
Reading –Alberts Chapter 8 p. 376-385 –Alberts Chapter 12 p. 561-567 –Lodish Chapter 11 p....
41
• Reading – Alberts Chapter 8 p. 376-385 – Alberts Chapter 12 p. 561-567 – Lodish Chapter 11 p. 426-436 RNA transport
-
Upload
alondra-farrier -
Category
Documents
-
view
227 -
download
0
Transcript of Reading –Alberts Chapter 8 p. 376-385 –Alberts Chapter 12 p. 561-567 –Lodish Chapter 11 p....

• Reading– Alberts Chapter 8 p. 376-385– Alberts Chapter 12 p. 561-567– Lodish Chapter 11 p. 426-436 RNA transport
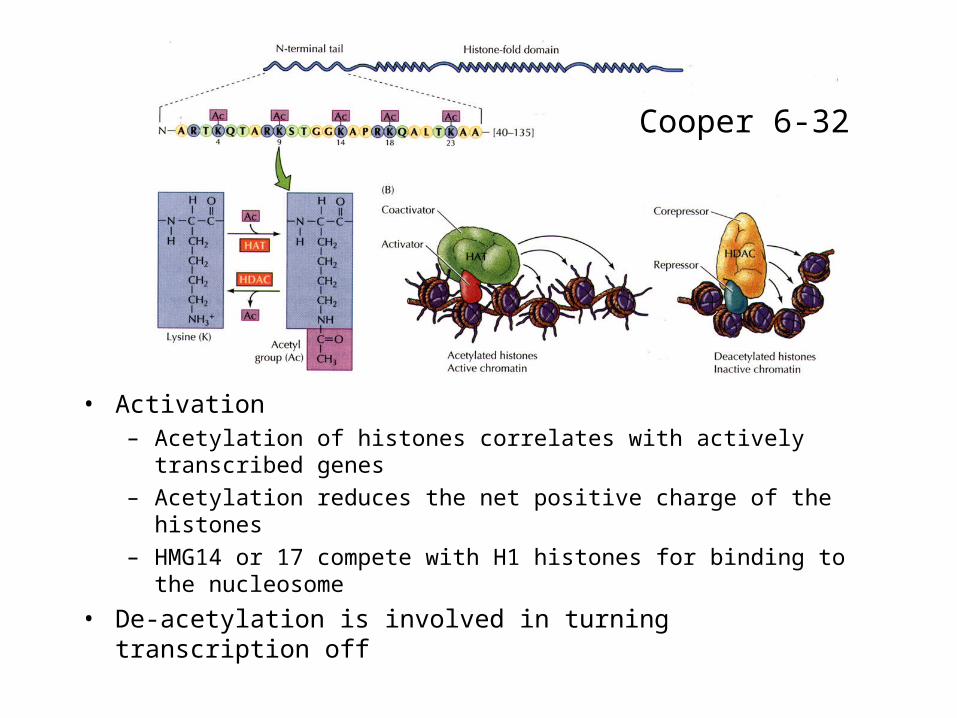
• Activation– Acetylation of histones correlates with actively transcribed genes
– Acetylation reduces the net positive charge of the histones
– HMG14 or 17 compete with H1 histones for binding to the nucleosome
• De-acetylation is involved in turning transcription off
Cooper 6-32

Chromosome organization

• Metaphase chromosome– Chromatids
– Telomeres
– Centromere• Holds sister chromatids
together
• Attachment of kinetochore during mitosis
Alberts 18-15
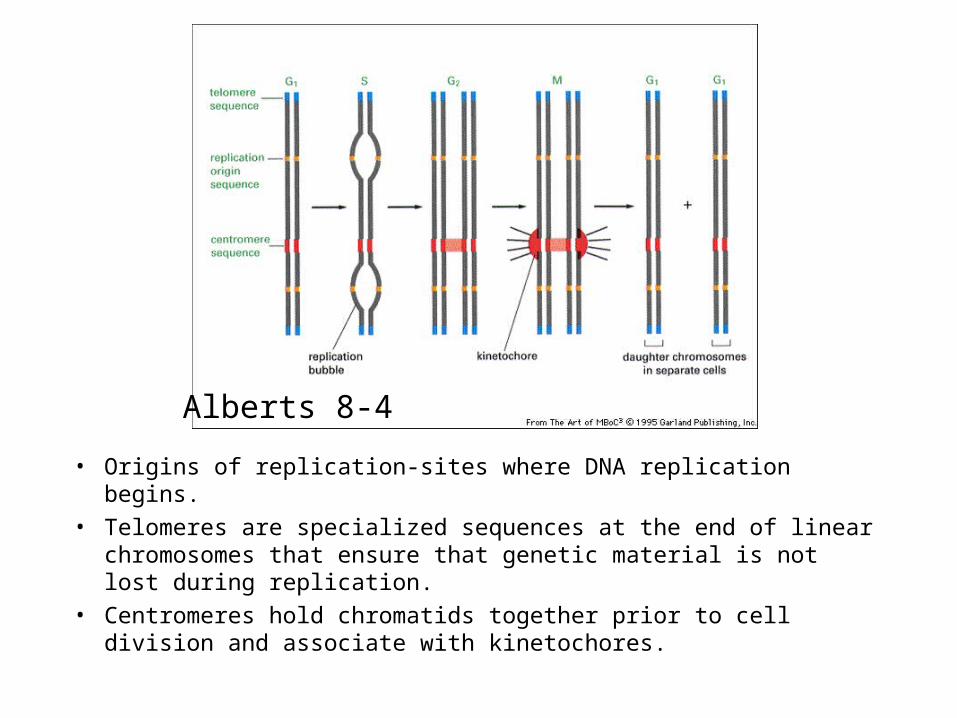
• Origins of replication-sites where DNA replication begins.
• Telomeres are specialized sequences at the end of linear chromosomes that ensure that genetic material is not lost during replication.
• Centromeres hold chromatids together prior to cell division and associate with kinetochores.
Alberts 8-4

