
Proposedthree-dimensional structure for the cellular … · Proposedthree-dimensional structure...
5
Proc. Natd. Acad. Sci. USA Vol. 91, pp. 7139-7143, July 1994 Biophysics Proposed three-dimensional structure for the cellular prion protein (protein conformation/secondary structure prediction/tertiary structure predictIon/computer modelng/prion diseases) ZIWEI HUANG*t, JEAN-MARC GABRIELtI, MICHAEL A. BALDWINt, ROBERT J. FLETTERICK*§, STANLEY B. PRUSINERt§, AND FRED E. COHEN*§1¶I Departments of *Pharmaceutical Chemistry, tNeurology, §Biochemistry and Biophysics, and NMedicine, University of California, San Francisco, CA 94143 Contributed by Stanley B. Prusiner, February 14, 1994 ABSTRACT Prion diseases are a group of neurodegenera- tive disorders in human and animals that seem to result from a conformatlonal change in the prion protein (PrP). Utilizing data obtained by circular dichroism and in spectroscopy, computational studies prdIe the tedi nal s ture of the cellular form of PrP (PrPC). A heuristic approach consistng of the prediction of so structures and of an evalnation of the pig of s y elements was used to search for plausible teriay strucures. After a series of exper- imental and theoretical ants were applied, four structural models of four-helix bundles emerged. A group of amino acids within the four predicted helices were Identified as important for tertiary interactions between helices. These amino acids could be essential for ma ining a stable tertiary suture of PrPC. Among four plausible s al models for PrPC, the X-bundle model seemed to correlate best with 5 of 11 known point mutations that segregate with the inherited prion diseases. These 5 mutions cluster around a central hydrophobic core in the X-bundle structure. Furthermore, these mutations occur at or near those amino acds which are predicted to e important for helix-helix interactions. The three-dimensonal structure of PrPc proposed here may not only provide a basis for rational- izing mutations of the PrP gene in the inherited prion diseases but also guide design of genticafly engineered NP molecules for further experimental studies. Prions are a novel class of "infectious" pathogens distinct from viroids and viruses with respect to both their structure and the neurodegenerative diseases that they -cause (1). Prion diseases are manifest as sporadic, inherited, and infectious disorders including scrapie, mink encephalopathy, chronic wasting disease, bovine spongiform encephalopathy, feline spongiform encephalopathy, and exotic ungulate encepha- lopathy of animals (2-4) as well as kuru, Creutzfeldt-Jakob disease (CJD), Gerstmann-Striussler-Scheinker syndrome, and fatal familial insomnia of humans (5-7). The prion protein (PrP) is the major, if not the only, component of prions (1). PrP exists in two isoforms: the normal cellular form (PrP9 and the abnormal disease (scrapie)-related form (PrPSc) (8, 9). The entire open reading frame of PrP genes is contained in a single exon, eliminating the possibility that PrPC and PrPSc arise from alternative RNA splicing (10). Attempts to identify a posttranslational chemical modification that features in the conversion of PrPC to PrPSc have been unsuccessful (11). Structural studies of PrPC and PrPSC using Fourier-transform infrared and circular dichroism spectroscopy indicated that PrPc and PrPSC differ from each other in their conformations (12-17). These findings suggest that prion diseases are dis- orders of protein conformation and result from a change in the structure of PrPC when it is converted into PrPsc. Elucidating the three-dimensional structures of PrPC and PrPSC and the conformational changes that occur during the production of PrPsc is central to understanding the molecular mechanisms of prion diseases. Because of the low level of expression of PrPC and insolubility of PrPSc, efforts to obtain crystals for x-ray structure determinations have been unsuc- cessful. Exploiting recent advances in protein structure pre- diction algorithms, we carried out computational studies to predict the three-dimensional structure of PrPC based on a family of homologous amino acid sequences. Our predictive studies were facilitated by spectroscopic findings showing that PrPC has a secondary structure which contains ==43% a-helix and is virtually devoid of 3-sheet (15). Although the problem of predicting protein structure from the amino acid sequence information alone remains unsolved, recent advances in secondary and tertiary structure predic- tion of proteins have shown that reasonable structures can often be proposed by using a heuristic approach in conjunc- tion with experimental data (18). For all-a-helical proteins, this approach has been applied to generate low-resolution structural models for a number of proteins (19-21). Some of the structural features proposed by these methods have been verified by subsequent x-ray or NMR experiments (22). In the study reported here, three-dimensional structures of PrPc were generated by applying the heuristic approach coupled with experimental data. In the absence of a three- dimensional structure of PrPC from either x-ray crystallog- raphy or NMR spectroscopy, these models provide a vehicle to rationalize much of the available data and help design further experimental studies. METHODS The computational procedures used for the prediction of the three-dimensional structures of PrPC involved four major steps: (i) alignment of a family of homologous sequences, (ii) prediction of secondary structures, (iii) packing of secondary elements to generate all plausible tertiary structures, and (iv) selection and refinement of final structural models. PrP amino acid sequences from 1 avian and 11 mammalian sources including chicken, cow, sheep, rat, mouse, hamster, mink, and human were used. The alignment of these se- quences was reported previously (23), using the Feng and Doolittle algorithm (24). Methods for secondary and tertiary structure prediction were applied independently to all 12 PrP sequences. As suggested by Benner and Gerloff (25), the use of a broad family of homologous sequences improves the accuracy of structure prediction. Secondary Structure Prediction. An initial attempt to pre- dict the secondary structure of PrPC was made by several different methods, including the Chou-Fasman method (26), GOR algorithm (27), and PHD program (28). The GOR algorithm Abbreviations: PrP, prion protein; PrPC, cellular isoform of PrP; PrPs, scrapie isoform of PrP; CJD, Creutzfeldt-Jakob disease. ITo whom reprint requests should be addressed at: Department of Pharmaceutical Chemistry, University of California, San Fran- cisco, CA 94143-0446. tPresent address: Department of Research, Neurobiology Labora- tory, Kantonsspital, CH-4031 Basel, Switzerland. 7139 The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Transcript of Proposedthree-dimensional structure for the cellular … · Proposedthree-dimensional structure...

