
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical Problems in Engineering
Hindawi Publishing Corporationhttpwwwhindawicom
Differential EquationsInternational Journal of
Volume 2014
Applied MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Probability and StatisticsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical PhysicsAdvances in
Complex AnalysisJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
OptimizationJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CombinatoricsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Operations ResearchAdvances in
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Function Spaces
Abstract and Applied AnalysisHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of Mathematics and Mathematical Sciences
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Algebra
Discrete Dynamics in Nature and Society
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Decision SciencesAdvances in
Discrete MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014 Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Stochastic AnalysisInternational Journal of

Abstract and Applied Analysis 3
Table 1 Parameter definitions values and source for model (4)
Parameter Definition Value Source119887 Rate of natural birth or death rate of the population (yearminus1) 002 [33]120573 Probability of transmission per contact (yearminus1) 1800 [33]120574 Natural recovery rate of the infective individuals (yearminus1) 100 [33]119901max Themaximal vaccination proportion (yearminus1) [0 1) [8 9]120579 The half saturation constant [0 1] Assumption119879 Period of pulse vaccination (year) [0 20] [8 9]
The formula for 119901(119905) plays a key role in investigating theeffect of the limited capacity for treatment on the spread ofinfectious disease To do this we assume without loss of gen-erality that the PVS is implemented in a developing countrybut one in which vaccines are relatively abundant Note thatthere are often major shortages of medical personnel in ruralareas and in certain specialities such as surgery paediatricsand obstetrics [14] Let vaccination pulses occur every 119879 timeunits When a pulse day occurs the medical personnel haveto go out into the rural population and vaccinate as manysusceptibles as they can find on that particular day If thereare very few susceptibles then it will be hard to find them sothe vaccination coverage will be low The more susceptiblesthere are the easier it is to find them so the coverage per pulsewill be greater But there is a limit to how fast the medicalteam can find and handle each susceptible and this causesthe vaccination rate 119901(119905) to saturate that is a saturationphenomenon of limited medical resources By employingthe Holling type II functional response for a predator-preymodel we can define 119901(119905) as follows
119901 (119905) =119901max119878 (119905)
119878 (119905) + 120579 0 le 119901max lt 1 (3)
where 119901max denotes the maximal vaccination proportion and120579 is the half saturation constant Note that the vaccinationcoverage is less than 100 and in practice a vaccine efficacyis also less than 100 which lead to the parameter 119901max lt 1Furthermore generalized formulae such as the Hill function[29] can be employed to characterize the saturation phe-nomenon of the limited medical resources
Taking into account the resource limitation and satura-tion effects for convenience and simplification then model(2) can be rewritten as
d119878 (119905)d119905
= 119887 minus 120573119878 (119905) 119868 (119905) minus 119887119878 (119905)
d119868 (119905)d119905
= 120573119878 (119905) 119868 (119905) minus 120574119868 (119905) minus 119887119868 (119905)
119905 = 119899119879 119899 isinN
119878 (119905+) =
(1 minus 119901max) 1198782
(119905) + 120579119878 (119905)
119878 (119905) + 120579
119868 (119905+) = 119868 (119905)
119905 = 119899119879 119899 isinN
(4)
where the parameters 119887 120573 120574 and 119901max 120579 are defined in mod-els (1) and (3) respectively Note that model (4) is reduced tothe classical SIRmodel with constant pulse vaccination when120579 = 0 which was investigated by Agur et al [7] Stone et al
[8] and Shulgin et al [9]The parameter definitions of model(4) are summarized in Table 1
3 Existence of the Disease-FreePeriodic Solution
We first demonstrate the existence of a disease-free periodicsolution of model (4) in which infectious individuals areentirely absent from the population permanently that is119868(119905) = 0 for a sufficiently long time Under some thresholdconditions we show below that the susceptible populationoscillates with period 119879 in synchronization with the periodicpulse vaccination and the infectious class will die out eventu-allyThus in this section we focus on determining the globalattraction of this disease-free periodic solution To addressthis we first consider the following submodel over the timeinterval 119899119879 lt 119905 le (119899 + 1)119879
d119878 (119905)d119905
= 119887 minus 119887119878 (119905) 119905 = 119899119879
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579 119905 = 119899119879
119878 (0+
) = 1198780
(5)
Integrating the first equation of model (5) between pulsesyields
119878 (119905) = [119878 (119899119879+
) minus 1] eminus119887(119905minus119899119879) + 1 (6)
for 119899119879 lt 119905 le (119899 + 1)119879 It follows from the second equation ofmodel (5) that
119878 ((119899 + 1) 119879+
) =(1 minus 119901max) 119878
2
((119899 + 1) 119879) + 120579119878 ((119899 + 1) 119879)
119878 ((119899 + 1) 119879) + 120579
(7)
Substitution of (6) into (7) gives
119878 ((119899 + 1) 119879+
) = ((1 minus 119901max) [119878 (119899119879+
) minus 1] eminus119887119879 + 12
+ 120579 [119878 (119899119879+
) minus 1] eminus119887119879 + 1)
times ([119878 (119899119879+
) minus 1] eminus119887119879 + 1 + 120579)minus1
(8)
4 Abstract and Applied Analysis
Denote 119878(119899119879+) = 119878119899then the previous equation can be
rewritten as the following difference equation
119878119899+1
=(1 minus 119901max) [(119878119899 minus 1) e
minus119887119879+ 1]2
+ 120579 [(119878119899minus 1) eminus119887119879 + 1]
(119878119899minus 1) eminus119887119879 + 1 + 120579
≐ 119891 (119878119899)
(9)
which is the so-called stroboscopic map of model (5) and itdescribes the relations of the number of susceptibles in thepopulation between any two successive pulse vaccinationsConsequently the existence of the positive steady state ofmodel (9) implies the existence of a positive periodic solutionof model (5) Therefore we first discuss the conditions whichguarantee the existence of a positive steady state of (9) Takingthe derivative of 119891(119878
119899) with respect to 119878
119899yields
1198911015840
(119878119899) = (1 minus 119901max) +
119901max1205792
[(119878119899minus 1) eminus119887119879 + 1 + 120579]2
eminus119887119879
(10)
and it is easy to see that 0 lt 1198911015840(119878119899) lt 1 holds true
For convenience we denote the positive fixed point ofthe stroboscopic map (9) by 119878 which satisfies the followingequation
11986011198782
+ 1198611119878 + 1198621= 0 (11)
where
1198601= eminus119887119879 [1 minus (1 minus 119901max) e
minus119887119879
] gt 0
1198611= (eminus119887119879 minus 1) [2 (1 minus 119901max) e
minus119887119879
minus (120579 + 1)]
1198621= minus [(1 minus 119901max) (1 minus eminus119887119879)
2
+ 120579 (1 minus eminus119887119879)] lt 0
(12)
Obviously (11) has a unique positive root that is
119878 =minus1198611+ radic11986121minus 411986011198621
21198601
(13)
which is stable due to 0 lt 1198911015840(119878) lt 1According to the above discussion on the relations
between a fixed point of the stroboscopic map (9) and theperiodic solution of model (5) we conclude that the sub-model (5) has a unique nontrivial positive periodic solutiondenoted by 119878lowast(119905) and
119878lowast
(119905) = (119878 minus 1) eminus119887(119905minus119899119879) + 1 for 119899119879 lt 119905 le (119899 + 1) 119879 (14)
and it follows from 0 lt 1198911015840(119878119899) lt 1 and [30] that we have the
following lemma
Lemma 1 Model (5) has a positive periodic solution 119878lowast(119905) andfor any solution 119878(119905) of (5) we have |119878(119905) minus 119878lowast(119905)| rarr 0 as119905 rarr +infin
0 2 4 6 8 100
1
2
3
4
5
6
T
T0max
120579 = 04
120579 = 02
120579 = 03
120579 = 01
120579 = 0
R1 0
Figure 1 The effects of resource limitation parameter 120579 and pulsevaccination period 119879 on the threshold value 1198771
0with 119901max = 075
All other parameter values are given in Table 1
By using Lemma 1 we have thus shown that the uniquefixed point 119878 of (9) is globally stable
In the following we will present the sufficient conditionfor the global attraction of the disease-free periodic solution(119878lowast(119905) 0) of model (4)
Theorem 2 The disease-free periodic solution (119878lowast(119905) 0) ofmodel (4) is globally asymptotically stable in the first quadrantprovided that 1198771
0lt 1 where
1198771
0=120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]
119887119879 (120574 + 119887) (15)
here 119878 is defined in (13)
The proof of Theorem 2 is given in Appendix ANote that although the threshold value 1198771
0depends on
all parameters of model (4) the most interesting parametershere are pulse vaccination period 119879 maximum proportion ofpulse vaccination 119901max and the parameter related to resourcelimitation that is 120579 In order to address how those factors (119879and 120579) affect the threshold value 1198771
0 we first note that
lim119879rarr0
1198771
0= lim119879rarr0
120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]119887119879 (120574 + 119887)
= 0 (16)
which indicates the importance of the frequency of vaccina-tion in eradicating an infectious diseaseThen letting 120579 and 119879vary and fixing all other parameters as those in Figure 1 wesee that the threshold value 1198771
0is a monotonically increasing
function of 119879 for fixed 120579 and it is a monotonically increasingfunction of 120579 for fixed period 119879 Moreover there are somecritical values of 119879 and 120579 such that 1198771
0= 1 which indicate
that there exists one maximum allowable period of pulsevaccination 119879120579max such that 119877
1
0lt 1 for all 119879 lt 119879120579max Moreover
the larger 120579 the smaller the value that119879120579max has Unfortunately
Abstract and Applied Analysis 5
we cannot get its analytical expression due to the complexityof 11987710 So we turn to find some approximations for 119879120579max in the
followingIf the PVS is applied frequently enough such that
d119868(119905)d119905 le 0 for all 119905 gt 0 in model (4) then the numberof infectious individuals is a decreasing function of time Itis possible to satisfy this condition if pulsing ensures that119878(119905) le 119878
119888≐ (119887 + 120574)120573 for all 119905 that is pulse vaccination
is applied every time once the 119878(119905) approaches the thresholdvalue 119878
119888(see [7 27] and the references therein)
It follows from the first equation of model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (17)
Consider the following comparison equation
d119909 (119905)d119905
= 119887 minus 119887119909 (119905) 119909 (0) =(1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
(18)
which gives
119909 (119905) = 1 + ((1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
minus 1) eminus119887119905 (19)
Hence
119878 (119905) le 119909 (119905) = 1 + ((1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
minus 1) eminus119887119905 le 119878119888
(20)
and solving the above inequality with respect to 119905 yields
0 le 119905 le1
119887ln(1 +
119901max1198782
119888
(119878119888+ 120579) (1 minus 119878
119888)) (21)
which indicates that
119879120579
max =1
119887ln(1 +
119901max1198782
119888
(119878119888+ 120579) (1 minus 119878
119888)) (22)
Thus if the nonlinear pulse vaccination strategy is appliedperiodically with a period less than 119879120579max then the numberof infectious in the population will decrease forever andeventually die out
We also note that if 120579 = 0 the maximum allowableperiod 119879120579max becomes 1198790max = (1119887) ln(1 + ((119901max119878119888)(1 minus119878119888))) and this is the case studied in [7 9] for a constant
proportion pulse vaccination strategy Obviously 119879120579max is amonotonically decreasing function with respect to 120579 whichgives that 119879120579max lt 119879
0
max This theoretical result indicates that ifwe aim to eradicate an infectious disease then it is necessaryto carry out pulse vaccination programs more frequentlyunder resource limitation than when necessary resourcesare available When various nonlinear factors including119901max 120579 are seriously considered our results show that it isgetting more and more difficult to eliminate the endemicdiseases It also confirms that getting vaccinated is by farthe most effective action that susceptibles can take to protect
themselves and their family from infectious disease Otheroptions available to health practitioners in response to limitedmedical resources include the release of prescriptive healtheducation training vaccinators and promoting the use ofvaccine-delivery patchwith dissolvingmicroneedles (see [14ndash16] for details)
A typical solution of the SIR model with a nonlinearpulse vaccination strategy is shown in Figure 2 where weobserve how the variable 119878(119905) oscillates in a stable cycle(as shown in Figure 2(a)) In contrast the infection 119868(119905)
rapidly decreases to zero (Figure 2(b)) if we set 120579 = 0That is without resource limitation the infectious diseaseis eventually eradicated (Figure 2(b)) However if there isresource limitation that is the parameter 120579 is larger thanzero then both susceptible and infected populations oscillateperiodically as shown in Figures 2(c) and 2(d) with 120579 = 02
Therefore if we have enough vaccine and vaccinators tocombat the infectious disease then it is reasonable that thesusceptible population vaccinated each time is assumed to beproportional to the number of susceptible individuals (here119901max) However this approximation cannot reflect the realcase if we aim to mitigate the emerging infectious diseaseUsually the number of susceptible individuals needing tobe vaccinated may exceed the capacity of local medicalconditions due to shortages of vaccine and doctors especiallyin rural areas in many developing countries where reachingall of the target population may be difficult For instancein 2010 in Lesotho diphtheria tetanus toxoid and pertussis(DTP3) immunization coverage among 1-year-olds was only83 (but an improvement on 78 in 2001 see [31]) and boththe measles and tuberculosis (BCG) vaccination programswere hampered by supplies of vaccine running out in someparts of the country and then allowing vaccinations only forchildren born in hospitals and not for those born elsewherein the communities Thus it is important to design pulsevaccination campaigns carefully For example how frequentlyshould the vaccination strategy be applied And how can thesize of the susceptible population be evaluatedThose are keyissues for evaluating vaccination coverages and the distribu-tion of vaccines Therefore with the help of mathematicalmodels the vaccination strategy can be properly designed
It is well known that in order to eradicate an infectiousdisease the vaccination strategy must be designed such thatthe threshold value 1198771
0is less than 1 which is equivalent to
1198772
0lt 0 where
1198772
0=120573 (119878 minus 1) (1 minus eminus119887119879)
119887+ (120573 minus 120574 minus 119887) 119879 (23)
In the following we employ the formula 11987720to investigate
the important factors which affect the threshold values mostsignificantly by using uncertainty and sensitivity analysis(see [21 22 32]) Sensitivity analysis of the most significantparameters including the birth or death rate (119887) the contactrate (120573) the maximum proportion vaccinated (119901max) and thepulse vaccination period (119879) was performed by evaluatingthe PRCCs for various input parameters against the thresholdcondition 1198772
0with the LHS method [21 22 32] By using
6 Abstract and Applied Analysis
0 10 20 30 400
002
004
006
008
01
S(t)
120579 = 0
(a)
0 10 20 30 40minus800
minus600
minus400
minus200
0
t
Log(I(t))
(b)
0 10 20 30 400
002
004
006
008
01
S(t)
120579 = 02
(c)
0 10 20 30 40minus100
minus80
minus60
minus40
minus20
0
t
Log(I(t))
(d)
Figure 2 The effects of resource limitation on dynamics of model (4) with 119901max = 05 119879 = 2 and all other parameter values are given inTable 1 (a and b) 120579 = 0 (c and d) 120579 = 02
LHS with 3000 samples uncertainty and sensitivity analysesfor all parameters in model (4) were determined A uniformdistribution function was used and tested for significantPRCCs for all parameters with wide ranges such as 119887 sim
119880(0001 01) 120573 sim 119880(1000 2500) 119879 sim 119880(01 20) and 120579 sim119880(0 1) and the baseline values of all parameters are givenin the figure legend of Figure 3 Note that for comparativepurposes we chose the same baseline parameter values asthose used in literatures [8 9 33] Unfortunately we do nothave the realistic ranges of parameter values and we chosethose parameter ranges just for illustrative purposes only andthen provided a scientific basis for disease control
Figure 3(a) shows the PRCC results which illustrate thedependence of 1198772
0on each parameter and PRCC scatter
plots for each parameter are given in Figures 3(b)ndash3(g)respectively We considered absolute values of PRCC valueswhich belong to (04 1) (02 04) and (0 02) as indicatingvery importantmoderate or correlations of little significancebetween input parameters and output variables respectivelyThe positive signs of their PRCCs indicate that if the param-eters are increased the value of 1198772
0increases (vice versa)
The negative sign suggests that if increased the value of 11987720
decreases (vice versa)
Therefore the parameters 119887 120573 120579 and 119879 are responsiblefor increasing the values of 1198772
0 so decreasing all those four
parameters can reduce the threshold value and consequentlyare beneficial for infectious disease control As the parametermaximum proportion of pulse vaccination 119901max changes itis negatively and significantly correlated with large PRCCsThus increasing this parameter will result in significantlyreducing the threshold value The most significant controlparameters are 119887 120573 119901max and 119879 (as shown in Figures 3(b)3(c) 3(e) and 3(g)) It is interesting to note that the period119879and the maximum proportion vaccinated 119901max are stronglycorrelated with the threshold value 1198772
0 which indicates that
carefully designing a pulse vaccination strategy is crucial forinfectious disease control
4 Existence of Endemic Periodic Solution andComplex Dynamics
An important issue is how the infectious disease develops ifthe threshold condition 1198771
0is larger than one In particular
what types of dynamic behavior maymodel (4) have once theperiod of pulse vaccination exceeds certain threshold levelsTherefore in this section we first focus on the existence of an
Abstract and Applied Analysis 7
minus1
minus05
0
05
1
b 120573 120574 120579 T
PRCC
s
pmax
(a)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [072346 0]
b
R2 0
(b)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [05492 0]
120573
R2 0
(c)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000
120574
R2 0
[minus012283 14766e minus 011]
(d)
minus2000 0 2000minus2000
minus1000
0
1000
2000 [minus071273 0]
R2 0
pmax
(e)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [004117 0024135]
R2 0
120579
(f)
minus2000 0 2000minus2000
minus1000
0
1000
2000 [091801 0]
T
R2 0
(g)
Figure 3 PRCC results and PRCC scatter plots of the baseline parameters as those shown in Table 1 Here we fix 119901max = 075 120579 = 02 119879 = 5(a) PRCC results (bndashg) PRCC scatter plots The title of each plot represents the PRCC value with the corresponding P value
endemic periodic solution that is to investigate under whatconditions will the infected population oscillate periodicallywith small amplitude and then we choose the period ofpulse vaccination as a bifurcation parameter to investigate thecomplex patterns of the infectious disease dynamics
Next we analyze the existence of an endemic periodicsolution of model (4) near the disease-free periodic solution(119878lowast(119905) 0) using bifurcation theory [34] and set the impulsive
period 119879 as bifurcation parameter For ease of narrative werewrite the model (4) as
d119878 (119905)d119905
= 119887 minus 120573119878 (119905) 119868 (119905) minus 119887119878 (119905) ≐ 1198651(119878 (119905) 119868 (119905))
d119868 (119905)d119905
= 120573119878 (119905) 119868 (119905) minus 120574119868 (119905) minus 119887119868 (119905) ≐ 1198652(119878 (119905) 119868 (119905))
119905 = 119899119879 119899 isinN
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579≐ Θ1(119878 (119905) 119868 (119905))
119868 (119905+
) = 119868 (119905) ≐ Θ2(119878 (119905) 119868 (119905))
119905 = 119899119879 119899 isinN
(24)According to the bifurcation theorem in [34] we can
obtain the following result
Theorem 3 If there exists a 1198790gt 0 satisfying 1198771
0(1198790) = 1 and
all the parameters in model (24) satisfy
120573 (119878 (1198790) minus 1) eminus1198871198790 (
11988711987901198603
1 minus 1198603eminus1198871198790
minus 1) lt 120573 minus 119903 minus 119887 (25)
where 1198603= 1 minus 119901max + ((120579
2119901max)(120579 + 119878
lowast(1198790))2
) then thereexists a supercritical branch at the point 119879
0
8 Abstract and Applied Analysis
5 10 15 20002
004
006
008
01
T
S(t)
(a)
2 4 6 8 10004
005
006
007
008
T
S(t)
(b)
5 6 7 8 9 10005
0055
006
0065
007
T
S(t)
(c)
7 75 8005
0055
006
0065
007
T
S(t)
(d)
Figure 4 Bifurcation diagrams of model (4) with respect to bifurcation parameter 119879 Here 119901max = 075 and all other parameter values aregiven in Table 1 (a) 120579 = 0 which are reproduced from [9] (b) 120579 = 02 (c and d) a magnified part of Figure 4(b)
The existence of 1198790has been indicated in Figure 1 and the
proof ofTheorem 3 is given in Appendix BTheorem 3 showsthat there exists an endemic periodic solution under someconditions providing that the disease-free periodic solutionbecomes unstable It follows from Theorem 3 that if 119879 gt 119879
0
and is close to 1198790 the endemic periodic solution of model (4)
is stableTo investigate the complex dynamics that model (4) can
have we chose the pulse period 119879 as a bifurcation parameterand fixed all other parameters as those in Figure 4 for twodifferent 120579 values Figure 4(a) is a bifurcation diagram with-out resource limitation that is 120579 = 0 which was obtainedby Shulgin et al [9] Figure 4(b) is a bifurcation diagramwith resource limitation that is 120579 = 02 Comparing thosetwo bifurcation diagrams we conclude that the nonlinearpulse vaccination can produce more complex dynamics thanthose for the model with linear pulse vaccination Once thepulse period 119879 exceeds 119879120579max defined in (22) both susceptibleand infected populations can oscillate periodically with alarge amplitude that corresponds to periodic outbreaks ofepidemics As the pulse period is further increased a series ofcomplex and interesting bifurcation phenomena are observed(see Figures 4(a) and 4(b)) Figures 4(b)ndash4(d) indicate thatthe dynamical behavior of model (4) is very complexincluding period doubling bifurcation chaotic attractors
multistability periodic-adding chaos crisis and periodicwindowsMeanwhile bifurcation analyses (ie Figures 4(b)ndash4(d)) also indicate that the model (4) has several differentattractors which can coexist for a wide range of parametersFor example Figure 5 provides an example of two attractorcoexistence when 119879 = 78 Multiple attractor coexistenceindicates that the infectious outbreak patterns depend onthe initial values which may cause difficulties for infectiousdisease control
5 Discussion and Biological Conclusions
In order to understand the effect of resource limitation inparticular lack of vaccine on the transmission of infectiousdisease we deliberately investigated the dynamical behaviorof an SIR epidemic model which incorporates a nonlinearpulse vaccination strategy To this end we introduced anonlinear form 119901(119905) = (119901max119878(119905))(119878(119905) + 120579) as vaccinationproportionThis resulted in interesting and dramatic changesin the dynamical behavior of solutions and they becamemoreand more complicated which means that it is a very difficulttask to control infectious diseases in particular emerginginfectious diseases under resource limitation We discussedthe control strategy based on the threshold value in moredetail through theoretical analysis numerical studies and
Abstract and Applied Analysis 9
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(a)
100 110 120 130 140 150minus13
minus12
minus11
minus10
minus9
minus8
minus7
t
Log(I(t)
)
(b)
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(c)
100 110 120 130 140 150minus16
minus14
minus12
minus10
minus8
minus6
t
Log(I(t)
)
(d)
Figure 5 Coexistence of two stable attractors of model (4) Here 119901max = 075 120579 = 02 119879 = 78 and all other parameter values are given inTable 1 (a and b) The initial value (119878
0 1198680) = (012 005) (c and d) the initial value (119878
0 1198680) = (01 002)
sensitivity analysis As pointed out byAgur et al [7] the costsrisks and the efficacy of the pulse vaccinations are key factorsin eliminating transmission of the measles virus In orderto depict those factors the nonlinear resource limitationsshould be taken into account [9] and the main purpose ofthe present work is to formulate the disease model to addressthe effects of resource limitations on disease control
The results indicate that vaccination as an approach tocontrolling epidemics must be committed to a long-termstrategy especially when resources arelimited To understandthe interactions amongst 119878(119905) and 119868(119905) which are importantfor successful control of the spread of infectious diseasewe have proposed detailed modeling methods involvingvaccination on 119878(119905) and determining the most significantparameters for the basic reproductive number 1198772
0by using
LHSPRCC uncertainty and sensitivity analysis techniques(see Figure 3) Our results indicate that we should paymore attention to the vaccination period and vaccinationproportion which reduce the threshold value 1198772
0or 11987710to
prevent the outbreak of disease Our results clarify that itis more significant to improve immunization programs (ievaccinations as mentioned in [15 16]) under conditions ofresource limitation
The results also indicate that the dynamic behavior ofmodel (4) may be dramatically affected by small changes in
the value of initial densities of susceptible and infected withresource limitation Bifurcation diagrams shown in Figure 4indicate that there are many forms of complexities in model(4) which are related to chaotic bands with periodic win-dows and attractor crises and Figure 5 reveals that theoccurrence of multiple attractors is a common property ofthe SIR model with resource limitation which can help usto further understand the application of a nonlinear pulsevaccination strategy in an SIR model (for more informationabout bifurcation diagrams of impulsive control strategiessee [35 36]) Some complexities are related to the long-termbehavior of population dynamics characterized either bywell behaving relatively simple or very complicated strangeattractors We also found that the routes to chaos are verycomplicated For instance with resource limitation there areseveral hidden factors that can adversely affect the controlstrategy The increasing number of potential complexitiespredicted by the theory does not seem to make this taskany easier Nevertheless identifying complicated possiblychaotic dynamics in population data has remained a majorchallenge in epidemiology studies In particular the sug-gestion that the dynamics of measles are chaotic has beenmuch debated (eg see [37]) and when seeking to establishwhether measles dynamics in New York City were chaoticor not Sugihara and May [38] omitted data from after 1963when vaccinations were introduced as the immunisations
10 Abstract and Applied Analysis
had altered the intrinsic dynamics However our resultsimply that even with immunisations disease dynamics couldbe chaotic under some circumstances and so a reanalysis ofthe New York data including the vaccination period might beworthwhile
In the present paper we have focused on studyingthe effects of dynamical behaviors on the SIR model withnonlinear pulse vaccination For our simple model we haveassumed that the maximal vaccination proportion 119901max isa constant and the pulse period 119879 can be varied Howeverdue to limited vaccine availability the maximal vaccinationproportion 119901max should depend on the vaccine obtained ineach pulse period Thus to describe the impact of a limitedvaccine stockpile a separate differential equation for theamount of vaccine denoted by 119881(119905) could be included inthe model and then 119901(119905) should be a function of 119881(119905) and119878(119905) Meanwhile it is essential to link the costs of developingand implementing controls to population dynamic modelingof disease epidemics in order to consider other resourcelimitations such as quantities of drugs availability of isolationunits numbers of hospital beds and equipmentThese topicswill be considered in further work in the future
Appendices
A Proof of Theorem 2
Firstly we prove the local stability of a periodic solution(119878lowast(119905) 0) which may be determined by considering the
behavior of small amplitude perturbations of the solutionDefine
119878 (119905) = 119880 (119905) + 119878lowast
(119905) 119868 (119905) = 119881 (119905) (A1)
here119880(119905) 119881(119905) are small perturbations whichmay be writtenas
(119880 (119905)
119881 (119905)) = Φ (119905) (
119880 (0)
119881 (0)) 0 le 119905 lt 119879 (A2)
where the fundamental matrixΦ(119905) satisfies
dΦ (119905)d119905
= (minus119887 minus120573119878
lowast
(119905)
0 120573119878lowast
(119905) minus 120574 minus 119887)Φ (119905) (A3)
and Φ(0) = 119868 is the identity matrix So
Φ (119879) = (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 ) (A4)
where 995333 is not necessarily computed in detail as it is notrequired in the following analysis
The resetting impulsive conditions of model (4) become
(119880 (119899119879
+)
119881 (119899119879+)) = (
1 minus 119901max +1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)(119880 (119899119879)
119881 (119899119879))
≐ 119861 (119899119879) (119880 (119899119879)
119881 (119899119879))
(A5)
Hence according to Floquet theory if the moduli of botheigenvalues of the matrix
119872 = 119861 (119879)Φ (119879)
= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)
times (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 )
(A6)
are less than one then the periodic solution (119878lowast(119905) 0) islocally stable In fact two Floquet expressions multiplied arethus
1205821= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119879))2) eminus119887119879
1205822= eint
119879
0(120573119878lowast(119905)minus120574minus119887)d119905
(A7)
It is obvious that 0 lt 1205821lt 1 Thus the stability of (119878lowast(119905) 0)
is decided by whether 1205822lt 1 or not So we conclude that
the disease-free periodic solution (119878lowast(119905) 0) is locally stable ifint119879
0(120573119878lowast(119905) minus 120574 minus 119887)d119905 lt 0 that is
1
119879int
119879
0
119878lowast
(119905) d119905 =(119878 minus 1) (1 minus eminus119887119879) + 119887119879
119887119879lt120574 + 119887
120573 (A8)
and by simple calculation we have
120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]119887119879 (120574 + 119887)
lt 1 (A9)
which holds true due to 11987710lt 1 Therefore we can draw
a conclusion from the above analysis that if 11987710lt 1 the
disease-free periodic solution (119878lowast(119905) 0) of model (4) is locallyasymptotically stable
Next we will prove the global attraction of the disease-free periodic solution (119878lowast(119905) 0) of model (4) Since 1198771
0lt 1
we can choose 1205761gt 0 sufficiently small such that
1
119879int
119879
0
(119878lowast
(119905) + 1205761) d119905 lt
120574 + 119887
120573 (A10)
It follows from model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) 119905 = 119899119879
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579 119905 = 119899119879
(A11)
Consider the comparison equation
d119885 (119905)d119905
= 119887 minus 119887119885 (119905) 119905 = 119899119879
119885 (119905+
) =(1 minus 119901max) 119885
2
(119905) + 120579119885 (119905)
119885 (119905) + 120579 119905 = 119899119879
(A12)
Abstract and Applied Analysis 11
From Lemma 1 and the comparison theorem on impul-sive differential equations we have 119878(119905) le 119885(119905) and 119885(119905) rarr119878lowast(119905) as 119905 rarr +infin Hence there exists a 119905
1gt 0 such that
119878 (119905) le 119885 (119905) lt 119878lowast
(119905) + 1205761 (A13)
for all 119905 ge 1199051 here 120576
1gt 0 is small enough
From the second equation of model (4) and (A13) we get
d119868 (119905)d119905
le (120573 (119878lowast
(119905) + 1205761) minus 120574 minus 119887) 119868 (119905) (A14)
for 119905 gt 1199051 then we consider the following comparison
equation with pulses
d1199101(119905)
d119905= (120573 (119878
lowast
(119905) + 1205761) minus 120574 minus 119887) 119910
1(119905) 119905 = 119899119879
1199101(119905+
) = 1199101(119905) 119905 = 119899119879
(A15)
Integrating model (A15) between pulses (119899119879 (119899 + 1)119879] wehave
1199101((119899 + 1) 119879) = 119910
1(119899119879) eint
(119899+1)119879
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101(119899119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A16)
Then by using step by step iterations
1199101(119899119879) = 119910
1((119899 minus 1) 119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101((119899 minus 2) 119879) e2 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= sdot sdot sdot
= 1199101(0) e119899 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A17)
where 1199101(0) = 119910
1(0+) gt 0 we obtain lim
119899rarrinfin1199101(119899119879) =
0 according to (A10) On the other hand integrating andsolving the first equation of model (A15) between pulses itgives
1199101(119905) = 119910
1(119899119879) eint
119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905 119905 isin (119899119879 (119899 + 1) 119879]
(A18)
incorporating into the boundedness of eint119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
due to 11987710lt 1 we have lim
119905rarrinfin1199101(119905) = 0
Let (119878(119905) 119868(119905)) be any solution of model (4) with initialvalue (119878
0 1198680) and 119878
0= 119878(0
+) gt 0 119868
0= 119868(0
+) =
1199101(0) gt 0 according to the comparison theorem we have
lim119905rarrinfin
sup 119868(119905) le lim119905rarrinfin
sup1199101(119905) = 0 Incorporating into
the positivity of 119868(119905) we know that lim119905rarrinfin
119868(119905) = 0Consequently there exists a 119905
2gt 1199051such that 0 lt 119868(119905) le 120576
2
for 119905 ge 1199052 and 120576
2is small enough Further we have
119887 minus (1205731205762+ 119887) 119878 (119905) le
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (A19)
for all 119905 gt 1199052 from which we can obtain the following
equation
d1199102(119905)
d119905= 119887 minus (120573120576
2+ 119887) 119910
2(119905) 119905 = 119899119879
1199102(119905+
) =(1 minus 119901max) 119910
2
2(119905) + 120579119910
2(119905)
1199102(119905) + 120579
119905 = 119899119879
(A20)
By employing the same methods as used for the proof ofLemma 1 we get that the model (A20) has a positive periodicsolution 119910lowast
2(119905) which is globally attractive where
119910lowast
2(119905) = (119910
2minus
119887
1205731205762+ 119887) eminus(1205731205762+119887)(119905minus119899119879) + 119887
1205731205762+ 119887
119905 isin (119899119879 (119899 + 1) 119879]
1199102= 1199102(119899119879+
) = 119910lowast
2(0+
) =minus1198612+ radic11986122minus 411986021198622