• Kinetochore– Attachment site for
spindle microtubules
– DNA/ protein structure
– Required for proper chromosome segregation
Alberts 18-16
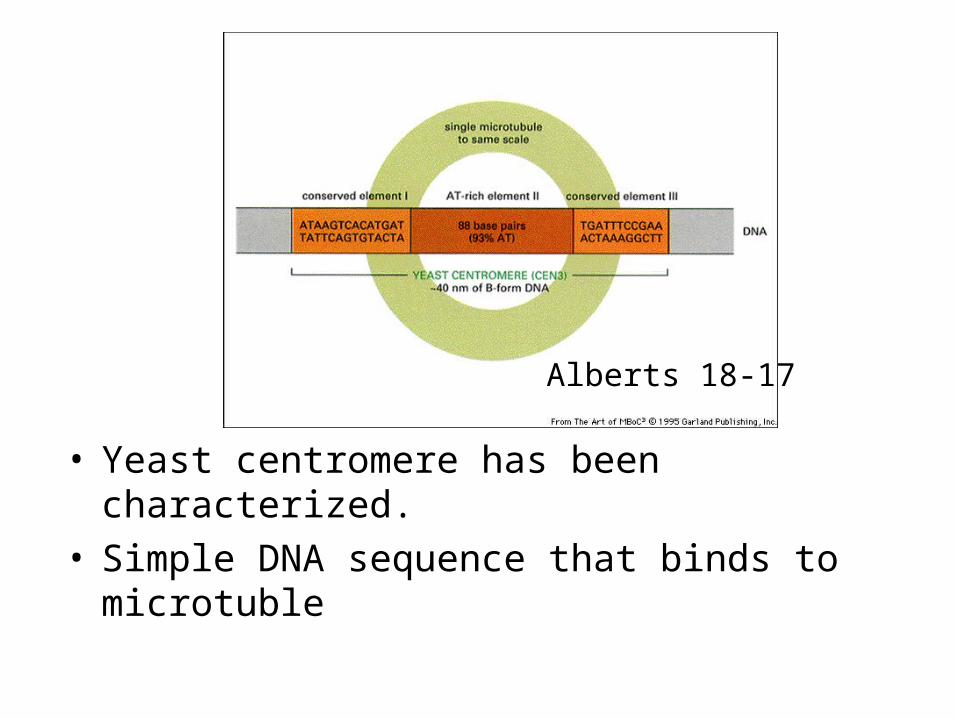
• Yeast centromere has been characterized.• Simple DNA sequence that binds to microtuble
Alberts 18-17
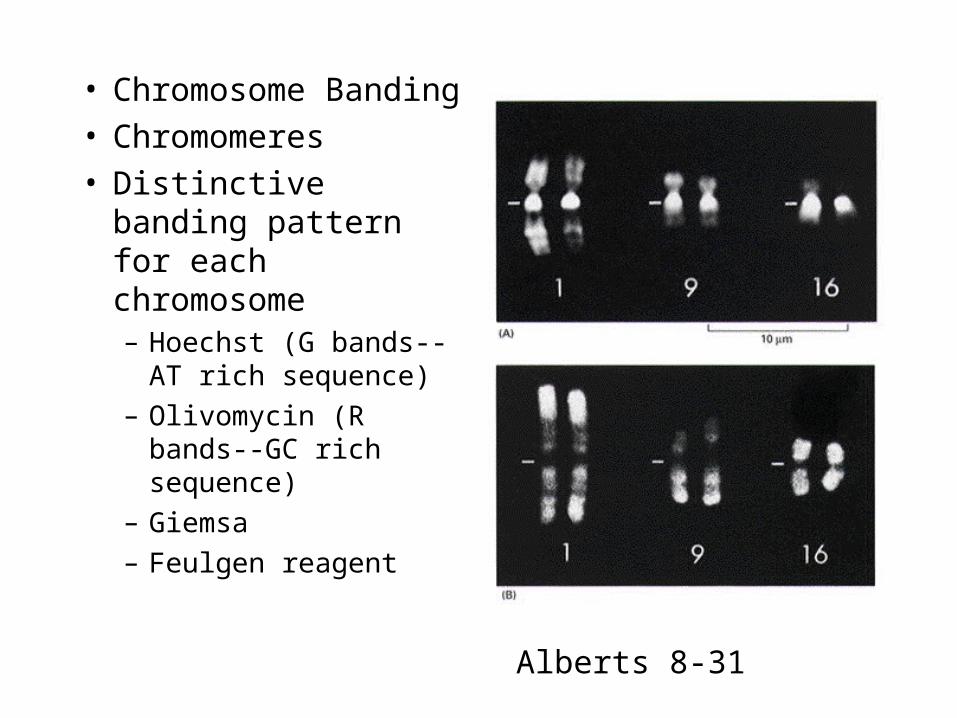
• Chromosome Banding• Chromomeres• Distinctive banding
pattern for each chromosome– Hoechst (G bands--AT
rich sequence)
– Olivomycin (R bands--GC rich sequence)
– Giemsa
– Feulgen reagent
Alberts 8-31
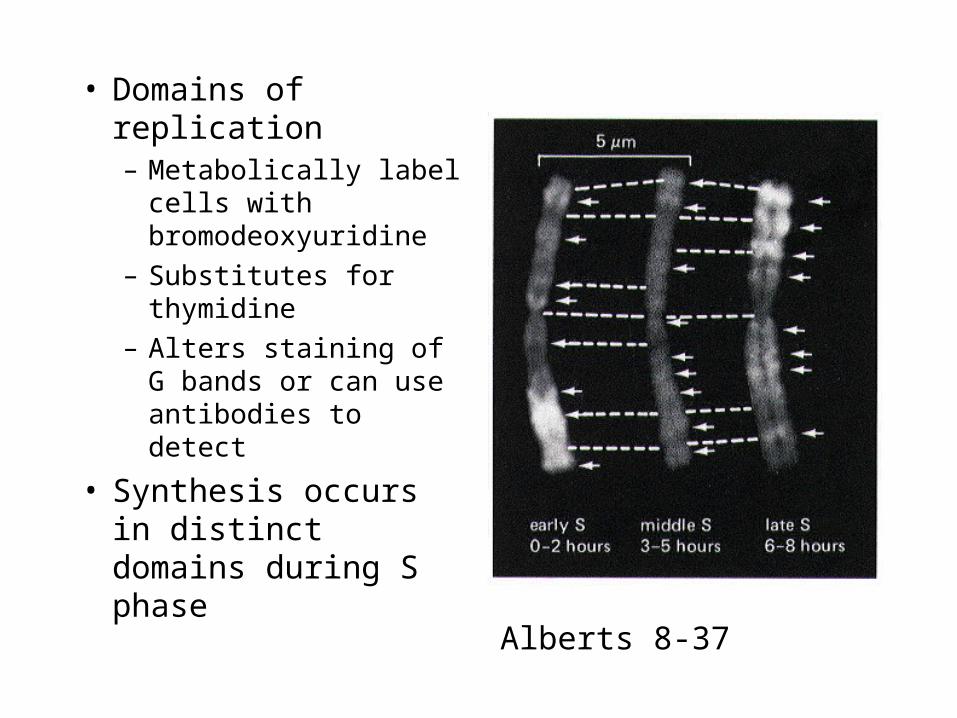
• Domains of replication– Metabolically label
cells with bromodeoxyuridine
– Substitutes for thymidine
– Alters staining of G bands or can use antibodies to detect
• Synthesis occurs in distinct domains during S phase
Alberts 8-37
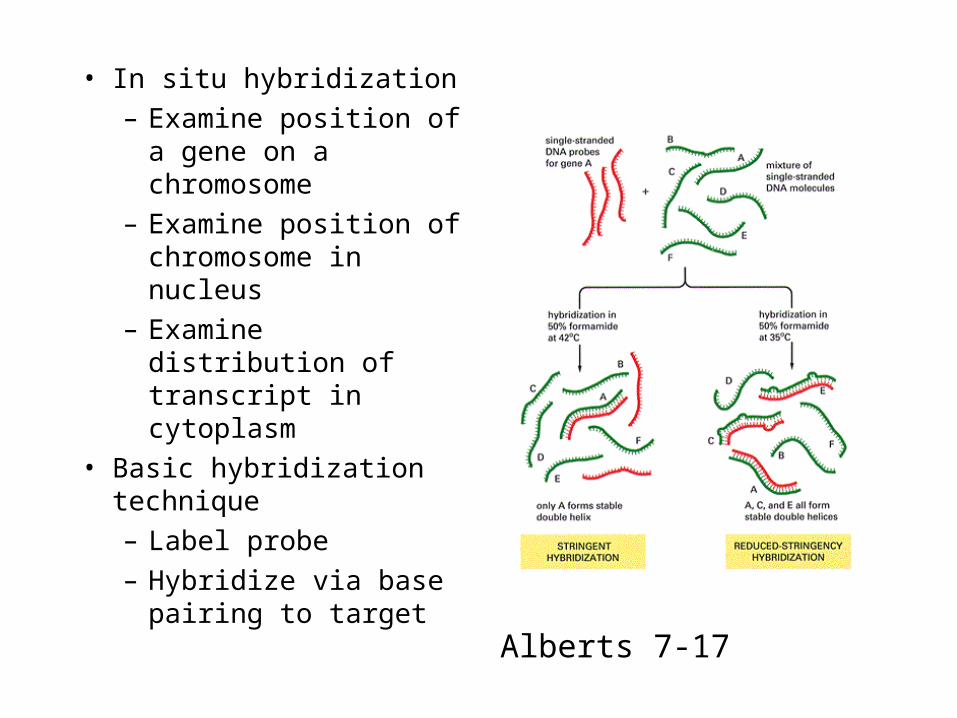
• In situ hybridization
– Examine position of a gene on a chromosome
– Examine position of chromosome in nucleus
– Examine distribution of transcript in cytoplasm
• Basic hybridization technique
– Label probe
– Hybridize via base pairing to target
Alberts 7-17
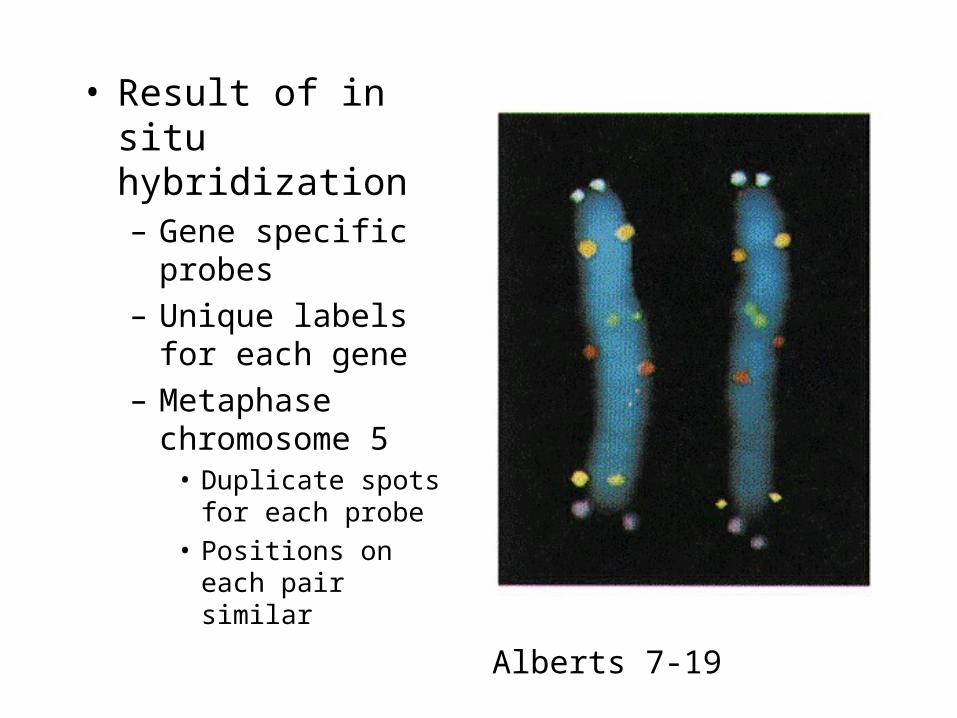
• Result of in situ hybridization– Gene specific probes
– Unique labels for each gene
– Metaphase chromosome 5
• Duplicate spots for each probe
• Positions on each pair similar
Alberts 7-19
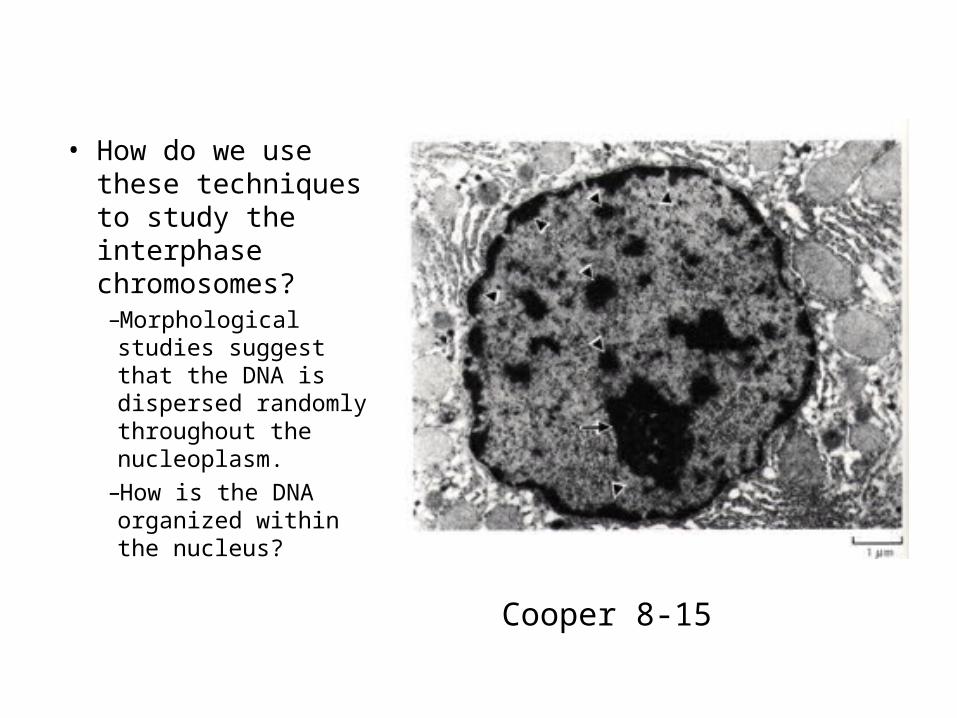
• How do we use these techniques to study the interphase chromosomes?–Morphological studies suggest that the DNA is dispersed randomly throughout the nucleoplasm.
–How is the DNA organized within the nucleus?
Cooper 8-15
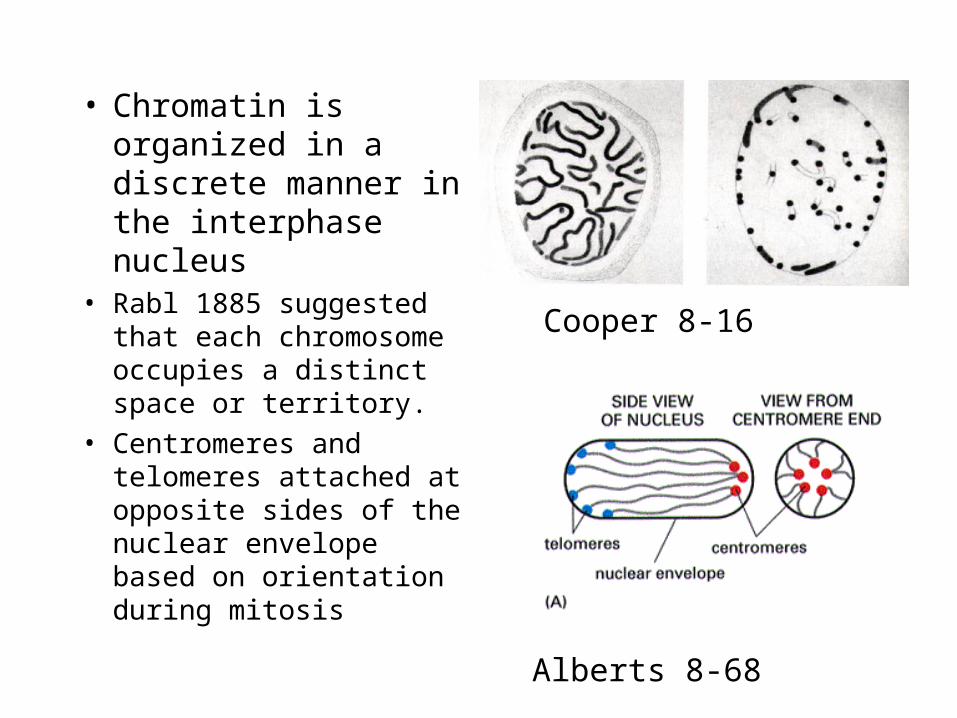
• Chromatin is organized in a discrete manner in the interphase nucleus
• Rabl 1885 suggested that each chromosome occupies a distinct space or territory.
• Centromeres and telomeres attached at opposite sides of the nuclear envelope based on orientation during mitosis
Alberts 8-68
Cooper 8-16