Proc. Natd. Acad. Sci. USAVol. 91, pp. 7139-7143, July 1994Biophysics
Proposed three-dimensional structure for the cellular prion protein(protein conformation/secondary structure prediction/tertiary structure predictIon/computer modelng/prion diseases)
ZIWEI HUANG*t, JEAN-MARC GABRIELtI, MICHAEL A. BALDWINt, ROBERT J. FLETTERICK*§,STANLEY B. PRUSINERt§, AND FRED E. COHEN*§1¶IDepartments of *Pharmaceutical Chemistry, tNeurology, §Biochemistry and Biophysics, and NMedicine, University of California, San Francisco, CA 94143
Contributed by Stanley B. Prusiner, February 14, 1994
ABSTRACT Prion diseases are a group of neurodegenera-tive disorders in human and animals that seem to result froma conformatlonal change in the prion protein (PrP). Utilizingdata obtained by circular dichroism and in spectroscopy,computational studies prdIe thetedi nal s tureof the cellular form of PrP (PrPC). A heuristic approachconsistng of the prediction ofso structures and of anevalnation of the pig of s y elements was used tosearch for plausible teriay strucures. After a series of exper-imental and theoretical ants were applied, four structuralmodels of four-helix bundles emerged. A group of amino acidswithin the four predicted helices were Identified as important fortertiary interactions between helices. These amino acids could beessential for ma ining a stable tertiary suture of PrPC.Among four plausible s al models for PrPC, the X-bundlemodel seemed to correlate best with 5 of 11 known pointmutations that segregate with the inherited prion diseases. These5 mutions cluster around a central hydrophobic core in theX-bundle structure. Furthermore, these mutations occur at ornear those amino acds which are predicted to e important forhelix-helix interactions. The three-dimensonal structure ofPrPc proposed here may not only provide a basis for rational-izing mutations of the PrP gene in the inherited prion diseasesbut also guide design ofgenticafly engineered NP molecules forfurther experimental studies.
Prions are a novel class of "infectious" pathogens distinctfrom viroids and viruses with respect to both their structureand the neurodegenerative diseases that they-cause (1). Priondiseases are manifest as sporadic, inherited, and infectiousdisorders including scrapie, mink encephalopathy, chronicwasting disease, bovine spongiform encephalopathy, felinespongiform encephalopathy, and exotic ungulate encepha-lopathy of animals (2-4) as well as kuru, Creutzfeldt-Jakobdisease (CJD), Gerstmann-Striussler-Scheinker syndrome,and fatal familial insomnia ofhumans (5-7). The prion protein(PrP) is the major, if not the only, component of prions (1).PrP exists in two isoforms: the normal cellular form (PrP9and the abnormal disease (scrapie)-related form (PrPSc) (8, 9).The entire open reading frame of PrP genes is contained in
a single exon, eliminating the possibility that PrPC and PrPScarise from alternative RNA splicing (10). Attempts to identifya posttranslational chemical modification that features in theconversion of PrPC to PrPSc have been unsuccessful (11).Structural studies of PrPC and PrPSC using Fourier-transforminfrared and circular dichroism spectroscopy indicated thatPrPc and PrPSC differ from each other in their conformations(12-17). These findings suggest that prion diseases are dis-orders of protein conformation and result from a change inthe structure of PrPC when it is converted into PrPsc.
Elucidating the three-dimensional structures of PrPC andPrPSC and the conformational changes that occur during the
production of PrPsc is central to understanding the molecularmechanisms of prion diseases. Because of the low level ofexpression ofPrPC and insolubility of PrPSc, efforts to obtaincrystals for x-ray structure determinations have been unsuc-cessful. Exploiting recent advances in protein structure pre-diction algorithms, we carried out computational studies topredict the three-dimensional structure of PrPC based on afamily of homologous amino acid sequences.Our predictive studies were facilitated by spectroscopic
findings showing that PrPC has a secondary structure whichcontains ==43% a-helix and is virtually devoid of 3-sheet (15).Although the problem ofpredicting protein structure from theamino acid sequence information alone remains unsolved,recent advances in secondary and tertiary structure predic-tion of proteins have shown that reasonable structures canoften be proposed by using a heuristic approach in conjunc-tion with experimental data (18). For all-a-helical proteins,this approach has been applied to generate low-resolutionstructural models for a number of proteins (19-21). Some ofthe structural features proposed by these methods have beenverified by subsequent x-ray or NMR experiments (22).
In the study reported here, three-dimensional structures ofPrPc were generated by applying the heuristic approachcoupled with experimental data. In the absence of a three-dimensional structure of PrPC from either x-ray crystallog-raphy or NMR spectroscopy, these models provide a vehicleto rationalize much of the available data and help designfurther experimental studies.
