
Proteolytic Cleavage of Covalently Linked Cell Wall Proteins by
12
EUKARYOTIC CELL, Jan. 2011, p. 98–109 Vol. 10, No. 1 1535-9778/11/$12.00 doi:10.1128/EC.00210-10 Copyright © 2011, American Society for Microbiology. All Rights Reserved. Proteolytic Cleavage of Covalently Linked Cell Wall Proteins by Candida albicans Sap9 and Sap10 † Lydia Schild, 1 Antje Heyken, 1 Piet W. J. de Groot, 2,3 Ekkehard Hiller, 4 Marlen Mock, 1 Chris de Koster, 2 Uwe Horn, 5 Steffen Rupp, 4 and Bernhard Hube 1,6 * Department of Microbial Pathogenicity Mechanisms, Leibniz Institute for Natural Product Research and Infection Biology, Hans-Knoell-Institute (HKI), Beutenbergstrasse 11a, 07745 Jena, Germany 1 ; Swammerdam Institute for Life Sciences, University of Amsterdam, Science Park 904, 1098 XH Amsterdam, Netherlands 2 ; Regional Centre for Biomedical Research, Albacete Science and Technology Park, University of Castilla—La Mancha, 02006 Albacete, Spain 3 ; Fraunhofer Institute for Interfacial Engineering and Biotechnology, Nobelstrasse 12, 70569 Stuttgart, Germany 4 ; Bio Pilot Plant, Leibniz Institute for Natural Product Research and Infection Biology, Hans-Knoell-Institute (HKI), Beutenbergstrasse 11a, 07745 Jena, Germany 5 ; and Friedrich Schiller University, Jena, Germany 6 Received 31 August 2010/Accepted 12 November 2010 The cell wall of the human-pathogenic fungus Candida albicans is a robust but also dynamic structure which mediates adaptation to changing environmental conditions during infection. Sap9 and Sap10 are cell surface- associated proteases which function in C. albicans cell wall integrity and interaction with human epithelial cells and neutrophils. In this study, we have analyzed the enzymatic properties of Sap9 and Sap10 and investigated whether these proteases cleave proteins on the fungal cell surface. We show that Sap9 and Sap10, in contrast to other aspartic proteases, exhibit a near-neutral pH optimum of proteolytic activity and prefer the processing of peptides containing basic or dibasic residues. However, both proteases also cleaved at nonbasic sites, and not all tested peptides with dibasic residues were processed. By digesting isolated cell walls with Sap9 or Sap10, we identified the covalently linked cell wall proteins (CWPs) Cht2, Ywp1, Als2, Rhd3, Rbt5, Ecm33, and Pga4 as in vitro protease substrates. Proteolytic cleavage of the chitinase Cht2 and the glucan-cross-linking protein Pir1 by Sap9 was verified using hemagglutinin (HA) epitope-tagged versions of both proteins. Deletion of the SAP9 and SAP10 genes resulted in a reduction of cell-associated chitinase activity similar to that upon deletion of CHT2, suggesting a direct influence of Sap9 and Sap10 on Cht2 function. In contrast, cell surface changes elicited by SAP9 and SAP10 deletion had no major impact on the phagocytosis and killing of C. albicans by human macrophages. We propose that Sap9 and Sap10 influence distinct cell wall functions by proteolytic cleavage of covalently linked cell wall proteins. The polymorphic fungus Candida albicans is the most fre- quent cause of disseminated candidiasis. This opportunistic human pathogen is a frequent colonizer of the gastrointestinal and urogenital tract and skin, where it exists as a member of the normal microbial flora in healthy individuals. However, even a mildly compromised immune system or a minor imbalance of the microbiota can be sufficient for C. albicans to cause super- ficial skin or mucosal infections (50). Furthermore, in cases of impaired immunity or disruption of natural barriers, C. albicans can cause fatal systemic disease, disseminating throughout the bloodstream and infecting internal organs (53, 60, 73). Several virulence attributes contribute to the pathogenic po- tential of C. albicans (7). For example, the ability to switch from yeast to hyphae permits tissue invasion and immune eva- sion (72). Furthermore, C. albicans secretes a multitude of hydrolytic enzymes, namely, lipases, phospholipases, and pro- teases (62). The Sap protein family of aspartic proteases, con- sisting of the 10 individual members Sap1 to Sap10, has been described as a key virulence determinant of C. albicans. Ex- tensive research on Sap1 to Sap6 has demonstrated the func- tional association of Saps with C. albicans pathogenicity by hydrolyzing host proteins. Sap functions affect a variety of processes, from tissue invasion to immune evasion (for a re- view see the work by Naglik et al. [46]). Recombinant Sap proteases have been used to determine the biochemical prop- erties of these enzymes, demonstrating substrate cleavage at acidic pH between larger hydrophobic amino acids (4, 31, 59). Another virulence attribute of C. albicans is its metabolic flexibility and the ability to rapidly adapt to changes in the extracellular environment during infection. The fungal cell wall, a robust but also dynamic structure, plays an important role in such environmental adaptation. The C. albicans cell wall is a bilayered structure. An inner polysaccharide network consists of tightly packed -1,3-glucan chains that are co- valently linked to -1,6-glucan molecules and underlying chitin. This inner polysaccharide layer is covered by an outer protein coat of often highly glycosylated mannoproteins (11, 61). Cell wall proteins (CWPs) are either covalently linked via a glycosylphosphatidylinositol (GPI) remnant to -1,6-glucan (GPI-CWPs) (57) or directly linked to -1,3-glucan via an alkali-sensitive linkage (non-GPI-CWPs or Pir-CWPs) (61). A third group, the SDS-soluble CWPs, are not covalently linked and represent mainly cell surface-associated cytosolic proteins (67). * Corresponding author. Mailing address: Department of Microbial Pathogenicity Mechanisms, Leibniz Institute for Natural Product Re- search and Infection Biology, Hans-Knoell-Institute (HKI), Beuten- bergstrasse 11a, 07745 Jena, Germany. Phone: (493641) 5321400. Fax: (493641) 6520810. E-mail: [email protected]. † Supplemental material for this article may be found at http://ec .asm.org/. Published ahead of print on 19 November 2010. 98 on January 4, 2019 by guest http://ec.asm.org/ Downloaded from
Transcript of Proteolytic Cleavage of Covalently Linked Cell Wall Proteins by

EUKARYOTIC CELL, Jan. 2011, p. 98–109 Vol. 10, No. 11535-9778/11/$12.00 doi:10.1128/EC.00210-10Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Proteolytic Cleavage of Covalently Linked Cell Wall Proteins byCandida albicans Sap9 and Sap10�†
Lydia Schild,1 Antje Heyken,1 Piet W. J. de Groot,2,3 Ekkehard Hiller,4 Marlen Mock,1Chris de Koster,2 Uwe Horn,5 Steffen Rupp,4 and Bernhard Hube1,6*
Department of Microbial Pathogenicity Mechanisms, Leibniz Institute for Natural Product Research and Infection Biology,Hans-Knoell-Institute (HKI), Beutenbergstrasse 11a, 07745 Jena, Germany1; Swammerdam Institute for Life Sciences,
University of Amsterdam, Science Park 904, 1098 XH Amsterdam, Netherlands2; Regional Centre for Biomedical Research,Albacete Science and Technology Park, University of Castilla—La Mancha, 02006 Albacete, Spain3; Fraunhofer Institute forInterfacial Engineering and Biotechnology, Nobelstrasse 12, 70569 Stuttgart, Germany4; Bio Pilot Plant, Leibniz Institute for
Natural Product Research and Infection Biology, Hans-Knoell-Institute (HKI), Beutenbergstrasse 11a,07745 Jena, Germany5; and Friedrich Schiller University, Jena, Germany6
Received 31 August 2010/Accepted 12 November 2010
The cell wall of the human-pathogenic fungus Candida albicans is a robust but also dynamic structure whichmediates adaptation to changing environmental conditions during infection. Sap9 and Sap10 are cell surface-associated proteases which function in C. albicans cell wall integrity and interaction with human epithelial cellsand neutrophils. In this study, we have analyzed the enzymatic properties of Sap9 and Sap10 and investigatedwhether these proteases cleave proteins on the fungal cell surface. We show that Sap9 and Sap10, in contrastto other aspartic proteases, exhibit a near-neutral pH optimum of proteolytic activity and prefer the processingof peptides containing basic or dibasic residues. However, both proteases also cleaved at nonbasic sites, andnot all tested peptides with dibasic residues were processed. By digesting isolated cell walls with Sap9 or Sap10,we identified the covalently linked cell wall proteins (CWPs) Cht2, Ywp1, Als2, Rhd3, Rbt5, Ecm33, and Pga4as in vitro protease substrates. Proteolytic cleavage of the chitinase Cht2 and the glucan-cross-linking proteinPir1 by Sap9 was verified using hemagglutinin (HA) epitope-tagged versions of both proteins. Deletion of theSAP9 and SAP10 genes resulted in a reduction of cell-associated chitinase activity similar to that upon deletionof CHT2, suggesting a direct influence of Sap9 and Sap10 on Cht2 function. In contrast, cell surface changeselicited by SAP9 and SAP10 deletion had no major impact on the phagocytosis and killing of C. albicans byhuman macrophages. We propose that Sap9 and Sap10 influence distinct cell wall functions by proteolyticcleavage of covalently linked cell wall proteins.
The polymorphic fungus Candida albicans is the most fre-quent cause of disseminated candidiasis. This opportunistichuman pathogen is a frequent colonizer of the gastrointestinaland urogenital tract and skin, where it exists as a member ofthe normal microbial flora in healthy individuals. However,even a mildly compromised immune system or a minor imbalanceof the microbiota can be sufficient for C. albicans to cause super-ficial skin or mucosal infections (50). Furthermore, in cases ofimpaired immunity or disruption of natural barriers, C. albicanscan cause fatal systemic disease, disseminating throughout thebloodstream and infecting internal organs (53, 60, 73).
Several virulence attributes contribute to the pathogenic po-tential of C. albicans (7). For example, the ability to switchfrom yeast to hyphae permits tissue invasion and immune eva-sion (72). Furthermore, C. albicans secretes a multitude ofhydrolytic enzymes, namely, lipases, phospholipases, and pro-teases (62). The Sap protein family of aspartic proteases, con-sisting of the 10 individual members Sap1 to Sap10, has been
described as a key virulence determinant of C. albicans. Ex-tensive research on Sap1 to Sap6 has demonstrated the func-tional association of Saps with C. albicans pathogenicity byhydrolyzing host proteins. Sap functions affect a variety ofprocesses, from tissue invasion to immune evasion (for a re-view see the work by Naglik et al. [46]). Recombinant Sapproteases have been used to determine the biochemical prop-erties of these enzymes, demonstrating substrate cleavage atacidic pH between larger hydrophobic amino acids (4, 31, 59).
