
Prevalence of a vertically transmitted single-stranded DNA ...
35
University of South Florida University of South Florida Scholar Commons Scholar Commons Marine Science Faculty Publications College of Marine Science 2019 Prevalence of a vertically transmitted single-stranded DNA virus in Prevalence of a vertically transmitted single-stranded DNA virus in spinybacked orbweavers ( spinybacked orbweavers (Gasteracantha cancriformis) from ) from Florida, USA Florida, USA Karyna Rosario University of South Florida, [email protected] Kaitlin A. Mettel University of South Florida Anthony M. Greco University of South Florida, [email protected] Mya Breitbart University of South Florida, [email protected] Follow this and additional works at: https://scholarcommons.usf.edu/msc_facpub Part of the Life Sciences Commons Scholar Commons Citation Scholar Commons Citation Rosario, Karyna; Mettel, Kaitlin A.; Greco, Anthony M.; and Breitbart, Mya, "Prevalence of a vertically transmitted single-stranded DNA virus in spinybacked orbweavers (Gasteracantha cancriformis) from Florida, USA" (2019). Marine Science Faculty Publications. 433. https://scholarcommons.usf.edu/msc_facpub/433 This Article is brought to you for free and open access by the College of Marine Science at Scholar Commons. It has been accepted for inclusion in Marine Science Faculty Publications by an authorized administrator of Scholar Commons. For more information, please contact [email protected].
Transcript of Prevalence of a vertically transmitted single-stranded DNA ...

University of South Florida University of South Florida
Scholar Commons Scholar Commons
Marine Science Faculty Publications College of Marine Science
2019
Prevalence of a vertically transmitted single-stranded DNA virus in Prevalence of a vertically transmitted single-stranded DNA virus in
spinybacked orbweavers (spinybacked orbweavers (Gasteracantha cancriformis) from ) from
Florida, USA Florida, USA
Karyna Rosario University of South Florida, [email protected]
Kaitlin A. Mettel University of South Florida
Anthony M. Greco University of South Florida, [email protected]
Mya Breitbart University of South Florida, [email protected]
Follow this and additional works at: https://scholarcommons.usf.edu/msc_facpub
Part of the Life Sciences Commons
Scholar Commons Citation Scholar Commons Citation Rosario, Karyna; Mettel, Kaitlin A.; Greco, Anthony M.; and Breitbart, Mya, "Prevalence of a vertically transmitted single-stranded DNA virus in spinybacked orbweavers (Gasteracantha cancriformis) from Florida, USA" (2019). Marine Science Faculty Publications. 433. https://scholarcommons.usf.edu/msc_facpub/433
This Article is brought to you for free and open access by the College of Marine Science at Scholar Commons. It has been accepted for inclusion in Marine Science Faculty Publications by an authorized administrator of Scholar Commons. For more information, please contact [email protected].

Prevalence of a vertically transmitted single-stranded DNA virus in spinybacked orbweavers 1
(Gasteracantha cancriformis) from Florida, USA 2
Karyna Rosario1*, Kaitlin A. Mettel1, Anthony M. Greco1, and Mya Breitbart1 3
4
1College of Marine Science, University of South Florida, 140 7th Avenue South, Saint Petersburg, 5
Florida, 33701, USA 6
7
8
9
*Corresponding author: Karyna Rosario, [email protected] 10
11
Keywords: CRESS DNA, spider, ssDNA, virus, vertical transmission, Araneae 12

Abstract 13
Spiders (Order Araneae, Class Arachnida) are an important group of predatory arthropods in 14
terrestrial ecosystems that have been recently identified as an untapped reservoir of single-stranded 15
(ss)DNA viruses. Specifically, spiders harbor a diversity of ssDNA viruses encoding a replication-16
associated protein (Rep) within a circular genome. However, little is known about the ecology of 17
novel circular Rep-encoding ssDNA (CRESS DNA) viruses. Here we investigated two CRESS 18
DNA viruses recently identified in spinybacked orbweavers (Gasteracantha cancriformis), 19
namely spinybacked orbweaver circular virus (SpOrbCV) 1 and 2. SpOrbCV-1 was detected in 20
the majority (>65%) of spider specimens from all life stages, including eggs, spiderlings, and 21
adults, demonstrating that this virus is active within spinybacked orbweavers. In contrast, 22
SpOrbCV-2 was only detected in adults at a lower (36%) prevalence. Since we also detected 23
SpOrbCV-2 in other spider species and this virus has been reported from a dragonfly, we suggest 24
that SpOrbCV-2 is accumulated in these predators through common insect prey. The prevalence 25
of SpOrbCV-1 in collected specimens allowed us to design assays to characterize this virus, which 26
represents a new group of CRESS DNA viruses, the “circularisviruses”. To our knowledge, 27
SpOrbCV-1 is the first example of a vertically transmitted virus in spiders, which may explain its 28
high prevalence in spinybacked orbweavers. Since vertically transmitted viruses infecting insects 29
(Class Insecta) can manipulate their host behavior and physiology, future studies should 30
investigate the ecological role of vertically transmitted viruses in spiders. 31

Introduction 32
Viruses encoding a replication-associated protein (Rep) within a covalently-closed, single-33
stranded (ss)DNA genome have been increasingly identified in organisms spanning the eukaryotic 34
tree of life, from unicellular organisms to major groups of vertebrate and invertebrate metazoans 35
[1]. Current classification schemes for eukaryotic circular Rep-encoding ssDNA (CRESS DNA) 36
viruses, which include six monophyletic families (Geminiviridae, Nanoviridae, Circoviridae, 37
Genomoviridae, Smacoviridae, Bacilladnaviridae) [40], lag behind the discovery rate of these 38
viruses. Many CRESS DNA viral species cannot be accommodated within established taxa and 39
remain unclassified, indicating that the number of CRESS DNA viral families will increase. Given 40
that many CRESS DNA viruses have only been identified through sequencing, little is known 41
about the ecology of a large number of CRESS DNA viruses, including the host species for both 42
classified (e.g., Genomoviridae and Smacoviridae) and unclassified viruses. Terrestrial arthropods 43
have been recently identified as an untapped reservoir of CRESS DNA viral diversity, highlighting 44
the need to further investigate viral prevalence and transmission in this dominant and 45
undersampled group of animals [2]. 46
Spiders (Order Araneae) are an important group of predatory arthropods [3] that have been 47
extensively studied due to their fascinating biology (e.g., [4-6]), potential as natural pest biocontrol 48
agents (e.g., [7, 8]), and the biotechnological potential of their silk and venom [9-11]. Previous 49
studies have investigated the influence of microbes, in particular bacteria, on spider biology [12-50
15]. Notably, studies investigating vertically transmitted bacteria in spiders indicate that these 51
‘reproductive parasites’ may lead to sex-ratio bias through behavior manipulation and male-killing 52
[16-18]. Although spiders harbor a diversity of viruses, there is scarce information regarding the 53
prevalence and ecological impact of viral infection in spiders. Endogenous viral sequences within 54
spider chromosomes indicate that these arachnids have been infected by both CRESS DNA viruses 55
and double-stranded DNA polyomaviruses [19, 20]. Studies within the past four years have also 56
revealed a diversity of novel extant viruses in multiple spider species, including more than 20 57
CRESS DNA [2] and 100 RNA viruses [21-24] encompassing various viral families and 58
unclassified species. Moreover, RNA viruses have been detected in multiple spider organs, 59
including brain, venom glands and silk glands, indicating that viral infection may play a role in 60
spider ecology [21]. 61

