
PROPERTIES OF THE ULTRAVIOLET-LIGHT-MEDIATED BINDING OF BOVINE SERUM ALBUMIN TO DNA
5
Phorochrmisrry ond Phorubiology. 1975. Vol. 21. pp. 243-241 Pergamon Press. Printed in Great Britain PROPERTIES OF THE ULTRAVIOLET-LIGHT-MEDIATED BINDING OF BOVINE SERUM ALBUMIN TO DNA* ANDREW BRAUN and BRUCE MERRICK Graduate Department of Biochemistry, Brandeis University, Waltham, Massachusetts 02154, U.S.A. (Received 8 March 1974; accepted 2 December 1974) Abstract-The binding of DNA to protein mediated by UV (254 nm) radiation has been investigated using binding of the complex to Millipore membrane filters as an assay technique. The reaction proceeds through an activated protein intermediate which then reacts with the DNA. The activated protein has a half-life ofabout 75 min at 0°C and about 18 min at 37°C. Short wavelengths are more efficient in form- ing the complex than wavelengths in the 25c280 nm range. N-ethyl maleimide treatment of protein before irradiation markedly inhibits the reaction. INTRODUCTION It has been known for some time that UVt irradiation of a concentrated mixture of protein and DNA results in a salt- and detergent-stable linkage between the two constituents (for a review, see Smith, 1975). The poss- ible significance of this interaction in uitro was demon- strated by Smith (1964), who found that such linkages could be observed in irradiated Escherichia coli and that these linkages seemed to be formed with a greater probability near the DNA replication point. This inter- action is often suggested as a cause of unusual action spectra, such as those for chromosome aberrations (Chu, 1965) and for killing (Todd et al., 1968) of Chinese hamster cells. In this communication, the UV-mediated binding of protein to DNA is measured by the simple filtration assay used previously by Markowitz (1972). Using this technique, we have found that the reaction can best be described as a two-step process: formation of an acti- vated state of protein by the UV radiation, followed by reaction of the activated protein with DNA to form a stable complex. MATERIALS AND METHODS C3H] 4X-174 RFI-DNA was isolated by the method of Schekman eta/. (1971).In general, we have omitted the final neutral sucrose gradient described in their method. This omission appears to leave the DNA contaminated with a substantial amount of unlabelled 260-nm-absorbing mater- ial, most probably ribonucleotides. We have been unable to detect any difference between this contaminated material and more purified DNA in the reactions described below. The labelled DNA had a specific activity of about 6 x lo4 cpm/nmol. Although the DNA is isolated as RFI, the DNA begins to deteriorate with time and is substantially ‘nicked‘ several months after isolation. We have not detected any dif- *Publication No. 1007 of the Graduate Department of Biochemistry, Brandeis University. Reprints will not be available to addresses in North America. t Abbreviations used: UV, Ultraviolet, 254 nm; BSA, Bovine serum albumin; NEM, N-ethyl maleimide; RFI, Replicative form I; ZXSSC, two-times saline citrate (0.3 M NaC1-0.03 M Na citrate). ference in the behaviour of ‘nicked’ circular DNA and in- tact, supercoiled, RFI in these assays. Booine serum albumin was obtained from Nutritional Bio- chemical Corp. and was dissolved in sterile water. N-etAyl maleimide (Calbiochem) was dissolved in sterile water im- mediately before use. Millipore HA WPJilters (25 mm) were stamped from large sheets. Just before use, the filters were wetted in water. Filtration was facilitated through the use of a 30-place filtration apparatus purchased from Millipore Corporation (Bedford, Mass.). Filtration assay. Assay mixtures were held at room tem- perature for 5 min after the end of the irradiation, then flooded with 5 mlp of ice-cold 2XSSC and held at 0°C for 5 rnin to allow salt-dissociable linkages to separate. The mixture was then poured over a membrane filter and filtered at a rate of about 10 mS?