21198602
1198602= eminus(1205731205762+119887)119879 [ 119887
1205731205762+ 119887
minus (1 minus 119901max) eminus(1205731205762+119887)119879]
1198612= (eminus(1205731205762+119887)119879 minus 119887
1205731205762+ 119887)
times [2 (1 minus 119901max) eminus(1205731205762+119887)119879minus (120579 +
119887
1205731205762+ 119887)]
1198622= minus[(1 minus 119901max) (
119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)2
+ 120579(119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)]
(A21)
It follows from the comparison theorem on impulsive differ-ential equations that 119910
2(119905) le 119878(119905) le 119885(119905) Moreover 119910
2(119905) rarr
119910lowast
2(119905) and 119885(119905) rarr 119878
lowast(119905) as 119905 rarr +infin Consequently there
exists a 1199053for 1205763small enough such that 119905
3ge 1199052and
119910lowast
2(119905) minus 120576
3lt 119878 (119905) lt 119878
lowast
(119905) + 1205763
(A22)
for 119905 ge 1199053 Let 120576
2rarr 0 Then
119878lowast
(119905) minus 1205763lt 119878 (119905) lt 119878
lowast
(119905) + 1205763 (A23)
Therefore 119878(119905) rarr 119878lowast(119905) as 119905 rarr +infin Thus we
have proved the global stability of the disease-free periodicsolution (119878lowast(119905) 0) of model (4) This completes the proof
B Proof of Theorem 3
In order to apply the bifurcation theory of [34] in the maintext we make the following calculations
1198891015840
0= 1 minus eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
(B1)
If 11988910158400= 0 then 119879
0satisfies the following equation
120573 [(119878 minus 1) (1 minus eminus1198871198790) + 1198871198790]
119887 (120574 + 119887) 1198790
= 1 (B2)
12 Abstract and Applied Analysis
which indicates that there exists a1198790such that 1198771
0= 1 and the
disease-free periodic solution 120575 = (119878lowast(119905) 0) loses its stabilityFurther
120597Φ1(1198790 1198830)
120597119878= eminus1198871198790 gt 0
120597Φ2(1198790 1198830)
120597119868= eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
120597Φ1(1198790 1198830)
120597119868= minusint
1198790
0
120573119878lowast
(V) eminus119887(1198790minusV)eintV
0(120573119878lowast(119905)minus120574minus119887)d119905dV
≐ 119860 lt 0
1198861015840
0= 1 minus 119860
3eminus1198871198790
1198871015840
0= 1 minus 119860
3119860
1205972Φ2(1198790 1198830)
120597119868120597119878= 1205731198790eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
1205972Φ2(1198790 1198830)
1205971198682
= minus1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
119878lowast
(120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585minus119887(Vminus120579)
d120579 dV
lt 0
1205972Φ2(1198790 1198830)
120597119868120597119879= (120573119878
lowast
(1198790) minus 120574 minus 119887) eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
120597Φ1(1198790 1198830)
120597119879= minus119887 (119878 minus 1) eminus1198871198790
119861 = 1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887)
times eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
119862 = 1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
eminus119887(119905minus120579)119878lowast (120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585
d120579 dV
+ 21205731198790
1 minus 1198603119860
1 minus 1198603eminus1198871198790
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
(B3)
It follows that 119861119862 lt 0 and we get the condition that is if theparameters satisfy condition 119861 lt 0 then the model (4) has asupercritical branch at 119879
0
In fact
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0 (B4)
Therefore 119861 lt 0 is equivalent to
1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887) lt 0
(B5)
Obviously 119861 lt 0 due to (25) The proof is completed
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thiswork is supported by theNational Natural Science Foun-dation of China (NSFCs 11171199 11371030 11301320) andthe Fundamental Research Funds for the Central Universities(GK201305010) The authors would like to thank the editorand the referees for their careful reading of the original paperand many valuable comments and suggestions that greatlyimproved the presentation of this paper
References
[1] R M Anderson and R M May ldquoPopulation biology of infec-tious diseases part Irdquo Nature vol 280 no 5721 pp 361ndash3671979
[2] E Hanert E Schumacher and E Deleersnijder ldquoFront dynam-ics in fractional-order epidemic modelsrdquo Journal of TheoreticalBiology vol 279 no 1 pp 9ndash16 2011
[3] H W Hethcote ldquoMathematics of infectious diseasesrdquo SIAMReview vol 42 no 4 pp 599ndash653 2000
[4] A E Fiore C B Bridges and N J Cox ldquoSeasonal influenzavaccinesrdquo Current Topics in Microbiology and Immunology vol333 no 1 pp 43ndash82 2009
[5] Y Chang N T Brewer A C Rinas K Schmitt and J S SmithldquoEvaluating the impact of human papillomavirus vaccinesrdquoVaccine vol 27 no 32 pp 4355ndash4362 2009
[6] T J Liesegang ldquoVaricella zoster virus vaccines effective butconcerns lingerrdquo Canadian Journal of Ophthalmology vol 44no 4 pp 379ndash384 2009
[7] Z Agur L Cojocaru G Mazor R M Anderson and Y LDanon ldquoPulse mass measles vaccination across age cohortsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 90 no 24 pp 11698ndash11702 1993
[8] L Stone B Shulgin and Z Agur ldquoTheoretical examinationof the pulse vaccination policy in the SIR epidemic modelrdquoMathematical and ComputerModelling vol 31 no 4-5 pp 207ndash215 2000
[9] B Shulgin L Stone and Z Agur ldquoPulse vaccination strategy inthe SIR epidemic modelrdquo Bulletin of Mathematical Biology vol60 no 6 pp 1123ndash1148 1998
[10] A DrsquoOnofrio ldquoStability properties of pulse vaccination strategyin SEIR epidemicmodelrdquoMathematical Biosciences vol 179 no1 pp 57ndash72 2002
[11] A Franceschetti and A Pugliese ldquoThreshold behaviour of a SIRepidemicmodel with age structure and immigrationrdquo Journal ofMathematical Biology vol 57 no 1 pp 1ndash27 2008
Abstract and Applied Analysis 13
[12] A J Terry ldquoPulse vaccination strategies in a metapopulationSIR modelrdquo Mathematical Biosciences and Engineering vol 7no 2 pp 455ndash477 2010
[13] S J Gao L S Chen J J Nieto and A Torres ldquoAnalysis of adelayed epidemic model with pulse vaccination and saturationincidencerdquo Vaccine vol 24 no 35-36 pp 6037ndash6045 2006
[14] L Matrajt M E Halloran and I M Longini Jr ldquoOptimal vac-cine allocation for the early mitigation of pandemic influenzardquoPLoS Computational Biology vol 9 no 3 Article ID e10029642013
[15] S P Sullivan D G Koutsonanos M D P Martin et alldquoDissolving polymer microneedle patches for influenza vacci-nationrdquo Nature Medicine vol 16 no 8 pp 915ndash920 2010
[16] Science Daily Web ldquoVaccine-delivery patch with dissolvingmicroneedles eliminates ldquosharpsrdquo boosts protectionrdquo httpwwwsciencedailycomreleases201007100718204733htm
[17] M J Reynolds-Hogland J S Hogland and M S MitchellldquoEvaluating intercepts fromdemographicmodels to understandresource limitation and resource thresholdsrdquo Ecological Mod-elling vol 211 no 3-4 pp 424ndash432 2008
[18] L Chow M Fan and Z L Feng ldquoDynamics of a multigroupepidemiological model with group-targeted vaccination strate-giesrdquo Journal of Theoretical Biology vol 291 no 1 pp 56ndash642011
[19] L H Zhou and M Fan ldquoDynamics of an SIR epidemic modelwith limited medical resources revisitedrdquo Nonlinear AnalysisReal World Applications vol 13 no 1 pp 312ndash324 2012
[20] V Lakshmikantham D D Bainov and P S Simeonov Theoryof Impulsive Differential Equations World Scientific Singapore1989
[21] S M Blower and H Dowlatabadi ldquoSensitivity and uncertaintyanalysis of complex-models of disease transmission An HIVmodel as an examplerdquo International Statistical Review vol 62no 2 pp 229ndash243 1994
[22] S Marino I B Hogue C J Ray and D E Kirschner ldquoA meth-odology for performing global uncertainty and sensitivityanalysis in systems biologyrdquo Journal of Theoretical Biology vol254 no 1 pp 178ndash196 2008
[23] S Y Tang J H Liang Y S Tan and R A Cheke ldquoThresholdconditions for integrated pest management models with pesti-cides that have residual effectsrdquo Journal ofMathematical Biologyvol 66 no 1-2 pp 1ndash35 2013
[24] F Brauer and C Castillo-ChavezMathematical Models in Pop-ulation Biology and Epidemiology Springer New York NYUSA 2000
[25] O Diekmann and J A P Heesterbeek Mathematical Epidemi-ology of Infectious Diseases JohnWiley amp Sons Chichester UK2000
[26] J D MurrayMathematical Biology Springer Berlin Germany1989
[27] R Anderson and R May Infectious Diseases of HumansDynamics and Control Oxford University Press Oxford UK1995
[28] M Hethcote ldquoThree basic epidemiological modelsrdquo in AppliedMathematical Ecology L Gross T G Hallam and S A LevinEds pp 119ndash144 Springer Berlin Germany 1989
[29] A V Hill ldquoThe possible effects of the aggregation of themolecules of haemoglobin on its dissociation curvesrdquo TheJournal of Physiology vol 40 no 4 pp 4ndash7 1910
[30] P Cull ldquoLocal and global stability for population modelsrdquoBiological Cybernetics vol 54 no 3 pp 141ndash149 1986
[31] WHOWeb ldquoGlobal health observatory data repositoryrdquo 2001httpappswhointghodatatheme=country
[32] M D McKay R J Beckman and W J Conover ldquoComparisonof three methods for selecting values of input variables in theanalysis of output from a computer coderdquo Technometrics vol21 no 2 pp 239ndash245 1979
[33] W M Schaffer L F Olsen G L Truty S L Fulmer andD J Graser ldquoPeriodic and chaotic dynamics in childhoodepidemicsrdquo in From Chemical to Biological Organization MMarkus S Muller and G Nicolis Eds Springer BerlinGermany 1988
[34] A Lakmeche and O Arino ldquoBifurcation of non trivial periodicsolutions of impulsive differential equations arising chemother-apeutic treatmentrdquo Dynamics of Continuous Discrete andImpulsive Systems B vol 7 no 2 pp 265ndash287 2000
[35] S Y Tang and L S Chen ldquoMultiple attractors in stage-structured population models with birth pulsesrdquo Bulletin ofMathematical Biology vol 65 no 3 pp 479ndash495 2003
[36] S Y Tang Y N Xiao and R A Cheke ldquoMultiple attractorsof host-parasitoid models with integrated pest managementstrategies eradication persistence and outbreakrdquo TheoreticalPopulation Biology vol 73 no 2 pp 181ndash197 2008
[37] L F Olsen andWM Schaffer ldquoChaos versus noisy periodicityalternative hypotheses for childhood epidemicsrdquo Science vol249 no 4968 pp 499ndash504 1990
[38] G Sugihara and R M May ldquoNonlinear forecasting as a way ofdistinguishing chaos from measurement error in time seriesrdquoNature vol 344 no 6268 pp 734ndash741 1990
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical Problems in Engineering
Hindawi Publishing Corporationhttpwwwhindawicom
Differential EquationsInternational Journal of
Volume 2014
Applied MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Probability and StatisticsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical PhysicsAdvances in
Complex AnalysisJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
OptimizationJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CombinatoricsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Operations ResearchAdvances in
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Function Spaces
Abstract and Applied AnalysisHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of Mathematics and Mathematical Sciences
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Algebra
Discrete Dynamics in Nature and Society
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Decision SciencesAdvances in
Discrete MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014 Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Stochastic AnalysisInternational Journal of

4 Abstract and Applied Analysis
Denote 119878(119899119879+) = 119878119899then the previous equation can be
rewritten as the following difference equation
119878119899+1
=(1 minus 119901max) [(119878119899 minus 1) e
minus119887119879+ 1]2
+ 120579 [(119878119899minus 1) eminus119887119879 + 1]
(119878119899minus 1) eminus119887119879 + 1 + 120579
≐ 119891 (119878119899)
(9)
which is the so-called stroboscopic map of model (5) and itdescribes the relations of the number of susceptibles in thepopulation between any two successive pulse vaccinationsConsequently the existence of the positive steady state ofmodel (9) implies the existence of a positive periodic solutionof model (5) Therefore we first discuss the conditions whichguarantee the existence of a positive steady state of (9) Takingthe derivative of 119891(119878
119899) with respect to 119878
119899yields
1198911015840
(119878119899) = (1 minus 119901max) +
119901max1205792
[(119878119899minus 1) eminus119887119879 + 1 + 120579]2
eminus119887119879
(10)
and it is easy to see that 0 lt 1198911015840(119878119899) lt 1 holds true
For convenience we denote the positive fixed point ofthe stroboscopic map (9) by 119878 which satisfies the followingequation
11986011198782
+ 1198611119878 + 1198621= 0 (11)
where
1198601= eminus119887119879 [1 minus (1 minus 119901max) e
minus119887119879
] gt 0
1198611= (eminus119887119879 minus 1) [2 (1 minus 119901max) e
minus119887119879
minus (120579 + 1)]
1198621= minus [(1 minus 119901max) (1 minus eminus119887119879)
2
+ 120579 (1 minus eminus119887119879)] lt 0
(12)
Obviously (11) has a unique positive root that is
119878 =minus1198611+ radic11986121minus 411986011198621
21198601
(13)
which is stable due to 0 lt 1198911015840(119878) lt 1According to the above discussion on the relations
between a fixed point of the stroboscopic map (9) and theperiodic solution of model (5) we conclude that the sub-model (5) has a unique nontrivial positive periodic solutiondenoted by 119878lowast(119905) and
119878lowast
(119905) = (119878 minus 1) eminus119887(119905minus119899119879) + 1 for 119899119879 lt 119905 le (119899 + 1) 119879 (14)
and it follows from 0 lt 1198911015840(119878119899) lt 1 and [30] that we have the
following lemma
Lemma 1 Model (5) has a positive periodic solution 119878lowast(119905) andfor any solution 119878(119905) of (5) we have |119878(119905) minus 119878lowast(119905)| rarr 0 as119905 rarr +infin
0 2 4 6 8 100
1
2
3
4
5
6
T
T0max
120579 = 04
120579 = 02
120579 = 03
120579 = 01
120579 = 0
R1 0
Figure 1 The effects of resource limitation parameter 120579 and pulsevaccination period 119879 on the threshold value 1198771
0with 119901max = 075
All other parameter values are given in Table 1
By using Lemma 1 we have thus shown that the uniquefixed point 119878 of (9) is globally stable
In the following we will present the sufficient conditionfor the global attraction of the disease-free periodic solution(119878lowast(119905) 0) of model (4)
Theorem 2 The disease-free periodic solution (119878lowast(119905) 0) ofmodel (4) is globally asymptotically stable in the first quadrantprovided that 1198771
0lt 1 where
1198771
0=120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]
119887119879 (120574 + 119887) (15)
here 119878 is defined in (13)
The proof of Theorem 2 is given in Appendix ANote that although the threshold value 1198771
0depends on
all parameters of model (4) the most interesting parametershere are pulse vaccination period 119879 maximum proportion ofpulse vaccination 119901max and the parameter related to resourcelimitation that is 120579 In order to address how those factors (119879and 120579) affect the threshold value 1198771
0 we first note that
lim119879rarr0
1198771
0= lim119879rarr0
120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]119887119879 (120574 + 119887)
= 0 (16)
which indicates the importance of the frequency of vaccina-tion in eradicating an infectious diseaseThen letting 120579 and 119879vary and fixing all other parameters as those in Figure 1 wesee that the threshold value 1198771
0is a monotonically increasing
function of 119879 for fixed 120579 and it is a monotonically increasingfunction of 120579 for fixed period 119879 Moreover there are somecritical values of 119879 and 120579 such that 1198771
0= 1 which indicate
that there exists one maximum allowable period of pulsevaccination 119879120579max such that 119877
1
0lt 1 for all 119879 lt 119879120579max Moreover
the larger 120579 the smaller the value that119879120579max has Unfortunately
Abstract and Applied Analysis 5
we cannot get its analytical expression due to the complexityof 11987710 So we turn to find some approximations for 119879120579max in the
followingIf the PVS is applied frequently enough such that
d119868(119905)d119905 le 0 for all 119905 gt 0 in model (4) then the numberof infectious individuals is a decreasing function of time Itis possible to satisfy this condition if pulsing ensures that119878(119905) le 119878
119888≐ (119887 + 120574)120573 for all 119905 that is pulse vaccination
is applied every time once the 119878(119905) approaches the thresholdvalue 119878
119888(see [7 27] and the references therein)
It follows from the first equation of model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (17)
Consider the following comparison equation
d119909 (119905)d119905
= 119887 minus 119887119909 (119905) 119909 (0) =(1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
(18)
which gives
119909 (119905) = 1 + ((1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
minus 1) eminus119887119905 (19)
Hence
119878 (119905) le 119909 (119905) = 1 + ((1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
minus 1) eminus119887119905 le 119878119888
(20)
and solving the above inequality with respect to 119905 yields
0 le 119905 le1
119887ln(1 +
119901max1198782
119888
(119878119888+ 120579) (1 minus 119878
119888)) (21)
which indicates that
119879120579
max =1
119887ln(1 +
119901max1198782
119888
(119878119888+ 120579) (1 minus 119878
119888)) (22)
Thus if the nonlinear pulse vaccination strategy is appliedperiodically with a period less than 119879120579max then the numberof infectious in the population will decrease forever andeventually die out
We also note that if 120579 = 0 the maximum allowableperiod 119879120579max becomes 1198790max = (1119887) ln(1 + ((119901max119878119888)(1 minus119878119888))) and this is the case studied in [7 9] for a constant
proportion pulse vaccination strategy Obviously 119879120579max is amonotonically decreasing function with respect to 120579 whichgives that 119879120579max lt 119879
0
max This theoretical result indicates that ifwe aim to eradicate an infectious disease then it is necessaryto carry out pulse vaccination programs more frequentlyunder resource limitation than when necessary resourcesare available When various nonlinear factors including119901max 120579 are seriously considered our results show that it isgetting more and more difficult to eliminate the endemicdiseases It also confirms that getting vaccinated is by farthe most effective action that susceptibles can take to protect
themselves and their family from infectious disease Otheroptions available to health practitioners in response to limitedmedical resources include the release of prescriptive healtheducation training vaccinators and promoting the use ofvaccine-delivery patchwith dissolvingmicroneedles (see [14ndash16] for details)
A typical solution of the SIR model with a nonlinearpulse vaccination strategy is shown in Figure 2 where weobserve how the variable 119878(119905) oscillates in a stable cycle(as shown in Figure 2(a)) In contrast the infection 119868(119905)
rapidly decreases to zero (Figure 2(b)) if we set 120579 = 0That is without resource limitation the infectious diseaseis eventually eradicated (Figure 2(b)) However if there isresource limitation that is the parameter 120579 is larger thanzero then both susceptible and infected populations oscillateperiodically as shown in Figures 2(c) and 2(d) with 120579 = 02
Therefore if we have enough vaccine and vaccinators tocombat the infectious disease then it is reasonable that thesusceptible population vaccinated each time is assumed to beproportional to the number of susceptible individuals (here119901max) However this approximation cannot reflect the realcase if we aim to mitigate the emerging infectious diseaseUsually the number of susceptible individuals needing tobe vaccinated may exceed the capacity of local medicalconditions due to shortages of vaccine and doctors especiallyin rural areas in many developing countries where reachingall of the target population may be difficult For instancein 2010 in Lesotho diphtheria tetanus toxoid and pertussis(DTP3) immunization coverage among 1-year-olds was only83 (but an improvement on 78 in 2001 see [31]) and boththe measles and tuberculosis (BCG) vaccination programswere hampered by supplies of vaccine running out in someparts of the country and then allowing vaccinations only forchildren born in hospitals and not for those born elsewherein the communities Thus it is important to design pulsevaccination campaigns carefully For example how frequentlyshould the vaccination strategy be applied And how can thesize of the susceptible population be evaluatedThose are keyissues for evaluating vaccination coverages and the distribu-tion of vaccines Therefore with the help of mathematicalmodels the vaccination strategy can be properly designed
It is well known that in order to eradicate an infectiousdisease the vaccination strategy must be designed such thatthe threshold value 1198771
0is less than 1 which is equivalent to
1198772
0lt 0 where
1198772
0=120573 (119878 minus 1) (1 minus eminus119887119879)
119887+ (120573 minus 120574 minus 119887) 119879 (23)
In the following we employ the formula 11987720to investigate
the important factors which affect the threshold values mostsignificantly by using uncertainty and sensitivity analysis(see [21 22 32]) Sensitivity analysis of the most significantparameters including the birth or death rate (119887) the contactrate (120573) the maximum proportion vaccinated (119901max) and thepulse vaccination period (119879) was performed by evaluatingthe PRCCs for various input parameters against the thresholdcondition 1198772
0with the LHS method [21 22 32] By using
6 Abstract and Applied Analysis
0 10 20 30 400
002
004
006
008
01
S(t)
120579 = 0
(a)
0 10 20 30 40minus800
minus600
minus400
minus200
0
t
Log(I(t))
(b)
0 10 20 30 400
002
004
006
008
01
S(t)
120579 = 02
(c)
0 10 20 30 40minus100
minus80
minus60
minus40
minus20
0
t
Log(I(t))
(d)
Figure 2 The effects of resource limitation on dynamics of model (4) with 119901max = 05 119879 = 2 and all other parameter values are given inTable 1 (a and b) 120579 = 0 (c and d) 120579 = 02
LHS with 3000 samples uncertainty and sensitivity analysesfor all parameters in model (4) were determined A uniformdistribution function was used and tested for significantPRCCs for all parameters with wide ranges such as 119887 sim
119880(0001 01) 120573 sim 119880(1000 2500) 119879 sim 119880(01 20) and 120579 sim119880(0 1) and the baseline values of all parameters are givenin the figure legend of Figure 3 Note that for comparativepurposes we chose the same baseline parameter values asthose used in literatures [8 9 33] Unfortunately we do nothave the realistic ranges of parameter values and we chosethose parameter ranges just for illustrative purposes only andthen provided a scientific basis for disease control
Figure 3(a) shows the PRCC results which illustrate thedependence of 1198772
0on each parameter and PRCC scatter
plots for each parameter are given in Figures 3(b)ndash3(g)respectively We considered absolute values of PRCC valueswhich belong to (04 1) (02 04) and (0 02) as indicatingvery importantmoderate or correlations of little significancebetween input parameters and output variables respectivelyThe positive signs of their PRCCs indicate that if the param-eters are increased the value of 1198772
0increases (vice versa)
The negative sign suggests that if increased the value of 11987720
decreases (vice versa)
Therefore the parameters 119887 120573 120579 and 119879 are responsiblefor increasing the values of 1198772
0 so decreasing all those four
parameters can reduce the threshold value and consequentlyare beneficial for infectious disease control As the parametermaximum proportion of pulse vaccination 119901max changes itis negatively and significantly correlated with large PRCCsThus increasing this parameter will result in significantlyreducing the threshold value The most significant controlparameters are 119887 120573 119901max and 119879 (as shown in Figures 3(b)3(c) 3(e) and 3(g)) It is interesting to note that the period119879and the maximum proportion vaccinated 119901max are stronglycorrelated with the threshold value 1198772
0 which indicates that
carefully designing a pulse vaccination strategy is crucial forinfectious disease control
4 Existence of Endemic Periodic Solution andComplex Dynamics
An important issue is how the infectious disease develops ifthe threshold condition 1198771
0is larger than one In particular
what types of dynamic behavior maymodel (4) have once theperiod of pulse vaccination exceeds certain threshold levelsTherefore in this section we first focus on the existence of an
Abstract and Applied Analysis 7
minus1
minus05
0
05
1
b 120573 120574 120579 T
PRCC
s
pmax
(a)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [072346 0]
b
R2 0
(b)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [05492 0]
120573
R2 0
(c)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000
120574
R2 0
[minus012283 14766e minus 011]
(d)
minus2000 0 2000minus2000
minus1000
0
1000
2000 [minus071273 0]
R2 0
pmax
(e)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [004117 0024135]
R2 0
120579
(f)
minus2000 0 2000minus2000
minus1000
0
1000
2000 [091801 0]
T
R2 0
(g)
Figure 3 PRCC results and PRCC scatter plots of the baseline parameters as those shown in Table 1 Here we fix 119901max = 075 120579 = 02 119879 = 5(a) PRCC results (bndashg) PRCC scatter plots The title of each plot represents the PRCC value with the corresponding P value
endemic periodic solution that is to investigate under whatconditions will the infected population oscillate periodicallywith small amplitude and then we choose the period ofpulse vaccination as a bifurcation parameter to investigate thecomplex patterns of the infectious disease dynamics
Next we analyze the existence of an endemic periodicsolution of model (4) near the disease-free periodic solution(119878lowast(119905) 0) using bifurcation theory [34] and set the impulsive
period 119879 as bifurcation parameter For ease of narrative werewrite the model (4) as
d119878 (119905)d119905
= 119887 minus 120573119878 (119905) 119868 (119905) minus 119887119878 (119905) ≐ 1198651(119878 (119905) 119868 (119905))
d119868 (119905)d119905
= 120573119878 (119905) 119868 (119905) minus 120574119868 (119905) minus 119887119868 (119905) ≐ 1198652(119878 (119905) 119868 (119905))
119905 = 119899119879 119899 isinN
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579≐ Θ1(119878 (119905) 119868 (119905))
119868 (119905+
) = 119868 (119905) ≐ Θ2(119878 (119905) 119868 (119905))
119905 = 119899119879 119899 isinN
(24)According to the bifurcation theorem in [34] we can
obtain the following result
Theorem 3 If there exists a 1198790gt 0 satisfying 1198771
0(1198790) = 1 and
all the parameters in model (24) satisfy
120573 (119878 (1198790) minus 1) eminus1198871198790 (
11988711987901198603
1 minus 1198603eminus1198871198790
minus 1) lt 120573 minus 119903 minus 119887 (25)
where 1198603= 1 minus 119901max + ((120579
2119901max)(120579 + 119878
lowast(1198790))2
) then thereexists a supercritical branch at the point 119879
0
8 Abstract and Applied Analysis
5 10 15 20002
004
006
008
01
T
S(t)
(a)
2 4 6 8 10004
005
006
007
008
T
S(t)
(b)
5 6 7 8 9 10005
0055
006
0065
007
T
S(t)
(c)
7 75 8005
0055
006
0065
007
T
S(t)
(d)
Figure 4 Bifurcation diagrams of model (4) with respect to bifurcation parameter 119879 Here 119901max = 075 and all other parameter values aregiven in Table 1 (a) 120579 = 0 which are reproduced from [9] (b) 120579 = 02 (c and d) a magnified part of Figure 4(b)
The existence of 1198790has been indicated in Figure 1 and the
proof ofTheorem 3 is given in Appendix BTheorem 3 showsthat there exists an endemic periodic solution under someconditions providing that the disease-free periodic solutionbecomes unstable It follows from Theorem 3 that if 119879 gt 119879
0
and is close to 1198790 the endemic periodic solution of model (4)
is stableTo investigate the complex dynamics that model (4) can
have we chose the pulse period 119879 as a bifurcation parameterand fixed all other parameters as those in Figure 4 for twodifferent 120579 values Figure 4(a) is a bifurcation diagram with-out resource limitation that is 120579 = 0 which was obtainedby Shulgin et al [9] Figure 4(b) is a bifurcation diagramwith resource limitation that is 120579 = 02 Comparing thosetwo bifurcation diagrams we conclude that the nonlinearpulse vaccination can produce more complex dynamics thanthose for the model with linear pulse vaccination Once thepulse period 119879 exceeds 119879120579max defined in (22) both susceptibleand infected populations can oscillate periodically with alarge amplitude that corresponds to periodic outbreaks ofepidemics As the pulse period is further increased a series ofcomplex and interesting bifurcation phenomena are observed(see Figures 4(a) and 4(b)) Figures 4(b)ndash4(d) indicate thatthe dynamical behavior of model (4) is very complexincluding period doubling bifurcation chaotic attractors
multistability periodic-adding chaos crisis and periodicwindowsMeanwhile bifurcation analyses (ie Figures 4(b)ndash4(d)) also indicate that the model (4) has several differentattractors which can coexist for a wide range of parametersFor example Figure 5 provides an example of two attractorcoexistence when 119879 = 78 Multiple attractor coexistenceindicates that the infectious outbreak patterns depend onthe initial values which may cause difficulties for infectiousdisease control
5 Discussion and Biological Conclusions
In order to understand the effect of resource limitation inparticular lack of vaccine on the transmission of infectiousdisease we deliberately investigated the dynamical behaviorof an SIR epidemic model which incorporates a nonlinearpulse vaccination strategy To this end we introduced anonlinear form 119901(119905) = (119901max119878(119905))(119878(119905) + 120579) as vaccinationproportionThis resulted in interesting and dramatic changesin the dynamical behavior of solutions and they becamemoreand more complicated which means that it is a very difficulttask to control infectious diseases in particular emerginginfectious diseases under resource limitation We discussedthe control strategy based on the threshold value in moredetail through theoretical analysis numerical studies and
Abstract and Applied Analysis 9
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(a)
100 110 120 130 140 150minus13
minus12
minus11
minus10
minus9
minus8
minus7
t
Log(I(t)
)
(b)
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(c)
100 110 120 130 140 150minus16
minus14
minus12
minus10
minus8
minus6
t
Log(I(t)
)
(d)
Figure 5 Coexistence of two stable attractors of model (4) Here 119901max = 075 120579 = 02 119879 = 78 and all other parameter values are given inTable 1 (a and b) The initial value (119878
0 1198680) = (012 005) (c and d) the initial value (119878
0 1198680) = (01 002)
sensitivity analysis As pointed out byAgur et al [7] the costsrisks and the efficacy of the pulse vaccinations are key factorsin eliminating transmission of the measles virus In orderto depict those factors the nonlinear resource limitationsshould be taken into account [9] and the main purpose ofthe present work is to formulate the disease model to addressthe effects of resource limitations on disease control
The results indicate that vaccination as an approach tocontrolling epidemics must be committed to a long-termstrategy especially when resources arelimited To understandthe interactions amongst 119878(119905) and 119868(119905) which are importantfor successful control of the spread of infectious diseasewe have proposed detailed modeling methods involvingvaccination on 119878(119905) and determining the most significantparameters for the basic reproductive number 1198772
0by using
LHSPRCC uncertainty and sensitivity analysis techniques(see Figure 3) Our results indicate that we should paymore attention to the vaccination period and vaccinationproportion which reduce the threshold value 1198772
0or 11987710to
prevent the outbreak of disease Our results clarify that itis more significant to improve immunization programs (ievaccinations as mentioned in [15 16]) under conditions ofresource limitation
The results also indicate that the dynamic behavior ofmodel (4) may be dramatically affected by small changes in
the value of initial densities of susceptible and infected withresource limitation Bifurcation diagrams shown in Figure 4indicate that there are many forms of complexities in model(4) which are related to chaotic bands with periodic win-dows and attractor crises and Figure 5 reveals that theoccurrence of multiple attractors is a common property ofthe SIR model with resource limitation which can help usto further understand the application of a nonlinear pulsevaccination strategy in an SIR model (for more informationabout bifurcation diagrams of impulsive control strategiessee [35 36]) Some complexities are related to the long-termbehavior of population dynamics characterized either bywell behaving relatively simple or very complicated strangeattractors We also found that the routes to chaos are verycomplicated For instance with resource limitation there areseveral hidden factors that can adversely affect the controlstrategy The increasing number of potential complexitiespredicted by the theory does not seem to make this taskany easier Nevertheless identifying complicated possiblychaotic dynamics in population data has remained a majorchallenge in epidemiology studies In particular the sug-gestion that the dynamics of measles are chaotic has beenmuch debated (eg see [37]) and when seeking to establishwhether measles dynamics in New York City were chaoticor not Sugihara and May [38] omitted data from after 1963when vaccinations were introduced as the immunisations
10 Abstract and Applied Analysis
had altered the intrinsic dynamics However our resultsimply that even with immunisations disease dynamics couldbe chaotic under some circumstances and so a reanalysis ofthe New York data including the vaccination period might beworthwhile
In the present paper we have focused on studyingthe effects of dynamical behaviors on the SIR model withnonlinear pulse vaccination For our simple model we haveassumed that the maximal vaccination proportion 119901max isa constant and the pulse period 119879 can be varied Howeverdue to limited vaccine availability the maximal vaccinationproportion 119901max should depend on the vaccine obtained ineach pulse period Thus to describe the impact of a limitedvaccine stockpile a separate differential equation for theamount of vaccine denoted by 119881(119905) could be included inthe model and then 119901(119905) should be a function of 119881(119905) and119878(119905) Meanwhile it is essential to link the costs of developingand implementing controls to population dynamic modelingof disease epidemics in order to consider other resourcelimitations such as quantities of drugs availability of isolationunits numbers of hospital beds and equipmentThese topicswill be considered in further work in the future
Appendices
A Proof of Theorem 2
Firstly we prove the local stability of a periodic solution(119878lowast(119905) 0) which may be determined by considering the
behavior of small amplitude perturbations of the solutionDefine
119878 (119905) = 119880 (119905) + 119878lowast
(119905) 119868 (119905) = 119881 (119905) (A1)
here119880(119905) 119881(119905) are small perturbations whichmay be writtenas
(119880 (119905)
119881 (119905)) = Φ (119905) (
119880 (0)
119881 (0)) 0 le 119905 lt 119879 (A2)
where the fundamental matrixΦ(119905) satisfies
dΦ (119905)d119905
= (minus119887 minus120573119878
lowast
(119905)
0 120573119878lowast
(119905) minus 120574 minus 119887)Φ (119905) (A3)
and Φ(0) = 119868 is the identity matrix So
Φ (119879) = (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 ) (A4)
