Prospective isolation of human erythroid lineage-committed ...

Receptors for Erythropoietin in Mouse and Human Erythroid Cells and Placenta
Blood, Vol 74, No 1 (July). 1989: pp 103-109 103
By Stephen T. Sawyer. Sanford B. Krantz, and Ken-ichi Sawada
High and lower affinity receptors for erythropoietin (EP)were initially identified on a very pure population of EP-responsive erythroblasts obtained from the spleens of miceinfected with anemia strain of Friend virus (FVA). The
structure of the receptor for EP in these cells was deter-
mined to be proteins of 100 and 85 Kd by cross-linking‘25l-EP. In this investigation. studies on the receptors for EPwere extended to other mouse erythroid cells and humanerythroid cells as well as to the placentas of mice and rats.Only lower affinity receptors for EP were detected onerythroblasts purified from the spleens of mice infected
with the polycythemia strain of Friend virus and a murineerythroleukemia cell line. both of which are not responsive
to EP in culture. Internalization of ‘�l-EP was observed inboth groups of cells. The structure of the receptor deter-mined by cross-linking ‘25l-EP was two equally labeledproteins of 100 Kd and 85 Kd molecular mass in all these
S PECIFIC BINDING of erythropoietin (EP) was first
observed in erythroblasts purified from spleens of mice
infected with the anemia strain of Friend virus (FVA).’3
These immature FVA-infected erythroid cells (FVA cells)
respond to physiological levels of EP in culture by pro-
gressing to near erythroid maturation.� ‘251-EP binds to
higher and lower affinity receptors on the surface of these
FVA cells and is subsequently internalized and degraded in
the lysosomal compartment. ‘In our initial investigation,
‘251EP binding was also observed in murine erythroleukemia
(MEL) cells, clone 745, but only to lower affinity receptors.’
In this study, we characterize the internalization of ‘25I-EP in
these MEL cells, which are not responsive to EP in culture,
and the binding and internalization of ‘25l-EP in erythroid
cells purified from the spleens of mice infected with the
polycythemia strain of Friend virus (FVP cells), which
spontaneously differentiate in vitro in the absence of exoge-
nously added EP. The existence of receptors for EP in
membranes prepared from placentas from mice and rats is
also identified.
When ‘251-EP bound to FVA cell membranes was cross-
linked by disuccinimydl suberate (DSS) to the receptor, two
labeled bands corresponding to proteins of molecular weights
of 100 and 85 Kd were observed on sodium dodecyl sulfate
polyacrylamide gel electrophoresis (SDS-PAGE).3 No evi-
dence of disulfide bridges between these two proteins was
found. More recently other investigators have presented
evidence that suggests the existence of additional lower
molecular weight proteins in the receptor7�’#{176} and have sug-
gested that the multiply labeled proteins may be subunits of a
very large complex that is bridged by disulfide bonds.8”#{176}We
have examined this possibility by cross-linking ‘25I-EP to the
receptor in MEL cells, FVP cells, human erythroid colony-
forming cells (CFU-E), and placentas from mice and rats.
The data presented here show a remarkable similarity of the
receptors for EP in human, mouse, and rat tissues and in EP
responsive and nonresponsive erythroid cells. No evidence of
additional subunits of the receptor or a larger complex of
subunits was detected in any of these sources of receptor.
mouse erythroid cells. The structure of the receptor wasfound to be very similar in human erythroid colony formingcells cultured from normal blood. These cells respond to EPwith erythroid maturation and were previously shown tohave high and lower affinity receptors. Placentas from miceand rats were found to have only lower affinity receptorsfor EP. and when placental membranes were cross-linkedto ‘25l-EP, the same 100 Kd and 85 Kd bands were found asseen in mouse and human erythroid cells. The structure ofthe receptor was similar in cells that have high affinityreceptors (FVA-infected and human erythroid colony-forming cells) and nonresponsive erythroid cells and pla-
centa that have lower affinity receptors. but only the cellswith the high affinity receptors respond to the addition ofEP with erythroid maturation.S 1989 by Grune Stratton, Inc.
MATERIALS AND METHODS
Human recombinant EP was purchased from AmGen Biologicals
(Thousand Oaks, CA). Na ‘25I was obtained from Amersham. DSS
and lODO-GEN (l,3,4,6-tetra-chloro-3a, 6a-diphenylglycouril)
were obtained from Pierce (Rockford, IL). Friend virus, pseudotype
SFFVA/FRE cl-3/MuLV (201) originally obtained from W.D.
Hankins (National Institutes of Health [NIH]) and FVP obtainedfrom R. Holdenreid (NIH) were maintained by the passage of
infectious plasma in BALB/c mice. MEL cells, clone 745, were
obtained from W. LeStourgeon, Vanderbilt University.
Cells and plasma membrane preparation. Immature erythroidcells were purified from the spleens of CD2F, mice infected with
FVA or FVP by velocity sedimentation at unit gravity through a
continuous gradient of bovine serum albumin as described previous-
ly.4’5 To prepare plasma membranes from FVA- or FVP-infected
erythroid cells, the total spleen was disrupted to a single cell
suspension and the erythrocytes were lysed by exposure to ISO
mmol/L NH4CI/15 mmol/L Tris HCI, pH 7.65, at 37#{176}Cfor 30
seconds. The NH4CI/Tris solution was diluted fourfold with Iscove’s
modified Dulbecco’s medium and the cells were pelleted by centrifu-
gation at 500 g for I 5 minutes. The pellet was resuspended in
NH4CI/Tris and the procedure was repeated. The cells were then
washed in 105 mmol/L NaCI and 10 mmol/L Tricine, pH 7.4, three
times and resuspended in 10 mmol/L KCI and 10 mmol/L Tricine,
From the Division of Hematology, Department of MedicineVanderbilt University School of Medicine and Veterans Adminis-
tration Medical Center, Nashville,
Submitted August 9, 1988; accepted March 3, 1989.
