
Production and Characterization Exopolysaccharide …aem.asm.org/content/60/11/4134.full.pdf ·...
8
Vol. 60, No. 11 APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Nov. 1994, p. 4134-4141 0099-2240/94/$04.00+0 Copyright ©D 1994, American Society for Microbiology Production and Characterization of an Exopolysaccharide Excreted by a Deep-Sea Hydrothermal Vent Bacterium Isolated from the Polychaete Annelid Alvinella pompejanat P. VINCENT,1 P. PIGNET,2 F. TALMONT,3 L. BOZZI,4 B. FOURNET,3 J. GUEZENNEC, C. JEANTHON,1 AND D. PRIEURl* LP 4601, Station Biologique, Centre National de la Recherche Scientifique (CNRS), 29682 Roscoff Cedex,1 IFREMER, DERO/EP, Centre de Brest, 29280 Plouzane,2 UMR 111, Laboratoire de Chimie Biologique, UST Lille, CNRS, 59650 Villeneuve d Ascq,3 and CERMAV, CNRS, 38041 Grenoble,4 France Received 9 March 1994/Accepted 30 August 1994 The heterotrophic and mesophilic marine bacterium HYD-1545 was isolated on a metal-amended medium from the dorsal integument of the hydrothermal vent polychaete Alvinella pompejana. This strain, which can be assigned to the genus Alteromonas on the basis of its G+C content and phenotypical features, produced large amounts of an acidic polysaccharide in batch cultures. The polysaccharide was excreted during the stationary phase of growth and contained glucose, galactose, glucuronic acid, galacturonic acid, and 4,6-O-(1-carboxyethi- lidene)-galactose as major components. This polysaccharide was a polyelectrolyte, and the viscosity of its solutions depended on the ionic strength. The decrease in viscosity with increasing NaCl concentrations and the effect of Ca21 in decreasing the viscosity at low Ca2+ concentrations support a model in which the polysaccharide carries anionic groups. However, an unusual behavior was observed at higher concentrations and could be related to intermolecular interactions involving Ca2+ ions. The adhesion of bacteria to solid surfaces in aquatic envi- ronments involving exocellular polysaccharides has been inves- tigated from an ecological and structural standpoint by using light and electron microscopy (10, 16, 35). Many authors have discussed the role and properties of these polymers in the attachment and survival of the cells and their interactions with ions, such as heavy metals (30, 49). More recently, consider- able attention has been focused on the chemical characteriza- tion of novel exopolysaccharides (EPS) excreted by marine bacteria (8, 33) in association with the study of their rheologi- cal properties (6) or antitumoral activities (32). Only a few interesting molecules, e.g., Marinactan (45), have yet been found, purified, and totally characterized. In the course of searches for new marine biopolymers of potential use to industry, 479 heterotrophic bacteria isolated from various deep-sea hydrothermal vent samples were screened for their ability to produce EPS (46). These ecosystems are character- ized by the existence of rich animal communities explained by the abundance of local chemosynthetic bacterial primary pro- duction (23), and several associations between bacteria and invertebrates have been described. The strain HYD-1545 examined in this study was isolated from the dorsal integument of the marine polychaete Alvinella pompejana (12), which builds mineralized organic tubes on the walls of sulfide diffus- ers. This worm lives in areas where hot, acidic, and metal-rich hydrothermal fluids are mixing with cold and well-oxygenated seawater (11). MATERIALS AND METHODS Microorganisms. The bacterium HYD-1545 was isolated from the epidermis of the hydrothermal vent polychaete A. pompejana, collected in 1987 during the "Hydronaut" cruise by * Corresponding author. Mailing address: CNRS. LP 4601, Station Biologique, B.P. 74, 29682 Roscoff Cedex, France. t This article is dedicated to B. Fournet. the submersible Nautile at 13°N on the East Pacific Rise (2,600 m depth, 12°48.56'N, 103°56.72'W). While we were searching for marine bacteria producing EPS, this strain was selected for its ability to show a mucoid phenotype on solid medium and to produce viscous broth. Vibrio natriegens (DMS 759) used as a reference for polyhydroxybutyrate synthesis was obtained from the Deutsche Sammlung von mikroorganismen und Zellkul- turen collection (RFA). Alteromonas haloplanktis used as a reference for the G+C content measurements was a gift of M. J. Gauthier Institut National de la Sante et de la Recherche Medicale, Unite 303, Nice, France). Media. The strain HYD-1545 was isolated on a metal- amended medium (24) prepared by adding to the 2216E medium (34) 1 mg of AsO43- ml-' and was tested for poly- saccharide production in shake flasks by using a sugar-contain- ing medium composed of the following: 5 g of peptone, 1 g of yeast extract, 250 ml of distilled water, 750 ml of filtered seawater, and 30 g of glucose (31). The medium was supple- mented with agar (20 g/liter) for culture on solid medium. The strain isolated in pure culture was preserved by being frozen in glycerol (20% [vol/vol]) at -20°C, and from these organisms precultures for each experiment were prepared. Medium 2216E was used for the determination of the optimal temper- ature for growth. Optimal pH was then determined by using the same medium supplemented with Tris-HCl (50 mM) in a pH range of 7.5 to 8.0, MOPS (morpholinepropanesulfonic acid; 50 mM) in a pH range of 7.0 to 7.5, and PIPES (piperazine-N,N'-bis(2-ethanesulfonic acid); 20 mM] in a pH range of 5.5 to 7.0. The optimal ionic strength was determined by using a modification of a medium described by Boyle and Reade (6) containing the following: MgCl2 6H20, 5.9 g; Na2SO4, 3.24 g; CaCl2 * 2H20, 1.8 g; KCl, 0.55 g; NaHCO3, 0.16 g; ferrous citrate, 0.1 g; tryptone, 0.7 g; yeast extract, 1 g; and distilled water, 1 liter. NaCl in a range of S to 70% was added. Culture conditions. In all experiments concerning EPS production, plates and flasks were incubated for 5 days at 23°C 4134 on May 23, 2018 by guest http://aem.asm.org/ Downloaded from
Transcript of Production and Characterization Exopolysaccharide …aem.asm.org/content/60/11/4134.full.pdf ·...

Vol. 60, No. 11APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Nov. 1994, p. 4134-41410099-2240/94/$04.00+0Copyright ©D 1994, American Society for Microbiology
Production and Characterization of an Exopolysaccharide Excreted bya Deep-Sea Hydrothermal Vent Bacterium Isolated from the
Polychaete Annelid Alvinella pompejanatP. VINCENT,1 P. PIGNET,2 F. TALMONT,3 L. BOZZI,4 B. FOURNET,3 J. GUEZENNEC,
C. JEANTHON,1 AND D. PRIEURl*
LP 4601, Station Biologique, Centre National de la Recherche Scientifique (CNRS), 29682 Roscoff Cedex,1IFREMER, DERO/EP, Centre de Brest, 29280 Plouzane,2 UMR 111, Laboratoire de Chimie Biologique,
UST Lille, CNRS, 59650 Villeneuve d Ascq,3 and CERMAV, CNRS, 38041 Grenoble,4 France
Received 9 March 1994/Accepted 30 August 1994
The heterotrophic and mesophilic marine bacterium HYD-1545 was isolated on a metal-amended mediumfrom the dorsal integument of the hydrothermal vent polychaete Alvinella pompejana. This strain, which can beassigned to the genus Alteromonas on the basis of its G+C content and phenotypical features, produced largeamounts of an acidic polysaccharide in batch cultures. The polysaccharide was excreted during the stationaryphase of growth and contained glucose, galactose, glucuronic acid, galacturonic acid, and 4,6-O-(1-carboxyethi-lidene)-galactose as major components. This polysaccharide was a polyelectrolyte, and the viscosity of itssolutions depended on the ionic strength. The decrease in viscosity with increasing NaCl concentrations andthe effect of Ca21 in decreasing the viscosity at low Ca2+ concentrations support a model in which thepolysaccharide carries anionic groups. However, an unusual behavior was observed at higher concentrationsand could be related to intermolecular interactions involving Ca2+ ions.