/min. The assay tube was washed with 5 mlp of cold 2XSSC and this was also filtered. The filter was dried and counted in a Packard scintillation counter using Liquiflor (New England Nuclear) as the scin- tillator. Controls of unirradiated protein and DNA gener- ally had less than 1 per cent of the DNA adhering to the filters. Irradiation. Solutions of protein and DNA were irra- diated in a 0.2-cm-thick quartz cuvette. UV radiation at 254 nm was from a Sylvania 15T8 bulb. Incident dose at the cuvette was measured by a “Blak-Ray” Ultraviolet Meter (Ultraviolet Products, Inc., San Gabriel, CA.). A Bausch & Lomb grating monochromator fitted with a high-pressure mercury lamp was used for irradiations at other wave- lengths. Incident UV dose was measured with an Eldorado photometer which had been calibrated with a chemical actinometer. For measurements of the action spectrum. the cuvette was placed in a Zeiss Q I1 spectrophotometer and the absorbance at the irradiation wavelength determined. Based upon this absorbance, an average dose to the sample was calculated according to the method of Morowitz (1950). The absorption of the cuvette was also considered and subtracted from the incident dose. Thus, dosage. as used in the action spectrum measurements, refers to the average dose in the sample. RESULTS When a BSA-C3H]-labelled-DNA mixture is UV- irradiated and then filtered through membrane filters. some of the labelled DNA adheres tightly to the filters. Figure 1 shows that the amount of DNA retained is di- rectly related to the incident UV dose and the protein concentration. The relationship between UV dose and retention is nonlinear at low doses. There is a linear 243
-
Upload
andrew-braun -
Category
Documents
-
view
213 -
download
0
Transcript of PROPERTIES OF THE ULTRAVIOLET-LIGHT-MEDIATED BINDING OF BOVINE SERUM ALBUMIN TO DNA

Phorochrmisrry ond Phorubiology. 1975. Vol. 21. pp. 243-241 Pergamon Press. Printed in Great Britain
PROPERTIES OF THE ULTRAVIOLET-LIGHT-MEDIATED BINDING OF BOVINE SERUM ALBUMIN TO DNA*
ANDREW BRAUN and BRUCE MERRICK Graduate Department of Biochemistry, Brandeis University, Waltham, Massachusetts 02154, U.S.A.
(Received 8 March 1974; accepted 2 December 1974)
Abstract-The binding of DNA to protein mediated by UV (254 nm) radiation has been investigated using binding of the complex to Millipore membrane filters as an assay technique. The reaction proceeds through an activated protein intermediate which then reacts with the DNA. The activated protein has a half-life ofabout 75 min at 0°C and about 18 min at 37°C. Short wavelengths are more efficient in form- ing the complex than wavelengths in the 25c280 nm range. N-ethyl maleimide treatment of protein before irradiation markedly inhibits the reaction.
INTRODUCTION
It has been known for some time that U V t irradiation of a concentrated mixture of protein and DNA results in a salt- and detergent-stable linkage between the two constituents (for a review, see Smith, 1975). The poss- ible significance of this interaction in uitro was demon- strated by Smith (1964), who found that such linkages could be observed in irradiated Escherichia coli and that these linkages seemed to be formed with a greater probability near the DNA replication point. This inter- action is often suggested as a cause of unusual action spectra, such as those for chromosome aberrations (Chu, 1965) and for killing (Todd et al., 1968) of Chinese hamster cells.
In this communication, the UV-mediated binding of protein to DNA is measured by the simple filtration assay used previously by Markowitz (1972). Using this technique, we have found that the reaction can best be described as a two-step process: formation of an acti- vated state of protein by the UV radiation, followed by reaction of the activated protein with DNA to form a stable complex.
MATERIALS AND METHODS
C3H] 4X-174 RFI-DNA was isolated by the method of Schekman eta/. (1971). In general, we have omitted the final neutral sucrose gradient described in their method. This omission appears to leave the DNA contaminated with a substantial amount of unlabelled 260-nm-absorbing mater- ial, most probably ribonucleotides. We have been unable to detect any difference between this contaminated material and more purified DNA in the reactions described below. The labelled DNA had a specific activity of about 6 x lo4 cpm/nmol. Although the DNA is isolated as RFI, the DNA begins to deteriorate with time and is substantially ‘nicked‘ several months after isolation. We have not detected any dif-
*Publication No. 1007 of the Graduate Department of Biochemistry, Brandeis University. Reprints will not be available to addresses in North America.
t Abbreviations used: UV, Ultraviolet, 254 nm; BSA, Bovine serum albumin; NEM, N-ethyl maleimide; RFI, Replicative form I; ZXSSC, two-times saline citrate (0.3 M NaC1-0.03 M Na citrate).
ference in the behaviour of ‘nicked’ circular DNA and in- tact, supercoiled, RFI in these assays.
Booine serum albumin was obtained from Nutritional Bio- chemical Corp. and was dissolved in sterile water. N-etAyl maleimide (Calbiochem) was dissolved in sterile water im- mediately before use.