Supported by grants from the National Institutes of Health,
DK-39781. AM-15555. T32 DK-07186 and VA Medical Research
Funds.Address reprint requests to Stephen T. Sawyer. PhD, Division of
Hematology, Room C-3101. Medical Center North, Vanderbilt
University School ofMedicine. Nashville, TN 37232.
The publication costs ofthis article were defrayed in part by page
charge payment. This article must therefore be hereby marked“advertisement” in accordance with 18 U.S.C. section 1734 solely to
indicate this fact.
© I 989 by Grune & Stratton, Inc.
0006-4971/89/7401-0034$3.OO/O
For personal use only.on January 3, 2019. by guest www.bloodjournal.orgFrom

104 SAWYER, KRANTZ. AND SAWADA
pH 7.4, containing a mixture of proteinase inhibitors (2 mmol/L
EGTA, 5 mmol/L EDTA, 1 �g leupeptin/mL, 5 mmol/L
ide, 1 mmol/L iodoacetamide, 10 �g/mL tosylamide-2-phenylethl-
chloromethyl ketone, 10 �g/mL p-tosyl-1-arginine methylester, 1
�g/mL n-d-tosyl-l-Tysine chloromethylketone, 2 �zg/mL aprotinin,
I zg/mL pepstatin, and 0.1 mmol/L penylmethyl-sulfonylfluorine).
Plasma membrane fractions of MEL cells, FVP cells, and FVAcells were prepared in identical fashion as described previously3
except that the above described mixture of proteinase inhibitors was
included at every step. Briefly, the cells were swollen in hypotonic
medium, and disrupted with ten strokes of a teflon, motor-driven
homogenizer. The cell debris was discarded after low speed centrifu-
gation, and a crude membrane pellet was obtained after one hour
centrifugation at I 50,000 g. This pellet was resuspended in a solution
containing a final concentration of 40% sucrose, which became the
bottom of a discontinuous sucrose gradient composed of solutions of
35%, 31%, 25%, and 8.5% sucrose. The light membrane fractions at
the 35%/3 1% interface and 3 l%/25% interface were collected as the
plasma membrane fraction.
Placenta membranes were prepared in a similar fashion. Placen-
tas from CD2F, mice were taken at days 14 and 18 of gestation, and
placentas from Sprague Dawley rats were taken at 1 8 to 20 days of
gestation. Placentas were washed in phosphate buffered saline
containing the above mentioned cocktail of protease inhibitors and
finely minced. The minced placenta was then homogenized, centri-
fuged, and crude membranes were fractionated on discontinuous
sucrose gradient exactly as described above for the erythroid cells.
Human CFU-E were obtained as described previously.” Briefly,
erythroid burst forming units were partially purified from normal
human blood. These cells were then cultured in the presence of EP to
the stage where the cells were not producing a significant level of
hemoglobin and were capable of forming colonies of eight to 49 cells
when further cultured in the presence of EP. At this stage the cells
were removed from culture and purified additionally by removing
adherent cells and by Ficoll-Hypaque density centrifugation. Purity
of the cells was >50% and receptors for EP have been found on the
cell surface at about 1 ,000 receptors per cell.’2
lodination of EP. EP was iodinated using IODO-GEN.’ Two
micrograms of lODO-GEN were coated on the walls of a conical
reaction vial. EP (50 units, -.4 �zg protein) and 20 �iCi 251 were
incubated in the reaction vial for five minutes at room temperature
in a final volume of 50 �L of 0.5 mol/L phosphate buffer, pH 7.0,
containing 0.02% Tween 20. After the incubation, the contents of the
reaction vial were transferred to a tube containing 10 mg KI in
phosphate buffered saline, 0.1% bovine serum albumin, and 0.02%
Tween 20; ‘251-EP was separated from the free 1251 by chroma-
tography over a Biogel P6 column. This procedure provided EP with
0.3 to I .0 molecule of 251 per molecule (25 to 75 �Ci/�zg) and with
full biological activity when assayed in these FVA-infected erythroid
cells.4
Binding ‘251-EP. ‘25I-EP was incubated with from 10 to 40 �gprotein of the plasma membrane fraction from FVA cells, FVP cells,
MEL cells, and placentas from mice and rats at 37#{176}Cfor the time
indicated in 100 mmol/L phosphate buffer, pH 7.4, containing I
mmol/L EGTA and 0.1% bovine serum albumin. The binding
mixture was then applied to 0.2 zm Millipore filters (EHWP) and
washed with 10 mL of phosphate buffered saline containing 0.1%
bovine serum albumin. The filters were then counted in a gamma
counter. Nonspecific binding was determined in the presence of 100
to 200 units of EP/mL and was subtracted.