The adhesion of bacteria to solid surfaces in aquatic envi-ronments involving exocellular polysaccharides has been inves-tigated from an ecological and structural standpoint by usinglight and electron microscopy (10, 16, 35). Many authors havediscussed the role and properties of these polymers in theattachment and survival of the cells and their interactions withions, such as heavy metals (30, 49). More recently, consider-able attention has been focused on the chemical characteriza-tion of novel exopolysaccharides (EPS) excreted by marinebacteria (8, 33) in association with the study of their rheologi-cal properties (6) or antitumoral activities (32). Only a fewinteresting molecules, e.g., Marinactan (45), have yet beenfound, purified, and totally characterized. In the course ofsearches for new marine biopolymers of potential use toindustry, 479 heterotrophic bacteria isolated from variousdeep-sea hydrothermal vent samples were screened for theirability to produce EPS (46). These ecosystems are character-ized by the existence of rich animal communities explained bythe abundance of local chemosynthetic bacterial primary pro-duction (23), and several associations between bacteria andinvertebrates have been described. The strain HYD-1545examined in this study was isolated from the dorsal integumentof the marine polychaete Alvinella pompejana (12), whichbuilds mineralized organic tubes on the walls of sulfide diffus-ers. This worm lives in areas where hot, acidic, and metal-richhydrothermal fluids are mixing with cold and well-oxygenatedseawater (11).
MATERIALS AND METHODS
Microorganisms. The bacterium HYD-1545 was isolatedfrom the epidermis of the hydrothermal vent polychaete A.pompejana, collected in 1987 during the "Hydronaut" cruise by
* Corresponding author. Mailing address: CNRS. LP 4601, StationBiologique, B.P. 74, 29682 Roscoff Cedex, France.
t This article is dedicated to B. Fournet.
the submersible Nautile at 13°N on the East Pacific Rise (2,600m depth, 12°48.56'N, 103°56.72'W). While we were searchingfor marine bacteria producing EPS, this strain was selected forits ability to show a mucoid phenotype on solid medium and toproduce viscous broth. Vibrio natriegens (DMS 759) used as areference for polyhydroxybutyrate synthesis was obtained fromthe Deutsche Sammlung von mikroorganismen und Zellkul-turen collection (RFA). Alteromonas haloplanktis used as areference for the G+C content measurements was a gift of M.J. Gauthier Institut National de la Sante et de la RechercheMedicale, Unite 303, Nice, France).
Media. The strain HYD-1545 was isolated on a metal-amended medium (24) prepared by adding to the 2216Emedium (34) 1 mg of AsO43- ml-' and was tested for poly-saccharide production in shake flasks by using a sugar-contain-ing medium composed of the following: 5 g of peptone, 1 g ofyeast extract, 250 ml of distilled water, 750 ml of filteredseawater, and 30 g of glucose (31). The medium was supple-mented with agar (20 g/liter) for culture on solid medium. Thestrain isolated in pure culture was preserved by being frozen inglycerol (20% [vol/vol]) at -20°C, and from these organismsprecultures for each experiment were prepared. Medium2216E was used for the determination of the optimal temper-ature for growth. Optimal pH was then determined by usingthe same medium supplemented with Tris-HCl (50 mM) in apH range of 7.5 to 8.0, MOPS (morpholinepropanesulfonicacid; 50 mM) in a pH range of 7.0 to 7.5, and PIPES(piperazine-N,N'-bis(2-ethanesulfonic acid); 20 mM] in a pHrange of 5.5 to 7.0. The optimal ionic strength was determinedby using a modification of a medium described by Boyle andReade (6) containing the following: MgCl2 6H20, 5.9 g;Na2SO4, 3.24 g; CaCl2 * 2H20, 1.8 g; KCl, 0.55 g; NaHCO3,0.16 g; ferrous citrate, 0.1 g; tryptone, 0.7 g; yeast extract, 1 g;and distilled water, 1 liter. NaCl in a range of S to 70% wasadded.
Culture conditions. In all experiments concerning EPSproduction, plates and flasks were incubated for 5 days at 23°C
4134
on May 23, 2018 by guest
http://aem.asm
.org/D
ownloaded from

EPS-PRODUCING BACTERIUM FROM VENT POLYCHAETE 4135
andEPS were obtained during the stationary phase of growth.Flasks (500 ml) containing 100 ml of glucose-containing me-dium were inoculated with a loopful of the primary bacterialculture and incubated at 150 rpm on a shaker. Optimal growthconditions were determined in 1-liter flasks containing 200 mlof the appropriate medium. Cultures were incubated for 36 h,and cell growth was monitored spectrophotometrically (A520).The media were inoculated with 4 ml of a bacterial suspensionspectrophotometrically standardized at an optical density at520 nm of 0.1 which was prepared from cells cultured overnighton the same medium. The doubling times were calculated fromthe slopes of the growth curves. The optimal temperature forgrowth was determined for cultures incubated at 16, 22, 25, 27,29, 31, 33, 35, and 39°C, at pH 7.2. The effect of pH on growthwas tested at pH 5.7, 6.0, 6.5, 7.0, 7.3, 7.5, and 7.8, at theoptimal temperature, which was 27°C. The optimal ionicstrength for growth was determined at 27°C and pH 7.3 byusing 11 different NaCl concentrations, 0, 5, 10, 20, 25, 30, 40,50, 60, and 70 g/liter.
Morphological, biochemical, and nutritional tests. Thepresence of a capsule was determined by using wet mountswith India ink and the staining technique of Allison andSutherland (1). Flagellation was characterized by the stainingmethod of Rhodes described by Conn et al. (9). The occur-rence of an oxidase reaction was determined by using one-daycultures according to the procedure of Kovacs (26). H2Sproduction, glucose fermentation, and Tween degradationwere recorded after 7 days by the methods described byHarrigan and McCance (22). Proteolytic activity on gelatin wastested as described by Kohn (25). The presence of a constitu-tive arginine dihydrolase system was determined by themethod of Baumann and Baumann (2). Amounts of indoleproduction, nitrate and nitrite reduction, starch hydrolysis, andurease, catalase, and 3-galactosidase activities were deter-mined by standard methods (27). Polyhydroxybutyrate accu-mulation was tested according to the method of Baumann andBaumann (2) for 4 days, with V natriegens (DSM 759) servingas a control strain. Chitinase activity was tested according tothe method of Reichenbach and Dworkin (38). The utilizationof 19 sugars and 16 other carbohydrate substrates as the solesource of carbon was tested according the method of Baumannand Baumann (2) by using a basal medium (BM) supple-mented with the substrate at a final concentration of 0.2%(wt/vol for solids; vol/vol for liquids) for sugars and 0.1%(wt/vol for solids; vol/vol for liquids) for the other carbohydratesubstrates. Most carbohydrate solutions were sterilized sepa-rately by being autoclaved at 120°C for 20 min, except xylose,fructose, and maltose solutions, which were sterilized byfiltration with 0.2-,um-pore-size filter units (Merck). DNA wasprepared as described by Beji et al. (4), and the GC contentwas determined by both high-performance liquid chromatog-raphy (HPLC) (36) and the method of Ulitzur (44).