Millipore H A WPJilters (25 mm) were stamped from large sheets. Just before use, the filters were wetted in water. Filtration was facilitated through the use of a 30-place filtration apparatus purchased from Millipore Corporation (Bedford, Mass.).
Filtration assay. Assay mixtures were held at room tem- perature for 5 min after the end of the irradiation, then flooded with 5 mlp of ice-cold 2XSSC and held at 0°C for 5 rnin to allow salt-dissociable linkages to separate. The mixture was then poured over a membrane filter and filtered at a rate of about 10 mS?/min. The assay tube was washed with 5 mlp of cold 2XSSC and this was also filtered. The filter was dried and counted in a Packard scintillation counter using Liquiflor (New England Nuclear) as the scin- tillator. Controls of unirradiated protein and DNA gener- ally had less than 1 per cent of the DNA adhering to the filters.
Irradiation. Solutions of protein and DNA were irra- diated in a 0.2-cm-thick quartz cuvette. UV radiation at 254 nm was from a Sylvania 15T8 bulb. Incident dose at the cuvette was measured by a “Blak-Ray” Ultraviolet Meter (Ultraviolet Products, Inc., San Gabriel, CA.). A Bausch & Lomb grating monochromator fitted with a high-pressure mercury lamp was used for irradiations at other wave- lengths. Incident UV dose was measured with an Eldorado photometer which had been calibrated with a chemical actinometer. For measurements of the action spectrum. the cuvette was placed in a Zeiss Q I1 spectrophotometer and the absorbance at the irradiation wavelength determined. Based upon this absorbance, an average dose to the sample was calculated according to the method of Morowitz (1950). The absorption of the cuvette was also considered and subtracted from the incident dose. Thus, dosage. as used in the action spectrum measurements, refers to the average dose in the sample.
RESULTS
When a BSA-C3H]-labelled-DNA mixture is UV- irradiated and then filtered through membrane filters. some of the labelled DNA adheres tightly to the filters. Figure 1 shows that the amount of DNA retained is di- rectly related to the incident UV dose and the protein concentration. The relationship between UV dose and retention is nonlinear a t low doses. There is a linear
243
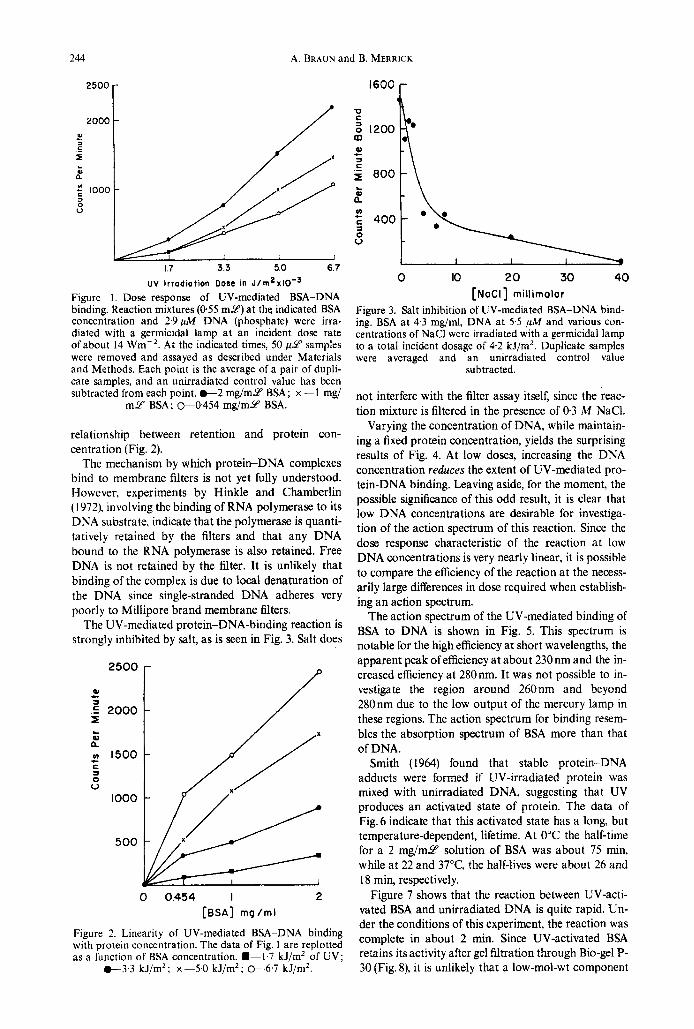
244 A. BRAUN and B. MERRICK
2500 r 2000
0)
3 c
E E c
a" ; 1000 = 0 V
UV Irradiation Dose in J / r n ' ~ l O - ~
Figure 1, Dose-response of UV-mediated BSA-DNA binding. Reaction mixtures (0.55 m u ) at the indicated BSA concentration and 2.9 pA4 DNA (phosphate) were irra- diated with a germicidal lamp at an incident dose rate ofabout 14 Wrn-'. At the indicated times, 50 p Y samples were removed and assayed as described under Materials and Methods. Each point is the average of a pair of dupli- cate samples, and an unirradiated control value has been subtracted from each point. 0 - 2 mg/m-Y BSA; x -1 mg/
m2' BSA; 0-0.454 mg/mY BSA.
