
Structure of Pisum sativumRubisco with bound ... - Loewen Labs
Upload
vuongnguyetCategory
view
214download
0
Plant Physiol. (1990) 93, 1579-15840032-0889/90/93/1 579/06/$01 .00/0
Received for publication October 25, 1989Accepted April 20, 1990
Regulation of Glutathione Synthesis by Cadmium inPisum sativum L.'
Adrian Ruegsegger, Daniel Schmutz, and Christian Brunold*Pflanzenphysiologisches Institut der Universitat Bern, Altenbergrain 21, 3013 Bern, Switzerland
ABSTRACT
In roots and shoots of pea plants (Pisum sativum L.) cultivatedwith CdCI2 concentrations up to 50 micromolar, growth, the con-tent of total acid soluble thiols, and the activity of glutathionesynthetase (EC 6.3.2.3) and of adenosine 5'-phosphosulfate sul-fotransferase were measured. In addition, the occurrence of Cd-binding peptides (phytochelatins) and the contents of glutathioneand cysteine were determined in roots of plants exposed to 20micromolar Cd and/or 1 millimolar buthionine sulfoximine, aninhibitor of glutathione synthesis. An appreciable increase inactivity of glutathione synthetase at 20 and 50 micromolar Cd andof adenosine 5'-phosphosulfate sulfotransferase at 5 micromolarand higher Cd concentrations was detected in the roots. Most ofthe additional thiols formed due to Cd treatment were eluted froma gel filtration HPLC column together with Cd, indicating thepresence of phytochelatins. In plants treated with buthioninesulfoximine and Cd, no phytochelatins could be detected but thecysteine content increased 21-fold. Additionally, a larger increasein both enzyme activities occurred than with Cd alone. Takentogether, our results are consistent with the hypothesis thatglutathione is a precursor for phytochelatin synthesis.
In plants, the general principle for complexing heavy metalions is the formation of peptides with the structure (y-glu-cys)n = 2-11), called phytochelatins (8, 10), poly(y-glutamyl-cysteinyl)glycines [(yEC)nG] (25), Cd-binding peptides (22),y-glutamyl metal-binding peptides (23), or class III metallo-thioneins (7). The capacity to produce such peptides due toCd-treatment seems to be widespread in the plant kingdom(8). Identical peptides, called cadystins, have been found inCd-treated fission yeast (14). Yeast mutants unable to synthe-size phytochelatins (17) and tomato (16, 29) and tobacco (21)cell cultures supplied with BSO2, a potent and specific inhib-itor of the enzyme catalyzing the first step of glutathionebiosynthesis, y-glutamylcysteine synthetase (EC 6.3.2.2), werehypersensitive towards Cd. This indicates that phytochelatinsplay a role in the detoxification of heavy metal ions (5, 8, 12).Because of the y-glutamic acid bonds in the peptide chains,phytochelatins cannot be primary gene products, and gluta-thione (y-glu-cys-gly), with its structure closely related to theone of phytochelatins, could be used as a precursor for the
Supported in part by the Swiss National Science Foundation andby the "Europaischer Forderpreis" of the Korber Foundation.
Abbreviations: BSO, buthionine sulfoximine; APS, adenosine5'-phosphosulfate; APSSTase, adenosine 5'-phosphosulfate sulfo-transferase.
synthesis of these compounds. Several findings are in accord-ance with this idea: Cd-treated Datura innoxia cells incorpo-rated 35S-radiolabel from glutathione into phytochelatinswithout any net loss of 35S from these two pools (1). In tomatocells supplied with both Cd and glutathione, more phytoche-latins were measured than in cells treated with Cd alone (16).Cd treatment caused lower glutathione contents (20, 26) andBSO suppressed the formation of phytochelatins (16, 21, 26,29). The capacity to synthesize phytochelatins was reestab-lished in BSO-treated tomato cells when glutathione wasadded to the culture medium (16, 26). Additionally, y-glutam-ylcysteine synthetase had a higher activity in selected cad-mium resistant tomato cells than in normal sensitive ones(28).These in vivo findings were very recently corroborated by
results from in vitro studies using extracts from Silene cucu-balus (10). They showed that phytochelatins were formedfrom glutathione in the presence of Cd2" by a -y-glutamyl-cysteine dipeptidyl transpeptidase.The extraordinarily high cysteine content of phytochelatins
makes an increased rate of assimilatory sulfate reductionprobable in plants exposed to Cd. We have tested this predic-tion in maize, and, indeed, both in vivo and in vitro sulfateassimilation were at higher levels due to Cd (19). Maize has amedium tolerance toward Cd (2), whereas pea plants areknown to be very sensitive (13), which could be due to aninsufficient capacity to enhance sulfate assimilation to thelevel required for effective phytochelatin synthesis. The pur-pose of the present study was to test this idea and to examinethe effect of Cd on glutathione synthesis and sulfate assimi-lation in pea plants.We therefore measured the activity of glutathione synthe-
tase, the enzyme catalyzing the second step of glutathionebiosynthesis, synthesizing glutathione from y-glutamyl-cys-teine and glycine. For comparison we determined the activityof APSSTase, which is considered to be the key enzyme ofassimilatory sulfate reduction (1 1, 19), catalyzing the transferof sulfate activated in APS to a carrier (car-SH), thus formingcar-S-SO32- (27).
