
Recent Res. Devel. Neurosci., 4 (2013): 89-107 ISBN: 978 ... A4.pdf · in retina grow faster than...
19
Research Signpost 37/661 (2), Fort P.O. Trivandrum-695 023 Kerala, India Original Article Recent Res. Devel. Neurosci., 4 (2013): 89-107 ISBN: 978-81-308-0525-2 4. Body and brain plasticity: Unraveling its principles through blindness Raquel Martínez-Méndez 1,* , Eduardo Martínez-Martínez 1 Horacio Villafán-Monroy 1 , Jessica Alejandra Guzmán-López 1 Alma Lilia Fuentes-Farías 2 , Tania Romo-González 3 Anahí Chavarría-Krauser 4 and Gabriel Gutiérrez-Ospina 1,* 1 Departamento de Biología Celular y Fisiología, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, México D.F., 04510, México; 2 Laboratorio de Ecofisiología, Facultad de Biología, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Michoacán, 58030, México 3 Área de Biología y Salud Integral, Instituto de Investigaciones Biológicas, Universidad Veracruzana Xalapa, Veracruz, 91190, México; 4 Departamento de Medicina Experimental, Facultad de Medicina Universidad Nacional Autónoma de México, México D.F., 06726, México Abstract. Phenotype plasticity is by far the most important property of living beings. In no other organ system plasticity has been the focus of such an intense scrutiny than in the nervous system. This is not surprising since constant neural reorganization is assumed to support, for instance, adaptive behavioral responses, individual differences of behavior and numerous neurological disease states (e.g., the phantom limb). With such a long history of research, the study of neural plasticity has already generated explanatory claims that many conceived as general principles that rule neural plastic responses. Information arising from developmental sensory neurobiology is, however, Correspondence/Reprint request: Dr. Raquel Martínez Méndez, Gabriel Gutierrez-Ospina, Departamento de Biología Celular y Fisiología, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, Ciudad Universitaria, Coyoacán, México D.F., 04510, México. E-mails: [email protected]; [email protected]
Transcript of Recent Res. Devel. Neurosci., 4 (2013): 89-107 ISBN: 978 ... A4.pdf · in retina grow faster than...

Research Signpost
37/661 (2), Fort P.O.
Trivandrum-695 023
Kerala, India
Original Article
Recent Res. Devel. Neurosci., 4 (2013): 89-107 ISBN: 978-81-308-0525-2
4. Body and brain plasticity: Unraveling its
principles through blindness
Raquel Martínez-Méndez1,*, Eduardo Martínez-Martínez1
Horacio Villafán-Monroy1, Jessica Alejandra Guzmán-López1
Alma Lilia Fuentes-Farías2, Tania Romo-González3
Anahí Chavarría-Krauser4 and Gabriel Gutiérrez-Ospina1,* 1Departamento de Biología Celular y Fisiología, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, México D.F., 04510, México; 2Laboratorio de Ecofisiología, Facultad de Biología, Universidad Michoacana de San Nicolás de Hidalgo, Morelia, Michoacán, 58030, México
3Área de Biología y Salud Integral, Instituto de Investigaciones Biológicas, Universidad Veracruzana Xalapa, Veracruz, 91190, México; 4Departamento de Medicina Experimental, Facultad de Medicina
Universidad Nacional Autónoma de México, México D.F., 06726, México
Abstract. Phenotype plasticity is by far the most important property of living beings. In no other organ system plasticity has been the focus of such an intense scrutiny than in the nervous system. This is not surprising since constant neural reorganization is assumed to support, for instance, adaptive behavioral responses, individual differences of behavior and numerous neurological disease states (e.g., the phantom limb). With such a long history of research, the study of neural plasticity has already generated explanatory claims that many conceived as general principles that rule neural plastic responses. Information arising from developmental sensory neurobiology is, however,
Correspondence/Reprint request: Dr. Raquel Martínez Méndez, Gabriel Gutierrez-Ospina, Departamento de
Biología Celular y Fisiología, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de
México, Ciudad Universitaria, Coyoacán, México D.F., 04510, México. E-mails: [email protected];

Raquel Martínez-Méndez et al. 90
challenging some of these conceptions. In this chapter, we comment new evidence
emerging from studies on blindness that clearly shift the way we think about neural
plasticity.
List of abbreviations
A1, primary auditory cortex
IC, inferior colliculus
LGN, lateral geniculate nucleus
LPN, lateral posterior nucleus
S1, primary somatosensory cortex
SC, superior colliculus
V1, primary visual cortex
Introduction
Phenotype plasticity is the most important property of biological systems. It may be defined as the constant reorganization of the structural and functional traits of the organism in response to internal and external drives. Although the entire body of complex organisms is subjected to quotidian reorganization, it is in the nervous system where phenotype plasticity has been extensively investigated. Hence, neurobiologists have long sustained that brain plastic responses follow well defined principles. It is believed, for instance, that neuronal circuits may reconfigure their geometry depending on whether they are being used or not. Under this assumption, used connections are strengthened whereas those less used or unused are weakened or eliminated entirely. Contact strengthening manifests by the preservation of connectivity, the increment of the number of neuronal processes or connections and/or the expansion of the representational area of a set of them. Contact weakening, in contrast, is assumed when the number of connections is reduced, the area covered by a set of them shrinkages or if they are literally wiped off the synaptic repertoire. Both processes are believed to result from competitive interactions prompted by the limited availability of target-derived neurotrophic factors, on one hand, and by the greater or lower probability of competing neuronal elements to fire or not together, on the other.
The example provided above is just one recount of numerous principles
postulated by the neurobiological community to explain the plasticity of the
brain. In spite of the abundance of empirical evidence that seems to support
each of these principles, emerging evidence arising from sensory
developmental neurobiology supports alternative principles on brain
plasticity that may shift significantly the way we conceive it.