METHODSThe computational procedures used for the prediction of thethree-dimensional structures of PrPC involved four majorsteps: (i) alignment ofa family ofhomologous sequences, (ii)prediction ofsecondary structures, (iii) packing of secondaryelements to generate all plausible tertiary structures, and (iv)selection and refinement of final structural models. PrPamino acid sequences from 1 avian and 11 mammaliansources including chicken, cow, sheep, rat, mouse, hamster,mink, and human were used. The alignment of these se-quences was reported previously (23), using the Feng andDoolittle algorithm (24). Methods for secondary and tertiarystructure prediction were applied independently to all 12 PrPsequences. As suggested by Benner and Gerloff (25), the useof a broad family of homologous sequences improves theaccuracy of structure prediction.
Secondary Structure Prediction. An initial attempt to pre-dict the secondary structure of PrPC was made by severaldifferent methods, including the Chou-Fasman method (26),GOR algorithm (27), and PHD program (28). The GOR algorithm
Abbreviations: PrP, prion protein; PrPC, cellular isoform of PrP;PrPs, scrapie isoform of PrP; CJD, Creutzfeldt-Jakob disease.ITo whom reprint requests should be addressed at: Department ofPharmaceutical Chemistry, University of California, San Fran-cisco, CA 94143-0446.
tPresent address: Department of Research, Neurobiology Labora-tory, Kantonsspital, CH-4031 Basel, Switzerland.
7139
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 91 (1994)
was used without setting a decision constant (27). Results ofthe PHD method were obtained from the European MolecularBiology Laboratories. As discussed later, these general-purpose prediction methods gave confusing assignments forsecondary structure. Given the experimental data indicatingthat PrP exists in two distinct conformations, the inconsis-tencies among various algorithms could reflect their intrinsiclimitations in predicting one structure for a protein with twoor more distinct conformational isoforms.
In an attempt to overcome the inherent limitations ofsecondary structure prediction methods, we considered in-formation derived from spectroscopic studies of PrPC. Fou-rier-transform infrared and circular dichroism spectroscopyindicated that PrPC contains 43% a-helices and virtually no(3-sheet (15). These data indicated that PrPc is a member ofthe structural class of proteins that are all-a-helical. With thisknowledge, we first applied a pattern-based turn predictionalgorithm (29) to identify sequence regions that could containsecondary structures, and then predicted a-helices or(-strands within these regions by using a neural network (30,31). While most general-purpose secondary structure predic-tion algorithms achieve only '65% accuracy due to the lackof tertiary information (32), the turn prediction algorithm andneural network method, having been optimized to exploit thecharacteristics of all-a-helical protein structures, are accu-rate 90%o and 79%6 of the time, respectively (29, 30). Foursequence regions were identified from the turn predictionalgorithm and these regions were subsequently predicted tocontain helices by the neural network method. Independentcalculations of polypeptide chain flexibility using the methodof Karplus and Schulz (33) also indicated that these regionswere the least flexible in the entire PrP sequence and thusmost likely to contain secondary structure.
Tertiary Structure Prediction. Tertiary structures weregenerated by investigating the plausible arrangements of thea-helices via a combinatorial packing approach (20, 34). Thisapproach was developed based on the original concept of thepacking of a-helices proposed by Crick (35). The generalgeometric features of helix-helix packing are followed in amanner compatible 'with x-ray data on the structures ofglobular proteins (36, 37). The algorithm is divided into threesteps: (i) the program AAPATCH identifies clusters of hydro-phobic residues within the putative helices that could mediatehelix-helix interactions, and these fall into three geometricsubclasses; (ii) AAFOLD generates all possible helix pairingsaccording to the location and geometric preferences of theinteraction sites; and (iii) AABUILD generates three-dimensional models of all possible structures from the list ofhelix pairings (from AAFOLD) subject to steric restrictions andgeometric constraints on chain folding. Additional physicaland chemical data for PrPc, such as the experimentallydetermined disulfide bridge (38) and glycosylation sites, aswell as general features of proteins, such as the expectedrelationship of surface area to molecular volume, were usedto reduce further the set of plausible alternative structures.At this stage, coordinates were specified for the core helicesof the protein. Loops connecting these core helices wereinserted by using the BLOOP program (39). In general theconformations of loops and side chains in proteins are moreflexible than the backbone of secondary elements; conse-quently, their conformations are less well defined in theproposed models.
RESULTSSecondary Structure of PrPC. Twelve PrP sequences were
aligned (23) and analyzed by use of a pattern-based turnprediction method and a neural network program. Results ofpattern-based turn predictions and neural network are avail-able from the authors via anonymous file transfer protocol.Four putative helical regions of PrPC were identified (Fig.