MATERIALS AND METHODS
Plant Material and Treatment
Pea seeds (Pisum sativum L. cv Fruehbusch, Vatter, Bern,Switzerland) were soaked for 1 d in aerated water at roomtemperature. After subsequent germination on damp paperin the dark at room temperature for 3 d, the seedlings were
1579
www.plantphysiol.orgon August 16, 2019 - Published by Downloaded from Copyright © 1990 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 93,1990
transferred on pots containing 300 mL nutrient solution (19).There were 15 seedlings on each pot for the 3-d kineticexperiments, 10 seedlings for all other experiments (exceptlabeling with "5Cd). BSO (1 mM) was added to the corre-sponding cultures. The plants were kept in continuous light(50 ,uE m-2 s-', 1 TL 40 W/33 and 2 TL 40 W/32, Philips)at 21 to 24°C and 55 to 60% RH. Three days later, 0.5 to 50AM CdCl2 was added.
Preparation of Extracts for Enzyme Assays
Root systems were rinsed extensively with tap water andcut off above the top lateral roots. All lateral roots wereremoved. Shoots were cut off below the lowest leaf. Plantmaterial was ground in an ice-cooled glass homogenizer andthe homogenate was filtered through one layer of 100%viscose fleece (Milette, Migros, Switzerland). For theAPSSTase assay the extraction was with 0.1 M Tris-HCl (pH8.0), 0.1 M KCI, 20 mM MgCl2, and 10 mM DTE, using onepart of plant material and 10 parts (w/v) of buffer. For themeasurement of glutathione synthetase the extraction waswith 0.1 M Tris-HCl (pH 8.2), 20 mm MgCl2, 10 mm KCI,and 4 mM EDTA, using one part of plant material and fiveparts (w/v) of buffer.
Enzyme Assays
APSSTase activity was measured as the production of [35S]sulfite, assayed as acid volatile radioactivity formed fromAP35S (specific activity 16-27 GBq/mol) in the presence ofDTE (4). The incubation mixture contained in 0.1 M Tris-HCl (pH 9.0), 4 mM DTE, 0.8 M MgSO4, 75M AP35S, 0.2MNa2SO3, and 130 ,L plant extract (50-250 Ag protein) in atotal volume of 500 ,AL. The assays were incubated for 30min at 37°C, due to the addition of sulfite they had not to bekept under argon (4). At the concentration added, sulfite hadno effect on the enzyme activity. Routinely, 400 to 5000 cpmwere measured per assay.
Glutathione synthetase (EC 6.3.2.3) activity was measuredby the formation of ['4C]glutathione using y-glutamylcysteineand ['4Cjglycine. The assay was a modification of the onedescribed by Law and Halliwell (15). The extract was centri-fuged for 20 min at 40,000g and 4°C. The incubation mixturecontained in 0.1 M Tris-HCl (pH 8.2), 20 mM MgCl2, 10 mMKCI, 10 mm ['4C]glycine (8.3 GBq/mol), 5 units creatine-kinase, and 5 mM creatine phosphate as an ATP-regeneratingsystem, 5 mM y-glutamylcysteine, 10 mM DTE, 5 mm ATP,3 mM EDTA, and 140 L extract (60-350 ug protein) in atotal volume of 200 ,uL. The assays were incubated for 90min at 37°C in closed Eppendorf tubes. The reaction wasstopped by the addition of 40 ,L 50% acetic acid containing3 mg reduced glutathione. ['4C]Glutathione was separatedfrom ['4C]glycine by high voltage electrophoresis of 20 ALsamples of the stopped assays. The running buffer wasH20:acetic acid:pyridine 389:10:1 (v:v:v), the separation tookplace at 4500 V for 30 min in a water cooled system (Camag,Muttenz, Switzerland). The electrophoresis paper was dryedat 105°C for 15 min and the glutathione spots were madevisible by spraying the paper with ninhydrin reagent and
heating to 1050C. The radioactivity was counted in a liquidscintillation system using Emulsifier Scintillation Cocktail(Packard, Zurich, Switzerland) and a liquid scintillation spec-trometer (MR 300, Kontron, Zurich, Switzerland). Blankscontained crude extract boiled for 10 min and centrifuged for5 min at l0,OOOg at room temperature before incubation.
ATP-sulfurylase (EC 2.7.7.4) and nitrate reductase (EC1.7.7.1) were measured according to Nussbaum et al. (19).
