
Effects of hypoxia on human cancer cell line chemosensitivity

Research ArticleMetabolic Effects of Hypoxia in Colorectal Cancer by13C NMR Isotopomer Analysis
Ana M Abrantes12 Ludgero C Tavares3 Salomeacute Pires14
Joatildeo Casalta-Lopes15 Cacircndida Mendes6 Marta Simotildees6 Manuela M Grazina67
Rui A Carvalho37 and Maria Filomena Botelho12
1 Biophysics Unit IBILI Faculty of Medicine University of Coimbra 3000-548 Coimbra Portugal2 Centre of Investigation on Environment Genetics and Oncobiology (CIMAGO) Faculty of Medicine University of Coimbra3001-301 Coimbra Portugal
3 Life Sciences Department Faculty of Sciences and Technology University of Coimbra 3000-456 Coimbra Portugal4 Faculty of Sciences and Technology University of Coimbra 3000-456 Coimbra Portugal5 Radiotherapy Department CHUC 3000-075 Coimbra Portugal6 Laboratory of Biochemical Genetics (CNCUC) Faculty of Medicine University of Coimbra 3000-548 Coimbra Portugal7 Center for Neuroscience and Cell Biology (CNC) 3004-517 Coimbra Portugal
Correspondence should be addressed to Ana M Abrantes mabrantesfmeducpt and Rui A Carvalho carvalhociucpt
Received 28 February 2014 Accepted 27 May 2014 Published 1 July 2014
Academic Editor Zhiqiang Meng
Copyright copy 2014 Ana M Abrantes et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
13C NMR isotopomer analysis was used to characterize intermediary metabolism in three colorectal cancer cell lines (WiDrLS1034 and C2BBe1) and determine the ldquometabolic remodelingrdquo that occurs under hypoxia Under normoxia the three colorectalcancer cell lines present high rates of lactate production and can be seen as ldquoWarburgrdquo like cancer cells independently of substrateavailability since such profile was dominant at both high and low glucose media contents The LS1034 was the less glycolytic of thethree cell lines and was the most affected by the event of hypoxia raising abruptly glucose consumption and lactate productionThe other two colorectal cell lines WiDr and C2BBe1 adapted better to hypoxia and were able to maintain their oxidative fluxeseven at the very low levels of oxygen These differential metabolic behaviors of the three colorectal cell lines show how importantan adequate knowledge of the ldquometabolic remodelingrdquo that follows a given cancer treatment is towards the correct (re)design oftherapeutic strategies against cancer
1 Introduction
Cancer cell alters its metabolism in response to a challengingenvironment by promoting cell growth and proliferationdiverging significantly from normal tissues According toOtto Warburg cancer cells maintain high aerobic glycolyticrates and produce high levels of lactate and pyruvate [1ndash3] tosustain cell proliferation and its high energy demands Whenthe pO
2is normal the oxidative phosphorylation process
occurs and pyruvate is directed towards theKrebs cycleThusmetabolism in ldquoWarburgrdquo like tumor cells could at first beseen as ldquowastefulrdquo when compared to normal cells or wecould say that tumor cells use a disproportionate nutrient
exchange with its environment This metabolic profile is infact prevalent inmany cancer cells and grants them advantageover normal cells by allowing them to proliferate at muchhigher rates By avoiding oxidative phosphorylation evenwhen oxygen is plentiful [4] and adopting aerobic glycolysiscarbon skeletons build up considerably in their cytosol andbiosynthetic pathways are efficiently activated Several studieshave demonstrated an increase in the contents of several gly-colytic enzymes such as 6-phosphofructo-2-kinasefructose26-biphosphatases The regulation of glycolysis by theseenzymes allows the alterations in glycolytic fluxes requiredto fulfill cancer cells bioenergetics and biosynthetic demandsThe glycolytic pathway is in fact becoming an increasing
Hindawi Publishing CorporationBioMed Research InternationalVolume 2014 Article ID 759791 10 pageshttpdxdoiorg1011552014759791
2 BioMed Research International
target in cancer therapy both by itself and in combinationwith other therapies such as immunotherapy This advancehelps overcoming drug resistance issues and improves theefficacy of current anticancer agents [3 5ndash7]
Besides that and having into account tumor microen-vironment due to the temporal and spatial heterogeneityof oxygenation that occurs in solid tumors the adaptationto the variability of its microenvironment is critical Oxy-gen supply is impaired in many tumors because there isimbalance between tissue growth and the development ofnew vasculature In solid tumors hypoxia is thus a commoncharacteristicmicroenvironment of tumoral cells becominga key factor for tumor progression and resistance to anti-cancer therapy [8] This decrease in oxygen pO
2influences
compensatory physiological events involving adaptations atall levels in order to maintain homeostasis between cellsenergetic requirements and supplies [8] Thus it appears thataerobic glycolysis is an adaptive mechanism that involvesseveral metabolic pathways coordinates which maintainthe morphological characteristics of tumor cells includingthe ability to survive hypoxic conditions the capacity ofmetastasis and evasion of death by apoptosis [5ndash8]
Solid tumors have also heterogeneous populations of cellsdue in part to a limited blood supply that provides reducedlevels of oxygen and prompts for acidic conditions and avidityof glucose [9]These changes in the tumormicroenvironmentmay represent physiological signals that activate cell survivalor death by apoptosis affecting the balance between growthand tumor suppression The mechanisms by which tumorcells adapt or die in the presence of low levels of oxygenare not well studied and understood However it is knownthat the expression of several transcription factors as wellas the modification of metabolic pathways interfere with theresponse to the lack of oxygen and nutrients by tumor cells[10]
The aim of this study is to characterize the metabolicprofile namely glycolysis and Krebs cycle fluxes of threecolorectal cancer cell lines using carbon-13 (13C) tracers andnuclear magnetic resonance (NMR) spectroscopy With thisapproach central metabolic pathways will be evaluated andmajor metabolic changes resulting from hypoxia and glucoseavailability will be determined towards depicting the possibleinvolvement of metabolic mechanisms [11] in some processesof chemotherapeutic resistance in colon cancer [12]
2 Materials and Methods
21 Cell Lines Colorectal cancer cell lines (WiDr LS1034 andC2BBe1) purchased from American Type Culture Collection(ATCC Rockville MD USA) were used to perform invitro studies The three cell lines are from different colonlocalizations WiDr from rectosigmoid location C2BBe1clone of the cell line Caco-2 (ATCC) from descendingcolon and LS1034 resistant to chemotherapy concerning P-glycoprotein overexpression from ascending colon [9ndash13]WiDr and C2BBe1 cell lines were maintained in DulbeccorsquosModified Eaglersquos cell culture medium (Sigma) and LS1034cell line was maintained in Roswell Park Memorial Institute(RPMI)medium All cell lines were cultured in high (25mM)
and low (5mM) glucose concentrations supplemented with10 fetal calf serum (Gibco) in 5 CO
2atmosphere at
37∘C To perform hypoxia studies cells were incubated at37∘C in 93 N
2 2 O
2 and 5 CO
2using a controlled-
environment cabinet (PlasLabs USA) Each of the mediawas acquired without glucose in order to be supplementedwith [U-13C]glucose for performing 13C NMR isotopomeranalysis
22 Cell Perchloric Acid Extracts Perchloric acid extractswere obtained using published methods [14 15] Briefly tothe cell pellet is added the equivalent (vw) of 2mL ice-cold perchloric acid 72gram wet weight of cells Theresulting supernatant is subsequently neutralized using ice-cold KOH solutions before lyophilization This procedureallows precipitation of KClO
4and a considerable reduction
of solution ionic strength towards the acquisition of highquality NMR spectra For kinetic analysis of lactate produc-tion aliquots of cell culture media (160 120583L) were drawn atspecific time intervals (0 4 and 8 hours) To each of thesealiquots 40 120583L of an internal standard consisting of sodiumfumarate (10mM) in phosphate buffer (100mM) was addedfor absolute quantification purposes
23 NMR Analyses of Intermediary Metabolism
231 Glycolytic Fluxes by Analysis of Cell Culture MediaProton (1H) nuclear magnetic resonance (NMR) spectraof cell culture media were acquired on a 600MHz VarianNMR spectrometer using a 3mm indirect detection probeTypical acquisition parameters included a 3 s acquisitiontime a radiofrequency pulse of 45∘ and an interpulse delayof 10 seconds to ensure full relaxation of all nuclei in thesample Due to direct ( 1119869HC) and long range (
2119869HC and 3119869HC)
heteronuclear scalar coupling it is possible to distinguish theresonances of [U-13C]lactate from those due to nonenriched[U-12C]lactate (Figure 1) [U-13C]lactate originates from [U-13C]glucose as a result of glycolysis followed by reductionof pyruvate by lactate dehydrogenase (LDH) Levels of [U-13C]lactate are thus used as an indirect measure of glycolyticactivity
232 Krebs Cycle Kinetics by 1H and 13C NMR Analysisof Cell Extracts 1H- and 13C-NMR spectra of cell extractswere acquired in a 600MHz Varian NMR spectrometerusing a 3mm indirect detection and a 3mm broadbandNMR probe respectively 1H-NMR spectra of cell extractsallow the monitoring of the levels of intracellular metabolicintermediates while 13C-NMR spectra allows tackling thedynamics of 13C incorporation in metabolic intermediatesthus warranting a dynamic vision of cell metabolism Ratesof 13C incorporation in glycolytic and Krebs cycle interme-diates are used as a measure of metabolic coupling betweenthese two major metabolic pathways and provide valuableclues about the ldquometabolic remodelingrdquo associated with thetumorigenic transformation and treatment 13C isotopomersof intracellular glutamate provide a window for monitoring
BioMed Research International 3
1 2 3 4 5 6
1
11
2
22
3
33
4 5
F1
F2
F3F4Alanine
OAACitrate
120572-Ketoglutarate
Succinyl-CoA Glutamate
Glucose
Lactate
3JHH
2JHC
3JHC
1st turn D45
2nd turn Q
D45D45
Q QQ Q
Figure 1 [U-13C]glucose as a carbon tracer for monitoring glycolysis and Krebs cycle Through glycolysis and lactic fermentation [U-13C]lactate is produced which is distinguishable (multiplets in each 13C satellite due to 2119869HC
3119869HC and
3119869HHmdashduplet of triplets since 2119869HC
and 3119869HC have similar magnitude) from tissue lactate or lactate originating from unenriched carbon sources [12-13C2]acetil-CoA originating
from [U-13C]pyruvate leads to incorporation of 13C atoms in Krebs cycle intermediates and other metabolites that exchange with those (egglutamate) It is possible to follow Krebs cycle kinetics by monitoring the 13C multiplet due to carbon 4 of glutamate The duplet component(D45) results from the simple entrance of [12-13C
2]acetil-CoA but the quartet requires further cycling and combination of [12-13C
2]acetil-
CoA with previously enriched oxaloacetate (OAA) A simple ratio of QD45is proportional to Krebs cycle flux and denotes higher rates of
oxidative metabolism in tissues as opposed to glycolysis Metabolic fluxes F1ndash4 denote the major pathways for [U-13C]glucose use
Krebs cycle kinetics (Figure 1) since the appearance of multi-13C-labeled species clearly indicate cycle turnover [14 16 17]and oxidative efficiency
24 Citrate Synthase Enzyme Activity The citrate synthaseenzyme (CS) is used as a marker enzyme of mitochondrialpreparations due to its stability and its regulatory role in theKrebs cycle Its activity can provide an estimate of the numberof mitochondria in a cell suspension and can be used in orderto standardize the results of the activities of the enzymes ofthe mitochondrial respiratory chain [15 18] The evaluationof the activity of CS was performed by spectrophotometry ata 120582 of 412 nmThis assay consists of the condensation reactionof acetyl-CoA (02mM) with oxaloacetate (8mM) catalyzedby CSThis reaction results in coenzymeA (CoA) release thatreacts with 551015840-dithiobis(-nitrobenzoic acid) (DNTB) addedto the medium at a concentration of 2mM thus enabling thereading at 412 nm To allow the access of substrates to theenzyme Triton X-100 was used at 01
25 Complex IV Activity To measure complex IV activity50120583g of protein was added to 1mL of buffer composed by10mM KH
2PO4 300mM sucrose and 5mgmL BSA with
pH 65 which was in a cuvette The analysis started with thecompletion of a baseline tracing at 120582 550 nm and then 10 120583Lwas added of detergent n-dodecyl-120573-D-maltoside whichpromotes the formation of pores in the outer membrane ofmitochondria that allow the entry of reduced cytochromec resulting in a concentration of 125mM This detergenttransfers its electrons to cytochrome c from the mitochon-drial respiratory chain located in the inner mitochondrialmembrane without the innermembrane commitment Uponaddition of reduced cytochrome c solution to make upthe concentration of 10mM it becomes possible to recordchanges in absorbance at 550 nm At the end 2 120583L of asolution of potassium cyanide (KCN) 80mM was added
to inhibit complex IV Measuring the ratio of complex IVactivity in the presence and absence of detergent allows theanalysis of mitochondrial fraction quality
26 Statistical Analysis Statistical analysis of the differentresults was performed using the IBM SPSS software v 200(IBM Corporation Armonk New York USA) Doublingtime results were analyzed by nonparametric Kruskal-Wallistest with multiple comparisons using Bonferroni correctionNMR results were analysed using parametric Student t-testComplex IV activity results were analyzed using nonparamet-ric Kruskal-Wallis test A significance of 5 was consideredfor all comparisons
3 Results and Discussion
31 Rates of Glycolysis Levels of lactate in cell culture mediawere monitored by 1H-NMR spectroscopy Figure 2 showsthe temporal evolution of lactate resonances for one ofthe cell lines (LS1034) This multiplet (duplet of triplets)represents one of the 13C-satellites of the methyl resonanceof lactate and constitutes half of the signal due to [U-13C]lactate Figure 3 shows the lactate production for all celllines under all experimental conditions highlow glucoseand normoxiahypoxia A linear evolution is seen for allexperimental conditions and for all colorectal cancer cell linesfor the first 8 h of incubation After this period there is aconsiderable reduction in lactate production which is alsoconsistent with a significant reduction in glucose levels in theculture media Under normoxia the three cell lines exhibita very pronounced glycolytic metabolic profile exportingconsiderable amounts of lactate to the culture media ThisldquoWarburgrdquo like metabolic behavior is characteristic of manytumor cells and was expectable for the colorectal cancer celllines under study From the three LS1034 is the less glycolytic
4 BioMed Research International
146 145 144 143 142 141
8h
4h
0h
(ppm)
(a)
8h
4h
0h
146 145 144 143 142 141
(ppm)
(b)
8h
4h
0h
146 145 144 143 142 141
(ppm)
(c)
8h
4h
0h
146 145 144 143 142 141
(ppm)
(d)
Figure 2 Expansions from 1H-NMR spectra of culture media of LS1034 cells showing the region of one of the satellites due to [U-13C]lactateat 0 4 and 8 hours of incubation in culture media (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d)hypoxialow glucose
followed by C2BBe1 and ultimatelyWiDr appears as themostglycolytic at both high and low glucose normoxic conditionsThis metabolic profile is dramatically altered under hypoxiaThis ldquoinsultrdquo has completely differential effects in the threecell lines and shows as well a dependence on the glucosecontent of the cell culture media The LS1034 cell line isthe most susceptible to hypoxia (Figure 3(b)) During thefirst 8 hours of incubation lactate production rates increasesignificantly more than triplicate while for other cell linesthe increases in those rates are much less notorious Thecolorectal cancer cell line with the lowest dependence onglycolysislactic fermentation under normoxia becomes themost glycolytic under hypoxia The amount of glucose inthe cell culture media also influences the rates of lactateproduction by the three cell lines The combination ofnormoxiahypoxia and highlow glucose forms a matrix thatseemingly affects differently each of the colorectal cancercell lines LS1034 shows a significant increase in lactateproduction due to hypoxia at both high and low glucosecontents while WiDr behaves opposingly for the two glucoselevels increasing glycolysis at high glucose but reducingglycolysis with significant decrease for low glucose assumingan adaptive behavior The C2BBe1 is the less affected by thehypoxic insult at both high and low glucose levels howeverwith significance at low levels of glucose
32 Glycolytic Flux versus Proliferative Activities A signif-icant glycolytic activity is expectable in high proliferativetissues for the sake of availability of carbons skeletons forbiosynthetic routes On this basis one would expect that
the more proliferative a given cell is the more glycolytic itsmetabolic profile should be Figure 4 shows the duplicationtimes for the three colorectal cell lines WiDr is the mostproliferative followed by the C2BBe1 and LS1034 appears asthe cancer linewith the lowest proliferation rate in agreementwith its lowest glycolytic activity as demonstrated by thelactate production rates measured under normoxia witheither high or low glucose in culturemediaDuplication timesand glycolytic activities fully match the concept that there isno ldquowastefulrdquo metabolic behavior by the cancer cell lines butsolely an adaptive behavior that grants them advantage overnormal neighboring cells
33 Krebs Cycle Kinetics Krebs cycle kinetics was evaluatedby 13C-NMR isotopomer analysis using cell perchloric acidextracts Figure 5 shows representative 13C-NMR spectra ofperchloric acid extracts derived from the three colorectal can-cer cell lines grown in normoxic conditions in the presenceof high glucose Some of the resonances are identified andthe expansions show themultiplet of glutamate C4 resonance(C4-Glu 342 ppm) a crucial carbon for performing Krebscycle kinetics analysis [14 16] Immediately perceptible is thefact that the resonance due to lactate methyl carbon (C3-Lac 208 ppm) dominates all three spectra corroborating therobust glycolyticfermentative character of these colorectalcancer cell lines even under normoxia as outlined abovefrom the rates of lactate production Other major differencesbetween cell lines are the amounts of alanine (C3-Ala170 ppm) present in cell extracts much higher in LS1034 and
BioMed Research International 5
00
10
20
30
40
50
60
70
80
90
100 WiDrC
once
ntra
tion
(mM
)
R2= 09945
R2= 09849
R2= 09571
lowast
0 2 4 6 8 10
HG-normoxiaHG-hypoxia LG-hypoxia
R2= 09922
lowast
LG-normoxia
(a)
00
10
20
30
40
50
60
70
80
90
100 LS1034
Con
cent
ratio
n (m
M)
R2= 09272
lowast
R2= 09608
lowast
R2= 07966
lowast
R2= 09998
lowast
0 2 4 6 8
HG-normoxiaHG-hypoxia LG-hypoxia
LG-normoxia
(b)
00
10
20
30
40
50
60
70
80
90
100 C2BBe1
R2= 09694
lowast
R2= 09952
R2= 09880
R2= 09720
lowast
0 2 4 6 8 10
HG-normoxiaHG-hypoxia LG-hypoxia
LG-normoxia
Con
cent
ratio
n (m
M)
(c)
Figure 3 [U-13C]lactate exported by colon cancer cell lines to the incubation media during the first 8 hours of incubation under normoxiaand hypoxia and low and high glucose media (a) WiDr (b) LS1034 (c) C2BBe1 Significant differences are indicated by lowast corresponding to119875 lt 005 (comparison between normoxia and hypoxia conditions) and corresponding to 119875 lt 005 (comparison between high and low cellmedia glucose concentrations (5 and 25mM))
300
200
100
0
WiDr LS1034 C2BBe1
lowastlowast
Dou
blin
g tim
e (h)
Figure 4 Duplication times (h) for the three colorectal cancer cell lines
the composition of the C4-Glu multiplet in terms of duplet45 (D
45) and quartet (Q) multiplets which as mentioned
correlate with Krebs cycle turnover ratesThe levels of alanineare intimately associated with the levels of pyruvate in thecells and can be used instead for deriving information con-cerning the cytosolic redox status [19] In fact C3-LacC3-Ala
is frequently used as an indirect measure of NADHNAD+ratio [20 21] The more the NADHH+ exists in cytosolthe more extensive the conversion of pyruvate is to lactatethrough lactic dehydrogenase (LDH) and the higher theratio is of those two metabolites In terms of C4-Glu theappearance of the Q multiplet is only possible after multiple
6 BioMed Research International
C4-Glu
C3-Glu
C3-Ala
C3-Ala
C3-Lac
C2-Ala
WiDr
LS1034
C2BBe1
(ppm)55 50 45 40 35 30 25 20
(a)
(b)
(c)
Q Q Q Q
D45 D45
Figure 5 13C NMR spectra of perchloric acid extracts for the three colorectal cancer cell lines under normoxia and high glucose media (a)WiDr (b) LS1034 (c) C2BBe1 Expanded is the multiplet due to glutamate carbon 4 (C4-Glu 342 ppm) composed by two major multipletsduplet 45 (D
45) due to glutamate isotopomers labeled in carbons 4 and 5 but not carbon 3 and quartet (Q) representing all glutamate
isotopomers with those three carbons enriched simultaneously Other resonances easily seen are the methyl carbons of lactate (C3-Lac208 ppm) and alanine (C3-Ala 170 ppm) and the carbon 3 of glutamate (C3-Glu 278 ppm)
0
10
20
30
40
WiDr LS1034 C2BBe1
lowastlowast
C3-L
acC3
-Ala
(a)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(b)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(c)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(d)Figure 6 Ratios of the methyl carbon of lactate (C3-Lac) to the methyl carbon of alanine (C3-Ala) for the three colorectal cancer cell linesunder all experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
BioMed Research International 7
00
05
10
15
20
25
WiDr LS1034 C2BBe1
C4Q
C4D45
(a)
WiDr LS1034 C2BBe1
lowast
00
05
10
15
20
25
C4Q
C4D45
(b)
WiDr LS1034 C2BBe100
05
10
15
20
25
C4Q
C4D45
(c)
WiDr LS1034 C2BBe1
lowast
00
05
10
15
20
25
C4Q
C4D45
(d)
Figure 7 Ratios of the glutamate C4 quartet (C4Q) to the duplet 45 (D45) multiplets for the three colorectal cancer cell lines under all
experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
turns of the Krebs cycle (Figure 1) and its contribution to theoverall C4-Glu multiplet is higher in cells with a more activeoxidative metabolism The ratio C4QC4D
45provides a way
to probe Krebs cycle kinetics and to compare different celllines in terms of their glycolyticoxidative metabolic charac-ters Another ratio of metabolic interest is the C3-LacC4-Glu since it reports how enriched the glycolyticoxidativemetabolite pools are and in such a way provides a measureof themetabolic coupling between glycolysis and Krebs cycleThe higher the ratio is the more uncoupled those two centralmetabolic pathways are and the more dependent the cell lineis on glycolysis for its bioenergetics requirements This ratiois frequently high in ldquoWarburgrdquo like cancer cell lines like theones used in this study
Figure 6 shows the ratios of C3-LacC3-Ala for thethree colon cancer cell lines in the four experimental con-ditions This ratio is significantly higher in WiDr undernormoxichigh glucose conditions than in the other twocell lines This correlates with a more pronounced glycolyticflux in this cancer cell line under such conditions Underhypoxia and high glucose there is a considerable attenuationof C3-LacC3-Ala ratio in WiDr only explainable by anoverall metabolic inhibition in these cells For LS1034 andC2BBe1 there is an increase in the C3-LacC3-Ala ratio underhypoxia explainable by a higher dependence on glycolysis forbioenergetics under anaerobic conditions
Figure 7 shows the ratios C4QC4D45
for the three col-orectal cancer cell lines in the four experimental conditionschosen This ratio clearly emphasizes that the three cell lineshave distinct Krebs cycle turnovers under hypoxiaWiDr andLS1034 cell lines suffer a considerable reduction in such ratioat both high and low glucose conditions The same does nothold however for the C2BBe1 which essentially shows noeffect in such ratio due to hypoxia This distinctive behavioremphasizes a distinct capacity of the cells for usage of oxygenat low concentration levels such as the ones prompted by thehypoxic episode and concomitantly justifies the distinctivebehavior of cells in tumors when subjected to hypoxia [22]
Figure 8 shows the C3-LacC4-Glu ratio for the threecolon cancer cell lines in the four experimental conditions Asexpected this ratio is higher formore glycolytic cell lines likeWiDr and suffers a slight increase under hypoxia Under nor-moxia andhigh glucose the LS1034 has the lowest C3-LacC4-Glu ratio in accordancewith amuchmore pronounced oxida-tive profile Upon hypoxia the LS1034 is the one sufferingthe more severe metabolic shift since the oxidative pathwayreduces its contribution to cell bioenergetics and has to becompensated by further glycolytic activity
34 Complex IV Activities In order to corroborate themetabolic alterations mentioned above for each of thecolorectal cancer cell lines and strengthen our findings
8 BioMed Research International
0
20
40
60
WiDr LS1034 C2BBe1
lowastlowast
C3-L
acC4
-Glu
(a)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(b)
WiDr LS1034 C2BBe10
20
40
60
C3-L
acC4
-Glu
(c)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(d)
Figure 8 Ratios of themethyl carbon of lactate (C3-Lac) to the carbon 4 of glutamate (C4-Glu) for the three colorectal cancer cell lines underall experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
concerning the metabolic (un)coupling observed for eachof the cell lines in each of the metabolic conditions theactivity of complex IV from the electron transport chain wasalso evaluatedThis activity adequately normalized for citratesynthase activity is presented in Figure 9 Under normoxiaand high glucose LS1034 cell line is the one possessingthe most active complex IV This is in accordance withall presented data which pointed these cells as the mostoxidativeless glycolytic under such incubation conditionsUnder normoxia and low glucose no significant changes weredetected in complex IV activities in accordance with theabsence of significant differences on the ratios C3-AlaC4-Glu and C4QC4D
45discussed above for this experimental
condition Low substrate concentration does not seem byitself to be able to distinguish the three colorectal cell linesunder analysis Hypoxia on the other hand prompts fordistinction even at low glucose levels thus emphasizing thedistinctive metabolic character of each colorectal cancer cellline This distinct responsiveness is a crucial parameter tobe evaluated aiming at the implementation of more efficientanticancer therapeutic strategies
The implemented methodology for analysis of both cul-ture media and perchloric acid cellular extracts providesunique and complimentary metabolic information which
affords a more complete picture of the metabolic transforma-tions associated with each of the specific actions undertakenfor challenging cancer cell metabolism This methodologycan naturally be extended to other metabolic pathways andmetabolic tracers towards widening the scopes of the currentmetabolic analysis
4 Conclusions
The three colorectal cancer cell lines show very distinctmetabolic profiles in the four experimental conditions testedGlycolysis is in every circumstance the most active metabolicpathway denoting the ldquoWarburgrdquo likemetabolic behavior thatcharacterizes many cancer cells Nevertheless oxidative fluxthrough the Krebs cycle assumes its importance in whole cel-lular bioenergetics and its extent is itself very distinct amongthe three colorectal cancer cell lines The interplay of oxygen(normoxia versus hypoxia) and nutrient (high versus lowglucose) availability also plays a crucial role in the definitionof cell capacity to adapt to distinct microenvironments Inthis study the (in)sensibility to the three colorectal cancercell lines was surprisingly different The less glycolytic undernormoxia LS1034 turns into the most glycolytic in hypoxiawhile themost glycolytic barely respond to the hypoxic insult
BioMed Research International 9
00
02
04
06
08
10
WiDr LS1034 C2BBe1
lowast lowast
Com
plex
IVac
tivity
(a)
WiDr LS1034 C2BBe1
lowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(b)
WiDr LS1034 C2BBe100
02
04
06
08
10
Com
plex
IVac
tivity
(c)
WiDr LS1034 C2BBe1
lowastlowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(d)
Figure 9 Complex IV activities for the three colorectal cancer cell lines under all experimental conditions (a) normoxiahigh glucose(b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
Also the alteration in nutrient availability interferes in themetabolic coupling between glycolysis and Krebs cycle anddemonstrates the distinct capacity that cells have to react tosuch interferences We understand that the whole picture ismuch more complex but would like to point out that thesimple management of O
2and glucose constitutes certainly a
mechanism to take into account to tackle tumor proliferationand devise more efficient therapeutic strategiesThe adequateknowledge of the metabolic behavior of cancer cells is thus amust on this ldquocrusaderdquo against cancer and the methodologyhere described is certainly pivotal for deriving this much-needed information
Conflict of Interests
The authors declare that there is no conflict of interests re-garding the publication of this paper
Authorsrsquo Contribution
Ana M Abrantes Ludgero C Tavares Salome Pires andCandidaMendes performed the experimentsManuela Graz-ina Rui A Carvalho and Maria Filomena Botelho analyzedresults andmade the figures AnaM Abrantes Manuela Gra-zina Rui A Carvalho andMaria Filomena Botelho designed
the research and wrote the paper Joao Casalta-Lopes per-formed statistical analyses
Acknowledgments
The authors are grateful to FCT (Fundacao para a Ciencia eTecnologiamdashPortugal) for financial support (PTDCEBB-EBI1158102009) The authors also acknowledge the ldquoRedeNacional de RMNrdquo (REDE1517RMN2005) for access tothe NMR spectrometer and facilities at Coimbra Uni-versity The authors thank Centre of Investigation onEnvironment Genetics and Oncobiology (CIMAGO) forfinancial support Salome Pires and Ludgero Tavares arerecipients of fellowships from the Portuguese Foundationfor Science and Technology (SFRHBD753002010) and(SFRHBD666002009) respectively
References
[1] OWarburg ldquoOrigin of cancer cellsrdquoOncologia vol 9 no 2 pp75ndash83 1956 (German)
[2] S Weinhouse O Warburg D Burk and A L Schade ldquoOn res-piratory impairment in cancer cellsrdquo Science vol 124 no 3215pp 267ndash272 1956
10 BioMed Research International
[3] Y Zhao E B Butler andMTan ldquoTargeting cellularmetabolismto improve cancer therapeuticsrdquo Cell Death and Disease vol 4no 3 article e532 2013
[4] C V Dang ldquoLinks between metabolism and cancerrdquo Genes ampDevelopment vol 26 no 9 pp 877ndash890 2012
[5] M Talekar S R Boreddy A Singh andMAmiji ldquoTumor aero-bic glycolysis new insights into therapeutic strategies with targ-eted deliveryrdquo Expert Opinion on Biological Therapy 2014
[6] R Xu H Pelicano Y Zhou et al ldquoInhibition of glycolysis incancer cells a novel strategy to overcome drug resistance asso-ciatedwithmitochondrial respiratory defect and hypoxiardquoCan-cer Research vol 65 no 2 pp 613ndash621 2005
[7] S Ros and A Schulze ldquoBalancing glycolytic flux the roleof 6-phosphofructo-2-kinasefructose 26-bisphosphatases incancer metabolismrdquoCancer ampMetabolism vol 1 article 8 2013
[8] E Roudier and A Perrin ldquoConsidering the role of pyruvate intumor cells during hypoxiardquo Biochimica et Biophysica Acta vol1796 no 2 pp 55ndash62 2009
[9] K J Williams B A Telfer R E Airley et al ldquoA protective rolefor HIF-1 in response to redox manipulation and glucose depr-ivation implications for tumorigenesisrdquo Oncogene vol 21 no2 pp 282ndash290 2002
[10] M Achison and T R Hupp ldquoHypoxia attenuates the p53 resp-onse to cellular damagerdquo Oncogene vol 22 no 22 pp 3431ndash3440 2003
[11] H L Kotze E G Armitage K J Sharkey J W AllwoodW B Dunn and K J Williams ldquoA novel untargeted metabol-omics correlation-based network analysis incorporating humanmetabolic reconstructionsrdquo BMC Systems Biology vol 7 article107 2013
[12] Y-A Wen P D Stevens M L Gasser R Andrei and T GaoldquoDownregulation of PHLPP expression contributes to hypoxia-induced resistance to chemotherapy in colon cancer cellsrdquoMolecular and Cellular Biology vol 33 no 22 pp 4594ndash46052013
[13] B Garcia-Barreno T Delgado B Akerlind-Stopner E Norrbyand J A Melero ldquoLocation of the epitope recognized by mono-clonal antibody 63G on the primary structure of human resp-iratory syncytial virus G glycoprotein and the ability of syn-thetic peptides containing this epitope to induce neutralizingantibodiesrdquo Journal of General Virology vol 73 no 10 pp 2625ndash2630 1992
[14] I F Duarte J Marques A F Ladeirinha et al ldquoAnalyticalapproaches toward successful human cell metabolome studiesby NMR spectroscopyrdquo Analytical Chemistry vol 81 no 12 pp5023ndash5032 2009
[15] S L Pereira J Ramalho-Santos A F Branco V A Sardao P JOliveira and R A Carvalho ldquoMetabolic remodeling duringH9c2 myoblast differentiation relevance for in vitro toxicitystudiesrdquo Cardiovascular Toxicology vol 11 no 2 pp 180ndash1902011
[16] MD Basson IMModlin and J AMadri ldquoHuman enterocyte(Caco-2)migration ismodulated in vitro by extracellularmatrixcomposition and epidermal growth factorrdquo Journal of ClinicalInvestigation vol 90 no 1 pp 15ndash23 1992
[17] M D Peterson and M S Mooseker ldquoCharacterization of theenterocyte-like brush border cytoskeleton of the C2BBe clonesof the human intestinal cell line Caco-2rdquo Journal of Cell Sciencevol 102 no 3 pp 581ndash600 1992
[18] E M Chance S H Seeholzer K Kobayashi and J R Will-iamson ldquoMathematical analysis of isotope labeling in the citric
acid cycle with applications to 13C NMR studies in perfusedrat heartsrdquo Journal of Biological Chemistry vol 258 no 22 pp13785ndash13794 1983
[19] D HWilliamson P Lund and H A Krebs ldquoThe redox state offree nicotinamide-adenine dinucleotide in the cytoplasm andmitochondria of rat liverrdquo Biochemical Journal vol 103 no 2pp 514ndash527 1967
[20] L Suardet A C Gaide J M Calmes et al ldquoResponsiveness ofthree newly established human colorectal cancer cell lines totransforming growth factors 1205731 and 12057321rdquo Cancer Research vol52 no 13 pp 3705ndash3712 1992
[21] R A Carvalho T B Rodrigues P Zhao F M H Jeffrey C RMalloy and A D Sherry ldquoA 13C isotopomer kinetic analysisof cardiac metabolism influence of altered cytosolic redoxand [Ca2+]
119900rdquo The American Journal of PhysiologymdashHeart and
Circulatory Physiology vol 287 no 2 pp H889ndashH895 2004[22] AM Abrantes M E S Serra A C Goncalves et al ldquoHypoxia-
induced redox alterations and their correlation with 99mTc-MIBI and 99mTc-HL-91 uptake in colon cancer cellsrdquo NuclearMedicine and Biology vol 37 no 2 pp 125ndash132 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 BioMed Research International
target in cancer therapy both by itself and in combinationwith other therapies such as immunotherapy This advancehelps overcoming drug resistance issues and improves theefficacy of current anticancer agents [3 5ndash7]
Besides that and having into account tumor microen-vironment due to the temporal and spatial heterogeneityof oxygenation that occurs in solid tumors the adaptationto the variability of its microenvironment is critical Oxy-gen supply is impaired in many tumors because there isimbalance between tissue growth and the development ofnew vasculature In solid tumors hypoxia is thus a commoncharacteristicmicroenvironment of tumoral cells becominga key factor for tumor progression and resistance to anti-cancer therapy [8] This decrease in oxygen pO
2influences
compensatory physiological events involving adaptations atall levels in order to maintain homeostasis between cellsenergetic requirements and supplies [8] Thus it appears thataerobic glycolysis is an adaptive mechanism that involvesseveral metabolic pathways coordinates which maintainthe morphological characteristics of tumor cells includingthe ability to survive hypoxic conditions the capacity ofmetastasis and evasion of death by apoptosis [5ndash8]
Solid tumors have also heterogeneous populations of cellsdue in part to a limited blood supply that provides reducedlevels of oxygen and prompts for acidic conditions and avidityof glucose [9]These changes in the tumormicroenvironmentmay represent physiological signals that activate cell survivalor death by apoptosis affecting the balance between growthand tumor suppression The mechanisms by which tumorcells adapt or die in the presence of low levels of oxygenare not well studied and understood However it is knownthat the expression of several transcription factors as wellas the modification of metabolic pathways interfere with theresponse to the lack of oxygen and nutrients by tumor cells[10]
The aim of this study is to characterize the metabolicprofile namely glycolysis and Krebs cycle fluxes of threecolorectal cancer cell lines using carbon-13 (13C) tracers andnuclear magnetic resonance (NMR) spectroscopy With thisapproach central metabolic pathways will be evaluated andmajor metabolic changes resulting from hypoxia and glucoseavailability will be determined towards depicting the possibleinvolvement of metabolic mechanisms [11] in some processesof chemotherapeutic resistance in colon cancer [12]
2 Materials and Methods
21 Cell Lines Colorectal cancer cell lines (WiDr LS1034 andC2BBe1) purchased from American Type Culture Collection(ATCC Rockville MD USA) were used to perform invitro studies The three cell lines are from different colonlocalizations WiDr from rectosigmoid location C2BBe1clone of the cell line Caco-2 (ATCC) from descendingcolon and LS1034 resistant to chemotherapy concerning P-glycoprotein overexpression from ascending colon [9ndash13]WiDr and C2BBe1 cell lines were maintained in DulbeccorsquosModified Eaglersquos cell culture medium (Sigma) and LS1034cell line was maintained in Roswell Park Memorial Institute(RPMI)medium All cell lines were cultured in high (25mM)
and low (5mM) glucose concentrations supplemented with10 fetal calf serum (Gibco) in 5 CO
2atmosphere at
37∘C To perform hypoxia studies cells were incubated at37∘C in 93 N
2 2 O
2 and 5 CO
2using a controlled-
environment cabinet (PlasLabs USA) Each of the mediawas acquired without glucose in order to be supplementedwith [U-13C]glucose for performing 13C NMR isotopomeranalysis
22 Cell Perchloric Acid Extracts Perchloric acid extractswere obtained using published methods [14 15] Briefly tothe cell pellet is added the equivalent (vw) of 2mL ice-cold perchloric acid 72gram wet weight of cells Theresulting supernatant is subsequently neutralized using ice-cold KOH solutions before lyophilization This procedureallows precipitation of KClO
4and a considerable reduction
of solution ionic strength towards the acquisition of highquality NMR spectra For kinetic analysis of lactate produc-tion aliquots of cell culture media (160 120583L) were drawn atspecific time intervals (0 4 and 8 hours) To each of thesealiquots 40 120583L of an internal standard consisting of sodiumfumarate (10mM) in phosphate buffer (100mM) was addedfor absolute quantification purposes
23 NMR Analyses of Intermediary Metabolism
231 Glycolytic Fluxes by Analysis of Cell Culture MediaProton (1H) nuclear magnetic resonance (NMR) spectraof cell culture media were acquired on a 600MHz VarianNMR spectrometer using a 3mm indirect detection probeTypical acquisition parameters included a 3 s acquisitiontime a radiofrequency pulse of 45∘ and an interpulse delayof 10 seconds to ensure full relaxation of all nuclei in thesample Due to direct ( 1119869HC) and long range (
2119869HC and 3119869HC)
heteronuclear scalar coupling it is possible to distinguish theresonances of [U-13C]lactate from those due to nonenriched[U-12C]lactate (Figure 1) [U-13C]lactate originates from [U-13C]glucose as a result of glycolysis followed by reductionof pyruvate by lactate dehydrogenase (LDH) Levels of [U-13C]lactate are thus used as an indirect measure of glycolyticactivity
232 Krebs Cycle Kinetics by 1H and 13C NMR Analysisof Cell Extracts 1H- and 13C-NMR spectra of cell extractswere acquired in a 600MHz Varian NMR spectrometerusing a 3mm indirect detection and a 3mm broadbandNMR probe respectively 1H-NMR spectra of cell extractsallow the monitoring of the levels of intracellular metabolicintermediates while 13C-NMR spectra allows tackling thedynamics of 13C incorporation in metabolic intermediatesthus warranting a dynamic vision of cell metabolism Ratesof 13C incorporation in glycolytic and Krebs cycle interme-diates are used as a measure of metabolic coupling betweenthese two major metabolic pathways and provide valuableclues about the ldquometabolic remodelingrdquo associated with thetumorigenic transformation and treatment 13C isotopomersof intracellular glutamate provide a window for monitoring
BioMed Research International 3
1 2 3 4 5 6
1
11
2
22
3
33
4 5
F1
F2
F3F4Alanine
OAACitrate
120572-Ketoglutarate
Succinyl-CoA Glutamate
Glucose
Lactate
3JHH
2JHC
3JHC
1st turn D45
2nd turn Q
D45D45
Q QQ Q
Figure 1 [U-13C]glucose as a carbon tracer for monitoring glycolysis and Krebs cycle Through glycolysis and lactic fermentation [U-13C]lactate is produced which is distinguishable (multiplets in each 13C satellite due to 2119869HC
3119869HC and
3119869HHmdashduplet of triplets since 2119869HC
and 3119869HC have similar magnitude) from tissue lactate or lactate originating from unenriched carbon sources [12-13C2]acetil-CoA originating
from [U-13C]pyruvate leads to incorporation of 13C atoms in Krebs cycle intermediates and other metabolites that exchange with those (egglutamate) It is possible to follow Krebs cycle kinetics by monitoring the 13C multiplet due to carbon 4 of glutamate The duplet component(D45) results from the simple entrance of [12-13C
2]acetil-CoA but the quartet requires further cycling and combination of [12-13C
2]acetil-
CoA with previously enriched oxaloacetate (OAA) A simple ratio of QD45is proportional to Krebs cycle flux and denotes higher rates of
oxidative metabolism in tissues as opposed to glycolysis Metabolic fluxes F1ndash4 denote the major pathways for [U-13C]glucose use
Krebs cycle kinetics (Figure 1) since the appearance of multi-13C-labeled species clearly indicate cycle turnover [14 16 17]and oxidative efficiency
24 Citrate Synthase Enzyme Activity The citrate synthaseenzyme (CS) is used as a marker enzyme of mitochondrialpreparations due to its stability and its regulatory role in theKrebs cycle Its activity can provide an estimate of the numberof mitochondria in a cell suspension and can be used in orderto standardize the results of the activities of the enzymes ofthe mitochondrial respiratory chain [15 18] The evaluationof the activity of CS was performed by spectrophotometry ata 120582 of 412 nmThis assay consists of the condensation reactionof acetyl-CoA (02mM) with oxaloacetate (8mM) catalyzedby CSThis reaction results in coenzymeA (CoA) release thatreacts with 551015840-dithiobis(-nitrobenzoic acid) (DNTB) addedto the medium at a concentration of 2mM thus enabling thereading at 412 nm To allow the access of substrates to theenzyme Triton X-100 was used at 01
25 Complex IV Activity To measure complex IV activity50120583g of protein was added to 1mL of buffer composed by10mM KH
2PO4 300mM sucrose and 5mgmL BSA with
pH 65 which was in a cuvette The analysis started with thecompletion of a baseline tracing at 120582 550 nm and then 10 120583Lwas added of detergent n-dodecyl-120573-D-maltoside whichpromotes the formation of pores in the outer membrane ofmitochondria that allow the entry of reduced cytochromec resulting in a concentration of 125mM This detergenttransfers its electrons to cytochrome c from the mitochon-drial respiratory chain located in the inner mitochondrialmembrane without the innermembrane commitment Uponaddition of reduced cytochrome c solution to make upthe concentration of 10mM it becomes possible to recordchanges in absorbance at 550 nm At the end 2 120583L of asolution of potassium cyanide (KCN) 80mM was added