The present study investigated two CRESS DNA viruses, namely spinybacked orbweaver 62
circular virus (SpOrbCV) 1 and 2, that were reported from specimens of Gasteracantha 63
cancriformis, commonly known as the spinybacked orbweaver [2]. Spinybacked orbwearvers are 64
generally considered beneficial spiders since they trap and consume insects considered pests, such 65
as whiteflies, and are typically found in wooded areas where there is abundant foliage and branches 66
[25]. However, spinybacked orbweavers are also commonly found in man-made structures, such 67
as porches and screen pool enclosures, where they may become a nuisance. These conspicuous 68
spiders, which can be easily spotted due to their contrasting colors and spine-like projections 69
around their abdomen, are widely distributed throughout the Caribbean, Central America, and 70
Southern USA [25, 26]. To our knowledge, SpOrbCV-1 and SpOrbCV-2 are the first viruses that 71
have been identified in this tropical spider species. 72
Here we examined the prevalence of SpOrbCV-1 and -2 in different spider life stages, 73
including eggs, spiderlings, and adults, to explore the association between spinybacked 74
orbweavers and these CRESS DNA viruses. Similar to known CRESS DNA viruses [1], both 75
SpOrbCVs have a small genome (< 2 kb) encoding the Rep and a putative capsid protein. 76
SpOrbCV-1 is an unclassified CRESS DNA virus that has persisted in the Tampa Bay, Florida, 77
USA spinybacked orbweaver population for at least four years (2014 through 2018). We detected 78
SpOrbCV-1 in the majority (>65%) of specimens from all life stages, demonstrating that this virus 79
is active within spinybacked orbweavers. In contrast, the detection of SpOrbCV-2 in adult 80
specimens alone, combined with its presence in various species of spiders (this study) and a 81
dragonfly [27], indicates that this cyclovirus (genus Cyclovirus, family Circoviridae) may 82
accumulate in predatory arthropods through common insect prey. The prevalence of SpOrbCV-1 83
in collected specimens allowed us to design assays to characterize this virus, which represents a 84
new cohesive group of CRESS DNA viruses, referred to here as circularisviruses. Finally, the 85
detection of SpOrbCV-1 in eggs represents the first evidence for vertical transmission of viruses 86
in spiders, which may explain its high prevalence in spinybacked orbweavers. 87

Materials and Methods 88
Analysis of viral sequences related to SpOrbCV-1 89 90
SpOrbCV-1 (GenBank accession number MH545519) and SpOrbCV-2 (GenBank 91
accession number MH545515) were recently described as a part of a CRESS DNA viral discovery 92
effort in terrestrial arthropods [2]. Since SpOrbCV-1 cannot be accommodated within established 93
CRESS DNA viral families, we further evaluated the characteristics of this virus and other 94
unclassified CRESS DNA viruses. CRESS DNA virus genomes with top BLASTn matches, 95
including reciprocal searches, to the SpOrbCV-1 genome were obtained from GenBank in 96
December 2018. SeqBuilder from the Lasergene software package version 11.2.1 (DNASTAR, 97
Madison, WI) was used to explore genomic features in the identified genomes, including: size, 98
open reading frames (ORFs), genome organization, and putative origin of replication (ori), which 99
is marked by a conserved nonanucleotide motif at the apex of a predicted stem-loop structure [28]. 100
Genome-wide and predicted amino acid sequence pairwise identities were calculated using the 101
Sequence Demarcation Tool version 1.2 [29]. Predicted Rep amino acid sequences from each 102
genome were aligned using the MUSCLE algorithm as implemented in MEGA7 [30, 31]. 103
Alignments were then used to create a graphical representation of conserved residues found within 104
CRESS DNA viral Rep domains, including endonuclease and superfamily 3 (SF3) helicase motifs, 105
in WebLogo 3 [32]. 106
107 Collection of spinybacked orbweavers to investigate SpOrbCVs 108 109
To further investigate SpOrbCVs, spinybacked orbweaver samples were collected from 110
vegetation and man-made structures found outdoors as well as screen pool enclosures in the Tampa 111
Bay area in Florida, USA in 2017 and 2018. The majority of samples, including adults (66%, n = 112
154), eggs (89%, n = 35) and spiderlings (100%, n = 14), were collected from a single screen pool 113
enclosure that was closely monitored. Based on size and the presence of abdominal spines [26], 114
99% of sampled adults were female. All eggs were collected by carefully scraping and collecting 115
complete egg sacs from surfaces without damaging the eggs. Spiderling collections only took place 116
in 2018. To do this, egg sacs were monitored until spiderlings emerged (~2 - 5 weeks) and 117
spiderlings were then collected before they dispersed from the silk surrounding the egg sac. All 118
specimens were collected using sterile 15 or 50 ml conical tubes and kept at room temperature 119
until taken to the lab within one hour of collection. Once in the lab, samples from adults were 120

either immediately stored as whole specimens or two legs were dissected off individual specimens 121
using a sterile scalpel and stored separately. Whole and dissected specimens as well as leg 122
subsamples were stored at -80°C until further processing. 123
124 PCR assays to evaluate the prevalence of SpOrbCVs 125 126
The prevalence of SpOrbCV-1 and SpOrbCV-2 was investigated in specimens representing 127
different spinybacked orbweaver life stages through PCR. The prevalence in adults was initially 128
assessed based on positive PCR products from virions isolated from whole specimens following a 129
method previously described for discovery of CRESS DNA viruses in arthropods (Rosario et al 130
2012, Rosario et al 2018). Briefly, spiders were rinsed with sterile suspension medium (SM) buffer 131
[0.1 M NaCl, 50 mM Tris/HCl (pH 7.4), 10 mM MgSO4] to remove debris, followed by 132
homogenization in 1 ml SM buffer through bead beating in a FisherbrandTM Bead Mill 4 133
homogenizer (Fisher Scientific, Hampton, NH) for one minute using 1 mm glass beads. 134
Homogenates were then centrifuged at 6,000 x g for 6 min and the supernatants containing viral 135
particles were filtered through a 0.45-μm Sterivex filter (MilliporeSigma, Burlington, MA) to 136
remove cells. DNA was extracted from 200 μl of the filtrate containing partially purified virus 137
particles using the QIAmp MinElute Virus Spin kit (Qiagen, Hilden, Germany). However, since 138
virus particles isolated from whole specimens may include viruses from dietary content, viral 139
prevalence was further assessed in adult spiders by testing legs. For this purpose, DNA was directly 140
extracted from two legs per specimen using the Extract-N-Amp Tissue PCR Kit (MilliporeSigma, 141
St. Louis, MO) following the manufacturer’s protocol. 142
Viral prevalence in eggs and spiderlings was assessed on a per egg sac basis rather than 143
individual eggs or spiderlings. For this purpose, the silk surrounding the eggs was removed using 144
sterile pipette tips and scapels and about a third of the eggs or two spiderlings from a given sac 145
were transferred to a 1.5 ml microcentrifuge tube. After rinsing the eggs and spiderlings with SM 146
buffer, DNA was directly extracted from tissues using the Extract-N-Amp Tissue PCR Kit 147
(MilliporeSigma, St. Louis, MO). 148
To assess the prevalence of each virus, primer pairs SpOrbCV1_F2/R2 and 149
SpOrbCV2_F2/R2 (Table 1) were used for PCR assays targeting partial SpOrbCV-1 and 150
SpOrbCV-2 genomes, respectively. For this purpose, 1 µl of DNA extract from adults, eggs, and 151
spiderlings was amplified through rolling circle amplification (RCA) to enrich for small circular 152