HPLC Analysis of Cd-Binding Compounds
Roots were pretreated and homogenized by the same pro-cedure as for the APSSTase assay, but HPLC running buffersaturated with Ar was used for extraction. The extracts werepassed through a filter with a pore size of 0.2 Mm previouslyflushed with Ar. Cd-binding components were separated byHPLC gel filtration using an UltroPac TSK G 2000 SW 7.5x 300 mm column, particle size 10 ± 2 ,um, mean pore size12.5 nm (LKB Producter AB, Bromma, Sweden). The run-ning buffer contained 10 mm Tris-HCl (pH 7.0) and 50 mMNaCl, saturated with Ar. A volume of 100 ML of extract wasseparated with a flow rate of 800 ML per min. The detectionwas at 254 nm, since the absorbance of Cd-mercaptide (cys-teine) chromophores is characteristically high at this wave-length. Total thiols were measured in 430 uL fractions usingEllman's reagent (6); Cd was detected using extracts of 11 dold plants, cultivated for 4 d on 10 mL 20 Mm Cd2" containingnutrient solution (19), labeled with "5Cd (0.9-1.4 GBq/molCd). Fractions of 860 uL were collected and the radioactivitywas counted in the same liquid scintillation system used for14C counting.
Measurement of Total Acid Soluble Thiols
Plant material was homogenized as described for the APSS-Tase assay, but 0.1 mM EDTA: 10% (w/v) TCA 10:2 (v:v) wasused as extraction medium. The measurement of thiols wasthat of Grill and Esterbauer (9) using Ellman's reagent (6).
Determination of Cysteine and Glutathione Contents
Roots were homogenized as for the APSSTase assay, butthe extraction was with the medium used for the separationby HPLC. The extracts were centrifuged for 20 min at 30,000gand 4°C. Supematants (100 ML) were used for the separationof thiols by isocratic reverse phase HPLC with a C18 4.6 x250 mm column, particle size 5 Mm (Stagroma, Wallisellen,Switzerland). The running buffer contained 96% 0.1 M mon-ochlor acetic acid, pH adjusted to 3.0 (NaOH), 2 mm sodiumoctyl sulfate, and 4% methanol. The flow rate was 1.5 mL/min. Reduced glutathione and cysteine were quantified (24)using an Au/Hg electrode and an amperometric detector (LC-4B, BAS, West Lafayette, IN). Separation and detection tookplace in a system made oxygen-free by flushing with helium.
Protein Determination
The protein content was measured according to Bradford(3) with bovine serum albumin as a standard.
ROEGSEGGER ET AL.1 580
www.plantphysiol.orgon August 16, 2019 - Published by Downloaded from Copyright © 1990 American Society of Plant Biologists. All rights reserved.
REGULATION OF GLUTHATHIONE SYNTHESIS BY CADMIUM
100
80I100
0'
E
60
40
20
8Cw-
E
a
00
LO.
E
6C
4C
2C
0 1 2 3 4 5 6 7 8 9
days
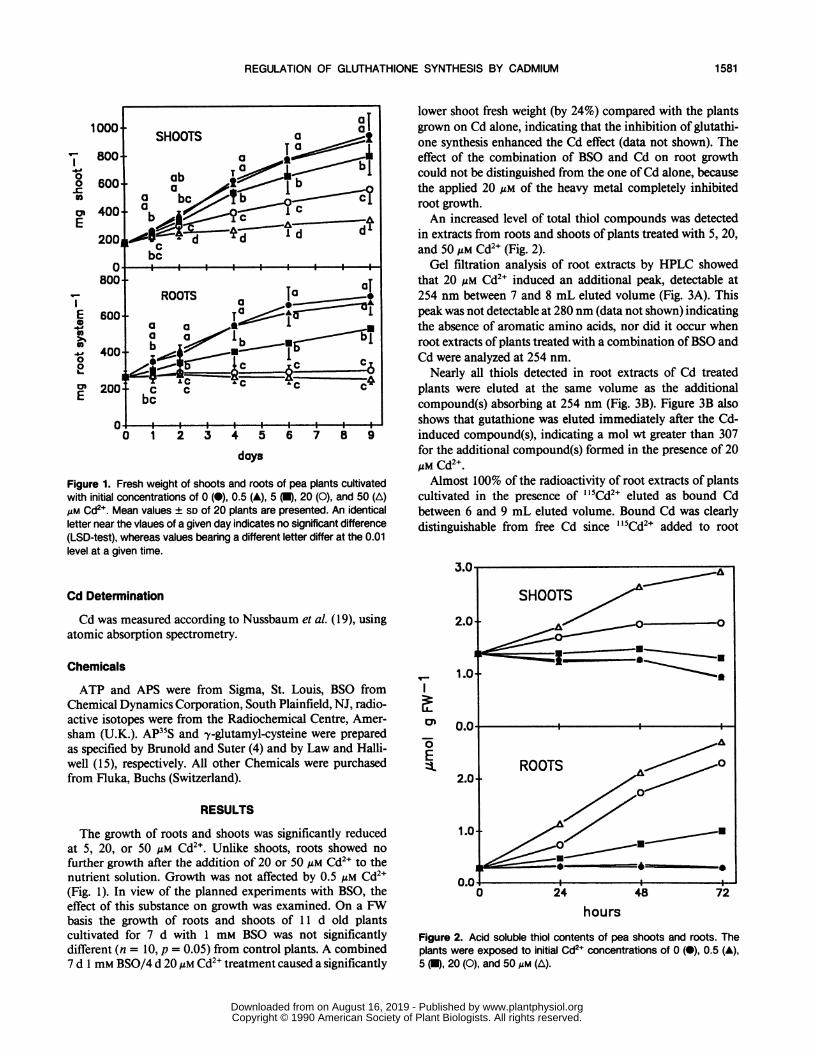
Figure 1. Fresh weight of shoots and roots of pea plants cultivatedwith initial concentrations of 0 (O), 0.5 (A), 5 (U), 20 (0), and 50 (A)AM Cd2 . Mean values ± SD of 20 plants are presented. An identicalletter near the vlaues of a given day indicates no significant difference(LSD-test), whereas values bearing a different letter differ at the 0.01level at a given time.