where 995333 is not necessarily computed in detail as it is notrequired in the following analysis
The resetting impulsive conditions of model (4) become
(119880 (119899119879
+)
119881 (119899119879+)) = (
1 minus 119901max +1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)(119880 (119899119879)
119881 (119899119879))
≐ 119861 (119899119879) (119880 (119899119879)
119881 (119899119879))
(A5)
Hence according to Floquet theory if the moduli of botheigenvalues of the matrix
119872 = 119861 (119879)Φ (119879)
= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)
times (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 )
(A6)
are less than one then the periodic solution (119878lowast(119905) 0) islocally stable In fact two Floquet expressions multiplied arethus
1205821= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119879))2) eminus119887119879
1205822= eint
119879
0(120573119878lowast(119905)minus120574minus119887)d119905
(A7)
It is obvious that 0 lt 1205821lt 1 Thus the stability of (119878lowast(119905) 0)
is decided by whether 1205822lt 1 or not So we conclude that
the disease-free periodic solution (119878lowast(119905) 0) is locally stable ifint119879
0(120573119878lowast(119905) minus 120574 minus 119887)d119905 lt 0 that is
1
119879int
119879
0
119878lowast
(119905) d119905 =(119878 minus 1) (1 minus eminus119887119879) + 119887119879
119887119879lt120574 + 119887
120573 (A8)
and by simple calculation we have
120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]119887119879 (120574 + 119887)
lt 1 (A9)
which holds true due to 11987710lt 1 Therefore we can draw
a conclusion from the above analysis that if 11987710lt 1 the
disease-free periodic solution (119878lowast(119905) 0) of model (4) is locallyasymptotically stable
Next we will prove the global attraction of the disease-free periodic solution (119878lowast(119905) 0) of model (4) Since 1198771
0lt 1
we can choose 1205761gt 0 sufficiently small such that
1
119879int
119879
0
(119878lowast
(119905) + 1205761) d119905 lt
120574 + 119887
120573 (A10)
It follows from model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) 119905 = 119899119879
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579 119905 = 119899119879
(A11)
Consider the comparison equation
d119885 (119905)d119905
= 119887 minus 119887119885 (119905) 119905 = 119899119879
119885 (119905+
) =(1 minus 119901max) 119885
2
(119905) + 120579119885 (119905)
119885 (119905) + 120579 119905 = 119899119879
(A12)
Abstract and Applied Analysis 11
From Lemma 1 and the comparison theorem on impul-sive differential equations we have 119878(119905) le 119885(119905) and 119885(119905) rarr119878lowast(119905) as 119905 rarr +infin Hence there exists a 119905
1gt 0 such that
119878 (119905) le 119885 (119905) lt 119878lowast
(119905) + 1205761 (A13)
for all 119905 ge 1199051 here 120576
1gt 0 is small enough
From the second equation of model (4) and (A13) we get
d119868 (119905)d119905
le (120573 (119878lowast
(119905) + 1205761) minus 120574 minus 119887) 119868 (119905) (A14)
for 119905 gt 1199051 then we consider the following comparison
equation with pulses
d1199101(119905)
d119905= (120573 (119878
lowast
(119905) + 1205761) minus 120574 minus 119887) 119910
1(119905) 119905 = 119899119879
1199101(119905+
) = 1199101(119905) 119905 = 119899119879
(A15)
Integrating model (A15) between pulses (119899119879 (119899 + 1)119879] wehave
1199101((119899 + 1) 119879) = 119910
1(119899119879) eint
(119899+1)119879
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101(119899119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A16)
Then by using step by step iterations
1199101(119899119879) = 119910
1((119899 minus 1) 119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101((119899 minus 2) 119879) e2 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= sdot sdot sdot
= 1199101(0) e119899 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A17)
where 1199101(0) = 119910
1(0+) gt 0 we obtain lim
119899rarrinfin1199101(119899119879) =
0 according to (A10) On the other hand integrating andsolving the first equation of model (A15) between pulses itgives
1199101(119905) = 119910
1(119899119879) eint
119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905 119905 isin (119899119879 (119899 + 1) 119879]
(A18)
incorporating into the boundedness of eint119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
due to 11987710lt 1 we have lim
119905rarrinfin1199101(119905) = 0
Let (119878(119905) 119868(119905)) be any solution of model (4) with initialvalue (119878
0 1198680) and 119878
0= 119878(0
+) gt 0 119868
0= 119868(0
+) =
1199101(0) gt 0 according to the comparison theorem we have
lim119905rarrinfin
sup 119868(119905) le lim119905rarrinfin
sup1199101(119905) = 0 Incorporating into
the positivity of 119868(119905) we know that lim119905rarrinfin
119868(119905) = 0Consequently there exists a 119905
2gt 1199051such that 0 lt 119868(119905) le 120576
2
for 119905 ge 1199052 and 120576
2is small enough Further we have
119887 minus (1205731205762+ 119887) 119878 (119905) le
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (A19)
for all 119905 gt 1199052 from which we can obtain the following
equation
d1199102(119905)
d119905= 119887 minus (120573120576
2+ 119887) 119910
2(119905) 119905 = 119899119879
1199102(119905+
) =(1 minus 119901max) 119910
2
2(119905) + 120579119910
2(119905)
1199102(119905) + 120579
119905 = 119899119879
(A20)
By employing the same methods as used for the proof ofLemma 1 we get that the model (A20) has a positive periodicsolution 119910lowast
2(119905) which is globally attractive where
119910lowast
2(119905) = (119910
2minus
119887
1205731205762+ 119887) eminus(1205731205762+119887)(119905minus119899119879) + 119887
1205731205762+ 119887
119905 isin (119899119879 (119899 + 1) 119879]
1199102= 1199102(119899119879+
) = 119910lowast
2(0+
) =minus1198612+ radic11986122minus 411986021198622
21198602
1198602= eminus(1205731205762+119887)119879 [ 119887
1205731205762+ 119887
minus (1 minus 119901max) eminus(1205731205762+119887)119879]
1198612= (eminus(1205731205762+119887)119879 minus 119887
1205731205762+ 119887)
times [2 (1 minus 119901max) eminus(1205731205762+119887)119879minus (120579 +
119887
1205731205762+ 119887)]
1198622= minus[(1 minus 119901max) (
119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)2
+ 120579(119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)]
(A21)
It follows from the comparison theorem on impulsive differ-ential equations that 119910
2(119905) le 119878(119905) le 119885(119905) Moreover 119910
2(119905) rarr
119910lowast
2(119905) and 119885(119905) rarr 119878
lowast(119905) as 119905 rarr +infin Consequently there
exists a 1199053for 1205763small enough such that 119905
3ge 1199052and
119910lowast
2(119905) minus 120576
3lt 119878 (119905) lt 119878
lowast
(119905) + 1205763
(A22)
for 119905 ge 1199053 Let 120576
2rarr 0 Then
119878lowast
(119905) minus 1205763lt 119878 (119905) lt 119878
lowast
(119905) + 1205763 (A23)
Therefore 119878(119905) rarr 119878lowast(119905) as 119905 rarr +infin Thus we
have proved the global stability of the disease-free periodicsolution (119878lowast(119905) 0) of model (4) This completes the proof
B Proof of Theorem 3
In order to apply the bifurcation theory of [34] in the maintext we make the following calculations
1198891015840
0= 1 minus eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
(B1)
If 11988910158400= 0 then 119879
0satisfies the following equation
120573 [(119878 minus 1) (1 minus eminus1198871198790) + 1198871198790]
119887 (120574 + 119887) 1198790
= 1 (B2)
12 Abstract and Applied Analysis
which indicates that there exists a1198790such that 1198771
0= 1 and the
disease-free periodic solution 120575 = (119878lowast(119905) 0) loses its stabilityFurther
120597Φ1(1198790 1198830)
120597119878= eminus1198871198790 gt 0
120597Φ2(1198790 1198830)
120597119868= eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
120597Φ1(1198790 1198830)
120597119868= minusint
1198790
0
120573119878lowast
(V) eminus119887(1198790minusV)eintV
0(120573119878lowast(119905)minus120574minus119887)d119905dV
≐ 119860 lt 0
1198861015840
0= 1 minus 119860
3eminus1198871198790
1198871015840
0= 1 minus 119860
3119860
1205972Φ2(1198790 1198830)
120597119868120597119878= 1205731198790eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
1205972Φ2(1198790 1198830)
1205971198682
= minus1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
119878lowast
(120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585minus119887(Vminus120579)
d120579 dV
lt 0
1205972Φ2(1198790 1198830)
120597119868120597119879= (120573119878
lowast
(1198790) minus 120574 minus 119887) eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
120597Φ1(1198790 1198830)
120597119879= minus119887 (119878 minus 1) eminus1198871198790
119861 = 1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887)
times eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
119862 = 1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
eminus119887(119905minus120579)119878lowast (120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585
d120579 dV
+ 21205731198790
1 minus 1198603119860
1 minus 1198603eminus1198871198790
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
(B3)
It follows that 119861119862 lt 0 and we get the condition that is if theparameters satisfy condition 119861 lt 0 then the model (4) has asupercritical branch at 119879
0
In fact
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0 (B4)
Therefore 119861 lt 0 is equivalent to
1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887) lt 0
(B5)
Obviously 119861 lt 0 due to (25) The proof is completed
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thiswork is supported by theNational Natural Science Foun-dation of China (NSFCs 11171199 11371030 11301320) andthe Fundamental Research Funds for the Central Universities(GK201305010) The authors would like to thank the editorand the referees for their careful reading of the original paperand many valuable comments and suggestions that greatlyimproved the presentation of this paper
References
[1] R M Anderson and R M May ldquoPopulation biology of infec-tious diseases part Irdquo Nature vol 280 no 5721 pp 361ndash3671979
[2] E Hanert E Schumacher and E Deleersnijder ldquoFront dynam-ics in fractional-order epidemic modelsrdquo Journal of TheoreticalBiology vol 279 no 1 pp 9ndash16 2011
[3] H W Hethcote ldquoMathematics of infectious diseasesrdquo SIAMReview vol 42 no 4 pp 599ndash653 2000
[4] A E Fiore C B Bridges and N J Cox ldquoSeasonal influenzavaccinesrdquo Current Topics in Microbiology and Immunology vol333 no 1 pp 43ndash82 2009
[5] Y Chang N T Brewer A C Rinas K Schmitt and J S SmithldquoEvaluating the impact of human papillomavirus vaccinesrdquoVaccine vol 27 no 32 pp 4355ndash4362 2009
[6] T J Liesegang ldquoVaricella zoster virus vaccines effective butconcerns lingerrdquo Canadian Journal of Ophthalmology vol 44no 4 pp 379ndash384 2009
[7] Z Agur L Cojocaru G Mazor R M Anderson and Y LDanon ldquoPulse mass measles vaccination across age cohortsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 90 no 24 pp 11698ndash11702 1993
[8] L Stone B Shulgin and Z Agur ldquoTheoretical examinationof the pulse vaccination policy in the SIR epidemic modelrdquoMathematical and ComputerModelling vol 31 no 4-5 pp 207ndash215 2000
[9] B Shulgin L Stone and Z Agur ldquoPulse vaccination strategy inthe SIR epidemic modelrdquo Bulletin of Mathematical Biology vol60 no 6 pp 1123ndash1148 1998
[10] A DrsquoOnofrio ldquoStability properties of pulse vaccination strategyin SEIR epidemicmodelrdquoMathematical Biosciences vol 179 no1 pp 57ndash72 2002
[11] A Franceschetti and A Pugliese ldquoThreshold behaviour of a SIRepidemicmodel with age structure and immigrationrdquo Journal ofMathematical Biology vol 57 no 1 pp 1ndash27 2008
Abstract and Applied Analysis 13
[12] A J Terry ldquoPulse vaccination strategies in a metapopulationSIR modelrdquo Mathematical Biosciences and Engineering vol 7no 2 pp 455ndash477 2010
[13] S J Gao L S Chen J J Nieto and A Torres ldquoAnalysis of adelayed epidemic model with pulse vaccination and saturationincidencerdquo Vaccine vol 24 no 35-36 pp 6037ndash6045 2006
[14] L Matrajt M E Halloran and I M Longini Jr ldquoOptimal vac-cine allocation for the early mitigation of pandemic influenzardquoPLoS Computational Biology vol 9 no 3 Article ID e10029642013
[15] S P Sullivan D G Koutsonanos M D P Martin et alldquoDissolving polymer microneedle patches for influenza vacci-nationrdquo Nature Medicine vol 16 no 8 pp 915ndash920 2010
[16] Science Daily Web ldquoVaccine-delivery patch with dissolvingmicroneedles eliminates ldquosharpsrdquo boosts protectionrdquo httpwwwsciencedailycomreleases201007100718204733htm
[17] M J Reynolds-Hogland J S Hogland and M S MitchellldquoEvaluating intercepts fromdemographicmodels to understandresource limitation and resource thresholdsrdquo Ecological Mod-elling vol 211 no 3-4 pp 424ndash432 2008
[18] L Chow M Fan and Z L Feng ldquoDynamics of a multigroupepidemiological model with group-targeted vaccination strate-giesrdquo Journal of Theoretical Biology vol 291 no 1 pp 56ndash642011
[19] L H Zhou and M Fan ldquoDynamics of an SIR epidemic modelwith limited medical resources revisitedrdquo Nonlinear AnalysisReal World Applications vol 13 no 1 pp 312ndash324 2012
[20] V Lakshmikantham D D Bainov and P S Simeonov Theoryof Impulsive Differential Equations World Scientific Singapore1989
[21] S M Blower and H Dowlatabadi ldquoSensitivity and uncertaintyanalysis of complex-models of disease transmission An HIVmodel as an examplerdquo International Statistical Review vol 62no 2 pp 229ndash243 1994
[22] S Marino I B Hogue C J Ray and D E Kirschner ldquoA meth-odology for performing global uncertainty and sensitivityanalysis in systems biologyrdquo Journal of Theoretical Biology vol254 no 1 pp 178ndash196 2008
[23] S Y Tang J H Liang Y S Tan and R A Cheke ldquoThresholdconditions for integrated pest management models with pesti-cides that have residual effectsrdquo Journal ofMathematical Biologyvol 66 no 1-2 pp 1ndash35 2013
[24] F Brauer and C Castillo-ChavezMathematical Models in Pop-ulation Biology and Epidemiology Springer New York NYUSA 2000
[25] O Diekmann and J A P Heesterbeek Mathematical Epidemi-ology of Infectious Diseases JohnWiley amp Sons Chichester UK2000
[26] J D MurrayMathematical Biology Springer Berlin Germany1989
[27] R Anderson and R May Infectious Diseases of HumansDynamics and Control Oxford University Press Oxford UK1995
[28] M Hethcote ldquoThree basic epidemiological modelsrdquo in AppliedMathematical Ecology L Gross T G Hallam and S A LevinEds pp 119ndash144 Springer Berlin Germany 1989
[29] A V Hill ldquoThe possible effects of the aggregation of themolecules of haemoglobin on its dissociation curvesrdquo TheJournal of Physiology vol 40 no 4 pp 4ndash7 1910
[30] P Cull ldquoLocal and global stability for population modelsrdquoBiological Cybernetics vol 54 no 3 pp 141ndash149 1986
[31] WHOWeb ldquoGlobal health observatory data repositoryrdquo 2001httpappswhointghodatatheme=country
[32] M D McKay R J Beckman and W J Conover ldquoComparisonof three methods for selecting values of input variables in theanalysis of output from a computer coderdquo Technometrics vol21 no 2 pp 239ndash245 1979
[33] W M Schaffer L F Olsen G L Truty S L Fulmer andD J Graser ldquoPeriodic and chaotic dynamics in childhoodepidemicsrdquo in From Chemical to Biological Organization MMarkus S Muller and G Nicolis Eds Springer BerlinGermany 1988
[34] A Lakmeche and O Arino ldquoBifurcation of non trivial periodicsolutions of impulsive differential equations arising chemother-apeutic treatmentrdquo Dynamics of Continuous Discrete andImpulsive Systems B vol 7 no 2 pp 265ndash287 2000
[35] S Y Tang and L S Chen ldquoMultiple attractors in stage-structured population models with birth pulsesrdquo Bulletin ofMathematical Biology vol 65 no 3 pp 479ndash495 2003
[36] S Y Tang Y N Xiao and R A Cheke ldquoMultiple attractorsof host-parasitoid models with integrated pest managementstrategies eradication persistence and outbreakrdquo TheoreticalPopulation Biology vol 73 no 2 pp 181ndash197 2008
[37] L F Olsen andWM Schaffer ldquoChaos versus noisy periodicityalternative hypotheses for childhood epidemicsrdquo Science vol249 no 4968 pp 499ndash504 1990
[38] G Sugihara and R M May ldquoNonlinear forecasting as a way ofdistinguishing chaos from measurement error in time seriesrdquoNature vol 344 no 6268 pp 734ndash741 1990
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical Problems in Engineering
Hindawi Publishing Corporationhttpwwwhindawicom
Differential EquationsInternational Journal of
Volume 2014
Applied MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Probability and StatisticsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical PhysicsAdvances in
Complex AnalysisJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
OptimizationJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CombinatoricsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Operations ResearchAdvances in
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Function Spaces
Abstract and Applied AnalysisHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of Mathematics and Mathematical Sciences
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Algebra
Discrete Dynamics in Nature and Society
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Decision SciencesAdvances in
Discrete MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014 Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Stochastic AnalysisInternational Journal of

Abstract and Applied Analysis 5
we cannot get its analytical expression due to the complexityof 11987710 So we turn to find some approximations for 119879120579max in the
followingIf the PVS is applied frequently enough such that
d119868(119905)d119905 le 0 for all 119905 gt 0 in model (4) then the numberof infectious individuals is a decreasing function of time Itis possible to satisfy this condition if pulsing ensures that119878(119905) le 119878
119888≐ (119887 + 120574)120573 for all 119905 that is pulse vaccination
is applied every time once the 119878(119905) approaches the thresholdvalue 119878
119888(see [7 27] and the references therein)
It follows from the first equation of model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (17)
Consider the following comparison equation
d119909 (119905)d119905
= 119887 minus 119887119909 (119905) 119909 (0) =(1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
(18)
which gives
119909 (119905) = 1 + ((1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
minus 1) eminus119887119905 (19)
Hence
119878 (119905) le 119909 (119905) = 1 + ((1 minus 119901max) 119878
2
119888+ 120579119878119888
119878119888+ 120579
minus 1) eminus119887119905 le 119878119888
(20)
and solving the above inequality with respect to 119905 yields
0 le 119905 le1
119887ln(1 +
119901max1198782
119888
(119878119888+ 120579) (1 minus 119878
119888)) (21)
which indicates that
119879120579
max =1
119887ln(1 +
119901max1198782
119888
(119878119888+ 120579) (1 minus 119878
119888)) (22)
Thus if the nonlinear pulse vaccination strategy is appliedperiodically with a period less than 119879120579max then the numberof infectious in the population will decrease forever andeventually die out
We also note that if 120579 = 0 the maximum allowableperiod 119879120579max becomes 1198790max = (1119887) ln(1 + ((119901max119878119888)(1 minus119878119888))) and this is the case studied in [7 9] for a constant
proportion pulse vaccination strategy Obviously 119879120579max is amonotonically decreasing function with respect to 120579 whichgives that 119879120579max lt 119879
0
max This theoretical result indicates that ifwe aim to eradicate an infectious disease then it is necessaryto carry out pulse vaccination programs more frequentlyunder resource limitation than when necessary resourcesare available When various nonlinear factors including119901max 120579 are seriously considered our results show that it isgetting more and more difficult to eliminate the endemicdiseases It also confirms that getting vaccinated is by farthe most effective action that susceptibles can take to protect
themselves and their family from infectious disease Otheroptions available to health practitioners in response to limitedmedical resources include the release of prescriptive healtheducation training vaccinators and promoting the use ofvaccine-delivery patchwith dissolvingmicroneedles (see [14ndash16] for details)
A typical solution of the SIR model with a nonlinearpulse vaccination strategy is shown in Figure 2 where weobserve how the variable 119878(119905) oscillates in a stable cycle(as shown in Figure 2(a)) In contrast the infection 119868(119905)
rapidly decreases to zero (Figure 2(b)) if we set 120579 = 0That is without resource limitation the infectious diseaseis eventually eradicated (Figure 2(b)) However if there isresource limitation that is the parameter 120579 is larger thanzero then both susceptible and infected populations oscillateperiodically as shown in Figures 2(c) and 2(d) with 120579 = 02
Therefore if we have enough vaccine and vaccinators tocombat the infectious disease then it is reasonable that thesusceptible population vaccinated each time is assumed to beproportional to the number of susceptible individuals (here119901max) However this approximation cannot reflect the realcase if we aim to mitigate the emerging infectious diseaseUsually the number of susceptible individuals needing tobe vaccinated may exceed the capacity of local medicalconditions due to shortages of vaccine and doctors especiallyin rural areas in many developing countries where reachingall of the target population may be difficult For instancein 2010 in Lesotho diphtheria tetanus toxoid and pertussis(DTP3) immunization coverage among 1-year-olds was only83 (but an improvement on 78 in 2001 see [31]) and boththe measles and tuberculosis (BCG) vaccination programswere hampered by supplies of vaccine running out in someparts of the country and then allowing vaccinations only forchildren born in hospitals and not for those born elsewherein the communities Thus it is important to design pulsevaccination campaigns carefully For example how frequentlyshould the vaccination strategy be applied And how can thesize of the susceptible population be evaluatedThose are keyissues for evaluating vaccination coverages and the distribu-tion of vaccines Therefore with the help of mathematicalmodels the vaccination strategy can be properly designed
It is well known that in order to eradicate an infectiousdisease the vaccination strategy must be designed such thatthe threshold value 1198771
0is less than 1 which is equivalent to
1198772
0lt 0 where
1198772
0=120573 (119878 minus 1) (1 minus eminus119887119879)
119887+ (120573 minus 120574 minus 119887) 119879 (23)
In the following we employ the formula 11987720to investigate
the important factors which affect the threshold values mostsignificantly by using uncertainty and sensitivity analysis(see [21 22 32]) Sensitivity analysis of the most significantparameters including the birth or death rate (119887) the contactrate (120573) the maximum proportion vaccinated (119901max) and thepulse vaccination period (119879) was performed by evaluatingthe PRCCs for various input parameters against the thresholdcondition 1198772
0with the LHS method [21 22 32] By using
6 Abstract and Applied Analysis
0 10 20 30 400
002
004
006
008
01
S(t)
120579 = 0
(a)
0 10 20 30 40minus800
minus600
minus400
minus200
0
t
Log(I(t))
(b)
0 10 20 30 400
002
004
006
008
01
S(t)
120579 = 02
(c)
0 10 20 30 40minus100
minus80
minus60
minus40
minus20
0
t
Log(I(t))
(d)
Figure 2 The effects of resource limitation on dynamics of model (4) with 119901max = 05 119879 = 2 and all other parameter values are given inTable 1 (a and b) 120579 = 0 (c and d) 120579 = 02
LHS with 3000 samples uncertainty and sensitivity analysesfor all parameters in model (4) were determined A uniformdistribution function was used and tested for significantPRCCs for all parameters with wide ranges such as 119887 sim
119880(0001 01) 120573 sim 119880(1000 2500) 119879 sim 119880(01 20) and 120579 sim119880(0 1) and the baseline values of all parameters are givenin the figure legend of Figure 3 Note that for comparativepurposes we chose the same baseline parameter values asthose used in literatures [8 9 33] Unfortunately we do nothave the realistic ranges of parameter values and we chosethose parameter ranges just for illustrative purposes only andthen provided a scientific basis for disease control
Figure 3(a) shows the PRCC results which illustrate thedependence of 1198772
0on each parameter and PRCC scatter
plots for each parameter are given in Figures 3(b)ndash3(g)respectively We considered absolute values of PRCC valueswhich belong to (04 1) (02 04) and (0 02) as indicatingvery importantmoderate or correlations of little significancebetween input parameters and output variables respectivelyThe positive signs of their PRCCs indicate that if the param-eters are increased the value of 1198772
0increases (vice versa)
The negative sign suggests that if increased the value of 11987720
decreases (vice versa)
Therefore the parameters 119887 120573 120579 and 119879 are responsiblefor increasing the values of 1198772
0 so decreasing all those four
parameters can reduce the threshold value and consequentlyare beneficial for infectious disease control As the parametermaximum proportion of pulse vaccination 119901max changes itis negatively and significantly correlated with large PRCCsThus increasing this parameter will result in significantlyreducing the threshold value The most significant controlparameters are 119887 120573 119901max and 119879 (as shown in Figures 3(b)3(c) 3(e) and 3(g)) It is interesting to note that the period119879and the maximum proportion vaccinated 119901max are stronglycorrelated with the threshold value 1198772
0 which indicates that
carefully designing a pulse vaccination strategy is crucial forinfectious disease control
4 Existence of Endemic Periodic Solution andComplex Dynamics
An important issue is how the infectious disease develops ifthe threshold condition 1198771
0is larger than one In particular
what types of dynamic behavior maymodel (4) have once theperiod of pulse vaccination exceeds certain threshold levelsTherefore in this section we first focus on the existence of an
Abstract and Applied Analysis 7
minus1
minus05
0
05
1
b 120573 120574 120579 T
PRCC
s
pmax
(a)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [072346 0]
b
R2 0
(b)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [05492 0]
120573
R2 0
(c)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000
120574
R2 0
[minus012283 14766e minus 011]
(d)
minus2000 0 2000minus2000
minus1000
0
1000
2000 [minus071273 0]
R2 0
pmax
(e)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [004117 0024135]
R2 0
120579
(f)
minus2000 0 2000minus2000
minus1000
0
1000
2000 [091801 0]
T
R2 0
(g)
Figure 3 PRCC results and PRCC scatter plots of the baseline parameters as those shown in Table 1 Here we fix 119901max = 075 120579 = 02 119879 = 5(a) PRCC results (bndashg) PRCC scatter plots The title of each plot represents the PRCC value with the corresponding P value
endemic periodic solution that is to investigate under whatconditions will the infected population oscillate periodicallywith small amplitude and then we choose the period ofpulse vaccination as a bifurcation parameter to investigate thecomplex patterns of the infectious disease dynamics
Next we analyze the existence of an endemic periodicsolution of model (4) near the disease-free periodic solution(119878lowast(119905) 0) using bifurcation theory [34] and set the impulsive
period 119879 as bifurcation parameter For ease of narrative werewrite the model (4) as
d119878 (119905)d119905
= 119887 minus 120573119878 (119905) 119868 (119905) minus 119887119878 (119905) ≐ 1198651(119878 (119905) 119868 (119905))
d119868 (119905)d119905
= 120573119878 (119905) 119868 (119905) minus 120574119868 (119905) minus 119887119868 (119905) ≐ 1198652(119878 (119905) 119868 (119905))
119905 = 119899119879 119899 isinN
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579≐ Θ1(119878 (119905) 119868 (119905))
119868 (119905+
) = 119868 (119905) ≐ Θ2(119878 (119905) 119868 (119905))
119905 = 119899119879 119899 isinN
(24)According to the bifurcation theorem in [34] we can
obtain the following result
Theorem 3 If there exists a 1198790gt 0 satisfying 1198771
0(1198790) = 1 and
all the parameters in model (24) satisfy
120573 (119878 (1198790) minus 1) eminus1198871198790 (
11988711987901198603
1 minus 1198603eminus1198871198790
minus 1) lt 120573 minus 119903 minus 119887 (25)
where 1198603= 1 minus 119901max + ((120579
2119901max)(120579 + 119878
lowast(1198790))2
) then thereexists a supercritical branch at the point 119879
0
8 Abstract and Applied Analysis
5 10 15 20002
004
006
008
01
T
S(t)
(a)
2 4 6 8 10004
005
006
007
008
T
S(t)
(b)
5 6 7 8 9 10005
0055
006
0065
007
T
S(t)
(c)
7 75 8005
0055
006
0065
007
T
S(t)
(d)
Figure 4 Bifurcation diagrams of model (4) with respect to bifurcation parameter 119879 Here 119901max = 075 and all other parameter values aregiven in Table 1 (a) 120579 = 0 which are reproduced from [9] (b) 120579 = 02 (c and d) a magnified part of Figure 4(b)
The existence of 1198790has been indicated in Figure 1 and the
proof ofTheorem 3 is given in Appendix BTheorem 3 showsthat there exists an endemic periodic solution under someconditions providing that the disease-free periodic solutionbecomes unstable It follows from Theorem 3 that if 119879 gt 119879
0
and is close to 1198790 the endemic periodic solution of model (4)
is stableTo investigate the complex dynamics that model (4) can
have we chose the pulse period 119879 as a bifurcation parameterand fixed all other parameters as those in Figure 4 for twodifferent 120579 values Figure 4(a) is a bifurcation diagram with-out resource limitation that is 120579 = 0 which was obtainedby Shulgin et al [9] Figure 4(b) is a bifurcation diagramwith resource limitation that is 120579 = 02 Comparing thosetwo bifurcation diagrams we conclude that the nonlinearpulse vaccination can produce more complex dynamics thanthose for the model with linear pulse vaccination Once thepulse period 119879 exceeds 119879120579max defined in (22) both susceptibleand infected populations can oscillate periodically with alarge amplitude that corresponds to periodic outbreaks ofepidemics As the pulse period is further increased a series ofcomplex and interesting bifurcation phenomena are observed(see Figures 4(a) and 4(b)) Figures 4(b)ndash4(d) indicate thatthe dynamical behavior of model (4) is very complexincluding period doubling bifurcation chaotic attractors
multistability periodic-adding chaos crisis and periodicwindowsMeanwhile bifurcation analyses (ie Figures 4(b)ndash4(d)) also indicate that the model (4) has several differentattractors which can coexist for a wide range of parametersFor example Figure 5 provides an example of two attractorcoexistence when 119879 = 78 Multiple attractor coexistenceindicates that the infectious outbreak patterns depend onthe initial values which may cause difficulties for infectiousdisease control
5 Discussion and Biological Conclusions
In order to understand the effect of resource limitation inparticular lack of vaccine on the transmission of infectiousdisease we deliberately investigated the dynamical behaviorof an SIR epidemic model which incorporates a nonlinearpulse vaccination strategy To this end we introduced anonlinear form 119901(119905) = (119901max119878(119905))(119878(119905) + 120579) as vaccinationproportionThis resulted in interesting and dramatic changesin the dynamical behavior of solutions and they becamemoreand more complicated which means that it is a very difficulttask to control infectious diseases in particular emerginginfectious diseases under resource limitation We discussedthe control strategy based on the threshold value in moredetail through theoretical analysis numerical studies and
Abstract and Applied Analysis 9
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(a)
100 110 120 130 140 150minus13
minus12
minus11
minus10
minus9
minus8
minus7
t
Log(I(t)
)
(b)
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(c)
100 110 120 130 140 150minus16
minus14
minus12
minus10
minus8
minus6
t
Log(I(t)
)
(d)
Figure 5 Coexistence of two stable attractors of model (4) Here 119901max = 075 120579 = 02 119879 = 78 and all other parameter values are given inTable 1 (a and b) The initial value (119878
0 1198680) = (012 005) (c and d) the initial value (119878
0 1198680) = (01 002)
sensitivity analysis As pointed out byAgur et al [7] the costsrisks and the efficacy of the pulse vaccinations are key factorsin eliminating transmission of the measles virus In orderto depict those factors the nonlinear resource limitationsshould be taken into account [9] and the main purpose ofthe present work is to formulate the disease model to addressthe effects of resource limitations on disease control
The results indicate that vaccination as an approach tocontrolling epidemics must be committed to a long-termstrategy especially when resources arelimited To understandthe interactions amongst 119878(119905) and 119868(119905) which are importantfor successful control of the spread of infectious diseasewe have proposed detailed modeling methods involvingvaccination on 119878(119905) and determining the most significantparameters for the basic reproductive number 1198772
0by using
LHSPRCC uncertainty and sensitivity analysis techniques(see Figure 3) Our results indicate that we should paymore attention to the vaccination period and vaccinationproportion which reduce the threshold value 1198772
0or 11987710to
prevent the outbreak of disease Our results clarify that itis more significant to improve immunization programs (ievaccinations as mentioned in [15 16]) under conditions ofresource limitation
The results also indicate that the dynamic behavior ofmodel (4) may be dramatically affected by small changes in
the value of initial densities of susceptible and infected withresource limitation Bifurcation diagrams shown in Figure 4indicate that there are many forms of complexities in model(4) which are related to chaotic bands with periodic win-dows and attractor crises and Figure 5 reveals that theoccurrence of multiple attractors is a common property ofthe SIR model with resource limitation which can help usto further understand the application of a nonlinear pulsevaccination strategy in an SIR model (for more informationabout bifurcation diagrams of impulsive control strategiessee [35 36]) Some complexities are related to the long-termbehavior of population dynamics characterized either bywell behaving relatively simple or very complicated strangeattractors We also found that the routes to chaos are verycomplicated For instance with resource limitation there areseveral hidden factors that can adversely affect the controlstrategy The increasing number of potential complexitiespredicted by the theory does not seem to make this taskany easier Nevertheless identifying complicated possiblychaotic dynamics in population data has remained a majorchallenge in epidemiology studies In particular the sug-gestion that the dynamics of measles are chaotic has beenmuch debated (eg see [37]) and when seeking to establishwhether measles dynamics in New York City were chaoticor not Sugihara and May [38] omitted data from after 1963when vaccinations were introduced as the immunisations
10 Abstract and Applied Analysis
had altered the intrinsic dynamics However our resultsimply that even with immunisations disease dynamics couldbe chaotic under some circumstances and so a reanalysis ofthe New York data including the vaccination period might beworthwhile