Blindness and body/brain plasticity 91
Blindness is a sensory impairment that leads to brain intermodal (also called cross-modal) reorganization. It is presumed that such shifts result from increased levels of use-dependent neuronal activity and assumed that they represent an adaptive response that “seeks” sensory compensation. In this chapter we will discuss evidence that supports otherwise. As we will show, neither intermodal reorganization is necessarily promoted by evoked sensory activity, nor does it have an intrinsic adaptive/compensatory value. We also comment on recent discoveries supporting that blindness is not a purely neurological condition, but that it is best viewed as an alternative phenotype of the whole body. All these concepts oblige us to revise conceptions on brain plasticity and their utility to design restorative neurological protocols.
Experimental models to study the biology of blindness
A great deal of the quality and usefulness of the information gathered on a given phenomenon relies on the experimental model and designs chosen to explore it. This is why in this section we briefly comment on the advantages and limitations of the models instrumented to study the phenomenology and the mechanics of brain plasticity in the blind. With this, we hope to provide the reader with a guide to judge objectively the reaching of the information gathered through these models to understand phenotype plasticity through blindness. The animal models used to describe the phenomenology and to understand the mechanics of blindness are summarized in Table 1. By far, the models most widely used commonly prevent the animals of having visual experience either by suturing their eyelids closed or by rearing them in darkness. Since these models retain the integrity of the visual pathway, they allow the experimenter to evaluate also the effects of visual restoration on the brain’s anatomy and function. A pitfall of both models is that they represent conditions of relative blindness. Indeed, they do not prevent intact retinas to convey instructional information to the central nervous system and eyelid suture does not prevent the sensing of environmental illumination. At best, these models may replicate visual impairments (i.e., relative blindness) associated with cataracts, extreme refractive problems and corneal opacities, but they will never provide information on the consequences of the absolute loss of vision on brain structure and function. Many diseased conditions (e.g., macular degeneration, glaucoma, retinitis pigmentosa) deeply compromise the functional anatomy of the retina leading to its partial or total degeneration. Then, those experimental models in which macular photocoagulation or photo-damage of the trabecular meshwork are used to render animals blind seem the most meaningful to replicate these clinical settings. Although these models have the advantage of producing partial or total blindness effectively while keeping the eyes in their anatomical position, they require the use of larger animals (e.g., monkeys or rabbits), the

Raquel Martínez-Méndez et al. 92
Table 1. Experimental animal models of blindness.
Model
of
blindness
Species Level of
deprivation
Modifications
in the eyes and
optic nerves
Modifications in
the brain Comparable to Ref
Contact lenses
cat
Absence of form stimulation and some reduction of light perception
Not observed Morphological changes in LGN
Cataract, corneal dystrophies
[1]
Eyelid suture
cat, guinea pig, monkey
Absence of form stimulation and reduction of light perception
No gross morphological modifications in retina and optic nerve
No obvious morphological changes in SC or V1, but changes in the auditory responses of the SC. Morphological changes in LGN and changes in the response of visual cortical cells.
Cataract, corneal dystrophies
[1–
4]
Dark reared
chicken, rat, turtle, tilapia, guinea pig, cat, monkey, human
Absence of light and form perception
Enlarged eyes and reduction in the curvature and thickness of the cornea. Dendritic trees in retina grow faster than normal. Larger fiber diameters of optic nerve axons.
Reduction of retinal fibers, enlarged auditory receptive fields and increase in the number of synaptic vesicles per nerve terminal in the SC. Increase in V1 activity and lack of orientation selective responses of neurons. Activation of V1 by tactile stimuli.
Amblyopia, glaucoma, retinopathy, amaurosis
[3,5
–15]
Enucleation
mice, rat, hamster, cat, monkey
Absence of visual and sensory perception from the eyes
Absence of ocular globe and degeneration of optic nerve
Reduction of the volume of the LGN. Changes in neuronal numbers in the V1 and A1. Projections from the IC neurons to the V1. The spontaneous firing rate of V1 neurons is significantly higher. Activation of V1 by tactile and auditive stimuli and expansion of S1 and A1.
Anophthalmia, retinopathy
[16–
27]
Ontological Bathyergidae family
Absence of light and form perception
Eyes reduced in size and covered by skin and fur. Degeneration of optic nerve.
Degeneration of visual structures and expansion of the S1.
Microphthalmia [28–
30]

Blindness and body/brain plasticity 93
Table 1. Continued
(Gnat−/−) Mice Functionally blind
Lack of functional rods
Inappropriate thalamocortical projections.
Nyctalopia, blindness
[31]
H1- ZRDCT-An anophthalmic
Mice
Absence of visual and sensory perception from the eyes
Absence of ocular globe and optic nerve
Decreased neuron number in various visual structures. Greater numbers of projections from the LPN reach V1.
Anophthalmia [32]
Nyx gene (Nob mutant)
Mice Refractive errors
Loss of visual transmission in the ON pathway
Not studied
Complete congenital stationary night blindness (CSNB1)
[33]
need to acquire expensive equipment (e.g., calibrated laser beam sources) and to have specialized training, circumstances all that preclude the sampling of large populations. This aspect becomes important since brain plasticity vary greatly among individuals. Transgenic or genetic animal models in which blindness is expressed as a physical attribute of a phenotype are also frequently used to approach the biology of blindness. Even though these models might provide valuable information on the molecular and cell biological underpinning of blindness, they all share a major drawback with regard to the interpretation of results. Indeed, the underlying genetic condition might influence the way the plastic response of the brain/body unit is organized. Furthermore, since the genetic mutations commonly are not restricted to the eye and may affect other phenotypic traits, we can never be sure whether the visual condition results mainly from ocular and/or extra-ocular factors. Another experimental approach regularly used to study the biology of blindness involves the unilateral or bilateral surgical removal of the eyes. Enucleation is not uncommon in the clinical practice since it is the elective procedure to end the life-threatening risk associated with, for instance, a retinoblastoma. Although undoubtedly this model is rather effective at preventing visual experience, enucleation jeopardizes ocular muscle and lachrymal gland interoception, thus contaminating the interpretation of the results since the absence of neighboring tissues must add up to the observed brain phenotype. Finally, naturally blind animals such as moles are also valuable sources of information on the biology of blindness. These natural models, however, may speak little on the plasticity mechanisms that are engaged when this condition is acquired by sighted animals, especially if blindness is acquired at adult ages. In sum, even though none of the animal models so far developed accommodate the variety of features that blindness imposes on the brain and body, their analyses have revealed some rules and principles that seem to underlie brain and body plasticity following the loss of vision. In the following sections we will revise these principles in more detail.

