
1 CS 501 Spring 2006 CS 501: Software Engineering Lectures 11 & 12 Usability.
description

Reinagel lectures 2006
Take home message about LGN1. Lateral geniculate nucleus transmits information from retina to cortex2. It is not known what computation if any occurs in the LGN3. For white noise stimuli, responses are precise and reliable4. PRECISION is the trial to trial jitter in spike TIMING (order 1msec)
feed forward inhibition may be the mechanism of precise timing5. RELIABILITY is the trial to trial variability in spike NUMBER (subpoisson)
refractoriness may be the mechanism of reliable spike count6. BURSTING in the LGN is a distinct biophysical phenomenon, of unknown
importance. The *right* question to ask is whether the burstingstate is visually primed and whether priming itself encodes information
7. We now have a visually behaving rodent prep to address all these questions
Take home message about efficient coding1. Natural scenes are full of spatial and temporal correlations2. This suggests WHY center-surround RF's are GOOD: redundancy reduction2. Test: LGN responses to natural scenes are decorrelated (whitened)3. More generally: are natural scenes optimal stimuli?
is this even the right question?
www-biology.ucsd.edu/labs/reinagel/

LGN
Retina
Cortex
Ramon y Cajal
Hubel 1960 (alert cat)Hubel & Wiesel 1961 (anesthetized)
Lateral Geniculate Nucleus

• spiking inputs• intrinsic properties• local circuits• cortical feedback
Gating? Attention? Binding? Prediction testing? Nothing?
What happens in the LGN?
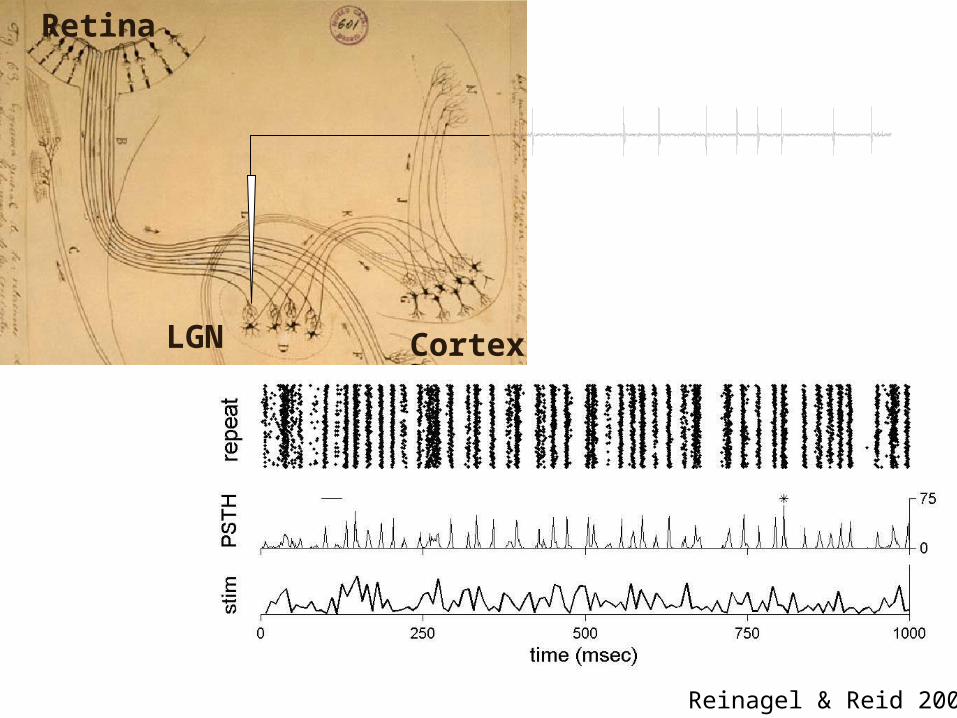
LGN
Retina
Cortex
Reinagel & Reid 2000

LGN response to purely temporal stimuli
Lum
inance
Repeat
Reinagel & Reid, 2000
Descriptive questions:• how precise is the timing?• how reliable is the number?• are there internal patterns?
In each case:• visual information?• mechanism of encoding?• mechanism of decoding?