templates [33], such as CRESS DNA genomes, using the Illustra TempliPhi Amplification kit (GE 153
Healthcare, Chicago, IL). Four microliters of DNA extract or 0.6 µl of RCA product were used as 154
template in 50 µl PCR reactions containing 1.5 mM MgCl2, 1X Apex NH4 buffer, 0.5 µM of each 155
primer, 0.2 mM of each dNTP, and 1U Apex Red Taq DNA Polymerase (Genesee Scientific, San 156
Diego, CA). Each PCR proceeded at 94ºC for 5 min, followed by 35 cycles of 94ºC for 1 min, 157
appropriate annealing temperature for 45 s (Table 1), and 72 ºC for 2.5 min, followed by a final 158
extension at 72 ºC for 8 min. Additionally, RCA products obtained from virus particles isolated 159
from a collection of 106 spiders from various families (Supplemental Table S1) were tested to 160
evaluate if SpOrbCVs could be detected in other spider species using the same PCR assay. This 161
spider collection included specimens from areas where we identified spinybacked orbweavers 162
positive for SpOrbCVs and specimens originally collected for CRESS DNA virus discovery 163
purposes [2]. 164
All PCRs included a negative process control composed of RCA product from SM buffer 165
DNA extracts processed alongside samples as well as a negative PCR control where sterile water 166
was used as template. DNA from the original spinybacked orbweavers where SpOrbCV-1 and 167
SpOrbCV-2 were detected served as positive controls. All PCR results were confirmed through 168
agarose gel electrophoresis and ethidium bromide staining. Templates resulting in strong, single 169
bands of the correct size relative to the positive control in agarose gels were considered positive, 170
whereas products producing weak bands were confirmed through Sanger sequencing with the 171
relevant PCR primers. 172
173 Egg surface decontamination to evaluate SpOrbCV-1 vertical transmission 174 175
Since eggs were positive for SpOrbCV-1, we evaluated if the virus could be detected in 176
individual eggs after surface decontamination. To test this, a total of 24 eggs from four egg sacs 177
known to be positive for SpOrbCV-1 were transferred individually to 1.5 ml microcentrifuge tubes 178
and processed through one out of four treatments. Each treatment was performed on three eggs 179
from each of two egg sacs for a total of six replicates per treatment. Individual eggs were rinsed 180
with SM buffer (untreated control) or decontaminated by submerging each egg in either 50 µl of 181
2% bleach for 5 min or 0.1% bleach for 30 mins [34]. For the remaining decontamination 182
treatment, each egg was submerged in 50 µl of 70% ethanol followed by 10% bleach for 1 min 183
each [35]. After bleach exposure, eggs were rinsed four times with 50 µl of SM buffer. The last 184

SM buffer wash was removed and DNA was immediately extracted from the rinsed egg using the 185
Extract-N-Amp Tissue PCR Kit (MilliporeSigma, St. Louis, MO). RCA products from each egg 186
were then tested using the PCR assay described above with SpOrbCV-1_F2/R2 primers. A 187
representative DNA extract from each treatment was spiked with a positive SpOrbCV-1 DNA 188
control and subjected to RCA followed by PCR to assess potential inhibition due to the 189
decontamination treatment. 190
191 Characterization of virion and tissue associated SpOrbCV-1 DNA 192 193
The single-stranded nature and polarity of the encapsidated SpOrbCV-1 DNA was 194
evaluated using approaches similar to those previously used to describe novel circular ssDNA 195
viruses [36]. All specimens selected for genome characterization assays were known to be positive 196
for SpOrbCV-1 based on leg subsamples. For virion strand or encapsidated DNA characterization 197
assays, virus particles were first purified from adult spinybacked orbweavers (n = 5) homogenized 198
in SM buffer. Virus particles were purified by filtering homogenates through a 0.22 µm Sterivex 199
filter and incubating 170 µl of filtrate at 37ºC for 2 hrs with a nuclease cocktail [37, 38] containing 200
1X Turbo DNAse Buffer (Invitrogen, Carlsbad, CA), 21U Turbo DNAse (Invitrogen, Carlsbad, 201
CA), 4.5U of Baseline-ZERO DNase (Epicentre, Madison, WI), and 112.5U Benzonase 202
(MilliporeSigma, Burlington, MA) to remove unencapsidated DNA. DNA was extracted from 203
200 µl of nuclease-treated viral particles using the QIAmp MinElute Virus Spin kit (Qiagen, 204
Hilden, Germany). The single-stranded nature of encapsidated SpOrbCV-1 DNA was then 205
assessed based on the susceptibility of the extracted DNA to S1 nuclease and restriction enzyme 206
digestion, which degrade single- and double-stranded DNA, respectively. 207
A controlled experiment using commercially available single- and double-stranded forms 208
of bacteriophage phiX174 circular DNA (New England Biolabs, Ipswich, MA) was conducted to 209
determine appropriate concentrations and incubation times for S1 nuclease treatment. Two PCR 210
assays with primer pairs, phiX174_F1/R1 and phiX174_F3/R3 (Table 1), were used to confirm 211
digestion. A total of 24U of S1 nuclease (Promega, Madison, WI) and incubation at 37ºC for 1 hr 212
was determined suitable to digest 120 ng of phiX174 ssDNA without completely degrading 120 213
ng of dsDNA. After appropriate S1 treatment conditions were determined, DNA extracted from 214
virus particles purified from spinybacked orbweavers were split into two 12 µl aliquots. One 215
aliquot was treated with 24U of S1 nuclease and incubated at 37ºC for 1 hr, while the other aliquot 216