to inhibit complex IV Measuring the ratio of complex IVactivity in the presence and absence of detergent allows theanalysis of mitochondrial fraction quality
26 Statistical Analysis Statistical analysis of the differentresults was performed using the IBM SPSS software v 200(IBM Corporation Armonk New York USA) Doublingtime results were analyzed by nonparametric Kruskal-Wallistest with multiple comparisons using Bonferroni correctionNMR results were analysed using parametric Student t-testComplex IV activity results were analyzed using nonparamet-ric Kruskal-Wallis test A significance of 5 was consideredfor all comparisons
3 Results and Discussion
31 Rates of Glycolysis Levels of lactate in cell culture mediawere monitored by 1H-NMR spectroscopy Figure 2 showsthe temporal evolution of lactate resonances for one ofthe cell lines (LS1034) This multiplet (duplet of triplets)represents one of the 13C-satellites of the methyl resonanceof lactate and constitutes half of the signal due to [U-13C]lactate Figure 3 shows the lactate production for all celllines under all experimental conditions highlow glucoseand normoxiahypoxia A linear evolution is seen for allexperimental conditions and for all colorectal cancer cell linesfor the first 8 h of incubation After this period there is aconsiderable reduction in lactate production which is alsoconsistent with a significant reduction in glucose levels in theculture media Under normoxia the three cell lines exhibita very pronounced glycolytic metabolic profile exportingconsiderable amounts of lactate to the culture media ThisldquoWarburgrdquo like metabolic behavior is characteristic of manytumor cells and was expectable for the colorectal cancer celllines under study From the three LS1034 is the less glycolytic
4 BioMed Research International
146 145 144 143 142 141
8h
4h
0h
(ppm)
(a)
8h
4h
0h
146 145 144 143 142 141
(ppm)
(b)
8h
4h
0h
146 145 144 143 142 141
(ppm)
(c)
8h
4h
0h
146 145 144 143 142 141
(ppm)
(d)
Figure 2 Expansions from 1H-NMR spectra of culture media of LS1034 cells showing the region of one of the satellites due to [U-13C]lactateat 0 4 and 8 hours of incubation in culture media (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d)hypoxialow glucose
followed by C2BBe1 and ultimatelyWiDr appears as themostglycolytic at both high and low glucose normoxic conditionsThis metabolic profile is dramatically altered under hypoxiaThis ldquoinsultrdquo has completely differential effects in the threecell lines and shows as well a dependence on the glucosecontent of the cell culture media The LS1034 cell line isthe most susceptible to hypoxia (Figure 3(b)) During thefirst 8 hours of incubation lactate production rates increasesignificantly more than triplicate while for other cell linesthe increases in those rates are much less notorious Thecolorectal cancer cell line with the lowest dependence onglycolysislactic fermentation under normoxia becomes themost glycolytic under hypoxia The amount of glucose inthe cell culture media also influences the rates of lactateproduction by the three cell lines The combination ofnormoxiahypoxia and highlow glucose forms a matrix thatseemingly affects differently each of the colorectal cancercell lines LS1034 shows a significant increase in lactateproduction due to hypoxia at both high and low glucosecontents while WiDr behaves opposingly for the two glucoselevels increasing glycolysis at high glucose but reducingglycolysis with significant decrease for low glucose assumingan adaptive behavior The C2BBe1 is the less affected by thehypoxic insult at both high and low glucose levels howeverwith significance at low levels of glucose
32 Glycolytic Flux versus Proliferative Activities A signif-icant glycolytic activity is expectable in high proliferativetissues for the sake of availability of carbons skeletons forbiosynthetic routes On this basis one would expect that
the more proliferative a given cell is the more glycolytic itsmetabolic profile should be Figure 4 shows the duplicationtimes for the three colorectal cell lines WiDr is the mostproliferative followed by the C2BBe1 and LS1034 appears asthe cancer linewith the lowest proliferation rate in agreementwith its lowest glycolytic activity as demonstrated by thelactate production rates measured under normoxia witheither high or low glucose in culturemediaDuplication timesand glycolytic activities fully match the concept that there isno ldquowastefulrdquo metabolic behavior by the cancer cell lines butsolely an adaptive behavior that grants them advantage overnormal neighboring cells
33 Krebs Cycle Kinetics Krebs cycle kinetics was evaluatedby 13C-NMR isotopomer analysis using cell perchloric acidextracts Figure 5 shows representative 13C-NMR spectra ofperchloric acid extracts derived from the three colorectal can-cer cell lines grown in normoxic conditions in the presenceof high glucose Some of the resonances are identified andthe expansions show themultiplet of glutamate C4 resonance(C4-Glu 342 ppm) a crucial carbon for performing Krebscycle kinetics analysis [14 16] Immediately perceptible is thefact that the resonance due to lactate methyl carbon (C3-Lac 208 ppm) dominates all three spectra corroborating therobust glycolyticfermentative character of these colorectalcancer cell lines even under normoxia as outlined abovefrom the rates of lactate production Other major differencesbetween cell lines are the amounts of alanine (C3-Ala170 ppm) present in cell extracts much higher in LS1034 and
BioMed Research International 5
00
10
20
30
40
50
60
70
80
90
100 WiDrC
once
ntra
tion
(mM
)
R2= 09945
R2= 09849
R2= 09571
lowast
0 2 4 6 8 10
HG-normoxiaHG-hypoxia LG-hypoxia
R2= 09922
lowast
LG-normoxia
(a)
00
10
20
30
40
50
60
70
80
90
100 LS1034
Con
cent
ratio
n (m
M)
R2= 09272
lowast
R2= 09608
lowast
R2= 07966
lowast
R2= 09998
lowast
0 2 4 6 8
HG-normoxiaHG-hypoxia LG-hypoxia
LG-normoxia
(b)
00
10
20
30
40
50
60
70
80
90
100 C2BBe1
R2= 09694
lowast
R2= 09952
R2= 09880
R2= 09720
lowast
0 2 4 6 8 10
HG-normoxiaHG-hypoxia LG-hypoxia
LG-normoxia
Con
cent
ratio
n (m
M)
(c)
Figure 3 [U-13C]lactate exported by colon cancer cell lines to the incubation media during the first 8 hours of incubation under normoxiaand hypoxia and low and high glucose media (a) WiDr (b) LS1034 (c) C2BBe1 Significant differences are indicated by lowast corresponding to119875 lt 005 (comparison between normoxia and hypoxia conditions) and corresponding to 119875 lt 005 (comparison between high and low cellmedia glucose concentrations (5 and 25mM))
300
200
100
0
WiDr LS1034 C2BBe1
lowastlowast
Dou
blin
g tim
e (h)
Figure 4 Duplication times (h) for the three colorectal cancer cell lines
the composition of the C4-Glu multiplet in terms of duplet45 (D
45) and quartet (Q) multiplets which as mentioned
correlate with Krebs cycle turnover ratesThe levels of alanineare intimately associated with the levels of pyruvate in thecells and can be used instead for deriving information con-cerning the cytosolic redox status [19] In fact C3-LacC3-Ala
is frequently used as an indirect measure of NADHNAD+ratio [20 21] The more the NADHH+ exists in cytosolthe more extensive the conversion of pyruvate is to lactatethrough lactic dehydrogenase (LDH) and the higher theratio is of those two metabolites In terms of C4-Glu theappearance of the Q multiplet is only possible after multiple
6 BioMed Research International
C4-Glu
C3-Glu
C3-Ala
C3-Ala
C3-Lac
C2-Ala
WiDr
LS1034
C2BBe1
(ppm)55 50 45 40 35 30 25 20
(a)
(b)
(c)
Q Q Q Q
D45 D45
Figure 5 13C NMR spectra of perchloric acid extracts for the three colorectal cancer cell lines under normoxia and high glucose media (a)WiDr (b) LS1034 (c) C2BBe1 Expanded is the multiplet due to glutamate carbon 4 (C4-Glu 342 ppm) composed by two major multipletsduplet 45 (D
45) due to glutamate isotopomers labeled in carbons 4 and 5 but not carbon 3 and quartet (Q) representing all glutamate
isotopomers with those three carbons enriched simultaneously Other resonances easily seen are the methyl carbons of lactate (C3-Lac208 ppm) and alanine (C3-Ala 170 ppm) and the carbon 3 of glutamate (C3-Glu 278 ppm)
0
10
20
30
40
WiDr LS1034 C2BBe1
lowastlowast
C3-L
acC3
-Ala
(a)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(b)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(c)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(d)Figure 6 Ratios of the methyl carbon of lactate (C3-Lac) to the methyl carbon of alanine (C3-Ala) for the three colorectal cancer cell linesunder all experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
BioMed Research International 7
00
05
10
15
20
25
WiDr LS1034 C2BBe1
C4Q
C4D45
(a)
WiDr LS1034 C2BBe1
lowast
00
05
10
15
20
25
C4Q
C4D45
(b)
WiDr LS1034 C2BBe100
05
10
15
20
25
C4Q
C4D45
(c)
WiDr LS1034 C2BBe1
lowast
00
05
10
15
20
25
C4Q
C4D45
(d)
Figure 7 Ratios of the glutamate C4 quartet (C4Q) to the duplet 45 (D45) multiplets for the three colorectal cancer cell lines under all
experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
turns of the Krebs cycle (Figure 1) and its contribution to theoverall C4-Glu multiplet is higher in cells with a more activeoxidative metabolism The ratio C4QC4D
45provides a way
to probe Krebs cycle kinetics and to compare different celllines in terms of their glycolyticoxidative metabolic charac-ters Another ratio of metabolic interest is the C3-LacC4-Glu since it reports how enriched the glycolyticoxidativemetabolite pools are and in such a way provides a measureof themetabolic coupling between glycolysis and Krebs cycleThe higher the ratio is the more uncoupled those two centralmetabolic pathways are and the more dependent the cell lineis on glycolysis for its bioenergetics requirements This ratiois frequently high in ldquoWarburgrdquo like cancer cell lines like theones used in this study
Figure 6 shows the ratios of C3-LacC3-Ala for thethree colon cancer cell lines in the four experimental con-ditions This ratio is significantly higher in WiDr undernormoxichigh glucose conditions than in the other twocell lines This correlates with a more pronounced glycolyticflux in this cancer cell line under such conditions Underhypoxia and high glucose there is a considerable attenuationof C3-LacC3-Ala ratio in WiDr only explainable by anoverall metabolic inhibition in these cells For LS1034 andC2BBe1 there is an increase in the C3-LacC3-Ala ratio underhypoxia explainable by a higher dependence on glycolysis forbioenergetics under anaerobic conditions
Figure 7 shows the ratios C4QC4D45
for the three col-orectal cancer cell lines in the four experimental conditionschosen This ratio clearly emphasizes that the three cell lineshave distinct Krebs cycle turnovers under hypoxiaWiDr andLS1034 cell lines suffer a considerable reduction in such ratioat both high and low glucose conditions The same does nothold however for the C2BBe1 which essentially shows noeffect in such ratio due to hypoxia This distinctive behavioremphasizes a distinct capacity of the cells for usage of oxygenat low concentration levels such as the ones prompted by thehypoxic episode and concomitantly justifies the distinctivebehavior of cells in tumors when subjected to hypoxia [22]
Figure 8 shows the C3-LacC4-Glu ratio for the threecolon cancer cell lines in the four experimental conditions Asexpected this ratio is higher formore glycolytic cell lines likeWiDr and suffers a slight increase under hypoxia Under nor-moxia andhigh glucose the LS1034 has the lowest C3-LacC4-Glu ratio in accordancewith amuchmore pronounced oxida-tive profile Upon hypoxia the LS1034 is the one sufferingthe more severe metabolic shift since the oxidative pathwayreduces its contribution to cell bioenergetics and has to becompensated by further glycolytic activity
34 Complex IV Activities In order to corroborate themetabolic alterations mentioned above for each of thecolorectal cancer cell lines and strengthen our findings
8 BioMed Research International
0
20
40
60
WiDr LS1034 C2BBe1
lowastlowast
C3-L
acC4
-Glu
(a)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(b)
WiDr LS1034 C2BBe10
20
40
60
C3-L
acC4
-Glu
(c)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(d)
Figure 8 Ratios of themethyl carbon of lactate (C3-Lac) to the carbon 4 of glutamate (C4-Glu) for the three colorectal cancer cell lines underall experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
concerning the metabolic (un)coupling observed for eachof the cell lines in each of the metabolic conditions theactivity of complex IV from the electron transport chain wasalso evaluatedThis activity adequately normalized for citratesynthase activity is presented in Figure 9 Under normoxiaand high glucose LS1034 cell line is the one possessingthe most active complex IV This is in accordance withall presented data which pointed these cells as the mostoxidativeless glycolytic under such incubation conditionsUnder normoxia and low glucose no significant changes weredetected in complex IV activities in accordance with theabsence of significant differences on the ratios C3-AlaC4-Glu and C4QC4D
45discussed above for this experimental
condition Low substrate concentration does not seem byitself to be able to distinguish the three colorectal cell linesunder analysis Hypoxia on the other hand prompts fordistinction even at low glucose levels thus emphasizing thedistinctive metabolic character of each colorectal cancer cellline This distinct responsiveness is a crucial parameter tobe evaluated aiming at the implementation of more efficientanticancer therapeutic strategies
The implemented methodology for analysis of both cul-ture media and perchloric acid cellular extracts providesunique and complimentary metabolic information which
affords a more complete picture of the metabolic transforma-tions associated with each of the specific actions undertakenfor challenging cancer cell metabolism This methodologycan naturally be extended to other metabolic pathways andmetabolic tracers towards widening the scopes of the currentmetabolic analysis
4 Conclusions
The three colorectal cancer cell lines show very distinctmetabolic profiles in the four experimental conditions testedGlycolysis is in every circumstance the most active metabolicpathway denoting the ldquoWarburgrdquo likemetabolic behavior thatcharacterizes many cancer cells Nevertheless oxidative fluxthrough the Krebs cycle assumes its importance in whole cel-lular bioenergetics and its extent is itself very distinct amongthe three colorectal cancer cell lines The interplay of oxygen(normoxia versus hypoxia) and nutrient (high versus lowglucose) availability also plays a crucial role in the definitionof cell capacity to adapt to distinct microenvironments Inthis study the (in)sensibility to the three colorectal cancercell lines was surprisingly different The less glycolytic undernormoxia LS1034 turns into the most glycolytic in hypoxiawhile themost glycolytic barely respond to the hypoxic insult
BioMed Research International 9
00
02
04
06
08
10
WiDr LS1034 C2BBe1
lowast lowast
Com
plex
IVac
tivity
(a)
WiDr LS1034 C2BBe1
lowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(b)
WiDr LS1034 C2BBe100
02
04
06
08
10
Com
plex
IVac
tivity
(c)
WiDr LS1034 C2BBe1
lowastlowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(d)
Figure 9 Complex IV activities for the three colorectal cancer cell lines under all experimental conditions (a) normoxiahigh glucose(b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
Also the alteration in nutrient availability interferes in themetabolic coupling between glycolysis and Krebs cycle anddemonstrates the distinct capacity that cells have to react tosuch interferences We understand that the whole picture ismuch more complex but would like to point out that thesimple management of O
2and glucose constitutes certainly a
mechanism to take into account to tackle tumor proliferationand devise more efficient therapeutic strategiesThe adequateknowledge of the metabolic behavior of cancer cells is thus amust on this ldquocrusaderdquo against cancer and the methodologyhere described is certainly pivotal for deriving this much-needed information
Conflict of Interests
The authors declare that there is no conflict of interests re-garding the publication of this paper
Authorsrsquo Contribution
Ana M Abrantes Ludgero C Tavares Salome Pires andCandidaMendes performed the experimentsManuela Graz-ina Rui A Carvalho and Maria Filomena Botelho analyzedresults andmade the figures AnaM Abrantes Manuela Gra-zina Rui A Carvalho andMaria Filomena Botelho designed
the research and wrote the paper Joao Casalta-Lopes per-formed statistical analyses
Acknowledgments
The authors are grateful to FCT (Fundacao para a Ciencia eTecnologiamdashPortugal) for financial support (PTDCEBB-EBI1158102009) The authors also acknowledge the ldquoRedeNacional de RMNrdquo (REDE1517RMN2005) for access tothe NMR spectrometer and facilities at Coimbra Uni-versity The authors thank Centre of Investigation onEnvironment Genetics and Oncobiology (CIMAGO) forfinancial support Salome Pires and Ludgero Tavares arerecipients of fellowships from the Portuguese Foundationfor Science and Technology (SFRHBD753002010) and(SFRHBD666002009) respectively
References
[1] OWarburg ldquoOrigin of cancer cellsrdquoOncologia vol 9 no 2 pp75ndash83 1956 (German)
[2] S Weinhouse O Warburg D Burk and A L Schade ldquoOn res-piratory impairment in cancer cellsrdquo Science vol 124 no 3215pp 267ndash272 1956
10 BioMed Research International
[3] Y Zhao E B Butler andMTan ldquoTargeting cellularmetabolismto improve cancer therapeuticsrdquo Cell Death and Disease vol 4no 3 article e532 2013
[4] C V Dang ldquoLinks between metabolism and cancerrdquo Genes ampDevelopment vol 26 no 9 pp 877ndash890 2012
[5] M Talekar S R Boreddy A Singh andMAmiji ldquoTumor aero-bic glycolysis new insights into therapeutic strategies with targ-eted deliveryrdquo Expert Opinion on Biological Therapy 2014
[6] R Xu H Pelicano Y Zhou et al ldquoInhibition of glycolysis incancer cells a novel strategy to overcome drug resistance asso-ciatedwithmitochondrial respiratory defect and hypoxiardquoCan-cer Research vol 65 no 2 pp 613ndash621 2005
[7] S Ros and A Schulze ldquoBalancing glycolytic flux the roleof 6-phosphofructo-2-kinasefructose 26-bisphosphatases incancer metabolismrdquoCancer ampMetabolism vol 1 article 8 2013
[8] E Roudier and A Perrin ldquoConsidering the role of pyruvate intumor cells during hypoxiardquo Biochimica et Biophysica Acta vol1796 no 2 pp 55ndash62 2009
[9] K J Williams B A Telfer R E Airley et al ldquoA protective rolefor HIF-1 in response to redox manipulation and glucose depr-ivation implications for tumorigenesisrdquo Oncogene vol 21 no2 pp 282ndash290 2002
[10] M Achison and T R Hupp ldquoHypoxia attenuates the p53 resp-onse to cellular damagerdquo Oncogene vol 22 no 22 pp 3431ndash3440 2003
[11] H L Kotze E G Armitage K J Sharkey J W AllwoodW B Dunn and K J Williams ldquoA novel untargeted metabol-omics correlation-based network analysis incorporating humanmetabolic reconstructionsrdquo BMC Systems Biology vol 7 article107 2013
[12] Y-A Wen P D Stevens M L Gasser R Andrei and T GaoldquoDownregulation of PHLPP expression contributes to hypoxia-induced resistance to chemotherapy in colon cancer cellsrdquoMolecular and Cellular Biology vol 33 no 22 pp 4594ndash46052013
[13] B Garcia-Barreno T Delgado B Akerlind-Stopner E Norrbyand J A Melero ldquoLocation of the epitope recognized by mono-clonal antibody 63G on the primary structure of human resp-iratory syncytial virus G glycoprotein and the ability of syn-thetic peptides containing this epitope to induce neutralizingantibodiesrdquo Journal of General Virology vol 73 no 10 pp 2625ndash2630 1992
[14] I F Duarte J Marques A F Ladeirinha et al ldquoAnalyticalapproaches toward successful human cell metabolome studiesby NMR spectroscopyrdquo Analytical Chemistry vol 81 no 12 pp5023ndash5032 2009
[15] S L Pereira J Ramalho-Santos A F Branco V A Sardao P JOliveira and R A Carvalho ldquoMetabolic remodeling duringH9c2 myoblast differentiation relevance for in vitro toxicitystudiesrdquo Cardiovascular Toxicology vol 11 no 2 pp 180ndash1902011
[16] MD Basson IMModlin and J AMadri ldquoHuman enterocyte(Caco-2)migration ismodulated in vitro by extracellularmatrixcomposition and epidermal growth factorrdquo Journal of ClinicalInvestigation vol 90 no 1 pp 15ndash23 1992
[17] M D Peterson and M S Mooseker ldquoCharacterization of theenterocyte-like brush border cytoskeleton of the C2BBe clonesof the human intestinal cell line Caco-2rdquo Journal of Cell Sciencevol 102 no 3 pp 581ndash600 1992
[18] E M Chance S H Seeholzer K Kobayashi and J R Will-iamson ldquoMathematical analysis of isotope labeling in the citric
acid cycle with applications to 13C NMR studies in perfusedrat heartsrdquo Journal of Biological Chemistry vol 258 no 22 pp13785ndash13794 1983
[19] D HWilliamson P Lund and H A Krebs ldquoThe redox state offree nicotinamide-adenine dinucleotide in the cytoplasm andmitochondria of rat liverrdquo Biochemical Journal vol 103 no 2pp 514ndash527 1967
[20] L Suardet A C Gaide J M Calmes et al ldquoResponsiveness ofthree newly established human colorectal cancer cell lines totransforming growth factors 1205731 and 12057321rdquo Cancer Research vol52 no 13 pp 3705ndash3712 1992
[21] R A Carvalho T B Rodrigues P Zhao F M H Jeffrey C RMalloy and A D Sherry ldquoA 13C isotopomer kinetic analysisof cardiac metabolism influence of altered cytosolic redoxand [Ca2+]
119900rdquo The American Journal of PhysiologymdashHeart and
Circulatory Physiology vol 287 no 2 pp H889ndashH895 2004[22] AM Abrantes M E S Serra A C Goncalves et al ldquoHypoxia-
induced redox alterations and their correlation with 99mTc-MIBI and 99mTc-HL-91 uptake in colon cancer cellsrdquo NuclearMedicine and Biology vol 37 no 2 pp 125ndash132 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 3
1 2 3 4 5 6
1
11
2
22
3
33
4 5
F1
F2
F3F4Alanine
OAACitrate
120572-Ketoglutarate
Succinyl-CoA Glutamate
Glucose
Lactate
3JHH
2JHC
3JHC
1st turn D45
2nd turn Q
D45D45
Q QQ Q
