
Rabies Virus Entry into Endosomes in IMR-32 Human Neuroblastoma Cells
9
Rabies Virus Entry into Endosomes in IMR-32 Human Neuroblastoma Cells Peter Lewis, Yiguang Fu, and Thomas L. Lentz 1 Department of Cell Biology, Yale University School of Medicine, New Haven, Connecticut 06520-8002 Received January 21, 1998; accepted June 18, 1998 Early events in rabies virus entry into cultured IMR-32 human neuroblastoma cells were investigated. After adsorption of rabies virus to the cell surface in the cold and warming to 37°C in the presence of tracers for early endosomes, rabies virus and tracers were localized by immunofluorescence microscopy. After 5 min, rabies virus colocalized with LuciferYellow, Texas Red-dextran, rhodamine-wheat germ agglutinin, and transferrin receptor in puncta in the cell body, neu- rites, and nerve terminals. Rabies virus did not colocal- ize with lysosomal glycoprotein. An acidotropic probe revealed that some of the virus-containing puncta were acidified. Rabies virus also colocalized with syn- apsin I, a synaptic vesicle marker, in swellings along processes, indicating some virus enters nerve termi- nals. Electron microscopy revealed the presence of rabies virus within irregular membrane compart- ments located near the cell surface in the cell body and neurites. The membrane of the virus particle was often continuous with that of the vacuole. It is concluded that rabies virus enters IMR-32 neuroblastoma cells by adsorptive endocytosis and that, shortly after entry, rabies virus is located within and fuses with acidic endosomes. r 1998 Academic Press Key Words: endosomes; IMR-32 neuroblastoma cells; rabies virus; synapsin; virus entry. INTRODUCTION Rabies virus is an enveloped, RNA virus belonging to the genus Lyssavirus. After attachment to the cell surface, enveloped viruses enter the cell by direct fusion with the plasma membrane or by adsorptive or receptor-mediated endocytosis (10, 16, 22). In adsorp- tive endocytosis, the virus is internalized in clathrin- coated pits that pinch off from the surface as vesicles, which in turn fuse with endosomes delivering the virus to the interior of the endosome. Endosomes (18) have an acidic interior generated by an ATP-driven proton pump. The acidic environment of the endosome triggers a conformational change in a viral fusion protein, which induces fusion of the viral membrane with the endosome membrane, allowing the viral genome to escape into the cytoplasm. Rabies virus appears to enter CER cells via this route because virus particles have been observed in coated pits and vesicles and lysosomotropic agents, which raise the internal pH of endosomes prevented infection (25). Electron-micro- scopic studies have shown endocytosis of virus particles by neurons (1, 3, 9). Vesicular stomatitis virus, another rhabdovirus, enters MDCK cells by endocytosis (17). In vivo, rabies virus is highly neurotropic and infects primarily neurons. Although it is often assumed that rabies virus enters neurons by the same mechanism other enveloped viruses enter other cell types, there is relatively little information on rabies virus entry into neurons and on subsequent steps after virus entry, for example, whether the virus enters an endosome com- partment and whether these are acidified. In addition, endosomes in neurons are not as well characterized as those in other cell types. Endosomes are generally considered to play a role in synaptic vesicle recycling (5), but the relationship of these endosomes to those involved in sorting and trafficking of endocytosed sub- stances is unknown. These studies were undertaken to investigate early events in rabies virus entry into IMR-32 human neuro- blastoma cells and the role of endosomes in uptake. IMR-32 cells (29) are of neural crest origin and express nicotinic and muscarinic acetylcholine receptors, a-bun- garotoxin-binding sites, and b-adrenergic receptors (4, 13). They form cholinergic synapses with skeletal muscle in vitro (26). Recently, we showed that IMR-32 cells are highly susceptible to rabies virus infection in vitro and that these cells might represent a useful model to investigate infection of human neurons by neurotropic viruses such as rabies virus (12). In the present study, we investigated early events in viral entry after adsorp- tion of rabies virus to IMR-32 cells. In particular, neurons were incubated with virus and tracers for early endosomes to determine whether internalized virus resides in an endosome compartment. It was found that 1 To whom correspondence should be addressed. Fax: (203) 785- 7226. E-mail: [email protected]. EXPERIMENTAL NEUROLOGY 153, 65–73 (1998) ARTICLE NO. EN986879 65 0014-4886/98 $25.00 Copyright r 1998 by Academic Press All rights of reproduction in any form reserved.