Viscosimetry. Solution viscosity was measured with a Brook-field viscosimeter model DV-II supplied with a small sampleadapter (SC4-18/13R) at 25°C and 30 rpm unless otherwiseindicated (Brookfield Engineering Laboratories, Stoughton,Mass.). Samples were maintained at the appropriate temper-ature for 5 min prior to measurements of viscosity.
Polysaccharide production. The EPS production by strainHYD-1545 was studied by using a 2-liter fermentor (LSLBiolafitte, St-Germain-en-Laye, France) initially containing 1.4liters of the appropriate medium. Batch fermentation wasstarted by inoculating a suspension of cells (2% [vol/vol])grown overnight in 2216E medium into the fermentor. Thetemperature was maintained at 25°C. Air was supplied at 40liters per volume of medium per h, and the culture was mixed
at an impeller speed within a range of 350 to 800 rpm. The pHwas adjusted to 7.2 at the beginning of the culture andmeasured but not controlled during the experiment. Samples(40 ml) were removed at regular intervals for growth andviscosity measurements, EPS isolation, and residual glucoseanalysis. The viscosity of the culture broth was measured asdescribed above, and the residual concentration of glucose inthe medium was determined enzymatically by using a reagentkit purchased from Boehringer Mannheim Biochemicals(Mannheim, Germany). Growth was measured by CFU countsand turbidimetrically at 520 nm, sterile medium being used asa blank. EPS that were analyzed with respect to compositionwere obtained during the stationary phase of growth, after 5days of culture.
Polysaccharide isolation. (i) Procedure 1. Culture brothswere diluted when necessary and centrifuged at 4°C and 4,000x g for 15 min in a Sorvall RC-5B centrifuge (Rotor GSA).WhenEPS was firmly bound to the cells as insoluble material,a loosely packed layer of cells covering a compact cell pelletwas observed. Both the layer of cells and the pellet were thenrecovered, and cell dry weight was determined. Cold ethanol (3volumes/1 volume) was added to the supernatant, and thesolution was maintained overnight at 4°C. The precipitatedpolymer was redissolved in a small volume of distilled water,and the precipitation procedure was repeated. The EPS wasthen redissolved in distilled water and dialyzed (molecularweight cutoff, 6,000 to 8,000) against distilled water for 3 daysat 4°C. The polysaccharide preparation was then dried at 40°C,freeze-dried, and stored at 4°C until required.
(ii) Procedure 2. The culture broth was diluted with water toyield a polymer concentration of about 1 g/liter. The solutionwas centrifuged for 2 h at 24,000 x g in order to removeinsoluble material and cell debris, and the supernatant washeated for 5 min at 90°C to denature the proteins and improvethe filtration. Finally, the solution was filtered through Sarto-rius membranes (3- to 0.45-,um pore diameters) and NaCl wasadded to a final concentration of 20 g/liter. To the solution wasadded ethanol (50% [vol/vol]), and the precipitate was washedwith 70 to 100% (vol/vol) ethyl alcohol (EtOH)-H20 mixturesto eliminate excess sodium chloride. The polysaccharide wasdried under vacuum at 30°C for 48 h.Chemical analysis. Protein content was determined by both
the method of Lowry et al. (28) with bovine serum albumin asthe standard (Sigma Chemical Co., St. Louis, Mo.) and by themethod of Bradford (7) by using a protein assay (LaboratoriesBio-Rad S.A., Paris, France). Uronic acid levels were deter-mined by both the carbazole-sulfuric acid reaction (13) and them-phenylphenol method (5), with glucuronic acid as the stan-dard. Hexosamine levels were determined by the Elson-Mor-gan reaction (15) with glucosamine as the standard. Neutralsugar levels were determined by the orcinol-sulfuric acidreaction of Tillmans and Philippi (42), as modified by Riming-ton (39), with 1 mannose molecule-1 galactose molecule as thestandard.
Polysaccharide hydrolysis. A freeze-dried polysaccharidesample (500 jig) was placed in a screw-cap glass tube, 4 Ntrifluoroacetic acid (0.5 ml) was added, and the tube was tightlycapped (Teflon-lined cap) and heated for 4 h at 100°C. Thehydrolyzed sample was then cooled to 25°C, uncapped, anddried under reduced pressure for 24 h. The residue wasresuspended in water, and the pH was adjusted to 9 with 0.1 Mammonia. Reduction was performed with sodium borohydride,and the mixture was left overnight at room temperature.Excess borohydride was removed by addition of acetic acid(pH 4). Borate ions were removed by codistillation withmethanol. The alditol acetates were prepared by adding pyri-
VOL. 60, 1994
on May 23, 2018 by guest
http://aem.asm
.org/D
ownloaded from

4136 VINCENT ET AL.
dine (300 ,ul) and acetic anhydride (300 ,ul) and incubating thesolution overnight at room temperature. The reaction mixtureswere evaporated to dryness, and the alditol acetate derivativeswere dissolved in dichloromethane for gas chromatographicanalysis.
Methanolysis, N acetylation, and trimethylsilylation. Freeze-dried sugar (500 pg) was dissolved in 0.5 ml of 0.5 M HCl inmethanol. After methanolysis for 24 h at 800, the acid solutionwas neutralized by the addition of silver carbonate. Re-Nacetylation was carried out by the addition of 20 pl. of aceticanhydride. This mixture was kept at room temperature for 24h in the dark. The precipitate was centrifuged (3000 x g for 5min), and the supernatant was collected. Lipids were removedwith heptane, and methanol was evaporated under a stream ofnitrogen. Finally the sample was trimethylsilylated with silylat-ing agent (40 p.l of pyridine plus 40 p.1 of BSTFA plus 1%TMCS) for 2 h at room temperature. The sample was driedunder a stream of nitrogen and dissolved in dichloromethanefor analysis.Gas chromatography. Gas chromatography was performed
on a Carlo Erba gas chromatograph equipped with a flameionization detector and an automatic injector "on column."The carrier gas was hydrogen, and the flow rate was 2 mlmin-'. A fused silica capillary column (CP-Sil 5 CB; 25 m;inside diameter, 0.25 mm) was operated with temperaturegradients (50 to 120°C, 20°C/min; and 120 to 250°C, 2°C/min).The initial temperature was maintained for 1 min at 50°C, andthe final temperature was maintained for 5 min at 250°C.