Figure 1 [U-13C]glucose as a carbon tracer for monitoring glycolysis and Krebs cycle Through glycolysis and lactic fermentation [U-13C]lactate is produced which is distinguishable (multiplets in each 13C satellite due to 2119869HC
3119869HC and
3119869HHmdashduplet of triplets since 2119869HC
and 3119869HC have similar magnitude) from tissue lactate or lactate originating from unenriched carbon sources [12-13C2]acetil-CoA originating
from [U-13C]pyruvate leads to incorporation of 13C atoms in Krebs cycle intermediates and other metabolites that exchange with those (egglutamate) It is possible to follow Krebs cycle kinetics by monitoring the 13C multiplet due to carbon 4 of glutamate The duplet component(D45) results from the simple entrance of [12-13C
2]acetil-CoA but the quartet requires further cycling and combination of [12-13C
2]acetil-
CoA with previously enriched oxaloacetate (OAA) A simple ratio of QD45is proportional to Krebs cycle flux and denotes higher rates of
oxidative metabolism in tissues as opposed to glycolysis Metabolic fluxes F1ndash4 denote the major pathways for [U-13C]glucose use
Krebs cycle kinetics (Figure 1) since the appearance of multi-13C-labeled species clearly indicate cycle turnover [14 16 17]and oxidative efficiency
24 Citrate Synthase Enzyme Activity The citrate synthaseenzyme (CS) is used as a marker enzyme of mitochondrialpreparations due to its stability and its regulatory role in theKrebs cycle Its activity can provide an estimate of the numberof mitochondria in a cell suspension and can be used in orderto standardize the results of the activities of the enzymes ofthe mitochondrial respiratory chain [15 18] The evaluationof the activity of CS was performed by spectrophotometry ata 120582 of 412 nmThis assay consists of the condensation reactionof acetyl-CoA (02mM) with oxaloacetate (8mM) catalyzedby CSThis reaction results in coenzymeA (CoA) release thatreacts with 551015840-dithiobis(-nitrobenzoic acid) (DNTB) addedto the medium at a concentration of 2mM thus enabling thereading at 412 nm To allow the access of substrates to theenzyme Triton X-100 was used at 01
25 Complex IV Activity To measure complex IV activity50120583g of protein was added to 1mL of buffer composed by10mM KH
2PO4 300mM sucrose and 5mgmL BSA with
pH 65 which was in a cuvette The analysis started with thecompletion of a baseline tracing at 120582 550 nm and then 10 120583Lwas added of detergent n-dodecyl-120573-D-maltoside whichpromotes the formation of pores in the outer membrane ofmitochondria that allow the entry of reduced cytochromec resulting in a concentration of 125mM This detergenttransfers its electrons to cytochrome c from the mitochon-drial respiratory chain located in the inner mitochondrialmembrane without the innermembrane commitment Uponaddition of reduced cytochrome c solution to make upthe concentration of 10mM it becomes possible to recordchanges in absorbance at 550 nm At the end 2 120583L of asolution of potassium cyanide (KCN) 80mM was added
to inhibit complex IV Measuring the ratio of complex IVactivity in the presence and absence of detergent allows theanalysis of mitochondrial fraction quality
26 Statistical Analysis Statistical analysis of the differentresults was performed using the IBM SPSS software v 200(IBM Corporation Armonk New York USA) Doublingtime results were analyzed by nonparametric Kruskal-Wallistest with multiple comparisons using Bonferroni correctionNMR results were analysed using parametric Student t-testComplex IV activity results were analyzed using nonparamet-ric Kruskal-Wallis test A significance of 5 was consideredfor all comparisons
3 Results and Discussion
31 Rates of Glycolysis Levels of lactate in cell culture mediawere monitored by 1H-NMR spectroscopy Figure 2 showsthe temporal evolution of lactate resonances for one ofthe cell lines (LS1034) This multiplet (duplet of triplets)represents one of the 13C-satellites of the methyl resonanceof lactate and constitutes half of the signal due to [U-13C]lactate Figure 3 shows the lactate production for all celllines under all experimental conditions highlow glucoseand normoxiahypoxia A linear evolution is seen for allexperimental conditions and for all colorectal cancer cell linesfor the first 8 h of incubation After this period there is aconsiderable reduction in lactate production which is alsoconsistent with a significant reduction in glucose levels in theculture media Under normoxia the three cell lines exhibita very pronounced glycolytic metabolic profile exportingconsiderable amounts of lactate to the culture media ThisldquoWarburgrdquo like metabolic behavior is characteristic of manytumor cells and was expectable for the colorectal cancer celllines under study From the three LS1034 is the less glycolytic
4 BioMed Research International
146 145 144 143 142 141
8h
4h
0h
(ppm)
(a)
8h
4h
0h
146 145 144 143 142 141
(ppm)
(b)
8h
4h
0h
146 145 144 143 142 141
(ppm)
(c)
8h
4h
0h
146 145 144 143 142 141
(ppm)
(d)
Figure 2 Expansions from 1H-NMR spectra of culture media of LS1034 cells showing the region of one of the satellites due to [U-13C]lactateat 0 4 and 8 hours of incubation in culture media (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d)hypoxialow glucose
followed by C2BBe1 and ultimatelyWiDr appears as themostglycolytic at both high and low glucose normoxic conditionsThis metabolic profile is dramatically altered under hypoxiaThis ldquoinsultrdquo has completely differential effects in the threecell lines and shows as well a dependence on the glucosecontent of the cell culture media The LS1034 cell line isthe most susceptible to hypoxia (Figure 3(b)) During thefirst 8 hours of incubation lactate production rates increasesignificantly more than triplicate while for other cell linesthe increases in those rates are much less notorious Thecolorectal cancer cell line with the lowest dependence onglycolysislactic fermentation under normoxia becomes themost glycolytic under hypoxia The amount of glucose inthe cell culture media also influences the rates of lactateproduction by the three cell lines The combination ofnormoxiahypoxia and highlow glucose forms a matrix thatseemingly affects differently each of the colorectal cancercell lines LS1034 shows a significant increase in lactateproduction due to hypoxia at both high and low glucosecontents while WiDr behaves opposingly for the two glucoselevels increasing glycolysis at high glucose but reducingglycolysis with significant decrease for low glucose assumingan adaptive behavior The C2BBe1 is the less affected by thehypoxic insult at both high and low glucose levels howeverwith significance at low levels of glucose
32 Glycolytic Flux versus Proliferative Activities A signif-icant glycolytic activity is expectable in high proliferativetissues for the sake of availability of carbons skeletons forbiosynthetic routes On this basis one would expect that
the more proliferative a given cell is the more glycolytic itsmetabolic profile should be Figure 4 shows the duplicationtimes for the three colorectal cell lines WiDr is the mostproliferative followed by the C2BBe1 and LS1034 appears asthe cancer linewith the lowest proliferation rate in agreementwith its lowest glycolytic activity as demonstrated by thelactate production rates measured under normoxia witheither high or low glucose in culturemediaDuplication timesand glycolytic activities fully match the concept that there isno ldquowastefulrdquo metabolic behavior by the cancer cell lines butsolely an adaptive behavior that grants them advantage overnormal neighboring cells
33 Krebs Cycle Kinetics Krebs cycle kinetics was evaluatedby 13C-NMR isotopomer analysis using cell perchloric acidextracts Figure 5 shows representative 13C-NMR spectra ofperchloric acid extracts derived from the three colorectal can-cer cell lines grown in normoxic conditions in the presenceof high glucose Some of the resonances are identified andthe expansions show themultiplet of glutamate C4 resonance(C4-Glu 342 ppm) a crucial carbon for performing Krebscycle kinetics analysis [14 16] Immediately perceptible is thefact that the resonance due to lactate methyl carbon (C3-Lac 208 ppm) dominates all three spectra corroborating therobust glycolyticfermentative character of these colorectalcancer cell lines even under normoxia as outlined abovefrom the rates of lactate production Other major differencesbetween cell lines are the amounts of alanine (C3-Ala170 ppm) present in cell extracts much higher in LS1034 and
BioMed Research International 5
00
10
20
30
40
50
60
70
80
90
100 WiDrC
once
ntra
tion
(mM
)
R2= 09945
R2= 09849
R2= 09571
lowast
0 2 4 6 8 10
HG-normoxiaHG-hypoxia LG-hypoxia
R2= 09922
lowast
LG-normoxia
(a)
00
10
20
30
40
50
60
70
80
90
100 LS1034
Con
cent
ratio
n (m
M)
R2= 09272
lowast
R2= 09608
lowast
R2= 07966
lowast
R2= 09998
lowast
0 2 4 6 8
HG-normoxiaHG-hypoxia LG-hypoxia
LG-normoxia
(b)
00
10
20
30
40
50
60
70
80
90
100 C2BBe1
R2= 09694
lowast
R2= 09952
R2= 09880
R2= 09720
lowast
0 2 4 6 8 10
HG-normoxiaHG-hypoxia LG-hypoxia
LG-normoxia
Con
cent
ratio
n (m
M)
(c)
Figure 3 [U-13C]lactate exported by colon cancer cell lines to the incubation media during the first 8 hours of incubation under normoxiaand hypoxia and low and high glucose media (a) WiDr (b) LS1034 (c) C2BBe1 Significant differences are indicated by lowast corresponding to119875 lt 005 (comparison between normoxia and hypoxia conditions) and corresponding to 119875 lt 005 (comparison between high and low cellmedia glucose concentrations (5 and 25mM))
300
200
100
0
WiDr LS1034 C2BBe1
lowastlowast
Dou
blin
g tim
e (h)
Figure 4 Duplication times (h) for the three colorectal cancer cell lines
the composition of the C4-Glu multiplet in terms of duplet45 (D
45) and quartet (Q) multiplets which as mentioned
correlate with Krebs cycle turnover ratesThe levels of alanineare intimately associated with the levels of pyruvate in thecells and can be used instead for deriving information con-cerning the cytosolic redox status [19] In fact C3-LacC3-Ala
is frequently used as an indirect measure of NADHNAD+ratio [20 21] The more the NADHH+ exists in cytosolthe more extensive the conversion of pyruvate is to lactatethrough lactic dehydrogenase (LDH) and the higher theratio is of those two metabolites In terms of C4-Glu theappearance of the Q multiplet is only possible after multiple
6 BioMed Research International
C4-Glu
C3-Glu
C3-Ala
C3-Ala
C3-Lac
C2-Ala
WiDr
LS1034
C2BBe1
(ppm)55 50 45 40 35 30 25 20
(a)
(b)
(c)
Q Q Q Q
D45 D45
Figure 5 13C NMR spectra of perchloric acid extracts for the three colorectal cancer cell lines under normoxia and high glucose media (a)WiDr (b) LS1034 (c) C2BBe1 Expanded is the multiplet due to glutamate carbon 4 (C4-Glu 342 ppm) composed by two major multipletsduplet 45 (D
45) due to glutamate isotopomers labeled in carbons 4 and 5 but not carbon 3 and quartet (Q) representing all glutamate
isotopomers with those three carbons enriched simultaneously Other resonances easily seen are the methyl carbons of lactate (C3-Lac208 ppm) and alanine (C3-Ala 170 ppm) and the carbon 3 of glutamate (C3-Glu 278 ppm)
0
10
20
30
40
WiDr LS1034 C2BBe1
lowastlowast
C3-L
acC3
-Ala
(a)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(b)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(c)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(d)Figure 6 Ratios of the methyl carbon of lactate (C3-Lac) to the methyl carbon of alanine (C3-Ala) for the three colorectal cancer cell linesunder all experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
BioMed Research International 7
00
05
10
15
20
25
WiDr LS1034 C2BBe1
C4Q
C4D45
(a)
WiDr LS1034 C2BBe1
lowast
00
05
10
15
20
25
C4Q
C4D45
(b)
WiDr LS1034 C2BBe100
05
10
15
20
25
C4Q
C4D45
(c)
WiDr LS1034 C2BBe1
lowast
00
05
10
15
20
25
C4Q
C4D45
(d)
Figure 7 Ratios of the glutamate C4 quartet (C4Q) to the duplet 45 (D45) multiplets for the three colorectal cancer cell lines under all
experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
turns of the Krebs cycle (Figure 1) and its contribution to theoverall C4-Glu multiplet is higher in cells with a more activeoxidative metabolism The ratio C4QC4D
45provides a way
to probe Krebs cycle kinetics and to compare different celllines in terms of their glycolyticoxidative metabolic charac-ters Another ratio of metabolic interest is the C3-LacC4-Glu since it reports how enriched the glycolyticoxidativemetabolite pools are and in such a way provides a measureof themetabolic coupling between glycolysis and Krebs cycleThe higher the ratio is the more uncoupled those two centralmetabolic pathways are and the more dependent the cell lineis on glycolysis for its bioenergetics requirements This ratiois frequently high in ldquoWarburgrdquo like cancer cell lines like theones used in this study
Figure 6 shows the ratios of C3-LacC3-Ala for thethree colon cancer cell lines in the four experimental con-ditions This ratio is significantly higher in WiDr undernormoxichigh glucose conditions than in the other twocell lines This correlates with a more pronounced glycolyticflux in this cancer cell line under such conditions Underhypoxia and high glucose there is a considerable attenuationof C3-LacC3-Ala ratio in WiDr only explainable by anoverall metabolic inhibition in these cells For LS1034 andC2BBe1 there is an increase in the C3-LacC3-Ala ratio underhypoxia explainable by a higher dependence on glycolysis forbioenergetics under anaerobic conditions
Figure 7 shows the ratios C4QC4D45
for the three col-orectal cancer cell lines in the four experimental conditionschosen This ratio clearly emphasizes that the three cell lineshave distinct Krebs cycle turnovers under hypoxiaWiDr andLS1034 cell lines suffer a considerable reduction in such ratioat both high and low glucose conditions The same does nothold however for the C2BBe1 which essentially shows noeffect in such ratio due to hypoxia This distinctive behavioremphasizes a distinct capacity of the cells for usage of oxygenat low concentration levels such as the ones prompted by thehypoxic episode and concomitantly justifies the distinctivebehavior of cells in tumors when subjected to hypoxia [22]
Figure 8 shows the C3-LacC4-Glu ratio for the threecolon cancer cell lines in the four experimental conditions Asexpected this ratio is higher formore glycolytic cell lines likeWiDr and suffers a slight increase under hypoxia Under nor-moxia andhigh glucose the LS1034 has the lowest C3-LacC4-Glu ratio in accordancewith amuchmore pronounced oxida-tive profile Upon hypoxia the LS1034 is the one sufferingthe more severe metabolic shift since the oxidative pathwayreduces its contribution to cell bioenergetics and has to becompensated by further glycolytic activity
34 Complex IV Activities In order to corroborate themetabolic alterations mentioned above for each of thecolorectal cancer cell lines and strengthen our findings
8 BioMed Research International
0
20
40
60
WiDr LS1034 C2BBe1
lowastlowast
C3-L
acC4
-Glu
(a)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(b)
WiDr LS1034 C2BBe10
20
40
60
C3-L
acC4
-Glu
(c)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(d)
Figure 8 Ratios of themethyl carbon of lactate (C3-Lac) to the carbon 4 of glutamate (C4-Glu) for the three colorectal cancer cell lines underall experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
concerning the metabolic (un)coupling observed for eachof the cell lines in each of the metabolic conditions theactivity of complex IV from the electron transport chain wasalso evaluatedThis activity adequately normalized for citratesynthase activity is presented in Figure 9 Under normoxiaand high glucose LS1034 cell line is the one possessingthe most active complex IV This is in accordance withall presented data which pointed these cells as the mostoxidativeless glycolytic under such incubation conditionsUnder normoxia and low glucose no significant changes weredetected in complex IV activities in accordance with theabsence of significant differences on the ratios C3-AlaC4-Glu and C4QC4D
45discussed above for this experimental
condition Low substrate concentration does not seem byitself to be able to distinguish the three colorectal cell linesunder analysis Hypoxia on the other hand prompts fordistinction even at low glucose levels thus emphasizing thedistinctive metabolic character of each colorectal cancer cellline This distinct responsiveness is a crucial parameter tobe evaluated aiming at the implementation of more efficientanticancer therapeutic strategies
The implemented methodology for analysis of both cul-ture media and perchloric acid cellular extracts providesunique and complimentary metabolic information which
affords a more complete picture of the metabolic transforma-tions associated with each of the specific actions undertakenfor challenging cancer cell metabolism This methodologycan naturally be extended to other metabolic pathways andmetabolic tracers towards widening the scopes of the currentmetabolic analysis
4 Conclusions
The three colorectal cancer cell lines show very distinctmetabolic profiles in the four experimental conditions testedGlycolysis is in every circumstance the most active metabolicpathway denoting the ldquoWarburgrdquo likemetabolic behavior thatcharacterizes many cancer cells Nevertheless oxidative fluxthrough the Krebs cycle assumes its importance in whole cel-lular bioenergetics and its extent is itself very distinct amongthe three colorectal cancer cell lines The interplay of oxygen(normoxia versus hypoxia) and nutrient (high versus lowglucose) availability also plays a crucial role in the definitionof cell capacity to adapt to distinct microenvironments Inthis study the (in)sensibility to the three colorectal cancercell lines was surprisingly different The less glycolytic undernormoxia LS1034 turns into the most glycolytic in hypoxiawhile themost glycolytic barely respond to the hypoxic insult
BioMed Research International 9
00
02
04
06
08
10
WiDr LS1034 C2BBe1
lowast lowast
Com
plex
IVac
tivity
(a)
WiDr LS1034 C2BBe1
lowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(b)
WiDr LS1034 C2BBe100
02
04
06
08
10
Com
plex
IVac
tivity
(c)
WiDr LS1034 C2BBe1
lowastlowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(d)
Figure 9 Complex IV activities for the three colorectal cancer cell lines under all experimental conditions (a) normoxiahigh glucose(b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
Also the alteration in nutrient availability interferes in themetabolic coupling between glycolysis and Krebs cycle anddemonstrates the distinct capacity that cells have to react tosuch interferences We understand that the whole picture ismuch more complex but would like to point out that thesimple management of O
2and glucose constitutes certainly a
mechanism to take into account to tackle tumor proliferationand devise more efficient therapeutic strategiesThe adequateknowledge of the metabolic behavior of cancer cells is thus amust on this ldquocrusaderdquo against cancer and the methodologyhere described is certainly pivotal for deriving this much-needed information
Conflict of Interests
The authors declare that there is no conflict of interests re-garding the publication of this paper
Authorsrsquo Contribution
Ana M Abrantes Ludgero C Tavares Salome Pires andCandidaMendes performed the experimentsManuela Graz-ina Rui A Carvalho and Maria Filomena Botelho analyzedresults andmade the figures AnaM Abrantes Manuela Gra-zina Rui A Carvalho andMaria Filomena Botelho designed
the research and wrote the paper Joao Casalta-Lopes per-formed statistical analyses
Acknowledgments
The authors are grateful to FCT (Fundacao para a Ciencia eTecnologiamdashPortugal) for financial support (PTDCEBB-EBI1158102009) The authors also acknowledge the ldquoRedeNacional de RMNrdquo (REDE1517RMN2005) for access tothe NMR spectrometer and facilities at Coimbra Uni-versity The authors thank Centre of Investigation onEnvironment Genetics and Oncobiology (CIMAGO) forfinancial support Salome Pires and Ludgero Tavares arerecipients of fellowships from the Portuguese Foundationfor Science and Technology (SFRHBD753002010) and(SFRHBD666002009) respectively
References
[1] OWarburg ldquoOrigin of cancer cellsrdquoOncologia vol 9 no 2 pp75ndash83 1956 (German)
[2] S Weinhouse O Warburg D Burk and A L Schade ldquoOn res-piratory impairment in cancer cellsrdquo Science vol 124 no 3215pp 267ndash272 1956
10 BioMed Research International
[3] Y Zhao E B Butler andMTan ldquoTargeting cellularmetabolismto improve cancer therapeuticsrdquo Cell Death and Disease vol 4no 3 article e532 2013
[4] C V Dang ldquoLinks between metabolism and cancerrdquo Genes ampDevelopment vol 26 no 9 pp 877ndash890 2012
[5] M Talekar S R Boreddy A Singh andMAmiji ldquoTumor aero-bic glycolysis new insights into therapeutic strategies with targ-eted deliveryrdquo Expert Opinion on Biological Therapy 2014
[6] R Xu H Pelicano Y Zhou et al ldquoInhibition of glycolysis incancer cells a novel strategy to overcome drug resistance asso-ciatedwithmitochondrial respiratory defect and hypoxiardquoCan-cer Research vol 65 no 2 pp 613ndash621 2005
[7] S Ros and A Schulze ldquoBalancing glycolytic flux the roleof 6-phosphofructo-2-kinasefructose 26-bisphosphatases incancer metabolismrdquoCancer ampMetabolism vol 1 article 8 2013
[8] E Roudier and A Perrin ldquoConsidering the role of pyruvate intumor cells during hypoxiardquo Biochimica et Biophysica Acta vol1796 no 2 pp 55ndash62 2009
[9] K J Williams B A Telfer R E Airley et al ldquoA protective rolefor HIF-1 in response to redox manipulation and glucose depr-ivation implications for tumorigenesisrdquo Oncogene vol 21 no2 pp 282ndash290 2002
[10] M Achison and T R Hupp ldquoHypoxia attenuates the p53 resp-onse to cellular damagerdquo Oncogene vol 22 no 22 pp 3431ndash3440 2003
[11] H L Kotze E G Armitage K J Sharkey J W AllwoodW B Dunn and K J Williams ldquoA novel untargeted metabol-omics correlation-based network analysis incorporating humanmetabolic reconstructionsrdquo BMC Systems Biology vol 7 article107 2013