Another virulence attribute of C. albicans is its metabolicflexibility and the ability to rapidly adapt to changes in theextracellular environment during infection. The fungal cellwall, a robust but also dynamic structure, plays an importantrole in such environmental adaptation. The C. albicans cellwall is a bilayered structure. An inner polysaccharide networkconsists of tightly packed �-1,3-glucan chains that are co-valently linked to �-1,6-glucan molecules and underlyingchitin. This inner polysaccharide layer is covered by an outerprotein coat of often highly glycosylated mannoproteins (11,61). Cell wall proteins (CWPs) are either covalently linked viaa glycosylphosphatidylinositol (GPI) remnant to �-1,6-glucan(GPI-CWPs) (57) or directly linked to �-1,3-glucan via analkali-sensitive linkage (non-GPI-CWPs or Pir-CWPs) (61). Athird group, the SDS-soluble CWPs, are not covalently linkedand represent mainly cell surface-associated cytosolic proteins(67).
* Corresponding author. Mailing address: Department of MicrobialPathogenicity Mechanisms, Leibniz Institute for Natural Product Re-search and Infection Biology, Hans-Knoell-Institute (HKI), Beuten-bergstrasse 11a, 07745 Jena, Germany. Phone: (493641) 5321400. Fax:(493641) 6520810. E-mail: [email protected].
† Supplemental material for this article may be found at http://ec.asm.org/.
� Published ahead of print on 19 November 2010.
98
on January 4, 2019 by guesthttp://ec.asm
.org/D
ownloaded from

The important role of covalently linked CWPs in C. albicansfitness and virulence has recently been reviewed by Klis et al.(30). CWPs mediate biofilm formation and promote adherenceto host cells and invasion into epithelial cell layers (e.g., the Alsprotein family of adhesins, Ywp1 and Ecm33) (24, 27, 37).Other CWPs mask cell wall �-1,3-glucan from recognition byhost immune cells (71) or participate in iron uptake (e.g., theiron acquisition protein Rbt5) (69). During adaptation tochanging environmental conditions, cell wall robustness andintegrity have to be maintained. Such processes require cellwall structural proteins like the glucan-cross-linking proteinPir1 (35) and remodeling enzymes, such as chitinases (e.g.,Cht2 [39]) or transglucosidases (e.g., Pga4 [56]). Cell wall re-modeling is thus an essential process for C. albicans viabilityand pathogenicity and has to be tightly regulated. On the geneexpression level, many CWP-encoding genes are regulated viaenvironmental sensing signaling pathways, such as the MAPkinase (Mkc1-mediated) cell wall integrity pathway (42). Onthe protein level, CWPs are often stable and not subject torapid turnover (30), suggesting additional, yet undefined, reg-ulatory events. The aspartic proteases Sap9 and Sap10 arepotential factors mediating such regulatory processes on thecell surface. Both proteases are, like other Sap family mem-bers, targeted to the cellular secretion machinery by an N-terminal signal peptide. However, only Sap9 and Sap10 containC-terminal GPI consensus sequences, which mediate localiza-tion in the cell membrane and cell wall (1), a feature typical formembers of the Saccharomyces cerevisiae yapsin family of reg-ulatory aspartic proteases (21). In agreement with such regu-latory functions, mutants lacking SAP9 or SAP10 exhibit pro-found cell wall and cell separation defects and are modified in
their interaction with epithelial cells and neutrophils (1, 26).Such phenotypes imply an important role for both proteases incell wall integrity, which influences C. albicans-host cell inter-action.
In the present study we aimed to characterize Sap9 andSap10 enzymatic functions, to identify substrates of these pro-teases among C. albicans cell surface proteins, and to definethe impact of cleavage events on substrate function. We usedrecombinant Sap9 and Sap10 to gain insight into the biochem-ical properties and cleavage preferences of both proteases. Todefine protease targets among CWPs, we analyzed solubleCWP profiles of the sap9� sap10� double mutant strain andtested proteolytic cleavage of proteins present in isolated fun-gal cell walls and of hemagglutinin (HA) epitope-tagged CWPsby Sap9 and Sap10. The activity of selected protease substratesin sap9� and sap10� single and double mutants was assayed toreveal a possible connection between proteolytic processingand distinct CWP function. Finally, we determined whetherSap9 and Sap10 are important for the phagocytosis and killingof C. albicans by human macrophages.
MATERIALS AND METHODS
Strains and growth conditions. Candida albicans and Pichia pastoris strainsused in this study are listed in Table 1. Strains were cultured at 30°C in yeastextract-peptone-dextrose (YPD)-rich medium (10 g/liter yeast extract, 20 g/literBacto peptone, 20 g/liter D-glucose) or synthetic defined (SD) medium (6.7 g/literyeast nitrogen base [YNB], YNB without amino acids [Becton Dickinson] sup-plemented with 75 mM ammonium sulfate, and 20 g/liter D-glucose). For growthon solid media plates, 2% agar was added. To avoid clumping of the sap9� andsap10� mutants, strains were precultured by streaking on YPD plates and incu-bating overnight at 30°C. Routinely, experiments were carried out with the
TABLE 1. Strains used in this study
Strain Description or genotype Reference orsource
Candida albicans strainsSC5314 Clinical isolate 23CAI4 SC5314, ura3�::imm434/ura3�::imm434 19CAI4 � pCIp10 SC5314, ura3�::imm434/ura3�::imm434 � pCIp10 (integration) 19, 45sap9� � pCIp10 CAI4, sap9�::hisG/sap9�::hisG � pCIp10 (integration) 1sap10� � pCIp10 CAI4, sap10�::hisG/sap10�::hisG � pCIp10 (integration) 1sap9� sap10� Ura� CAI4, sap10�::hisG/sap10�::hisG sap9�::hisG/sap9�::hisG-URA3-hisG 1sap9� sap10� CAI4, sap10�::hisG/sap10�::hisG sap9�::his/sap9�::hisG This studysap9� sap10� � pCIp10 CAI4, sap10�::hisG/sap10�::hisG sap9�::his/sap9�::hisG � pCIp10 (integration) This studyCAI4 Cht2-HA CAI4, CHT2/CHT2::HA This studysap9� sap10� Cht2-HA sap9� sap10�, CHT2/CHT2::HA This studyCAI4 Ywp1-HA CAI4, YWP1/YWP1::HA This studysap9� sap10� Ywp1-HA sap9� sap10�, YWP1/YWP1::HA This studyCAI4 Rbt5-HA CAI4, RBT5/RBT5::HA This studysap9� sap10� Rbt5-HA sap9� sap10�, RBT5/RBT5::HA This studyCAI4 Ecm33-HA CAI4, ECM33/ECM33::HA This studysap9� sap10� Ecm33-HA sap9� sap10�, ECM33/ECM33::HA This studyCAI4 Pir1-HA CAI4, PIR1/PIR1::HA This studysap9� sap10� Pir1-HA sap9� sap10�, PIR1/PIR1::HA This studycht2� CAI4, cht2�::hisG/cht2�::hisG-URA3-hisG 63rbt5� CAI4, rbt5�::hisG/rbt5�::hisG-URA3-hisG 6
Pichia pastoris strainspCA2 GS115 � pKJ113 with C. albicans SAP2 integration 4pCA6 GS115 � pKJ113 with C. albicans SAP6 integration 4pCA9 GS115 � pKJ113 with C. albicans SAP9 integration M. Monod; 1pCA10 GS115 � pKJ113 with C. albicans SAP10 integration M. Monod; 1
VOL. 10, 2011 C. ALBICANS Sap9 AND Sap10 CLEAVE CELL WALL PROTEINS 99
on January 4, 2019 by guesthttp://ec.asm
.org/D
ownloaded from

pCIp10-containing sap9� and sap10� mutant strains and the CAI4 � pCIp10strain as the wild-type (wt) control.
Plasmid construction. Plasmid pHA-URA3-HA was constructed by insertingtwo direct repeats of the 6�HA epitope into the vector pCIp10 (45) (see Fig.3A). 6�HA was amplified from plasmid pOM10 (22) (kindly provided byEUROSCARF; http://www.uni-frankfurt.de/fb15/mikro/EUROSCARF), wherebyPCR primers introduced terminal restriction sites and the 24-bp linker (GA)6. ASacI/NotI 6�HA fragment was amplified with primers HA-cassette for1 and rev1and cloned 5� of URA3, while a SpeI/SpeI 6�HA fragment was amplified withprimers HA-cassette for2 and rev2 and cloned 3� of URA3. Primers are listed inTable 2.
Strain construction. For the chromosomal integration of pCIp10 into thesap9� sap10� double mutant, the Ura-positive mutant strain (sap9� sap10�Ura�) was streaked on 25 �g/ml 5-fluoroorotic acid (FOA)-containing plates toselect for loss of the URA3 gene. Next, strains were transformed with StuI-linearized pCIp10, and Ura-positive integrants were selected on SD plates.
Strains containing insertions of an internal HA epitope tag into open readingframes were obtained as follows (see Fig. 3A). A tag-URA3-tag cassette wasamplified from pHA-URA3-HA with T1 for and rev primers, such that the endsof the PCR fragments possessed homology to the gene of interest. After trans-formation, Ura� clones with tagging cassette integration were selected and thenplated on FOA to obtain cells in which recombination between the epitoperepeats had removed the URA3 gene and generated a single copy of the 6�HAepitope. Primers are listed in Table 2.
C. albicans transformation. C. albicans strains were transformed with a mod-ified lithium acetate procedure (68). Uridine-prototrophic transformants wereselected on SD agar plates. Correct integration was verified by PCR.
Recombinant proteases and peptide cleavage assay. The Pichia pastoris ex-pression system (Invitrogen) was used to express recombinant C. albicans aspar-tic proteases rSap2, rSap6, rSap9, and rSap10 as previously described (4). Pichia-expressed rSap9 and rSap10 proteins lacked C-terminal GPI anchor consensussequences (rSap9, 50 C-terminal amino acids; rSap10, 66 C-terminal aminoacids). Proteins were recovered from P. pastoris culture supernatants. Proteaseswere purified via anion-exchange chromatography on Q-Sepharose columns(rSap2, rSap9, and rSap10) or via cation-exchange chromatography on SP-Sepharose columns (rSap6) and desalted by passage through a Sephadex G25
column, using 0.1 M sodium citrate buffer, pH 4.5 (rSap2 and rSap6) or pH 5.5(rSap9 and rSap10).
General proteolytic activity of recombinant proteases in the presence or ab-sence of pepstatin A (Sigma Aldrich) was tested with a microplate fluorescence-based casein assay (Enzchek protease assay kit; Molecular Probes) according to
the manufacturer’s instructions. Reactions were performed in 0.1 M sodiumcitrate buffer (pH 3 to 6) or 0.1 M potassium phosphate buffer (pH 7 to 8) at37°C. For all subsequently described rSap9 and rSap10 cleavage assays, 0.1 Msodium citrate buffer, pH 6, was used. Specific activity of rSap preparations wasdetermined by comparing their activity to the defined activity of 5 U trypsin (0.29�g; sequencing grade, modified; Promega) in the fluorescence-quenched caseinassay. An average of 1.5 �g rSap9, 15 �g rSap10, 3 �g rSap2, and 0.2 �g rSap6corresponded to the activity of 5 U trypsin.
The activity of 1 �g rSap9 and rSap10 against a protease substrate set of 360internally quenched peptides was tested according to the manufacturer’s instruc-tions (JPT Peptide Technologies GmbH). Screened peptides are listed on themanufacturer’s homepage (http://www.jpt.com). To evaluate peptide cleavage,fluorescence was quantified for every peptide after a 2-h reaction at 37°C, and thepeptide with the highest fluorescent value was determined. A peptide was se-lected as processed if its fluorescence reached at least 40% of the highest valuepeptide, in at least two out of three independent experiments.