• Rabl observations were confirmed 100 years later by studies of Drosophila embryos– Chromosomal order at
the end of mitosis
– Indication that interphase chromatin is not entangled mass.
– Ordered structure.
Alberts 8-68
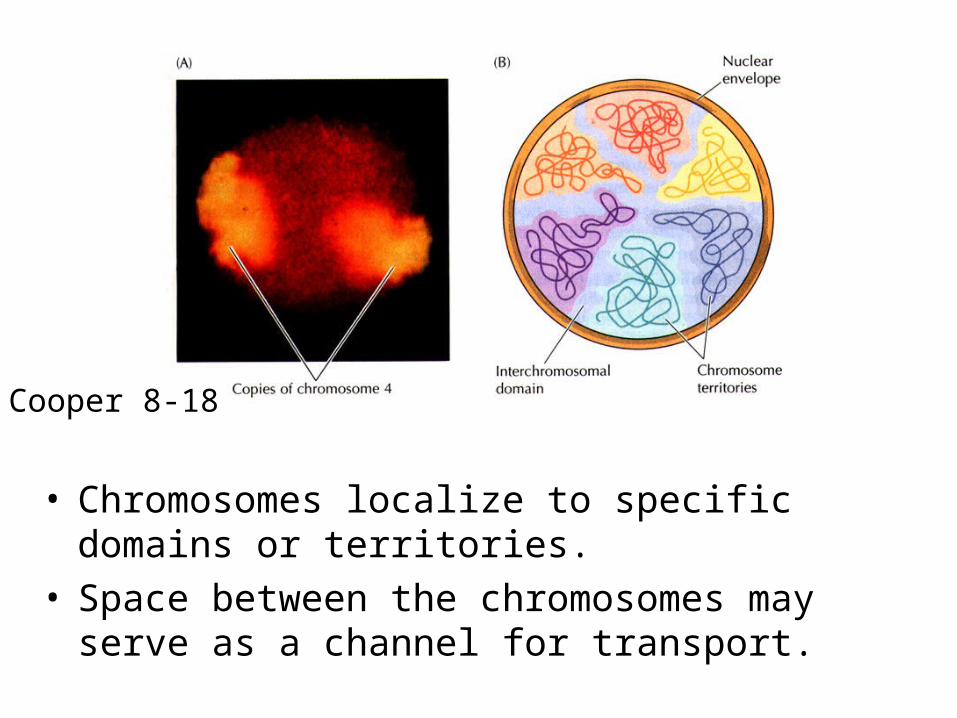
• Chromosomes localize to specific domains or territories.
• Space between the chromosomes may serve as a channel for transport.
Cooper 8-18