Raquel Martínez-Méndez et al. 94
Blindness and brain plasticity: Lessons and a decalogue of
principles
Human and animal functional anatomical studies have revealed that the brain of the blind is profoundly reorganized (Figure 1). Indeed, we now know that information other than visual, especially auditory and somatosensory, is processed in former visual cortical [16,21,34–39] and subcortical centers [40]. At the same time, auditory and somatosensory relays increase their representational areas across the nervous system [24,25,27,30,38,41–43]. This clearly is a major plastic rearrangement if compared with the functional anatomy of the normal brain. What then can blindness tell us about the principles governing neural reorganization? It has long been claimed that the expansion of the primary somatosensory
(S1) and auditory (A1) cortices observed in the blind’s brain results from
increments in the amount of evoked neuronal activity [24,27,44]. We have
shown, nonetheless, that levels of spontaneous activity in S1 are reduced in
slices obtained from neonatal enucleated rats by the time S1 enlargement
is actively ongoing ([45]; Figure 2). This feature coincides with a decrease of
glutamate and GABA availability [46], both excitatory neurotransmitters at
early developmental stages [47–49]. In addition, no major differences in
cytochrome oxidase activity or H3 2-deoxyglucose uptake were found in S1
when sighted and enucleated rats were compared along postnatal development
Figure 1. Intermdodal plasticity in the brain of blind individuals. At the cortical level, intermodal reorganization has two basic components. The recruitment of former visual centers (blue) by somatosensory (green) and auditory (purple) inputs is one of them. The other is the expansion of cortical territories dedicated to process somatosensory and auditory information. These shifts do no generate functional redundancies. Instead, they truly reflect the relocation and transfer of somatosensory and auditory functions to the former visual cortex.

Blindness and body/brain plasticity 95
Figure 2. The number and activity of nerve terminals in columns of the rat primary somatosensory cortex (S1) at postnatal day 7 (PD7) is reduced following
enucleation. A. Photomicrograph illustrating the pattern of S1 staining following the incubation of a representative brain section with FM1-43, a dye used to monitor in vivo synaptic vesicle recycling in nerve terminals. The asterisks indicate S1 layer IV columns. Notice that the relative amount and the intensity of staining appear to be reduced in neonatally enucleated, PD7 rats (C) as compared with their age-matched, sighted littermates (B). The quantification of nerve terminals supports these qualitative results (D).
[50]. Also, whisker cauterization did not prevent the enlargement of S1 in sighted
rats [50]. Interestingly, the enlargement of S1 in neonatal enucleated rats occurs
during the first few days of life [50], well before the rat pups explore actively
their surroundings. Hence, the first lesson learned from studying the role of the
spontaneous and evoked activity on brain plasticity in the blind is that the
increased levels of them are not the primary cause of intermodal plasticity. This
must not be surprising because a wealth of experimental evidence has shown that
reduced neuronal activation promotes the rapid remodeling of sensory afferents
[51,52] and may facilitate axonal elongation through the somatosensory pathway
[53,54]. The contrary is true when evoked neuronal activity increases or when
glutamate is released to the extracellular space. In both cases the axon growth
cone collapse, synapse elimination ensues [55–58] and S1 gets smaller [52].
In the blind, it is assumed that the lack of visual information plays a
central role in promoting the expansion of both S1 and A1, as a well as the
reactivation of the former visual cortex by somatosensory and auditory
inputs. We have found, nonetheless, that S1 expansion does not occur if rats
are dark-reared since birth [50]. This suggests that it is not the loss of visual
information what triggers S1 intermodal plasticity, but is the presence or
absence of the eyes what constitutes the key factor for setting on or not
intermodal plasticity. At first glance, this conclusion might be puzzling but it
is not. It is well known that sensory organs provide trophic information to the
brain. During development this information is used by the cerebral cortex to