In the present paper we have focused on studyingthe effects of dynamical behaviors on the SIR model withnonlinear pulse vaccination For our simple model we haveassumed that the maximal vaccination proportion 119901max isa constant and the pulse period 119879 can be varied Howeverdue to limited vaccine availability the maximal vaccinationproportion 119901max should depend on the vaccine obtained ineach pulse period Thus to describe the impact of a limitedvaccine stockpile a separate differential equation for theamount of vaccine denoted by 119881(119905) could be included inthe model and then 119901(119905) should be a function of 119881(119905) and119878(119905) Meanwhile it is essential to link the costs of developingand implementing controls to population dynamic modelingof disease epidemics in order to consider other resourcelimitations such as quantities of drugs availability of isolationunits numbers of hospital beds and equipmentThese topicswill be considered in further work in the future
Appendices
A Proof of Theorem 2
Firstly we prove the local stability of a periodic solution(119878lowast(119905) 0) which may be determined by considering the
behavior of small amplitude perturbations of the solutionDefine
119878 (119905) = 119880 (119905) + 119878lowast
(119905) 119868 (119905) = 119881 (119905) (A1)
here119880(119905) 119881(119905) are small perturbations whichmay be writtenas
(119880 (119905)
119881 (119905)) = Φ (119905) (
119880 (0)
119881 (0)) 0 le 119905 lt 119879 (A2)
where the fundamental matrixΦ(119905) satisfies
dΦ (119905)d119905
= (minus119887 minus120573119878
lowast
(119905)
0 120573119878lowast
(119905) minus 120574 minus 119887)Φ (119905) (A3)
and Φ(0) = 119868 is the identity matrix So
Φ (119879) = (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 ) (A4)
where 995333 is not necessarily computed in detail as it is notrequired in the following analysis
The resetting impulsive conditions of model (4) become
(119880 (119899119879
+)
119881 (119899119879+)) = (
1 minus 119901max +1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)(119880 (119899119879)
119881 (119899119879))
≐ 119861 (119899119879) (119880 (119899119879)
119881 (119899119879))
(A5)
Hence according to Floquet theory if the moduli of botheigenvalues of the matrix
119872 = 119861 (119879)Φ (119879)
= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)
times (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 )
(A6)
are less than one then the periodic solution (119878lowast(119905) 0) islocally stable In fact two Floquet expressions multiplied arethus
1205821= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119879))2) eminus119887119879
1205822= eint
119879
0(120573119878lowast(119905)minus120574minus119887)d119905
(A7)
It is obvious that 0 lt 1205821lt 1 Thus the stability of (119878lowast(119905) 0)
is decided by whether 1205822lt 1 or not So we conclude that
the disease-free periodic solution (119878lowast(119905) 0) is locally stable ifint119879
0(120573119878lowast(119905) minus 120574 minus 119887)d119905 lt 0 that is
1
119879int
119879
0
119878lowast
(119905) d119905 =(119878 minus 1) (1 minus eminus119887119879) + 119887119879
119887119879lt120574 + 119887
120573 (A8)
and by simple calculation we have
120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]119887119879 (120574 + 119887)
lt 1 (A9)
which holds true due to 11987710lt 1 Therefore we can draw
a conclusion from the above analysis that if 11987710lt 1 the
disease-free periodic solution (119878lowast(119905) 0) of model (4) is locallyasymptotically stable
Next we will prove the global attraction of the disease-free periodic solution (119878lowast(119905) 0) of model (4) Since 1198771
0lt 1
we can choose 1205761gt 0 sufficiently small such that
1
119879int
119879
0
(119878lowast
(119905) + 1205761) d119905 lt
120574 + 119887
120573 (A10)
It follows from model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) 119905 = 119899119879
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579 119905 = 119899119879
(A11)
Consider the comparison equation
d119885 (119905)d119905
= 119887 minus 119887119885 (119905) 119905 = 119899119879
119885 (119905+
) =(1 minus 119901max) 119885
2
(119905) + 120579119885 (119905)
119885 (119905) + 120579 119905 = 119899119879
(A12)
Abstract and Applied Analysis 11
From Lemma 1 and the comparison theorem on impul-sive differential equations we have 119878(119905) le 119885(119905) and 119885(119905) rarr119878lowast(119905) as 119905 rarr +infin Hence there exists a 119905
1gt 0 such that
119878 (119905) le 119885 (119905) lt 119878lowast
(119905) + 1205761 (A13)
for all 119905 ge 1199051 here 120576
1gt 0 is small enough
From the second equation of model (4) and (A13) we get
d119868 (119905)d119905
le (120573 (119878lowast
(119905) + 1205761) minus 120574 minus 119887) 119868 (119905) (A14)
for 119905 gt 1199051 then we consider the following comparison
equation with pulses
d1199101(119905)
d119905= (120573 (119878
lowast
(119905) + 1205761) minus 120574 minus 119887) 119910
1(119905) 119905 = 119899119879
1199101(119905+
) = 1199101(119905) 119905 = 119899119879
(A15)
Integrating model (A15) between pulses (119899119879 (119899 + 1)119879] wehave
1199101((119899 + 1) 119879) = 119910
1(119899119879) eint
(119899+1)119879
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101(119899119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A16)
Then by using step by step iterations
1199101(119899119879) = 119910
1((119899 minus 1) 119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101((119899 minus 2) 119879) e2 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= sdot sdot sdot
= 1199101(0) e119899 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A17)
where 1199101(0) = 119910
1(0+) gt 0 we obtain lim
119899rarrinfin1199101(119899119879) =
0 according to (A10) On the other hand integrating andsolving the first equation of model (A15) between pulses itgives
1199101(119905) = 119910
1(119899119879) eint
119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905 119905 isin (119899119879 (119899 + 1) 119879]
(A18)
incorporating into the boundedness of eint119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
due to 11987710lt 1 we have lim
119905rarrinfin1199101(119905) = 0
Let (119878(119905) 119868(119905)) be any solution of model (4) with initialvalue (119878
0 1198680) and 119878
0= 119878(0
+) gt 0 119868
0= 119868(0
+) =
1199101(0) gt 0 according to the comparison theorem we have
lim119905rarrinfin
sup 119868(119905) le lim119905rarrinfin
sup1199101(119905) = 0 Incorporating into
the positivity of 119868(119905) we know that lim119905rarrinfin
119868(119905) = 0Consequently there exists a 119905
2gt 1199051such that 0 lt 119868(119905) le 120576
2
for 119905 ge 1199052 and 120576
2is small enough Further we have
119887 minus (1205731205762+ 119887) 119878 (119905) le
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (A19)
for all 119905 gt 1199052 from which we can obtain the following
equation
d1199102(119905)
d119905= 119887 minus (120573120576
2+ 119887) 119910
2(119905) 119905 = 119899119879
1199102(119905+
) =(1 minus 119901max) 119910
2
2(119905) + 120579119910
2(119905)
1199102(119905) + 120579
119905 = 119899119879
(A20)
By employing the same methods as used for the proof ofLemma 1 we get that the model (A20) has a positive periodicsolution 119910lowast
2(119905) which is globally attractive where
119910lowast
2(119905) = (119910
2minus
119887
1205731205762+ 119887) eminus(1205731205762+119887)(119905minus119899119879) + 119887
1205731205762+ 119887
119905 isin (119899119879 (119899 + 1) 119879]
1199102= 1199102(119899119879+
) = 119910lowast
2(0+
) =minus1198612+ radic11986122minus 411986021198622
21198602
1198602= eminus(1205731205762+119887)119879 [ 119887
1205731205762+ 119887
minus (1 minus 119901max) eminus(1205731205762+119887)119879]
1198612= (eminus(1205731205762+119887)119879 minus 119887
1205731205762+ 119887)
times [2 (1 minus 119901max) eminus(1205731205762+119887)119879minus (120579 +
119887
1205731205762+ 119887)]
1198622= minus[(1 minus 119901max) (
119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)2
+ 120579(119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)]
(A21)
It follows from the comparison theorem on impulsive differ-ential equations that 119910
2(119905) le 119878(119905) le 119885(119905) Moreover 119910
2(119905) rarr
119910lowast
2(119905) and 119885(119905) rarr 119878
lowast(119905) as 119905 rarr +infin Consequently there
exists a 1199053for 1205763small enough such that 119905
3ge 1199052and
119910lowast
2(119905) minus 120576
3lt 119878 (119905) lt 119878
lowast
(119905) + 1205763
(A22)
for 119905 ge 1199053 Let 120576
2rarr 0 Then
119878lowast
(119905) minus 1205763lt 119878 (119905) lt 119878
lowast
(119905) + 1205763 (A23)
Therefore 119878(119905) rarr 119878lowast(119905) as 119905 rarr +infin Thus we
have proved the global stability of the disease-free periodicsolution (119878lowast(119905) 0) of model (4) This completes the proof
B Proof of Theorem 3
In order to apply the bifurcation theory of [34] in the maintext we make the following calculations
1198891015840
0= 1 minus eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
(B1)
If 11988910158400= 0 then 119879
0satisfies the following equation
120573 [(119878 minus 1) (1 minus eminus1198871198790) + 1198871198790]
119887 (120574 + 119887) 1198790
= 1 (B2)
12 Abstract and Applied Analysis
which indicates that there exists a1198790such that 1198771
0= 1 and the
disease-free periodic solution 120575 = (119878lowast(119905) 0) loses its stabilityFurther
120597Φ1(1198790 1198830)
120597119878= eminus1198871198790 gt 0
120597Φ2(1198790 1198830)
120597119868= eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
120597Φ1(1198790 1198830)
120597119868= minusint
1198790
0
120573119878lowast
(V) eminus119887(1198790minusV)eintV
0(120573119878lowast(119905)minus120574minus119887)d119905dV
≐ 119860 lt 0
1198861015840
0= 1 minus 119860
3eminus1198871198790
1198871015840
0= 1 minus 119860
3119860
1205972Φ2(1198790 1198830)
120597119868120597119878= 1205731198790eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
1205972Φ2(1198790 1198830)
1205971198682
= minus1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
119878lowast
(120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585minus119887(Vminus120579)
d120579 dV
lt 0
1205972Φ2(1198790 1198830)
120597119868120597119879= (120573119878
lowast
(1198790) minus 120574 minus 119887) eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
120597Φ1(1198790 1198830)
120597119879= minus119887 (119878 minus 1) eminus1198871198790
119861 = 1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887)
times eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
119862 = 1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
eminus119887(119905minus120579)119878lowast (120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585
d120579 dV
+ 21205731198790
1 minus 1198603119860
1 minus 1198603eminus1198871198790
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
(B3)
It follows that 119861119862 lt 0 and we get the condition that is if theparameters satisfy condition 119861 lt 0 then the model (4) has asupercritical branch at 119879
0
In fact
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0 (B4)
Therefore 119861 lt 0 is equivalent to
1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887) lt 0
(B5)
Obviously 119861 lt 0 due to (25) The proof is completed
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thiswork is supported by theNational Natural Science Foun-dation of China (NSFCs 11171199 11371030 11301320) andthe Fundamental Research Funds for the Central Universities(GK201305010) The authors would like to thank the editorand the referees for their careful reading of the original paperand many valuable comments and suggestions that greatlyimproved the presentation of this paper
References
[1] R M Anderson and R M May ldquoPopulation biology of infec-tious diseases part Irdquo Nature vol 280 no 5721 pp 361ndash3671979
[2] E Hanert E Schumacher and E Deleersnijder ldquoFront dynam-ics in fractional-order epidemic modelsrdquo Journal of TheoreticalBiology vol 279 no 1 pp 9ndash16 2011
[3] H W Hethcote ldquoMathematics of infectious diseasesrdquo SIAMReview vol 42 no 4 pp 599ndash653 2000
[4] A E Fiore C B Bridges and N J Cox ldquoSeasonal influenzavaccinesrdquo Current Topics in Microbiology and Immunology vol333 no 1 pp 43ndash82 2009
[5] Y Chang N T Brewer A C Rinas K Schmitt and J S SmithldquoEvaluating the impact of human papillomavirus vaccinesrdquoVaccine vol 27 no 32 pp 4355ndash4362 2009
[6] T J Liesegang ldquoVaricella zoster virus vaccines effective butconcerns lingerrdquo Canadian Journal of Ophthalmology vol 44no 4 pp 379ndash384 2009
[7] Z Agur L Cojocaru G Mazor R M Anderson and Y LDanon ldquoPulse mass measles vaccination across age cohortsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 90 no 24 pp 11698ndash11702 1993
[8] L Stone B Shulgin and Z Agur ldquoTheoretical examinationof the pulse vaccination policy in the SIR epidemic modelrdquoMathematical and ComputerModelling vol 31 no 4-5 pp 207ndash215 2000
[9] B Shulgin L Stone and Z Agur ldquoPulse vaccination strategy inthe SIR epidemic modelrdquo Bulletin of Mathematical Biology vol60 no 6 pp 1123ndash1148 1998
[10] A DrsquoOnofrio ldquoStability properties of pulse vaccination strategyin SEIR epidemicmodelrdquoMathematical Biosciences vol 179 no1 pp 57ndash72 2002
[11] A Franceschetti and A Pugliese ldquoThreshold behaviour of a SIRepidemicmodel with age structure and immigrationrdquo Journal ofMathematical Biology vol 57 no 1 pp 1ndash27 2008
Abstract and Applied Analysis 13
[12] A J Terry ldquoPulse vaccination strategies in a metapopulationSIR modelrdquo Mathematical Biosciences and Engineering vol 7no 2 pp 455ndash477 2010
[13] S J Gao L S Chen J J Nieto and A Torres ldquoAnalysis of adelayed epidemic model with pulse vaccination and saturationincidencerdquo Vaccine vol 24 no 35-36 pp 6037ndash6045 2006
[14] L Matrajt M E Halloran and I M Longini Jr ldquoOptimal vac-cine allocation for the early mitigation of pandemic influenzardquoPLoS Computational Biology vol 9 no 3 Article ID e10029642013
[15] S P Sullivan D G Koutsonanos M D P Martin et alldquoDissolving polymer microneedle patches for influenza vacci-nationrdquo Nature Medicine vol 16 no 8 pp 915ndash920 2010
[16] Science Daily Web ldquoVaccine-delivery patch with dissolvingmicroneedles eliminates ldquosharpsrdquo boosts protectionrdquo httpwwwsciencedailycomreleases201007100718204733htm
[17] M J Reynolds-Hogland J S Hogland and M S MitchellldquoEvaluating intercepts fromdemographicmodels to understandresource limitation and resource thresholdsrdquo Ecological Mod-elling vol 211 no 3-4 pp 424ndash432 2008
[18] L Chow M Fan and Z L Feng ldquoDynamics of a multigroupepidemiological model with group-targeted vaccination strate-giesrdquo Journal of Theoretical Biology vol 291 no 1 pp 56ndash642011
[19] L H Zhou and M Fan ldquoDynamics of an SIR epidemic modelwith limited medical resources revisitedrdquo Nonlinear AnalysisReal World Applications vol 13 no 1 pp 312ndash324 2012
[20] V Lakshmikantham D D Bainov and P S Simeonov Theoryof Impulsive Differential Equations World Scientific Singapore1989
[21] S M Blower and H Dowlatabadi ldquoSensitivity and uncertaintyanalysis of complex-models of disease transmission An HIVmodel as an examplerdquo International Statistical Review vol 62no 2 pp 229ndash243 1994
[22] S Marino I B Hogue C J Ray and D E Kirschner ldquoA meth-odology for performing global uncertainty and sensitivityanalysis in systems biologyrdquo Journal of Theoretical Biology vol254 no 1 pp 178ndash196 2008
[23] S Y Tang J H Liang Y S Tan and R A Cheke ldquoThresholdconditions for integrated pest management models with pesti-cides that have residual effectsrdquo Journal ofMathematical Biologyvol 66 no 1-2 pp 1ndash35 2013
[24] F Brauer and C Castillo-ChavezMathematical Models in Pop-ulation Biology and Epidemiology Springer New York NYUSA 2000
[25] O Diekmann and J A P Heesterbeek Mathematical Epidemi-ology of Infectious Diseases JohnWiley amp Sons Chichester UK2000
[26] J D MurrayMathematical Biology Springer Berlin Germany1989
[27] R Anderson and R May Infectious Diseases of HumansDynamics and Control Oxford University Press Oxford UK1995
[28] M Hethcote ldquoThree basic epidemiological modelsrdquo in AppliedMathematical Ecology L Gross T G Hallam and S A LevinEds pp 119ndash144 Springer Berlin Germany 1989
[29] A V Hill ldquoThe possible effects of the aggregation of themolecules of haemoglobin on its dissociation curvesrdquo TheJournal of Physiology vol 40 no 4 pp 4ndash7 1910
[30] P Cull ldquoLocal and global stability for population modelsrdquoBiological Cybernetics vol 54 no 3 pp 141ndash149 1986
[31] WHOWeb ldquoGlobal health observatory data repositoryrdquo 2001httpappswhointghodatatheme=country
[32] M D McKay R J Beckman and W J Conover ldquoComparisonof three methods for selecting values of input variables in theanalysis of output from a computer coderdquo Technometrics vol21 no 2 pp 239ndash245 1979
[33] W M Schaffer L F Olsen G L Truty S L Fulmer andD J Graser ldquoPeriodic and chaotic dynamics in childhoodepidemicsrdquo in From Chemical to Biological Organization MMarkus S Muller and G Nicolis Eds Springer BerlinGermany 1988
[34] A Lakmeche and O Arino ldquoBifurcation of non trivial periodicsolutions of impulsive differential equations arising chemother-apeutic treatmentrdquo Dynamics of Continuous Discrete andImpulsive Systems B vol 7 no 2 pp 265ndash287 2000
[35] S Y Tang and L S Chen ldquoMultiple attractors in stage-structured population models with birth pulsesrdquo Bulletin ofMathematical Biology vol 65 no 3 pp 479ndash495 2003
[36] S Y Tang Y N Xiao and R A Cheke ldquoMultiple attractorsof host-parasitoid models with integrated pest managementstrategies eradication persistence and outbreakrdquo TheoreticalPopulation Biology vol 73 no 2 pp 181ndash197 2008
[37] L F Olsen andWM Schaffer ldquoChaos versus noisy periodicityalternative hypotheses for childhood epidemicsrdquo Science vol249 no 4968 pp 499ndash504 1990
[38] G Sugihara and R M May ldquoNonlinear forecasting as a way ofdistinguishing chaos from measurement error in time seriesrdquoNature vol 344 no 6268 pp 734ndash741 1990
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical Problems in Engineering
Hindawi Publishing Corporationhttpwwwhindawicom
Differential EquationsInternational Journal of
Volume 2014
Applied MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Probability and StatisticsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical PhysicsAdvances in
Complex AnalysisJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
OptimizationJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CombinatoricsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Operations ResearchAdvances in
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Function Spaces
Abstract and Applied AnalysisHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of Mathematics and Mathematical Sciences
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Algebra
Discrete Dynamics in Nature and Society
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Decision SciencesAdvances in
Discrete MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014 Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Stochastic AnalysisInternational Journal of

6 Abstract and Applied Analysis
0 10 20 30 400
002
004
006
008
01
S(t)
120579 = 0
(a)
0 10 20 30 40minus800
minus600
minus400
minus200
0
t
Log(I(t))
(b)
0 10 20 30 400
002
004
006
008
01
S(t)
120579 = 02
(c)
0 10 20 30 40minus100
minus80
minus60
minus40
minus20
0
t
Log(I(t))
(d)
Figure 2 The effects of resource limitation on dynamics of model (4) with 119901max = 05 119879 = 2 and all other parameter values are given inTable 1 (a and b) 120579 = 0 (c and d) 120579 = 02
LHS with 3000 samples uncertainty and sensitivity analysesfor all parameters in model (4) were determined A uniformdistribution function was used and tested for significantPRCCs for all parameters with wide ranges such as 119887 sim
119880(0001 01) 120573 sim 119880(1000 2500) 119879 sim 119880(01 20) and 120579 sim119880(0 1) and the baseline values of all parameters are givenin the figure legend of Figure 3 Note that for comparativepurposes we chose the same baseline parameter values asthose used in literatures [8 9 33] Unfortunately we do nothave the realistic ranges of parameter values and we chosethose parameter ranges just for illustrative purposes only andthen provided a scientific basis for disease control
Figure 3(a) shows the PRCC results which illustrate thedependence of 1198772
0on each parameter and PRCC scatter
plots for each parameter are given in Figures 3(b)ndash3(g)respectively We considered absolute values of PRCC valueswhich belong to (04 1) (02 04) and (0 02) as indicatingvery importantmoderate or correlations of little significancebetween input parameters and output variables respectivelyThe positive signs of their PRCCs indicate that if the param-eters are increased the value of 1198772
0increases (vice versa)
The negative sign suggests that if increased the value of 11987720
decreases (vice versa)
Therefore the parameters 119887 120573 120579 and 119879 are responsiblefor increasing the values of 1198772
0 so decreasing all those four
parameters can reduce the threshold value and consequentlyare beneficial for infectious disease control As the parametermaximum proportion of pulse vaccination 119901max changes itis negatively and significantly correlated with large PRCCsThus increasing this parameter will result in significantlyreducing the threshold value The most significant controlparameters are 119887 120573 119901max and 119879 (as shown in Figures 3(b)3(c) 3(e) and 3(g)) It is interesting to note that the period119879and the maximum proportion vaccinated 119901max are stronglycorrelated with the threshold value 1198772
0 which indicates that
carefully designing a pulse vaccination strategy is crucial forinfectious disease control
4 Existence of Endemic Periodic Solution andComplex Dynamics
An important issue is how the infectious disease develops ifthe threshold condition 1198771
0is larger than one In particular
what types of dynamic behavior maymodel (4) have once theperiod of pulse vaccination exceeds certain threshold levelsTherefore in this section we first focus on the existence of an
Abstract and Applied Analysis 7
minus1
minus05
0
05
1
b 120573 120574 120579 T
PRCC
s
pmax
(a)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [072346 0]
b
R2 0
(b)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [05492 0]
120573
R2 0
(c)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000
120574
R2 0
[minus012283 14766e minus 011]
(d)
minus2000 0 2000minus2000
minus1000
0
1000
2000 [minus071273 0]
R2 0
pmax
(e)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [004117 0024135]
R2 0
120579
(f)
minus2000 0 2000minus2000
minus1000
0
1000
2000 [091801 0]
T
R2 0
(g)
Figure 3 PRCC results and PRCC scatter plots of the baseline parameters as those shown in Table 1 Here we fix 119901max = 075 120579 = 02 119879 = 5(a) PRCC results (bndashg) PRCC scatter plots The title of each plot represents the PRCC value with the corresponding P value
endemic periodic solution that is to investigate under whatconditions will the infected population oscillate periodicallywith small amplitude and then we choose the period ofpulse vaccination as a bifurcation parameter to investigate thecomplex patterns of the infectious disease dynamics
Next we analyze the existence of an endemic periodicsolution of model (4) near the disease-free periodic solution(119878lowast(119905) 0) using bifurcation theory [34] and set the impulsive
period 119879 as bifurcation parameter For ease of narrative werewrite the model (4) as
d119878 (119905)d119905
= 119887 minus 120573119878 (119905) 119868 (119905) minus 119887119878 (119905) ≐ 1198651(119878 (119905) 119868 (119905))
d119868 (119905)d119905
= 120573119878 (119905) 119868 (119905) minus 120574119868 (119905) minus 119887119868 (119905) ≐ 1198652(119878 (119905) 119868 (119905))
119905 = 119899119879 119899 isinN
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579≐ Θ1(119878 (119905) 119868 (119905))
119868 (119905+
) = 119868 (119905) ≐ Θ2(119878 (119905) 119868 (119905))
119905 = 119899119879 119899 isinN
(24)According to the bifurcation theorem in [34] we can
obtain the following result
Theorem 3 If there exists a 1198790gt 0 satisfying 1198771
0(1198790) = 1 and
all the parameters in model (24) satisfy
120573 (119878 (1198790) minus 1) eminus1198871198790 (
11988711987901198603
1 minus 1198603eminus1198871198790
minus 1) lt 120573 minus 119903 minus 119887 (25)
where 1198603= 1 minus 119901max + ((120579
2119901max)(120579 + 119878
lowast(1198790))2
) then thereexists a supercritical branch at the point 119879
0
8 Abstract and Applied Analysis
5 10 15 20002
004
006
008
01
T
S(t)
(a)
2 4 6 8 10004
005
006
007
008
T
S(t)
(b)
5 6 7 8 9 10005
0055
006
0065
007
T
S(t)
(c)
7 75 8005
0055
006
0065
007
T
S(t)
(d)
Figure 4 Bifurcation diagrams of model (4) with respect to bifurcation parameter 119879 Here 119901max = 075 and all other parameter values aregiven in Table 1 (a) 120579 = 0 which are reproduced from [9] (b) 120579 = 02 (c and d) a magnified part of Figure 4(b)
The existence of 1198790has been indicated in Figure 1 and the
proof ofTheorem 3 is given in Appendix BTheorem 3 showsthat there exists an endemic periodic solution under someconditions providing that the disease-free periodic solutionbecomes unstable It follows from Theorem 3 that if 119879 gt 119879
0
and is close to 1198790 the endemic periodic solution of model (4)
is stableTo investigate the complex dynamics that model (4) can
have we chose the pulse period 119879 as a bifurcation parameterand fixed all other parameters as those in Figure 4 for twodifferent 120579 values Figure 4(a) is a bifurcation diagram with-out resource limitation that is 120579 = 0 which was obtainedby Shulgin et al [9] Figure 4(b) is a bifurcation diagramwith resource limitation that is 120579 = 02 Comparing thosetwo bifurcation diagrams we conclude that the nonlinearpulse vaccination can produce more complex dynamics thanthose for the model with linear pulse vaccination Once thepulse period 119879 exceeds 119879120579max defined in (22) both susceptibleand infected populations can oscillate periodically with alarge amplitude that corresponds to periodic outbreaks ofepidemics As the pulse period is further increased a series ofcomplex and interesting bifurcation phenomena are observed(see Figures 4(a) and 4(b)) Figures 4(b)ndash4(d) indicate thatthe dynamical behavior of model (4) is very complexincluding period doubling bifurcation chaotic attractors
multistability periodic-adding chaos crisis and periodicwindowsMeanwhile bifurcation analyses (ie Figures 4(b)ndash4(d)) also indicate that the model (4) has several differentattractors which can coexist for a wide range of parametersFor example Figure 5 provides an example of two attractorcoexistence when 119879 = 78 Multiple attractor coexistenceindicates that the infectious outbreak patterns depend onthe initial values which may cause difficulties for infectiousdisease control
5 Discussion and Biological Conclusions
In order to understand the effect of resource limitation inparticular lack of vaccine on the transmission of infectiousdisease we deliberately investigated the dynamical behaviorof an SIR epidemic model which incorporates a nonlinearpulse vaccination strategy To this end we introduced anonlinear form 119901(119905) = (119901max119878(119905))(119878(119905) + 120579) as vaccinationproportionThis resulted in interesting and dramatic changesin the dynamical behavior of solutions and they becamemoreand more complicated which means that it is a very difficulttask to control infectious diseases in particular emerginginfectious diseases under resource limitation We discussedthe control strategy based on the threshold value in moredetail through theoretical analysis numerical studies and
Abstract and Applied Analysis 9
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(a)
100 110 120 130 140 150minus13
minus12
minus11
minus10
minus9
minus8
minus7
t
Log(I(t)
)
(b)
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(c)
100 110 120 130 140 150minus16
minus14
minus12
minus10
minus8
minus6
t
Log(I(t)
)
(d)
Figure 5 Coexistence of two stable attractors of model (4) Here 119901max = 075 120579 = 02 119879 = 78 and all other parameter values are given inTable 1 (a and b) The initial value (119878
0 1198680) = (012 005) (c and d) the initial value (119878
0 1198680) = (01 002)
sensitivity analysis As pointed out byAgur et al [7] the costsrisks and the efficacy of the pulse vaccinations are key factorsin eliminating transmission of the measles virus In orderto depict those factors the nonlinear resource limitationsshould be taken into account [9] and the main purpose ofthe present work is to formulate the disease model to addressthe effects of resource limitations on disease control
The results indicate that vaccination as an approach tocontrolling epidemics must be committed to a long-termstrategy especially when resources arelimited To understandthe interactions amongst 119878(119905) and 119868(119905) which are importantfor successful control of the spread of infectious diseasewe have proposed detailed modeling methods involvingvaccination on 119878(119905) and determining the most significantparameters for the basic reproductive number 1198772
0by using
LHSPRCC uncertainty and sensitivity analysis techniques(see Figure 3) Our results indicate that we should paymore attention to the vaccination period and vaccinationproportion which reduce the threshold value 1198772
0or 11987710to
prevent the outbreak of disease Our results clarify that itis more significant to improve immunization programs (ievaccinations as mentioned in [15 16]) under conditions ofresource limitation
The results also indicate that the dynamic behavior ofmodel (4) may be dramatically affected by small changes in
the value of initial densities of susceptible and infected withresource limitation Bifurcation diagrams shown in Figure 4indicate that there are many forms of complexities in model(4) which are related to chaotic bands with periodic win-dows and attractor crises and Figure 5 reveals that theoccurrence of multiple attractors is a common property ofthe SIR model with resource limitation which can help usto further understand the application of a nonlinear pulsevaccination strategy in an SIR model (for more informationabout bifurcation diagrams of impulsive control strategiessee [35 36]) Some complexities are related to the long-termbehavior of population dynamics characterized either bywell behaving relatively simple or very complicated strangeattractors We also found that the routes to chaos are verycomplicated For instance with resource limitation there areseveral hidden factors that can adversely affect the controlstrategy The increasing number of potential complexitiespredicted by the theory does not seem to make this taskany easier Nevertheless identifying complicated possiblychaotic dynamics in population data has remained a majorchallenge in epidemiology studies In particular the sug-gestion that the dynamics of measles are chaotic has beenmuch debated (eg see [37]) and when seeking to establishwhether measles dynamics in New York City were chaoticor not Sugihara and May [38] omitted data from after 1963when vaccinations were introduced as the immunisations
10 Abstract and Applied Analysis
had altered the intrinsic dynamics However our resultsimply that even with immunisations disease dynamics couldbe chaotic under some circumstances and so a reanalysis ofthe New York data including the vaccination period might beworthwhile
In the present paper we have focused on studyingthe effects of dynamical behaviors on the SIR model withnonlinear pulse vaccination For our simple model we haveassumed that the maximal vaccination proportion 119901max isa constant and the pulse period 119879 can be varied Howeverdue to limited vaccine availability the maximal vaccinationproportion 119901max should depend on the vaccine obtained ineach pulse period Thus to describe the impact of a limitedvaccine stockpile a separate differential equation for theamount of vaccine denoted by 119881(119905) could be included inthe model and then 119901(119905) should be a function of 119881(119905) and119878(119905) Meanwhile it is essential to link the costs of developingand implementing controls to population dynamic modelingof disease epidemics in order to consider other resourcelimitations such as quantities of drugs availability of isolationunits numbers of hospital beds and equipmentThese topicswill be considered in further work in the future
Appendices
A Proof of Theorem 2
Firstly we prove the local stability of a periodic solution(119878lowast(119905) 0) which may be determined by considering the
behavior of small amplitude perturbations of the solutionDefine
119878 (119905) = 119880 (119905) + 119878lowast
(119905) 119868 (119905) = 119881 (119905) (A1)
here119880(119905) 119881(119905) are small perturbations whichmay be writtenas
(119880 (119905)
119881 (119905)) = Φ (119905) (
119880 (0)
119881 (0)) 0 le 119905 lt 119879 (A2)
where the fundamental matrixΦ(119905) satisfies
dΦ (119905)d119905
= (minus119887 minus120573119878
lowast
(119905)
0 120573119878lowast
(119905) minus 120574 minus 119887)Φ (119905) (A3)
and Φ(0) = 119868 is the identity matrix So
Φ (119879) = (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 ) (A4)
where 995333 is not necessarily computed in detail as it is notrequired in the following analysis
The resetting impulsive conditions of model (4) become
(119880 (119899119879
+)
119881 (119899119879+)) = (
1 minus 119901max +1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)(119880 (119899119879)
119881 (119899119879))
≐ 119861 (119899119879) (119880 (119899119879)
119881 (119899119879))
(A5)
Hence according to Floquet theory if the moduli of botheigenvalues of the matrix
119872 = 119861 (119879)Φ (119879)
= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)
times (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 )
(A6)
are less than one then the periodic solution (119878lowast(119905) 0) islocally stable In fact two Floquet expressions multiplied arethus
1205821= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119879))2) eminus119887119879
1205822= eint
119879
0(120573119878lowast(119905)minus120574minus119887)d119905
(A7)
It is obvious that 0 lt 1205821lt 1 Thus the stability of (119878lowast(119905) 0)
is decided by whether 1205822lt 1 or not So we conclude that
the disease-free periodic solution (119878lowast(119905) 0) is locally stable ifint119879
0(120573119878lowast(119905) minus 120574 minus 119887)d119905 lt 0 that is
1
119879int
119879
0
119878lowast
(119905) d119905 =(119878 minus 1) (1 minus eminus119887119879) + 119887119879
119887119879lt120574 + 119887
120573 (A8)
and by simple calculation we have
120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]119887119879 (120574 + 119887)
lt 1 (A9)
which holds true due to 11987710lt 1 Therefore we can draw
a conclusion from the above analysis that if 11987710lt 1 the
disease-free periodic solution (119878lowast(119905) 0) of model (4) is locallyasymptotically stable
Next we will prove the global attraction of the disease-free periodic solution (119878lowast(119905) 0) of model (4) Since 1198771
0lt 1
we can choose 1205761gt 0 sufficiently small such that
1
119879int
119879
0
(119878lowast
(119905) + 1205761) d119905 lt
120574 + 119887
120573 (A10)
It follows from model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) 119905 = 119899119879
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579 119905 = 119899119879
(A11)
Consider the comparison equation
d119885 (119905)d119905
= 119887 minus 119887119885 (119905) 119905 = 119899119879
119885 (119905+
) =(1 minus 119901max) 119885
2
(119905) + 120579119885 (119905)
119885 (119905) + 120579 119905 = 119899119879
(A12)
Abstract and Applied Analysis 11
From Lemma 1 and the comparison theorem on impul-sive differential equations we have 119878(119905) le 119885(119905) and 119885(119905) rarr119878lowast(119905) as 119905 rarr +infin Hence there exists a 119905
1gt 0 such that
119878 (119905) le 119885 (119905) lt 119878lowast
(119905) + 1205761 (A13)
for all 119905 ge 1199051 here 120576
1gt 0 is small enough
From the second equation of model (4) and (A13) we get
d119868 (119905)d119905
le (120573 (119878lowast
(119905) + 1205761) minus 120574 minus 119887) 119868 (119905) (A14)
for 119905 gt 1199051 then we consider the following comparison