FVA cells, FVP ceils, and MEL cells were washed and resus-
pended in binding medium (Iscove’s modified Dulbecco’s medium)
supplemented to contain 20 mmol/L HEPES, pH 7.4, and 2%
bovine serum albumin. The cells were allowed to stand for one hour
at 37#{176}Cin a 5% CO2 atmosphere before the initiation of binding
studies. For binding at 37#{176}Clabeled EP was added to the cells in thebinding medium at varying concentrations for different times in the
incubator. The concentration of cells was iO� cells/mL or less.
Binding was terminated by sedimenting the cells through dibutyl
phthalate oil (0.5 mL) for one minute in a minifuge (8,000 g). The
tube was frozen at -80#{176}C, and the tip containing the cell pellet was
cut off. Radioactivity in the tip was determined by counting the tip in
a gamma counter. Nonspecific binding of the labeled EP was
determined by adding a 20 to 100-fold excess of unlabeled EP in the
binding assay (100 to 200 units EP/mL).
For binding at 0#{176}C,the protocol was essentially the same as at
37#{176}Cexcept that the cells were cooled for one hour in an ice bath at
4#{176}C,and the incubation was carried out in sealed 1 .5 mL minifuge
tubes in the ice bath.
For determination of the binding affinities by the method of
Scatchard,’3 the cells were incubated for 20 hours at 4#{176}C.This time
is sufficient for the binding to plateau, the cell viability as measured
by trypan blue exclusion to remain >90% viable, and bound EP to be
substantially released. However the binding of ‘251-EP is not totally
reversible at 20 hours (60% of ‘25I-EP bound was released). ‘251-EP
bound to FVA cells was not totally reversible during a reasonable
period at 4#{176}C.Extrapolation of the second order release curve
showed 90% of bound ‘25I-EP was released at 72 hours. However,binding at 4#{176}Cfor this length of time or greater is not feasible since
the cells are not stable and they disintegrate. Nevertheless, binding
at 48 hours at 4#{176}Cdid not affect the binding constant for FVA cells.
Therefore these studies were carried out at 20 hours for conve-
nience.
Determination of surface-bound and internalized EP. An acidwash, which has been used previously to remove surface-bound
ligands,’’4 was used to remove surface-bound labeled EP from the
cell surface. At the indicated times after labeled EP was added to the
cells, the cells were cooled to 0#{176}Cby the addition of a ninefold excess
of coldbinding medium and transferred to an ice bath. After three
washes to remove unbound ligand, the cells were pelleted by
centrifugation in a minifuge for 30 seconds and resuspended in cold
0.5 mmol/L NaCI and to 0.25 mmol/L acetic acid, pH 2.5. Afterthree minutes, the cells were centrifuged through dibutyl phthalate
oil. The tube was frozen, and the tip containing the pellet was cut off
and counted. The aqueous phase also was counted. Parallel experi-
ments were performed with a large excess of unlabeled EP in the
binding assay, and the cells were treated with the high salt, pH 2.5,
wash to determine the distribution of the nonspecifically bound EP
between the acid labile and stable radioactivity. After nonspecific
radioactivity was subtracted, the labeled EP in the acid wash was
considered surface bound, while the labeled EP resistant to the acid
wash was considered internalized.
Cross-linking of251-EP. ‘251-EP was cross-linked to the surface
of human erythroid colony-forming cells with DSS in a similar
fashion as described previously for FVA cells.3 25I-EP was added to10’ cells for 30 minutes at 37#{176}C.DSS was added at 500 zm for 15
minutes at 0#{176}C.The cells were washed three times with 150 mmol/L
Tris-HC1, pH 7.4, to quench the cross-linking reaction and remove
unbound EP. The cells were then extracted for one minute at 0#{176}C
with a solution containing 1.0% Triton X-lOO, 20 mmol/L HEPES,
pH 7.4, containing the mixture of 1 1 protease inhibitors described
above. The disrupted cell preparation was then centrifuged at 1,000
g for one minute to remove nuclei and cell debris; the supernatant
and pellet were both analyzed by SDS-PAGE.