Rheological analysis. The polysaccharide 1545 was purifiedas described above (procedure 2) but without heating. Theaverage molecular weight was determined by using a Chro-matix KMX6 light scattering detector. The measurements werecarried out in 0.1 M NaCl with different polymer concentra-tions (0.05 to 0.4 g liter-'). The solutions were filtered through0.2-p.m-pore-size Sartorius membranes. The dn/dc value wasdetermined on the same solutions at 25°C by using a Brice-Phoenix differential refractometer. The polymer characteriza-tion, e.g., determination of the molecular weight distribution,was carried out by size exclusion chromatography (columns,Shodex OH-Pak 804 and 805) by using a 1545 solution (0.625g/liter) in 0.1 M NH4NO3. Three detectors were used on-line,a low angle light scattering detector (Chromatix CMX100), acapillary viscometer, and a refractometer, as previously de-scribed (43). Viscosity measurements were performed at 25 ±0.01°C by using a Contraves Low Shear 30 viscometer in arange of shear rate from 10-2 to 128 s- . For high viscosities,a Carrimed CS-50 rheometer, equipped with a Rheo 1000 Csystem and 5.0 software which allows direct viscosity shear ratedetermination by angular speed control, was used. For thesemeasurements a cone with a 6-cm diameter and a 10 angle wasused. Polymer solutions were obtained by dissolving the po-lysaccharide in distilled water and adding 1 M NaCl solution inorder to reach the desired NaCl concentration. A potentiomet-ric titration was performed by using a Tacussel (Minisis 6000)potentiometer. The protonated form of the polymer wasobtained by passage of a sodium salt solution through anion-exchange column (Amberlite IR120-H+ form). The titra-tion was carried out at 25 ± 0.1°C by using a 24 mM NaOHsolution.
RESULTS
Strain HYD-1545 appeared as a motile, nonfermentative,gram-negative rod (0.5 by 1.3 to 1.5 p.m) with a single polarflagellum when cultured on 2216E medium for 36 h. Five-day-old colonies on 2216E medium supplemented with glucose
160
140
120
100
80
60
40
20
70
65
60
55
50
45
40
b4)
,00n
10 15 20 25 30 35 40 45 50
a - Temperatue (°C)
5.5 6 6.5
b -pH7S
70
65
60
55
SO
45
40
7 7.5 8
0 10 20 30 40 50 60 70 80
c - NaCI concentration (gA)FIG. 1. Influence of incubation temperature, pH (0, initial pH; El,
final pH), and NaCl concentration on growth of strain HYD-1545. Thedoubling times were calculated from the slopes of the growth curves(not shown).
were viscous, glistening, and irregular shaped, with a lobateedge, and they were about 1.0 to 1.2 cm in diameter.The optimal temperature for growth was between 25 and
29°C, with a doubling time of 38 min at 27°C (Fig. la). Theoptimal pH was between 7.0 and 7.3. For initial pHs of 6.5, 7.0,and 7.3, the final values after 36 h of culture were, respectively,pH 7.2, 7.3, and 7.5 (Fig. lb). The optimal ionic strength was 30g of NaCl per liter (Fig. lc), with a longer doubling time (45
APPL. ENVIRON. MICROBIOL.
c35
on May 23, 2018 by guest
http://aem.asm
.org/D
ownloaded from
EPS-PRODUCING BACTERIUM FROM VENT POLYCHAETE
10
0-%
4)
'0
lo
C*)
UX
4 .-
0
0
2 -
0
100
ooW4)_
O
'0
U)
to
goo_
P4
90
80
70
60
50
40
30
20
10
*. 1-4
250
225
200 j
175 ._00
15 .
125 f0
100
75 _
U
50
25
0
5
4
.3 .=00
...3
2 D0-4
C)
0 5 10 15 20 25 30 50 70
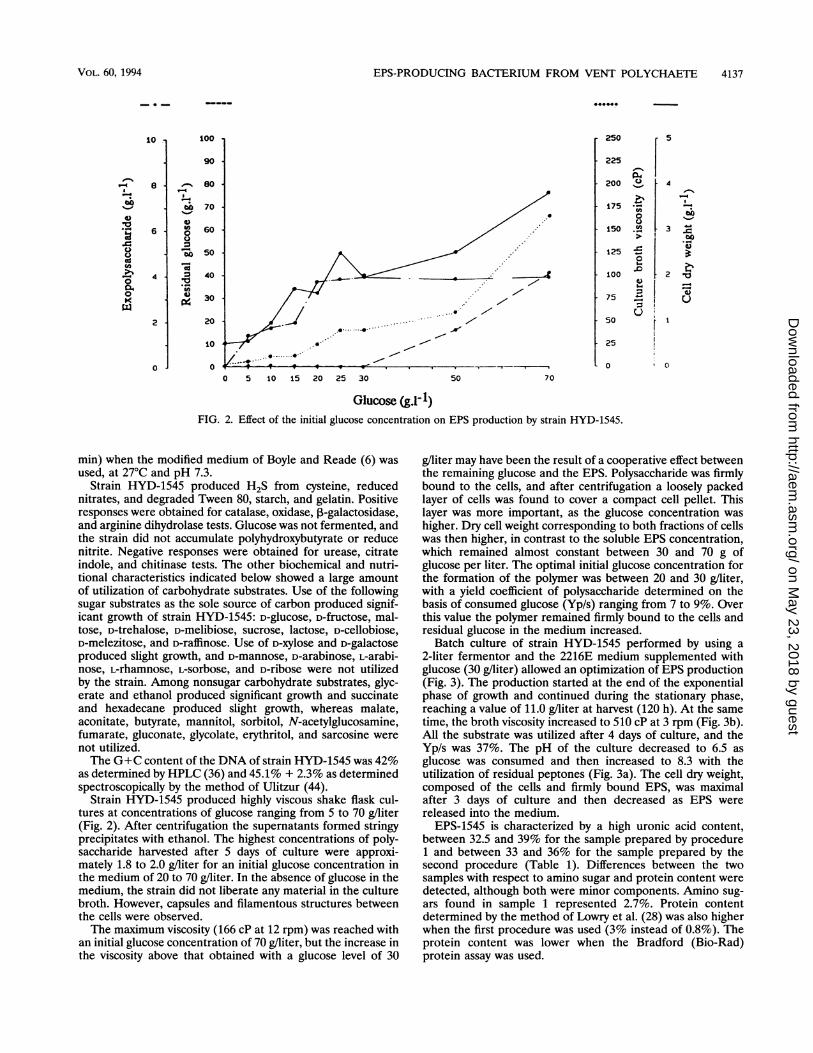
Glucose (g.1-1)FIG. 2. Effect of the initial glucose concentration on EPS production by strain HYD-1545.
min) when the modified medium of Boyle and Reade (6) wasused, at 27°C and pH 7.3.
Strain HYD-1545 produced H2S from cysteine, reducednitrates, and degraded Tween 80, starch, and gelatin. Positiveresponses were obtained for catalase, oxidase, P-galactosidase,and arginine dihydrolase tests. Glucose was not fermented, andthe strain did not accumulate polyhydroxybutyrate or reducenitrite. Negative responses were obtained for urease, citrateindole, and chitinase tests. The other biochemical and nutri-tional characteristics indicated below showed a large amountof utilization of carbohydrate substrates. Use of the followingsugar substrates as the sole source of carbon produced signif-icant growth of strain HYD-1545: D-glucose, D-fructose, mal-tose, D-trehalose, D-melibiose, sucrose, lactose, D-cellobiose,D-melezitose, and D-raffinose. Use of D-xylose and D-galactoseproduced slight growth, and D-mannose, D-arabinose, L-arabi-nose, L-rhamnose, L-sorbose, and D-ribose were not utilizedby the strain. Among nonsugar carbohydrate substrates, glyc-erate and ethanol produced significant growth and succinateand hexadecane produced slight growth, whereas malate,aconitate, butyrate, mannitol, sorbitol, N-acetylglucosamine,fumarate, gluconate, glycolate, erythritol, and sarcosine werenot utilized.The G+C content of the DNA of strain HYD-1545 was 42%
as determined by HPLC (36) and 45.1% + 2.3% as determinedspectroscopically by the method of Ulitzur (44).
