
Comparative Transcriptome Analysis of Climacteric Fruit of ...
Upload
trannguyetCategory
view
217download
0
Research ArticleComparative Transcriptome Analysis Reveals Effects ofExogenous Hematin on Anthocyanin Biosynthesis duringStrawberry Fruit Ripening
Yi Li12 Huayin Li1 Fengde Wang1 Jingjuan Li1 Yihui Zhang1
Liangju Wang2 and Jianwei Gao1
1 Institute of Vegetables and Flowers Shandong Academy of Agricultural Sciences and Shandong Key Laboratory ofGreenhouse Vegetable Biology and Shandong Branch of National Vegetable Improvement Center Jinan 250100 China2College of Horticulture Nanjing Agricultural University Nanjing 210095 China
Correspondence should be addressed to Liangju Wang wangliangju0909sohucom and Jianwei Gao jianweigao3qqcom
Received 15 July 2016 Revised 30 September 2016 Accepted 25 October 2016
Academic Editor Quanhu Sheng
Copyright copy 2016 Yi Li et al This is an open access article distributed under the Creative Commons Attribution License whichpermits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Anthocyanin in strawberries has a positive effect on fruit coloration In this study the role of exogenous hematin on anthocyaninbiosynthesis was investigated Our result showed that the white stage of strawberries treated with exogenous hematin had higheranthocyanin content compared to the control group Among all treatments 5120583Mof hematin was the optimal condition to promotecolor development In order to explore the molecular mechanism of fruit coloring regulated by hematin transcriptomes in thehematin- and non-hematin-treated fruit were analyzed A large number of differentially expressed genes (DEGs) were identifiedin regulating anthocyanin synthesis including the DEGs involved in anthocyanin biosynthesis hormone signaling transductionphytochrome signaling starch and sucrose degradation and transcriptional pathways These regulatory networks may play animportant role in regulating the color process of strawberries treated with hematin In summary exogenous hematin could promotefruit coloring by increasing anthocyanin content in the white stage of strawberries Furthermore transcriptome analysis suggeststhat hematin-promoted fruit coloring occurs through multiple related metabolic pathways which provides valuable informationfor regulating fruit color via anthocyanin biosynthesis in strawberries
1 Introduction
Strawberry (Fragaria times ananassa Duch) is one of the mostpopular fruits with global economic importance [1] Becauseof its appealing red coloration and abundant nutritionstrawberries are highly sought after by consumers [2 3]These qualities are partially due to the high anthocyanincontent in strawberry Anthocyanins have a high antioxidantactivity [4] Research suggests that anthocyanins have poten-tial health benefits for a variety of conditions including car-diovascular disorders advanced age-induced oxidative stressinflammatory response [5] and diverse degenerative diseases[6 7] Increasing anthocyanin content in strawberries hasbeen a relevant research topic in recent years
Biosynthesis of anthocyanins is a complex biologicalprocess which is affected by genetic developmental and
environmental factors [8] Over the past few years moststructural genes encoding enzymes in the anthocyaninbiosynthetic pathways have been isolated and character-ized in strawberries The first group of structural genesinvolved in these pathways includes phenylalanine ammo-nia lyase (PAL) chalcone synthase (CHS) flavanone 3-hydroxylase (F3H) dihydroflavonol-4-reductase (DFR) leu-coanthocyanidin reductase (LAR)anthocyanidin synthase(ANS) and UDP-glucose flavonoid 3-O-glucosyltransferase(UFGT) These structural genes comprise the pathwaysneeded for the synthesis of anthocyanins [9] These genesare regulated primarily by the MybbHLHWD40 (MBW)complex in many plants [10 11]
It has been shown that anthocyanin synthesis increasesrapidly after the white stage in strawberries [12] Phytohor-mones such as abscisic acid (ABA) cytokinin (CTK) and
Hindawi Publishing CorporationInternational Journal of GenomicsVolume 2016 Article ID 6762731 14 pageshttpdxdoiorg10115520166762731
2 International Journal of Genomics
ethylene and methyl jasmonate (JA) also play an importantrole in regulating the color development process in strawber-ries by increasing anthocyanin accumulation [12 13] Auxinand gibberellins (GAs) are known to reduce anthocyaninbiosynthesis during the color development in fruit [8]
Light is one of the most important environmental fac-tors regulating anthocyanin biosynthesis [14] Phytochromeswhich act as photoreceptors play an important role in lightstimulation during the development of strawberries [15]Phytochromes are homodimeric chromoproteins where eachholophytochrome is composed of a phytochrome proteincovalently bound to a linear tetrapyrrole chromophore phy-tochromobilin (PΦB) PΦB acts as a light-receiving antennafor phytochrome PΦB is synthesized in the plastid fromheme catalyzed by heme oxygenase (HO) and subsequentlyphytochromobilin synthase [16] Heme oxygenase 1 (HO1)is crucial to this process and acts as a rate-limiting enzymein the biosynthesis of PΦB HO1 catalyzes the oxygenationof heme to carbon monoxide Fe2+ and biliverdin (BV)in plants [17] Additionally HO1 has been shown to playan important role in anthocyanin accumulation in plantsFor example tomato [18 19] and Arabidopsis [20ndash22] HO1deficientmutants which can not synthesize the phytochromechromophore have a major reduction in anthocyanin accu-mulation
Hematin (C34H33O5N4Fe) a protoporphyrin complex
is an inducer and substrate of HO1 in animals and plants[17 23] Exogenous hematin was shown to alleviate mercury-induced oxidative damage in the roots of Medicago sativa[17] induce adventitious root numbers and root length ofcucumbers [24] regulate Brassica nigra seed germinationunder nanosilver stress and relieve etiolation in the leavesof wheat seedlings under complete darkness [25 26] Theseeffects might be derived from a hematin induced HO1enzymatic reaction product
In this study the effect of exogenous hematin on thewhite stage of strawberries was investigated by comparing theanthocyanin contents in the hematin-treated fruit and thecontrol group In order to gain insight into the underlyingmolecular mechanisms regulating fruit coloring in responseto the hematin treatment an analysis of mRNA expressionprofiles was performed using high-throughput sequencingThe results demonstrated that exogenous hematin couldpromote fruit coloring Comparative transcriptome analysesmay give us a better understanding of the mechanism of thecoloring process in strawberry fruit
2 Materials and Methods
21 Plant Materials Growth Condition and Hematin Treat-ments Strawberry (Fragaria times ananassa Duch cv Beni-hoppe) an octoploid (2119899 = 8119883 = 56) species was plantedunder standard culture conditions in a greenhouse (3015∘C1410 h daynight relative humidity 50ndash80) The maximumlight intensity inside the greenhouse was 55300 lux Thedevelopmental stage of the strawberry fruit was dividedinto seven visual stages small green (SG) big green (BG)degreening (DG) white (Wt) initial red (IR) partial red(PR) and full red (FR) [12] Strawberry plants at the white
stage (about 25 d after anthesis) were chosen to study theeffect of hematin on fruit coloration in the study To studythe effect of different concentration of hematin on fruitcoloration white stage strawberry plants (119899 ge 50 for eachtreatment) were sprayed with 0 1 10 or 100 120583M hematin(H3281 Sigma-Aldrich St Louis MO USA) respectively[26] Strawberry fruits (119899 ge 30 for each treatment) wereharvestedwhen the fruit entered the PR stage (48 h after treat-ment) The fruits were immediately frozen in liquid nitrogenand stored at minus80∘C for further analysis We found the 10 120583Mhematin-treated strawberries accumulatedmost anthocyaninamong all treatments (Figure S1 in Supplementary Materialavailable online at httpdxdoiorg10115520166762731) Byusing the same method to treat the fruits with 0 5 10 or 15-120583M hematin we found the treatment by 5 120583M hematin wasthe optimal condition for increasing anthocyanin productionin strawberry (Figure 1) (119901 lt 001) The treatment with 5120583Mhematinwas used for subsequent analysisThree independentreplicates were prepared for each treatment
22 Determination of the Anthocyanin Content The totalanthocyanin content of the strawberries was determinedusing the method previously reported [6] Absorbance wasrecorded on a BeckmanDU640B spectrophotometer (Fuller-ton CA USA) at 510 and 700 nm for anthocyanin solutionsin a pH 10 and pH 45 buffer respectively The calculatedabsorbance was obtained according to
119860 = [(119860510minus 119860700)pH 10 minus (119860510 minus 119860700)pH 45] (1)
The molar extinction coefficient is 26900 as describedin other studies (eg [6]) Anthocyanin concentrations wereexpressed in milligrams of cyanidin-3-galactoside equivalentper gram of fresh weight Three independent replicates wereconducted for each treatment
23 BV Preparation and Assay Strawberry fruit (1 g) washomogenized in a Potter-Elvehjemhomogenizer using 12mLice-cold 025M sucrose solution containing 1mM phenyl-methyl sulfonyl fluoride 02mM EDTA and 50mM potas-sium phosphate buffer (pH 74) Homogenates were cen-trifuged at 20000timesg for 20min and chloroplasts were usedfor activity determination BV was assayed as previouslydescribed [27] The concentration of BV was estimated usinga molar absorption coefficient at 650 nm of 625mMminus1 cmminus1in 01M HEPES-NaOH buffer (pH 72) Three independentreplicates were conducted for each treatment
24 RNA Isolation and cDNA Library Construction TotalRNA isolation was carried out as previously described [28]The procedure is briefly presented below Strawberry fruitswere ground into powder and mixed at a ratio of 05 gpowder to 20mL extraction buffer (200mM Tris-HCl (pH82) 100mM LiCl 50mM ETA 15 SDS 2 PVP (SigmaPCP40) 2 BSA (Sigma) and 10mM DTT (Sigma)) Atotal of 200120583L 10mgmL proteinase K (Merck DarmstadtGermany) was added to remove contaminating proteinsTotal RNA was extracted using phenolchloroformisoamylalcohol (25 24 1) and precipitated in a sodium acetate
International Journal of Genomics 3
CK Treated with 5120583M hematin
(a)
0
50
100
150
200
250
ControlHematin treatment
C
B
B
A
15120583M10120583M5120583MA
ntho
cyan
in co
nten
t (m
gmiddotgminus
1middotF
W)
(b)
Figure 1 Fruit coloration and the anthocyanin content in the strawberry fruit The color of the 5 120583M hematin-treated strawberry fruit wasredder than the control (CK) at 48 h after treatment (a) The anthocyanin content in the strawberry fruit was measured after treatment withdifferent concentrations of hematin (0 5 10 and 15120583M) (b) Vertical bars represent standard errors values with different letter are significantlydifferent at 119901 lt 001
and ethanol mixture The mixture was resuspended in anappropriate volume of DEPC-treated distilled water andthen stored at minus80∘C for the next step The quality andquantity of the total RNA were measured using a NanoDropND-1000 Spectrophotometer (NanoDrop Wilmington DEUSA) Only samples that met the criteria of 18 leOD260280le D20 and OD260230 ge 18 and concentration ge 200 ng120583Lwere used for sequencing RNA samples of 30 strawberryfruits harvested in the same treatment group were pooledtogether for subsequent experiments
RNA samples from two biological replicates were usedfor cDNA library construction and RNA-Seq at the BeijingGenomics Institute (BGI Shenzhen China) Total RNAsamples were treated with DNase I (TaKaRa Dalian China)to remove any possible DNA contaminationThemRNA wasenriched by using oligo (dT) magnetic beads (Illumina SanDiegoCAUSA) and cut into short fragments (about 200 bp)The first-strand cDNA was synthesized using a SuperscriptPreamplification System Kit (Gibco-BRL Grand Island NYUSA) as described in the manufacturerrsquos instructions Thedouble stranded cDNAwas purified with the oligo (dT)mag-netic beads following the manufacturerrsquos instructions Endrepair was performed and adaptors were ligated to the endsof these fragments Ligation products were purified usingTAE-agarose gel electrophoresis The fragments were thenenriched by PCR amplificationwith an initial denaturing stepat 98∘C for 30 s followed by 15 cycles of amplification (98∘Cfor 10 s 65∘C for 30 s and 72∘C for 5min) and a final extensionat 72∘C for 5min The PCR products were then purified
using the oligo (dT) magnetic beads DNA size purity andconcentration were checked on an Agilent 2100 Bioanalyzer(Agilent Santa Clara CA USA)
25 RNA Sequencing and Identification of DifferentiallyExpressed Genes RNA sequencing was performed using theion proton platform at the Beijing Genomics Institute Theoriginal sequence data were filtered to obtain clean reads forfurther analyses by removing short reads (less than 30 bp) andtrimming adapters Adapter reads were trimmed by first cal-culating the average quality of the first 15 bases from 31015840-enduntil the average quality was larger than 10 and then removingthe bases that were counted The high-quality clean readswere mapped against the strawberry reference genome(httpstrawberry-gardenkazusaorjp) using Ion Torrentrsquosmapping program (TMAP version 023 httpsgithubcomiontorrentTMAP) No more than two mismatches wereallowed in the sequence alignment Quality assessment ofreads statistics of alignment sequencing saturation analysisand randomness assessments were carried out subsequentlyto assess the quality of sequencing
Gene expression levels were quantified by the softwareSailfish [29] Raw counts were normalized to Reads PerKilobase of exon model per Million mapped reads (RPKM)Differential expression analysis was performed using theEBSeq package [30]119876 valuelt 005 and |log
2(fold change)| gt
1were set as the threshold to identify significant differentiallyexpressed genes In statistics correction for false positiveerrors was carried out using the FDR statistic [31]
4 International Journal of Genomics
26 Gene Ontology and Pathway Enrichment Analysis ofDifferentially Expressed Genes Gene functions of strawberrywere annotated according to the Gene Ontology (GO) stand-ardized terms for molecular function cellular componentand biological process using Blast2GO (httpswwwblast2gocom) Gene function was annotated using the followingdatabases NCBI nonredundant protein sequences NCBInonredundant nucleotide sequences protein family Clustersof Orthologous Groups of proteins the manually annotatedand reviewed protein sequence database KEGG Orthologdatabase and Gene Ontology After GO annotation forDEGs we performed GO functional classification for DEGsby the WEGO software [32] and analyzed the distributionof gene functions We used the GO Term Finder tool(httpwwwyeastgenomeorghelpanalyzego-term-finder)to search for significant shared GO terms
119901 = 1 minus119898minus1
sum119894=0
(119872119894) (119873minus119872119899minus119894)
(119873119899) (2)
where 119873 is the number of all genes with GO annotation119899 is the number of DEGs in 119873 119872 is the number of allgenes that are annotated to certain GO terms and 119898 is thenumber of DEGs in119872 The calculated 119901 value was subjectedto Bonferroni Correction [33] The corrected 119901 value lt 005was set as the threshold GO terms fulfilling this conditionare defined as significantly enriched GO terms in DEGsThe pathway enrichment analyses using the KEGG database(httpwwwgenomejpkegg) were conducted subsequentlyto study the functions of differentially expressed genes identi-fied between the control and the hematin-treated strawberrygroups The formula is the same as that in GO analysis Here119873 is the number of all genes with KEGG annotation 119899 is thenumber ofDEGs in119873119872 is the number of all genes annotatedto specific pathways and119898 is the number of DEGs in119872
27 Quantitative Real-Time PCR Analysis Quantitative real-time PCR (qRT-PCR) experiments were conducted to assessthe reliability of the RNA-Seq data [34] Eleven pairs of genespecific primers were designed to verify the DEGs in thestrawberry fruit (Table S1) Strawberry RNA was extractedfrom the fruit according to the previously detailed methodTotal RNA was digested with DNase I for 30min at 25∘C toremove DNA contamination according to the manufacturerrsquosinstructions The qRT-PCR was performed using a SYBRPCR master mix (TaKaRa Dalian China) on a Bio-Rad IQ-5 thermal cycler (Bio-Rad Philadelphia PA USA) Threereplicates of each sample were conducted to calculate theaverage Ct valuesThe relative expression level was calculatedby the comparative 2minusΔΔCt method [35 36] The significancewas determined with the SPSS software (SPSS 170 IBMChicago IL USA) (119901 lt 005)
3 Results and Discussion
31 Effects of Exogenous Hematin on Anthocyanin andBiliverdin Accumulation Hematin is an inducer and sub-strate of HO1 in animals and plants [17 23] In this studystrawberries at theWt stage were treated with 0 120583M(control)
00
05
10
15
20
25
ControlHematin treatment
B
A
5120583M hematin
Biliv
erdi
n co
nten
t (120583
Mmiddotg
minus1middotF
W)
Figure 2 The BV content in strawberry fruit was measured aftertreatment with 0 and 5 120583Mhematin Vertical bars represent standarderrors values with different letter are significantly different at 119901 lt001
5 10 or 15 120583M hematin The results indicated that the antho-cyanin content in the hematin-treated strawberries was morethan the control after 48 h The treatment by 5120583M hematinwas found to be the optimal condition for increasing antho-cyanin production (Figure 1) (119901 lt 001) The anthocyanincontent in the 5 120583M hematin-treated strawberries was 25times higher than that in the control This is the first reportthat hematin could increase the anthocyanin production infruit Wemeasured the expression of FaHO-1 and the contentof biliverdin (BV) which is themetabolite of heme oxygenaseWe found that hematin could also significantly increase theexpression of FaHO-1 in strawberry fruit (Figure 4) andpromote the accumulation of BV (Figure 2) These resultsindicate that hematin promotes anthocyanin accumulationthrough the heme metabolism pathway
32 Sequences Assembly Mapping and Functional Anno-tation High-throughput sequencing technology is utilizedwidely in analyzing gene expression in many organisms[37 38] In this study approximately 108 Gbp raw tagswere generated for each library After eliminating adaptersambiguous nucleotides and low-quality sequences a total of34618832 and 45464123 clean reads between 150 and 200nucleotides in length were obtained (Table 1) Over 85 ofthe clean tags from each library mapped to reference geneswith less than 2 bp mismatches Less than 29 of the cleantags from each library could not be aligned to any referencegenes because of incomplete sequences and these tags weredesignated as unknown More than 673 of the clean tagsin each library were mapped to a single gene while less than47mapped to multiple reference genes Unknown tags andtags mapped to multigenes were filtered out and the unique
International Journal of Genomics 5
Table 1 Overview of the sequencing and assembly
Biological replicate 1 Biological replicate 2Sample Control Hematin-treated Control Hematin-treatedClean reads 17808924 26809908 19235452 26228671Genome map rate () 9658 9652 9665 9674Gene map rate () 8646 8627 8704 8563Perfect match () 4677273 (2432) 6510668 (2428) 4414050 (2479) 6149508 (2345)Unique match () 15075148 (8465) 22628939 (8441) 16409557 (8531) 21986962 (8383)Expressed genes 27976 28713 28999 28211Mismatch () 11321162 (6357) 17101590 (6379) 12456871 (6476) 16697280 (6366)Total unmapped reads 645135 (335) 932213 (348) 608914 (342) 856028 (326)
clean tags that mapped to a single gene were retained forfurther DEG analysis
In addition the perfect match rates were 2432 24282479 and 2435 in control 1 control 2 hematin 1 andhematin 2 respectively
The saturation of the libraries with and without thehematin treatment was analyzedThe number of the detectedgenes became saturated at about 2 million reads (FigureS2) This indicates that the obtained reads are sufficient forcomplete transcriptome coverage The randomness of RNAfragmentation in the four libraries was assessed for subse-quent bioinformatics analysis The results showed that thereads in each position of the reference gene were distributedevenly and demonstrated highly similar tendencies in alllibraries (Figure S3)
33 Genes Differentially Regulated in Response to ExogenousHematin Based on sequencing results of the four mRNAlibraries approximately 29000 genes (about 70 of thereference genes in the octoploid strawberry genome) weredetected in each library Additionally correlations amonggenes based on RPKM between the two biological replicateswere analyzed the correlation coefficients (1198772) were high(093 and 095 for the control and hematin-treated groupresp) Genes expressed in both replicates were screened Atotal of 28713 28999 27976 and 28211 genes (Table 1) wereexpressed in the control and hematin-treated samples
To reveal the molecular pathways regulating strawberrycoloring in response to hematin the DEGs between the con-trol and hematin-treated fruit were analyzed We comparedthe gene expression profiles between the control and thetreated samples in both biological replicates DEGs detectedin both biological replicates were screened for subsequentanalysis A total of 1080 (402 up- and 678 downregulated)genes were differentially expressed in the 5120583M hematin-treated groups comparedwith the control groupsMore geneswere downregulated in the hematin-treated group comparedto the control group
To facilitate the global analysis of gene expression aGO analysis was performed by mapping each differentiallyexpressed gene into the records of the GO database GeneOntology (GO) enrichment analysis of the DEGs in thecontrol and hematin-treated groups was performed to revealthe possible mechanisms under which hematin promotes
anthocyanin biosynthesis GO terms with 119901 lt 005 wererepresented among genes with significant changes in expres-sion over a given interval In this study only three termsincluding anatomical structure arrangementmeristem struc-tural organization radial pattern formationwere significantlyenriched in biological process (Supplementary Table S2)AlthoughmostGo termswere not significantly enriched theycould provide a reference for further study According to theGO cellular components a large number of the DEGs wereclassified intomain cell organelles such as vacuole nucleolusmitochondrion apoplast chloroplast plastid cytoplasm andmembrane In particular many DEGs were assigned to theplastid and chloroplast (Figure 3) indicating that hematinhas an important impact on the expression of plastid andchloroplast genes It is probably because heme oxygenase 1 isa soluble plastid protein [22] According to the GO molec-ular function a large number of the DEGs were classifiedintoUDP-glucosyltransferase sucrose transmembrane trans-porter activity nucleic acid binding and DNA binding termswhich are closely related to anthocyanin biosynthesis ManyDEGs were classified into heme binding peroxidase activityand tetrapyrrole binding (Figure 3) It is probably becausehematin can increase the activity of HO1 and promote thedegradation of hemoglobin [17] In addition anthocyaninaccumulation in fruit is closely related to hormone [8] andcarbohydrate biosynthesis [9] According to the GO biologi-cal process a large number of the DEGs were classified intoregulation of hormone levels cytokinin metabolic processresponse to hormone response to abscisic acid response toauxin and response to ethylene terms indicating that thehormone-related DEGs are involved in regulating the antho-cyanin biosynthesis in the hematin-treated fruit Many DEGswere classified into starch and sucrose biosynthetic and trans-port process and phenylpropanoid metabolic process termsindicating that hematin can promote anthocyanin accumu-lation via the phenylpropanoid metabolic and carbohydratemetabolism In addition many DEGs were classified into theprocess for response to light stimulus term indicating thathematin may also participate in the light stimulus system
Pathway enrichment analysis was performed to under-stand the biological functions of the DEGs by identify-ing significantly enriched signal transduction pathways ormetabolic pathways [39] In the study a number of alteredbiological pathways associated with the hematin treatment
6 International Journal of Genomics
0 10 20 30 40 50 60 70 80Starch biosynthetic process
Cytokinin metabolic processResponse to abscisic acid
Response to auxinResponse to ethylene
Phenylpropanoid metabolic processSucrose transport
Regulation of hormone levelsResponse to light stimulus
Response to hormonePeroxidase activity
Sucrose transmembrane transporter activityUDP-glucosyltransferase
Heme bindingTetrapyrrole binding
DNA bindingNucleic acid binding
VacuoleNucleolusApoplast
MitochondrionChloroplast
PlastidCytoplasmMembrane
Biol
ogic
al p
rogr
ess
Mol
ecul
ar fu
nctio
nC
ellu
lar c
ompo
nent
Number of DEGs
Figure 3 Gene classification based on Gene Ontology (GO) for DEGs in the strawberries treated by hematin
were identified and three pathways including RNA poly-merase pyrimidine metabolism and purine metabolismwere significantly enriched (119876-value lt 005) (Table 2)Although the majority of the pathway terms were not sig-nificantly enriched the pathway enrichment analysis helpsus to further understand the biological functions of theDEGs and the molecular mechanisms that regulate fruitcoloring in response to the hematin treatmentThese pathwayterms included metabolic pathways biosynthesis of sec-ondary metabolites isoflavonoid biosynthesis phenylalaninemetabolism phenylalanine tyrosine and tryptophan biosyn-thesis starch and sucrose metabolism plant hormone signaltransduction flavone and flavonol biosynthesis and phenyl-propanoid biosynthesis terms (Supplementary Table S2)
34 Analysis of DEGs Involved in Anthocyanin Biosynthesis
341 Transcription Factors Transcription factors (TFs) areproteins that regulate the expression of downstream targetgenes Many TFs such as v-myb avian myeloblastosis viraloncogene homolog (MYB) basic helix-loop-helix (bHLH)WD40-repeats protein (WD40) and MADS-box (MADS)directly regulating the expression of the structural genes inanthocyanin biosynthesis have been identified from manyspecies [8] Therefore the enhanced expression of key TFs
might regulate the anthocyanin biosynthesis In our studysome anthocyanin biosynthesis-related TFs were found to beuniquely present in the DEG profiling of the hematin-treated fruit for example MYB bHLH PIF3 MADs-boxAP2-EREBP and ABI3VP1 (Table 3) The anthocyanin bio-synthesis genes are regulated primarily by the MBW tran-scription factor which is a ternary transcription factor com-plex [10 11] Most of the MYBs involved in regulating antho-cyanin biosynthesis are positive regulators of transcription[8] However MYBs can act as repressors too such as straw-berry FaMYB1 and FaMYB9 and grapevine VvMYB4 whichcan significantly suppress the biosynthesis of anthocyaninsand flavonols [40] In this study three unigenes belong-ing to MYB transcription factors were differentiallyexpressed and two MYB transcription factors were signifi-cantly upregulated Of these the unigene ldquoFANhybrscf000001461g000071rdquo from theMYB family of R2R3MYBtranscription factors was significantly upregulatedMoreoverincreasing evidence indicates that the expression of thebHLHs promotes anthocyanin accumulation in fruit [41] Inour study all of the unigenes from the bHLH family weredownregulated among which the expression level of unigeneFANhyb rscf000012921g000031 was significantly decreased6250-fold (Table 3) FANhyb rscf000022101g000011 en-codes a predicted MADS-box transcription factor and it was
International Journal of Genomics 7
Control Hematin-treated
FANhyb_icon00002410_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_rscf000049691g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00027696_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00004395_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_rscf000063641g000011
lowastlowast
lowast
Control Hematin-treated
FANhyb_rscf000001411g000161
Control Hematin-treated
FANhyb_icon00006295_a1g000011lowastlowast
Control Hematin-treated
FANhyb_rscf000031551g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00001444_a1g000011
lowastlowast
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
0
05
1
15
2
25
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
005
115
225
335
445
5
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
Control Hematin-treated
FANhyb_rscf000023111g000011
lowastlowast
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
Control Hematin-treated
FANhyb_rscf000002951g000031
lowastlowast
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
Figure 4 qRT-PCR validation of the RNA-Seq based gene expression The values indicate means of three biological replicates plusmn SD Starindicates that the expression level is significantly different between the hematin-treated and the control group (lowast119901 lt 005 lowastlowast119901 lt 001)
8 International Journal of Genomics
Table 2 Pathway enrichment analysis of differentially expressed genes
Pathway DEGs with pathwayannotation (451)
All genes with pathwayannotation (15877)
Corrected 119901 value 119876-value Pathway ID
RNA polymerase 16 183 688119864 minus 5 00068107 ko03020Pyrimidine metabolism 20 317 00007651746 00320 ko00240Purine metabolism 21 347 00009710371 0032044224 ko00230
significantly downregulated There were three unigenes inthe AP2-EREBP family with only one upregulated and theothers downregulated These results suggest that exogenoushematin is involved in developmental transcriptionalregulation of anthocyanin biosynthesis
342 DEGs in Anthocyanin Biosynthesis The anthocyaninbiosynthetic pathway has been studied intensively Theanthocyanin biosynthetic pathway via the phenylpropanoidpathway is well known [8] In this study the transcriptomedata showed a significant increase in the expression of genesin the anthocyanin biosynthesis pathway when exposedto exogenous hematin The structural genes involved inanthocyanin biosynthesis including DFR LAR and UDP-glycosyltransferase exhibited significant differential expres-sion in response to the hematin treatment (Table 4) Forexample two DFR genes and one LAR gene were signifi-cantly upregulated (the expression increased 18- 30- and3-fold in the hematin-treated fruit resp) In addition allsix genes encodingUDP-glycosyltransferasewere significantlyupregulated Among these the expression of one gene(Fanhy icon15742070o1g000011) was upregulated 18305-fold The results suggest that exogenous hematin may havean important role in promoting anthocyanin biosynthesis
343 DEGs in Hormone Signaling Transduction in Responseto Hematin Auxin has been shown to negatively regulatethe expression of the anthocyanin biosynthesis genes [42] Inthis study there were three unigenes involved in the auxinsignaling pathway and all were downregulated includingone gene encoding auxin efflux carrier component 1-like(AUX1-like) and two genes encoding auxin response factors(ARF) These results suggest that hematin promotes antho-cyanin accumulation by regulating auxin in the colorationprocess of strawberries It has shown that auxin suppressesanthocyanin biosynthesis in the red-fleshed apple callus [43]Endogenous expression of auxin [44 45] has been foundto impede anthocyanin accumulation in strawberries Inadditional cytokinins are also known to play an importantrole in anthocyanin biosynthesis It has shown to enhanceanthocyanin accumulation in Zea mays and regulate antho-cyanin production and composition in suspension culturesof strawberry cells [46] In this study two unigenes relatedto the cytokinin signaling pathway were identified and theywere downregulated as well One gene was identified ascytokinin dehydrogenase 5-like which catalyzes the irre-versible degradation of cytokinins This result indicates thathematin promotes anthocyanin accumulation by regulatingthe cytokinin signaling pathway
Extensive researches have shown that ABA plays animportant role in the regulation of anthocyanin biosynthesisin nonclimacteric fruit [47] In this study a total of five uni-genes related to the ABA signaling pathway were identifiedamong which two were upregulated and three were down-regulated Two genes which are involved in the abscisic acidresponse were upregulated These two unigenes encode 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) and ABA overlysensitive 5 (ABO5) respectively ABA is synthesized fromcarotenoids via several enzymatic reactions in the plastidThe rate-limiting step in these reactions is catalyzed by 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) [48] Mutations inthe FaNCED1 gene result in colorless strawberry fruit whichcan become colored through the application of exogenousABA [12] Another unigene was identified to encode abscisicacid 81015840-hydroxylase 1-like a key enzyme in the oxidativecatabolism of abscisic acid which was downregulated Appli-cation of exogenous ABA regulates phenylalanine ammonialyase activity and increases the phenolic and anthocyanincontent of strawberry fruit [12 49] Our results suggestthat hematin promotes ABA biosynthesis and inhibits ABAdisintegration in the hematin-treated fruit
In Arabidopsis JAs can affect anthocyanin accumulationvia the interaction of negative regulators with the MBWcomplex of transcription factors involved in anthocyaninbiosynthesis [50] Preharvest application of JA to ldquoFujirdquo applesenhances red coloration [51] JA vapor treatment can alsoenhance anthocyanins in strawberry fruit [52] In this studyjasmonate O-methyltransferase-like exhibited significantlydifferential expression Jasmonate O-methyltransferase-likeis also known to be a key enzyme for jasmonate-regulatedplant responses to stimuli [53 54] These results suggest thathematin increases the activity of the JA-regulated antho-cyanin biosynthesis
344 DEGs in the Phytochrome Signaling Pathway in Responseto Hematin Phytochromes that act as photoreceptors playan important role in anthocyanin regulation [15] Phy-tochromes are homodimeric chromoproteins where eachholophytochrome is composed of a phytochrome protein(apophytochrome) covalently bound to a linear tetrapyrrolePΦB HO1 is crucial to this process and acts as a rate-limiting enzyme in the biosynthesis of PΦB [55] Exogenoushematin can also induce HO-1 expression in many plants[56 57] In this study we found exogenous hematin signif-icantly increased the expression of FaHO1 (Table 5) Thisresult implies that hematin can promote PΦB biosynthesisRecently it has been reported that hematin could inducethe accumulation of far-red phytochrome and phytochrome
International Journal of Genomics 9
Table 3 DEGs acting as transcription factors in response to exogenous hematin
Annotation transcription factors Gene IDReal
fold-changevalues
Upregulationdownregulation FDR
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb icon00006295 a1g000011 938 Up 120119864 minus 05
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb rscf000031551g000011 4 Down 480119864 minus 16
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL5-like
FANhyb rscf000000291g000101 4 Down 320119864 minus 08
MYBdomain protein 66 FANhyb rscf000001461g000071 654 Up 780119864 minus 07MYB-related protein 3R-1-like partial FANhyb rscf000001411g000161 282 Up 830119864 minus 04PREDICTED transcription factorGAMYB-like FANhyb rscf000006491g000061 303 Down 150119864 minus 13
PREDICTED probable WRKYtranscription factor 53-like FANhyb rscf000019731g000021 217 Down 200119864 minus 08
Transcription factor bHLH80-like FANhyb rscf000012921g000031 6250 Down 150119864 minus 06Transcription factor bHLH70-like FANhyb rscf000002951g000031 526 Down 660119864 minus 12Transcription factor ICE1-like FANhyb rscf000001701g000061 244 Down 450119864 minus 18C2H2-like zinc finger protein FANhyb rscf000011431g000021 282 Up 590119864 minus 04Zinc finger protein MAGPIE-like FANhyb rscf000030241g000011 204 Up 150119864 minus 04PREDICTED axial regulator YABBY5-like FANhyb rscf000016671g000011 417 Down 730119864 minus 05
PREDICTED axial regulator YABBY1-like FANhyb rscf000068991g000011 357 Down 620119864 minus 06
PREDICTED dof zinc finger proteinDOF37-like isoform 1 FANhyb rscf000068561g000011 263 Down 280119864 minus 04
ZF-HD homeobox protein FANhyb rscf000000441g000221 385 Down 420119864 minus 08ZF-HD homeobox proteinAt4g24660-like FANhyb rscf000052411g000011 357 Down 470119864 minus 09
PREDICTED B3 domain-containingprotein REM14-like FANhyb rscf000010091g000021 243 Up 530119864 minus 04
PREDICTED B3 domain-containingtranscription factor ABI3-like FANhyb rscf000012711g000021 455 Down 130119864 minus 28
Growth-regulating factor 6 FANhyb rscf000000241g000291 345 Down 330119864 minus 34Growth-regulating factor 5 FANhyb icon00009704 a1g000011 27 Down 190119864 minus 06GATA type zinc finger transcriptionfactor family protein FANhyb rscf000003931g000131 1111 Down 110119864 minus 05
PREDICTED zinc finger CCCHdomain-containing protein 2-like FANhyb icon00006770 a1g000011 313 Down 680119864 minus 49
PREDICTED E2F transcriptionfactor-like E2FE-like FANhyb icon00002702 a1g000011 278 Down 240119864 minus 04
PREDICTED uncharacterized protein FANhyb rscf000010661g000011 263 Down 490119864 minus 06PREDICTED nuclear transcriptionfactor Y subunit A-3-like isoform 1 FANhyb rscf000000081g000351 385 Down 500119864 minus 20
PREDICTED homeobox-leucinezipper protein ATHB-8-like FANhyb rscf000000151g000051 263 Down 100119864 minus 06
PREDICTED MADS-boxtranscription factor 18-like FANhyb rscf000003231g000161 243 Down 180119864 minus 06
PREDICTED zinc finger protein132-like isoform 1 FANhyb rscf000070151g000011 243 Down 180119864 minus 08
PREDICTED transcription factorPIF3-like FANhyb rscf000006691g000021 346 Up 140119864 minus 07
10 International Journal of Genomics
Table 4 Regulation of the DEGs in anthocyanin biosynthesis in response to exogenous hematin
Genes in anthocyaninbiosynthesis Gene ID
Realfold-change
valuesRegulation level FDR
Dihydroflavonol-4-reductase FANhyb icon00002410 a1g000011 3091 Up 910119864 minus 06
Flavonol synthase FANhyb icon00015354 a1g000011 345 down 180119864 minus 05Isoflavone reductasehomolog FANhyb icon20341135 s1g000011 334 Up 750119864 minus 14
UDP-glycosyltransferase FANhyb rscf000049691g000011 307 Up 640119864 minus 08UDP-glycosyltransferaseactivity FANhyb icon15742070 o1g000011 18 30563 Up 510119864 minus 09
UDP-glycosyltransferase73C1 FANhyb icon00027696 a1g000011 298 Up 560119864 minus 04
UDP-glycosyltransferaseactivity FANhyb icon00008064 a1g000011 249 Up 310119864 minus 04
PREDICTEDUDP-glucose flavonoid3-O-glucosyltransferase7-like
FANhyb rscf000055631g000011 212 Up 110119864 minus 19
PREDICTEDUDP-glycosyltransferase76F1-like
FANhyb rscf000021771g000031 2 Up 720119864 minus 29
A transcripts in the etiolated leaves of wheat seedling phy-tochromes [26] However our study shows that phytochromeA was not differentially expressed It is probably becausethe hematin-treated strawberries were not shaded during theexperiment while the function of phytochrome A is light-dependent In addition we found that a downstream gene ofphytochrome PIF3 (FANhyb rscf000006691g000021) wassignificantly upregulated PIF3 is thought to be a positive reg-ulator of phytochrome B mediated by light-dependent signaltransduction [58]These results suggest that the phytochromepathways may be involved in the anthocyanin biosynthesispromoted by hematin
345 DEGs in Starch and Sucrose Degradation Proteomicapproaches have revealed that starch degradation contributesto anthocyanin accumulation in tuberous roots of the purplesweet potato variety [59] Starch and sucrose biosynthesis isimportant for anthocyanin accumulation in strawberry fruit[60] In this study we found the DEGs involved in starchand sucrose synthesis in the hematin-treated fruit were alldownregulated (Table 5) For example the expression of theunigenes encoding soluble starch synthase 3 sucrose trans-port protein SUC2-like and a probable sucrose-phosphatesynthase 4-like enzyme were decreased 23- 46- 32- and2-fold respectively This suggests that exogenous hematinregulates the biosynthesis of starch and sucrose and henceaffects fruit coloring
346 DEGs in the Calcium Pathway in Response to HematinCalcium can also increase the transcription levels of keystructural genes F3H DFR ANT and UFGT in the whitestage of strawberries [61] In this study we found thatthree DEGs in calcium biosynthesis and transport were
downregulated in the hematin-treated fruit (Table 5) Onlythe unigene encoding predicted cationcalcium exchanger5-like was upregulated The unigenes encoding a calpain-type cysteine protease and a predicted calcium-binding pro-tein PBP1-like were downregulated respectively Exogenousapplication of calcium can promote apple coloring [62] inaddition to variation in anthocyanin content [61] Our resultsindicate that the biosynthesis and transport of calcium areinvolved in the development of coloring in hematin-treatedstrawberries
35 Validation of Selected DEGs by qRT-PCR To validate theexpression of the DEGs obtained from RNA-Seq 11 DEGswere selected for qRT-PCR including structural genes (DFRand UDFGs) transcription factor genes (MYB and bHLH)and phytochrome chromophore-related gene (HO-1) Theprimers used for qRT-PCR are listed in Supplementary TableS1 The qRT-PCR results were consistent with the RNA-Seqdata (Figure 4) except for FANhyb rscf000001411g000161which had a higher log
2ratio (hematin-treatedcontrol) in
the transcriptome data than in qRT-PCR These results indi-cate the RNA-Seq data from the strawberry transcriptome isreproducible and accurate
4 Conclusions
In this study the anthocyanin content in the strawberryfruit was elevated by the application of exogenous hematinThis is the first report that hematin could increase theanthocyanin production in fruit Furthermore we exploredthe effects of the exogenous hematin on metabolic path-ways using genome-wide transcriptome analysis The resultsindicate that the expression levels of many genes involved
International Journal of Genomics 11
Table 5 Other DEGs identified in anthocyanin biosynthesis-related pathways
Anthocyaninbiosynthesis-relatedpathways
Annotation genes Gene IDReal fold-changevalues
Upregulationdownregulation FDR
Calcium ion binding
Cationcalciumexchanger 5-like
FANhyb rscf000019061g000011 203 Up 64119864 minus 06
Calpain-type cysteineprotease family
FANhyb icon00011755 a1g000011 345 Down 18119864 minus 09
Calcium-bindingprotein PBP1-like
FANhyb rscf000007501g000081 323 Down 97119864 minus 04
Calpain-type cysteineprotease family
FANhyb icon00023658 a1g000011 233 Down 60119864 minus 08
CytokininCytokinin
dehydrogenase 5-likeFANhyb iscf00325393 1 s1g000011 667 Down 62119864 minus 06
Cytochrome P450714A1
FANhyb rscf000005921g000031 217 Down 98119864 minus 10
JasmonateJasmonate
O-methyltransferase-like
FANhyb rscf000000041g000131 201 Up 70119864 minus 04
Abscisic acid
Protein ABSCISICACID-INSENSITIVE
5-likeFANhyb rscf000021641g000011 4 Down 16119864 minus 18
ABA overly sensitive 5 FANhyb icon00051144 a1g000011 291 Up 24119864 minus 059-cis-
epoxycarotenoiddioxygenase NCED1
FANhyb icon18399909 o1g000011 441 Up 43119864 minus 06
Abscisic acid81015840-hydroxylase 1-like
FANhyb icon00000938 a1g000011 256 Down 14119864 minus 06
Abscisic acid receptorPYR1-like
FANhyb icon00020426 a1g000011 208 Down 30119864 minus 06
Auxin
Auxin efflux carriercomponent 1-like
FANhyb rscf000010081g000031 384 Down 23119864 minus 06
Auxin response factor8
FANhyb rscf000013061g000021 263 Down 61119864 minus 17
Auxin response factor17-like
FANhyb rscf000060741g000011 217 Down 11119864 minus 36
Starch and sucrose
Glycosyltransferasefamily 35
FANhyb rscf000000341g000081 384 Down 11119864 minus 05
Soluble starchsynthase 3
FANhyb rscf000000451g000041 232 Down 38119864 minus 12
Sucrose transportprotein SUC2-like
FANhyb icon00036727 a1g000011 455 Down 98119864 minus 04
Sucrose transportprotein SUC2-like
FANhyb rscf000007551g000021 323 Down 66119864 minus 10
Probablesucrose-phosphatesynthase 4-like
FANhyb rscf000013501g000021 213 Down 11119864 minus 14
Phytochrome
PREDICTEDtranscription factor
PIF3-likeFANhyb rscf000006691g000021 346 up 14119864 minus 07
Phytochrome E-like FANhyb rscf000004361g000041 4 Down 33119864 minus 05
Heme oxygenase 1 FANhyb icon00004395 a1g000011 258 27119864 minus 07 27119864 minus 07
12 International Journal of Genomics
Hematin
Endoplasmicreticulum
WD40 bLHLMYB
Structural genes
TFs
Nuclear
JA
ABA Sucrose
Plastid
G-cis-neoxanthin
Apophytochrome
StarchHeme
BVSucroseABA
Holophytochrome
PfrPIF3
Flavonoids Anthocyanins
Vacuole
Cytoplasm
Cell wall
①
②③
④⑤
⑥
⑦
PΦB
PΦB
CTKAuxin
Ca2+
Figure 5 Genes and biological pathways that were involved in regulation of anthocyanin accumulation promoted by hematin Outermostbox represents the cell wall Bilayer oval shape represents the plastidThe red-brown ring and network represent the nuclear and endoplasmicreticulum respectively The red crescent represents the vacuole The box in the nucleus represents transcription factors (TFs) Yellow MYBgreenWD40 and red bHLH represent a ternary transcription factor complex which transcribes anthocyanin biosynthesis genes The circlednumbers represent biological pathways involved in the regulation of anthocyanin accumulation promoted by hematin A represents thephytochrome regulation pathwayB represents the starch and sucrose pathwayCDE andF represent phytohormones ABA JA AuxinandCTK regulation pathways respectivelyG represents theCa2+ regulation pathway Pfr represents the far-red phytochrome PIF3 representsphytochrome-interacting factor Each anthocyanin biosynthesis regulatory pathway related gene is in Table 5
in anthocyanin biosynthesis were significantly altered withthe hematin treatment This suggests that the physiologicalprocess of fruit color development is regulated through com-plex interactions among anthocyanin biosynthesis pathwaysplant hormone signal transduction pathways phytochromesignal transduction pathways starch and sugar metabolicpathways calcium pathways and transcription factors (Fig-ure 5) This study adds to the in-depth understanding of thefruit coloration process in strawberry
Competing Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contributions
Jianwei Gao and Liangju Wang conceived and designed theresearch Yi Li Huayin Li Fengde Wang Jingjuan Li andYihui Zhang conducted experiments Yi Li analyzed the dataYi Li and Fengde Wang wrote the manuscript All authorsread and approved the manuscript Yi Li and Huayin Licontributed equally to this work
Acknowledgments
This work was supported by the Modern Agricultural Indus-trial Technology System Funding of Shandong ProvinceChina (SDAIT-02-022-04) the China agriculture researchsystem (CARS-25) and the National Science Foundation ofChina (31401820)
References
[1] K Aaby SMazur A Nes andG Skrede ldquoPhenolic compoundsin strawberry (Fragaria x ananassa Duch) fruits compositionin 27 cultivars and changes during ripeningrdquo Food Chemistryvol 132 no 1 pp 86ndash97 2012
[2] T A Colquhoun L A Levin H RMoskowitz VMWhitakerD G Clark and K M Folta ldquoFraming the perfect strawberryan exercise in consumer-assisted selection of fruit cropsrdquoJournal of Berry Research vol 2 no 1 pp 45ndash61 2012
[3] F Giampieri J M Alvarez-Suarez L Mazzoni et al ldquoThepotential impact of strawberry on human healthrdquo NaturalProduct Research vol 27 no 4 pp 448ndash455 2013
[4] Y S Velioglu GMazza L Gao and B D Oomah ldquoAntioxidantactivity and total phenolics in selected fruits vegetables and
International Journal of Genomics 13
grain productsrdquo Journal of Agricultural amp Food Chemistry vol46 no 10 pp 4113ndash4117 1998
[5] S S Hassellund A Flaa S E Kjeldsen et al ldquoEffects of antho-cyanins on cardiovascular risk factors and inflammation inpre-hypertensive men a double-blind randomized placebo-controlled crossover studyrdquo Journal of Human Hypertensionvol 27 no 2 pp 100ndash106 2013
[6] K J Meyers C B Watkins M P Pritts and R H Liu ldquoAnti-oxidant and antiproliferative activities of strawberriesrdquo Journalof Agricultural amp Food Chemistry vol 51 no 23 pp 6887ndash68922003
[7] S Zafra-Stone T YasminM Bagchi AChatterjee J AVinsonand D Bagchi ldquoBerry anthocyanins as novel antioxidants inhuman health and disease preventionrdquo Molecular Nutrition ampFood Research vol 51 no 6 pp 675ndash683 2007
[8] L Jaakola ldquoNew insights into the regulation of anthocyanin bio-synthesis in fruitsrdquoTrends in Plant Science vol 18 no 9 pp 477ndash483 2013
[9] K Springob J-I Nakajima M Yamazaki and K Saito ldquoRecentadvances in the biosynthesis and accumulation of antho-cyaninsrdquo Natural Product Reports vol 20 no 3 pp 288ndash3032003
[10] A Gonzalez M Zhao J M Leavitt and A M Lloyd ldquoRegu-lation of the anthocyanin biosynthetic pathway by the TTG1bHLHMyb transcriptional complex in Arabidopsis seedlingsrdquoPlant Journal vol 53 no 5 pp 814ndash827 2008
[11] K Petroni and C Tonelli ldquoRecent advances on the regulationof anthocyanin synthesis in reproductive organsrdquo Plant Sciencevol 181 no 3 pp 219ndash229 2011
[12] H-F Jia Y-M Chai C-L Li et al ldquoAbscisic acid plays animportant role in the regulation of strawberry fruit ripeningrdquoPlant Physiology vol 157 no 1 pp 188ndash199 2011
[13] C Merchante J G Vallarino S Osorio et al ldquoEthylene isinvolved in strawberry fruit ripening in an organ-specific man-nerrdquo Journal of Experimental Botany vol 64 no 14 pp 4421ndash4439 2013
[14] A Maier A Schrader L Kokkelink et al ldquoLight and the E3ubiquitin ligase COP1SPA control the protein stability of theMYB transcription factors PAP1 and PAP2 involved in antho-cyanin accumulation in Arabidopsisrdquo Plant Journal vol 74 no4 pp 638ndash651 2013
[15] H Lange W Shropshire and H Mohr ldquoAn analysis of phyto-chrome-mediated anthocyanin synthesisrdquo Plant Physiology vol47 no 5 pp 649ndash655 1971
[16] T D Elich and J C Lagarias ldquoPhytochrome chromophore bio-synthesis both 5-aminolevulinic acid and biliverdin overcomeinhibition by gabaculine in etiolated Avena sativa L seedlingsrdquoPlant Physiology vol 84 no 2 pp 304ndash310 1987
[17] Y Han W Xuan T Yu et al ldquoExogenous hematin alleviatesmercury-induced oxidative damage in the roots of Medicagosativardquo Journal of Integrative Plant Biology vol 49 no 12 pp1703ndash1713 2007
[18] M Koornneef J Cone R Dekens E OrsquoHerne-Robers CSpruit and R Kendrick ldquoPhotomorphogenic responses of longhypocotyl mutants of tomatordquo Journal of Plant Physiology vol120 no 2 pp 153ndash165 1985
[19] M J Terry and R E Kendrick ldquoFeedback inhibition of chloro-phyll synthesis in the phytochrome chromophore-deficientaurea and yellow-green-2 mutants of tomatordquo Plant Physiologyvol 119 no 1 pp 143ndash152 1999
[20] B M Parks and P H Quail ldquoPhytochrome-deficient hy1 andhy2 long hypocotyl mutants of Arabidopsis are defective inphytochrome chromophore biosynthesisrdquo Plant Cell vol 3 no11 pp 1177ndash1186 1991
[21] M J Terry ldquoPhytochrome chromophore-deficient mutantsrdquoPlant Cell amp Environment vol 20 no 6 pp 740ndash745 1997
[22] T Muramoto T Kohchi A Yokota I Hwang and H MGoodman ldquoThe Arabidopsis photomorphogenic mutant hy1 isdeficient in phytochrome chromophore biosynthesis as a resultof a mutation in a plastid heme oxygenaserdquo Plant Cell vol 11no 3 pp 335ndash347 1999
[23] M D Maines ldquoThe heme oxygenase system a regulator ofsecond messenger gasesrdquo Annual Review of Pharmacology andToxicology vol 37 no 1 pp 517ndash554 1997
[24] W Xuan F-Y Zhu S Xu et al ldquoThe heme oxygenasecarbonmonoxide system is involved in the auxin-induced cucumberadventitious rooting processrdquo Plant Physiology vol 148 no 2pp 881ndash893 2008
[25] R Amooaghaie F Tabatabaei and A-M Ahadi ldquoRole ofhematin and sodium nitroprusside in regulating Brassica nigraseed germination under nanosilver and silver nitrate stressesrdquoEcotoxicology and Environmental Safety vol 113 pp 259ndash2702015
[26] Y Liu X Li L Xu and W Shen ldquoDe-etiolation of wheatseedling leaves cross talk between heme oxygenasecarbonmonoxide and nitric oxiderdquo PLoS ONE vol 8 no 12 ArticleID e81470 2013
[27] T Muramoto N Tsurui M J Terry A Yokota and T KohchildquoExpression and biochemical properties of a ferredoxin-dependent heme oxygenase required for phytochrome chro-mophore synthesisrdquo Plant Physiology vol 130 no 4 pp 1958ndash1966 2002
[28] J-W Gao J Liu B Li and Z Li ldquoIsolation and purificationof functional total RNA from blue-grained wheat endospermtissues containing high levels of starches and flavonoidsrdquo PlantMolecular Biology Reporter vol 19 no 2 pp 185ndash186 2001
[29] R Patro S M Mount and C Kingsford ldquoSailfish enablesalignment-free isoform quantification from RNA-seq readsusing lightweight algorithmsrdquoNature Biotechnology vol 32 no5 pp 462ndash464 2014
[30] Y-X Qi Y-B Liu and W-H Rong ldquoRNA-Seq and its applica-tions a new technology for transcriptomicsrdquo Hereditas vol 33no 11 pp 1191ndash1202 2011
[31] C R Genovese N A Lazar and T Nichols ldquoThresholdingof statistical maps in functional neuroimaging using the falsediscovery raterdquo NeuroImage vol 15 no 4 pp 870ndash878 2002
[32] J Ye L Fang H Zheng et al ldquoWEGO a web tool for plottingGO annotationsrdquoNucleic Acids Research vol 34 supplement 2pp W293ndashW297 2006
[33] H Abdi ldquoThe Bonferonni and Sidak corrections for multiplecomparisonrdquo Encyclopedia of Measurement and Statistics vol 3pp 103ndash107 2007
[34] J Li Q Ding F Wang Y Zhang H Li and J Gao ldquoIntegrativeanalysis of mRNA and miRNA expression profiles of thetuberous root development at seedling stages in turnipsrdquo PLoSONE vol 10 no 9 Article ID e0137983 2015
[35] J Bennett D Hondred and J C Register ldquoKeeping qRT-PCRrigorous and biologically relevantrdquo Plant Cell Reports vol 34no 1 pp 1ndash3 2015
[36] F Wang L Li H Li et al ldquoTranscriptome analysis of rosetteand folding leaves in Chinese cabbage using high-throughputRNA sequencingrdquo Genomics vol 99 no 5 pp 299ndash307 2012
14 International Journal of Genomics
[37] T Shi Z Gao L Wang et al ldquoIdentification of differentially-expressed genes associated with pistil abortion in Japaneseapricot by genome-wide transcriptional analysisrdquo PLoS ONEvol 7 no 10 Article ID e47810 2012
[38] F Wang H Li Y Zhang et al ldquoMicroRNA expression analysisof rosette and folding leaves in Chinese cabbage using high-throughput Solexa sequencingrdquo Gene vol 532 no 2 pp 222ndash229 2013
[39] X Gu Y Chen Z Gao Y Qiao and X Wang ldquoTranscriptionfactors and anthocyanin genes related to low-temperature toler-ance in rd29A RdreB1BI transgenic strawberryrdquo Plant Physiol-ogy and Biochemistry vol 89 pp 31ndash43 2015
[40] J G Schaart C Dubos I R De La Fuente et al ldquoIdentificationand characterization of MYB-bHLH-WD40 regulatory com-plexes controlling proanthocyanidin biosynthesis in strawberry(Fragaria times ananassa) fruitsrdquoNew Phytologist vol 197 no 2 pp454ndash467 2013
[41] L Xie Z H Wang X H Cheng J J Gao Z P Zhangand L J Wang ldquo5-Aminolevulinic acid promotes anthocyaninaccumulation in Fuji applesrdquo Plant Growth Regulation vol 69no 3 pp 295ndash303 2013
[42] M Daminato F Guzzo and G Casadoro ldquoASHATTERPROOF-like gene controls ripening in non-climac-teric strawberries and auxin and abscisic acid antagonisticallyaffect its expressionrdquo Journal of Experimental Botany vol 64no 12 pp 3775ndash3786 2013
[43] X-H Ji R Zhang N Wang L Yang and X-S Chen ldquoTran-scriptome profiling reveals auxin suppressed anthocyanin bio-synthesis in red-fleshed apple callus (Malus sieversii f niedzwet-zkyana)rdquo Plant Cell Tissue and Organ Culture vol 123 no 2pp 389ndash404 2015
[44] J Chen LMaoW Lu T Ying and Z Luo ldquoTranscriptome pro-filing of postharvest strawberry fruit in response to exogenousauxin and abscisic acidrdquo Planta vol 243 no 1 pp 183ndash197 2016
[45] G A Martınez A R Chaves and M C Anon ldquoEffect of gib-berellic acid on ripening of strawberry fruits (Fragaria annan-assa Duch)rdquo Journal of Plant Growth Regulation vol 13 no 2pp 87ndash91 1994