1A). The different assignments of a-helix or (3-strand withinthese four helical regions made by other methods such as theChou-Fasman and GOR algorithms were also shown (Fig.1A). Ten of 11 point mutations that segregate with theinherited prion diseases (45) lie either within or adjacent tothese four regions of secondary structure (Fig. 1B). Theseobservations are consistent with the findings presented herewhich argue that the four predicted a-helices of PrPC arestructurally and functionally important regions.Tertary Structural Models of PrPC. The combinatorial
helix-helix packing approach was applied to search forplausible tertiary structures of PrPC. Within the four pre-dicted helical regions, 15 residues were identified as potentialsites that could mediate helix-helix interactions. Table 1shows the location of these amino acid residues and theirrelative conservation over 12 PrP sequences. Except forresidue 184 in helix 3, all potential helix-helix interactionresidues were highly conserved. Even though residue 184 ismore variable, further analysis has revealed that either Ile orVal at this position can maintain the helix-helix interaction atthis site (36). An exhaustive search of plausible packingarrangements of four helices mediated by their possiblehelix-helix interaction sites gave rise to a total of -300,000structures. Among this large number of structures, only asmall number of structures were sterically reasonable, with asufficient number of residues in the loops between thesehelices to create a stereochemically sensible structure andstill form the experimentally determined disulfide bond be-tween Cys-179 in helix 3 and Cys-214 in helix 4 (38). Thesestructures were clustered into w200 structural families. Theremaining structures were then rank ordered by solvent-accessible surface area. Four structures were chosen as theplausible models of PrPC (Fig. 2). These structures weresignificantly more compact than the rest as judged by theratio of accessible surface area to molecular volume.
A ! i:'. IN IlChN .;
I1\lixL~~~~~~~~~~~~~~~~(i l) R
Ncural NcLtwrk iormIiI .-hXelical protcliri _
B
FI.. 1. Structural and mutational features of the PrP sequence.(A) Assignments of secondary structures within the four helicalregions by means of the Chou-Fasman method (26), GOR algorithm(27), and PHD program (28). These four regions were identified asthose likely to contain secondary structures using an approach ofcombining the pattern-based turn prediction (29) and polypeptidechain flexibility calculation (33). The final assignment of a-heliceswithin these regions was based on results from the neural networkprogram optimized for all a-helical proteins (30, 31). a-Helices aredesignated by stippling and (3-sheets by hatching. (B) Point mutationsthat segregate with inherited prion diseases in humans. Four of thesemutations have been genetically linked: P1O2L, D178N, F198S, andE200K (6, 40-42). Polymorphisms M129V and E219K in the humanPrP gene are also shown (43, 44).
7140 Biophysics: Huang et al.

Proc. Nadl. Acad. Sci. USA 91 (1994) 7141
Table 1. Predicted a-helices and helix-helix interaction sites
Helix1
234
Resi- Sequence identity*,dues* Helix interaction sitest %109-122 MKHMAGAAAAGAW 100; 100; 100; 100;
100129-141 MLGSAMSRPIIHF 92; 100178-191 DCVNITIKQHTVTT 100; 100; 58202-218 DVKMMERVVEQMCITQY 92; 100; 92; 100; 100
*Residue numbers correspond to those in human PrP. The corre-sponding numbers in the published alignment of the 12 PrP se-quences are as follows: helix 1, 122-135; helix 2, 142-154; helix 3,191-204; helix 4, 225-241 (23).tHuman PrP amino acid sequences for the four predicted helices areshown. Helix-helix interaction sites are hih ted by boldfacetype.tPercent sequence identity is shown in order corresponding torespective helix-helix interaction residues. The percentage wascalculated as the number of identical residues divided by the numberof total sequences.
These four structures do not resemble the commonly ob-served square four-helix bundles studied by Presnell andCohen (47); however, they are compatible with a more diversecollection of four-helix bundles (46). Harris et al. (46) foundthat helical bundles adopted a number of different topologicalarrangements in addition to the square bundle motif. Accord-ing to their definitions, the four possible structures predictedfor PrPc are classified as bicornate, X, and splinter bundles.Both models I and II belong to the same bicornate type butdisplay different relative locations for helices 3 and 4. Bicor-nate bundles have only been seen as part of a larger proteinstructure (46) while X and splinter bundles seem to be stablein isolation without additional structural elements. For exam-ple, granulocyte/macrophage-colony-stimulatng factor is asplinter bundle (48). The POU-specific DNA-binding domainof Oct-i forms an X-bundle conformation (49).
Model I: Bicornate Bundle Model II: Bicornate Bundle
Model III: X-Bundle Model IV: Splinter Bundle
FiG. 2. Schematic drawings of the four plausible structures ofPrPC. These tertiary structural models were selected from -300,000possible structures based on the experimentally determined andtheoretical constraints. The definitions for these models follow thoseused by Harris et al. (46).