relationship between retention and protein con- centration (Fig. 2).
The mechanism by which protein-DNA complexes bind to membrane filters is not yet fully understood. However, experiments by Hinkle and Chamberlin ( 1 972), involving the binding of RNA polymerase to its DNA substrate, indicate that the polymerase is quanti- tatively retained by the filters and that any DNA bound to the RNA polymerase is also retained. Free DNA is not retained by the filter. It is unlikely that binding of the complex is due to local denaturation of the DNA since single-stranded DNA adheres very poorly to Millipore brand membrane filters.
The UV-mediated protein-DNA-binding reaction is strongly inhibited by salt, as is seen in Fig. 3. Salt does
2500
al a t.
.E 2000 ' I
n L al
v) 1500 t. c a 0 0
I000
500
0 0.454 I 2 [BSA] m g / m l
Figure 2. Linearity of UV-mediated BSA-DNA binding with protein concentration. The data of Fig. 1 are replotted as a function of BSA concentration. .--1.7 kJ/mZ of UV;
t 3 . 3 kJ/m2; x-5.0 kJ/m2; -6.7 kJ/m2.
1600 r
0 10 20 30 40 [NaCI] millimolar
Figure 3. Salt inhibition of UV-mediated BSA-DNA bind- ing. BSA at 4.3 mg/ml, DNA at 5.5 p M and various con- centrations of NaCl were irradiated with a germicidal lamp to a total incident dosage of 4 2 kJ/mz. Duplicate samples were averaged and an unirradiated control value
subtracted.
not interfere with the filter assay itself, since the reac- tion mixture is filtered in the presence of 0.3 M NaCl.
Varying the concentration of DNA, while maintain- ing a fixed protein concentration, yields the surprising results of Fig. 4. At low doses, increasing the DNA concentration reduces the extent of UV-mediated pro- tein-DNA binding. Leaving aside, for the moment, the possible significance of this odd result, it is clear that low DNA concentrations are desirable for investiga- tion of the action spectrum of this reaction. Since the dose response characteristic of the reaction a t low DNA concentrations is very nearly linear, it is possible to compare the efficiency of the reaction at the necess- arily large differences in dose required when establish- ing a n action spectrum.
The action spectrum of the UV-mediated binding of BSA to DNA is shown in Fig. 5. This spectrum is notable for the high efficiency at short wavelengths, the apparent peak of efficiency a t about 230 nm and the in- creased efficiency at 280nm. It was not possible to in- vestigate the region around 260nm and beyond 280nm due to the low output of the mercury lamp in these regions. The action spectrum for binding resem- bles the absorption spectrum of BSA more than that of DNA.
Smith (1964) found that stable protein-DNA adducts were formed if UV-irradiated protein was mixed with unirradiated DNA, suggesting that UV produces a n activated state of protein. The data of Fig. 6 indicate that this activated state has a long, but temperature-dependent, lifetime. At 0°C the half-time for a 2 m g / m 9 solution of BSA was about 75 min, while a t 22 and 3 7 T , the half-lives were about 26 and 18 min, respectively.