[46] P K Das D H Shin S-B Choi S-D Yoo G Choi and Y-IPark ldquoCytokinins enhance sugar-induced anthocyanin biosyn-thesis in Arabidopsisrdquo Molecules amp Cells vol 34 no 1 pp 93ndash101 2012
[47] P McAtee S Karim R Schaffer and K David ldquoA dynamicinterplay between phytohormones is required for fruit develop-ment maturation and ripeningrdquo Frontiers in Plant Science vol4 no 3 article 79 7 pages 2013
[48] E Nambara andAMarion-Poll ldquoAbscisic acid biosynthesis andcatabolismrdquoAnnual Review of Plant Biology vol 56 pp 165ndash1852005
[49] Y Jiang and D C Joyce ldquoABA effects on ethylene productionPAL activity anthocyanin and phenolic contents of strawberryfruitrdquo Plant Growth Regulation vol 39 no 2 pp 171ndash174 2003
[50] S Li ldquoTranscriptional control of flavonoid biosynthesis fine-tuning of the MYB-bHLH-WD40 (MBW) complexrdquo PlantSignaling amp Behavior vol 9 Article ID e27522 2014
[51] D R Rudell J K Fellman and J P Mattheis ldquoPreharvest appli-cation of methyl jasmonate to rsquoFujirsquo apples enhances red col-oration and affects fruit size splitting and bitter pit incidencerdquoHortScience vol 40 no 6 pp 1760ndash1762 2005
[52] A Belhadj N Telef C Saigne et al ldquoEffect of methyl jasmonatein combination with carbohydrates on gene expression of PR
proteins stilbene and anthocyanin accumulation in grapevinecell culturesrdquo Plant Physiology and Biochemistry vol 46 no 4pp 493ndash499 2008
[53] E Loreti G Povero G Novi C Solfanelli A Alpi and PPerata ldquoGibberellins jasmonate and abscisic acid modulate thesucrose-induced expression of anthocyanin biosynthetic genesin Arabidopsisrdquo New Phytologist vol 179 no 4 pp 1004ndash10162008
[54] H S Seo J T Song J-J Cheong et al ldquoJasmonic acid carboxylmethyltransferase A key enzyme for jasmonate-regulated plantresponsesrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 98 no 8 pp 4788ndash4793 2001
[55] M T McDowell and J C Lagarias ldquoPurification and biochemi-cal properties of phytochromobilin synthase from etiolated oatseedlingsrdquo Plant Physiology vol 126 no 4 pp 1546ndash1554 2001
[56] G-Q Fu S Xu Y-J Xie et al ldquoMolecular cloning characteri-zation and expression of an alfalfa (Medicago sativa L) hemeoxygenase-1 gene MsHO1 which is pro-oxidants-regulatedrdquoPlant Physiology and Biochemistry vol 49 no 7 pp 792ndash7992011
[57] X-Y Chen X Ding S Xu et al ldquoEndogenous hydrogen per-oxide plays a positive role in the upregulation of heme oxy-genase and acclimation to oxidative stress in wheat seedlingleavesrdquo Journal of Integrative Plant Biology vol 51 no 10 pp951ndash960 2009
[58] J Shin E Park and G Choi ldquoPIF3 regulates anthocyaninbiosynthesis in an HY5-dependent manner with both factorsdirectly binding anthocyanin biosynthetic gene promoters inArabidopsisrdquo Plant Journal vol 49 no 6 pp 981ndash994 2007
[59] S Wang D Pan X Lv et al ldquoProteomic approach reveals thatstarch degradation contributes to anthocyanin accumulation intuberous root of purple sweet potatordquo Journal of Proteomics vol143 pp 298ndash305 2016
[60] H-F Jia C-L Li Y-M Chai Y Xing and Y Shen ldquoSucrosepromotes strawberry fruit ripening by stimulation of abscisicacid biosynthesisrdquo Pakistan Journal of Botany vol 45 no 1 pp169ndash176 2013
[61] W Xu H Peng T Yang et al ldquoEffect of calcium on strawberryfruit flavonoid pathway gene expression and anthocyanin accu-mulationrdquo Plant Physiology amp Biochemistry vol 82 pp 289ndash298 2014
[62] Z-W-SWan and Z Singh ldquoExogenous application of prohexa-dione-calcium promotes fruit colour development of lsquoCrippsPinkrsquo applerdquo Acta Horticulturae vol 1012 pp 219ndash225 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 International Journal of Genomics
ethylene and methyl jasmonate (JA) also play an importantrole in regulating the color development process in strawber-ries by increasing anthocyanin accumulation [12 13] Auxinand gibberellins (GAs) are known to reduce anthocyaninbiosynthesis during the color development in fruit [8]
Light is one of the most important environmental fac-tors regulating anthocyanin biosynthesis [14] Phytochromeswhich act as photoreceptors play an important role in lightstimulation during the development of strawberries [15]Phytochromes are homodimeric chromoproteins where eachholophytochrome is composed of a phytochrome proteincovalently bound to a linear tetrapyrrole chromophore phy-tochromobilin (PΦB) PΦB acts as a light-receiving antennafor phytochrome PΦB is synthesized in the plastid fromheme catalyzed by heme oxygenase (HO) and subsequentlyphytochromobilin synthase [16] Heme oxygenase 1 (HO1)is crucial to this process and acts as a rate-limiting enzymein the biosynthesis of PΦB HO1 catalyzes the oxygenationof heme to carbon monoxide Fe2+ and biliverdin (BV)in plants [17] Additionally HO1 has been shown to playan important role in anthocyanin accumulation in plantsFor example tomato [18 19] and Arabidopsis [20ndash22] HO1deficientmutants which can not synthesize the phytochromechromophore have a major reduction in anthocyanin accu-mulation
Hematin (C34H33O5N4Fe) a protoporphyrin complex
is an inducer and substrate of HO1 in animals and plants[17 23] Exogenous hematin was shown to alleviate mercury-induced oxidative damage in the roots of Medicago sativa[17] induce adventitious root numbers and root length ofcucumbers [24] regulate Brassica nigra seed germinationunder nanosilver stress and relieve etiolation in the leavesof wheat seedlings under complete darkness [25 26] Theseeffects might be derived from a hematin induced HO1enzymatic reaction product
In this study the effect of exogenous hematin on thewhite stage of strawberries was investigated by comparing theanthocyanin contents in the hematin-treated fruit and thecontrol group In order to gain insight into the underlyingmolecular mechanisms regulating fruit coloring in responseto the hematin treatment an analysis of mRNA expressionprofiles was performed using high-throughput sequencingThe results demonstrated that exogenous hematin couldpromote fruit coloring Comparative transcriptome analysesmay give us a better understanding of the mechanism of thecoloring process in strawberry fruit
2 Materials and Methods
21 Plant Materials Growth Condition and Hematin Treat-ments Strawberry (Fragaria times ananassa Duch cv Beni-hoppe) an octoploid (2119899 = 8119883 = 56) species was plantedunder standard culture conditions in a greenhouse (3015∘C1410 h daynight relative humidity 50ndash80) The maximumlight intensity inside the greenhouse was 55300 lux Thedevelopmental stage of the strawberry fruit was dividedinto seven visual stages small green (SG) big green (BG)degreening (DG) white (Wt) initial red (IR) partial red(PR) and full red (FR) [12] Strawberry plants at the white
stage (about 25 d after anthesis) were chosen to study theeffect of hematin on fruit coloration in the study To studythe effect of different concentration of hematin on fruitcoloration white stage strawberry plants (119899 ge 50 for eachtreatment) were sprayed with 0 1 10 or 100 120583M hematin(H3281 Sigma-Aldrich St Louis MO USA) respectively[26] Strawberry fruits (119899 ge 30 for each treatment) wereharvestedwhen the fruit entered the PR stage (48 h after treat-ment) The fruits were immediately frozen in liquid nitrogenand stored at minus80∘C for further analysis We found the 10 120583Mhematin-treated strawberries accumulatedmost anthocyaninamong all treatments (Figure S1 in Supplementary Materialavailable online at httpdxdoiorg10115520166762731) Byusing the same method to treat the fruits with 0 5 10 or 15-120583M hematin we found the treatment by 5 120583M hematin wasthe optimal condition for increasing anthocyanin productionin strawberry (Figure 1) (119901 lt 001) The treatment with 5120583Mhematinwas used for subsequent analysisThree independentreplicates were prepared for each treatment
22 Determination of the Anthocyanin Content The totalanthocyanin content of the strawberries was determinedusing the method previously reported [6] Absorbance wasrecorded on a BeckmanDU640B spectrophotometer (Fuller-ton CA USA) at 510 and 700 nm for anthocyanin solutionsin a pH 10 and pH 45 buffer respectively The calculatedabsorbance was obtained according to
119860 = [(119860510minus 119860700)pH 10 minus (119860510 minus 119860700)pH 45] (1)
The molar extinction coefficient is 26900 as describedin other studies (eg [6]) Anthocyanin concentrations wereexpressed in milligrams of cyanidin-3-galactoside equivalentper gram of fresh weight Three independent replicates wereconducted for each treatment
23 BV Preparation and Assay Strawberry fruit (1 g) washomogenized in a Potter-Elvehjemhomogenizer using 12mLice-cold 025M sucrose solution containing 1mM phenyl-methyl sulfonyl fluoride 02mM EDTA and 50mM potas-sium phosphate buffer (pH 74) Homogenates were cen-trifuged at 20000timesg for 20min and chloroplasts were usedfor activity determination BV was assayed as previouslydescribed [27] The concentration of BV was estimated usinga molar absorption coefficient at 650 nm of 625mMminus1 cmminus1in 01M HEPES-NaOH buffer (pH 72) Three independentreplicates were conducted for each treatment
24 RNA Isolation and cDNA Library Construction TotalRNA isolation was carried out as previously described [28]The procedure is briefly presented below Strawberry fruitswere ground into powder and mixed at a ratio of 05 gpowder to 20mL extraction buffer (200mM Tris-HCl (pH82) 100mM LiCl 50mM ETA 15 SDS 2 PVP (SigmaPCP40) 2 BSA (Sigma) and 10mM DTT (Sigma)) Atotal of 200120583L 10mgmL proteinase K (Merck DarmstadtGermany) was added to remove contaminating proteinsTotal RNA was extracted using phenolchloroformisoamylalcohol (25 24 1) and precipitated in a sodium acetate
International Journal of Genomics 3
CK Treated with 5120583M hematin
(a)
0
50
100
150
200
250
ControlHematin treatment
C
B
B
A
15120583M10120583M5120583MA
ntho
cyan
in co
nten
t (m
gmiddotgminus
1middotF
W)
(b)
Figure 1 Fruit coloration and the anthocyanin content in the strawberry fruit The color of the 5 120583M hematin-treated strawberry fruit wasredder than the control (CK) at 48 h after treatment (a) The anthocyanin content in the strawberry fruit was measured after treatment withdifferent concentrations of hematin (0 5 10 and 15120583M) (b) Vertical bars represent standard errors values with different letter are significantlydifferent at 119901 lt 001
and ethanol mixture The mixture was resuspended in anappropriate volume of DEPC-treated distilled water andthen stored at minus80∘C for the next step The quality andquantity of the total RNA were measured using a NanoDropND-1000 Spectrophotometer (NanoDrop Wilmington DEUSA) Only samples that met the criteria of 18 leOD260280le D20 and OD260230 ge 18 and concentration ge 200 ng120583Lwere used for sequencing RNA samples of 30 strawberryfruits harvested in the same treatment group were pooledtogether for subsequent experiments
RNA samples from two biological replicates were usedfor cDNA library construction and RNA-Seq at the BeijingGenomics Institute (BGI Shenzhen China) Total RNAsamples were treated with DNase I (TaKaRa Dalian China)to remove any possible DNA contaminationThemRNA wasenriched by using oligo (dT) magnetic beads (Illumina SanDiegoCAUSA) and cut into short fragments (about 200 bp)The first-strand cDNA was synthesized using a SuperscriptPreamplification System Kit (Gibco-BRL Grand Island NYUSA) as described in the manufacturerrsquos instructions Thedouble stranded cDNAwas purified with the oligo (dT)mag-netic beads following the manufacturerrsquos instructions Endrepair was performed and adaptors were ligated to the endsof these fragments Ligation products were purified usingTAE-agarose gel electrophoresis The fragments were thenenriched by PCR amplificationwith an initial denaturing stepat 98∘C for 30 s followed by 15 cycles of amplification (98∘Cfor 10 s 65∘C for 30 s and 72∘C for 5min) and a final extensionat 72∘C for 5min The PCR products were then purified
using the oligo (dT) magnetic beads DNA size purity andconcentration were checked on an Agilent 2100 Bioanalyzer(Agilent Santa Clara CA USA)
25 RNA Sequencing and Identification of DifferentiallyExpressed Genes RNA sequencing was performed using theion proton platform at the Beijing Genomics Institute Theoriginal sequence data were filtered to obtain clean reads forfurther analyses by removing short reads (less than 30 bp) andtrimming adapters Adapter reads were trimmed by first cal-culating the average quality of the first 15 bases from 31015840-enduntil the average quality was larger than 10 and then removingthe bases that were counted The high-quality clean readswere mapped against the strawberry reference genome(httpstrawberry-gardenkazusaorjp) using Ion Torrentrsquosmapping program (TMAP version 023 httpsgithubcomiontorrentTMAP) No more than two mismatches wereallowed in the sequence alignment Quality assessment ofreads statistics of alignment sequencing saturation analysisand randomness assessments were carried out subsequentlyto assess the quality of sequencing
Gene expression levels were quantified by the softwareSailfish [29] Raw counts were normalized to Reads PerKilobase of exon model per Million mapped reads (RPKM)Differential expression analysis was performed using theEBSeq package [30]119876 valuelt 005 and |log
2(fold change)| gt
1were set as the threshold to identify significant differentiallyexpressed genes In statistics correction for false positiveerrors was carried out using the FDR statistic [31]
4 International Journal of Genomics
26 Gene Ontology and Pathway Enrichment Analysis ofDifferentially Expressed Genes Gene functions of strawberrywere annotated according to the Gene Ontology (GO) stand-ardized terms for molecular function cellular componentand biological process using Blast2GO (httpswwwblast2gocom) Gene function was annotated using the followingdatabases NCBI nonredundant protein sequences NCBInonredundant nucleotide sequences protein family Clustersof Orthologous Groups of proteins the manually annotatedand reviewed protein sequence database KEGG Orthologdatabase and Gene Ontology After GO annotation forDEGs we performed GO functional classification for DEGsby the WEGO software [32] and analyzed the distributionof gene functions We used the GO Term Finder tool(httpwwwyeastgenomeorghelpanalyzego-term-finder)to search for significant shared GO terms
119901 = 1 minus119898minus1
sum119894=0
(119872119894) (119873minus119872119899minus119894)
(119873119899) (2)
where 119873 is the number of all genes with GO annotation119899 is the number of DEGs in 119873 119872 is the number of allgenes that are annotated to certain GO terms and 119898 is thenumber of DEGs in119872 The calculated 119901 value was subjectedto Bonferroni Correction [33] The corrected 119901 value lt 005was set as the threshold GO terms fulfilling this conditionare defined as significantly enriched GO terms in DEGsThe pathway enrichment analyses using the KEGG database(httpwwwgenomejpkegg) were conducted subsequentlyto study the functions of differentially expressed genes identi-fied between the control and the hematin-treated strawberrygroups The formula is the same as that in GO analysis Here119873 is the number of all genes with KEGG annotation 119899 is thenumber ofDEGs in119873119872 is the number of all genes annotatedto specific pathways and119898 is the number of DEGs in119872
27 Quantitative Real-Time PCR Analysis Quantitative real-time PCR (qRT-PCR) experiments were conducted to assessthe reliability of the RNA-Seq data [34] Eleven pairs of genespecific primers were designed to verify the DEGs in thestrawberry fruit (Table S1) Strawberry RNA was extractedfrom the fruit according to the previously detailed methodTotal RNA was digested with DNase I for 30min at 25∘C toremove DNA contamination according to the manufacturerrsquosinstructions The qRT-PCR was performed using a SYBRPCR master mix (TaKaRa Dalian China) on a Bio-Rad IQ-5 thermal cycler (Bio-Rad Philadelphia PA USA) Threereplicates of each sample were conducted to calculate theaverage Ct valuesThe relative expression level was calculatedby the comparative 2minusΔΔCt method [35 36] The significancewas determined with the SPSS software (SPSS 170 IBMChicago IL USA) (119901 lt 005)
3 Results and Discussion
31 Effects of Exogenous Hematin on Anthocyanin andBiliverdin Accumulation Hematin is an inducer and sub-strate of HO1 in animals and plants [17 23] In this studystrawberries at theWt stage were treated with 0 120583M(control)
00
05
10
15
20
25
ControlHematin treatment
B
A
5120583M hematin
Biliv
erdi
n co
nten
t (120583
Mmiddotg
minus1middotF
W)
Figure 2 The BV content in strawberry fruit was measured aftertreatment with 0 and 5 120583Mhematin Vertical bars represent standarderrors values with different letter are significantly different at 119901 lt001
5 10 or 15 120583M hematin The results indicated that the antho-cyanin content in the hematin-treated strawberries was morethan the control after 48 h The treatment by 5120583M hematinwas found to be the optimal condition for increasing antho-cyanin production (Figure 1) (119901 lt 001) The anthocyanincontent in the 5 120583M hematin-treated strawberries was 25times higher than that in the control This is the first reportthat hematin could increase the anthocyanin production infruit Wemeasured the expression of FaHO-1 and the contentof biliverdin (BV) which is themetabolite of heme oxygenaseWe found that hematin could also significantly increase theexpression of FaHO-1 in strawberry fruit (Figure 4) andpromote the accumulation of BV (Figure 2) These resultsindicate that hematin promotes anthocyanin accumulationthrough the heme metabolism pathway
32 Sequences Assembly Mapping and Functional Anno-tation High-throughput sequencing technology is utilizedwidely in analyzing gene expression in many organisms[37 38] In this study approximately 108 Gbp raw tagswere generated for each library After eliminating adaptersambiguous nucleotides and low-quality sequences a total of34618832 and 45464123 clean reads between 150 and 200nucleotides in length were obtained (Table 1) Over 85 ofthe clean tags from each library mapped to reference geneswith less than 2 bp mismatches Less than 29 of the cleantags from each library could not be aligned to any referencegenes because of incomplete sequences and these tags weredesignated as unknown More than 673 of the clean tagsin each library were mapped to a single gene while less than47mapped to multiple reference genes Unknown tags andtags mapped to multigenes were filtered out and the unique
International Journal of Genomics 5
Table 1 Overview of the sequencing and assembly
Biological replicate 1 Biological replicate 2Sample Control Hematin-treated Control Hematin-treatedClean reads 17808924 26809908 19235452 26228671Genome map rate () 9658 9652 9665 9674Gene map rate () 8646 8627 8704 8563Perfect match () 4677273 (2432) 6510668 (2428) 4414050 (2479) 6149508 (2345)Unique match () 15075148 (8465) 22628939 (8441) 16409557 (8531) 21986962 (8383)Expressed genes 27976 28713 28999 28211Mismatch () 11321162 (6357) 17101590 (6379) 12456871 (6476) 16697280 (6366)Total unmapped reads 645135 (335) 932213 (348) 608914 (342) 856028 (326)
clean tags that mapped to a single gene were retained forfurther DEG analysis
In addition the perfect match rates were 2432 24282479 and 2435 in control 1 control 2 hematin 1 andhematin 2 respectively
The saturation of the libraries with and without thehematin treatment was analyzedThe number of the detectedgenes became saturated at about 2 million reads (FigureS2) This indicates that the obtained reads are sufficient forcomplete transcriptome coverage The randomness of RNAfragmentation in the four libraries was assessed for subse-quent bioinformatics analysis The results showed that thereads in each position of the reference gene were distributedevenly and demonstrated highly similar tendencies in alllibraries (Figure S3)
33 Genes Differentially Regulated in Response to ExogenousHematin Based on sequencing results of the four mRNAlibraries approximately 29000 genes (about 70 of thereference genes in the octoploid strawberry genome) weredetected in each library Additionally correlations amonggenes based on RPKM between the two biological replicateswere analyzed the correlation coefficients (1198772) were high(093 and 095 for the control and hematin-treated groupresp) Genes expressed in both replicates were screened Atotal of 28713 28999 27976 and 28211 genes (Table 1) wereexpressed in the control and hematin-treated samples
To reveal the molecular pathways regulating strawberrycoloring in response to hematin the DEGs between the con-trol and hematin-treated fruit were analyzed We comparedthe gene expression profiles between the control and thetreated samples in both biological replicates DEGs detectedin both biological replicates were screened for subsequentanalysis A total of 1080 (402 up- and 678 downregulated)genes were differentially expressed in the 5120583M hematin-treated groups comparedwith the control groupsMore geneswere downregulated in the hematin-treated group comparedto the control group
To facilitate the global analysis of gene expression aGO analysis was performed by mapping each differentiallyexpressed gene into the records of the GO database GeneOntology (GO) enrichment analysis of the DEGs in thecontrol and hematin-treated groups was performed to revealthe possible mechanisms under which hematin promotes
anthocyanin biosynthesis GO terms with 119901 lt 005 wererepresented among genes with significant changes in expres-sion over a given interval In this study only three termsincluding anatomical structure arrangementmeristem struc-tural organization radial pattern formationwere significantlyenriched in biological process (Supplementary Table S2)AlthoughmostGo termswere not significantly enriched theycould provide a reference for further study According to theGO cellular components a large number of the DEGs wereclassified intomain cell organelles such as vacuole nucleolusmitochondrion apoplast chloroplast plastid cytoplasm andmembrane In particular many DEGs were assigned to theplastid and chloroplast (Figure 3) indicating that hematinhas an important impact on the expression of plastid andchloroplast genes It is probably because heme oxygenase 1 isa soluble plastid protein [22] According to the GO molec-ular function a large number of the DEGs were classifiedintoUDP-glucosyltransferase sucrose transmembrane trans-porter activity nucleic acid binding and DNA binding termswhich are closely related to anthocyanin biosynthesis ManyDEGs were classified into heme binding peroxidase activityand tetrapyrrole binding (Figure 3) It is probably becausehematin can increase the activity of HO1 and promote thedegradation of hemoglobin [17] In addition anthocyaninaccumulation in fruit is closely related to hormone [8] andcarbohydrate biosynthesis [9] According to the GO biologi-cal process a large number of the DEGs were classified intoregulation of hormone levels cytokinin metabolic processresponse to hormone response to abscisic acid response toauxin and response to ethylene terms indicating that thehormone-related DEGs are involved in regulating the antho-cyanin biosynthesis in the hematin-treated fruit Many DEGswere classified into starch and sucrose biosynthetic and trans-port process and phenylpropanoid metabolic process termsindicating that hematin can promote anthocyanin accumu-lation via the phenylpropanoid metabolic and carbohydratemetabolism In addition many DEGs were classified into theprocess for response to light stimulus term indicating thathematin may also participate in the light stimulus system
Pathway enrichment analysis was performed to under-stand the biological functions of the DEGs by identify-ing significantly enriched signal transduction pathways ormetabolic pathways [39] In the study a number of alteredbiological pathways associated with the hematin treatment
6 International Journal of Genomics
0 10 20 30 40 50 60 70 80Starch biosynthetic process
Cytokinin metabolic processResponse to abscisic acid
Response to auxinResponse to ethylene
Phenylpropanoid metabolic processSucrose transport
Regulation of hormone levelsResponse to light stimulus
Response to hormonePeroxidase activity
Sucrose transmembrane transporter activityUDP-glucosyltransferase
Heme bindingTetrapyrrole binding
DNA bindingNucleic acid binding
VacuoleNucleolusApoplast
MitochondrionChloroplast
PlastidCytoplasmMembrane
Biol
ogic
al p
rogr
ess
Mol
ecul
ar fu
nctio
nC
ellu
lar c
ompo
nent
Number of DEGs
Figure 3 Gene classification based on Gene Ontology (GO) for DEGs in the strawberries treated by hematin
were identified and three pathways including RNA poly-merase pyrimidine metabolism and purine metabolismwere significantly enriched (119876-value lt 005) (Table 2)Although the majority of the pathway terms were not sig-nificantly enriched the pathway enrichment analysis helpsus to further understand the biological functions of theDEGs and the molecular mechanisms that regulate fruitcoloring in response to the hematin treatmentThese pathwayterms included metabolic pathways biosynthesis of sec-ondary metabolites isoflavonoid biosynthesis phenylalaninemetabolism phenylalanine tyrosine and tryptophan biosyn-thesis starch and sucrose metabolism plant hormone signaltransduction flavone and flavonol biosynthesis and phenyl-propanoid biosynthesis terms (Supplementary Table S2)
34 Analysis of DEGs Involved in Anthocyanin Biosynthesis
341 Transcription Factors Transcription factors (TFs) areproteins that regulate the expression of downstream targetgenes Many TFs such as v-myb avian myeloblastosis viraloncogene homolog (MYB) basic helix-loop-helix (bHLH)WD40-repeats protein (WD40) and MADS-box (MADS)directly regulating the expression of the structural genes inanthocyanin biosynthesis have been identified from manyspecies [8] Therefore the enhanced expression of key TFs
might regulate the anthocyanin biosynthesis In our studysome anthocyanin biosynthesis-related TFs were found to beuniquely present in the DEG profiling of the hematin-treated fruit for example MYB bHLH PIF3 MADs-boxAP2-EREBP and ABI3VP1 (Table 3) The anthocyanin bio-synthesis genes are regulated primarily by the MBW tran-scription factor which is a ternary transcription factor com-plex [10 11] Most of the MYBs involved in regulating antho-cyanin biosynthesis are positive regulators of transcription[8] However MYBs can act as repressors too such as straw-berry FaMYB1 and FaMYB9 and grapevine VvMYB4 whichcan significantly suppress the biosynthesis of anthocyaninsand flavonols [40] In this study three unigenes belong-ing to MYB transcription factors were differentiallyexpressed and two MYB transcription factors were signifi-cantly upregulated Of these the unigene ldquoFANhybrscf000001461g000071rdquo from theMYB family of R2R3MYBtranscription factors was significantly upregulatedMoreoverincreasing evidence indicates that the expression of thebHLHs promotes anthocyanin accumulation in fruit [41] Inour study all of the unigenes from the bHLH family weredownregulated among which the expression level of unigeneFANhyb rscf000012921g000031 was significantly decreased6250-fold (Table 3) FANhyb rscf000022101g000011 en-codes a predicted MADS-box transcription factor and it was
International Journal of Genomics 7
Control Hematin-treated
FANhyb_icon00002410_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_rscf000049691g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00027696_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00004395_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_rscf000063641g000011
lowastlowast
lowast
Control Hematin-treated
FANhyb_rscf000001411g000161
Control Hematin-treated
FANhyb_icon00006295_a1g000011lowastlowast
Control Hematin-treated
FANhyb_rscf000031551g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00001444_a1g000011
lowastlowast
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
0
05
1
15
2
25
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
005
115
225
335
445
5
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
Control Hematin-treated
FANhyb_rscf000023111g000011
lowastlowast
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
Control Hematin-treated
FANhyb_rscf000002951g000031
lowastlowast
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
Figure 4 qRT-PCR validation of the RNA-Seq based gene expression The values indicate means of three biological replicates plusmn SD Starindicates that the expression level is significantly different between the hematin-treated and the control group (lowast119901 lt 005 lowastlowast119901 lt 001)
8 International Journal of Genomics
Table 2 Pathway enrichment analysis of differentially expressed genes
Pathway DEGs with pathwayannotation (451)
All genes with pathwayannotation (15877)
Corrected 119901 value 119876-value Pathway ID
RNA polymerase 16 183 688119864 minus 5 00068107 ko03020Pyrimidine metabolism 20 317 00007651746 00320 ko00240Purine metabolism 21 347 00009710371 0032044224 ko00230
significantly downregulated There were three unigenes inthe AP2-EREBP family with only one upregulated and theothers downregulated These results suggest that exogenoushematin is involved in developmental transcriptionalregulation of anthocyanin biosynthesis
342 DEGs in Anthocyanin Biosynthesis The anthocyaninbiosynthetic pathway has been studied intensively Theanthocyanin biosynthetic pathway via the phenylpropanoidpathway is well known [8] In this study the transcriptomedata showed a significant increase in the expression of genesin the anthocyanin biosynthesis pathway when exposedto exogenous hematin The structural genes involved inanthocyanin biosynthesis including DFR LAR and UDP-glycosyltransferase exhibited significant differential expres-sion in response to the hematin treatment (Table 4) Forexample two DFR genes and one LAR gene were signifi-cantly upregulated (the expression increased 18- 30- and3-fold in the hematin-treated fruit resp) In addition allsix genes encodingUDP-glycosyltransferasewere significantlyupregulated Among these the expression of one gene(Fanhy icon15742070o1g000011) was upregulated 18305-fold The results suggest that exogenous hematin may havean important role in promoting anthocyanin biosynthesis
343 DEGs in Hormone Signaling Transduction in Responseto Hematin Auxin has been shown to negatively regulatethe expression of the anthocyanin biosynthesis genes [42] Inthis study there were three unigenes involved in the auxinsignaling pathway and all were downregulated includingone gene encoding auxin efflux carrier component 1-like(AUX1-like) and two genes encoding auxin response factors(ARF) These results suggest that hematin promotes antho-cyanin accumulation by regulating auxin in the colorationprocess of strawberries It has shown that auxin suppressesanthocyanin biosynthesis in the red-fleshed apple callus [43]Endogenous expression of auxin [44 45] has been foundto impede anthocyanin accumulation in strawberries Inadditional cytokinins are also known to play an importantrole in anthocyanin biosynthesis It has shown to enhanceanthocyanin accumulation in Zea mays and regulate antho-cyanin production and composition in suspension culturesof strawberry cells [46] In this study two unigenes relatedto the cytokinin signaling pathway were identified and theywere downregulated as well One gene was identified ascytokinin dehydrogenase 5-like which catalyzes the irre-versible degradation of cytokinins This result indicates thathematin promotes anthocyanin accumulation by regulatingthe cytokinin signaling pathway
Extensive researches have shown that ABA plays animportant role in the regulation of anthocyanin biosynthesisin nonclimacteric fruit [47] In this study a total of five uni-genes related to the ABA signaling pathway were identifiedamong which two were upregulated and three were down-regulated Two genes which are involved in the abscisic acidresponse were upregulated These two unigenes encode 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) and ABA overlysensitive 5 (ABO5) respectively ABA is synthesized fromcarotenoids via several enzymatic reactions in the plastidThe rate-limiting step in these reactions is catalyzed by 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) [48] Mutations inthe FaNCED1 gene result in colorless strawberry fruit whichcan become colored through the application of exogenousABA [12] Another unigene was identified to encode abscisicacid 81015840-hydroxylase 1-like a key enzyme in the oxidativecatabolism of abscisic acid which was downregulated Appli-cation of exogenous ABA regulates phenylalanine ammonialyase activity and increases the phenolic and anthocyanincontent of strawberry fruit [12 49] Our results suggestthat hematin promotes ABA biosynthesis and inhibits ABAdisintegration in the hematin-treated fruit
In Arabidopsis JAs can affect anthocyanin accumulationvia the interaction of negative regulators with the MBWcomplex of transcription factors involved in anthocyaninbiosynthesis [50] Preharvest application of JA to ldquoFujirdquo applesenhances red coloration [51] JA vapor treatment can alsoenhance anthocyanins in strawberry fruit [52] In this studyjasmonate O-methyltransferase-like exhibited significantlydifferential expression Jasmonate O-methyltransferase-likeis also known to be a key enzyme for jasmonate-regulatedplant responses to stimuli [53 54] These results suggest thathematin increases the activity of the JA-regulated antho-cyanin biosynthesis
344 DEGs in the Phytochrome Signaling Pathway in Responseto Hematin Phytochromes that act as photoreceptors playan important role in anthocyanin regulation [15] Phy-tochromes are homodimeric chromoproteins where eachholophytochrome is composed of a phytochrome protein(apophytochrome) covalently bound to a linear tetrapyrrolePΦB HO1 is crucial to this process and acts as a rate-limiting enzyme in the biosynthesis of PΦB [55] Exogenoushematin can also induce HO-1 expression in many plants[56 57] In this study we found exogenous hematin signif-icantly increased the expression of FaHO1 (Table 5) Thisresult implies that hematin can promote PΦB biosynthesisRecently it has been reported that hematin could inducethe accumulation of far-red phytochrome and phytochrome
International Journal of Genomics 9
Table 3 DEGs acting as transcription factors in response to exogenous hematin
Annotation transcription factors Gene IDReal
fold-changevalues
Upregulationdownregulation FDR
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb icon00006295 a1g000011 938 Up 120119864 minus 05
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb rscf000031551g000011 4 Down 480119864 minus 16
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL5-like
FANhyb rscf000000291g000101 4 Down 320119864 minus 08
MYBdomain protein 66 FANhyb rscf000001461g000071 654 Up 780119864 minus 07MYB-related protein 3R-1-like partial FANhyb rscf000001411g000161 282 Up 830119864 minus 04PREDICTED transcription factorGAMYB-like FANhyb rscf000006491g000061 303 Down 150119864 minus 13
PREDICTED probable WRKYtranscription factor 53-like FANhyb rscf000019731g000021 217 Down 200119864 minus 08
Transcription factor bHLH80-like FANhyb rscf000012921g000031 6250 Down 150119864 minus 06Transcription factor bHLH70-like FANhyb rscf000002951g000031 526 Down 660119864 minus 12Transcription factor ICE1-like FANhyb rscf000001701g000061 244 Down 450119864 minus 18C2H2-like zinc finger protein FANhyb rscf000011431g000021 282 Up 590119864 minus 04Zinc finger protein MAGPIE-like FANhyb rscf000030241g000011 204 Up 150119864 minus 04PREDICTED axial regulator YABBY5-like FANhyb rscf000016671g000011 417 Down 730119864 minus 05
PREDICTED axial regulator YABBY1-like FANhyb rscf000068991g000011 357 Down 620119864 minus 06
PREDICTED dof zinc finger proteinDOF37-like isoform 1 FANhyb rscf000068561g000011 263 Down 280119864 minus 04
ZF-HD homeobox protein FANhyb rscf000000441g000221 385 Down 420119864 minus 08ZF-HD homeobox proteinAt4g24660-like FANhyb rscf000052411g000011 357 Down 470119864 minus 09
PREDICTED B3 domain-containingprotein REM14-like FANhyb rscf000010091g000021 243 Up 530119864 minus 04
PREDICTED B3 domain-containingtranscription factor ABI3-like FANhyb rscf000012711g000021 455 Down 130119864 minus 28
Growth-regulating factor 6 FANhyb rscf000000241g000291 345 Down 330119864 minus 34Growth-regulating factor 5 FANhyb icon00009704 a1g000011 27 Down 190119864 minus 06GATA type zinc finger transcriptionfactor family protein FANhyb rscf000003931g000131 1111 Down 110119864 minus 05
PREDICTED zinc finger CCCHdomain-containing protein 2-like FANhyb icon00006770 a1g000011 313 Down 680119864 minus 49
PREDICTED E2F transcriptionfactor-like E2FE-like FANhyb icon00002702 a1g000011 278 Down 240119864 minus 04
PREDICTED uncharacterized protein FANhyb rscf000010661g000011 263 Down 490119864 minus 06PREDICTED nuclear transcriptionfactor Y subunit A-3-like isoform 1 FANhyb rscf000000081g000351 385 Down 500119864 minus 20
PREDICTED homeobox-leucinezipper protein ATHB-8-like FANhyb rscf000000151g000051 263 Down 100119864 minus 06
PREDICTED MADS-boxtranscription factor 18-like FANhyb rscf000003231g000161 243 Down 180119864 minus 06
PREDICTED zinc finger protein132-like isoform 1 FANhyb rscf000070151g000011 243 Down 180119864 minus 08
PREDICTED transcription factorPIF3-like FANhyb rscf000006691g000021 346 Up 140119864 minus 07
10 International Journal of Genomics
Table 4 Regulation of the DEGs in anthocyanin biosynthesis in response to exogenous hematin
Genes in anthocyaninbiosynthesis Gene ID
Realfold-change
valuesRegulation level FDR
Dihydroflavonol-4-reductase FANhyb icon00002410 a1g000011 3091 Up 910119864 minus 06
Flavonol synthase FANhyb icon00015354 a1g000011 345 down 180119864 minus 05Isoflavone reductasehomolog FANhyb icon20341135 s1g000011 334 Up 750119864 minus 14
UDP-glycosyltransferase FANhyb rscf000049691g000011 307 Up 640119864 minus 08UDP-glycosyltransferaseactivity FANhyb icon15742070 o1g000011 18 30563 Up 510119864 minus 09
UDP-glycosyltransferase73C1 FANhyb icon00027696 a1g000011 298 Up 560119864 minus 04
UDP-glycosyltransferaseactivity FANhyb icon00008064 a1g000011 249 Up 310119864 minus 04
PREDICTEDUDP-glucose flavonoid3-O-glucosyltransferase7-like
FANhyb rscf000055631g000011 212 Up 110119864 minus 19
PREDICTEDUDP-glycosyltransferase76F1-like
FANhyb rscf000021771g000031 2 Up 720119864 minus 29
A transcripts in the etiolated leaves of wheat seedling phy-tochromes [26] However our study shows that phytochromeA was not differentially expressed It is probably becausethe hematin-treated strawberries were not shaded during theexperiment while the function of phytochrome A is light-dependent In addition we found that a downstream gene ofphytochrome PIF3 (FANhyb rscf000006691g000021) wassignificantly upregulated PIF3 is thought to be a positive reg-ulator of phytochrome B mediated by light-dependent signaltransduction [58]These results suggest that the phytochromepathways may be involved in the anthocyanin biosynthesispromoted by hematin
345 DEGs in Starch and Sucrose Degradation Proteomicapproaches have revealed that starch degradation contributesto anthocyanin accumulation in tuberous roots of the purplesweet potato variety [59] Starch and sucrose biosynthesis isimportant for anthocyanin accumulation in strawberry fruit[60] In this study we found the DEGs involved in starchand sucrose synthesis in the hematin-treated fruit were alldownregulated (Table 5) For example the expression of theunigenes encoding soluble starch synthase 3 sucrose trans-port protein SUC2-like and a probable sucrose-phosphatesynthase 4-like enzyme were decreased 23- 46- 32- and2-fold respectively This suggests that exogenous hematinregulates the biosynthesis of starch and sucrose and henceaffects fruit coloring
346 DEGs in the Calcium Pathway in Response to HematinCalcium can also increase the transcription levels of keystructural genes F3H DFR ANT and UFGT in the whitestage of strawberries [61] In this study we found thatthree DEGs in calcium biosynthesis and transport were
downregulated in the hematin-treated fruit (Table 5) Onlythe unigene encoding predicted cationcalcium exchanger5-like was upregulated The unigenes encoding a calpain-type cysteine protease and a predicted calcium-binding pro-tein PBP1-like were downregulated respectively Exogenousapplication of calcium can promote apple coloring [62] inaddition to variation in anthocyanin content [61] Our resultsindicate that the biosynthesis and transport of calcium areinvolved in the development of coloring in hematin-treatedstrawberries
35 Validation of Selected DEGs by qRT-PCR To validate theexpression of the DEGs obtained from RNA-Seq 11 DEGswere selected for qRT-PCR including structural genes (DFRand UDFGs) transcription factor genes (MYB and bHLH)and phytochrome chromophore-related gene (HO-1) Theprimers used for qRT-PCR are listed in Supplementary TableS1 The qRT-PCR results were consistent with the RNA-Seqdata (Figure 4) except for FANhyb rscf000001411g000161which had a higher log
2ratio (hematin-treatedcontrol) in
the transcriptome data than in qRT-PCR These results indi-cate the RNA-Seq data from the strawberry transcriptome isreproducible and accurate
4 Conclusions
In this study the anthocyanin content in the strawberryfruit was elevated by the application of exogenous hematinThis is the first report that hematin could increase theanthocyanin production in fruit Furthermore we exploredthe effects of the exogenous hematin on metabolic path-ways using genome-wide transcriptome analysis The resultsindicate that the expression levels of many genes involved
International Journal of Genomics 11
Table 5 Other DEGs identified in anthocyanin biosynthesis-related pathways
Anthocyaninbiosynthesis-relatedpathways
Annotation genes Gene IDReal fold-changevalues
Upregulationdownregulation FDR
Calcium ion binding
Cationcalciumexchanger 5-like
FANhyb rscf000019061g000011 203 Up 64119864 minus 06
Calpain-type cysteineprotease family
FANhyb icon00011755 a1g000011 345 Down 18119864 minus 09
Calcium-bindingprotein PBP1-like
FANhyb rscf000007501g000081 323 Down 97119864 minus 04
Calpain-type cysteineprotease family
FANhyb icon00023658 a1g000011 233 Down 60119864 minus 08
CytokininCytokinin
dehydrogenase 5-likeFANhyb iscf00325393 1 s1g000011 667 Down 62119864 minus 06
Cytochrome P450714A1
FANhyb rscf000005921g000031 217 Down 98119864 minus 10
JasmonateJasmonate
O-methyltransferase-like
FANhyb rscf000000041g000131 201 Up 70119864 minus 04
Abscisic acid
Protein ABSCISICACID-INSENSITIVE
5-likeFANhyb rscf000021641g000011 4 Down 16119864 minus 18
ABA overly sensitive 5 FANhyb icon00051144 a1g000011 291 Up 24119864 minus 059-cis-
epoxycarotenoiddioxygenase NCED1
FANhyb icon18399909 o1g000011 441 Up 43119864 minus 06
Abscisic acid81015840-hydroxylase 1-like
FANhyb icon00000938 a1g000011 256 Down 14119864 minus 06
Abscisic acid receptorPYR1-like
FANhyb icon00020426 a1g000011 208 Down 30119864 minus 06
Auxin
Auxin efflux carriercomponent 1-like
FANhyb rscf000010081g000031 384 Down 23119864 minus 06
Auxin response factor8
FANhyb rscf000013061g000021 263 Down 61119864 minus 17
Auxin response factor17-like
FANhyb rscf000060741g000011 217 Down 11119864 minus 36
Starch and sucrose
Glycosyltransferasefamily 35
FANhyb rscf000000341g000081 384 Down 11119864 minus 05
Soluble starchsynthase 3
FANhyb rscf000000451g000041 232 Down 38119864 minus 12
Sucrose transportprotein SUC2-like
FANhyb icon00036727 a1g000011 455 Down 98119864 minus 04
Sucrose transportprotein SUC2-like
FANhyb rscf000007551g000021 323 Down 66119864 minus 10
Probablesucrose-phosphatesynthase 4-like
FANhyb rscf000013501g000021 213 Down 11119864 minus 14
Phytochrome
PREDICTEDtranscription factor
PIF3-likeFANhyb rscf000006691g000021 346 up 14119864 minus 07
Phytochrome E-like FANhyb rscf000004361g000041 4 Down 33119864 minus 05
Heme oxygenase 1 FANhyb icon00004395 a1g000011 258 27119864 minus 07 27119864 minus 07
12 International Journal of Genomics
Hematin
Endoplasmicreticulum
WD40 bLHLMYB
Structural genes
TFs
Nuclear
JA
ABA Sucrose
Plastid
G-cis-neoxanthin
Apophytochrome
StarchHeme
BVSucroseABA
Holophytochrome
PfrPIF3
Flavonoids Anthocyanins
Vacuole
Cytoplasm
Cell wall
①
②③
④⑤
⑥
⑦
PΦB
PΦB
CTKAuxin
Ca2+
Figure 5 Genes and biological pathways that were involved in regulation of anthocyanin accumulation promoted by hematin Outermostbox represents the cell wall Bilayer oval shape represents the plastidThe red-brown ring and network represent the nuclear and endoplasmicreticulum respectively The red crescent represents the vacuole The box in the nucleus represents transcription factors (TFs) Yellow MYBgreenWD40 and red bHLH represent a ternary transcription factor complex which transcribes anthocyanin biosynthesis genes The circlednumbers represent biological pathways involved in the regulation of anthocyanin accumulation promoted by hematin A represents thephytochrome regulation pathwayB represents the starch and sucrose pathwayCDE andF represent phytohormones ABA JA AuxinandCTK regulation pathways respectivelyG represents theCa2+ regulation pathway Pfr represents the far-red phytochrome PIF3 representsphytochrome-interacting factor Each anthocyanin biosynthesis regulatory pathway related gene is in Table 5
in anthocyanin biosynthesis were significantly altered withthe hematin treatment This suggests that the physiologicalprocess of fruit color development is regulated through com-plex interactions among anthocyanin biosynthesis pathwaysplant hormone signal transduction pathways phytochromesignal transduction pathways starch and sugar metabolicpathways calcium pathways and transcription factors (Fig-ure 5) This study adds to the in-depth understanding of thefruit coloration process in strawberry
Competing Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contributions
Jianwei Gao and Liangju Wang conceived and designed theresearch Yi Li Huayin Li Fengde Wang Jingjuan Li andYihui Zhang conducted experiments Yi Li analyzed the dataYi Li and Fengde Wang wrote the manuscript All authorsread and approved the manuscript Yi Li and Huayin Licontributed equally to this work
Acknowledgments
This work was supported by the Modern Agricultural Indus-trial Technology System Funding of Shandong ProvinceChina (SDAIT-02-022-04) the China agriculture researchsystem (CARS-25) and the National Science Foundation ofChina (31401820)
References
[1] K Aaby SMazur A Nes andG Skrede ldquoPhenolic compoundsin strawberry (Fragaria x ananassa Duch) fruits compositionin 27 cultivars and changes during ripeningrdquo Food Chemistryvol 132 no 1 pp 86ndash97 2012
[2] T A Colquhoun L A Levin H RMoskowitz VMWhitakerD G Clark and K M Folta ldquoFraming the perfect strawberryan exercise in consumer-assisted selection of fruit cropsrdquoJournal of Berry Research vol 2 no 1 pp 45ndash61 2012
[3] F Giampieri J M Alvarez-Suarez L Mazzoni et al ldquoThepotential impact of strawberry on human healthrdquo NaturalProduct Research vol 27 no 4 pp 448ndash455 2013
[4] Y S Velioglu GMazza L Gao and B D Oomah ldquoAntioxidantactivity and total phenolics in selected fruits vegetables and
International Journal of Genomics 13
grain productsrdquo Journal of Agricultural amp Food Chemistry vol46 no 10 pp 4113ndash4117 1998
[5] S S Hassellund A Flaa S E Kjeldsen et al ldquoEffects of antho-cyanins on cardiovascular risk factors and inflammation inpre-hypertensive men a double-blind randomized placebo-controlled crossover studyrdquo Journal of Human Hypertensionvol 27 no 2 pp 100ndash106 2013
[6] K J Meyers C B Watkins M P Pritts and R H Liu ldquoAnti-oxidant and antiproliferative activities of strawberriesrdquo Journalof Agricultural amp Food Chemistry vol 51 no 23 pp 6887ndash68922003
[7] S Zafra-Stone T YasminM Bagchi AChatterjee J AVinsonand D Bagchi ldquoBerry anthocyanins as novel antioxidants inhuman health and disease preventionrdquo Molecular Nutrition ampFood Research vol 51 no 6 pp 675ndash683 2007
[8] L Jaakola ldquoNew insights into the regulation of anthocyanin bio-synthesis in fruitsrdquoTrends in Plant Science vol 18 no 9 pp 477ndash483 2013
[9] K Springob J-I Nakajima M Yamazaki and K Saito ldquoRecentadvances in the biosynthesis and accumulation of antho-cyaninsrdquo Natural Product Reports vol 20 no 3 pp 288ndash3032003
[10] A Gonzalez M Zhao J M Leavitt and A M Lloyd ldquoRegu-lation of the anthocyanin biosynthetic pathway by the TTG1bHLHMyb transcriptional complex in Arabidopsis seedlingsrdquoPlant Journal vol 53 no 5 pp 814ndash827 2008
[11] K Petroni and C Tonelli ldquoRecent advances on the regulationof anthocyanin synthesis in reproductive organsrdquo Plant Sciencevol 181 no 3 pp 219ndash229 2011
[12] H-F Jia Y-M Chai C-L Li et al ldquoAbscisic acid plays animportant role in the regulation of strawberry fruit ripeningrdquoPlant Physiology vol 157 no 1 pp 188ndash199 2011
[13] C Merchante J G Vallarino S Osorio et al ldquoEthylene isinvolved in strawberry fruit ripening in an organ-specific man-nerrdquo Journal of Experimental Botany vol 64 no 14 pp 4421ndash4439 2013
[14] A Maier A Schrader L Kokkelink et al ldquoLight and the E3ubiquitin ligase COP1SPA control the protein stability of theMYB transcription factors PAP1 and PAP2 involved in antho-cyanin accumulation in Arabidopsisrdquo Plant Journal vol 74 no4 pp 638ndash651 2013
[15] H Lange W Shropshire and H Mohr ldquoAn analysis of phyto-chrome-mediated anthocyanin synthesisrdquo Plant Physiology vol47 no 5 pp 649ndash655 1971
[16] T D Elich and J C Lagarias ldquoPhytochrome chromophore bio-synthesis both 5-aminolevulinic acid and biliverdin overcomeinhibition by gabaculine in etiolated Avena sativa L seedlingsrdquoPlant Physiology vol 84 no 2 pp 304ndash310 1987
[17] Y Han W Xuan T Yu et al ldquoExogenous hematin alleviatesmercury-induced oxidative damage in the roots of Medicagosativardquo Journal of Integrative Plant Biology vol 49 no 12 pp1703ndash1713 2007
[18] M Koornneef J Cone R Dekens E OrsquoHerne-Robers CSpruit and R Kendrick ldquoPhotomorphogenic responses of longhypocotyl mutants of tomatordquo Journal of Plant Physiology vol120 no 2 pp 153ndash165 1985
[19] M J Terry and R E Kendrick ldquoFeedback inhibition of chloro-phyll synthesis in the phytochrome chromophore-deficientaurea and yellow-green-2 mutants of tomatordquo Plant Physiologyvol 119 no 1 pp 143ndash152 1999
[20] B M Parks and P H Quail ldquoPhytochrome-deficient hy1 andhy2 long hypocotyl mutants of Arabidopsis are defective inphytochrome chromophore biosynthesisrdquo Plant Cell vol 3 no11 pp 1177ndash1186 1991
[21] M J Terry ldquoPhytochrome chromophore-deficient mutantsrdquoPlant Cell amp Environment vol 20 no 6 pp 740ndash745 1997
[22] T Muramoto T Kohchi A Yokota I Hwang and H MGoodman ldquoThe Arabidopsis photomorphogenic mutant hy1 isdeficient in phytochrome chromophore biosynthesis as a resultof a mutation in a plastid heme oxygenaserdquo Plant Cell vol 11no 3 pp 335ndash347 1999
[23] M D Maines ldquoThe heme oxygenase system a regulator ofsecond messenger gasesrdquo Annual Review of Pharmacology andToxicology vol 37 no 1 pp 517ndash554 1997
[24] W Xuan F-Y Zhu S Xu et al ldquoThe heme oxygenasecarbonmonoxide system is involved in the auxin-induced cucumberadventitious rooting processrdquo Plant Physiology vol 148 no 2pp 881ndash893 2008
[25] R Amooaghaie F Tabatabaei and A-M Ahadi ldquoRole ofhematin and sodium nitroprusside in regulating Brassica nigraseed germination under nanosilver and silver nitrate stressesrdquoEcotoxicology and Environmental Safety vol 113 pp 259ndash2702015
[26] Y Liu X Li L Xu and W Shen ldquoDe-etiolation of wheatseedling leaves cross talk between heme oxygenasecarbonmonoxide and nitric oxiderdquo PLoS ONE vol 8 no 12 ArticleID e81470 2013
[27] T Muramoto N Tsurui M J Terry A Yokota and T KohchildquoExpression and biochemical properties of a ferredoxin-dependent heme oxygenase required for phytochrome chro-mophore synthesisrdquo Plant Physiology vol 130 no 4 pp 1958ndash1966 2002
[28] J-W Gao J Liu B Li and Z Li ldquoIsolation and purificationof functional total RNA from blue-grained wheat endospermtissues containing high levels of starches and flavonoidsrdquo PlantMolecular Biology Reporter vol 19 no 2 pp 185ndash186 2001
[29] R Patro S M Mount and C Kingsford ldquoSailfish enablesalignment-free isoform quantification from RNA-seq readsusing lightweight algorithmsrdquoNature Biotechnology vol 32 no5 pp 462ndash464 2014
[30] Y-X Qi Y-B Liu and W-H Rong ldquoRNA-Seq and its applica-tions a new technology for transcriptomicsrdquo Hereditas vol 33no 11 pp 1191ndash1202 2011
[31] C R Genovese N A Lazar and T Nichols ldquoThresholdingof statistical maps in functional neuroimaging using the falsediscovery raterdquo NeuroImage vol 15 no 4 pp 870ndash878 2002
[32] J Ye L Fang H Zheng et al ldquoWEGO a web tool for plottingGO annotationsrdquoNucleic Acids Research vol 34 supplement 2pp W293ndashW297 2006
[33] H Abdi ldquoThe Bonferonni and Sidak corrections for multiplecomparisonrdquo Encyclopedia of Measurement and Statistics vol 3pp 103ndash107 2007
[34] J Li Q Ding F Wang Y Zhang H Li and J Gao ldquoIntegrativeanalysis of mRNA and miRNA expression profiles of thetuberous root development at seedling stages in turnipsrdquo PLoSONE vol 10 no 9 Article ID e0137983 2015
[35] J Bennett D Hondred and J C Register ldquoKeeping qRT-PCRrigorous and biologically relevantrdquo Plant Cell Reports vol 34no 1 pp 1ndash3 2015
[36] F Wang L Li H Li et al ldquoTranscriptome analysis of rosetteand folding leaves in Chinese cabbage using high-throughputRNA sequencingrdquo Genomics vol 99 no 5 pp 299ndash307 2012
14 International Journal of Genomics
[37] T Shi Z Gao L Wang et al ldquoIdentification of differentially-expressed genes associated with pistil abortion in Japaneseapricot by genome-wide transcriptional analysisrdquo PLoS ONEvol 7 no 10 Article ID e47810 2012
[38] F Wang H Li Y Zhang et al ldquoMicroRNA expression analysisof rosette and folding leaves in Chinese cabbage using high-throughput Solexa sequencingrdquo Gene vol 532 no 2 pp 222ndash229 2013
[39] X Gu Y Chen Z Gao Y Qiao and X Wang ldquoTranscriptionfactors and anthocyanin genes related to low-temperature toler-ance in rd29A RdreB1BI transgenic strawberryrdquo Plant Physiol-ogy and Biochemistry vol 89 pp 31ndash43 2015
[40] J G Schaart C Dubos I R De La Fuente et al ldquoIdentificationand characterization of MYB-bHLH-WD40 regulatory com-plexes controlling proanthocyanidin biosynthesis in strawberry(Fragaria times ananassa) fruitsrdquoNew Phytologist vol 197 no 2 pp454ndash467 2013
[41] L Xie Z H Wang X H Cheng J J Gao Z P Zhangand L J Wang ldquo5-Aminolevulinic acid promotes anthocyaninaccumulation in Fuji applesrdquo Plant Growth Regulation vol 69no 3 pp 295ndash303 2013
[42] M Daminato F Guzzo and G Casadoro ldquoASHATTERPROOF-like gene controls ripening in non-climac-teric strawberries and auxin and abscisic acid antagonisticallyaffect its expressionrdquo Journal of Experimental Botany vol 64no 12 pp 3775ndash3786 2013
[43] X-H Ji R Zhang N Wang L Yang and X-S Chen ldquoTran-scriptome profiling reveals auxin suppressed anthocyanin bio-synthesis in red-fleshed apple callus (Malus sieversii f niedzwet-zkyana)rdquo Plant Cell Tissue and Organ Culture vol 123 no 2pp 389ndash404 2015
[44] J Chen LMaoW Lu T Ying and Z Luo ldquoTranscriptome pro-filing of postharvest strawberry fruit in response to exogenousauxin and abscisic acidrdquo Planta vol 243 no 1 pp 183ndash197 2016
[45] G A Martınez A R Chaves and M C Anon ldquoEffect of gib-berellic acid on ripening of strawberry fruits (Fragaria annan-assa Duch)rdquo Journal of Plant Growth Regulation vol 13 no 2pp 87ndash91 1994
[46] P K Das D H Shin S-B Choi S-D Yoo G Choi and Y-IPark ldquoCytokinins enhance sugar-induced anthocyanin biosyn-thesis in Arabidopsisrdquo Molecules amp Cells vol 34 no 1 pp 93ndash101 2012