�25��p was bound to receptors on the plasma membrane fraction
and was cross-linked in a similar fashion as described previously.3
Membranes were incubated with 7.5 units of ‘25I-EP per milliliter in
100 mmol/L phosphate buffer, pH 7.4, containing I mmol/L EGTA
and 0.1% bovine serum albumin for 15 minutes at 37#{176}C.The mixture
was transferred to an ice bath, and DSS was added to a final
For personal use only.on January 3, 2019. by guest www.bloodjournal.orgFrom
TIME, minutes
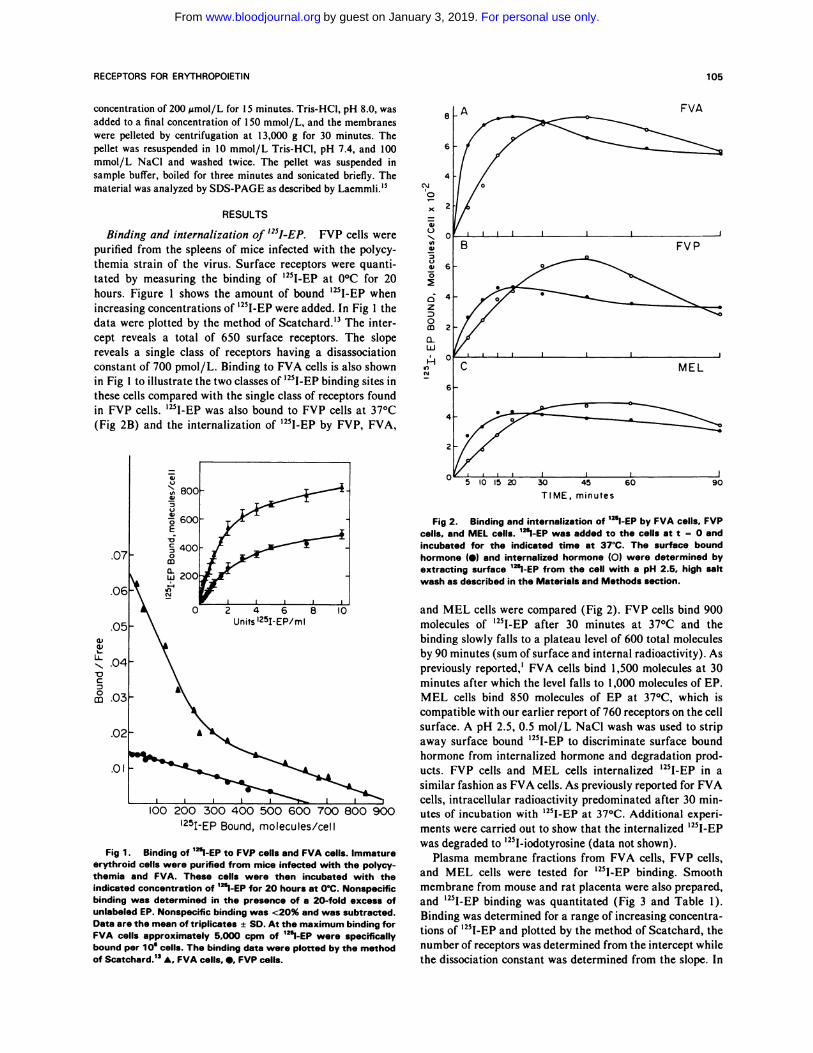
‘251-EP Bound, molecules/cell
Fig 1 . Binding of �l-EP to FVP cells and FVA cells. Immatureerythroid cells were purified from mice infected with the polycy-themia and FVA. These cells were then incubated with theindicated concentration of ‘�l-EP for 20 hours at O’C. Nonspecific
binding was determined in the presence of a 20-fold excess ofunlabeled EP. Nonspecific binding was <20% and was subtracted.Data are the mean of triplicates ± SD. At the maximum binding for
FVA cells approximately 5.000 cpm of ‘�l-EP were specificallybound per 1 0 cells. The binding data were plotted by the methodof Scatchard.’3 A, FVA cells, �, FVP cells.
RECEPTORS FOR ERYTHROPOIETIN 105
concentration of 200 �imol/L for 15 minutes. Tris-HCI, pH 8.0, was
added to a final concentration of 1 50 mmol/L, and the membranes
were pelleted by centrifugation at 13,000 g for 30 minutes. The
pellet was resuspended in 10 mmol/L Tris-HCI, pH 7.4, and 100
mmol/L NaC1 and washed twice. The pellet was suspended in
sample buffer, boiled for three minutes and sonicated briefly. The
material was analyzed by SDS-PAGE as described by Laemmli.’5
RESULTS
Binding and internalization of ‘251-EP. FVP cells were
purified from the spleens of mice infected with the polycy- .�
themia strain of the virus. Surface receptors were quanti- �
tated by measuring the binding of ‘25I-EP at 0#{176}Cfor 20 �
hours. Figure 1 shows the amount of bound ‘25I-EP when
increasing concentrations of ‘25I-EP were added. In Fig 1 the
data were plotted by the method of Scatchard.’3 The inter- �
cept reveals a total of 650 surface receptors. The slope o
reveals a single class of receptors having a disassociation �
constant of 700 pmol/L. Binding to FVA cells is also shown
in Fig 1 to illustrate the two classes of ‘251-EP binding sites in
these cells compared with the single class of receptors found
in FVP cells. ‘25I-EP was also bound to FVP cells at 37#{176}C
(Fig 2B) and the internalization of ‘25I-EP by FVP, FVA,
Fig 2. Binding and internalization of ‘251-EP by FVA cells. FVPcells. and MEL cells. 1�l-EP was added to the cells at t = 0 andincubated for the indicated time at 37’C. The surface bound
hormone (#{149})and internalized hormone (0) were determined byextracting surface 12�l-EP from the cell with a pH 2.5. high saltwash as described in the Materials and Methods section.
and MEL cells were compared (Fig 2). FVP cells bind 900
molecules of ‘251-EP after 30 minutes at 37#{176}Cand the
� binding slowly falls to a plateau level of 600 total molecules
L� by 90 minutes (sum ofsurface and internal radioactivity). As:� previously reported,’ FVA cells bind 1,500 molecules at 30� minutes after which the level falls to I ,000 molecules of EP.
c� MEL cells bind 850 molecules of EP at 37#{176}C,which iscompatible with our earlier report of 760 receptors on the cell
surface. A pH 2.5, 0.5 mol/L NaC1 wash was used to strip
away surface bound ‘251-EP to discriminate surface bound
hormone from internalized hormone and degradation prod-
ucts. FVP cells and MEL cells internalized ‘25I-EP in a
similar fashion as FVA cells. As previously reported for FVA
cells, intracellular radioactivity predominated after 30 mm-
utes of incubation with ‘251-EP at 37#{176}C.Additional experi-
ments were carried out to show that the internalized ‘251-EP
was degraded to ‘25I-iodotyrosine (data not shown).