equation with pulses
d1199101(119905)
d119905= (120573 (119878
lowast
(119905) + 1205761) minus 120574 minus 119887) 119910
1(119905) 119905 = 119899119879
1199101(119905+
) = 1199101(119905) 119905 = 119899119879
(A15)
Integrating model (A15) between pulses (119899119879 (119899 + 1)119879] wehave
1199101((119899 + 1) 119879) = 119910
1(119899119879) eint
(119899+1)119879
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101(119899119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A16)
Then by using step by step iterations
1199101(119899119879) = 119910
1((119899 minus 1) 119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101((119899 minus 2) 119879) e2 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= sdot sdot sdot
= 1199101(0) e119899 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A17)
where 1199101(0) = 119910
1(0+) gt 0 we obtain lim
119899rarrinfin1199101(119899119879) =
0 according to (A10) On the other hand integrating andsolving the first equation of model (A15) between pulses itgives
1199101(119905) = 119910
1(119899119879) eint
119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905 119905 isin (119899119879 (119899 + 1) 119879]
(A18)
incorporating into the boundedness of eint119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
due to 11987710lt 1 we have lim
119905rarrinfin1199101(119905) = 0
Let (119878(119905) 119868(119905)) be any solution of model (4) with initialvalue (119878
0 1198680) and 119878
0= 119878(0
+) gt 0 119868
0= 119868(0
+) =
1199101(0) gt 0 according to the comparison theorem we have
lim119905rarrinfin
sup 119868(119905) le lim119905rarrinfin
sup1199101(119905) = 0 Incorporating into
the positivity of 119868(119905) we know that lim119905rarrinfin
119868(119905) = 0Consequently there exists a 119905
2gt 1199051such that 0 lt 119868(119905) le 120576
2
for 119905 ge 1199052 and 120576
2is small enough Further we have
119887 minus (1205731205762+ 119887) 119878 (119905) le
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (A19)
for all 119905 gt 1199052 from which we can obtain the following
equation
d1199102(119905)
d119905= 119887 minus (120573120576
2+ 119887) 119910
2(119905) 119905 = 119899119879
1199102(119905+
) =(1 minus 119901max) 119910
2
2(119905) + 120579119910
2(119905)
1199102(119905) + 120579
119905 = 119899119879
(A20)
By employing the same methods as used for the proof ofLemma 1 we get that the model (A20) has a positive periodicsolution 119910lowast
2(119905) which is globally attractive where
119910lowast
2(119905) = (119910
2minus
119887
1205731205762+ 119887) eminus(1205731205762+119887)(119905minus119899119879) + 119887
1205731205762+ 119887
119905 isin (119899119879 (119899 + 1) 119879]
1199102= 1199102(119899119879+
) = 119910lowast
2(0+
) =minus1198612+ radic11986122minus 411986021198622
21198602
1198602= eminus(1205731205762+119887)119879 [ 119887
1205731205762+ 119887
minus (1 minus 119901max) eminus(1205731205762+119887)119879]
1198612= (eminus(1205731205762+119887)119879 minus 119887
1205731205762+ 119887)
times [2 (1 minus 119901max) eminus(1205731205762+119887)119879minus (120579 +
119887
1205731205762+ 119887)]
1198622= minus[(1 minus 119901max) (
119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)2
+ 120579(119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)]
(A21)
It follows from the comparison theorem on impulsive differ-ential equations that 119910
2(119905) le 119878(119905) le 119885(119905) Moreover 119910
2(119905) rarr
119910lowast
2(119905) and 119885(119905) rarr 119878
lowast(119905) as 119905 rarr +infin Consequently there
exists a 1199053for 1205763small enough such that 119905
3ge 1199052and
119910lowast
2(119905) minus 120576
3lt 119878 (119905) lt 119878
lowast
(119905) + 1205763
(A22)
for 119905 ge 1199053 Let 120576
2rarr 0 Then
119878lowast
(119905) minus 1205763lt 119878 (119905) lt 119878
lowast
(119905) + 1205763 (A23)
Therefore 119878(119905) rarr 119878lowast(119905) as 119905 rarr +infin Thus we
have proved the global stability of the disease-free periodicsolution (119878lowast(119905) 0) of model (4) This completes the proof
B Proof of Theorem 3
In order to apply the bifurcation theory of [34] in the maintext we make the following calculations
1198891015840
0= 1 minus eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
(B1)
If 11988910158400= 0 then 119879
0satisfies the following equation
120573 [(119878 minus 1) (1 minus eminus1198871198790) + 1198871198790]
119887 (120574 + 119887) 1198790
= 1 (B2)
12 Abstract and Applied Analysis
which indicates that there exists a1198790such that 1198771
0= 1 and the
disease-free periodic solution 120575 = (119878lowast(119905) 0) loses its stabilityFurther
120597Φ1(1198790 1198830)
120597119878= eminus1198871198790 gt 0
120597Φ2(1198790 1198830)
120597119868= eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
120597Φ1(1198790 1198830)
120597119868= minusint
1198790
0
120573119878lowast
(V) eminus119887(1198790minusV)eintV
0(120573119878lowast(119905)minus120574minus119887)d119905dV
≐ 119860 lt 0
1198861015840
0= 1 minus 119860
3eminus1198871198790
1198871015840
0= 1 minus 119860
3119860
1205972Φ2(1198790 1198830)
120597119868120597119878= 1205731198790eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
1205972Φ2(1198790 1198830)
1205971198682
= minus1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
119878lowast
(120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585minus119887(Vminus120579)
d120579 dV
lt 0
1205972Φ2(1198790 1198830)
120597119868120597119879= (120573119878
lowast
(1198790) minus 120574 minus 119887) eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
120597Φ1(1198790 1198830)
120597119879= minus119887 (119878 minus 1) eminus1198871198790
119861 = 1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887)
times eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
119862 = 1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
eminus119887(119905minus120579)119878lowast (120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585
d120579 dV
+ 21205731198790
1 minus 1198603119860
1 minus 1198603eminus1198871198790
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
(B3)
It follows that 119861119862 lt 0 and we get the condition that is if theparameters satisfy condition 119861 lt 0 then the model (4) has asupercritical branch at 119879
0
In fact
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0 (B4)
Therefore 119861 lt 0 is equivalent to
1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887) lt 0
(B5)
Obviously 119861 lt 0 due to (25) The proof is completed
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thiswork is supported by theNational Natural Science Foun-dation of China (NSFCs 11171199 11371030 11301320) andthe Fundamental Research Funds for the Central Universities(GK201305010) The authors would like to thank the editorand the referees for their careful reading of the original paperand many valuable comments and suggestions that greatlyimproved the presentation of this paper
References
[1] R M Anderson and R M May ldquoPopulation biology of infec-tious diseases part Irdquo Nature vol 280 no 5721 pp 361ndash3671979
[2] E Hanert E Schumacher and E Deleersnijder ldquoFront dynam-ics in fractional-order epidemic modelsrdquo Journal of TheoreticalBiology vol 279 no 1 pp 9ndash16 2011
[3] H W Hethcote ldquoMathematics of infectious diseasesrdquo SIAMReview vol 42 no 4 pp 599ndash653 2000
[4] A E Fiore C B Bridges and N J Cox ldquoSeasonal influenzavaccinesrdquo Current Topics in Microbiology and Immunology vol333 no 1 pp 43ndash82 2009
[5] Y Chang N T Brewer A C Rinas K Schmitt and J S SmithldquoEvaluating the impact of human papillomavirus vaccinesrdquoVaccine vol 27 no 32 pp 4355ndash4362 2009
[6] T J Liesegang ldquoVaricella zoster virus vaccines effective butconcerns lingerrdquo Canadian Journal of Ophthalmology vol 44no 4 pp 379ndash384 2009
[7] Z Agur L Cojocaru G Mazor R M Anderson and Y LDanon ldquoPulse mass measles vaccination across age cohortsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 90 no 24 pp 11698ndash11702 1993
[8] L Stone B Shulgin and Z Agur ldquoTheoretical examinationof the pulse vaccination policy in the SIR epidemic modelrdquoMathematical and ComputerModelling vol 31 no 4-5 pp 207ndash215 2000
[9] B Shulgin L Stone and Z Agur ldquoPulse vaccination strategy inthe SIR epidemic modelrdquo Bulletin of Mathematical Biology vol60 no 6 pp 1123ndash1148 1998
[10] A DrsquoOnofrio ldquoStability properties of pulse vaccination strategyin SEIR epidemicmodelrdquoMathematical Biosciences vol 179 no1 pp 57ndash72 2002
[11] A Franceschetti and A Pugliese ldquoThreshold behaviour of a SIRepidemicmodel with age structure and immigrationrdquo Journal ofMathematical Biology vol 57 no 1 pp 1ndash27 2008
Abstract and Applied Analysis 13
[12] A J Terry ldquoPulse vaccination strategies in a metapopulationSIR modelrdquo Mathematical Biosciences and Engineering vol 7no 2 pp 455ndash477 2010
[13] S J Gao L S Chen J J Nieto and A Torres ldquoAnalysis of adelayed epidemic model with pulse vaccination and saturationincidencerdquo Vaccine vol 24 no 35-36 pp 6037ndash6045 2006
[14] L Matrajt M E Halloran and I M Longini Jr ldquoOptimal vac-cine allocation for the early mitigation of pandemic influenzardquoPLoS Computational Biology vol 9 no 3 Article ID e10029642013
[15] S P Sullivan D G Koutsonanos M D P Martin et alldquoDissolving polymer microneedle patches for influenza vacci-nationrdquo Nature Medicine vol 16 no 8 pp 915ndash920 2010
[16] Science Daily Web ldquoVaccine-delivery patch with dissolvingmicroneedles eliminates ldquosharpsrdquo boosts protectionrdquo httpwwwsciencedailycomreleases201007100718204733htm
[17] M J Reynolds-Hogland J S Hogland and M S MitchellldquoEvaluating intercepts fromdemographicmodels to understandresource limitation and resource thresholdsrdquo Ecological Mod-elling vol 211 no 3-4 pp 424ndash432 2008
[18] L Chow M Fan and Z L Feng ldquoDynamics of a multigroupepidemiological model with group-targeted vaccination strate-giesrdquo Journal of Theoretical Biology vol 291 no 1 pp 56ndash642011
[19] L H Zhou and M Fan ldquoDynamics of an SIR epidemic modelwith limited medical resources revisitedrdquo Nonlinear AnalysisReal World Applications vol 13 no 1 pp 312ndash324 2012
[20] V Lakshmikantham D D Bainov and P S Simeonov Theoryof Impulsive Differential Equations World Scientific Singapore1989
[21] S M Blower and H Dowlatabadi ldquoSensitivity and uncertaintyanalysis of complex-models of disease transmission An HIVmodel as an examplerdquo International Statistical Review vol 62no 2 pp 229ndash243 1994
[22] S Marino I B Hogue C J Ray and D E Kirschner ldquoA meth-odology for performing global uncertainty and sensitivityanalysis in systems biologyrdquo Journal of Theoretical Biology vol254 no 1 pp 178ndash196 2008
[23] S Y Tang J H Liang Y S Tan and R A Cheke ldquoThresholdconditions for integrated pest management models with pesti-cides that have residual effectsrdquo Journal ofMathematical Biologyvol 66 no 1-2 pp 1ndash35 2013
[24] F Brauer and C Castillo-ChavezMathematical Models in Pop-ulation Biology and Epidemiology Springer New York NYUSA 2000
[25] O Diekmann and J A P Heesterbeek Mathematical Epidemi-ology of Infectious Diseases JohnWiley amp Sons Chichester UK2000
[26] J D MurrayMathematical Biology Springer Berlin Germany1989
[27] R Anderson and R May Infectious Diseases of HumansDynamics and Control Oxford University Press Oxford UK1995
[28] M Hethcote ldquoThree basic epidemiological modelsrdquo in AppliedMathematical Ecology L Gross T G Hallam and S A LevinEds pp 119ndash144 Springer Berlin Germany 1989
[29] A V Hill ldquoThe possible effects of the aggregation of themolecules of haemoglobin on its dissociation curvesrdquo TheJournal of Physiology vol 40 no 4 pp 4ndash7 1910
[30] P Cull ldquoLocal and global stability for population modelsrdquoBiological Cybernetics vol 54 no 3 pp 141ndash149 1986
[31] WHOWeb ldquoGlobal health observatory data repositoryrdquo 2001httpappswhointghodatatheme=country
[32] M D McKay R J Beckman and W J Conover ldquoComparisonof three methods for selecting values of input variables in theanalysis of output from a computer coderdquo Technometrics vol21 no 2 pp 239ndash245 1979
[33] W M Schaffer L F Olsen G L Truty S L Fulmer andD J Graser ldquoPeriodic and chaotic dynamics in childhoodepidemicsrdquo in From Chemical to Biological Organization MMarkus S Muller and G Nicolis Eds Springer BerlinGermany 1988
[34] A Lakmeche and O Arino ldquoBifurcation of non trivial periodicsolutions of impulsive differential equations arising chemother-apeutic treatmentrdquo Dynamics of Continuous Discrete andImpulsive Systems B vol 7 no 2 pp 265ndash287 2000
[35] S Y Tang and L S Chen ldquoMultiple attractors in stage-structured population models with birth pulsesrdquo Bulletin ofMathematical Biology vol 65 no 3 pp 479ndash495 2003
[36] S Y Tang Y N Xiao and R A Cheke ldquoMultiple attractorsof host-parasitoid models with integrated pest managementstrategies eradication persistence and outbreakrdquo TheoreticalPopulation Biology vol 73 no 2 pp 181ndash197 2008
[37] L F Olsen andWM Schaffer ldquoChaos versus noisy periodicityalternative hypotheses for childhood epidemicsrdquo Science vol249 no 4968 pp 499ndash504 1990
[38] G Sugihara and R M May ldquoNonlinear forecasting as a way ofdistinguishing chaos from measurement error in time seriesrdquoNature vol 344 no 6268 pp 734ndash741 1990
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical Problems in Engineering
Hindawi Publishing Corporationhttpwwwhindawicom
Differential EquationsInternational Journal of
Volume 2014
Applied MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Probability and StatisticsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical PhysicsAdvances in
Complex AnalysisJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
OptimizationJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CombinatoricsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Operations ResearchAdvances in
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Function Spaces
Abstract and Applied AnalysisHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of Mathematics and Mathematical Sciences
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Algebra
Discrete Dynamics in Nature and Society
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Decision SciencesAdvances in
Discrete MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014 Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Stochastic AnalysisInternational Journal of

Abstract and Applied Analysis 7
minus1
minus05
0
05
1
b 120573 120574 120579 T
PRCC
s
pmax
(a)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [072346 0]
b
R2 0
(b)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [05492 0]
120573
R2 0
(c)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000
120574
R2 0
[minus012283 14766e minus 011]
(d)
minus2000 0 2000minus2000
minus1000
0
1000
2000 [minus071273 0]
R2 0
pmax
(e)
minus2000 minus1000 0 1000minus2000
minus1000
0
1000
2000 [004117 0024135]
R2 0
120579
(f)
minus2000 0 2000minus2000
minus1000
0
1000
2000 [091801 0]
T
R2 0
(g)
Figure 3 PRCC results and PRCC scatter plots of the baseline parameters as those shown in Table 1 Here we fix 119901max = 075 120579 = 02 119879 = 5(a) PRCC results (bndashg) PRCC scatter plots The title of each plot represents the PRCC value with the corresponding P value
endemic periodic solution that is to investigate under whatconditions will the infected population oscillate periodicallywith small amplitude and then we choose the period ofpulse vaccination as a bifurcation parameter to investigate thecomplex patterns of the infectious disease dynamics
Next we analyze the existence of an endemic periodicsolution of model (4) near the disease-free periodic solution(119878lowast(119905) 0) using bifurcation theory [34] and set the impulsive
period 119879 as bifurcation parameter For ease of narrative werewrite the model (4) as
d119878 (119905)d119905
= 119887 minus 120573119878 (119905) 119868 (119905) minus 119887119878 (119905) ≐ 1198651(119878 (119905) 119868 (119905))
d119868 (119905)d119905
= 120573119878 (119905) 119868 (119905) minus 120574119868 (119905) minus 119887119868 (119905) ≐ 1198652(119878 (119905) 119868 (119905))
119905 = 119899119879 119899 isinN
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579≐ Θ1(119878 (119905) 119868 (119905))
119868 (119905+
) = 119868 (119905) ≐ Θ2(119878 (119905) 119868 (119905))
119905 = 119899119879 119899 isinN
(24)According to the bifurcation theorem in [34] we can
obtain the following result
Theorem 3 If there exists a 1198790gt 0 satisfying 1198771
0(1198790) = 1 and
all the parameters in model (24) satisfy
120573 (119878 (1198790) minus 1) eminus1198871198790 (
11988711987901198603
1 minus 1198603eminus1198871198790
minus 1) lt 120573 minus 119903 minus 119887 (25)
where 1198603= 1 minus 119901max + ((120579
2119901max)(120579 + 119878
lowast(1198790))2
) then thereexists a supercritical branch at the point 119879
0
8 Abstract and Applied Analysis
5 10 15 20002
004
006
008
01
T
S(t)
(a)
2 4 6 8 10004
005
006
007
008
T
S(t)
(b)
5 6 7 8 9 10005
0055
006
0065
007
T
S(t)
(c)
7 75 8005
0055
006
0065
007
T
S(t)
(d)
Figure 4 Bifurcation diagrams of model (4) with respect to bifurcation parameter 119879 Here 119901max = 075 and all other parameter values aregiven in Table 1 (a) 120579 = 0 which are reproduced from [9] (b) 120579 = 02 (c and d) a magnified part of Figure 4(b)
The existence of 1198790has been indicated in Figure 1 and the
proof ofTheorem 3 is given in Appendix BTheorem 3 showsthat there exists an endemic periodic solution under someconditions providing that the disease-free periodic solutionbecomes unstable It follows from Theorem 3 that if 119879 gt 119879
0
and is close to 1198790 the endemic periodic solution of model (4)
is stableTo investigate the complex dynamics that model (4) can
have we chose the pulse period 119879 as a bifurcation parameterand fixed all other parameters as those in Figure 4 for twodifferent 120579 values Figure 4(a) is a bifurcation diagram with-out resource limitation that is 120579 = 0 which was obtainedby Shulgin et al [9] Figure 4(b) is a bifurcation diagramwith resource limitation that is 120579 = 02 Comparing thosetwo bifurcation diagrams we conclude that the nonlinearpulse vaccination can produce more complex dynamics thanthose for the model with linear pulse vaccination Once thepulse period 119879 exceeds 119879120579max defined in (22) both susceptibleand infected populations can oscillate periodically with alarge amplitude that corresponds to periodic outbreaks ofepidemics As the pulse period is further increased a series ofcomplex and interesting bifurcation phenomena are observed(see Figures 4(a) and 4(b)) Figures 4(b)ndash4(d) indicate thatthe dynamical behavior of model (4) is very complexincluding period doubling bifurcation chaotic attractors
multistability periodic-adding chaos crisis and periodicwindowsMeanwhile bifurcation analyses (ie Figures 4(b)ndash4(d)) also indicate that the model (4) has several differentattractors which can coexist for a wide range of parametersFor example Figure 5 provides an example of two attractorcoexistence when 119879 = 78 Multiple attractor coexistenceindicates that the infectious outbreak patterns depend onthe initial values which may cause difficulties for infectiousdisease control
5 Discussion and Biological Conclusions
In order to understand the effect of resource limitation inparticular lack of vaccine on the transmission of infectiousdisease we deliberately investigated the dynamical behaviorof an SIR epidemic model which incorporates a nonlinearpulse vaccination strategy To this end we introduced anonlinear form 119901(119905) = (119901max119878(119905))(119878(119905) + 120579) as vaccinationproportionThis resulted in interesting and dramatic changesin the dynamical behavior of solutions and they becamemoreand more complicated which means that it is a very difficulttask to control infectious diseases in particular emerginginfectious diseases under resource limitation We discussedthe control strategy based on the threshold value in moredetail through theoretical analysis numerical studies and
Abstract and Applied Analysis 9
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(a)
100 110 120 130 140 150minus13
minus12
minus11
minus10
minus9
minus8
minus7
t
Log(I(t)
)
(b)
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(c)
100 110 120 130 140 150minus16
minus14
minus12
minus10
minus8
minus6
t
Log(I(t)
)
(d)
Figure 5 Coexistence of two stable attractors of model (4) Here 119901max = 075 120579 = 02 119879 = 78 and all other parameter values are given inTable 1 (a and b) The initial value (119878
0 1198680) = (012 005) (c and d) the initial value (119878
0 1198680) = (01 002)
sensitivity analysis As pointed out byAgur et al [7] the costsrisks and the efficacy of the pulse vaccinations are key factorsin eliminating transmission of the measles virus In orderto depict those factors the nonlinear resource limitationsshould be taken into account [9] and the main purpose ofthe present work is to formulate the disease model to addressthe effects of resource limitations on disease control
The results indicate that vaccination as an approach tocontrolling epidemics must be committed to a long-termstrategy especially when resources arelimited To understandthe interactions amongst 119878(119905) and 119868(119905) which are importantfor successful control of the spread of infectious diseasewe have proposed detailed modeling methods involvingvaccination on 119878(119905) and determining the most significantparameters for the basic reproductive number 1198772
0by using
LHSPRCC uncertainty and sensitivity analysis techniques(see Figure 3) Our results indicate that we should paymore attention to the vaccination period and vaccinationproportion which reduce the threshold value 1198772
0or 11987710to
prevent the outbreak of disease Our results clarify that itis more significant to improve immunization programs (ievaccinations as mentioned in [15 16]) under conditions ofresource limitation
The results also indicate that the dynamic behavior ofmodel (4) may be dramatically affected by small changes in
the value of initial densities of susceptible and infected withresource limitation Bifurcation diagrams shown in Figure 4indicate that there are many forms of complexities in model(4) which are related to chaotic bands with periodic win-dows and attractor crises and Figure 5 reveals that theoccurrence of multiple attractors is a common property ofthe SIR model with resource limitation which can help usto further understand the application of a nonlinear pulsevaccination strategy in an SIR model (for more informationabout bifurcation diagrams of impulsive control strategiessee [35 36]) Some complexities are related to the long-termbehavior of population dynamics characterized either bywell behaving relatively simple or very complicated strangeattractors We also found that the routes to chaos are verycomplicated For instance with resource limitation there areseveral hidden factors that can adversely affect the controlstrategy The increasing number of potential complexitiespredicted by the theory does not seem to make this taskany easier Nevertheless identifying complicated possiblychaotic dynamics in population data has remained a majorchallenge in epidemiology studies In particular the sug-gestion that the dynamics of measles are chaotic has beenmuch debated (eg see [37]) and when seeking to establishwhether measles dynamics in New York City were chaoticor not Sugihara and May [38] omitted data from after 1963when vaccinations were introduced as the immunisations
10 Abstract and Applied Analysis
had altered the intrinsic dynamics However our resultsimply that even with immunisations disease dynamics couldbe chaotic under some circumstances and so a reanalysis ofthe New York data including the vaccination period might beworthwhile
In the present paper we have focused on studyingthe effects of dynamical behaviors on the SIR model withnonlinear pulse vaccination For our simple model we haveassumed that the maximal vaccination proportion 119901max isa constant and the pulse period 119879 can be varied Howeverdue to limited vaccine availability the maximal vaccinationproportion 119901max should depend on the vaccine obtained ineach pulse period Thus to describe the impact of a limitedvaccine stockpile a separate differential equation for theamount of vaccine denoted by 119881(119905) could be included inthe model and then 119901(119905) should be a function of 119881(119905) and119878(119905) Meanwhile it is essential to link the costs of developingand implementing controls to population dynamic modelingof disease epidemics in order to consider other resourcelimitations such as quantities of drugs availability of isolationunits numbers of hospital beds and equipmentThese topicswill be considered in further work in the future
Appendices
A Proof of Theorem 2
Firstly we prove the local stability of a periodic solution(119878lowast(119905) 0) which may be determined by considering the
behavior of small amplitude perturbations of the solutionDefine
119878 (119905) = 119880 (119905) + 119878lowast
(119905) 119868 (119905) = 119881 (119905) (A1)
here119880(119905) 119881(119905) are small perturbations whichmay be writtenas
(119880 (119905)
119881 (119905)) = Φ (119905) (
119880 (0)
119881 (0)) 0 le 119905 lt 119879 (A2)
where the fundamental matrixΦ(119905) satisfies
dΦ (119905)d119905
= (minus119887 minus120573119878
lowast
(119905)
0 120573119878lowast
(119905) minus 120574 minus 119887)Φ (119905) (A3)
and Φ(0) = 119868 is the identity matrix So
Φ (119879) = (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 ) (A4)
where 995333 is not necessarily computed in detail as it is notrequired in the following analysis
The resetting impulsive conditions of model (4) become
(119880 (119899119879
+)
119881 (119899119879+)) = (
1 minus 119901max +1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)(119880 (119899119879)
119881 (119899119879))
≐ 119861 (119899119879) (119880 (119899119879)
119881 (119899119879))
(A5)
Hence according to Floquet theory if the moduli of botheigenvalues of the matrix
119872 = 119861 (119879)Φ (119879)
= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)
times (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 )
(A6)
are less than one then the periodic solution (119878lowast(119905) 0) islocally stable In fact two Floquet expressions multiplied arethus
1205821= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119879))2) eminus119887119879
1205822= eint
119879
0(120573119878lowast(119905)minus120574minus119887)d119905
(A7)
It is obvious that 0 lt 1205821lt 1 Thus the stability of (119878lowast(119905) 0)
is decided by whether 1205822lt 1 or not So we conclude that
the disease-free periodic solution (119878lowast(119905) 0) is locally stable ifint119879
0(120573119878lowast(119905) minus 120574 minus 119887)d119905 lt 0 that is
1
119879int
119879
0
119878lowast
(119905) d119905 =(119878 minus 1) (1 minus eminus119887119879) + 119887119879
119887119879lt120574 + 119887
120573 (A8)
and by simple calculation we have
120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]119887119879 (120574 + 119887)
lt 1 (A9)
which holds true due to 11987710lt 1 Therefore we can draw
a conclusion from the above analysis that if 11987710lt 1 the
disease-free periodic solution (119878lowast(119905) 0) of model (4) is locallyasymptotically stable
Next we will prove the global attraction of the disease-free periodic solution (119878lowast(119905) 0) of model (4) Since 1198771
0lt 1
we can choose 1205761gt 0 sufficiently small such that
1
119879int
119879
0
(119878lowast
(119905) + 1205761) d119905 lt
120574 + 119887
120573 (A10)
It follows from model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) 119905 = 119899119879
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579 119905 = 119899119879
(A11)
Consider the comparison equation
d119885 (119905)d119905
= 119887 minus 119887119885 (119905) 119905 = 119899119879
119885 (119905+
) =(1 minus 119901max) 119885
2
(119905) + 120579119885 (119905)
119885 (119905) + 120579 119905 = 119899119879
(A12)
Abstract and Applied Analysis 11
From Lemma 1 and the comparison theorem on impul-sive differential equations we have 119878(119905) le 119885(119905) and 119885(119905) rarr119878lowast(119905) as 119905 rarr +infin Hence there exists a 119905
1gt 0 such that
119878 (119905) le 119885 (119905) lt 119878lowast
(119905) + 1205761 (A13)
for all 119905 ge 1199051 here 120576
1gt 0 is small enough
From the second equation of model (4) and (A13) we get
d119868 (119905)d119905
le (120573 (119878lowast
(119905) + 1205761) minus 120574 minus 119887) 119868 (119905) (A14)
for 119905 gt 1199051 then we consider the following comparison
equation with pulses
d1199101(119905)
d119905= (120573 (119878
lowast
(119905) + 1205761) minus 120574 minus 119887) 119910
1(119905) 119905 = 119899119879
1199101(119905+
) = 1199101(119905) 119905 = 119899119879
(A15)
Integrating model (A15) between pulses (119899119879 (119899 + 1)119879] wehave
1199101((119899 + 1) 119879) = 119910
1(119899119879) eint
(119899+1)119879
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101(119899119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A16)
Then by using step by step iterations
1199101(119899119879) = 119910
1((119899 minus 1) 119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101((119899 minus 2) 119879) e2 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= sdot sdot sdot
= 1199101(0) e119899 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A17)
where 1199101(0) = 119910
1(0+) gt 0 we obtain lim
119899rarrinfin1199101(119899119879) =
0 according to (A10) On the other hand integrating andsolving the first equation of model (A15) between pulses itgives
1199101(119905) = 119910
1(119899119879) eint
119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905 119905 isin (119899119879 (119899 + 1) 119879]
(A18)
incorporating into the boundedness of eint119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
due to 11987710lt 1 we have lim
119905rarrinfin1199101(119905) = 0
Let (119878(119905) 119868(119905)) be any solution of model (4) with initialvalue (119878
0 1198680) and 119878
0= 119878(0
+) gt 0 119868
0= 119868(0
+) =
1199101(0) gt 0 according to the comparison theorem we have
lim119905rarrinfin
sup 119868(119905) le lim119905rarrinfin
sup1199101(119905) = 0 Incorporating into
the positivity of 119868(119905) we know that lim119905rarrinfin
119868(119905) = 0Consequently there exists a 119905
2gt 1199051such that 0 lt 119868(119905) le 120576
2
for 119905 ge 1199052 and 120576
2is small enough Further we have
119887 minus (1205731205762+ 119887) 119878 (119905) le
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (A19)
for all 119905 gt 1199052 from which we can obtain the following
equation
d1199102(119905)
d119905= 119887 minus (120573120576
2+ 119887) 119910
2(119905) 119905 = 119899119879
1199102(119905+
) =(1 minus 119901max) 119910
2
2(119905) + 120579119910
2(119905)
1199102(119905) + 120579
119905 = 119899119879
(A20)
By employing the same methods as used for the proof ofLemma 1 we get that the model (A20) has a positive periodicsolution 119910lowast
2(119905) which is globally attractive where
119910lowast
2(119905) = (119910
2minus
119887
1205731205762+ 119887) eminus(1205731205762+119887)(119905minus119899119879) + 119887
1205731205762+ 119887
119905 isin (119899119879 (119899 + 1) 119879]
1199102= 1199102(119899119879+
) = 119910lowast
2(0+
) =minus1198612+ radic11986122minus 411986021198622
21198602
1198602= eminus(1205731205762+119887)119879 [ 119887
1205731205762+ 119887
minus (1 minus 119901max) eminus(1205731205762+119887)119879]
1198612= (eminus(1205731205762+119887)119879 minus 119887
1205731205762+ 119887)
times [2 (1 minus 119901max) eminus(1205731205762+119887)119879minus (120579 +
119887
1205731205762+ 119887)]
1198622= minus[(1 minus 119901max) (
119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)2
+ 120579(119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)]
(A21)
It follows from the comparison theorem on impulsive differ-ential equations that 119910
2(119905) le 119878(119905) le 119885(119905) Moreover 119910
2(119905) rarr
119910lowast
2(119905) and 119885(119905) rarr 119878
lowast(119905) as 119905 rarr +infin Consequently there
exists a 1199053for 1205763small enough such that 119905
3ge 1199052and
119910lowast
2(119905) minus 120576
3lt 119878 (119905) lt 119878
lowast
(119905) + 1205763
(A22)
for 119905 ge 1199053 Let 120576
2rarr 0 Then
119878lowast
(119905) minus 1205763lt 119878 (119905) lt 119878
lowast
(119905) + 1205763 (A23)
Therefore 119878(119905) rarr 119878lowast(119905) as 119905 rarr +infin Thus we
have proved the global stability of the disease-free periodicsolution (119878lowast(119905) 0) of model (4) This completes the proof
B Proof of Theorem 3
In order to apply the bifurcation theory of [34] in the maintext we make the following calculations
1198891015840
0= 1 minus eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
(B1)
If 11988910158400= 0 then 119879
0satisfies the following equation
120573 [(119878 minus 1) (1 minus eminus1198871198790) + 1198871198790]
119887 (120574 + 119887) 1198790
= 1 (B2)
12 Abstract and Applied Analysis
which indicates that there exists a1198790such that 1198771
0= 1 and the
disease-free periodic solution 120575 = (119878lowast(119905) 0) loses its stabilityFurther
120597Φ1(1198790 1198830)
120597119878= eminus1198871198790 gt 0
120597Φ2(1198790 1198830)
120597119868= eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
120597Φ1(1198790 1198830)
120597119868= minusint
1198790
0
120573119878lowast
(V) eminus119887(1198790minusV)eintV
0(120573119878lowast(119905)minus120574minus119887)d119905dV
≐ 119860 lt 0
1198861015840
0= 1 minus 119860
3eminus1198871198790
1198871015840
0= 1 minus 119860
3119860
1205972Φ2(1198790 1198830)
120597119868120597119878= 1205731198790eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
1205972Φ2(1198790 1198830)
1205971198682
= minus1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
119878lowast
(120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585minus119887(Vminus120579)
d120579 dV
lt 0
1205972Φ2(1198790 1198830)
120597119868120597119879= (120573119878
lowast
(1198790) minus 120574 minus 119887) eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
120597Φ1(1198790 1198830)
120597119879= minus119887 (119878 minus 1) eminus1198871198790
119861 = 1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887)
times eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
119862 = 1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
eminus119887(119905minus120579)119878lowast (120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585
d120579 dV
+ 21205731198790
1 minus 1198603119860
1 minus 1198603eminus1198871198790
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
(B3)
It follows that 119861119862 lt 0 and we get the condition that is if theparameters satisfy condition 119861 lt 0 then the model (4) has asupercritical branch at 119879
0
In fact
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0 (B4)
Therefore 119861 lt 0 is equivalent to
1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887) lt 0
(B5)
Obviously 119861 lt 0 due to (25) The proof is completed
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thiswork is supported by theNational Natural Science Foun-dation of China (NSFCs 11171199 11371030 11301320) andthe Fundamental Research Funds for the Central Universities(GK201305010) The authors would like to thank the editorand the referees for their careful reading of the original paperand many valuable comments and suggestions that greatlyimproved the presentation of this paper
References
[1] R M Anderson and R M May ldquoPopulation biology of infec-tious diseases part Irdquo Nature vol 280 no 5721 pp 361ndash3671979
[2] E Hanert E Schumacher and E Deleersnijder ldquoFront dynam-ics in fractional-order epidemic modelsrdquo Journal of TheoreticalBiology vol 279 no 1 pp 9ndash16 2011
[3] H W Hethcote ldquoMathematics of infectious diseasesrdquo SIAMReview vol 42 no 4 pp 599ndash653 2000