was treated with 1 µl FastDigest EcoRI (Thermo Fisher Scientific, Waltham, MA) in 30 µl 217
reactions following manufacturer’s instructions. Products from the S1 nuclease treatment were 218
cleaned with the DNA Clean & Concentrator kit (Zymo Research, Irvine, CA) and eluted with 12 219
µl of sterile water. Cleaned digested products (2.5 µl) were used as template for a 25 µl PCR 220
reaction using SpOrbCV-1_F3/R3 primers (Table 1), which flank an EcoRI restriction site in the 221
SpOrbCV-1 genome, to confirm nuclease digestions. 222
A very similar nuclease treatment approach was used to determine if a dsDNA replicative 223
form (RF) of the SpOrbCV-1 genome could be detected in spinybacked orbweaver tissues. To do 224
this, the hard, shell-like abdomen was removed from five adult spinybacked orbweavers and DNA 225
was directly extracted from the cephalothorax. Spiderlings from six egg sacs were also investigated 226
for the presence of a RF. Total DNA was extracted from individual adults and twenty spiderlings 227
per egg sac using the ZR Tissue & Insect DNA Kit (Zymo Research, Irvine, CA) following the 228
manufacturer’s protocol. DNA extracts were treated with 24U of S1 nuclease in 30 µl reactions 229
and incubated at 37ºC for 1 hr. S1 nuclease digested products were cleaned using the DNA Clean 230
& Concentrator kit (Zymo Research, Irvine, CA) and eluted with 12 µl of sterile water. Clean S1 231
digested products (2.5 µl) were then treated with 1 µl FastDigest EcoRI following manufacturer’s 232
instructions in 10 µl reactions. Cleaned 10-fold diluted S1 digested products and products treated 233
with both S1 and EcoRI were used as template (2.5 µl) for PCR using SpOrbCV-1_F3/R3 primers 234
(Table 1) in 25 µl reactions. 235
The polarity of the encapsidated SpOrbCV-1 DNA was determined using strand-specific, 236
linear amplification with either SpOrbCV-1_F3 or SpOrbCV-1_R3 primers adapting a method 237
previously used to characterize circular ssDNA viral genomes [39]. For this purpose, 2.5 µl of 238
100-fold diluted DNA extracted from virus particles purified from adult spinybacked orbweavers 239
(n = 2) was linearly amplified for 50 cycles in 25 µl reactions containing 1.5 mM MgCl2, 1X Buffer 240
II, 0.4 µM of either primer, 0.2 mM of each dNTP, and 0.6U AmpliTaq Gold DNA Polymerase 241
(Applied Biosystems, Foster City, CA). A mock reaction containing template and all the reagents, 242
except for primer, was also included alongside strand-specific reactions to account for background. 243
Each amplification reaction proceeded at 94ºC for 5 min, followed by 50 cycles of 94ºC for 45 s, 244
52ºC for 45 s (Table 1), and 72ºC for 1 min, then a final extension at 72ºC for 8 min. One microliter 245
of linearly amplified products and mock reactions were then exponentially amplified in 25 µl PCR 246
reactions using primer pairs SpOrbCV1_F3n1/R3n1 and SpOrbCV1_F3n2/R3n2 (Table 1), which 247

are nested relative to strand-specific primers. PCR reactions contained 1.5 mM MgCl2, 1X Buffer 248
II, 0.4 µM of each primer, 0.2 mM of each dNTP, and 0.6U AmpliTaq Gold DNA Polymerase 249
(Applied Biosystems, Foster City, CA). Each reaction proceeded for 25 cycles using the same 250
thermocycling conditions listed for strand-specific linear amplification. Note that the encapsidated 251
viral DNA that is complementary to a given strand-specific primer will produce a strong signal 252
after exponential amplification. 253
254
Transmission electron microscopy analysis of SpOrbCV-1 positive specimens 255 256
Transmission electron microscopy (TEM) was used to observe virus-like particles (VLPs) 257
within spinybacked orbweaver tissues. For this purpose, legs were dissected from newly collected 258
adult spinybacked orbweavers and immediately fixed in a mixture of 4% glutaraldehyde and 2% 259
formaldehyde in 0.1M sodium cacodylate buffer, pH=7.3 using a mild vacuum. In addition, two 260
leg subsamples from each specimen were left unfixed and processed with the Extract-N-Amp 261
Tissue PCR Kit (MilliporeSigma, St. Louis, MO) to screen for SpOrbCV-1 through PCR using 262
methods outlined above. Legs from five positive specimens were chosen for TEM analysis. For 263
this purpose, fixed legs were rinsed for two hours with 0.1M sodium cacodylate buffer, pH=7.3 264
three times followed by an overnight rinse in the refrigerator and a final water rinse for 1 hour. 265
Rinsed samples were secondarily fixed in 2% aqueous OsO4 for 2 hours and rinsed in water for 30 266
minutes followed by overnight storage in water in the refrigerator. Tissue was enbloc stained in 267
0.5% aqueous uranyl acetate in the dark for 2 hours and then rinsed in water three times for 30 268
minutes each. Samples were dehydrated in an ethanol series, including 70% and 95% ethanol, for 269
15 minutes each, then two changes in 100% ethanol for 15 minutes each. A final dehydration 270
involved two 30 minute changes in 100% acetone. Leg tissues were embedded in Spurr’s resin and 271
cured in the oven. Ultrathin sections were cut and placed on copper grids for observation on a 272
Hitachi 7100 TEM at 100 kV. A Gatan Orius high-resolution digital camera was used to record 273
images. 274
275

Results and Discussion 276
SpOrbCV-1 represents a novel group of CRESS DNA viruses 277 278 SpOrbCV-1 and SpOrbCV-2 represent two distinct CRESS DNA viral groups identified 279
in spinybacked orbweavers. SpOrbCV-1 cannot be classified within established taxa, whereas 280
SpOrbCV-2 is an isolate of a species classified within the family Circoviridae (genus Cyclovirus) 281
known as dragonfly associated cyclovirus 3 (DfACyV-3; accession number JX185424) [2]. Since 282
SpOrbCV-1 shares similar genomic characteristics with unclassified viruses reported from other 283
spider species, dragonflies, and bat feces [2], we further explored the genomic sequences of 284
circularisviruses, named after the first virus reported from this group (dragonfly circularisvirus) 285
[27], and identified common features among this novel group of CRESS DNA viruses. Reciprocal 286
BLAST searches led to the identification of panicum ecklonii-associated virus (accession number 287
MH425571), a virus reported from the leaf tissue of a perennial grass [41], as an additional 288
circularisvirus. Therefore, circularisviruses include viruses identified in terrestrial arthropods, 289
animal feces, and plants (Table 2). 290
Circularisviruses contain small (~1.9 – 2 kb), circular genomes with two major ORFs 291
encoding Rep and capsid proteins in the same strand (Fig. 1). This genomic architecture was 292
deemed novel back in 2009 when metagenomic analyses uncovered small, circular ssDNA viral 293
genomes with gene arrangements that had only been observed in linear ssDNA viruses [42]. 294
Although similar genome organizations have been observed in CRESS DNA viruses reported from 295
various environments and organisms since then [28, 43-46], circularisviruses are the first 296
phylogenetic group whose members contain genomes with Rep and capsid protein coding 297
sequences on the same strand. Circularisviruses form a monophylogenetic clade when comparing 298
CRESS DNA viruses based on the Rep [2] and share > 57% genome-wide pairwise identity (PI) 299
among each other (Fig. 1), which is congruent with the lower limit of PIs reported for established 300
CRESS DNA viral families [47-49]. Similar to most CRESS DNA viral groups, the circularisvirus 301
Rep is more conserved than the putative capsid based on amino acid pairwise identities (Fig. 1). 302
Based on the lowest species demarcation threshold (75% genome-wide pairwise identity) for 303
established CRESS DNA viral families [50], currently there are eight distinct circularisvirus 304
species sharing <70% genome-wide PI and two genomes representing variants of SpOrbCV-1, 305
which share 91% genome-wide PI (Fig. 1). The distribution of genome-wide PIs among 306