-
Upload
peter-lewis -
Category
Documents
-
view
212 -
download
0
Transcript of Rabies Virus Entry into Endosomes in IMR-32 Human Neuroblastoma Cells

Rabies Virus Entry into Endosomes in IMR-32 HumanNeuroblastoma Cells
Peter Lewis, Yiguang Fu, and Thomas L. Lentz1
Department of Cell Biology, Yale University School of Medicine, New Haven, Connecticut 06520-8002
Received January 21, 1998; accepted June 18, 1998
Early events in rabies virus entry into culturedIMR-32 human neuroblastoma cells were investigated.After adsorption of rabies virus to the cell surface inthe cold and warming to 37°C in the presence of tracersfor early endosomes, rabies virus and tracers werelocalized by immunofluorescence microscopy. After 5min, rabies virus colocalized with Lucifer Yellow, TexasRed-dextran, rhodamine-wheat germ agglutinin, andtransferrin receptor in puncta in the cell body, neu-rites, and nerve terminals. Rabies virus did not colocal-ize with lysosomal glycoprotein. An acidotropic proberevealed that some of the virus-containing punctawere acidified. Rabies virus also colocalized with syn-apsin I, a synaptic vesicle marker, in swellings alongprocesses, indicating some virus enters nerve termi-nals. Electron microscopy revealed the presence ofrabies virus within irregular membrane compart-ments located near the cell surface in the cell body andneurites. The membrane of the virus particle was oftencontinuous with that of the vacuole. It is concludedthat rabies virus enters IMR-32 neuroblastoma cells byadsorptive endocytosis and that, shortly after entry,rabies virus is located within and fuses with acidicendosomes. r 1998 Academic Press
Key Words: endosomes; IMR-32 neuroblastoma cells;rabies virus; synapsin; virus entry.
INTRODUCTION
Rabies virus is an enveloped, RNA virus belonging tothe genus Lyssavirus. After attachment to the cellsurface, enveloped viruses enter the cell by directfusion with the plasma membrane or by adsorptive orreceptor-mediated endocytosis (10, 16, 22). In adsorp-tive endocytosis, the virus is internalized in clathrin-coated pits that pinch off from the surface as vesicles,which in turn fuse with endosomes delivering the virusto the interior of the endosome. Endosomes (18) havean acidic interior generated by an ATP-driven proton
pump. The acidic environment of the endosome triggersa conformational change in a viral fusion protein,which induces fusion of the viral membrane with theendosome membrane, allowing the viral genome toescape into the cytoplasm. Rabies virus appears toenter CER cells via this route because virus particleshave been observed in coated pits and vesicles andlysosomotropic agents, which raise the internal pH ofendosomes prevented infection (25). Electron-micro-scopic studies have shown endocytosis of virus particlesby neurons (1, 3, 9). Vesicular stomatitis virus, anotherrhabdovirus, enters MDCK cells by endocytosis (17).
In vivo, rabies virus is highly neurotropic and infectsprimarily neurons. Although it is often assumed thatrabies virus enters neurons by the same mechanismother enveloped viruses enter other cell types, there isrelatively little information on rabies virus entry intoneurons and on subsequent steps after virus entry, forexample, whether the virus enters an endosome com-partment and whether these are acidified. In addition,endosomes in neurons are not as well characterized asthose in other cell types. Endosomes are generallyconsidered to play a role in synaptic vesicle recycling(5), but the relationship of these endosomes to thoseinvolved in sorting and trafficking of endocytosed sub-stances is unknown.
These studies were undertaken to investigate earlyevents in rabies virus entry into IMR-32 human neuro-blastoma cells and the role of endosomes in uptake.IMR-32 cells (29) are of neural crest origin and expressnicotinic and muscarinic acetylcholine receptors, a-bun-garotoxin-binding sites, and b-adrenergic receptors (4,13). They form cholinergic synapses with skeletal musclein vitro (26). Recently, we showed that IMR-32 cells arehighly susceptible to rabies virus infection in vitro andthat these cells might represent a useful model toinvestigate infection of human neurons by neurotropicviruses such as rabies virus (12). In the present study,we investigated early events in viral entry after adsorp-tion of rabies virus to IMR-32 cells. In particular,neurons were incubated with virus and tracers for earlyendosomes to determine whether internalized virusresides in an endosome compartment. It was found that
1 To whom correspondence should be addressed. Fax: (203) 785-7226. E-mail: [email protected].
EXPERIMENTAL NEUROLOGY 153, 65–73 (1998)ARTICLE NO. EN986879
65 0014-4886/98 $25.00Copyright r 1998 by Academic Press
All rights of reproduction in any form reserved.

within 5–10 min rabies virus and endosome markerswere colocalized and that by electron microscopy rabiesvirus was located in structures with the morphologicalcharacteristics of endosomes.