Analysis of soluble cell wall proteins. For a description of biotinylation andaffinity purification of cell surface proteins, 2-dimensional (2D) gel electrophore-sis, and matrix-assisted laser desorption ionization–time of flight tandem massspectrometry (MALDI-TOF MS/MS) analysis, see the supplemental material.
Cell wall isolation and cell wall digestion. For cell wall isolation, C. albicanssap9� sap10� double mutant cultures were grown in liquid SD medium bufferedwith 0.1 M sodium citrate buffer to pH 4.0 or with 0.1 M potassium phosphatebuffer to pH 6.5. Two hundred optical density (OD) units (e.g., 100-ml culturewith an OD at 600 nm [OD600] of 2) of cells were harvested after 8 h (latelogarithmic phase) or 24 h (stationary phase) of growth at 30°C. Isolation ofSDS-treated cell walls was performed as previously described (15). Six milligramsof freeze-dried cell walls were incubated with 2.75 �g rSap9 or 40 �g rSap10 orwithout protease. After overnight incubation at 37°C, supernatants were col-lected and freeze-dried, and released peptides were identified by liquid chroma-tography (LC)-MS/MS analysis.
LC-MS/MS analysis. LC-MS/MS analysis was performed as described previ-ously (15). Samples were analyzed with and without digestion with trypsin. Priorto analysis, peptides were desalted and collected on Omix C18 pipette tips(Varian). Protein spectra were processed with MASCOT software (Matrix Sci-ence), and MASCOT scores were used to evaluate the identified peptides andproteins. A P value of �0.05 was considered significant for peptide identification.Control samples without addition of rSaps but with tryptic digestion yielded nopeptide identifications.
Western blot analysis of HA epitope-tagged proteins. Strains expressing HA-tagged CWPs were grown in SD liquid medium for 24 h at 30°C. Cultures of
TABLE 2. Primers used in this study
Name Sequence 5�–3�a
HA-cassette for1 .............ATCACAGATCGAGCTCGGTGCTGGCGCAGGTGCTTCCGGTTCTGCTGCTAGHA-cassette rev1.............ATGACAGATCGCGGCCGCATCATTGCTAGTGCTAGAAGCGTAATCHA-cassette for2 .............ATCAGACATCACTAGTGCTAGATCCGGTTCTGCTGCTAGHA-cassette rev2.............ATGAGACATCACTAGTAGCGCCTGCACCAGCGCCGCTAGTGCTAGAAGCGTAATCCHT2 T1 for ...................GTCTTTTAAATCATTATTAGCTGCTGCAGTTGTAGCATCATCAGCTTTGGCCAGTGCCTCTAATCAAGTTG
CTCTTTACTGGGGTCAAAATGGTGCTGGTGGTGCTGGCGCAGGTGCTCHT2 T1 rev ...................CAAAATTAACGTTCAATGGATCTGGAAACAAATTCAAGAATGACAATAAAACAATGTCAACGTCAGTTT
CTTGACAGTATTGTGCCAATCTTTCTTGACCAGCGCCTGCACCAGCGCCYWP1 T1 for...................AAAACAATTAGAAGCTAATTCTACTGCTGCTATTAGAAGAAGATTAATGGGTGAAACTCCAATTGTTAA
AAGAGATCAAATTGATGATTTTATTGCTTCTGGTGCTGGCGCAGGTGCTYWP1 T1 rev ..................GAGAAGCAGAACCCGATGGGGAAGCAGAACTTGAACCTGGAACGTAATCAACAACTTCAAGAGTAGAA
CCTTCAAGAGCAGTACCTTCAGTATTTTCAATAGCGCCTGCACCAGCGCCRBT5 T1 for....................TAATACTACAATGCTCGCCTTATCCTTATTGTCAATCGTTTCCATTGCTTCAGCTGCTGGTGTCACTGCTA
TCCCAGAAGGTGATAATCCATACACTATTGGTGCTGGCGCAGGTGCTRBT5 T1 rev ...................TAGTACTTTGCTTGACACATTCTTTGGCACATTCTGGCAATTGGTCGTAGATTCTGTCAGCGAAACCGTT
GATGGAAGCGGTTTTAGCAACACTTGGGAAAGCGCCTGCACCAGCGCCECM33 T1 for.................CAAAACAGAATAATTGAATCAACAACAGCAATAACAACCCATGGTTTAATTTCAATATACTAACAGCTTA
TTATTAGACTCAACTTTGACTACTGCTACTGGTGCTGGCGCAGGTGCTECM33 T1 rev ................TGATGTCACCAACAGCAGTGGCACAAGCAGCAACTTGTTGAACTTGGTTGGAAGCAGTAGCGGTGAAAT
CTTTGAATGAACATTCAGATTTGACACTTGGAGCGCCTGCACCAGCGCCPIR1 T1 for .....................CAATAACAATTAAATCATGAAGTATTCTACACTTGTTAGTATTGCTGCTTTTATTAGCACCTCTTTAGCTG
CTACTGTTCCTGATGAACATTATTCAACAGGTGCTGGCGCAGGTGCTPIR1 T1 rev.....................TTGAAAGAGCAGAAGCTGATTCAACAGTAACAACTTGAATACCAAAAGTACCACTGAAATCTGTACTAG
CACCAGATGGAATTTTAGCTGAAGGTGATAAAGCGCCTGCACCAGCGCC
a Restriction sites are marked in bold, the (GA)6 linker is in italics, and the 6�HA-tag-homologous sequences (in HA cassette primers) and cassette-homologoussequences are underlined.
100 SCHILD ET AL. EUKARYOT. CELL
on January 4, 2019 by guesthttp://ec.asm
.org/D
ownloaded from

Rbt5-HA-expressing strains were supplemented with 500 �M iron chelatorbathophenanthroline disulfonic acid disodium salt (BPS; Sigma-Aldrich) to in-duce Rbt5 expression by iron starvation (69). Subsequently, cells were removedfrom the culture medium by centrifugation, and proteins were precipitated out of50-ml supernatants with methanol-chloroform (70). For comparison of wt andmutant samples, equal amounts (30 to 50 �g) of precipitated proteins weresubjected to de-N-glycosylation by incubation with 100 U N-glycosidase F(PNGaseF; New England Biolabs) according to the manufacturer’s instructions.Alternatively, 20 to 40 �g protein was digested with 0.7 �g rSap9 or 8.6 �grSap10 recombinant proteases or buffer only for 2 h at 37°C. Enzymaticallytreated supernatant proteins were separated using sodium dodecyl sulfate-poly-acrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidenedifluoride membranes (Millipore). Membranes were blocked with 5% (wt/vol)milk powder and probed with a rat anti-HA high-affinity antibody (clone 3F10;Roche Applied Science) and a goat anti-rat IgG-horseradish peroxidase (HRP)(Santa Cruz Biotechnology) secondary antibody. Signals were visualized withhighly sensitive enhanced chemiluminescence (ECL) Western blotting detectionreagents (GE Healthcare or Pierce) according to the manufacturer’s instructions.At least three independent experiments were performed.
Fluorescence microscopy analysis. For immunofluorescence staining of HAepitope-tagged proteins, C. albicans cells from logarithmic liquid cultures werefixed in 4% paraformaldehyde, blocked with 5% (wt/vol) bovine serum albumin,and incubated with a rat anti-HA high-affinity antibody (clone 3F10; RocheApplied Science) and the secondary antibody anti-rat IgG-Alexa Fluor 488(Molecular Probes). Stained C. albicans cells were visualized with epifluores-cence (Leica DM 5500B microscope and Leica DFC 350 FX camera), andimages were processed using the Adobe Photoshop software.
Measurement of chitinase activity. Endochitinase activity was determined witha fluorescence-based assay as previously described (63), with minor modifications.Briefly, the cell-bound activity of 2 � 107 phosphate-buffered saline (PBS)-washedC. albicans cells precultured on YPD agar plates was assayed with 50 �M substrate4-methylumbelliferyl-�-D-N-triacetylchitotrioside (4-MU-[GlcNAc]3; Sigma Al-drich). Fluorescence at 340-nm excitation/465-nm emission and an OD600 weremonitored in a microplate reader at the reaction start and endpoint (30 min). Inparallel, the OD600 was monitored. The relative chitinase activity of C. albicansmutants was calculated as a percentage of wt values, normalized against the OD600.
Macrophage assays. The human acute monocytic leukemia cell line (THP-1) wasmaintained in RPMI 1640 supplemented with 10% fetal bovine serum (PAA) at37°C, 5% CO2. Monocytes were differentiated to macrophages by overnight incu-bation with 10 ng/ml phorbol 12-myristate 13-acetate (PMA; Enzo Life Sciences).Killing and phagocytosis assays were performed with 2 � 105 differentiated macro-phages in 24-well polystyrene plates with RPMI 1640 without serum.
C. albicans killing by macrophages was assessed using a modified endpointdilution assay (58). C. albicans cells precultured on YPD agar plates (150 cells)were coincubated with macrophages for 2 days. The percentage of killing wascalculated by comparing the numbers of C. albicans CFU in wells with andwithout macrophages.
To analyze phagocytosis of C. albicans, macrophages grown on glass coverslips,were infected with C. albicans in a fungus-to-macrophage ratio of 2:1. After 1.5 hof coincubation, unbound fungal cells were removed, and cells were fixed with4% paraformaldehyde. Noningested macrophage-associated fungal cells werestained with a rabbit anti-C. albicans polyclonal antibody (Acris Antibodies) anda secondary anti-rabbit IgG-Alexa Fluor 488 antibody (Molecular Probes). Mac-rophage cells were then permeabilized in 0.5% (vol/vol) Triton X-100, beforeingested fungal cells and macrophage-associated cells were stained with 35 �g/mlcalcofluor white (Sigma Aldrich). Coverslips were then mounted onto slides, andstained cells were visualized with epifluorescence (see above). The percentage ofphagocytosed C. albicans cells was determined by relating the number of ingestedfungal cells to the total number of stained cells. At least 200 fungal cells werecounted on each coverslip.
Statistical analysis. Mean values and standard deviations (error bars) werecalculated from at least three independent experiments. The heteroscedasticStudent t test was used for the significance calculation of two groups. P valuesof �0.05 were considered significant differences.
RESULTS
Recombinant Sap9 and Sap10 exhibit an unusually broadpH spectrum, with activities above neutral pH and an incom-plete inhibition by pepstatin A. Secreted Sap proteases displaytypical features of aspartic proteases, including acidic pH op-
tima (“acid proteases”), a bell-shaped (Gaussian) pH depen-dence of activity (55), and sensitivity to the inhibitor pepstatinA (43). To test whether Sap9 and Sap10 follow this prototypicscheme, we measured the proteolytic activities of recombinantSap9 and Sap10 (rSap9 and rSap10) against the fluorescence-quenched substrate casein and compared these to Sap2 andSap6 (representing the two main subfamilies of Sap1 to 3 andSap4 to 6). The activity of recombinant Sap2 (rSap2) washighest at pH 3 or 4, with significant reduction at pH 6 to 8, asexpected from previous studies (4, 31, 59) (Fig. 1A). Recom-binant Sap6 (rSap6) showed a narrow pH optimum at pH 6,with activity sharply declining at pH values higher than pH 6 orlower than pH 5 (Fig. 1B), confirming and extending previousdata (4). Strikingly, rSap9 and rSap10 exhibited high activityover a broad pH range, even above neutral pH (pH 5 to 8).Both proteases displayed a pH optimum between pH 6 and pH7 and showed a significantly reduced activity at lower acidic pHvalues (pH 3 or 4) (Fig. 1C and D).