[47] P McAtee S Karim R Schaffer and K David ldquoA dynamicinterplay between phytohormones is required for fruit develop-ment maturation and ripeningrdquo Frontiers in Plant Science vol4 no 3 article 79 7 pages 2013
[48] E Nambara andAMarion-Poll ldquoAbscisic acid biosynthesis andcatabolismrdquoAnnual Review of Plant Biology vol 56 pp 165ndash1852005
[49] Y Jiang and D C Joyce ldquoABA effects on ethylene productionPAL activity anthocyanin and phenolic contents of strawberryfruitrdquo Plant Growth Regulation vol 39 no 2 pp 171ndash174 2003
[50] S Li ldquoTranscriptional control of flavonoid biosynthesis fine-tuning of the MYB-bHLH-WD40 (MBW) complexrdquo PlantSignaling amp Behavior vol 9 Article ID e27522 2014
[51] D R Rudell J K Fellman and J P Mattheis ldquoPreharvest appli-cation of methyl jasmonate to rsquoFujirsquo apples enhances red col-oration and affects fruit size splitting and bitter pit incidencerdquoHortScience vol 40 no 6 pp 1760ndash1762 2005
[52] A Belhadj N Telef C Saigne et al ldquoEffect of methyl jasmonatein combination with carbohydrates on gene expression of PR
proteins stilbene and anthocyanin accumulation in grapevinecell culturesrdquo Plant Physiology and Biochemistry vol 46 no 4pp 493ndash499 2008
[53] E Loreti G Povero G Novi C Solfanelli A Alpi and PPerata ldquoGibberellins jasmonate and abscisic acid modulate thesucrose-induced expression of anthocyanin biosynthetic genesin Arabidopsisrdquo New Phytologist vol 179 no 4 pp 1004ndash10162008
[54] H S Seo J T Song J-J Cheong et al ldquoJasmonic acid carboxylmethyltransferase A key enzyme for jasmonate-regulated plantresponsesrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 98 no 8 pp 4788ndash4793 2001
[55] M T McDowell and J C Lagarias ldquoPurification and biochemi-cal properties of phytochromobilin synthase from etiolated oatseedlingsrdquo Plant Physiology vol 126 no 4 pp 1546ndash1554 2001
[56] G-Q Fu S Xu Y-J Xie et al ldquoMolecular cloning characteri-zation and expression of an alfalfa (Medicago sativa L) hemeoxygenase-1 gene MsHO1 which is pro-oxidants-regulatedrdquoPlant Physiology and Biochemistry vol 49 no 7 pp 792ndash7992011
[57] X-Y Chen X Ding S Xu et al ldquoEndogenous hydrogen per-oxide plays a positive role in the upregulation of heme oxy-genase and acclimation to oxidative stress in wheat seedlingleavesrdquo Journal of Integrative Plant Biology vol 51 no 10 pp951ndash960 2009
[58] J Shin E Park and G Choi ldquoPIF3 regulates anthocyaninbiosynthesis in an HY5-dependent manner with both factorsdirectly binding anthocyanin biosynthetic gene promoters inArabidopsisrdquo Plant Journal vol 49 no 6 pp 981ndash994 2007
[59] S Wang D Pan X Lv et al ldquoProteomic approach reveals thatstarch degradation contributes to anthocyanin accumulation intuberous root of purple sweet potatordquo Journal of Proteomics vol143 pp 298ndash305 2016
[60] H-F Jia C-L Li Y-M Chai Y Xing and Y Shen ldquoSucrosepromotes strawberry fruit ripening by stimulation of abscisicacid biosynthesisrdquo Pakistan Journal of Botany vol 45 no 1 pp169ndash176 2013
[61] W Xu H Peng T Yang et al ldquoEffect of calcium on strawberryfruit flavonoid pathway gene expression and anthocyanin accu-mulationrdquo Plant Physiology amp Biochemistry vol 82 pp 289ndash298 2014
[62] Z-W-SWan and Z Singh ldquoExogenous application of prohexa-dione-calcium promotes fruit colour development of lsquoCrippsPinkrsquo applerdquo Acta Horticulturae vol 1012 pp 219ndash225 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
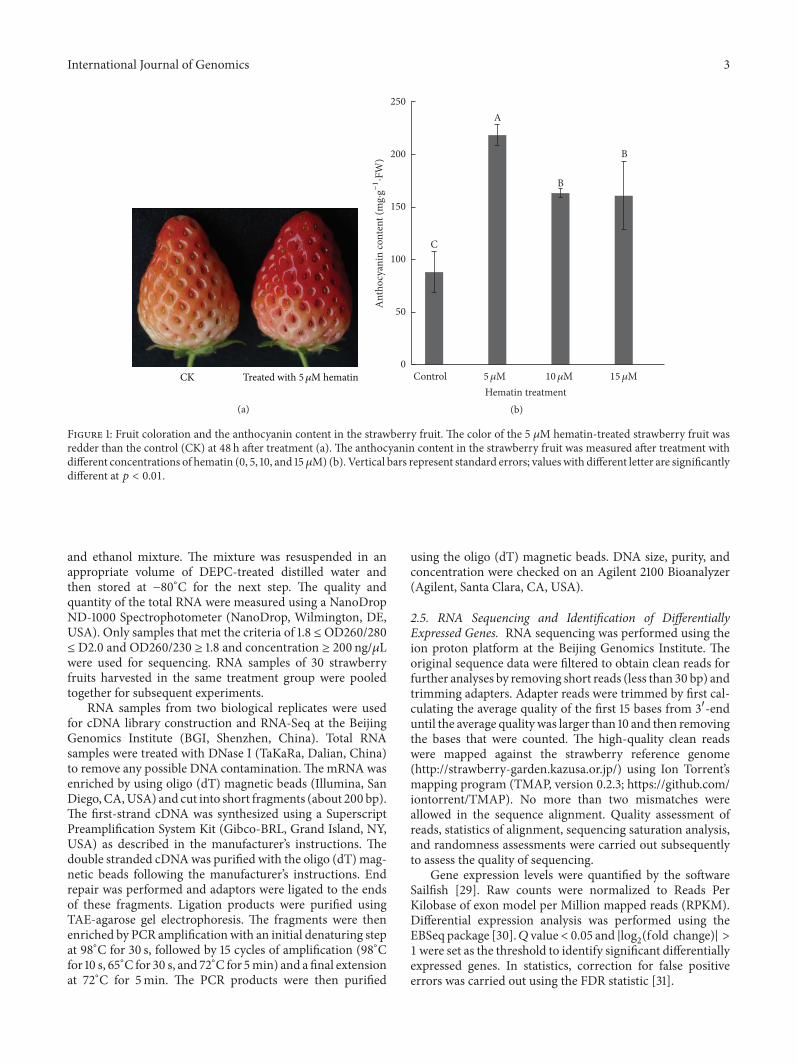
International Journal of Genomics 3
CK Treated with 5120583M hematin
(a)
0
50
100
150
200
250
ControlHematin treatment
C
B
B
A
15120583M10120583M5120583MA
ntho
cyan
in co
nten
t (m
gmiddotgminus
1middotF
W)
(b)
Figure 1 Fruit coloration and the anthocyanin content in the strawberry fruit The color of the 5 120583M hematin-treated strawberry fruit wasredder than the control (CK) at 48 h after treatment (a) The anthocyanin content in the strawberry fruit was measured after treatment withdifferent concentrations of hematin (0 5 10 and 15120583M) (b) Vertical bars represent standard errors values with different letter are significantlydifferent at 119901 lt 001
and ethanol mixture The mixture was resuspended in anappropriate volume of DEPC-treated distilled water andthen stored at minus80∘C for the next step The quality andquantity of the total RNA were measured using a NanoDropND-1000 Spectrophotometer (NanoDrop Wilmington DEUSA) Only samples that met the criteria of 18 leOD260280le D20 and OD260230 ge 18 and concentration ge 200 ng120583Lwere used for sequencing RNA samples of 30 strawberryfruits harvested in the same treatment group were pooledtogether for subsequent experiments
RNA samples from two biological replicates were usedfor cDNA library construction and RNA-Seq at the BeijingGenomics Institute (BGI Shenzhen China) Total RNAsamples were treated with DNase I (TaKaRa Dalian China)to remove any possible DNA contaminationThemRNA wasenriched by using oligo (dT) magnetic beads (Illumina SanDiegoCAUSA) and cut into short fragments (about 200 bp)The first-strand cDNA was synthesized using a SuperscriptPreamplification System Kit (Gibco-BRL Grand Island NYUSA) as described in the manufacturerrsquos instructions Thedouble stranded cDNAwas purified with the oligo (dT)mag-netic beads following the manufacturerrsquos instructions Endrepair was performed and adaptors were ligated to the endsof these fragments Ligation products were purified usingTAE-agarose gel electrophoresis The fragments were thenenriched by PCR amplificationwith an initial denaturing stepat 98∘C for 30 s followed by 15 cycles of amplification (98∘Cfor 10 s 65∘C for 30 s and 72∘C for 5min) and a final extensionat 72∘C for 5min The PCR products were then purified
using the oligo (dT) magnetic beads DNA size purity andconcentration were checked on an Agilent 2100 Bioanalyzer(Agilent Santa Clara CA USA)
25 RNA Sequencing and Identification of DifferentiallyExpressed Genes RNA sequencing was performed using theion proton platform at the Beijing Genomics Institute Theoriginal sequence data were filtered to obtain clean reads forfurther analyses by removing short reads (less than 30 bp) andtrimming adapters Adapter reads were trimmed by first cal-culating the average quality of the first 15 bases from 31015840-enduntil the average quality was larger than 10 and then removingthe bases that were counted The high-quality clean readswere mapped against the strawberry reference genome(httpstrawberry-gardenkazusaorjp) using Ion Torrentrsquosmapping program (TMAP version 023 httpsgithubcomiontorrentTMAP) No more than two mismatches wereallowed in the sequence alignment Quality assessment ofreads statistics of alignment sequencing saturation analysisand randomness assessments were carried out subsequentlyto assess the quality of sequencing
Gene expression levels were quantified by the softwareSailfish [29] Raw counts were normalized to Reads PerKilobase of exon model per Million mapped reads (RPKM)Differential expression analysis was performed using theEBSeq package [30]119876 valuelt 005 and |log
2(fold change)| gt
1were set as the threshold to identify significant differentiallyexpressed genes In statistics correction for false positiveerrors was carried out using the FDR statistic [31]
4 International Journal of Genomics
26 Gene Ontology and Pathway Enrichment Analysis ofDifferentially Expressed Genes Gene functions of strawberrywere annotated according to the Gene Ontology (GO) stand-ardized terms for molecular function cellular componentand biological process using Blast2GO (httpswwwblast2gocom) Gene function was annotated using the followingdatabases NCBI nonredundant protein sequences NCBInonredundant nucleotide sequences protein family Clustersof Orthologous Groups of proteins the manually annotatedand reviewed protein sequence database KEGG Orthologdatabase and Gene Ontology After GO annotation forDEGs we performed GO functional classification for DEGsby the WEGO software [32] and analyzed the distributionof gene functions We used the GO Term Finder tool(httpwwwyeastgenomeorghelpanalyzego-term-finder)to search for significant shared GO terms
119901 = 1 minus119898minus1
sum119894=0
(119872119894) (119873minus119872119899minus119894)
(119873119899) (2)
where 119873 is the number of all genes with GO annotation119899 is the number of DEGs in 119873 119872 is the number of allgenes that are annotated to certain GO terms and 119898 is thenumber of DEGs in119872 The calculated 119901 value was subjectedto Bonferroni Correction [33] The corrected 119901 value lt 005was set as the threshold GO terms fulfilling this conditionare defined as significantly enriched GO terms in DEGsThe pathway enrichment analyses using the KEGG database(httpwwwgenomejpkegg) were conducted subsequentlyto study the functions of differentially expressed genes identi-fied between the control and the hematin-treated strawberrygroups The formula is the same as that in GO analysis Here119873 is the number of all genes with KEGG annotation 119899 is thenumber ofDEGs in119873119872 is the number of all genes annotatedto specific pathways and119898 is the number of DEGs in119872
27 Quantitative Real-Time PCR Analysis Quantitative real-time PCR (qRT-PCR) experiments were conducted to assessthe reliability of the RNA-Seq data [34] Eleven pairs of genespecific primers were designed to verify the DEGs in thestrawberry fruit (Table S1) Strawberry RNA was extractedfrom the fruit according to the previously detailed methodTotal RNA was digested with DNase I for 30min at 25∘C toremove DNA contamination according to the manufacturerrsquosinstructions The qRT-PCR was performed using a SYBRPCR master mix (TaKaRa Dalian China) on a Bio-Rad IQ-5 thermal cycler (Bio-Rad Philadelphia PA USA) Threereplicates of each sample were conducted to calculate theaverage Ct valuesThe relative expression level was calculatedby the comparative 2minusΔΔCt method [35 36] The significancewas determined with the SPSS software (SPSS 170 IBMChicago IL USA) (119901 lt 005)
3 Results and Discussion
31 Effects of Exogenous Hematin on Anthocyanin andBiliverdin Accumulation Hematin is an inducer and sub-strate of HO1 in animals and plants [17 23] In this studystrawberries at theWt stage were treated with 0 120583M(control)
00
05
10
15
20
25
ControlHematin treatment
B
A
5120583M hematin
Biliv
erdi
n co
nten
t (120583
Mmiddotg
minus1middotF
W)
Figure 2 The BV content in strawberry fruit was measured aftertreatment with 0 and 5 120583Mhematin Vertical bars represent standarderrors values with different letter are significantly different at 119901 lt001
5 10 or 15 120583M hematin The results indicated that the antho-cyanin content in the hematin-treated strawberries was morethan the control after 48 h The treatment by 5120583M hematinwas found to be the optimal condition for increasing antho-cyanin production (Figure 1) (119901 lt 001) The anthocyanincontent in the 5 120583M hematin-treated strawberries was 25times higher than that in the control This is the first reportthat hematin could increase the anthocyanin production infruit Wemeasured the expression of FaHO-1 and the contentof biliverdin (BV) which is themetabolite of heme oxygenaseWe found that hematin could also significantly increase theexpression of FaHO-1 in strawberry fruit (Figure 4) andpromote the accumulation of BV (Figure 2) These resultsindicate that hematin promotes anthocyanin accumulationthrough the heme metabolism pathway
32 Sequences Assembly Mapping and Functional Anno-tation High-throughput sequencing technology is utilizedwidely in analyzing gene expression in many organisms[37 38] In this study approximately 108 Gbp raw tagswere generated for each library After eliminating adaptersambiguous nucleotides and low-quality sequences a total of34618832 and 45464123 clean reads between 150 and 200nucleotides in length were obtained (Table 1) Over 85 ofthe clean tags from each library mapped to reference geneswith less than 2 bp mismatches Less than 29 of the cleantags from each library could not be aligned to any referencegenes because of incomplete sequences and these tags weredesignated as unknown More than 673 of the clean tagsin each library were mapped to a single gene while less than47mapped to multiple reference genes Unknown tags andtags mapped to multigenes were filtered out and the unique
International Journal of Genomics 5
Table 1 Overview of the sequencing and assembly
Biological replicate 1 Biological replicate 2Sample Control Hematin-treated Control Hematin-treatedClean reads 17808924 26809908 19235452 26228671Genome map rate () 9658 9652 9665 9674Gene map rate () 8646 8627 8704 8563Perfect match () 4677273 (2432) 6510668 (2428) 4414050 (2479) 6149508 (2345)Unique match () 15075148 (8465) 22628939 (8441) 16409557 (8531) 21986962 (8383)Expressed genes 27976 28713 28999 28211Mismatch () 11321162 (6357) 17101590 (6379) 12456871 (6476) 16697280 (6366)Total unmapped reads 645135 (335) 932213 (348) 608914 (342) 856028 (326)
clean tags that mapped to a single gene were retained forfurther DEG analysis
In addition the perfect match rates were 2432 24282479 and 2435 in control 1 control 2 hematin 1 andhematin 2 respectively
The saturation of the libraries with and without thehematin treatment was analyzedThe number of the detectedgenes became saturated at about 2 million reads (FigureS2) This indicates that the obtained reads are sufficient forcomplete transcriptome coverage The randomness of RNAfragmentation in the four libraries was assessed for subse-quent bioinformatics analysis The results showed that thereads in each position of the reference gene were distributedevenly and demonstrated highly similar tendencies in alllibraries (Figure S3)
33 Genes Differentially Regulated in Response to ExogenousHematin Based on sequencing results of the four mRNAlibraries approximately 29000 genes (about 70 of thereference genes in the octoploid strawberry genome) weredetected in each library Additionally correlations amonggenes based on RPKM between the two biological replicateswere analyzed the correlation coefficients (1198772) were high(093 and 095 for the control and hematin-treated groupresp) Genes expressed in both replicates were screened Atotal of 28713 28999 27976 and 28211 genes (Table 1) wereexpressed in the control and hematin-treated samples
To reveal the molecular pathways regulating strawberrycoloring in response to hematin the DEGs between the con-trol and hematin-treated fruit were analyzed We comparedthe gene expression profiles between the control and thetreated samples in both biological replicates DEGs detectedin both biological replicates were screened for subsequentanalysis A total of 1080 (402 up- and 678 downregulated)genes were differentially expressed in the 5120583M hematin-treated groups comparedwith the control groupsMore geneswere downregulated in the hematin-treated group comparedto the control group
To facilitate the global analysis of gene expression aGO analysis was performed by mapping each differentiallyexpressed gene into the records of the GO database GeneOntology (GO) enrichment analysis of the DEGs in thecontrol and hematin-treated groups was performed to revealthe possible mechanisms under which hematin promotes
anthocyanin biosynthesis GO terms with 119901 lt 005 wererepresented among genes with significant changes in expres-sion over a given interval In this study only three termsincluding anatomical structure arrangementmeristem struc-tural organization radial pattern formationwere significantlyenriched in biological process (Supplementary Table S2)AlthoughmostGo termswere not significantly enriched theycould provide a reference for further study According to theGO cellular components a large number of the DEGs wereclassified intomain cell organelles such as vacuole nucleolusmitochondrion apoplast chloroplast plastid cytoplasm andmembrane In particular many DEGs were assigned to theplastid and chloroplast (Figure 3) indicating that hematinhas an important impact on the expression of plastid andchloroplast genes It is probably because heme oxygenase 1 isa soluble plastid protein [22] According to the GO molec-ular function a large number of the DEGs were classifiedintoUDP-glucosyltransferase sucrose transmembrane trans-porter activity nucleic acid binding and DNA binding termswhich are closely related to anthocyanin biosynthesis ManyDEGs were classified into heme binding peroxidase activityand tetrapyrrole binding (Figure 3) It is probably becausehematin can increase the activity of HO1 and promote thedegradation of hemoglobin [17] In addition anthocyaninaccumulation in fruit is closely related to hormone [8] andcarbohydrate biosynthesis [9] According to the GO biologi-cal process a large number of the DEGs were classified intoregulation of hormone levels cytokinin metabolic processresponse to hormone response to abscisic acid response toauxin and response to ethylene terms indicating that thehormone-related DEGs are involved in regulating the antho-cyanin biosynthesis in the hematin-treated fruit Many DEGswere classified into starch and sucrose biosynthetic and trans-port process and phenylpropanoid metabolic process termsindicating that hematin can promote anthocyanin accumu-lation via the phenylpropanoid metabolic and carbohydratemetabolism In addition many DEGs were classified into theprocess for response to light stimulus term indicating thathematin may also participate in the light stimulus system
Pathway enrichment analysis was performed to under-stand the biological functions of the DEGs by identify-ing significantly enriched signal transduction pathways ormetabolic pathways [39] In the study a number of alteredbiological pathways associated with the hematin treatment
6 International Journal of Genomics
0 10 20 30 40 50 60 70 80Starch biosynthetic process
Cytokinin metabolic processResponse to abscisic acid
Response to auxinResponse to ethylene
Phenylpropanoid metabolic processSucrose transport
Regulation of hormone levelsResponse to light stimulus
Response to hormonePeroxidase activity
Sucrose transmembrane transporter activityUDP-glucosyltransferase
Heme bindingTetrapyrrole binding
DNA bindingNucleic acid binding
VacuoleNucleolusApoplast
MitochondrionChloroplast
PlastidCytoplasmMembrane
Biol
ogic
al p
rogr
ess
Mol
ecul
ar fu
nctio
nC
ellu
lar c
ompo
nent
Number of DEGs
Figure 3 Gene classification based on Gene Ontology (GO) for DEGs in the strawberries treated by hematin
were identified and three pathways including RNA poly-merase pyrimidine metabolism and purine metabolismwere significantly enriched (119876-value lt 005) (Table 2)Although the majority of the pathway terms were not sig-nificantly enriched the pathway enrichment analysis helpsus to further understand the biological functions of theDEGs and the molecular mechanisms that regulate fruitcoloring in response to the hematin treatmentThese pathwayterms included metabolic pathways biosynthesis of sec-ondary metabolites isoflavonoid biosynthesis phenylalaninemetabolism phenylalanine tyrosine and tryptophan biosyn-thesis starch and sucrose metabolism plant hormone signaltransduction flavone and flavonol biosynthesis and phenyl-propanoid biosynthesis terms (Supplementary Table S2)
34 Analysis of DEGs Involved in Anthocyanin Biosynthesis
341 Transcription Factors Transcription factors (TFs) areproteins that regulate the expression of downstream targetgenes Many TFs such as v-myb avian myeloblastosis viraloncogene homolog (MYB) basic helix-loop-helix (bHLH)WD40-repeats protein (WD40) and MADS-box (MADS)directly regulating the expression of the structural genes inanthocyanin biosynthesis have been identified from manyspecies [8] Therefore the enhanced expression of key TFs
might regulate the anthocyanin biosynthesis In our studysome anthocyanin biosynthesis-related TFs were found to beuniquely present in the DEG profiling of the hematin-treated fruit for example MYB bHLH PIF3 MADs-boxAP2-EREBP and ABI3VP1 (Table 3) The anthocyanin bio-synthesis genes are regulated primarily by the MBW tran-scription factor which is a ternary transcription factor com-plex [10 11] Most of the MYBs involved in regulating antho-cyanin biosynthesis are positive regulators of transcription[8] However MYBs can act as repressors too such as straw-berry FaMYB1 and FaMYB9 and grapevine VvMYB4 whichcan significantly suppress the biosynthesis of anthocyaninsand flavonols [40] In this study three unigenes belong-ing to MYB transcription factors were differentiallyexpressed and two MYB transcription factors were signifi-cantly upregulated Of these the unigene ldquoFANhybrscf000001461g000071rdquo from theMYB family of R2R3MYBtranscription factors was significantly upregulatedMoreoverincreasing evidence indicates that the expression of thebHLHs promotes anthocyanin accumulation in fruit [41] Inour study all of the unigenes from the bHLH family weredownregulated among which the expression level of unigeneFANhyb rscf000012921g000031 was significantly decreased6250-fold (Table 3) FANhyb rscf000022101g000011 en-codes a predicted MADS-box transcription factor and it was
International Journal of Genomics 7
Control Hematin-treated
FANhyb_icon00002410_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_rscf000049691g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00027696_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00004395_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_rscf000063641g000011
lowastlowast
lowast
Control Hematin-treated
FANhyb_rscf000001411g000161
Control Hematin-treated
FANhyb_icon00006295_a1g000011lowastlowast
Control Hematin-treated
FANhyb_rscf000031551g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00001444_a1g000011
lowastlowast
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
0
05
1
15
2
25
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
005
115
225
335
445
5
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
Control Hematin-treated
FANhyb_rscf000023111g000011
lowastlowast
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
Control Hematin-treated
FANhyb_rscf000002951g000031
lowastlowast
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
Figure 4 qRT-PCR validation of the RNA-Seq based gene expression The values indicate means of three biological replicates plusmn SD Starindicates that the expression level is significantly different between the hematin-treated and the control group (lowast119901 lt 005 lowastlowast119901 lt 001)
8 International Journal of Genomics
Table 2 Pathway enrichment analysis of differentially expressed genes
Pathway DEGs with pathwayannotation (451)
All genes with pathwayannotation (15877)
Corrected 119901 value 119876-value Pathway ID
RNA polymerase 16 183 688119864 minus 5 00068107 ko03020Pyrimidine metabolism 20 317 00007651746 00320 ko00240Purine metabolism 21 347 00009710371 0032044224 ko00230
significantly downregulated There were three unigenes inthe AP2-EREBP family with only one upregulated and theothers downregulated These results suggest that exogenoushematin is involved in developmental transcriptionalregulation of anthocyanin biosynthesis
342 DEGs in Anthocyanin Biosynthesis The anthocyaninbiosynthetic pathway has been studied intensively Theanthocyanin biosynthetic pathway via the phenylpropanoidpathway is well known [8] In this study the transcriptomedata showed a significant increase in the expression of genesin the anthocyanin biosynthesis pathway when exposedto exogenous hematin The structural genes involved inanthocyanin biosynthesis including DFR LAR and UDP-glycosyltransferase exhibited significant differential expres-sion in response to the hematin treatment (Table 4) Forexample two DFR genes and one LAR gene were signifi-cantly upregulated (the expression increased 18- 30- and3-fold in the hematin-treated fruit resp) In addition allsix genes encodingUDP-glycosyltransferasewere significantlyupregulated Among these the expression of one gene(Fanhy icon15742070o1g000011) was upregulated 18305-fold The results suggest that exogenous hematin may havean important role in promoting anthocyanin biosynthesis
343 DEGs in Hormone Signaling Transduction in Responseto Hematin Auxin has been shown to negatively regulatethe expression of the anthocyanin biosynthesis genes [42] Inthis study there were three unigenes involved in the auxinsignaling pathway and all were downregulated includingone gene encoding auxin efflux carrier component 1-like(AUX1-like) and two genes encoding auxin response factors(ARF) These results suggest that hematin promotes antho-cyanin accumulation by regulating auxin in the colorationprocess of strawberries It has shown that auxin suppressesanthocyanin biosynthesis in the red-fleshed apple callus [43]Endogenous expression of auxin [44 45] has been foundto impede anthocyanin accumulation in strawberries Inadditional cytokinins are also known to play an importantrole in anthocyanin biosynthesis It has shown to enhanceanthocyanin accumulation in Zea mays and regulate antho-cyanin production and composition in suspension culturesof strawberry cells [46] In this study two unigenes relatedto the cytokinin signaling pathway were identified and theywere downregulated as well One gene was identified ascytokinin dehydrogenase 5-like which catalyzes the irre-versible degradation of cytokinins This result indicates thathematin promotes anthocyanin accumulation by regulatingthe cytokinin signaling pathway
Extensive researches have shown that ABA plays animportant role in the regulation of anthocyanin biosynthesisin nonclimacteric fruit [47] In this study a total of five uni-genes related to the ABA signaling pathway were identifiedamong which two were upregulated and three were down-regulated Two genes which are involved in the abscisic acidresponse were upregulated These two unigenes encode 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) and ABA overlysensitive 5 (ABO5) respectively ABA is synthesized fromcarotenoids via several enzymatic reactions in the plastidThe rate-limiting step in these reactions is catalyzed by 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) [48] Mutations inthe FaNCED1 gene result in colorless strawberry fruit whichcan become colored through the application of exogenousABA [12] Another unigene was identified to encode abscisicacid 81015840-hydroxylase 1-like a key enzyme in the oxidativecatabolism of abscisic acid which was downregulated Appli-cation of exogenous ABA regulates phenylalanine ammonialyase activity and increases the phenolic and anthocyanincontent of strawberry fruit [12 49] Our results suggestthat hematin promotes ABA biosynthesis and inhibits ABAdisintegration in the hematin-treated fruit
In Arabidopsis JAs can affect anthocyanin accumulationvia the interaction of negative regulators with the MBWcomplex of transcription factors involved in anthocyaninbiosynthesis [50] Preharvest application of JA to ldquoFujirdquo applesenhances red coloration [51] JA vapor treatment can alsoenhance anthocyanins in strawberry fruit [52] In this studyjasmonate O-methyltransferase-like exhibited significantlydifferential expression Jasmonate O-methyltransferase-likeis also known to be a key enzyme for jasmonate-regulatedplant responses to stimuli [53 54] These results suggest thathematin increases the activity of the JA-regulated antho-cyanin biosynthesis
344 DEGs in the Phytochrome Signaling Pathway in Responseto Hematin Phytochromes that act as photoreceptors playan important role in anthocyanin regulation [15] Phy-tochromes are homodimeric chromoproteins where eachholophytochrome is composed of a phytochrome protein(apophytochrome) covalently bound to a linear tetrapyrrolePΦB HO1 is crucial to this process and acts as a rate-limiting enzyme in the biosynthesis of PΦB [55] Exogenoushematin can also induce HO-1 expression in many plants[56 57] In this study we found exogenous hematin signif-icantly increased the expression of FaHO1 (Table 5) Thisresult implies that hematin can promote PΦB biosynthesisRecently it has been reported that hematin could inducethe accumulation of far-red phytochrome and phytochrome
International Journal of Genomics 9
Table 3 DEGs acting as transcription factors in response to exogenous hematin
Annotation transcription factors Gene IDReal
fold-changevalues
Upregulationdownregulation FDR
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb icon00006295 a1g000011 938 Up 120119864 minus 05
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb rscf000031551g000011 4 Down 480119864 minus 16
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL5-like
FANhyb rscf000000291g000101 4 Down 320119864 minus 08
MYBdomain protein 66 FANhyb rscf000001461g000071 654 Up 780119864 minus 07MYB-related protein 3R-1-like partial FANhyb rscf000001411g000161 282 Up 830119864 minus 04PREDICTED transcription factorGAMYB-like FANhyb rscf000006491g000061 303 Down 150119864 minus 13
PREDICTED probable WRKYtranscription factor 53-like FANhyb rscf000019731g000021 217 Down 200119864 minus 08
Transcription factor bHLH80-like FANhyb rscf000012921g000031 6250 Down 150119864 minus 06Transcription factor bHLH70-like FANhyb rscf000002951g000031 526 Down 660119864 minus 12Transcription factor ICE1-like FANhyb rscf000001701g000061 244 Down 450119864 minus 18C2H2-like zinc finger protein FANhyb rscf000011431g000021 282 Up 590119864 minus 04Zinc finger protein MAGPIE-like FANhyb rscf000030241g000011 204 Up 150119864 minus 04PREDICTED axial regulator YABBY5-like FANhyb rscf000016671g000011 417 Down 730119864 minus 05
PREDICTED axial regulator YABBY1-like FANhyb rscf000068991g000011 357 Down 620119864 minus 06
PREDICTED dof zinc finger proteinDOF37-like isoform 1 FANhyb rscf000068561g000011 263 Down 280119864 minus 04
ZF-HD homeobox protein FANhyb rscf000000441g000221 385 Down 420119864 minus 08ZF-HD homeobox proteinAt4g24660-like FANhyb rscf000052411g000011 357 Down 470119864 minus 09
PREDICTED B3 domain-containingprotein REM14-like FANhyb rscf000010091g000021 243 Up 530119864 minus 04
PREDICTED B3 domain-containingtranscription factor ABI3-like FANhyb rscf000012711g000021 455 Down 130119864 minus 28
Growth-regulating factor 6 FANhyb rscf000000241g000291 345 Down 330119864 minus 34Growth-regulating factor 5 FANhyb icon00009704 a1g000011 27 Down 190119864 minus 06GATA type zinc finger transcriptionfactor family protein FANhyb rscf000003931g000131 1111 Down 110119864 minus 05
PREDICTED zinc finger CCCHdomain-containing protein 2-like FANhyb icon00006770 a1g000011 313 Down 680119864 minus 49
PREDICTED E2F transcriptionfactor-like E2FE-like FANhyb icon00002702 a1g000011 278 Down 240119864 minus 04
PREDICTED uncharacterized protein FANhyb rscf000010661g000011 263 Down 490119864 minus 06PREDICTED nuclear transcriptionfactor Y subunit A-3-like isoform 1 FANhyb rscf000000081g000351 385 Down 500119864 minus 20
PREDICTED homeobox-leucinezipper protein ATHB-8-like FANhyb rscf000000151g000051 263 Down 100119864 minus 06
PREDICTED MADS-boxtranscription factor 18-like FANhyb rscf000003231g000161 243 Down 180119864 minus 06
PREDICTED zinc finger protein132-like isoform 1 FANhyb rscf000070151g000011 243 Down 180119864 minus 08
PREDICTED transcription factorPIF3-like FANhyb rscf000006691g000021 346 Up 140119864 minus 07
10 International Journal of Genomics
Table 4 Regulation of the DEGs in anthocyanin biosynthesis in response to exogenous hematin
Genes in anthocyaninbiosynthesis Gene ID
Realfold-change
valuesRegulation level FDR
Dihydroflavonol-4-reductase FANhyb icon00002410 a1g000011 3091 Up 910119864 minus 06
Flavonol synthase FANhyb icon00015354 a1g000011 345 down 180119864 minus 05Isoflavone reductasehomolog FANhyb icon20341135 s1g000011 334 Up 750119864 minus 14
UDP-glycosyltransferase FANhyb rscf000049691g000011 307 Up 640119864 minus 08UDP-glycosyltransferaseactivity FANhyb icon15742070 o1g000011 18 30563 Up 510119864 minus 09
UDP-glycosyltransferase73C1 FANhyb icon00027696 a1g000011 298 Up 560119864 minus 04
UDP-glycosyltransferaseactivity FANhyb icon00008064 a1g000011 249 Up 310119864 minus 04
PREDICTEDUDP-glucose flavonoid3-O-glucosyltransferase7-like
FANhyb rscf000055631g000011 212 Up 110119864 minus 19
PREDICTEDUDP-glycosyltransferase76F1-like
FANhyb rscf000021771g000031 2 Up 720119864 minus 29
A transcripts in the etiolated leaves of wheat seedling phy-tochromes [26] However our study shows that phytochromeA was not differentially expressed It is probably becausethe hematin-treated strawberries were not shaded during theexperiment while the function of phytochrome A is light-dependent In addition we found that a downstream gene ofphytochrome PIF3 (FANhyb rscf000006691g000021) wassignificantly upregulated PIF3 is thought to be a positive reg-ulator of phytochrome B mediated by light-dependent signaltransduction [58]These results suggest that the phytochromepathways may be involved in the anthocyanin biosynthesispromoted by hematin
345 DEGs in Starch and Sucrose Degradation Proteomicapproaches have revealed that starch degradation contributesto anthocyanin accumulation in tuberous roots of the purplesweet potato variety [59] Starch and sucrose biosynthesis isimportant for anthocyanin accumulation in strawberry fruit[60] In this study we found the DEGs involved in starchand sucrose synthesis in the hematin-treated fruit were alldownregulated (Table 5) For example the expression of theunigenes encoding soluble starch synthase 3 sucrose trans-port protein SUC2-like and a probable sucrose-phosphatesynthase 4-like enzyme were decreased 23- 46- 32- and2-fold respectively This suggests that exogenous hematinregulates the biosynthesis of starch and sucrose and henceaffects fruit coloring
346 DEGs in the Calcium Pathway in Response to HematinCalcium can also increase the transcription levels of keystructural genes F3H DFR ANT and UFGT in the whitestage of strawberries [61] In this study we found thatthree DEGs in calcium biosynthesis and transport were
downregulated in the hematin-treated fruit (Table 5) Onlythe unigene encoding predicted cationcalcium exchanger5-like was upregulated The unigenes encoding a calpain-type cysteine protease and a predicted calcium-binding pro-tein PBP1-like were downregulated respectively Exogenousapplication of calcium can promote apple coloring [62] inaddition to variation in anthocyanin content [61] Our resultsindicate that the biosynthesis and transport of calcium areinvolved in the development of coloring in hematin-treatedstrawberries
35 Validation of Selected DEGs by qRT-PCR To validate theexpression of the DEGs obtained from RNA-Seq 11 DEGswere selected for qRT-PCR including structural genes (DFRand UDFGs) transcription factor genes (MYB and bHLH)and phytochrome chromophore-related gene (HO-1) Theprimers used for qRT-PCR are listed in Supplementary TableS1 The qRT-PCR results were consistent with the RNA-Seqdata (Figure 4) except for FANhyb rscf000001411g000161which had a higher log
2ratio (hematin-treatedcontrol) in
the transcriptome data than in qRT-PCR These results indi-cate the RNA-Seq data from the strawberry transcriptome isreproducible and accurate
4 Conclusions
In this study the anthocyanin content in the strawberryfruit was elevated by the application of exogenous hematinThis is the first report that hematin could increase theanthocyanin production in fruit Furthermore we exploredthe effects of the exogenous hematin on metabolic path-ways using genome-wide transcriptome analysis The resultsindicate that the expression levels of many genes involved
International Journal of Genomics 11
Table 5 Other DEGs identified in anthocyanin biosynthesis-related pathways
Anthocyaninbiosynthesis-relatedpathways
Annotation genes Gene IDReal fold-changevalues
Upregulationdownregulation FDR
Calcium ion binding
Cationcalciumexchanger 5-like
FANhyb rscf000019061g000011 203 Up 64119864 minus 06
Calpain-type cysteineprotease family
FANhyb icon00011755 a1g000011 345 Down 18119864 minus 09
Calcium-bindingprotein PBP1-like
FANhyb rscf000007501g000081 323 Down 97119864 minus 04
Calpain-type cysteineprotease family
FANhyb icon00023658 a1g000011 233 Down 60119864 minus 08
CytokininCytokinin
dehydrogenase 5-likeFANhyb iscf00325393 1 s1g000011 667 Down 62119864 minus 06
Cytochrome P450714A1
FANhyb rscf000005921g000031 217 Down 98119864 minus 10
JasmonateJasmonate
O-methyltransferase-like
FANhyb rscf000000041g000131 201 Up 70119864 minus 04
Abscisic acid
Protein ABSCISICACID-INSENSITIVE
5-likeFANhyb rscf000021641g000011 4 Down 16119864 minus 18
ABA overly sensitive 5 FANhyb icon00051144 a1g000011 291 Up 24119864 minus 059-cis-
epoxycarotenoiddioxygenase NCED1
FANhyb icon18399909 o1g000011 441 Up 43119864 minus 06
Abscisic acid81015840-hydroxylase 1-like
FANhyb icon00000938 a1g000011 256 Down 14119864 minus 06
Abscisic acid receptorPYR1-like
FANhyb icon00020426 a1g000011 208 Down 30119864 minus 06
Auxin
Auxin efflux carriercomponent 1-like
FANhyb rscf000010081g000031 384 Down 23119864 minus 06
Auxin response factor8
FANhyb rscf000013061g000021 263 Down 61119864 minus 17
Auxin response factor17-like
FANhyb rscf000060741g000011 217 Down 11119864 minus 36
Starch and sucrose
Glycosyltransferasefamily 35
FANhyb rscf000000341g000081 384 Down 11119864 minus 05
Soluble starchsynthase 3
FANhyb rscf000000451g000041 232 Down 38119864 minus 12
Sucrose transportprotein SUC2-like
FANhyb icon00036727 a1g000011 455 Down 98119864 minus 04
Sucrose transportprotein SUC2-like
FANhyb rscf000007551g000021 323 Down 66119864 minus 10
Probablesucrose-phosphatesynthase 4-like
FANhyb rscf000013501g000021 213 Down 11119864 minus 14
Phytochrome
PREDICTEDtranscription factor
PIF3-likeFANhyb rscf000006691g000021 346 up 14119864 minus 07
Phytochrome E-like FANhyb rscf000004361g000041 4 Down 33119864 minus 05
Heme oxygenase 1 FANhyb icon00004395 a1g000011 258 27119864 minus 07 27119864 minus 07
12 International Journal of Genomics
Hematin
Endoplasmicreticulum
WD40 bLHLMYB
Structural genes
TFs
Nuclear
JA
ABA Sucrose
Plastid
G-cis-neoxanthin
Apophytochrome
StarchHeme
BVSucroseABA
Holophytochrome
PfrPIF3
Flavonoids Anthocyanins
Vacuole
Cytoplasm
Cell wall
①
②③
④⑤
⑥
⑦
PΦB
PΦB
CTKAuxin
Ca2+
Figure 5 Genes and biological pathways that were involved in regulation of anthocyanin accumulation promoted by hematin Outermostbox represents the cell wall Bilayer oval shape represents the plastidThe red-brown ring and network represent the nuclear and endoplasmicreticulum respectively The red crescent represents the vacuole The box in the nucleus represents transcription factors (TFs) Yellow MYBgreenWD40 and red bHLH represent a ternary transcription factor complex which transcribes anthocyanin biosynthesis genes The circlednumbers represent biological pathways involved in the regulation of anthocyanin accumulation promoted by hematin A represents thephytochrome regulation pathwayB represents the starch and sucrose pathwayCDE andF represent phytohormones ABA JA AuxinandCTK regulation pathways respectivelyG represents theCa2+ regulation pathway Pfr represents the far-red phytochrome PIF3 representsphytochrome-interacting factor Each anthocyanin biosynthesis regulatory pathway related gene is in Table 5
in anthocyanin biosynthesis were significantly altered withthe hematin treatment This suggests that the physiologicalprocess of fruit color development is regulated through com-plex interactions among anthocyanin biosynthesis pathwaysplant hormone signal transduction pathways phytochromesignal transduction pathways starch and sugar metabolicpathways calcium pathways and transcription factors (Fig-ure 5) This study adds to the in-depth understanding of thefruit coloration process in strawberry
Competing Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contributions
Jianwei Gao and Liangju Wang conceived and designed theresearch Yi Li Huayin Li Fengde Wang Jingjuan Li andYihui Zhang conducted experiments Yi Li analyzed the dataYi Li and Fengde Wang wrote the manuscript All authorsread and approved the manuscript Yi Li and Huayin Licontributed equally to this work
Acknowledgments
This work was supported by the Modern Agricultural Indus-trial Technology System Funding of Shandong ProvinceChina (SDAIT-02-022-04) the China agriculture researchsystem (CARS-25) and the National Science Foundation ofChina (31401820)
References
[1] K Aaby SMazur A Nes andG Skrede ldquoPhenolic compoundsin strawberry (Fragaria x ananassa Duch) fruits compositionin 27 cultivars and changes during ripeningrdquo Food Chemistryvol 132 no 1 pp 86ndash97 2012
[2] T A Colquhoun L A Levin H RMoskowitz VMWhitakerD G Clark and K M Folta ldquoFraming the perfect strawberryan exercise in consumer-assisted selection of fruit cropsrdquoJournal of Berry Research vol 2 no 1 pp 45ndash61 2012
[3] F Giampieri J M Alvarez-Suarez L Mazzoni et al ldquoThepotential impact of strawberry on human healthrdquo NaturalProduct Research vol 27 no 4 pp 448ndash455 2013
[4] Y S Velioglu GMazza L Gao and B D Oomah ldquoAntioxidantactivity and total phenolics in selected fruits vegetables and
International Journal of Genomics 13
grain productsrdquo Journal of Agricultural amp Food Chemistry vol46 no 10 pp 4113ndash4117 1998
[5] S S Hassellund A Flaa S E Kjeldsen et al ldquoEffects of antho-cyanins on cardiovascular risk factors and inflammation inpre-hypertensive men a double-blind randomized placebo-controlled crossover studyrdquo Journal of Human Hypertensionvol 27 no 2 pp 100ndash106 2013
[6] K J Meyers C B Watkins M P Pritts and R H Liu ldquoAnti-oxidant and antiproliferative activities of strawberriesrdquo Journalof Agricultural amp Food Chemistry vol 51 no 23 pp 6887ndash68922003
[7] S Zafra-Stone T YasminM Bagchi AChatterjee J AVinsonand D Bagchi ldquoBerry anthocyanins as novel antioxidants inhuman health and disease preventionrdquo Molecular Nutrition ampFood Research vol 51 no 6 pp 675ndash683 2007
[8] L Jaakola ldquoNew insights into the regulation of anthocyanin bio-synthesis in fruitsrdquoTrends in Plant Science vol 18 no 9 pp 477ndash483 2013
[9] K Springob J-I Nakajima M Yamazaki and K Saito ldquoRecentadvances in the biosynthesis and accumulation of antho-cyaninsrdquo Natural Product Reports vol 20 no 3 pp 288ndash3032003
[10] A Gonzalez M Zhao J M Leavitt and A M Lloyd ldquoRegu-lation of the anthocyanin biosynthetic pathway by the TTG1bHLHMyb transcriptional complex in Arabidopsis seedlingsrdquoPlant Journal vol 53 no 5 pp 814ndash827 2008
[11] K Petroni and C Tonelli ldquoRecent advances on the regulationof anthocyanin synthesis in reproductive organsrdquo Plant Sciencevol 181 no 3 pp 219ndash229 2011
[12] H-F Jia Y-M Chai C-L Li et al ldquoAbscisic acid plays animportant role in the regulation of strawberry fruit ripeningrdquoPlant Physiology vol 157 no 1 pp 188ndash199 2011
[13] C Merchante J G Vallarino S Osorio et al ldquoEthylene isinvolved in strawberry fruit ripening in an organ-specific man-nerrdquo Journal of Experimental Botany vol 64 no 14 pp 4421ndash4439 2013
[14] A Maier A Schrader L Kokkelink et al ldquoLight and the E3ubiquitin ligase COP1SPA control the protein stability of theMYB transcription factors PAP1 and PAP2 involved in antho-cyanin accumulation in Arabidopsisrdquo Plant Journal vol 74 no4 pp 638ndash651 2013
[15] H Lange W Shropshire and H Mohr ldquoAn analysis of phyto-chrome-mediated anthocyanin synthesisrdquo Plant Physiology vol47 no 5 pp 649ndash655 1971
[16] T D Elich and J C Lagarias ldquoPhytochrome chromophore bio-synthesis both 5-aminolevulinic acid and biliverdin overcomeinhibition by gabaculine in etiolated Avena sativa L seedlingsrdquoPlant Physiology vol 84 no 2 pp 304ndash310 1987
[17] Y Han W Xuan T Yu et al ldquoExogenous hematin alleviatesmercury-induced oxidative damage in the roots of Medicagosativardquo Journal of Integrative Plant Biology vol 49 no 12 pp1703ndash1713 2007
[18] M Koornneef J Cone R Dekens E OrsquoHerne-Robers CSpruit and R Kendrick ldquoPhotomorphogenic responses of longhypocotyl mutants of tomatordquo Journal of Plant Physiology vol120 no 2 pp 153ndash165 1985
[19] M J Terry and R E Kendrick ldquoFeedback inhibition of chloro-phyll synthesis in the phytochrome chromophore-deficientaurea and yellow-green-2 mutants of tomatordquo Plant Physiologyvol 119 no 1 pp 143ndash152 1999
[20] B M Parks and P H Quail ldquoPhytochrome-deficient hy1 andhy2 long hypocotyl mutants of Arabidopsis are defective inphytochrome chromophore biosynthesisrdquo Plant Cell vol 3 no11 pp 1177ndash1186 1991
[21] M J Terry ldquoPhytochrome chromophore-deficient mutantsrdquoPlant Cell amp Environment vol 20 no 6 pp 740ndash745 1997
[22] T Muramoto T Kohchi A Yokota I Hwang and H MGoodman ldquoThe Arabidopsis photomorphogenic mutant hy1 isdeficient in phytochrome chromophore biosynthesis as a resultof a mutation in a plastid heme oxygenaserdquo Plant Cell vol 11no 3 pp 335ndash347 1999
[23] M D Maines ldquoThe heme oxygenase system a regulator ofsecond messenger gasesrdquo Annual Review of Pharmacology andToxicology vol 37 no 1 pp 517ndash554 1997
[24] W Xuan F-Y Zhu S Xu et al ldquoThe heme oxygenasecarbonmonoxide system is involved in the auxin-induced cucumberadventitious rooting processrdquo Plant Physiology vol 148 no 2pp 881ndash893 2008
[25] R Amooaghaie F Tabatabaei and A-M Ahadi ldquoRole ofhematin and sodium nitroprusside in regulating Brassica nigraseed germination under nanosilver and silver nitrate stressesrdquoEcotoxicology and Environmental Safety vol 113 pp 259ndash2702015
[26] Y Liu X Li L Xu and W Shen ldquoDe-etiolation of wheatseedling leaves cross talk between heme oxygenasecarbonmonoxide and nitric oxiderdquo PLoS ONE vol 8 no 12 ArticleID e81470 2013
[27] T Muramoto N Tsurui M J Terry A Yokota and T KohchildquoExpression and biochemical properties of a ferredoxin-dependent heme oxygenase required for phytochrome chro-mophore synthesisrdquo Plant Physiology vol 130 no 4 pp 1958ndash1966 2002
[28] J-W Gao J Liu B Li and Z Li ldquoIsolation and purificationof functional total RNA from blue-grained wheat endospermtissues containing high levels of starches and flavonoidsrdquo PlantMolecular Biology Reporter vol 19 no 2 pp 185ndash186 2001
[29] R Patro S M Mount and C Kingsford ldquoSailfish enablesalignment-free isoform quantification from RNA-seq readsusing lightweight algorithmsrdquoNature Biotechnology vol 32 no5 pp 462ndash464 2014
[30] Y-X Qi Y-B Liu and W-H Rong ldquoRNA-Seq and its applica-tions a new technology for transcriptomicsrdquo Hereditas vol 33no 11 pp 1191ndash1202 2011
[31] C R Genovese N A Lazar and T Nichols ldquoThresholdingof statistical maps in functional neuroimaging using the falsediscovery raterdquo NeuroImage vol 15 no 4 pp 870ndash878 2002
[32] J Ye L Fang H Zheng et al ldquoWEGO a web tool for plottingGO annotationsrdquoNucleic Acids Research vol 34 supplement 2pp W293ndashW297 2006
[33] H Abdi ldquoThe Bonferonni and Sidak corrections for multiplecomparisonrdquo Encyclopedia of Measurement and Statistics vol 3pp 103ndash107 2007
[34] J Li Q Ding F Wang Y Zhang H Li and J Gao ldquoIntegrativeanalysis of mRNA and miRNA expression profiles of thetuberous root development at seedling stages in turnipsrdquo PLoSONE vol 10 no 9 Article ID e0137983 2015
[35] J Bennett D Hondred and J C Register ldquoKeeping qRT-PCRrigorous and biologically relevantrdquo Plant Cell Reports vol 34no 1 pp 1ndash3 2015
[36] F Wang L Li H Li et al ldquoTranscriptome analysis of rosetteand folding leaves in Chinese cabbage using high-throughputRNA sequencingrdquo Genomics vol 99 no 5 pp 299ndash307 2012
14 International Journal of Genomics
[37] T Shi Z Gao L Wang et al ldquoIdentification of differentially-expressed genes associated with pistil abortion in Japaneseapricot by genome-wide transcriptional analysisrdquo PLoS ONEvol 7 no 10 Article ID e47810 2012
[38] F Wang H Li Y Zhang et al ldquoMicroRNA expression analysisof rosette and folding leaves in Chinese cabbage using high-throughput Solexa sequencingrdquo Gene vol 532 no 2 pp 222ndash229 2013
[39] X Gu Y Chen Z Gao Y Qiao and X Wang ldquoTranscriptionfactors and anthocyanin genes related to low-temperature toler-ance in rd29A RdreB1BI transgenic strawberryrdquo Plant Physiol-ogy and Biochemistry vol 89 pp 31ndash43 2015
[40] J G Schaart C Dubos I R De La Fuente et al ldquoIdentificationand characterization of MYB-bHLH-WD40 regulatory com-plexes controlling proanthocyanidin biosynthesis in strawberry(Fragaria times ananassa) fruitsrdquoNew Phytologist vol 197 no 2 pp454ndash467 2013
[41] L Xie Z H Wang X H Cheng J J Gao Z P Zhangand L J Wang ldquo5-Aminolevulinic acid promotes anthocyaninaccumulation in Fuji applesrdquo Plant Growth Regulation vol 69no 3 pp 295ndash303 2013
[42] M Daminato F Guzzo and G Casadoro ldquoASHATTERPROOF-like gene controls ripening in non-climac-teric strawberries and auxin and abscisic acid antagonisticallyaffect its expressionrdquo Journal of Experimental Botany vol 64no 12 pp 3775ndash3786 2013
[43] X-H Ji R Zhang N Wang L Yang and X-S Chen ldquoTran-scriptome profiling reveals auxin suppressed anthocyanin bio-synthesis in red-fleshed apple callus (Malus sieversii f niedzwet-zkyana)rdquo Plant Cell Tissue and Organ Culture vol 123 no 2pp 389ndash404 2015
[44] J Chen LMaoW Lu T Ying and Z Luo ldquoTranscriptome pro-filing of postharvest strawberry fruit in response to exogenousauxin and abscisic acidrdquo Planta vol 243 no 1 pp 183ndash197 2016
[45] G A Martınez A R Chaves and M C Anon ldquoEffect of gib-berellic acid on ripening of strawberry fruits (Fragaria annan-assa Duch)rdquo Journal of Plant Growth Regulation vol 13 no 2pp 87ndash91 1994
[46] P K Das D H Shin S-B Choi S-D Yoo G Choi and Y-IPark ldquoCytokinins enhance sugar-induced anthocyanin biosyn-thesis in Arabidopsisrdquo Molecules amp Cells vol 34 no 1 pp 93ndash101 2012
[47] P McAtee S Karim R Schaffer and K David ldquoA dynamicinterplay between phytohormones is required for fruit develop-ment maturation and ripeningrdquo Frontiers in Plant Science vol4 no 3 article 79 7 pages 2013
[48] E Nambara andAMarion-Poll ldquoAbscisic acid biosynthesis andcatabolismrdquoAnnual Review of Plant Biology vol 56 pp 165ndash1852005
[49] Y Jiang and D C Joyce ldquoABA effects on ethylene productionPAL activity anthocyanin and phenolic contents of strawberryfruitrdquo Plant Growth Regulation vol 39 no 2 pp 171ndash174 2003
[50] S Li ldquoTranscriptional control of flavonoid biosynthesis fine-tuning of the MYB-bHLH-WD40 (MBW) complexrdquo PlantSignaling amp Behavior vol 9 Article ID e27522 2014
[51] D R Rudell J K Fellman and J P Mattheis ldquoPreharvest appli-cation of methyl jasmonate to rsquoFujirsquo apples enhances red col-oration and affects fruit size splitting and bitter pit incidencerdquoHortScience vol 40 no 6 pp 1760ndash1762 2005
[52] A Belhadj N Telef C Saigne et al ldquoEffect of methyl jasmonatein combination with carbohydrates on gene expression of PR
proteins stilbene and anthocyanin accumulation in grapevinecell culturesrdquo Plant Physiology and Biochemistry vol 46 no 4pp 493ndash499 2008
[53] E Loreti G Povero G Novi C Solfanelli A Alpi and PPerata ldquoGibberellins jasmonate and abscisic acid modulate thesucrose-induced expression of anthocyanin biosynthetic genesin Arabidopsisrdquo New Phytologist vol 179 no 4 pp 1004ndash10162008
[54] H S Seo J T Song J-J Cheong et al ldquoJasmonic acid carboxylmethyltransferase A key enzyme for jasmonate-regulated plantresponsesrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 98 no 8 pp 4788ndash4793 2001
[55] M T McDowell and J C Lagarias ldquoPurification and biochemi-cal properties of phytochromobilin synthase from etiolated oatseedlingsrdquo Plant Physiology vol 126 no 4 pp 1546ndash1554 2001
[56] G-Q Fu S Xu Y-J Xie et al ldquoMolecular cloning characteri-zation and expression of an alfalfa (Medicago sativa L) hemeoxygenase-1 gene MsHO1 which is pro-oxidants-regulatedrdquoPlant Physiology and Biochemistry vol 49 no 7 pp 792ndash7992011
[57] X-Y Chen X Ding S Xu et al ldquoEndogenous hydrogen per-oxide plays a positive role in the upregulation of heme oxy-genase and acclimation to oxidative stress in wheat seedlingleavesrdquo Journal of Integrative Plant Biology vol 51 no 10 pp951ndash960 2009
[58] J Shin E Park and G Choi ldquoPIF3 regulates anthocyaninbiosynthesis in an HY5-dependent manner with both factorsdirectly binding anthocyanin biosynthetic gene promoters inArabidopsisrdquo Plant Journal vol 49 no 6 pp 981ndash994 2007
[59] S Wang D Pan X Lv et al ldquoProteomic approach reveals thatstarch degradation contributes to anthocyanin accumulation intuberous root of purple sweet potatordquo Journal of Proteomics vol143 pp 298ndash305 2016
[60] H-F Jia C-L Li Y-M Chai Y Xing and Y Shen ldquoSucrosepromotes strawberry fruit ripening by stimulation of abscisicacid biosynthesisrdquo Pakistan Journal of Botany vol 45 no 1 pp169ndash176 2013
[61] W Xu H Peng T Yang et al ldquoEffect of calcium on strawberryfruit flavonoid pathway gene expression and anthocyanin accu-mulationrdquo Plant Physiology amp Biochemistry vol 82 pp 289ndash298 2014
[62] Z-W-SWan and Z Singh ldquoExogenous application of prohexa-dione-calcium promotes fruit colour development of lsquoCrippsPinkrsquo applerdquo Acta Horticulturae vol 1012 pp 219ndash225 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 International Journal of Genomics
26 Gene Ontology and Pathway Enrichment Analysis ofDifferentially Expressed Genes Gene functions of strawberrywere annotated according to the Gene Ontology (GO) stand-ardized terms for molecular function cellular componentand biological process using Blast2GO (httpswwwblast2gocom) Gene function was annotated using the followingdatabases NCBI nonredundant protein sequences NCBInonredundant nucleotide sequences protein family Clustersof Orthologous Groups of proteins the manually annotatedand reviewed protein sequence database KEGG Orthologdatabase and Gene Ontology After GO annotation forDEGs we performed GO functional classification for DEGsby the WEGO software [32] and analyzed the distributionof gene functions We used the GO Term Finder tool(httpwwwyeastgenomeorghelpanalyzego-term-finder)to search for significant shared GO terms
119901 = 1 minus119898minus1
sum119894=0
(119872119894) (119873minus119872119899minus119894)
(119873119899) (2)
where 119873 is the number of all genes with GO annotation119899 is the number of DEGs in 119873 119872 is the number of allgenes that are annotated to certain GO terms and 119898 is thenumber of DEGs in119872 The calculated 119901 value was subjectedto Bonferroni Correction [33] The corrected 119901 value lt 005was set as the threshold GO terms fulfilling this conditionare defined as significantly enriched GO terms in DEGsThe pathway enrichment analyses using the KEGG database(httpwwwgenomejpkegg) were conducted subsequentlyto study the functions of differentially expressed genes identi-fied between the control and the hematin-treated strawberrygroups The formula is the same as that in GO analysis Here119873 is the number of all genes with KEGG annotation 119899 is thenumber ofDEGs in119873119872 is the number of all genes annotatedto specific pathways and119898 is the number of DEGs in119872
27 Quantitative Real-Time PCR Analysis Quantitative real-time PCR (qRT-PCR) experiments were conducted to assessthe reliability of the RNA-Seq data [34] Eleven pairs of genespecific primers were designed to verify the DEGs in thestrawberry fruit (Table S1) Strawberry RNA was extractedfrom the fruit according to the previously detailed methodTotal RNA was digested with DNase I for 30min at 25∘C toremove DNA contamination according to the manufacturerrsquosinstructions The qRT-PCR was performed using a SYBRPCR master mix (TaKaRa Dalian China) on a Bio-Rad IQ-5 thermal cycler (Bio-Rad Philadelphia PA USA) Threereplicates of each sample were conducted to calculate theaverage Ct valuesThe relative expression level was calculatedby the comparative 2minusΔΔCt method [35 36] The significancewas determined with the SPSS software (SPSS 170 IBMChicago IL USA) (119901 lt 005)
3 Results and Discussion