Raquel Martínez-Méndez et al. 96
map the sensory organs and segregate sensory cortices (nicely reviewed in a
classic book by Purves [59]). Accordingly, neuronal assembles in the
developing retina engage into spontaneous waves of activity [60] that may
inform the brain about the presence or absence of the eyes. When the eyes are
removed or the retina hypotrophies or degenerates before or by eye opening,
the lack of this trophic information might be the primary trigger of
intermodal plasticity. So, the second lesson learned from studying plasticity
of the blind is that the sensory deprivation alone is not sufficient to drive an
intermodal structural plastic response during early development, but it is
necessary to compromise the anatomical integrity of the sensory organ and
of its related sensory pathways. In support of this contention stand the
magnetic resonance imaging studies that have documented that humans with
early onset blindness exhibit a significant reduction of the volume of the
optic nerves, tract and radiation, the dorsal lateral geniculate nucleus (LGN),
the primary visual cortex (V1) and the posterior segment of the corpus
callosum [61,62]. Similar modifications have been reported in the mole rat
[28,29] and in enucleated rodents (mice [17], rats [63] and hamsters [19,64]),
cats [16,20] and monkeys [65]. Also congenital blindness is associated with
decreased neuron number in LGN and fewer projections from this nucleus to
V1 in mice [32].
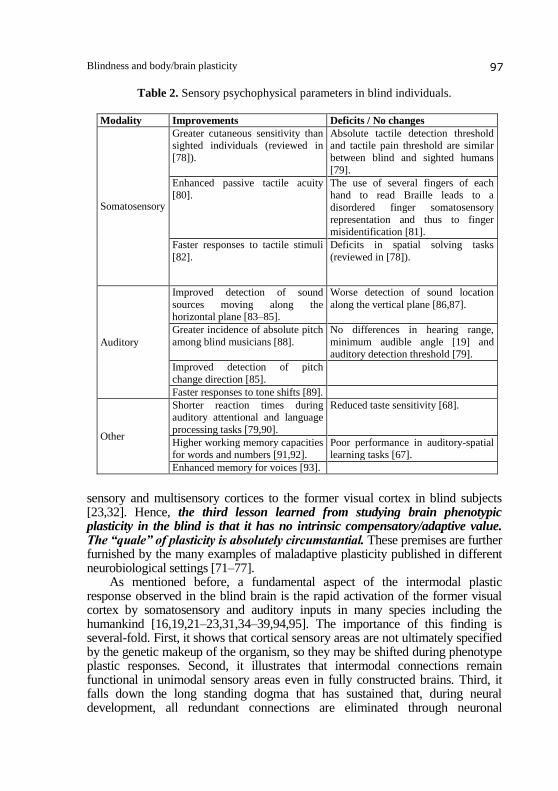
Laymen and neurobiologist alike have known for a long time that blind
people perform better that sighted people on certain tactile and auditory
psychophysical tasks. The blind´s enhanced somatosensory/auditory abilities
are attributed to the recruitment of former visual structures into
somatosensory/auditory processing [37,38,66]. These findings are compatible
with the idea that intermodal plasticity “seeks” sensory compensation.
However, not all sensory shifts lead to improved abilities in the blind [67].
For instance, blind people perform poorly in taste discrimination tasks [68],
as they do when localizing sounds that move along the vertical plane and
during spatial task solving (Table 2). Although the connectivity underlying
this maladaptive/dysfunctional plasticity is unclear, anatomical evidence
suggest that the extensive reorganization of neural circuitry within and
between/among sensory and non-sensory regions of the brain may explain in
part these behavioral impairments in the blind. For instance, abnormal
projections from the inferior colliculus (normally an auditory relay) to the
lateral geniculate nucleus (normally a visual relay) and to the former visual
cortex have been documented in enucleated hamsters [19], anophthalmic
mice [22] and mole rats [69]. Overrepresented connectivity between former
visual areas and associative and speech regions have been shown in blind
humans [70]. Also, studies have traced abnormal afferents from non-visual
thalamic nuclei to former visual cortex and a greater connectivity from other
Blindness and body/brain plasticity 97
Table 2. Sensory psychophysical parameters in blind individuals.
Modality Improvements Deficits / No changes
Somatosensory
Greater cutaneous sensitivity than sighted individuals (reviewed in
[78]).
Absolute tactile detection threshold and tactile pain threshold are similar
between blind and sighted humans
[79].
Enhanced passive tactile acuity [80].
The use of several fingers of each hand to read Braille leads to a
disordered finger somatosensory
representation and thus to finger misidentification [81].
Faster responses to tactile stimuli
[82].
Deficits in spatial solving tasks
(reviewed in [78]).
Auditory
Improved detection of sound
sources moving along the horizontal plane [83–85].
Worse detection of sound location
along the vertical plane [86,87].
Greater incidence of absolute pitch
among blind musicians [88].
No differences in hearing range,
minimum audible angle [19] and auditory detection threshold [79].
Improved detection of pitch
change direction [85].
Faster responses to tone shifts [89].
Other
Shorter reaction times during auditory attentional and language
processing tasks [79,90].
Reduced taste sensitivity [68].
Higher working memory capacities for words and numbers [91,92].
Poor performance in auditory-spatial learning tasks [67].
Enhanced memory for voices [93].
sensory and multisensory cortices to the former visual cortex in blind subjects [23,32]. Hence, the third lesson learned from studying brain phenotypic plasticity in the blind is that it has no intrinsic compensatory/adaptive value. The “quale” of plasticity is absolutely circumstantial. These premises are further furnished by the many examples of maladaptive plasticity published in different neurobiological settings [71–77]. As mentioned before, a fundamental aspect of the intermodal plastic response observed in the blind brain is the rapid activation of the former visual cortex by somatosensory and auditory inputs in many species including the humankind [16,19,21–23,31,34–39,94,95]. The importance of this finding is several-fold. First, it shows that cortical sensory areas are not ultimately specified by the genetic makeup of the organism, so they may be shifted during phenotype plastic responses. Second, it illustrates that intermodal connections remain functional in unimodal sensory areas even in fully constructed brains. Third, it falls down the long standing dogma that has sustained that, during neural development, all redundant connections are eliminated through neuronal

Raquel Martínez-Méndez et al. 98
competition. Fourth, it indicates that not only the sprouting of connectivity but also its redundancy, the synaptic unmasking and the adjustments of functional hierarchies (i.e., balance of different input weights) are central events governing brain plasticity and its outcome. And fifth, it documents that sensory functions can be truly transferred/exchange among sensory cortices even in the mature brain. Therefore, the fourth lesson sustains that the neocortex is not parceled in unimodal and multimodal sensory processing fields but that unimodal areas hide, likely through establishing functional hierarchies, their multisensory nature. This view is further supported by anatomical and physiological data showing multisensory integration properties of unimodal neurons (for a recent review see [96]). “Use it or lose it” is an aphorism that captures a central tenet in developmental neuroscience that predicts the retention, reinforcement or strengthening of neuronal connectivity that is being preferentially used and the weakening or eradication of connectivity that is relatively inactive. Then, it is intriguing that the functional reactivation of former visual areas by somatosensory and auditory inputs is unable to prevent neuronal death and the elimination of connectivity along the reactivated former visual relays [1,16,17,19,20,28,29,32,61–65]. In addition, the “use it or lose it” aphorism has also led to the concept that neuronal circuitry is shaped early in development primarily through synaptic, dendritic and axon collaterals elimination. Studies support, nonetheless, that in the rodent S1 neuronal circuits are constructed by adding precisely the very same neuronal elements in a way that is dependent upon the increased availability of neurotrophic factors [97,98]. Since S1 and its constituent parts increase in size following enucleation, S1 expansion in the enucleated rat brain undoubtedly results from constructive rather than from regressive events or retention of transitory connectivity. Hence, the fifth lesson says that the “use it or lose it” aphorism is not an “all or nothing” rule when plastic responses are taking place in the brain and that constructive events underlie S1 intermodal plasticity in the blind. It is generally believed that neural plasticity is a feature characteristic of the “young maturing brain”. In contrast, the adult brain is commonly thought to be an essentially immutable entity. However, a few years ago, work published by Merzenich and others proved this last assumption wrong [99]. Accordingly, research conducted in blind subjects has shown that the adult brain reorganizes its circuitry within periods of hours of having been deprived of vision [100,101]. So, it seems that “the old monkey readily learns new tricks”, as the proverb sentences. These results lead to the sixth lesson; the mature brain is capable of engaging large-scale plastic responses. As discussed before, a recent publication of our authoring supports that the expansion of S1 in enucleated rats is not promoted by increments in evoked neuronal activity. In the same text, we showed that neonatal enucleation accelerates the development and specification of S1 leading to its early expansion. The accelerated rate of S1 development in the blinded rats is associated with an earlier process of histone deacetylation suggesting that the