circularisviruses should be evaluated as more representative genomes are discovered, which may 307
lead to the creation of a new CRESS DNA family. 308
To confirm the single-stranded nature of the SpOrbCV-1 genome, we took advantage of 309
S1 nuclease and EcoRI restriction endonuclease, which degrade ssDNA templates and dsDNA at 310
a specific site, respectively (Fig. 2). DNA extracts from purified virions were susceptible to S1 311
nuclease treatment but resistant to EcoRI digestion, confirming that SpOrbCV-1 virions 312
encapsidate circular ssDNA. Based on the putative ori of many CRESS DNA viruses, which 313
contains a conserved nonanucleotide motif (‘NANTATTAC’) at the apex of a stem-loop structure 314
[28], it was predicted that the protein-encoding strand of SpOrbCV-1 is the virion strand. A strand-315
specific amplification assay on DNA extracted from virions purified from two individual 316
spinybacked orbweavers confirmed that the Rep- and capsid-encoding strand containing the 317
nonanucleotide motif ‘CAGTATTAC’ is indeed the virion strand of this circularisvirus. The 318
majority of circularisvirus genomes contain a putative ori in the protein-coding strand with the 319
conserved nonanucleotide motif ‘NANTATTAC’ (Fig. 1). However, two genomes, one reported 320
from a spider (accession number MH545522) and the other from grass (accession number 321
MH425571), have a substitution in the nonamer, namely ‘NANTGTTAC’ and ‘NANTACTAC’, 322
respectively. Similar nonanucleotide variants have been observed in other CRESS DNA viruses, 323
such as members of the family Smacoviridae and some members of the family Genomoviridae 324
[47, 48]. 325
The circularisvirus Rep contains the two-domain organization that is distinctive of 326
eukaryotic CRESS DNA viruses, including endonuclease and superfamily 3 (SF3) helicase 327
domains that are important for rolling circle replication (RCR) (Fig. 1) [51, 52]. RCR motifs found 328
within the endonuclease domain are important for RCR initiation and termination, whereas the 329
helicase domain may be involved in DNA unwinding during elongation (reviewed by [28]). The 330
circularisvirus Rep endonuclease domain contains conserved residues observed in other CRESS 331
DNA viral Reps, including an arginine finger motif found within the SF3 helicase domain of most 332
CRESS DNA viral groups [53]. However, the Walker-A motif within the SF3 helicase domain 333
contains a unique substitution in comparison to all other CRESS DNA viral groups that is 334
consistently found in all circularisvirus Reps. The Walker-A motif, which may be involved in 335
substrate binding [54], is characterized by the sequence ‘GxxxxGK(S/T)’ in most CRESS DNA 336
viral groups, with the exception of members of the families Smacoviridae and Nanoviridae which 337

contain the sequence ‘GxxGK(S/T)’ [53]. However, the circularisvirus Walker-A motif contains 338
the sequence ‘GxxxxGKD’ (Fig. 1). The biochemical implications of this acidic amino acid 339
substitution within the Walker-A motif are unknown, but it seems to be a distinguishing feature of 340
circularisvirus Reps. Interestingly, BLAST analysis revealed an endogenous circularisvirus-like 341
SF3 helicase sequence in a genome scaffold from Parasteatoda tepidariorum (accession 342
XP_015919340), known as the common house spider. Since SF3 helicases have been mainly 343
identified in small DNA and RNA viruses [54, 55] and the P. tepidariorum has significant matches 344
(BLASTp, e-value < 10-50) to the helicase domain of circularisvirus Reps, including the 345
‘GxxxxGKD’ domain, it is possible that this spider genome acquired the helicase from a CRESS 346
DNA virus. 347
348
SpOrbCV-1 is highly prevalent in spinybacked orbweavers 349 350
SpOrbCV-1 and SpOrbCV-2 were originally identified in spinybacked orbweavers [2]; 351
however, SpOrbCV-2 shares 99% genome-wide pairwise identity with DfACyV-3, a cyclovirus 352
species originally discovered in dragonflies collected in Florida in 2010 [27]. Since spiders and 353
dragonflies are top insect predators, we hypothesized that SpOrbCV-2 is of dietary origin whereas 354
SpOrbCV-1 is specifically associated with spinybacked orbweavers. To further explore the 355
association between these CRESS DNA viruses and spinybacked orbweavers, we compared the 356
prevalence of SpOrbCV-1 and SpOrbCV-2 in specimens collected in 2017 and 2018 through PCR. 357
PCR assays indicated that SpOrbCV-1 was more prevalent in spinybacked orbweavers than 358
SpOrbCV-2 (Fig. 3). Notably, SpOrbV-1 was detected in at least 65% of specimens representing 359
various spinybacked orbweaver life stages, including adults, eggs, and spiderlings. In contrast, 360
SpOrbCV-2 was only detected in adults and the highest prevalence (36%) was detected among 361
samples composed of virus particles isolated from whole specimens compared to 2% based on leg 362
tissues alone. The vast majority (85%) of samples that were positive for SpOrbCV-2 (n = 13) were 363
also positive for SpOrbCV-1. In addition, SpOrbCV-1 was more abundant in leg samples than 364
SpOrbCV-2 given that the former was detected without RCA pre-amplification, whereas 365
SpOrbCV-2 was only detected in PCR reactions containing RCA product as template. A similar 366
trend was observed for virus particles isolated from whole bodies since 40% and 20% of 367
SpOrbCV-1 and SpOrbCV-2 positive samples, respectively, were directly identified from DNA 368
extracts, rather than RCA products. 369