31 Effects of Exogenous Hematin on Anthocyanin andBiliverdin Accumulation Hematin is an inducer and sub-strate of HO1 in animals and plants [17 23] In this studystrawberries at theWt stage were treated with 0 120583M(control)
00
05
10
15
20
25
ControlHematin treatment
B
A
5120583M hematin
Biliv
erdi
n co
nten
t (120583
Mmiddotg
minus1middotF
W)
Figure 2 The BV content in strawberry fruit was measured aftertreatment with 0 and 5 120583Mhematin Vertical bars represent standarderrors values with different letter are significantly different at 119901 lt001
5 10 or 15 120583M hematin The results indicated that the antho-cyanin content in the hematin-treated strawberries was morethan the control after 48 h The treatment by 5120583M hematinwas found to be the optimal condition for increasing antho-cyanin production (Figure 1) (119901 lt 001) The anthocyanincontent in the 5 120583M hematin-treated strawberries was 25times higher than that in the control This is the first reportthat hematin could increase the anthocyanin production infruit Wemeasured the expression of FaHO-1 and the contentof biliverdin (BV) which is themetabolite of heme oxygenaseWe found that hematin could also significantly increase theexpression of FaHO-1 in strawberry fruit (Figure 4) andpromote the accumulation of BV (Figure 2) These resultsindicate that hematin promotes anthocyanin accumulationthrough the heme metabolism pathway
32 Sequences Assembly Mapping and Functional Anno-tation High-throughput sequencing technology is utilizedwidely in analyzing gene expression in many organisms[37 38] In this study approximately 108 Gbp raw tagswere generated for each library After eliminating adaptersambiguous nucleotides and low-quality sequences a total of34618832 and 45464123 clean reads between 150 and 200nucleotides in length were obtained (Table 1) Over 85 ofthe clean tags from each library mapped to reference geneswith less than 2 bp mismatches Less than 29 of the cleantags from each library could not be aligned to any referencegenes because of incomplete sequences and these tags weredesignated as unknown More than 673 of the clean tagsin each library were mapped to a single gene while less than47mapped to multiple reference genes Unknown tags andtags mapped to multigenes were filtered out and the unique
International Journal of Genomics 5
Table 1 Overview of the sequencing and assembly
Biological replicate 1 Biological replicate 2Sample Control Hematin-treated Control Hematin-treatedClean reads 17808924 26809908 19235452 26228671Genome map rate () 9658 9652 9665 9674Gene map rate () 8646 8627 8704 8563Perfect match () 4677273 (2432) 6510668 (2428) 4414050 (2479) 6149508 (2345)Unique match () 15075148 (8465) 22628939 (8441) 16409557 (8531) 21986962 (8383)Expressed genes 27976 28713 28999 28211Mismatch () 11321162 (6357) 17101590 (6379) 12456871 (6476) 16697280 (6366)Total unmapped reads 645135 (335) 932213 (348) 608914 (342) 856028 (326)
clean tags that mapped to a single gene were retained forfurther DEG analysis
In addition the perfect match rates were 2432 24282479 and 2435 in control 1 control 2 hematin 1 andhematin 2 respectively
The saturation of the libraries with and without thehematin treatment was analyzedThe number of the detectedgenes became saturated at about 2 million reads (FigureS2) This indicates that the obtained reads are sufficient forcomplete transcriptome coverage The randomness of RNAfragmentation in the four libraries was assessed for subse-quent bioinformatics analysis The results showed that thereads in each position of the reference gene were distributedevenly and demonstrated highly similar tendencies in alllibraries (Figure S3)
33 Genes Differentially Regulated in Response to ExogenousHematin Based on sequencing results of the four mRNAlibraries approximately 29000 genes (about 70 of thereference genes in the octoploid strawberry genome) weredetected in each library Additionally correlations amonggenes based on RPKM between the two biological replicateswere analyzed the correlation coefficients (1198772) were high(093 and 095 for the control and hematin-treated groupresp) Genes expressed in both replicates were screened Atotal of 28713 28999 27976 and 28211 genes (Table 1) wereexpressed in the control and hematin-treated samples
To reveal the molecular pathways regulating strawberrycoloring in response to hematin the DEGs between the con-trol and hematin-treated fruit were analyzed We comparedthe gene expression profiles between the control and thetreated samples in both biological replicates DEGs detectedin both biological replicates were screened for subsequentanalysis A total of 1080 (402 up- and 678 downregulated)genes were differentially expressed in the 5120583M hematin-treated groups comparedwith the control groupsMore geneswere downregulated in the hematin-treated group comparedto the control group
To facilitate the global analysis of gene expression aGO analysis was performed by mapping each differentiallyexpressed gene into the records of the GO database GeneOntology (GO) enrichment analysis of the DEGs in thecontrol and hematin-treated groups was performed to revealthe possible mechanisms under which hematin promotes
anthocyanin biosynthesis GO terms with 119901 lt 005 wererepresented among genes with significant changes in expres-sion over a given interval In this study only three termsincluding anatomical structure arrangementmeristem struc-tural organization radial pattern formationwere significantlyenriched in biological process (Supplementary Table S2)AlthoughmostGo termswere not significantly enriched theycould provide a reference for further study According to theGO cellular components a large number of the DEGs wereclassified intomain cell organelles such as vacuole nucleolusmitochondrion apoplast chloroplast plastid cytoplasm andmembrane In particular many DEGs were assigned to theplastid and chloroplast (Figure 3) indicating that hematinhas an important impact on the expression of plastid andchloroplast genes It is probably because heme oxygenase 1 isa soluble plastid protein [22] According to the GO molec-ular function a large number of the DEGs were classifiedintoUDP-glucosyltransferase sucrose transmembrane trans-porter activity nucleic acid binding and DNA binding termswhich are closely related to anthocyanin biosynthesis ManyDEGs were classified into heme binding peroxidase activityand tetrapyrrole binding (Figure 3) It is probably becausehematin can increase the activity of HO1 and promote thedegradation of hemoglobin [17] In addition anthocyaninaccumulation in fruit is closely related to hormone [8] andcarbohydrate biosynthesis [9] According to the GO biologi-cal process a large number of the DEGs were classified intoregulation of hormone levels cytokinin metabolic processresponse to hormone response to abscisic acid response toauxin and response to ethylene terms indicating that thehormone-related DEGs are involved in regulating the antho-cyanin biosynthesis in the hematin-treated fruit Many DEGswere classified into starch and sucrose biosynthetic and trans-port process and phenylpropanoid metabolic process termsindicating that hematin can promote anthocyanin accumu-lation via the phenylpropanoid metabolic and carbohydratemetabolism In addition many DEGs were classified into theprocess for response to light stimulus term indicating thathematin may also participate in the light stimulus system
Pathway enrichment analysis was performed to under-stand the biological functions of the DEGs by identify-ing significantly enriched signal transduction pathways ormetabolic pathways [39] In the study a number of alteredbiological pathways associated with the hematin treatment
6 International Journal of Genomics
0 10 20 30 40 50 60 70 80Starch biosynthetic process
Cytokinin metabolic processResponse to abscisic acid
Response to auxinResponse to ethylene
Phenylpropanoid metabolic processSucrose transport
Regulation of hormone levelsResponse to light stimulus
Response to hormonePeroxidase activity
Sucrose transmembrane transporter activityUDP-glucosyltransferase
Heme bindingTetrapyrrole binding
DNA bindingNucleic acid binding
VacuoleNucleolusApoplast
MitochondrionChloroplast
PlastidCytoplasmMembrane
Biol
ogic
al p
rogr
ess
Mol
ecul
ar fu
nctio
nC
ellu
lar c
ompo
nent
Number of DEGs
Figure 3 Gene classification based on Gene Ontology (GO) for DEGs in the strawberries treated by hematin
were identified and three pathways including RNA poly-merase pyrimidine metabolism and purine metabolismwere significantly enriched (119876-value lt 005) (Table 2)Although the majority of the pathway terms were not sig-nificantly enriched the pathway enrichment analysis helpsus to further understand the biological functions of theDEGs and the molecular mechanisms that regulate fruitcoloring in response to the hematin treatmentThese pathwayterms included metabolic pathways biosynthesis of sec-ondary metabolites isoflavonoid biosynthesis phenylalaninemetabolism phenylalanine tyrosine and tryptophan biosyn-thesis starch and sucrose metabolism plant hormone signaltransduction flavone and flavonol biosynthesis and phenyl-propanoid biosynthesis terms (Supplementary Table S2)
34 Analysis of DEGs Involved in Anthocyanin Biosynthesis
341 Transcription Factors Transcription factors (TFs) areproteins that regulate the expression of downstream targetgenes Many TFs such as v-myb avian myeloblastosis viraloncogene homolog (MYB) basic helix-loop-helix (bHLH)WD40-repeats protein (WD40) and MADS-box (MADS)directly regulating the expression of the structural genes inanthocyanin biosynthesis have been identified from manyspecies [8] Therefore the enhanced expression of key TFs
might regulate the anthocyanin biosynthesis In our studysome anthocyanin biosynthesis-related TFs were found to beuniquely present in the DEG profiling of the hematin-treated fruit for example MYB bHLH PIF3 MADs-boxAP2-EREBP and ABI3VP1 (Table 3) The anthocyanin bio-synthesis genes are regulated primarily by the MBW tran-scription factor which is a ternary transcription factor com-plex [10 11] Most of the MYBs involved in regulating antho-cyanin biosynthesis are positive regulators of transcription[8] However MYBs can act as repressors too such as straw-berry FaMYB1 and FaMYB9 and grapevine VvMYB4 whichcan significantly suppress the biosynthesis of anthocyaninsand flavonols [40] In this study three unigenes belong-ing to MYB transcription factors were differentiallyexpressed and two MYB transcription factors were signifi-cantly upregulated Of these the unigene ldquoFANhybrscf000001461g000071rdquo from theMYB family of R2R3MYBtranscription factors was significantly upregulatedMoreoverincreasing evidence indicates that the expression of thebHLHs promotes anthocyanin accumulation in fruit [41] Inour study all of the unigenes from the bHLH family weredownregulated among which the expression level of unigeneFANhyb rscf000012921g000031 was significantly decreased6250-fold (Table 3) FANhyb rscf000022101g000011 en-codes a predicted MADS-box transcription factor and it was
International Journal of Genomics 7
Control Hematin-treated
FANhyb_icon00002410_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_rscf000049691g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00027696_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00004395_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_rscf000063641g000011
lowastlowast
lowast
Control Hematin-treated
FANhyb_rscf000001411g000161
Control Hematin-treated
FANhyb_icon00006295_a1g000011lowastlowast
Control Hematin-treated
FANhyb_rscf000031551g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00001444_a1g000011
lowastlowast
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
0
05
1
15
2
25
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
005
115
225
335
445
5
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
Control Hematin-treated
FANhyb_rscf000023111g000011
lowastlowast
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
Control Hematin-treated
FANhyb_rscf000002951g000031
lowastlowast
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
Figure 4 qRT-PCR validation of the RNA-Seq based gene expression The values indicate means of three biological replicates plusmn SD Starindicates that the expression level is significantly different between the hematin-treated and the control group (lowast119901 lt 005 lowastlowast119901 lt 001)
8 International Journal of Genomics
Table 2 Pathway enrichment analysis of differentially expressed genes
Pathway DEGs with pathwayannotation (451)
All genes with pathwayannotation (15877)
Corrected 119901 value 119876-value Pathway ID
RNA polymerase 16 183 688119864 minus 5 00068107 ko03020Pyrimidine metabolism 20 317 00007651746 00320 ko00240Purine metabolism 21 347 00009710371 0032044224 ko00230
significantly downregulated There were three unigenes inthe AP2-EREBP family with only one upregulated and theothers downregulated These results suggest that exogenoushematin is involved in developmental transcriptionalregulation of anthocyanin biosynthesis
342 DEGs in Anthocyanin Biosynthesis The anthocyaninbiosynthetic pathway has been studied intensively Theanthocyanin biosynthetic pathway via the phenylpropanoidpathway is well known [8] In this study the transcriptomedata showed a significant increase in the expression of genesin the anthocyanin biosynthesis pathway when exposedto exogenous hematin The structural genes involved inanthocyanin biosynthesis including DFR LAR and UDP-glycosyltransferase exhibited significant differential expres-sion in response to the hematin treatment (Table 4) Forexample two DFR genes and one LAR gene were signifi-cantly upregulated (the expression increased 18- 30- and3-fold in the hematin-treated fruit resp) In addition allsix genes encodingUDP-glycosyltransferasewere significantlyupregulated Among these the expression of one gene(Fanhy icon15742070o1g000011) was upregulated 18305-fold The results suggest that exogenous hematin may havean important role in promoting anthocyanin biosynthesis
343 DEGs in Hormone Signaling Transduction in Responseto Hematin Auxin has been shown to negatively regulatethe expression of the anthocyanin biosynthesis genes [42] Inthis study there were three unigenes involved in the auxinsignaling pathway and all were downregulated includingone gene encoding auxin efflux carrier component 1-like(AUX1-like) and two genes encoding auxin response factors(ARF) These results suggest that hematin promotes antho-cyanin accumulation by regulating auxin in the colorationprocess of strawberries It has shown that auxin suppressesanthocyanin biosynthesis in the red-fleshed apple callus [43]Endogenous expression of auxin [44 45] has been foundto impede anthocyanin accumulation in strawberries Inadditional cytokinins are also known to play an importantrole in anthocyanin biosynthesis It has shown to enhanceanthocyanin accumulation in Zea mays and regulate antho-cyanin production and composition in suspension culturesof strawberry cells [46] In this study two unigenes relatedto the cytokinin signaling pathway were identified and theywere downregulated as well One gene was identified ascytokinin dehydrogenase 5-like which catalyzes the irre-versible degradation of cytokinins This result indicates thathematin promotes anthocyanin accumulation by regulatingthe cytokinin signaling pathway
Extensive researches have shown that ABA plays animportant role in the regulation of anthocyanin biosynthesisin nonclimacteric fruit [47] In this study a total of five uni-genes related to the ABA signaling pathway were identifiedamong which two were upregulated and three were down-regulated Two genes which are involved in the abscisic acidresponse were upregulated These two unigenes encode 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) and ABA overlysensitive 5 (ABO5) respectively ABA is synthesized fromcarotenoids via several enzymatic reactions in the plastidThe rate-limiting step in these reactions is catalyzed by 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) [48] Mutations inthe FaNCED1 gene result in colorless strawberry fruit whichcan become colored through the application of exogenousABA [12] Another unigene was identified to encode abscisicacid 81015840-hydroxylase 1-like a key enzyme in the oxidativecatabolism of abscisic acid which was downregulated Appli-cation of exogenous ABA regulates phenylalanine ammonialyase activity and increases the phenolic and anthocyanincontent of strawberry fruit [12 49] Our results suggestthat hematin promotes ABA biosynthesis and inhibits ABAdisintegration in the hematin-treated fruit
In Arabidopsis JAs can affect anthocyanin accumulationvia the interaction of negative regulators with the MBWcomplex of transcription factors involved in anthocyaninbiosynthesis [50] Preharvest application of JA to ldquoFujirdquo applesenhances red coloration [51] JA vapor treatment can alsoenhance anthocyanins in strawberry fruit [52] In this studyjasmonate O-methyltransferase-like exhibited significantlydifferential expression Jasmonate O-methyltransferase-likeis also known to be a key enzyme for jasmonate-regulatedplant responses to stimuli [53 54] These results suggest thathematin increases the activity of the JA-regulated antho-cyanin biosynthesis
344 DEGs in the Phytochrome Signaling Pathway in Responseto Hematin Phytochromes that act as photoreceptors playan important role in anthocyanin regulation [15] Phy-tochromes are homodimeric chromoproteins where eachholophytochrome is composed of a phytochrome protein(apophytochrome) covalently bound to a linear tetrapyrrolePΦB HO1 is crucial to this process and acts as a rate-limiting enzyme in the biosynthesis of PΦB [55] Exogenoushematin can also induce HO-1 expression in many plants[56 57] In this study we found exogenous hematin signif-icantly increased the expression of FaHO1 (Table 5) Thisresult implies that hematin can promote PΦB biosynthesisRecently it has been reported that hematin could inducethe accumulation of far-red phytochrome and phytochrome
International Journal of Genomics 9
Table 3 DEGs acting as transcription factors in response to exogenous hematin
Annotation transcription factors Gene IDReal
fold-changevalues
Upregulationdownregulation FDR
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb icon00006295 a1g000011 938 Up 120119864 minus 05
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb rscf000031551g000011 4 Down 480119864 minus 16
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL5-like
FANhyb rscf000000291g000101 4 Down 320119864 minus 08
MYBdomain protein 66 FANhyb rscf000001461g000071 654 Up 780119864 minus 07MYB-related protein 3R-1-like partial FANhyb rscf000001411g000161 282 Up 830119864 minus 04PREDICTED transcription factorGAMYB-like FANhyb rscf000006491g000061 303 Down 150119864 minus 13
PREDICTED probable WRKYtranscription factor 53-like FANhyb rscf000019731g000021 217 Down 200119864 minus 08
Transcription factor bHLH80-like FANhyb rscf000012921g000031 6250 Down 150119864 minus 06Transcription factor bHLH70-like FANhyb rscf000002951g000031 526 Down 660119864 minus 12Transcription factor ICE1-like FANhyb rscf000001701g000061 244 Down 450119864 minus 18C2H2-like zinc finger protein FANhyb rscf000011431g000021 282 Up 590119864 minus 04Zinc finger protein MAGPIE-like FANhyb rscf000030241g000011 204 Up 150119864 minus 04PREDICTED axial regulator YABBY5-like FANhyb rscf000016671g000011 417 Down 730119864 minus 05
PREDICTED axial regulator YABBY1-like FANhyb rscf000068991g000011 357 Down 620119864 minus 06
PREDICTED dof zinc finger proteinDOF37-like isoform 1 FANhyb rscf000068561g000011 263 Down 280119864 minus 04
ZF-HD homeobox protein FANhyb rscf000000441g000221 385 Down 420119864 minus 08ZF-HD homeobox proteinAt4g24660-like FANhyb rscf000052411g000011 357 Down 470119864 minus 09
PREDICTED B3 domain-containingprotein REM14-like FANhyb rscf000010091g000021 243 Up 530119864 minus 04
PREDICTED B3 domain-containingtranscription factor ABI3-like FANhyb rscf000012711g000021 455 Down 130119864 minus 28
Growth-regulating factor 6 FANhyb rscf000000241g000291 345 Down 330119864 minus 34Growth-regulating factor 5 FANhyb icon00009704 a1g000011 27 Down 190119864 minus 06GATA type zinc finger transcriptionfactor family protein FANhyb rscf000003931g000131 1111 Down 110119864 minus 05
PREDICTED zinc finger CCCHdomain-containing protein 2-like FANhyb icon00006770 a1g000011 313 Down 680119864 minus 49
PREDICTED E2F transcriptionfactor-like E2FE-like FANhyb icon00002702 a1g000011 278 Down 240119864 minus 04
PREDICTED uncharacterized protein FANhyb rscf000010661g000011 263 Down 490119864 minus 06PREDICTED nuclear transcriptionfactor Y subunit A-3-like isoform 1 FANhyb rscf000000081g000351 385 Down 500119864 minus 20
PREDICTED homeobox-leucinezipper protein ATHB-8-like FANhyb rscf000000151g000051 263 Down 100119864 minus 06
PREDICTED MADS-boxtranscription factor 18-like FANhyb rscf000003231g000161 243 Down 180119864 minus 06
PREDICTED zinc finger protein132-like isoform 1 FANhyb rscf000070151g000011 243 Down 180119864 minus 08
PREDICTED transcription factorPIF3-like FANhyb rscf000006691g000021 346 Up 140119864 minus 07
10 International Journal of Genomics
Table 4 Regulation of the DEGs in anthocyanin biosynthesis in response to exogenous hematin
Genes in anthocyaninbiosynthesis Gene ID
Realfold-change
valuesRegulation level FDR
Dihydroflavonol-4-reductase FANhyb icon00002410 a1g000011 3091 Up 910119864 minus 06
Flavonol synthase FANhyb icon00015354 a1g000011 345 down 180119864 minus 05Isoflavone reductasehomolog FANhyb icon20341135 s1g000011 334 Up 750119864 minus 14
UDP-glycosyltransferase FANhyb rscf000049691g000011 307 Up 640119864 minus 08UDP-glycosyltransferaseactivity FANhyb icon15742070 o1g000011 18 30563 Up 510119864 minus 09
UDP-glycosyltransferase73C1 FANhyb icon00027696 a1g000011 298 Up 560119864 minus 04
UDP-glycosyltransferaseactivity FANhyb icon00008064 a1g000011 249 Up 310119864 minus 04
PREDICTEDUDP-glucose flavonoid3-O-glucosyltransferase7-like
FANhyb rscf000055631g000011 212 Up 110119864 minus 19
PREDICTEDUDP-glycosyltransferase76F1-like
FANhyb rscf000021771g000031 2 Up 720119864 minus 29
A transcripts in the etiolated leaves of wheat seedling phy-tochromes [26] However our study shows that phytochromeA was not differentially expressed It is probably becausethe hematin-treated strawberries were not shaded during theexperiment while the function of phytochrome A is light-dependent In addition we found that a downstream gene ofphytochrome PIF3 (FANhyb rscf000006691g000021) wassignificantly upregulated PIF3 is thought to be a positive reg-ulator of phytochrome B mediated by light-dependent signaltransduction [58]These results suggest that the phytochromepathways may be involved in the anthocyanin biosynthesispromoted by hematin
345 DEGs in Starch and Sucrose Degradation Proteomicapproaches have revealed that starch degradation contributesto anthocyanin accumulation in tuberous roots of the purplesweet potato variety [59] Starch and sucrose biosynthesis isimportant for anthocyanin accumulation in strawberry fruit[60] In this study we found the DEGs involved in starchand sucrose synthesis in the hematin-treated fruit were alldownregulated (Table 5) For example the expression of theunigenes encoding soluble starch synthase 3 sucrose trans-port protein SUC2-like and a probable sucrose-phosphatesynthase 4-like enzyme were decreased 23- 46- 32- and2-fold respectively This suggests that exogenous hematinregulates the biosynthesis of starch and sucrose and henceaffects fruit coloring
346 DEGs in the Calcium Pathway in Response to HematinCalcium can also increase the transcription levels of keystructural genes F3H DFR ANT and UFGT in the whitestage of strawberries [61] In this study we found thatthree DEGs in calcium biosynthesis and transport were
downregulated in the hematin-treated fruit (Table 5) Onlythe unigene encoding predicted cationcalcium exchanger5-like was upregulated The unigenes encoding a calpain-type cysteine protease and a predicted calcium-binding pro-tein PBP1-like were downregulated respectively Exogenousapplication of calcium can promote apple coloring [62] inaddition to variation in anthocyanin content [61] Our resultsindicate that the biosynthesis and transport of calcium areinvolved in the development of coloring in hematin-treatedstrawberries
35 Validation of Selected DEGs by qRT-PCR To validate theexpression of the DEGs obtained from RNA-Seq 11 DEGswere selected for qRT-PCR including structural genes (DFRand UDFGs) transcription factor genes (MYB and bHLH)and phytochrome chromophore-related gene (HO-1) Theprimers used for qRT-PCR are listed in Supplementary TableS1 The qRT-PCR results were consistent with the RNA-Seqdata (Figure 4) except for FANhyb rscf000001411g000161which had a higher log
2ratio (hematin-treatedcontrol) in
the transcriptome data than in qRT-PCR These results indi-cate the RNA-Seq data from the strawberry transcriptome isreproducible and accurate
4 Conclusions
In this study the anthocyanin content in the strawberryfruit was elevated by the application of exogenous hematinThis is the first report that hematin could increase theanthocyanin production in fruit Furthermore we exploredthe effects of the exogenous hematin on metabolic path-ways using genome-wide transcriptome analysis The resultsindicate that the expression levels of many genes involved
International Journal of Genomics 11
Table 5 Other DEGs identified in anthocyanin biosynthesis-related pathways
Anthocyaninbiosynthesis-relatedpathways
Annotation genes Gene IDReal fold-changevalues
Upregulationdownregulation FDR
Calcium ion binding
Cationcalciumexchanger 5-like
FANhyb rscf000019061g000011 203 Up 64119864 minus 06
Calpain-type cysteineprotease family
FANhyb icon00011755 a1g000011 345 Down 18119864 minus 09
Calcium-bindingprotein PBP1-like
FANhyb rscf000007501g000081 323 Down 97119864 minus 04
Calpain-type cysteineprotease family
FANhyb icon00023658 a1g000011 233 Down 60119864 minus 08
CytokininCytokinin
dehydrogenase 5-likeFANhyb iscf00325393 1 s1g000011 667 Down 62119864 minus 06
Cytochrome P450714A1
FANhyb rscf000005921g000031 217 Down 98119864 minus 10
JasmonateJasmonate
O-methyltransferase-like
FANhyb rscf000000041g000131 201 Up 70119864 minus 04
Abscisic acid
Protein ABSCISICACID-INSENSITIVE
5-likeFANhyb rscf000021641g000011 4 Down 16119864 minus 18
ABA overly sensitive 5 FANhyb icon00051144 a1g000011 291 Up 24119864 minus 059-cis-
epoxycarotenoiddioxygenase NCED1
FANhyb icon18399909 o1g000011 441 Up 43119864 minus 06
Abscisic acid81015840-hydroxylase 1-like
FANhyb icon00000938 a1g000011 256 Down 14119864 minus 06
Abscisic acid receptorPYR1-like
FANhyb icon00020426 a1g000011 208 Down 30119864 minus 06
Auxin
Auxin efflux carriercomponent 1-like
FANhyb rscf000010081g000031 384 Down 23119864 minus 06
Auxin response factor8
FANhyb rscf000013061g000021 263 Down 61119864 minus 17
Auxin response factor17-like
FANhyb rscf000060741g000011 217 Down 11119864 minus 36
Starch and sucrose
Glycosyltransferasefamily 35
FANhyb rscf000000341g000081 384 Down 11119864 minus 05
Soluble starchsynthase 3
FANhyb rscf000000451g000041 232 Down 38119864 minus 12
Sucrose transportprotein SUC2-like
FANhyb icon00036727 a1g000011 455 Down 98119864 minus 04
Sucrose transportprotein SUC2-like
FANhyb rscf000007551g000021 323 Down 66119864 minus 10
Probablesucrose-phosphatesynthase 4-like
FANhyb rscf000013501g000021 213 Down 11119864 minus 14
Phytochrome
PREDICTEDtranscription factor
PIF3-likeFANhyb rscf000006691g000021 346 up 14119864 minus 07
Phytochrome E-like FANhyb rscf000004361g000041 4 Down 33119864 minus 05
Heme oxygenase 1 FANhyb icon00004395 a1g000011 258 27119864 minus 07 27119864 minus 07
12 International Journal of Genomics
Hematin
Endoplasmicreticulum
WD40 bLHLMYB
Structural genes
TFs
Nuclear
JA
ABA Sucrose
Plastid
G-cis-neoxanthin
Apophytochrome
StarchHeme
BVSucroseABA
Holophytochrome
PfrPIF3
Flavonoids Anthocyanins
Vacuole
Cytoplasm
Cell wall
①
②③
④⑤
⑥
⑦
PΦB
PΦB
CTKAuxin
Ca2+
Figure 5 Genes and biological pathways that were involved in regulation of anthocyanin accumulation promoted by hematin Outermostbox represents the cell wall Bilayer oval shape represents the plastidThe red-brown ring and network represent the nuclear and endoplasmicreticulum respectively The red crescent represents the vacuole The box in the nucleus represents transcription factors (TFs) Yellow MYBgreenWD40 and red bHLH represent a ternary transcription factor complex which transcribes anthocyanin biosynthesis genes The circlednumbers represent biological pathways involved in the regulation of anthocyanin accumulation promoted by hematin A represents thephytochrome regulation pathwayB represents the starch and sucrose pathwayCDE andF represent phytohormones ABA JA AuxinandCTK regulation pathways respectivelyG represents theCa2+ regulation pathway Pfr represents the far-red phytochrome PIF3 representsphytochrome-interacting factor Each anthocyanin biosynthesis regulatory pathway related gene is in Table 5
in anthocyanin biosynthesis were significantly altered withthe hematin treatment This suggests that the physiologicalprocess of fruit color development is regulated through com-plex interactions among anthocyanin biosynthesis pathwaysplant hormone signal transduction pathways phytochromesignal transduction pathways starch and sugar metabolicpathways calcium pathways and transcription factors (Fig-ure 5) This study adds to the in-depth understanding of thefruit coloration process in strawberry
Competing Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contributions
Jianwei Gao and Liangju Wang conceived and designed theresearch Yi Li Huayin Li Fengde Wang Jingjuan Li andYihui Zhang conducted experiments Yi Li analyzed the dataYi Li and Fengde Wang wrote the manuscript All authorsread and approved the manuscript Yi Li and Huayin Licontributed equally to this work
Acknowledgments
This work was supported by the Modern Agricultural Indus-trial Technology System Funding of Shandong ProvinceChina (SDAIT-02-022-04) the China agriculture researchsystem (CARS-25) and the National Science Foundation ofChina (31401820)
References
[1] K Aaby SMazur A Nes andG Skrede ldquoPhenolic compoundsin strawberry (Fragaria x ananassa Duch) fruits compositionin 27 cultivars and changes during ripeningrdquo Food Chemistryvol 132 no 1 pp 86ndash97 2012
[2] T A Colquhoun L A Levin H RMoskowitz VMWhitakerD G Clark and K M Folta ldquoFraming the perfect strawberryan exercise in consumer-assisted selection of fruit cropsrdquoJournal of Berry Research vol 2 no 1 pp 45ndash61 2012
[3] F Giampieri J M Alvarez-Suarez L Mazzoni et al ldquoThepotential impact of strawberry on human healthrdquo NaturalProduct Research vol 27 no 4 pp 448ndash455 2013
[4] Y S Velioglu GMazza L Gao and B D Oomah ldquoAntioxidantactivity and total phenolics in selected fruits vegetables and
International Journal of Genomics 13
grain productsrdquo Journal of Agricultural amp Food Chemistry vol46 no 10 pp 4113ndash4117 1998
[5] S S Hassellund A Flaa S E Kjeldsen et al ldquoEffects of antho-cyanins on cardiovascular risk factors and inflammation inpre-hypertensive men a double-blind randomized placebo-controlled crossover studyrdquo Journal of Human Hypertensionvol 27 no 2 pp 100ndash106 2013
[6] K J Meyers C B Watkins M P Pritts and R H Liu ldquoAnti-oxidant and antiproliferative activities of strawberriesrdquo Journalof Agricultural amp Food Chemistry vol 51 no 23 pp 6887ndash68922003
[7] S Zafra-Stone T YasminM Bagchi AChatterjee J AVinsonand D Bagchi ldquoBerry anthocyanins as novel antioxidants inhuman health and disease preventionrdquo Molecular Nutrition ampFood Research vol 51 no 6 pp 675ndash683 2007
[8] L Jaakola ldquoNew insights into the regulation of anthocyanin bio-synthesis in fruitsrdquoTrends in Plant Science vol 18 no 9 pp 477ndash483 2013
[9] K Springob J-I Nakajima M Yamazaki and K Saito ldquoRecentadvances in the biosynthesis and accumulation of antho-cyaninsrdquo Natural Product Reports vol 20 no 3 pp 288ndash3032003
[10] A Gonzalez M Zhao J M Leavitt and A M Lloyd ldquoRegu-lation of the anthocyanin biosynthetic pathway by the TTG1bHLHMyb transcriptional complex in Arabidopsis seedlingsrdquoPlant Journal vol 53 no 5 pp 814ndash827 2008
[11] K Petroni and C Tonelli ldquoRecent advances on the regulationof anthocyanin synthesis in reproductive organsrdquo Plant Sciencevol 181 no 3 pp 219ndash229 2011
[12] H-F Jia Y-M Chai C-L Li et al ldquoAbscisic acid plays animportant role in the regulation of strawberry fruit ripeningrdquoPlant Physiology vol 157 no 1 pp 188ndash199 2011
[13] C Merchante J G Vallarino S Osorio et al ldquoEthylene isinvolved in strawberry fruit ripening in an organ-specific man-nerrdquo Journal of Experimental Botany vol 64 no 14 pp 4421ndash4439 2013
[14] A Maier A Schrader L Kokkelink et al ldquoLight and the E3ubiquitin ligase COP1SPA control the protein stability of theMYB transcription factors PAP1 and PAP2 involved in antho-cyanin accumulation in Arabidopsisrdquo Plant Journal vol 74 no4 pp 638ndash651 2013
[15] H Lange W Shropshire and H Mohr ldquoAn analysis of phyto-chrome-mediated anthocyanin synthesisrdquo Plant Physiology vol47 no 5 pp 649ndash655 1971
[16] T D Elich and J C Lagarias ldquoPhytochrome chromophore bio-synthesis both 5-aminolevulinic acid and biliverdin overcomeinhibition by gabaculine in etiolated Avena sativa L seedlingsrdquoPlant Physiology vol 84 no 2 pp 304ndash310 1987
[17] Y Han W Xuan T Yu et al ldquoExogenous hematin alleviatesmercury-induced oxidative damage in the roots of Medicagosativardquo Journal of Integrative Plant Biology vol 49 no 12 pp1703ndash1713 2007
[18] M Koornneef J Cone R Dekens E OrsquoHerne-Robers CSpruit and R Kendrick ldquoPhotomorphogenic responses of longhypocotyl mutants of tomatordquo Journal of Plant Physiology vol120 no 2 pp 153ndash165 1985
[19] M J Terry and R E Kendrick ldquoFeedback inhibition of chloro-phyll synthesis in the phytochrome chromophore-deficientaurea and yellow-green-2 mutants of tomatordquo Plant Physiologyvol 119 no 1 pp 143ndash152 1999
[20] B M Parks and P H Quail ldquoPhytochrome-deficient hy1 andhy2 long hypocotyl mutants of Arabidopsis are defective inphytochrome chromophore biosynthesisrdquo Plant Cell vol 3 no11 pp 1177ndash1186 1991
[21] M J Terry ldquoPhytochrome chromophore-deficient mutantsrdquoPlant Cell amp Environment vol 20 no 6 pp 740ndash745 1997
[22] T Muramoto T Kohchi A Yokota I Hwang and H MGoodman ldquoThe Arabidopsis photomorphogenic mutant hy1 isdeficient in phytochrome chromophore biosynthesis as a resultof a mutation in a plastid heme oxygenaserdquo Plant Cell vol 11no 3 pp 335ndash347 1999
[23] M D Maines ldquoThe heme oxygenase system a regulator ofsecond messenger gasesrdquo Annual Review of Pharmacology andToxicology vol 37 no 1 pp 517ndash554 1997
[24] W Xuan F-Y Zhu S Xu et al ldquoThe heme oxygenasecarbonmonoxide system is involved in the auxin-induced cucumberadventitious rooting processrdquo Plant Physiology vol 148 no 2pp 881ndash893 2008
[25] R Amooaghaie F Tabatabaei and A-M Ahadi ldquoRole ofhematin and sodium nitroprusside in regulating Brassica nigraseed germination under nanosilver and silver nitrate stressesrdquoEcotoxicology and Environmental Safety vol 113 pp 259ndash2702015
[26] Y Liu X Li L Xu and W Shen ldquoDe-etiolation of wheatseedling leaves cross talk between heme oxygenasecarbonmonoxide and nitric oxiderdquo PLoS ONE vol 8 no 12 ArticleID e81470 2013
[27] T Muramoto N Tsurui M J Terry A Yokota and T KohchildquoExpression and biochemical properties of a ferredoxin-dependent heme oxygenase required for phytochrome chro-mophore synthesisrdquo Plant Physiology vol 130 no 4 pp 1958ndash1966 2002
[28] J-W Gao J Liu B Li and Z Li ldquoIsolation and purificationof functional total RNA from blue-grained wheat endospermtissues containing high levels of starches and flavonoidsrdquo PlantMolecular Biology Reporter vol 19 no 2 pp 185ndash186 2001
[29] R Patro S M Mount and C Kingsford ldquoSailfish enablesalignment-free isoform quantification from RNA-seq readsusing lightweight algorithmsrdquoNature Biotechnology vol 32 no5 pp 462ndash464 2014
[30] Y-X Qi Y-B Liu and W-H Rong ldquoRNA-Seq and its applica-tions a new technology for transcriptomicsrdquo Hereditas vol 33no 11 pp 1191ndash1202 2011
[31] C R Genovese N A Lazar and T Nichols ldquoThresholdingof statistical maps in functional neuroimaging using the falsediscovery raterdquo NeuroImage vol 15 no 4 pp 870ndash878 2002
[32] J Ye L Fang H Zheng et al ldquoWEGO a web tool for plottingGO annotationsrdquoNucleic Acids Research vol 34 supplement 2pp W293ndashW297 2006
[33] H Abdi ldquoThe Bonferonni and Sidak corrections for multiplecomparisonrdquo Encyclopedia of Measurement and Statistics vol 3pp 103ndash107 2007
[34] J Li Q Ding F Wang Y Zhang H Li and J Gao ldquoIntegrativeanalysis of mRNA and miRNA expression profiles of thetuberous root development at seedling stages in turnipsrdquo PLoSONE vol 10 no 9 Article ID e0137983 2015
[35] J Bennett D Hondred and J C Register ldquoKeeping qRT-PCRrigorous and biologically relevantrdquo Plant Cell Reports vol 34no 1 pp 1ndash3 2015
[36] F Wang L Li H Li et al ldquoTranscriptome analysis of rosetteand folding leaves in Chinese cabbage using high-throughputRNA sequencingrdquo Genomics vol 99 no 5 pp 299ndash307 2012
14 International Journal of Genomics
[37] T Shi Z Gao L Wang et al ldquoIdentification of differentially-expressed genes associated with pistil abortion in Japaneseapricot by genome-wide transcriptional analysisrdquo PLoS ONEvol 7 no 10 Article ID e47810 2012
[38] F Wang H Li Y Zhang et al ldquoMicroRNA expression analysisof rosette and folding leaves in Chinese cabbage using high-throughput Solexa sequencingrdquo Gene vol 532 no 2 pp 222ndash229 2013
[39] X Gu Y Chen Z Gao Y Qiao and X Wang ldquoTranscriptionfactors and anthocyanin genes related to low-temperature toler-ance in rd29A RdreB1BI transgenic strawberryrdquo Plant Physiol-ogy and Biochemistry vol 89 pp 31ndash43 2015
[40] J G Schaart C Dubos I R De La Fuente et al ldquoIdentificationand characterization of MYB-bHLH-WD40 regulatory com-plexes controlling proanthocyanidin biosynthesis in strawberry(Fragaria times ananassa) fruitsrdquoNew Phytologist vol 197 no 2 pp454ndash467 2013
[41] L Xie Z H Wang X H Cheng J J Gao Z P Zhangand L J Wang ldquo5-Aminolevulinic acid promotes anthocyaninaccumulation in Fuji applesrdquo Plant Growth Regulation vol 69no 3 pp 295ndash303 2013
[42] M Daminato F Guzzo and G Casadoro ldquoASHATTERPROOF-like gene controls ripening in non-climac-teric strawberries and auxin and abscisic acid antagonisticallyaffect its expressionrdquo Journal of Experimental Botany vol 64no 12 pp 3775ndash3786 2013
[43] X-H Ji R Zhang N Wang L Yang and X-S Chen ldquoTran-scriptome profiling reveals auxin suppressed anthocyanin bio-synthesis in red-fleshed apple callus (Malus sieversii f niedzwet-zkyana)rdquo Plant Cell Tissue and Organ Culture vol 123 no 2pp 389ndash404 2015
[44] J Chen LMaoW Lu T Ying and Z Luo ldquoTranscriptome pro-filing of postharvest strawberry fruit in response to exogenousauxin and abscisic acidrdquo Planta vol 243 no 1 pp 183ndash197 2016
[45] G A Martınez A R Chaves and M C Anon ldquoEffect of gib-berellic acid on ripening of strawberry fruits (Fragaria annan-assa Duch)rdquo Journal of Plant Growth Regulation vol 13 no 2pp 87ndash91 1994
[46] P K Das D H Shin S-B Choi S-D Yoo G Choi and Y-IPark ldquoCytokinins enhance sugar-induced anthocyanin biosyn-thesis in Arabidopsisrdquo Molecules amp Cells vol 34 no 1 pp 93ndash101 2012
[47] P McAtee S Karim R Schaffer and K David ldquoA dynamicinterplay between phytohormones is required for fruit develop-ment maturation and ripeningrdquo Frontiers in Plant Science vol4 no 3 article 79 7 pages 2013
[48] E Nambara andAMarion-Poll ldquoAbscisic acid biosynthesis andcatabolismrdquoAnnual Review of Plant Biology vol 56 pp 165ndash1852005
[49] Y Jiang and D C Joyce ldquoABA effects on ethylene productionPAL activity anthocyanin and phenolic contents of strawberryfruitrdquo Plant Growth Regulation vol 39 no 2 pp 171ndash174 2003
[50] S Li ldquoTranscriptional control of flavonoid biosynthesis fine-tuning of the MYB-bHLH-WD40 (MBW) complexrdquo PlantSignaling amp Behavior vol 9 Article ID e27522 2014
[51] D R Rudell J K Fellman and J P Mattheis ldquoPreharvest appli-cation of methyl jasmonate to rsquoFujirsquo apples enhances red col-oration and affects fruit size splitting and bitter pit incidencerdquoHortScience vol 40 no 6 pp 1760ndash1762 2005
[52] A Belhadj N Telef C Saigne et al ldquoEffect of methyl jasmonatein combination with carbohydrates on gene expression of PR
proteins stilbene and anthocyanin accumulation in grapevinecell culturesrdquo Plant Physiology and Biochemistry vol 46 no 4pp 493ndash499 2008
[53] E Loreti G Povero G Novi C Solfanelli A Alpi and PPerata ldquoGibberellins jasmonate and abscisic acid modulate thesucrose-induced expression of anthocyanin biosynthetic genesin Arabidopsisrdquo New Phytologist vol 179 no 4 pp 1004ndash10162008
[54] H S Seo J T Song J-J Cheong et al ldquoJasmonic acid carboxylmethyltransferase A key enzyme for jasmonate-regulated plantresponsesrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 98 no 8 pp 4788ndash4793 2001
[55] M T McDowell and J C Lagarias ldquoPurification and biochemi-cal properties of phytochromobilin synthase from etiolated oatseedlingsrdquo Plant Physiology vol 126 no 4 pp 1546ndash1554 2001
[56] G-Q Fu S Xu Y-J Xie et al ldquoMolecular cloning characteri-zation and expression of an alfalfa (Medicago sativa L) hemeoxygenase-1 gene MsHO1 which is pro-oxidants-regulatedrdquoPlant Physiology and Biochemistry vol 49 no 7 pp 792ndash7992011
[57] X-Y Chen X Ding S Xu et al ldquoEndogenous hydrogen per-oxide plays a positive role in the upregulation of heme oxy-genase and acclimation to oxidative stress in wheat seedlingleavesrdquo Journal of Integrative Plant Biology vol 51 no 10 pp951ndash960 2009
[58] J Shin E Park and G Choi ldquoPIF3 regulates anthocyaninbiosynthesis in an HY5-dependent manner with both factorsdirectly binding anthocyanin biosynthetic gene promoters inArabidopsisrdquo Plant Journal vol 49 no 6 pp 981ndash994 2007
[59] S Wang D Pan X Lv et al ldquoProteomic approach reveals thatstarch degradation contributes to anthocyanin accumulation intuberous root of purple sweet potatordquo Journal of Proteomics vol143 pp 298ndash305 2016
[60] H-F Jia C-L Li Y-M Chai Y Xing and Y Shen ldquoSucrosepromotes strawberry fruit ripening by stimulation of abscisicacid biosynthesisrdquo Pakistan Journal of Botany vol 45 no 1 pp169ndash176 2013
[61] W Xu H Peng T Yang et al ldquoEffect of calcium on strawberryfruit flavonoid pathway gene expression and anthocyanin accu-mulationrdquo Plant Physiology amp Biochemistry vol 82 pp 289ndash298 2014
[62] Z-W-SWan and Z Singh ldquoExogenous application of prohexa-dione-calcium promotes fruit colour development of lsquoCrippsPinkrsquo applerdquo Acta Horticulturae vol 1012 pp 219ndash225 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Genomics 5
Table 1 Overview of the sequencing and assembly
Biological replicate 1 Biological replicate 2Sample Control Hematin-treated Control Hematin-treatedClean reads 17808924 26809908 19235452 26228671Genome map rate () 9658 9652 9665 9674Gene map rate () 8646 8627 8704 8563Perfect match () 4677273 (2432) 6510668 (2428) 4414050 (2479) 6149508 (2345)Unique match () 15075148 (8465) 22628939 (8441) 16409557 (8531) 21986962 (8383)Expressed genes 27976 28713 28999 28211Mismatch () 11321162 (6357) 17101590 (6379) 12456871 (6476) 16697280 (6366)Total unmapped reads 645135 (335) 932213 (348) 608914 (342) 856028 (326)
clean tags that mapped to a single gene were retained forfurther DEG analysis
In addition the perfect match rates were 2432 24282479 and 2435 in control 1 control 2 hematin 1 andhematin 2 respectively
The saturation of the libraries with and without thehematin treatment was analyzedThe number of the detectedgenes became saturated at about 2 million reads (FigureS2) This indicates that the obtained reads are sufficient forcomplete transcriptome coverage The randomness of RNAfragmentation in the four libraries was assessed for subse-quent bioinformatics analysis The results showed that thereads in each position of the reference gene were distributedevenly and demonstrated highly similar tendencies in alllibraries (Figure S3)
33 Genes Differentially Regulated in Response to ExogenousHematin Based on sequencing results of the four mRNAlibraries approximately 29000 genes (about 70 of thereference genes in the octoploid strawberry genome) weredetected in each library Additionally correlations amonggenes based on RPKM between the two biological replicateswere analyzed the correlation coefficients (1198772) were high(093 and 095 for the control and hematin-treated groupresp) Genes expressed in both replicates were screened Atotal of 28713 28999 27976 and 28211 genes (Table 1) wereexpressed in the control and hematin-treated samples
To reveal the molecular pathways regulating strawberrycoloring in response to hematin the DEGs between the con-trol and hematin-treated fruit were analyzed We comparedthe gene expression profiles between the control and thetreated samples in both biological replicates DEGs detectedin both biological replicates were screened for subsequentanalysis A total of 1080 (402 up- and 678 downregulated)genes were differentially expressed in the 5120583M hematin-treated groups comparedwith the control groupsMore geneswere downregulated in the hematin-treated group comparedto the control group
To facilitate the global analysis of gene expression aGO analysis was performed by mapping each differentiallyexpressed gene into the records of the GO database GeneOntology (GO) enrichment analysis of the DEGs in thecontrol and hematin-treated groups was performed to revealthe possible mechanisms under which hematin promotes
anthocyanin biosynthesis GO terms with 119901 lt 005 wererepresented among genes with significant changes in expres-sion over a given interval In this study only three termsincluding anatomical structure arrangementmeristem struc-tural organization radial pattern formationwere significantlyenriched in biological process (Supplementary Table S2)AlthoughmostGo termswere not significantly enriched theycould provide a reference for further study According to theGO cellular components a large number of the DEGs wereclassified intomain cell organelles such as vacuole nucleolusmitochondrion apoplast chloroplast plastid cytoplasm andmembrane In particular many DEGs were assigned to theplastid and chloroplast (Figure 3) indicating that hematinhas an important impact on the expression of plastid andchloroplast genes It is probably because heme oxygenase 1 isa soluble plastid protein [22] According to the GO molec-ular function a large number of the DEGs were classifiedintoUDP-glucosyltransferase sucrose transmembrane trans-porter activity nucleic acid binding and DNA binding termswhich are closely related to anthocyanin biosynthesis ManyDEGs were classified into heme binding peroxidase activityand tetrapyrrole binding (Figure 3) It is probably becausehematin can increase the activity of HO1 and promote thedegradation of hemoglobin [17] In addition anthocyaninaccumulation in fruit is closely related to hormone [8] andcarbohydrate biosynthesis [9] According to the GO biologi-cal process a large number of the DEGs were classified intoregulation of hormone levels cytokinin metabolic processresponse to hormone response to abscisic acid response toauxin and response to ethylene terms indicating that thehormone-related DEGs are involved in regulating the antho-cyanin biosynthesis in the hematin-treated fruit Many DEGswere classified into starch and sucrose biosynthetic and trans-port process and phenylpropanoid metabolic process termsindicating that hematin can promote anthocyanin accumu-lation via the phenylpropanoid metabolic and carbohydratemetabolism In addition many DEGs were classified into theprocess for response to light stimulus term indicating thathematin may also participate in the light stimulus system
Pathway enrichment analysis was performed to under-stand the biological functions of the DEGs by identify-ing significantly enriched signal transduction pathways ormetabolic pathways [39] In the study a number of alteredbiological pathways associated with the hematin treatment
6 International Journal of Genomics
0 10 20 30 40 50 60 70 80Starch biosynthetic process
Cytokinin metabolic processResponse to abscisic acid
Response to auxinResponse to ethylene
Phenylpropanoid metabolic processSucrose transport
Regulation of hormone levelsResponse to light stimulus
Response to hormonePeroxidase activity
Sucrose transmembrane transporter activityUDP-glucosyltransferase
Heme bindingTetrapyrrole binding
DNA bindingNucleic acid binding
VacuoleNucleolusApoplast
MitochondrionChloroplast
PlastidCytoplasmMembrane
Biol
ogic
al p
rogr
ess
Mol
ecul
ar fu
nctio
nC
ellu
lar c
ompo
nent
Number of DEGs
Figure 3 Gene classification based on Gene Ontology (GO) for DEGs in the strawberries treated by hematin
were identified and three pathways including RNA poly-merase pyrimidine metabolism and purine metabolismwere significantly enriched (119876-value lt 005) (Table 2)Although the majority of the pathway terms were not sig-nificantly enriched the pathway enrichment analysis helpsus to further understand the biological functions of theDEGs and the molecular mechanisms that regulate fruitcoloring in response to the hematin treatmentThese pathwayterms included metabolic pathways biosynthesis of sec-ondary metabolites isoflavonoid biosynthesis phenylalaninemetabolism phenylalanine tyrosine and tryptophan biosyn-thesis starch and sucrose metabolism plant hormone signaltransduction flavone and flavonol biosynthesis and phenyl-propanoid biosynthesis terms (Supplementary Table S2)
34 Analysis of DEGs Involved in Anthocyanin Biosynthesis
341 Transcription Factors Transcription factors (TFs) areproteins that regulate the expression of downstream targetgenes Many TFs such as v-myb avian myeloblastosis viraloncogene homolog (MYB) basic helix-loop-helix (bHLH)WD40-repeats protein (WD40) and MADS-box (MADS)directly regulating the expression of the structural genes inanthocyanin biosynthesis have been identified from manyspecies [8] Therefore the enhanced expression of key TFs
might regulate the anthocyanin biosynthesis In our studysome anthocyanin biosynthesis-related TFs were found to beuniquely present in the DEG profiling of the hematin-treated fruit for example MYB bHLH PIF3 MADs-boxAP2-EREBP and ABI3VP1 (Table 3) The anthocyanin bio-synthesis genes are regulated primarily by the MBW tran-scription factor which is a ternary transcription factor com-plex [10 11] Most of the MYBs involved in regulating antho-cyanin biosynthesis are positive regulators of transcription[8] However MYBs can act as repressors too such as straw-berry FaMYB1 and FaMYB9 and grapevine VvMYB4 whichcan significantly suppress the biosynthesis of anthocyaninsand flavonols [40] In this study three unigenes belong-ing to MYB transcription factors were differentiallyexpressed and two MYB transcription factors were signifi-cantly upregulated Of these the unigene ldquoFANhybrscf000001461g000071rdquo from theMYB family of R2R3MYBtranscription factors was significantly upregulatedMoreoverincreasing evidence indicates that the expression of thebHLHs promotes anthocyanin accumulation in fruit [41] Inour study all of the unigenes from the bHLH family weredownregulated among which the expression level of unigeneFANhyb rscf000012921g000031 was significantly decreased6250-fold (Table 3) FANhyb rscf000022101g000011 en-codes a predicted MADS-box transcription factor and it was
International Journal of Genomics 7
Control Hematin-treated
FANhyb_icon00002410_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_rscf000049691g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00027696_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00004395_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_rscf000063641g000011
lowastlowast
lowast
Control Hematin-treated
FANhyb_rscf000001411g000161
Control Hematin-treated
FANhyb_icon00006295_a1g000011lowastlowast
Control Hematin-treated
FANhyb_rscf000031551g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00001444_a1g000011
lowastlowast
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
0
05
1
15
2
25
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
005
115
225
335
445
5
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
Control Hematin-treated
FANhyb_rscf000023111g000011
lowastlowast
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
Control Hematin-treated
FANhyb_rscf000002951g000031
lowastlowast
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
Figure 4 qRT-PCR validation of the RNA-Seq based gene expression The values indicate means of three biological replicates plusmn SD Starindicates that the expression level is significantly different between the hematin-treated and the control group (lowast119901 lt 005 lowastlowast119901 lt 001)
8 International Journal of Genomics
Table 2 Pathway enrichment analysis of differentially expressed genes
Pathway DEGs with pathwayannotation (451)
All genes with pathwayannotation (15877)
Corrected 119901 value 119876-value Pathway ID
RNA polymerase 16 183 688119864 minus 5 00068107 ko03020Pyrimidine metabolism 20 317 00007651746 00320 ko00240Purine metabolism 21 347 00009710371 0032044224 ko00230
significantly downregulated There were three unigenes inthe AP2-EREBP family with only one upregulated and theothers downregulated These results suggest that exogenoushematin is involved in developmental transcriptionalregulation of anthocyanin biosynthesis
342 DEGs in Anthocyanin Biosynthesis The anthocyaninbiosynthetic pathway has been studied intensively Theanthocyanin biosynthetic pathway via the phenylpropanoidpathway is well known [8] In this study the transcriptomedata showed a significant increase in the expression of genesin the anthocyanin biosynthesis pathway when exposedto exogenous hematin The structural genes involved inanthocyanin biosynthesis including DFR LAR and UDP-glycosyltransferase exhibited significant differential expres-sion in response to the hematin treatment (Table 4) Forexample two DFR genes and one LAR gene were signifi-cantly upregulated (the expression increased 18- 30- and3-fold in the hematin-treated fruit resp) In addition allsix genes encodingUDP-glycosyltransferasewere significantlyupregulated Among these the expression of one gene(Fanhy icon15742070o1g000011) was upregulated 18305-fold The results suggest that exogenous hematin may havean important role in promoting anthocyanin biosynthesis
343 DEGs in Hormone Signaling Transduction in Responseto Hematin Auxin has been shown to negatively regulatethe expression of the anthocyanin biosynthesis genes [42] Inthis study there were three unigenes involved in the auxinsignaling pathway and all were downregulated includingone gene encoding auxin efflux carrier component 1-like(AUX1-like) and two genes encoding auxin response factors(ARF) These results suggest that hematin promotes antho-cyanin accumulation by regulating auxin in the colorationprocess of strawberries It has shown that auxin suppressesanthocyanin biosynthesis in the red-fleshed apple callus [43]Endogenous expression of auxin [44 45] has been foundto impede anthocyanin accumulation in strawberries Inadditional cytokinins are also known to play an importantrole in anthocyanin biosynthesis It has shown to enhanceanthocyanin accumulation in Zea mays and regulate antho-cyanin production and composition in suspension culturesof strawberry cells [46] In this study two unigenes relatedto the cytokinin signaling pathway were identified and theywere downregulated as well One gene was identified ascytokinin dehydrogenase 5-like which catalyzes the irre-versible degradation of cytokinins This result indicates thathematin promotes anthocyanin accumulation by regulatingthe cytokinin signaling pathway