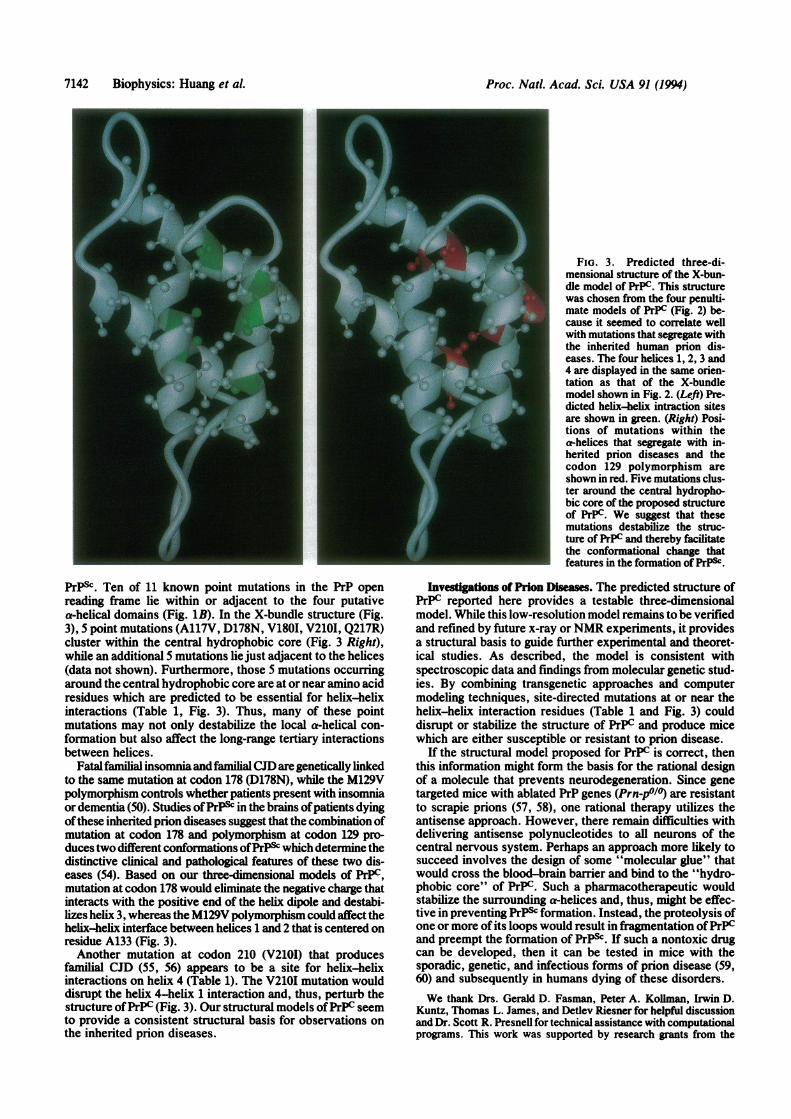
Each of the four-helix bundle models was analyzed withrespect to the point mutations found in the inherited priondiseases. The X-bundle structure was most intriguing withrespect to the spatial clustering of mutations. The averagedistance between mutations on the various helices was sig-nificantly smaller for the X-bundle structure (9.3 A) than forthe other three models (I, 14.2 A; 11, 14.5 A; IV, 12.6 A). Thethree-dimensional arrangement ofthe four putative helices inthe X-bundle structure is shown in Fig. 3. The sites ofhelix-helix interactions on the hydrophobic faces of eachhelix are lighted in green (Fig. 3 Left). The five pointmutations (A117V, D178N, V1801, V2101, Q217R) clusteraround a central hydrophobic core (Fig. 3 Right), while anadditional five mutations lie just adjacent to the helices (datanot shown). The sites of the mutations clustering around thehydrophobic core are shown in red as is the M129V poly-morphism which controls whether the D178N mutationcauses neurodegeneration presenting with insomnia or de-mentia (50); residue 129 lies at the beginning of helix 2.
DISCUSSIONA igu 1 ofoPredico S re: A Different
Perspcive? In our initial analysis ofthe secondary structureofPrPC, general-purpose methods such as the Chou-Fasmanand Garnier algorithms were used (26, 27). These methodsgave a confusing view of the secondary structural elements.In particular, these methods disagreed as to the preferredconformation for each of the four putative structural regions(Fig. 1A). While the conflicting predictions could reflect theintrinsic limitations ofcurrent secondary structure predictionalgorithms, it is also plausible that apparent inconsistenciesreflect an attempt to predict one structure for a protein withtwo or more quite distinct conformational isoforms. Thislatter explanation is supported by spectroscopic studiesdemonstrating that PrPC is an all-a-helical protein whereasPrPsC possesses a high (-sheet content (15).These results suggest that different conformations of the
four structural regions predicted by classical algorithmscould be indicative of the alternative conformations that PrPcan adopt. This formulation is consistent with experimentalstudies with synthetic peptides corresponding to the fourputative regions of secondary structure for PrP. Three ofthefour peptides were found to adopt a 3-sheet conformation inaqueous buffers (51). This is reminiscent of the conforma-tions of the pre-pro-parathyroid hormone precursor peptide.The Chou-Fasman algorithm predicted two structures, onelargely a-helical and the other rich in P-strands (52). Circulardichroism studies demonstrated that this peptide could adopttwo distinct conformations under different solvent condi-tions. Although it is widely accepted that some domains ofproteins can adopt distinct conformations in different envi-ronments (53), structure prediction analyses generally at-tempt to identify a consensus conformation. While we realizethe importance of seeking a consensus among various meth-ods used for prediction of protein structure, we suggest thatthis conventional approach should probably be revised forproteins exhibiting conformational heterogeneity such asPrP. The results of studies on PrP summarized here providea different perspective into the use and interpretation ofsecondary structure prediction methods.Strucural Imp of Mutations in PrP Genes. Since
the first study demonstrating genetic linkage between amutation in codon 102 and development of Gerstmann-StrAussler-Scheinker syndrome (6), 17 other mutations in thePrP gene have been identified that segregate with the inher-ited prion diseases (45). In both humans and transgenic mice,these mutations produce neurodegeneration and often prionsas demonstrated by transmission of disease to experimentalanimals. Presumably, these amino acid substitutions desta-bilize the conformation ofPrPc and facilitate its refolding into
Biophysics: Huang et A
Proc. Natl. Acad. Sci. USA 91 (1994)
FIG. 3. Predicted three-di-mensional structure of the X-bun-dle model of PrPC. This structurewas chosen from the four penulti-mate models of PrPC (Fig. 2) be-cause it seemed to correlate wellwith mutations that segregate withthe inherited human prion dis-eases. The four helices 1, 2, 3 and4 are displayed in the same orien-tation as that of the X-bundlemodel shown in Fig. 2. (Left) Pre-dicted helix-helix intraction sitesare shown in green. (Right) Posi-tions of mutations within thea-helices that segregate with in-herited prion diseases and thecodon 129 polymorphism areshown in red. Five mutations clus-ter around the central hydropho-bic core of the proposed structureof PrPC. We suggest that thesemutations destabilize the struc-ture of PrPC and thereby facilitatethe conformational change thatfeatures in the formation of PrP&.