Blindness and body/brain plasticity 99
expansion of S1 is subjected to epigenetic modulation and/or caused by epigenetic influences independent of evoked neuronal activity and associated with the absence of the eyes. Interestingly, when S1 deacetylation was inhibited by administering valproic acid through the mother´s milk, the time of S1 specification was normalized and its expansion was prevented. So, the seventh lesson predicts that the epigenetic control of the temporal organization of postnatal ontogenetic events causes brain phenotypic plasticity. The body is the material mean by which the organism interacts with the environment. It also provides a great deal of trophic information that brings and sustains brain growth, organization, maturation, maintenance and possible aging [102]. Then, if the brain shifts in the blind, his body must do it accordingly, and this is precisely what happens. We have shown that adult neonatally enucleated rats have whisker muscles with increased oxidative activity, expanded motor plates and facilitated neuromuscular transmission [103]. Hence, the message of the eighth lesson is that blindness is not a condition that triggers plastic responses exclusively along central structures of the nervous system, but that it also does it in neural elements placed at peripheral locations of the body. If this phenomenon is also observed following other types of neural plastic responses remains to be elucidated. The body, nonetheless, is not only formed by neural elements, but by a variety of distinct cell types arranged in complex tissues and organs. Would it be possible for blindness to shift the anatomy and physiology of the body outside the neural domain? The answer to this question seems to be positive. We have data showing that the epidermis of seven day old, birth enucleated rats and of adult, birth enucleated mice show increased thickness and hypertrophied keratinocytes as compared with their control littermates (Figure 3; [104]). Structural epidermal shifts parallel those observed in the availability of neurotrophin-3 [103] and in the number of Merkel’s mechanoreceptors [104] across the whisker pad. Thus the ninth lesson is that blindness modifies, in the long run, trophic brain-body interdependencies. This principle is further supported by evidence showing an association between retinal and optic nerve micro-structural changes with larger visual fields in the deaf [105]. The influence of blindness on the body and its functions is even more
pervasive and profound. We have conducted studies that show that neonatal
enucleation leads to an enhanced adult inflammatory response [106] to skin-
contact hypersensitivity allergens. This response is in part due to a reduction in
the number of intraepidermal T lymphocytes (Figure 4) that down regulate
contact hypersensitivity responses. Shifts of the immunological abilities in the
blind are not restricted to the skin, since other studies have shown increased
phagocytic activity of liver Kupffer cells [107] and protracted semi-activation of
microglial cells in enucleated rats and mice, several weeks after enucleation
[108]. Hence, the tenth lesson foretells that blindness is not a purely
neurological condition, but a state of the whole body.

Raquel Martínez-Méndez et al. 100
Figure 3. Rats enucleated at birth have increased epidermal thickness. Photomicrographs showing tangential sections of skin samples of the whisker pad in
7 days old sighted (A) and enucleated (B) rats stained with Hematoxylin-Eosin.
Notice that the epidermis is thicker in enucleated rats likely due to the presence of
hypertrophied keratinocytes located in intermediate strata.
Figure 4. Rats enucleated at birth have reduced intra-epidermal dendritic
T lymphocytes when adults. Photomicrographs showing ear epidermal
sheets immunocytochemically stained for CD3 , a specific marker of epidermal
T lymphocytes, obtained from adult sighted (A) and enucleated (B) rats.
Scale bar = 50 µm.
In sum, the evidence commented supports that both the brain and the
body reorganize following visual loss. Since brain reorganization occurs soon
after enucleation and motor and immunological shifts proceed weeks or
months later (see for instance [103]), we suspect that a process of top-down
plasticity underlies the reorganization of the brain/body whole.

Blindness and body/brain plasticity 101
Clinical considerations
Basic science not only has important implications on the way we view
and understand nature, it also impacts the way we design protocols aimed at
solving practical problems. Here, we will speak a bit on these matters with
regard to visual restoration.
Visual deficits constitute a major social and financial burden to health
systems worldwide. The World Health Organization estimates that 39 million
people experience visual impairments and blindness (total loss of vision)
associated with cataract, glaucoma, age-related macular degeneration, corneal
opacities, diabetic retinopathy and trachoma [109,110]. Given the magnitude of
the problem, major efforts have been made to restore vision by using
bioelectronic prostheses, highly specialized surgical procedures (e.g., corneal
replacement or refractive surgery) and cell regenerative therapies. Unfortunately,
visual restoration has proven difficult because, we believe, therapeutic attempts
have been focused primarily at trying to reinstall the eyes function without
considering that blindness leads to a large-scale reorganization of the brain/body
whole. So, in the future, we advise new efforts to restoring visual functions in the
blind to consider the influence of ontogenetic and epigenetic elements on the
desired outcome, while keeping in mind that blindness is a condition that
involves the modulation of the entire body’s plasticity.
Fungal, viral and bacterial skin diseases are frequent ailments suffered by
blind children [111,112]. Although all these infections have been suspected
to be secondary to skin injuries and/or the result of poor hygiene habits, the
observations discussed before support other possibilities, namely, that they
might be consequence of essential differences in the way the cutaneous
immune system is organized in the blind.
Lastly, recent studies have demonstrated the participation of T
lymphocytes in hippocampal learning and memory processes [113]. Sightless
rodents, on the other hand, learn faster and solve better than normal spatial
auditory and tactile recognition tasks (mentioned in [114]). Since activated
microglial cells release cytokines that attract T lymphocytes, the fact that
adult neonatally enucleated rats have semi-activated microglial hippocampal
and septal cells [108] could explain in part the differences in learning abilities
observed between enucleated and control rodents.
Conclusions
The plasticity of the brain, whether adult or still maturing, has long
fascinated neuroscientists. Principles governing it have come and gone over
the years as technical and conceptual advances and new experimental models