[12] Y-A Wen P D Stevens M L Gasser R Andrei and T GaoldquoDownregulation of PHLPP expression contributes to hypoxia-induced resistance to chemotherapy in colon cancer cellsrdquoMolecular and Cellular Biology vol 33 no 22 pp 4594ndash46052013
[13] B Garcia-Barreno T Delgado B Akerlind-Stopner E Norrbyand J A Melero ldquoLocation of the epitope recognized by mono-clonal antibody 63G on the primary structure of human resp-iratory syncytial virus G glycoprotein and the ability of syn-thetic peptides containing this epitope to induce neutralizingantibodiesrdquo Journal of General Virology vol 73 no 10 pp 2625ndash2630 1992
[14] I F Duarte J Marques A F Ladeirinha et al ldquoAnalyticalapproaches toward successful human cell metabolome studiesby NMR spectroscopyrdquo Analytical Chemistry vol 81 no 12 pp5023ndash5032 2009
[15] S L Pereira J Ramalho-Santos A F Branco V A Sardao P JOliveira and R A Carvalho ldquoMetabolic remodeling duringH9c2 myoblast differentiation relevance for in vitro toxicitystudiesrdquo Cardiovascular Toxicology vol 11 no 2 pp 180ndash1902011
[16] MD Basson IMModlin and J AMadri ldquoHuman enterocyte(Caco-2)migration ismodulated in vitro by extracellularmatrixcomposition and epidermal growth factorrdquo Journal of ClinicalInvestigation vol 90 no 1 pp 15ndash23 1992
[17] M D Peterson and M S Mooseker ldquoCharacterization of theenterocyte-like brush border cytoskeleton of the C2BBe clonesof the human intestinal cell line Caco-2rdquo Journal of Cell Sciencevol 102 no 3 pp 581ndash600 1992
[18] E M Chance S H Seeholzer K Kobayashi and J R Will-iamson ldquoMathematical analysis of isotope labeling in the citric
acid cycle with applications to 13C NMR studies in perfusedrat heartsrdquo Journal of Biological Chemistry vol 258 no 22 pp13785ndash13794 1983
[19] D HWilliamson P Lund and H A Krebs ldquoThe redox state offree nicotinamide-adenine dinucleotide in the cytoplasm andmitochondria of rat liverrdquo Biochemical Journal vol 103 no 2pp 514ndash527 1967
[20] L Suardet A C Gaide J M Calmes et al ldquoResponsiveness ofthree newly established human colorectal cancer cell lines totransforming growth factors 1205731 and 12057321rdquo Cancer Research vol52 no 13 pp 3705ndash3712 1992
[21] R A Carvalho T B Rodrigues P Zhao F M H Jeffrey C RMalloy and A D Sherry ldquoA 13C isotopomer kinetic analysisof cardiac metabolism influence of altered cytosolic redoxand [Ca2+]
119900rdquo The American Journal of PhysiologymdashHeart and
Circulatory Physiology vol 287 no 2 pp H889ndashH895 2004[22] AM Abrantes M E S Serra A C Goncalves et al ldquoHypoxia-
induced redox alterations and their correlation with 99mTc-MIBI and 99mTc-HL-91 uptake in colon cancer cellsrdquo NuclearMedicine and Biology vol 37 no 2 pp 125ndash132 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 BioMed Research International
146 145 144 143 142 141
8h
4h
0h
(ppm)
(a)
8h
4h
0h
146 145 144 143 142 141
(ppm)
(b)
8h
4h
0h
146 145 144 143 142 141
(ppm)
(c)
8h
4h
0h
146 145 144 143 142 141
(ppm)
(d)
Figure 2 Expansions from 1H-NMR spectra of culture media of LS1034 cells showing the region of one of the satellites due to [U-13C]lactateat 0 4 and 8 hours of incubation in culture media (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d)hypoxialow glucose
followed by C2BBe1 and ultimatelyWiDr appears as themostglycolytic at both high and low glucose normoxic conditionsThis metabolic profile is dramatically altered under hypoxiaThis ldquoinsultrdquo has completely differential effects in the threecell lines and shows as well a dependence on the glucosecontent of the cell culture media The LS1034 cell line isthe most susceptible to hypoxia (Figure 3(b)) During thefirst 8 hours of incubation lactate production rates increasesignificantly more than triplicate while for other cell linesthe increases in those rates are much less notorious Thecolorectal cancer cell line with the lowest dependence onglycolysislactic fermentation under normoxia becomes themost glycolytic under hypoxia The amount of glucose inthe cell culture media also influences the rates of lactateproduction by the three cell lines The combination ofnormoxiahypoxia and highlow glucose forms a matrix thatseemingly affects differently each of the colorectal cancercell lines LS1034 shows a significant increase in lactateproduction due to hypoxia at both high and low glucosecontents while WiDr behaves opposingly for the two glucoselevels increasing glycolysis at high glucose but reducingglycolysis with significant decrease for low glucose assumingan adaptive behavior The C2BBe1 is the less affected by thehypoxic insult at both high and low glucose levels howeverwith significance at low levels of glucose
32 Glycolytic Flux versus Proliferative Activities A signif-icant glycolytic activity is expectable in high proliferativetissues for the sake of availability of carbons skeletons forbiosynthetic routes On this basis one would expect that
the more proliferative a given cell is the more glycolytic itsmetabolic profile should be Figure 4 shows the duplicationtimes for the three colorectal cell lines WiDr is the mostproliferative followed by the C2BBe1 and LS1034 appears asthe cancer linewith the lowest proliferation rate in agreementwith its lowest glycolytic activity as demonstrated by thelactate production rates measured under normoxia witheither high or low glucose in culturemediaDuplication timesand glycolytic activities fully match the concept that there isno ldquowastefulrdquo metabolic behavior by the cancer cell lines butsolely an adaptive behavior that grants them advantage overnormal neighboring cells
33 Krebs Cycle Kinetics Krebs cycle kinetics was evaluatedby 13C-NMR isotopomer analysis using cell perchloric acidextracts Figure 5 shows representative 13C-NMR spectra ofperchloric acid extracts derived from the three colorectal can-cer cell lines grown in normoxic conditions in the presenceof high glucose Some of the resonances are identified andthe expansions show themultiplet of glutamate C4 resonance(C4-Glu 342 ppm) a crucial carbon for performing Krebscycle kinetics analysis [14 16] Immediately perceptible is thefact that the resonance due to lactate methyl carbon (C3-Lac 208 ppm) dominates all three spectra corroborating therobust glycolyticfermentative character of these colorectalcancer cell lines even under normoxia as outlined abovefrom the rates of lactate production Other major differencesbetween cell lines are the amounts of alanine (C3-Ala170 ppm) present in cell extracts much higher in LS1034 and
BioMed Research International 5
00
10
20
30
40
50
60
70
80
90
100 WiDrC
once
ntra
tion
(mM
)
R2= 09945
R2= 09849
R2= 09571
lowast
0 2 4 6 8 10
HG-normoxiaHG-hypoxia LG-hypoxia
R2= 09922
lowast
LG-normoxia
(a)
00
10
20
30
40
50
60
70
80
90
100 LS1034
Con
cent
ratio
n (m
M)
R2= 09272
lowast
R2= 09608
lowast
R2= 07966
lowast
R2= 09998
lowast
0 2 4 6 8
HG-normoxiaHG-hypoxia LG-hypoxia
LG-normoxia
(b)
00
10
20
30
40
50
60
70
80
90
100 C2BBe1
R2= 09694
lowast
R2= 09952
R2= 09880
R2= 09720
lowast
0 2 4 6 8 10
HG-normoxiaHG-hypoxia LG-hypoxia
LG-normoxia
Con
cent
ratio
n (m
M)
(c)
Figure 3 [U-13C]lactate exported by colon cancer cell lines to the incubation media during the first 8 hours of incubation under normoxiaand hypoxia and low and high glucose media (a) WiDr (b) LS1034 (c) C2BBe1 Significant differences are indicated by lowast corresponding to119875 lt 005 (comparison between normoxia and hypoxia conditions) and corresponding to 119875 lt 005 (comparison between high and low cellmedia glucose concentrations (5 and 25mM))
300
200
100
0
WiDr LS1034 C2BBe1
lowastlowast
Dou
blin
g tim
e (h)
Figure 4 Duplication times (h) for the three colorectal cancer cell lines
the composition of the C4-Glu multiplet in terms of duplet45 (D
45) and quartet (Q) multiplets which as mentioned
correlate with Krebs cycle turnover ratesThe levels of alanineare intimately associated with the levels of pyruvate in thecells and can be used instead for deriving information con-cerning the cytosolic redox status [19] In fact C3-LacC3-Ala
is frequently used as an indirect measure of NADHNAD+ratio [20 21] The more the NADHH+ exists in cytosolthe more extensive the conversion of pyruvate is to lactatethrough lactic dehydrogenase (LDH) and the higher theratio is of those two metabolites In terms of C4-Glu theappearance of the Q multiplet is only possible after multiple
6 BioMed Research International
C4-Glu
C3-Glu
C3-Ala
C3-Ala
C3-Lac
C2-Ala
WiDr
LS1034
C2BBe1
(ppm)55 50 45 40 35 30 25 20
(a)
(b)
(c)
Q Q Q Q
D45 D45
Figure 5 13C NMR spectra of perchloric acid extracts for the three colorectal cancer cell lines under normoxia and high glucose media (a)WiDr (b) LS1034 (c) C2BBe1 Expanded is the multiplet due to glutamate carbon 4 (C4-Glu 342 ppm) composed by two major multipletsduplet 45 (D
45) due to glutamate isotopomers labeled in carbons 4 and 5 but not carbon 3 and quartet (Q) representing all glutamate
isotopomers with those three carbons enriched simultaneously Other resonances easily seen are the methyl carbons of lactate (C3-Lac208 ppm) and alanine (C3-Ala 170 ppm) and the carbon 3 of glutamate (C3-Glu 278 ppm)
0
10
20
30
40
WiDr LS1034 C2BBe1
lowastlowast
C3-L
acC3
-Ala
(a)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(b)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(c)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(d)Figure 6 Ratios of the methyl carbon of lactate (C3-Lac) to the methyl carbon of alanine (C3-Ala) for the three colorectal cancer cell linesunder all experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
BioMed Research International 7
00
05
10
15
20
25
WiDr LS1034 C2BBe1
C4Q
C4D45
(a)
WiDr LS1034 C2BBe1
lowast
00
05
10
15
20
25
C4Q
C4D45
(b)
WiDr LS1034 C2BBe100
05
10
15
20
25
C4Q
C4D45
(c)
WiDr LS1034 C2BBe1
lowast
00
05
10
15
20
25
C4Q
C4D45
(d)
Figure 7 Ratios of the glutamate C4 quartet (C4Q) to the duplet 45 (D45) multiplets for the three colorectal cancer cell lines under all
experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
turns of the Krebs cycle (Figure 1) and its contribution to theoverall C4-Glu multiplet is higher in cells with a more activeoxidative metabolism The ratio C4QC4D
45provides a way
to probe Krebs cycle kinetics and to compare different celllines in terms of their glycolyticoxidative metabolic charac-ters Another ratio of metabolic interest is the C3-LacC4-Glu since it reports how enriched the glycolyticoxidativemetabolite pools are and in such a way provides a measureof themetabolic coupling between glycolysis and Krebs cycleThe higher the ratio is the more uncoupled those two centralmetabolic pathways are and the more dependent the cell lineis on glycolysis for its bioenergetics requirements This ratiois frequently high in ldquoWarburgrdquo like cancer cell lines like theones used in this study
Figure 6 shows the ratios of C3-LacC3-Ala for thethree colon cancer cell lines in the four experimental con-ditions This ratio is significantly higher in WiDr undernormoxichigh glucose conditions than in the other twocell lines This correlates with a more pronounced glycolyticflux in this cancer cell line under such conditions Underhypoxia and high glucose there is a considerable attenuationof C3-LacC3-Ala ratio in WiDr only explainable by anoverall metabolic inhibition in these cells For LS1034 andC2BBe1 there is an increase in the C3-LacC3-Ala ratio underhypoxia explainable by a higher dependence on glycolysis forbioenergetics under anaerobic conditions
Figure 7 shows the ratios C4QC4D45
for the three col-orectal cancer cell lines in the four experimental conditionschosen This ratio clearly emphasizes that the three cell lineshave distinct Krebs cycle turnovers under hypoxiaWiDr andLS1034 cell lines suffer a considerable reduction in such ratioat both high and low glucose conditions The same does nothold however for the C2BBe1 which essentially shows noeffect in such ratio due to hypoxia This distinctive behavioremphasizes a distinct capacity of the cells for usage of oxygenat low concentration levels such as the ones prompted by thehypoxic episode and concomitantly justifies the distinctivebehavior of cells in tumors when subjected to hypoxia [22]
Figure 8 shows the C3-LacC4-Glu ratio for the threecolon cancer cell lines in the four experimental conditions Asexpected this ratio is higher formore glycolytic cell lines likeWiDr and suffers a slight increase under hypoxia Under nor-moxia andhigh glucose the LS1034 has the lowest C3-LacC4-Glu ratio in accordancewith amuchmore pronounced oxida-tive profile Upon hypoxia the LS1034 is the one sufferingthe more severe metabolic shift since the oxidative pathwayreduces its contribution to cell bioenergetics and has to becompensated by further glycolytic activity
34 Complex IV Activities In order to corroborate themetabolic alterations mentioned above for each of thecolorectal cancer cell lines and strengthen our findings
8 BioMed Research International
0
20
40
60
WiDr LS1034 C2BBe1
lowastlowast
C3-L
acC4
-Glu
(a)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(b)
WiDr LS1034 C2BBe10
20
40
60
C3-L
acC4
-Glu
(c)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(d)
Figure 8 Ratios of themethyl carbon of lactate (C3-Lac) to the carbon 4 of glutamate (C4-Glu) for the three colorectal cancer cell lines underall experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
concerning the metabolic (un)coupling observed for eachof the cell lines in each of the metabolic conditions theactivity of complex IV from the electron transport chain wasalso evaluatedThis activity adequately normalized for citratesynthase activity is presented in Figure 9 Under normoxiaand high glucose LS1034 cell line is the one possessingthe most active complex IV This is in accordance withall presented data which pointed these cells as the mostoxidativeless glycolytic under such incubation conditionsUnder normoxia and low glucose no significant changes weredetected in complex IV activities in accordance with theabsence of significant differences on the ratios C3-AlaC4-Glu and C4QC4D
45discussed above for this experimental
condition Low substrate concentration does not seem byitself to be able to distinguish the three colorectal cell linesunder analysis Hypoxia on the other hand prompts fordistinction even at low glucose levels thus emphasizing thedistinctive metabolic character of each colorectal cancer cellline This distinct responsiveness is a crucial parameter tobe evaluated aiming at the implementation of more efficientanticancer therapeutic strategies
The implemented methodology for analysis of both cul-ture media and perchloric acid cellular extracts providesunique and complimentary metabolic information which
affords a more complete picture of the metabolic transforma-tions associated with each of the specific actions undertakenfor challenging cancer cell metabolism This methodologycan naturally be extended to other metabolic pathways andmetabolic tracers towards widening the scopes of the currentmetabolic analysis
4 Conclusions
The three colorectal cancer cell lines show very distinctmetabolic profiles in the four experimental conditions testedGlycolysis is in every circumstance the most active metabolicpathway denoting the ldquoWarburgrdquo likemetabolic behavior thatcharacterizes many cancer cells Nevertheless oxidative fluxthrough the Krebs cycle assumes its importance in whole cel-lular bioenergetics and its extent is itself very distinct amongthe three colorectal cancer cell lines The interplay of oxygen(normoxia versus hypoxia) and nutrient (high versus lowglucose) availability also plays a crucial role in the definitionof cell capacity to adapt to distinct microenvironments Inthis study the (in)sensibility to the three colorectal cancercell lines was surprisingly different The less glycolytic undernormoxia LS1034 turns into the most glycolytic in hypoxiawhile themost glycolytic barely respond to the hypoxic insult
BioMed Research International 9
00
02
04
06
08
10
WiDr LS1034 C2BBe1
lowast lowast
Com
plex
IVac
tivity
(a)
WiDr LS1034 C2BBe1
lowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(b)
WiDr LS1034 C2BBe100
02
04
06
08
10
Com
plex
IVac
tivity
(c)
WiDr LS1034 C2BBe1
lowastlowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(d)
Figure 9 Complex IV activities for the three colorectal cancer cell lines under all experimental conditions (a) normoxiahigh glucose(b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
Also the alteration in nutrient availability interferes in themetabolic coupling between glycolysis and Krebs cycle anddemonstrates the distinct capacity that cells have to react tosuch interferences We understand that the whole picture ismuch more complex but would like to point out that thesimple management of O
2and glucose constitutes certainly a
mechanism to take into account to tackle tumor proliferationand devise more efficient therapeutic strategiesThe adequateknowledge of the metabolic behavior of cancer cells is thus amust on this ldquocrusaderdquo against cancer and the methodologyhere described is certainly pivotal for deriving this much-needed information
Conflict of Interests
The authors declare that there is no conflict of interests re-garding the publication of this paper
Authorsrsquo Contribution
Ana M Abrantes Ludgero C Tavares Salome Pires andCandidaMendes performed the experimentsManuela Graz-ina Rui A Carvalho and Maria Filomena Botelho analyzedresults andmade the figures AnaM Abrantes Manuela Gra-zina Rui A Carvalho andMaria Filomena Botelho designed
the research and wrote the paper Joao Casalta-Lopes per-formed statistical analyses
Acknowledgments
The authors are grateful to FCT (Fundacao para a Ciencia eTecnologiamdashPortugal) for financial support (PTDCEBB-EBI1158102009) The authors also acknowledge the ldquoRedeNacional de RMNrdquo (REDE1517RMN2005) for access tothe NMR spectrometer and facilities at Coimbra Uni-versity The authors thank Centre of Investigation onEnvironment Genetics and Oncobiology (CIMAGO) forfinancial support Salome Pires and Ludgero Tavares arerecipients of fellowships from the Portuguese Foundationfor Science and Technology (SFRHBD753002010) and(SFRHBD666002009) respectively
References
[1] OWarburg ldquoOrigin of cancer cellsrdquoOncologia vol 9 no 2 pp75ndash83 1956 (German)
[2] S Weinhouse O Warburg D Burk and A L Schade ldquoOn res-piratory impairment in cancer cellsrdquo Science vol 124 no 3215pp 267ndash272 1956
10 BioMed Research International
[3] Y Zhao E B Butler andMTan ldquoTargeting cellularmetabolismto improve cancer therapeuticsrdquo Cell Death and Disease vol 4no 3 article e532 2013
[4] C V Dang ldquoLinks between metabolism and cancerrdquo Genes ampDevelopment vol 26 no 9 pp 877ndash890 2012
[5] M Talekar S R Boreddy A Singh andMAmiji ldquoTumor aero-bic glycolysis new insights into therapeutic strategies with targ-eted deliveryrdquo Expert Opinion on Biological Therapy 2014
[6] R Xu H Pelicano Y Zhou et al ldquoInhibition of glycolysis incancer cells a novel strategy to overcome drug resistance asso-ciatedwithmitochondrial respiratory defect and hypoxiardquoCan-cer Research vol 65 no 2 pp 613ndash621 2005
[7] S Ros and A Schulze ldquoBalancing glycolytic flux the roleof 6-phosphofructo-2-kinasefructose 26-bisphosphatases incancer metabolismrdquoCancer ampMetabolism vol 1 article 8 2013
[8] E Roudier and A Perrin ldquoConsidering the role of pyruvate intumor cells during hypoxiardquo Biochimica et Biophysica Acta vol1796 no 2 pp 55ndash62 2009
[9] K J Williams B A Telfer R E Airley et al ldquoA protective rolefor HIF-1 in response to redox manipulation and glucose depr-ivation implications for tumorigenesisrdquo Oncogene vol 21 no2 pp 282ndash290 2002
[10] M Achison and T R Hupp ldquoHypoxia attenuates the p53 resp-onse to cellular damagerdquo Oncogene vol 22 no 22 pp 3431ndash3440 2003
[11] H L Kotze E G Armitage K J Sharkey J W AllwoodW B Dunn and K J Williams ldquoA novel untargeted metabol-omics correlation-based network analysis incorporating humanmetabolic reconstructionsrdquo BMC Systems Biology vol 7 article107 2013
[12] Y-A Wen P D Stevens M L Gasser R Andrei and T GaoldquoDownregulation of PHLPP expression contributes to hypoxia-induced resistance to chemotherapy in colon cancer cellsrdquoMolecular and Cellular Biology vol 33 no 22 pp 4594ndash46052013
[13] B Garcia-Barreno T Delgado B Akerlind-Stopner E Norrbyand J A Melero ldquoLocation of the epitope recognized by mono-clonal antibody 63G on the primary structure of human resp-iratory syncytial virus G glycoprotein and the ability of syn-thetic peptides containing this epitope to induce neutralizingantibodiesrdquo Journal of General Virology vol 73 no 10 pp 2625ndash2630 1992
[14] I F Duarte J Marques A F Ladeirinha et al ldquoAnalyticalapproaches toward successful human cell metabolome studiesby NMR spectroscopyrdquo Analytical Chemistry vol 81 no 12 pp5023ndash5032 2009
[15] S L Pereira J Ramalho-Santos A F Branco V A Sardao P JOliveira and R A Carvalho ldquoMetabolic remodeling duringH9c2 myoblast differentiation relevance for in vitro toxicitystudiesrdquo Cardiovascular Toxicology vol 11 no 2 pp 180ndash1902011
[16] MD Basson IMModlin and J AMadri ldquoHuman enterocyte(Caco-2)migration ismodulated in vitro by extracellularmatrixcomposition and epidermal growth factorrdquo Journal of ClinicalInvestigation vol 90 no 1 pp 15ndash23 1992
[17] M D Peterson and M S Mooseker ldquoCharacterization of theenterocyte-like brush border cytoskeleton of the C2BBe clonesof the human intestinal cell line Caco-2rdquo Journal of Cell Sciencevol 102 no 3 pp 581ndash600 1992
[18] E M Chance S H Seeholzer K Kobayashi and J R Will-iamson ldquoMathematical analysis of isotope labeling in the citric
acid cycle with applications to 13C NMR studies in perfusedrat heartsrdquo Journal of Biological Chemistry vol 258 no 22 pp13785ndash13794 1983
[19] D HWilliamson P Lund and H A Krebs ldquoThe redox state offree nicotinamide-adenine dinucleotide in the cytoplasm andmitochondria of rat liverrdquo Biochemical Journal vol 103 no 2pp 514ndash527 1967
[20] L Suardet A C Gaide J M Calmes et al ldquoResponsiveness ofthree newly established human colorectal cancer cell lines totransforming growth factors 1205731 and 12057321rdquo Cancer Research vol52 no 13 pp 3705ndash3712 1992