Although most specimens were collected from a single screen pool enclosure, both 370
SpOrbCV-1 and SpOrbCV-2 were also detected in samples collected outside of this closely 371
monitored area. While SpOrbCV-2 was only detected in spiders from two screen pool enclosures, 372
SpOrbCV-1 was detected in samples collected from all tested sites, including six pool enclosures 373
and three outdoor areas. Therefore, SpOrbCV-1 is more widely associated with spinybacked 374
orbweavers than SpOrbCV-2. 375
Despite the high prevalence of SpOrbCV-1 in spinybacked orbweavers, including 376
SpOrbCV-2 positive specimens, this virus was not detected in any of the 106 specimens tested 377
representing various species from at least twelve families of spiders (Supplemental Table S1). 378
However, SpOrbCV-2 was detected in two specimens representing distinct spider species. One of 379
the SpOrbCV-2 positive specimens was collected within the closely monitored area where some 380
spinybacked orbweavers were also positive for this virus. Therefore, the PCR data support that 381
SpOrbCV-1 is highly prevalent and specifically associated with spinybacked orbweavers, whereas 382
SpOrbCV-2 is less prevalent and can be detected in different species of predatory arthropods 383
including insects (dragonflies) and at least three species of spiders. Although these data suggest 384
that SpOrbCV-2 is likely associated with dietary contents, at this time, we cannot rule out the 385
possibility that SpOrbCV-2 infects spinybacked orbweavers. For example, it is possible that 386
SpOrbCV-2 has different tissue tropism than SpOrbCV-1 and, thus, cannot be readily detected in 387
legs. Additionally, little is known about latent infections, a type of persistent infection where 388
virions are not produced, in arthropods [56]. PCR assays used to target viruses associated with 389
whole spiders only tested virus particles and, thus, prevalence data based on whole bodies did not 390
account for potential latent infections. 391
392 Similar VLPs are found in SpOrbCV-1 positive specimens 393 394
TEM was used to demonstrate the presence of VLPs within spider leg tissues that were 395
positive for SpOrbCV-1 by PCR. Similar icosahedral VLPs, 25 nm in diameter, were observed in 396
leg muscle tissue from each of the five specimens analyzed (Fig. 4). Although this morphology 397
and VLP size resembles that of well-characterized CRESS DNA viruses, such as circoviruses [57], 398
we cannot conclude that the observed VLPs represent SpOrbCV-1 without further research. It 399
should be noted that one of the specimens also contained a second type of VLPs with icosahedral 400

morphology, averaging 66 nm in diameter, indicating that some spinybacked orbweavers may be 401
co-infected with multiple viruses (Supplemental Fig. S1). 402
403 SpOrbCV-1 is active within spinybacked orbweavers and is vertically transmitted 404 405
Since SpOrbCV-1 was prevalent in spinybacked orbweavers we set out to determine if the 406
virus was replicating within spiders by screening tissues for a dsDNA replicative form (RF) form. 407
S1 nuclease treatment followed by EcoRI digestion of total DNA extracted from spider tissues was 408
used to detect SpOrbCV-1 dsDNA, keeping in mind that dsDNA will be resistant to S1 treatment 409
but not EcoRI digestions (Fig. 2). The five total DNA extracts from adult specimens subjected to 410
the enzyme treatments resulted in the detection of dsDNA. The dsDNA RF was also detected in 411
total DNA extracts from spiderlings, albeit with a lower detection rate (two out of six). These 412
findings confirm that SpOrbCV-1 replicates within spinybacked orbweavers, including spiderling 413
and adult life stages. 414
The prevalence of SpOrbCV-1 in eggs and spiderlings that had not emerged from the egg 415
sac indicates the vertical transmission of this virus among spinybacked orbweavers. To distinguish 416
between a transovum route of transmission, where contaminated egg surfaces lead to infected 417
offspring, and a transmission pathway where viruses are passed to offspring within eggs [34, 56], 418
we evaluated if the virus could still be detected after decontaminating egg surfaces. In at least five 419
out of six replicates, SpOrbCV-1 was detected in individual eggs after decontaminating the egg 420
surfaces using strategies from insect rearing studies, including 0.1% and 2% bleach treatments 421
(Kukan 1999). SpOrbCV-1 was also detected in three out of six eggs treated with 70% ethanol 422
followed by 10% bleach, which has been used as an effective surface decontamination strategy for 423
molecular analysis [35]. However, this may underestimate the number of positive eggs since PCR 424
inhibition was documented in the failed amplification of positive controls spiked into DNA 425
extracts from samples treated with 10% bleach. Nevertheless, the detection of SpOrbCV-1 in eggs 426
regardless of surface decontamination strategy indicates that this virus can be transmitted within 427
eggs. Based on the high prevalence of SpOrbCV-1 in female spinybacked orbweavers, we 428
speculate that this virus is maternally transmitted, which is a common form of parasite vertical 429
transmission in arthropods [58], through a transovarial route. However, the role of male spiders in 430
virus transmission cannot be excluded since the vast majority of specimens in this study were 431
female. 432

Elucidating viral transmission strategies is critical to understand how a given virus persists 433
in natural populations [59, 60]. To our knowledge, SpOrbCV-1 is the first virus found to be 434
vertically transmitted in spiders, since most studies investigating viruses in spiders have focused 435
on adults [21]. Moreover, although CRESS DNA viruses have been identified in all major lineages 436
of terrestrial arthropods [2], SpOrbCV-1 is the first CRESS DNA virus shown to be vertically 437
transmitted outside of fungal [61] and plant [62] viruses that are known to be vertically transmitted 438
within their insect vector population. Vertically transmitted viruses are expected to have low 439
virulence given that the fitness of the virus is tightly linked to that of its host [58, 60]. The high 440
prevalence of SpOrbCV-1 in seemingly healthy spinybacked orbweavers seems to follow this 441
general notion of low virulence. However, it remains to be determined if SpOrbCV-1 can also be 442
transmitted horizontally. Several DNA and RNA viruses have mixed-modes of transmission, 443
where the prevalence of either a vertical or horizontal route may provide a selective advantage 444
depending on host densities [56, 59]. Considering that vertically transmitted viruses may play a 445
major role in insect biology by manipulating their host’s physiology, behavior, and offspring sex-446
ratios to increase transmission [59, 63-65], future work should investigate the ecological role of 447
vertically transmitted viruses in spiders. 448
449