Plasma membrane fractions from FVA cells, FVP cells,
and MEL cells were tested for ‘251-EP binding. Smooth
membrane from mouse and rat placenta were also prepared,
and ‘25I-EP binding was quantitated (Fig 3 and Table 1).
Binding was determined for a range of increasing concentra-
tions of ‘25I-EP and plotted by the method of Scatchard, the
number of receptors was determined from the intercept while
the dissociation constant was determined from the slope. In
For personal use only.on January 3, 2019. by guest www.bloodjournal.orgFrom
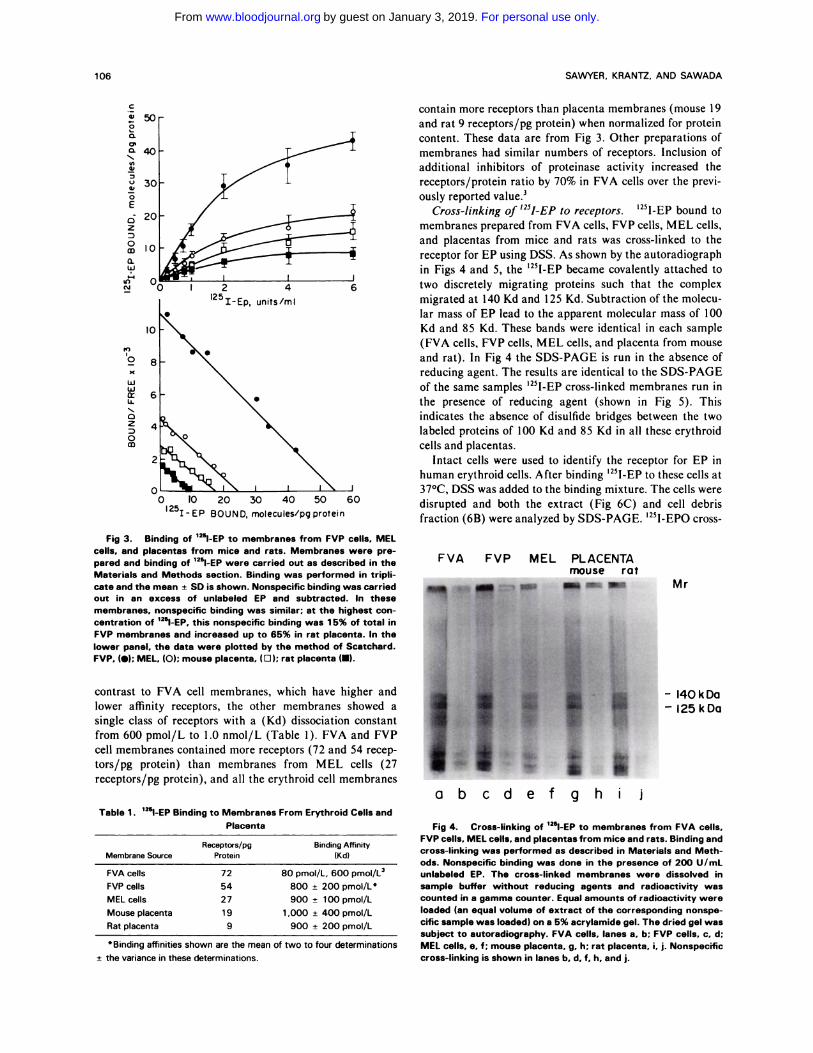
0 10 20 30 40 50 60l25� � EP BOUND, molecules/pg protein
- l4OkDa- 125 kDa
abcd
± the variance in these determinations.
106 SAWYER, KRANTZ, AND SAWADA
C
a,0
a’a
.%..‘I,0)
‘3
0�
0
E
0z:20
0.Ui‘.4
In
‘I)
0
UiUia:L�.
0z
0
Fig 3. Binding of ‘251-EP to membranes from FVP cells. MELcells. and placentas from mice and rats. Membranes were pre-pared and binding of 125l-EP were carried out as described in theMaterials and Methods section. Binding was performed in tripli-cate and the mean ± SD is shown. Nonspecific binding was carriedout in an excess of unlabeled EP and subtracted. In thesemembranes. nonspecific binding was similar; at the highest con-centration of lThl�EP, this nonspecific binding was 1 5% of total inFVP membranes and increased up to 65% in rat placenta. In the
lower panel. the data were plotted by the method of Scatchard.
FVP. (#{149});MEL. (0); mouse placenta. (D); rat placenta (s).