[21] R A Carvalho T B Rodrigues P Zhao F M H Jeffrey C RMalloy and A D Sherry ldquoA 13C isotopomer kinetic analysisof cardiac metabolism influence of altered cytosolic redoxand [Ca2+]
119900rdquo The American Journal of PhysiologymdashHeart and
Circulatory Physiology vol 287 no 2 pp H889ndashH895 2004[22] AM Abrantes M E S Serra A C Goncalves et al ldquoHypoxia-
induced redox alterations and their correlation with 99mTc-MIBI and 99mTc-HL-91 uptake in colon cancer cellsrdquo NuclearMedicine and Biology vol 37 no 2 pp 125ndash132 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 5
00
10
20
30
40
50
60
70
80
90
100 WiDrC
once
ntra
tion
(mM
)
R2= 09945
R2= 09849
R2= 09571
lowast
0 2 4 6 8 10
HG-normoxiaHG-hypoxia LG-hypoxia
R2= 09922
lowast
LG-normoxia
(a)
00
10
20
30
40
50
60
70
80
90
100 LS1034
Con
cent
ratio
n (m
M)
R2= 09272
lowast
R2= 09608
lowast
R2= 07966
lowast
R2= 09998
lowast
0 2 4 6 8
HG-normoxiaHG-hypoxia LG-hypoxia
LG-normoxia
(b)
00
10
20
30
40
50
60
70
80
90
100 C2BBe1
R2= 09694
lowast
R2= 09952
R2= 09880
R2= 09720
lowast
0 2 4 6 8 10
HG-normoxiaHG-hypoxia LG-hypoxia
LG-normoxia
Con
cent
ratio
n (m
M)
(c)
Figure 3 [U-13C]lactate exported by colon cancer cell lines to the incubation media during the first 8 hours of incubation under normoxiaand hypoxia and low and high glucose media (a) WiDr (b) LS1034 (c) C2BBe1 Significant differences are indicated by lowast corresponding to119875 lt 005 (comparison between normoxia and hypoxia conditions) and corresponding to 119875 lt 005 (comparison between high and low cellmedia glucose concentrations (5 and 25mM))
300
200
100
0
WiDr LS1034 C2BBe1
lowastlowast
Dou
blin
g tim
e (h)
Figure 4 Duplication times (h) for the three colorectal cancer cell lines
the composition of the C4-Glu multiplet in terms of duplet45 (D
45) and quartet (Q) multiplets which as mentioned
correlate with Krebs cycle turnover ratesThe levels of alanineare intimately associated with the levels of pyruvate in thecells and can be used instead for deriving information con-cerning the cytosolic redox status [19] In fact C3-LacC3-Ala
is frequently used as an indirect measure of NADHNAD+ratio [20 21] The more the NADHH+ exists in cytosolthe more extensive the conversion of pyruvate is to lactatethrough lactic dehydrogenase (LDH) and the higher theratio is of those two metabolites In terms of C4-Glu theappearance of the Q multiplet is only possible after multiple
6 BioMed Research International
C4-Glu
C3-Glu
C3-Ala
C3-Ala
C3-Lac
C2-Ala
WiDr
LS1034
C2BBe1
(ppm)55 50 45 40 35 30 25 20
(a)
(b)
(c)
Q Q Q Q
D45 D45
Figure 5 13C NMR spectra of perchloric acid extracts for the three colorectal cancer cell lines under normoxia and high glucose media (a)WiDr (b) LS1034 (c) C2BBe1 Expanded is the multiplet due to glutamate carbon 4 (C4-Glu 342 ppm) composed by two major multipletsduplet 45 (D
45) due to glutamate isotopomers labeled in carbons 4 and 5 but not carbon 3 and quartet (Q) representing all glutamate
isotopomers with those three carbons enriched simultaneously Other resonances easily seen are the methyl carbons of lactate (C3-Lac208 ppm) and alanine (C3-Ala 170 ppm) and the carbon 3 of glutamate (C3-Glu 278 ppm)
0
10
20
30
40
WiDr LS1034 C2BBe1
lowastlowast
C3-L
acC3
-Ala
(a)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(b)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(c)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(d)Figure 6 Ratios of the methyl carbon of lactate (C3-Lac) to the methyl carbon of alanine (C3-Ala) for the three colorectal cancer cell linesunder all experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
BioMed Research International 7
00
05
10
15
20
25
WiDr LS1034 C2BBe1
C4Q
C4D45
(a)
WiDr LS1034 C2BBe1
lowast
00
05
10
15
20
25
C4Q
C4D45
(b)
WiDr LS1034 C2BBe100
05
10
15
20
25
C4Q
C4D45
(c)
WiDr LS1034 C2BBe1
lowast
00
05
10
15
20
25
C4Q
C4D45
(d)
Figure 7 Ratios of the glutamate C4 quartet (C4Q) to the duplet 45 (D45) multiplets for the three colorectal cancer cell lines under all
experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
turns of the Krebs cycle (Figure 1) and its contribution to theoverall C4-Glu multiplet is higher in cells with a more activeoxidative metabolism The ratio C4QC4D
45provides a way
to probe Krebs cycle kinetics and to compare different celllines in terms of their glycolyticoxidative metabolic charac-ters Another ratio of metabolic interest is the C3-LacC4-Glu since it reports how enriched the glycolyticoxidativemetabolite pools are and in such a way provides a measureof themetabolic coupling between glycolysis and Krebs cycleThe higher the ratio is the more uncoupled those two centralmetabolic pathways are and the more dependent the cell lineis on glycolysis for its bioenergetics requirements This ratiois frequently high in ldquoWarburgrdquo like cancer cell lines like theones used in this study
Figure 6 shows the ratios of C3-LacC3-Ala for thethree colon cancer cell lines in the four experimental con-ditions This ratio is significantly higher in WiDr undernormoxichigh glucose conditions than in the other twocell lines This correlates with a more pronounced glycolyticflux in this cancer cell line under such conditions Underhypoxia and high glucose there is a considerable attenuationof C3-LacC3-Ala ratio in WiDr only explainable by anoverall metabolic inhibition in these cells For LS1034 andC2BBe1 there is an increase in the C3-LacC3-Ala ratio underhypoxia explainable by a higher dependence on glycolysis forbioenergetics under anaerobic conditions
Figure 7 shows the ratios C4QC4D45
for the three col-orectal cancer cell lines in the four experimental conditionschosen This ratio clearly emphasizes that the three cell lineshave distinct Krebs cycle turnovers under hypoxiaWiDr andLS1034 cell lines suffer a considerable reduction in such ratioat both high and low glucose conditions The same does nothold however for the C2BBe1 which essentially shows noeffect in such ratio due to hypoxia This distinctive behavioremphasizes a distinct capacity of the cells for usage of oxygenat low concentration levels such as the ones prompted by thehypoxic episode and concomitantly justifies the distinctivebehavior of cells in tumors when subjected to hypoxia [22]
Figure 8 shows the C3-LacC4-Glu ratio for the threecolon cancer cell lines in the four experimental conditions Asexpected this ratio is higher formore glycolytic cell lines likeWiDr and suffers a slight increase under hypoxia Under nor-moxia andhigh glucose the LS1034 has the lowest C3-LacC4-Glu ratio in accordancewith amuchmore pronounced oxida-tive profile Upon hypoxia the LS1034 is the one sufferingthe more severe metabolic shift since the oxidative pathwayreduces its contribution to cell bioenergetics and has to becompensated by further glycolytic activity
34 Complex IV Activities In order to corroborate themetabolic alterations mentioned above for each of thecolorectal cancer cell lines and strengthen our findings
8 BioMed Research International
0
20
40
60
WiDr LS1034 C2BBe1
lowastlowast
C3-L
acC4
-Glu
(a)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(b)
WiDr LS1034 C2BBe10
20
40
60
C3-L
acC4
-Glu
(c)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(d)
Figure 8 Ratios of themethyl carbon of lactate (C3-Lac) to the carbon 4 of glutamate (C4-Glu) for the three colorectal cancer cell lines underall experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
concerning the metabolic (un)coupling observed for eachof the cell lines in each of the metabolic conditions theactivity of complex IV from the electron transport chain wasalso evaluatedThis activity adequately normalized for citratesynthase activity is presented in Figure 9 Under normoxiaand high glucose LS1034 cell line is the one possessingthe most active complex IV This is in accordance withall presented data which pointed these cells as the mostoxidativeless glycolytic under such incubation conditionsUnder normoxia and low glucose no significant changes weredetected in complex IV activities in accordance with theabsence of significant differences on the ratios C3-AlaC4-Glu and C4QC4D
45discussed above for this experimental
condition Low substrate concentration does not seem byitself to be able to distinguish the three colorectal cell linesunder analysis Hypoxia on the other hand prompts fordistinction even at low glucose levels thus emphasizing thedistinctive metabolic character of each colorectal cancer cellline This distinct responsiveness is a crucial parameter tobe evaluated aiming at the implementation of more efficientanticancer therapeutic strategies
The implemented methodology for analysis of both cul-ture media and perchloric acid cellular extracts providesunique and complimentary metabolic information which
affords a more complete picture of the metabolic transforma-tions associated with each of the specific actions undertakenfor challenging cancer cell metabolism This methodologycan naturally be extended to other metabolic pathways andmetabolic tracers towards widening the scopes of the currentmetabolic analysis
4 Conclusions
The three colorectal cancer cell lines show very distinctmetabolic profiles in the four experimental conditions testedGlycolysis is in every circumstance the most active metabolicpathway denoting the ldquoWarburgrdquo likemetabolic behavior thatcharacterizes many cancer cells Nevertheless oxidative fluxthrough the Krebs cycle assumes its importance in whole cel-lular bioenergetics and its extent is itself very distinct amongthe three colorectal cancer cell lines The interplay of oxygen(normoxia versus hypoxia) and nutrient (high versus lowglucose) availability also plays a crucial role in the definitionof cell capacity to adapt to distinct microenvironments Inthis study the (in)sensibility to the three colorectal cancercell lines was surprisingly different The less glycolytic undernormoxia LS1034 turns into the most glycolytic in hypoxiawhile themost glycolytic barely respond to the hypoxic insult
BioMed Research International 9
00
02
04
06
08
10
WiDr LS1034 C2BBe1
lowast lowast
Com
plex
IVac
tivity
(a)
WiDr LS1034 C2BBe1
lowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(b)
WiDr LS1034 C2BBe100
02
04
06
08
10
Com
plex
IVac
tivity
(c)
WiDr LS1034 C2BBe1
lowastlowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(d)
Figure 9 Complex IV activities for the three colorectal cancer cell lines under all experimental conditions (a) normoxiahigh glucose(b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
Also the alteration in nutrient availability interferes in themetabolic coupling between glycolysis and Krebs cycle anddemonstrates the distinct capacity that cells have to react tosuch interferences We understand that the whole picture ismuch more complex but would like to point out that thesimple management of O
2and glucose constitutes certainly a
mechanism to take into account to tackle tumor proliferationand devise more efficient therapeutic strategiesThe adequateknowledge of the metabolic behavior of cancer cells is thus amust on this ldquocrusaderdquo against cancer and the methodologyhere described is certainly pivotal for deriving this much-needed information
Conflict of Interests
The authors declare that there is no conflict of interests re-garding the publication of this paper
Authorsrsquo Contribution
Ana M Abrantes Ludgero C Tavares Salome Pires andCandidaMendes performed the experimentsManuela Graz-ina Rui A Carvalho and Maria Filomena Botelho analyzedresults andmade the figures AnaM Abrantes Manuela Gra-zina Rui A Carvalho andMaria Filomena Botelho designed
the research and wrote the paper Joao Casalta-Lopes per-formed statistical analyses
Acknowledgments
The authors are grateful to FCT (Fundacao para a Ciencia eTecnologiamdashPortugal) for financial support (PTDCEBB-EBI1158102009) The authors also acknowledge the ldquoRedeNacional de RMNrdquo (REDE1517RMN2005) for access tothe NMR spectrometer and facilities at Coimbra Uni-versity The authors thank Centre of Investigation onEnvironment Genetics and Oncobiology (CIMAGO) forfinancial support Salome Pires and Ludgero Tavares arerecipients of fellowships from the Portuguese Foundationfor Science and Technology (SFRHBD753002010) and(SFRHBD666002009) respectively
References
[1] OWarburg ldquoOrigin of cancer cellsrdquoOncologia vol 9 no 2 pp75ndash83 1956 (German)
[2] S Weinhouse O Warburg D Burk and A L Schade ldquoOn res-piratory impairment in cancer cellsrdquo Science vol 124 no 3215pp 267ndash272 1956
10 BioMed Research International
[3] Y Zhao E B Butler andMTan ldquoTargeting cellularmetabolismto improve cancer therapeuticsrdquo Cell Death and Disease vol 4no 3 article e532 2013
[4] C V Dang ldquoLinks between metabolism and cancerrdquo Genes ampDevelopment vol 26 no 9 pp 877ndash890 2012
[5] M Talekar S R Boreddy A Singh andMAmiji ldquoTumor aero-bic glycolysis new insights into therapeutic strategies with targ-eted deliveryrdquo Expert Opinion on Biological Therapy 2014
[6] R Xu H Pelicano Y Zhou et al ldquoInhibition of glycolysis incancer cells a novel strategy to overcome drug resistance asso-ciatedwithmitochondrial respiratory defect and hypoxiardquoCan-cer Research vol 65 no 2 pp 613ndash621 2005
[7] S Ros and A Schulze ldquoBalancing glycolytic flux the roleof 6-phosphofructo-2-kinasefructose 26-bisphosphatases incancer metabolismrdquoCancer ampMetabolism vol 1 article 8 2013
[8] E Roudier and A Perrin ldquoConsidering the role of pyruvate intumor cells during hypoxiardquo Biochimica et Biophysica Acta vol1796 no 2 pp 55ndash62 2009
[9] K J Williams B A Telfer R E Airley et al ldquoA protective rolefor HIF-1 in response to redox manipulation and glucose depr-ivation implications for tumorigenesisrdquo Oncogene vol 21 no2 pp 282ndash290 2002
[10] M Achison and T R Hupp ldquoHypoxia attenuates the p53 resp-onse to cellular damagerdquo Oncogene vol 22 no 22 pp 3431ndash3440 2003
[11] H L Kotze E G Armitage K J Sharkey J W AllwoodW B Dunn and K J Williams ldquoA novel untargeted metabol-omics correlation-based network analysis incorporating humanmetabolic reconstructionsrdquo BMC Systems Biology vol 7 article107 2013
[12] Y-A Wen P D Stevens M L Gasser R Andrei and T GaoldquoDownregulation of PHLPP expression contributes to hypoxia-induced resistance to chemotherapy in colon cancer cellsrdquoMolecular and Cellular Biology vol 33 no 22 pp 4594ndash46052013
[13] B Garcia-Barreno T Delgado B Akerlind-Stopner E Norrbyand J A Melero ldquoLocation of the epitope recognized by mono-clonal antibody 63G on the primary structure of human resp-iratory syncytial virus G glycoprotein and the ability of syn-thetic peptides containing this epitope to induce neutralizingantibodiesrdquo Journal of General Virology vol 73 no 10 pp 2625ndash2630 1992
[14] I F Duarte J Marques A F Ladeirinha et al ldquoAnalyticalapproaches toward successful human cell metabolome studiesby NMR spectroscopyrdquo Analytical Chemistry vol 81 no 12 pp5023ndash5032 2009
[15] S L Pereira J Ramalho-Santos A F Branco V A Sardao P JOliveira and R A Carvalho ldquoMetabolic remodeling duringH9c2 myoblast differentiation relevance for in vitro toxicitystudiesrdquo Cardiovascular Toxicology vol 11 no 2 pp 180ndash1902011
[16] MD Basson IMModlin and J AMadri ldquoHuman enterocyte(Caco-2)migration ismodulated in vitro by extracellularmatrixcomposition and epidermal growth factorrdquo Journal of ClinicalInvestigation vol 90 no 1 pp 15ndash23 1992
[17] M D Peterson and M S Mooseker ldquoCharacterization of theenterocyte-like brush border cytoskeleton of the C2BBe clonesof the human intestinal cell line Caco-2rdquo Journal of Cell Sciencevol 102 no 3 pp 581ndash600 1992
[18] E M Chance S H Seeholzer K Kobayashi and J R Will-iamson ldquoMathematical analysis of isotope labeling in the citric
acid cycle with applications to 13C NMR studies in perfusedrat heartsrdquo Journal of Biological Chemistry vol 258 no 22 pp13785ndash13794 1983
[19] D HWilliamson P Lund and H A Krebs ldquoThe redox state offree nicotinamide-adenine dinucleotide in the cytoplasm andmitochondria of rat liverrdquo Biochemical Journal vol 103 no 2pp 514ndash527 1967
[20] L Suardet A C Gaide J M Calmes et al ldquoResponsiveness ofthree newly established human colorectal cancer cell lines totransforming growth factors 1205731 and 12057321rdquo Cancer Research vol52 no 13 pp 3705ndash3712 1992
[21] R A Carvalho T B Rodrigues P Zhao F M H Jeffrey C RMalloy and A D Sherry ldquoA 13C isotopomer kinetic analysisof cardiac metabolism influence of altered cytosolic redoxand [Ca2+]
119900rdquo The American Journal of PhysiologymdashHeart and
Circulatory Physiology vol 287 no 2 pp H889ndashH895 2004[22] AM Abrantes M E S Serra A C Goncalves et al ldquoHypoxia-
induced redox alterations and their correlation with 99mTc-MIBI and 99mTc-HL-91 uptake in colon cancer cellsrdquo NuclearMedicine and Biology vol 37 no 2 pp 125ndash132 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 BioMed Research International
C4-Glu
C3-Glu
C3-Ala
C3-Ala
C3-Lac
C2-Ala
WiDr
LS1034
C2BBe1
(ppm)55 50 45 40 35 30 25 20
(a)
(b)
(c)
Q Q Q Q
D45 D45
Figure 5 13C NMR spectra of perchloric acid extracts for the three colorectal cancer cell lines under normoxia and high glucose media (a)WiDr (b) LS1034 (c) C2BBe1 Expanded is the multiplet due to glutamate carbon 4 (C4-Glu 342 ppm) composed by two major multipletsduplet 45 (D
45) due to glutamate isotopomers labeled in carbons 4 and 5 but not carbon 3 and quartet (Q) representing all glutamate
isotopomers with those three carbons enriched simultaneously Other resonances easily seen are the methyl carbons of lactate (C3-Lac208 ppm) and alanine (C3-Ala 170 ppm) and the carbon 3 of glutamate (C3-Glu 278 ppm)
0
10
20
30
40
WiDr LS1034 C2BBe1
lowastlowast
C3-L
acC3
-Ala
(a)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(b)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(c)
WiDr LS1034 C2BBe10
10
20
30
40
C3-L
acC3
-Ala
(d)Figure 6 Ratios of the methyl carbon of lactate (C3-Lac) to the methyl carbon of alanine (C3-Ala) for the three colorectal cancer cell linesunder all experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
BioMed Research International 7
00
05
10
15
20
25
WiDr LS1034 C2BBe1
C4Q
C4D45
(a)
WiDr LS1034 C2BBe1
lowast
00
05
10
15
20
25
C4Q
C4D45
(b)
WiDr LS1034 C2BBe100
05
10
15
20
25
C4Q
C4D45
(c)
WiDr LS1034 C2BBe1
lowast
00
05
10
15
20
25
C4Q
C4D45
(d)
Figure 7 Ratios of the glutamate C4 quartet (C4Q) to the duplet 45 (D45) multiplets for the three colorectal cancer cell lines under all
experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
turns of the Krebs cycle (Figure 1) and its contribution to theoverall C4-Glu multiplet is higher in cells with a more activeoxidative metabolism The ratio C4QC4D
45provides a way
to probe Krebs cycle kinetics and to compare different celllines in terms of their glycolyticoxidative metabolic charac-ters Another ratio of metabolic interest is the C3-LacC4-Glu since it reports how enriched the glycolyticoxidativemetabolite pools are and in such a way provides a measureof themetabolic coupling between glycolysis and Krebs cycleThe higher the ratio is the more uncoupled those two centralmetabolic pathways are and the more dependent the cell lineis on glycolysis for its bioenergetics requirements This ratiois frequently high in ldquoWarburgrdquo like cancer cell lines like theones used in this study
Figure 6 shows the ratios of C3-LacC3-Ala for thethree colon cancer cell lines in the four experimental con-ditions This ratio is significantly higher in WiDr undernormoxichigh glucose conditions than in the other twocell lines This correlates with a more pronounced glycolyticflux in this cancer cell line under such conditions Underhypoxia and high glucose there is a considerable attenuationof C3-LacC3-Ala ratio in WiDr only explainable by anoverall metabolic inhibition in these cells For LS1034 andC2BBe1 there is an increase in the C3-LacC3-Ala ratio underhypoxia explainable by a higher dependence on glycolysis forbioenergetics under anaerobic conditions
Figure 7 shows the ratios C4QC4D45
for the three col-orectal cancer cell lines in the four experimental conditionschosen This ratio clearly emphasizes that the three cell lineshave distinct Krebs cycle turnovers under hypoxiaWiDr andLS1034 cell lines suffer a considerable reduction in such ratioat both high and low glucose conditions The same does nothold however for the C2BBe1 which essentially shows noeffect in such ratio due to hypoxia This distinctive behavioremphasizes a distinct capacity of the cells for usage of oxygenat low concentration levels such as the ones prompted by thehypoxic episode and concomitantly justifies the distinctivebehavior of cells in tumors when subjected to hypoxia [22]
Figure 8 shows the C3-LacC4-Glu ratio for the threecolon cancer cell lines in the four experimental conditions Asexpected this ratio is higher formore glycolytic cell lines likeWiDr and suffers a slight increase under hypoxia Under nor-moxia andhigh glucose the LS1034 has the lowest C3-LacC4-Glu ratio in accordancewith amuchmore pronounced oxida-tive profile Upon hypoxia the LS1034 is the one sufferingthe more severe metabolic shift since the oxidative pathwayreduces its contribution to cell bioenergetics and has to becompensated by further glycolytic activity
34 Complex IV Activities In order to corroborate themetabolic alterations mentioned above for each of thecolorectal cancer cell lines and strengthen our findings
8 BioMed Research International
0
20
40
60
WiDr LS1034 C2BBe1
lowastlowast
C3-L
acC4
-Glu
(a)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(b)
WiDr LS1034 C2BBe10
20
40
60
C3-L
acC4
-Glu
(c)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(d)
Figure 8 Ratios of themethyl carbon of lactate (C3-Lac) to the carbon 4 of glutamate (C4-Glu) for the three colorectal cancer cell lines underall experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