METHODS
Materials
Human neuroblastoma cell line IMR-32 was ob-tained from American Type Culture Collection. Cellswere maintained in Eagles minimum essential mediumwith nonessential amino acids, 10% fetal bovine se-rum/1% glutamine/0.5% penicillin-streptomycin. Cellswere grown at 37°C in 5% CO2 for 2 or more days beforeuse. The challenge virus standard (CVS) strain ofrabies virus was used in these studies. Virus waspropagated in mouse N18 neuroblastoma cells as de-scribed (11). Infected tissue culture supernatants (infec-tion potency 5 3 105 fluorescent focus units/ml) andpartially purified virus (infection potency 5 3 107 fluo-rescent focus units/ml) were prepared as previouslydescribed (11, 12).
Immunofluorescence Microscopy
IMR-32 cells were transferred from 35-mm culturedishes to Lab Tek 8-chamber tissue culture slides(Nunc, Naperville, IL), which had been coated withpolylysine and laminin and cultured for 1 day. Toadsorb virus to the cell surface, IMR-32 cells wereincubated in culture medium containing a high concen-tration of rabies virus (1:200 dilution of partially-purified preparation) for 1 h at 4°C. Cycloheximide (100µg/ml) was included to prevent viral replication. Lyso-Tracker Red DND-99 (Molecular probes, Eugene, OR)was added along with the virus to a concentration of 2µM. Other tracers were added at the end of theadsorption period: Lucifer Yellow CH (Sigma ChemicalCo., St. Louis, MO), 1–2 mg/ml; Texas Red-conjugateddextran (Molecular Probes), 0.2–2 mg/ml; and rhoda-mine-conjugated wheat germ agglutinin (Vector Labora-tories, Burlingame, CA), 10–25 µg/ml. Cells were thenwarmed to 37°C in the cell culture incubator for incre-mental time periods and then rinsed quickly with threechanges of phosphate-buffered saline and fixed for20–30 min in freshly prepared 4% paraformaldehyde in120 mM sodium phosphate buffer, pH 7.4, containing0.12 M sucrose. Except for the LysoTracker Red-labeledcells, fixed preparations were washed three times for 5min each in each of the following solutions: 120 mMsodium phosphate buffer, pH 7.4; low salt buffer (10mM sodium phosphate, pH 7.4, 150 mM NaCl); andhigh salt buffer (20 mM sodium phosphate,pH 7.4, 500mM NaCl). Cultures were then blocked for 30 min atroom temperature in Triton buffer (0.3% Triton X-100,450 mM NaCl, 20 mM sodium phosphate, pH 7.4, and
16% normal goat serum). For detection of lysosomalglycoprotein, 0.01% saponin was used instead of TritonX-100.
Cells were incubated for at least 2 h at room tempera-ture with primary antibodies [fluorescein isothiocya-nate-labeled anti-rabies globulin (Becton-Dickinson,Cockeysville, MD), rabbit anti-Lucifer Yellow (Molecu-lar Probes), mouse monoclonal antibody against humanlysosomal glycoprotein (gift of Dr. Norma Andrews),mouse monoclonal antibody against transferrin recep-tor (gift of Dr. D. Sheff), rabbit polyclonal antibodyagainst synapsin I (gift of Dr. P. De Camilli)] diluted1:200 in Triton buffer. Detection of rabies virus byimmunofluorescence has been reviewed (28). LuciferYellow was detected by immunofluorescence to improvesensitivity and to avoid spectral overlap with thefluorescein-labeled rabies antibody. Secondary antibod-ies [Texas Red-conjugated donkey anti-rabbit antibody(Jackson ImmunoResearch, West Grove, PA); TexasRed-conjugated goat anti-mouse antibody (Jackson Im-munoResearch)] were used to detect anti-Lucifer Yel-low, anti-lysosomal glycoprotein, anti-transferrin recep-tor, and anti-synapsin I antibodies. The cells werewashed for 30–60 min in three changes of high saltbuffer and incubated at least 2 h with the secondaryantibody diluted 1:100 in Triton buffer. Cells were thenwashed three times in high salt buffer; then two timesin low salt buffer; once for 5 min in 120 mM sodiumphosphate, pH 7.4; and, finally, briefly in 5 mM sodiumphosphate before coverslipping with FluorSave (Calbio-chem, San Diego, CA). Cultures were then viewed andphotographed by fluorescence microscopy with a ZeissAxiophot microscope at appropriate wavelengths formarker and rabies virus.
Because LysoTracker Red fluorescence was partiallylost to diffusion of dye during processing, after adsorp-tion of virus, incubation with the dye, and fixation, thecells were rinsed with phosphate-buffered saline andcoverslipped. Photographs were taken of the Lyso-Tracker fluorescence, the coverslip removed, and theremainder of the procedure for immunofluorescentdetection of viral antigen completed. The cells showingLysoTracker Red fluorescence were relocated and photo-graphed for rabies virus fluorescence.