Cd Determination
Cd was measured according to Nussbaum et al. ( 19), usingatomic absorption spectrometry.
Chemicals
ATP and APS were from Sigma, St. Louis, BSO fromChemical Dynamics Corporation, South Plainfield, NJ, radio-active isotopes were from the Radiochemical Centre, Amer-sham (U.K.). AP35S and y-glutamyl-cysteine were preparedas specified by Brunold and Suter (4) and by Law and Halli-well (15), respectively. All other Chemicals were purchasedfrom Fluka, Buchs (Switzerland).
RESULTS
The growth of roots and shoots was significantly reducedat 5, 20, or 50 gM Cd2 . Unlike shoots, roots showed no
further growth after the addition of 20 or 50 uM Cd2+ to thenutrient solution. Growth was not affected by 0.5 gM Cd2+(Fig. 1). In view of the planned experiments with BSO, theeffect of this substance on growth was examined. On a FWbasis the growth of roots and shoots of 11 d old plantscultivated for 7 d with 1 mM BSO was not significantlydifferent (n = 10, p = 0.05) from control plants. A combined7 d 1 mm BSO/4 d 20uM Cd2" treatment caused a significantly
lower shoot fresh weight (by 24%) compared with the plantsgrown on Cd alone, indicating that the inhibition of glutathi-one synthesis enhanced the Cd effect (data not shown). Theeffect of the combination of BSO and Cd on root growthcould not be distinguished from the one of Cd alone, becausethe applied 20 Mm of the heavy metal completely inhibitedroot growth.An increased level of total thiol compounds was detected
in extracts from roots and shoots of plants treated with 5, 20,and 50 Mm Cd2+ (Fig. 2).Gel filtration analysis of root extracts by HPLC showed
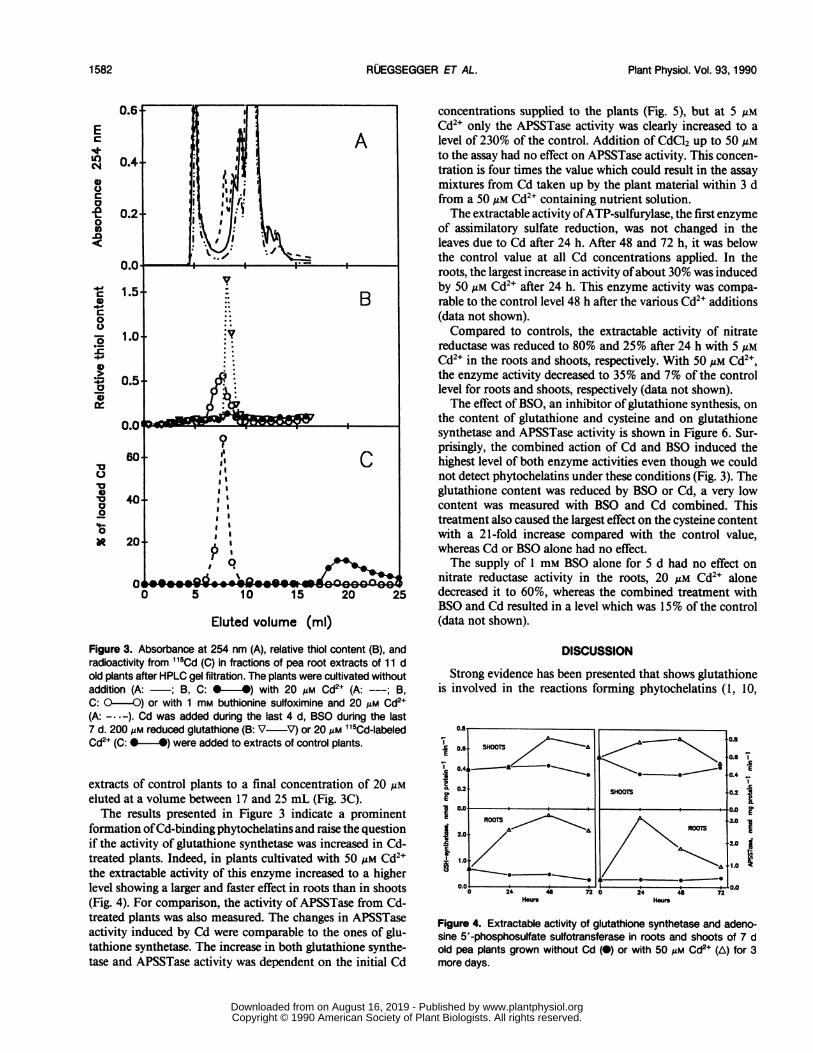
that 20 gM Cd2" induced an additional peak, detectable at254 nm between 7 and 8 mL eluted volume (Fig. 3A). Thispeak was not detectable at 280 nm (data not shown) indicatingthe absence of aromatic amino acids, nor did it occur whenroot extracts of plants treated with a combination of BSO andCd were analyzed at 254 nm.