References 525
1. Zhao L, Rosario K, Breitbart M, Duffy S. Eukaryotic circular Rep-encoding single-526
stranded DNA (CRESS DNA) viruses: Ubiquitous viruses with small genomes and a diverse host 527
range. In: Kielian M, Mettenleiter TC, Roossinck MJ (editors). Advances in Virus Research: 528
Academic Press; 2019. pp. 71-133. 529
2. Rosario K, Mettel KA, Benner BE, Johnson R, Scott C et al. Virus discovery in all three 530
major lineages of terrestrial arthropods highlights the diversity of single-stranded DNA viruses 531
associated with invertebrates. PeerJ 2018;6:e5761. 532
3. Nyffeler M, Birkhofer K. An estimated 400–800 million tons of prey are annually killed 533
by the global spider community. The Science of Nature 2017;104(3):30. 534
4. Chen Z, Corlett RT, Jiao X, Liu S-J, Charles-Dominique T et al. Prolonged milk 535
provisioning in a jumping spider. Science 2018;362(6418):1052-1055. 536
5. Girard MB, Kasumovic MM, Elias DO. Multi-modal courtship in the peacock spider, 537
Maratus volans (O.P.-Cambridge, 1874). PLOS ONE 2011;6(9):e25390. 538
6. Mammola S, Michalik P, Hebets EA, Isaia M. Record breaking achievements by spiders 539
and the scientists who study them. PeerJ 2017;5:e3972. 540
7. Michalko R, Pekár S. The biocontrol potential of Philodromus (Araneae, Philodromidae) 541
spiders for the suppression of pome fruit orchard pests. Biol Control 2015;82:13-20. 542
8. Rypstra Ann L, Buddle Christopher M. Spider silk reduces insect herbivory. Biol Lett 543
2013;9(1):20120948. 544
9. Agnarsson I, Kuntner M, Blackledge TA. Bioprospecting finds the toughest biological 545
material: Extraordinary silk from a giant riverine orb spider. PLOS ONE 2010;5(9):e11234. 546
10. Saez NJ, Senff S, Jensen JE, Er SY, Herzig V et al. Spider-venom peptides as 547
therapeutics. Toxins 2010;2(12):2851. 548
11. King GF, Hardy MC. Spider-venom peptides: Structure, pharmacology, and potential for 549
control of insect pests. Annu Rev Entomol 2013;58(1):475-496. 550
12. Curry MM, Paliulis LV, Welch KD, Harwood JD, White JA. Multiple endosymbiont 551
infections and reproductive manipulations in a linyphiid spider population. Heredity 552
2015;115(2):146-152. 553
13. Zhang L, Yun Y, Hu G, Peng Y. Insights into the bacterial symbiont diversity in spiders. 554
Ecol Evol 2018;8(10):4899-4906. 555

14. Goodacre SL, Martin OY, Bonte D, Hutchings L, Woolley C et al. Microbial 556
modification of host long-distance dispersal capacity. BMC Biol 2009;7(1):32. 557
15. Goodacre SL, Martin OY. Modification of insect and arachnid behaviours by vertically 558
transmitted endosymbionts: Infections as drivers of behavioural change and evolutionary novelty. 559
Insects 2012;3(1):246-261. 560
16. Gunnarsson B, Hewitt GM, Goodacre SL. Sex ratio, mating behaviour and Wolbachia 561
infections in a sheetweb spider. Biol J Linnean Soc 2009;98(1):181-186. 562
17. Vanthournout B, Swaegers J, Hendrickx F. Spiders do not escape reproductive 563
manipulations by Wolbachia. BMC Evol Biol 2011;11(1):15. 564
18. Goodacre SL. Endosymbiont infections in spiders. In: Casas J (editor). Advances in Insect 565
Physiology: Academic Press; 2011. pp. 137-153. 566
19. Buck CB, Van Doorslaer K, Peretti A, Geoghegan EM, Tisza MJ et al. The ancient 567
evolutionary history of polyomaviruses. PLOS Pathog 2016;12(4):e1005574. 568
20. Dennis TPW, Flynn PJ, Marciel de Souza W, Singer JB, Moreau CS et al. Insights into 569
circovirus host range from the genomic fossil record. J Virol 2018;92(16): e00145-00118. 570
21. Debat HJ. An RNA virome associated to the golden orb-weaver spider Nephila clavipes. 571
Front Microbiol 2017;8:2097. 572
22. Shi M, Lin XD, Tian JH, Chen LJ, Chen X et al. Redefining the invertebrate RNA 573
virosphere. Nature 2016(540):539–543. 574
23. Li CX, Shi M, Tian JH, Lin XD, Kang YJ et al. Unprecedented genomic diversity of 575
RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. Elife 576
2015;4:e05378. 577
24. Shean RC, Makhsous N, Crawford RL, Jerome KR, Greninger AL. Draft genome 578
sequences of six novel picorna-like viruses from Washington State spiders. Genome Announc 579
2017;5(9):e01705-01716. 580
25. Muma MH. Biological and behavioral notes on Gasteracantha cancriformis (Arachnida: 581
Araneidae). Fla Entomol 1971;54:345-352. 582
26. Levi HW. The American orb-weaver genera Colphepeira, Micrathena, and Gasteracantha 583
north of Mexico (Araneae, Araneidae). Bull Mus Comp Zool 1978;148:417-442. 584

27. Rosario K, Dayaram A, Marinov M, Ware J, Kraberger S et al. Diverse circular 585
ssDNA viruses discovered in dragonflies (Odonata: Epiprocta). J Gen Virol 2012;93(Pt 12):2668-586
2681. 587
28. Rosario K, Duffy S, Breitbart M. A field guide to eukaryotic circular single-stranded 588
DNA viruses: Insights gained from metagenomics. Arch Virol 2012;157(10):1851-1871. 589
29. Muhire BM, Varsani A, Martin DP. SDT: A virus classification tool based on pairwise 590
sequence alignment and identity calculation. PLOS ONE 2014;9(9):e108277. 591
30. Kumar S, Stecher G, Tamura K. MEGA7: Molecular Evolutionary Genetics Analysis 592
version 7.0 for bigger datasets. Mol Biol Evol 2016;33(7):1870-1874. 593
31. Edgar RC. MUSCLE: multiple sequence alignment with high accuracy and high 594
throughput. Nucleic Acids Res 2004;32(5):1792-1797. 595
32. Crooks GE, Hon G, Chandonia JM, Brenner SE. WebLogo: a sequence logo generator. 596
Genome Res 2004;14(6):1188-1190. 597
33. Kim KH, Bae JW. Amplification methods bias metagenomic libraries of uncultured 598
single-stranded and double-stranded DNA viruses. Appl Environ Microb 2011;77(21):7663-7668. 599
34. Kukan B. Vertical transmission of nucleopolyhedrovirus in insects. J Invertebr Pathol 600
1999;74(2):103-111. 601
35. Hammer TJ, Dickerson JC, Fierer N. Evidence-based recommendations on storing and 602
handling specimens for analyses of insect microbiota. PeerJ 2015;3:e1190. 603
36. Ng TFF, Manire C, Borrowman K, Langer T, Ehrhart L et al. Discovery of a novel 604
single-stranded DNA virus from a sea turtle fibropapilloma by using viral metagenomics. J Virol 605
2009;83(6):2500-2509. 606
37. Victoria JG, Kapoor A, Li L, Blinkova O, Slikas B et al. Metagenomic analyses of 607
viruses in stool samples from children with acute flaccid paralysis. J Virol 2009;83(9):4642-4651. 608
38. Ng TF, Marine R, Wang C, Simmonds P, Kapusinszky B et al. High variety of known 609
and new RNA and DNA viruses of diverse origins in untreated sewage. J Virol 610
2012;86(22):12161-12175. 611
39. Okamoto H, Ukita M, Nishizawa T, Kishimoto J, Hoshi Y et al. Circular double-612
stranded forms of TT virus DNA in the liver. J Virol 2000;74(11):5161-5167. 613
40. Kazlauskas D, Varsani A, Krupovic M. Pervasive chimerism in the replication-614
associated proteins of uncultured single-stranded DNA viruses. Viruses 2018;10(4):187. 615