Electron Microscopy
For electron microscopy, virus was adsorbed for 1 h at4°C to the surface of cells grown on coated coverslipsand cells warmed to 37°C for 10 min. Other cells oncoverslips were infected with tissue culture superna-tant (1:1000 dilution in culture medium) and fixed after3 days of infection. Cells were rinsed and fixed with1.5% glutaraldehyde, 4% sucrose in 100 mM cacodylatebuffer, pH 7.2, for 60 min. Coverslips were rinsed incacodylate buffer and postfixed for 60 min with cold 2%osmium tetroxide. Subsequent processing, sectioning,
66 LEWIS, FU, AND LENTZ

and staining were performed as described by Mundiglet al. (20). Sections were examined with a Philips 410electron microscope.
RESULTS
Localization of Virus Adsorbed to Cell Surface
IMR-32 neuroblastoma cells are spindle- to polygon-shaped with interconnecting processes or neurites(Fig. 1). The neurites have small swellings or varicosi-ties along their length and at points of contact with cellbodies and other neurites (Fig. 1F). After adsorption ofvirus at 4°C and 5–10 min at 37°C, rabies virus waslocalized to small punctate structures in IMR-32 neuro-blastoma cells (Figs. 1A and 1D). These were located inthe perikaryon and along the cell processes. In someplaces in the cell body, the small, fluorescent structureswere present in clusters which appeared as fluorescentmasses when out of the plane of focus. Fainter, diffusefluorescence occurred in places on the cell body andalong the processes. The small swellings along neuritesand at points of contact contained viral antigen (Fig.1D). There was little difference in the localization ofrabies virus antigen at time periods up to 2 h.
Colocalization of Rabies Virus and Endocytic Markers
The tracers or markers Lucifer Yellow (Fig. 1B),Texas Red-dextran, rhodamine-wheat germ agglutinin,and transferrin receptor (Fig. 1E) were localized byfluorescence microscopy after adsorption of virus andwarming of cells. These tracers are markers for earlyendosomes at short incubation time periods. The trac-ers showed closely similar patterns of localization inIMR-32 cells. All were present in small, punctatestructures in the cell body and along the processes attimes from 5 min to 2 h (Figs. 1B and 1E). The largestnumber of fluorescent puncta were observed afterstaining for the transferrin receptor (Fig. 1E). All ofthese tracers colocalized with virus antigen in thepuncta in the cell bodies and in swellings along neu-rites. Puncta in the cell body were often located nearthe cell surface (Fig. 1B). Some of the structurescontaining tracer did not contain rabies virus, but moststructures fluorescent for virus also contained tracer.Lysosomes were demonstrated by incubating with anti-lysosomal glycoprotein antibody. These were present assmall dots most numerous in the central region of thecell body (Fig. 2B). The lysosomal glycoprotein did notcolocalize with the structures containing rabies virusantigen (Fig. 2A).
In order to determine whether structures containingrabies virus were acidified, cells were incubated withvirus and LysoTracker Red, a fluorescent probe forlabeling acidic compartments and which is reported toretain its staining pattern after fixation with aldehydes
(Molecular Probes). The suitability of LysoTracker Redwas determined in preliminary experiments in which itwas found that LysoTracker Red colocalized with acri-dine orange, a commonly used acidotropic probe. Thelatter, however, was not used along with rabies virusbecause it does not remain localized after fixation.Because slow diffusion of LysoTracker Red occurredafter fixation, cultures incubated with virus and Lyso-Tracker Red were warmed, fixed, and immediatelyphotographed with fluorescence for LysoTracker Red.They were then processed with labeled anti-rabiesvirus antibody and the same fields reexamined underfluorescein–fluorescence microscopy. It was noted thatcells underwent slight changes in shape during process-ing for virus localization. However, it could be deter-mined that puncta containing rabies antigen also con-tained LysoTracker Red (Fig. 3). Extensive colocalizationoccurred in the dense clusters of puncta in the cell body.
Colocalization of Rabies Virus and Synapsin I
In order to determine whether any virus entered cellsat nerve terminals, cultures were exposed to virus andincubated with antibody against synapsin I, followedby Texas Red-labeled second antibody. Synapsin I waslocalized to many small puncta distributed throughoutthe cell body and processes of IMR-32 cells (Fig. 4).Most of the puncta positive for synapsin I in the cellbody did not contain rabies virus antigen. However,rabies virus and synapsin I were present in the varicosi-ties where neurites contacted another cell or crossedone another.