Nearly all thiols detected in root extracts of Cd treatedplants were eluted at the same volume as the additionalcompound(s) absorbing at 254 nm (Fig. 3B). Figure 3B alsoshows that gutathione was eluted immediately after the Cd-induced compound(s), indicating a mol wt greater than 307for the additional compound(s) formed in the presence of 20Mm Cd2+.Almost 100% of the radioactivity of root extracts of plants
cultivated in the presence of "5Cd2+ eluted as bound Cdbetween 6 and 9 mL eluted volume. Bound Cd was clearlydistinguishable from free Cd since "5Cd2' added to root
v 0.0 I I
E2.01 ROOTS2.0-
00
1.0T
0 24 48 72
hours
Figure 2. Acid soluble thiol contents of pea shoots and roots. Theplants were exposed to initial Cd2+ concentrations of 0 (0), 0.5 (A),5 (U), 20 (0), and 50 AM (A).
SHOOTS aa
i~T
10a 0
JT
a
)0-.a
cc
bc
[1. a a I i a I
1581
0%
www.plantphysiol.orgon August 16, 2019 - Published by Downloaded from Copyright © 1990 American Society of Plant Biologists. All rights reserved.
Plant Physiol. Vol. 93,1990
ECA
E i0.4-l
o .10.20*I
.0
0.0-
%~~~~
N 20w ,
o 5 10 15 20 25
Eluted volume (ml)Figure 3. Absorbance at 254 nm (A), relative thiol content (B), andradioactivity from 115Cd (C) in fractions of pea root extracts of 1 1 dold plants after HPLC gel filtration. The plants were cultivated withoutaddition (A: ~; B, C: 0-*) with 20 Am Cd"+(A: ; B,C: O -O) or with 1 mm buthionine sulfoximine and 20 lim Cd2+(A: - ---). Cd was added during the last 4 d, BSO during the last7 d. 200OAm reduced glutathione (B: V ~V) or 20 Am 'l5Cd-labeledCd2+ (C: v *) were added to extracts of control plants.
extracts of control plants to a final concentration of 20 gmeluted at a volume between 17 and 25 mL (Fig. 3C).The results presented in Figure 3 indicate a prominent
formation ofCd-binding phytochelatins and raise the questionif the activity of glutathione synthetase was increased in Cd-treated plants. Indeed, in plants cultivated with 50 jAm Cd2+the extractable activity of this enzyme increased to a higherlevel showing a larger and faster effect in roots than in shoots(Fig. 4). For comparison, the activity of APSSTase from Cd-treated plants was also measured. The changes in APSSTaseactivity induced by Cd were comparable to the ones of glu-tathione synthetase. The increase in both glutathione synthe-tase and APSSTase activity was dependent on the initial Cd
concentrations supplied to the plants (Fig. 5), but at 5 uMCd2" only the APSSTase activity was clearly increased to alevel of 230% of the control. Addition of CdCl2 up to 50 Mmto the assay had no effect on APSSTase activity. This concen-tration is four times the value which could result in the assaymixtures from Cd taken up by the plant material within 3 dfrom a 50 Mm Cd2+ containing nutrient solution.The extractable activity ofATP-sulfurylase, the first enzyme
of assimilatory sulfate reduction, was not changed in theleaves due to Cd after 24 h. After 48 and 72 h, it was belowthe control value at all Cd concentrations applied. In theroots, the largest increase in activity ofabout 30% was inducedby 50 Mm Cd2' after 24 h. This enzyme activity was compa-rable to the control level 48 h after the various Cd2' additions(data not shown).Compared to controls, the extractable activity of nitrate