PrPSc. Ten of 11 known point mutations in the PrP openreading frame lie within or adjacent to the four putativea-helical domains (Fig. 1B). In the X-bundle structure (Fig.3), 5 point mutations (A117V, D178N, V180I, V210I, Q217R)cluster within the central hydrophobic core (Fig. 3 Right),while an additional 5 mutations liejust adjacent to the helices(data not shown). Furthermore, those S mutations occurringaround the central hydrophobic core are at or near amino acidresidues which are predicted to be essential for helix-helixinteractions (Table 1, Fig. 3). Thus, many of these pointmutations may not only destabilize the local a-helical con-formation but also affect the long-range tertiary interactionsbetween helices.
Fatalfamilialinsomniaandfamilial CJD are genetically linkedto the same mutation at codon 178 (D178N), while the M129Vpolymorphism controls whether patients present with insomniaor dementia (50). Studies ofPrP& in the brains ofpatients dyingofthese inherited prion diseases suggest that the combination ofmutation at codon 178 and polymorphism at codon 129 pro-duces two different conformations ofPrP& which determine thedistinctive clinical and pathological features of these two dis-eases (54). Based on our three-dimensional models of PrPC,mutation at codon 178 would eliminate the negative charge thatinteracts with the positive end of the helix dipole and destabi-lizes helix 3, whereas the M129V polymorphism could affect thehelix-helix interface between helices 1 and 2 that is centered onresidue A133 (Fig. 3).Another mutation at codon 210 (V2101) that produces
familial CJD (55, 56) appears to be a site for helix-helixinteractions on helix 4 (Table 1). The V210I mutation woulddisrupt the helix 4-helix 1 interaction and, thus, perturb thestructure ofPrPC (Fig. 3). Our structural models ofPrPC seemto provide a consistent structural basis for observations onthe inherited prion diseases.
Investigations of Prion Diseases. The predicted structure ofPrPc reported here provides a testable three-dimensionalmodel. While this low-resolution model remains to be verifiedand refined by future x-ray orNMR experiments, it providesa structural basis to guide further experimental and theoret-ical studies. As described, the model is consistent withspectroscopic data and findings from molecular genetic stud-ies. By combining transgenetic approaches and computermodeling techniques, site-directed mutations at or near thehelix-helix interaction residues (Table 1 and Fig. 3) coulddisrupt or stabilize the structure of PrPC and produce micewhich are either susceptible or resistant to prion disease.
If the structural model proposed for PrPC is correct, thenthis information might form the basis for the rational designof a molecule that prevents neurodegeneration. Since genetargeted mice with ablated PrP genes (Prn-p0/0) are resistantto scrapie prions (57, 58), one rational therapy utilizes theantisense approach. However, there remain difficulties withdelivering antisense polynucleotides to all neurons of thecentral nervous system. Perhaps an approach more likely tosucceed involves the design of some "molecular glue" thatwould cross the blood-brain barrier and bind to the "hydro-phobic core" of PrPC. Such a pharmacotherapeutic wouldstabilize the surrounding a-helices and, thus, might be effec-tive in preventing PrPsc formation. Instead, the proteolysis ofone or more of its loops would result in fiagmentation ofPrPCand preempt the formation of PrPsc. If such a nontoxic drugcan be developed, then it can be tested in mice with thesporadic, genetic, and infectious forms of prion disease (59,60) and subsequently in humans dying of these disorders.We thank Drs. Gerald D. Fasman, Peter A. Kollman, Irwin D.
Kuntz, Thomas L. James, and Detlev Riesner for helpful discussionand Dr. Scott R. Presnell for technical assistance with computationalprograms. This work was supported by research grants from the
7142 Biophysics: Huang et aL

Proc. Natl. Acad. Sci. USA 91 (1994) 7143
National Institutes of Health and a gift from the Sherman FairchildFoundation. Z.H. is supported by a postdoctoral training grant fromthe National Institutes of Health. J.-M.G. was supported by afellowship from the Swiss National Science Foundation.
1. Prusiner, S. B. (1991) Science 252, 1515-1522.2. Wilesmith, J. & Wells, G. A. H. (1991) Curr. Top. Microbiol.
Immunol. 172, 21-38.3. Marsh, R. F., Bessen, R. A., Lehmann, S. & Hartsough, G. R.
(1991) J. Gen. Virol. 72, 589-594.4. Westaway, D., Zuliani, V., Mirenda Cooper, C., Da Costa, M.,
Neuman, S., Jenny, A. L., Detwiler, L. & Prusiner, S. B.(1994) Genes Dev. 8, 959-969.