Electron Microscopy
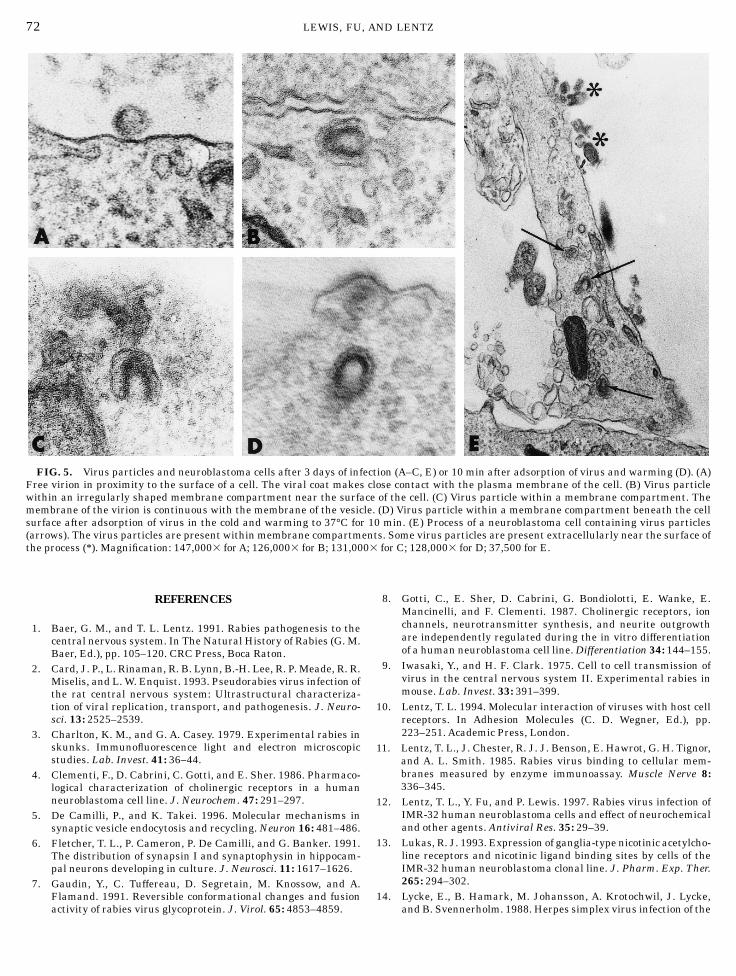
By electron microscopy, cells infected for 3 dayscontained large masses of matrix material with associ-ated virus particles budding into membrane compart-ments and from the cell surface. Free virus particleswere present extracellularly. Some were located inclose proximity to the cell surface with the viral coat incontact with the cell plasma membrane (Fig. 5A).Single virus particles were present within irregularlyshaped membrane compartments immediately beneaththe cell surface (Fig. 5B). These were present in bothcell bodies and processes and were not associated withmatrix material. In many cases, the membrane of thevirus particle was continuous with that of the vesicle(Fig. 5C). These structures were present in both in-fected cells and those warmed to 37°C after adsorptionof virus (Fig. 5D).
Processes of cells came into close proximity to eachother or cell bodies. Morphological specializations char-acteristic of differentiated synapses were not observed[see also Gotti et al. (8)]. Processes were similar instructure and no distinction could be made betweenaxons and dendrites. Small vesicles or dense core
67RABIES VIRUS AND ENDOSOMES

68 LEWIS, FU, AND LENTZ

vesicles were present in processes. Processes containedvirus particles within membrane compartments (Fig.5E).
DISCUSSION
Rabies virus and markers for endosomes were local-ized by immunofluorescence microscopy after adsorp-tion of virus in the cold to the surface of IMR-32neuroblastoma cells and incubation with endocytictracers. Within 5–10 min of warming to 37°C, endocytictracers (Lucifer Yellow, Texas Red-dextran, rhodamine-wheat germ agglutinin, and transferrin receptor) werelocalized to puncta in the perikaryon and along cellprocesses. Tracers also occurred in varicosities alongneurites, representing contacts between neurites. Simi-larly, in cultured hippocampal neurons, endocytic mark-ers label punctate structures in the cell body, dendrites,and axon terminals (20, 21). Rabies virus colocalizedwith the endocytic tracers in the puncta and neuritecontacts in IMR-32 cells.Although previous morphologi-cal studies indicate virus enters neurons by endocyto-sis, these studies showing colocalization of virus andtracers for early endosomes provides functional evi-dence that virus enters neurons by adsorptive endocyto-sis and resides in an endosome compartment shortlyafter uptake. It appears that entry can take place atany region of the nerve cell, including cell body andprocesses. Based on colocalization of rabies virus andLysoTracker Red, a probe for acidic compartments, atleast some of the virus-containing compartments areacidified. Little or no viral antigen was present inlysosomes, indicating virus is not delivered to lyso-somes at time periods up to 2 h after entry.