Raquel Martínez-Méndez et al. 102
and approaches appear. In this vein, blindness provides with a unique
opportunity to asses several of the principles assumed to underlie brain
plasticity that have endure the passing of time. As commented throughout the
manuscript the study of blindness is making us re-think the role of evoked
activity in brain plasticity, while it introduces the notion that the body itself
provides information that if lost early in life triggers brain plasticity in a way
independent of sensory experiences. Blindness also offers information that
challenges conventional views on whether brain plasticity is always adaptive /
compensatory or if the mammalian neocortex really contains unimodal
sensory fields that are genetically specified during early development and
remain untouched once adult age is reached. The understanding of
epigenetics of brain plasticity by studying it through blindness is fundamental
if one aspires to modulate large scale neural plasticity to restore visual
function. Finally, blindness has taught us that brain plasticity is not only
neural, but that it involves significant shifts across the body outside of the
neurobiological whereabouts. In sum, the blind is not only a blind, it is a
completely new phenotype, an alternative one, in which the body/brain whole
departs from norm providing new ways to understand who we are in
biological terms.
Authors’ contributions
RMM carried out the biochemical studies aimed at evaluating the
neurochemical parameters in the primary somatosensory cortex of neonatally
enucleated rats [46]. She also participated during the drafting and production
of the final version of the manuscript.
EMM carried out the experiments that revealed neuromuscular and skin
plasticity in neonatally enucleated rats ([103,104]; Figure 3).
HVM performed the experiments that documented deviant immune skin
response in neonatally enucleated mice ([106]; Figure 4).
JAGL performed the experiments that revealed reduced levels of
synaptic activity in brain slices of neonatally enucleated rats ([45]; Figure 2).
ALFF drafted the manuscript, elaborated Figure 1 and participated in the
final editing and production of the manuscript.
TRG/ACK designed and conducted the studies that documented
protracted activation of microglial cells in neonatally enucleated mice
([108]).
GGO drafted, edited and produced the final version of the manuscript.
All authors read and approved the final manuscript. Similarly, all authors
declare to have no conflict of interest of any sort with regard to the data and
concepts discussed here.

Blindness and body/brain plasticity 103
Acknowledgements
The authors thank Patricia Padilla Cortés, Jesus Ramírez Santos and
Marcela Palomero Rivero for technical support, and Dr. Gabriel Manjarrez
Gutiérrez for permitting access to HPLC equipment. We are also indebted to
Raymundo Reyes, Ivonne Mora, Martha Carrasco and Dra. Marta Elinos for
providing administrative support. This work was financed by grants from the
Consejo Nacional de Ciencia y Tecnología (38615N and P45872-M),
Dirección General de Asuntos del Personal Académico, Universidad
Nacional Autónoma de México (PAPIIT IX232604). RMM was fellow of the
Consejo Nacional de Ciencia y Tecnología and Ph.D. student of the Programa
de Posgrado en Ciencias Biomédicas, Universidad Nacional Autónoma de
México.
References
1. Wiesel, T.N., and Hubel, D.H. 1963, J Neurophysiol, 26, 978.
2. Withington, D.J. 1992, Neurosci Lett, 136, 153.
3. Guyton, D.L., Greene, P.R., and Scholz, R.T. 1989, Invest Ophthalmol Vis Sci,
30, 761.
4. Wiesel, T.N., and Hubel, D.H. 1965, J Neurophysiol, 28, 1029.
5. Jenkins, R.L., Ivey, W.D., McDaniel, G.R., and Albert, R.A. 1979, Poult Sci,
58, 55.
6. Gottlieb, M.D., Fugate-Wentzek, L.A., and Wallman, J. 1987, Invest Ophthalmol
Vis Sci, 28, 1225.
7. Lee, E.J., Merwine, D.K., Mann, L.B., and Grzywacz, N.M. 2005, Brain Res,
1060, 40.
8. Fukui, Y., Hayasaka, S., Bedi, K.S., Ozaki, H.S., and Takeuchi, Y. 1991, J Anat,
174, 37.
9. Zeutzius, I., Probst, W., and Rahmann, H. 1984, Exp Biol, 43, 87.
10. Prichard, J.R., Armacanqui, H.S., Benca, R.M., and Behan, M. 2007, Anat Rec
(Hoboken), 290, 341.
11. Withington-Wray, D.J., Binns, K.E., and Keating, M.J. 1990, Eur J Neurosci,
2, 682.
12. Cynader, M., Berman, N., and Hein, A. 1976, Exp Brain Res, 25, 139.
13. Papadopoulos, G.C. 1993, J Hirnforsch, 34, 539.
14. Pitskel, N.B., Merabet, L.B., Ramos-Estebanez, C., Kauffman, T., and Pascual-
Leone, A. 2007, Neuroreport, 18, 1703.
15. Merabet, L.B., Hamilton, R., Schlaug, G., Swisher, J.D., Kiriakopoulos, E.T.,
Pitskel, N.B., Kauffman, T., and Pascual-Leone, A. 2008, PLoS One, 3, e3046.
16. Yaka, R., Yinon, U., and Wollberg, Z. 1999, Eur J Neurosci, 11, 1301.
17. Heumann, D., and Rabinowicz, T. 1980, Exp Brain Res, 38, 75.
18. Heumann, D., and Rabinowicz, T. 1982, Exp Brain Res, 46, 99.