reductase was reduced to 80% and 25% after 24 h with 5 gMCd2+ in the roots and shoots, respectively. With 50 Mm Cd2 ,the enzyme activity decreased to 35% and 7% of the controllevel for roots and shoots, respectively (data not shown).The effect of BSO, an inhibitor of glutathione synthesis, on
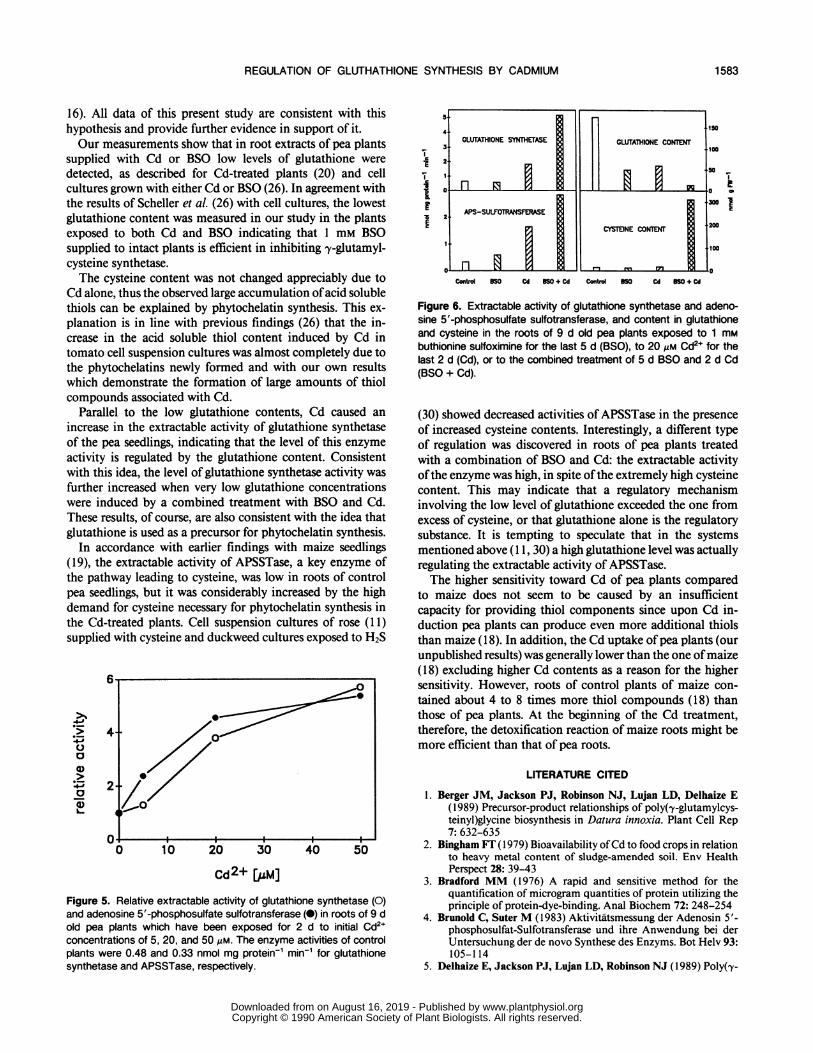
the content of glutathione and cysteine and on glutathionesynthetase and APSSTase activity is shown in Figure 6. Sur-prisingly, the combined action of Cd and BSO induced thehighest level of both enzyme activities even though we couldnot detect phytochelatins under these conditions (Fig. 3). Theglutathione content was reduced by BSO or Cd, a very lowcontent was measured with BSO and Cd combined. Thistreatment also caused the largest effect on the cysteine contentwith a 21-fold increase compared with the control value,whereas Cd or BSO alone had no effect.The supply of 1 mm BSO alone for 5 d had no effect on
nitrate reductase activity in the roots, 20 gM Cd2" alonedecreased it to 60%, whereas the combined treatment withBSO and Cd resulted in a level which was 15% of the control(data not shown).
DISCUSSION
Strong evidence has been presented that shows glutathioneis involved in the reactions forming phytochelatins (1, 10,
E9IE
cE
i
Houm
EOI9
Hourm
Figure 4. Extractable activity of glutathione synthetase and adeno-sine 5'-phosphosulfate sulfotransferase in roots and shoots of 7 dold pea plants grown without Cd (0) or with 50 uM Cd2+ (A) for 3more days.
RUEGSEGGER ET AL.1 582
www.plantphysiol.orgon August 16, 2019 - Published by Downloaded from Copyright © 1990 American Society of Plant Biologists. All rights reserved.
REGULATION OF GLUTHATHIONE SYNTHESIS BY CADMIUM
16). All data of this present study are consistent with thishypothesis and provide further evidence in support of it.Our measurements show that in root extracts of pea plants
supplied with Cd or BSO low levels of glutathione weredetected, as described for Cd-treated plants (20) and cellcultures grown with either Cd or BSO (26). In agreement withthe results of Scheller et al. (26) with cell cultures, the lowestglutathione content was measured in our study in the plantsexposed to both Cd and BSO indicating that 1 mm BSOsupplied to intact plants is efficient in inhibiting y-glutamyl-cysteine synthetase.The cysteine content was not changed appreciably due to
Cd alone, thus the observed large accumulation ofacid solublethiols can be explained by phytochelatin synthesis. This ex-planation is in line with previous findings (26) that the in-crease in the acid soluble thiol content induced by Cd intomato cell suspension cultures was almost completely due tothe phytochelatins newly formed and with our own resultswhich demonstrate the formation of large amounts of thiolcompounds associated with Cd.
Parallel to the low glutathione contents, Cd caused anincrease in the extractable activity of glutathione synthetaseof the pea seedlings, indicating that the level of this enzymeactivity is regulated by the glutathione content. Consistentwith this idea, the level of glutathione synthetase activity wasfurther increased when very low glutathione concentrationswere induced by a combined treatment with BSO and Cd.These results, of course, are also consistent with the idea thatglutathione is used as a precursor for phytochelatin synthesis.