[4] A E Fiore C B Bridges and N J Cox ldquoSeasonal influenzavaccinesrdquo Current Topics in Microbiology and Immunology vol333 no 1 pp 43ndash82 2009
[5] Y Chang N T Brewer A C Rinas K Schmitt and J S SmithldquoEvaluating the impact of human papillomavirus vaccinesrdquoVaccine vol 27 no 32 pp 4355ndash4362 2009
[6] T J Liesegang ldquoVaricella zoster virus vaccines effective butconcerns lingerrdquo Canadian Journal of Ophthalmology vol 44no 4 pp 379ndash384 2009
[7] Z Agur L Cojocaru G Mazor R M Anderson and Y LDanon ldquoPulse mass measles vaccination across age cohortsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 90 no 24 pp 11698ndash11702 1993
[8] L Stone B Shulgin and Z Agur ldquoTheoretical examinationof the pulse vaccination policy in the SIR epidemic modelrdquoMathematical and ComputerModelling vol 31 no 4-5 pp 207ndash215 2000
[9] B Shulgin L Stone and Z Agur ldquoPulse vaccination strategy inthe SIR epidemic modelrdquo Bulletin of Mathematical Biology vol60 no 6 pp 1123ndash1148 1998
[10] A DrsquoOnofrio ldquoStability properties of pulse vaccination strategyin SEIR epidemicmodelrdquoMathematical Biosciences vol 179 no1 pp 57ndash72 2002
[11] A Franceschetti and A Pugliese ldquoThreshold behaviour of a SIRepidemicmodel with age structure and immigrationrdquo Journal ofMathematical Biology vol 57 no 1 pp 1ndash27 2008
Abstract and Applied Analysis 13
[12] A J Terry ldquoPulse vaccination strategies in a metapopulationSIR modelrdquo Mathematical Biosciences and Engineering vol 7no 2 pp 455ndash477 2010
[13] S J Gao L S Chen J J Nieto and A Torres ldquoAnalysis of adelayed epidemic model with pulse vaccination and saturationincidencerdquo Vaccine vol 24 no 35-36 pp 6037ndash6045 2006
[14] L Matrajt M E Halloran and I M Longini Jr ldquoOptimal vac-cine allocation for the early mitigation of pandemic influenzardquoPLoS Computational Biology vol 9 no 3 Article ID e10029642013
[15] S P Sullivan D G Koutsonanos M D P Martin et alldquoDissolving polymer microneedle patches for influenza vacci-nationrdquo Nature Medicine vol 16 no 8 pp 915ndash920 2010
[16] Science Daily Web ldquoVaccine-delivery patch with dissolvingmicroneedles eliminates ldquosharpsrdquo boosts protectionrdquo httpwwwsciencedailycomreleases201007100718204733htm
[17] M J Reynolds-Hogland J S Hogland and M S MitchellldquoEvaluating intercepts fromdemographicmodels to understandresource limitation and resource thresholdsrdquo Ecological Mod-elling vol 211 no 3-4 pp 424ndash432 2008
[18] L Chow M Fan and Z L Feng ldquoDynamics of a multigroupepidemiological model with group-targeted vaccination strate-giesrdquo Journal of Theoretical Biology vol 291 no 1 pp 56ndash642011
[19] L H Zhou and M Fan ldquoDynamics of an SIR epidemic modelwith limited medical resources revisitedrdquo Nonlinear AnalysisReal World Applications vol 13 no 1 pp 312ndash324 2012
[20] V Lakshmikantham D D Bainov and P S Simeonov Theoryof Impulsive Differential Equations World Scientific Singapore1989
[21] S M Blower and H Dowlatabadi ldquoSensitivity and uncertaintyanalysis of complex-models of disease transmission An HIVmodel as an examplerdquo International Statistical Review vol 62no 2 pp 229ndash243 1994
[22] S Marino I B Hogue C J Ray and D E Kirschner ldquoA meth-odology for performing global uncertainty and sensitivityanalysis in systems biologyrdquo Journal of Theoretical Biology vol254 no 1 pp 178ndash196 2008
[23] S Y Tang J H Liang Y S Tan and R A Cheke ldquoThresholdconditions for integrated pest management models with pesti-cides that have residual effectsrdquo Journal ofMathematical Biologyvol 66 no 1-2 pp 1ndash35 2013
[24] F Brauer and C Castillo-ChavezMathematical Models in Pop-ulation Biology and Epidemiology Springer New York NYUSA 2000
[25] O Diekmann and J A P Heesterbeek Mathematical Epidemi-ology of Infectious Diseases JohnWiley amp Sons Chichester UK2000
[26] J D MurrayMathematical Biology Springer Berlin Germany1989
[27] R Anderson and R May Infectious Diseases of HumansDynamics and Control Oxford University Press Oxford UK1995
[28] M Hethcote ldquoThree basic epidemiological modelsrdquo in AppliedMathematical Ecology L Gross T G Hallam and S A LevinEds pp 119ndash144 Springer Berlin Germany 1989
[29] A V Hill ldquoThe possible effects of the aggregation of themolecules of haemoglobin on its dissociation curvesrdquo TheJournal of Physiology vol 40 no 4 pp 4ndash7 1910
[30] P Cull ldquoLocal and global stability for population modelsrdquoBiological Cybernetics vol 54 no 3 pp 141ndash149 1986
[31] WHOWeb ldquoGlobal health observatory data repositoryrdquo 2001httpappswhointghodatatheme=country
[32] M D McKay R J Beckman and W J Conover ldquoComparisonof three methods for selecting values of input variables in theanalysis of output from a computer coderdquo Technometrics vol21 no 2 pp 239ndash245 1979
[33] W M Schaffer L F Olsen G L Truty S L Fulmer andD J Graser ldquoPeriodic and chaotic dynamics in childhoodepidemicsrdquo in From Chemical to Biological Organization MMarkus S Muller and G Nicolis Eds Springer BerlinGermany 1988
[34] A Lakmeche and O Arino ldquoBifurcation of non trivial periodicsolutions of impulsive differential equations arising chemother-apeutic treatmentrdquo Dynamics of Continuous Discrete andImpulsive Systems B vol 7 no 2 pp 265ndash287 2000
[35] S Y Tang and L S Chen ldquoMultiple attractors in stage-structured population models with birth pulsesrdquo Bulletin ofMathematical Biology vol 65 no 3 pp 479ndash495 2003
[36] S Y Tang Y N Xiao and R A Cheke ldquoMultiple attractorsof host-parasitoid models with integrated pest managementstrategies eradication persistence and outbreakrdquo TheoreticalPopulation Biology vol 73 no 2 pp 181ndash197 2008
[37] L F Olsen andWM Schaffer ldquoChaos versus noisy periodicityalternative hypotheses for childhood epidemicsrdquo Science vol249 no 4968 pp 499ndash504 1990
[38] G Sugihara and R M May ldquoNonlinear forecasting as a way ofdistinguishing chaos from measurement error in time seriesrdquoNature vol 344 no 6268 pp 734ndash741 1990
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical Problems in Engineering
Hindawi Publishing Corporationhttpwwwhindawicom
Differential EquationsInternational Journal of
Volume 2014
Applied MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Probability and StatisticsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical PhysicsAdvances in
Complex AnalysisJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
OptimizationJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CombinatoricsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Operations ResearchAdvances in
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Function Spaces
Abstract and Applied AnalysisHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of Mathematics and Mathematical Sciences
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Algebra
Discrete Dynamics in Nature and Society
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Decision SciencesAdvances in
Discrete MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014 Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Stochastic AnalysisInternational Journal of
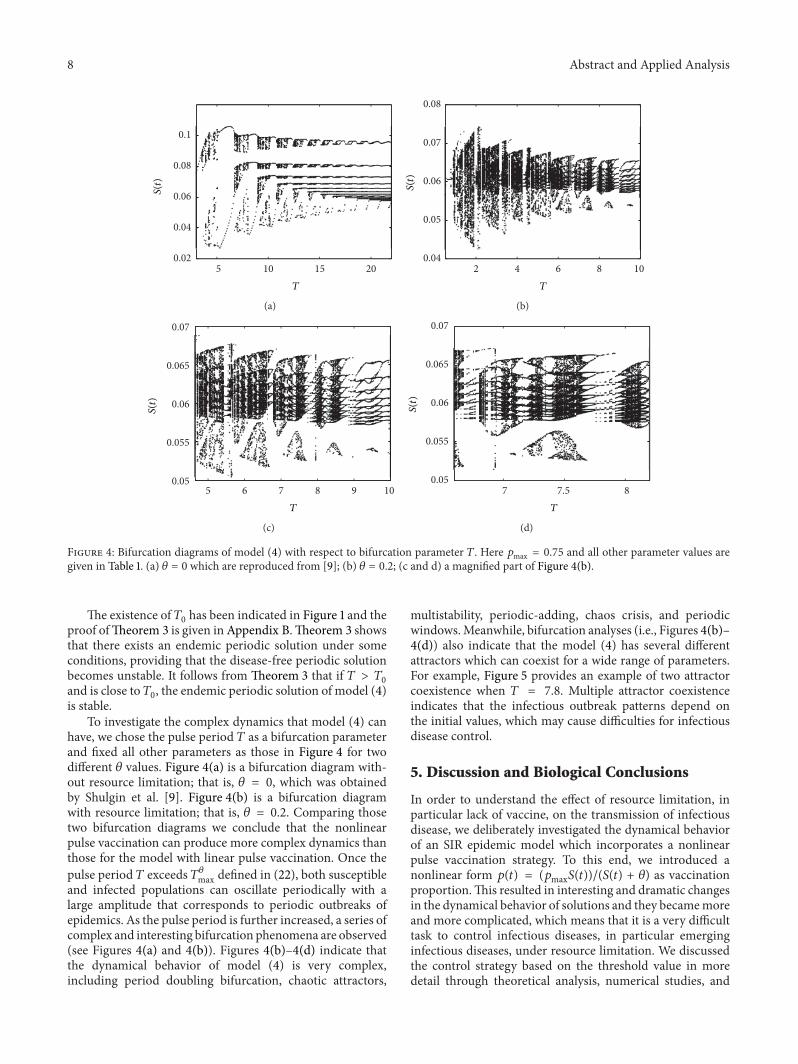
8 Abstract and Applied Analysis
5 10 15 20002
004
006
008
01
T
S(t)
(a)
2 4 6 8 10004
005
006
007
008
T
S(t)
(b)
5 6 7 8 9 10005
0055
006
0065
007
T
S(t)
(c)
7 75 8005
0055
006
0065
007
T
S(t)
(d)
Figure 4 Bifurcation diagrams of model (4) with respect to bifurcation parameter 119879 Here 119901max = 075 and all other parameter values aregiven in Table 1 (a) 120579 = 0 which are reproduced from [9] (b) 120579 = 02 (c and d) a magnified part of Figure 4(b)
The existence of 1198790has been indicated in Figure 1 and the
proof ofTheorem 3 is given in Appendix BTheorem 3 showsthat there exists an endemic periodic solution under someconditions providing that the disease-free periodic solutionbecomes unstable It follows from Theorem 3 that if 119879 gt 119879
0
and is close to 1198790 the endemic periodic solution of model (4)
is stableTo investigate the complex dynamics that model (4) can
have we chose the pulse period 119879 as a bifurcation parameterand fixed all other parameters as those in Figure 4 for twodifferent 120579 values Figure 4(a) is a bifurcation diagram with-out resource limitation that is 120579 = 0 which was obtainedby Shulgin et al [9] Figure 4(b) is a bifurcation diagramwith resource limitation that is 120579 = 02 Comparing thosetwo bifurcation diagrams we conclude that the nonlinearpulse vaccination can produce more complex dynamics thanthose for the model with linear pulse vaccination Once thepulse period 119879 exceeds 119879120579max defined in (22) both susceptibleand infected populations can oscillate periodically with alarge amplitude that corresponds to periodic outbreaks ofepidemics As the pulse period is further increased a series ofcomplex and interesting bifurcation phenomena are observed(see Figures 4(a) and 4(b)) Figures 4(b)ndash4(d) indicate thatthe dynamical behavior of model (4) is very complexincluding period doubling bifurcation chaotic attractors
multistability periodic-adding chaos crisis and periodicwindowsMeanwhile bifurcation analyses (ie Figures 4(b)ndash4(d)) also indicate that the model (4) has several differentattractors which can coexist for a wide range of parametersFor example Figure 5 provides an example of two attractorcoexistence when 119879 = 78 Multiple attractor coexistenceindicates that the infectious outbreak patterns depend onthe initial values which may cause difficulties for infectiousdisease control
5 Discussion and Biological Conclusions
In order to understand the effect of resource limitation inparticular lack of vaccine on the transmission of infectiousdisease we deliberately investigated the dynamical behaviorof an SIR epidemic model which incorporates a nonlinearpulse vaccination strategy To this end we introduced anonlinear form 119901(119905) = (119901max119878(119905))(119878(119905) + 120579) as vaccinationproportionThis resulted in interesting and dramatic changesin the dynamical behavior of solutions and they becamemoreand more complicated which means that it is a very difficulttask to control infectious diseases in particular emerginginfectious diseases under resource limitation We discussedthe control strategy based on the threshold value in moredetail through theoretical analysis numerical studies and
Abstract and Applied Analysis 9
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(a)
100 110 120 130 140 150minus13
minus12
minus11
minus10
minus9
minus8
minus7
t
Log(I(t)
)
(b)
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(c)
100 110 120 130 140 150minus16
minus14
minus12
minus10
minus8
minus6
t
Log(I(t)
)
(d)
Figure 5 Coexistence of two stable attractors of model (4) Here 119901max = 075 120579 = 02 119879 = 78 and all other parameter values are given inTable 1 (a and b) The initial value (119878
0 1198680) = (012 005) (c and d) the initial value (119878
0 1198680) = (01 002)
sensitivity analysis As pointed out byAgur et al [7] the costsrisks and the efficacy of the pulse vaccinations are key factorsin eliminating transmission of the measles virus In orderto depict those factors the nonlinear resource limitationsshould be taken into account [9] and the main purpose ofthe present work is to formulate the disease model to addressthe effects of resource limitations on disease control
The results indicate that vaccination as an approach tocontrolling epidemics must be committed to a long-termstrategy especially when resources arelimited To understandthe interactions amongst 119878(119905) and 119868(119905) which are importantfor successful control of the spread of infectious diseasewe have proposed detailed modeling methods involvingvaccination on 119878(119905) and determining the most significantparameters for the basic reproductive number 1198772
0by using
LHSPRCC uncertainty and sensitivity analysis techniques(see Figure 3) Our results indicate that we should paymore attention to the vaccination period and vaccinationproportion which reduce the threshold value 1198772
0or 11987710to
prevent the outbreak of disease Our results clarify that itis more significant to improve immunization programs (ievaccinations as mentioned in [15 16]) under conditions ofresource limitation
The results also indicate that the dynamic behavior ofmodel (4) may be dramatically affected by small changes in
the value of initial densities of susceptible and infected withresource limitation Bifurcation diagrams shown in Figure 4indicate that there are many forms of complexities in model(4) which are related to chaotic bands with periodic win-dows and attractor crises and Figure 5 reveals that theoccurrence of multiple attractors is a common property ofthe SIR model with resource limitation which can help usto further understand the application of a nonlinear pulsevaccination strategy in an SIR model (for more informationabout bifurcation diagrams of impulsive control strategiessee [35 36]) Some complexities are related to the long-termbehavior of population dynamics characterized either bywell behaving relatively simple or very complicated strangeattractors We also found that the routes to chaos are verycomplicated For instance with resource limitation there areseveral hidden factors that can adversely affect the controlstrategy The increasing number of potential complexitiespredicted by the theory does not seem to make this taskany easier Nevertheless identifying complicated possiblychaotic dynamics in population data has remained a majorchallenge in epidemiology studies In particular the sug-gestion that the dynamics of measles are chaotic has beenmuch debated (eg see [37]) and when seeking to establishwhether measles dynamics in New York City were chaoticor not Sugihara and May [38] omitted data from after 1963when vaccinations were introduced as the immunisations
10 Abstract and Applied Analysis
had altered the intrinsic dynamics However our resultsimply that even with immunisations disease dynamics couldbe chaotic under some circumstances and so a reanalysis ofthe New York data including the vaccination period might beworthwhile
In the present paper we have focused on studyingthe effects of dynamical behaviors on the SIR model withnonlinear pulse vaccination For our simple model we haveassumed that the maximal vaccination proportion 119901max isa constant and the pulse period 119879 can be varied Howeverdue to limited vaccine availability the maximal vaccinationproportion 119901max should depend on the vaccine obtained ineach pulse period Thus to describe the impact of a limitedvaccine stockpile a separate differential equation for theamount of vaccine denoted by 119881(119905) could be included inthe model and then 119901(119905) should be a function of 119881(119905) and119878(119905) Meanwhile it is essential to link the costs of developingand implementing controls to population dynamic modelingof disease epidemics in order to consider other resourcelimitations such as quantities of drugs availability of isolationunits numbers of hospital beds and equipmentThese topicswill be considered in further work in the future
Appendices
A Proof of Theorem 2
Firstly we prove the local stability of a periodic solution(119878lowast(119905) 0) which may be determined by considering the
behavior of small amplitude perturbations of the solutionDefine
119878 (119905) = 119880 (119905) + 119878lowast
(119905) 119868 (119905) = 119881 (119905) (A1)
here119880(119905) 119881(119905) are small perturbations whichmay be writtenas
(119880 (119905)
119881 (119905)) = Φ (119905) (
119880 (0)
119881 (0)) 0 le 119905 lt 119879 (A2)
where the fundamental matrixΦ(119905) satisfies
dΦ (119905)d119905
= (minus119887 minus120573119878
lowast
(119905)
0 120573119878lowast
(119905) minus 120574 minus 119887)Φ (119905) (A3)
and Φ(0) = 119868 is the identity matrix So
Φ (119879) = (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 ) (A4)
where 995333 is not necessarily computed in detail as it is notrequired in the following analysis
The resetting impulsive conditions of model (4) become
(119880 (119899119879
+)
119881 (119899119879+)) = (
1 minus 119901max +1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)(119880 (119899119879)
119881 (119899119879))
≐ 119861 (119899119879) (119880 (119899119879)
119881 (119899119879))
(A5)
Hence according to Floquet theory if the moduli of botheigenvalues of the matrix
119872 = 119861 (119879)Φ (119879)
= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)
times (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 )
(A6)
are less than one then the periodic solution (119878lowast(119905) 0) islocally stable In fact two Floquet expressions multiplied arethus
1205821= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119879))2) eminus119887119879
1205822= eint
119879
0(120573119878lowast(119905)minus120574minus119887)d119905
(A7)
It is obvious that 0 lt 1205821lt 1 Thus the stability of (119878lowast(119905) 0)
is decided by whether 1205822lt 1 or not So we conclude that
the disease-free periodic solution (119878lowast(119905) 0) is locally stable ifint119879
0(120573119878lowast(119905) minus 120574 minus 119887)d119905 lt 0 that is
1
119879int
119879
0
119878lowast
(119905) d119905 =(119878 minus 1) (1 minus eminus119887119879) + 119887119879
119887119879lt120574 + 119887
120573 (A8)
and by simple calculation we have
120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]119887119879 (120574 + 119887)
lt 1 (A9)
which holds true due to 11987710lt 1 Therefore we can draw
a conclusion from the above analysis that if 11987710lt 1 the
disease-free periodic solution (119878lowast(119905) 0) of model (4) is locallyasymptotically stable
Next we will prove the global attraction of the disease-free periodic solution (119878lowast(119905) 0) of model (4) Since 1198771
0lt 1
we can choose 1205761gt 0 sufficiently small such that
1
119879int
119879
0
(119878lowast
(119905) + 1205761) d119905 lt
120574 + 119887
120573 (A10)
It follows from model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) 119905 = 119899119879
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579 119905 = 119899119879
(A11)
Consider the comparison equation
d119885 (119905)d119905
= 119887 minus 119887119885 (119905) 119905 = 119899119879
119885 (119905+
) =(1 minus 119901max) 119885
2
(119905) + 120579119885 (119905)
119885 (119905) + 120579 119905 = 119899119879
(A12)
Abstract and Applied Analysis 11
From Lemma 1 and the comparison theorem on impul-sive differential equations we have 119878(119905) le 119885(119905) and 119885(119905) rarr119878lowast(119905) as 119905 rarr +infin Hence there exists a 119905
1gt 0 such that
119878 (119905) le 119885 (119905) lt 119878lowast
(119905) + 1205761 (A13)
for all 119905 ge 1199051 here 120576
1gt 0 is small enough
From the second equation of model (4) and (A13) we get
d119868 (119905)d119905
le (120573 (119878lowast
(119905) + 1205761) minus 120574 minus 119887) 119868 (119905) (A14)
for 119905 gt 1199051 then we consider the following comparison
equation with pulses
d1199101(119905)
d119905= (120573 (119878
lowast
(119905) + 1205761) minus 120574 minus 119887) 119910
1(119905) 119905 = 119899119879
1199101(119905+
) = 1199101(119905) 119905 = 119899119879
(A15)
Integrating model (A15) between pulses (119899119879 (119899 + 1)119879] wehave
1199101((119899 + 1) 119879) = 119910
1(119899119879) eint
(119899+1)119879
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101(119899119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A16)
Then by using step by step iterations
1199101(119899119879) = 119910
1((119899 minus 1) 119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101((119899 minus 2) 119879) e2 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= sdot sdot sdot
= 1199101(0) e119899 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A17)
where 1199101(0) = 119910
1(0+) gt 0 we obtain lim
119899rarrinfin1199101(119899119879) =
0 according to (A10) On the other hand integrating andsolving the first equation of model (A15) between pulses itgives
1199101(119905) = 119910
1(119899119879) eint
119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905 119905 isin (119899119879 (119899 + 1) 119879]
(A18)
incorporating into the boundedness of eint119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
due to 11987710lt 1 we have lim
119905rarrinfin1199101(119905) = 0
Let (119878(119905) 119868(119905)) be any solution of model (4) with initialvalue (119878
0 1198680) and 119878
0= 119878(0
+) gt 0 119868
0= 119868(0
+) =
1199101(0) gt 0 according to the comparison theorem we have
lim119905rarrinfin
sup 119868(119905) le lim119905rarrinfin
sup1199101(119905) = 0 Incorporating into
the positivity of 119868(119905) we know that lim119905rarrinfin
119868(119905) = 0Consequently there exists a 119905
2gt 1199051such that 0 lt 119868(119905) le 120576
2
for 119905 ge 1199052 and 120576
2is small enough Further we have
119887 minus (1205731205762+ 119887) 119878 (119905) le
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (A19)
for all 119905 gt 1199052 from which we can obtain the following
equation
d1199102(119905)
d119905= 119887 minus (120573120576
2+ 119887) 119910
2(119905) 119905 = 119899119879
1199102(119905+
) =(1 minus 119901max) 119910
2
2(119905) + 120579119910
2(119905)
1199102(119905) + 120579
119905 = 119899119879
(A20)
By employing the same methods as used for the proof ofLemma 1 we get that the model (A20) has a positive periodicsolution 119910lowast
2(119905) which is globally attractive where
119910lowast
2(119905) = (119910
2minus
119887
1205731205762+ 119887) eminus(1205731205762+119887)(119905minus119899119879) + 119887
1205731205762+ 119887
119905 isin (119899119879 (119899 + 1) 119879]
1199102= 1199102(119899119879+
) = 119910lowast
2(0+
) =minus1198612+ radic11986122minus 411986021198622
21198602
1198602= eminus(1205731205762+119887)119879 [ 119887
1205731205762+ 119887
minus (1 minus 119901max) eminus(1205731205762+119887)119879]
1198612= (eminus(1205731205762+119887)119879 minus 119887
1205731205762+ 119887)
times [2 (1 minus 119901max) eminus(1205731205762+119887)119879minus (120579 +
119887
1205731205762+ 119887)]
1198622= minus[(1 minus 119901max) (
119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)2
+ 120579(119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)]
(A21)
It follows from the comparison theorem on impulsive differ-ential equations that 119910
2(119905) le 119878(119905) le 119885(119905) Moreover 119910
2(119905) rarr
119910lowast
2(119905) and 119885(119905) rarr 119878
lowast(119905) as 119905 rarr +infin Consequently there
exists a 1199053for 1205763small enough such that 119905
3ge 1199052and
119910lowast
2(119905) minus 120576
3lt 119878 (119905) lt 119878
lowast
(119905) + 1205763
(A22)
for 119905 ge 1199053 Let 120576
2rarr 0 Then
119878lowast
(119905) minus 1205763lt 119878 (119905) lt 119878
lowast
(119905) + 1205763 (A23)
Therefore 119878(119905) rarr 119878lowast(119905) as 119905 rarr +infin Thus we
have proved the global stability of the disease-free periodicsolution (119878lowast(119905) 0) of model (4) This completes the proof
B Proof of Theorem 3
In order to apply the bifurcation theory of [34] in the maintext we make the following calculations
1198891015840
0= 1 minus eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
(B1)
If 11988910158400= 0 then 119879
0satisfies the following equation
120573 [(119878 minus 1) (1 minus eminus1198871198790) + 1198871198790]
119887 (120574 + 119887) 1198790
= 1 (B2)
12 Abstract and Applied Analysis
which indicates that there exists a1198790such that 1198771
0= 1 and the
disease-free periodic solution 120575 = (119878lowast(119905) 0) loses its stabilityFurther
120597Φ1(1198790 1198830)
120597119878= eminus1198871198790 gt 0
120597Φ2(1198790 1198830)
120597119868= eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
120597Φ1(1198790 1198830)
120597119868= minusint
1198790
0
120573119878lowast
(V) eminus119887(1198790minusV)eintV
0(120573119878lowast(119905)minus120574minus119887)d119905dV
≐ 119860 lt 0
1198861015840
0= 1 minus 119860
3eminus1198871198790
1198871015840
0= 1 minus 119860
3119860
1205972Φ2(1198790 1198830)
120597119868120597119878= 1205731198790eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
1205972Φ2(1198790 1198830)
1205971198682
= minus1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
119878lowast
(120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585minus119887(Vminus120579)
d120579 dV
lt 0
1205972Φ2(1198790 1198830)
120597119868120597119879= (120573119878
lowast
(1198790) minus 120574 minus 119887) eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
120597Φ1(1198790 1198830)
120597119879= minus119887 (119878 minus 1) eminus1198871198790
119861 = 1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887)
times eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
119862 = 1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
eminus119887(119905minus120579)119878lowast (120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585
d120579 dV
+ 21205731198790
1 minus 1198603119860
1 minus 1198603eminus1198871198790
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
(B3)
It follows that 119861119862 lt 0 and we get the condition that is if theparameters satisfy condition 119861 lt 0 then the model (4) has asupercritical branch at 119879
0
In fact
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0 (B4)
Therefore 119861 lt 0 is equivalent to
1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887) lt 0
(B5)
Obviously 119861 lt 0 due to (25) The proof is completed
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thiswork is supported by theNational Natural Science Foun-dation of China (NSFCs 11171199 11371030 11301320) andthe Fundamental Research Funds for the Central Universities(GK201305010) The authors would like to thank the editorand the referees for their careful reading of the original paperand many valuable comments and suggestions that greatlyimproved the presentation of this paper
References
[1] R M Anderson and R M May ldquoPopulation biology of infec-tious diseases part Irdquo Nature vol 280 no 5721 pp 361ndash3671979
[2] E Hanert E Schumacher and E Deleersnijder ldquoFront dynam-ics in fractional-order epidemic modelsrdquo Journal of TheoreticalBiology vol 279 no 1 pp 9ndash16 2011
[3] H W Hethcote ldquoMathematics of infectious diseasesrdquo SIAMReview vol 42 no 4 pp 599ndash653 2000
[4] A E Fiore C B Bridges and N J Cox ldquoSeasonal influenzavaccinesrdquo Current Topics in Microbiology and Immunology vol333 no 1 pp 43ndash82 2009
[5] Y Chang N T Brewer A C Rinas K Schmitt and J S SmithldquoEvaluating the impact of human papillomavirus vaccinesrdquoVaccine vol 27 no 32 pp 4355ndash4362 2009
[6] T J Liesegang ldquoVaricella zoster virus vaccines effective butconcerns lingerrdquo Canadian Journal of Ophthalmology vol 44no 4 pp 379ndash384 2009
[7] Z Agur L Cojocaru G Mazor R M Anderson and Y LDanon ldquoPulse mass measles vaccination across age cohortsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 90 no 24 pp 11698ndash11702 1993
[8] L Stone B Shulgin and Z Agur ldquoTheoretical examinationof the pulse vaccination policy in the SIR epidemic modelrdquoMathematical and ComputerModelling vol 31 no 4-5 pp 207ndash215 2000
[9] B Shulgin L Stone and Z Agur ldquoPulse vaccination strategy inthe SIR epidemic modelrdquo Bulletin of Mathematical Biology vol60 no 6 pp 1123ndash1148 1998
[10] A DrsquoOnofrio ldquoStability properties of pulse vaccination strategyin SEIR epidemicmodelrdquoMathematical Biosciences vol 179 no1 pp 57ndash72 2002
[11] A Franceschetti and A Pugliese ldquoThreshold behaviour of a SIRepidemicmodel with age structure and immigrationrdquo Journal ofMathematical Biology vol 57 no 1 pp 1ndash27 2008
Abstract and Applied Analysis 13
[12] A J Terry ldquoPulse vaccination strategies in a metapopulationSIR modelrdquo Mathematical Biosciences and Engineering vol 7no 2 pp 455ndash477 2010
[13] S J Gao L S Chen J J Nieto and A Torres ldquoAnalysis of adelayed epidemic model with pulse vaccination and saturationincidencerdquo Vaccine vol 24 no 35-36 pp 6037ndash6045 2006
[14] L Matrajt M E Halloran and I M Longini Jr ldquoOptimal vac-cine allocation for the early mitigation of pandemic influenzardquoPLoS Computational Biology vol 9 no 3 Article ID e10029642013
[15] S P Sullivan D G Koutsonanos M D P Martin et alldquoDissolving polymer microneedle patches for influenza vacci-nationrdquo Nature Medicine vol 16 no 8 pp 915ndash920 2010
[16] Science Daily Web ldquoVaccine-delivery patch with dissolvingmicroneedles eliminates ldquosharpsrdquo boosts protectionrdquo httpwwwsciencedailycomreleases201007100718204733htm
[17] M J Reynolds-Hogland J S Hogland and M S MitchellldquoEvaluating intercepts fromdemographicmodels to understandresource limitation and resource thresholdsrdquo Ecological Mod-elling vol 211 no 3-4 pp 424ndash432 2008
[18] L Chow M Fan and Z L Feng ldquoDynamics of a multigroupepidemiological model with group-targeted vaccination strate-giesrdquo Journal of Theoretical Biology vol 291 no 1 pp 56ndash642011
[19] L H Zhou and M Fan ldquoDynamics of an SIR epidemic modelwith limited medical resources revisitedrdquo Nonlinear AnalysisReal World Applications vol 13 no 1 pp 312ndash324 2012
[20] V Lakshmikantham D D Bainov and P S Simeonov Theoryof Impulsive Differential Equations World Scientific Singapore1989
[21] S M Blower and H Dowlatabadi ldquoSensitivity and uncertaintyanalysis of complex-models of disease transmission An HIVmodel as an examplerdquo International Statistical Review vol 62no 2 pp 229ndash243 1994
[22] S Marino I B Hogue C J Ray and D E Kirschner ldquoA meth-odology for performing global uncertainty and sensitivityanalysis in systems biologyrdquo Journal of Theoretical Biology vol254 no 1 pp 178ndash196 2008
[23] S Y Tang J H Liang Y S Tan and R A Cheke ldquoThresholdconditions for integrated pest management models with pesti-cides that have residual effectsrdquo Journal ofMathematical Biologyvol 66 no 1-2 pp 1ndash35 2013
[24] F Brauer and C Castillo-ChavezMathematical Models in Pop-ulation Biology and Epidemiology Springer New York NYUSA 2000
[25] O Diekmann and J A P Heesterbeek Mathematical Epidemi-ology of Infectious Diseases JohnWiley amp Sons Chichester UK2000
[26] J D MurrayMathematical Biology Springer Berlin Germany1989
[27] R Anderson and R May Infectious Diseases of HumansDynamics and Control Oxford University Press Oxford UK1995
[28] M Hethcote ldquoThree basic epidemiological modelsrdquo in AppliedMathematical Ecology L Gross T G Hallam and S A LevinEds pp 119ndash144 Springer Berlin Germany 1989
[29] A V Hill ldquoThe possible effects of the aggregation of themolecules of haemoglobin on its dissociation curvesrdquo TheJournal of Physiology vol 40 no 4 pp 4ndash7 1910
[30] P Cull ldquoLocal and global stability for population modelsrdquoBiological Cybernetics vol 54 no 3 pp 141ndash149 1986
[31] WHOWeb ldquoGlobal health observatory data repositoryrdquo 2001httpappswhointghodatatheme=country
[32] M D McKay R J Beckman and W J Conover ldquoComparisonof three methods for selecting values of input variables in theanalysis of output from a computer coderdquo Technometrics vol21 no 2 pp 239ndash245 1979
[33] W M Schaffer L F Olsen G L Truty S L Fulmer andD J Graser ldquoPeriodic and chaotic dynamics in childhoodepidemicsrdquo in From Chemical to Biological Organization MMarkus S Muller and G Nicolis Eds Springer BerlinGermany 1988
[34] A Lakmeche and O Arino ldquoBifurcation of non trivial periodicsolutions of impulsive differential equations arising chemother-apeutic treatmentrdquo Dynamics of Continuous Discrete andImpulsive Systems B vol 7 no 2 pp 265ndash287 2000
[35] S Y Tang and L S Chen ldquoMultiple attractors in stage-structured population models with birth pulsesrdquo Bulletin ofMathematical Biology vol 65 no 3 pp 479ndash495 2003
[36] S Y Tang Y N Xiao and R A Cheke ldquoMultiple attractorsof host-parasitoid models with integrated pest managementstrategies eradication persistence and outbreakrdquo TheoreticalPopulation Biology vol 73 no 2 pp 181ndash197 2008
[37] L F Olsen andWM Schaffer ldquoChaos versus noisy periodicityalternative hypotheses for childhood epidemicsrdquo Science vol249 no 4968 pp 499ndash504 1990
[38] G Sugihara and R M May ldquoNonlinear forecasting as a way ofdistinguishing chaos from measurement error in time seriesrdquoNature vol 344 no 6268 pp 734ndash741 1990
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical Problems in Engineering
Hindawi Publishing Corporationhttpwwwhindawicom
Differential EquationsInternational Journal of
Volume 2014
Applied MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Probability and StatisticsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical PhysicsAdvances in
Complex AnalysisJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
OptimizationJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CombinatoricsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Operations ResearchAdvances in
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Function Spaces
Abstract and Applied AnalysisHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of Mathematics and Mathematical Sciences
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Algebra
Discrete Dynamics in Nature and Society
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Decision SciencesAdvances in
Discrete MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014 Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Stochastic AnalysisInternational Journal of

Abstract and Applied Analysis 9
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(a)
100 110 120 130 140 150minus13
minus12
minus11
minus10
minus9
minus8
minus7
t
Log(I(t)
)
(b)
100 110 120 130 140 150004
0045
005
0055
006
0065
t
S(t)
(c)
100 110 120 130 140 150minus16
minus14
minus12
minus10
minus8
minus6
t
Log(I(t)
)
(d)
Figure 5 Coexistence of two stable attractors of model (4) Here 119901max = 075 120579 = 02 119879 = 78 and all other parameter values are given inTable 1 (a and b) The initial value (119878
0 1198680) = (012 005) (c and d) the initial value (119878
0 1198680) = (01 002)
sensitivity analysis As pointed out byAgur et al [7] the costsrisks and the efficacy of the pulse vaccinations are key factorsin eliminating transmission of the measles virus In orderto depict those factors the nonlinear resource limitationsshould be taken into account [9] and the main purpose ofthe present work is to formulate the disease model to addressthe effects of resource limitations on disease control
The results indicate that vaccination as an approach tocontrolling epidemics must be committed to a long-termstrategy especially when resources arelimited To understandthe interactions amongst 119878(119905) and 119868(119905) which are importantfor successful control of the spread of infectious diseasewe have proposed detailed modeling methods involvingvaccination on 119878(119905) and determining the most significantparameters for the basic reproductive number 1198772
0by using
LHSPRCC uncertainty and sensitivity analysis techniques(see Figure 3) Our results indicate that we should paymore attention to the vaccination period and vaccinationproportion which reduce the threshold value 1198772
0or 11987710to
prevent the outbreak of disease Our results clarify that itis more significant to improve immunization programs (ievaccinations as mentioned in [15 16]) under conditions ofresource limitation
The results also indicate that the dynamic behavior ofmodel (4) may be dramatically affected by small changes in
the value of initial densities of susceptible and infected withresource limitation Bifurcation diagrams shown in Figure 4indicate that there are many forms of complexities in model(4) which are related to chaotic bands with periodic win-dows and attractor crises and Figure 5 reveals that theoccurrence of multiple attractors is a common property ofthe SIR model with resource limitation which can help usto further understand the application of a nonlinear pulsevaccination strategy in an SIR model (for more informationabout bifurcation diagrams of impulsive control strategiessee [35 36]) Some complexities are related to the long-termbehavior of population dynamics characterized either bywell behaving relatively simple or very complicated strangeattractors We also found that the routes to chaos are verycomplicated For instance with resource limitation there areseveral hidden factors that can adversely affect the controlstrategy The increasing number of potential complexitiespredicted by the theory does not seem to make this taskany easier Nevertheless identifying complicated possiblychaotic dynamics in population data has remained a majorchallenge in epidemiology studies In particular the sug-gestion that the dynamics of measles are chaotic has beenmuch debated (eg see [37]) and when seeking to establishwhether measles dynamics in New York City were chaoticor not Sugihara and May [38] omitted data from after 1963when vaccinations were introduced as the immunisations