Nucleolus
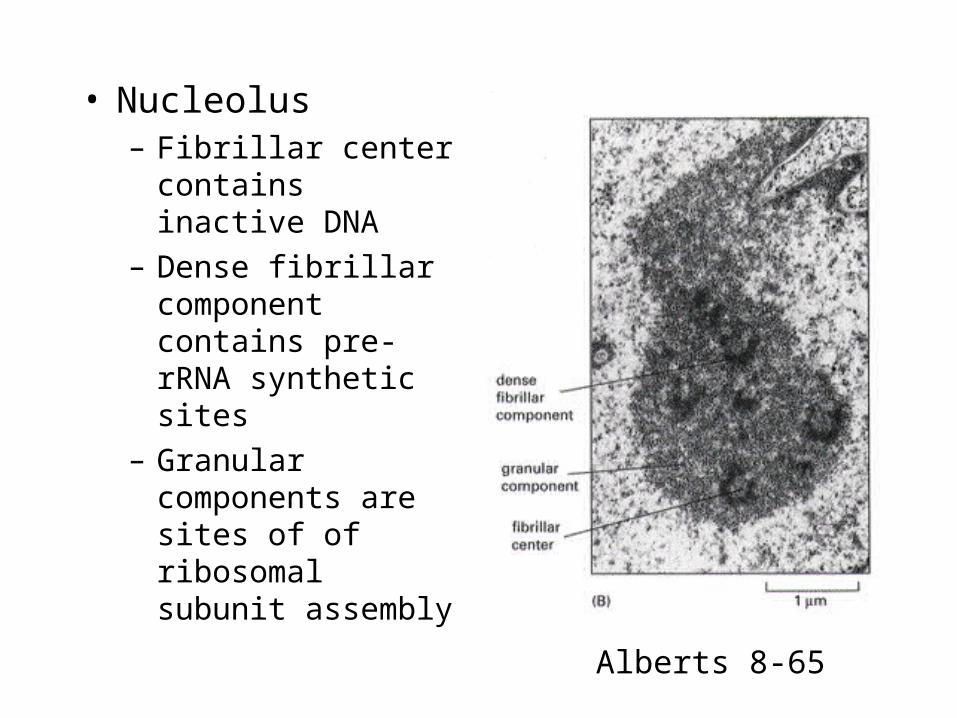
• Nucleolus– Fibrillar center
contains inactive DNA
– Dense fibrillar component contains pre-rRNA synthetic sites
– Granular components are sites of of ribosomal subunit assembly
Alberts 8-65

• The genes encoding the rRNAs are located on 5 different chromosomes
• rRNA genes are tandem repeats encoding a 45S precursor RNA
• Nucleolar organizing centers.– Contain ribosomal
genes
Alberts 8-63
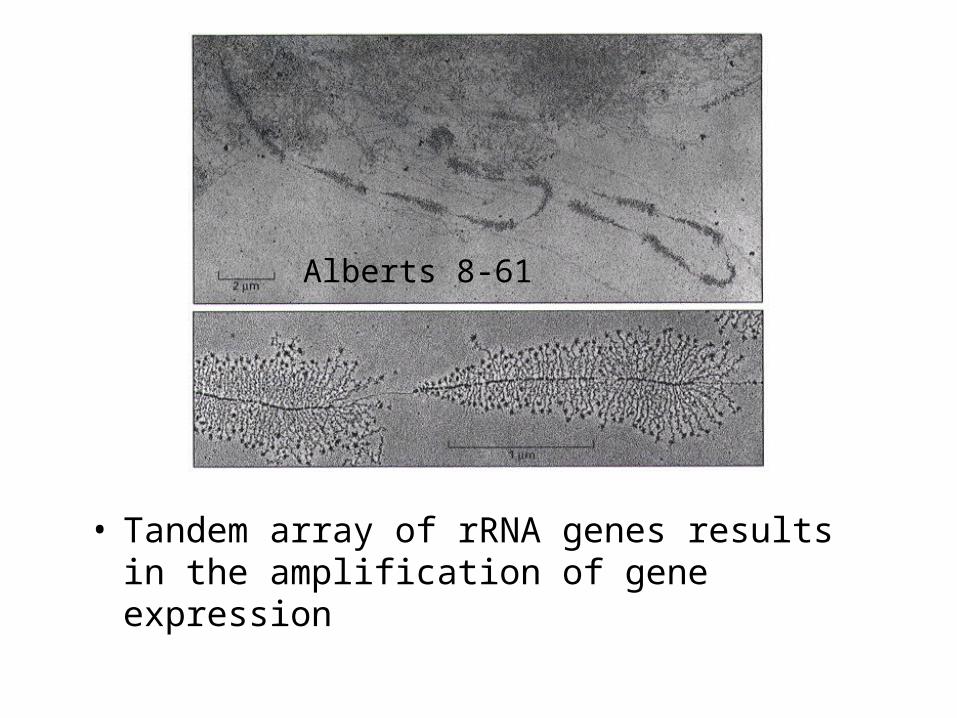
• Tandem array of rRNA genes results in the amplification of gene expression
Alberts 8-61