Extensive researches have shown that ABA plays animportant role in the regulation of anthocyanin biosynthesisin nonclimacteric fruit [47] In this study a total of five uni-genes related to the ABA signaling pathway were identifiedamong which two were upregulated and three were down-regulated Two genes which are involved in the abscisic acidresponse were upregulated These two unigenes encode 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) and ABA overlysensitive 5 (ABO5) respectively ABA is synthesized fromcarotenoids via several enzymatic reactions in the plastidThe rate-limiting step in these reactions is catalyzed by 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) [48] Mutations inthe FaNCED1 gene result in colorless strawberry fruit whichcan become colored through the application of exogenousABA [12] Another unigene was identified to encode abscisicacid 81015840-hydroxylase 1-like a key enzyme in the oxidativecatabolism of abscisic acid which was downregulated Appli-cation of exogenous ABA regulates phenylalanine ammonialyase activity and increases the phenolic and anthocyanincontent of strawberry fruit [12 49] Our results suggestthat hematin promotes ABA biosynthesis and inhibits ABAdisintegration in the hematin-treated fruit
In Arabidopsis JAs can affect anthocyanin accumulationvia the interaction of negative regulators with the MBWcomplex of transcription factors involved in anthocyaninbiosynthesis [50] Preharvest application of JA to ldquoFujirdquo applesenhances red coloration [51] JA vapor treatment can alsoenhance anthocyanins in strawberry fruit [52] In this studyjasmonate O-methyltransferase-like exhibited significantlydifferential expression Jasmonate O-methyltransferase-likeis also known to be a key enzyme for jasmonate-regulatedplant responses to stimuli [53 54] These results suggest thathematin increases the activity of the JA-regulated antho-cyanin biosynthesis
344 DEGs in the Phytochrome Signaling Pathway in Responseto Hematin Phytochromes that act as photoreceptors playan important role in anthocyanin regulation [15] Phy-tochromes are homodimeric chromoproteins where eachholophytochrome is composed of a phytochrome protein(apophytochrome) covalently bound to a linear tetrapyrrolePΦB HO1 is crucial to this process and acts as a rate-limiting enzyme in the biosynthesis of PΦB [55] Exogenoushematin can also induce HO-1 expression in many plants[56 57] In this study we found exogenous hematin signif-icantly increased the expression of FaHO1 (Table 5) Thisresult implies that hematin can promote PΦB biosynthesisRecently it has been reported that hematin could inducethe accumulation of far-red phytochrome and phytochrome
International Journal of Genomics 9
Table 3 DEGs acting as transcription factors in response to exogenous hematin
Annotation transcription factors Gene IDReal
fold-changevalues
Upregulationdownregulation FDR
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb icon00006295 a1g000011 938 Up 120119864 minus 05
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb rscf000031551g000011 4 Down 480119864 minus 16
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL5-like
FANhyb rscf000000291g000101 4 Down 320119864 minus 08
MYBdomain protein 66 FANhyb rscf000001461g000071 654 Up 780119864 minus 07MYB-related protein 3R-1-like partial FANhyb rscf000001411g000161 282 Up 830119864 minus 04PREDICTED transcription factorGAMYB-like FANhyb rscf000006491g000061 303 Down 150119864 minus 13
PREDICTED probable WRKYtranscription factor 53-like FANhyb rscf000019731g000021 217 Down 200119864 minus 08
Transcription factor bHLH80-like FANhyb rscf000012921g000031 6250 Down 150119864 minus 06Transcription factor bHLH70-like FANhyb rscf000002951g000031 526 Down 660119864 minus 12Transcription factor ICE1-like FANhyb rscf000001701g000061 244 Down 450119864 minus 18C2H2-like zinc finger protein FANhyb rscf000011431g000021 282 Up 590119864 minus 04Zinc finger protein MAGPIE-like FANhyb rscf000030241g000011 204 Up 150119864 minus 04PREDICTED axial regulator YABBY5-like FANhyb rscf000016671g000011 417 Down 730119864 minus 05
PREDICTED axial regulator YABBY1-like FANhyb rscf000068991g000011 357 Down 620119864 minus 06
PREDICTED dof zinc finger proteinDOF37-like isoform 1 FANhyb rscf000068561g000011 263 Down 280119864 minus 04
ZF-HD homeobox protein FANhyb rscf000000441g000221 385 Down 420119864 minus 08ZF-HD homeobox proteinAt4g24660-like FANhyb rscf000052411g000011 357 Down 470119864 minus 09
PREDICTED B3 domain-containingprotein REM14-like FANhyb rscf000010091g000021 243 Up 530119864 minus 04
PREDICTED B3 domain-containingtranscription factor ABI3-like FANhyb rscf000012711g000021 455 Down 130119864 minus 28
Growth-regulating factor 6 FANhyb rscf000000241g000291 345 Down 330119864 minus 34Growth-regulating factor 5 FANhyb icon00009704 a1g000011 27 Down 190119864 minus 06GATA type zinc finger transcriptionfactor family protein FANhyb rscf000003931g000131 1111 Down 110119864 minus 05
PREDICTED zinc finger CCCHdomain-containing protein 2-like FANhyb icon00006770 a1g000011 313 Down 680119864 minus 49
PREDICTED E2F transcriptionfactor-like E2FE-like FANhyb icon00002702 a1g000011 278 Down 240119864 minus 04
PREDICTED uncharacterized protein FANhyb rscf000010661g000011 263 Down 490119864 minus 06PREDICTED nuclear transcriptionfactor Y subunit A-3-like isoform 1 FANhyb rscf000000081g000351 385 Down 500119864 minus 20
PREDICTED homeobox-leucinezipper protein ATHB-8-like FANhyb rscf000000151g000051 263 Down 100119864 minus 06
PREDICTED MADS-boxtranscription factor 18-like FANhyb rscf000003231g000161 243 Down 180119864 minus 06
PREDICTED zinc finger protein132-like isoform 1 FANhyb rscf000070151g000011 243 Down 180119864 minus 08
PREDICTED transcription factorPIF3-like FANhyb rscf000006691g000021 346 Up 140119864 minus 07
10 International Journal of Genomics
Table 4 Regulation of the DEGs in anthocyanin biosynthesis in response to exogenous hematin
Genes in anthocyaninbiosynthesis Gene ID
Realfold-change
valuesRegulation level FDR
Dihydroflavonol-4-reductase FANhyb icon00002410 a1g000011 3091 Up 910119864 minus 06
Flavonol synthase FANhyb icon00015354 a1g000011 345 down 180119864 minus 05Isoflavone reductasehomolog FANhyb icon20341135 s1g000011 334 Up 750119864 minus 14
UDP-glycosyltransferase FANhyb rscf000049691g000011 307 Up 640119864 minus 08UDP-glycosyltransferaseactivity FANhyb icon15742070 o1g000011 18 30563 Up 510119864 minus 09
UDP-glycosyltransferase73C1 FANhyb icon00027696 a1g000011 298 Up 560119864 minus 04
UDP-glycosyltransferaseactivity FANhyb icon00008064 a1g000011 249 Up 310119864 minus 04
PREDICTEDUDP-glucose flavonoid3-O-glucosyltransferase7-like
FANhyb rscf000055631g000011 212 Up 110119864 minus 19
PREDICTEDUDP-glycosyltransferase76F1-like
FANhyb rscf000021771g000031 2 Up 720119864 minus 29
A transcripts in the etiolated leaves of wheat seedling phy-tochromes [26] However our study shows that phytochromeA was not differentially expressed It is probably becausethe hematin-treated strawberries were not shaded during theexperiment while the function of phytochrome A is light-dependent In addition we found that a downstream gene ofphytochrome PIF3 (FANhyb rscf000006691g000021) wassignificantly upregulated PIF3 is thought to be a positive reg-ulator of phytochrome B mediated by light-dependent signaltransduction [58]These results suggest that the phytochromepathways may be involved in the anthocyanin biosynthesispromoted by hematin
345 DEGs in Starch and Sucrose Degradation Proteomicapproaches have revealed that starch degradation contributesto anthocyanin accumulation in tuberous roots of the purplesweet potato variety [59] Starch and sucrose biosynthesis isimportant for anthocyanin accumulation in strawberry fruit[60] In this study we found the DEGs involved in starchand sucrose synthesis in the hematin-treated fruit were alldownregulated (Table 5) For example the expression of theunigenes encoding soluble starch synthase 3 sucrose trans-port protein SUC2-like and a probable sucrose-phosphatesynthase 4-like enzyme were decreased 23- 46- 32- and2-fold respectively This suggests that exogenous hematinregulates the biosynthesis of starch and sucrose and henceaffects fruit coloring
346 DEGs in the Calcium Pathway in Response to HematinCalcium can also increase the transcription levels of keystructural genes F3H DFR ANT and UFGT in the whitestage of strawberries [61] In this study we found thatthree DEGs in calcium biosynthesis and transport were
downregulated in the hematin-treated fruit (Table 5) Onlythe unigene encoding predicted cationcalcium exchanger5-like was upregulated The unigenes encoding a calpain-type cysteine protease and a predicted calcium-binding pro-tein PBP1-like were downregulated respectively Exogenousapplication of calcium can promote apple coloring [62] inaddition to variation in anthocyanin content [61] Our resultsindicate that the biosynthesis and transport of calcium areinvolved in the development of coloring in hematin-treatedstrawberries
35 Validation of Selected DEGs by qRT-PCR To validate theexpression of the DEGs obtained from RNA-Seq 11 DEGswere selected for qRT-PCR including structural genes (DFRand UDFGs) transcription factor genes (MYB and bHLH)and phytochrome chromophore-related gene (HO-1) Theprimers used for qRT-PCR are listed in Supplementary TableS1 The qRT-PCR results were consistent with the RNA-Seqdata (Figure 4) except for FANhyb rscf000001411g000161which had a higher log
2ratio (hematin-treatedcontrol) in
the transcriptome data than in qRT-PCR These results indi-cate the RNA-Seq data from the strawberry transcriptome isreproducible and accurate
4 Conclusions
In this study the anthocyanin content in the strawberryfruit was elevated by the application of exogenous hematinThis is the first report that hematin could increase theanthocyanin production in fruit Furthermore we exploredthe effects of the exogenous hematin on metabolic path-ways using genome-wide transcriptome analysis The resultsindicate that the expression levels of many genes involved
International Journal of Genomics 11
Table 5 Other DEGs identified in anthocyanin biosynthesis-related pathways
Anthocyaninbiosynthesis-relatedpathways
Annotation genes Gene IDReal fold-changevalues
Upregulationdownregulation FDR
Calcium ion binding
Cationcalciumexchanger 5-like
FANhyb rscf000019061g000011 203 Up 64119864 minus 06
Calpain-type cysteineprotease family
FANhyb icon00011755 a1g000011 345 Down 18119864 minus 09
Calcium-bindingprotein PBP1-like
FANhyb rscf000007501g000081 323 Down 97119864 minus 04
Calpain-type cysteineprotease family
FANhyb icon00023658 a1g000011 233 Down 60119864 minus 08
CytokininCytokinin
dehydrogenase 5-likeFANhyb iscf00325393 1 s1g000011 667 Down 62119864 minus 06
Cytochrome P450714A1
FANhyb rscf000005921g000031 217 Down 98119864 minus 10
JasmonateJasmonate
O-methyltransferase-like
FANhyb rscf000000041g000131 201 Up 70119864 minus 04
Abscisic acid
Protein ABSCISICACID-INSENSITIVE
5-likeFANhyb rscf000021641g000011 4 Down 16119864 minus 18
ABA overly sensitive 5 FANhyb icon00051144 a1g000011 291 Up 24119864 minus 059-cis-
epoxycarotenoiddioxygenase NCED1
FANhyb icon18399909 o1g000011 441 Up 43119864 minus 06
Abscisic acid81015840-hydroxylase 1-like
FANhyb icon00000938 a1g000011 256 Down 14119864 minus 06
Abscisic acid receptorPYR1-like
FANhyb icon00020426 a1g000011 208 Down 30119864 minus 06
Auxin
Auxin efflux carriercomponent 1-like
FANhyb rscf000010081g000031 384 Down 23119864 minus 06
Auxin response factor8
FANhyb rscf000013061g000021 263 Down 61119864 minus 17
Auxin response factor17-like
FANhyb rscf000060741g000011 217 Down 11119864 minus 36
Starch and sucrose
Glycosyltransferasefamily 35
FANhyb rscf000000341g000081 384 Down 11119864 minus 05
Soluble starchsynthase 3
FANhyb rscf000000451g000041 232 Down 38119864 minus 12
Sucrose transportprotein SUC2-like
FANhyb icon00036727 a1g000011 455 Down 98119864 minus 04
Sucrose transportprotein SUC2-like
FANhyb rscf000007551g000021 323 Down 66119864 minus 10
Probablesucrose-phosphatesynthase 4-like
FANhyb rscf000013501g000021 213 Down 11119864 minus 14
Phytochrome
PREDICTEDtranscription factor
PIF3-likeFANhyb rscf000006691g000021 346 up 14119864 minus 07
Phytochrome E-like FANhyb rscf000004361g000041 4 Down 33119864 minus 05
Heme oxygenase 1 FANhyb icon00004395 a1g000011 258 27119864 minus 07 27119864 minus 07
12 International Journal of Genomics
Hematin
Endoplasmicreticulum
WD40 bLHLMYB
Structural genes
TFs
Nuclear
JA
ABA Sucrose
Plastid
G-cis-neoxanthin
Apophytochrome
StarchHeme
BVSucroseABA
Holophytochrome
PfrPIF3
Flavonoids Anthocyanins
Vacuole
Cytoplasm
Cell wall
①
②③
④⑤
⑥
⑦
PΦB
PΦB
CTKAuxin
Ca2+
Figure 5 Genes and biological pathways that were involved in regulation of anthocyanin accumulation promoted by hematin Outermostbox represents the cell wall Bilayer oval shape represents the plastidThe red-brown ring and network represent the nuclear and endoplasmicreticulum respectively The red crescent represents the vacuole The box in the nucleus represents transcription factors (TFs) Yellow MYBgreenWD40 and red bHLH represent a ternary transcription factor complex which transcribes anthocyanin biosynthesis genes The circlednumbers represent biological pathways involved in the regulation of anthocyanin accumulation promoted by hematin A represents thephytochrome regulation pathwayB represents the starch and sucrose pathwayCDE andF represent phytohormones ABA JA AuxinandCTK regulation pathways respectivelyG represents theCa2+ regulation pathway Pfr represents the far-red phytochrome PIF3 representsphytochrome-interacting factor Each anthocyanin biosynthesis regulatory pathway related gene is in Table 5
in anthocyanin biosynthesis were significantly altered withthe hematin treatment This suggests that the physiologicalprocess of fruit color development is regulated through com-plex interactions among anthocyanin biosynthesis pathwaysplant hormone signal transduction pathways phytochromesignal transduction pathways starch and sugar metabolicpathways calcium pathways and transcription factors (Fig-ure 5) This study adds to the in-depth understanding of thefruit coloration process in strawberry
Competing Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contributions
Jianwei Gao and Liangju Wang conceived and designed theresearch Yi Li Huayin Li Fengde Wang Jingjuan Li andYihui Zhang conducted experiments Yi Li analyzed the dataYi Li and Fengde Wang wrote the manuscript All authorsread and approved the manuscript Yi Li and Huayin Licontributed equally to this work
Acknowledgments
This work was supported by the Modern Agricultural Indus-trial Technology System Funding of Shandong ProvinceChina (SDAIT-02-022-04) the China agriculture researchsystem (CARS-25) and the National Science Foundation ofChina (31401820)
References
[1] K Aaby SMazur A Nes andG Skrede ldquoPhenolic compoundsin strawberry (Fragaria x ananassa Duch) fruits compositionin 27 cultivars and changes during ripeningrdquo Food Chemistryvol 132 no 1 pp 86ndash97 2012
[2] T A Colquhoun L A Levin H RMoskowitz VMWhitakerD G Clark and K M Folta ldquoFraming the perfect strawberryan exercise in consumer-assisted selection of fruit cropsrdquoJournal of Berry Research vol 2 no 1 pp 45ndash61 2012
[3] F Giampieri J M Alvarez-Suarez L Mazzoni et al ldquoThepotential impact of strawberry on human healthrdquo NaturalProduct Research vol 27 no 4 pp 448ndash455 2013
[4] Y S Velioglu GMazza L Gao and B D Oomah ldquoAntioxidantactivity and total phenolics in selected fruits vegetables and
International Journal of Genomics 13
grain productsrdquo Journal of Agricultural amp Food Chemistry vol46 no 10 pp 4113ndash4117 1998
[5] S S Hassellund A Flaa S E Kjeldsen et al ldquoEffects of antho-cyanins on cardiovascular risk factors and inflammation inpre-hypertensive men a double-blind randomized placebo-controlled crossover studyrdquo Journal of Human Hypertensionvol 27 no 2 pp 100ndash106 2013
[6] K J Meyers C B Watkins M P Pritts and R H Liu ldquoAnti-oxidant and antiproliferative activities of strawberriesrdquo Journalof Agricultural amp Food Chemistry vol 51 no 23 pp 6887ndash68922003
[7] S Zafra-Stone T YasminM Bagchi AChatterjee J AVinsonand D Bagchi ldquoBerry anthocyanins as novel antioxidants inhuman health and disease preventionrdquo Molecular Nutrition ampFood Research vol 51 no 6 pp 675ndash683 2007
[8] L Jaakola ldquoNew insights into the regulation of anthocyanin bio-synthesis in fruitsrdquoTrends in Plant Science vol 18 no 9 pp 477ndash483 2013
[9] K Springob J-I Nakajima M Yamazaki and K Saito ldquoRecentadvances in the biosynthesis and accumulation of antho-cyaninsrdquo Natural Product Reports vol 20 no 3 pp 288ndash3032003
[10] A Gonzalez M Zhao J M Leavitt and A M Lloyd ldquoRegu-lation of the anthocyanin biosynthetic pathway by the TTG1bHLHMyb transcriptional complex in Arabidopsis seedlingsrdquoPlant Journal vol 53 no 5 pp 814ndash827 2008
[11] K Petroni and C Tonelli ldquoRecent advances on the regulationof anthocyanin synthesis in reproductive organsrdquo Plant Sciencevol 181 no 3 pp 219ndash229 2011
[12] H-F Jia Y-M Chai C-L Li et al ldquoAbscisic acid plays animportant role in the regulation of strawberry fruit ripeningrdquoPlant Physiology vol 157 no 1 pp 188ndash199 2011
[13] C Merchante J G Vallarino S Osorio et al ldquoEthylene isinvolved in strawberry fruit ripening in an organ-specific man-nerrdquo Journal of Experimental Botany vol 64 no 14 pp 4421ndash4439 2013
[14] A Maier A Schrader L Kokkelink et al ldquoLight and the E3ubiquitin ligase COP1SPA control the protein stability of theMYB transcription factors PAP1 and PAP2 involved in antho-cyanin accumulation in Arabidopsisrdquo Plant Journal vol 74 no4 pp 638ndash651 2013
[15] H Lange W Shropshire and H Mohr ldquoAn analysis of phyto-chrome-mediated anthocyanin synthesisrdquo Plant Physiology vol47 no 5 pp 649ndash655 1971
[16] T D Elich and J C Lagarias ldquoPhytochrome chromophore bio-synthesis both 5-aminolevulinic acid and biliverdin overcomeinhibition by gabaculine in etiolated Avena sativa L seedlingsrdquoPlant Physiology vol 84 no 2 pp 304ndash310 1987
[17] Y Han W Xuan T Yu et al ldquoExogenous hematin alleviatesmercury-induced oxidative damage in the roots of Medicagosativardquo Journal of Integrative Plant Biology vol 49 no 12 pp1703ndash1713 2007
[18] M Koornneef J Cone R Dekens E OrsquoHerne-Robers CSpruit and R Kendrick ldquoPhotomorphogenic responses of longhypocotyl mutants of tomatordquo Journal of Plant Physiology vol120 no 2 pp 153ndash165 1985
[19] M J Terry and R E Kendrick ldquoFeedback inhibition of chloro-phyll synthesis in the phytochrome chromophore-deficientaurea and yellow-green-2 mutants of tomatordquo Plant Physiologyvol 119 no 1 pp 143ndash152 1999
[20] B M Parks and P H Quail ldquoPhytochrome-deficient hy1 andhy2 long hypocotyl mutants of Arabidopsis are defective inphytochrome chromophore biosynthesisrdquo Plant Cell vol 3 no11 pp 1177ndash1186 1991
[21] M J Terry ldquoPhytochrome chromophore-deficient mutantsrdquoPlant Cell amp Environment vol 20 no 6 pp 740ndash745 1997
[22] T Muramoto T Kohchi A Yokota I Hwang and H MGoodman ldquoThe Arabidopsis photomorphogenic mutant hy1 isdeficient in phytochrome chromophore biosynthesis as a resultof a mutation in a plastid heme oxygenaserdquo Plant Cell vol 11no 3 pp 335ndash347 1999
[23] M D Maines ldquoThe heme oxygenase system a regulator ofsecond messenger gasesrdquo Annual Review of Pharmacology andToxicology vol 37 no 1 pp 517ndash554 1997
[24] W Xuan F-Y Zhu S Xu et al ldquoThe heme oxygenasecarbonmonoxide system is involved in the auxin-induced cucumberadventitious rooting processrdquo Plant Physiology vol 148 no 2pp 881ndash893 2008
[25] R Amooaghaie F Tabatabaei and A-M Ahadi ldquoRole ofhematin and sodium nitroprusside in regulating Brassica nigraseed germination under nanosilver and silver nitrate stressesrdquoEcotoxicology and Environmental Safety vol 113 pp 259ndash2702015
[26] Y Liu X Li L Xu and W Shen ldquoDe-etiolation of wheatseedling leaves cross talk between heme oxygenasecarbonmonoxide and nitric oxiderdquo PLoS ONE vol 8 no 12 ArticleID e81470 2013
[27] T Muramoto N Tsurui M J Terry A Yokota and T KohchildquoExpression and biochemical properties of a ferredoxin-dependent heme oxygenase required for phytochrome chro-mophore synthesisrdquo Plant Physiology vol 130 no 4 pp 1958ndash1966 2002
[28] J-W Gao J Liu B Li and Z Li ldquoIsolation and purificationof functional total RNA from blue-grained wheat endospermtissues containing high levels of starches and flavonoidsrdquo PlantMolecular Biology Reporter vol 19 no 2 pp 185ndash186 2001
[29] R Patro S M Mount and C Kingsford ldquoSailfish enablesalignment-free isoform quantification from RNA-seq readsusing lightweight algorithmsrdquoNature Biotechnology vol 32 no5 pp 462ndash464 2014
[30] Y-X Qi Y-B Liu and W-H Rong ldquoRNA-Seq and its applica-tions a new technology for transcriptomicsrdquo Hereditas vol 33no 11 pp 1191ndash1202 2011
[31] C R Genovese N A Lazar and T Nichols ldquoThresholdingof statistical maps in functional neuroimaging using the falsediscovery raterdquo NeuroImage vol 15 no 4 pp 870ndash878 2002
[32] J Ye L Fang H Zheng et al ldquoWEGO a web tool for plottingGO annotationsrdquoNucleic Acids Research vol 34 supplement 2pp W293ndashW297 2006
[33] H Abdi ldquoThe Bonferonni and Sidak corrections for multiplecomparisonrdquo Encyclopedia of Measurement and Statistics vol 3pp 103ndash107 2007
[34] J Li Q Ding F Wang Y Zhang H Li and J Gao ldquoIntegrativeanalysis of mRNA and miRNA expression profiles of thetuberous root development at seedling stages in turnipsrdquo PLoSONE vol 10 no 9 Article ID e0137983 2015
[35] J Bennett D Hondred and J C Register ldquoKeeping qRT-PCRrigorous and biologically relevantrdquo Plant Cell Reports vol 34no 1 pp 1ndash3 2015
[36] F Wang L Li H Li et al ldquoTranscriptome analysis of rosetteand folding leaves in Chinese cabbage using high-throughputRNA sequencingrdquo Genomics vol 99 no 5 pp 299ndash307 2012
14 International Journal of Genomics
[37] T Shi Z Gao L Wang et al ldquoIdentification of differentially-expressed genes associated with pistil abortion in Japaneseapricot by genome-wide transcriptional analysisrdquo PLoS ONEvol 7 no 10 Article ID e47810 2012
[38] F Wang H Li Y Zhang et al ldquoMicroRNA expression analysisof rosette and folding leaves in Chinese cabbage using high-throughput Solexa sequencingrdquo Gene vol 532 no 2 pp 222ndash229 2013
[39] X Gu Y Chen Z Gao Y Qiao and X Wang ldquoTranscriptionfactors and anthocyanin genes related to low-temperature toler-ance in rd29A RdreB1BI transgenic strawberryrdquo Plant Physiol-ogy and Biochemistry vol 89 pp 31ndash43 2015
[40] J G Schaart C Dubos I R De La Fuente et al ldquoIdentificationand characterization of MYB-bHLH-WD40 regulatory com-plexes controlling proanthocyanidin biosynthesis in strawberry(Fragaria times ananassa) fruitsrdquoNew Phytologist vol 197 no 2 pp454ndash467 2013
[41] L Xie Z H Wang X H Cheng J J Gao Z P Zhangand L J Wang ldquo5-Aminolevulinic acid promotes anthocyaninaccumulation in Fuji applesrdquo Plant Growth Regulation vol 69no 3 pp 295ndash303 2013
[42] M Daminato F Guzzo and G Casadoro ldquoASHATTERPROOF-like gene controls ripening in non-climac-teric strawberries and auxin and abscisic acid antagonisticallyaffect its expressionrdquo Journal of Experimental Botany vol 64no 12 pp 3775ndash3786 2013
[43] X-H Ji R Zhang N Wang L Yang and X-S Chen ldquoTran-scriptome profiling reveals auxin suppressed anthocyanin bio-synthesis in red-fleshed apple callus (Malus sieversii f niedzwet-zkyana)rdquo Plant Cell Tissue and Organ Culture vol 123 no 2pp 389ndash404 2015
[44] J Chen LMaoW Lu T Ying and Z Luo ldquoTranscriptome pro-filing of postharvest strawberry fruit in response to exogenousauxin and abscisic acidrdquo Planta vol 243 no 1 pp 183ndash197 2016
[45] G A Martınez A R Chaves and M C Anon ldquoEffect of gib-berellic acid on ripening of strawberry fruits (Fragaria annan-assa Duch)rdquo Journal of Plant Growth Regulation vol 13 no 2pp 87ndash91 1994
[46] P K Das D H Shin S-B Choi S-D Yoo G Choi and Y-IPark ldquoCytokinins enhance sugar-induced anthocyanin biosyn-thesis in Arabidopsisrdquo Molecules amp Cells vol 34 no 1 pp 93ndash101 2012
[47] P McAtee S Karim R Schaffer and K David ldquoA dynamicinterplay between phytohormones is required for fruit develop-ment maturation and ripeningrdquo Frontiers in Plant Science vol4 no 3 article 79 7 pages 2013
[48] E Nambara andAMarion-Poll ldquoAbscisic acid biosynthesis andcatabolismrdquoAnnual Review of Plant Biology vol 56 pp 165ndash1852005
[49] Y Jiang and D C Joyce ldquoABA effects on ethylene productionPAL activity anthocyanin and phenolic contents of strawberryfruitrdquo Plant Growth Regulation vol 39 no 2 pp 171ndash174 2003
[50] S Li ldquoTranscriptional control of flavonoid biosynthesis fine-tuning of the MYB-bHLH-WD40 (MBW) complexrdquo PlantSignaling amp Behavior vol 9 Article ID e27522 2014
[51] D R Rudell J K Fellman and J P Mattheis ldquoPreharvest appli-cation of methyl jasmonate to rsquoFujirsquo apples enhances red col-oration and affects fruit size splitting and bitter pit incidencerdquoHortScience vol 40 no 6 pp 1760ndash1762 2005
[52] A Belhadj N Telef C Saigne et al ldquoEffect of methyl jasmonatein combination with carbohydrates on gene expression of PR
proteins stilbene and anthocyanin accumulation in grapevinecell culturesrdquo Plant Physiology and Biochemistry vol 46 no 4pp 493ndash499 2008
[53] E Loreti G Povero G Novi C Solfanelli A Alpi and PPerata ldquoGibberellins jasmonate and abscisic acid modulate thesucrose-induced expression of anthocyanin biosynthetic genesin Arabidopsisrdquo New Phytologist vol 179 no 4 pp 1004ndash10162008
[54] H S Seo J T Song J-J Cheong et al ldquoJasmonic acid carboxylmethyltransferase A key enzyme for jasmonate-regulated plantresponsesrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 98 no 8 pp 4788ndash4793 2001
[55] M T McDowell and J C Lagarias ldquoPurification and biochemi-cal properties of phytochromobilin synthase from etiolated oatseedlingsrdquo Plant Physiology vol 126 no 4 pp 1546ndash1554 2001
[56] G-Q Fu S Xu Y-J Xie et al ldquoMolecular cloning characteri-zation and expression of an alfalfa (Medicago sativa L) hemeoxygenase-1 gene MsHO1 which is pro-oxidants-regulatedrdquoPlant Physiology and Biochemistry vol 49 no 7 pp 792ndash7992011
[57] X-Y Chen X Ding S Xu et al ldquoEndogenous hydrogen per-oxide plays a positive role in the upregulation of heme oxy-genase and acclimation to oxidative stress in wheat seedlingleavesrdquo Journal of Integrative Plant Biology vol 51 no 10 pp951ndash960 2009
[58] J Shin E Park and G Choi ldquoPIF3 regulates anthocyaninbiosynthesis in an HY5-dependent manner with both factorsdirectly binding anthocyanin biosynthetic gene promoters inArabidopsisrdquo Plant Journal vol 49 no 6 pp 981ndash994 2007
[59] S Wang D Pan X Lv et al ldquoProteomic approach reveals thatstarch degradation contributes to anthocyanin accumulation intuberous root of purple sweet potatordquo Journal of Proteomics vol143 pp 298ndash305 2016
[60] H-F Jia C-L Li Y-M Chai Y Xing and Y Shen ldquoSucrosepromotes strawberry fruit ripening by stimulation of abscisicacid biosynthesisrdquo Pakistan Journal of Botany vol 45 no 1 pp169ndash176 2013
[61] W Xu H Peng T Yang et al ldquoEffect of calcium on strawberryfruit flavonoid pathway gene expression and anthocyanin accu-mulationrdquo Plant Physiology amp Biochemistry vol 82 pp 289ndash298 2014
[62] Z-W-SWan and Z Singh ldquoExogenous application of prohexa-dione-calcium promotes fruit colour development of lsquoCrippsPinkrsquo applerdquo Acta Horticulturae vol 1012 pp 219ndash225 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
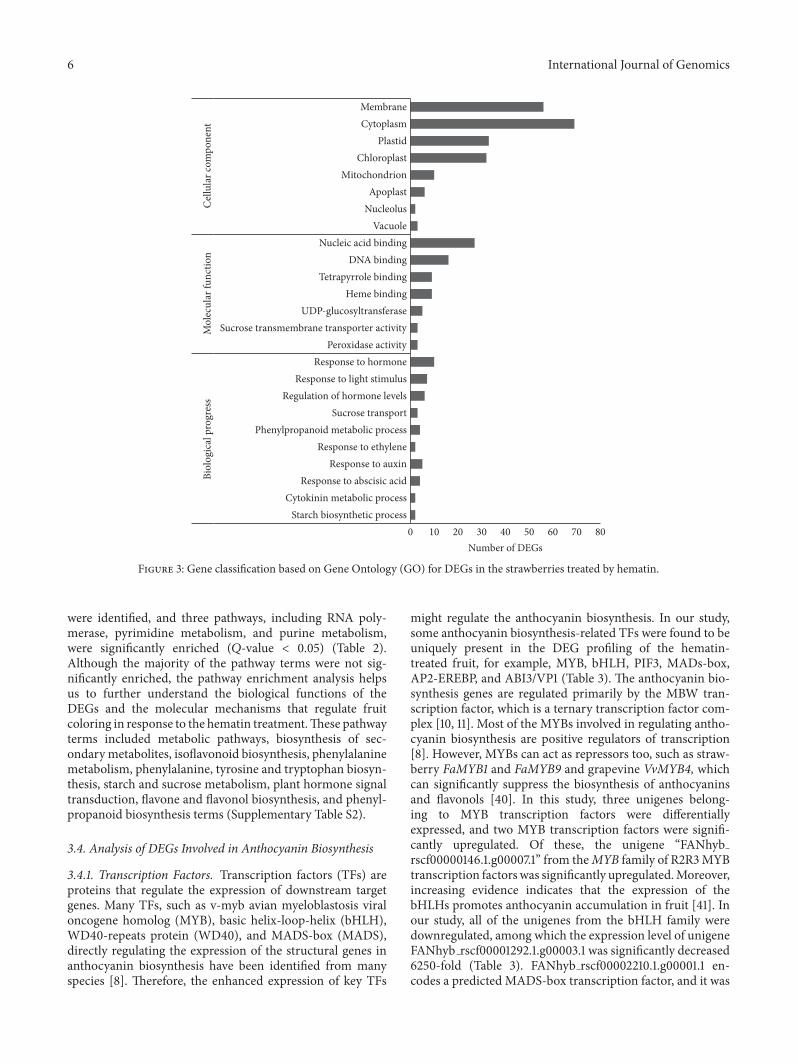
6 International Journal of Genomics
0 10 20 30 40 50 60 70 80Starch biosynthetic process
Cytokinin metabolic processResponse to abscisic acid
Response to auxinResponse to ethylene
Phenylpropanoid metabolic processSucrose transport
Regulation of hormone levelsResponse to light stimulus
Response to hormonePeroxidase activity
Sucrose transmembrane transporter activityUDP-glucosyltransferase
Heme bindingTetrapyrrole binding
DNA bindingNucleic acid binding
VacuoleNucleolusApoplast
MitochondrionChloroplast
PlastidCytoplasmMembrane
Biol
ogic
al p
rogr
ess
Mol
ecul
ar fu
nctio
nC
ellu
lar c
ompo
nent
Number of DEGs
Figure 3 Gene classification based on Gene Ontology (GO) for DEGs in the strawberries treated by hematin
were identified and three pathways including RNA poly-merase pyrimidine metabolism and purine metabolismwere significantly enriched (119876-value lt 005) (Table 2)Although the majority of the pathway terms were not sig-nificantly enriched the pathway enrichment analysis helpsus to further understand the biological functions of theDEGs and the molecular mechanisms that regulate fruitcoloring in response to the hematin treatmentThese pathwayterms included metabolic pathways biosynthesis of sec-ondary metabolites isoflavonoid biosynthesis phenylalaninemetabolism phenylalanine tyrosine and tryptophan biosyn-thesis starch and sucrose metabolism plant hormone signaltransduction flavone and flavonol biosynthesis and phenyl-propanoid biosynthesis terms (Supplementary Table S2)
34 Analysis of DEGs Involved in Anthocyanin Biosynthesis
341 Transcription Factors Transcription factors (TFs) areproteins that regulate the expression of downstream targetgenes Many TFs such as v-myb avian myeloblastosis viraloncogene homolog (MYB) basic helix-loop-helix (bHLH)WD40-repeats protein (WD40) and MADS-box (MADS)directly regulating the expression of the structural genes inanthocyanin biosynthesis have been identified from manyspecies [8] Therefore the enhanced expression of key TFs
might regulate the anthocyanin biosynthesis In our studysome anthocyanin biosynthesis-related TFs were found to beuniquely present in the DEG profiling of the hematin-treated fruit for example MYB bHLH PIF3 MADs-boxAP2-EREBP and ABI3VP1 (Table 3) The anthocyanin bio-synthesis genes are regulated primarily by the MBW tran-scription factor which is a ternary transcription factor com-plex [10 11] Most of the MYBs involved in regulating antho-cyanin biosynthesis are positive regulators of transcription[8] However MYBs can act as repressors too such as straw-berry FaMYB1 and FaMYB9 and grapevine VvMYB4 whichcan significantly suppress the biosynthesis of anthocyaninsand flavonols [40] In this study three unigenes belong-ing to MYB transcription factors were differentiallyexpressed and two MYB transcription factors were signifi-cantly upregulated Of these the unigene ldquoFANhybrscf000001461g000071rdquo from theMYB family of R2R3MYBtranscription factors was significantly upregulatedMoreoverincreasing evidence indicates that the expression of thebHLHs promotes anthocyanin accumulation in fruit [41] Inour study all of the unigenes from the bHLH family weredownregulated among which the expression level of unigeneFANhyb rscf000012921g000031 was significantly decreased6250-fold (Table 3) FANhyb rscf000022101g000011 en-codes a predicted MADS-box transcription factor and it was
International Journal of Genomics 7
Control Hematin-treated
FANhyb_icon00002410_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_rscf000049691g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00027696_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00004395_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_rscf000063641g000011
lowastlowast
lowast
Control Hematin-treated
FANhyb_rscf000001411g000161
Control Hematin-treated
FANhyb_icon00006295_a1g000011lowastlowast
Control Hematin-treated
FANhyb_rscf000031551g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00001444_a1g000011
lowastlowast
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
0
05
1
15
2
25
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
005
115
225
335
445
5
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
Control Hematin-treated
FANhyb_rscf000023111g000011
lowastlowast
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
Control Hematin-treated
FANhyb_rscf000002951g000031
lowastlowast
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
Figure 4 qRT-PCR validation of the RNA-Seq based gene expression The values indicate means of three biological replicates plusmn SD Starindicates that the expression level is significantly different between the hematin-treated and the control group (lowast119901 lt 005 lowastlowast119901 lt 001)
8 International Journal of Genomics
Table 2 Pathway enrichment analysis of differentially expressed genes
Pathway DEGs with pathwayannotation (451)
All genes with pathwayannotation (15877)
Corrected 119901 value 119876-value Pathway ID
RNA polymerase 16 183 688119864 minus 5 00068107 ko03020Pyrimidine metabolism 20 317 00007651746 00320 ko00240Purine metabolism 21 347 00009710371 0032044224 ko00230
significantly downregulated There were three unigenes inthe AP2-EREBP family with only one upregulated and theothers downregulated These results suggest that exogenoushematin is involved in developmental transcriptionalregulation of anthocyanin biosynthesis
342 DEGs in Anthocyanin Biosynthesis The anthocyaninbiosynthetic pathway has been studied intensively Theanthocyanin biosynthetic pathway via the phenylpropanoidpathway is well known [8] In this study the transcriptomedata showed a significant increase in the expression of genesin the anthocyanin biosynthesis pathway when exposedto exogenous hematin The structural genes involved inanthocyanin biosynthesis including DFR LAR and UDP-glycosyltransferase exhibited significant differential expres-sion in response to the hematin treatment (Table 4) Forexample two DFR genes and one LAR gene were signifi-cantly upregulated (the expression increased 18- 30- and3-fold in the hematin-treated fruit resp) In addition allsix genes encodingUDP-glycosyltransferasewere significantlyupregulated Among these the expression of one gene(Fanhy icon15742070o1g000011) was upregulated 18305-fold The results suggest that exogenous hematin may havean important role in promoting anthocyanin biosynthesis
343 DEGs in Hormone Signaling Transduction in Responseto Hematin Auxin has been shown to negatively regulatethe expression of the anthocyanin biosynthesis genes [42] Inthis study there were three unigenes involved in the auxinsignaling pathway and all were downregulated includingone gene encoding auxin efflux carrier component 1-like(AUX1-like) and two genes encoding auxin response factors(ARF) These results suggest that hematin promotes antho-cyanin accumulation by regulating auxin in the colorationprocess of strawberries It has shown that auxin suppressesanthocyanin biosynthesis in the red-fleshed apple callus [43]Endogenous expression of auxin [44 45] has been foundto impede anthocyanin accumulation in strawberries Inadditional cytokinins are also known to play an importantrole in anthocyanin biosynthesis It has shown to enhanceanthocyanin accumulation in Zea mays and regulate antho-cyanin production and composition in suspension culturesof strawberry cells [46] In this study two unigenes relatedto the cytokinin signaling pathway were identified and theywere downregulated as well One gene was identified ascytokinin dehydrogenase 5-like which catalyzes the irre-versible degradation of cytokinins This result indicates thathematin promotes anthocyanin accumulation by regulatingthe cytokinin signaling pathway
Extensive researches have shown that ABA plays animportant role in the regulation of anthocyanin biosynthesisin nonclimacteric fruit [47] In this study a total of five uni-genes related to the ABA signaling pathway were identifiedamong which two were upregulated and three were down-regulated Two genes which are involved in the abscisic acidresponse were upregulated These two unigenes encode 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) and ABA overlysensitive 5 (ABO5) respectively ABA is synthesized fromcarotenoids via several enzymatic reactions in the plastidThe rate-limiting step in these reactions is catalyzed by 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) [48] Mutations inthe FaNCED1 gene result in colorless strawberry fruit whichcan become colored through the application of exogenousABA [12] Another unigene was identified to encode abscisicacid 81015840-hydroxylase 1-like a key enzyme in the oxidativecatabolism of abscisic acid which was downregulated Appli-cation of exogenous ABA regulates phenylalanine ammonialyase activity and increases the phenolic and anthocyanincontent of strawberry fruit [12 49] Our results suggestthat hematin promotes ABA biosynthesis and inhibits ABAdisintegration in the hematin-treated fruit
In Arabidopsis JAs can affect anthocyanin accumulationvia the interaction of negative regulators with the MBWcomplex of transcription factors involved in anthocyaninbiosynthesis [50] Preharvest application of JA to ldquoFujirdquo applesenhances red coloration [51] JA vapor treatment can alsoenhance anthocyanins in strawberry fruit [52] In this studyjasmonate O-methyltransferase-like exhibited significantlydifferential expression Jasmonate O-methyltransferase-likeis also known to be a key enzyme for jasmonate-regulatedplant responses to stimuli [53 54] These results suggest thathematin increases the activity of the JA-regulated antho-cyanin biosynthesis
344 DEGs in the Phytochrome Signaling Pathway in Responseto Hematin Phytochromes that act as photoreceptors playan important role in anthocyanin regulation [15] Phy-tochromes are homodimeric chromoproteins where eachholophytochrome is composed of a phytochrome protein(apophytochrome) covalently bound to a linear tetrapyrrolePΦB HO1 is crucial to this process and acts as a rate-limiting enzyme in the biosynthesis of PΦB [55] Exogenoushematin can also induce HO-1 expression in many plants[56 57] In this study we found exogenous hematin signif-icantly increased the expression of FaHO1 (Table 5) Thisresult implies that hematin can promote PΦB biosynthesisRecently it has been reported that hematin could inducethe accumulation of far-red phytochrome and phytochrome
International Journal of Genomics 9
Table 3 DEGs acting as transcription factors in response to exogenous hematin
Annotation transcription factors Gene IDReal
fold-changevalues
Upregulationdownregulation FDR
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb icon00006295 a1g000011 938 Up 120119864 minus 05
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb rscf000031551g000011 4 Down 480119864 minus 16
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL5-like
FANhyb rscf000000291g000101 4 Down 320119864 minus 08
MYBdomain protein 66 FANhyb rscf000001461g000071 654 Up 780119864 minus 07MYB-related protein 3R-1-like partial FANhyb rscf000001411g000161 282 Up 830119864 minus 04PREDICTED transcription factorGAMYB-like FANhyb rscf000006491g000061 303 Down 150119864 minus 13
PREDICTED probable WRKYtranscription factor 53-like FANhyb rscf000019731g000021 217 Down 200119864 minus 08
Transcription factor bHLH80-like FANhyb rscf000012921g000031 6250 Down 150119864 minus 06Transcription factor bHLH70-like FANhyb rscf000002951g000031 526 Down 660119864 minus 12Transcription factor ICE1-like FANhyb rscf000001701g000061 244 Down 450119864 minus 18C2H2-like zinc finger protein FANhyb rscf000011431g000021 282 Up 590119864 minus 04Zinc finger protein MAGPIE-like FANhyb rscf000030241g000011 204 Up 150119864 minus 04PREDICTED axial regulator YABBY5-like FANhyb rscf000016671g000011 417 Down 730119864 minus 05
PREDICTED axial regulator YABBY1-like FANhyb rscf000068991g000011 357 Down 620119864 minus 06
PREDICTED dof zinc finger proteinDOF37-like isoform 1 FANhyb rscf000068561g000011 263 Down 280119864 minus 04
ZF-HD homeobox protein FANhyb rscf000000441g000221 385 Down 420119864 minus 08ZF-HD homeobox proteinAt4g24660-like FANhyb rscf000052411g000011 357 Down 470119864 minus 09
PREDICTED B3 domain-containingprotein REM14-like FANhyb rscf000010091g000021 243 Up 530119864 minus 04
PREDICTED B3 domain-containingtranscription factor ABI3-like FANhyb rscf000012711g000021 455 Down 130119864 minus 28
Growth-regulating factor 6 FANhyb rscf000000241g000291 345 Down 330119864 minus 34Growth-regulating factor 5 FANhyb icon00009704 a1g000011 27 Down 190119864 minus 06GATA type zinc finger transcriptionfactor family protein FANhyb rscf000003931g000131 1111 Down 110119864 minus 05
PREDICTED zinc finger CCCHdomain-containing protein 2-like FANhyb icon00006770 a1g000011 313 Down 680119864 minus 49
PREDICTED E2F transcriptionfactor-like E2FE-like FANhyb icon00002702 a1g000011 278 Down 240119864 minus 04
PREDICTED uncharacterized protein FANhyb rscf000010661g000011 263 Down 490119864 minus 06PREDICTED nuclear transcriptionfactor Y subunit A-3-like isoform 1 FANhyb rscf000000081g000351 385 Down 500119864 minus 20
PREDICTED homeobox-leucinezipper protein ATHB-8-like FANhyb rscf000000151g000051 263 Down 100119864 minus 06
PREDICTED MADS-boxtranscription factor 18-like FANhyb rscf000003231g000161 243 Down 180119864 minus 06
PREDICTED zinc finger protein132-like isoform 1 FANhyb rscf000070151g000011 243 Down 180119864 minus 08
PREDICTED transcription factorPIF3-like FANhyb rscf000006691g000021 346 Up 140119864 minus 07
10 International Journal of Genomics
Table 4 Regulation of the DEGs in anthocyanin biosynthesis in response to exogenous hematin
Genes in anthocyaninbiosynthesis Gene ID
Realfold-change
valuesRegulation level FDR
Dihydroflavonol-4-reductase FANhyb icon00002410 a1g000011 3091 Up 910119864 minus 06
Flavonol synthase FANhyb icon00015354 a1g000011 345 down 180119864 minus 05Isoflavone reductasehomolog FANhyb icon20341135 s1g000011 334 Up 750119864 minus 14
UDP-glycosyltransferase FANhyb rscf000049691g000011 307 Up 640119864 minus 08UDP-glycosyltransferaseactivity FANhyb icon15742070 o1g000011 18 30563 Up 510119864 minus 09
UDP-glycosyltransferase73C1 FANhyb icon00027696 a1g000011 298 Up 560119864 minus 04
UDP-glycosyltransferaseactivity FANhyb icon00008064 a1g000011 249 Up 310119864 minus 04
PREDICTEDUDP-glucose flavonoid3-O-glucosyltransferase7-like
FANhyb rscf000055631g000011 212 Up 110119864 minus 19
PREDICTEDUDP-glycosyltransferase76F1-like
FANhyb rscf000021771g000031 2 Up 720119864 minus 29
A transcripts in the etiolated leaves of wheat seedling phy-tochromes [26] However our study shows that phytochromeA was not differentially expressed It is probably becausethe hematin-treated strawberries were not shaded during theexperiment while the function of phytochrome A is light-dependent In addition we found that a downstream gene ofphytochrome PIF3 (FANhyb rscf000006691g000021) wassignificantly upregulated PIF3 is thought to be a positive reg-ulator of phytochrome B mediated by light-dependent signaltransduction [58]These results suggest that the phytochromepathways may be involved in the anthocyanin biosynthesispromoted by hematin
345 DEGs in Starch and Sucrose Degradation Proteomicapproaches have revealed that starch degradation contributesto anthocyanin accumulation in tuberous roots of the purplesweet potato variety [59] Starch and sucrose biosynthesis isimportant for anthocyanin accumulation in strawberry fruit[60] In this study we found the DEGs involved in starchand sucrose synthesis in the hematin-treated fruit were alldownregulated (Table 5) For example the expression of theunigenes encoding soluble starch synthase 3 sucrose trans-port protein SUC2-like and a probable sucrose-phosphatesynthase 4-like enzyme were decreased 23- 46- 32- and2-fold respectively This suggests that exogenous hematinregulates the biosynthesis of starch and sucrose and henceaffects fruit coloring
346 DEGs in the Calcium Pathway in Response to HematinCalcium can also increase the transcription levels of keystructural genes F3H DFR ANT and UFGT in the whitestage of strawberries [61] In this study we found thatthree DEGs in calcium biosynthesis and transport were
downregulated in the hematin-treated fruit (Table 5) Onlythe unigene encoding predicted cationcalcium exchanger5-like was upregulated The unigenes encoding a calpain-type cysteine protease and a predicted calcium-binding pro-tein PBP1-like were downregulated respectively Exogenousapplication of calcium can promote apple coloring [62] inaddition to variation in anthocyanin content [61] Our resultsindicate that the biosynthesis and transport of calcium areinvolved in the development of coloring in hematin-treatedstrawberries
35 Validation of Selected DEGs by qRT-PCR To validate theexpression of the DEGs obtained from RNA-Seq 11 DEGswere selected for qRT-PCR including structural genes (DFRand UDFGs) transcription factor genes (MYB and bHLH)and phytochrome chromophore-related gene (HO-1) Theprimers used for qRT-PCR are listed in Supplementary TableS1 The qRT-PCR results were consistent with the RNA-Seqdata (Figure 4) except for FANhyb rscf000001411g000161which had a higher log
2ratio (hematin-treatedcontrol) in
the transcriptome data than in qRT-PCR These results indi-cate the RNA-Seq data from the strawberry transcriptome isreproducible and accurate
4 Conclusions
In this study the anthocyanin content in the strawberryfruit was elevated by the application of exogenous hematinThis is the first report that hematin could increase theanthocyanin production in fruit Furthermore we exploredthe effects of the exogenous hematin on metabolic path-ways using genome-wide transcriptome analysis The resultsindicate that the expression levels of many genes involved
International Journal of Genomics 11
Table 5 Other DEGs identified in anthocyanin biosynthesis-related pathways
Anthocyaninbiosynthesis-relatedpathways
Annotation genes Gene IDReal fold-changevalues
Upregulationdownregulation FDR
Calcium ion binding
Cationcalciumexchanger 5-like
FANhyb rscf000019061g000011 203 Up 64119864 minus 06
Calpain-type cysteineprotease family
FANhyb icon00011755 a1g000011 345 Down 18119864 minus 09
Calcium-bindingprotein PBP1-like
FANhyb rscf000007501g000081 323 Down 97119864 minus 04
Calpain-type cysteineprotease family
FANhyb icon00023658 a1g000011 233 Down 60119864 minus 08
CytokininCytokinin
dehydrogenase 5-likeFANhyb iscf00325393 1 s1g000011 667 Down 62119864 minus 06
Cytochrome P450714A1
FANhyb rscf000005921g000031 217 Down 98119864 minus 10
JasmonateJasmonate
O-methyltransferase-like
FANhyb rscf000000041g000131 201 Up 70119864 minus 04
Abscisic acid
Protein ABSCISICACID-INSENSITIVE
5-likeFANhyb rscf000021641g000011 4 Down 16119864 minus 18
ABA overly sensitive 5 FANhyb icon00051144 a1g000011 291 Up 24119864 minus 059-cis-
epoxycarotenoiddioxygenase NCED1
FANhyb icon18399909 o1g000011 441 Up 43119864 minus 06
Abscisic acid81015840-hydroxylase 1-like
FANhyb icon00000938 a1g000011 256 Down 14119864 minus 06
Abscisic acid receptorPYR1-like
FANhyb icon00020426 a1g000011 208 Down 30119864 minus 06
Auxin
Auxin efflux carriercomponent 1-like
FANhyb rscf000010081g000031 384 Down 23119864 minus 06
Auxin response factor8
FANhyb rscf000013061g000021 263 Down 61119864 minus 17
Auxin response factor17-like
FANhyb rscf000060741g000011 217 Down 11119864 minus 36
Starch and sucrose
Glycosyltransferasefamily 35
FANhyb rscf000000341g000081 384 Down 11119864 minus 05
Soluble starchsynthase 3
FANhyb rscf000000451g000041 232 Down 38119864 minus 12
Sucrose transportprotein SUC2-like
FANhyb icon00036727 a1g000011 455 Down 98119864 minus 04
Sucrose transportprotein SUC2-like
FANhyb rscf000007551g000021 323 Down 66119864 minus 10
Probablesucrose-phosphatesynthase 4-like
FANhyb rscf000013501g000021 213 Down 11119864 minus 14
Phytochrome
PREDICTEDtranscription factor
PIF3-likeFANhyb rscf000006691g000021 346 up 14119864 minus 07
Phytochrome E-like FANhyb rscf000004361g000041 4 Down 33119864 minus 05
Heme oxygenase 1 FANhyb icon00004395 a1g000011 258 27119864 minus 07 27119864 minus 07
12 International Journal of Genomics
Hematin
Endoplasmicreticulum
WD40 bLHLMYB
Structural genes
TFs
Nuclear
JA
ABA Sucrose
Plastid
G-cis-neoxanthin
Apophytochrome
StarchHeme
BVSucroseABA
Holophytochrome
PfrPIF3
Flavonoids Anthocyanins
Vacuole
Cytoplasm
Cell wall
①
②③
④⑤
⑥
⑦
PΦB
PΦB
CTKAuxin
Ca2+
Figure 5 Genes and biological pathways that were involved in regulation of anthocyanin accumulation promoted by hematin Outermostbox represents the cell wall Bilayer oval shape represents the plastidThe red-brown ring and network represent the nuclear and endoplasmicreticulum respectively The red crescent represents the vacuole The box in the nucleus represents transcription factors (TFs) Yellow MYBgreenWD40 and red bHLH represent a ternary transcription factor complex which transcribes anthocyanin biosynthesis genes The circlednumbers represent biological pathways involved in the regulation of anthocyanin accumulation promoted by hematin A represents thephytochrome regulation pathwayB represents the starch and sucrose pathwayCDE andF represent phytohormones ABA JA AuxinandCTK regulation pathways respectivelyG represents theCa2+ regulation pathway Pfr represents the far-red phytochrome PIF3 representsphytochrome-interacting factor Each anthocyanin biosynthesis regulatory pathway related gene is in Table 5
in anthocyanin biosynthesis were significantly altered withthe hematin treatment This suggests that the physiologicalprocess of fruit color development is regulated through com-plex interactions among anthocyanin biosynthesis pathwaysplant hormone signal transduction pathways phytochromesignal transduction pathways starch and sugar metabolicpathways calcium pathways and transcription factors (Fig-ure 5) This study adds to the in-depth understanding of thefruit coloration process in strawberry
Competing Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contributions
Jianwei Gao and Liangju Wang conceived and designed theresearch Yi Li Huayin Li Fengde Wang Jingjuan Li andYihui Zhang conducted experiments Yi Li analyzed the dataYi Li and Fengde Wang wrote the manuscript All authorsread and approved the manuscript Yi Li and Huayin Licontributed equally to this work
Acknowledgments
This work was supported by the Modern Agricultural Indus-trial Technology System Funding of Shandong ProvinceChina (SDAIT-02-022-04) the China agriculture researchsystem (CARS-25) and the National Science Foundation ofChina (31401820)
References
[1] K Aaby SMazur A Nes andG Skrede ldquoPhenolic compoundsin strawberry (Fragaria x ananassa Duch) fruits compositionin 27 cultivars and changes during ripeningrdquo Food Chemistryvol 132 no 1 pp 86ndash97 2012
[2] T A Colquhoun L A Levin H RMoskowitz VMWhitakerD G Clark and K M Folta ldquoFraming the perfect strawberryan exercise in consumer-assisted selection of fruit cropsrdquoJournal of Berry Research vol 2 no 1 pp 45ndash61 2012
[3] F Giampieri J M Alvarez-Suarez L Mazzoni et al ldquoThepotential impact of strawberry on human healthrdquo NaturalProduct Research vol 27 no 4 pp 448ndash455 2013
[4] Y S Velioglu GMazza L Gao and B D Oomah ldquoAntioxidantactivity and total phenolics in selected fruits vegetables and
International Journal of Genomics 13
grain productsrdquo Journal of Agricultural amp Food Chemistry vol46 no 10 pp 4113ndash4117 1998
[5] S S Hassellund A Flaa S E Kjeldsen et al ldquoEffects of antho-cyanins on cardiovascular risk factors and inflammation inpre-hypertensive men a double-blind randomized placebo-controlled crossover studyrdquo Journal of Human Hypertensionvol 27 no 2 pp 100ndash106 2013
[6] K J Meyers C B Watkins M P Pritts and R H Liu ldquoAnti-oxidant and antiproliferative activities of strawberriesrdquo Journalof Agricultural amp Food Chemistry vol 51 no 23 pp 6887ndash68922003
[7] S Zafra-Stone T YasminM Bagchi AChatterjee J AVinsonand D Bagchi ldquoBerry anthocyanins as novel antioxidants inhuman health and disease preventionrdquo Molecular Nutrition ampFood Research vol 51 no 6 pp 675ndash683 2007
[8] L Jaakola ldquoNew insights into the regulation of anthocyanin bio-synthesis in fruitsrdquoTrends in Plant Science vol 18 no 9 pp 477ndash483 2013
[9] K Springob J-I Nakajima M Yamazaki and K Saito ldquoRecentadvances in the biosynthesis and accumulation of antho-cyaninsrdquo Natural Product Reports vol 20 no 3 pp 288ndash3032003
[10] A Gonzalez M Zhao J M Leavitt and A M Lloyd ldquoRegu-lation of the anthocyanin biosynthetic pathway by the TTG1bHLHMyb transcriptional complex in Arabidopsis seedlingsrdquoPlant Journal vol 53 no 5 pp 814ndash827 2008
[11] K Petroni and C Tonelli ldquoRecent advances on the regulationof anthocyanin synthesis in reproductive organsrdquo Plant Sciencevol 181 no 3 pp 219ndash229 2011
[12] H-F Jia Y-M Chai C-L Li et al ldquoAbscisic acid plays animportant role in the regulation of strawberry fruit ripeningrdquoPlant Physiology vol 157 no 1 pp 188ndash199 2011
[13] C Merchante J G Vallarino S Osorio et al ldquoEthylene isinvolved in strawberry fruit ripening in an organ-specific man-nerrdquo Journal of Experimental Botany vol 64 no 14 pp 4421ndash4439 2013
[14] A Maier A Schrader L Kokkelink et al ldquoLight and the E3ubiquitin ligase COP1SPA control the protein stability of theMYB transcription factors PAP1 and PAP2 involved in antho-cyanin accumulation in Arabidopsisrdquo Plant Journal vol 74 no4 pp 638ndash651 2013
[15] H Lange W Shropshire and H Mohr ldquoAn analysis of phyto-chrome-mediated anthocyanin synthesisrdquo Plant Physiology vol47 no 5 pp 649ndash655 1971
[16] T D Elich and J C Lagarias ldquoPhytochrome chromophore bio-synthesis both 5-aminolevulinic acid and biliverdin overcomeinhibition by gabaculine in etiolated Avena sativa L seedlingsrdquoPlant Physiology vol 84 no 2 pp 304ndash310 1987
[17] Y Han W Xuan T Yu et al ldquoExogenous hematin alleviatesmercury-induced oxidative damage in the roots of Medicagosativardquo Journal of Integrative Plant Biology vol 49 no 12 pp1703ndash1713 2007
[18] M Koornneef J Cone R Dekens E OrsquoHerne-Robers CSpruit and R Kendrick ldquoPhotomorphogenic responses of longhypocotyl mutants of tomatordquo Journal of Plant Physiology vol120 no 2 pp 153ndash165 1985
[19] M J Terry and R E Kendrick ldquoFeedback inhibition of chloro-phyll synthesis in the phytochrome chromophore-deficientaurea and yellow-green-2 mutants of tomatordquo Plant Physiologyvol 119 no 1 pp 143ndash152 1999
[20] B M Parks and P H Quail ldquoPhytochrome-deficient hy1 andhy2 long hypocotyl mutants of Arabidopsis are defective inphytochrome chromophore biosynthesisrdquo Plant Cell vol 3 no11 pp 1177ndash1186 1991
[21] M J Terry ldquoPhytochrome chromophore-deficient mutantsrdquoPlant Cell amp Environment vol 20 no 6 pp 740ndash745 1997
[22] T Muramoto T Kohchi A Yokota I Hwang and H MGoodman ldquoThe Arabidopsis photomorphogenic mutant hy1 isdeficient in phytochrome chromophore biosynthesis as a resultof a mutation in a plastid heme oxygenaserdquo Plant Cell vol 11no 3 pp 335ndash347 1999
[23] M D Maines ldquoThe heme oxygenase system a regulator ofsecond messenger gasesrdquo Annual Review of Pharmacology andToxicology vol 37 no 1 pp 517ndash554 1997
[24] W Xuan F-Y Zhu S Xu et al ldquoThe heme oxygenasecarbonmonoxide system is involved in the auxin-induced cucumberadventitious rooting processrdquo Plant Physiology vol 148 no 2pp 881ndash893 2008
[25] R Amooaghaie F Tabatabaei and A-M Ahadi ldquoRole ofhematin and sodium nitroprusside in regulating Brassica nigraseed germination under nanosilver and silver nitrate stressesrdquoEcotoxicology and Environmental Safety vol 113 pp 259ndash2702015
[26] Y Liu X Li L Xu and W Shen ldquoDe-etiolation of wheatseedling leaves cross talk between heme oxygenasecarbonmonoxide and nitric oxiderdquo PLoS ONE vol 8 no 12 ArticleID e81470 2013
[27] T Muramoto N Tsurui M J Terry A Yokota and T KohchildquoExpression and biochemical properties of a ferredoxin-dependent heme oxygenase required for phytochrome chro-mophore synthesisrdquo Plant Physiology vol 130 no 4 pp 1958ndash1966 2002
[28] J-W Gao J Liu B Li and Z Li ldquoIsolation and purificationof functional total RNA from blue-grained wheat endospermtissues containing high levels of starches and flavonoidsrdquo PlantMolecular Biology Reporter vol 19 no 2 pp 185ndash186 2001