Strain HYD-1545 produced highly viscous shake flask cul-tures at concentrations of glucose ranging from 5 to 70 g/liter(Fig. 2). After centrifugation the supernatants formed stringyprecipitates with ethanol. The highest concentrations of poly-saccharide harvested after 5 days of culture were approxi-mately 1.8 to 2.0 g/liter for an initial glucose concentration inthe medium of 20 to 70 g/liter. In the absence of glucose in themedium, the strain did not liberate any material in the culturebroth. However, capsules and filamentous structures betweenthe cells were observed.The maximum viscosity (166 cP at 12 rpm) was reached with
an initial glucose concentration of 70 g/liter, but the increase inthe viscosity above that obtained with a glucose level of 30
g/liter may have been the result of a cooperative effect betweenthe remaining glucose and the EPS. Polysaccharide was firmlybound to the cells, and after centrifugation a loosely packedlayer of cells was found to cover a compact cell pellet. Thislayer was more important, as the glucose concentration washigher. Dry cell weight corresponding to both fractions of cellswas then higher, in contrast to the soluble EPS concentration,which remained almost constant between 30 and 70 g ofglucose per liter. The optimal initial glucose concentration forthe formation of the polymer was between 20 and 30 g/liter,with a yield coefficient of polysaccharide determined on thebasis of consumed glucose (Yp/s) ranging from 7 to 9%. Overthis value the polymer remained firmly bound to the cells andresidual glucose in the medium increased.
Batch culture of strain HYD-1545 performed by using a2-liter fermentor and the 2216E medium supplemented withglucose (30 g/liter) allowed an optimization of EPS production(Fig. 3). The production started at the end of the exponentialphase of growth and continued during the stationary phase,reaching a value of 11.0 g/liter at harvest (120 h). At the sametime, the broth viscosity increased to 510 cP at 3 rpm (Fig. 3b).All the substrate was utilized after 4 days of culture, and theYp/s was 37%. The pH of the culture decreased to 6.5 asglucose was consumed and then increased to 8.3 with theutilization of residual peptones (Fig. 3a). The cell dry weight,composed of the cells and firmly bound EPS, was maximalafter 3 days of culture and then decreased as EPS werereleased into the medium.EPS-1545 is characterized by a high uronic acid content,
between 32.5 and 39% for the sample prepared by procedure1 and between 33 and 36% for the sample prepared by thesecond procedure (Table 1). Differences between the twosamples with respect to amino sugar and protein content weredetected, although both were minor components. Amino sug-ars found in sample 1 represented 2.7%. Protein contentdetermined by the method of Lowry et al. (28) was also higherwhen the first procedure was used (3% instead of 0.8%). Theprotein content was lower when the Bradford (Bio-Rad)protein assay was used.
VOL. 60, 1994 4137
on May 23, 2018 by guest
http://aem.asm
.org/D
ownloaded from

4138 VINCENT ET AL.
a b
_D--Cd
0 20 40 60 80 100 120
Time (hours)
-8
-7v
- 6
20
16-
5 t;o15 tb 12-
0~ ~ ~
_
= ~~~~~~~~~.)
10 cc%
-- 0
10 R 805 o
5 X 4-
OJ
FIG. 3. EPS production by strain HYD-1545 during an aerobic batch culture in a 1.4-liter-working-volume reactor.
The percentage of monomers recovered after hydrolysis or
methanolysis was low, between 20 and 34% (Table 2), probablybecause of the stability of osidic linkage in this acidic polymer.Rhamnose, glucose, galactose, and mannose were found in allthe samples as glucuronic acid, galacturonic acid, and 4,6-0-(1-carboxyethylidene)-galactose after methanolysis (Table 2).However, the values obtained for mannose and rhamnose were
very low. The percentage of recovery for monosaccharides was
lower when the sample was prepared by procedure 1 (20.4 to30.8%), and the highest value (33.9%) was obtained for sample2, after methanolysis. After hydrolysis, molar glucose/galactoseratios were, respectively, 3:5.1 and 3:3 for samples 1 and 2.After methanolysis, molar glucose/galactose/glucuronic acid/galacturonic acid ratios were, respectively, 3:1.2:0.9:0.5 and3:1.7:0.7:0.4 for samples 1 and 2. These molar ratios are onlyindicative, because of the low degree of hydrolysis of thepolymer.
The average molecular mass of the polymer was 1.8 x 106 gmol-', and the dn/dc value was 0.146 ml g-.
Potentiometry. The mass per anionic equivalent found bypH measurements was 542 ± 50 g mol-'. The pKo valueextrapolated for a null net charge, aT = 0 (LT = tN + cxH,
where aN is the degree of neutralization and aH = [H+]I/cP)was 3.2 ± 0.2.
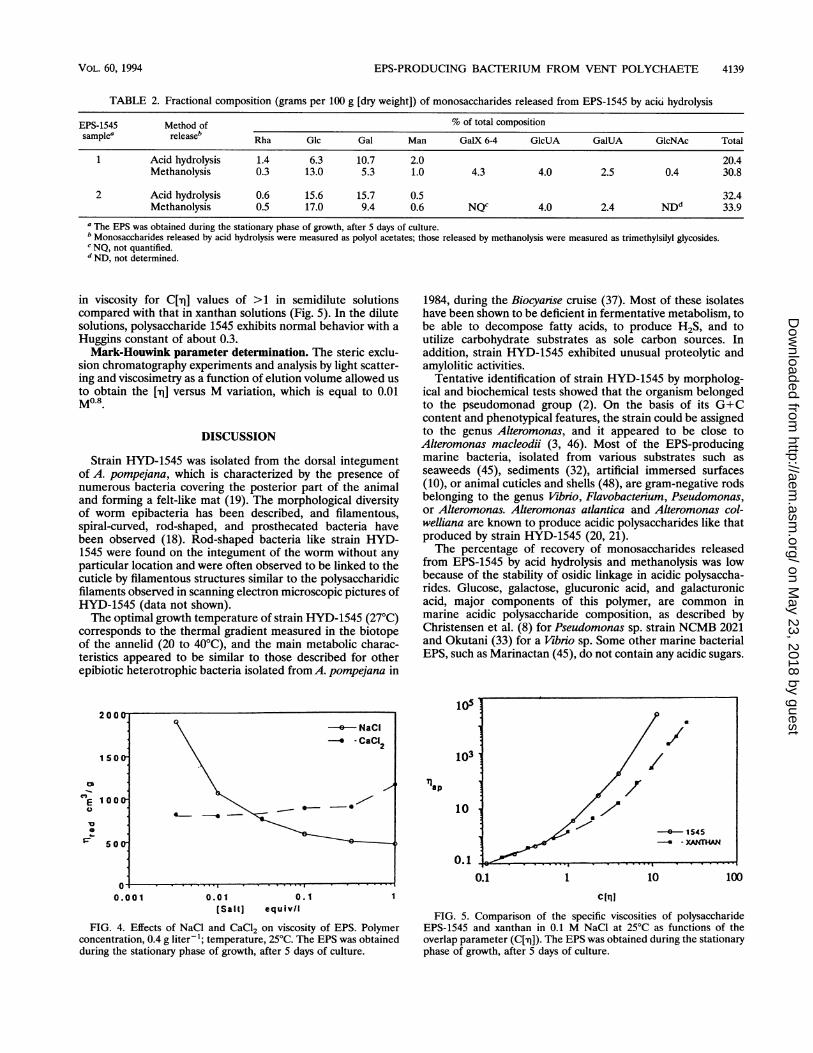
Viscosimetry. Figure 4 shows the influence of ionic strengthon the viscosities of EPS solutions prepared from EPS-1545. Asalt excess screens the electrostatic repulsions between thecharges and leads to a decrease in the rigidity of the polymericchain and consequently in viscosity. However, in the presenceof CaCl2, a stabilization and even an increase in viscosity are
observed. The influence of the polymer concentration on thesolution viscosities was also tested in O.1M NaCl. When -q
(specific viscosity) is plotted against C[-q] (overlap parameters([rj], intrinsic viscosity), the curve shows an abnormal increase
TABLE 1. Fractional analysis (grams per 100 g [dry weight]) of crude EPS-1545
% of total compositionEPS-1545sample' Neutral Hexuronic acids Hexosamines Total Proteins
sugars Carbazol-sulfuric acidb m-Phenylphenol' carbohydrates Lowryd Bradforde
1 51 39 32.5 2.7 927, 86.2 3.0 NDf2* 49 35 36 0.2 84.2, 85.2 0.8 0.22** 51 33 33 0.2 84.2 0.8 0.22*** 55 33 33 0.2 88.2 0.8 0.2
a The polysaccharide was obtained during the stationary phase of growth, after 5 days of culture. Samples 2*, 2**, and 2*** were prepared by procedure 2 with 1Gal molecule, 1 Glc molecule and 1 Gal molecule, and 2 Glc molecules and 1 Gal molecule, respectively, as the reference for neutral sugar determination.