contrast to FVA cell membranes, which have higher and
lower affinity receptors, the other membranes showed a
single class of receptors with a (Kd) dissociation constant
from 600 pmol/L to 1.0 nmol/L (Table 1). FVA and FVP
cell membranes contained more receptors (72 and 54 recep-
tors/pg protein) than membranes from MEL cells (27
receptors/pg protein), and all the erythroid cell membranes
Table 1 . ‘�I-EP Binding to Membranes From Erythroid Cells and
Placenta
Membrane SourceReceptors/pg
ProteinBinding Affinity
(Kd)
FVA cells 72 80 pmol/L, 600 pmol/L3
FVP cells 54 800 ± 200 pmol/L’
MEL cells 27 900 ± 100 pmol/L
Mouse placenta 1 9 1 .000 ± 400 pmol/L
Rat placenta 9 900 ± 200 pmol/L
Binding affinities hown are the mea n of two to four determinations
contain more receptors than placenta membranes (mouse I 9
and rat 9 receptors/pg protein) when normalized for protein
content. These data are from Fig 3. Other preparations of
membranes had similar numbers of receptors. Inclusion of
additional inhibitors of proteinase activity increased the
receptors/protein ratio by 70% in FVA cells over the previ-
ously reported value.3
Cross-linking of ‘251-EP to receptors. ‘25l-EP bound to
membranes prepared from FVA cells, FVP cells, MEL cells,
and placentas from mice and rats was cross-linked to the
receptor for EP using DSS. As shown by the autoradiograph
in Figs 4 and 5, the ‘251-EP became covalently attached to
two discretely migrating proteins such that the complex
migrated at 140 Kd and I 25 Kd. Subtraction of the molecu-
lar mass of EP lead to the apparent molecular mass of 100
Kd and 85 Kd. These bands were identical in each sample
(FVA cells, FVP cells, MEL cells, and placenta from mouse
and rat). In Fig 4 the SDS-PAGE is run in the absence of
reducing agent. The results are identical to the SDS-PAGE
of the same samples ‘251-EP cross-linked membranes run in
the presence of reducing agent (shown in Fig 5). This
indicates the absence of disulfide bridges between the two
labeled proteins of 100 Kd and 85 Kd in all these erythroid
cells and placentas.
Intact cells were used to identify the receptor for EP in
human erythroid cells. After binding ‘25I-EP to these cells at
37#{176}C,DSS was added to the binding mixture. The cells were
disrupted and both the extract (Fig 6C) and cell debris
fraction (6B) were analyzed by SDS-PAGE. ‘25I-EPO cross-
FVA FVP MEL PLACENTAmouse rat
efghi j
Mr
Fig 4. Cross-linking of 1251-EP to membranes from FVA cells.FVP cells, MEL cells. and placentas from mice and rats. Binding andcross-linking was performed as described in Materials and Meth-ods. Nonspecific binding was done in the presence of 200 U/mLunlabeled EP. The cross-linked membranes were dissolved insample buffer without reducing agents and radioactivity wascounted in a gamma counter. Equal amounts of radioactivity wereloaded (an equal volume of extract of the corresponding nonspe-cific sample was loaded) on a 5% acrylamide gel. The dried gel wassubject to autoradiography. FVA cells. lanes a. b; FVP cells. c. d;MEL cells. e. f; mouse placenta. g. h; rat placenta. i. j. Nonspecificcross-linking is shown in lanes b, d. f. h. and j.
For personal use only.on January 3, 2019. by guest www.bloodjournal.orgFrom

RECEPTORS FOR ERVTHROPOIETIN 107
Mr
- 140 kDa- 125 kDa
Mr
k Do
200
-140-130-I--. 25-‘- I 10
92
ABCDE
Fig 5. Cross-linking of ‘25l-EP to membranes from FVA cells.FVP cells. MEL cells. and placentas from mice and rats. Onemilligram of membrane protein was incubated with 5 units of‘�‘l-EP for 1 5 minutes and cross-linked with 0.2 mmol/L DSS. Thesamples were analyzed by SDS-PAGE in the presence of fi-
mercaptoethanol and autoradiography. Lane A. FVA cells; B. FVPcells; C. MEL cells; D. mouse placenta; E. rat placenta.
linked proteins were found to migrate in the gel at a position
corresponding to a molecular mass of 140 Kd and approxi-
mately 130 Kd. While the 140 Kd band is identical to that
found in mouse tissues, the lesser molecular weight band
appears just slightly larger than the corresponding band in
mouse FVA cells (Fig 6D). A minor band that migrates at
1 10 Kd was also seen in the cell extract but not in the cell
debris fraction. The presence of this band was variable from
one experiment to another and may be a proteolytic fragment
of the larger cross-linked bands. The gel pattern was identi-
cal in the presence and absence of reducing agents.
DISCUSSION
Following the appearance of our report of the structure of
the receptor in FVA cells as two proteins of 100 Kd and 85
Kd molecular mass,3 Todakaro et a17 reported that the
receptor in an EP responsive cell line was composed of three
subunits of I I 9, 94, and 63 Kd, which were not connected
through disulfide bridges. In addition, three erythroid cell
lines unresponsive to EP (including clone 745 MEL cells)
were found to have only the 63 Kd band cross-linked to
‘25IEP However, Sasaki et aO#{176}reported identical 100 Kd
and 85 Kd subunits of the receptor for EP in EP-responsive
TSA8 erythroid cells, FVA cells, mouse fetal liver cells, and
spleens from anemic mice. Tojo et al reported the EP
receptor as bands of I 10 and 95 Kd in fetal mouse liver,’6
while Mayeux et al reported two proteins of molecular mass
of 94 and 78 Kd in fetal rat liver8; and Pekonen et al
described two proteins of 85 Kd and 41 Kd in human fetal
liver.9
A B C D E F
68
Fig 6. Cross-linking of ‘�l-EP to human erythroid colony-
forming cells and FVA cells. 1�l-EP was bound to the cells andcross-linked as described in Materials and Methods. Nonspecificbinding was determined in the presence of 100 units of unlabeled
EP. lanes A and F. The cells were extracted with detergent and
centrifuged at 1 .000 g. Cell extract. lanes A. C. D, and F; celldebris. B lanes and E. Human erythroid colony-forming cells. lanesA. B. and C; FVA cells. lanes D. E. and F.