concerning the metabolic (un)coupling observed for eachof the cell lines in each of the metabolic conditions theactivity of complex IV from the electron transport chain wasalso evaluatedThis activity adequately normalized for citratesynthase activity is presented in Figure 9 Under normoxiaand high glucose LS1034 cell line is the one possessingthe most active complex IV This is in accordance withall presented data which pointed these cells as the mostoxidativeless glycolytic under such incubation conditionsUnder normoxia and low glucose no significant changes weredetected in complex IV activities in accordance with theabsence of significant differences on the ratios C3-AlaC4-Glu and C4QC4D
45discussed above for this experimental
condition Low substrate concentration does not seem byitself to be able to distinguish the three colorectal cell linesunder analysis Hypoxia on the other hand prompts fordistinction even at low glucose levels thus emphasizing thedistinctive metabolic character of each colorectal cancer cellline This distinct responsiveness is a crucial parameter tobe evaluated aiming at the implementation of more efficientanticancer therapeutic strategies
The implemented methodology for analysis of both cul-ture media and perchloric acid cellular extracts providesunique and complimentary metabolic information which
affords a more complete picture of the metabolic transforma-tions associated with each of the specific actions undertakenfor challenging cancer cell metabolism This methodologycan naturally be extended to other metabolic pathways andmetabolic tracers towards widening the scopes of the currentmetabolic analysis
4 Conclusions
The three colorectal cancer cell lines show very distinctmetabolic profiles in the four experimental conditions testedGlycolysis is in every circumstance the most active metabolicpathway denoting the ldquoWarburgrdquo likemetabolic behavior thatcharacterizes many cancer cells Nevertheless oxidative fluxthrough the Krebs cycle assumes its importance in whole cel-lular bioenergetics and its extent is itself very distinct amongthe three colorectal cancer cell lines The interplay of oxygen(normoxia versus hypoxia) and nutrient (high versus lowglucose) availability also plays a crucial role in the definitionof cell capacity to adapt to distinct microenvironments Inthis study the (in)sensibility to the three colorectal cancercell lines was surprisingly different The less glycolytic undernormoxia LS1034 turns into the most glycolytic in hypoxiawhile themost glycolytic barely respond to the hypoxic insult
BioMed Research International 9
00
02
04
06
08
10
WiDr LS1034 C2BBe1
lowast lowast
Com
plex
IVac
tivity
(a)
WiDr LS1034 C2BBe1
lowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(b)
WiDr LS1034 C2BBe100
02
04
06
08
10
Com
plex
IVac
tivity
(c)
WiDr LS1034 C2BBe1
lowastlowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(d)
Figure 9 Complex IV activities for the three colorectal cancer cell lines under all experimental conditions (a) normoxiahigh glucose(b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
Also the alteration in nutrient availability interferes in themetabolic coupling between glycolysis and Krebs cycle anddemonstrates the distinct capacity that cells have to react tosuch interferences We understand that the whole picture ismuch more complex but would like to point out that thesimple management of O
2and glucose constitutes certainly a
mechanism to take into account to tackle tumor proliferationand devise more efficient therapeutic strategiesThe adequateknowledge of the metabolic behavior of cancer cells is thus amust on this ldquocrusaderdquo against cancer and the methodologyhere described is certainly pivotal for deriving this much-needed information
Conflict of Interests
The authors declare that there is no conflict of interests re-garding the publication of this paper
Authorsrsquo Contribution
Ana M Abrantes Ludgero C Tavares Salome Pires andCandidaMendes performed the experimentsManuela Graz-ina Rui A Carvalho and Maria Filomena Botelho analyzedresults andmade the figures AnaM Abrantes Manuela Gra-zina Rui A Carvalho andMaria Filomena Botelho designed
the research and wrote the paper Joao Casalta-Lopes per-formed statistical analyses
Acknowledgments
The authors are grateful to FCT (Fundacao para a Ciencia eTecnologiamdashPortugal) for financial support (PTDCEBB-EBI1158102009) The authors also acknowledge the ldquoRedeNacional de RMNrdquo (REDE1517RMN2005) for access tothe NMR spectrometer and facilities at Coimbra Uni-versity The authors thank Centre of Investigation onEnvironment Genetics and Oncobiology (CIMAGO) forfinancial support Salome Pires and Ludgero Tavares arerecipients of fellowships from the Portuguese Foundationfor Science and Technology (SFRHBD753002010) and(SFRHBD666002009) respectively
References
[1] OWarburg ldquoOrigin of cancer cellsrdquoOncologia vol 9 no 2 pp75ndash83 1956 (German)
[2] S Weinhouse O Warburg D Burk and A L Schade ldquoOn res-piratory impairment in cancer cellsrdquo Science vol 124 no 3215pp 267ndash272 1956
10 BioMed Research International
[3] Y Zhao E B Butler andMTan ldquoTargeting cellularmetabolismto improve cancer therapeuticsrdquo Cell Death and Disease vol 4no 3 article e532 2013
[4] C V Dang ldquoLinks between metabolism and cancerrdquo Genes ampDevelopment vol 26 no 9 pp 877ndash890 2012
[5] M Talekar S R Boreddy A Singh andMAmiji ldquoTumor aero-bic glycolysis new insights into therapeutic strategies with targ-eted deliveryrdquo Expert Opinion on Biological Therapy 2014
[6] R Xu H Pelicano Y Zhou et al ldquoInhibition of glycolysis incancer cells a novel strategy to overcome drug resistance asso-ciatedwithmitochondrial respiratory defect and hypoxiardquoCan-cer Research vol 65 no 2 pp 613ndash621 2005
[7] S Ros and A Schulze ldquoBalancing glycolytic flux the roleof 6-phosphofructo-2-kinasefructose 26-bisphosphatases incancer metabolismrdquoCancer ampMetabolism vol 1 article 8 2013
[8] E Roudier and A Perrin ldquoConsidering the role of pyruvate intumor cells during hypoxiardquo Biochimica et Biophysica Acta vol1796 no 2 pp 55ndash62 2009
[9] K J Williams B A Telfer R E Airley et al ldquoA protective rolefor HIF-1 in response to redox manipulation and glucose depr-ivation implications for tumorigenesisrdquo Oncogene vol 21 no2 pp 282ndash290 2002
[10] M Achison and T R Hupp ldquoHypoxia attenuates the p53 resp-onse to cellular damagerdquo Oncogene vol 22 no 22 pp 3431ndash3440 2003
[11] H L Kotze E G Armitage K J Sharkey J W AllwoodW B Dunn and K J Williams ldquoA novel untargeted metabol-omics correlation-based network analysis incorporating humanmetabolic reconstructionsrdquo BMC Systems Biology vol 7 article107 2013
[12] Y-A Wen P D Stevens M L Gasser R Andrei and T GaoldquoDownregulation of PHLPP expression contributes to hypoxia-induced resistance to chemotherapy in colon cancer cellsrdquoMolecular and Cellular Biology vol 33 no 22 pp 4594ndash46052013
[13] B Garcia-Barreno T Delgado B Akerlind-Stopner E Norrbyand J A Melero ldquoLocation of the epitope recognized by mono-clonal antibody 63G on the primary structure of human resp-iratory syncytial virus G glycoprotein and the ability of syn-thetic peptides containing this epitope to induce neutralizingantibodiesrdquo Journal of General Virology vol 73 no 10 pp 2625ndash2630 1992
[14] I F Duarte J Marques A F Ladeirinha et al ldquoAnalyticalapproaches toward successful human cell metabolome studiesby NMR spectroscopyrdquo Analytical Chemistry vol 81 no 12 pp5023ndash5032 2009
[15] S L Pereira J Ramalho-Santos A F Branco V A Sardao P JOliveira and R A Carvalho ldquoMetabolic remodeling duringH9c2 myoblast differentiation relevance for in vitro toxicitystudiesrdquo Cardiovascular Toxicology vol 11 no 2 pp 180ndash1902011
[16] MD Basson IMModlin and J AMadri ldquoHuman enterocyte(Caco-2)migration ismodulated in vitro by extracellularmatrixcomposition and epidermal growth factorrdquo Journal of ClinicalInvestigation vol 90 no 1 pp 15ndash23 1992
[17] M D Peterson and M S Mooseker ldquoCharacterization of theenterocyte-like brush border cytoskeleton of the C2BBe clonesof the human intestinal cell line Caco-2rdquo Journal of Cell Sciencevol 102 no 3 pp 581ndash600 1992
[18] E M Chance S H Seeholzer K Kobayashi and J R Will-iamson ldquoMathematical analysis of isotope labeling in the citric
acid cycle with applications to 13C NMR studies in perfusedrat heartsrdquo Journal of Biological Chemistry vol 258 no 22 pp13785ndash13794 1983
[19] D HWilliamson P Lund and H A Krebs ldquoThe redox state offree nicotinamide-adenine dinucleotide in the cytoplasm andmitochondria of rat liverrdquo Biochemical Journal vol 103 no 2pp 514ndash527 1967
[20] L Suardet A C Gaide J M Calmes et al ldquoResponsiveness ofthree newly established human colorectal cancer cell lines totransforming growth factors 1205731 and 12057321rdquo Cancer Research vol52 no 13 pp 3705ndash3712 1992
[21] R A Carvalho T B Rodrigues P Zhao F M H Jeffrey C RMalloy and A D Sherry ldquoA 13C isotopomer kinetic analysisof cardiac metabolism influence of altered cytosolic redoxand [Ca2+]
119900rdquo The American Journal of PhysiologymdashHeart and
Circulatory Physiology vol 287 no 2 pp H889ndashH895 2004[22] AM Abrantes M E S Serra A C Goncalves et al ldquoHypoxia-
induced redox alterations and their correlation with 99mTc-MIBI and 99mTc-HL-91 uptake in colon cancer cellsrdquo NuclearMedicine and Biology vol 37 no 2 pp 125ndash132 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

BioMed Research International 7
00
05
10
15
20
25
WiDr LS1034 C2BBe1
C4Q
C4D45
(a)
WiDr LS1034 C2BBe1
lowast
00
05
10
15
20
25
C4Q
C4D45
(b)
WiDr LS1034 C2BBe100
05
10
15
20
25
C4Q
C4D45
(c)
WiDr LS1034 C2BBe1
lowast
00
05
10
15
20
25
C4Q
C4D45
(d)
Figure 7 Ratios of the glutamate C4 quartet (C4Q) to the duplet 45 (D45) multiplets for the three colorectal cancer cell lines under all
experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
turns of the Krebs cycle (Figure 1) and its contribution to theoverall C4-Glu multiplet is higher in cells with a more activeoxidative metabolism The ratio C4QC4D
45provides a way
to probe Krebs cycle kinetics and to compare different celllines in terms of their glycolyticoxidative metabolic charac-ters Another ratio of metabolic interest is the C3-LacC4-Glu since it reports how enriched the glycolyticoxidativemetabolite pools are and in such a way provides a measureof themetabolic coupling between glycolysis and Krebs cycleThe higher the ratio is the more uncoupled those two centralmetabolic pathways are and the more dependent the cell lineis on glycolysis for its bioenergetics requirements This ratiois frequently high in ldquoWarburgrdquo like cancer cell lines like theones used in this study
Figure 6 shows the ratios of C3-LacC3-Ala for thethree colon cancer cell lines in the four experimental con-ditions This ratio is significantly higher in WiDr undernormoxichigh glucose conditions than in the other twocell lines This correlates with a more pronounced glycolyticflux in this cancer cell line under such conditions Underhypoxia and high glucose there is a considerable attenuationof C3-LacC3-Ala ratio in WiDr only explainable by anoverall metabolic inhibition in these cells For LS1034 andC2BBe1 there is an increase in the C3-LacC3-Ala ratio underhypoxia explainable by a higher dependence on glycolysis forbioenergetics under anaerobic conditions
Figure 7 shows the ratios C4QC4D45
for the three col-orectal cancer cell lines in the four experimental conditionschosen This ratio clearly emphasizes that the three cell lineshave distinct Krebs cycle turnovers under hypoxiaWiDr andLS1034 cell lines suffer a considerable reduction in such ratioat both high and low glucose conditions The same does nothold however for the C2BBe1 which essentially shows noeffect in such ratio due to hypoxia This distinctive behavioremphasizes a distinct capacity of the cells for usage of oxygenat low concentration levels such as the ones prompted by thehypoxic episode and concomitantly justifies the distinctivebehavior of cells in tumors when subjected to hypoxia [22]
Figure 8 shows the C3-LacC4-Glu ratio for the threecolon cancer cell lines in the four experimental conditions Asexpected this ratio is higher formore glycolytic cell lines likeWiDr and suffers a slight increase under hypoxia Under nor-moxia andhigh glucose the LS1034 has the lowest C3-LacC4-Glu ratio in accordancewith amuchmore pronounced oxida-tive profile Upon hypoxia the LS1034 is the one sufferingthe more severe metabolic shift since the oxidative pathwayreduces its contribution to cell bioenergetics and has to becompensated by further glycolytic activity
34 Complex IV Activities In order to corroborate themetabolic alterations mentioned above for each of thecolorectal cancer cell lines and strengthen our findings
8 BioMed Research International
0
20
40
60
WiDr LS1034 C2BBe1
lowastlowast
C3-L
acC4
-Glu
(a)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(b)
WiDr LS1034 C2BBe10
20
40
60
C3-L
acC4
-Glu
(c)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(d)
Figure 8 Ratios of themethyl carbon of lactate (C3-Lac) to the carbon 4 of glutamate (C4-Glu) for the three colorectal cancer cell lines underall experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
concerning the metabolic (un)coupling observed for eachof the cell lines in each of the metabolic conditions theactivity of complex IV from the electron transport chain wasalso evaluatedThis activity adequately normalized for citratesynthase activity is presented in Figure 9 Under normoxiaand high glucose LS1034 cell line is the one possessingthe most active complex IV This is in accordance withall presented data which pointed these cells as the mostoxidativeless glycolytic under such incubation conditionsUnder normoxia and low glucose no significant changes weredetected in complex IV activities in accordance with theabsence of significant differences on the ratios C3-AlaC4-Glu and C4QC4D
45discussed above for this experimental
condition Low substrate concentration does not seem byitself to be able to distinguish the three colorectal cell linesunder analysis Hypoxia on the other hand prompts fordistinction even at low glucose levels thus emphasizing thedistinctive metabolic character of each colorectal cancer cellline This distinct responsiveness is a crucial parameter tobe evaluated aiming at the implementation of more efficientanticancer therapeutic strategies
The implemented methodology for analysis of both cul-ture media and perchloric acid cellular extracts providesunique and complimentary metabolic information which
affords a more complete picture of the metabolic transforma-tions associated with each of the specific actions undertakenfor challenging cancer cell metabolism This methodologycan naturally be extended to other metabolic pathways andmetabolic tracers towards widening the scopes of the currentmetabolic analysis
4 Conclusions
The three colorectal cancer cell lines show very distinctmetabolic profiles in the four experimental conditions testedGlycolysis is in every circumstance the most active metabolicpathway denoting the ldquoWarburgrdquo likemetabolic behavior thatcharacterizes many cancer cells Nevertheless oxidative fluxthrough the Krebs cycle assumes its importance in whole cel-lular bioenergetics and its extent is itself very distinct amongthe three colorectal cancer cell lines The interplay of oxygen(normoxia versus hypoxia) and nutrient (high versus lowglucose) availability also plays a crucial role in the definitionof cell capacity to adapt to distinct microenvironments Inthis study the (in)sensibility to the three colorectal cancercell lines was surprisingly different The less glycolytic undernormoxia LS1034 turns into the most glycolytic in hypoxiawhile themost glycolytic barely respond to the hypoxic insult
BioMed Research International 9
00
02
04
06
08
10
WiDr LS1034 C2BBe1
lowast lowast
Com
plex
IVac
tivity
(a)
WiDr LS1034 C2BBe1
lowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(b)
WiDr LS1034 C2BBe100
02
04
06
08
10
Com
plex
IVac
tivity
(c)
WiDr LS1034 C2BBe1
lowastlowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(d)
Figure 9 Complex IV activities for the three colorectal cancer cell lines under all experimental conditions (a) normoxiahigh glucose(b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
Also the alteration in nutrient availability interferes in themetabolic coupling between glycolysis and Krebs cycle anddemonstrates the distinct capacity that cells have to react tosuch interferences We understand that the whole picture ismuch more complex but would like to point out that thesimple management of O
2and glucose constitutes certainly a
mechanism to take into account to tackle tumor proliferationand devise more efficient therapeutic strategiesThe adequateknowledge of the metabolic behavior of cancer cells is thus amust on this ldquocrusaderdquo against cancer and the methodologyhere described is certainly pivotal for deriving this much-needed information
Conflict of Interests
The authors declare that there is no conflict of interests re-garding the publication of this paper
Authorsrsquo Contribution
Ana M Abrantes Ludgero C Tavares Salome Pires andCandidaMendes performed the experimentsManuela Graz-ina Rui A Carvalho and Maria Filomena Botelho analyzedresults andmade the figures AnaM Abrantes Manuela Gra-zina Rui A Carvalho andMaria Filomena Botelho designed
the research and wrote the paper Joao Casalta-Lopes per-formed statistical analyses
Acknowledgments
The authors are grateful to FCT (Fundacao para a Ciencia eTecnologiamdashPortugal) for financial support (PTDCEBB-EBI1158102009) The authors also acknowledge the ldquoRedeNacional de RMNrdquo (REDE1517RMN2005) for access tothe NMR spectrometer and facilities at Coimbra Uni-versity The authors thank Centre of Investigation onEnvironment Genetics and Oncobiology (CIMAGO) forfinancial support Salome Pires and Ludgero Tavares arerecipients of fellowships from the Portuguese Foundationfor Science and Technology (SFRHBD753002010) and(SFRHBD666002009) respectively
References
[1] OWarburg ldquoOrigin of cancer cellsrdquoOncologia vol 9 no 2 pp75ndash83 1956 (German)
[2] S Weinhouse O Warburg D Burk and A L Schade ldquoOn res-piratory impairment in cancer cellsrdquo Science vol 124 no 3215pp 267ndash272 1956
10 BioMed Research International
[3] Y Zhao E B Butler andMTan ldquoTargeting cellularmetabolismto improve cancer therapeuticsrdquo Cell Death and Disease vol 4no 3 article e532 2013
[4] C V Dang ldquoLinks between metabolism and cancerrdquo Genes ampDevelopment vol 26 no 9 pp 877ndash890 2012
[5] M Talekar S R Boreddy A Singh andMAmiji ldquoTumor aero-bic glycolysis new insights into therapeutic strategies with targ-eted deliveryrdquo Expert Opinion on Biological Therapy 2014
[6] R Xu H Pelicano Y Zhou et al ldquoInhibition of glycolysis incancer cells a novel strategy to overcome drug resistance asso-ciatedwithmitochondrial respiratory defect and hypoxiardquoCan-cer Research vol 65 no 2 pp 613ndash621 2005
[7] S Ros and A Schulze ldquoBalancing glycolytic flux the roleof 6-phosphofructo-2-kinasefructose 26-bisphosphatases incancer metabolismrdquoCancer ampMetabolism vol 1 article 8 2013
[8] E Roudier and A Perrin ldquoConsidering the role of pyruvate intumor cells during hypoxiardquo Biochimica et Biophysica Acta vol1796 no 2 pp 55ndash62 2009
[9] K J Williams B A Telfer R E Airley et al ldquoA protective rolefor HIF-1 in response to redox manipulation and glucose depr-ivation implications for tumorigenesisrdquo Oncogene vol 21 no2 pp 282ndash290 2002
[10] M Achison and T R Hupp ldquoHypoxia attenuates the p53 resp-onse to cellular damagerdquo Oncogene vol 22 no 22 pp 3431ndash3440 2003
[11] H L Kotze E G Armitage K J Sharkey J W AllwoodW B Dunn and K J Williams ldquoA novel untargeted metabol-omics correlation-based network analysis incorporating humanmetabolic reconstructionsrdquo BMC Systems Biology vol 7 article107 2013
[12] Y-A Wen P D Stevens M L Gasser R Andrei and T GaoldquoDownregulation of PHLPP expression contributes to hypoxia-induced resistance to chemotherapy in colon cancer cellsrdquoMolecular and Cellular Biology vol 33 no 22 pp 4594ndash46052013
[13] B Garcia-Barreno T Delgado B Akerlind-Stopner E Norrbyand J A Melero ldquoLocation of the epitope recognized by mono-clonal antibody 63G on the primary structure of human resp-iratory syncytial virus G glycoprotein and the ability of syn-thetic peptides containing this epitope to induce neutralizingantibodiesrdquo Journal of General Virology vol 73 no 10 pp 2625ndash2630 1992
[14] I F Duarte J Marques A F Ladeirinha et al ldquoAnalyticalapproaches toward successful human cell metabolome studiesby NMR spectroscopyrdquo Analytical Chemistry vol 81 no 12 pp5023ndash5032 2009
[15] S L Pereira J Ramalho-Santos A F Branco V A Sardao P JOliveira and R A Carvalho ldquoMetabolic remodeling duringH9c2 myoblast differentiation relevance for in vitro toxicitystudiesrdquo Cardiovascular Toxicology vol 11 no 2 pp 180ndash1902011
[16] MD Basson IMModlin and J AMadri ldquoHuman enterocyte(Caco-2)migration ismodulated in vitro by extracellularmatrixcomposition and epidermal growth factorrdquo Journal of ClinicalInvestigation vol 90 no 1 pp 15ndash23 1992
[17] M D Peterson and M S Mooseker ldquoCharacterization of theenterocyte-like brush border cytoskeleton of the C2BBe clonesof the human intestinal cell line Caco-2rdquo Journal of Cell Sciencevol 102 no 3 pp 581ndash600 1992
[18] E M Chance S H Seeholzer K Kobayashi and J R Will-iamson ldquoMathematical analysis of isotope labeling in the citric
acid cycle with applications to 13C NMR studies in perfusedrat heartsrdquo Journal of Biological Chemistry vol 258 no 22 pp13785ndash13794 1983
[19] D HWilliamson P Lund and H A Krebs ldquoThe redox state offree nicotinamide-adenine dinucleotide in the cytoplasm andmitochondria of rat liverrdquo Biochemical Journal vol 103 no 2pp 514ndash527 1967
[20] L Suardet A C Gaide J M Calmes et al ldquoResponsiveness ofthree newly established human colorectal cancer cell lines totransforming growth factors 1205731 and 12057321rdquo Cancer Research vol52 no 13 pp 3705ndash3712 1992