Proteins of the nucleolus
• RNA polymerase I• Nucleolin
– RNA binding motifs
– Highly phosphorylated
• B23– NO38/numatrin/
nucleo-phosmin
– Highly phosphorylated
• Fibrillarin– Associates with
snoRNPS
• snoRNP– Small nucleolar RNPs
– over 150 different ones
– RNA chaperones, ribonucleases, and guides for rRNA modifications
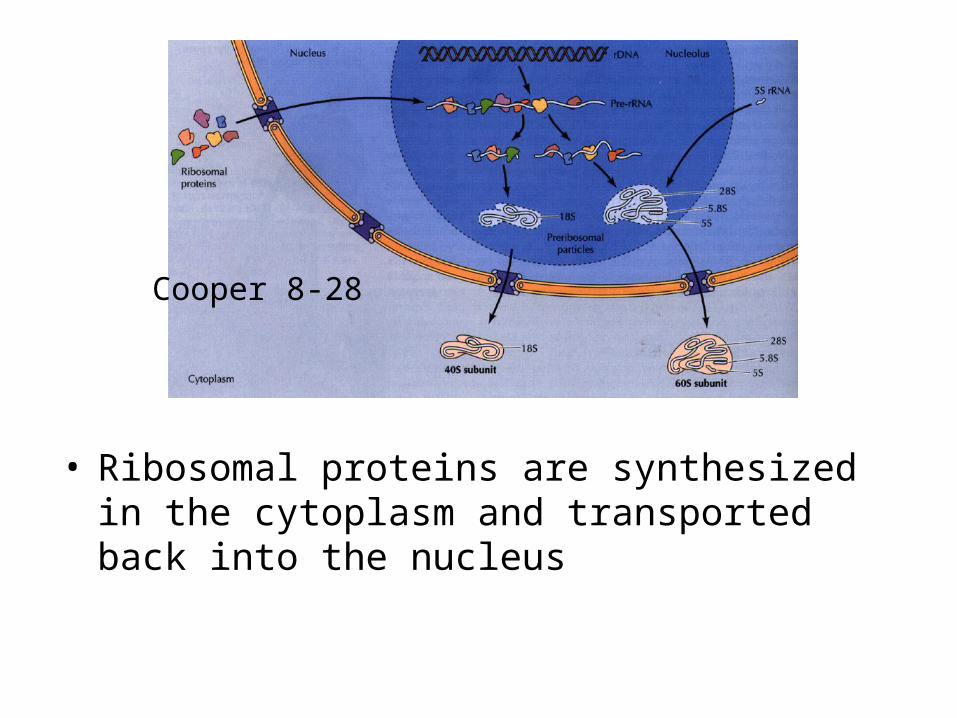
• Ribosomal proteins are synthesized in the cytoplasm and transported back into the nucleus
Cooper 8-28

• The nucleolus is a dynamic structure.– Changes shape in a cell cycle dependent manner.
Alberts 8-67
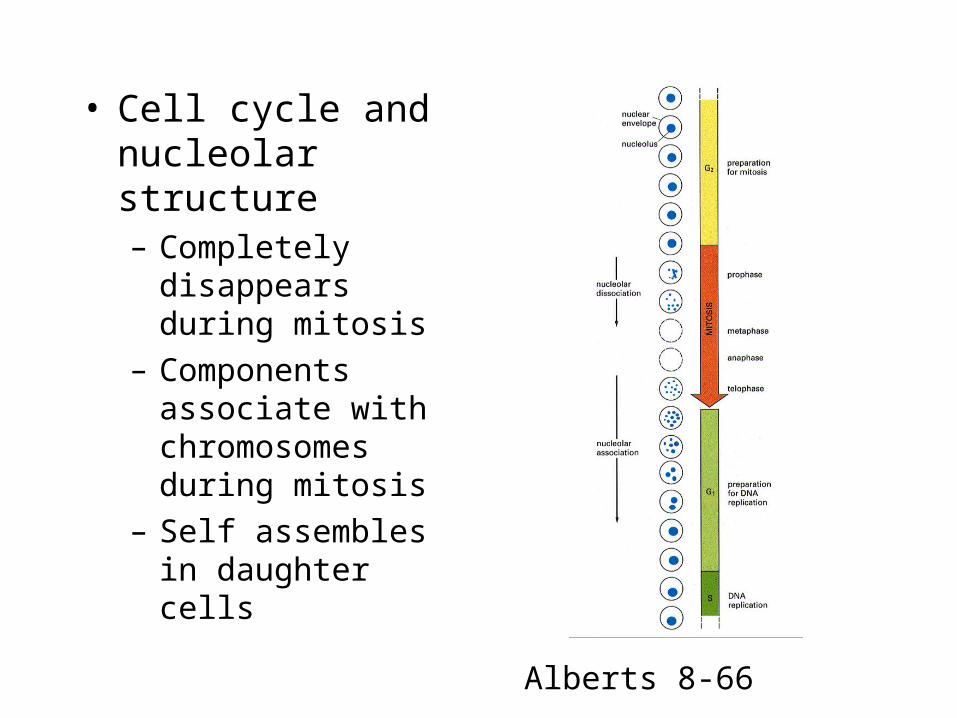
• Cell cycle and nucleolar structure– Completely disappears
during mitosis
– Components associate with chromosomes during mitosis
– Self assembles in daughter cells
Alberts 8-66

Nuclear pores
Alberts 8-72

• Everything that enters or exists the nucleus must pass through the nuclear pores– Passive diffusion
• Small proteins (less than 50 kD
– Active transport• Utilizes energy
Cooper 8-5
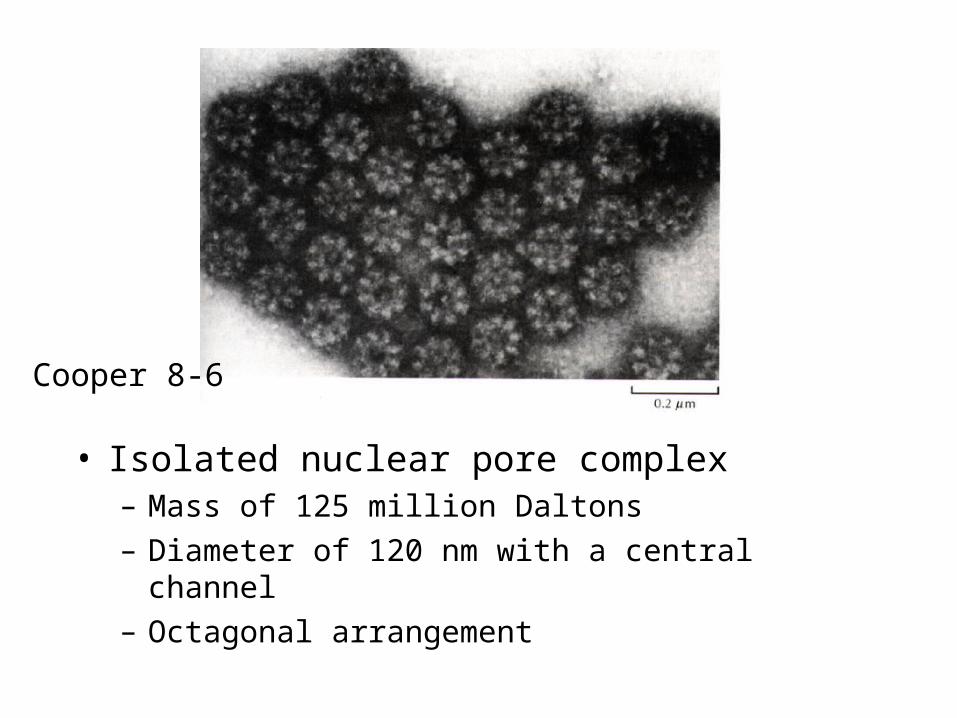
• Isolated nuclear pore complex– Mass of 125 million Daltons
– Diameter of 120 nm with a central channel
– Octagonal arrangement
Cooper 8-6