b Determined by the carbazol-sulfuric acid reaction.'Determined by the m-phenylphenol method.d Determined by the method of Lowry et al. (28).e Determined by the method of Bradford (7).f ND, not determined.
2:
1:
00:
C.)C
,0Dg - 1:0
-2J
Co
CL)
0
uo
CZ
rAVL)W.
500
-400 _C)4P1-
300 oC0
200 ,.
100 U
- 0
Time (hours)
APPL. ENVIRON. MICROBIOL.
O
on May 23, 2018 by guest
http://aem.asm
.org/D
ownloaded from
EPS-PRODUCING BACTERIUM FROM VENT POLYCHAETE 4139
TABLE 2. Fractional composition (grams per 100 g [dry weight]) of monosaccharides released from EPS-1545 by acici hydrolysis
EPS-1545 Method of % of total compositionsamplea releaseb Rha Glc Gal Man GalX 6-4 GlcUA GalUA GlcNAc Total
1 Acid hydrolysis 1.4 6.3 10.7 2.0 20.4Methanolysis 0.3 13.0 5.3 1.0 4.3 4.0 2.5 0.4 30.8
2 Acid hydrolysis 0.6 15.6 15.7 0.5 32.4Methanolysis 0.5 17.0 9.4 0.6 NQc 4.0 2.4 NDd 33.9
a The EPS was obtained during the stationary phase of growth, after 5 days of culture.b Monosaccharides released by acid hydrolysis were measured as polyol acetates; those released by methanolysis were measured as trimethylsilyl glycosides.c NQ, not quantified.d ND, not determined.
in viscosity for C['r] values of >1 in semidilute solutionscompared with that in xanthan solutions (Fig. 5). In the dilutesolutions, polysaccharide 1545 exhibits normal behavior with a
Huggins constant of about 0.3.Mark-Houwink parameter determination. The steric exclu-
sion chromatography experiments and analysis by light scatter-ing and viscosimetry as a function of elution volume allowed us
to obtain the [-q] versus M variation, which is equal to 0.01WO8.
DISCUSSION
Strain HYD-1545 was isolated from the dorsal integumentof A. pompejana, which is characterized by the presence ofnumerous bacteria covering the posterior part of the animaland forming a felt-like mat (19). The morphological diversityof worm epibacteria has been described, and filamentous,spiral-curved, rod-shaped, and prosthecated bacteria havebeen observed (18). Rod-shaped bacteria like strain HYD-1545 were found on the integument of the worm without anyparticular location and were often observed to be linked to thecuticle by filamentous structures similar to the polysaccharidicfilaments observed in scanning electron microscopic pictures ofHYD-1545 (data not shown).The optimal growth temperature of strain HYD-1545 (27°C)
corresponds to the thermal gradient measured in the biotopeof the annelid (20 to 40°C), and the main metabolic charac-teristics appeared to be similar to those described for otherepibiotic heterotrophic bacteria isolated from A. pompejana in
2000
-0 NaCI\ ~~~~~- -Cad2
100l _ X >.
a~~~~~~~~~~~~~~~~cc
F 500
0
0.001 0.01 0.1 1
[Salt) equiv/1
FIG. 4. Effects of NaCl and CaCl2 on viscosity of EPS. Polymerconcentration, 0.4 g liter-'; temperature, 25°C. The EPS was obtainedduring the stationary phase of growth, after 5 days of culture.
1984, during the Biocyarise cruise (37). Most of these isolateshave been shown to be deficient in fermentative metabolism, tobe able to decompose fatty acids, to produce H2S, and toutilize carbohydrate substrates as sole carbon sources. Inaddition, strain HYD-1545 exhibited unusual proteolytic andamylolitic activities.
Tentative identification of strain HYD-1545 by morpholog-ical and biochemical tests showed that the organism belongedto the pseudomonad group (2). On the basis of its G+Ccontent and phenotypical features, the strain could be assignedto the genus Alteromonas, and it appeared to be close toAlteromonas macleodii (3, 46). Most of the EPS-producingmarine bacteria, isolated from various substrates such asseaweeds (45), sediments (32), artificial immersed surfaces(10), or animal cuticles and shells (48), are gram-negative rodsbelonging to the genus Vibrio, Flavobacterium, Pseudomonas,or Alteromonas. Alteromonas atlantica and Alteromonas col-welliana are known to produce acidic polysaccharides like thatproduced by strain HYD-1545 (20, 21).The percentage of recovery of monosaccharides released
from EPS-1545 by acid hydrolysis and methanolysis was lowbecause of the stability of osidic linkage in acidic polysaccha-rides. Glucose, galactose, glucuronic acid, and galacturonicacid, major components of this polymer, are common inmarine acidic polysaccharide composition, as described byChristensen et al. (8) for Pseudomonas sp. strain NCMB 2021and Okutani (33) for a Vibrio sp. Some other marine bacterialEPS, such as Marinactan (45), do not contain any acidic sugars.
/y103
Xi ~ ~~~~1545- XANTHW
0.10.1 1 10 100
c[rIFIG. 5. Comparison of the specific viscosities of polysaccharide
EPS-1545 and xanthan in 0.1 M NaCl at 25°C as functions of theoverlap parameter (C[-q]). The EPS was obtained during the stationaryphase of growth, after 5 days of culture.