Further evidence for entry of virus by receptor-mediated endocytosis are previous findings that lysoso-motropic agents inhibited infection of IMR-32 cells (12).Chloroquine, quinine, and NH4Cl inhibited infection byrabies virus. Similarly, chloroquine and NH4Cl were
FIG. 1. Colocalization of rabies virus with Lucifer Yellow andtransferrin receptor. (A–C) Rabies virus and Lucifer Yellow. Cellswere adsorbed with rabies virus and warmed to 37°C for 10 min. (A)Localization of rabies virus. Viral antigen is present within smallpuncta located near the cell surface (arrows). (B) Localization ofLucifer Yellow. Lucifer Yellow shows colocalization with virus antigenin the puncta in the cell bodies (arrows). (C) Corresponding phasemicrograph. (D–F) Rabies virus and transferrin receptor. Cells wereadsorbed with rabies virus and warmed to 37°C for 5 min. Rabiesvirus was localized with fluorescein-labeled rabies antibody andtransferrin receptor with mouse antibody against transferrin recep-tor and Texas Red-conjugated goat anti-mouse antibody. (D) Rabiesvirus is localized in the cell body and along neurites (circles). (E)Transferrin receptor is colocalized with rabies virus in the cell bodyand processes (circles). Some puncta show localization of transferrinreceptor only. (F) Corresponding phase micrograph. Regions of colocal-ization along processes (circles) often appear as moderately denseswellings. Magnification: 9003 for A–C; 5253 for D–F.
FIG. 2. Localization of rabies virus and lysosomal glycoproteinafter adsorption of virus to cells and warming to 37°C for 90 min. (A)Rabies virus is localized to puncta most numerous at the cellperiphery. (B) Lysosomal glycoprotein is localized to clusters ofpuncta most abundant in the central regions of the cells. There islittle colocalization with rabies virus. (C) Corresponding phasemicrograph. Magnification: 5753.
69RABIES VIRUS AND ENDOSOMES

shown to inhibit infection of CER cells (25) and NS20murine neuroblastoma cells (27). These agents raisethe pH of endosomes and lysosomes, preventing fusionof the viral envelope with the endosomal membrane.Fusion of rabies virus to membranes has been observedat low pH (19, 23). Fusion with membranes is mediatedby a conformational change in the rabies virus glycopro-tein (7).
Rabies virus and synapsin I, a synaptic vesicleprotein, were localized in IMR-32 cells to determine
whether virus enter cells at synapses. In mature hippo-campal neurons in culture, synapsin I is primarilylocalized to presynaptic boutons (6). In immature neu-rons, synapsin is localized to puncta in the cell body andprocesses. IMR-32 cells showed both patterns of local-ization. Rabies virus was colocalized with synapsin I inthe varicosities along neurites considered to representpoints of contact, but much less so in the cell body. Thisis not surprising because much of the synapsin in thecell body is most likely newly synthesized synapsin and
FIG. 3. Localization of rabies virus and LysoTracker Red. Cells were adsorbed with rabies virus, incubated with LysoTracker Red, andfixed 5 min after warming to 37°C. Cells were then photographed with fluorescence for LysoTracker Red and by phase microscopy. They werethen processed with fluorescein-labeled anti-rabies virus antibody and the same fields reexamined with fluorescence microscopy forfluorescein and phase microscopy. (A) Rabies virus is localized in puncta (circles) and clusters of small puncta (arrows). (B) Correspondingphase micrograph. (C) LysoTracker Red shows colocalization with rabies virus in puncta (circles) and clusters of puncta (arrows). (D)Corresponding phase micrograph. Cells underwent slight changes in shape during processing for rabies virus localization (B and D).Magnification: 8253.
70 LEWIS, FU, AND LENTZ

not associated with synapses. These results indicatethat at least some virus enters neurons at nerveterminals. In addition, because these structures showedcolocalization of virus and endocytic tracers, the virusresides in endosomes in the nerve terminal. Electronmicroscopic observations of infected skunk brain haveshown budding of virus from dendrites into adjacentaxon terminals (3). Uptake of virus by nerve terminalsrepresents a mechanism for cell to cell spread of virusin the central nervous system.