As predicted, activity of rSap2 and rSap6 was almost com-pletely inhibited by the addition of the potent aspartic proteaseinhibitor pepstatin A (20 �M) at all tested pH values (Fig. 1Aand B). In contrast, pepstatin A only partially inhibited rSap9and rSap10 (Fig. 1C and D), and high residual activity wasobserved. The residual rSap9, but not rSap10, activity could befurther reduced at higher pepstatin A concentrations, with40% and 27% residual activity at pH 6 when applying 50 and100 �M pepstatin A, respectively. These data demonstrate thatalthough Sap9 and Sap10 do possess characteristics of asparticproteases, they also exhibit distinct features, such as decreasedsensitivity to pepstatin A and near-neutral pH optima.
Sap9 and Sap10 cleavage preference for basic or dibasicresidues is not exclusive. In a previous study on a limited set ofsynthetic peptides we showed that rSap9 and rSap10 cleave atbasic lysine or arginine (K or R) or dibasic residues, similar tothe S. cerevisiae yapsins, and that rSap10 cleavage can also
FIG. 1. pH dependence and pepstatin A inhibition of rSap proteo-lytic activity against BODIPY FL fluorescence-quenched casein. Val-ues show percentage (%) of the activity at the pH optimum withoutinhibitor addition (100%). Black bars, no inhibitor added; gray bars, 20�M aspartic protease inhibitor pepstatin A added. (A) rSap2;(B) rSap6; (C) rSap9; (D) rSap10.
VOL. 10, 2011 C. ALBICANS Sap9 AND Sap10 CLEAVE CELL WALL PROTEINS 101
on January 4, 2019 by guesthttp://ec.asm
.org/D
ownloaded from

occur at yapsin-atypical amino acids F and H (1, 32, 51). Inorder to gain more detailed knowledge of Sap9 and Sap10substrate cleavage sites, we applied a systematic screen, digest-ing a set of 360 fluorescence-quenched peptides with rSap9 orrSap10 (protease substrate set; JPT Peptide Technologies).The peptides represented the cleavage sequences of a large setof known protease substrates. Sequences of peptides cleavedby rSap9 and rSap10 are listed in Table 3.
Out of the 360 peptides tested, almost 25% were processedexclusively by rSap9, only a minor fraction of peptides wascleaved exclusively by rSap10, and an overlapping fraction of31 peptides was processed by both proteases (Fig. 2A). Pep-tides with dibasic residues were clearly enriched amongcleaved peptides, with 45% among rSap9-cleaved peptides (51out of 113) and 41% among rSap10-cleaved peptides (16 out of39) compared to 23% in the whole protease substrate set (82out of 360). However, 29 dibasic residue-containing peptideswere not processed by rSap9 or rSap10 (Fig. 2A). This could bedue to the influence of residues surrounding basic amino acidson cleavage preferences, as certain amino acids were enrichedat these positions among cleaved peptides (Fig. 2B; summa-rized in Fig. 2C). As expected, basic residues were in theenriched fraction, but other residues were also present. Hy-drophobic amino acids (A, I, L, Y, and V) were enrichedN-terminal to K and R residues among rSap10 cleaved pep-tides and were also present in the rSap9-cleaved fraction. C-terminal to basic amino acids, acidic amino acids seem to betolerated, as D and E were enriched in rSap9-cleaved peptidesand were also present in the rSap10-cleaved fraction. Thus,basic or dibasic residues are common but not unique amongcleaved peptides, and residues neighboring basic amino acidshave an impact on cleavage. Eighteen rSap9-cleaved peptidesand four rSap10-cleaved peptides contained no basic residues.Consequently, cleavage at amino acids other than lysine or
arginine must have occurred not only by rSap10 (1) but also byrSap9.
A pattern-matching search of the cleaved peptides againstC. albicans-translated ORF sequences using the CandidaGenome Database (http://www.candidagenome.org) generatedno hits. However, one of the peptides cleaved by rSap9 andrSap10, AKRAASQI, represents a sequence present in the S.cerevisiae covalently linked cell wall protein (CWP) ScPir2,which is important for ScPir2 propeptide cleavage by Kex2(44). Interestingly, the C. albicans homolog CaPir1, possessesdibasic residues at a similar position in the protein sequence(N74KK) (see Fig. 5), suggesting that this protein is a potentialSap9 and Sap10 substrate in C. albicans.
Minor influence of Sap9 and Sap10 on the soluble cell wallproteome. Localization of Sap9 and Sap10 at the C. albicanscell surface suggested that both proteases cleave fungal CWPs.We therefore examined the influence of Sap9 and Sap10 cleav-age events on noncovalently linked SDS-soluble CWPs by com-paring soluble CWP profiles of the wild type (wt) with those ofa sap9� sap10� double mutant. Cell surface proteins that havebeen labeled with a membrane-impermeable biotin derivativewere solubilized using detergents, affinity purified, and sepa-rated via 2D-PAGE (for gel images and a list of proteinsidentified by mass spectrometry see Fig. S1 and Table S1 in thesupplemental material).
Comparison of protein patterns of the protease double mu-tant and wt demonstrated high similarity. After the matchingof 318 spots and quantification of spot volume, only sevenprotein spots with low abundance were shown to be differen-tially regulated, with one protein upregulated and six proteinsdownregulated in the sap9� sap10� mutant. One protein spotwith less abundance was identified as the aldehyde dehydro-genase Ald5, a protein previously detected in cytosolic proteinfractions (5) but also among soluble CWPs (8). Other proteins
TABLE 3. Sequences of peptides of the protease substrate set cleaved by Sap9 or Sap10
Protease usedfor cleavage Peptide sequencea
Sap9 1b 14 27 40 53 66 79 92 105GKKRDMSS RVRRDVEQ RARRELAP AEPDYGAL GKRSEQDA FANEFAEG PQRFGRNT RAKRSVHF EKQRIIGGKTRREAED AATRRQAV RKRRAVLT MSPKALQQ ALADSLGK QSPRSFQK DPQSIRSQ GPLGIAGI GLQRALEIVKRRVQDV RGARARLG ARRHSDGT KTPRVVGG EAGRNAQV TEPKVKLP DPLEFKSH KELGLGRH DDRDSANGEFKRELTG RQRYGKRS GGKRAMSD RHQREITR TEIDGRSI ELPTVSQE RDAQAHPG ETLDMMKK YYRADDANRTKREAGA QRYGKRDG KGANKKGL RSRREADK AEIRATSE RARRDAGP TVADGLKK PGLKIPKE EIRAGEKRSEVKMDAE RVKRYRQS KPVGKKRR ILRDKDNT EVKMDAEF AQRDSHLG GLARSNLD DSLDGRSL LDRRGIQRFMKKDAEE HLADSPAV APQRFGKK RRRRELDP ERKKSEHP DLRDDKDT REEMGKGY SSLDAREV DDSDAATFMGKKSTGE YVKASELP TQFDAAHP RKRRAPLA RSKRALKQ LERKEVLV DEEDILSH RKLDNTKF DYSLVAPPRNKRAVQG RVKRQVRS VSKRSADD LDRKEVLG RRKRSTRE KESDLSHV SKGRSLIG IVLDGTDN DEVDGMAGEEMDFRSG IQPFATNG LEKKSPLG LLSRSEED RVKRRVKR DGVDLKTQ DRHDSGLD DEMDEKSEPGPKRGTE RHQRELSA RKKRSTKK LKSNEIPD RTKRDYDS LQKRGIVE RAIDALRE DLPDMKETVHRDMAAR VMRDPASK RYMRADEA DTVDGKEI SLLDELES AAGVAPLS RNKRSKSN LSPDLLTLAKRAASQI AKRRTKRD VEMDSLSE VARRKLPT SPMGESAV MGKKSLEP RKKRSTSA MTPRSRGS
Sap10 1b 6 11 16 21 26 31 36RVKRQVRS IQPFATNG LKSNEIPD DPLEFKSH VEMDSLSE DTVDGKEI MTAKQFRK MSPKALQQYYRADDAN VKRRVQDV VARRKLPT VSKRSADD IEPRSFSQ ELQRSANS TEIDGRSI GLQRALEIRNKRAVQG EFKRELTG FMKKDAEE RSKRALKQ RKRRAVLT LQKRGIVE RVKRRVKR AKRAASQIGGKRAMSD YVKASELP RVKRYRQS EPLDLASY LERKEVLV GPYELKAL RGLTSLAD DGVDLKTQAEIRATSE VHRDMAAR HPSAFSEA RYMRADEA SEVKMDAE DPQSIRSQ QSPRSFQK
a Peptides cleaved by both Sap9 and Sap10 are underlined.b Lists start with peptides with the highest cleavage efficiency.
102 SCHILD ET AL. EUKARYOT. CELL
on January 4, 2019 by guesthttp://ec.asm
.org/D
ownloaded from

characteristic of the soluble cell surface proteome (8, 36, 54)were found in similar patterns in the wt and sap9� sap10�mutant: Atp2, Ssb1, Pdc11, Rpp0, Eno1, Eft2, Cdc19, Atp1,Tdh3, Fba1, and Gpm1. Consequently, the lack of Sap9 andSap10 had only a minor influence on soluble CWPs.
Similarly, deletion of SAP9 and SAP10 appears to have aminor influence on the presence and migration of major se-creted proteins, as suggested from 2D-PAGE experiments anddetermination of protein concentration in culture supernatantsof the protease double mutant (data not shown).
Recombinant Sap9 and Sap10 proteases cleave GPI-CWPson isolated C. albicans cell walls. We next applied a morespecific approach, monitoring cleavage events by rSap9 andrSap10 targeting covalently linked CWPs. Isolated cell walls ofthe sap9� sap10� mutant were incubated with rSap9 andrSap10 proteases. Peptides released from cell walls after diges-tion were analyzed by LC-MS/MS.
Incubation with rSap9 and rSap10 specifically liberated pro-tein species from cell walls, while no peptides were detected insamples without protease treatment. Identified proteins be-longed to the class of covalently linked GPI-modified cell wall
proteins (GPI-CWPs), with functions from cell wall remodel-ing (Cht2, Ecm33, Pga4 and Rhd3) (14, 37, 39, 56) to host cellinteraction and virulence (Ywp1, Als2, Ecm33, and Rhd3) (14,24, 37, 64) and iron metabolism (Rbt5) (69) (Table 4). Cellwalls were isolated from logarithmic- or stationary-phase cul-tures grown at pH 4 or pH 6.5. Release of peptides afterprotease digest varied mainly with the growth phase ratherthan with the culture pH. These data provide the first in vitroevidence of proteolytic targets of Sap9 and Sap10 among co-valently linked CWPs. This reinforces the view that Sap9 andSap10 activity may regulate CWP functions and supports ourearlier observations that deletion of SAP9 and SAP10 influ-ences cell wall integrity and adhesion to host cells (1).