• Scanning electron micrographs of the nuclear pore complex– Cytoplasmic face
– Nucleoplasmic face
– Nucleoplasmic face with nuclear envelope stripped off.
Lodish 11-28
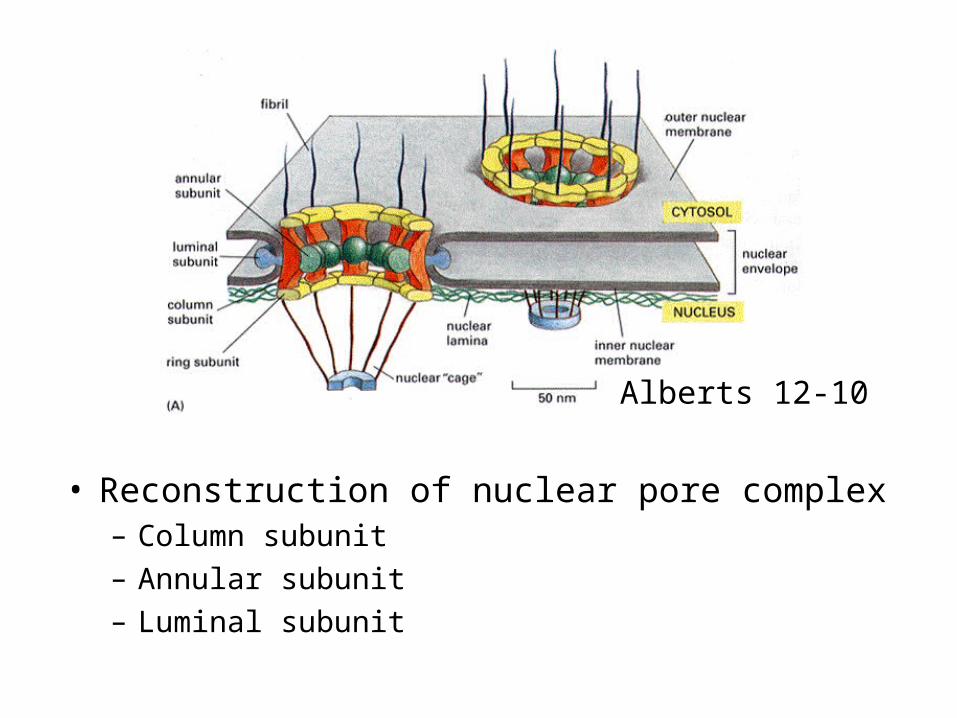
• Reconstruction of nuclear pore complex– Column subunit– Annular subunit – Luminal subunit
Alberts 12-10

• How big is the pore– Experimental results indicate diameter is 9 nm
– EM data my be missing a key component• Central transporter or plug
Alberts 12-12

So how do proteins get through the pore?
• NLS– Nuclear localization signal
• NES– Nuclear exclusion signal
• NRS– Nuclear retention signal
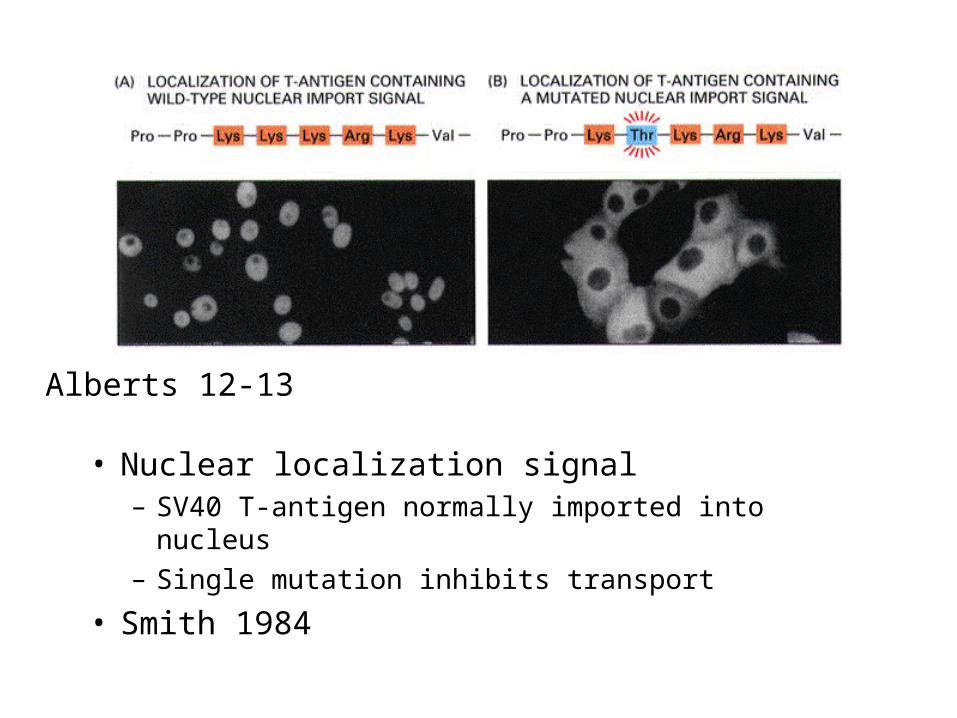
• Nuclear localization signal– SV40 T-antigen normally imported into nucleus
– Single mutation inhibits transport
• Smith 1984
Alberts 12-13
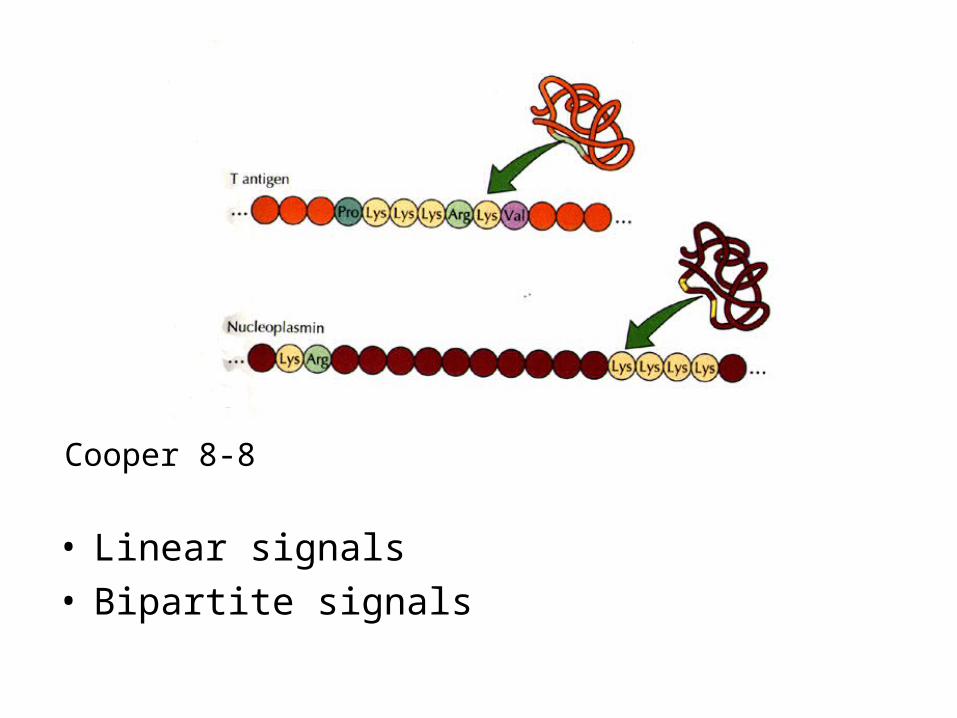
• Linear signals• Bipartite signals
Cooper 8-8