10 Abstract and Applied Analysis
had altered the intrinsic dynamics However our resultsimply that even with immunisations disease dynamics couldbe chaotic under some circumstances and so a reanalysis ofthe New York data including the vaccination period might beworthwhile
In the present paper we have focused on studyingthe effects of dynamical behaviors on the SIR model withnonlinear pulse vaccination For our simple model we haveassumed that the maximal vaccination proportion 119901max isa constant and the pulse period 119879 can be varied Howeverdue to limited vaccine availability the maximal vaccinationproportion 119901max should depend on the vaccine obtained ineach pulse period Thus to describe the impact of a limitedvaccine stockpile a separate differential equation for theamount of vaccine denoted by 119881(119905) could be included inthe model and then 119901(119905) should be a function of 119881(119905) and119878(119905) Meanwhile it is essential to link the costs of developingand implementing controls to population dynamic modelingof disease epidemics in order to consider other resourcelimitations such as quantities of drugs availability of isolationunits numbers of hospital beds and equipmentThese topicswill be considered in further work in the future
Appendices
A Proof of Theorem 2
Firstly we prove the local stability of a periodic solution(119878lowast(119905) 0) which may be determined by considering the
behavior of small amplitude perturbations of the solutionDefine
119878 (119905) = 119880 (119905) + 119878lowast
(119905) 119868 (119905) = 119881 (119905) (A1)
here119880(119905) 119881(119905) are small perturbations whichmay be writtenas
(119880 (119905)
119881 (119905)) = Φ (119905) (
119880 (0)
119881 (0)) 0 le 119905 lt 119879 (A2)
where the fundamental matrixΦ(119905) satisfies
dΦ (119905)d119905
= (minus119887 minus120573119878
lowast
(119905)
0 120573119878lowast
(119905) minus 120574 minus 119887)Φ (119905) (A3)
and Φ(0) = 119868 is the identity matrix So
Φ (119879) = (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 ) (A4)
where 995333 is not necessarily computed in detail as it is notrequired in the following analysis
The resetting impulsive conditions of model (4) become
(119880 (119899119879
+)
119881 (119899119879+)) = (
1 minus 119901max +1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)(119880 (119899119879)
119881 (119899119879))
≐ 119861 (119899119879) (119880 (119899119879)
119881 (119899119879))
(A5)
Hence according to Floquet theory if the moduli of botheigenvalues of the matrix
119872 = 119861 (119879)Φ (119879)
= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)
times (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 )
(A6)
are less than one then the periodic solution (119878lowast(119905) 0) islocally stable In fact two Floquet expressions multiplied arethus
1205821= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119879))2) eminus119887119879
1205822= eint
119879
0(120573119878lowast(119905)minus120574minus119887)d119905
(A7)
It is obvious that 0 lt 1205821lt 1 Thus the stability of (119878lowast(119905) 0)
is decided by whether 1205822lt 1 or not So we conclude that
the disease-free periodic solution (119878lowast(119905) 0) is locally stable ifint119879
0(120573119878lowast(119905) minus 120574 minus 119887)d119905 lt 0 that is
1
119879int
119879
0
119878lowast
(119905) d119905 =(119878 minus 1) (1 minus eminus119887119879) + 119887119879
119887119879lt120574 + 119887
120573 (A8)
and by simple calculation we have
120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]119887119879 (120574 + 119887)
lt 1 (A9)
which holds true due to 11987710lt 1 Therefore we can draw
a conclusion from the above analysis that if 11987710lt 1 the
disease-free periodic solution (119878lowast(119905) 0) of model (4) is locallyasymptotically stable
Next we will prove the global attraction of the disease-free periodic solution (119878lowast(119905) 0) of model (4) Since 1198771
0lt 1
we can choose 1205761gt 0 sufficiently small such that
1
119879int
119879
0
(119878lowast
(119905) + 1205761) d119905 lt
120574 + 119887
120573 (A10)
It follows from model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) 119905 = 119899119879
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579 119905 = 119899119879
(A11)
Consider the comparison equation
d119885 (119905)d119905
= 119887 minus 119887119885 (119905) 119905 = 119899119879
119885 (119905+
) =(1 minus 119901max) 119885
2
(119905) + 120579119885 (119905)
119885 (119905) + 120579 119905 = 119899119879
(A12)
Abstract and Applied Analysis 11
From Lemma 1 and the comparison theorem on impul-sive differential equations we have 119878(119905) le 119885(119905) and 119885(119905) rarr119878lowast(119905) as 119905 rarr +infin Hence there exists a 119905
1gt 0 such that
119878 (119905) le 119885 (119905) lt 119878lowast
(119905) + 1205761 (A13)
for all 119905 ge 1199051 here 120576
1gt 0 is small enough
From the second equation of model (4) and (A13) we get
d119868 (119905)d119905
le (120573 (119878lowast
(119905) + 1205761) minus 120574 minus 119887) 119868 (119905) (A14)
for 119905 gt 1199051 then we consider the following comparison
equation with pulses
d1199101(119905)
d119905= (120573 (119878
lowast
(119905) + 1205761) minus 120574 minus 119887) 119910
1(119905) 119905 = 119899119879
1199101(119905+
) = 1199101(119905) 119905 = 119899119879
(A15)
Integrating model (A15) between pulses (119899119879 (119899 + 1)119879] wehave
1199101((119899 + 1) 119879) = 119910
1(119899119879) eint
(119899+1)119879
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101(119899119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A16)
Then by using step by step iterations
1199101(119899119879) = 119910
1((119899 minus 1) 119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101((119899 minus 2) 119879) e2 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= sdot sdot sdot
= 1199101(0) e119899 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A17)
where 1199101(0) = 119910
1(0+) gt 0 we obtain lim
119899rarrinfin1199101(119899119879) =
0 according to (A10) On the other hand integrating andsolving the first equation of model (A15) between pulses itgives
1199101(119905) = 119910
1(119899119879) eint
119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905 119905 isin (119899119879 (119899 + 1) 119879]
(A18)
incorporating into the boundedness of eint119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
due to 11987710lt 1 we have lim
119905rarrinfin1199101(119905) = 0
Let (119878(119905) 119868(119905)) be any solution of model (4) with initialvalue (119878
0 1198680) and 119878
0= 119878(0
+) gt 0 119868
0= 119868(0
+) =
1199101(0) gt 0 according to the comparison theorem we have
lim119905rarrinfin
sup 119868(119905) le lim119905rarrinfin
sup1199101(119905) = 0 Incorporating into
the positivity of 119868(119905) we know that lim119905rarrinfin
119868(119905) = 0Consequently there exists a 119905
2gt 1199051such that 0 lt 119868(119905) le 120576
2
for 119905 ge 1199052 and 120576
2is small enough Further we have
119887 minus (1205731205762+ 119887) 119878 (119905) le
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (A19)
for all 119905 gt 1199052 from which we can obtain the following
equation
d1199102(119905)
d119905= 119887 minus (120573120576
2+ 119887) 119910
2(119905) 119905 = 119899119879
1199102(119905+
) =(1 minus 119901max) 119910
2
2(119905) + 120579119910
2(119905)
1199102(119905) + 120579
119905 = 119899119879
(A20)
By employing the same methods as used for the proof ofLemma 1 we get that the model (A20) has a positive periodicsolution 119910lowast
2(119905) which is globally attractive where
119910lowast
2(119905) = (119910
2minus
119887
1205731205762+ 119887) eminus(1205731205762+119887)(119905minus119899119879) + 119887
1205731205762+ 119887
119905 isin (119899119879 (119899 + 1) 119879]
1199102= 1199102(119899119879+
) = 119910lowast
2(0+
) =minus1198612+ radic11986122minus 411986021198622
21198602
1198602= eminus(1205731205762+119887)119879 [ 119887
1205731205762+ 119887
minus (1 minus 119901max) eminus(1205731205762+119887)119879]
1198612= (eminus(1205731205762+119887)119879 minus 119887
1205731205762+ 119887)
times [2 (1 minus 119901max) eminus(1205731205762+119887)119879minus (120579 +
119887
1205731205762+ 119887)]
1198622= minus[(1 minus 119901max) (
119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)2
+ 120579(119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)]
(A21)
It follows from the comparison theorem on impulsive differ-ential equations that 119910
2(119905) le 119878(119905) le 119885(119905) Moreover 119910
2(119905) rarr
119910lowast
2(119905) and 119885(119905) rarr 119878
lowast(119905) as 119905 rarr +infin Consequently there
exists a 1199053for 1205763small enough such that 119905
3ge 1199052and
119910lowast
2(119905) minus 120576
3lt 119878 (119905) lt 119878
lowast
(119905) + 1205763
(A22)
for 119905 ge 1199053 Let 120576
2rarr 0 Then
119878lowast
(119905) minus 1205763lt 119878 (119905) lt 119878
lowast
(119905) + 1205763 (A23)
Therefore 119878(119905) rarr 119878lowast(119905) as 119905 rarr +infin Thus we
have proved the global stability of the disease-free periodicsolution (119878lowast(119905) 0) of model (4) This completes the proof
B Proof of Theorem 3
In order to apply the bifurcation theory of [34] in the maintext we make the following calculations
1198891015840
0= 1 minus eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
(B1)
If 11988910158400= 0 then 119879
0satisfies the following equation
120573 [(119878 minus 1) (1 minus eminus1198871198790) + 1198871198790]
119887 (120574 + 119887) 1198790
= 1 (B2)
12 Abstract and Applied Analysis
which indicates that there exists a1198790such that 1198771
0= 1 and the
disease-free periodic solution 120575 = (119878lowast(119905) 0) loses its stabilityFurther
120597Φ1(1198790 1198830)
120597119878= eminus1198871198790 gt 0
120597Φ2(1198790 1198830)
120597119868= eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
120597Φ1(1198790 1198830)
120597119868= minusint
1198790
0
120573119878lowast
(V) eminus119887(1198790minusV)eintV
0(120573119878lowast(119905)minus120574minus119887)d119905dV
≐ 119860 lt 0
1198861015840
0= 1 minus 119860
3eminus1198871198790
1198871015840
0= 1 minus 119860
3119860
1205972Φ2(1198790 1198830)
120597119868120597119878= 1205731198790eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
1205972Φ2(1198790 1198830)
1205971198682
= minus1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
119878lowast
(120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585minus119887(Vminus120579)
d120579 dV
lt 0
1205972Φ2(1198790 1198830)
120597119868120597119879= (120573119878
lowast
(1198790) minus 120574 minus 119887) eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
120597Φ1(1198790 1198830)
120597119879= minus119887 (119878 minus 1) eminus1198871198790
119861 = 1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887)
times eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
119862 = 1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
eminus119887(119905minus120579)119878lowast (120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585
d120579 dV
+ 21205731198790
1 minus 1198603119860
1 minus 1198603eminus1198871198790
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
(B3)
It follows that 119861119862 lt 0 and we get the condition that is if theparameters satisfy condition 119861 lt 0 then the model (4) has asupercritical branch at 119879
0
In fact
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0 (B4)
Therefore 119861 lt 0 is equivalent to
1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887) lt 0
(B5)
Obviously 119861 lt 0 due to (25) The proof is completed
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thiswork is supported by theNational Natural Science Foun-dation of China (NSFCs 11171199 11371030 11301320) andthe Fundamental Research Funds for the Central Universities(GK201305010) The authors would like to thank the editorand the referees for their careful reading of the original paperand many valuable comments and suggestions that greatlyimproved the presentation of this paper
References
[1] R M Anderson and R M May ldquoPopulation biology of infec-tious diseases part Irdquo Nature vol 280 no 5721 pp 361ndash3671979
[2] E Hanert E Schumacher and E Deleersnijder ldquoFront dynam-ics in fractional-order epidemic modelsrdquo Journal of TheoreticalBiology vol 279 no 1 pp 9ndash16 2011
[3] H W Hethcote ldquoMathematics of infectious diseasesrdquo SIAMReview vol 42 no 4 pp 599ndash653 2000
[4] A E Fiore C B Bridges and N J Cox ldquoSeasonal influenzavaccinesrdquo Current Topics in Microbiology and Immunology vol333 no 1 pp 43ndash82 2009
[5] Y Chang N T Brewer A C Rinas K Schmitt and J S SmithldquoEvaluating the impact of human papillomavirus vaccinesrdquoVaccine vol 27 no 32 pp 4355ndash4362 2009
[6] T J Liesegang ldquoVaricella zoster virus vaccines effective butconcerns lingerrdquo Canadian Journal of Ophthalmology vol 44no 4 pp 379ndash384 2009
[7] Z Agur L Cojocaru G Mazor R M Anderson and Y LDanon ldquoPulse mass measles vaccination across age cohortsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 90 no 24 pp 11698ndash11702 1993
[8] L Stone B Shulgin and Z Agur ldquoTheoretical examinationof the pulse vaccination policy in the SIR epidemic modelrdquoMathematical and ComputerModelling vol 31 no 4-5 pp 207ndash215 2000
[9] B Shulgin L Stone and Z Agur ldquoPulse vaccination strategy inthe SIR epidemic modelrdquo Bulletin of Mathematical Biology vol60 no 6 pp 1123ndash1148 1998
[10] A DrsquoOnofrio ldquoStability properties of pulse vaccination strategyin SEIR epidemicmodelrdquoMathematical Biosciences vol 179 no1 pp 57ndash72 2002
[11] A Franceschetti and A Pugliese ldquoThreshold behaviour of a SIRepidemicmodel with age structure and immigrationrdquo Journal ofMathematical Biology vol 57 no 1 pp 1ndash27 2008
Abstract and Applied Analysis 13
[12] A J Terry ldquoPulse vaccination strategies in a metapopulationSIR modelrdquo Mathematical Biosciences and Engineering vol 7no 2 pp 455ndash477 2010
[13] S J Gao L S Chen J J Nieto and A Torres ldquoAnalysis of adelayed epidemic model with pulse vaccination and saturationincidencerdquo Vaccine vol 24 no 35-36 pp 6037ndash6045 2006
[14] L Matrajt M E Halloran and I M Longini Jr ldquoOptimal vac-cine allocation for the early mitigation of pandemic influenzardquoPLoS Computational Biology vol 9 no 3 Article ID e10029642013
[15] S P Sullivan D G Koutsonanos M D P Martin et alldquoDissolving polymer microneedle patches for influenza vacci-nationrdquo Nature Medicine vol 16 no 8 pp 915ndash920 2010
[16] Science Daily Web ldquoVaccine-delivery patch with dissolvingmicroneedles eliminates ldquosharpsrdquo boosts protectionrdquo httpwwwsciencedailycomreleases201007100718204733htm
[17] M J Reynolds-Hogland J S Hogland and M S MitchellldquoEvaluating intercepts fromdemographicmodels to understandresource limitation and resource thresholdsrdquo Ecological Mod-elling vol 211 no 3-4 pp 424ndash432 2008
[18] L Chow M Fan and Z L Feng ldquoDynamics of a multigroupepidemiological model with group-targeted vaccination strate-giesrdquo Journal of Theoretical Biology vol 291 no 1 pp 56ndash642011
[19] L H Zhou and M Fan ldquoDynamics of an SIR epidemic modelwith limited medical resources revisitedrdquo Nonlinear AnalysisReal World Applications vol 13 no 1 pp 312ndash324 2012
[20] V Lakshmikantham D D Bainov and P S Simeonov Theoryof Impulsive Differential Equations World Scientific Singapore1989
[21] S M Blower and H Dowlatabadi ldquoSensitivity and uncertaintyanalysis of complex-models of disease transmission An HIVmodel as an examplerdquo International Statistical Review vol 62no 2 pp 229ndash243 1994
[22] S Marino I B Hogue C J Ray and D E Kirschner ldquoA meth-odology for performing global uncertainty and sensitivityanalysis in systems biologyrdquo Journal of Theoretical Biology vol254 no 1 pp 178ndash196 2008
[23] S Y Tang J H Liang Y S Tan and R A Cheke ldquoThresholdconditions for integrated pest management models with pesti-cides that have residual effectsrdquo Journal ofMathematical Biologyvol 66 no 1-2 pp 1ndash35 2013
[24] F Brauer and C Castillo-ChavezMathematical Models in Pop-ulation Biology and Epidemiology Springer New York NYUSA 2000
[25] O Diekmann and J A P Heesterbeek Mathematical Epidemi-ology of Infectious Diseases JohnWiley amp Sons Chichester UK2000
[26] J D MurrayMathematical Biology Springer Berlin Germany1989
[27] R Anderson and R May Infectious Diseases of HumansDynamics and Control Oxford University Press Oxford UK1995
[28] M Hethcote ldquoThree basic epidemiological modelsrdquo in AppliedMathematical Ecology L Gross T G Hallam and S A LevinEds pp 119ndash144 Springer Berlin Germany 1989
[29] A V Hill ldquoThe possible effects of the aggregation of themolecules of haemoglobin on its dissociation curvesrdquo TheJournal of Physiology vol 40 no 4 pp 4ndash7 1910
[30] P Cull ldquoLocal and global stability for population modelsrdquoBiological Cybernetics vol 54 no 3 pp 141ndash149 1986
[31] WHOWeb ldquoGlobal health observatory data repositoryrdquo 2001httpappswhointghodatatheme=country
[32] M D McKay R J Beckman and W J Conover ldquoComparisonof three methods for selecting values of input variables in theanalysis of output from a computer coderdquo Technometrics vol21 no 2 pp 239ndash245 1979
[33] W M Schaffer L F Olsen G L Truty S L Fulmer andD J Graser ldquoPeriodic and chaotic dynamics in childhoodepidemicsrdquo in From Chemical to Biological Organization MMarkus S Muller and G Nicolis Eds Springer BerlinGermany 1988
[34] A Lakmeche and O Arino ldquoBifurcation of non trivial periodicsolutions of impulsive differential equations arising chemother-apeutic treatmentrdquo Dynamics of Continuous Discrete andImpulsive Systems B vol 7 no 2 pp 265ndash287 2000
[35] S Y Tang and L S Chen ldquoMultiple attractors in stage-structured population models with birth pulsesrdquo Bulletin ofMathematical Biology vol 65 no 3 pp 479ndash495 2003
[36] S Y Tang Y N Xiao and R A Cheke ldquoMultiple attractorsof host-parasitoid models with integrated pest managementstrategies eradication persistence and outbreakrdquo TheoreticalPopulation Biology vol 73 no 2 pp 181ndash197 2008
[37] L F Olsen andWM Schaffer ldquoChaos versus noisy periodicityalternative hypotheses for childhood epidemicsrdquo Science vol249 no 4968 pp 499ndash504 1990
[38] G Sugihara and R M May ldquoNonlinear forecasting as a way ofdistinguishing chaos from measurement error in time seriesrdquoNature vol 344 no 6268 pp 734ndash741 1990
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical Problems in Engineering
Hindawi Publishing Corporationhttpwwwhindawicom
Differential EquationsInternational Journal of
Volume 2014
Applied MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Probability and StatisticsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical PhysicsAdvances in
Complex AnalysisJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
OptimizationJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CombinatoricsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Operations ResearchAdvances in
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Function Spaces
Abstract and Applied AnalysisHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of Mathematics and Mathematical Sciences
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Algebra
Discrete Dynamics in Nature and Society
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Decision SciencesAdvances in
Discrete MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014 Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Stochastic AnalysisInternational Journal of

10 Abstract and Applied Analysis
had altered the intrinsic dynamics However our resultsimply that even with immunisations disease dynamics couldbe chaotic under some circumstances and so a reanalysis ofthe New York data including the vaccination period might beworthwhile
In the present paper we have focused on studyingthe effects of dynamical behaviors on the SIR model withnonlinear pulse vaccination For our simple model we haveassumed that the maximal vaccination proportion 119901max isa constant and the pulse period 119879 can be varied Howeverdue to limited vaccine availability the maximal vaccinationproportion 119901max should depend on the vaccine obtained ineach pulse period Thus to describe the impact of a limitedvaccine stockpile a separate differential equation for theamount of vaccine denoted by 119881(119905) could be included inthe model and then 119901(119905) should be a function of 119881(119905) and119878(119905) Meanwhile it is essential to link the costs of developingand implementing controls to population dynamic modelingof disease epidemics in order to consider other resourcelimitations such as quantities of drugs availability of isolationunits numbers of hospital beds and equipmentThese topicswill be considered in further work in the future
Appendices
A Proof of Theorem 2
Firstly we prove the local stability of a periodic solution(119878lowast(119905) 0) which may be determined by considering the
behavior of small amplitude perturbations of the solutionDefine
119878 (119905) = 119880 (119905) + 119878lowast
(119905) 119868 (119905) = 119881 (119905) (A1)
here119880(119905) 119881(119905) are small perturbations whichmay be writtenas
(119880 (119905)
119881 (119905)) = Φ (119905) (
119880 (0)
119881 (0)) 0 le 119905 lt 119879 (A2)
where the fundamental matrixΦ(119905) satisfies
dΦ (119905)d119905
= (minus119887 minus120573119878
lowast
(119905)
0 120573119878lowast
(119905) minus 120574 minus 119887)Φ (119905) (A3)
and Φ(0) = 119868 is the identity matrix So
Φ (119879) = (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 ) (A4)
where 995333 is not necessarily computed in detail as it is notrequired in the following analysis
The resetting impulsive conditions of model (4) become
(119880 (119899119879
+)
119881 (119899119879+)) = (
1 minus 119901max +1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)(119880 (119899119879)
119881 (119899119879))
≐ 119861 (119899119879) (119880 (119899119879)
119881 (119899119879))
(A5)
Hence according to Floquet theory if the moduli of botheigenvalues of the matrix
119872 = 119861 (119879)Φ (119879)
= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119899119879))20
0 1
)
times (eminus119887119879 995333
0 eint119879
0(120573119878lowast(119905)minus120574minus119887)d119905 )
(A6)
are less than one then the periodic solution (119878lowast(119905) 0) islocally stable In fact two Floquet expressions multiplied arethus
1205821= (1 minus 119901max +
1205792119901max
(120579 + 119878lowast (119879))2) eminus119887119879
1205822= eint
119879
0(120573119878lowast(119905)minus120574minus119887)d119905
(A7)
It is obvious that 0 lt 1205821lt 1 Thus the stability of (119878lowast(119905) 0)
is decided by whether 1205822lt 1 or not So we conclude that
the disease-free periodic solution (119878lowast(119905) 0) is locally stable ifint119879
0(120573119878lowast(119905) minus 120574 minus 119887)d119905 lt 0 that is
1
119879int
119879
0
119878lowast
(119905) d119905 =(119878 minus 1) (1 minus eminus119887119879) + 119887119879
119887119879lt120574 + 119887
120573 (A8)
and by simple calculation we have
120573 [(119878 minus 1) (1 minus eminus119887119879) + 119887119879]119887119879 (120574 + 119887)
lt 1 (A9)
which holds true due to 11987710lt 1 Therefore we can draw
a conclusion from the above analysis that if 11987710lt 1 the
disease-free periodic solution (119878lowast(119905) 0) of model (4) is locallyasymptotically stable
Next we will prove the global attraction of the disease-free periodic solution (119878lowast(119905) 0) of model (4) Since 1198771
0lt 1
we can choose 1205761gt 0 sufficiently small such that
1
119879int
119879
0
(119878lowast
(119905) + 1205761) d119905 lt
120574 + 119887
120573 (A10)
It follows from model (4) that
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) 119905 = 119899119879
119878 (119905+
) =(1 minus 119901max) 119878
2
(119905) + 120579119878 (119905)
119878 (119905) + 120579 119905 = 119899119879
(A11)
Consider the comparison equation
d119885 (119905)d119905
= 119887 minus 119887119885 (119905) 119905 = 119899119879
119885 (119905+
) =(1 minus 119901max) 119885
2
(119905) + 120579119885 (119905)
119885 (119905) + 120579 119905 = 119899119879
(A12)
Abstract and Applied Analysis 11
From Lemma 1 and the comparison theorem on impul-sive differential equations we have 119878(119905) le 119885(119905) and 119885(119905) rarr119878lowast(119905) as 119905 rarr +infin Hence there exists a 119905
1gt 0 such that
119878 (119905) le 119885 (119905) lt 119878lowast
(119905) + 1205761 (A13)
for all 119905 ge 1199051 here 120576
1gt 0 is small enough
From the second equation of model (4) and (A13) we get
d119868 (119905)d119905
le (120573 (119878lowast
(119905) + 1205761) minus 120574 minus 119887) 119868 (119905) (A14)
for 119905 gt 1199051 then we consider the following comparison
equation with pulses
d1199101(119905)
d119905= (120573 (119878
lowast
(119905) + 1205761) minus 120574 minus 119887) 119910
1(119905) 119905 = 119899119879
1199101(119905+
) = 1199101(119905) 119905 = 119899119879
(A15)
Integrating model (A15) between pulses (119899119879 (119899 + 1)119879] wehave
1199101((119899 + 1) 119879) = 119910
1(119899119879) eint
(119899+1)119879
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101(119899119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A16)
Then by using step by step iterations
1199101(119899119879) = 119910
1((119899 minus 1) 119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101((119899 minus 2) 119879) e2 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= sdot sdot sdot
= 1199101(0) e119899 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A17)
where 1199101(0) = 119910
1(0+) gt 0 we obtain lim
119899rarrinfin1199101(119899119879) =
0 according to (A10) On the other hand integrating andsolving the first equation of model (A15) between pulses itgives
1199101(119905) = 119910
1(119899119879) eint
119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905 119905 isin (119899119879 (119899 + 1) 119879]
(A18)
incorporating into the boundedness of eint119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
due to 11987710lt 1 we have lim
119905rarrinfin1199101(119905) = 0
Let (119878(119905) 119868(119905)) be any solution of model (4) with initialvalue (119878
0 1198680) and 119878
0= 119878(0
+) gt 0 119868
0= 119868(0
+) =
1199101(0) gt 0 according to the comparison theorem we have
lim119905rarrinfin
sup 119868(119905) le lim119905rarrinfin
sup1199101(119905) = 0 Incorporating into
the positivity of 119868(119905) we know that lim119905rarrinfin
119868(119905) = 0Consequently there exists a 119905
2gt 1199051such that 0 lt 119868(119905) le 120576
2
for 119905 ge 1199052 and 120576
2is small enough Further we have
119887 minus (1205731205762+ 119887) 119878 (119905) le
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (A19)
for all 119905 gt 1199052 from which we can obtain the following
equation
d1199102(119905)
d119905= 119887 minus (120573120576
2+ 119887) 119910
2(119905) 119905 = 119899119879
1199102(119905+
) =(1 minus 119901max) 119910
2
2(119905) + 120579119910
2(119905)
1199102(119905) + 120579
119905 = 119899119879
(A20)
By employing the same methods as used for the proof ofLemma 1 we get that the model (A20) has a positive periodicsolution 119910lowast
2(119905) which is globally attractive where
119910lowast
2(119905) = (119910
2minus
119887
1205731205762+ 119887) eminus(1205731205762+119887)(119905minus119899119879) + 119887
1205731205762+ 119887
119905 isin (119899119879 (119899 + 1) 119879]
1199102= 1199102(119899119879+
) = 119910lowast
2(0+
) =minus1198612+ radic11986122minus 411986021198622
21198602
1198602= eminus(1205731205762+119887)119879 [ 119887
1205731205762+ 119887
minus (1 minus 119901max) eminus(1205731205762+119887)119879]
1198612= (eminus(1205731205762+119887)119879 minus 119887
1205731205762+ 119887)
times [2 (1 minus 119901max) eminus(1205731205762+119887)119879minus (120579 +
119887
1205731205762+ 119887)]
1198622= minus[(1 minus 119901max) (
119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)2
+ 120579(119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)]
(A21)
It follows from the comparison theorem on impulsive differ-ential equations that 119910
2(119905) le 119878(119905) le 119885(119905) Moreover 119910
2(119905) rarr
119910lowast
2(119905) and 119885(119905) rarr 119878
lowast(119905) as 119905 rarr +infin Consequently there
exists a 1199053for 1205763small enough such that 119905
3ge 1199052and
119910lowast
2(119905) minus 120576
3lt 119878 (119905) lt 119878
lowast
(119905) + 1205763
(A22)
for 119905 ge 1199053 Let 120576
2rarr 0 Then
119878lowast
(119905) minus 1205763lt 119878 (119905) lt 119878
lowast
(119905) + 1205763 (A23)
Therefore 119878(119905) rarr 119878lowast(119905) as 119905 rarr +infin Thus we
have proved the global stability of the disease-free periodicsolution (119878lowast(119905) 0) of model (4) This completes the proof
B Proof of Theorem 3
In order to apply the bifurcation theory of [34] in the maintext we make the following calculations
1198891015840
0= 1 minus eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
(B1)
If 11988910158400= 0 then 119879
0satisfies the following equation
120573 [(119878 minus 1) (1 minus eminus1198871198790) + 1198871198790]
119887 (120574 + 119887) 1198790
= 1 (B2)
12 Abstract and Applied Analysis
which indicates that there exists a1198790such that 1198771
0= 1 and the
disease-free periodic solution 120575 = (119878lowast(119905) 0) loses its stabilityFurther
120597Φ1(1198790 1198830)
120597119878= eminus1198871198790 gt 0
120597Φ2(1198790 1198830)
120597119868= eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
120597Φ1(1198790 1198830)
120597119868= minusint
1198790
0
120573119878lowast
(V) eminus119887(1198790minusV)eintV
0(120573119878lowast(119905)minus120574minus119887)d119905dV
≐ 119860 lt 0
1198861015840
0= 1 minus 119860
3eminus1198871198790
1198871015840
0= 1 minus 119860
3119860
1205972Φ2(1198790 1198830)
120597119868120597119878= 1205731198790eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
1205972Φ2(1198790 1198830)
1205971198682
= minus1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
119878lowast
(120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585minus119887(Vminus120579)
d120579 dV
lt 0
1205972Φ2(1198790 1198830)
120597119868120597119879= (120573119878
lowast
(1198790) minus 120574 minus 119887) eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
120597Φ1(1198790 1198830)
120597119879= minus119887 (119878 minus 1) eminus1198871198790
119861 = 1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887)
times eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
119862 = 1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
eminus119887(119905minus120579)119878lowast (120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585
d120579 dV
+ 21205731198790
1 minus 1198603119860
1 minus 1198603eminus1198871198790
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
(B3)
It follows that 119861119862 lt 0 and we get the condition that is if theparameters satisfy condition 119861 lt 0 then the model (4) has asupercritical branch at 119879
0
In fact
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0 (B4)
Therefore 119861 lt 0 is equivalent to
1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887) lt 0
(B5)
Obviously 119861 lt 0 due to (25) The proof is completed
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thiswork is supported by theNational Natural Science Foun-dation of China (NSFCs 11171199 11371030 11301320) andthe Fundamental Research Funds for the Central Universities(GK201305010) The authors would like to thank the editorand the referees for their careful reading of the original paperand many valuable comments and suggestions that greatlyimproved the presentation of this paper
References
[1] R M Anderson and R M May ldquoPopulation biology of infec-tious diseases part Irdquo Nature vol 280 no 5721 pp 361ndash3671979
[2] E Hanert E Schumacher and E Deleersnijder ldquoFront dynam-ics in fractional-order epidemic modelsrdquo Journal of TheoreticalBiology vol 279 no 1 pp 9ndash16 2011
[3] H W Hethcote ldquoMathematics of infectious diseasesrdquo SIAMReview vol 42 no 4 pp 599ndash653 2000
[4] A E Fiore C B Bridges and N J Cox ldquoSeasonal influenzavaccinesrdquo Current Topics in Microbiology and Immunology vol333 no 1 pp 43ndash82 2009
[5] Y Chang N T Brewer A C Rinas K Schmitt and J S SmithldquoEvaluating the impact of human papillomavirus vaccinesrdquoVaccine vol 27 no 32 pp 4355ndash4362 2009
[6] T J Liesegang ldquoVaricella zoster virus vaccines effective butconcerns lingerrdquo Canadian Journal of Ophthalmology vol 44no 4 pp 379ndash384 2009
[7] Z Agur L Cojocaru G Mazor R M Anderson and Y LDanon ldquoPulse mass measles vaccination across age cohortsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 90 no 24 pp 11698ndash11702 1993
[8] L Stone B Shulgin and Z Agur ldquoTheoretical examinationof the pulse vaccination policy in the SIR epidemic modelrdquoMathematical and ComputerModelling vol 31 no 4-5 pp 207ndash215 2000
[9] B Shulgin L Stone and Z Agur ldquoPulse vaccination strategy inthe SIR epidemic modelrdquo Bulletin of Mathematical Biology vol60 no 6 pp 1123ndash1148 1998
[10] A DrsquoOnofrio ldquoStability properties of pulse vaccination strategyin SEIR epidemicmodelrdquoMathematical Biosciences vol 179 no1 pp 57ndash72 2002
[11] A Franceschetti and A Pugliese ldquoThreshold behaviour of a SIRepidemicmodel with age structure and immigrationrdquo Journal ofMathematical Biology vol 57 no 1 pp 1ndash27 2008
Abstract and Applied Analysis 13
[12] A J Terry ldquoPulse vaccination strategies in a metapopulationSIR modelrdquo Mathematical Biosciences and Engineering vol 7no 2 pp 455ndash477 2010
[13] S J Gao L S Chen J J Nieto and A Torres ldquoAnalysis of adelayed epidemic model with pulse vaccination and saturationincidencerdquo Vaccine vol 24 no 35-36 pp 6037ndash6045 2006
[14] L Matrajt M E Halloran and I M Longini Jr ldquoOptimal vac-cine allocation for the early mitigation of pandemic influenzardquoPLoS Computational Biology vol 9 no 3 Article ID e10029642013
[15] S P Sullivan D G Koutsonanos M D P Martin et alldquoDissolving polymer microneedle patches for influenza vacci-nationrdquo Nature Medicine vol 16 no 8 pp 915ndash920 2010
[16] Science Daily Web ldquoVaccine-delivery patch with dissolvingmicroneedles eliminates ldquosharpsrdquo boosts protectionrdquo httpwwwsciencedailycomreleases201007100718204733htm
[17] M J Reynolds-Hogland J S Hogland and M S MitchellldquoEvaluating intercepts fromdemographicmodels to understandresource limitation and resource thresholdsrdquo Ecological Mod-elling vol 211 no 3-4 pp 424ndash432 2008
[18] L Chow M Fan and Z L Feng ldquoDynamics of a multigroupepidemiological model with group-targeted vaccination strate-giesrdquo Journal of Theoretical Biology vol 291 no 1 pp 56ndash642011
[19] L H Zhou and M Fan ldquoDynamics of an SIR epidemic modelwith limited medical resources revisitedrdquo Nonlinear AnalysisReal World Applications vol 13 no 1 pp 312ndash324 2012
[20] V Lakshmikantham D D Bainov and P S Simeonov Theoryof Impulsive Differential Equations World Scientific Singapore1989
[21] S M Blower and H Dowlatabadi ldquoSensitivity and uncertaintyanalysis of complex-models of disease transmission An HIVmodel as an examplerdquo International Statistical Review vol 62no 2 pp 229ndash243 1994