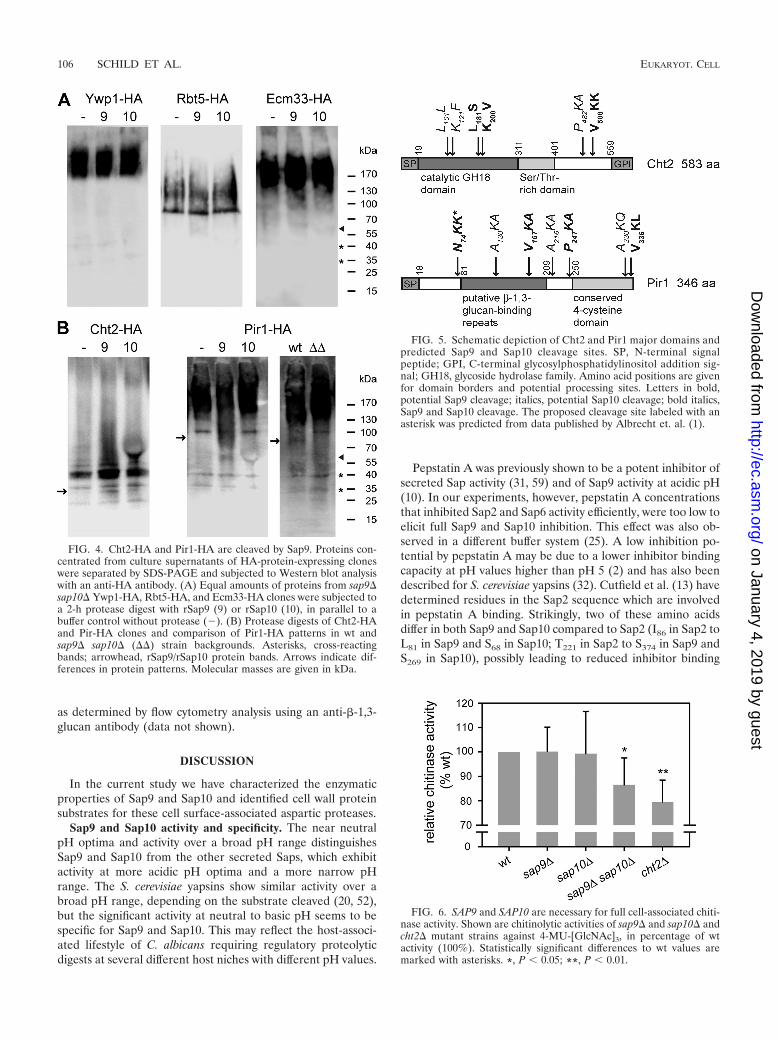
Processing of epitope-tagged Cht2 and Pir1 by Sap9. Tofurther characterize Sap9 and Sap10 cleavage events on CWPs,we integrated the small internal hemagglutinin epitope tag(HA) into the protein sequences of the previously identifiedproteolytic targets Cht2, Ywp1, Rbt5, and Ecm33 (Fig. 3A).Pir1, a non-GPI-CWP with a proposed role in �-1,3-glucancross-linking (28) was also included for analysis because pep-tide cleavage data had suggested processing of this CWP by
FIG. 2. Systematic determination of rSap9 and rSap10 cleavage preferences using a set of synthetic peptides (protease substrate set; JPTPeptide Technologies). (A) Venn diagram indicates, out of the total 360 peptides, the number of rSap9-cleaved peptides in comparison torSap10-cleaved peptides and dibasic residue-containing peptides (KR, KK, RR, and RK). (B) Frequency of residues N-terminal and C-terminalto basic amino acids in rSap9- or rSap10-cleaved peptides. Sequences of the total 360 peptides (whole set) were compared to the subset of rSap9-and rSap10-cleaved peptides, to analyze amino acid preferences for cleavage. Residues with a relative frequency that was �2% higher in thecleaved subset than that of the whole peptide set are marked with a “#” (rSap9-cleaved peptides) or an asterisk (rSap10-cleaved peptides).(C) Overview of enriched amino acids according to data shown in panel B).
VOL. 10, 2011 C. ALBICANS Sap9 AND Sap10 CLEAVE CELL WALL PROTEINS 103
on January 4, 2019 by guesthttp://ec.asm
.org/D
ownloaded from

Sap9 and Sap10 (see above). The HA tag was inserted close tothe protein N terminus, leaving protein N- and C-terminalregions unmodified to ensure proper secretion and cell surfacelocalization. This approach allows the detection of HA-taggedmature proteins or N-terminal cleavage fragments with anepitope-specific antibody (Fig. 3B) and was used for compar-ison of epitope-tagged protein characteristics in wt and sap9�sap10� double mutant strain backgrounds.
Successful epitope tagging and surface localization of taggedproteins was confirmed by immunofluorescence staining withan HA-specific antibody (Fig. 3C). The presence of HA-taggedfragments in C. albicans culture supernatants was then exam-ined by Western blot analysis. All tested proteins were not onlypresent on the cell surface (Fig. 3C) but were also partlyreleased into the extracellular space (Fig. 4A). The presence ofwall-related proteins in the secretome of C. albicans is notuncommon, as has been shown previously (34, 65, 66). Ywp1,for example, has been detected in culture supernatants of sta-tionary-phase cells (24). We therefore used proteins concen-trated from culture supernatants for Western blot analyses(Fig. 4A), avoiding further isolation steps for protein liberationfrom the cell wall. Surprisingly, apart from Pir1-HA (see be-low), none of the HA-epitope-tagged proteins showed differ-ences in protein migration in strains lacking SAP9 and SAP10compared with the wt (data not shown). We therefore directlysubjected sap9� sap10� supernatant proteins to digestion withrSap9 or rSap10.
Proteolytic digestion with recombinant rSaps did not re-lease defined cleavage fragments from Ywp1-HA, Rbt5-HA,or Ecm33-HA (Fig. 4A). In contrast, the Cht2-HA proteinpattern changed after protease digestion: a 30-kDa fragmentwas enriched in the rSap9-digested sample and also slightly inthe rSap10-digested sample. In addition, several distinct bandsin the molecular weight range of 25 to 45 kDa were detected(Fig. 4B). A large portion of Pir1-HA migrated with a high mo-lecular mass (180 kDa) (35). Proteolytic digestion with rSap9reduced the amount of a distinct 100-kDa band and gener-ated a slightly smaller fragment (Fig. 4B). Interestingly, wtsamples showed an 90-kDa band, while an 100-kDa bandwas present in protease mutant samples (Fig. 4B). The ob-served differences in patterns of Cht2-HA and Pir1-HA furthersuggested that these two CWPs are in vitro proteolytic targetsof Sap9 and that Cht2 may also constitute a Sap10 target.
In silico prediction of Sap9 and Sap10 cleavage sites in Cht2and Pir1. In order to assign cleavage fragments detected inanti-HA-Western blot analysis to potential cleavage sites, weanalyzed the Cht2 and Pir1 protein sequences for the presenceof the following sequence motifs: (i) a basic residue flanked bytwo amino acids that were found enriched when comparingrSap9 and rSap10 cleaved peptides with noncleaved peptidesof the protease substrate set (Fig. 2B and C); (ii) a three-aminoacid motif identically present in rSap9 and rSap10 cleavedsynthetic peptides (Table 3); (iii) a three-amino acid motifidentified during previous Sap9 and Sap10 peptide cleavage
TABLE 4. Mass spectrometric analysis of GPI-CWPs released from isolated cell walls after digestion with Sap9 or Sap10
Protein Descriptiona Identified peptide (1*sequence*�1) (position)
pH at whichprotein
released by:b
Sap9 Sap10
Logarithmic-phase cellwalls
Cht2 Chitinase; required for normal filamentousgrowth
L*LSLGGGVGDYGFSDVASATK*F(102–121)
4; 6.5
Cht2 L*SLGGGVGDYGFSDVASATK*F(103–121)
4; 6.5
Cht2 L*SAAPQCPYPDASLGDLLSK*V(182–200)
4; 6.5
Cht2 S*AAPQCPYPDASLGDLLSK*V(183–200)
4
Ywp1 Cell wall and secreted protein with suggestedrole in dispersal in host
R*DQIDDFIASIENTEGTALEGSTL*E(134–156)
4
Als2 (or Als9)c ALS family protein; role in adhesion, biofilmformation, germ tube induction
I*TGVFNSFDSLTWTR*S (21–34) 4
Als10d ALS family protein F*NVGGTGSLVDLESSK*C (135–149) 6.5
Stationary-phase cellwalls
Rhd3/Pga29 Unknown function; contributes to virulence TISSIQLFAK*S (16–25)e 4 4Rbt5 (or Pga10, Csa1) Hemoglobin acquisition R*IYDQLPECAK*E (47–56) 6.5 6.5Rbt5 (or Csa2) K*QSTSSTPCPYWDTGCL*C (61–76) 4Rbt5 L*CVMPQFAGAVGNCVAK*N (77–92) 4; 6.5Rbt5 Q*FAGAVGNCVAK*N (82–92) 4; 6.5Ecm33 Important for cell wall integrity K*TIGGALQISDNSELR*S (276–290) 4Pga4 Transglucosidase K*AGIYVILDVNTPHSSITR*S (99–116) 6.5
a According to Candida Genome Database (http://www.candidagenome.org/).b Cell wall preparations from logarithmic or stationary cultures at pH 4/pH 6.5. Boldface indicates peptides that were identified without trypsin digestion prior to
LC-MS/MS analysis. All peptides shown have Mascot P values of �0.05.c Als2 but not Als9 has previously been found in cell walls (14).d orf19.2355/orf19.9891 deleted from Candida Genome Database (http://www.candidagenome.org).e N terminus of mature protein.
104 SCHILD ET AL. EUKARYOT. CELL
on January 4, 2019 by guesthttp://ec.asm
.org/D
ownloaded from

assays (1); and (iv) N- and C-terminal two-amino acid motifs ofpeptides released by Sap9 or Sap10 from isolated cell wallswithout tryptic digestion, which likely represent specific cleav-age sites (Table 4). This analysis led to the prediction of sixCht2 and seven Pir1 cleavage sites (Fig. 5). Four predictedcleavage sites are located in the catalytic glycoside hydrolase(GH18) domain of Cht2. Cleavage at one of these positionscould have generated the 30-kDa Cht2-HA fragment (Fig.4B). Further, two cleavage sites are located close to the GPIaddition region of Cht2. Interestingly, two of the potentialSap9 and Sap10 cleavage sites of Pir1 lie between repeats thatare possibly involved in �-1,3-glucan-binding and cross-linkingof glucan chains by Pir1 (18, 30). The 90-kDa Pir1-HA frag-ment (Fig. 4B) may have been generated by processing at oneof these sites. However, the prediction of Pir1 cleavage sitesbased on the molecular weight is hindered by the fact that thisprotein is highly O glycosylated (35). In addition, it is possiblethat Pir1 is processed at N74KK by Sap9 and Sap10 (seeabove).