Figure 7 shows that the reaction between UV-acti- vated BSA and unirradiated DNA is quite rapid. Un- der the conditions of this experiment, the reaction was complete in about 2 min. Since UV-activated BSA retains its activity after gel filtration through Bio-gel P- 30 (Fig. 8), it is unlikely that a low-mol-wt component
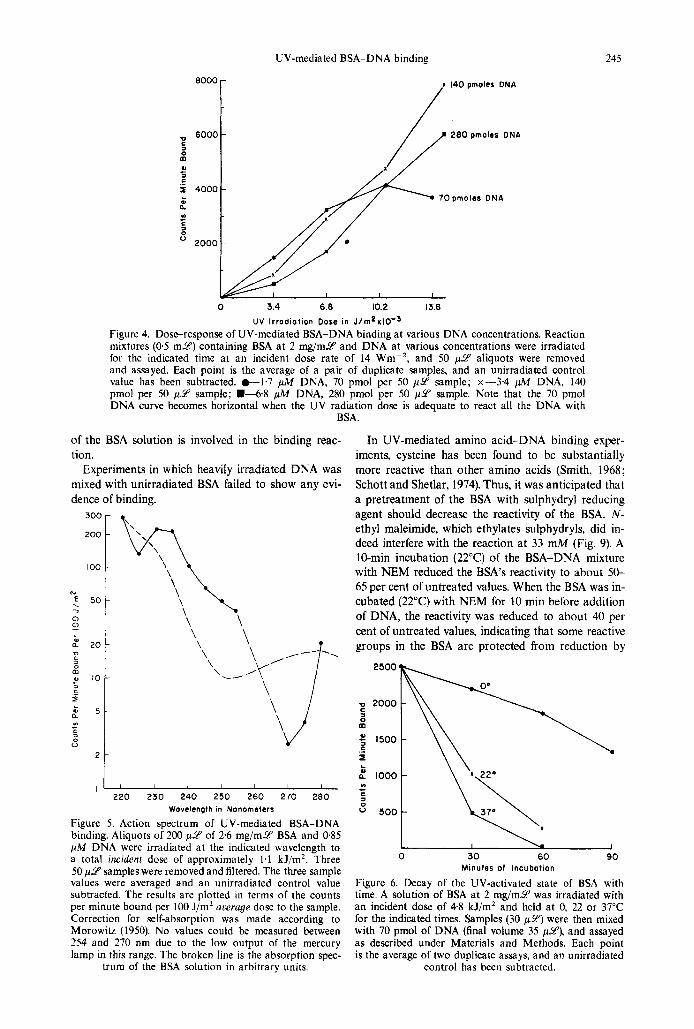
UV-mediated BSA-DNA binding 245
8oool 6000
C =- 2
x 140 pmolrr DNA
/ 8000 -
i 140 pmolrr DNA
280 pmolcs ONA C =-
as
c
6 0 0 0 -
2 .- 5 4000- L as
In C
70pmolas DNA n
a c
2000 -
0 3.4 6.8 10.2 I3 6
UV lrrodiotion Dose in J / ~ n ~ x l O - ~ Figure 4. Doseresponse of UV-mediated BSA-DNA binding at various DNA concentrations. Reaction mixtures (0.5 m y ) containing BSA a t 2 mg/mU and DNA at various concentrations were irradiated for the indicated time at an incident dose rate of 14 Wm-*, and 50 p2' aliquots were removed and assayed. Each point is the average of a pair of duplicate samples, and an unirradiated control value has been subtracted. t 1 . 7 pA4 DNA, 70 pmol per 50 p2' sample; x-3.4 pA4 DNA, 140 pmol per 50 p 9 sample; L 6 . 8 fl DNA, 280 pmol per 50 p 9 sample. Note that the 70 pmol DNA curve becomes horizontal when the UV radiation dose is adequate to react all the DNA with
BSA.
of the BSA solution is involved in the binding reac- tion.
Experiments in which heavily irradiated DNA was mixed with unirradiated BSA failed to show any evi- dence of binding.
300
zoo
too
N 5 50
0 P
7
L
2 2 0 D
3 0 m 42 10
3
3
$ 5 Y)
3 0 u
- 2
I
1,
\\\ \ \ \
\ \
\ \ \ \
\
\
220 230 240 250 260 270 280 Wovelength in Nonometers
Figure 5. Action spectrum of UV-mediated BSA-DNA binding. Aliquots of 200 p Y of 2 6 mg/mY BSA and 0.85 ptM DNA were irradiated at the indicated wavelength to a total incident dose of approximately 1.1 kJ/mZ. Three 50 p6p samples were removed and filtered. The three sample values were averaged and an unirradiated control value subtracted. The results are plotted in terms of the counts per minute bound per 100 J/m' average dose to the sample. Correction for self-absorption was made according to Morowitz (1950). No values could be measured between 254 and 270 nm due to the low output of the mercury lamp in this range. The broken line is the absorption spec-
trum of the BSA solution in arbitrary units.