PSTH peaks are milliseconds wide
Reinagel & Reid, 2002

1500 1600 1700 1800 1900 20000
0.5
A
0.2 0.1 -0.1 0.6 0.9 0.20.2
-0.90.3
-0.3-0.1
PS
TH
Norm'dt
Time (ms)
Reinagel & Reid 2002
Temporal patterns conserved across animals

Precision of spike times used (ms)
0.5 1 2 4 8 16 32 64 1280
50
100
Mu
tual In
form
ati
on
(b
its/
s)a b c de
Temporal precision of visual information
Theory of Shannon, 1948Method of Strong et al., 1998Result of Reinagel & Reid, 2000

Mechanisms Underlying Precise Timing
Pouille & Scanzian 2001


Mean 4Variance 0
Mean 4Variance 4
Deterministic Poisson

Spike Count: Trial to Trial Variability
Variance in Spike #Mean Spike #
Random Deterministic(Poisson)
= 1 = 0
Measure of variability

PSTH
LGN vs. Poisson Model
PSTH

0 100 500 10000
0.25
0.5
0.75
1
bin size T (msec)
Fano F
act
or
LGN Variability << Poisson
LGN
Poisson

Variability increases from retina to cortexF
ano
Fa
cto
r a
t ~ 4
0 H
z
0
1
RGC LGN V1
Kara, Reinagel & Reid, 2000

FF
FiringRate
200
0
0
1
RGC LGN V1
0 500
Time (ms)
When firing rate is high, variability is low
Kara, Reinagel & Reid, 2000

Refractoriness Regularizes?
PSTH
Poisson model
Poisson withRefractory Period

0 10 20 30 40 50 60 70 80 90 100
prob
abili
ty
ISI
data
Estimating refractoriness from data
Method:Berry & Meister1998
model

0 5 10 15 20 25 30 350
0.5
1
time since last spike (ms)
reco
very
fun
ctio
nRecovery Function
absolute andrelativerefractoriness

0 500 10000
100
200
300
400
500
time (ms)
firin
g ra
te (
sp/s
)
observed
free
Free Firing Rate

Refractory models for all cell types
0
1
2
Fan
o F
acto
r
0 200 0
Time (ms)
V1LGNRGC
Kara, Reinagel & Reid, 2000
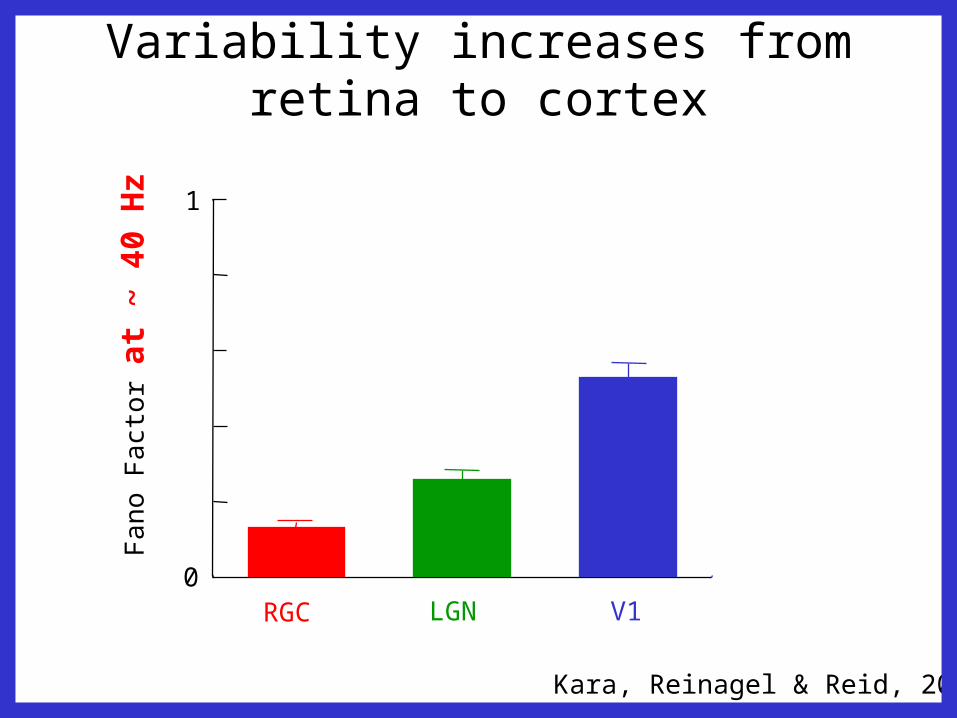
Variability increases from retina to cortexF
ano
Fa
cto
r at
~ 4
0 H
z
0
1
RGC LGN V1
Kara, Reinagel & Reid, 2000