VOL. 60, 1994
on May 23, 2018 by guest
http://aem.asm
.org/D
ownloaded from

4140 VINCENT ET AL.
Mannose and rhamnose were also found in marine EPSexcreted by gram-negative rods such as Vibrio spp. and Pseudo-monas spp., but these small amounts could be attributed tocontamination by components of the culture medium, such asmannose, or cell wall constituents, such as rhamnose (14).Procedure 2 (see Materials and Methods) for isolation of thepolysaccharide provides preparations with the lower proteincontent.The EPS-1545 prepared by procedure 1 was purified by
Talmont et al. (41) by using a DEAE Trisacryl ion-exchangecolumn and a Sepharose 4B gel filtration column. The majorfraction represented 60% of the crude extract; was made ofneutral sugars (50%), uronic acids (40%), and 4,6-O-(1-car-boxyethylidene)-galactose (GalX; X as pyruvate) (10%); andwas composed of galactose, glucose, glucuronic acid, galactu-ronic acid, and GalX in a molar ratio of 2.5:3:2:2:1. Pyruvatewas found in some marine EPS produced by gram-negativerods, including Pseudomonas atlantica, renamed A. atlantica(6).The presence of acidic sugars in the EPS may be important,
considering the heavy-metal-binding properties of this poly-mer. A. pompejana lives in large colonies on black and whiteactive smokers and diffusers at 13°N hydrothermal vents. Theanimal is therefore exposed to high concentrations of chemi-cals (e.g., metallic sulfides), and the polysaccharide coatings ofsuch microorganisms may interact with heavy metals occurringin the environment of the worm. Numerous rod-shaped bac-teria which, on metal-amended media, produced EPS wereisolated from alvinellids (47). During this screening, about30% of the isolated strains showed mucoid phenotypes on2216E plates and 9% produced viscous broth as they werecultured on 2216E medium supplemented with glucose. Thesusceptibility of those heterotrophic bacteria to heavy metalshas been studied by Jeanthon and Prieur (24), who showed anadaptation of alvinellid-associated microflora to this metal-richenvironment. Numerous strains (92.3% of the isolates) dis-played resistance to cadmium, zinc, arsenate, and/or silver andtolerated large amounts of copper. However, no clear relation-ship between EPS production and heavy metal resistance wasfound.Many data about the composition of EPS produced by
marine bacteria are available, but little is known about theamounts produced by these organisms. The production andrelease of EPS during starvation has been studied by Wrang-stadh et al. (49). In an attempt to increase the production,strain HYD-1545 was however cultured in a rich nutrientmedium. Compared with those of other EPS-producing marinebacteria, the production rate of EPS by strain HYD-1545following harvest of 11 g/liter during the 2-liter fermentorexperiment, with a yield coefficient (Yp/s) of 37%, seems to beparticularly significant.The value of the mass per anionic equivalent was in agree-
ment with the presence of one uronic acid for every two neutralsugars in the polymer structure. The pKo value found is typicalfor carboxyl groups (40), excluding the presence of somestrong acidic groups like sulfates or sulfonates on the polymer.The ionic strength had an influence on the viscosities of
EPS-1545 solutions, and this behavior is common for polyelec-trolyte solutions (17). The stabilization and the increase ob-served in the presence of CaCl2 may be due to intermolecularbonds or to a polymer conformational change (29). However,it is not certain that all divalent ions were removed duringpurification of the polysaccharide, and ions may have beenresponsible for the observed aggregation. Nevertheless, nogelation was produced either by adding an excess of divalentions or by using acidic medium. The 1545 polymer did not
exhibit any conformational change when monovalent saltconcentrations and temperatures were varied. The value foundfor the Mark-Houwink exponent permits classification of the1545 polysaccharide among the semirigid polymers.The polysaccharide HYD-1545 is a polyelectrolyte, and the
viscosity of its solutions depends strongly on their ionicstrength. However, an unusual behavior is observed at higherconcentrations, and this behavior could be related to intermo-lecular interactions in semidilute solutions. In order to betterunderstand these physicochemical properties, more completestudies should be carried out.
REFERENCES1. Allison, D. G., and I. W. Sutherland. 1984. A staining technique
for attached bacteria and its correlation to extracellular carbohy-drate production. J. Microbiol. Methods 2:93-99.
2. Baumann, P., and L. Baumann. 1981. The marine gram-negativeeubacteria: genera Photobacterium, Beneckea, Alteromonas, Pseu-domonas, and Alcaligenes, p. 1302-1331. In M. P. Starr, H. Stolp,H. G. Truper, A. Balows, and H. G. Schlegel (ed.), The pro-karyotes. Springer-Verlag KG, Berlin.
3. Baumann, P., M. J. Gauthier, and L. Baumann. 1984. GenusAlteromonas, p. 342-352. In N. R. Krieg, and J. G. Holt (ed.),Bergey's manual of systematic bacteriology, vol. 1. The Williamsand Wilkins Co., Baltimore.
4. Beji, A., D. Izard, F. Gavini, H. Leclerc, M. Leseine-Delstanche,and J. Krembel. 1987. A rapid chemical procedure for isolationand purification of chromosomal DNA from gram-negative bacilli.Anal. Biochem. 162:18-23.
5. Blumenkrantz, N., and G. Asboe-Hansen. 1973. New method forquantitative determination of uronic acids. Anal. Biochem. 54:484-489.
6. Boyle, C. D., and A. E. Reade. 1983. Characterization of twoextracellular polysaccharides from marine bacteria. Appl. Environ.Microbiol. 46:392-399.
7. Bradford, M. M. 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 72:248-254.
8. Christensen, B. E., J. Kjosbakken, and 0. Smidsrod. 1985. Partialchemical and physical characterization of two extracellular poly-saccharides produced by marine, periphytic Pseudomonas sp.strain NCMB 2021. Appl. Environ. Microbiol. 50:837-845.
9. Conn, H. J., J. W. Bartholomew, and M. W. Jennison. 1957.Staining methods, p. 10-36. In Society of American Bacteriologists(ed.), Manual of microbiological methods. McGraw-Hill BookCompany, Inc., New York.
10. Corpe, W. A. 1970. Attachment of marine bacteria to solidsurfaces, p. 73-87. In Adhesion in biological systems. AcademicPress, Inc., New York.
11. Desbruyeres, D., F. Gaill, L. Laubier, and Y. Fouquet. 1986.Polychaetous annelids from hydrothermal vent ecosystems: anecological overview. In M. I. Jones (ed.), The hydrothermal ventsof the eastern Pacific. Bull. Biol. Soc. Wash. 6:103-116.
12. Desbruyires, D., and L. Laubier. 1980. Alvinella pompejana gen.sp. nov., Ampharatidae aberrant des sources hydrothermales de laride Est-Pacifique. Oceanol. Acta 3:267-274.
13. Dische, Z. 1947. A new specific color reaction of hexuronic acids.J. Biol. Chem. 167:189-198.
14. Eagon, R. G. 1956. Studies on polysaccharide formation by Pseudo-monas fluorescens. Can. J. Microbiol. 2:673-676.
15. Elson, L. A., and W. T. J. Morgan. 1933. Colorimetric method forthe determination of glucosamine and chondrosamine. Biochem.J. 27:1824-1828.
16. Fletcher, M., and G. D. Floodgate. 1973. An electron-microscopicdemonstration of acidic polysaccharide involved in the adhesion ofa marine bacterium to solid surfaces. J. Gen. Microbiol. 74:325-334.
17. Fouissac, E., M. Milas, M. Rinaudo, and R. Borsali. 1992.Influence of the ionic strength on the dimensions of sodiumhyaluronate. Macromolecules 25:5613-5617.