By electron microscopy, single virus particles werelocated within small, membranous vacuoles near thecell surface. Morphologically, these structures resembleendosomes observed in other cell types. In most cases,the membrane of the virus was continuous with that ofthe vacuole. After 3 days of infection when virus isreplicating, some of these images such as those withinmasses of matrix material could represent virus bud-ding into a membrane compartment. However, thosepresent 10 min after virus adsorption, and probablythose in infected cells located near the cell surface andnot in proximity to matrix material, most likely repre-sent internalized virus fusing with the endosome.These structures were observed in both the cell bodyand processes. Fusion must take place quickly afterendocytosis because fused particles were present 10min after adsorption and few unfused particles wereseen within membrane compartments. It appears, there-fore, that in IMR-32 cells, at least some virus fuses withthe endosome near the site of uptake. If fusion takesplace in axon terminals of neurons, the viral nucleocap-sid would have to be transported retrogradely forreplication to occur, as has been observed with swinea-herpesvirus and herpes simplex virus (2, 14, 15, 24).It cannot be ruled out, however, that some virusparticles were transported in the endosome prior tofusion.
ACKNOWLEDGMENTS
We thank Dr. Olaf Mundigl and Dr. Pietro De Camilli for helpfuldiscussions and suggestions and Jack Valentijn for photographicassistance. This work was supported by Grant NS34275 from theNational Institutes of Health.
FIG. 4. Colocalization of rabies virus and synapsin I. Cells wereadsorbed with rabies virus and warmed to 37°C for 5 min. Rabiesvirus was localized with fluorescein-labeled rabies antibody andsynapsin I with rabbit anti-synapsin antibody and Texas Red-conjugated donkey anti-rabbit antibody. (A) Rabies virus is localizedin puncta in cell bodies and in swellings along processes (circles). (B)Synapsin I is colocalized with rabies virus in varicosities alongprocesses (circles). Some puncta in the cell body show localization ofsynapsin only. (C) Corresponding phase micrograph. Some regionsshowing colocalization are circled. Magnification: 4253.
71RABIES VIRUS AND ENDOSOMES
REFERENCES
1. Baer, G. M., and T. L. Lentz. 1991. Rabies pathogenesis to thecentral nervous system. In The Natural History of Rabies (G. M.Baer, Ed.), pp. 105–120. CRC Press, Boca Raton.
2. Card, J. P., L. Rinaman, R. B. Lynn, B.-H. Lee, R. P. Meade, R. R.Miselis, and L. W. Enquist. 1993. Pseudorabies virus infection ofthe rat central nervous system: Ultrastructural characteriza-tion of viral replication, transport, and pathogenesis. J. Neuro-sci. 13: 2525–2539.
3. Charlton, K. M., and G. A. Casey. 1979. Experimental rabies inskunks. Immunofluorescence light and electron microscopicstudies. Lab. Invest. 41: 36–44.
4. Clementi, F., D. Cabrini, C. Gotti, and E. Sher. 1986. Pharmaco-logical characterization of cholinergic receptors in a humanneuroblastoma cell line. J. Neurochem. 47: 291–297.
5. De Camilli, P., and K. Takei. 1996. Molecular mechanisms insynaptic vesicle endocytosis and recycling. Neuron 16: 481–486.
6. Fletcher, T. L., P. Cameron, P. De Camilli, and G. Banker. 1991.The distribution of synapsin I and synaptophysin in hippocam-pal neurons developing in culture. J. Neurosci. 11: 1617–1626.
7. Gaudin, Y., C. Tuffereau, D. Segretain, M. Knossow, and A.Flamand. 1991. Reversible conformational changes and fusionactivity of rabies virus glycoprotein. J. Virol. 65: 4853–4859.
8. Gotti, C., E. Sher, D. Cabrini, G. Bondiolotti, E. Wanke, E.Mancinelli, and F. Clementi. 1987. Cholinergic receptors, ionchannels, neurotransmitter synthesis, and neurite outgrowthare independently regulated during the in vitro differentiationof a human neuroblastoma cell line. Differentiation 34: 144–155.
9. Iwasaki, Y., and H. F. Clark. 1975. Cell to cell transmission ofvirus in the central nervous system II. Experimental rabies inmouse. Lab. Invest. 33: 391–399.
10. Lentz, T. L. 1994. Molecular interaction of viruses with host cellreceptors. In Adhesion Molecules (C. D. Wegner, Ed.), pp.223–251. Academic Press, London.
11. Lentz, T. L., J. Chester, R. J. J. Benson, E. Hawrot, G. H. Tignor,and A. L. Smith. 1985. Rabies virus binding to cellular mem-branes measured by enzyme immunoassay. Muscle Nerve 8:336–345.
12. Lentz, T. L., Y. Fu, and P. Lewis. 1997. Rabies virus infection ofIMR-32 human neuroblastoma cells and effect of neurochemicaland other agents. Antiviral Res. 35: 29–39.