Refractoriness decreasesfrom retina to cortex
Rec
over
y F
unct
ion
0 10 20 30
Time (ms)
0.0
1.0
0.5
V1
RGC
Kara, Reinagel & Reid, 2000

Summary of Reliability
• Spike count has sub-Poisson variability
• High FR High Reliability
• Refractoriness completely explains
• Noise is low, but doubling each synapse
- firing rate is decreasing
- refractoriness is decreasing


Thalamic Bursts (It)
Jahnsen and Llinas (1984)
Hubel and Wiesel (1961)

Bursting in the LGN
• dominate during sleep, when vision is suppressed
• frequent under anesthesia, when vision is absent
• almost never seen in alert animals, when vision is happening
ERGO
Bursts are irrelevant to vision
• not rhythmic or synchronous in anesthetized animals
• visual in anesthetized animals
• synapses prefer bursts
• do occur in alert animals, and rare signals can be important
• cool computational ideas
ERGO
Bursts are crucial to vision

-0.6 -0.4 -0.2 0
-0.1
0
0.1
0.2
Time before spike (s)
Before a burst
Before a tonic spike
Optimal Guess of Stimulus
Visual inputs trigger bursts
0
0.5
1
1.5
Bit
s/event
0
0.05
0.1
0.15
0.2
Codin
g E
ffici
ency
Burst BurstTonic Tonic
Reinagel, Godwin, Sherman & Koch 1999

Bursts: Triggering vs. Priming
time
active
inactiveLT-C
a+
+
channel
state
A
P
tim
es
Tri
gger
synapti
cin
put •• • ••• • •
*Ca
++
spik
e
observable

Bursts in LGN are distinct code words
Denning & Reinagel 2005
Alitto, Weyand & Usrey 2005 Lesica & Stanley 2004

Summary: Bursting• LGN neurons have 2 states
• Visual inputs trigger responses in both states
• Visual inputs also control the state
BUT All this is under anesthesia
What about alert?
- Stimulus ensemble matters
- Behavioral state may also
- Triggering and priming

• spiking inputs • intrinsic properties • local circuits • cortical feedback
What happens in the LGN?

Directions
• Do bursts occur and are they visual in alert animals?
• Function of cortical feedback to the LGN?
• Does precision in the LGN matter for perception?

Flister, Meier , Conway & Reinagel (unpub)
An awake behaving rodent prep for vision
Thanks to collaborators at CSHL

Bursts in LGN in the awake, behaving rat
Flister, Meier & Reinagel (unpub)

[break]

Center Surround Opponent RFs
Kuffler 1958

Natural scenes are spatially correlated

Spatial correlations in unnatural images

Natural Image
-40 -20 0 20 400.2
0.4
0.6
0.8
1
distance
Correlation
100
10210
-2
100
102
Power spectrum
cycles/degree
Spatial correlation in natural images
(cf. Field 1987; Tadmore & Tolhurst; Ruderman & Bialek; van Hateren)

Natural Image
-20 0 200
0.2
0.4
0.6
0.8
1
Distance (pixels)
Correlation
10-2
100
102
10-2
100
102
104
Power spectrum
Spatial frequency
-20 0 200
0.2
0.4
0.6
0.8
1
10-2
100
102
10-2
100
102
104
(cf. Barlow 1961)

0 1 2 3 410-2
10-1
100
time (s)
lum
ina
nce
-1 0 10
0.2
0.4
0.6
0.8
1
Distance (sec)
Correlation
10-1
100
101
102
103
10-6
10-4
10-2
100
102
Power Spectrum
Temporal frequency (Hz)
Natural temporal stimulus
(cf. Dong & Atick 1995; van Hateren 1997)