In UV-mediated amino acid-DNA binding exper- iments, cysteine has been found to be substantially more reactive than other amino acids (Smith, 1968; Schott and Shetlar, 1974). Thus, it was anticipated that a pretreatment of the BSA with sulphydryl reducing agent should decrease the reactivity of the BSA. N - ethyl maleimide, which ethylates sulphydryls, did in- deed interfere with the reaction at 33 mM (Fig. 9). A 10-min incubation (22°C) of the BSA-DNA mixture with NEM reduced the BSA's reactivity to about 50- 65 per cent of untreated values. When the BSA was in- cubated (22°C) with NEM for 10 min before addition of DNA, the reactivity was reduced to about 40 per cent of untreated values, indicating that some reactive groups in the BSA are protected from reduction by
~~~~
0 30 60 90 Minutes of Incubation
Figure 6. Decay of the UV-activated state of BSA with time. A solution of BSA a t 2 mg/mY was irradiated with an incident dose of 4.8 kJ/m2 and held at 0, 22 or 37°C for the indicated times. Samples (30 p2') were then mixed with 70 pmol of DNA (final volume 35 p y ) , and assayed as described under Materials and Methods. Each point is the average of two duplicate assays, and an unirradiated
control has been subtracted.
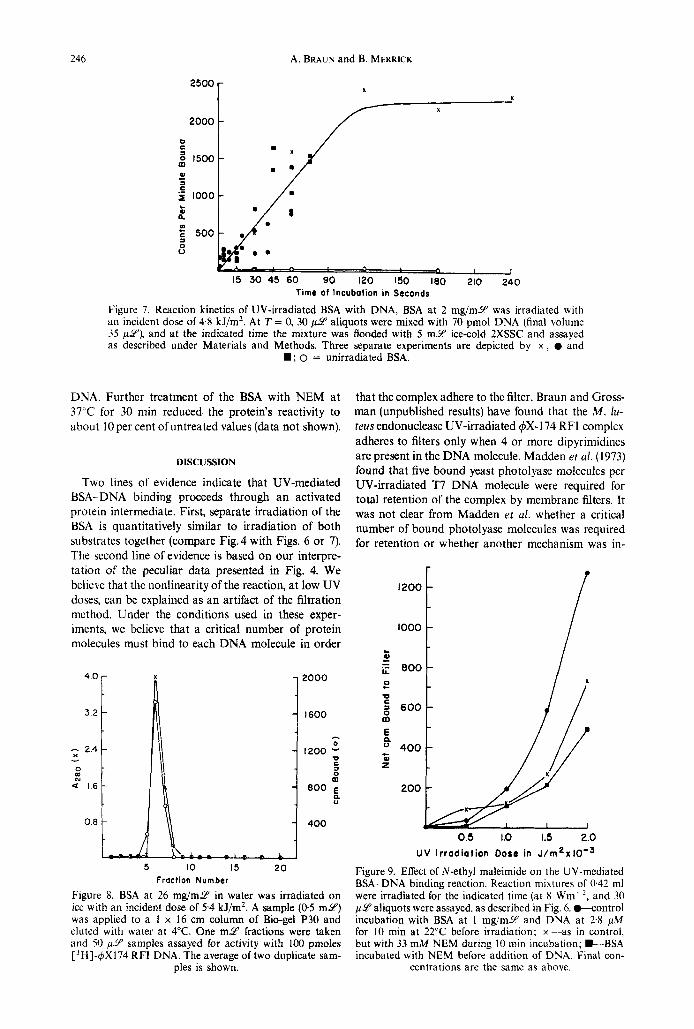
246 A. BRAUN and B. MERRICK
2000 25001
a
c
A 0 I I 15 30 45 60 90 120 60 180 210 240
Time of lncubalion in Seconds
Figure 7. Reaction kinetics of UV-irradiated BSA with DNA, BSA at 2 mg/mY was irradiated with an incident dose of 4.8 kJ/m2. At T = 0, 30 pcY aliquots were mixed with 70 pmol DNA (final volume 35 p iu ) , and at the indicated time the mixture was flooded with 5 m Y ice-cold ZXSSC and assayed as described under Materials and Methods. Three separate experiments are depicted by x , 0 and
D; 0 = unirradiated BSA.
DNA. Further treatment of the BSA with NEM at 3 7 T for 30 min reduced the protein's reactivity to about 10 per cent ofuntreated values (data not shown).