5. Gajdusek, D. C. (1977) Science 197, 943-960.6. Hsiao, K., Baker, H. F., Crow, T. J., Poulter, M., Owen, F.,
Terwilliger, J. D., Westaway, D., Ott, J. & Prusiner, S. B.(1989) Nature (London) 338, 342-345.
7. Medori, R., Tritschler, H.-J., LeBlanc, A., Villare, F., Man-etto, V., Chen, H. Y., Xue, R., Leal, S., Montagna, P.,Cortelli, P., Tinuper, P., Avoni, P., Mochi, M., Baruzzi, A.,Hauw, J. J., Ott, J., Lugaresi, E., Autilio-Gambetti, L. &Gambetti, P. (1992) N. Engl. J. Med. 326, 444-449.
8. Oesch, B., Westaway, D., WAlchli, M., McKinley, M. P.,Kent, S. B. H., Aebersold, R., Barry, R. A., Tempst, P.,Teplow, D. B., Hood, L. E., Prusiner, S. B. & Weissmann, C.(1985) Cell 40, 735-746.
9. Meyer, R. K., McKinley, M. P., Bowman, K. A., Braunfeld,M. B., Barry, R. A. & Prusiner, S. B. (1986) Proc. Nati. Acad.Sci. USA 83, 2310-2314.
10. Basler, K., Oesch, B., Scott, M., Westaway, D., Walchli, M.,Groth, D. F. McKinley, M. P., Prusiner, S. B. & Weissmann,C. (1986) Cell 46, 417-428.
11. Stahl, N., Baldwin, M. A., Teplow, D. B., Hood, L., Gibson,B. W., Burlingame, A. L. & Prusiner, S. B. (1993) Biochem-istry 32, 1991-2002.
12. Prusiner, S. B., McKinley, M. P., Bowman, K. A., Bolton,D. C., Bendheim, P. E., Groth, D. F. & Glenner, G. G. (1983)Cell 35, 349-358.
13. Caughey, B. W., Dong, A., Bhat, K. S., Ernst, D., Hayes,S. F. & Caughey, W. S. (1991) Biochemistry 30, 7672-7680.
14. Gasset, M., Baldwin, M. A., Fletterick, R. J. & Prusiner, S. B.(1993) Proc. Natd. Acad. Sci. USA 90, 1-5.
15. Pan, K.-M., Baldwin, M., Nguyen, J., Gasset, M., Serban, A.,Groth, D., Mehlhorn, I., Huang, Z., Fletterick, R. J., Cohen,F. E. & Prusiner, S. B. (1993) Proc. Natd. Acad. Sci. USA 90,10962-10966.
16. Safar, J., Roller, P. P., Gajdusek, D. C. & Gibbs, C. J. J.(1993) Protein Sci. 2, 2206-2216.
17. Safar, J., Roller, P. P., Gajdusek, D. C. & Gibbs, C. J., Jr.(1993) J. Biol. Chem. 268, 20276-20284.
18. Ring, C. S. & Cohen, F. E. (1993) FASEB J. 7, 783-790.19. Cohen, F. E., Kosen, P. A., Kuntz, I. D., Epstein, L. B.,
Ciardelli, T. L. & Smith, K. A. (1986) Science 234, 349-352.20. Curtis, B. M., Presnell, S. R., Srnivasan, S., Sassenfeld, H.0,
Klinke, R., Jeffrey, E., Cosman, D., March, C. J. & Cohen,F. E. (1991) Proteins Struct. Funct. Genet. 11, 111-119.
21. Boissel, J. P., Lee, W. R., Presnell, S. R., Cohen, F. E. &Bunn, H. F. (1993) J. Biol. Chem. 268, 15983-15993.
22. Smith, L. J., Redfield, A. C., Boyd, A. J., Lawrence, G. M.,Edwards, R., Smith, R. A. & Dobson, C. M. (1992) J. Mol.Biol. 224,899-904.
23. Gabriel, J.-M., Oesch, B., Kretzschmar, H., Scott, M. &Prusiner, S. B. (1992) Proc. Natl. Acad. Sci. USA 89, 9097-9101.
24. Feng, D. F. & Doolittle, R. F. (1987) J. Mol. Evol. 25,351-360.25. Benner, S. A. & Gerloff, D. (1991) Adv. Enzyme Regul. 31,
121-181.26. Chou, P. Y. & Fasman, G. D. (1978) Annu. Rev. Biochem. 47,
251-276.27. Garnier, J., Osguthorpe, D. J. & Robson, B. (1978) J. Mol.
Biol. 120, 97-120.28. Rost, B. & Sander, C. (1992) Nature (London) 360, 540.29. Cohen, F. E., Abarbanel, R. M., Kuntz, I. D. & Fletterick,
R. J. (1986) Biochemistry 25, 266-275.30. Kneller, D. G., Cohen, F. E. & Langridge, R. (1990) J. Mol.
Biol. 214, 171-182.
31. Presnell, S. R., Cohen, B. I. & Cohen, F. E. (1993) Comput.Appl. Biosci. 9, 373-374.
32. Gamier, J. (1990) Biochimie 72, 513-524.33. Karplus, P. A. & Schulz, G. E. (1985) Naturwissenschaften 72,
212-213.34. Cohen, F. E., Richmond, T. J. & Richards, F. M. (1979) J.
Mol. Biol. 132, 275-288.35. Crick, F. H. C. (1953) Acta Crystallogr. 6, 689-697.36. Richmond, T. J. & Richards, F. M. (1978) J. Mol. Biol. 119,
537-555.37. Chothia, C., Levitt, M. & Richardson, D. (1977) Proc. Natl.