0 1 2 3 410
-2
10-1
100
time (s)
lum
ina
nce
-2 -1 0 1 20
0.2
0.4
0.6
0.8
1
distance (sec)
Correlation
10-1
100
101
103
Power Spectrum
10-5
100
102
Temporal frequency (Hz)
(cf. Dan Atick & Reid 1996)

+
Barlow 1961 Redundancy Reduction Hypothesis
+
Sensory neurons decorrelate natural inputs to reduce redundancy

Dan, Atick & Reid 1996
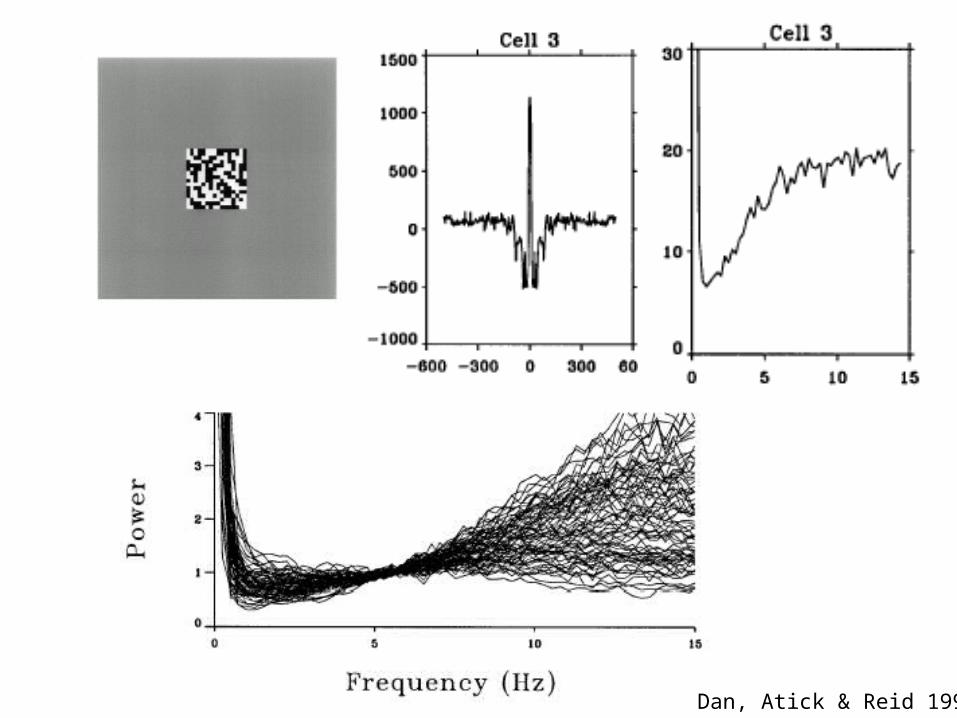
Dan, Atick & Reid 1996

Whitening in the fly
Van Hateren 1997

Summary: Redundancy Reduction
• Shannon 1948: Optimal codes lack redundancy
• Kuffler 1958: Center-surround receptive fields in retinaHubel 1960: Center-surround RFs in LGN
• Barlow 1961: Center-surround RFs reduce redundancy for natural scenes
• Dan, Atick & Reid 1996: Responses in LGN are less redundant for natural scenes

white natural0
50
100
150
200
250
300A
Info
rma
tion
(b
its/s
)
white natural0
1
2
3
4
5
6
7B
Info
rma
tion
(b
its/s
pk)
white natural0
0.2
0.4
0.6
0.8
1C
Eff
icie
ncy
(b
its/b
it)
Bullfrog Auditory Neuron: Natural Stimulus is ‘Optimal’
Rieke, Bodnar & Bialek 1995

Cat LGN Neuron: Opposite result
white natural0
20
40
60
80
Info
rma
tion
(bits
/s)
A
white natural0
0.5
1
1.5
2
2.5
3
Info
rma
tion
(bits
/spk
)
B
white natural0
0.1
0.2
0.3
0.4
0.5
0.6
Eff
icie
ncy
(bits
/bit)
C
Analog LED Stimuli
Reinagel & Reid, in prep.

10 0 101 102 103101
102
103
104
105
temporal frequency (Hz)
Po
we
rLGN Responses to full field temporal stimuli
Natural visual stimulus
Random visual stimulus
(replication ofDan et al 1996)


