13. Lukas, R. J. 1993. Expression of ganglia-type nicotinic acetylcho-line receptors and nicotinic ligand binding sites by cells of theIMR-32 human neuroblastoma clonal line. J. Pharm. Exp. Ther.265: 294–302.
14. Lycke, E., B. Hamark, M. Johansson, A. Krotochwil, J. Lycke,and B. Svennerholm. 1988. Herpes simplex virus infection of the
FIG. 5. Virus particles and neuroblastoma cells after 3 days of infection (A–C, E) or 10 min after adsorption of virus and warming (D). (A)Free virion in proximity to the surface of a cell. The viral coat makes close contact with the plasma membrane of the cell. (B) Virus particlewithin an irregularly shaped membrane compartment near the surface of the cell. (C) Virus particle within a membrane compartment. Themembrane of the virion is continuous with the membrane of the vesicle. (D) Virus particle within a membrane compartment beneath the cellsurface after adsorption of virus in the cold and warming to 37°C for 10 min. (E) Process of a neuroblastoma cell containing virus particles(arrows). The virus particles are present within membrane compartments. Some virus particles are present extracellularly near the surface ofthe process (*). Magnification: 147,0003 for A; 126,0003 for B; 131,0003 for C; 128,0003 for D; 37,500 for E.
72 LEWIS, FU, AND LENTZ

human sensory neuron. An electron microscopy study. Arch.Virol. 101: 87–104.
15. Marchand, C. F., and M. E. Schwab. 1986. Binding, uptake andretrograde axonal transport of herpes virus suis in sympatheticneurons. Brain Res. 383: 262–270.
16. Marsh, M., and A. Helenius. 1989. Virus entry into animal cells.Adv. Virus Res. 36: 107–151.
17. Matlin, K. S., H. Reggio, A. Helenius, and K. Simons. 1982.Pathway of vesicular stomatitis entry leading to infection. J.Mol. Biol. 156: 609–631.
18. Mellman, I. 1996. Endocytosis and molecular sorting. Annu.Rev. Cell Dev. Biol. 12: 575–625.
19. Mifune, K., M. Ohuchi, and K. Mannen. 1982. Hemolysis andcell fusion by rhabdoviruses. FEBS Lett. 137: 293–297.
20. Mundigl, O., M. Matteoli, L. Daniell, A. Thomas-Reetz, A.Metcalf, R. Jahn, and P. De Camilli. 1993. Synaptic vesicleproteins and early endosomes in cultured hippocampal neurons:Differential effects of Brefeldin A in axons and dendrites. J. CellBiol. 122: 1207–1221.
21. Parton, R. G., K. Simons, and C. G. Dotti. 1992. Axonal anddendritic endocytic pathways in cultured neurons. J. Cell Biol.119: 123–137.
22. Paulson, J. C. 1985. Interactions of animal viruses with cell
surface receptors. In The Receptors (P. M. Conn, Ed.), Vol. II, pp.131–219. Academic Press, Orlando.
23. Perrin, P., D. Portnoı, and P. Sureau. 1982. Etude de l’adsorptionet de la penetration du virus rabique: Interactions avec lescellules BHK21 et des membranes artificielles. Ann. Virol. (Inst.Pasteur) 133 E: 403–422.
24. Sodeik, B., M. W. Ebersold, and A. Helenius. 1997. Microtubule-mediated transport of incoming herpes simplex virus 1 capsidsto the nucleus. J. Cell Biol. 136: 1007–1021.
25. Superti, F., M. Derer, and H. Tsiang. 1984. Mechanism of rabiesvirus entry in CER cells. J. Gen. Virol. 65: 781–789.
26. Thompson, J. M., E. D. London, and J. E. Johnson, Jr. 1982.Ultrastructural, functional and biochemical characteristics ofmouse and human neuroblastoma cell lines. Neuroscience 7:1807–1815.
27. Tsiang, H., and F. Superti. 1984. Ammonium chloride andchloroquine inhibit rabies virus infection in neuroblastomacells. Arch. Virol 81: 377–382.
28. Trimarchi, C. V., and J. G. Debbie. 1991. The fluorescentantibody in rabies. In The Natural History of Rabies (G. M.Baer, Ed.), pp. 219–233. CRC Press, Boca Raton.
29. Tumilowicz, J. J., W. W. Nichols, J. J. Cholon, and A. E. Greene.1970. Definition of a continuous human cell line derived fromneuroblastoma. Cancer Res. 30: 2110–2118.
73RABIES VIRUS AND ENDOSOMES


