• How do we know this is actually do to transport through the pores?– Experimentally!
– Inject radiolabeled protein and determine what gets into nucleus
– Gold label nuclear targeted protein and examine by EM
Alberts 12-14
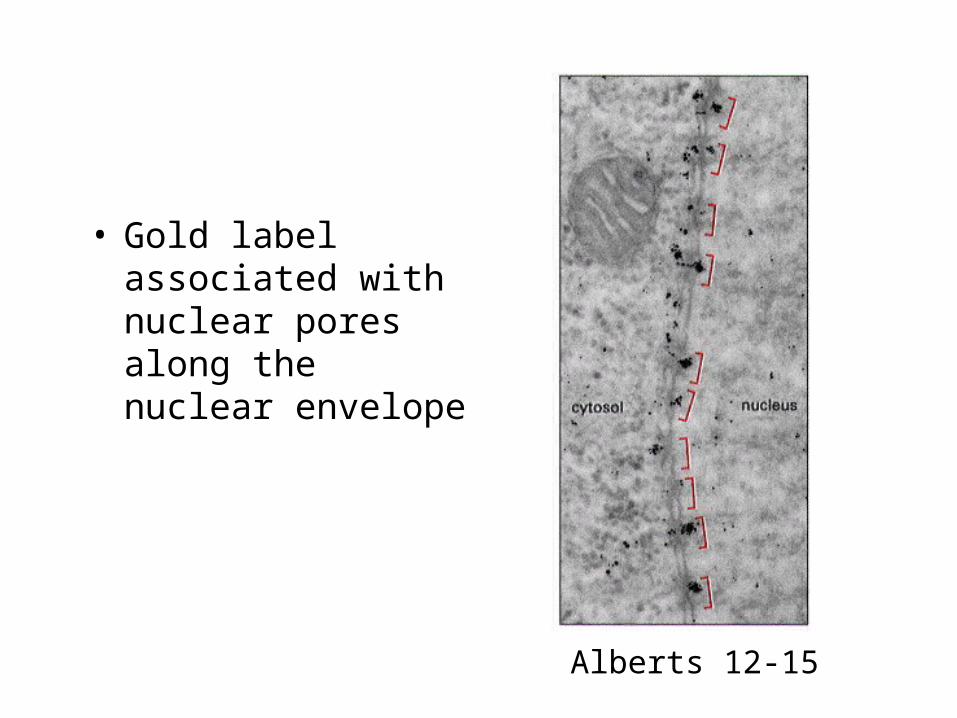
• Gold label associated with nuclear pores along the nuclear envelope
Alberts 12-15
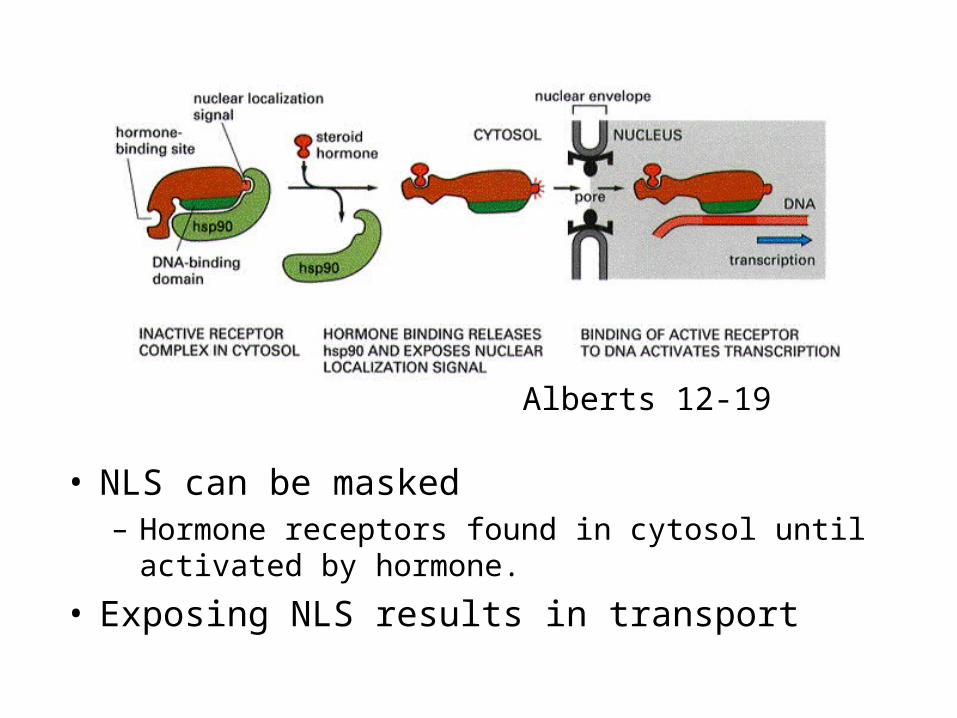
• NLS can be masked– Hormone receptors found in cytosol until activated by
hormone.
• Exposing NLS results in transport
Alberts 12-19

What is happening at the molecular level?
• Step 1: Importins– Import receptors and – Importin bind to NLS– Importin-cargo complex binds to the
cytoplasmic fibrils of the nuclear pores– Does not expend energy