In view of the possibility that different subunits of the
receptor might exist in different animal species or different
cell lines, we compared the structure of receptors for EP in
different erythroid cells. In contrast to the finding of oth-
ers,7”#{176}we found that the structure of the receptor for EP in
the MEL cells consisted of the same 100 Kd and 85 Kd
proteins found in FVA cells and all other tissues tested in this
study. In addition, the rat and human receptor were similar
to the mouse receptor, in contrast to reports of lower molecu-
lar weight receptors.8’9 We suspect that the lower molecular
weight proteins cross-linked to ‘25I-EP in these earlier reports
are proteolytic fragments of the larger molecular weight
proteins. In early experiments we observed a band of 70 Kd
when ‘25I-EP was cross-linked to intact MEL cells and the
cross-linked complex was extracted with Triton X-lOO. In
other studies to be published elsewhere, the receptor for EP
in FVA cells was observed to be degraded to a series of lower
molecular weight protein after cross-linking to ‘251-EP to
intact cells. This could be prevented by the addition of
proteinase inhibitors. While the cross-linked receptor is very
insoluble in nonionic detergents, the degradation fragments
are more easily extracted. Earlier reports on the EP receptor
in MEL and fetal rat liver cells used Triton X-I00 extraction
of cross-linked receptor from intact cells. Likewise, an addi-
tional minor band was seen in the soluble extract of cross-
For personal use only.on January 3, 2019. by guest www.bloodjournal.orgFrom

108 SAWYER, KRANTZ. AND SAWADA
linked receptor from human CFU-E but not in the insoluble
fraction (Fig 6). Preparation of plasma membranes from
cells eliminates the need to extract the receptor from the
intact cell and lowers the ratio of receptor to proteinase
activity during the cross-linking reaction.
In contrast to our work and the work of Todokoro et al.,
Sasaki et al showed that the cross-linked bands mostly did
not enter the gel or a minor band was observed at 210 Kd in
the absence of reducing agent during SDS-PAGE.’#{176} Mayeux
et al reported the existence of 94 and 78 Kd proteins
cross-linked to ‘251-EP in fetal rat liver cells and also
suggested that these proteins were subunits of a large molec-
ular weight complex bridged by disulfide bonds.8 In this
study no evidence of disulfide bridging between the 100 and
85 Kd proteins of the EP receptor was detected in any cell or
tissue examined. The observations of large molecular weight
proteins cross-linked to ‘251-EP on SDS-PAGE in the
absence of reducing agents may be due to the incomplete
solubilization of the cross-linked membranes. Mild treat-
ment of ‘251-EP cross-linked to FVA cell membranes and
SDS-PAGE in the absence of reducing agents leads to a
weakly labeled band at 230 Kd and most of the radioactivity
at the top of the running gel and stacking gel. This radioac-
tive material was cut out of an unfixed gel and rerun on
SDS-PAGE in the presence of/3-mercaptoethanol. Only 1%
of this radioactivity was found in the 100 and 85 Kd
cross-linked bands while 99% was noncross-linked ‘251-EP
(data not shown). Mild sonication of the cross-linked mem-
brane in sample buffer for SDS-PAGE containing no reduc-
ing agent converted the higher molecular weight cross-linked
material into the clear 100 and 85 Kd proteins. This strongly
suggested the absence of disulfide bridges between the 100
and 85 Kd proteins.
This study reports the structure of the normal human
erythroid receptor for EP. The human receptor is virtually
identical to the mouse and rat receptor, however a slight
increase in the molecular mass of the lesser protein of the
human receptor compared with mouse and rat may occur.
Other experiments show the very slight difference in migra-
tion of the two proteins of the human receptor compared with
that in FVA cells. We suspect that the lower band of the
receptor for EP may be due to the cleavage of the 100 Kd
band by proteinase. If this is the case, a subtle difference in
the amino acid sequence of the human 100 Kd receptor
protein could lead to a different site of cleavage and therefore
a fragment of slightly larger molecular mass.
It is of interest that the receptor for EP is remarkably
similar in mouse, rat, and human tissue. Furthermore, the
similarity of receptor in erythroid cells and placenta is also of
interest in that the function of the receptor in the placenta is
not proven. This is the first report of the existence of
receptors for EP in placenta. The finding in this laboratory
that ‘25I-EP crossed from maternal circulation to the fetus in
pregnant mice’7 led us to investigate the existence of recep-
tors for EP in the placenta. We think that the placental
receptors for EP may be involved in the transplacental
transfer of ‘251-EP but additional experiments are necessary
to prove the physiological relevance of the transfer of EP to
the fetus and the receptor mediated nature of the transfer.
The nearly identical structure of the receptors in the
normal human erythroid cells and normal placentas com-
pared with the receptors in mouse erythroid cells that
proliferate in response to infection with the Friend virus
indicates that the Friend virus does not stimulate erythroid
development through a gross modification of the structure of
the receptor for EP. The two strains of Friend virus, FVA,
and FVP, lead to different responses when either mice or
bone marrow cells are infected. FVA infection generates
erythroid cells comparable with CFU-E in that the cells are
responsive to EP and respond by undergoing erythroid matu-
ration and growth. In contrast, FVP infection results in a
proliferation of erythroid cells that are apparently undergo-
ing erythroid maturation independently of EP. In culture
these cells do not respond to EP.4”8 It is possible that
autocrine production of EP by FVP cells might be responsi-
ble for this apparent independence of regulation by EP, but
preliminary experiments in our laboratory have failed to
detect production of mRNA coding for EP in FVP cells.