[21] R A Carvalho T B Rodrigues P Zhao F M H Jeffrey C RMalloy and A D Sherry ldquoA 13C isotopomer kinetic analysisof cardiac metabolism influence of altered cytosolic redoxand [Ca2+]
119900rdquo The American Journal of PhysiologymdashHeart and
Circulatory Physiology vol 287 no 2 pp H889ndashH895 2004[22] AM Abrantes M E S Serra A C Goncalves et al ldquoHypoxia-
induced redox alterations and their correlation with 99mTc-MIBI and 99mTc-HL-91 uptake in colon cancer cellsrdquo NuclearMedicine and Biology vol 37 no 2 pp 125ndash132 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 BioMed Research International
0
20
40
60
WiDr LS1034 C2BBe1
lowastlowast
C3-L
acC4
-Glu
(a)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(b)
WiDr LS1034 C2BBe10
20
40
60
C3-L
acC4
-Glu
(c)
WiDr LS1034 C2BBe1
lowast
0
20
40
60
C3-L
acC4
-Glu
(d)
Figure 8 Ratios of themethyl carbon of lactate (C3-Lac) to the carbon 4 of glutamate (C4-Glu) for the three colorectal cancer cell lines underall experimental conditions (a) normoxiahigh glucose (b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
concerning the metabolic (un)coupling observed for eachof the cell lines in each of the metabolic conditions theactivity of complex IV from the electron transport chain wasalso evaluatedThis activity adequately normalized for citratesynthase activity is presented in Figure 9 Under normoxiaand high glucose LS1034 cell line is the one possessingthe most active complex IV This is in accordance withall presented data which pointed these cells as the mostoxidativeless glycolytic under such incubation conditionsUnder normoxia and low glucose no significant changes weredetected in complex IV activities in accordance with theabsence of significant differences on the ratios C3-AlaC4-Glu and C4QC4D
45discussed above for this experimental
condition Low substrate concentration does not seem byitself to be able to distinguish the three colorectal cell linesunder analysis Hypoxia on the other hand prompts fordistinction even at low glucose levels thus emphasizing thedistinctive metabolic character of each colorectal cancer cellline This distinct responsiveness is a crucial parameter tobe evaluated aiming at the implementation of more efficientanticancer therapeutic strategies
The implemented methodology for analysis of both cul-ture media and perchloric acid cellular extracts providesunique and complimentary metabolic information which
affords a more complete picture of the metabolic transforma-tions associated with each of the specific actions undertakenfor challenging cancer cell metabolism This methodologycan naturally be extended to other metabolic pathways andmetabolic tracers towards widening the scopes of the currentmetabolic analysis
4 Conclusions
The three colorectal cancer cell lines show very distinctmetabolic profiles in the four experimental conditions testedGlycolysis is in every circumstance the most active metabolicpathway denoting the ldquoWarburgrdquo likemetabolic behavior thatcharacterizes many cancer cells Nevertheless oxidative fluxthrough the Krebs cycle assumes its importance in whole cel-lular bioenergetics and its extent is itself very distinct amongthe three colorectal cancer cell lines The interplay of oxygen(normoxia versus hypoxia) and nutrient (high versus lowglucose) availability also plays a crucial role in the definitionof cell capacity to adapt to distinct microenvironments Inthis study the (in)sensibility to the three colorectal cancercell lines was surprisingly different The less glycolytic undernormoxia LS1034 turns into the most glycolytic in hypoxiawhile themost glycolytic barely respond to the hypoxic insult
BioMed Research International 9
00
02
04
06
08
10
WiDr LS1034 C2BBe1
lowast lowast
Com
plex
IVac
tivity
(a)
WiDr LS1034 C2BBe1
lowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(b)
WiDr LS1034 C2BBe100
02
04
06
08
10
Com
plex
IVac
tivity
(c)
WiDr LS1034 C2BBe1
lowastlowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(d)
Figure 9 Complex IV activities for the three colorectal cancer cell lines under all experimental conditions (a) normoxiahigh glucose(b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
Also the alteration in nutrient availability interferes in themetabolic coupling between glycolysis and Krebs cycle anddemonstrates the distinct capacity that cells have to react tosuch interferences We understand that the whole picture ismuch more complex but would like to point out that thesimple management of O
2and glucose constitutes certainly a
mechanism to take into account to tackle tumor proliferationand devise more efficient therapeutic strategiesThe adequateknowledge of the metabolic behavior of cancer cells is thus amust on this ldquocrusaderdquo against cancer and the methodologyhere described is certainly pivotal for deriving this much-needed information
Conflict of Interests
The authors declare that there is no conflict of interests re-garding the publication of this paper
Authorsrsquo Contribution
Ana M Abrantes Ludgero C Tavares Salome Pires andCandidaMendes performed the experimentsManuela Graz-ina Rui A Carvalho and Maria Filomena Botelho analyzedresults andmade the figures AnaM Abrantes Manuela Gra-zina Rui A Carvalho andMaria Filomena Botelho designed
the research and wrote the paper Joao Casalta-Lopes per-formed statistical analyses
Acknowledgments
The authors are grateful to FCT (Fundacao para a Ciencia eTecnologiamdashPortugal) for financial support (PTDCEBB-EBI1158102009) The authors also acknowledge the ldquoRedeNacional de RMNrdquo (REDE1517RMN2005) for access tothe NMR spectrometer and facilities at Coimbra Uni-versity The authors thank Centre of Investigation onEnvironment Genetics and Oncobiology (CIMAGO) forfinancial support Salome Pires and Ludgero Tavares arerecipients of fellowships from the Portuguese Foundationfor Science and Technology (SFRHBD753002010) and(SFRHBD666002009) respectively
References
[1] OWarburg ldquoOrigin of cancer cellsrdquoOncologia vol 9 no 2 pp75ndash83 1956 (German)
[2] S Weinhouse O Warburg D Burk and A L Schade ldquoOn res-piratory impairment in cancer cellsrdquo Science vol 124 no 3215pp 267ndash272 1956
10 BioMed Research International
[3] Y Zhao E B Butler andMTan ldquoTargeting cellularmetabolismto improve cancer therapeuticsrdquo Cell Death and Disease vol 4no 3 article e532 2013
[4] C V Dang ldquoLinks between metabolism and cancerrdquo Genes ampDevelopment vol 26 no 9 pp 877ndash890 2012
[5] M Talekar S R Boreddy A Singh andMAmiji ldquoTumor aero-bic glycolysis new insights into therapeutic strategies with targ-eted deliveryrdquo Expert Opinion on Biological Therapy 2014
[6] R Xu H Pelicano Y Zhou et al ldquoInhibition of glycolysis incancer cells a novel strategy to overcome drug resistance asso-ciatedwithmitochondrial respiratory defect and hypoxiardquoCan-cer Research vol 65 no 2 pp 613ndash621 2005
[7] S Ros and A Schulze ldquoBalancing glycolytic flux the roleof 6-phosphofructo-2-kinasefructose 26-bisphosphatases incancer metabolismrdquoCancer ampMetabolism vol 1 article 8 2013
[8] E Roudier and A Perrin ldquoConsidering the role of pyruvate intumor cells during hypoxiardquo Biochimica et Biophysica Acta vol1796 no 2 pp 55ndash62 2009
[9] K J Williams B A Telfer R E Airley et al ldquoA protective rolefor HIF-1 in response to redox manipulation and glucose depr-ivation implications for tumorigenesisrdquo Oncogene vol 21 no2 pp 282ndash290 2002
[10] M Achison and T R Hupp ldquoHypoxia attenuates the p53 resp-onse to cellular damagerdquo Oncogene vol 22 no 22 pp 3431ndash3440 2003
[11] H L Kotze E G Armitage K J Sharkey J W AllwoodW B Dunn and K J Williams ldquoA novel untargeted metabol-omics correlation-based network analysis incorporating humanmetabolic reconstructionsrdquo BMC Systems Biology vol 7 article107 2013
[12] Y-A Wen P D Stevens M L Gasser R Andrei and T GaoldquoDownregulation of PHLPP expression contributes to hypoxia-induced resistance to chemotherapy in colon cancer cellsrdquoMolecular and Cellular Biology vol 33 no 22 pp 4594ndash46052013
[13] B Garcia-Barreno T Delgado B Akerlind-Stopner E Norrbyand J A Melero ldquoLocation of the epitope recognized by mono-clonal antibody 63G on the primary structure of human resp-iratory syncytial virus G glycoprotein and the ability of syn-thetic peptides containing this epitope to induce neutralizingantibodiesrdquo Journal of General Virology vol 73 no 10 pp 2625ndash2630 1992
[14] I F Duarte J Marques A F Ladeirinha et al ldquoAnalyticalapproaches toward successful human cell metabolome studiesby NMR spectroscopyrdquo Analytical Chemistry vol 81 no 12 pp5023ndash5032 2009
[15] S L Pereira J Ramalho-Santos A F Branco V A Sardao P JOliveira and R A Carvalho ldquoMetabolic remodeling duringH9c2 myoblast differentiation relevance for in vitro toxicitystudiesrdquo Cardiovascular Toxicology vol 11 no 2 pp 180ndash1902011
[16] MD Basson IMModlin and J AMadri ldquoHuman enterocyte(Caco-2)migration ismodulated in vitro by extracellularmatrixcomposition and epidermal growth factorrdquo Journal of ClinicalInvestigation vol 90 no 1 pp 15ndash23 1992
[17] M D Peterson and M S Mooseker ldquoCharacterization of theenterocyte-like brush border cytoskeleton of the C2BBe clonesof the human intestinal cell line Caco-2rdquo Journal of Cell Sciencevol 102 no 3 pp 581ndash600 1992
[18] E M Chance S H Seeholzer K Kobayashi and J R Will-iamson ldquoMathematical analysis of isotope labeling in the citric
acid cycle with applications to 13C NMR studies in perfusedrat heartsrdquo Journal of Biological Chemistry vol 258 no 22 pp13785ndash13794 1983
[19] D HWilliamson P Lund and H A Krebs ldquoThe redox state offree nicotinamide-adenine dinucleotide in the cytoplasm andmitochondria of rat liverrdquo Biochemical Journal vol 103 no 2pp 514ndash527 1967
[20] L Suardet A C Gaide J M Calmes et al ldquoResponsiveness ofthree newly established human colorectal cancer cell lines totransforming growth factors 1205731 and 12057321rdquo Cancer Research vol52 no 13 pp 3705ndash3712 1992
[21] R A Carvalho T B Rodrigues P Zhao F M H Jeffrey C RMalloy and A D Sherry ldquoA 13C isotopomer kinetic analysisof cardiac metabolism influence of altered cytosolic redoxand [Ca2+]
119900rdquo The American Journal of PhysiologymdashHeart and
Circulatory Physiology vol 287 no 2 pp H889ndashH895 2004[22] AM Abrantes M E S Serra A C Goncalves et al ldquoHypoxia-
induced redox alterations and their correlation with 99mTc-MIBI and 99mTc-HL-91 uptake in colon cancer cellsrdquo NuclearMedicine and Biology vol 37 no 2 pp 125ndash132 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
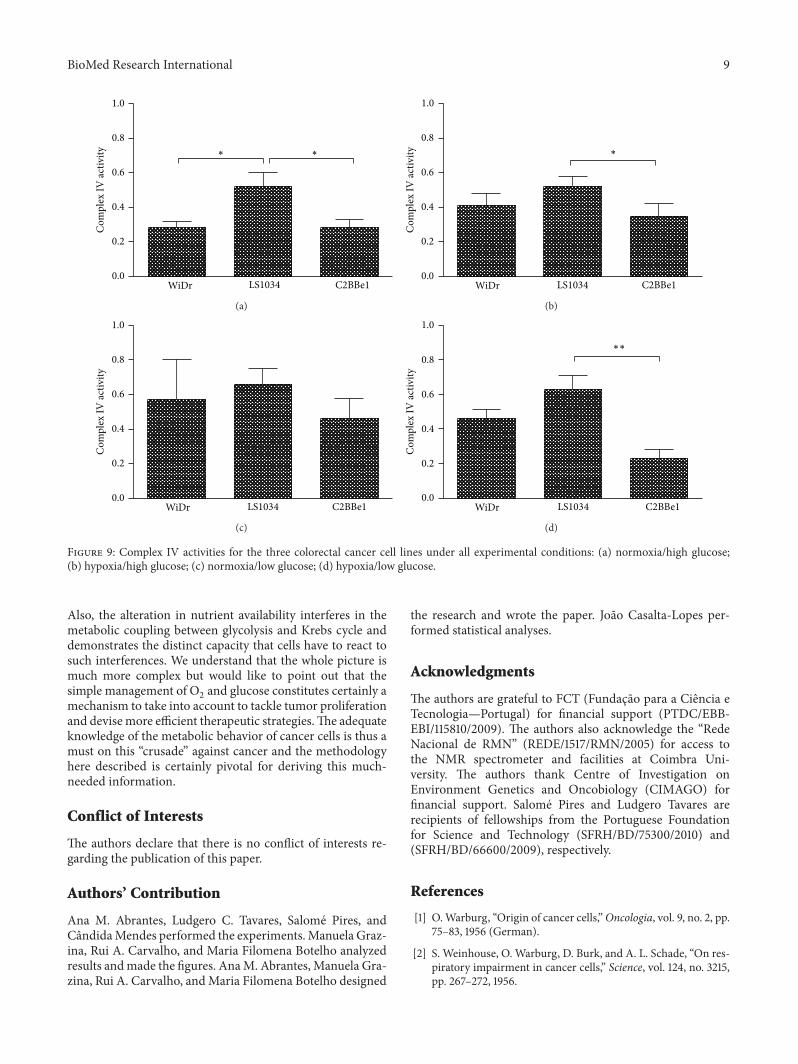
BioMed Research International 9
00
02
04
06
08
10
WiDr LS1034 C2BBe1
lowast lowast
Com
plex
IVac
tivity
(a)
WiDr LS1034 C2BBe1
lowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(b)
WiDr LS1034 C2BBe100
02
04
06
08
10
Com
plex
IVac
tivity
(c)
WiDr LS1034 C2BBe1
lowastlowast
00
02
04
06
08
10
Com
plex
IVac
tivity
(d)
Figure 9 Complex IV activities for the three colorectal cancer cell lines under all experimental conditions (a) normoxiahigh glucose(b) hypoxiahigh glucose (c) normoxialow glucose (d) hypoxialow glucose
Also the alteration in nutrient availability interferes in themetabolic coupling between glycolysis and Krebs cycle anddemonstrates the distinct capacity that cells have to react tosuch interferences We understand that the whole picture ismuch more complex but would like to point out that thesimple management of O
2and glucose constitutes certainly a
mechanism to take into account to tackle tumor proliferationand devise more efficient therapeutic strategiesThe adequateknowledge of the metabolic behavior of cancer cells is thus amust on this ldquocrusaderdquo against cancer and the methodologyhere described is certainly pivotal for deriving this much-needed information
Conflict of Interests
The authors declare that there is no conflict of interests re-garding the publication of this paper
Authorsrsquo Contribution
Ana M Abrantes Ludgero C Tavares Salome Pires andCandidaMendes performed the experimentsManuela Graz-ina Rui A Carvalho and Maria Filomena Botelho analyzedresults andmade the figures AnaM Abrantes Manuela Gra-zina Rui A Carvalho andMaria Filomena Botelho designed
the research and wrote the paper Joao Casalta-Lopes per-formed statistical analyses
Acknowledgments
The authors are grateful to FCT (Fundacao para a Ciencia eTecnologiamdashPortugal) for financial support (PTDCEBB-EBI1158102009) The authors also acknowledge the ldquoRedeNacional de RMNrdquo (REDE1517RMN2005) for access tothe NMR spectrometer and facilities at Coimbra Uni-versity The authors thank Centre of Investigation onEnvironment Genetics and Oncobiology (CIMAGO) forfinancial support Salome Pires and Ludgero Tavares arerecipients of fellowships from the Portuguese Foundationfor Science and Technology (SFRHBD753002010) and(SFRHBD666002009) respectively
References
[1] OWarburg ldquoOrigin of cancer cellsrdquoOncologia vol 9 no 2 pp75ndash83 1956 (German)
[2] S Weinhouse O Warburg D Burk and A L Schade ldquoOn res-piratory impairment in cancer cellsrdquo Science vol 124 no 3215pp 267ndash272 1956
10 BioMed Research International
[3] Y Zhao E B Butler andMTan ldquoTargeting cellularmetabolismto improve cancer therapeuticsrdquo Cell Death and Disease vol 4no 3 article e532 2013
[4] C V Dang ldquoLinks between metabolism and cancerrdquo Genes ampDevelopment vol 26 no 9 pp 877ndash890 2012
[5] M Talekar S R Boreddy A Singh andMAmiji ldquoTumor aero-bic glycolysis new insights into therapeutic strategies with targ-eted deliveryrdquo Expert Opinion on Biological Therapy 2014
[6] R Xu H Pelicano Y Zhou et al ldquoInhibition of glycolysis incancer cells a novel strategy to overcome drug resistance asso-ciatedwithmitochondrial respiratory defect and hypoxiardquoCan-cer Research vol 65 no 2 pp 613ndash621 2005
[7] S Ros and A Schulze ldquoBalancing glycolytic flux the roleof 6-phosphofructo-2-kinasefructose 26-bisphosphatases incancer metabolismrdquoCancer ampMetabolism vol 1 article 8 2013
[8] E Roudier and A Perrin ldquoConsidering the role of pyruvate intumor cells during hypoxiardquo Biochimica et Biophysica Acta vol1796 no 2 pp 55ndash62 2009
[9] K J Williams B A Telfer R E Airley et al ldquoA protective rolefor HIF-1 in response to redox manipulation and glucose depr-ivation implications for tumorigenesisrdquo Oncogene vol 21 no2 pp 282ndash290 2002
[10] M Achison and T R Hupp ldquoHypoxia attenuates the p53 resp-onse to cellular damagerdquo Oncogene vol 22 no 22 pp 3431ndash3440 2003
[11] H L Kotze E G Armitage K J Sharkey J W AllwoodW B Dunn and K J Williams ldquoA novel untargeted metabol-omics correlation-based network analysis incorporating humanmetabolic reconstructionsrdquo BMC Systems Biology vol 7 article107 2013
[12] Y-A Wen P D Stevens M L Gasser R Andrei and T GaoldquoDownregulation of PHLPP expression contributes to hypoxia-induced resistance to chemotherapy in colon cancer cellsrdquoMolecular and Cellular Biology vol 33 no 22 pp 4594ndash46052013
[13] B Garcia-Barreno T Delgado B Akerlind-Stopner E Norrbyand J A Melero ldquoLocation of the epitope recognized by mono-clonal antibody 63G on the primary structure of human resp-iratory syncytial virus G glycoprotein and the ability of syn-thetic peptides containing this epitope to induce neutralizingantibodiesrdquo Journal of General Virology vol 73 no 10 pp 2625ndash2630 1992
[14] I F Duarte J Marques A F Ladeirinha et al ldquoAnalyticalapproaches toward successful human cell metabolome studiesby NMR spectroscopyrdquo Analytical Chemistry vol 81 no 12 pp5023ndash5032 2009
[15] S L Pereira J Ramalho-Santos A F Branco V A Sardao P JOliveira and R A Carvalho ldquoMetabolic remodeling duringH9c2 myoblast differentiation relevance for in vitro toxicitystudiesrdquo Cardiovascular Toxicology vol 11 no 2 pp 180ndash1902011
[16] MD Basson IMModlin and J AMadri ldquoHuman enterocyte(Caco-2)migration ismodulated in vitro by extracellularmatrixcomposition and epidermal growth factorrdquo Journal of ClinicalInvestigation vol 90 no 1 pp 15ndash23 1992
[17] M D Peterson and M S Mooseker ldquoCharacterization of theenterocyte-like brush border cytoskeleton of the C2BBe clonesof the human intestinal cell line Caco-2rdquo Journal of Cell Sciencevol 102 no 3 pp 581ndash600 1992
[18] E M Chance S H Seeholzer K Kobayashi and J R Will-iamson ldquoMathematical analysis of isotope labeling in the citric
acid cycle with applications to 13C NMR studies in perfusedrat heartsrdquo Journal of Biological Chemistry vol 258 no 22 pp13785ndash13794 1983
[19] D HWilliamson P Lund and H A Krebs ldquoThe redox state offree nicotinamide-adenine dinucleotide in the cytoplasm andmitochondria of rat liverrdquo Biochemical Journal vol 103 no 2pp 514ndash527 1967
[20] L Suardet A C Gaide J M Calmes et al ldquoResponsiveness ofthree newly established human colorectal cancer cell lines totransforming growth factors 1205731 and 12057321rdquo Cancer Research vol52 no 13 pp 3705ndash3712 1992
[21] R A Carvalho T B Rodrigues P Zhao F M H Jeffrey C RMalloy and A D Sherry ldquoA 13C isotopomer kinetic analysisof cardiac metabolism influence of altered cytosolic redoxand [Ca2+]
119900rdquo The American Journal of PhysiologymdashHeart and
Circulatory Physiology vol 287 no 2 pp H889ndashH895 2004[22] AM Abrantes M E S Serra A C Goncalves et al ldquoHypoxia-
induced redox alterations and their correlation with 99mTc-MIBI and 99mTc-HL-91 uptake in colon cancer cellsrdquo NuclearMedicine and Biology vol 37 no 2 pp 125ndash132 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 BioMed Research International
[3] Y Zhao E B Butler andMTan ldquoTargeting cellularmetabolismto improve cancer therapeuticsrdquo Cell Death and Disease vol 4no 3 article e532 2013
[4] C V Dang ldquoLinks between metabolism and cancerrdquo Genes ampDevelopment vol 26 no 9 pp 877ndash890 2012
[5] M Talekar S R Boreddy A Singh andMAmiji ldquoTumor aero-bic glycolysis new insights into therapeutic strategies with targ-eted deliveryrdquo Expert Opinion on Biological Therapy 2014
[6] R Xu H Pelicano Y Zhou et al ldquoInhibition of glycolysis incancer cells a novel strategy to overcome drug resistance asso-ciatedwithmitochondrial respiratory defect and hypoxiardquoCan-cer Research vol 65 no 2 pp 613ndash621 2005
[7] S Ros and A Schulze ldquoBalancing glycolytic flux the roleof 6-phosphofructo-2-kinasefructose 26-bisphosphatases incancer metabolismrdquoCancer ampMetabolism vol 1 article 8 2013
[8] E Roudier and A Perrin ldquoConsidering the role of pyruvate intumor cells during hypoxiardquo Biochimica et Biophysica Acta vol1796 no 2 pp 55ndash62 2009
[9] K J Williams B A Telfer R E Airley et al ldquoA protective rolefor HIF-1 in response to redox manipulation and glucose depr-ivation implications for tumorigenesisrdquo Oncogene vol 21 no2 pp 282ndash290 2002
[10] M Achison and T R Hupp ldquoHypoxia attenuates the p53 resp-onse to cellular damagerdquo Oncogene vol 22 no 22 pp 3431ndash3440 2003
[11] H L Kotze E G Armitage K J Sharkey J W AllwoodW B Dunn and K J Williams ldquoA novel untargeted metabol-omics correlation-based network analysis incorporating humanmetabolic reconstructionsrdquo BMC Systems Biology vol 7 article107 2013
[12] Y-A Wen P D Stevens M L Gasser R Andrei and T GaoldquoDownregulation of PHLPP expression contributes to hypoxia-induced resistance to chemotherapy in colon cancer cellsrdquoMolecular and Cellular Biology vol 33 no 22 pp 4594ndash46052013
[13] B Garcia-Barreno T Delgado B Akerlind-Stopner E Norrbyand J A Melero ldquoLocation of the epitope recognized by mono-clonal antibody 63G on the primary structure of human resp-iratory syncytial virus G glycoprotein and the ability of syn-thetic peptides containing this epitope to induce neutralizingantibodiesrdquo Journal of General Virology vol 73 no 10 pp 2625ndash2630 1992
[14] I F Duarte J Marques A F Ladeirinha et al ldquoAnalyticalapproaches toward successful human cell metabolome studiesby NMR spectroscopyrdquo Analytical Chemistry vol 81 no 12 pp5023ndash5032 2009
[15] S L Pereira J Ramalho-Santos A F Branco V A Sardao P JOliveira and R A Carvalho ldquoMetabolic remodeling duringH9c2 myoblast differentiation relevance for in vitro toxicitystudiesrdquo Cardiovascular Toxicology vol 11 no 2 pp 180ndash1902011
[16] MD Basson IMModlin and J AMadri ldquoHuman enterocyte(Caco-2)migration ismodulated in vitro by extracellularmatrixcomposition and epidermal growth factorrdquo Journal of ClinicalInvestigation vol 90 no 1 pp 15ndash23 1992
[17] M D Peterson and M S Mooseker ldquoCharacterization of theenterocyte-like brush border cytoskeleton of the C2BBe clonesof the human intestinal cell line Caco-2rdquo Journal of Cell Sciencevol 102 no 3 pp 581ndash600 1992
[18] E M Chance S H Seeholzer K Kobayashi and J R Will-iamson ldquoMathematical analysis of isotope labeling in the citric
acid cycle with applications to 13C NMR studies in perfusedrat heartsrdquo Journal of Biological Chemistry vol 258 no 22 pp13785ndash13794 1983
[19] D HWilliamson P Lund and H A Krebs ldquoThe redox state offree nicotinamide-adenine dinucleotide in the cytoplasm andmitochondria of rat liverrdquo Biochemical Journal vol 103 no 2pp 514ndash527 1967
[20] L Suardet A C Gaide J M Calmes et al ldquoResponsiveness ofthree newly established human colorectal cancer cell lines totransforming growth factors 1205731 and 12057321rdquo Cancer Research vol52 no 13 pp 3705ndash3712 1992
[21] R A Carvalho T B Rodrigues P Zhao F M H Jeffrey C RMalloy and A D Sherry ldquoA 13C isotopomer kinetic analysisof cardiac metabolism influence of altered cytosolic redoxand [Ca2+]
119900rdquo The American Journal of PhysiologymdashHeart and
Circulatory Physiology vol 287 no 2 pp H889ndashH895 2004[22] AM Abrantes M E S Serra A C Goncalves et al ldquoHypoxia-
induced redox alterations and their correlation with 99mTc-MIBI and 99mTc-HL-91 uptake in colon cancer cellsrdquo NuclearMedicine and Biology vol 37 no 2 pp 125ndash132 2010
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom


