[29] R Patro S M Mount and C Kingsford ldquoSailfish enablesalignment-free isoform quantification from RNA-seq readsusing lightweight algorithmsrdquoNature Biotechnology vol 32 no5 pp 462ndash464 2014
[30] Y-X Qi Y-B Liu and W-H Rong ldquoRNA-Seq and its applica-tions a new technology for transcriptomicsrdquo Hereditas vol 33no 11 pp 1191ndash1202 2011
[31] C R Genovese N A Lazar and T Nichols ldquoThresholdingof statistical maps in functional neuroimaging using the falsediscovery raterdquo NeuroImage vol 15 no 4 pp 870ndash878 2002
[32] J Ye L Fang H Zheng et al ldquoWEGO a web tool for plottingGO annotationsrdquoNucleic Acids Research vol 34 supplement 2pp W293ndashW297 2006
[33] H Abdi ldquoThe Bonferonni and Sidak corrections for multiplecomparisonrdquo Encyclopedia of Measurement and Statistics vol 3pp 103ndash107 2007
[34] J Li Q Ding F Wang Y Zhang H Li and J Gao ldquoIntegrativeanalysis of mRNA and miRNA expression profiles of thetuberous root development at seedling stages in turnipsrdquo PLoSONE vol 10 no 9 Article ID e0137983 2015
[35] J Bennett D Hondred and J C Register ldquoKeeping qRT-PCRrigorous and biologically relevantrdquo Plant Cell Reports vol 34no 1 pp 1ndash3 2015
[36] F Wang L Li H Li et al ldquoTranscriptome analysis of rosetteand folding leaves in Chinese cabbage using high-throughputRNA sequencingrdquo Genomics vol 99 no 5 pp 299ndash307 2012
14 International Journal of Genomics
[37] T Shi Z Gao L Wang et al ldquoIdentification of differentially-expressed genes associated with pistil abortion in Japaneseapricot by genome-wide transcriptional analysisrdquo PLoS ONEvol 7 no 10 Article ID e47810 2012
[38] F Wang H Li Y Zhang et al ldquoMicroRNA expression analysisof rosette and folding leaves in Chinese cabbage using high-throughput Solexa sequencingrdquo Gene vol 532 no 2 pp 222ndash229 2013
[39] X Gu Y Chen Z Gao Y Qiao and X Wang ldquoTranscriptionfactors and anthocyanin genes related to low-temperature toler-ance in rd29A RdreB1BI transgenic strawberryrdquo Plant Physiol-ogy and Biochemistry vol 89 pp 31ndash43 2015
[40] J G Schaart C Dubos I R De La Fuente et al ldquoIdentificationand characterization of MYB-bHLH-WD40 regulatory com-plexes controlling proanthocyanidin biosynthesis in strawberry(Fragaria times ananassa) fruitsrdquoNew Phytologist vol 197 no 2 pp454ndash467 2013
[41] L Xie Z H Wang X H Cheng J J Gao Z P Zhangand L J Wang ldquo5-Aminolevulinic acid promotes anthocyaninaccumulation in Fuji applesrdquo Plant Growth Regulation vol 69no 3 pp 295ndash303 2013
[42] M Daminato F Guzzo and G Casadoro ldquoASHATTERPROOF-like gene controls ripening in non-climac-teric strawberries and auxin and abscisic acid antagonisticallyaffect its expressionrdquo Journal of Experimental Botany vol 64no 12 pp 3775ndash3786 2013
[43] X-H Ji R Zhang N Wang L Yang and X-S Chen ldquoTran-scriptome profiling reveals auxin suppressed anthocyanin bio-synthesis in red-fleshed apple callus (Malus sieversii f niedzwet-zkyana)rdquo Plant Cell Tissue and Organ Culture vol 123 no 2pp 389ndash404 2015
[44] J Chen LMaoW Lu T Ying and Z Luo ldquoTranscriptome pro-filing of postharvest strawberry fruit in response to exogenousauxin and abscisic acidrdquo Planta vol 243 no 1 pp 183ndash197 2016
[45] G A Martınez A R Chaves and M C Anon ldquoEffect of gib-berellic acid on ripening of strawberry fruits (Fragaria annan-assa Duch)rdquo Journal of Plant Growth Regulation vol 13 no 2pp 87ndash91 1994
[46] P K Das D H Shin S-B Choi S-D Yoo G Choi and Y-IPark ldquoCytokinins enhance sugar-induced anthocyanin biosyn-thesis in Arabidopsisrdquo Molecules amp Cells vol 34 no 1 pp 93ndash101 2012
[47] P McAtee S Karim R Schaffer and K David ldquoA dynamicinterplay between phytohormones is required for fruit develop-ment maturation and ripeningrdquo Frontiers in Plant Science vol4 no 3 article 79 7 pages 2013
[48] E Nambara andAMarion-Poll ldquoAbscisic acid biosynthesis andcatabolismrdquoAnnual Review of Plant Biology vol 56 pp 165ndash1852005
[49] Y Jiang and D C Joyce ldquoABA effects on ethylene productionPAL activity anthocyanin and phenolic contents of strawberryfruitrdquo Plant Growth Regulation vol 39 no 2 pp 171ndash174 2003
[50] S Li ldquoTranscriptional control of flavonoid biosynthesis fine-tuning of the MYB-bHLH-WD40 (MBW) complexrdquo PlantSignaling amp Behavior vol 9 Article ID e27522 2014
[51] D R Rudell J K Fellman and J P Mattheis ldquoPreharvest appli-cation of methyl jasmonate to rsquoFujirsquo apples enhances red col-oration and affects fruit size splitting and bitter pit incidencerdquoHortScience vol 40 no 6 pp 1760ndash1762 2005
[52] A Belhadj N Telef C Saigne et al ldquoEffect of methyl jasmonatein combination with carbohydrates on gene expression of PR
proteins stilbene and anthocyanin accumulation in grapevinecell culturesrdquo Plant Physiology and Biochemistry vol 46 no 4pp 493ndash499 2008
[53] E Loreti G Povero G Novi C Solfanelli A Alpi and PPerata ldquoGibberellins jasmonate and abscisic acid modulate thesucrose-induced expression of anthocyanin biosynthetic genesin Arabidopsisrdquo New Phytologist vol 179 no 4 pp 1004ndash10162008
[54] H S Seo J T Song J-J Cheong et al ldquoJasmonic acid carboxylmethyltransferase A key enzyme for jasmonate-regulated plantresponsesrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 98 no 8 pp 4788ndash4793 2001
[55] M T McDowell and J C Lagarias ldquoPurification and biochemi-cal properties of phytochromobilin synthase from etiolated oatseedlingsrdquo Plant Physiology vol 126 no 4 pp 1546ndash1554 2001
[56] G-Q Fu S Xu Y-J Xie et al ldquoMolecular cloning characteri-zation and expression of an alfalfa (Medicago sativa L) hemeoxygenase-1 gene MsHO1 which is pro-oxidants-regulatedrdquoPlant Physiology and Biochemistry vol 49 no 7 pp 792ndash7992011
[57] X-Y Chen X Ding S Xu et al ldquoEndogenous hydrogen per-oxide plays a positive role in the upregulation of heme oxy-genase and acclimation to oxidative stress in wheat seedlingleavesrdquo Journal of Integrative Plant Biology vol 51 no 10 pp951ndash960 2009
[58] J Shin E Park and G Choi ldquoPIF3 regulates anthocyaninbiosynthesis in an HY5-dependent manner with both factorsdirectly binding anthocyanin biosynthetic gene promoters inArabidopsisrdquo Plant Journal vol 49 no 6 pp 981ndash994 2007
[59] S Wang D Pan X Lv et al ldquoProteomic approach reveals thatstarch degradation contributes to anthocyanin accumulation intuberous root of purple sweet potatordquo Journal of Proteomics vol143 pp 298ndash305 2016
[60] H-F Jia C-L Li Y-M Chai Y Xing and Y Shen ldquoSucrosepromotes strawberry fruit ripening by stimulation of abscisicacid biosynthesisrdquo Pakistan Journal of Botany vol 45 no 1 pp169ndash176 2013
[61] W Xu H Peng T Yang et al ldquoEffect of calcium on strawberryfruit flavonoid pathway gene expression and anthocyanin accu-mulationrdquo Plant Physiology amp Biochemistry vol 82 pp 289ndash298 2014
[62] Z-W-SWan and Z Singh ldquoExogenous application of prohexa-dione-calcium promotes fruit colour development of lsquoCrippsPinkrsquo applerdquo Acta Horticulturae vol 1012 pp 219ndash225 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
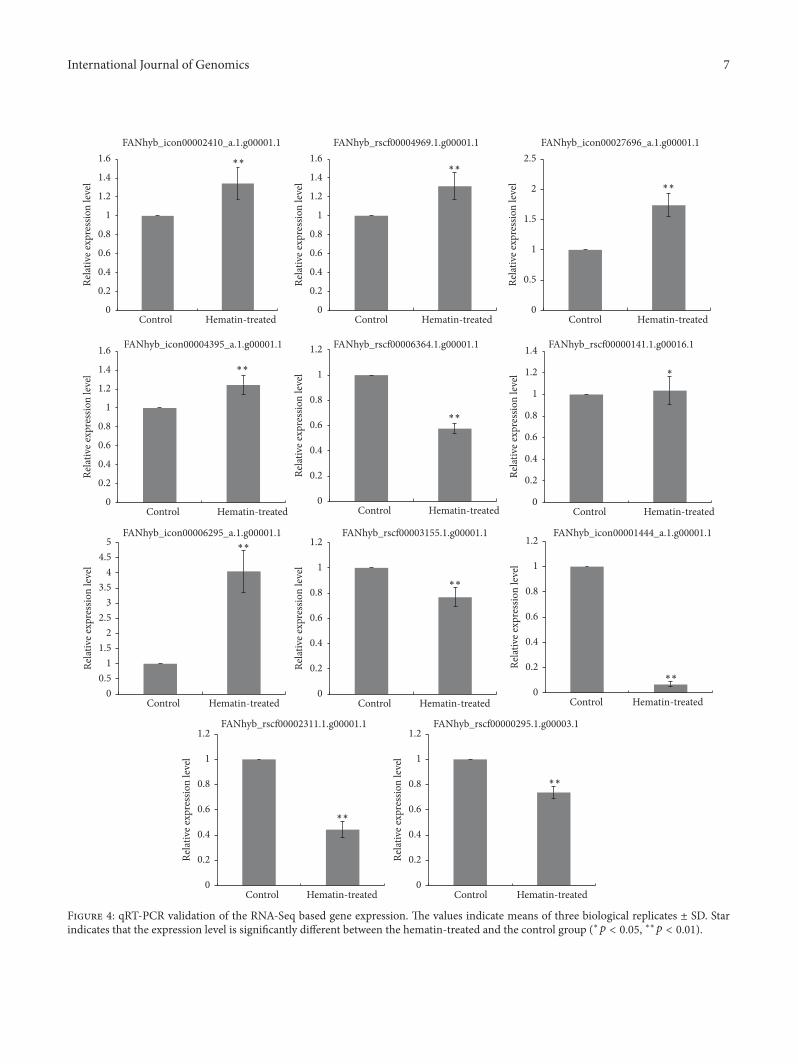
International Journal of Genomics 7
Control Hematin-treated
FANhyb_icon00002410_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_rscf000049691g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00027696_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00004395_a1g000011
lowastlowast
Control Hematin-treated
FANhyb_rscf000063641g000011
lowastlowast
lowast
Control Hematin-treated
FANhyb_rscf000001411g000161
Control Hematin-treated
FANhyb_icon00006295_a1g000011lowastlowast
Control Hematin-treated
FANhyb_rscf000031551g000011
lowastlowast
Control Hematin-treated
FANhyb_icon00001444_a1g000011
lowastlowast
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
0
05
1
15
2
25
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
005
115
225
335
445
5
Relat
ive e
xpre
ssio
n le
vel
0
02
04
06
08
1
12
14
16
Relat
ive e
xpre
ssio
n le
vel
Control Hematin-treated
FANhyb_rscf000023111g000011
lowastlowast
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
Control Hematin-treated
FANhyb_rscf000002951g000031
lowastlowast
0
02
04
06
08
1
12
Relat
ive e
xpre
ssio
n le
vel
Figure 4 qRT-PCR validation of the RNA-Seq based gene expression The values indicate means of three biological replicates plusmn SD Starindicates that the expression level is significantly different between the hematin-treated and the control group (lowast119901 lt 005 lowastlowast119901 lt 001)
8 International Journal of Genomics
Table 2 Pathway enrichment analysis of differentially expressed genes
Pathway DEGs with pathwayannotation (451)
All genes with pathwayannotation (15877)
Corrected 119901 value 119876-value Pathway ID
RNA polymerase 16 183 688119864 minus 5 00068107 ko03020Pyrimidine metabolism 20 317 00007651746 00320 ko00240Purine metabolism 21 347 00009710371 0032044224 ko00230
significantly downregulated There were three unigenes inthe AP2-EREBP family with only one upregulated and theothers downregulated These results suggest that exogenoushematin is involved in developmental transcriptionalregulation of anthocyanin biosynthesis
342 DEGs in Anthocyanin Biosynthesis The anthocyaninbiosynthetic pathway has been studied intensively Theanthocyanin biosynthetic pathway via the phenylpropanoidpathway is well known [8] In this study the transcriptomedata showed a significant increase in the expression of genesin the anthocyanin biosynthesis pathway when exposedto exogenous hematin The structural genes involved inanthocyanin biosynthesis including DFR LAR and UDP-glycosyltransferase exhibited significant differential expres-sion in response to the hematin treatment (Table 4) Forexample two DFR genes and one LAR gene were signifi-cantly upregulated (the expression increased 18- 30- and3-fold in the hematin-treated fruit resp) In addition allsix genes encodingUDP-glycosyltransferasewere significantlyupregulated Among these the expression of one gene(Fanhy icon15742070o1g000011) was upregulated 18305-fold The results suggest that exogenous hematin may havean important role in promoting anthocyanin biosynthesis
343 DEGs in Hormone Signaling Transduction in Responseto Hematin Auxin has been shown to negatively regulatethe expression of the anthocyanin biosynthesis genes [42] Inthis study there were three unigenes involved in the auxinsignaling pathway and all were downregulated includingone gene encoding auxin efflux carrier component 1-like(AUX1-like) and two genes encoding auxin response factors(ARF) These results suggest that hematin promotes antho-cyanin accumulation by regulating auxin in the colorationprocess of strawberries It has shown that auxin suppressesanthocyanin biosynthesis in the red-fleshed apple callus [43]Endogenous expression of auxin [44 45] has been foundto impede anthocyanin accumulation in strawberries Inadditional cytokinins are also known to play an importantrole in anthocyanin biosynthesis It has shown to enhanceanthocyanin accumulation in Zea mays and regulate antho-cyanin production and composition in suspension culturesof strawberry cells [46] In this study two unigenes relatedto the cytokinin signaling pathway were identified and theywere downregulated as well One gene was identified ascytokinin dehydrogenase 5-like which catalyzes the irre-versible degradation of cytokinins This result indicates thathematin promotes anthocyanin accumulation by regulatingthe cytokinin signaling pathway
Extensive researches have shown that ABA plays animportant role in the regulation of anthocyanin biosynthesisin nonclimacteric fruit [47] In this study a total of five uni-genes related to the ABA signaling pathway were identifiedamong which two were upregulated and three were down-regulated Two genes which are involved in the abscisic acidresponse were upregulated These two unigenes encode 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) and ABA overlysensitive 5 (ABO5) respectively ABA is synthesized fromcarotenoids via several enzymatic reactions in the plastidThe rate-limiting step in these reactions is catalyzed by 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) [48] Mutations inthe FaNCED1 gene result in colorless strawberry fruit whichcan become colored through the application of exogenousABA [12] Another unigene was identified to encode abscisicacid 81015840-hydroxylase 1-like a key enzyme in the oxidativecatabolism of abscisic acid which was downregulated Appli-cation of exogenous ABA regulates phenylalanine ammonialyase activity and increases the phenolic and anthocyanincontent of strawberry fruit [12 49] Our results suggestthat hematin promotes ABA biosynthesis and inhibits ABAdisintegration in the hematin-treated fruit
In Arabidopsis JAs can affect anthocyanin accumulationvia the interaction of negative regulators with the MBWcomplex of transcription factors involved in anthocyaninbiosynthesis [50] Preharvest application of JA to ldquoFujirdquo applesenhances red coloration [51] JA vapor treatment can alsoenhance anthocyanins in strawberry fruit [52] In this studyjasmonate O-methyltransferase-like exhibited significantlydifferential expression Jasmonate O-methyltransferase-likeis also known to be a key enzyme for jasmonate-regulatedplant responses to stimuli [53 54] These results suggest thathematin increases the activity of the JA-regulated antho-cyanin biosynthesis
344 DEGs in the Phytochrome Signaling Pathway in Responseto Hematin Phytochromes that act as photoreceptors playan important role in anthocyanin regulation [15] Phy-tochromes are homodimeric chromoproteins where eachholophytochrome is composed of a phytochrome protein(apophytochrome) covalently bound to a linear tetrapyrrolePΦB HO1 is crucial to this process and acts as a rate-limiting enzyme in the biosynthesis of PΦB [55] Exogenoushematin can also induce HO-1 expression in many plants[56 57] In this study we found exogenous hematin signif-icantly increased the expression of FaHO1 (Table 5) Thisresult implies that hematin can promote PΦB biosynthesisRecently it has been reported that hematin could inducethe accumulation of far-red phytochrome and phytochrome
International Journal of Genomics 9
Table 3 DEGs acting as transcription factors in response to exogenous hematin
Annotation transcription factors Gene IDReal
fold-changevalues
Upregulationdownregulation FDR
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb icon00006295 a1g000011 938 Up 120119864 minus 05
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb rscf000031551g000011 4 Down 480119864 minus 16
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL5-like
FANhyb rscf000000291g000101 4 Down 320119864 minus 08
MYBdomain protein 66 FANhyb rscf000001461g000071 654 Up 780119864 minus 07MYB-related protein 3R-1-like partial FANhyb rscf000001411g000161 282 Up 830119864 minus 04PREDICTED transcription factorGAMYB-like FANhyb rscf000006491g000061 303 Down 150119864 minus 13
PREDICTED probable WRKYtranscription factor 53-like FANhyb rscf000019731g000021 217 Down 200119864 minus 08
Transcription factor bHLH80-like FANhyb rscf000012921g000031 6250 Down 150119864 minus 06Transcription factor bHLH70-like FANhyb rscf000002951g000031 526 Down 660119864 minus 12Transcription factor ICE1-like FANhyb rscf000001701g000061 244 Down 450119864 minus 18C2H2-like zinc finger protein FANhyb rscf000011431g000021 282 Up 590119864 minus 04Zinc finger protein MAGPIE-like FANhyb rscf000030241g000011 204 Up 150119864 minus 04PREDICTED axial regulator YABBY5-like FANhyb rscf000016671g000011 417 Down 730119864 minus 05
PREDICTED axial regulator YABBY1-like FANhyb rscf000068991g000011 357 Down 620119864 minus 06
PREDICTED dof zinc finger proteinDOF37-like isoform 1 FANhyb rscf000068561g000011 263 Down 280119864 minus 04
ZF-HD homeobox protein FANhyb rscf000000441g000221 385 Down 420119864 minus 08ZF-HD homeobox proteinAt4g24660-like FANhyb rscf000052411g000011 357 Down 470119864 minus 09
PREDICTED B3 domain-containingprotein REM14-like FANhyb rscf000010091g000021 243 Up 530119864 minus 04
PREDICTED B3 domain-containingtranscription factor ABI3-like FANhyb rscf000012711g000021 455 Down 130119864 minus 28
Growth-regulating factor 6 FANhyb rscf000000241g000291 345 Down 330119864 minus 34Growth-regulating factor 5 FANhyb icon00009704 a1g000011 27 Down 190119864 minus 06GATA type zinc finger transcriptionfactor family protein FANhyb rscf000003931g000131 1111 Down 110119864 minus 05
PREDICTED zinc finger CCCHdomain-containing protein 2-like FANhyb icon00006770 a1g000011 313 Down 680119864 minus 49
PREDICTED E2F transcriptionfactor-like E2FE-like FANhyb icon00002702 a1g000011 278 Down 240119864 minus 04
PREDICTED uncharacterized protein FANhyb rscf000010661g000011 263 Down 490119864 minus 06PREDICTED nuclear transcriptionfactor Y subunit A-3-like isoform 1 FANhyb rscf000000081g000351 385 Down 500119864 minus 20
PREDICTED homeobox-leucinezipper protein ATHB-8-like FANhyb rscf000000151g000051 263 Down 100119864 minus 06
PREDICTED MADS-boxtranscription factor 18-like FANhyb rscf000003231g000161 243 Down 180119864 minus 06
PREDICTED zinc finger protein132-like isoform 1 FANhyb rscf000070151g000011 243 Down 180119864 minus 08
PREDICTED transcription factorPIF3-like FANhyb rscf000006691g000021 346 Up 140119864 minus 07
10 International Journal of Genomics
Table 4 Regulation of the DEGs in anthocyanin biosynthesis in response to exogenous hematin
Genes in anthocyaninbiosynthesis Gene ID
Realfold-change
valuesRegulation level FDR
Dihydroflavonol-4-reductase FANhyb icon00002410 a1g000011 3091 Up 910119864 minus 06
Flavonol synthase FANhyb icon00015354 a1g000011 345 down 180119864 minus 05Isoflavone reductasehomolog FANhyb icon20341135 s1g000011 334 Up 750119864 minus 14
UDP-glycosyltransferase FANhyb rscf000049691g000011 307 Up 640119864 minus 08UDP-glycosyltransferaseactivity FANhyb icon15742070 o1g000011 18 30563 Up 510119864 minus 09
UDP-glycosyltransferase73C1 FANhyb icon00027696 a1g000011 298 Up 560119864 minus 04
UDP-glycosyltransferaseactivity FANhyb icon00008064 a1g000011 249 Up 310119864 minus 04
PREDICTEDUDP-glucose flavonoid3-O-glucosyltransferase7-like
FANhyb rscf000055631g000011 212 Up 110119864 minus 19
PREDICTEDUDP-glycosyltransferase76F1-like
FANhyb rscf000021771g000031 2 Up 720119864 minus 29
A transcripts in the etiolated leaves of wheat seedling phy-tochromes [26] However our study shows that phytochromeA was not differentially expressed It is probably becausethe hematin-treated strawberries were not shaded during theexperiment while the function of phytochrome A is light-dependent In addition we found that a downstream gene ofphytochrome PIF3 (FANhyb rscf000006691g000021) wassignificantly upregulated PIF3 is thought to be a positive reg-ulator of phytochrome B mediated by light-dependent signaltransduction [58]These results suggest that the phytochromepathways may be involved in the anthocyanin biosynthesispromoted by hematin
345 DEGs in Starch and Sucrose Degradation Proteomicapproaches have revealed that starch degradation contributesto anthocyanin accumulation in tuberous roots of the purplesweet potato variety [59] Starch and sucrose biosynthesis isimportant for anthocyanin accumulation in strawberry fruit[60] In this study we found the DEGs involved in starchand sucrose synthesis in the hematin-treated fruit were alldownregulated (Table 5) For example the expression of theunigenes encoding soluble starch synthase 3 sucrose trans-port protein SUC2-like and a probable sucrose-phosphatesynthase 4-like enzyme were decreased 23- 46- 32- and2-fold respectively This suggests that exogenous hematinregulates the biosynthesis of starch and sucrose and henceaffects fruit coloring
346 DEGs in the Calcium Pathway in Response to HematinCalcium can also increase the transcription levels of keystructural genes F3H DFR ANT and UFGT in the whitestage of strawberries [61] In this study we found thatthree DEGs in calcium biosynthesis and transport were
downregulated in the hematin-treated fruit (Table 5) Onlythe unigene encoding predicted cationcalcium exchanger5-like was upregulated The unigenes encoding a calpain-type cysteine protease and a predicted calcium-binding pro-tein PBP1-like were downregulated respectively Exogenousapplication of calcium can promote apple coloring [62] inaddition to variation in anthocyanin content [61] Our resultsindicate that the biosynthesis and transport of calcium areinvolved in the development of coloring in hematin-treatedstrawberries
35 Validation of Selected DEGs by qRT-PCR To validate theexpression of the DEGs obtained from RNA-Seq 11 DEGswere selected for qRT-PCR including structural genes (DFRand UDFGs) transcription factor genes (MYB and bHLH)and phytochrome chromophore-related gene (HO-1) Theprimers used for qRT-PCR are listed in Supplementary TableS1 The qRT-PCR results were consistent with the RNA-Seqdata (Figure 4) except for FANhyb rscf000001411g000161which had a higher log
2ratio (hematin-treatedcontrol) in
the transcriptome data than in qRT-PCR These results indi-cate the RNA-Seq data from the strawberry transcriptome isreproducible and accurate
4 Conclusions
In this study the anthocyanin content in the strawberryfruit was elevated by the application of exogenous hematinThis is the first report that hematin could increase theanthocyanin production in fruit Furthermore we exploredthe effects of the exogenous hematin on metabolic path-ways using genome-wide transcriptome analysis The resultsindicate that the expression levels of many genes involved
International Journal of Genomics 11
Table 5 Other DEGs identified in anthocyanin biosynthesis-related pathways
Anthocyaninbiosynthesis-relatedpathways
Annotation genes Gene IDReal fold-changevalues
Upregulationdownregulation FDR
Calcium ion binding
Cationcalciumexchanger 5-like
FANhyb rscf000019061g000011 203 Up 64119864 minus 06
Calpain-type cysteineprotease family
FANhyb icon00011755 a1g000011 345 Down 18119864 minus 09
Calcium-bindingprotein PBP1-like
FANhyb rscf000007501g000081 323 Down 97119864 minus 04
Calpain-type cysteineprotease family
FANhyb icon00023658 a1g000011 233 Down 60119864 minus 08
CytokininCytokinin
dehydrogenase 5-likeFANhyb iscf00325393 1 s1g000011 667 Down 62119864 minus 06
Cytochrome P450714A1
FANhyb rscf000005921g000031 217 Down 98119864 minus 10
JasmonateJasmonate
O-methyltransferase-like
FANhyb rscf000000041g000131 201 Up 70119864 minus 04
Abscisic acid
Protein ABSCISICACID-INSENSITIVE
5-likeFANhyb rscf000021641g000011 4 Down 16119864 minus 18
ABA overly sensitive 5 FANhyb icon00051144 a1g000011 291 Up 24119864 minus 059-cis-
epoxycarotenoiddioxygenase NCED1
FANhyb icon18399909 o1g000011 441 Up 43119864 minus 06
Abscisic acid81015840-hydroxylase 1-like
FANhyb icon00000938 a1g000011 256 Down 14119864 minus 06
Abscisic acid receptorPYR1-like
FANhyb icon00020426 a1g000011 208 Down 30119864 minus 06
Auxin
Auxin efflux carriercomponent 1-like
FANhyb rscf000010081g000031 384 Down 23119864 minus 06
Auxin response factor8
FANhyb rscf000013061g000021 263 Down 61119864 minus 17
Auxin response factor17-like
FANhyb rscf000060741g000011 217 Down 11119864 minus 36
Starch and sucrose
Glycosyltransferasefamily 35
FANhyb rscf000000341g000081 384 Down 11119864 minus 05
Soluble starchsynthase 3
FANhyb rscf000000451g000041 232 Down 38119864 minus 12
Sucrose transportprotein SUC2-like
FANhyb icon00036727 a1g000011 455 Down 98119864 minus 04
Sucrose transportprotein SUC2-like
FANhyb rscf000007551g000021 323 Down 66119864 minus 10
Probablesucrose-phosphatesynthase 4-like
FANhyb rscf000013501g000021 213 Down 11119864 minus 14
Phytochrome
PREDICTEDtranscription factor
PIF3-likeFANhyb rscf000006691g000021 346 up 14119864 minus 07
Phytochrome E-like FANhyb rscf000004361g000041 4 Down 33119864 minus 05
Heme oxygenase 1 FANhyb icon00004395 a1g000011 258 27119864 minus 07 27119864 minus 07
12 International Journal of Genomics
Hematin
Endoplasmicreticulum
WD40 bLHLMYB
Structural genes
TFs
Nuclear
JA
ABA Sucrose
Plastid
G-cis-neoxanthin
Apophytochrome
StarchHeme
BVSucroseABA
Holophytochrome
PfrPIF3
Flavonoids Anthocyanins
Vacuole
Cytoplasm
Cell wall
①
②③
④⑤
⑥
⑦
PΦB
PΦB
CTKAuxin
Ca2+
Figure 5 Genes and biological pathways that were involved in regulation of anthocyanin accumulation promoted by hematin Outermostbox represents the cell wall Bilayer oval shape represents the plastidThe red-brown ring and network represent the nuclear and endoplasmicreticulum respectively The red crescent represents the vacuole The box in the nucleus represents transcription factors (TFs) Yellow MYBgreenWD40 and red bHLH represent a ternary transcription factor complex which transcribes anthocyanin biosynthesis genes The circlednumbers represent biological pathways involved in the regulation of anthocyanin accumulation promoted by hematin A represents thephytochrome regulation pathwayB represents the starch and sucrose pathwayCDE andF represent phytohormones ABA JA AuxinandCTK regulation pathways respectivelyG represents theCa2+ regulation pathway Pfr represents the far-red phytochrome PIF3 representsphytochrome-interacting factor Each anthocyanin biosynthesis regulatory pathway related gene is in Table 5
in anthocyanin biosynthesis were significantly altered withthe hematin treatment This suggests that the physiologicalprocess of fruit color development is regulated through com-plex interactions among anthocyanin biosynthesis pathwaysplant hormone signal transduction pathways phytochromesignal transduction pathways starch and sugar metabolicpathways calcium pathways and transcription factors (Fig-ure 5) This study adds to the in-depth understanding of thefruit coloration process in strawberry
Competing Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contributions
Jianwei Gao and Liangju Wang conceived and designed theresearch Yi Li Huayin Li Fengde Wang Jingjuan Li andYihui Zhang conducted experiments Yi Li analyzed the dataYi Li and Fengde Wang wrote the manuscript All authorsread and approved the manuscript Yi Li and Huayin Licontributed equally to this work
Acknowledgments
This work was supported by the Modern Agricultural Indus-trial Technology System Funding of Shandong ProvinceChina (SDAIT-02-022-04) the China agriculture researchsystem (CARS-25) and the National Science Foundation ofChina (31401820)
References
[1] K Aaby SMazur A Nes andG Skrede ldquoPhenolic compoundsin strawberry (Fragaria x ananassa Duch) fruits compositionin 27 cultivars and changes during ripeningrdquo Food Chemistryvol 132 no 1 pp 86ndash97 2012
[2] T A Colquhoun L A Levin H RMoskowitz VMWhitakerD G Clark and K M Folta ldquoFraming the perfect strawberryan exercise in consumer-assisted selection of fruit cropsrdquoJournal of Berry Research vol 2 no 1 pp 45ndash61 2012
[3] F Giampieri J M Alvarez-Suarez L Mazzoni et al ldquoThepotential impact of strawberry on human healthrdquo NaturalProduct Research vol 27 no 4 pp 448ndash455 2013
[4] Y S Velioglu GMazza L Gao and B D Oomah ldquoAntioxidantactivity and total phenolics in selected fruits vegetables and
International Journal of Genomics 13
grain productsrdquo Journal of Agricultural amp Food Chemistry vol46 no 10 pp 4113ndash4117 1998
[5] S S Hassellund A Flaa S E Kjeldsen et al ldquoEffects of antho-cyanins on cardiovascular risk factors and inflammation inpre-hypertensive men a double-blind randomized placebo-controlled crossover studyrdquo Journal of Human Hypertensionvol 27 no 2 pp 100ndash106 2013
[6] K J Meyers C B Watkins M P Pritts and R H Liu ldquoAnti-oxidant and antiproliferative activities of strawberriesrdquo Journalof Agricultural amp Food Chemistry vol 51 no 23 pp 6887ndash68922003
[7] S Zafra-Stone T YasminM Bagchi AChatterjee J AVinsonand D Bagchi ldquoBerry anthocyanins as novel antioxidants inhuman health and disease preventionrdquo Molecular Nutrition ampFood Research vol 51 no 6 pp 675ndash683 2007
[8] L Jaakola ldquoNew insights into the regulation of anthocyanin bio-synthesis in fruitsrdquoTrends in Plant Science vol 18 no 9 pp 477ndash483 2013
[9] K Springob J-I Nakajima M Yamazaki and K Saito ldquoRecentadvances in the biosynthesis and accumulation of antho-cyaninsrdquo Natural Product Reports vol 20 no 3 pp 288ndash3032003
[10] A Gonzalez M Zhao J M Leavitt and A M Lloyd ldquoRegu-lation of the anthocyanin biosynthetic pathway by the TTG1bHLHMyb transcriptional complex in Arabidopsis seedlingsrdquoPlant Journal vol 53 no 5 pp 814ndash827 2008
[11] K Petroni and C Tonelli ldquoRecent advances on the regulationof anthocyanin synthesis in reproductive organsrdquo Plant Sciencevol 181 no 3 pp 219ndash229 2011
[12] H-F Jia Y-M Chai C-L Li et al ldquoAbscisic acid plays animportant role in the regulation of strawberry fruit ripeningrdquoPlant Physiology vol 157 no 1 pp 188ndash199 2011
[13] C Merchante J G Vallarino S Osorio et al ldquoEthylene isinvolved in strawberry fruit ripening in an organ-specific man-nerrdquo Journal of Experimental Botany vol 64 no 14 pp 4421ndash4439 2013
[14] A Maier A Schrader L Kokkelink et al ldquoLight and the E3ubiquitin ligase COP1SPA control the protein stability of theMYB transcription factors PAP1 and PAP2 involved in antho-cyanin accumulation in Arabidopsisrdquo Plant Journal vol 74 no4 pp 638ndash651 2013
[15] H Lange W Shropshire and H Mohr ldquoAn analysis of phyto-chrome-mediated anthocyanin synthesisrdquo Plant Physiology vol47 no 5 pp 649ndash655 1971
[16] T D Elich and J C Lagarias ldquoPhytochrome chromophore bio-synthesis both 5-aminolevulinic acid and biliverdin overcomeinhibition by gabaculine in etiolated Avena sativa L seedlingsrdquoPlant Physiology vol 84 no 2 pp 304ndash310 1987
[17] Y Han W Xuan T Yu et al ldquoExogenous hematin alleviatesmercury-induced oxidative damage in the roots of Medicagosativardquo Journal of Integrative Plant Biology vol 49 no 12 pp1703ndash1713 2007
[18] M Koornneef J Cone R Dekens E OrsquoHerne-Robers CSpruit and R Kendrick ldquoPhotomorphogenic responses of longhypocotyl mutants of tomatordquo Journal of Plant Physiology vol120 no 2 pp 153ndash165 1985
[19] M J Terry and R E Kendrick ldquoFeedback inhibition of chloro-phyll synthesis in the phytochrome chromophore-deficientaurea and yellow-green-2 mutants of tomatordquo Plant Physiologyvol 119 no 1 pp 143ndash152 1999
[20] B M Parks and P H Quail ldquoPhytochrome-deficient hy1 andhy2 long hypocotyl mutants of Arabidopsis are defective inphytochrome chromophore biosynthesisrdquo Plant Cell vol 3 no11 pp 1177ndash1186 1991
[21] M J Terry ldquoPhytochrome chromophore-deficient mutantsrdquoPlant Cell amp Environment vol 20 no 6 pp 740ndash745 1997
[22] T Muramoto T Kohchi A Yokota I Hwang and H MGoodman ldquoThe Arabidopsis photomorphogenic mutant hy1 isdeficient in phytochrome chromophore biosynthesis as a resultof a mutation in a plastid heme oxygenaserdquo Plant Cell vol 11no 3 pp 335ndash347 1999
[23] M D Maines ldquoThe heme oxygenase system a regulator ofsecond messenger gasesrdquo Annual Review of Pharmacology andToxicology vol 37 no 1 pp 517ndash554 1997
[24] W Xuan F-Y Zhu S Xu et al ldquoThe heme oxygenasecarbonmonoxide system is involved in the auxin-induced cucumberadventitious rooting processrdquo Plant Physiology vol 148 no 2pp 881ndash893 2008
[25] R Amooaghaie F Tabatabaei and A-M Ahadi ldquoRole ofhematin and sodium nitroprusside in regulating Brassica nigraseed germination under nanosilver and silver nitrate stressesrdquoEcotoxicology and Environmental Safety vol 113 pp 259ndash2702015
[26] Y Liu X Li L Xu and W Shen ldquoDe-etiolation of wheatseedling leaves cross talk between heme oxygenasecarbonmonoxide and nitric oxiderdquo PLoS ONE vol 8 no 12 ArticleID e81470 2013
[27] T Muramoto N Tsurui M J Terry A Yokota and T KohchildquoExpression and biochemical properties of a ferredoxin-dependent heme oxygenase required for phytochrome chro-mophore synthesisrdquo Plant Physiology vol 130 no 4 pp 1958ndash1966 2002
[28] J-W Gao J Liu B Li and Z Li ldquoIsolation and purificationof functional total RNA from blue-grained wheat endospermtissues containing high levels of starches and flavonoidsrdquo PlantMolecular Biology Reporter vol 19 no 2 pp 185ndash186 2001
[29] R Patro S M Mount and C Kingsford ldquoSailfish enablesalignment-free isoform quantification from RNA-seq readsusing lightweight algorithmsrdquoNature Biotechnology vol 32 no5 pp 462ndash464 2014
[30] Y-X Qi Y-B Liu and W-H Rong ldquoRNA-Seq and its applica-tions a new technology for transcriptomicsrdquo Hereditas vol 33no 11 pp 1191ndash1202 2011
[31] C R Genovese N A Lazar and T Nichols ldquoThresholdingof statistical maps in functional neuroimaging using the falsediscovery raterdquo NeuroImage vol 15 no 4 pp 870ndash878 2002
[32] J Ye L Fang H Zheng et al ldquoWEGO a web tool for plottingGO annotationsrdquoNucleic Acids Research vol 34 supplement 2pp W293ndashW297 2006
[33] H Abdi ldquoThe Bonferonni and Sidak corrections for multiplecomparisonrdquo Encyclopedia of Measurement and Statistics vol 3pp 103ndash107 2007
[34] J Li Q Ding F Wang Y Zhang H Li and J Gao ldquoIntegrativeanalysis of mRNA and miRNA expression profiles of thetuberous root development at seedling stages in turnipsrdquo PLoSONE vol 10 no 9 Article ID e0137983 2015
[35] J Bennett D Hondred and J C Register ldquoKeeping qRT-PCRrigorous and biologically relevantrdquo Plant Cell Reports vol 34no 1 pp 1ndash3 2015
[36] F Wang L Li H Li et al ldquoTranscriptome analysis of rosetteand folding leaves in Chinese cabbage using high-throughputRNA sequencingrdquo Genomics vol 99 no 5 pp 299ndash307 2012
14 International Journal of Genomics
[37] T Shi Z Gao L Wang et al ldquoIdentification of differentially-expressed genes associated with pistil abortion in Japaneseapricot by genome-wide transcriptional analysisrdquo PLoS ONEvol 7 no 10 Article ID e47810 2012
[38] F Wang H Li Y Zhang et al ldquoMicroRNA expression analysisof rosette and folding leaves in Chinese cabbage using high-throughput Solexa sequencingrdquo Gene vol 532 no 2 pp 222ndash229 2013
[39] X Gu Y Chen Z Gao Y Qiao and X Wang ldquoTranscriptionfactors and anthocyanin genes related to low-temperature toler-ance in rd29A RdreB1BI transgenic strawberryrdquo Plant Physiol-ogy and Biochemistry vol 89 pp 31ndash43 2015
[40] J G Schaart C Dubos I R De La Fuente et al ldquoIdentificationand characterization of MYB-bHLH-WD40 regulatory com-plexes controlling proanthocyanidin biosynthesis in strawberry(Fragaria times ananassa) fruitsrdquoNew Phytologist vol 197 no 2 pp454ndash467 2013
[41] L Xie Z H Wang X H Cheng J J Gao Z P Zhangand L J Wang ldquo5-Aminolevulinic acid promotes anthocyaninaccumulation in Fuji applesrdquo Plant Growth Regulation vol 69no 3 pp 295ndash303 2013
[42] M Daminato F Guzzo and G Casadoro ldquoASHATTERPROOF-like gene controls ripening in non-climac-teric strawberries and auxin and abscisic acid antagonisticallyaffect its expressionrdquo Journal of Experimental Botany vol 64no 12 pp 3775ndash3786 2013
[43] X-H Ji R Zhang N Wang L Yang and X-S Chen ldquoTran-scriptome profiling reveals auxin suppressed anthocyanin bio-synthesis in red-fleshed apple callus (Malus sieversii f niedzwet-zkyana)rdquo Plant Cell Tissue and Organ Culture vol 123 no 2pp 389ndash404 2015
[44] J Chen LMaoW Lu T Ying and Z Luo ldquoTranscriptome pro-filing of postharvest strawberry fruit in response to exogenousauxin and abscisic acidrdquo Planta vol 243 no 1 pp 183ndash197 2016
[45] G A Martınez A R Chaves and M C Anon ldquoEffect of gib-berellic acid on ripening of strawberry fruits (Fragaria annan-assa Duch)rdquo Journal of Plant Growth Regulation vol 13 no 2pp 87ndash91 1994
[46] P K Das D H Shin S-B Choi S-D Yoo G Choi and Y-IPark ldquoCytokinins enhance sugar-induced anthocyanin biosyn-thesis in Arabidopsisrdquo Molecules amp Cells vol 34 no 1 pp 93ndash101 2012
[47] P McAtee S Karim R Schaffer and K David ldquoA dynamicinterplay between phytohormones is required for fruit develop-ment maturation and ripeningrdquo Frontiers in Plant Science vol4 no 3 article 79 7 pages 2013
[48] E Nambara andAMarion-Poll ldquoAbscisic acid biosynthesis andcatabolismrdquoAnnual Review of Plant Biology vol 56 pp 165ndash1852005
[49] Y Jiang and D C Joyce ldquoABA effects on ethylene productionPAL activity anthocyanin and phenolic contents of strawberryfruitrdquo Plant Growth Regulation vol 39 no 2 pp 171ndash174 2003
[50] S Li ldquoTranscriptional control of flavonoid biosynthesis fine-tuning of the MYB-bHLH-WD40 (MBW) complexrdquo PlantSignaling amp Behavior vol 9 Article ID e27522 2014
[51] D R Rudell J K Fellman and J P Mattheis ldquoPreharvest appli-cation of methyl jasmonate to rsquoFujirsquo apples enhances red col-oration and affects fruit size splitting and bitter pit incidencerdquoHortScience vol 40 no 6 pp 1760ndash1762 2005
[52] A Belhadj N Telef C Saigne et al ldquoEffect of methyl jasmonatein combination with carbohydrates on gene expression of PR
proteins stilbene and anthocyanin accumulation in grapevinecell culturesrdquo Plant Physiology and Biochemistry vol 46 no 4pp 493ndash499 2008
[53] E Loreti G Povero G Novi C Solfanelli A Alpi and PPerata ldquoGibberellins jasmonate and abscisic acid modulate thesucrose-induced expression of anthocyanin biosynthetic genesin Arabidopsisrdquo New Phytologist vol 179 no 4 pp 1004ndash10162008
[54] H S Seo J T Song J-J Cheong et al ldquoJasmonic acid carboxylmethyltransferase A key enzyme for jasmonate-regulated plantresponsesrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 98 no 8 pp 4788ndash4793 2001
[55] M T McDowell and J C Lagarias ldquoPurification and biochemi-cal properties of phytochromobilin synthase from etiolated oatseedlingsrdquo Plant Physiology vol 126 no 4 pp 1546ndash1554 2001
[56] G-Q Fu S Xu Y-J Xie et al ldquoMolecular cloning characteri-zation and expression of an alfalfa (Medicago sativa L) hemeoxygenase-1 gene MsHO1 which is pro-oxidants-regulatedrdquoPlant Physiology and Biochemistry vol 49 no 7 pp 792ndash7992011
[57] X-Y Chen X Ding S Xu et al ldquoEndogenous hydrogen per-oxide plays a positive role in the upregulation of heme oxy-genase and acclimation to oxidative stress in wheat seedlingleavesrdquo Journal of Integrative Plant Biology vol 51 no 10 pp951ndash960 2009
[58] J Shin E Park and G Choi ldquoPIF3 regulates anthocyaninbiosynthesis in an HY5-dependent manner with both factorsdirectly binding anthocyanin biosynthetic gene promoters inArabidopsisrdquo Plant Journal vol 49 no 6 pp 981ndash994 2007
[59] S Wang D Pan X Lv et al ldquoProteomic approach reveals thatstarch degradation contributes to anthocyanin accumulation intuberous root of purple sweet potatordquo Journal of Proteomics vol143 pp 298ndash305 2016
[60] H-F Jia C-L Li Y-M Chai Y Xing and Y Shen ldquoSucrosepromotes strawberry fruit ripening by stimulation of abscisicacid biosynthesisrdquo Pakistan Journal of Botany vol 45 no 1 pp169ndash176 2013
[61] W Xu H Peng T Yang et al ldquoEffect of calcium on strawberryfruit flavonoid pathway gene expression and anthocyanin accu-mulationrdquo Plant Physiology amp Biochemistry vol 82 pp 289ndash298 2014
[62] Z-W-SWan and Z Singh ldquoExogenous application of prohexa-dione-calcium promotes fruit colour development of lsquoCrippsPinkrsquo applerdquo Acta Horticulturae vol 1012 pp 219ndash225 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 International Journal of Genomics
Table 2 Pathway enrichment analysis of differentially expressed genes
Pathway DEGs with pathwayannotation (451)
All genes with pathwayannotation (15877)
Corrected 119901 value 119876-value Pathway ID
RNA polymerase 16 183 688119864 minus 5 00068107 ko03020Pyrimidine metabolism 20 317 00007651746 00320 ko00240Purine metabolism 21 347 00009710371 0032044224 ko00230
significantly downregulated There were three unigenes inthe AP2-EREBP family with only one upregulated and theothers downregulated These results suggest that exogenoushematin is involved in developmental transcriptionalregulation of anthocyanin biosynthesis
342 DEGs in Anthocyanin Biosynthesis The anthocyaninbiosynthetic pathway has been studied intensively Theanthocyanin biosynthetic pathway via the phenylpropanoidpathway is well known [8] In this study the transcriptomedata showed a significant increase in the expression of genesin the anthocyanin biosynthesis pathway when exposedto exogenous hematin The structural genes involved inanthocyanin biosynthesis including DFR LAR and UDP-glycosyltransferase exhibited significant differential expres-sion in response to the hematin treatment (Table 4) Forexample two DFR genes and one LAR gene were signifi-cantly upregulated (the expression increased 18- 30- and3-fold in the hematin-treated fruit resp) In addition allsix genes encodingUDP-glycosyltransferasewere significantlyupregulated Among these the expression of one gene(Fanhy icon15742070o1g000011) was upregulated 18305-fold The results suggest that exogenous hematin may havean important role in promoting anthocyanin biosynthesis
343 DEGs in Hormone Signaling Transduction in Responseto Hematin Auxin has been shown to negatively regulatethe expression of the anthocyanin biosynthesis genes [42] Inthis study there were three unigenes involved in the auxinsignaling pathway and all were downregulated includingone gene encoding auxin efflux carrier component 1-like(AUX1-like) and two genes encoding auxin response factors(ARF) These results suggest that hematin promotes antho-cyanin accumulation by regulating auxin in the colorationprocess of strawberries It has shown that auxin suppressesanthocyanin biosynthesis in the red-fleshed apple callus [43]Endogenous expression of auxin [44 45] has been foundto impede anthocyanin accumulation in strawberries Inadditional cytokinins are also known to play an importantrole in anthocyanin biosynthesis It has shown to enhanceanthocyanin accumulation in Zea mays and regulate antho-cyanin production and composition in suspension culturesof strawberry cells [46] In this study two unigenes relatedto the cytokinin signaling pathway were identified and theywere downregulated as well One gene was identified ascytokinin dehydrogenase 5-like which catalyzes the irre-versible degradation of cytokinins This result indicates thathematin promotes anthocyanin accumulation by regulatingthe cytokinin signaling pathway
Extensive researches have shown that ABA plays animportant role in the regulation of anthocyanin biosynthesisin nonclimacteric fruit [47] In this study a total of five uni-genes related to the ABA signaling pathway were identifiedamong which two were upregulated and three were down-regulated Two genes which are involved in the abscisic acidresponse were upregulated These two unigenes encode 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) and ABA overlysensitive 5 (ABO5) respectively ABA is synthesized fromcarotenoids via several enzymatic reactions in the plastidThe rate-limiting step in these reactions is catalyzed by 9-cis-epoxycarotenoid dioxygenase 1 (NCED1) [48] Mutations inthe FaNCED1 gene result in colorless strawberry fruit whichcan become colored through the application of exogenousABA [12] Another unigene was identified to encode abscisicacid 81015840-hydroxylase 1-like a key enzyme in the oxidativecatabolism of abscisic acid which was downregulated Appli-cation of exogenous ABA regulates phenylalanine ammonialyase activity and increases the phenolic and anthocyanincontent of strawberry fruit [12 49] Our results suggestthat hematin promotes ABA biosynthesis and inhibits ABAdisintegration in the hematin-treated fruit
In Arabidopsis JAs can affect anthocyanin accumulationvia the interaction of negative regulators with the MBWcomplex of transcription factors involved in anthocyaninbiosynthesis [50] Preharvest application of JA to ldquoFujirdquo applesenhances red coloration [51] JA vapor treatment can alsoenhance anthocyanins in strawberry fruit [52] In this studyjasmonate O-methyltransferase-like exhibited significantlydifferential expression Jasmonate O-methyltransferase-likeis also known to be a key enzyme for jasmonate-regulatedplant responses to stimuli [53 54] These results suggest thathematin increases the activity of the JA-regulated antho-cyanin biosynthesis
344 DEGs in the Phytochrome Signaling Pathway in Responseto Hematin Phytochromes that act as photoreceptors playan important role in anthocyanin regulation [15] Phy-tochromes are homodimeric chromoproteins where eachholophytochrome is composed of a phytochrome protein(apophytochrome) covalently bound to a linear tetrapyrrolePΦB HO1 is crucial to this process and acts as a rate-limiting enzyme in the biosynthesis of PΦB [55] Exogenoushematin can also induce HO-1 expression in many plants[56 57] In this study we found exogenous hematin signif-icantly increased the expression of FaHO1 (Table 5) Thisresult implies that hematin can promote PΦB biosynthesisRecently it has been reported that hematin could inducethe accumulation of far-red phytochrome and phytochrome
International Journal of Genomics 9
Table 3 DEGs acting as transcription factors in response to exogenous hematin
Annotation transcription factors Gene IDReal
fold-changevalues
Upregulationdownregulation FDR
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb icon00006295 a1g000011 938 Up 120119864 minus 05
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb rscf000031551g000011 4 Down 480119864 minus 16
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL5-like
FANhyb rscf000000291g000101 4 Down 320119864 minus 08
MYBdomain protein 66 FANhyb rscf000001461g000071 654 Up 780119864 minus 07MYB-related protein 3R-1-like partial FANhyb rscf000001411g000161 282 Up 830119864 minus 04PREDICTED transcription factorGAMYB-like FANhyb rscf000006491g000061 303 Down 150119864 minus 13
PREDICTED probable WRKYtranscription factor 53-like FANhyb rscf000019731g000021 217 Down 200119864 minus 08
Transcription factor bHLH80-like FANhyb rscf000012921g000031 6250 Down 150119864 minus 06Transcription factor bHLH70-like FANhyb rscf000002951g000031 526 Down 660119864 minus 12Transcription factor ICE1-like FANhyb rscf000001701g000061 244 Down 450119864 minus 18C2H2-like zinc finger protein FANhyb rscf000011431g000021 282 Up 590119864 minus 04Zinc finger protein MAGPIE-like FANhyb rscf000030241g000011 204 Up 150119864 minus 04PREDICTED axial regulator YABBY5-like FANhyb rscf000016671g000011 417 Down 730119864 minus 05
PREDICTED axial regulator YABBY1-like FANhyb rscf000068991g000011 357 Down 620119864 minus 06
PREDICTED dof zinc finger proteinDOF37-like isoform 1 FANhyb rscf000068561g000011 263 Down 280119864 minus 04
ZF-HD homeobox protein FANhyb rscf000000441g000221 385 Down 420119864 minus 08ZF-HD homeobox proteinAt4g24660-like FANhyb rscf000052411g000011 357 Down 470119864 minus 09
PREDICTED B3 domain-containingprotein REM14-like FANhyb rscf000010091g000021 243 Up 530119864 minus 04
PREDICTED B3 domain-containingtranscription factor ABI3-like FANhyb rscf000012711g000021 455 Down 130119864 minus 28
Growth-regulating factor 6 FANhyb rscf000000241g000291 345 Down 330119864 minus 34Growth-regulating factor 5 FANhyb icon00009704 a1g000011 27 Down 190119864 minus 06GATA type zinc finger transcriptionfactor family protein FANhyb rscf000003931g000131 1111 Down 110119864 minus 05
PREDICTED zinc finger CCCHdomain-containing protein 2-like FANhyb icon00006770 a1g000011 313 Down 680119864 minus 49
PREDICTED E2F transcriptionfactor-like E2FE-like FANhyb icon00002702 a1g000011 278 Down 240119864 minus 04
PREDICTED uncharacterized protein FANhyb rscf000010661g000011 263 Down 490119864 minus 06PREDICTED nuclear transcriptionfactor Y subunit A-3-like isoform 1 FANhyb rscf000000081g000351 385 Down 500119864 minus 20
PREDICTED homeobox-leucinezipper protein ATHB-8-like FANhyb rscf000000151g000051 263 Down 100119864 minus 06
PREDICTED MADS-boxtranscription factor 18-like FANhyb rscf000003231g000161 243 Down 180119864 minus 06
PREDICTED zinc finger protein132-like isoform 1 FANhyb rscf000070151g000011 243 Down 180119864 minus 08
PREDICTED transcription factorPIF3-like FANhyb rscf000006691g000021 346 Up 140119864 minus 07
10 International Journal of Genomics
Table 4 Regulation of the DEGs in anthocyanin biosynthesis in response to exogenous hematin
Genes in anthocyaninbiosynthesis Gene ID
Realfold-change
valuesRegulation level FDR
Dihydroflavonol-4-reductase FANhyb icon00002410 a1g000011 3091 Up 910119864 minus 06
Flavonol synthase FANhyb icon00015354 a1g000011 345 down 180119864 minus 05Isoflavone reductasehomolog FANhyb icon20341135 s1g000011 334 Up 750119864 minus 14
UDP-glycosyltransferase FANhyb rscf000049691g000011 307 Up 640119864 minus 08UDP-glycosyltransferaseactivity FANhyb icon15742070 o1g000011 18 30563 Up 510119864 minus 09
UDP-glycosyltransferase73C1 FANhyb icon00027696 a1g000011 298 Up 560119864 minus 04
UDP-glycosyltransferaseactivity FANhyb icon00008064 a1g000011 249 Up 310119864 minus 04
PREDICTEDUDP-glucose flavonoid3-O-glucosyltransferase7-like
FANhyb rscf000055631g000011 212 Up 110119864 minus 19
PREDICTEDUDP-glycosyltransferase76F1-like
FANhyb rscf000021771g000031 2 Up 720119864 minus 29
A transcripts in the etiolated leaves of wheat seedling phy-tochromes [26] However our study shows that phytochromeA was not differentially expressed It is probably becausethe hematin-treated strawberries were not shaded during theexperiment while the function of phytochrome A is light-dependent In addition we found that a downstream gene ofphytochrome PIF3 (FANhyb rscf000006691g000021) wassignificantly upregulated PIF3 is thought to be a positive reg-ulator of phytochrome B mediated by light-dependent signaltransduction [58]These results suggest that the phytochromepathways may be involved in the anthocyanin biosynthesispromoted by hematin
345 DEGs in Starch and Sucrose Degradation Proteomicapproaches have revealed that starch degradation contributesto anthocyanin accumulation in tuberous roots of the purplesweet potato variety [59] Starch and sucrose biosynthesis isimportant for anthocyanin accumulation in strawberry fruit[60] In this study we found the DEGs involved in starchand sucrose synthesis in the hematin-treated fruit were alldownregulated (Table 5) For example the expression of theunigenes encoding soluble starch synthase 3 sucrose trans-port protein SUC2-like and a probable sucrose-phosphatesynthase 4-like enzyme were decreased 23- 46- 32- and2-fold respectively This suggests that exogenous hematinregulates the biosynthesis of starch and sucrose and henceaffects fruit coloring
346 DEGs in the Calcium Pathway in Response to HematinCalcium can also increase the transcription levels of keystructural genes F3H DFR ANT and UFGT in the whitestage of strawberries [61] In this study we found thatthree DEGs in calcium biosynthesis and transport were
downregulated in the hematin-treated fruit (Table 5) Onlythe unigene encoding predicted cationcalcium exchanger5-like was upregulated The unigenes encoding a calpain-type cysteine protease and a predicted calcium-binding pro-tein PBP1-like were downregulated respectively Exogenousapplication of calcium can promote apple coloring [62] inaddition to variation in anthocyanin content [61] Our resultsindicate that the biosynthesis and transport of calcium areinvolved in the development of coloring in hematin-treatedstrawberries
35 Validation of Selected DEGs by qRT-PCR To validate theexpression of the DEGs obtained from RNA-Seq 11 DEGswere selected for qRT-PCR including structural genes (DFRand UDFGs) transcription factor genes (MYB and bHLH)and phytochrome chromophore-related gene (HO-1) Theprimers used for qRT-PCR are listed in Supplementary TableS1 The qRT-PCR results were consistent with the RNA-Seqdata (Figure 4) except for FANhyb rscf000001411g000161which had a higher log
2ratio (hematin-treatedcontrol) in
the transcriptome data than in qRT-PCR These results indi-cate the RNA-Seq data from the strawberry transcriptome isreproducible and accurate
4 Conclusions
In this study the anthocyanin content in the strawberryfruit was elevated by the application of exogenous hematinThis is the first report that hematin could increase theanthocyanin production in fruit Furthermore we exploredthe effects of the exogenous hematin on metabolic path-ways using genome-wide transcriptome analysis The resultsindicate that the expression levels of many genes involved
International Journal of Genomics 11
Table 5 Other DEGs identified in anthocyanin biosynthesis-related pathways
Anthocyaninbiosynthesis-relatedpathways
Annotation genes Gene IDReal fold-changevalues
Upregulationdownregulation FDR
Calcium ion binding
Cationcalciumexchanger 5-like
FANhyb rscf000019061g000011 203 Up 64119864 minus 06
Calpain-type cysteineprotease family
FANhyb icon00011755 a1g000011 345 Down 18119864 minus 09
Calcium-bindingprotein PBP1-like
FANhyb rscf000007501g000081 323 Down 97119864 minus 04
Calpain-type cysteineprotease family
FANhyb icon00023658 a1g000011 233 Down 60119864 minus 08
CytokininCytokinin
dehydrogenase 5-likeFANhyb iscf00325393 1 s1g000011 667 Down 62119864 minus 06
Cytochrome P450714A1
FANhyb rscf000005921g000031 217 Down 98119864 minus 10
JasmonateJasmonate
O-methyltransferase-like
FANhyb rscf000000041g000131 201 Up 70119864 minus 04
Abscisic acid
Protein ABSCISICACID-INSENSITIVE
5-likeFANhyb rscf000021641g000011 4 Down 16119864 minus 18
ABA overly sensitive 5 FANhyb icon00051144 a1g000011 291 Up 24119864 minus 059-cis-
epoxycarotenoiddioxygenase NCED1
FANhyb icon18399909 o1g000011 441 Up 43119864 minus 06
Abscisic acid81015840-hydroxylase 1-like
FANhyb icon00000938 a1g000011 256 Down 14119864 minus 06
Abscisic acid receptorPYR1-like
FANhyb icon00020426 a1g000011 208 Down 30119864 minus 06
Auxin
Auxin efflux carriercomponent 1-like
FANhyb rscf000010081g000031 384 Down 23119864 minus 06
Auxin response factor8
FANhyb rscf000013061g000021 263 Down 61119864 minus 17
Auxin response factor17-like
FANhyb rscf000060741g000011 217 Down 11119864 minus 36
Starch and sucrose
Glycosyltransferasefamily 35
FANhyb rscf000000341g000081 384 Down 11119864 minus 05
Soluble starchsynthase 3
FANhyb rscf000000451g000041 232 Down 38119864 minus 12
Sucrose transportprotein SUC2-like
FANhyb icon00036727 a1g000011 455 Down 98119864 minus 04
Sucrose transportprotein SUC2-like
FANhyb rscf000007551g000021 323 Down 66119864 minus 10
Probablesucrose-phosphatesynthase 4-like
FANhyb rscf000013501g000021 213 Down 11119864 minus 14
Phytochrome
PREDICTEDtranscription factor
PIF3-likeFANhyb rscf000006691g000021 346 up 14119864 minus 07
Phytochrome E-like FANhyb rscf000004361g000041 4 Down 33119864 minus 05
Heme oxygenase 1 FANhyb icon00004395 a1g000011 258 27119864 minus 07 27119864 minus 07
12 International Journal of Genomics
Hematin
Endoplasmicreticulum
WD40 bLHLMYB
Structural genes
TFs
Nuclear
JA
ABA Sucrose
Plastid
G-cis-neoxanthin
Apophytochrome
StarchHeme
BVSucroseABA
Holophytochrome
PfrPIF3
Flavonoids Anthocyanins
Vacuole
Cytoplasm
Cell wall
①
②③
④⑤
⑥
⑦
PΦB
PΦB
CTKAuxin
Ca2+
Figure 5 Genes and biological pathways that were involved in regulation of anthocyanin accumulation promoted by hematin Outermostbox represents the cell wall Bilayer oval shape represents the plastidThe red-brown ring and network represent the nuclear and endoplasmicreticulum respectively The red crescent represents the vacuole The box in the nucleus represents transcription factors (TFs) Yellow MYBgreenWD40 and red bHLH represent a ternary transcription factor complex which transcribes anthocyanin biosynthesis genes The circlednumbers represent biological pathways involved in the regulation of anthocyanin accumulation promoted by hematin A represents thephytochrome regulation pathwayB represents the starch and sucrose pathwayCDE andF represent phytohormones ABA JA AuxinandCTK regulation pathways respectivelyG represents theCa2+ regulation pathway Pfr represents the far-red phytochrome PIF3 representsphytochrome-interacting factor Each anthocyanin biosynthesis regulatory pathway related gene is in Table 5