Chitinase activity is impaired in a sap9� sap10� mutant.Together, our data demonstrated in vitro cleavage of CWPs bySap9 and Sap10. To evaluate if such cleavage may affect func-tions of CWPs in living C. albicans cells, we applied activityassays testing the function of two potential protease substratesin sap9� and sap10� single and double mutant strains. First,we examined cell-associated chitinase activity with a fluores-cence-based enzymatic assay (38). The sap9� sap10� doublemutant, but not the sap9� or sap10� single mutants, exhibitedsignificantly decreased chitinase activity, with 13.5% reductioncompared to that of the wt (Fig. 6). This reduction was similarto that of a cht2� deletion mutant, which also showed a smallbut significant reduction in activity, as described previously(63).
Second, we tested the function of Rbt5. This protein isnecessary for utilization of iron from hemoglobin. While anrbt5� mutant exhibited reduced growth with hemoglobin as thesole iron source (69), single and double protease mutants grewas well as the wt under these conditions (data not shown),indicating that Rbt5 function is not impaired in the absence ofSAP9 and SAP10. Consequently, Sap9 and Sap10 in combina-tion are necessary for full chitinase activity but not for Rbt5function.
Sap9 and Sap10 functions are dispensable for survival dur-ing interaction with macrophages. The importance of Sap9and Sap10 for cell wall integrity and an intact cell wall structure(1) suggested that these proteases might influence the recog-nition of C. albicans by macrophages or fungal survival afterphagocytosis. To test this hypothesis, we analyzed the interac-tion of C. albicans wt and protease mutants with human mac-rophages, using the monocytic cell line THP-1. Uptake offungal cells into macrophages as well as survival after coincu-bation with macrophages were quantified. Neither uptake norsurvival rates were altered for sap9� and sap10� single anddouble mutants compared with the wt (phagocytosis: wt, 52.9%[12.9], sap9� sap10� mutant, 51.2% [17.4]; killing: wt,35.4% [11.6], sap9� sap10� mutant, 35.7% [5.1]). Proteasemutants were able to form wt-like filaments inside macro-phages and to escape from macrophages like the wt. Similarresults were obtained with primary human blood monocyte-derived macrophages (data not shown).
Thus, cell surface changes in the absence of SAP9 andSAP10 do not seem to cause major alterations in cell surfaceimmunogenicity. This observation is further supported by thefinding that accessibility of �-1,3-glucan, a molecule importantfor immune recognition of C. albicans by macrophages (48),was similar in wt and sap9� sap10� single or double mutants,
FIG. 3. Internal hemagglutinin (HA) epitope tagging of potential protease substrates. (A) Two-step PCR-based epitope-tagging procedure asused in this study. Tagging constructs with URA3 marker flanked by two direct repeats of the HA epitope were introduced into the target geneopen reading frames. Growth on FOA (5-fluoroorotic acid) selected for clones in which recombination between the HA epitope repeats hadgenerated one intact HA epitope tag. (B) GPI-CWP with an internal HA epitope tag close to the N terminus and a possible cleavage product. SP,N-terminal signal peptide for secretion; GPI, C-terminal glycosylphosphatidylinositol addition signal; asterisks, serine- and threonine-enrichedregion, often highly O glycosylated. (C) HA-epitope-tagged CWPs localize to the cell surface as shown by immunofluorescence staining with ananti-HA antibody for wt and sap9� sap10� (��) HA clones. Bars, 10 �m.
VOL. 10, 2011 C. ALBICANS Sap9 AND Sap10 CLEAVE CELL WALL PROTEINS 105
on January 4, 2019 by guesthttp://ec.asm
.org/D
ownloaded from
as determined by flow cytometry analysis using an anti-�-1,3-glucan antibody (data not shown).
DISCUSSION
In the current study we have characterized the enzymaticproperties of Sap9 and Sap10 and identified cell wall proteinsubstrates for these cell surface-associated aspartic proteases.
Sap9 and Sap10 activity and specificity. The near neutralpH optima and activity over a broad pH range distinguishesSap9 and Sap10 from the other secreted Saps, which exhibitactivity at more acidic pH optima and a more narrow pHrange. The S. cerevisiae yapsins show similar activity over abroad pH range, depending on the substrate cleaved (20, 52),but the significant activity at neutral to basic pH seems to bespecific for Sap9 and Sap10. This may reflect the host-associ-ated lifestyle of C. albicans requiring regulatory proteolyticdigests at several different host niches with different pH values.
Pepstatin A was previously shown to be a potent inhibitor ofsecreted Sap activity (31, 59) and of Sap9 activity at acidic pH(10). In our experiments, however, pepstatin A concentrationsthat inhibited Sap2 and Sap6 activity efficiently, were too low toelicit full Sap9 and Sap10 inhibition. This effect was also ob-served in a different buffer system (25). A low inhibition po-tential by pepstatin A may be due to a lower inhibitor bindingcapacity at pH values higher than pH 5 (2) and has also beendescribed for S. cerevisiae yapsins (32). Cutfield et al. (13) havedetermined residues in the Sap2 sequence which are involvedin pepstatin A binding. Strikingly, two of these amino acidsdiffer in both Sap9 and Sap10 compared to Sap2 (I86 in Sap2 toL81 in Sap9 and S68 in Sap10; T221 in Sap2 to S374 in Sap9 andS269 in Sap10), possibly leading to reduced inhibitor binding
FIG. 4. Cht2-HA and Pir1-HA are cleaved by Sap9. Proteins con-centrated from culture supernatants of HA-protein-expressing cloneswere separated by SDS-PAGE and subjected to Western blot analysiswith an anti-HA antibody. (A) Equal amounts of proteins from sap9�sap10� Ywp1-HA, Rbt5-HA, and Ecm33-HA clones were subjected toa 2-h protease digest with rSap9 (9) or rSap10 (10), in parallel to abuffer control without protease (�). (B) Protease digests of Cht2-HAand Pir-HA clones and comparison of Pir1-HA patterns in wt andsap9� sap10� (��) strain backgrounds. Asterisks, cross-reactingbands; arrowhead, rSap9/rSap10 protein bands. Arrows indicate dif-ferences in protein patterns. Molecular masses are given in kDa.
FIG. 5. Schematic depiction of Cht2 and Pir1 major domains andpredicted Sap9 and Sap10 cleavage sites. SP, N-terminal signalpeptide; GPI, C-terminal glycosylphosphatidylinositol addition sig-nal; GH18, glycoside hydrolase family. Amino acid positions are givenfor domain borders and potential processing sites. Letters in bold,potential Sap9 cleavage; italics, potential Sap10 cleavage; bold italics,Sap9 and Sap10 cleavage. The proposed cleavage site labeled with anasterisk was predicted from data published by Albrecht et. al. (1).
FIG. 6. SAP9 and SAP10 are necessary for full cell-associated chiti-nase activity. Shown are chitinolytic activities of sap9� and sap10� andcht2� mutant strains against 4-MU-[GlcNAc]3, in percentage of wtactivity (100%). Statistically significant differences to wt values aremarked with asterisks. *, P � 0.05; **, P � 0.01.
106 SCHILD ET AL. EUKARYOT. CELL
on January 4, 2019 by guesthttp://ec.asm
.org/D
ownloaded from

independent of the pH. Our data imply that experiments usingpepstatin A to address the importance of Saps for C. albicanspathogenicity should be interpreted with caution. For example,near-neutral pH values used in most cell culture systems couldallow substantial residual Sap9 and Sap10 activity despite pep-statin A treatment.
A systematic screen of synthetic peptides for Sap9 and Sap10cleavage confirmed our previous observation that both pro-teases prefer cleavage at basic lysine or arginine or dibasicresidues, similar to S. cerevisiae yapsins (1, 21). However, bothSap9 and Sap10 cleavage events also occurred independent ofbasic residues, an ability that seems to be specific for the C.albicans yapsin-like proteases. Furthermore, several peptidescontaining dibasic residues were not digested, suggesting thatthe presence of such sites alone is not a definite indication forcleavage. Thus, the generation of a cleavage consensus se-quence for systematic prediction of proteolytic substrates bysearching the C. albicans genome is not possible on the basis ofour data. Comparing Sap9 and Sap10 behavior, our cleavageassays suggest common as well as specific processing sites forthe two proteases. This confirms our previous observation thatSap9 can only partially substitute Sap10 and vice versa (1). Ofnote was the absence of certain amino acid combinationsamong Sap10-cleaved peptides (e.g., DK, DR, EK, TK, andTR) and the lower number of Sap10-cleaved peptides comparedto Sap9-cleaved peptides. This may indicate a more specific ac-tion of Sap10 compared to Sap9 but may also be due to a lowerspecific activity of our Sap10 preparation (see Materials andMethods). However, increasing the amount of Sap10 protein inthe protease substrate set assay did not significantly enhancethe number of processed peptides (data not shown), suggestingrather that Sap10 may indeed have a more limited number ofpotential substrates.
Sap9 and Sap10 in vitro substrates and suggested role ofprocessing. Cleavage experiments with isolated C. albicans cellwalls and synthetic peptides facilitated the identification ofpotential Sap9 and Sap10 substrates among previously de-scribed covalently linked C. albicans CWPs: Cht2, Ywp1, Als2,Rhd3, Rbt5, Ecm33, Pga4 and Pir1 (14, 15). Interestingly,mutants lacking these proteins showed sap9� sap10�-similarphenotypes, such as altered adhesion (ywp1� and als2� mu-tants), cell wall structural or integrity defects (rhd3�, ecm33�,and pir1� mutants), and cell separation defects (ecm33� andpir1� mutants) (14, 24, 35, 37, 64), suggesting a functionalconnection between these cell wall proteins and the two pro-teases. Surprisingly, of the eight potential substrates, it ap-peared that only Cht2-HA and Pir1-HA were cleaved by Sap9and Sap10 following epitope tagging. One reason for this un-expected result may be that cleavage sites, which were accessiblein isolated cell walls or synthetic peptides, were masked in livingfungal cells, e.g., due to glycosylation or protein folding. Mostof the tagged proteins were detected in Western blots as highlydiffuse bands, as expected for heavily N- and O-glycosylatedCWPs (30). Thus, slight molecular weight shifts of proteinbands may not have been detectable. Notable, epitope-taggedCWPs were detected not only on the cell surface, but also inthe culture medium. GPI-anchored proteins can be eithermembrane localized or covalently linked to the cell wall �-1,6-glucan after cleavage of the GPI anchor (28). As discussed byGranger et al. (24), an intermediate form, lacking the GPI
anchor may exist that could possibly diffuse into the me-dium. Noncovalently linked, soluble surface proteins alsorepresent possible proteolytic targets for Sap9 and Sap10.However, our experiments did not detect a major influenceof Sap9 and Sap10 on this class of proteins. The reducedabundance of the aldehyde dehydrogenase Ald5 in thesap9� sap10� mutant compared with the wt may rather bedue to secondary effects (5).