DISCUSSION
Two lines of evidence indicate that UV-mediated BSA-DNA binding proceeds through a n activated protein intermediate. First, separate irradiation of the BSA is quantitatively similar to irradiation of both substrates together (compare Fig.4 with Figs. 6 or 7). The second line of evidence is based on our interpre- tation of the peculiar data presented in Fig. 4. We believe that the nonlinearity of the reaction, a t low UV doses, can be explained as an artifact of the filtration method. Under the conditions used in these exper- iments, we believe that a critical number of protein molecules must bind to each DNA molecule in order
401 3 2 I - 0
1200 ;;
h
C
0 m 800
400
5 10 15 20 Fraction Number
Figure 8. BSA at 26 mg/mY in water was irradiated on ice with an incident dose of 5.4 kJ/m2. A sample (05 m y ) was applied to a 1 x 16 cm column of Bio-gel P30 and eluted with water at 4°C. One m Y fractions were taken arid 50 p 9 samples assayed for activity with 100 pmoles ['H]-q5X174 RFI DNA. The average of two duplicate sam-
ples is shown.
that the complex adhere to the filter. Braun and Gross- man (unpublished results) have found that the M . lu- teus endonuclease UV-irradiated $X-I 74 RFI complex adheres to filters only when 4 or more dipyrimidines are present in the DNA molecule. Madden et a/ . (1973) found that five bound yeast photolyase molecules per UV-irradiated T7 DNA molecule were required for total retention of the complex by membrane filters. It was not clear from Madden et a[. whether a critical number of bound photolyase molecules was required for retention or whether another mechanism was in-
I2O0 t
U V I r rad ia t ion Dose in J / 1 n ~ x I 0 - ~
Figure 9. Effect of N-ethyl maleimide on the UV-mediated BSA-DNA binding reaction. Reaction mixtures of 0.42 ml were irradiated for the indicated time (at 8 Wm-', and 30 p Y aliquots were assayed, as described in Fig. 6. w o n t r o l incubation with BSA at 1 mg/mY and DNA at 2.8 pA4 for 10 min at 22°C before irradiation; x--as in control, but with 33 mM NEM during 10 min incubation; D-BSA incubated with NEM before addition of DNA. Final con-
centrations are the same as above.
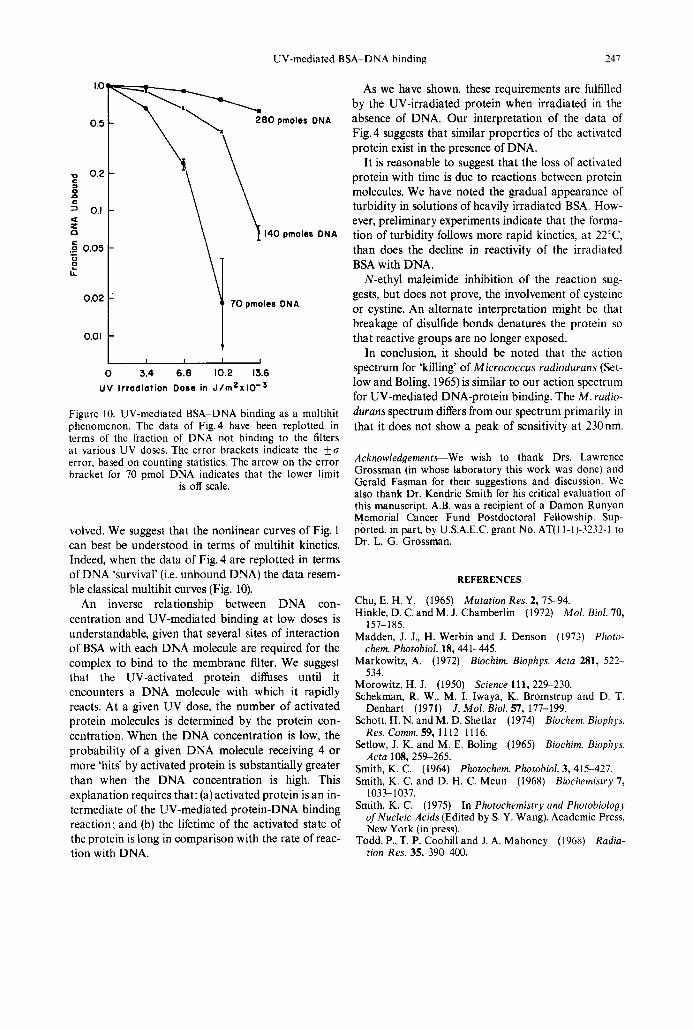
UV-mediated BSA-DNA binding 247
I .o
0.5
0.2 C
c1 C
a
= 0.1 a n
.E 0.05
e
z
c
LL
0.02
0.0 I
0 3.4 6.8 10.2 13.6 U V Irradiation Dose in J / m Z x I O - s
Figure 10. UV-mediated BSA-DNA binding as a multihit phenomenon. The data of Fig.4 have been replotted in terms of the fraction of DNA not binding to the filters a t various UV doses. The error brackets indicate the ku error, based on counting statistics. The arrow on the error bracket for 70 pmol DNA indicates that the lower limit
is off scale.