In accordance with earlier findings with maize seedlings(19), the extractable activity of APSSTase, a key enzyme ofthe pathway leading to cysteine, was low in roots of controlpea seedlings, but it was considerably increased by the highdemand for cysteine necessary for phytochelatin synthesis inthe Cd-treated plants. Cell suspension cultures of rose (11)supplied with cysteine and duckweed cultures exposed to H2S
-4._
4-,-
C,
0
L.
2t
10 20 30 40 50
Cd2+ [,sM]
Figure 5. Relative extractable activity of glutathione synthetase (0)and adenosine 5'-phosphosulfate sulfotransferase (0) in roots of 9 dold pea plants which have been exposed for 2 d to initial Cd2+concentrations of 5, 20, and 50 Mm. The enzyme activities of controlplants were 0.48 and 0.33 nmol mg protein-' min-' for glutathionesynthetase and APSSTase, respectively.
I
cE
I
Conftrol 30 Cd BSO+Cd Control 130 Cd BSO+Cd
Figure 6. Extractable activity of glutathione synthetase and adeno-sine 5'-phosphosulfate sulfotransferase, and content in glutathioneand cysteine in the roots of 9 d old pea plants exposed to 1 mMbuthionine sulfoximine for the last 5 d (BSO), to 20 Mm Cd2+ for thelast 2 d (Cd), or to the combined treatment of 5 d BSO and 2 d Cd(BSO + Cd).
(30) showed decreased activities of APSSTase in the presenceof increased cysteine contents. Interestingly, a different typeof regulation was discovered in roots of pea plants treatedwith a combination of BSO and Cd: the extractable activityofthe enzyme was high, in spite ofthe extremely high cysteinecontent. This may indicate that a regulatory mechanisminvolving the low level of glutathione exceeded the one fromexcess of cysteine, or that glutathione alone is the regulatorysubstance. It is tempting to speculate that in the systemsmentioned above (1 1, 30) a high glutathione level was actuallyregulating the extractable activity of APSSTase.The higher sensitivity toward Cd of pea plants compared
to maize does not seem to be caused by an insufficientcapacity for providing thiol components since upon Cd in-duction pea plants can produce even more additional thiolsthan maize (18). In addition, the Cd uptake ofpea plants (ourunpublished results) was generally lower than the one ofmaize(18) excluding higher Cd contents as a reason for the highersensitivity. However, roots of control plants of maize con-tained about 4 to 8 times more thiol compounds (18) thanthose of pea plants. At the beginning of the Cd treatment,therefore, the detoxification reaction of maize roots might bemore efficient than that of pea roots.
LITERATURE CITED
1. Berger JM, Jackson PJ, Robinson NJ, Lujan LD, Delhaize E(1989) Precursor-product relationships of poly(y-glutamylcys-teinyl)glycine biosynthesis in Datura innoxia. Plant Cell Rep7: 632-635
2. Bingham F' (1979) Bioavailability ofCd to food crops in relationto heavy metal content of sludge-amended soil. Env HealthPerspect 28: 39-43
3. Bradford MM (1976) A rapid and sensitive method for thequantification of microgram quantities of protein utilizing theprinciple of protein-dye-binding. Anal Biochem 72: 248-254
4. Brunold C, Suter M (1983) Aktivititsmessung der Adenosin 5'-phosphosulfat-Sulfotransferase und ihre Anwendung bei derUntersuchung der de novo Synthese des Enzyms. Bot Helv 93:105-114
5. Delhaize E, Jackson PJ, Lujan LD, Robinson NJ (1989) Poly(y-
10~~~~~~~~~~~10I-~~ ~ ~ ~ ~ ~ -
b I
LI ||.