18. Gaill, F., D. Desbruyeres, and D. Prieur. 1987. Bacterial commu-nities associated with "Pompei Worms" from the East Pacific Rise
APPL. ENVIRON. MICROBIOL.
on May 23, 2018 by guest
http://aem.asm
.org/D
ownloaded from

EPS-PRODUCING BACTERIUM FROM VENT POLYCHAETE 4141
hydrothermal vents: SEM. TEM observations. Microb. Ecol. 13:129-139.
19. Gaill, F., D. Desbruyeres, D. Prieur, and J.-P. Gourret. 1984. Miseen evidence de communautes bacteriennes epibiontes du "Ver dePompei" (Alvinella pompejana). C. R. Acad. Sci. 298:553-558.
20. Gauthier, M. J., and V. M. Breittmayer. 1990. Genera Alteromonasand Marinomonas. In A. Balows, H. G. Truper, M. Dworkin, W.Harder, and K. H. Schleifer (ed.), The prokaryotes, 2nd ed.Springer-Verlag, New York.
21. Geesey, G. G., P. J. Bremer, J. J. Smith, M. Muegger, and L. K.Jang. 1992. Two-phase model for describing the interactionsbetween copper ions and exopolymers from Alteromonas atlantica.Can. J. Microbiol. 38:785-793.
22. Harrigan, W. F., and M. E. McCance. 1966. Laboratory methodsin microbiology. Academic Press, London.
23. Jannasch, H. W., and C. 0. Wirsen. 1979. Chemosynthetic primaryproduction at East Pacific Rise sea floor spreading centers.BioScience 29:592-598.
24. Jeanthon, C., and D. Prieur. 1990. Susceptibility to heavy metalsand characterization of heterotrophic bacteria isolated from twohydrothermal vent polychaete annelids, Alvinella pompejana andAlvinella caudata. Appl. Environ. Microbiol. 56:3308-3314.
25. Kohn, J. 1953. A preliminary report of a new gelatin liquefactionmethod. J. Clin. Pathol. 6:249.
26. Kovacs, N. 1956. Identification of Pseudomonas pyocyanae by theoxydase reaction. Nature (London) 178:703.
27. Lanyi, B. 1987. Classical and rapid identification methods formedically important bacteria, p. 1-67. In R. R. Colwell and R.Grigorova (ed.), Methods in microbiology, vol. 19. AcademicPress, Inc., New York.
28. Lowry, 0. H., N. J. Rosenbrough, A. L. Farr, and R. J. Randall.1951. Protein measurement with the Folin phenol reagent. J. Biol.Chem. 193:265-275.
29. Milas, M., and M. Rinaudo. 1986. Properties of xanthan gum inaqueous solutions: role of the conformational transition. Carbo-hydr. Res. 158:191-204.
30. Mittelman, M. W., and G. G. Geesey. 1985. Copper-bindingcharacteristics of exopolymers from a freshwater sediment bacte-rium. Appl. Environ. Microbiol. 49:846-851.
31. Okutani, K. 1982. Structural investigation of the fructan frommarine bacterium NAM-1. Bull. Jpn. Soc. Sci. Fish. 48:1621-1625.
32. Okutani, K. 1984. Antitumor and immunostimulant activities ofpolysaccharide produced by a marine bacterium of the genusVibrio. Bull. Jpn. Soc. Sci. Fish. 50:1035-1037.
33. Okutani, K. 1985. Isolation and fractionation of an extracellularpolysaccharide from marine Vibrio. Bull. Jpn. Soc. Sci. Fish.51:493-496.
34. Oppenheimer, C. E., and C. E. Zobell. 1952. The growth andviability of sixty-three species of marine bacteria as influenced byhydrostatic pressure. J. Mar. Res. 11:10-18.
35. Pearl, H. W. 1975. Microbial attachment to particles in marine and
freshwater ecosystems. Microb. Ecol. 2:73-83.36. Peyret, M., J. Freney, H. Meugnier, and J. Fleurette. 1989.
Determination of G+C content of DNA using high-liquid perfor-mance chromatography for the identification of staphylococci andmicrococci. Res. Microbiol. 140:467-475.
37. Prieur, D., and C. Jeanthon. 1987. Preliminary study of heterotro-phic bacteria isolated from two deep-sea hydrothermal ventinvertebrates: Alvinella pompejana (polychaete) and Bathymodio-lus thermophilus (bivalve). Symbiosis 4:87-98.
38. Reichenbach, H., and M. Dworkin. 1981. The order Cytophagales,p. 356. In M. P. Starr, H. Stolp, H. G. Truper, A. Balows, and H. G.Schlegel (ed.), The prokaryotes, Vol. I. Springer-Verlag, NewYork.
39. Rimington, C. 1931. The carbohydrate complex of serum protein.II. Improved method for isolation and redetermination of struc-ture. Isolation of glucoaminodimannose from protein of ox blood.Biochem. J. 25:1062-1071.
40. Rinaudo, M., and M. Milas. 1974. Interaction of monovalent anddivalent counterions with some carboxylic polysaccharides. J.Polym. Sci. 12:2073-2081.
41. Talmont, F., P. Vincent, T. Fontaine, J. Guezennec, D. Prieur, andB. Fournet. 1991. Structural investigation of an acidic exopolysac-charide from a deep-sea hydrothermal vent marine bacteria. FoodHydrocolloids 5:171-172.
42. Tillmans, J., and K. Philippi. 1929. Uber den Gehalt der wichtig-sten Protein der Nahrungsmittel an Kohlehydrat and uber einkolorimetrisches Verfahren zur quantitativen Bestimmung vonstockstoffreiem Zucker in Elweiss. Biochem. Z. 215:36-60.
43. Tinland, B., J. Mazet, and M. Rinaudo. 1988. Characterization ofwater-soluble polymers by multidetection size-exclusion chroma-tography. Makromol. Chem. 9:69-73.
44. Ulitzur, S. 1972. Rapid determination of DNA base compositionby ultraviolet spectroscopy. Biochim. Biophys. Acta 272:1-11.
45. Umezawa, H. Y., Y. Okami, S. Kurasawa, T. Ohnuki, M. Ishizuka,T. Takeushi, T. Shiio, and Y. Yugari. 1983. Marinactan, antitumorpolysaccharide produced by marine bacteria. J. Antibiot. 5:471-477.
46. Vincent, P. 1993. Etude d'eubact6ries productrices d'exopoly-saccharides, originaires d'un site hydrothermal profond (13°N).Ph.D. thesis. Universite de Bretagne Occidentale, Brest, France.
47. Vincent, P., C. Jeanthon, and D. Prieur. 1991. Production ofexopolysaccharides by bacteria from deep-sea hydrothermal vents.Kiel. Meeresforsch. 8:188-192.
48. Weiner, R. M., R. R. Colwell, R. N. Jarman, D. C. Stein, C. C.Somerville, and D. B. Bonar. 1985. Applications of biotechnologyto the production, recovery and use of marine polysaccharides.Bio/Technology 3:899-902.
49. Wrangstadh, M., P. L. Conway, and S. Kjelleberg. 1986. Theproduction and release of an extracellular polysaccharide duringstarvation of a marine Pseudomonas sp. and the effect thereof onadhesion. Arch. Microbiol. 145:220-227.
VOL. 60, 1994
on May 23, 2018 by guest
http://aem.asm
.org/D
ownloaded from


