[22] S Marino I B Hogue C J Ray and D E Kirschner ldquoA meth-odology for performing global uncertainty and sensitivityanalysis in systems biologyrdquo Journal of Theoretical Biology vol254 no 1 pp 178ndash196 2008
[23] S Y Tang J H Liang Y S Tan and R A Cheke ldquoThresholdconditions for integrated pest management models with pesti-cides that have residual effectsrdquo Journal ofMathematical Biologyvol 66 no 1-2 pp 1ndash35 2013
[24] F Brauer and C Castillo-ChavezMathematical Models in Pop-ulation Biology and Epidemiology Springer New York NYUSA 2000
[25] O Diekmann and J A P Heesterbeek Mathematical Epidemi-ology of Infectious Diseases JohnWiley amp Sons Chichester UK2000
[26] J D MurrayMathematical Biology Springer Berlin Germany1989
[27] R Anderson and R May Infectious Diseases of HumansDynamics and Control Oxford University Press Oxford UK1995
[28] M Hethcote ldquoThree basic epidemiological modelsrdquo in AppliedMathematical Ecology L Gross T G Hallam and S A LevinEds pp 119ndash144 Springer Berlin Germany 1989
[29] A V Hill ldquoThe possible effects of the aggregation of themolecules of haemoglobin on its dissociation curvesrdquo TheJournal of Physiology vol 40 no 4 pp 4ndash7 1910
[30] P Cull ldquoLocal and global stability for population modelsrdquoBiological Cybernetics vol 54 no 3 pp 141ndash149 1986
[31] WHOWeb ldquoGlobal health observatory data repositoryrdquo 2001httpappswhointghodatatheme=country
[32] M D McKay R J Beckman and W J Conover ldquoComparisonof three methods for selecting values of input variables in theanalysis of output from a computer coderdquo Technometrics vol21 no 2 pp 239ndash245 1979
[33] W M Schaffer L F Olsen G L Truty S L Fulmer andD J Graser ldquoPeriodic and chaotic dynamics in childhoodepidemicsrdquo in From Chemical to Biological Organization MMarkus S Muller and G Nicolis Eds Springer BerlinGermany 1988
[34] A Lakmeche and O Arino ldquoBifurcation of non trivial periodicsolutions of impulsive differential equations arising chemother-apeutic treatmentrdquo Dynamics of Continuous Discrete andImpulsive Systems B vol 7 no 2 pp 265ndash287 2000
[35] S Y Tang and L S Chen ldquoMultiple attractors in stage-structured population models with birth pulsesrdquo Bulletin ofMathematical Biology vol 65 no 3 pp 479ndash495 2003
[36] S Y Tang Y N Xiao and R A Cheke ldquoMultiple attractorsof host-parasitoid models with integrated pest managementstrategies eradication persistence and outbreakrdquo TheoreticalPopulation Biology vol 73 no 2 pp 181ndash197 2008
[37] L F Olsen andWM Schaffer ldquoChaos versus noisy periodicityalternative hypotheses for childhood epidemicsrdquo Science vol249 no 4968 pp 499ndash504 1990
[38] G Sugihara and R M May ldquoNonlinear forecasting as a way ofdistinguishing chaos from measurement error in time seriesrdquoNature vol 344 no 6268 pp 734ndash741 1990
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical Problems in Engineering
Hindawi Publishing Corporationhttpwwwhindawicom
Differential EquationsInternational Journal of
Volume 2014
Applied MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Probability and StatisticsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical PhysicsAdvances in
Complex AnalysisJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
OptimizationJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CombinatoricsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Operations ResearchAdvances in
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Function Spaces
Abstract and Applied AnalysisHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of Mathematics and Mathematical Sciences
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Algebra
Discrete Dynamics in Nature and Society
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Decision SciencesAdvances in
Discrete MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014 Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Stochastic AnalysisInternational Journal of

Abstract and Applied Analysis 11
From Lemma 1 and the comparison theorem on impul-sive differential equations we have 119878(119905) le 119885(119905) and 119885(119905) rarr119878lowast(119905) as 119905 rarr +infin Hence there exists a 119905
1gt 0 such that
119878 (119905) le 119885 (119905) lt 119878lowast
(119905) + 1205761 (A13)
for all 119905 ge 1199051 here 120576
1gt 0 is small enough
From the second equation of model (4) and (A13) we get
d119868 (119905)d119905
le (120573 (119878lowast
(119905) + 1205761) minus 120574 minus 119887) 119868 (119905) (A14)
for 119905 gt 1199051 then we consider the following comparison
equation with pulses
d1199101(119905)
d119905= (120573 (119878
lowast
(119905) + 1205761) minus 120574 minus 119887) 119910
1(119905) 119905 = 119899119879
1199101(119905+
) = 1199101(119905) 119905 = 119899119879
(A15)
Integrating model (A15) between pulses (119899119879 (119899 + 1)119879] wehave
1199101((119899 + 1) 119879) = 119910
1(119899119879) eint
(119899+1)119879
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101(119899119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A16)
Then by using step by step iterations
1199101(119899119879) = 119910
1((119899 minus 1) 119879) eint
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= 1199101((119899 minus 2) 119879) e2 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
= sdot sdot sdot
= 1199101(0) e119899 int
119879
0[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
(A17)
where 1199101(0) = 119910
1(0+) gt 0 we obtain lim
119899rarrinfin1199101(119899119879) =
0 according to (A10) On the other hand integrating andsolving the first equation of model (A15) between pulses itgives
1199101(119905) = 119910
1(119899119879) eint
119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905 119905 isin (119899119879 (119899 + 1) 119879]
(A18)
incorporating into the boundedness of eint119905
119899119879[120573(119878lowast(119905)+1205761)minus120574minus119887]d119905
due to 11987710lt 1 we have lim
119905rarrinfin1199101(119905) = 0
Let (119878(119905) 119868(119905)) be any solution of model (4) with initialvalue (119878
0 1198680) and 119878
0= 119878(0
+) gt 0 119868
0= 119868(0
+) =
1199101(0) gt 0 according to the comparison theorem we have
lim119905rarrinfin
sup 119868(119905) le lim119905rarrinfin
sup1199101(119905) = 0 Incorporating into
the positivity of 119868(119905) we know that lim119905rarrinfin
119868(119905) = 0Consequently there exists a 119905
2gt 1199051such that 0 lt 119868(119905) le 120576
2
for 119905 ge 1199052 and 120576
2is small enough Further we have
119887 minus (1205731205762+ 119887) 119878 (119905) le
d119878 (119905)d119905
le 119887 minus 119887119878 (119905) (A19)
for all 119905 gt 1199052 from which we can obtain the following
equation
d1199102(119905)
d119905= 119887 minus (120573120576
2+ 119887) 119910
2(119905) 119905 = 119899119879
1199102(119905+
) =(1 minus 119901max) 119910
2
2(119905) + 120579119910
2(119905)
1199102(119905) + 120579
119905 = 119899119879
(A20)
By employing the same methods as used for the proof ofLemma 1 we get that the model (A20) has a positive periodicsolution 119910lowast
2(119905) which is globally attractive where
119910lowast
2(119905) = (119910
2minus
119887
1205731205762+ 119887) eminus(1205731205762+119887)(119905minus119899119879) + 119887
1205731205762+ 119887
119905 isin (119899119879 (119899 + 1) 119879]
1199102= 1199102(119899119879+
) = 119910lowast
2(0+
) =minus1198612+ radic11986122minus 411986021198622
21198602
1198602= eminus(1205731205762+119887)119879 [ 119887
1205731205762+ 119887
minus (1 minus 119901max) eminus(1205731205762+119887)119879]
1198612= (eminus(1205731205762+119887)119879 minus 119887
1205731205762+ 119887)
times [2 (1 minus 119901max) eminus(1205731205762+119887)119879minus (120579 +
119887
1205731205762+ 119887)]
1198622= minus[(1 minus 119901max) (
119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)2
+ 120579(119887
1205731205762+ 119887
minus eminus(1205731205762+119887)119879)]
(A21)
It follows from the comparison theorem on impulsive differ-ential equations that 119910
2(119905) le 119878(119905) le 119885(119905) Moreover 119910
2(119905) rarr
119910lowast
2(119905) and 119885(119905) rarr 119878
lowast(119905) as 119905 rarr +infin Consequently there
exists a 1199053for 1205763small enough such that 119905
3ge 1199052and
119910lowast
2(119905) minus 120576
3lt 119878 (119905) lt 119878
lowast
(119905) + 1205763
(A22)
for 119905 ge 1199053 Let 120576
2rarr 0 Then
119878lowast
(119905) minus 1205763lt 119878 (119905) lt 119878
lowast
(119905) + 1205763 (A23)
Therefore 119878(119905) rarr 119878lowast(119905) as 119905 rarr +infin Thus we
have proved the global stability of the disease-free periodicsolution (119878lowast(119905) 0) of model (4) This completes the proof
B Proof of Theorem 3
In order to apply the bifurcation theory of [34] in the maintext we make the following calculations
1198891015840
0= 1 minus eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
(B1)
If 11988910158400= 0 then 119879
0satisfies the following equation
120573 [(119878 minus 1) (1 minus eminus1198871198790) + 1198871198790]
119887 (120574 + 119887) 1198790
= 1 (B2)
12 Abstract and Applied Analysis
which indicates that there exists a1198790such that 1198771
0= 1 and the
disease-free periodic solution 120575 = (119878lowast(119905) 0) loses its stabilityFurther
120597Φ1(1198790 1198830)
120597119878= eminus1198871198790 gt 0
120597Φ2(1198790 1198830)
120597119868= eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
120597Φ1(1198790 1198830)
120597119868= minusint
1198790
0
120573119878lowast
(V) eminus119887(1198790minusV)eintV
0(120573119878lowast(119905)minus120574minus119887)d119905dV
≐ 119860 lt 0
1198861015840
0= 1 minus 119860
3eminus1198871198790
1198871015840
0= 1 minus 119860
3119860
1205972Φ2(1198790 1198830)
120597119868120597119878= 1205731198790eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
1205972Φ2(1198790 1198830)
1205971198682
= minus1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
119878lowast
(120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585minus119887(Vminus120579)
d120579 dV
lt 0
1205972Φ2(1198790 1198830)
120597119868120597119879= (120573119878
lowast
(1198790) minus 120574 minus 119887) eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
120597Φ1(1198790 1198830)
120597119879= minus119887 (119878 minus 1) eminus1198871198790
119861 = 1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887)
times eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
119862 = 1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
eminus119887(119905minus120579)119878lowast (120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585
d120579 dV
+ 21205731198790
1 minus 1198603119860
1 minus 1198603eminus1198871198790
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
(B3)
It follows that 119861119862 lt 0 and we get the condition that is if theparameters satisfy condition 119861 lt 0 then the model (4) has asupercritical branch at 119879
0
In fact
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0 (B4)
Therefore 119861 lt 0 is equivalent to
1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887) lt 0
(B5)
Obviously 119861 lt 0 due to (25) The proof is completed
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thiswork is supported by theNational Natural Science Foun-dation of China (NSFCs 11171199 11371030 11301320) andthe Fundamental Research Funds for the Central Universities(GK201305010) The authors would like to thank the editorand the referees for their careful reading of the original paperand many valuable comments and suggestions that greatlyimproved the presentation of this paper
References
[1] R M Anderson and R M May ldquoPopulation biology of infec-tious diseases part Irdquo Nature vol 280 no 5721 pp 361ndash3671979
[2] E Hanert E Schumacher and E Deleersnijder ldquoFront dynam-ics in fractional-order epidemic modelsrdquo Journal of TheoreticalBiology vol 279 no 1 pp 9ndash16 2011
[3] H W Hethcote ldquoMathematics of infectious diseasesrdquo SIAMReview vol 42 no 4 pp 599ndash653 2000
[4] A E Fiore C B Bridges and N J Cox ldquoSeasonal influenzavaccinesrdquo Current Topics in Microbiology and Immunology vol333 no 1 pp 43ndash82 2009
[5] Y Chang N T Brewer A C Rinas K Schmitt and J S SmithldquoEvaluating the impact of human papillomavirus vaccinesrdquoVaccine vol 27 no 32 pp 4355ndash4362 2009
[6] T J Liesegang ldquoVaricella zoster virus vaccines effective butconcerns lingerrdquo Canadian Journal of Ophthalmology vol 44no 4 pp 379ndash384 2009
[7] Z Agur L Cojocaru G Mazor R M Anderson and Y LDanon ldquoPulse mass measles vaccination across age cohortsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 90 no 24 pp 11698ndash11702 1993
[8] L Stone B Shulgin and Z Agur ldquoTheoretical examinationof the pulse vaccination policy in the SIR epidemic modelrdquoMathematical and ComputerModelling vol 31 no 4-5 pp 207ndash215 2000
[9] B Shulgin L Stone and Z Agur ldquoPulse vaccination strategy inthe SIR epidemic modelrdquo Bulletin of Mathematical Biology vol60 no 6 pp 1123ndash1148 1998
[10] A DrsquoOnofrio ldquoStability properties of pulse vaccination strategyin SEIR epidemicmodelrdquoMathematical Biosciences vol 179 no1 pp 57ndash72 2002
[11] A Franceschetti and A Pugliese ldquoThreshold behaviour of a SIRepidemicmodel with age structure and immigrationrdquo Journal ofMathematical Biology vol 57 no 1 pp 1ndash27 2008
Abstract and Applied Analysis 13
[12] A J Terry ldquoPulse vaccination strategies in a metapopulationSIR modelrdquo Mathematical Biosciences and Engineering vol 7no 2 pp 455ndash477 2010
[13] S J Gao L S Chen J J Nieto and A Torres ldquoAnalysis of adelayed epidemic model with pulse vaccination and saturationincidencerdquo Vaccine vol 24 no 35-36 pp 6037ndash6045 2006
[14] L Matrajt M E Halloran and I M Longini Jr ldquoOptimal vac-cine allocation for the early mitigation of pandemic influenzardquoPLoS Computational Biology vol 9 no 3 Article ID e10029642013
[15] S P Sullivan D G Koutsonanos M D P Martin et alldquoDissolving polymer microneedle patches for influenza vacci-nationrdquo Nature Medicine vol 16 no 8 pp 915ndash920 2010
[16] Science Daily Web ldquoVaccine-delivery patch with dissolvingmicroneedles eliminates ldquosharpsrdquo boosts protectionrdquo httpwwwsciencedailycomreleases201007100718204733htm
[17] M J Reynolds-Hogland J S Hogland and M S MitchellldquoEvaluating intercepts fromdemographicmodels to understandresource limitation and resource thresholdsrdquo Ecological Mod-elling vol 211 no 3-4 pp 424ndash432 2008
[18] L Chow M Fan and Z L Feng ldquoDynamics of a multigroupepidemiological model with group-targeted vaccination strate-giesrdquo Journal of Theoretical Biology vol 291 no 1 pp 56ndash642011
[19] L H Zhou and M Fan ldquoDynamics of an SIR epidemic modelwith limited medical resources revisitedrdquo Nonlinear AnalysisReal World Applications vol 13 no 1 pp 312ndash324 2012
[20] V Lakshmikantham D D Bainov and P S Simeonov Theoryof Impulsive Differential Equations World Scientific Singapore1989
[21] S M Blower and H Dowlatabadi ldquoSensitivity and uncertaintyanalysis of complex-models of disease transmission An HIVmodel as an examplerdquo International Statistical Review vol 62no 2 pp 229ndash243 1994
[22] S Marino I B Hogue C J Ray and D E Kirschner ldquoA meth-odology for performing global uncertainty and sensitivityanalysis in systems biologyrdquo Journal of Theoretical Biology vol254 no 1 pp 178ndash196 2008
[23] S Y Tang J H Liang Y S Tan and R A Cheke ldquoThresholdconditions for integrated pest management models with pesti-cides that have residual effectsrdquo Journal ofMathematical Biologyvol 66 no 1-2 pp 1ndash35 2013
[24] F Brauer and C Castillo-ChavezMathematical Models in Pop-ulation Biology and Epidemiology Springer New York NYUSA 2000
[25] O Diekmann and J A P Heesterbeek Mathematical Epidemi-ology of Infectious Diseases JohnWiley amp Sons Chichester UK2000
[26] J D MurrayMathematical Biology Springer Berlin Germany1989
[27] R Anderson and R May Infectious Diseases of HumansDynamics and Control Oxford University Press Oxford UK1995
[28] M Hethcote ldquoThree basic epidemiological modelsrdquo in AppliedMathematical Ecology L Gross T G Hallam and S A LevinEds pp 119ndash144 Springer Berlin Germany 1989
[29] A V Hill ldquoThe possible effects of the aggregation of themolecules of haemoglobin on its dissociation curvesrdquo TheJournal of Physiology vol 40 no 4 pp 4ndash7 1910
[30] P Cull ldquoLocal and global stability for population modelsrdquoBiological Cybernetics vol 54 no 3 pp 141ndash149 1986
[31] WHOWeb ldquoGlobal health observatory data repositoryrdquo 2001httpappswhointghodatatheme=country
[32] M D McKay R J Beckman and W J Conover ldquoComparisonof three methods for selecting values of input variables in theanalysis of output from a computer coderdquo Technometrics vol21 no 2 pp 239ndash245 1979
[33] W M Schaffer L F Olsen G L Truty S L Fulmer andD J Graser ldquoPeriodic and chaotic dynamics in childhoodepidemicsrdquo in From Chemical to Biological Organization MMarkus S Muller and G Nicolis Eds Springer BerlinGermany 1988
[34] A Lakmeche and O Arino ldquoBifurcation of non trivial periodicsolutions of impulsive differential equations arising chemother-apeutic treatmentrdquo Dynamics of Continuous Discrete andImpulsive Systems B vol 7 no 2 pp 265ndash287 2000
[35] S Y Tang and L S Chen ldquoMultiple attractors in stage-structured population models with birth pulsesrdquo Bulletin ofMathematical Biology vol 65 no 3 pp 479ndash495 2003
[36] S Y Tang Y N Xiao and R A Cheke ldquoMultiple attractorsof host-parasitoid models with integrated pest managementstrategies eradication persistence and outbreakrdquo TheoreticalPopulation Biology vol 73 no 2 pp 181ndash197 2008
[37] L F Olsen andWM Schaffer ldquoChaos versus noisy periodicityalternative hypotheses for childhood epidemicsrdquo Science vol249 no 4968 pp 499ndash504 1990
[38] G Sugihara and R M May ldquoNonlinear forecasting as a way ofdistinguishing chaos from measurement error in time seriesrdquoNature vol 344 no 6268 pp 734ndash741 1990
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical Problems in Engineering
Hindawi Publishing Corporationhttpwwwhindawicom
Differential EquationsInternational Journal of
Volume 2014
Applied MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Probability and StatisticsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical PhysicsAdvances in
Complex AnalysisJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
OptimizationJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CombinatoricsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Operations ResearchAdvances in
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Function Spaces
Abstract and Applied AnalysisHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of Mathematics and Mathematical Sciences
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Algebra
Discrete Dynamics in Nature and Society
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Decision SciencesAdvances in
Discrete MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014 Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Stochastic AnalysisInternational Journal of

12 Abstract and Applied Analysis
which indicates that there exists a1198790such that 1198771
0= 1 and the
disease-free periodic solution 120575 = (119878lowast(119905) 0) loses its stabilityFurther
120597Φ1(1198790 1198830)
120597119878= eminus1198871198790 gt 0
120597Φ2(1198790 1198830)
120597119868= eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
120597Φ1(1198790 1198830)
120597119868= minusint
1198790
0
120573119878lowast
(V) eminus119887(1198790minusV)eintV
0(120573119878lowast(119905)minus120574minus119887)d119905dV
≐ 119860 lt 0
1198861015840
0= 1 minus 119860
3eminus1198871198790
1198871015840
0= 1 minus 119860
3119860
1205972Φ2(1198790 1198830)
120597119868120597119878= 1205731198790eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
1205972Φ2(1198790 1198830)
1205971198682
= minus1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
119878lowast
(120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585minus119887(Vminus120579)
d120579 dV
lt 0
1205972Φ2(1198790 1198830)
120597119868120597119879= (120573119878
lowast
(1198790) minus 120574 minus 119887) eint
1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
120597Φ1(1198790 1198830)
120597119879= minus119887 (119878 minus 1) eminus1198871198790
119861 = 1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887)
times eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
119862 = 1205732
int
1198790
0
eint1198790
V(120573119878lowast(120585)minus120574minus119887)d120585
times int
V
0
eminus119887(119905minus120579)119878lowast (120579) eint120579
0(120573119878lowast(120585)minus120574minus119887)d120585
d120579 dV
+ 21205731198790
1 minus 1198603119860
1 minus 1198603eminus1198871198790
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0
(B3)
It follows that 119861119862 lt 0 and we get the condition that is if theparameters satisfy condition 119861 lt 0 then the model (4) has asupercritical branch at 119879
0
In fact
eint1198790
0(120573119878lowast(119905)minus120574minus119887)d119905
gt 0 (B4)
Therefore 119861 lt 0 is equivalent to
1205731198871198790
1198603eminus1198871198790
1 minus 1198603eminus1198871198790
(119878 (1198790) minus 1) minus (120573119878
lowast
(1198790) minus 120574 minus 119887) lt 0
(B5)
Obviously 119861 lt 0 due to (25) The proof is completed
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thiswork is supported by theNational Natural Science Foun-dation of China (NSFCs 11171199 11371030 11301320) andthe Fundamental Research Funds for the Central Universities(GK201305010) The authors would like to thank the editorand the referees for their careful reading of the original paperand many valuable comments and suggestions that greatlyimproved the presentation of this paper
References
[1] R M Anderson and R M May ldquoPopulation biology of infec-tious diseases part Irdquo Nature vol 280 no 5721 pp 361ndash3671979
[2] E Hanert E Schumacher and E Deleersnijder ldquoFront dynam-ics in fractional-order epidemic modelsrdquo Journal of TheoreticalBiology vol 279 no 1 pp 9ndash16 2011
[3] H W Hethcote ldquoMathematics of infectious diseasesrdquo SIAMReview vol 42 no 4 pp 599ndash653 2000
[4] A E Fiore C B Bridges and N J Cox ldquoSeasonal influenzavaccinesrdquo Current Topics in Microbiology and Immunology vol333 no 1 pp 43ndash82 2009
[5] Y Chang N T Brewer A C Rinas K Schmitt and J S SmithldquoEvaluating the impact of human papillomavirus vaccinesrdquoVaccine vol 27 no 32 pp 4355ndash4362 2009
[6] T J Liesegang ldquoVaricella zoster virus vaccines effective butconcerns lingerrdquo Canadian Journal of Ophthalmology vol 44no 4 pp 379ndash384 2009
[7] Z Agur L Cojocaru G Mazor R M Anderson and Y LDanon ldquoPulse mass measles vaccination across age cohortsrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 90 no 24 pp 11698ndash11702 1993
[8] L Stone B Shulgin and Z Agur ldquoTheoretical examinationof the pulse vaccination policy in the SIR epidemic modelrdquoMathematical and ComputerModelling vol 31 no 4-5 pp 207ndash215 2000
[9] B Shulgin L Stone and Z Agur ldquoPulse vaccination strategy inthe SIR epidemic modelrdquo Bulletin of Mathematical Biology vol60 no 6 pp 1123ndash1148 1998
[10] A DrsquoOnofrio ldquoStability properties of pulse vaccination strategyin SEIR epidemicmodelrdquoMathematical Biosciences vol 179 no1 pp 57ndash72 2002
[11] A Franceschetti and A Pugliese ldquoThreshold behaviour of a SIRepidemicmodel with age structure and immigrationrdquo Journal ofMathematical Biology vol 57 no 1 pp 1ndash27 2008
Abstract and Applied Analysis 13
[12] A J Terry ldquoPulse vaccination strategies in a metapopulationSIR modelrdquo Mathematical Biosciences and Engineering vol 7no 2 pp 455ndash477 2010
[13] S J Gao L S Chen J J Nieto and A Torres ldquoAnalysis of adelayed epidemic model with pulse vaccination and saturationincidencerdquo Vaccine vol 24 no 35-36 pp 6037ndash6045 2006
[14] L Matrajt M E Halloran and I M Longini Jr ldquoOptimal vac-cine allocation for the early mitigation of pandemic influenzardquoPLoS Computational Biology vol 9 no 3 Article ID e10029642013
[15] S P Sullivan D G Koutsonanos M D P Martin et alldquoDissolving polymer microneedle patches for influenza vacci-nationrdquo Nature Medicine vol 16 no 8 pp 915ndash920 2010
[16] Science Daily Web ldquoVaccine-delivery patch with dissolvingmicroneedles eliminates ldquosharpsrdquo boosts protectionrdquo httpwwwsciencedailycomreleases201007100718204733htm
[17] M J Reynolds-Hogland J S Hogland and M S MitchellldquoEvaluating intercepts fromdemographicmodels to understandresource limitation and resource thresholdsrdquo Ecological Mod-elling vol 211 no 3-4 pp 424ndash432 2008
[18] L Chow M Fan and Z L Feng ldquoDynamics of a multigroupepidemiological model with group-targeted vaccination strate-giesrdquo Journal of Theoretical Biology vol 291 no 1 pp 56ndash642011
[19] L H Zhou and M Fan ldquoDynamics of an SIR epidemic modelwith limited medical resources revisitedrdquo Nonlinear AnalysisReal World Applications vol 13 no 1 pp 312ndash324 2012
[20] V Lakshmikantham D D Bainov and P S Simeonov Theoryof Impulsive Differential Equations World Scientific Singapore1989
[21] S M Blower and H Dowlatabadi ldquoSensitivity and uncertaintyanalysis of complex-models of disease transmission An HIVmodel as an examplerdquo International Statistical Review vol 62no 2 pp 229ndash243 1994
[22] S Marino I B Hogue C J Ray and D E Kirschner ldquoA meth-odology for performing global uncertainty and sensitivityanalysis in systems biologyrdquo Journal of Theoretical Biology vol254 no 1 pp 178ndash196 2008
[23] S Y Tang J H Liang Y S Tan and R A Cheke ldquoThresholdconditions for integrated pest management models with pesti-cides that have residual effectsrdquo Journal ofMathematical Biologyvol 66 no 1-2 pp 1ndash35 2013
[24] F Brauer and C Castillo-ChavezMathematical Models in Pop-ulation Biology and Epidemiology Springer New York NYUSA 2000
[25] O Diekmann and J A P Heesterbeek Mathematical Epidemi-ology of Infectious Diseases JohnWiley amp Sons Chichester UK2000
[26] J D MurrayMathematical Biology Springer Berlin Germany1989
[27] R Anderson and R May Infectious Diseases of HumansDynamics and Control Oxford University Press Oxford UK1995
[28] M Hethcote ldquoThree basic epidemiological modelsrdquo in AppliedMathematical Ecology L Gross T G Hallam and S A LevinEds pp 119ndash144 Springer Berlin Germany 1989
[29] A V Hill ldquoThe possible effects of the aggregation of themolecules of haemoglobin on its dissociation curvesrdquo TheJournal of Physiology vol 40 no 4 pp 4ndash7 1910
[30] P Cull ldquoLocal and global stability for population modelsrdquoBiological Cybernetics vol 54 no 3 pp 141ndash149 1986
[31] WHOWeb ldquoGlobal health observatory data repositoryrdquo 2001httpappswhointghodatatheme=country
[32] M D McKay R J Beckman and W J Conover ldquoComparisonof three methods for selecting values of input variables in theanalysis of output from a computer coderdquo Technometrics vol21 no 2 pp 239ndash245 1979
[33] W M Schaffer L F Olsen G L Truty S L Fulmer andD J Graser ldquoPeriodic and chaotic dynamics in childhoodepidemicsrdquo in From Chemical to Biological Organization MMarkus S Muller and G Nicolis Eds Springer BerlinGermany 1988
[34] A Lakmeche and O Arino ldquoBifurcation of non trivial periodicsolutions of impulsive differential equations arising chemother-apeutic treatmentrdquo Dynamics of Continuous Discrete andImpulsive Systems B vol 7 no 2 pp 265ndash287 2000
[35] S Y Tang and L S Chen ldquoMultiple attractors in stage-structured population models with birth pulsesrdquo Bulletin ofMathematical Biology vol 65 no 3 pp 479ndash495 2003
[36] S Y Tang Y N Xiao and R A Cheke ldquoMultiple attractorsof host-parasitoid models with integrated pest managementstrategies eradication persistence and outbreakrdquo TheoreticalPopulation Biology vol 73 no 2 pp 181ndash197 2008
[37] L F Olsen andWM Schaffer ldquoChaos versus noisy periodicityalternative hypotheses for childhood epidemicsrdquo Science vol249 no 4968 pp 499ndash504 1990
[38] G Sugihara and R M May ldquoNonlinear forecasting as a way ofdistinguishing chaos from measurement error in time seriesrdquoNature vol 344 no 6268 pp 734ndash741 1990
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical Problems in Engineering
Hindawi Publishing Corporationhttpwwwhindawicom
Differential EquationsInternational Journal of
Volume 2014
Applied MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Probability and StatisticsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical PhysicsAdvances in
Complex AnalysisJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
OptimizationJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CombinatoricsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Operations ResearchAdvances in
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Function Spaces
Abstract and Applied AnalysisHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of Mathematics and Mathematical Sciences
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Algebra
Discrete Dynamics in Nature and Society
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Decision SciencesAdvances in
Discrete MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014 Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Stochastic AnalysisInternational Journal of

Abstract and Applied Analysis 13
[12] A J Terry ldquoPulse vaccination strategies in a metapopulationSIR modelrdquo Mathematical Biosciences and Engineering vol 7no 2 pp 455ndash477 2010
[13] S J Gao L S Chen J J Nieto and A Torres ldquoAnalysis of adelayed epidemic model with pulse vaccination and saturationincidencerdquo Vaccine vol 24 no 35-36 pp 6037ndash6045 2006
[14] L Matrajt M E Halloran and I M Longini Jr ldquoOptimal vac-cine allocation for the early mitigation of pandemic influenzardquoPLoS Computational Biology vol 9 no 3 Article ID e10029642013
[15] S P Sullivan D G Koutsonanos M D P Martin et alldquoDissolving polymer microneedle patches for influenza vacci-nationrdquo Nature Medicine vol 16 no 8 pp 915ndash920 2010
[16] Science Daily Web ldquoVaccine-delivery patch with dissolvingmicroneedles eliminates ldquosharpsrdquo boosts protectionrdquo httpwwwsciencedailycomreleases201007100718204733htm
[17] M J Reynolds-Hogland J S Hogland and M S MitchellldquoEvaluating intercepts fromdemographicmodels to understandresource limitation and resource thresholdsrdquo Ecological Mod-elling vol 211 no 3-4 pp 424ndash432 2008
[18] L Chow M Fan and Z L Feng ldquoDynamics of a multigroupepidemiological model with group-targeted vaccination strate-giesrdquo Journal of Theoretical Biology vol 291 no 1 pp 56ndash642011
[19] L H Zhou and M Fan ldquoDynamics of an SIR epidemic modelwith limited medical resources revisitedrdquo Nonlinear AnalysisReal World Applications vol 13 no 1 pp 312ndash324 2012
[20] V Lakshmikantham D D Bainov and P S Simeonov Theoryof Impulsive Differential Equations World Scientific Singapore1989
[21] S M Blower and H Dowlatabadi ldquoSensitivity and uncertaintyanalysis of complex-models of disease transmission An HIVmodel as an examplerdquo International Statistical Review vol 62no 2 pp 229ndash243 1994
[22] S Marino I B Hogue C J Ray and D E Kirschner ldquoA meth-odology for performing global uncertainty and sensitivityanalysis in systems biologyrdquo Journal of Theoretical Biology vol254 no 1 pp 178ndash196 2008
[23] S Y Tang J H Liang Y S Tan and R A Cheke ldquoThresholdconditions for integrated pest management models with pesti-cides that have residual effectsrdquo Journal ofMathematical Biologyvol 66 no 1-2 pp 1ndash35 2013
[24] F Brauer and C Castillo-ChavezMathematical Models in Pop-ulation Biology and Epidemiology Springer New York NYUSA 2000
[25] O Diekmann and J A P Heesterbeek Mathematical Epidemi-ology of Infectious Diseases JohnWiley amp Sons Chichester UK2000
[26] J D MurrayMathematical Biology Springer Berlin Germany1989
[27] R Anderson and R May Infectious Diseases of HumansDynamics and Control Oxford University Press Oxford UK1995
[28] M Hethcote ldquoThree basic epidemiological modelsrdquo in AppliedMathematical Ecology L Gross T G Hallam and S A LevinEds pp 119ndash144 Springer Berlin Germany 1989
[29] A V Hill ldquoThe possible effects of the aggregation of themolecules of haemoglobin on its dissociation curvesrdquo TheJournal of Physiology vol 40 no 4 pp 4ndash7 1910
[30] P Cull ldquoLocal and global stability for population modelsrdquoBiological Cybernetics vol 54 no 3 pp 141ndash149 1986
[31] WHOWeb ldquoGlobal health observatory data repositoryrdquo 2001httpappswhointghodatatheme=country
[32] M D McKay R J Beckman and W J Conover ldquoComparisonof three methods for selecting values of input variables in theanalysis of output from a computer coderdquo Technometrics vol21 no 2 pp 239ndash245 1979
[33] W M Schaffer L F Olsen G L Truty S L Fulmer andD J Graser ldquoPeriodic and chaotic dynamics in childhoodepidemicsrdquo in From Chemical to Biological Organization MMarkus S Muller and G Nicolis Eds Springer BerlinGermany 1988
[34] A Lakmeche and O Arino ldquoBifurcation of non trivial periodicsolutions of impulsive differential equations arising chemother-apeutic treatmentrdquo Dynamics of Continuous Discrete andImpulsive Systems B vol 7 no 2 pp 265ndash287 2000
[35] S Y Tang and L S Chen ldquoMultiple attractors in stage-structured population models with birth pulsesrdquo Bulletin ofMathematical Biology vol 65 no 3 pp 479ndash495 2003
[36] S Y Tang Y N Xiao and R A Cheke ldquoMultiple attractorsof host-parasitoid models with integrated pest managementstrategies eradication persistence and outbreakrdquo TheoreticalPopulation Biology vol 73 no 2 pp 181ndash197 2008
[37] L F Olsen andWM Schaffer ldquoChaos versus noisy periodicityalternative hypotheses for childhood epidemicsrdquo Science vol249 no 4968 pp 499ndash504 1990
[38] G Sugihara and R M May ldquoNonlinear forecasting as a way ofdistinguishing chaos from measurement error in time seriesrdquoNature vol 344 no 6268 pp 734ndash741 1990
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical Problems in Engineering
Hindawi Publishing Corporationhttpwwwhindawicom
Differential EquationsInternational Journal of
Volume 2014
Applied MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Probability and StatisticsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical PhysicsAdvances in
Complex AnalysisJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
OptimizationJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CombinatoricsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Operations ResearchAdvances in
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Function Spaces
Abstract and Applied AnalysisHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of Mathematics and Mathematical Sciences
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Algebra
Discrete Dynamics in Nature and Society
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Decision SciencesAdvances in
Discrete MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014 Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Stochastic AnalysisInternational Journal of

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical Problems in Engineering
Hindawi Publishing Corporationhttpwwwhindawicom
Differential EquationsInternational Journal of
Volume 2014
Applied MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Probability and StatisticsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Mathematical PhysicsAdvances in
Complex AnalysisJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
OptimizationJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CombinatoricsHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Operations ResearchAdvances in
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Function Spaces
Abstract and Applied AnalysisHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of Mathematics and Mathematical Sciences
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Algebra
Discrete Dynamics in Nature and Society
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Decision SciencesAdvances in
Discrete MathematicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014 Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Stochastic AnalysisInternational Journal of


