1 583
www.plantphysiol.orgon August 16, 2019 - Published by Downloaded from Copyright © 1990 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 93,1990
glutamylcysteinyl)glycine synthesis on Datura innoxia andbinding with cadmium. Plant Physiol 89: 700-706
6. Ellman GL (1959) Tissue sulfhydryl groups. Arch Biochem Bio-phys 82: 70-77
7. Fowles BA, Hildrebrand CD, Koyima Y, Webb M (1987) No-menclature of metallothionein. In JHR Kagi, Y Koyima, eds,Metallothionein II. Birkhauser Verlag, Basel, pp 19-22
8. Gekeler W, Grill E, Winnacker E-L, Zenk MH (1989) Surveyof the plant kingdom for the ability to bind heavy metalsthrough phytochelatins. Z Naturforsch 44c: 361-369
9. Grill D, Esterbauer H (1973) Quantitative Bestimmung wasser-loslicher Sulfhydrylverbindungen in gesunden und geschadig-ten Nadeln von Picea abies. Phyton 15: 87-101
10. Grill E, LUffler S, Winnacker EL, Zenk MH (1989) Phytoche-latins, the heavy-metal-binding peptides of plants, are synthe-sized from glutathione by a specific -y-glutamylcysteine dipep-tidyl transpeptidase (phytochelatin synthase). Proc Natl AcadSci USA 86: 6838-6842
11. Haller E, Suter M, Brunold C (1986) Regulation of ATP-sulfur-ylase and adenosine 5'-phosphosulfate sulfotransferase by thesulfur and the nitrogen source in heterotropic cell suspensioncultures of Paul's Scarlet Rose. J Plant Physiol 125: 275-283
12. Jackson PJ, Unkefer CJ, Doolen JA, Watt K, Robinson NJ(1987) Poly(y-glutamylcysteinyl)glycine: its role in cadmiumresistance in plant cells. Proc Natl Acad Sci USA 84: 6619-6623
13. Kloke A, Schenke HD (1979) Influence of cadmium in soil tothe yield of various plant species and their cadmium content.Z Pflanzenernahr Bodenkd 142: 131-136
14. Kondo N, Imai K, Isobe M, Goto T (1984) Cadystin A and B,major unit peptides comprising cadmium binding peptidesinduced in a fission yeast-separation, revision of structures andsynthesis. Tetrahedron Lett 25: 3869-3872
15. Law MY, Halliwell B (1986) Purification and properties ofglutathione synthetase from spinach (Spinacia oleracea) leaves.Plant Sci 43: 185-191
16. Mendum ML, Gupta SC, Goldsbrough PB (1989) Effect ofglutathione on phytochelatin synthesis and cadmium tolerancein tomato cells (abstract No. 718). Plant Physiol 89: S- 120
17. Mutoh N, Hayashi Y (1988) Isolation of mutants of Schizosac-charomyces pombe unable to synthesize cadystin, small cad-
mium-binding peptides. Biochem Biophys Res Commun 151:32-39
18. Nussbaum S (1987) Effekt von Cadmium auf Enzyme der assi-milatorischen Sulfatreduktion von Zea mays L., Phaseolusvulgaris L. und Triticum aestivum L. Diplomarbeit, Universi-tAt Bern, Switzerland
19. Nussbaum S, Schmutz D, Brunold C (1988) Regulation ofassim-ilatory sulfate reduction by cadmium in Zea mays L. PlantPhysiol 88: 1407-14 10
20. Rauser WE (1987) Changes in glutathione content of maizeseedlings exposed to cadmium. Plant Sci 51: 171-175
21. Reese RN, Wagner GJ (1987) Effects of buthionine sulfoximineon Cd-binding peptide levels in suspension-cultured tobaccocells treated with Cd, Zn, or Cu. Plant Physiol 84: 574-577
22. Reese RN, Wagner GJ (1987) Properties of tobacco (Nicotianatabacum) cadmium-binding peptide(s). Biochem J 241: 641-647
23. Reese RN, Mehra RK, Tarbet EB, Winge DR (1988) Studies onthe -y-glutamyl Cu-binding peptide from Schizosaccharomycespombe. J Biol Chem 263: 4186-4192
24. Richie JP, Lang CA (1987) The determination of glutathione,cyst(e)ine, and other thiols and disulfides in biological samplesusing high-performance liquid chromatography with dual elec-trochemical detection. Anal Biochem 163: 9-15
25. Robinson NJ, Ratliff RL, Anderson PJ, Delhaize E, Berger JM(1988) Biosynthesis of Poly(y-glutamylcysteinyl)glycines incadmium-tolerant Datura innoxia (Mill.) cells. Plant Sci 56:197-204
26. Scheller HV, Huang B, Hatch E, Goldsbrough PB (1987) Phy-tochelatin synthesis and glutathione levels in response to heavymetals in tomato cells. Plant Physiol 85: 1031-1035
27. Schiff JA (1983) Reduction and other metabolic reactions ofsulfate. In A Lauchli, RL Bielski, eds. Enzyclopedia of PlantPhysiology, Vol 15A. Springer-Verlag, Berlin, pp 401-421
28. Steffens JC, Williams BG (1987) Increased activity of y-glutam-ylcysteine synthetase in cadmium resistant tomato cells (ab-stract No. 666). Plant Physiol 83: S-llO
29. Steffens JC, Hunt DF, Williams BG (1986) Accumulation ofnon-protein metal-binding polypeptides (-y-glutamylcystei-nyl),-glycine in selected cadmium resistant tomato cells. J BiolChem 261: 13879-13882
30. Von Arb C, Brunold C (1980) Analysis of the regulation ofadenosine 5 '-phosphosulfate sulfotransferase activity in Lemnaminor L. using '5N-density labeling. Planta 149: 355-360
RUEGSEGGER ET AL.1 584
www.plantphysiol.orgon August 16, 2019 - Published by Downloaded from Copyright © 1990 American Society of Plant Biologists. All rights reserved.











![Review Article Role of Glutathione in Cancer Progression ...downloads.hindawi.com/journals/omcl/2013/972913.pdf · GCL and glutathione S-transferases [ ]. 2. GSH Biosynthesis Glutathione](https://static.fdocuments.us/doc/165x107/5edbd12aad6a402d666637cd/review-article-role-of-glutathione-in-cancer-progression-gcl-and-glutathione.jpg)