What is happening at the molecular level?
• Step 2: Ran GTPase cycle– Ran relative of Ras is a small GTPase– Activity depends on whether GDP or GTP is
bound– Ran GTP in the nucleus
• Binds importins inside the nucleus
• Releases importins from cargo
– Ran GDP in the cytoplasm
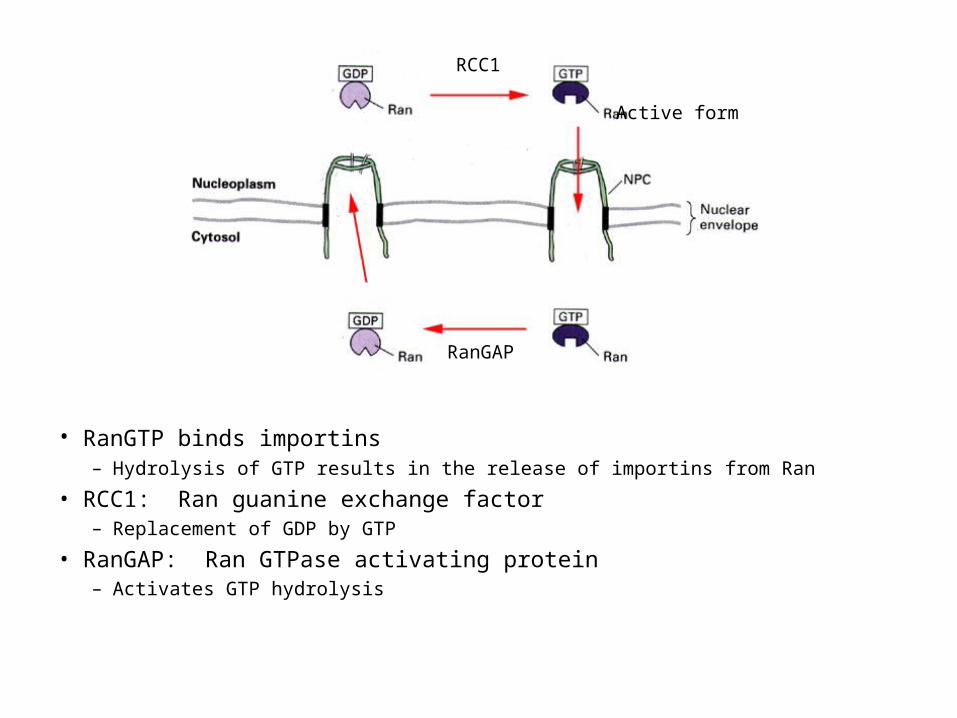
• RanGTP binds importins – Hydrolysis of GTP results in the release of importins from Ran
• RCC1: Ran guanine exchange factor– Replacement of GDP by GTP
• RanGAP: Ran GTPase activating protein– Activates GTP hydrolysis
RCC1
RanGAP
Active form
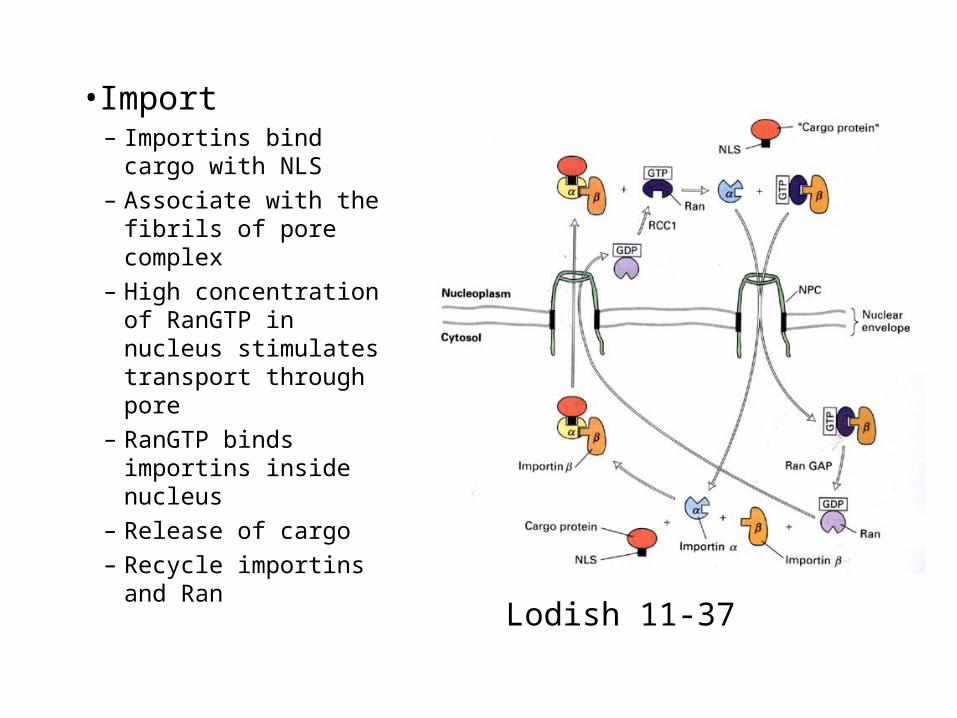
•Import– Importins bind cargo
with NLS
– Associate with the fibrils of pore complex
– High concentration of RanGTP in nucleus stimulates transport through pore
– RanGTP binds importins inside nucleus
– Release of cargo
– Recycle importins and Ran
Lodish 11-37
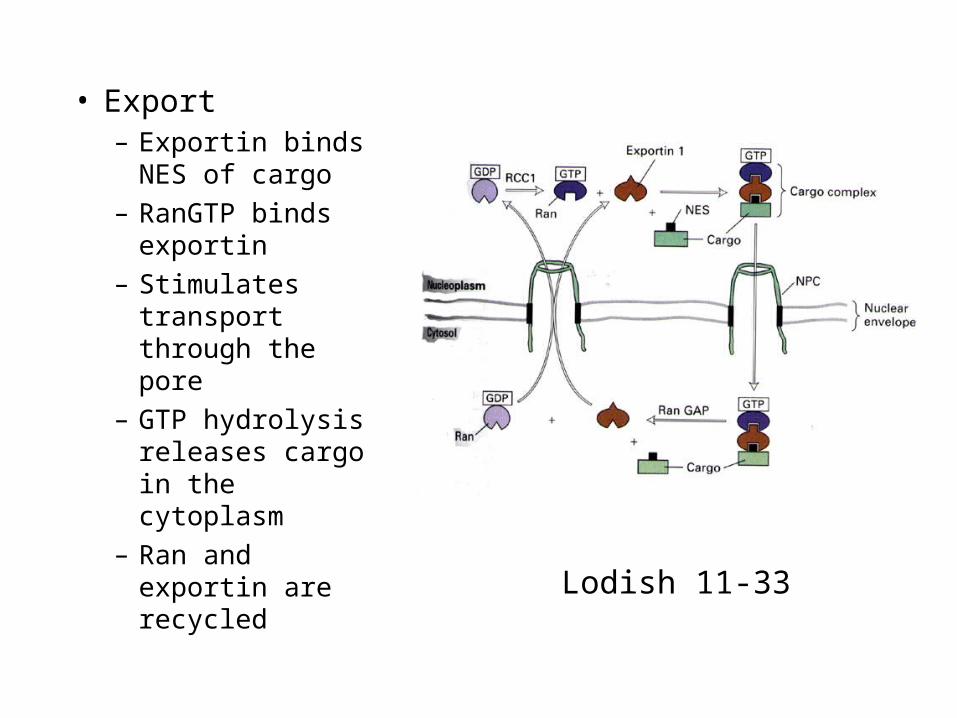
• Export– Exportin binds
NES of cargo
– RanGTP binds exportin
– Stimulates transport through the pore
– GTP hydrolysis releases cargo in the cytoplasm
– Ran and exportin are recycled
Lodish 11-33
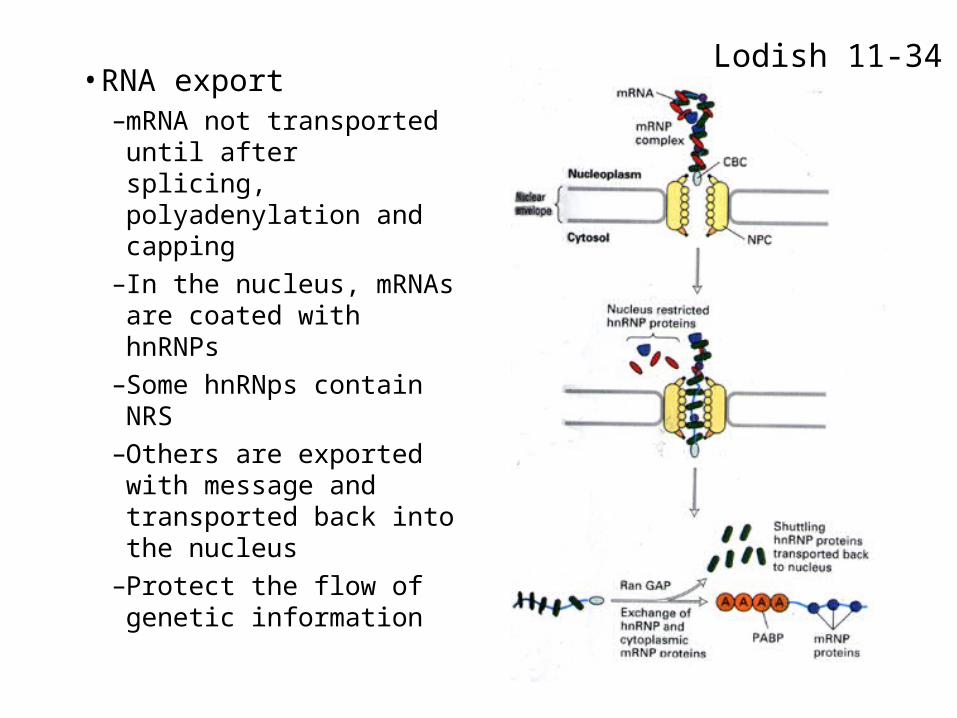
• RNA export–mRNA not transported
until after splicing, polyadenylation and capping
–In the nucleus, mRNAs are coated with hnRNPs
–Some hnRNps contain NRS
–Others are exported with message and transported back into the nucleus
–Protect the flow of genetic information
Lodish 11-34


