The presence of receptors for EP on MEL and FVP cells
on which EP has no apparent effect suggests that the normal
mechanism by which EP triggers these cells has been altered.
In this regard, we studied internalization of ‘25I-EP by MEL
and FVP cells. ‘251-EP was internalized in these cells in
exactly the same manner as the EP-responsive FVA cells. In
earlier work, the higher affinity receptors on the FVA cells
appeared to be preferentially endocytosed’; however, this
study shows equal endocytosis of higher and lower affinity
receptor since FVP and MEL have only lower affinity
receptor for EP. The presence of higher affinity receptors on
FVA cells and human CFU-E’2 may be responsible for their
capacity to respond to EP with erythroid maturation. How-
ever, the molecular mechanism for this effect is still unknown
since the present study shows that the structure of the
receptor for EP in cells containing either higher and lower
affinity receptor or only affinity receptors is identical in gross
structure.
REFERENCES
1 . Sawyer ST. Krantz SB, Goldwasser E: Binding and receptor
mediated endocytosis of erythropoietin in Friend virus-infected
erythroid cells. J Biol Chem 262:5554, 1987
2. Krantz SB, Goldwasser E: Specific binding oferythropoietin in
spleen cells infected with the anemia strain of Friend virus. Proc
Natl Acad Sci USA 81:7574, 1984
3. Sawyer ST. Krantz SB, Luna J: Identification of the receptor
for erythropoietin by cross-linking to Friend virus-infected erythroid
cells. Proc Natl Acad Sci USA 84:3690, 1987
4. Koury Mi, Sawyer ST, Bondurant MC: Splenic erythroblasts
in anemia-inducing Friend disease: A source of cells for studies oferythropoietin-mediated differentiation. J Cell Physiol 121:526,
1984
5. Sawyer ST, Koury Mi, Bondurant MC: Large-scale procure-
ment of erythropoietin-responsive erythroid cells. Methods Enzymol
147:340, 1987
6. Koury Mi, Bondurant MC, Atkinson JB: Erythropoietin con-
trol of terminal erythroid differentiation: Maintenance of cell viabil-
For personal use only.on January 3, 2019. by guest www.bloodjournal.orgFrom

RECEPTORS FOR ERVTHROPOIETIN 109
ity, production of hemoglobin, and development of the erythrocyte
membrane. Blood Cells 13:217, 1987
7. Todokoro K, Kanazawa S, Amanuma H, Ikawa Y: Specific
binding of erythropoietin to its receptor on responsive mouseerythroleukemia cells. Proc Natl Acad Sci USA 84:4126, 1987
8. Mayeux D, Billat C, Jacquot R: The erythropoietin receptor of
rat erythroid progenitor cells. J Biol Chem 262: 1 3985, 1987
9. Pekonen F, Rosenlof K, Rutanen E-M, Fyhrquist F: Erythro-
poietin binding sites in human foetal tissues. Acta Endocriniol
(Copenh) 116:561, 1987
10. Sasaki R, Yanagawa 5, Hitomi K, Chiba H: Characteriza-tion of erythropoietin receptor of murine erythroid cells. Eur JBiochem 168:43, 1987
1 1 . Sawada K, Krantz SB, Kans J, Dessypris EN, Sawyer ST.
Glick AD, Civin CI: Purification ofhuman erythroid colony-forming
units and demonstration of specific binding of erythropoietin. J Clin
Invest 80:347, 1987
12. Sawada K, Krantz SB, Sawyer ST, Civin CI: Quantitation of
specific binding of erythropoietin to human erythroid colony-
forming cells. J Cell Physiol 137:337, 1988
13. Scatchard G: The attraction of proteins for small molecules
and ions. Ann NY Acad Sci 51:660, 1949
I 4. Sawyer ST, Krantz SB: Transferrin receptor number, synthe-
sis, and endocytosis during erythropoietin-induced maturation of
Friend virus-infected erythroid cells. J Biol Chem 261 :91 87, 1986
I 5. Laemmli UK: Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227:680, 1970
16. Tojo A, Fukamachi H, Kasuga M, Urabe A, Takaku F:
Identification of erythropoietin receptors on fetal liver erythroid
cells. Biochem Biophys Res Commun 148:443, 1987
17. Koury Mi, Bondurant MC, Graber SE, Sawyer ST: Erythro-
poietin messenger RNA levels in developing mice and transfer of
‘25I-erythropoietin by the placenta. J Clin Invest 82:154, 1988
18. Ruscetti 5, Wolff L: Spleen focus-forming virus: Relationship
of an altered envelope gene to the development of a rapid erythroleu-
kemia. Curr Top Microbiol Immunol 112:21, 1984
For personal use only.on January 3, 2019. by guest www.bloodjournal.orgFrom

1989 74: 103-109
ST Sawyer, SB Krantz and K Sawada placentaReceptors for erythropoietin in mouse and human erythroid cells and
http://www.bloodjournal.org/content/74/1/103.full.htmlUpdated information and services can be found at:
Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society of
For personal use only.on January 3, 2019. by guest www.bloodjournal.orgFrom