volved. We suggest that the nonlinear curves of Fig. 1 can best be understood in terms of multihit kinetics. Indeed, when the data of Fig. 4 are replotted in terms of DNA ‘survival’ (i.e. unbound DNA) the data resem- ble classical multihit curves (Fig. 10).
An inverse relationship between DNA con- centration and UV-mediated binding at low doses is understandable, given that several sites of interaction of BSA with each DNA molecule are required for the complex to bind to the membrane filter. We suggest that the UV-activated protein diffuses until it encounters a DNA molecule with which it rapidly reacts. At a given UV dose, the number of activated protein molecules is determined by the protein con- centration. When the DNA concentration is low, the probability of a given DNA molecule receiving 4 or more ‘hits’ by activated protein is substantially greater than when the DNA concentration is high. This explanation requires that: (a) activated protein is an in- termediate of the UV-mediated protein-DNA binding reaction; and (b) the lifetime of the activated state of the protein is long in comparison with the rate of reac- tion with DNA.
As we have shown, these requirements are fulfilled by the UV-irradiated protein when irradiated in the absence of DNA. Our interpretation of the data of Fig.4 suggests that similar properties of the activated protein exist in the presence of DNA.
It is reasonable to suggest that the loss of activated protein with time is due to reactions between protein molecules. We have noted the gradual appearance of turbidity in solutions of heavily irradiated BSA. How- ever, preliminary experiments indicate that the forma- tion of turbidity follows more rapid kinetics, a t 2 2 T , than does the decline in reactivity of the irradiated BSA with DNA.
N-ethyl maleimide inhibition of the reaction sug- gests, but does not prove, the involvement of cysteine or cystine. An alternate interpretation might be that breakage of disulfide bonds denatures the protein so that reactive groups are no longer exposed.
In conclusion, it should be noted that the action spectrum for ‘killing’ of Micrococcus rudiodurans (Set- low and Boling, 1965) is similar to our action spectrum for UV-mediated DNA-protein binding. The M . rudio- durans spectrum differs from our spectrum primarily in that it does not show a peak of sensitivity a t 230nm.
Acknowledgements-We wish to thank Drs. Lawrence Grossman (in whose laboratory this work was done) and Gerald Fasman for their suggestions and discussion. We also thank Dr. Kendric Smith for his critical evaluation of this manuscript. A.B. was a recipient of a Damon Runyon Memorial Cancer Fund Postdoctoral Fellowship. Sup- ported. in part. by U.S.A.E.C. grant No. AT(I 1-1)-3232-l to Dr. L. G. Grossman.
REFERENCES
Chu, E. H. Y. (1965) Mutation Res. 2, 75-94. Hinkle, D. C. and M. J. Chamberlin (1972) Mol. B id . 70,
Madden, J. J., H. Werbin and J. Denson (1973) Photo-
Markowitz, A. (1972) Eiochim. Biophys. Acta 281, 522-
Morowitz, H. J. (1950) Science 111,229-230. Schekman, R. W . , M. I. Iwaya, K. Bromstrup and D. T.
Denhart (1971) J. Mol. Biol. 57, 177-199. Schott, H. N. and M. D. Shetlar (1974) Biochem. Biophys.
Res. Comm. 59, 1 112-1 116. Setlow, J. K. and M. E. Boling (1965) Biochirn. Biophys.
Acta 108,259-265. Smith, K. C. (1964) Photochem. Photohiol. 3, 415-427. Smith, K. C. and D. H. C. Meun (1968) Biochemistry 7,
Smith, K. C. (1975) In Photochemistry and Photobiology of Nucleic Acids (Edited by S. Y. Wang). Academic Press, New York (in press).
Radia- tion Res. 35. 390-400.
157- 185.
chem. Photobiol. 18,441-445.
534.
1033-1037.
Todd. P.. T. P. Coohill and J . A. Mahoney (1968)


