Acad. Sci. USA 74, 4130-4134.38. Turk, E., Teplow, D. B., Hood, L. E. & Prusiner, S. B. (1988)
Eur. J. Biochem. 176, 21-30.39. Ring, C. S. & Cohen, F. E. (1994) Isr. J. Chem., in press.40. Dlouhy, S. R., Hsiao, K., Farlow, M. R., Foroud, T., Con-
neally, P. M., Johnson, P., Prusiner, S. B., Hodes, M. E. &Ghetti, B. (1992) Nat. Genet. 1, 64-67.
41. Petersen, R. B., Tabaton, M., Berg, L., Schrank, B., Torack,R. M., Leal, S., Julien, J., Vital, C., Deleplanque, B., Pend-lebury, W. W., Drachman, D., Smith, T. W., Martin, J. J.,Oda, M., Montagna, P., Ott, J., Autilio-Gambetti, L., Lugaresi,E. & Gambetti, P. (1992) Neurology 42, 1859-1863.
42. Gabizon, R., Rosenmann, H., Meiner, Z., Kahana, I., Kahana,E., Shugart, Y., Ott, J. & Prusiner, S. B. (1993) Am. J. Hum.Genet. 33, 828-835.
43. Owen, F., Poulter, M., Collinge, J. & Crow, T. J. (1990) Am.J. Hum. Genet. 46, 1215-1216.
44. Kitamoto, T. & Tateishi, J. (1994) Phil. Trans. R. Soc. LondonB 343, 391-398.
45. Prusiner, S. B. & Hsiao, K. K. (1994) Ann. Neurol. 35, 385-395.
46. Harris, N. L., Presnell, S. R. & Cohen, F. E. (1994) J. Mol.Biol. 236, 1356-1368.
47. Presnell, S. R. & Cohen, F. E. (1989) Proc. Natd. Acad. Sci.USA 86, 6592-6596.
48. Diederichs, K., Boone, T. & Karplus, P. A. (1991) Science 254,1779-1782.
49. Dekker, N., Cox, M., Boelens, R., Vernier, C. P., Vandervliet,P. C. & Kaptein, R. (1993) Nature (London) 362, 852-855.
50. Goldfarb, L. G., Petersen, R. B., Tabaton, M., Brown, P.,LeBlanc, A. C., Montagna, P., Cortelli, P., Julien, J., Vital, C.,Pendelbury, W. W., Haltia, M., Wills, P. R., Hauw, J. J.,McKeever, P. E., Monari, L., Schrank, B., Swergold, G. D.,Autilio-Gambetti, L., Gajdusek, D. C., Lugaresi, E. & Gam-betti, P. (1992) Science 258, 806-808.
51. Gasset, M., Baldwin, M. A., Lloyd, D., Gabriel, J.-M., Holtz-man, D. M., Cohen, F., Fletterick, R. & Prusiner, S. B. (1992)Proc. Natl. Acad. Sci. USA 89, 10940-10944.
52. Rosenblatt, M., Beaudette, N. V. & Fasman, G. D. (1980)Proc. Natl. Acad. Sci. USA 77, 3983-3987.
53. Mottonen, J., Strand, A., Symersky, J., Sweet, R. M., Danley,D. E., Geoghegan, K. F., Gerard, R. D. & Goldsmith, E. J.(1992) Nature (London) 355, 270-273.
54. Monari, L., Chen, S. G., Brown, P., Parchi, P., Petersen,R. B., Mikol, J., Gray, F., Cortelli, P., Montagna, P., Ghetti,B., Goldfarb, L. G., Gaidusek, D. C., Lugaresi, E., Gambetti,P. & Autilio-Gambetti, L. (1994) Proc. Natl. Acad. Sci. USA91, 2839-2842.
55. Ripoll, L., Laplanche, J.-L., Salzmann, M., Jouvet, A.,Planques, B., Dussaucy, M., Chatelain, J., Beaudry, P. &Launay, J.-M. (1993) Neurology 43, 1934-1938.
56. Pocchiari, M., Salvatore, M., Cutruzzola, F., Genuardi, M.,Travaglini Allocatelli, C., Masullo, C., Macchi, G., Alema, G.,Galgani, S., Xi, Y. G., Petraroli, P., Silvestrini, M. C. &Brunori, M. (1993) Ann. Neurol. 34, 802-807.
57. Bfleler, H., Aguzzi, A., Sailer, A., Greiner, R.-A., Autenried,P., Aguet, M. & Weissmann, C. (1993) Cell 73, 1339-1347.
58. Prusiner, S. B., Groth, D., Serban, A., Koehler, R., Foster, D.,Torchia, M., Burton, D., Yang, S.-L. & DeArmond, S. J.(1993) Proc. Natl. Acad. Sci. USA 90, 10608-10612.
59. Hsiao, K. K., Scott, M., Foster, D., Groth, D. F., DeArmond,S. J. & Prusiner, S. B. (1990) Science 250, 1587-1590.
60. Westaway, D., DeArmond, S. J., Cayetano-Canlas, J., Groth,D., Foster, D., Yang, S.-L., Torchia, M., Carlson, G. A. &Prusiner, S. B. (1994) Cell 76, 117-129.
Biophysics: Huang et al.


