Raquel Martínez-Méndez et al. 104
19. Izraeli, R., Koay, G., Lamish, M., Heicklen-Klein, A.J., Heffner, H.E., Heffner,
R.S., and Wollberg, Z. 2002, Eur J Neurosci, 15, 693.
20. Berman, N.E. 1991, Brain Res Dev Brain Res, 63, 163.
21. Newton, J.R., Sikes, R.W., and Skavenski, A.A. 2002, Exp Brain Res, 144, 423.
22. Laemle, L.K., Strominger, N.L., and Carpenter, D.O. 2006, Neurosci Lett,
396, 108.
23. Karlen, S.J., Kahn, D.M., and Krubitzer, L. 2006, Neuroscience, 142, 843.
24. Rauschecker, J.P., Tian, B., Korte, M., and Egert, U. 1992, Proc Natl Acad Sci
U S A, 89, 5063.
25. Rauschecker, J.P. 1995, Trends Neurosci, 18, 36.
26. Toldi, J., Rojik, I., and Feher, O. 1994, Neuroscience, 62, 105.
27. Bronchti, G., Schonenberger, N., Welker, E., and Van der Loos, H. 1992,
Neuroreport, 3, 489.
28. Bronchti, G., Rado, R., Terkel, J., and Wollberg, Z. 1991, Brain Res Dev Brain
Res, 58, 159.
29. Cooper, H.M., Herbin, M., and Nevo, E. 1993, J Comp Neurol, 328, 313.
30. Catania, K.C., and Remple, M.S. 2002, Proc Natl Acad Sci U S A, 99, 5692.
31. Larsen, D.D., Luu, J.D., Burns, M.E., and Krubitzer, L. 2009, Front Neuroanat,
3, 30.
32. Godement, P., Saillour, P., and Imbert, M. 1979, Neurosci Lett, 13, 271.
33. Pardue, M.T., Faulkner, A.E., Fernandes, A., Yin, H., Schaeffel, F., Williams,
R.W., Pozdeyev, N., and Iuvone, P.M. 2008, Invest Ophthalmol Vis Sci, 49, 706.
34. Bronchti, G., Heil, P., Sadka, R., Hess, A., Scheich, H., and Wollberg, Z. 2002,
Eur J Neurosci, 16, 311.
35. Kupers, R., Beaulieu-Lefebvre, M., Schneider, F.C., Kassuba, T., Paulson, O.B.,
Siebner, H.R., and Ptito, M. 2011, Neuropsychologia, 49, 2037.
36. Gougoux, F., Zatorre, R.J., Lassonde, M., Voss, P., and Lepore, F. 2005, PLoS
Biol, 3, e27.
37. Sadato, N., Pascual-Leone, A., Grafman, J., Ibanez, V., Deiber, M.P., Dold, G.,
and Hallett, M. 1996, Nature, 380, 526.
38. Burton, H., Sinclair, R.J., and McLaren, D.G. 2004, Hum Brain Mapp, 23, 210.
39. Bedny, M., Pascual-Leone, A., Dodell-Feder, D., Fedorenko, E., and Saxe, R.
2011, Proc Natl Acad Sci U S A, 108, 4429.
40. Bronchti, G., Heil, P., Scheich, H., and Wollberg, Z. 1989, J Comp Neurol,
284, 253.
41. Necker, R., Rehkamper, G., and Nevo, E. 1992, Neuroreport, 3, 505.
42. Mann, M.D., Rehkamper, G., Reinke, H., Frahm, H.D., Necker, R., and Nevo, E.
1997, J Hirnforsch, 38, 47.
43. Elbert, T., Sterr, A., Rockstroh, B., Pantev, C., Muller, M.M., and Taub, E. 2002,
J Neurosci, 22, 9941.
44. Zheng, D., and Purves, D. 1995, Proc Natl Acad Sci U S A, 92, 1802.
45. Guzmán-López, J.A. 2004, B.Sc. Thesis.
46. Martínez Méndez, R. 2008, B.Sc. Thesis.
47. Gao, X.B., and van den Pol, A.N. 2001, Journal of neurophysiology, 85, 425.
48. Cherubini, E., Gaiarsa, J.L., and Ben-Ari, Y. 1991, Trends Neurosci, 14, 515.

Blindness and body/brain plasticity 105
49. Ben-Ari, Y. 2002, Nature reviews. Neuroscience, 3, 728.
50. Fetter-Pruneda, I., Geovannini-Acuña, H., Santiago, C., Ibarrarán-Viniegra, A.S.,
Martínez-Martínez, E., Sandoval-Velasco, M., Uribe-Figueroa, L., Padilla-
Cortés, P., Mercado-Célis, G., and Gutiérrez-Ospina, G. 2013, PLoS One, 8,
e54940.
51. Antonini, A., and Stryker, M.P. 1996, J Comp Neurol, 369, 64.
52. Penschuck, S., Giorgetta, O., and Fritschy, J.M. 1999, Brain Res Dev Brain Res,
112, 117.
53. Jensen, K.F., and Killackey, H.P. 1987, J Neurosci, 7, 3529.
54. Lee, L.-J., Iwasato, T., Itohara, S., and Erzurumlu, R.S. 2005, J Comp Neurol,
485, 280.
55. Fields, R.D., Neale, E.A., and Nelson, P.G. 1990, J Neurosci, 10, 2950.
56. McCobb, D.P., and Kater, S.B. 1988, Dev Biol, 130, 599.
57. Cohan, C.S., and Kater, S.B. 1986, Science, 232, 1638.
58. Chang, S., and De Camilli, P. 2001, Nat Neurosci, 4, 787.
59. Purves, D. 1988, Body and Brain: A Trophic Theory of Neural Connections,
Harvard University Press, Cambridge.
60. Meister, M., Wong, R.O., Baylor, D.A., and Shatz, C.J. 1991, Science
(New York, N.Y.), 252, 939.
61. Pan, W.J., Wu, G., Li, C.X., Lin, F., Sun, J., and Lei, H. 2007, Neuroimage,
37, 212.
62. Ptito, M., Schneider, F.C., Paulson, O.B., and Kupers, R. 2008, Exp Brain Res,
187, 41.
63. Fukuda, Y., and Hsiao, C.F. 1984, Brain Res, 301, 13.
64. Trevelyan, A.J., and Thompson, I.D. 1995, Vis Neurosci, 12, 971.
65. Dehay, C., Giroud, P., Berland, M., Killackey, H., and Kennedy, H. 1996, J
Comp Neurol, 367, 70.
66. Cohen, L.G., Celnik, P., Pascual-Leone, A., Corwell, B., Falz, L., Dambrosia, J.,
Honda, M., Sadato, N., Gerloff, C., Catala, M.D., and Hallett, M. 1997, Nature,
389, 180.
67. Spigelman, M.N. 1969, Canadian journal of psychology, 23, 292.
68. Gagnon, L., Kupers, R., and Ptito, M. 2013, Chem Senses, 38, 509.
69. Doron, N., and Wollberg, Z. 1994, Neuroreport, 5, 2697.
70. Liu, Y., Yu, C., Liang, M., Li, J., Tian, L., Zhou, Y., Qin, W., Li, K., and Jiang,
T. 2007, Brain, 130, 2085.
71. Costigan, M., Scholz, J., and Woolf, C.J. 2009, Annu Rev Neurosci, 32, 1.
72. Giummarra, M.J., Georgiou-Karistianis, N., Nicholls, M.E., Gibson, S.J., Chou,
M., and Bradshaw, J.L. 2011, Clin J Pain, 27, 691.
73. Wang, H., Brozoski, T.J., and Caspary, D.M. 2011, Hear Res, 279, 111.
74. Nava, E., and Roder, B. 2011, Prog Brain Res, 191, 177.
75. Elbert, T., and Rockstroh, B. 2004, Neuroscientist, 10, 129.
76. Voss, P., Gougoux, F., Zatorre, R.J., Lassonde, M., and Lepore, F. 2008,
NeuroImage, 40, 746.
77. Mao, Y.-T., and Pallas, S.L. 2012, J Neurosci, 32, 10338.