Cell wall integrity and cell separation defects of mutantslacking SAP9 and SAP10 (1), as well as upregulation of SAP9expression during cell wall stress (12) and cell wall regenera-tion (9), suggest that proteolytic cleavage has a direct impacton CWP functions. This is similar to S. cerevisiae, as yapsindeletion mutants develop cell wall integrity defects (33) andScYps1 was shown to cleave the cell wall remodeling CWPsScPir4 and ScGas1 (20). However, the consequences of pro-teolytic processing of distinct CWPs for their function remainsunclear. We propose three possible consequences of cleavage:activation, structural rearrangements, and shedding, based onthe following findings. Our assays revealed that C. albicanscell-bound chitinase activity is reduced in the absence of SAP9and SAP10. C. albicans possesses at least four chitinase-encod-ing genes, CHT1 to -4. Besides Cht2, Cht3 has been shownpreviously to contribute to yeast chitinase activity, while Cht1and Cht4 likely play a minor role (17, 63). Constitutive CHT3expression did not reconstitute wt-like chitinase activity in asap9� sap10� mutant (data not shown). Thus, a Cht2 defectrather than a dysfunction of Cht3 is likely to be the main causeof reduced chitinase activity in the sap9�sap10� mutant, al-though the influence of further factors cannot be ruled out.Therefore, a direct activating effect of Sap9 and Sap10 proteo-lytic cleavage on Cht2 function is proposed. A connectionbetween chitinase activity and proteolytic cleavage has alreadybeen described for a C. albicans microsomal zymogenic chiti-nase (16).
In silico analysis of the Pir1 sequence predicted cleavagesites between repeats that are likely involved in �-1,3-glucan-binding and cross-linking of glucan chains (18, 30). It is possi-ble that cleavage in this region alters the degree of glucancross-linking by Pir1, thus assigning Sap9 and Sap10 a functionin rearranging cell wall components. A third function of cleav-age could be the shedding of CWPs or CWP fragments into thesurrounding medium. Shedding of a substrate by Sap9 hasbeen suggested to induce chemotaxis of polymorphonuclearneutrophils toward C. albicans (26).
Depending on environmental conditions and fungal mor-phology, the cell wall proteome, and thus the proteins poten-tially exposed to Sap9 and Sap10, can change dramatically. Inaddition, target protein localization might be restricted to cer-tain cell surface compartments, such as bud sites (30). Theconstitutive expression of SAP9 and SAP10 under both yeast-and hypha-inducing growth conditions (47) and the apparentlyeven distribution of Sap9 and Sap10 over the C. albicans cellsurface (1) suggest that proteolytic activities are importantduring both yeast and hyphal growth and are not restricted tocertain cell surface components. The experiments in this studyfocused on yeast cells since the most prominent phenotype ofdeletion mutants, a cell separation defect, was observed foryeast cells, while no alteration in hyphal induction, growth, orappearance was evident.
VOL. 10, 2011 C. ALBICANS Sap9 AND Sap10 CLEAVE CELL WALL PROTEINS 107
on January 4, 2019 by guesthttp://ec.asm
.org/D
ownloaded from

Many of our experiments comparing the wt with the sap9�and/or sap10� mutants showed no differences in protein pat-terns or processing. It may be possible that lack of Sap9 and/orSap10 activity can be complemented by other proteases such asthe proprotein-converting serine protease Kex2 (3, 49). Asdiscussed by Albrecht et al. (1), Kex2, which uses processingsites similar to those of Sap9 and Sap10, is active in the trans-Golgi network and may process target proteins of Sap9 andSap10 that are transported to the surface via the secretorypathway. Two observations from the present study indicateoverlapping functions between Kex2 and Sap9 and Sap10.First, the Ywp1-fragment KR133*DQIDDFIASIENTEGTALEGSTL*E157 was detected following Sap9 digestion of isolatedcell walls. Interestingly, cleavage at KR133 has been suggestedto be responsible for generation of an alternative 12-kDaYwp1 propeptide in the absence of Kex2. In wt and sap9� andsap10� mutant cells, an 11-kDa propeptide is generated, likelyby processing at a triple arginine site, I120RRRLM, by Kex2(24; Bruce L. Granger, personal communication). Second, thesuggested Pir1 processing at N74KK by Sap9 and Sap10 maygenerate a Pir1 propeptide independent of Kex2.
Interaction with macrophages. Despite their importance forcell wall structure and integrity, Sap9 and Sap10 were shown tobe dispensable during interaction of C. albicans with humanmacrophages in this study. McKenzie et al. (40) suggest thatCWP defects alone have a minor impact on macrophagephagocytosis and killing, as seen for the CWP hwp1� and als3�mutants. Fungal yapsins, however, have the potential to influ-ence fungus-macrophage interactions, as shown for C. glabrata,in which mutants lacking yapsins exhibit reduced survival inmurine macrophages (29). Sap9 has recently been shown todegrade and inactivate the oral salivary antimicrobial peptidehistatin-5 (41), and further, yet undefined, protease targets onthe host side may be possible.
Conclusions. In summary, our data demonstrate that Sap9and Sap10 are distinct C. albicans aspartic proteases: theydiffer from other Sap members (i) in exhibiting enzymaticactivity over a broad range of pH, even at neutral (physiolog-ical) pH, (ii) in a limited inhibition potential by pepstatin A,and (iii) in distinct cleavage preferences. We also, for the firsttime, provide evidence for Sap9/Sap10 processing of a set of C.albicans cell wall proteins in vitro. Finally, the deletion of SAP9and SAP10 genes in C. albicans resulted in a reduction inchitinase activity similar to that upon deletion of the chitinase-encoding gene CHT2. This result indicates that Sap9/Sap10cleavage has the potential to directly influence cell wall proteinfunctions. It opens up perspectives for future experiments fo-cusing on the characterization of the particular consequence ofproteolytic cleavage for the functions of Cht2 and other cellwall protein targets.
ACKNOWLEDGMENTS
We acknowledge the financial support by the Deutsche Forschungs-gemeinschaft DFG project Hu 528/14-1.
We are grateful to Olaf Kniemeyer and Maria Poetsch (HKI) forsupporting 2D-PAGE analyses. We thank Frans Klis and Henk Dek-ker (Swammerdam Institute for Life Sciences, Netherlands) for helpfuldiscussions regarding potential Sap targets and support in LC-MS/MSanalyses, respectively, and Duncan Wilson (HKI) for critical reading ofthe manuscript. C. albicans cht2� and rbt5� strains were kindly pro-vided by Carol Munro (University of Aberdeen, United Kingdom) and
Daniel Kornitzer (Rappaport Institute for Research in the MedicalSciences, Haifa, Israel), and Sap-producing Pichia pastoris strains werekindly provided by Michel Monod (CHUV Lausanne, Switzerland).Furthermore, we thank Betty Wachtler, Katja Seider, and SaschaBrunke (HKI) for fruitful discussions.
REFERENCES
1. Albrecht, A., et al. 2006. Glycosylphosphatidylinositol-anchored proteases ofCandida albicans target proteins necessary for both cellular processes andhost-pathogen interactions. J. Biol. Chem. 281:688–694.
2. Backman, D., and U. H. Danielson. 2003. Kinetic and mechanistic analysis ofthe association and dissociation of inhibitors interacting with secreted aspar-tic acid proteases 1 and 2 from Candida albicans. Biochim. Biophys. Acta1646:184–195.
3. Bader, O., Y. Krauke, and B. Hube. 2008. Processing of predicted substratesof fungal Kex2 proteinases from Candida albicans, C. glabrata, Saccharomy-ces cerevisiae and Pichia pastoris. BMC Microbiol. 8:116.
4. Borg-von Zepelin, M., S. Beggah, K. Boggian, D. Sanglard, and M. Monod.1998. The expression of the secreted aspartyl proteinases Sap4 to Sap6 fromCandida albicans in murine macrophages. Mol. Microbiol. 28:543–554.
5. Brand, A., D. M. MacCallum, A. J. Brown, N. A. Gow, and F. C. Odds. 2004.Ectopic expression of URA3 can influence the virulence phenotypes andproteome of Candida albicans but can be overcome by targeted reintegrationof URA3 at the RPS10 locus. Eukaryot. Cell 3:900–909.
6. Braun, B. R., W. S. Head, M. X. Wang, and A. D. Johnson. 2000. Identifi-cation and characterization of TUP1-regulated genes in Candida albicans.Genetics 156:31–44.
7. Calderone, R. A., and W. A. Fonzi. 2001. Virulence factors of Candidaalbicans. Trends Microbiol. 9:327–335.
8. Castillo, L., et al. 2008. A study of the Candida albicans cell wall proteome.Proteomics 8:3871–3881.
9. Castillo, L., et al. 2006. Genomic response programs of Candida albicansfollowing protoplasting and regeneration. Fungal Genet. Biol. 43:124–134.
10. Cawley, N. X., et al. 2003. Synthesis and characterization of the first potentinhibitor of yapsin 1. Implications for the study of yapsin-like enzymes.J. Biol. Chem. 278:5523–5530.
11. Chaffin, W. L. 2008. Candida albicans cell wall proteins. Microbiol. Mol. Biol.Rev. 72:495–544.
12. Copping, V. M., et al. 2005. Exposure of Candida albicans to antifungalagents affects expression of SAP2 and SAP9 secreted proteinase genes. J.Antimicrob. Chemother. 55:645–654.
13. Cutfield, S. M., et al. 1995. The crystal structure of a major secreted asparticproteinase from Candida albicans in complexes with two inhibitors. Structure3:1261–1271.
14. de Boer, A. D., et al. 2010. The Candida albicans cell wall protein Rhd3/Pga29 is abundant in the yeast form and contributes to virulence. Yeast27:611–624.
15. de Groot, P. W. J., et al. 2004. Proteomic analysis of Candida albicans cellwalls reveals covalently bound carbohydrate-active enzymes and adhesins.Eukaryot. Cell 3:955–965.
16. Dickinson, K., V. Keer, C. A. Hitchcock, and D. J. Adams. 1991. Microsomalchitinase activity from Candida albicans. Biochim. Biophys. Acta 1073:177–182.
17. Dunkler, A., A. Walther, C. A. Specht, and J. Wendland. 2005. Candidaalbicans CHT3 encodes the functional homolog of the Cts1 chitinase ofSaccharomyces cerevisiae. Fungal Genet. Biol. 42:935–947.
18. Ecker, M., R. Deutzmann, L. Lehle, V. Mrsa, and W. Tanner. 2006. Pirproteins of Saccharomyces cerevisiae are attached to beta-1,3-glucan by a newprotein-carbohydrate linkage. J. Biol. Chem. 281:11523–11529.
19. Fonzi, W. A., and M. Y. Irwin. 1993. Isogenic strain construction and genemapping in Candida albicans. Genetics 134:717–728.
20. Gagnon-Arsenault, I., L. Parise, J. Tremblay, and Y. Bourbonnais. 2008.Activation mechanism, functional role and shedding of glycosylphosphatidy-linositol-anchored Yps1p at the Saccharomyces cerevisiae cell surface. Mol.Microbiol. 69:982–993.
21. Gagnon-Arsenault, I., J. Tremblay, and Y. Bourbonnais. 2006. Fungalyapsins and cell wall: a unique family of aspartic peptidases for a distinctivecellular function. FEMS Yeast Res. 6:966–978.