in anthocyanin biosynthesis were significantly altered withthe hematin treatment This suggests that the physiologicalprocess of fruit color development is regulated through com-plex interactions among anthocyanin biosynthesis pathwaysplant hormone signal transduction pathways phytochromesignal transduction pathways starch and sugar metabolicpathways calcium pathways and transcription factors (Fig-ure 5) This study adds to the in-depth understanding of thefruit coloration process in strawberry
Competing Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contributions
Jianwei Gao and Liangju Wang conceived and designed theresearch Yi Li Huayin Li Fengde Wang Jingjuan Li andYihui Zhang conducted experiments Yi Li analyzed the dataYi Li and Fengde Wang wrote the manuscript All authorsread and approved the manuscript Yi Li and Huayin Licontributed equally to this work
Acknowledgments
This work was supported by the Modern Agricultural Indus-trial Technology System Funding of Shandong ProvinceChina (SDAIT-02-022-04) the China agriculture researchsystem (CARS-25) and the National Science Foundation ofChina (31401820)
References
[1] K Aaby SMazur A Nes andG Skrede ldquoPhenolic compoundsin strawberry (Fragaria x ananassa Duch) fruits compositionin 27 cultivars and changes during ripeningrdquo Food Chemistryvol 132 no 1 pp 86ndash97 2012
[2] T A Colquhoun L A Levin H RMoskowitz VMWhitakerD G Clark and K M Folta ldquoFraming the perfect strawberryan exercise in consumer-assisted selection of fruit cropsrdquoJournal of Berry Research vol 2 no 1 pp 45ndash61 2012
[3] F Giampieri J M Alvarez-Suarez L Mazzoni et al ldquoThepotential impact of strawberry on human healthrdquo NaturalProduct Research vol 27 no 4 pp 448ndash455 2013
[4] Y S Velioglu GMazza L Gao and B D Oomah ldquoAntioxidantactivity and total phenolics in selected fruits vegetables and
International Journal of Genomics 13
grain productsrdquo Journal of Agricultural amp Food Chemistry vol46 no 10 pp 4113ndash4117 1998
[5] S S Hassellund A Flaa S E Kjeldsen et al ldquoEffects of antho-cyanins on cardiovascular risk factors and inflammation inpre-hypertensive men a double-blind randomized placebo-controlled crossover studyrdquo Journal of Human Hypertensionvol 27 no 2 pp 100ndash106 2013
[6] K J Meyers C B Watkins M P Pritts and R H Liu ldquoAnti-oxidant and antiproliferative activities of strawberriesrdquo Journalof Agricultural amp Food Chemistry vol 51 no 23 pp 6887ndash68922003
[7] S Zafra-Stone T YasminM Bagchi AChatterjee J AVinsonand D Bagchi ldquoBerry anthocyanins as novel antioxidants inhuman health and disease preventionrdquo Molecular Nutrition ampFood Research vol 51 no 6 pp 675ndash683 2007
[8] L Jaakola ldquoNew insights into the regulation of anthocyanin bio-synthesis in fruitsrdquoTrends in Plant Science vol 18 no 9 pp 477ndash483 2013
[9] K Springob J-I Nakajima M Yamazaki and K Saito ldquoRecentadvances in the biosynthesis and accumulation of antho-cyaninsrdquo Natural Product Reports vol 20 no 3 pp 288ndash3032003
[10] A Gonzalez M Zhao J M Leavitt and A M Lloyd ldquoRegu-lation of the anthocyanin biosynthetic pathway by the TTG1bHLHMyb transcriptional complex in Arabidopsis seedlingsrdquoPlant Journal vol 53 no 5 pp 814ndash827 2008
[11] K Petroni and C Tonelli ldquoRecent advances on the regulationof anthocyanin synthesis in reproductive organsrdquo Plant Sciencevol 181 no 3 pp 219ndash229 2011
[12] H-F Jia Y-M Chai C-L Li et al ldquoAbscisic acid plays animportant role in the regulation of strawberry fruit ripeningrdquoPlant Physiology vol 157 no 1 pp 188ndash199 2011
[13] C Merchante J G Vallarino S Osorio et al ldquoEthylene isinvolved in strawberry fruit ripening in an organ-specific man-nerrdquo Journal of Experimental Botany vol 64 no 14 pp 4421ndash4439 2013
[14] A Maier A Schrader L Kokkelink et al ldquoLight and the E3ubiquitin ligase COP1SPA control the protein stability of theMYB transcription factors PAP1 and PAP2 involved in antho-cyanin accumulation in Arabidopsisrdquo Plant Journal vol 74 no4 pp 638ndash651 2013
[15] H Lange W Shropshire and H Mohr ldquoAn analysis of phyto-chrome-mediated anthocyanin synthesisrdquo Plant Physiology vol47 no 5 pp 649ndash655 1971
[16] T D Elich and J C Lagarias ldquoPhytochrome chromophore bio-synthesis both 5-aminolevulinic acid and biliverdin overcomeinhibition by gabaculine in etiolated Avena sativa L seedlingsrdquoPlant Physiology vol 84 no 2 pp 304ndash310 1987
[17] Y Han W Xuan T Yu et al ldquoExogenous hematin alleviatesmercury-induced oxidative damage in the roots of Medicagosativardquo Journal of Integrative Plant Biology vol 49 no 12 pp1703ndash1713 2007
[18] M Koornneef J Cone R Dekens E OrsquoHerne-Robers CSpruit and R Kendrick ldquoPhotomorphogenic responses of longhypocotyl mutants of tomatordquo Journal of Plant Physiology vol120 no 2 pp 153ndash165 1985
[19] M J Terry and R E Kendrick ldquoFeedback inhibition of chloro-phyll synthesis in the phytochrome chromophore-deficientaurea and yellow-green-2 mutants of tomatordquo Plant Physiologyvol 119 no 1 pp 143ndash152 1999
[20] B M Parks and P H Quail ldquoPhytochrome-deficient hy1 andhy2 long hypocotyl mutants of Arabidopsis are defective inphytochrome chromophore biosynthesisrdquo Plant Cell vol 3 no11 pp 1177ndash1186 1991
[21] M J Terry ldquoPhytochrome chromophore-deficient mutantsrdquoPlant Cell amp Environment vol 20 no 6 pp 740ndash745 1997
[22] T Muramoto T Kohchi A Yokota I Hwang and H MGoodman ldquoThe Arabidopsis photomorphogenic mutant hy1 isdeficient in phytochrome chromophore biosynthesis as a resultof a mutation in a plastid heme oxygenaserdquo Plant Cell vol 11no 3 pp 335ndash347 1999
[23] M D Maines ldquoThe heme oxygenase system a regulator ofsecond messenger gasesrdquo Annual Review of Pharmacology andToxicology vol 37 no 1 pp 517ndash554 1997
[24] W Xuan F-Y Zhu S Xu et al ldquoThe heme oxygenasecarbonmonoxide system is involved in the auxin-induced cucumberadventitious rooting processrdquo Plant Physiology vol 148 no 2pp 881ndash893 2008
[25] R Amooaghaie F Tabatabaei and A-M Ahadi ldquoRole ofhematin and sodium nitroprusside in regulating Brassica nigraseed germination under nanosilver and silver nitrate stressesrdquoEcotoxicology and Environmental Safety vol 113 pp 259ndash2702015
[26] Y Liu X Li L Xu and W Shen ldquoDe-etiolation of wheatseedling leaves cross talk between heme oxygenasecarbonmonoxide and nitric oxiderdquo PLoS ONE vol 8 no 12 ArticleID e81470 2013
[27] T Muramoto N Tsurui M J Terry A Yokota and T KohchildquoExpression and biochemical properties of a ferredoxin-dependent heme oxygenase required for phytochrome chro-mophore synthesisrdquo Plant Physiology vol 130 no 4 pp 1958ndash1966 2002
[28] J-W Gao J Liu B Li and Z Li ldquoIsolation and purificationof functional total RNA from blue-grained wheat endospermtissues containing high levels of starches and flavonoidsrdquo PlantMolecular Biology Reporter vol 19 no 2 pp 185ndash186 2001
[29] R Patro S M Mount and C Kingsford ldquoSailfish enablesalignment-free isoform quantification from RNA-seq readsusing lightweight algorithmsrdquoNature Biotechnology vol 32 no5 pp 462ndash464 2014
[30] Y-X Qi Y-B Liu and W-H Rong ldquoRNA-Seq and its applica-tions a new technology for transcriptomicsrdquo Hereditas vol 33no 11 pp 1191ndash1202 2011
[31] C R Genovese N A Lazar and T Nichols ldquoThresholdingof statistical maps in functional neuroimaging using the falsediscovery raterdquo NeuroImage vol 15 no 4 pp 870ndash878 2002
[32] J Ye L Fang H Zheng et al ldquoWEGO a web tool for plottingGO annotationsrdquoNucleic Acids Research vol 34 supplement 2pp W293ndashW297 2006
[33] H Abdi ldquoThe Bonferonni and Sidak corrections for multiplecomparisonrdquo Encyclopedia of Measurement and Statistics vol 3pp 103ndash107 2007
[34] J Li Q Ding F Wang Y Zhang H Li and J Gao ldquoIntegrativeanalysis of mRNA and miRNA expression profiles of thetuberous root development at seedling stages in turnipsrdquo PLoSONE vol 10 no 9 Article ID e0137983 2015
[35] J Bennett D Hondred and J C Register ldquoKeeping qRT-PCRrigorous and biologically relevantrdquo Plant Cell Reports vol 34no 1 pp 1ndash3 2015
[36] F Wang L Li H Li et al ldquoTranscriptome analysis of rosetteand folding leaves in Chinese cabbage using high-throughputRNA sequencingrdquo Genomics vol 99 no 5 pp 299ndash307 2012
14 International Journal of Genomics
[37] T Shi Z Gao L Wang et al ldquoIdentification of differentially-expressed genes associated with pistil abortion in Japaneseapricot by genome-wide transcriptional analysisrdquo PLoS ONEvol 7 no 10 Article ID e47810 2012
[38] F Wang H Li Y Zhang et al ldquoMicroRNA expression analysisof rosette and folding leaves in Chinese cabbage using high-throughput Solexa sequencingrdquo Gene vol 532 no 2 pp 222ndash229 2013
[39] X Gu Y Chen Z Gao Y Qiao and X Wang ldquoTranscriptionfactors and anthocyanin genes related to low-temperature toler-ance in rd29A RdreB1BI transgenic strawberryrdquo Plant Physiol-ogy and Biochemistry vol 89 pp 31ndash43 2015
[40] J G Schaart C Dubos I R De La Fuente et al ldquoIdentificationand characterization of MYB-bHLH-WD40 regulatory com-plexes controlling proanthocyanidin biosynthesis in strawberry(Fragaria times ananassa) fruitsrdquoNew Phytologist vol 197 no 2 pp454ndash467 2013
[41] L Xie Z H Wang X H Cheng J J Gao Z P Zhangand L J Wang ldquo5-Aminolevulinic acid promotes anthocyaninaccumulation in Fuji applesrdquo Plant Growth Regulation vol 69no 3 pp 295ndash303 2013
[42] M Daminato F Guzzo and G Casadoro ldquoASHATTERPROOF-like gene controls ripening in non-climac-teric strawberries and auxin and abscisic acid antagonisticallyaffect its expressionrdquo Journal of Experimental Botany vol 64no 12 pp 3775ndash3786 2013
[43] X-H Ji R Zhang N Wang L Yang and X-S Chen ldquoTran-scriptome profiling reveals auxin suppressed anthocyanin bio-synthesis in red-fleshed apple callus (Malus sieversii f niedzwet-zkyana)rdquo Plant Cell Tissue and Organ Culture vol 123 no 2pp 389ndash404 2015
[44] J Chen LMaoW Lu T Ying and Z Luo ldquoTranscriptome pro-filing of postharvest strawberry fruit in response to exogenousauxin and abscisic acidrdquo Planta vol 243 no 1 pp 183ndash197 2016
[45] G A Martınez A R Chaves and M C Anon ldquoEffect of gib-berellic acid on ripening of strawberry fruits (Fragaria annan-assa Duch)rdquo Journal of Plant Growth Regulation vol 13 no 2pp 87ndash91 1994
[46] P K Das D H Shin S-B Choi S-D Yoo G Choi and Y-IPark ldquoCytokinins enhance sugar-induced anthocyanin biosyn-thesis in Arabidopsisrdquo Molecules amp Cells vol 34 no 1 pp 93ndash101 2012
[47] P McAtee S Karim R Schaffer and K David ldquoA dynamicinterplay between phytohormones is required for fruit develop-ment maturation and ripeningrdquo Frontiers in Plant Science vol4 no 3 article 79 7 pages 2013
[48] E Nambara andAMarion-Poll ldquoAbscisic acid biosynthesis andcatabolismrdquoAnnual Review of Plant Biology vol 56 pp 165ndash1852005
[49] Y Jiang and D C Joyce ldquoABA effects on ethylene productionPAL activity anthocyanin and phenolic contents of strawberryfruitrdquo Plant Growth Regulation vol 39 no 2 pp 171ndash174 2003
[50] S Li ldquoTranscriptional control of flavonoid biosynthesis fine-tuning of the MYB-bHLH-WD40 (MBW) complexrdquo PlantSignaling amp Behavior vol 9 Article ID e27522 2014
[51] D R Rudell J K Fellman and J P Mattheis ldquoPreharvest appli-cation of methyl jasmonate to rsquoFujirsquo apples enhances red col-oration and affects fruit size splitting and bitter pit incidencerdquoHortScience vol 40 no 6 pp 1760ndash1762 2005
[52] A Belhadj N Telef C Saigne et al ldquoEffect of methyl jasmonatein combination with carbohydrates on gene expression of PR
proteins stilbene and anthocyanin accumulation in grapevinecell culturesrdquo Plant Physiology and Biochemistry vol 46 no 4pp 493ndash499 2008
[53] E Loreti G Povero G Novi C Solfanelli A Alpi and PPerata ldquoGibberellins jasmonate and abscisic acid modulate thesucrose-induced expression of anthocyanin biosynthetic genesin Arabidopsisrdquo New Phytologist vol 179 no 4 pp 1004ndash10162008
[54] H S Seo J T Song J-J Cheong et al ldquoJasmonic acid carboxylmethyltransferase A key enzyme for jasmonate-regulated plantresponsesrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 98 no 8 pp 4788ndash4793 2001
[55] M T McDowell and J C Lagarias ldquoPurification and biochemi-cal properties of phytochromobilin synthase from etiolated oatseedlingsrdquo Plant Physiology vol 126 no 4 pp 1546ndash1554 2001
[56] G-Q Fu S Xu Y-J Xie et al ldquoMolecular cloning characteri-zation and expression of an alfalfa (Medicago sativa L) hemeoxygenase-1 gene MsHO1 which is pro-oxidants-regulatedrdquoPlant Physiology and Biochemistry vol 49 no 7 pp 792ndash7992011
[57] X-Y Chen X Ding S Xu et al ldquoEndogenous hydrogen per-oxide plays a positive role in the upregulation of heme oxy-genase and acclimation to oxidative stress in wheat seedlingleavesrdquo Journal of Integrative Plant Biology vol 51 no 10 pp951ndash960 2009
[58] J Shin E Park and G Choi ldquoPIF3 regulates anthocyaninbiosynthesis in an HY5-dependent manner with both factorsdirectly binding anthocyanin biosynthetic gene promoters inArabidopsisrdquo Plant Journal vol 49 no 6 pp 981ndash994 2007
[59] S Wang D Pan X Lv et al ldquoProteomic approach reveals thatstarch degradation contributes to anthocyanin accumulation intuberous root of purple sweet potatordquo Journal of Proteomics vol143 pp 298ndash305 2016
[60] H-F Jia C-L Li Y-M Chai Y Xing and Y Shen ldquoSucrosepromotes strawberry fruit ripening by stimulation of abscisicacid biosynthesisrdquo Pakistan Journal of Botany vol 45 no 1 pp169ndash176 2013
[61] W Xu H Peng T Yang et al ldquoEffect of calcium on strawberryfruit flavonoid pathway gene expression and anthocyanin accu-mulationrdquo Plant Physiology amp Biochemistry vol 82 pp 289ndash298 2014
[62] Z-W-SWan and Z Singh ldquoExogenous application of prohexa-dione-calcium promotes fruit colour development of lsquoCrippsPinkrsquo applerdquo Acta Horticulturae vol 1012 pp 219ndash225 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Genomics 9
Table 3 DEGs acting as transcription factors in response to exogenous hematin
Annotation transcription factors Gene IDReal
fold-changevalues
Upregulationdownregulation FDR
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb icon00006295 a1g000011 938 Up 120119864 minus 05
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL1-like
FANhyb rscf000031551g000011 4 Down 480119864 minus 16
PREDICTED AP2-likeethylene-responsive transcriptionfactor AIL5-like
FANhyb rscf000000291g000101 4 Down 320119864 minus 08
MYBdomain protein 66 FANhyb rscf000001461g000071 654 Up 780119864 minus 07MYB-related protein 3R-1-like partial FANhyb rscf000001411g000161 282 Up 830119864 minus 04PREDICTED transcription factorGAMYB-like FANhyb rscf000006491g000061 303 Down 150119864 minus 13
PREDICTED probable WRKYtranscription factor 53-like FANhyb rscf000019731g000021 217 Down 200119864 minus 08
Transcription factor bHLH80-like FANhyb rscf000012921g000031 6250 Down 150119864 minus 06Transcription factor bHLH70-like FANhyb rscf000002951g000031 526 Down 660119864 minus 12Transcription factor ICE1-like FANhyb rscf000001701g000061 244 Down 450119864 minus 18C2H2-like zinc finger protein FANhyb rscf000011431g000021 282 Up 590119864 minus 04Zinc finger protein MAGPIE-like FANhyb rscf000030241g000011 204 Up 150119864 minus 04PREDICTED axial regulator YABBY5-like FANhyb rscf000016671g000011 417 Down 730119864 minus 05
PREDICTED axial regulator YABBY1-like FANhyb rscf000068991g000011 357 Down 620119864 minus 06
PREDICTED dof zinc finger proteinDOF37-like isoform 1 FANhyb rscf000068561g000011 263 Down 280119864 minus 04
ZF-HD homeobox protein FANhyb rscf000000441g000221 385 Down 420119864 minus 08ZF-HD homeobox proteinAt4g24660-like FANhyb rscf000052411g000011 357 Down 470119864 minus 09
PREDICTED B3 domain-containingprotein REM14-like FANhyb rscf000010091g000021 243 Up 530119864 minus 04
PREDICTED B3 domain-containingtranscription factor ABI3-like FANhyb rscf000012711g000021 455 Down 130119864 minus 28
Growth-regulating factor 6 FANhyb rscf000000241g000291 345 Down 330119864 minus 34Growth-regulating factor 5 FANhyb icon00009704 a1g000011 27 Down 190119864 minus 06GATA type zinc finger transcriptionfactor family protein FANhyb rscf000003931g000131 1111 Down 110119864 minus 05
PREDICTED zinc finger CCCHdomain-containing protein 2-like FANhyb icon00006770 a1g000011 313 Down 680119864 minus 49
PREDICTED E2F transcriptionfactor-like E2FE-like FANhyb icon00002702 a1g000011 278 Down 240119864 minus 04
PREDICTED uncharacterized protein FANhyb rscf000010661g000011 263 Down 490119864 minus 06PREDICTED nuclear transcriptionfactor Y subunit A-3-like isoform 1 FANhyb rscf000000081g000351 385 Down 500119864 minus 20
PREDICTED homeobox-leucinezipper protein ATHB-8-like FANhyb rscf000000151g000051 263 Down 100119864 minus 06
PREDICTED MADS-boxtranscription factor 18-like FANhyb rscf000003231g000161 243 Down 180119864 minus 06
PREDICTED zinc finger protein132-like isoform 1 FANhyb rscf000070151g000011 243 Down 180119864 minus 08
PREDICTED transcription factorPIF3-like FANhyb rscf000006691g000021 346 Up 140119864 minus 07
10 International Journal of Genomics
Table 4 Regulation of the DEGs in anthocyanin biosynthesis in response to exogenous hematin
Genes in anthocyaninbiosynthesis Gene ID
Realfold-change
valuesRegulation level FDR
Dihydroflavonol-4-reductase FANhyb icon00002410 a1g000011 3091 Up 910119864 minus 06
Flavonol synthase FANhyb icon00015354 a1g000011 345 down 180119864 minus 05Isoflavone reductasehomolog FANhyb icon20341135 s1g000011 334 Up 750119864 minus 14
UDP-glycosyltransferase FANhyb rscf000049691g000011 307 Up 640119864 minus 08UDP-glycosyltransferaseactivity FANhyb icon15742070 o1g000011 18 30563 Up 510119864 minus 09
UDP-glycosyltransferase73C1 FANhyb icon00027696 a1g000011 298 Up 560119864 minus 04
UDP-glycosyltransferaseactivity FANhyb icon00008064 a1g000011 249 Up 310119864 minus 04
PREDICTEDUDP-glucose flavonoid3-O-glucosyltransferase7-like
FANhyb rscf000055631g000011 212 Up 110119864 minus 19
PREDICTEDUDP-glycosyltransferase76F1-like
FANhyb rscf000021771g000031 2 Up 720119864 minus 29
A transcripts in the etiolated leaves of wheat seedling phy-tochromes [26] However our study shows that phytochromeA was not differentially expressed It is probably becausethe hematin-treated strawberries were not shaded during theexperiment while the function of phytochrome A is light-dependent In addition we found that a downstream gene ofphytochrome PIF3 (FANhyb rscf000006691g000021) wassignificantly upregulated PIF3 is thought to be a positive reg-ulator of phytochrome B mediated by light-dependent signaltransduction [58]These results suggest that the phytochromepathways may be involved in the anthocyanin biosynthesispromoted by hematin
345 DEGs in Starch and Sucrose Degradation Proteomicapproaches have revealed that starch degradation contributesto anthocyanin accumulation in tuberous roots of the purplesweet potato variety [59] Starch and sucrose biosynthesis isimportant for anthocyanin accumulation in strawberry fruit[60] In this study we found the DEGs involved in starchand sucrose synthesis in the hematin-treated fruit were alldownregulated (Table 5) For example the expression of theunigenes encoding soluble starch synthase 3 sucrose trans-port protein SUC2-like and a probable sucrose-phosphatesynthase 4-like enzyme were decreased 23- 46- 32- and2-fold respectively This suggests that exogenous hematinregulates the biosynthesis of starch and sucrose and henceaffects fruit coloring
346 DEGs in the Calcium Pathway in Response to HematinCalcium can also increase the transcription levels of keystructural genes F3H DFR ANT and UFGT in the whitestage of strawberries [61] In this study we found thatthree DEGs in calcium biosynthesis and transport were
downregulated in the hematin-treated fruit (Table 5) Onlythe unigene encoding predicted cationcalcium exchanger5-like was upregulated The unigenes encoding a calpain-type cysteine protease and a predicted calcium-binding pro-tein PBP1-like were downregulated respectively Exogenousapplication of calcium can promote apple coloring [62] inaddition to variation in anthocyanin content [61] Our resultsindicate that the biosynthesis and transport of calcium areinvolved in the development of coloring in hematin-treatedstrawberries
35 Validation of Selected DEGs by qRT-PCR To validate theexpression of the DEGs obtained from RNA-Seq 11 DEGswere selected for qRT-PCR including structural genes (DFRand UDFGs) transcription factor genes (MYB and bHLH)and phytochrome chromophore-related gene (HO-1) Theprimers used for qRT-PCR are listed in Supplementary TableS1 The qRT-PCR results were consistent with the RNA-Seqdata (Figure 4) except for FANhyb rscf000001411g000161which had a higher log
2ratio (hematin-treatedcontrol) in
the transcriptome data than in qRT-PCR These results indi-cate the RNA-Seq data from the strawberry transcriptome isreproducible and accurate
4 Conclusions
In this study the anthocyanin content in the strawberryfruit was elevated by the application of exogenous hematinThis is the first report that hematin could increase theanthocyanin production in fruit Furthermore we exploredthe effects of the exogenous hematin on metabolic path-ways using genome-wide transcriptome analysis The resultsindicate that the expression levels of many genes involved
International Journal of Genomics 11
Table 5 Other DEGs identified in anthocyanin biosynthesis-related pathways
Anthocyaninbiosynthesis-relatedpathways
Annotation genes Gene IDReal fold-changevalues
Upregulationdownregulation FDR
Calcium ion binding
Cationcalciumexchanger 5-like
FANhyb rscf000019061g000011 203 Up 64119864 minus 06
Calpain-type cysteineprotease family
FANhyb icon00011755 a1g000011 345 Down 18119864 minus 09
Calcium-bindingprotein PBP1-like
FANhyb rscf000007501g000081 323 Down 97119864 minus 04
Calpain-type cysteineprotease family
FANhyb icon00023658 a1g000011 233 Down 60119864 minus 08
CytokininCytokinin
dehydrogenase 5-likeFANhyb iscf00325393 1 s1g000011 667 Down 62119864 minus 06
Cytochrome P450714A1
FANhyb rscf000005921g000031 217 Down 98119864 minus 10
JasmonateJasmonate
O-methyltransferase-like
FANhyb rscf000000041g000131 201 Up 70119864 minus 04
Abscisic acid
Protein ABSCISICACID-INSENSITIVE
5-likeFANhyb rscf000021641g000011 4 Down 16119864 minus 18
ABA overly sensitive 5 FANhyb icon00051144 a1g000011 291 Up 24119864 minus 059-cis-
epoxycarotenoiddioxygenase NCED1
FANhyb icon18399909 o1g000011 441 Up 43119864 minus 06
Abscisic acid81015840-hydroxylase 1-like
FANhyb icon00000938 a1g000011 256 Down 14119864 minus 06
Abscisic acid receptorPYR1-like
FANhyb icon00020426 a1g000011 208 Down 30119864 minus 06
Auxin
Auxin efflux carriercomponent 1-like
FANhyb rscf000010081g000031 384 Down 23119864 minus 06
Auxin response factor8
FANhyb rscf000013061g000021 263 Down 61119864 minus 17
Auxin response factor17-like
FANhyb rscf000060741g000011 217 Down 11119864 minus 36
Starch and sucrose
Glycosyltransferasefamily 35
FANhyb rscf000000341g000081 384 Down 11119864 minus 05
Soluble starchsynthase 3
FANhyb rscf000000451g000041 232 Down 38119864 minus 12
Sucrose transportprotein SUC2-like
FANhyb icon00036727 a1g000011 455 Down 98119864 minus 04
Sucrose transportprotein SUC2-like
FANhyb rscf000007551g000021 323 Down 66119864 minus 10
Probablesucrose-phosphatesynthase 4-like
FANhyb rscf000013501g000021 213 Down 11119864 minus 14
Phytochrome
PREDICTEDtranscription factor
PIF3-likeFANhyb rscf000006691g000021 346 up 14119864 minus 07
Phytochrome E-like FANhyb rscf000004361g000041 4 Down 33119864 minus 05
Heme oxygenase 1 FANhyb icon00004395 a1g000011 258 27119864 minus 07 27119864 minus 07
12 International Journal of Genomics
Hematin
Endoplasmicreticulum
WD40 bLHLMYB
Structural genes
TFs
Nuclear
JA
ABA Sucrose
Plastid
G-cis-neoxanthin
Apophytochrome
StarchHeme
BVSucroseABA
Holophytochrome
PfrPIF3
Flavonoids Anthocyanins
Vacuole
Cytoplasm
Cell wall
①
②③
④⑤
⑥
⑦
PΦB
PΦB
CTKAuxin
Ca2+
Figure 5 Genes and biological pathways that were involved in regulation of anthocyanin accumulation promoted by hematin Outermostbox represents the cell wall Bilayer oval shape represents the plastidThe red-brown ring and network represent the nuclear and endoplasmicreticulum respectively The red crescent represents the vacuole The box in the nucleus represents transcription factors (TFs) Yellow MYBgreenWD40 and red bHLH represent a ternary transcription factor complex which transcribes anthocyanin biosynthesis genes The circlednumbers represent biological pathways involved in the regulation of anthocyanin accumulation promoted by hematin A represents thephytochrome regulation pathwayB represents the starch and sucrose pathwayCDE andF represent phytohormones ABA JA AuxinandCTK regulation pathways respectivelyG represents theCa2+ regulation pathway Pfr represents the far-red phytochrome PIF3 representsphytochrome-interacting factor Each anthocyanin biosynthesis regulatory pathway related gene is in Table 5
in anthocyanin biosynthesis were significantly altered withthe hematin treatment This suggests that the physiologicalprocess of fruit color development is regulated through com-plex interactions among anthocyanin biosynthesis pathwaysplant hormone signal transduction pathways phytochromesignal transduction pathways starch and sugar metabolicpathways calcium pathways and transcription factors (Fig-ure 5) This study adds to the in-depth understanding of thefruit coloration process in strawberry
Competing Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contributions
Jianwei Gao and Liangju Wang conceived and designed theresearch Yi Li Huayin Li Fengde Wang Jingjuan Li andYihui Zhang conducted experiments Yi Li analyzed the dataYi Li and Fengde Wang wrote the manuscript All authorsread and approved the manuscript Yi Li and Huayin Licontributed equally to this work
Acknowledgments
This work was supported by the Modern Agricultural Indus-trial Technology System Funding of Shandong ProvinceChina (SDAIT-02-022-04) the China agriculture researchsystem (CARS-25) and the National Science Foundation ofChina (31401820)
References
[1] K Aaby SMazur A Nes andG Skrede ldquoPhenolic compoundsin strawberry (Fragaria x ananassa Duch) fruits compositionin 27 cultivars and changes during ripeningrdquo Food Chemistryvol 132 no 1 pp 86ndash97 2012
[2] T A Colquhoun L A Levin H RMoskowitz VMWhitakerD G Clark and K M Folta ldquoFraming the perfect strawberryan exercise in consumer-assisted selection of fruit cropsrdquoJournal of Berry Research vol 2 no 1 pp 45ndash61 2012
[3] F Giampieri J M Alvarez-Suarez L Mazzoni et al ldquoThepotential impact of strawberry on human healthrdquo NaturalProduct Research vol 27 no 4 pp 448ndash455 2013
[4] Y S Velioglu GMazza L Gao and B D Oomah ldquoAntioxidantactivity and total phenolics in selected fruits vegetables and
International Journal of Genomics 13
grain productsrdquo Journal of Agricultural amp Food Chemistry vol46 no 10 pp 4113ndash4117 1998
[5] S S Hassellund A Flaa S E Kjeldsen et al ldquoEffects of antho-cyanins on cardiovascular risk factors and inflammation inpre-hypertensive men a double-blind randomized placebo-controlled crossover studyrdquo Journal of Human Hypertensionvol 27 no 2 pp 100ndash106 2013
[6] K J Meyers C B Watkins M P Pritts and R H Liu ldquoAnti-oxidant and antiproliferative activities of strawberriesrdquo Journalof Agricultural amp Food Chemistry vol 51 no 23 pp 6887ndash68922003
[7] S Zafra-Stone T YasminM Bagchi AChatterjee J AVinsonand D Bagchi ldquoBerry anthocyanins as novel antioxidants inhuman health and disease preventionrdquo Molecular Nutrition ampFood Research vol 51 no 6 pp 675ndash683 2007
[8] L Jaakola ldquoNew insights into the regulation of anthocyanin bio-synthesis in fruitsrdquoTrends in Plant Science vol 18 no 9 pp 477ndash483 2013
[9] K Springob J-I Nakajima M Yamazaki and K Saito ldquoRecentadvances in the biosynthesis and accumulation of antho-cyaninsrdquo Natural Product Reports vol 20 no 3 pp 288ndash3032003
[10] A Gonzalez M Zhao J M Leavitt and A M Lloyd ldquoRegu-lation of the anthocyanin biosynthetic pathway by the TTG1bHLHMyb transcriptional complex in Arabidopsis seedlingsrdquoPlant Journal vol 53 no 5 pp 814ndash827 2008
[11] K Petroni and C Tonelli ldquoRecent advances on the regulationof anthocyanin synthesis in reproductive organsrdquo Plant Sciencevol 181 no 3 pp 219ndash229 2011
[12] H-F Jia Y-M Chai C-L Li et al ldquoAbscisic acid plays animportant role in the regulation of strawberry fruit ripeningrdquoPlant Physiology vol 157 no 1 pp 188ndash199 2011
[13] C Merchante J G Vallarino S Osorio et al ldquoEthylene isinvolved in strawberry fruit ripening in an organ-specific man-nerrdquo Journal of Experimental Botany vol 64 no 14 pp 4421ndash4439 2013
[14] A Maier A Schrader L Kokkelink et al ldquoLight and the E3ubiquitin ligase COP1SPA control the protein stability of theMYB transcription factors PAP1 and PAP2 involved in antho-cyanin accumulation in Arabidopsisrdquo Plant Journal vol 74 no4 pp 638ndash651 2013
[15] H Lange W Shropshire and H Mohr ldquoAn analysis of phyto-chrome-mediated anthocyanin synthesisrdquo Plant Physiology vol47 no 5 pp 649ndash655 1971
[16] T D Elich and J C Lagarias ldquoPhytochrome chromophore bio-synthesis both 5-aminolevulinic acid and biliverdin overcomeinhibition by gabaculine in etiolated Avena sativa L seedlingsrdquoPlant Physiology vol 84 no 2 pp 304ndash310 1987
[17] Y Han W Xuan T Yu et al ldquoExogenous hematin alleviatesmercury-induced oxidative damage in the roots of Medicagosativardquo Journal of Integrative Plant Biology vol 49 no 12 pp1703ndash1713 2007
[18] M Koornneef J Cone R Dekens E OrsquoHerne-Robers CSpruit and R Kendrick ldquoPhotomorphogenic responses of longhypocotyl mutants of tomatordquo Journal of Plant Physiology vol120 no 2 pp 153ndash165 1985
[19] M J Terry and R E Kendrick ldquoFeedback inhibition of chloro-phyll synthesis in the phytochrome chromophore-deficientaurea and yellow-green-2 mutants of tomatordquo Plant Physiologyvol 119 no 1 pp 143ndash152 1999
[20] B M Parks and P H Quail ldquoPhytochrome-deficient hy1 andhy2 long hypocotyl mutants of Arabidopsis are defective inphytochrome chromophore biosynthesisrdquo Plant Cell vol 3 no11 pp 1177ndash1186 1991
[21] M J Terry ldquoPhytochrome chromophore-deficient mutantsrdquoPlant Cell amp Environment vol 20 no 6 pp 740ndash745 1997
[22] T Muramoto T Kohchi A Yokota I Hwang and H MGoodman ldquoThe Arabidopsis photomorphogenic mutant hy1 isdeficient in phytochrome chromophore biosynthesis as a resultof a mutation in a plastid heme oxygenaserdquo Plant Cell vol 11no 3 pp 335ndash347 1999
[23] M D Maines ldquoThe heme oxygenase system a regulator ofsecond messenger gasesrdquo Annual Review of Pharmacology andToxicology vol 37 no 1 pp 517ndash554 1997
[24] W Xuan F-Y Zhu S Xu et al ldquoThe heme oxygenasecarbonmonoxide system is involved in the auxin-induced cucumberadventitious rooting processrdquo Plant Physiology vol 148 no 2pp 881ndash893 2008
[25] R Amooaghaie F Tabatabaei and A-M Ahadi ldquoRole ofhematin and sodium nitroprusside in regulating Brassica nigraseed germination under nanosilver and silver nitrate stressesrdquoEcotoxicology and Environmental Safety vol 113 pp 259ndash2702015
[26] Y Liu X Li L Xu and W Shen ldquoDe-etiolation of wheatseedling leaves cross talk between heme oxygenasecarbonmonoxide and nitric oxiderdquo PLoS ONE vol 8 no 12 ArticleID e81470 2013
[27] T Muramoto N Tsurui M J Terry A Yokota and T KohchildquoExpression and biochemical properties of a ferredoxin-dependent heme oxygenase required for phytochrome chro-mophore synthesisrdquo Plant Physiology vol 130 no 4 pp 1958ndash1966 2002
[28] J-W Gao J Liu B Li and Z Li ldquoIsolation and purificationof functional total RNA from blue-grained wheat endospermtissues containing high levels of starches and flavonoidsrdquo PlantMolecular Biology Reporter vol 19 no 2 pp 185ndash186 2001
[29] R Patro S M Mount and C Kingsford ldquoSailfish enablesalignment-free isoform quantification from RNA-seq readsusing lightweight algorithmsrdquoNature Biotechnology vol 32 no5 pp 462ndash464 2014
[30] Y-X Qi Y-B Liu and W-H Rong ldquoRNA-Seq and its applica-tions a new technology for transcriptomicsrdquo Hereditas vol 33no 11 pp 1191ndash1202 2011
[31] C R Genovese N A Lazar and T Nichols ldquoThresholdingof statistical maps in functional neuroimaging using the falsediscovery raterdquo NeuroImage vol 15 no 4 pp 870ndash878 2002
[32] J Ye L Fang H Zheng et al ldquoWEGO a web tool for plottingGO annotationsrdquoNucleic Acids Research vol 34 supplement 2pp W293ndashW297 2006
[33] H Abdi ldquoThe Bonferonni and Sidak corrections for multiplecomparisonrdquo Encyclopedia of Measurement and Statistics vol 3pp 103ndash107 2007
[34] J Li Q Ding F Wang Y Zhang H Li and J Gao ldquoIntegrativeanalysis of mRNA and miRNA expression profiles of thetuberous root development at seedling stages in turnipsrdquo PLoSONE vol 10 no 9 Article ID e0137983 2015
[35] J Bennett D Hondred and J C Register ldquoKeeping qRT-PCRrigorous and biologically relevantrdquo Plant Cell Reports vol 34no 1 pp 1ndash3 2015
[36] F Wang L Li H Li et al ldquoTranscriptome analysis of rosetteand folding leaves in Chinese cabbage using high-throughputRNA sequencingrdquo Genomics vol 99 no 5 pp 299ndash307 2012
14 International Journal of Genomics
[37] T Shi Z Gao L Wang et al ldquoIdentification of differentially-expressed genes associated with pistil abortion in Japaneseapricot by genome-wide transcriptional analysisrdquo PLoS ONEvol 7 no 10 Article ID e47810 2012
[38] F Wang H Li Y Zhang et al ldquoMicroRNA expression analysisof rosette and folding leaves in Chinese cabbage using high-throughput Solexa sequencingrdquo Gene vol 532 no 2 pp 222ndash229 2013
[39] X Gu Y Chen Z Gao Y Qiao and X Wang ldquoTranscriptionfactors and anthocyanin genes related to low-temperature toler-ance in rd29A RdreB1BI transgenic strawberryrdquo Plant Physiol-ogy and Biochemistry vol 89 pp 31ndash43 2015
[40] J G Schaart C Dubos I R De La Fuente et al ldquoIdentificationand characterization of MYB-bHLH-WD40 regulatory com-plexes controlling proanthocyanidin biosynthesis in strawberry(Fragaria times ananassa) fruitsrdquoNew Phytologist vol 197 no 2 pp454ndash467 2013
[41] L Xie Z H Wang X H Cheng J J Gao Z P Zhangand L J Wang ldquo5-Aminolevulinic acid promotes anthocyaninaccumulation in Fuji applesrdquo Plant Growth Regulation vol 69no 3 pp 295ndash303 2013
[42] M Daminato F Guzzo and G Casadoro ldquoASHATTERPROOF-like gene controls ripening in non-climac-teric strawberries and auxin and abscisic acid antagonisticallyaffect its expressionrdquo Journal of Experimental Botany vol 64no 12 pp 3775ndash3786 2013
[43] X-H Ji R Zhang N Wang L Yang and X-S Chen ldquoTran-scriptome profiling reveals auxin suppressed anthocyanin bio-synthesis in red-fleshed apple callus (Malus sieversii f niedzwet-zkyana)rdquo Plant Cell Tissue and Organ Culture vol 123 no 2pp 389ndash404 2015
[44] J Chen LMaoW Lu T Ying and Z Luo ldquoTranscriptome pro-filing of postharvest strawberry fruit in response to exogenousauxin and abscisic acidrdquo Planta vol 243 no 1 pp 183ndash197 2016
[45] G A Martınez A R Chaves and M C Anon ldquoEffect of gib-berellic acid on ripening of strawberry fruits (Fragaria annan-assa Duch)rdquo Journal of Plant Growth Regulation vol 13 no 2pp 87ndash91 1994
[46] P K Das D H Shin S-B Choi S-D Yoo G Choi and Y-IPark ldquoCytokinins enhance sugar-induced anthocyanin biosyn-thesis in Arabidopsisrdquo Molecules amp Cells vol 34 no 1 pp 93ndash101 2012
[47] P McAtee S Karim R Schaffer and K David ldquoA dynamicinterplay between phytohormones is required for fruit develop-ment maturation and ripeningrdquo Frontiers in Plant Science vol4 no 3 article 79 7 pages 2013
[48] E Nambara andAMarion-Poll ldquoAbscisic acid biosynthesis andcatabolismrdquoAnnual Review of Plant Biology vol 56 pp 165ndash1852005
[49] Y Jiang and D C Joyce ldquoABA effects on ethylene productionPAL activity anthocyanin and phenolic contents of strawberryfruitrdquo Plant Growth Regulation vol 39 no 2 pp 171ndash174 2003
[50] S Li ldquoTranscriptional control of flavonoid biosynthesis fine-tuning of the MYB-bHLH-WD40 (MBW) complexrdquo PlantSignaling amp Behavior vol 9 Article ID e27522 2014
[51] D R Rudell J K Fellman and J P Mattheis ldquoPreharvest appli-cation of methyl jasmonate to rsquoFujirsquo apples enhances red col-oration and affects fruit size splitting and bitter pit incidencerdquoHortScience vol 40 no 6 pp 1760ndash1762 2005
[52] A Belhadj N Telef C Saigne et al ldquoEffect of methyl jasmonatein combination with carbohydrates on gene expression of PR
proteins stilbene and anthocyanin accumulation in grapevinecell culturesrdquo Plant Physiology and Biochemistry vol 46 no 4pp 493ndash499 2008
[53] E Loreti G Povero G Novi C Solfanelli A Alpi and PPerata ldquoGibberellins jasmonate and abscisic acid modulate thesucrose-induced expression of anthocyanin biosynthetic genesin Arabidopsisrdquo New Phytologist vol 179 no 4 pp 1004ndash10162008
[54] H S Seo J T Song J-J Cheong et al ldquoJasmonic acid carboxylmethyltransferase A key enzyme for jasmonate-regulated plantresponsesrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 98 no 8 pp 4788ndash4793 2001
[55] M T McDowell and J C Lagarias ldquoPurification and biochemi-cal properties of phytochromobilin synthase from etiolated oatseedlingsrdquo Plant Physiology vol 126 no 4 pp 1546ndash1554 2001
[56] G-Q Fu S Xu Y-J Xie et al ldquoMolecular cloning characteri-zation and expression of an alfalfa (Medicago sativa L) hemeoxygenase-1 gene MsHO1 which is pro-oxidants-regulatedrdquoPlant Physiology and Biochemistry vol 49 no 7 pp 792ndash7992011
[57] X-Y Chen X Ding S Xu et al ldquoEndogenous hydrogen per-oxide plays a positive role in the upregulation of heme oxy-genase and acclimation to oxidative stress in wheat seedlingleavesrdquo Journal of Integrative Plant Biology vol 51 no 10 pp951ndash960 2009
[58] J Shin E Park and G Choi ldquoPIF3 regulates anthocyaninbiosynthesis in an HY5-dependent manner with both factorsdirectly binding anthocyanin biosynthetic gene promoters inArabidopsisrdquo Plant Journal vol 49 no 6 pp 981ndash994 2007
[59] S Wang D Pan X Lv et al ldquoProteomic approach reveals thatstarch degradation contributes to anthocyanin accumulation intuberous root of purple sweet potatordquo Journal of Proteomics vol143 pp 298ndash305 2016
[60] H-F Jia C-L Li Y-M Chai Y Xing and Y Shen ldquoSucrosepromotes strawberry fruit ripening by stimulation of abscisicacid biosynthesisrdquo Pakistan Journal of Botany vol 45 no 1 pp169ndash176 2013
[61] W Xu H Peng T Yang et al ldquoEffect of calcium on strawberryfruit flavonoid pathway gene expression and anthocyanin accu-mulationrdquo Plant Physiology amp Biochemistry vol 82 pp 289ndash298 2014
[62] Z-W-SWan and Z Singh ldquoExogenous application of prohexa-dione-calcium promotes fruit colour development of lsquoCrippsPinkrsquo applerdquo Acta Horticulturae vol 1012 pp 219ndash225 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
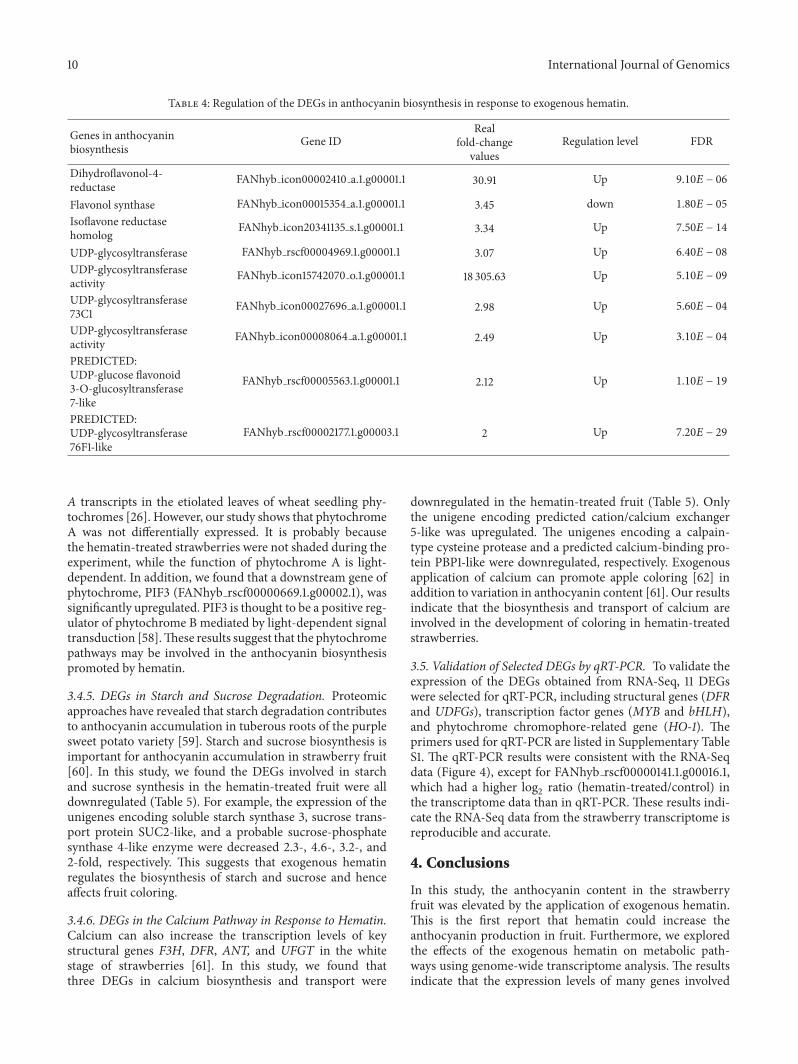
10 International Journal of Genomics
Table 4 Regulation of the DEGs in anthocyanin biosynthesis in response to exogenous hematin
Genes in anthocyaninbiosynthesis Gene ID
Realfold-change
valuesRegulation level FDR
Dihydroflavonol-4-reductase FANhyb icon00002410 a1g000011 3091 Up 910119864 minus 06
Flavonol synthase FANhyb icon00015354 a1g000011 345 down 180119864 minus 05Isoflavone reductasehomolog FANhyb icon20341135 s1g000011 334 Up 750119864 minus 14
UDP-glycosyltransferase FANhyb rscf000049691g000011 307 Up 640119864 minus 08UDP-glycosyltransferaseactivity FANhyb icon15742070 o1g000011 18 30563 Up 510119864 minus 09
UDP-glycosyltransferase73C1 FANhyb icon00027696 a1g000011 298 Up 560119864 minus 04
UDP-glycosyltransferaseactivity FANhyb icon00008064 a1g000011 249 Up 310119864 minus 04
PREDICTEDUDP-glucose flavonoid3-O-glucosyltransferase7-like
FANhyb rscf000055631g000011 212 Up 110119864 minus 19
PREDICTEDUDP-glycosyltransferase76F1-like
FANhyb rscf000021771g000031 2 Up 720119864 minus 29
A transcripts in the etiolated leaves of wheat seedling phy-tochromes [26] However our study shows that phytochromeA was not differentially expressed It is probably becausethe hematin-treated strawberries were not shaded during theexperiment while the function of phytochrome A is light-dependent In addition we found that a downstream gene ofphytochrome PIF3 (FANhyb rscf000006691g000021) wassignificantly upregulated PIF3 is thought to be a positive reg-ulator of phytochrome B mediated by light-dependent signaltransduction [58]These results suggest that the phytochromepathways may be involved in the anthocyanin biosynthesispromoted by hematin
345 DEGs in Starch and Sucrose Degradation Proteomicapproaches have revealed that starch degradation contributesto anthocyanin accumulation in tuberous roots of the purplesweet potato variety [59] Starch and sucrose biosynthesis isimportant for anthocyanin accumulation in strawberry fruit[60] In this study we found the DEGs involved in starchand sucrose synthesis in the hematin-treated fruit were alldownregulated (Table 5) For example the expression of theunigenes encoding soluble starch synthase 3 sucrose trans-port protein SUC2-like and a probable sucrose-phosphatesynthase 4-like enzyme were decreased 23- 46- 32- and2-fold respectively This suggests that exogenous hematinregulates the biosynthesis of starch and sucrose and henceaffects fruit coloring
346 DEGs in the Calcium Pathway in Response to HematinCalcium can also increase the transcription levels of keystructural genes F3H DFR ANT and UFGT in the whitestage of strawberries [61] In this study we found thatthree DEGs in calcium biosynthesis and transport were
downregulated in the hematin-treated fruit (Table 5) Onlythe unigene encoding predicted cationcalcium exchanger5-like was upregulated The unigenes encoding a calpain-type cysteine protease and a predicted calcium-binding pro-tein PBP1-like were downregulated respectively Exogenousapplication of calcium can promote apple coloring [62] inaddition to variation in anthocyanin content [61] Our resultsindicate that the biosynthesis and transport of calcium areinvolved in the development of coloring in hematin-treatedstrawberries
35 Validation of Selected DEGs by qRT-PCR To validate theexpression of the DEGs obtained from RNA-Seq 11 DEGswere selected for qRT-PCR including structural genes (DFRand UDFGs) transcription factor genes (MYB and bHLH)and phytochrome chromophore-related gene (HO-1) Theprimers used for qRT-PCR are listed in Supplementary TableS1 The qRT-PCR results were consistent with the RNA-Seqdata (Figure 4) except for FANhyb rscf000001411g000161which had a higher log
2ratio (hematin-treatedcontrol) in
the transcriptome data than in qRT-PCR These results indi-cate the RNA-Seq data from the strawberry transcriptome isreproducible and accurate
4 Conclusions
In this study the anthocyanin content in the strawberryfruit was elevated by the application of exogenous hematinThis is the first report that hematin could increase theanthocyanin production in fruit Furthermore we exploredthe effects of the exogenous hematin on metabolic path-ways using genome-wide transcriptome analysis The resultsindicate that the expression levels of many genes involved
International Journal of Genomics 11
Table 5 Other DEGs identified in anthocyanin biosynthesis-related pathways
Anthocyaninbiosynthesis-relatedpathways
Annotation genes Gene IDReal fold-changevalues
Upregulationdownregulation FDR
Calcium ion binding
Cationcalciumexchanger 5-like
FANhyb rscf000019061g000011 203 Up 64119864 minus 06
Calpain-type cysteineprotease family
FANhyb icon00011755 a1g000011 345 Down 18119864 minus 09
Calcium-bindingprotein PBP1-like
FANhyb rscf000007501g000081 323 Down 97119864 minus 04
Calpain-type cysteineprotease family
FANhyb icon00023658 a1g000011 233 Down 60119864 minus 08
CytokininCytokinin
dehydrogenase 5-likeFANhyb iscf00325393 1 s1g000011 667 Down 62119864 minus 06
Cytochrome P450714A1
FANhyb rscf000005921g000031 217 Down 98119864 minus 10
JasmonateJasmonate
O-methyltransferase-like
FANhyb rscf000000041g000131 201 Up 70119864 minus 04
Abscisic acid
Protein ABSCISICACID-INSENSITIVE
5-likeFANhyb rscf000021641g000011 4 Down 16119864 minus 18
ABA overly sensitive 5 FANhyb icon00051144 a1g000011 291 Up 24119864 minus 059-cis-
epoxycarotenoiddioxygenase NCED1
FANhyb icon18399909 o1g000011 441 Up 43119864 minus 06
Abscisic acid81015840-hydroxylase 1-like
FANhyb icon00000938 a1g000011 256 Down 14119864 minus 06
Abscisic acid receptorPYR1-like
FANhyb icon00020426 a1g000011 208 Down 30119864 minus 06
Auxin
Auxin efflux carriercomponent 1-like
FANhyb rscf000010081g000031 384 Down 23119864 minus 06
Auxin response factor8
FANhyb rscf000013061g000021 263 Down 61119864 minus 17
Auxin response factor17-like
FANhyb rscf000060741g000011 217 Down 11119864 minus 36
Starch and sucrose
Glycosyltransferasefamily 35
FANhyb rscf000000341g000081 384 Down 11119864 minus 05
Soluble starchsynthase 3
FANhyb rscf000000451g000041 232 Down 38119864 minus 12
Sucrose transportprotein SUC2-like
FANhyb icon00036727 a1g000011 455 Down 98119864 minus 04
Sucrose transportprotein SUC2-like
FANhyb rscf000007551g000021 323 Down 66119864 minus 10
Probablesucrose-phosphatesynthase 4-like
FANhyb rscf000013501g000021 213 Down 11119864 minus 14
Phytochrome
PREDICTEDtranscription factor
PIF3-likeFANhyb rscf000006691g000021 346 up 14119864 minus 07
Phytochrome E-like FANhyb rscf000004361g000041 4 Down 33119864 minus 05
Heme oxygenase 1 FANhyb icon00004395 a1g000011 258 27119864 minus 07 27119864 minus 07
12 International Journal of Genomics
Hematin
Endoplasmicreticulum
WD40 bLHLMYB
Structural genes
TFs
Nuclear
JA
ABA Sucrose
Plastid
G-cis-neoxanthin
Apophytochrome
StarchHeme
BVSucroseABA
Holophytochrome
PfrPIF3
Flavonoids Anthocyanins
Vacuole
Cytoplasm
Cell wall
①
②③
④⑤
⑥
⑦
PΦB
PΦB
CTKAuxin
Ca2+
Figure 5 Genes and biological pathways that were involved in regulation of anthocyanin accumulation promoted by hematin Outermostbox represents the cell wall Bilayer oval shape represents the plastidThe red-brown ring and network represent the nuclear and endoplasmicreticulum respectively The red crescent represents the vacuole The box in the nucleus represents transcription factors (TFs) Yellow MYBgreenWD40 and red bHLH represent a ternary transcription factor complex which transcribes anthocyanin biosynthesis genes The circlednumbers represent biological pathways involved in the regulation of anthocyanin accumulation promoted by hematin A represents thephytochrome regulation pathwayB represents the starch and sucrose pathwayCDE andF represent phytohormones ABA JA AuxinandCTK regulation pathways respectivelyG represents theCa2+ regulation pathway Pfr represents the far-red phytochrome PIF3 representsphytochrome-interacting factor Each anthocyanin biosynthesis regulatory pathway related gene is in Table 5
in anthocyanin biosynthesis were significantly altered withthe hematin treatment This suggests that the physiologicalprocess of fruit color development is regulated through com-plex interactions among anthocyanin biosynthesis pathwaysplant hormone signal transduction pathways phytochromesignal transduction pathways starch and sugar metabolicpathways calcium pathways and transcription factors (Fig-ure 5) This study adds to the in-depth understanding of thefruit coloration process in strawberry
Competing Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contributions
Jianwei Gao and Liangju Wang conceived and designed theresearch Yi Li Huayin Li Fengde Wang Jingjuan Li andYihui Zhang conducted experiments Yi Li analyzed the dataYi Li and Fengde Wang wrote the manuscript All authorsread and approved the manuscript Yi Li and Huayin Licontributed equally to this work
Acknowledgments
This work was supported by the Modern Agricultural Indus-trial Technology System Funding of Shandong ProvinceChina (SDAIT-02-022-04) the China agriculture researchsystem (CARS-25) and the National Science Foundation ofChina (31401820)
References
[1] K Aaby SMazur A Nes andG Skrede ldquoPhenolic compoundsin strawberry (Fragaria x ananassa Duch) fruits compositionin 27 cultivars and changes during ripeningrdquo Food Chemistryvol 132 no 1 pp 86ndash97 2012
[2] T A Colquhoun L A Levin H RMoskowitz VMWhitakerD G Clark and K M Folta ldquoFraming the perfect strawberryan exercise in consumer-assisted selection of fruit cropsrdquoJournal of Berry Research vol 2 no 1 pp 45ndash61 2012
[3] F Giampieri J M Alvarez-Suarez L Mazzoni et al ldquoThepotential impact of strawberry on human healthrdquo NaturalProduct Research vol 27 no 4 pp 448ndash455 2013
[4] Y S Velioglu GMazza L Gao and B D Oomah ldquoAntioxidantactivity and total phenolics in selected fruits vegetables and
International Journal of Genomics 13
grain productsrdquo Journal of Agricultural amp Food Chemistry vol46 no 10 pp 4113ndash4117 1998
[5] S S Hassellund A Flaa S E Kjeldsen et al ldquoEffects of antho-cyanins on cardiovascular risk factors and inflammation inpre-hypertensive men a double-blind randomized placebo-controlled crossover studyrdquo Journal of Human Hypertensionvol 27 no 2 pp 100ndash106 2013
[6] K J Meyers C B Watkins M P Pritts and R H Liu ldquoAnti-oxidant and antiproliferative activities of strawberriesrdquo Journalof Agricultural amp Food Chemistry vol 51 no 23 pp 6887ndash68922003
[7] S Zafra-Stone T YasminM Bagchi AChatterjee J AVinsonand D Bagchi ldquoBerry anthocyanins as novel antioxidants inhuman health and disease preventionrdquo Molecular Nutrition ampFood Research vol 51 no 6 pp 675ndash683 2007
[8] L Jaakola ldquoNew insights into the regulation of anthocyanin bio-synthesis in fruitsrdquoTrends in Plant Science vol 18 no 9 pp 477ndash483 2013
[9] K Springob J-I Nakajima M Yamazaki and K Saito ldquoRecentadvances in the biosynthesis and accumulation of antho-cyaninsrdquo Natural Product Reports vol 20 no 3 pp 288ndash3032003
[10] A Gonzalez M Zhao J M Leavitt and A M Lloyd ldquoRegu-lation of the anthocyanin biosynthetic pathway by the TTG1bHLHMyb transcriptional complex in Arabidopsis seedlingsrdquoPlant Journal vol 53 no 5 pp 814ndash827 2008
[11] K Petroni and C Tonelli ldquoRecent advances on the regulationof anthocyanin synthesis in reproductive organsrdquo Plant Sciencevol 181 no 3 pp 219ndash229 2011
[12] H-F Jia Y-M Chai C-L Li et al ldquoAbscisic acid plays animportant role in the regulation of strawberry fruit ripeningrdquoPlant Physiology vol 157 no 1 pp 188ndash199 2011
[13] C Merchante J G Vallarino S Osorio et al ldquoEthylene isinvolved in strawberry fruit ripening in an organ-specific man-nerrdquo Journal of Experimental Botany vol 64 no 14 pp 4421ndash4439 2013
[14] A Maier A Schrader L Kokkelink et al ldquoLight and the E3ubiquitin ligase COP1SPA control the protein stability of theMYB transcription factors PAP1 and PAP2 involved in antho-cyanin accumulation in Arabidopsisrdquo Plant Journal vol 74 no4 pp 638ndash651 2013
[15] H Lange W Shropshire and H Mohr ldquoAn analysis of phyto-chrome-mediated anthocyanin synthesisrdquo Plant Physiology vol47 no 5 pp 649ndash655 1971
[16] T D Elich and J C Lagarias ldquoPhytochrome chromophore bio-synthesis both 5-aminolevulinic acid and biliverdin overcomeinhibition by gabaculine in etiolated Avena sativa L seedlingsrdquoPlant Physiology vol 84 no 2 pp 304ndash310 1987
[17] Y Han W Xuan T Yu et al ldquoExogenous hematin alleviatesmercury-induced oxidative damage in the roots of Medicagosativardquo Journal of Integrative Plant Biology vol 49 no 12 pp1703ndash1713 2007
[18] M Koornneef J Cone R Dekens E OrsquoHerne-Robers CSpruit and R Kendrick ldquoPhotomorphogenic responses of longhypocotyl mutants of tomatordquo Journal of Plant Physiology vol120 no 2 pp 153ndash165 1985
[19] M J Terry and R E Kendrick ldquoFeedback inhibition of chloro-phyll synthesis in the phytochrome chromophore-deficientaurea and yellow-green-2 mutants of tomatordquo Plant Physiologyvol 119 no 1 pp 143ndash152 1999
[20] B M Parks and P H Quail ldquoPhytochrome-deficient hy1 andhy2 long hypocotyl mutants of Arabidopsis are defective inphytochrome chromophore biosynthesisrdquo Plant Cell vol 3 no11 pp 1177ndash1186 1991
[21] M J Terry ldquoPhytochrome chromophore-deficient mutantsrdquoPlant Cell amp Environment vol 20 no 6 pp 740ndash745 1997
[22] T Muramoto T Kohchi A Yokota I Hwang and H MGoodman ldquoThe Arabidopsis photomorphogenic mutant hy1 isdeficient in phytochrome chromophore biosynthesis as a resultof a mutation in a plastid heme oxygenaserdquo Plant Cell vol 11no 3 pp 335ndash347 1999
[23] M D Maines ldquoThe heme oxygenase system a regulator ofsecond messenger gasesrdquo Annual Review of Pharmacology andToxicology vol 37 no 1 pp 517ndash554 1997
[24] W Xuan F-Y Zhu S Xu et al ldquoThe heme oxygenasecarbonmonoxide system is involved in the auxin-induced cucumberadventitious rooting processrdquo Plant Physiology vol 148 no 2pp 881ndash893 2008
[25] R Amooaghaie F Tabatabaei and A-M Ahadi ldquoRole ofhematin and sodium nitroprusside in regulating Brassica nigraseed germination under nanosilver and silver nitrate stressesrdquoEcotoxicology and Environmental Safety vol 113 pp 259ndash2702015
[26] Y Liu X Li L Xu and W Shen ldquoDe-etiolation of wheatseedling leaves cross talk between heme oxygenasecarbonmonoxide and nitric oxiderdquo PLoS ONE vol 8 no 12 ArticleID e81470 2013
[27] T Muramoto N Tsurui M J Terry A Yokota and T KohchildquoExpression and biochemical properties of a ferredoxin-dependent heme oxygenase required for phytochrome chro-mophore synthesisrdquo Plant Physiology vol 130 no 4 pp 1958ndash1966 2002
[28] J-W Gao J Liu B Li and Z Li ldquoIsolation and purificationof functional total RNA from blue-grained wheat endospermtissues containing high levels of starches and flavonoidsrdquo PlantMolecular Biology Reporter vol 19 no 2 pp 185ndash186 2001
[29] R Patro S M Mount and C Kingsford ldquoSailfish enablesalignment-free isoform quantification from RNA-seq readsusing lightweight algorithmsrdquoNature Biotechnology vol 32 no5 pp 462ndash464 2014
[30] Y-X Qi Y-B Liu and W-H Rong ldquoRNA-Seq and its applica-tions a new technology for transcriptomicsrdquo Hereditas vol 33no 11 pp 1191ndash1202 2011
[31] C R Genovese N A Lazar and T Nichols ldquoThresholdingof statistical maps in functional neuroimaging using the falsediscovery raterdquo NeuroImage vol 15 no 4 pp 870ndash878 2002
[32] J Ye L Fang H Zheng et al ldquoWEGO a web tool for plottingGO annotationsrdquoNucleic Acids Research vol 34 supplement 2pp W293ndashW297 2006
[33] H Abdi ldquoThe Bonferonni and Sidak corrections for multiplecomparisonrdquo Encyclopedia of Measurement and Statistics vol 3pp 103ndash107 2007
[34] J Li Q Ding F Wang Y Zhang H Li and J Gao ldquoIntegrativeanalysis of mRNA and miRNA expression profiles of thetuberous root development at seedling stages in turnipsrdquo PLoSONE vol 10 no 9 Article ID e0137983 2015
[35] J Bennett D Hondred and J C Register ldquoKeeping qRT-PCRrigorous and biologically relevantrdquo Plant Cell Reports vol 34no 1 pp 1ndash3 2015
[36] F Wang L Li H Li et al ldquoTranscriptome analysis of rosetteand folding leaves in Chinese cabbage using high-throughputRNA sequencingrdquo Genomics vol 99 no 5 pp 299ndash307 2012
14 International Journal of Genomics
[37] T Shi Z Gao L Wang et al ldquoIdentification of differentially-expressed genes associated with pistil abortion in Japaneseapricot by genome-wide transcriptional analysisrdquo PLoS ONEvol 7 no 10 Article ID e47810 2012
[38] F Wang H Li Y Zhang et al ldquoMicroRNA expression analysisof rosette and folding leaves in Chinese cabbage using high-throughput Solexa sequencingrdquo Gene vol 532 no 2 pp 222ndash229 2013
[39] X Gu Y Chen Z Gao Y Qiao and X Wang ldquoTranscriptionfactors and anthocyanin genes related to low-temperature toler-ance in rd29A RdreB1BI transgenic strawberryrdquo Plant Physiol-ogy and Biochemistry vol 89 pp 31ndash43 2015