Raquel Martínez-Méndez et al. 106
78. Burnstine, T., Greenough, W., and Tees, R. 1984, Early Brain Damage. Volume
1, C. Almli and S. Finger (Eds.), Academic Press, New York, 3.
79. Collignon, O., Renier, L., Bruyer, R., Tranduy, D., and Veraart, C. 2006, Brain
Res, 1075, 175.
80. Goldreich, D., and Kanics, I.M. 2003, J Neurosci, 23, 3439.
81. Sterr, A., Muller, M.M., Elbert, T., Rockstroh, B., Pantev, C., and Taub, E. 1998,
J Neurosci, 18, 4417.
82. Forster, B., Eardley, A.F., and Eimer, M. 2007, Brain Res, 1131, 149.
83. Rauschecker, J.P., and Kniepert, U. 1994, Eur J Neurosci, 6, 149.
84. Lessard, N., Pare, M., Lepore, F., and Lassonde, M. 1998, Nature, 395, 278.
85. Röder, B., Teder-Salejarvi, W., Sterr, A., Rosler, F., Hillyard, S.A., and Neville,
H.J. 1999, Nature, 400, 162.
86. Zwiers, M.P., Van Opstal, A.J., and Cruysberg, J.R. 2001, J Neurosci, 21,
RC142: 1.
87. Lewald, J. 2002, Neuropsychologia, 40, 1868.
88. Hamilton, R.H., Pascual-Leone, A., and Schlaug, G. 2004, Neuroreport, 15, 803.
89. Röder, B., Rosler, F., and Neville, H.J. 1999, Neurosci Lett, 264, 53.
90. Röder Demuth, L., Streb, J. and Rösler, F., B. 2003, Lang Cogn Process, 18, 1.
91. Röder, B., Rosler, F., and Neville, H.J. 2001, Brain Res Cogn Brain Res, 11, 289.
92. Hull, T., and Mason, H. 1995, J Vis Impair Blind, 89, 166.
93. Bull, R., Rathborn, H., and Clifford, B.R. 1983, Perception, 12, 223.
94. Kujala, T., Alho, K., Kekoni, J., Hämäläinen, H., Reinikainen, K., Salonen, O.,
Standertskjöld-Nordenstam, C.G., and Näätänen, R. 1995, Exp Brain Res,
104, 519.
95. Weeks, R., Horwitz, B., Aziz-Sultan, A., Tian, B., Wessinger, C.M., Cohen,
L.G., Hallett, M., and Rauschecker, J.P. 2000, J Neurosci, 20, 2664.
96. Ghazanfar, A.A., and Schroeder, C.E. 2006, Trends in cognitive sciences,
10, 278.
97. Gutiérrez-Ospina, G., Calikoglu, A.S., Ye, P., and D’Ercole, A.J. 1996,
Endocrinology, 137, 5484.
98. Gutiérrez-Ospina, G., Uribe-Querol, E., Sánchez, N., Geovannini, H., Padilla, P.,
and Hernández-Echeagaray, E. 2004, Brain, behavior and evolution, 64, 61.
99. Merzenich, M.M., Kaas, J.H., Wall, J., Nelson, R.J., Sur, M., and Felleman, D.
1983, Neuroscience, 8, 33.
100. Weisser, V., Stilla, R., Peltier, S., Hu, X., and Sathian, K. 2005, Exp Brain Res,
166, 572.
101. Lewald, J. 2007, Neuropsychologia, 45, 1215.
102. Purves, D., Snider, W.D., and Voyvodic, J.T. 1988, Nature, 336, 123.
103. Toscano-Márquez, B., Martínez-Martínez, E., Manjarrez, E., Martínez, L.,
Mendoza-Torreblanca, J., and Gutiérrez-Ospina, G. 2008, Proc Natl Acad Sci
U S A, 105, 15973.
104. Martínez Martínez, E. 2004, B.Sc. Thesis.
105. Codina, C., Pascalis, O., Mody, C., Toomey, P., Rose, J., Gummer, L., and
Buckley, D. 2011, PLoS One, 6, e20417.

Blindness and body/brain plasticity 107
106. Villafán-Monroy, H., and Gutiérrez-Ospina, G. 2006, Sociedad Mexicana de
Bioquimica, 1, MA126.
107. Jansa, P., Urbánek, K., and Riegrová, D. 1993, Cesk Patol, 29, 157.
108. Escalante-Martínez, D., Chavarría, A., Gutiérrez-Ospina, G., and Romo-
González, T. 2009, Revista Medica de la Universidad Veracruzana, 9, 13.
109. Visual impairment and blindness, [http://www.who.int/mediacentre/factsheets/
fs282/en/].
110. Blindness, [http://www.who.int/topics/blindness/en/].
111. Abolfotouh, M.A., and Bahamdan, K. 2000, Ann Saudi Med, 20, 161.
112. Fathy, H., El-Mongy, S., Baker, N.I., Abdel-Azim, Z., and El-Gilany, A. 2004,
East Mediterr Health J, 10, 416.
113. Ziv, Y., Ron, N., Butovsky, O., Landa, G., Sudai, E., Greenberg, N., Cohen, H.,
Kipnis, J., and Schwartz, M. 2006, Nature neuroscience, 9, 268.
114. Théoret, H., Merabet, L., and Pascual-Leone, A. Journal of physiology, Paris,
98, 221.


