22. Gauss, R., M. Trautwein, T. Sommer, and A. Spang. 2005. New modules forthe repeated internal and N-terminal epitope tagging of genes in Saccharo-myces cerevisiae. Yeast 22:1–12.
23. Gillum, A. M., E. Y. Tsay, and D. R. Kirsch. 1984. Isolation of the Candidaalbicans gene for orotidine-5�-phosphate decarboxylase by complementationof S. cerevisiae ura3 and E. coli pyrF mutations. Mol. Gen. Genet. 198:179–182.
24. Granger, B. L., M. L. Flenniken, D. A. Davis, A. P. Mitchell, and J. E. Cutler.2005. Yeast wall protein 1 of Candida albicans. Microbiology 151:1631–1644.
25. Gropp, K., et al. 2009. The yeast Candida albicans evades human comple-ment attack by secretion of aspartic proteases. Mol. Immunol. 47:465–475.
26. Hornbach, A., et al. 2009. The glycosylphosphatidylinositol-anchored pro-
108 SCHILD ET AL. EUKARYOT. CELL
on January 4, 2019 by guesthttp://ec.asm
.org/D
ownloaded from

tease Sap9 modulates the interaction of Candida albicans with human neu-trophils. Infect. Immun. 77:5216–5224.
27. Hoyer, L. L., C. B. Green, S. H. Oh, and X. Zhao. 2008. Discovering thesecrets of the Candida albicans agglutinin-like sequence (ALS) gene fami-ly—a sticky pursuit. Med. Mycol. 46:1–15.
28. Kapteyn, J. C., et al. 2000. The cell wall architecture of Candida albicanswild-type cells and cell wall-defective mutants. Mol. Microbiol. 35:601–611.
29. Kaur, R., B. Ma, and B. P. Cormack. 2007. A family of glycosylphosphati-dylinositol-linked aspartyl proteases is required for virulence of Candidaglabrata. Proc. Natl. Acad. Sci. U. S. A. 104:7628–7633.
30. Klis, F. M., G. J. Sosinska, P. W. J. de Groot, and S. Brul. 2009. Covalentlylinked cell wall proteins of Candida albicans and their role in fitness andvirulence. FEMS Yeast Res. 9:1013–1028.
31. Koelsch, G., et al. 2000. Enzymic characteristics of secreted aspartic pro-teases of Candida albicans. Biochim. Biophys. Acta 1480:117–131.
32. Komano, H., N. Rockwell, G. T. Wang, G. A. Krafft, and R. S. Fuller. 1999.Purification and characterization of the yeast glycosylphosphatidylinositol-anchored, monobasic-specific aspartyl protease yapsin 2 (Mkc7p). J. Biol.Chem. 274:24431–24437.
33. Krysan, D. J., E. L. Ting, C. Abeijon, L. Kroos, and R. S. Fuller. 2005.Yapsins are a family of aspartyl proteases required for cell wall integrity inSaccharomyces cerevisiae. Eukaryot. Cell 4:1364–1374.
34. Maddi, A., S. M. Bowman, and S. J. Free. 2009. Trifluoromethanesulfonicacid-based proteomic analysis of cell wall and secreted proteins of the asco-mycetous fungi Neurospora crassa and Candida albicans. Fungal Genet. Biol.46:768–781.
35. Martínez, A. I., et al. 2004. Role of Pir1 in the construction of the Candidaalbicans cell wall. Microbiology 150:3151–3161.
36. Martínez-Lopez, R., C. Nombela, R. Diez-Orejas, L. Monteoliva, and C. Gil.2008. Immunoproteomic analysis of the protective response obtained fromvaccination with Candida albicans ecm33 cell wall mutant in mice. Proteom-ics 8:2651–2664.
37. Martinez-Lopez, R., H. Park, C. L. Myers, C. Gil, and S. G. Filler. 2006.Candida albicans Ecm33p is important for normal cell wall architecture andinteractions with host cells. Eukaryot. Cell 5:140–147.
38. McCreath, K. J., and G. W. Gooday. 1992. A rapid and sensitive microassayfor determination of chitinolytic activity. J. Microbiol. Methods 14:229–237.
39. McCreath, K. J., C. A. Specht, and P. W. Robbins. 1995. Molecular cloningand characterization of chitinase genes from Candida albicans. Proc. Natl.Acad. Sci. U. S. A. 92:2544–2548.
40. McKenzie, C. G., et al. 2010. Contribution of Candida albicans cell wallcomponents to recognition by and escape from murine macrophages. Infect.Immun. 78:1650–1658.
41. Meiller, T. F., et al. 2009. A novel immune evasion strategy of Candidaalbicans: proteolytic cleavage of a salivary antimicrobial peptide. PLoS One4:e5039.
42. Monge, R. A., E. Roman, C. Nombela, and J. Pla. 2006. The MAP kinasesignal transduction network in Candida albicans. Microbiology 152:905–912.
43. Morishima, H., T. Takita, T. Aoyagi, T. Takeuchi, and H. Umezawa. 1970.The structure of pepstatin. J. Antibiot. (Tokyo) 23:263–265.
44. Mrsa, V., T. Seidl, M. Gentzsch, and W. Tanner. 1997. Specific labelling ofcell wall proteins by biotinylation. Identification of four covalently linkedO-mannosylated proteins of Saccharomyces cerevisiae. Yeast 13:1145–1154.
45. Murad, A. M., P. R. Lee, I. D. Broadbent, C. J. Barelle, and A. J. Brown.2000. CIp10, an efficient and convenient integrating vector for Candidaalbicans. Yeast 16:325–327.
46. Naglik, J. R., S. J. Challacombe, and B. Hube. 2003. Candida albicanssecreted aspartyl proteinases in virulence and pathogenesis. Microbiol. Mol.Biol. Rev. 67:400–428.
47. Naglik, J. R., et al. 2008. Quantitative expression of the Candida albicanssecreted aspartyl proteinase gene family in human oral and vaginal candidi-asis. Microbiology 154:3266–3280.
48. Netea, M. G., G. D. Brown, B. J. Kullberg, and N. A. Gow. 2008. Anintegrated model of the recognition of Candida albicans by the innate im-mune system. Nat. Rev. Microbiol. 6:67–78.
49. Newport, G., and N. Agabian. 1997. KEX2 influences Candida albicans pro-teinase secretion and hyphal formation. J. Biol. Chem. 272:28954–28961.
50. Odds, F. C. 1988. Candida and candidosis. Bailliere Tindall, London, UnitedKingdom.
51. Olsen, V., N. X. Cawley, J. Brandt, M. Egel-Mitani, and Y. P. Loh. 1999.Identification and characterization of Saccharomyces cerevisiae yapsin 3, anew member of the yapsin family of aspartic proteases encoded by the YPS3gene. Biochem. J. 339(2):407–411.
52. Olsen, V., et al. 1998. Cleavage efficiency of the novel aspartic proteaseyapsin 1 (Yap3p) enhanced for substrates with arginine residues flanking theP1 site: correlation with electronegative active-site pockets predicted bymolecular modeling. Biochemistry 37:2768–2777.
53. Perlroth, J., B. Choi, and B. Spellberg. 2007. Nosocomial fungal infections:epidemiology, diagnosis, and treatment. Med. Mycol. 45:321–346.
54. Pitarch, A., M. Sanchez, C. Nombela, and C. Gil. 2002. Sequential fraction-ation and two-dimensional gel analysis unravels the complexity of the dimor-phic fungus Candida albicans cell wall proteome. Mol. Cell. Proteomics1:967–982.
55. Polgar, L., Z. Szeltner, and I. Boros. 1994. Substrate-dependent mechanismsin the catalysis of human immunodeficiency virus protease. Biochemistry33:9351–9357.
56. Popolo, L., and M. Vai. 1999. The Gas1 glycoprotein, a putative wall polymercross-linker. Biochim. Biophys. Acta 1426:385–400.
57. Richard, M. L., and A. Plaine. 2007. Comprehensive analysis of glycosylphos-phatidylinositol-anchored proteins in Candida albicans. Eukaryot. Cell6:119–133.
58. Rocha, C. R., et al. 2001. Signaling through adenylyl cyclase is essential forhyphal growth and virulence in the pathogenic fungus Candida albicans. Mol.Biol. Cell 12:3631–3643.
59. Ruchel, R. 1981. Properties of a purified proteinase from the yeast Candidaalbicans. Biochim. Biophys. Acta 659:99–113.
60. Ruhnke, M., and G. Maschmeyer. 2002. Management of mycoses in patientswith hematologic disease and cancer—review of the literature. Eur. J. Med.Res. 7:227–235.
61. Ruiz-Herrera, J., M. V. Elorza, E. Valentin, and R. Sentandreu. 2006. Mo-lecular organization of the cell wall of Candida albicans and its relation topathogenicity. FEMS Yeast Res. 6:14–29.
62. Schaller, M., C. Borelli, H. C. Korting, and B. Hube. 2005. Hydrolyticenzymes as virulence factors of Candida albicans. Mycoses 48:365–377.
63. Selvaggini, S., C. A. Munro, S. Paschoud, D. Sanglard, and N. A. Gow. 2004.Independent regulation of chitin synthase and chitinase activity in Candidaalbicans and Saccharomyces cerevisiae. Microbiology 150:921–928.
64. Sheppard, D. C., et al. 2004. Functional and structural diversity in the Alsprotein family of Candida albicans. J. Biol. Chem. 279:30480–30489.
65. Sorgo, A. G., et al. 2010. Mass spectrometric analysis of the secretome ofCandida albicans. Yeast 27:661–672.
66. Thomas, D. P., S. P. Bachmann, and J. L. Lopez-Ribot. 2006. Proteomics forthe analysis of the Candida albicans biofilm lifestyle. Proteomics 6:5795–5804.
67. Urban, C., K. Sohn, F. Lottspeich, H. Brunner, and S. Rupp. 2003. Identi-fication of cell surface determinants in Candida albicans reveals Tsa1p, aprotein differentially localized in the cell. FEBS Lett. 544:228–235.
68. Walther, A., and J. Wendland. 2003. An improved transformation protocolfor the human fungal pathogen Candida albicans. Curr. Genet. 42:339–343.
69. Weissman, Z., and D. Kornitzer. 2004. A family of Candida cell surfacehaem-binding proteins involved in haemin and haemoglobin-iron utilization.Mol. Microbiol. 53:1209–1220.
70. Wessel, D., and U. I. Flugge. 1984. A method for the quantitative recovery ofprotein in dilute solution in the presence of detergents and lipids. Anal.Biochem. 138:141–143.
71. Wheeler, R. T., and G. R. Fink. 2006. A drug-sensitive genetic network masksfungi from the immune system. PLoS Pathog. 2:e35.
72. Whiteway, M., and C. Bachewich. 2007. Morphogenesis in Candida albicans.Annu. Rev. Microbiol. 61:529–553.
73. Wilson, D., et al. 2009. Identifying infection-associated genes of Candidaalbicans in the postgenomic era. FEMS Yeast Res. 9:688–700.
VOL. 10, 2011 C. ALBICANS Sap9 AND Sap10 CLEAVE CELL WALL PROTEINS 109
on January 4, 2019 by guesthttp://ec.asm
.org/D
ownloaded from


