[40] J G Schaart C Dubos I R De La Fuente et al ldquoIdentificationand characterization of MYB-bHLH-WD40 regulatory com-plexes controlling proanthocyanidin biosynthesis in strawberry(Fragaria times ananassa) fruitsrdquoNew Phytologist vol 197 no 2 pp454ndash467 2013
[41] L Xie Z H Wang X H Cheng J J Gao Z P Zhangand L J Wang ldquo5-Aminolevulinic acid promotes anthocyaninaccumulation in Fuji applesrdquo Plant Growth Regulation vol 69no 3 pp 295ndash303 2013
[42] M Daminato F Guzzo and G Casadoro ldquoASHATTERPROOF-like gene controls ripening in non-climac-teric strawberries and auxin and abscisic acid antagonisticallyaffect its expressionrdquo Journal of Experimental Botany vol 64no 12 pp 3775ndash3786 2013
[43] X-H Ji R Zhang N Wang L Yang and X-S Chen ldquoTran-scriptome profiling reveals auxin suppressed anthocyanin bio-synthesis in red-fleshed apple callus (Malus sieversii f niedzwet-zkyana)rdquo Plant Cell Tissue and Organ Culture vol 123 no 2pp 389ndash404 2015
[44] J Chen LMaoW Lu T Ying and Z Luo ldquoTranscriptome pro-filing of postharvest strawberry fruit in response to exogenousauxin and abscisic acidrdquo Planta vol 243 no 1 pp 183ndash197 2016
[45] G A Martınez A R Chaves and M C Anon ldquoEffect of gib-berellic acid on ripening of strawberry fruits (Fragaria annan-assa Duch)rdquo Journal of Plant Growth Regulation vol 13 no 2pp 87ndash91 1994
[46] P K Das D H Shin S-B Choi S-D Yoo G Choi and Y-IPark ldquoCytokinins enhance sugar-induced anthocyanin biosyn-thesis in Arabidopsisrdquo Molecules amp Cells vol 34 no 1 pp 93ndash101 2012
[47] P McAtee S Karim R Schaffer and K David ldquoA dynamicinterplay between phytohormones is required for fruit develop-ment maturation and ripeningrdquo Frontiers in Plant Science vol4 no 3 article 79 7 pages 2013
[48] E Nambara andAMarion-Poll ldquoAbscisic acid biosynthesis andcatabolismrdquoAnnual Review of Plant Biology vol 56 pp 165ndash1852005
[49] Y Jiang and D C Joyce ldquoABA effects on ethylene productionPAL activity anthocyanin and phenolic contents of strawberryfruitrdquo Plant Growth Regulation vol 39 no 2 pp 171ndash174 2003
[50] S Li ldquoTranscriptional control of flavonoid biosynthesis fine-tuning of the MYB-bHLH-WD40 (MBW) complexrdquo PlantSignaling amp Behavior vol 9 Article ID e27522 2014
[51] D R Rudell J K Fellman and J P Mattheis ldquoPreharvest appli-cation of methyl jasmonate to rsquoFujirsquo apples enhances red col-oration and affects fruit size splitting and bitter pit incidencerdquoHortScience vol 40 no 6 pp 1760ndash1762 2005
[52] A Belhadj N Telef C Saigne et al ldquoEffect of methyl jasmonatein combination with carbohydrates on gene expression of PR
proteins stilbene and anthocyanin accumulation in grapevinecell culturesrdquo Plant Physiology and Biochemistry vol 46 no 4pp 493ndash499 2008
[53] E Loreti G Povero G Novi C Solfanelli A Alpi and PPerata ldquoGibberellins jasmonate and abscisic acid modulate thesucrose-induced expression of anthocyanin biosynthetic genesin Arabidopsisrdquo New Phytologist vol 179 no 4 pp 1004ndash10162008
[54] H S Seo J T Song J-J Cheong et al ldquoJasmonic acid carboxylmethyltransferase A key enzyme for jasmonate-regulated plantresponsesrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 98 no 8 pp 4788ndash4793 2001
[55] M T McDowell and J C Lagarias ldquoPurification and biochemi-cal properties of phytochromobilin synthase from etiolated oatseedlingsrdquo Plant Physiology vol 126 no 4 pp 1546ndash1554 2001
[56] G-Q Fu S Xu Y-J Xie et al ldquoMolecular cloning characteri-zation and expression of an alfalfa (Medicago sativa L) hemeoxygenase-1 gene MsHO1 which is pro-oxidants-regulatedrdquoPlant Physiology and Biochemistry vol 49 no 7 pp 792ndash7992011
[57] X-Y Chen X Ding S Xu et al ldquoEndogenous hydrogen per-oxide plays a positive role in the upregulation of heme oxy-genase and acclimation to oxidative stress in wheat seedlingleavesrdquo Journal of Integrative Plant Biology vol 51 no 10 pp951ndash960 2009
[58] J Shin E Park and G Choi ldquoPIF3 regulates anthocyaninbiosynthesis in an HY5-dependent manner with both factorsdirectly binding anthocyanin biosynthetic gene promoters inArabidopsisrdquo Plant Journal vol 49 no 6 pp 981ndash994 2007
[59] S Wang D Pan X Lv et al ldquoProteomic approach reveals thatstarch degradation contributes to anthocyanin accumulation intuberous root of purple sweet potatordquo Journal of Proteomics vol143 pp 298ndash305 2016
[60] H-F Jia C-L Li Y-M Chai Y Xing and Y Shen ldquoSucrosepromotes strawberry fruit ripening by stimulation of abscisicacid biosynthesisrdquo Pakistan Journal of Botany vol 45 no 1 pp169ndash176 2013
[61] W Xu H Peng T Yang et al ldquoEffect of calcium on strawberryfruit flavonoid pathway gene expression and anthocyanin accu-mulationrdquo Plant Physiology amp Biochemistry vol 82 pp 289ndash298 2014
[62] Z-W-SWan and Z Singh ldquoExogenous application of prohexa-dione-calcium promotes fruit colour development of lsquoCrippsPinkrsquo applerdquo Acta Horticulturae vol 1012 pp 219ndash225 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Genomics 11
Table 5 Other DEGs identified in anthocyanin biosynthesis-related pathways
Anthocyaninbiosynthesis-relatedpathways
Annotation genes Gene IDReal fold-changevalues
Upregulationdownregulation FDR
Calcium ion binding
Cationcalciumexchanger 5-like
FANhyb rscf000019061g000011 203 Up 64119864 minus 06
Calpain-type cysteineprotease family
FANhyb icon00011755 a1g000011 345 Down 18119864 minus 09
Calcium-bindingprotein PBP1-like
FANhyb rscf000007501g000081 323 Down 97119864 minus 04
Calpain-type cysteineprotease family
FANhyb icon00023658 a1g000011 233 Down 60119864 minus 08
CytokininCytokinin
dehydrogenase 5-likeFANhyb iscf00325393 1 s1g000011 667 Down 62119864 minus 06
Cytochrome P450714A1
FANhyb rscf000005921g000031 217 Down 98119864 minus 10
JasmonateJasmonate
O-methyltransferase-like
FANhyb rscf000000041g000131 201 Up 70119864 minus 04
Abscisic acid
Protein ABSCISICACID-INSENSITIVE
5-likeFANhyb rscf000021641g000011 4 Down 16119864 minus 18
ABA overly sensitive 5 FANhyb icon00051144 a1g000011 291 Up 24119864 minus 059-cis-
epoxycarotenoiddioxygenase NCED1
FANhyb icon18399909 o1g000011 441 Up 43119864 minus 06
Abscisic acid81015840-hydroxylase 1-like
FANhyb icon00000938 a1g000011 256 Down 14119864 minus 06
Abscisic acid receptorPYR1-like
FANhyb icon00020426 a1g000011 208 Down 30119864 minus 06
Auxin
Auxin efflux carriercomponent 1-like
FANhyb rscf000010081g000031 384 Down 23119864 minus 06
Auxin response factor8
FANhyb rscf000013061g000021 263 Down 61119864 minus 17
Auxin response factor17-like
FANhyb rscf000060741g000011 217 Down 11119864 minus 36
Starch and sucrose
Glycosyltransferasefamily 35
FANhyb rscf000000341g000081 384 Down 11119864 minus 05
Soluble starchsynthase 3
FANhyb rscf000000451g000041 232 Down 38119864 minus 12
Sucrose transportprotein SUC2-like
FANhyb icon00036727 a1g000011 455 Down 98119864 minus 04
Sucrose transportprotein SUC2-like
FANhyb rscf000007551g000021 323 Down 66119864 minus 10
Probablesucrose-phosphatesynthase 4-like
FANhyb rscf000013501g000021 213 Down 11119864 minus 14
Phytochrome
PREDICTEDtranscription factor
PIF3-likeFANhyb rscf000006691g000021 346 up 14119864 minus 07
Phytochrome E-like FANhyb rscf000004361g000041 4 Down 33119864 minus 05
Heme oxygenase 1 FANhyb icon00004395 a1g000011 258 27119864 minus 07 27119864 minus 07
12 International Journal of Genomics
Hematin
Endoplasmicreticulum
WD40 bLHLMYB
Structural genes
TFs
Nuclear
JA
ABA Sucrose
Plastid
G-cis-neoxanthin
Apophytochrome
StarchHeme
BVSucroseABA
Holophytochrome
PfrPIF3
Flavonoids Anthocyanins
Vacuole
Cytoplasm
Cell wall
①
②③
④⑤
⑥
⑦
PΦB
PΦB
CTKAuxin
Ca2+
Figure 5 Genes and biological pathways that were involved in regulation of anthocyanin accumulation promoted by hematin Outermostbox represents the cell wall Bilayer oval shape represents the plastidThe red-brown ring and network represent the nuclear and endoplasmicreticulum respectively The red crescent represents the vacuole The box in the nucleus represents transcription factors (TFs) Yellow MYBgreenWD40 and red bHLH represent a ternary transcription factor complex which transcribes anthocyanin biosynthesis genes The circlednumbers represent biological pathways involved in the regulation of anthocyanin accumulation promoted by hematin A represents thephytochrome regulation pathwayB represents the starch and sucrose pathwayCDE andF represent phytohormones ABA JA AuxinandCTK regulation pathways respectivelyG represents theCa2+ regulation pathway Pfr represents the far-red phytochrome PIF3 representsphytochrome-interacting factor Each anthocyanin biosynthesis regulatory pathway related gene is in Table 5
in anthocyanin biosynthesis were significantly altered withthe hematin treatment This suggests that the physiologicalprocess of fruit color development is regulated through com-plex interactions among anthocyanin biosynthesis pathwaysplant hormone signal transduction pathways phytochromesignal transduction pathways starch and sugar metabolicpathways calcium pathways and transcription factors (Fig-ure 5) This study adds to the in-depth understanding of thefruit coloration process in strawberry
Competing Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contributions
Jianwei Gao and Liangju Wang conceived and designed theresearch Yi Li Huayin Li Fengde Wang Jingjuan Li andYihui Zhang conducted experiments Yi Li analyzed the dataYi Li and Fengde Wang wrote the manuscript All authorsread and approved the manuscript Yi Li and Huayin Licontributed equally to this work
Acknowledgments
This work was supported by the Modern Agricultural Indus-trial Technology System Funding of Shandong ProvinceChina (SDAIT-02-022-04) the China agriculture researchsystem (CARS-25) and the National Science Foundation ofChina (31401820)
References
[1] K Aaby SMazur A Nes andG Skrede ldquoPhenolic compoundsin strawberry (Fragaria x ananassa Duch) fruits compositionin 27 cultivars and changes during ripeningrdquo Food Chemistryvol 132 no 1 pp 86ndash97 2012
[2] T A Colquhoun L A Levin H RMoskowitz VMWhitakerD G Clark and K M Folta ldquoFraming the perfect strawberryan exercise in consumer-assisted selection of fruit cropsrdquoJournal of Berry Research vol 2 no 1 pp 45ndash61 2012
[3] F Giampieri J M Alvarez-Suarez L Mazzoni et al ldquoThepotential impact of strawberry on human healthrdquo NaturalProduct Research vol 27 no 4 pp 448ndash455 2013
[4] Y S Velioglu GMazza L Gao and B D Oomah ldquoAntioxidantactivity and total phenolics in selected fruits vegetables and
International Journal of Genomics 13
grain productsrdquo Journal of Agricultural amp Food Chemistry vol46 no 10 pp 4113ndash4117 1998
[5] S S Hassellund A Flaa S E Kjeldsen et al ldquoEffects of antho-cyanins on cardiovascular risk factors and inflammation inpre-hypertensive men a double-blind randomized placebo-controlled crossover studyrdquo Journal of Human Hypertensionvol 27 no 2 pp 100ndash106 2013
[6] K J Meyers C B Watkins M P Pritts and R H Liu ldquoAnti-oxidant and antiproliferative activities of strawberriesrdquo Journalof Agricultural amp Food Chemistry vol 51 no 23 pp 6887ndash68922003
[7] S Zafra-Stone T YasminM Bagchi AChatterjee J AVinsonand D Bagchi ldquoBerry anthocyanins as novel antioxidants inhuman health and disease preventionrdquo Molecular Nutrition ampFood Research vol 51 no 6 pp 675ndash683 2007
[8] L Jaakola ldquoNew insights into the regulation of anthocyanin bio-synthesis in fruitsrdquoTrends in Plant Science vol 18 no 9 pp 477ndash483 2013
[9] K Springob J-I Nakajima M Yamazaki and K Saito ldquoRecentadvances in the biosynthesis and accumulation of antho-cyaninsrdquo Natural Product Reports vol 20 no 3 pp 288ndash3032003
[10] A Gonzalez M Zhao J M Leavitt and A M Lloyd ldquoRegu-lation of the anthocyanin biosynthetic pathway by the TTG1bHLHMyb transcriptional complex in Arabidopsis seedlingsrdquoPlant Journal vol 53 no 5 pp 814ndash827 2008
[11] K Petroni and C Tonelli ldquoRecent advances on the regulationof anthocyanin synthesis in reproductive organsrdquo Plant Sciencevol 181 no 3 pp 219ndash229 2011
[12] H-F Jia Y-M Chai C-L Li et al ldquoAbscisic acid plays animportant role in the regulation of strawberry fruit ripeningrdquoPlant Physiology vol 157 no 1 pp 188ndash199 2011
[13] C Merchante J G Vallarino S Osorio et al ldquoEthylene isinvolved in strawberry fruit ripening in an organ-specific man-nerrdquo Journal of Experimental Botany vol 64 no 14 pp 4421ndash4439 2013
[14] A Maier A Schrader L Kokkelink et al ldquoLight and the E3ubiquitin ligase COP1SPA control the protein stability of theMYB transcription factors PAP1 and PAP2 involved in antho-cyanin accumulation in Arabidopsisrdquo Plant Journal vol 74 no4 pp 638ndash651 2013
[15] H Lange W Shropshire and H Mohr ldquoAn analysis of phyto-chrome-mediated anthocyanin synthesisrdquo Plant Physiology vol47 no 5 pp 649ndash655 1971
[16] T D Elich and J C Lagarias ldquoPhytochrome chromophore bio-synthesis both 5-aminolevulinic acid and biliverdin overcomeinhibition by gabaculine in etiolated Avena sativa L seedlingsrdquoPlant Physiology vol 84 no 2 pp 304ndash310 1987
[17] Y Han W Xuan T Yu et al ldquoExogenous hematin alleviatesmercury-induced oxidative damage in the roots of Medicagosativardquo Journal of Integrative Plant Biology vol 49 no 12 pp1703ndash1713 2007
[18] M Koornneef J Cone R Dekens E OrsquoHerne-Robers CSpruit and R Kendrick ldquoPhotomorphogenic responses of longhypocotyl mutants of tomatordquo Journal of Plant Physiology vol120 no 2 pp 153ndash165 1985
[19] M J Terry and R E Kendrick ldquoFeedback inhibition of chloro-phyll synthesis in the phytochrome chromophore-deficientaurea and yellow-green-2 mutants of tomatordquo Plant Physiologyvol 119 no 1 pp 143ndash152 1999
[20] B M Parks and P H Quail ldquoPhytochrome-deficient hy1 andhy2 long hypocotyl mutants of Arabidopsis are defective inphytochrome chromophore biosynthesisrdquo Plant Cell vol 3 no11 pp 1177ndash1186 1991
[21] M J Terry ldquoPhytochrome chromophore-deficient mutantsrdquoPlant Cell amp Environment vol 20 no 6 pp 740ndash745 1997
[22] T Muramoto T Kohchi A Yokota I Hwang and H MGoodman ldquoThe Arabidopsis photomorphogenic mutant hy1 isdeficient in phytochrome chromophore biosynthesis as a resultof a mutation in a plastid heme oxygenaserdquo Plant Cell vol 11no 3 pp 335ndash347 1999
[23] M D Maines ldquoThe heme oxygenase system a regulator ofsecond messenger gasesrdquo Annual Review of Pharmacology andToxicology vol 37 no 1 pp 517ndash554 1997
[24] W Xuan F-Y Zhu S Xu et al ldquoThe heme oxygenasecarbonmonoxide system is involved in the auxin-induced cucumberadventitious rooting processrdquo Plant Physiology vol 148 no 2pp 881ndash893 2008
[25] R Amooaghaie F Tabatabaei and A-M Ahadi ldquoRole ofhematin and sodium nitroprusside in regulating Brassica nigraseed germination under nanosilver and silver nitrate stressesrdquoEcotoxicology and Environmental Safety vol 113 pp 259ndash2702015
[26] Y Liu X Li L Xu and W Shen ldquoDe-etiolation of wheatseedling leaves cross talk between heme oxygenasecarbonmonoxide and nitric oxiderdquo PLoS ONE vol 8 no 12 ArticleID e81470 2013
[27] T Muramoto N Tsurui M J Terry A Yokota and T KohchildquoExpression and biochemical properties of a ferredoxin-dependent heme oxygenase required for phytochrome chro-mophore synthesisrdquo Plant Physiology vol 130 no 4 pp 1958ndash1966 2002
[28] J-W Gao J Liu B Li and Z Li ldquoIsolation and purificationof functional total RNA from blue-grained wheat endospermtissues containing high levels of starches and flavonoidsrdquo PlantMolecular Biology Reporter vol 19 no 2 pp 185ndash186 2001
[29] R Patro S M Mount and C Kingsford ldquoSailfish enablesalignment-free isoform quantification from RNA-seq readsusing lightweight algorithmsrdquoNature Biotechnology vol 32 no5 pp 462ndash464 2014
[30] Y-X Qi Y-B Liu and W-H Rong ldquoRNA-Seq and its applica-tions a new technology for transcriptomicsrdquo Hereditas vol 33no 11 pp 1191ndash1202 2011
[31] C R Genovese N A Lazar and T Nichols ldquoThresholdingof statistical maps in functional neuroimaging using the falsediscovery raterdquo NeuroImage vol 15 no 4 pp 870ndash878 2002
[32] J Ye L Fang H Zheng et al ldquoWEGO a web tool for plottingGO annotationsrdquoNucleic Acids Research vol 34 supplement 2pp W293ndashW297 2006
[33] H Abdi ldquoThe Bonferonni and Sidak corrections for multiplecomparisonrdquo Encyclopedia of Measurement and Statistics vol 3pp 103ndash107 2007
[34] J Li Q Ding F Wang Y Zhang H Li and J Gao ldquoIntegrativeanalysis of mRNA and miRNA expression profiles of thetuberous root development at seedling stages in turnipsrdquo PLoSONE vol 10 no 9 Article ID e0137983 2015
[35] J Bennett D Hondred and J C Register ldquoKeeping qRT-PCRrigorous and biologically relevantrdquo Plant Cell Reports vol 34no 1 pp 1ndash3 2015
[36] F Wang L Li H Li et al ldquoTranscriptome analysis of rosetteand folding leaves in Chinese cabbage using high-throughputRNA sequencingrdquo Genomics vol 99 no 5 pp 299ndash307 2012
14 International Journal of Genomics
[37] T Shi Z Gao L Wang et al ldquoIdentification of differentially-expressed genes associated with pistil abortion in Japaneseapricot by genome-wide transcriptional analysisrdquo PLoS ONEvol 7 no 10 Article ID e47810 2012
[38] F Wang H Li Y Zhang et al ldquoMicroRNA expression analysisof rosette and folding leaves in Chinese cabbage using high-throughput Solexa sequencingrdquo Gene vol 532 no 2 pp 222ndash229 2013
[39] X Gu Y Chen Z Gao Y Qiao and X Wang ldquoTranscriptionfactors and anthocyanin genes related to low-temperature toler-ance in rd29A RdreB1BI transgenic strawberryrdquo Plant Physiol-ogy and Biochemistry vol 89 pp 31ndash43 2015
[40] J G Schaart C Dubos I R De La Fuente et al ldquoIdentificationand characterization of MYB-bHLH-WD40 regulatory com-plexes controlling proanthocyanidin biosynthesis in strawberry(Fragaria times ananassa) fruitsrdquoNew Phytologist vol 197 no 2 pp454ndash467 2013
[41] L Xie Z H Wang X H Cheng J J Gao Z P Zhangand L J Wang ldquo5-Aminolevulinic acid promotes anthocyaninaccumulation in Fuji applesrdquo Plant Growth Regulation vol 69no 3 pp 295ndash303 2013
[42] M Daminato F Guzzo and G Casadoro ldquoASHATTERPROOF-like gene controls ripening in non-climac-teric strawberries and auxin and abscisic acid antagonisticallyaffect its expressionrdquo Journal of Experimental Botany vol 64no 12 pp 3775ndash3786 2013
[43] X-H Ji R Zhang N Wang L Yang and X-S Chen ldquoTran-scriptome profiling reveals auxin suppressed anthocyanin bio-synthesis in red-fleshed apple callus (Malus sieversii f niedzwet-zkyana)rdquo Plant Cell Tissue and Organ Culture vol 123 no 2pp 389ndash404 2015
[44] J Chen LMaoW Lu T Ying and Z Luo ldquoTranscriptome pro-filing of postharvest strawberry fruit in response to exogenousauxin and abscisic acidrdquo Planta vol 243 no 1 pp 183ndash197 2016
[45] G A Martınez A R Chaves and M C Anon ldquoEffect of gib-berellic acid on ripening of strawberry fruits (Fragaria annan-assa Duch)rdquo Journal of Plant Growth Regulation vol 13 no 2pp 87ndash91 1994
[46] P K Das D H Shin S-B Choi S-D Yoo G Choi and Y-IPark ldquoCytokinins enhance sugar-induced anthocyanin biosyn-thesis in Arabidopsisrdquo Molecules amp Cells vol 34 no 1 pp 93ndash101 2012
[47] P McAtee S Karim R Schaffer and K David ldquoA dynamicinterplay between phytohormones is required for fruit develop-ment maturation and ripeningrdquo Frontiers in Plant Science vol4 no 3 article 79 7 pages 2013
[48] E Nambara andAMarion-Poll ldquoAbscisic acid biosynthesis andcatabolismrdquoAnnual Review of Plant Biology vol 56 pp 165ndash1852005
[49] Y Jiang and D C Joyce ldquoABA effects on ethylene productionPAL activity anthocyanin and phenolic contents of strawberryfruitrdquo Plant Growth Regulation vol 39 no 2 pp 171ndash174 2003
[50] S Li ldquoTranscriptional control of flavonoid biosynthesis fine-tuning of the MYB-bHLH-WD40 (MBW) complexrdquo PlantSignaling amp Behavior vol 9 Article ID e27522 2014
[51] D R Rudell J K Fellman and J P Mattheis ldquoPreharvest appli-cation of methyl jasmonate to rsquoFujirsquo apples enhances red col-oration and affects fruit size splitting and bitter pit incidencerdquoHortScience vol 40 no 6 pp 1760ndash1762 2005
[52] A Belhadj N Telef C Saigne et al ldquoEffect of methyl jasmonatein combination with carbohydrates on gene expression of PR
proteins stilbene and anthocyanin accumulation in grapevinecell culturesrdquo Plant Physiology and Biochemistry vol 46 no 4pp 493ndash499 2008
[53] E Loreti G Povero G Novi C Solfanelli A Alpi and PPerata ldquoGibberellins jasmonate and abscisic acid modulate thesucrose-induced expression of anthocyanin biosynthetic genesin Arabidopsisrdquo New Phytologist vol 179 no 4 pp 1004ndash10162008
[54] H S Seo J T Song J-J Cheong et al ldquoJasmonic acid carboxylmethyltransferase A key enzyme for jasmonate-regulated plantresponsesrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 98 no 8 pp 4788ndash4793 2001
[55] M T McDowell and J C Lagarias ldquoPurification and biochemi-cal properties of phytochromobilin synthase from etiolated oatseedlingsrdquo Plant Physiology vol 126 no 4 pp 1546ndash1554 2001
[56] G-Q Fu S Xu Y-J Xie et al ldquoMolecular cloning characteri-zation and expression of an alfalfa (Medicago sativa L) hemeoxygenase-1 gene MsHO1 which is pro-oxidants-regulatedrdquoPlant Physiology and Biochemistry vol 49 no 7 pp 792ndash7992011
[57] X-Y Chen X Ding S Xu et al ldquoEndogenous hydrogen per-oxide plays a positive role in the upregulation of heme oxy-genase and acclimation to oxidative stress in wheat seedlingleavesrdquo Journal of Integrative Plant Biology vol 51 no 10 pp951ndash960 2009
[58] J Shin E Park and G Choi ldquoPIF3 regulates anthocyaninbiosynthesis in an HY5-dependent manner with both factorsdirectly binding anthocyanin biosynthetic gene promoters inArabidopsisrdquo Plant Journal vol 49 no 6 pp 981ndash994 2007
[59] S Wang D Pan X Lv et al ldquoProteomic approach reveals thatstarch degradation contributes to anthocyanin accumulation intuberous root of purple sweet potatordquo Journal of Proteomics vol143 pp 298ndash305 2016
[60] H-F Jia C-L Li Y-M Chai Y Xing and Y Shen ldquoSucrosepromotes strawberry fruit ripening by stimulation of abscisicacid biosynthesisrdquo Pakistan Journal of Botany vol 45 no 1 pp169ndash176 2013
[61] W Xu H Peng T Yang et al ldquoEffect of calcium on strawberryfruit flavonoid pathway gene expression and anthocyanin accu-mulationrdquo Plant Physiology amp Biochemistry vol 82 pp 289ndash298 2014
[62] Z-W-SWan and Z Singh ldquoExogenous application of prohexa-dione-calcium promotes fruit colour development of lsquoCrippsPinkrsquo applerdquo Acta Horticulturae vol 1012 pp 219ndash225 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

12 International Journal of Genomics
Hematin
Endoplasmicreticulum
WD40 bLHLMYB
Structural genes
TFs
Nuclear
JA
ABA Sucrose
Plastid
G-cis-neoxanthin
Apophytochrome
StarchHeme
BVSucroseABA
Holophytochrome
PfrPIF3
Flavonoids Anthocyanins
Vacuole
Cytoplasm
Cell wall
①
②③
④⑤
⑥
⑦
PΦB
PΦB
CTKAuxin
Ca2+
Figure 5 Genes and biological pathways that were involved in regulation of anthocyanin accumulation promoted by hematin Outermostbox represents the cell wall Bilayer oval shape represents the plastidThe red-brown ring and network represent the nuclear and endoplasmicreticulum respectively The red crescent represents the vacuole The box in the nucleus represents transcription factors (TFs) Yellow MYBgreenWD40 and red bHLH represent a ternary transcription factor complex which transcribes anthocyanin biosynthesis genes The circlednumbers represent biological pathways involved in the regulation of anthocyanin accumulation promoted by hematin A represents thephytochrome regulation pathwayB represents the starch and sucrose pathwayCDE andF represent phytohormones ABA JA AuxinandCTK regulation pathways respectivelyG represents theCa2+ regulation pathway Pfr represents the far-red phytochrome PIF3 representsphytochrome-interacting factor Each anthocyanin biosynthesis regulatory pathway related gene is in Table 5
in anthocyanin biosynthesis were significantly altered withthe hematin treatment This suggests that the physiologicalprocess of fruit color development is regulated through com-plex interactions among anthocyanin biosynthesis pathwaysplant hormone signal transduction pathways phytochromesignal transduction pathways starch and sugar metabolicpathways calcium pathways and transcription factors (Fig-ure 5) This study adds to the in-depth understanding of thefruit coloration process in strawberry
Competing Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contributions
Jianwei Gao and Liangju Wang conceived and designed theresearch Yi Li Huayin Li Fengde Wang Jingjuan Li andYihui Zhang conducted experiments Yi Li analyzed the dataYi Li and Fengde Wang wrote the manuscript All authorsread and approved the manuscript Yi Li and Huayin Licontributed equally to this work
Acknowledgments
This work was supported by the Modern Agricultural Indus-trial Technology System Funding of Shandong ProvinceChina (SDAIT-02-022-04) the China agriculture researchsystem (CARS-25) and the National Science Foundation ofChina (31401820)
References
[1] K Aaby SMazur A Nes andG Skrede ldquoPhenolic compoundsin strawberry (Fragaria x ananassa Duch) fruits compositionin 27 cultivars and changes during ripeningrdquo Food Chemistryvol 132 no 1 pp 86ndash97 2012
[2] T A Colquhoun L A Levin H RMoskowitz VMWhitakerD G Clark and K M Folta ldquoFraming the perfect strawberryan exercise in consumer-assisted selection of fruit cropsrdquoJournal of Berry Research vol 2 no 1 pp 45ndash61 2012
[3] F Giampieri J M Alvarez-Suarez L Mazzoni et al ldquoThepotential impact of strawberry on human healthrdquo NaturalProduct Research vol 27 no 4 pp 448ndash455 2013
[4] Y S Velioglu GMazza L Gao and B D Oomah ldquoAntioxidantactivity and total phenolics in selected fruits vegetables and
International Journal of Genomics 13
grain productsrdquo Journal of Agricultural amp Food Chemistry vol46 no 10 pp 4113ndash4117 1998
[5] S S Hassellund A Flaa S E Kjeldsen et al ldquoEffects of antho-cyanins on cardiovascular risk factors and inflammation inpre-hypertensive men a double-blind randomized placebo-controlled crossover studyrdquo Journal of Human Hypertensionvol 27 no 2 pp 100ndash106 2013
[6] K J Meyers C B Watkins M P Pritts and R H Liu ldquoAnti-oxidant and antiproliferative activities of strawberriesrdquo Journalof Agricultural amp Food Chemistry vol 51 no 23 pp 6887ndash68922003
[7] S Zafra-Stone T YasminM Bagchi AChatterjee J AVinsonand D Bagchi ldquoBerry anthocyanins as novel antioxidants inhuman health and disease preventionrdquo Molecular Nutrition ampFood Research vol 51 no 6 pp 675ndash683 2007
[8] L Jaakola ldquoNew insights into the regulation of anthocyanin bio-synthesis in fruitsrdquoTrends in Plant Science vol 18 no 9 pp 477ndash483 2013
[9] K Springob J-I Nakajima M Yamazaki and K Saito ldquoRecentadvances in the biosynthesis and accumulation of antho-cyaninsrdquo Natural Product Reports vol 20 no 3 pp 288ndash3032003
[10] A Gonzalez M Zhao J M Leavitt and A M Lloyd ldquoRegu-lation of the anthocyanin biosynthetic pathway by the TTG1bHLHMyb transcriptional complex in Arabidopsis seedlingsrdquoPlant Journal vol 53 no 5 pp 814ndash827 2008
[11] K Petroni and C Tonelli ldquoRecent advances on the regulationof anthocyanin synthesis in reproductive organsrdquo Plant Sciencevol 181 no 3 pp 219ndash229 2011
[12] H-F Jia Y-M Chai C-L Li et al ldquoAbscisic acid plays animportant role in the regulation of strawberry fruit ripeningrdquoPlant Physiology vol 157 no 1 pp 188ndash199 2011
[13] C Merchante J G Vallarino S Osorio et al ldquoEthylene isinvolved in strawberry fruit ripening in an organ-specific man-nerrdquo Journal of Experimental Botany vol 64 no 14 pp 4421ndash4439 2013
[14] A Maier A Schrader L Kokkelink et al ldquoLight and the E3ubiquitin ligase COP1SPA control the protein stability of theMYB transcription factors PAP1 and PAP2 involved in antho-cyanin accumulation in Arabidopsisrdquo Plant Journal vol 74 no4 pp 638ndash651 2013
[15] H Lange W Shropshire and H Mohr ldquoAn analysis of phyto-chrome-mediated anthocyanin synthesisrdquo Plant Physiology vol47 no 5 pp 649ndash655 1971
[16] T D Elich and J C Lagarias ldquoPhytochrome chromophore bio-synthesis both 5-aminolevulinic acid and biliverdin overcomeinhibition by gabaculine in etiolated Avena sativa L seedlingsrdquoPlant Physiology vol 84 no 2 pp 304ndash310 1987
[17] Y Han W Xuan T Yu et al ldquoExogenous hematin alleviatesmercury-induced oxidative damage in the roots of Medicagosativardquo Journal of Integrative Plant Biology vol 49 no 12 pp1703ndash1713 2007
[18] M Koornneef J Cone R Dekens E OrsquoHerne-Robers CSpruit and R Kendrick ldquoPhotomorphogenic responses of longhypocotyl mutants of tomatordquo Journal of Plant Physiology vol120 no 2 pp 153ndash165 1985
[19] M J Terry and R E Kendrick ldquoFeedback inhibition of chloro-phyll synthesis in the phytochrome chromophore-deficientaurea and yellow-green-2 mutants of tomatordquo Plant Physiologyvol 119 no 1 pp 143ndash152 1999
[20] B M Parks and P H Quail ldquoPhytochrome-deficient hy1 andhy2 long hypocotyl mutants of Arabidopsis are defective inphytochrome chromophore biosynthesisrdquo Plant Cell vol 3 no11 pp 1177ndash1186 1991
[21] M J Terry ldquoPhytochrome chromophore-deficient mutantsrdquoPlant Cell amp Environment vol 20 no 6 pp 740ndash745 1997
[22] T Muramoto T Kohchi A Yokota I Hwang and H MGoodman ldquoThe Arabidopsis photomorphogenic mutant hy1 isdeficient in phytochrome chromophore biosynthesis as a resultof a mutation in a plastid heme oxygenaserdquo Plant Cell vol 11no 3 pp 335ndash347 1999
[23] M D Maines ldquoThe heme oxygenase system a regulator ofsecond messenger gasesrdquo Annual Review of Pharmacology andToxicology vol 37 no 1 pp 517ndash554 1997
[24] W Xuan F-Y Zhu S Xu et al ldquoThe heme oxygenasecarbonmonoxide system is involved in the auxin-induced cucumberadventitious rooting processrdquo Plant Physiology vol 148 no 2pp 881ndash893 2008
[25] R Amooaghaie F Tabatabaei and A-M Ahadi ldquoRole ofhematin and sodium nitroprusside in regulating Brassica nigraseed germination under nanosilver and silver nitrate stressesrdquoEcotoxicology and Environmental Safety vol 113 pp 259ndash2702015
[26] Y Liu X Li L Xu and W Shen ldquoDe-etiolation of wheatseedling leaves cross talk between heme oxygenasecarbonmonoxide and nitric oxiderdquo PLoS ONE vol 8 no 12 ArticleID e81470 2013
[27] T Muramoto N Tsurui M J Terry A Yokota and T KohchildquoExpression and biochemical properties of a ferredoxin-dependent heme oxygenase required for phytochrome chro-mophore synthesisrdquo Plant Physiology vol 130 no 4 pp 1958ndash1966 2002
[28] J-W Gao J Liu B Li and Z Li ldquoIsolation and purificationof functional total RNA from blue-grained wheat endospermtissues containing high levels of starches and flavonoidsrdquo PlantMolecular Biology Reporter vol 19 no 2 pp 185ndash186 2001
[29] R Patro S M Mount and C Kingsford ldquoSailfish enablesalignment-free isoform quantification from RNA-seq readsusing lightweight algorithmsrdquoNature Biotechnology vol 32 no5 pp 462ndash464 2014
[30] Y-X Qi Y-B Liu and W-H Rong ldquoRNA-Seq and its applica-tions a new technology for transcriptomicsrdquo Hereditas vol 33no 11 pp 1191ndash1202 2011
[31] C R Genovese N A Lazar and T Nichols ldquoThresholdingof statistical maps in functional neuroimaging using the falsediscovery raterdquo NeuroImage vol 15 no 4 pp 870ndash878 2002
[32] J Ye L Fang H Zheng et al ldquoWEGO a web tool for plottingGO annotationsrdquoNucleic Acids Research vol 34 supplement 2pp W293ndashW297 2006
[33] H Abdi ldquoThe Bonferonni and Sidak corrections for multiplecomparisonrdquo Encyclopedia of Measurement and Statistics vol 3pp 103ndash107 2007
[34] J Li Q Ding F Wang Y Zhang H Li and J Gao ldquoIntegrativeanalysis of mRNA and miRNA expression profiles of thetuberous root development at seedling stages in turnipsrdquo PLoSONE vol 10 no 9 Article ID e0137983 2015
[35] J Bennett D Hondred and J C Register ldquoKeeping qRT-PCRrigorous and biologically relevantrdquo Plant Cell Reports vol 34no 1 pp 1ndash3 2015
[36] F Wang L Li H Li et al ldquoTranscriptome analysis of rosetteand folding leaves in Chinese cabbage using high-throughputRNA sequencingrdquo Genomics vol 99 no 5 pp 299ndash307 2012
14 International Journal of Genomics
[37] T Shi Z Gao L Wang et al ldquoIdentification of differentially-expressed genes associated with pistil abortion in Japaneseapricot by genome-wide transcriptional analysisrdquo PLoS ONEvol 7 no 10 Article ID e47810 2012
[38] F Wang H Li Y Zhang et al ldquoMicroRNA expression analysisof rosette and folding leaves in Chinese cabbage using high-throughput Solexa sequencingrdquo Gene vol 532 no 2 pp 222ndash229 2013
[39] X Gu Y Chen Z Gao Y Qiao and X Wang ldquoTranscriptionfactors and anthocyanin genes related to low-temperature toler-ance in rd29A RdreB1BI transgenic strawberryrdquo Plant Physiol-ogy and Biochemistry vol 89 pp 31ndash43 2015
[40] J G Schaart C Dubos I R De La Fuente et al ldquoIdentificationand characterization of MYB-bHLH-WD40 regulatory com-plexes controlling proanthocyanidin biosynthesis in strawberry(Fragaria times ananassa) fruitsrdquoNew Phytologist vol 197 no 2 pp454ndash467 2013
[41] L Xie Z H Wang X H Cheng J J Gao Z P Zhangand L J Wang ldquo5-Aminolevulinic acid promotes anthocyaninaccumulation in Fuji applesrdquo Plant Growth Regulation vol 69no 3 pp 295ndash303 2013
[42] M Daminato F Guzzo and G Casadoro ldquoASHATTERPROOF-like gene controls ripening in non-climac-teric strawberries and auxin and abscisic acid antagonisticallyaffect its expressionrdquo Journal of Experimental Botany vol 64no 12 pp 3775ndash3786 2013
[43] X-H Ji R Zhang N Wang L Yang and X-S Chen ldquoTran-scriptome profiling reveals auxin suppressed anthocyanin bio-synthesis in red-fleshed apple callus (Malus sieversii f niedzwet-zkyana)rdquo Plant Cell Tissue and Organ Culture vol 123 no 2pp 389ndash404 2015
[44] J Chen LMaoW Lu T Ying and Z Luo ldquoTranscriptome pro-filing of postharvest strawberry fruit in response to exogenousauxin and abscisic acidrdquo Planta vol 243 no 1 pp 183ndash197 2016
[45] G A Martınez A R Chaves and M C Anon ldquoEffect of gib-berellic acid on ripening of strawberry fruits (Fragaria annan-assa Duch)rdquo Journal of Plant Growth Regulation vol 13 no 2pp 87ndash91 1994
[46] P K Das D H Shin S-B Choi S-D Yoo G Choi and Y-IPark ldquoCytokinins enhance sugar-induced anthocyanin biosyn-thesis in Arabidopsisrdquo Molecules amp Cells vol 34 no 1 pp 93ndash101 2012
[47] P McAtee S Karim R Schaffer and K David ldquoA dynamicinterplay between phytohormones is required for fruit develop-ment maturation and ripeningrdquo Frontiers in Plant Science vol4 no 3 article 79 7 pages 2013
[48] E Nambara andAMarion-Poll ldquoAbscisic acid biosynthesis andcatabolismrdquoAnnual Review of Plant Biology vol 56 pp 165ndash1852005
[49] Y Jiang and D C Joyce ldquoABA effects on ethylene productionPAL activity anthocyanin and phenolic contents of strawberryfruitrdquo Plant Growth Regulation vol 39 no 2 pp 171ndash174 2003
[50] S Li ldquoTranscriptional control of flavonoid biosynthesis fine-tuning of the MYB-bHLH-WD40 (MBW) complexrdquo PlantSignaling amp Behavior vol 9 Article ID e27522 2014
[51] D R Rudell J K Fellman and J P Mattheis ldquoPreharvest appli-cation of methyl jasmonate to rsquoFujirsquo apples enhances red col-oration and affects fruit size splitting and bitter pit incidencerdquoHortScience vol 40 no 6 pp 1760ndash1762 2005
[52] A Belhadj N Telef C Saigne et al ldquoEffect of methyl jasmonatein combination with carbohydrates on gene expression of PR
proteins stilbene and anthocyanin accumulation in grapevinecell culturesrdquo Plant Physiology and Biochemistry vol 46 no 4pp 493ndash499 2008
[53] E Loreti G Povero G Novi C Solfanelli A Alpi and PPerata ldquoGibberellins jasmonate and abscisic acid modulate thesucrose-induced expression of anthocyanin biosynthetic genesin Arabidopsisrdquo New Phytologist vol 179 no 4 pp 1004ndash10162008
[54] H S Seo J T Song J-J Cheong et al ldquoJasmonic acid carboxylmethyltransferase A key enzyme for jasmonate-regulated plantresponsesrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 98 no 8 pp 4788ndash4793 2001
[55] M T McDowell and J C Lagarias ldquoPurification and biochemi-cal properties of phytochromobilin synthase from etiolated oatseedlingsrdquo Plant Physiology vol 126 no 4 pp 1546ndash1554 2001
[56] G-Q Fu S Xu Y-J Xie et al ldquoMolecular cloning characteri-zation and expression of an alfalfa (Medicago sativa L) hemeoxygenase-1 gene MsHO1 which is pro-oxidants-regulatedrdquoPlant Physiology and Biochemistry vol 49 no 7 pp 792ndash7992011
[57] X-Y Chen X Ding S Xu et al ldquoEndogenous hydrogen per-oxide plays a positive role in the upregulation of heme oxy-genase and acclimation to oxidative stress in wheat seedlingleavesrdquo Journal of Integrative Plant Biology vol 51 no 10 pp951ndash960 2009
[58] J Shin E Park and G Choi ldquoPIF3 regulates anthocyaninbiosynthesis in an HY5-dependent manner with both factorsdirectly binding anthocyanin biosynthetic gene promoters inArabidopsisrdquo Plant Journal vol 49 no 6 pp 981ndash994 2007
[59] S Wang D Pan X Lv et al ldquoProteomic approach reveals thatstarch degradation contributes to anthocyanin accumulation intuberous root of purple sweet potatordquo Journal of Proteomics vol143 pp 298ndash305 2016
[60] H-F Jia C-L Li Y-M Chai Y Xing and Y Shen ldquoSucrosepromotes strawberry fruit ripening by stimulation of abscisicacid biosynthesisrdquo Pakistan Journal of Botany vol 45 no 1 pp169ndash176 2013
[61] W Xu H Peng T Yang et al ldquoEffect of calcium on strawberryfruit flavonoid pathway gene expression and anthocyanin accu-mulationrdquo Plant Physiology amp Biochemistry vol 82 pp 289ndash298 2014
[62] Z-W-SWan and Z Singh ldquoExogenous application of prohexa-dione-calcium promotes fruit colour development of lsquoCrippsPinkrsquo applerdquo Acta Horticulturae vol 1012 pp 219ndash225 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Genomics 13
grain productsrdquo Journal of Agricultural amp Food Chemistry vol46 no 10 pp 4113ndash4117 1998
[5] S S Hassellund A Flaa S E Kjeldsen et al ldquoEffects of antho-cyanins on cardiovascular risk factors and inflammation inpre-hypertensive men a double-blind randomized placebo-controlled crossover studyrdquo Journal of Human Hypertensionvol 27 no 2 pp 100ndash106 2013
[6] K J Meyers C B Watkins M P Pritts and R H Liu ldquoAnti-oxidant and antiproliferative activities of strawberriesrdquo Journalof Agricultural amp Food Chemistry vol 51 no 23 pp 6887ndash68922003
[7] S Zafra-Stone T YasminM Bagchi AChatterjee J AVinsonand D Bagchi ldquoBerry anthocyanins as novel antioxidants inhuman health and disease preventionrdquo Molecular Nutrition ampFood Research vol 51 no 6 pp 675ndash683 2007
[8] L Jaakola ldquoNew insights into the regulation of anthocyanin bio-synthesis in fruitsrdquoTrends in Plant Science vol 18 no 9 pp 477ndash483 2013
[9] K Springob J-I Nakajima M Yamazaki and K Saito ldquoRecentadvances in the biosynthesis and accumulation of antho-cyaninsrdquo Natural Product Reports vol 20 no 3 pp 288ndash3032003
[10] A Gonzalez M Zhao J M Leavitt and A M Lloyd ldquoRegu-lation of the anthocyanin biosynthetic pathway by the TTG1bHLHMyb transcriptional complex in Arabidopsis seedlingsrdquoPlant Journal vol 53 no 5 pp 814ndash827 2008
[11] K Petroni and C Tonelli ldquoRecent advances on the regulationof anthocyanin synthesis in reproductive organsrdquo Plant Sciencevol 181 no 3 pp 219ndash229 2011
[12] H-F Jia Y-M Chai C-L Li et al ldquoAbscisic acid plays animportant role in the regulation of strawberry fruit ripeningrdquoPlant Physiology vol 157 no 1 pp 188ndash199 2011
[13] C Merchante J G Vallarino S Osorio et al ldquoEthylene isinvolved in strawberry fruit ripening in an organ-specific man-nerrdquo Journal of Experimental Botany vol 64 no 14 pp 4421ndash4439 2013
[14] A Maier A Schrader L Kokkelink et al ldquoLight and the E3ubiquitin ligase COP1SPA control the protein stability of theMYB transcription factors PAP1 and PAP2 involved in antho-cyanin accumulation in Arabidopsisrdquo Plant Journal vol 74 no4 pp 638ndash651 2013
[15] H Lange W Shropshire and H Mohr ldquoAn analysis of phyto-chrome-mediated anthocyanin synthesisrdquo Plant Physiology vol47 no 5 pp 649ndash655 1971
[16] T D Elich and J C Lagarias ldquoPhytochrome chromophore bio-synthesis both 5-aminolevulinic acid and biliverdin overcomeinhibition by gabaculine in etiolated Avena sativa L seedlingsrdquoPlant Physiology vol 84 no 2 pp 304ndash310 1987
[17] Y Han W Xuan T Yu et al ldquoExogenous hematin alleviatesmercury-induced oxidative damage in the roots of Medicagosativardquo Journal of Integrative Plant Biology vol 49 no 12 pp1703ndash1713 2007
[18] M Koornneef J Cone R Dekens E OrsquoHerne-Robers CSpruit and R Kendrick ldquoPhotomorphogenic responses of longhypocotyl mutants of tomatordquo Journal of Plant Physiology vol120 no 2 pp 153ndash165 1985
[19] M J Terry and R E Kendrick ldquoFeedback inhibition of chloro-phyll synthesis in the phytochrome chromophore-deficientaurea and yellow-green-2 mutants of tomatordquo Plant Physiologyvol 119 no 1 pp 143ndash152 1999
[20] B M Parks and P H Quail ldquoPhytochrome-deficient hy1 andhy2 long hypocotyl mutants of Arabidopsis are defective inphytochrome chromophore biosynthesisrdquo Plant Cell vol 3 no11 pp 1177ndash1186 1991
[21] M J Terry ldquoPhytochrome chromophore-deficient mutantsrdquoPlant Cell amp Environment vol 20 no 6 pp 740ndash745 1997
[22] T Muramoto T Kohchi A Yokota I Hwang and H MGoodman ldquoThe Arabidopsis photomorphogenic mutant hy1 isdeficient in phytochrome chromophore biosynthesis as a resultof a mutation in a plastid heme oxygenaserdquo Plant Cell vol 11no 3 pp 335ndash347 1999
[23] M D Maines ldquoThe heme oxygenase system a regulator ofsecond messenger gasesrdquo Annual Review of Pharmacology andToxicology vol 37 no 1 pp 517ndash554 1997
[24] W Xuan F-Y Zhu S Xu et al ldquoThe heme oxygenasecarbonmonoxide system is involved in the auxin-induced cucumberadventitious rooting processrdquo Plant Physiology vol 148 no 2pp 881ndash893 2008
[25] R Amooaghaie F Tabatabaei and A-M Ahadi ldquoRole ofhematin and sodium nitroprusside in regulating Brassica nigraseed germination under nanosilver and silver nitrate stressesrdquoEcotoxicology and Environmental Safety vol 113 pp 259ndash2702015
[26] Y Liu X Li L Xu and W Shen ldquoDe-etiolation of wheatseedling leaves cross talk between heme oxygenasecarbonmonoxide and nitric oxiderdquo PLoS ONE vol 8 no 12 ArticleID e81470 2013
[27] T Muramoto N Tsurui M J Terry A Yokota and T KohchildquoExpression and biochemical properties of a ferredoxin-dependent heme oxygenase required for phytochrome chro-mophore synthesisrdquo Plant Physiology vol 130 no 4 pp 1958ndash1966 2002
[28] J-W Gao J Liu B Li and Z Li ldquoIsolation and purificationof functional total RNA from blue-grained wheat endospermtissues containing high levels of starches and flavonoidsrdquo PlantMolecular Biology Reporter vol 19 no 2 pp 185ndash186 2001
[29] R Patro S M Mount and C Kingsford ldquoSailfish enablesalignment-free isoform quantification from RNA-seq readsusing lightweight algorithmsrdquoNature Biotechnology vol 32 no5 pp 462ndash464 2014
[30] Y-X Qi Y-B Liu and W-H Rong ldquoRNA-Seq and its applica-tions a new technology for transcriptomicsrdquo Hereditas vol 33no 11 pp 1191ndash1202 2011
[31] C R Genovese N A Lazar and T Nichols ldquoThresholdingof statistical maps in functional neuroimaging using the falsediscovery raterdquo NeuroImage vol 15 no 4 pp 870ndash878 2002
[32] J Ye L Fang H Zheng et al ldquoWEGO a web tool for plottingGO annotationsrdquoNucleic Acids Research vol 34 supplement 2pp W293ndashW297 2006
[33] H Abdi ldquoThe Bonferonni and Sidak corrections for multiplecomparisonrdquo Encyclopedia of Measurement and Statistics vol 3pp 103ndash107 2007
[34] J Li Q Ding F Wang Y Zhang H Li and J Gao ldquoIntegrativeanalysis of mRNA and miRNA expression profiles of thetuberous root development at seedling stages in turnipsrdquo PLoSONE vol 10 no 9 Article ID e0137983 2015
[35] J Bennett D Hondred and J C Register ldquoKeeping qRT-PCRrigorous and biologically relevantrdquo Plant Cell Reports vol 34no 1 pp 1ndash3 2015
[36] F Wang L Li H Li et al ldquoTranscriptome analysis of rosetteand folding leaves in Chinese cabbage using high-throughputRNA sequencingrdquo Genomics vol 99 no 5 pp 299ndash307 2012
14 International Journal of Genomics
[37] T Shi Z Gao L Wang et al ldquoIdentification of differentially-expressed genes associated with pistil abortion in Japaneseapricot by genome-wide transcriptional analysisrdquo PLoS ONEvol 7 no 10 Article ID e47810 2012
[38] F Wang H Li Y Zhang et al ldquoMicroRNA expression analysisof rosette and folding leaves in Chinese cabbage using high-throughput Solexa sequencingrdquo Gene vol 532 no 2 pp 222ndash229 2013
[39] X Gu Y Chen Z Gao Y Qiao and X Wang ldquoTranscriptionfactors and anthocyanin genes related to low-temperature toler-ance in rd29A RdreB1BI transgenic strawberryrdquo Plant Physiol-ogy and Biochemistry vol 89 pp 31ndash43 2015
[40] J G Schaart C Dubos I R De La Fuente et al ldquoIdentificationand characterization of MYB-bHLH-WD40 regulatory com-plexes controlling proanthocyanidin biosynthesis in strawberry(Fragaria times ananassa) fruitsrdquoNew Phytologist vol 197 no 2 pp454ndash467 2013
[41] L Xie Z H Wang X H Cheng J J Gao Z P Zhangand L J Wang ldquo5-Aminolevulinic acid promotes anthocyaninaccumulation in Fuji applesrdquo Plant Growth Regulation vol 69no 3 pp 295ndash303 2013
[42] M Daminato F Guzzo and G Casadoro ldquoASHATTERPROOF-like gene controls ripening in non-climac-teric strawberries and auxin and abscisic acid antagonisticallyaffect its expressionrdquo Journal of Experimental Botany vol 64no 12 pp 3775ndash3786 2013
[43] X-H Ji R Zhang N Wang L Yang and X-S Chen ldquoTran-scriptome profiling reveals auxin suppressed anthocyanin bio-synthesis in red-fleshed apple callus (Malus sieversii f niedzwet-zkyana)rdquo Plant Cell Tissue and Organ Culture vol 123 no 2pp 389ndash404 2015
[44] J Chen LMaoW Lu T Ying and Z Luo ldquoTranscriptome pro-filing of postharvest strawberry fruit in response to exogenousauxin and abscisic acidrdquo Planta vol 243 no 1 pp 183ndash197 2016
[45] G A Martınez A R Chaves and M C Anon ldquoEffect of gib-berellic acid on ripening of strawberry fruits (Fragaria annan-assa Duch)rdquo Journal of Plant Growth Regulation vol 13 no 2pp 87ndash91 1994
[46] P K Das D H Shin S-B Choi S-D Yoo G Choi and Y-IPark ldquoCytokinins enhance sugar-induced anthocyanin biosyn-thesis in Arabidopsisrdquo Molecules amp Cells vol 34 no 1 pp 93ndash101 2012
[47] P McAtee S Karim R Schaffer and K David ldquoA dynamicinterplay between phytohormones is required for fruit develop-ment maturation and ripeningrdquo Frontiers in Plant Science vol4 no 3 article 79 7 pages 2013
[48] E Nambara andAMarion-Poll ldquoAbscisic acid biosynthesis andcatabolismrdquoAnnual Review of Plant Biology vol 56 pp 165ndash1852005
[49] Y Jiang and D C Joyce ldquoABA effects on ethylene productionPAL activity anthocyanin and phenolic contents of strawberryfruitrdquo Plant Growth Regulation vol 39 no 2 pp 171ndash174 2003
[50] S Li ldquoTranscriptional control of flavonoid biosynthesis fine-tuning of the MYB-bHLH-WD40 (MBW) complexrdquo PlantSignaling amp Behavior vol 9 Article ID e27522 2014
[51] D R Rudell J K Fellman and J P Mattheis ldquoPreharvest appli-cation of methyl jasmonate to rsquoFujirsquo apples enhances red col-oration and affects fruit size splitting and bitter pit incidencerdquoHortScience vol 40 no 6 pp 1760ndash1762 2005
[52] A Belhadj N Telef C Saigne et al ldquoEffect of methyl jasmonatein combination with carbohydrates on gene expression of PR
proteins stilbene and anthocyanin accumulation in grapevinecell culturesrdquo Plant Physiology and Biochemistry vol 46 no 4pp 493ndash499 2008
[53] E Loreti G Povero G Novi C Solfanelli A Alpi and PPerata ldquoGibberellins jasmonate and abscisic acid modulate thesucrose-induced expression of anthocyanin biosynthetic genesin Arabidopsisrdquo New Phytologist vol 179 no 4 pp 1004ndash10162008
[54] H S Seo J T Song J-J Cheong et al ldquoJasmonic acid carboxylmethyltransferase A key enzyme for jasmonate-regulated plantresponsesrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 98 no 8 pp 4788ndash4793 2001
[55] M T McDowell and J C Lagarias ldquoPurification and biochemi-cal properties of phytochromobilin synthase from etiolated oatseedlingsrdquo Plant Physiology vol 126 no 4 pp 1546ndash1554 2001
[56] G-Q Fu S Xu Y-J Xie et al ldquoMolecular cloning characteri-zation and expression of an alfalfa (Medicago sativa L) hemeoxygenase-1 gene MsHO1 which is pro-oxidants-regulatedrdquoPlant Physiology and Biochemistry vol 49 no 7 pp 792ndash7992011
[57] X-Y Chen X Ding S Xu et al ldquoEndogenous hydrogen per-oxide plays a positive role in the upregulation of heme oxy-genase and acclimation to oxidative stress in wheat seedlingleavesrdquo Journal of Integrative Plant Biology vol 51 no 10 pp951ndash960 2009
[58] J Shin E Park and G Choi ldquoPIF3 regulates anthocyaninbiosynthesis in an HY5-dependent manner with both factorsdirectly binding anthocyanin biosynthetic gene promoters inArabidopsisrdquo Plant Journal vol 49 no 6 pp 981ndash994 2007
[59] S Wang D Pan X Lv et al ldquoProteomic approach reveals thatstarch degradation contributes to anthocyanin accumulation intuberous root of purple sweet potatordquo Journal of Proteomics vol143 pp 298ndash305 2016
[60] H-F Jia C-L Li Y-M Chai Y Xing and Y Shen ldquoSucrosepromotes strawberry fruit ripening by stimulation of abscisicacid biosynthesisrdquo Pakistan Journal of Botany vol 45 no 1 pp169ndash176 2013
[61] W Xu H Peng T Yang et al ldquoEffect of calcium on strawberryfruit flavonoid pathway gene expression and anthocyanin accu-mulationrdquo Plant Physiology amp Biochemistry vol 82 pp 289ndash298 2014
[62] Z-W-SWan and Z Singh ldquoExogenous application of prohexa-dione-calcium promotes fruit colour development of lsquoCrippsPinkrsquo applerdquo Acta Horticulturae vol 1012 pp 219ndash225 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

14 International Journal of Genomics
[37] T Shi Z Gao L Wang et al ldquoIdentification of differentially-expressed genes associated with pistil abortion in Japaneseapricot by genome-wide transcriptional analysisrdquo PLoS ONEvol 7 no 10 Article ID e47810 2012
[38] F Wang H Li Y Zhang et al ldquoMicroRNA expression analysisof rosette and folding leaves in Chinese cabbage using high-throughput Solexa sequencingrdquo Gene vol 532 no 2 pp 222ndash229 2013
[39] X Gu Y Chen Z Gao Y Qiao and X Wang ldquoTranscriptionfactors and anthocyanin genes related to low-temperature toler-ance in rd29A RdreB1BI transgenic strawberryrdquo Plant Physiol-ogy and Biochemistry vol 89 pp 31ndash43 2015
[40] J G Schaart C Dubos I R De La Fuente et al ldquoIdentificationand characterization of MYB-bHLH-WD40 regulatory com-plexes controlling proanthocyanidin biosynthesis in strawberry(Fragaria times ananassa) fruitsrdquoNew Phytologist vol 197 no 2 pp454ndash467 2013
[41] L Xie Z H Wang X H Cheng J J Gao Z P Zhangand L J Wang ldquo5-Aminolevulinic acid promotes anthocyaninaccumulation in Fuji applesrdquo Plant Growth Regulation vol 69no 3 pp 295ndash303 2013
[42] M Daminato F Guzzo and G Casadoro ldquoASHATTERPROOF-like gene controls ripening in non-climac-teric strawberries and auxin and abscisic acid antagonisticallyaffect its expressionrdquo Journal of Experimental Botany vol 64no 12 pp 3775ndash3786 2013
[43] X-H Ji R Zhang N Wang L Yang and X-S Chen ldquoTran-scriptome profiling reveals auxin suppressed anthocyanin bio-synthesis in red-fleshed apple callus (Malus sieversii f niedzwet-zkyana)rdquo Plant Cell Tissue and Organ Culture vol 123 no 2pp 389ndash404 2015
[44] J Chen LMaoW Lu T Ying and Z Luo ldquoTranscriptome pro-filing of postharvest strawberry fruit in response to exogenousauxin and abscisic acidrdquo Planta vol 243 no 1 pp 183ndash197 2016
[45] G A Martınez A R Chaves and M C Anon ldquoEffect of gib-berellic acid on ripening of strawberry fruits (Fragaria annan-assa Duch)rdquo Journal of Plant Growth Regulation vol 13 no 2pp 87ndash91 1994
[46] P K Das D H Shin S-B Choi S-D Yoo G Choi and Y-IPark ldquoCytokinins enhance sugar-induced anthocyanin biosyn-thesis in Arabidopsisrdquo Molecules amp Cells vol 34 no 1 pp 93ndash101 2012
[47] P McAtee S Karim R Schaffer and K David ldquoA dynamicinterplay between phytohormones is required for fruit develop-ment maturation and ripeningrdquo Frontiers in Plant Science vol4 no 3 article 79 7 pages 2013
[48] E Nambara andAMarion-Poll ldquoAbscisic acid biosynthesis andcatabolismrdquoAnnual Review of Plant Biology vol 56 pp 165ndash1852005
[49] Y Jiang and D C Joyce ldquoABA effects on ethylene productionPAL activity anthocyanin and phenolic contents of strawberryfruitrdquo Plant Growth Regulation vol 39 no 2 pp 171ndash174 2003
[50] S Li ldquoTranscriptional control of flavonoid biosynthesis fine-tuning of the MYB-bHLH-WD40 (MBW) complexrdquo PlantSignaling amp Behavior vol 9 Article ID e27522 2014
[51] D R Rudell J K Fellman and J P Mattheis ldquoPreharvest appli-cation of methyl jasmonate to rsquoFujirsquo apples enhances red col-oration and affects fruit size splitting and bitter pit incidencerdquoHortScience vol 40 no 6 pp 1760ndash1762 2005
[52] A Belhadj N Telef C Saigne et al ldquoEffect of methyl jasmonatein combination with carbohydrates on gene expression of PR
proteins stilbene and anthocyanin accumulation in grapevinecell culturesrdquo Plant Physiology and Biochemistry vol 46 no 4pp 493ndash499 2008
[53] E Loreti G Povero G Novi C Solfanelli A Alpi and PPerata ldquoGibberellins jasmonate and abscisic acid modulate thesucrose-induced expression of anthocyanin biosynthetic genesin Arabidopsisrdquo New Phytologist vol 179 no 4 pp 1004ndash10162008
[54] H S Seo J T Song J-J Cheong et al ldquoJasmonic acid carboxylmethyltransferase A key enzyme for jasmonate-regulated plantresponsesrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 98 no 8 pp 4788ndash4793 2001
[55] M T McDowell and J C Lagarias ldquoPurification and biochemi-cal properties of phytochromobilin synthase from etiolated oatseedlingsrdquo Plant Physiology vol 126 no 4 pp 1546ndash1554 2001
[56] G-Q Fu S Xu Y-J Xie et al ldquoMolecular cloning characteri-zation and expression of an alfalfa (Medicago sativa L) hemeoxygenase-1 gene MsHO1 which is pro-oxidants-regulatedrdquoPlant Physiology and Biochemistry vol 49 no 7 pp 792ndash7992011
[57] X-Y Chen X Ding S Xu et al ldquoEndogenous hydrogen per-oxide plays a positive role in the upregulation of heme oxy-genase and acclimation to oxidative stress in wheat seedlingleavesrdquo Journal of Integrative Plant Biology vol 51 no 10 pp951ndash960 2009
[58] J Shin E Park and G Choi ldquoPIF3 regulates anthocyaninbiosynthesis in an HY5-dependent manner with both factorsdirectly binding anthocyanin biosynthetic gene promoters inArabidopsisrdquo Plant Journal vol 49 no 6 pp 981ndash994 2007
[59] S Wang D Pan X Lv et al ldquoProteomic approach reveals thatstarch degradation contributes to anthocyanin accumulation intuberous root of purple sweet potatordquo Journal of Proteomics vol143 pp 298ndash305 2016
[60] H-F Jia C-L Li Y-M Chai Y Xing and Y Shen ldquoSucrosepromotes strawberry fruit ripening by stimulation of abscisicacid biosynthesisrdquo Pakistan Journal of Botany vol 45 no 1 pp169ndash176 2013
[61] W Xu H Peng T Yang et al ldquoEffect of calcium on strawberryfruit flavonoid pathway gene expression and anthocyanin accu-mulationrdquo Plant Physiology amp Biochemistry vol 82 pp 289ndash298 2014
[62] Z-W-SWan and Z Singh ldquoExogenous application of prohexa-dione-calcium promotes fruit colour development of lsquoCrippsPinkrsquo applerdquo Acta Horticulturae vol 1012 pp 219ndash225 2012
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology


