41. Richet C, Kraberger S, Filloux D, Bernardo P, Harkins GW et al. Novel circular DNA 616
viruses associated with Apiaceae and Poaceae from South Africa and New Zealand. Arch Virol 617
2019;164(1):237-242. 618
42. Rosario K, Duffy S, Breitbart M. Diverse circovirus-like genome architectures revealed 619
by environmental metagenomics. J Gen Virol 2009;90:2418-2424. 620
43. Pham HT, Bergoin M, Tijssen P. Acheta domesticus volvovirus, a novel single-stranded 621
circular DNA virus of the house cricket. Genome Announc 2013;1(2):e00079. 622
44. Dayaram A, Galatowitsch ML, Argüello-Astorga GR, van Bysterveldt K, Kraberger 623
S et al. Diverse circular replication-associated protein encoding viruses circulating in invertebrates 624
within a lake ecosystem. Infect Genet Evol 2016;39:304-316. 625
45. Dayaram A, Goldstien S, Argüello-Astorga GR, Zawar-Reza P, Gomez C et al. 626
Diverse small circular DNA viruses circulating amongst estuarine molluscs. Infect Genet Evol 627
2015;31:284-295. 628
46. Dayaram A, Potter KA, Pailes R, Marinov M, Rosenstein DD et al. Identification of 629
diverse circular single-stranded DNA viruses in adult dragonflies and damselflies (Insecta: 630
Odonata) of Arizona and Oklahoma, USA. Infect Genet Evol 2015;30:278-287. 631
47. Varsani A, Krupovic M. Sequence-based taxonomic framework for the classification of 632
uncultured single-stranded DNA viruses of the family Genomoviridae. Virus Evol 633
2017;3(1):vew037. 634
48. Varsani A, Krupovic M. Smacoviridae: A new family of animal-associated single-635
stranded DNA viruses. Arch Virol 2018;163(7):2005-2015. 636
49. Rosario K, Breitbart M, Harrach B, Segalés J, Delwart E et al. Revisiting the taxonomy 637
of the family Circoviridae: establishment of the genus Cyclovirus and removal of the genus 638
Gyrovirus. Arch Virol 2017;162(5):1447-1463. 639
50. Lefkowitz EJ, Dempsey DM, Hendrickson RC, Orton RJ, Siddell SG et al. Virus 640
taxonomy: the database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic 641
Acids Res 2018;46(Database issue):D708-D717. 642
51. Koonin EV, Dolja VV, Krupovic M. Origins and evolution of viruses of eukaryotes: The 643
ultimate modularity. Virology 2015;479–480:2-25. 644
52. Koonin EV, Ilyina TV. Computer-assisted dissection of rolling circle DNA replication. 645
Biosystems 1993;30(1):241-268. 646

53. Kazlauskas D, Dayaram A, Kraberger S, Goldstien S, Varsani A et al. Evolutionary 647
history of ssDNA bacilladnaviruses features horizontal acquisition of the capsid gene from ssRNA 648
nodaviruses. Virology 2017;504:114-121. 649
54. Gorbalenya AE, Koonin EV. Helicases: amino acid sequence comparisons and structure-650
function relationships. Curr Opin Struc Biol 1993;3(3):419-429. 651
55. Gorbalenya AE, Koonin EV, Wolf YI. A new superfamily of putative NTP-binding 652
domains encoded by genomes of small DNA and RNA viruses. FEBS Lett 1990;262(1):145-148. 653
56. Williams T, Virto C, Murillo R, Caballero P. Covert infection of insects by 654
baculoviruses. Front Microbiol 2017;8:1337-1337. 655
57. Breitbart M, Delwart E, Rosario K, Segalés J, Varsani A et al. ICTV virus taxonomy 656
profile: Circoviridae. J Gen Virol 2017;98(8):1997-1998. 657
58. Longdon B, Jiggins FM. Vertically transmitted viral endosymbionts of insects: Do sigma 658
viruses walk alone? Proc Biol Sci 2012;279(1744):3889-3898. 659
59. Cory JS. Insect virus transmission: Different routes to persistence. Curr Opin Insect Sci 660
2015;8:130-135. 661
60. Fine PEM. Vectors and vertical transmission: An epidemiologic perspective. Ann NY Acad 662
Sci 1975;266(1):173-194. 663
61. Liu S, Xie J, Cheng J, Li B, Chen T et al. Fungal DNA virus infects a mycophagous 664
insect and utilizes it as a transmission vector. Proc Natl Acad Sci USA 2016;113(45):12803–12808. 665
62. Wei J, He Y-Z, Guo Q, Guo T, Liu Y-Q et al. Vector development and vitellogenin 666
determine the transovarial transmission of begomoviruses. Proc Natl Acad Sci USA 667
2017;114(26):6746-6751. 668
63. Liu B, Preisser EL, Chu D, Pan H, Xie W et al. Multiple forms of vector manipulation 669
by a plant-infecting virus: Bemisia tabaci and Tomato yellow leaf curl virus. J Virol 670
2013;87(9):4929-4937. 671
64. Clem RJ, Passarelli AL. Baculoviruses: sophisticated pathogens of insects. PLOS Pathog 672
2013;9(11):e1003729. 673
65. Wang F, Fang Q, Wang B, Yan Z, Hong J et al. A novel negative-stranded RNA virus 674
mediates sex ratio in its parasitoid host. PLOS Pathog 2017;13(3):e1006201. 675

66. Male MF, Kraberger S, Stainton D, Kami V, Varsani A. Cycloviruses, 676
gemycircularviruses and other novel replication-associated protein encoding circular viruses in 677
Pacific flying fox (Pteropus tonganus) faeces. Infect Genet Evol 2016;39:279-292. 678

Author Statements 450
Authors and contributions 451
KR and MB designed study. KR, KAM, AMG, and MB collected specimens. KR and KAM 452
performed experimental work and data analysis. AMG performed transmission electron 453
microscopy analysis. KR wrote the manuscript and prepared figures. KAM and MB reviewed and 454
edited manuscript. All authors approved final draft. 455
456
Conflicts of interest 457 The authors declare that there are no conflicts of interest. 458
459
Funding information 460
This work was funded through NSF Assembling the Tree of Life Program grant DEB-1239976 to 461
KR and MB. 462
463
Acknowledgements 464
Authors would like to thank Alexandria Creasy, Catherine Scott, and William Landry for providing 465
samples, Mason Kerr for help with sample processing, and Bentley Fane for advice on nuclease 466
treatment assays. 467

Tables and Figure Legends 468 469 Table 1. Primers used for PCR assays performed during this study. 470 471 Table 2. Summary of viral isolates representing circularisviruses. 472 473 Figure 1. Genome organization (A), conserved motifs (B) and pairwise sequence comparisons (C) 474
for circularisviruses. Panel A shows a genome schematic of major open reading frames found 475
within circularisvirus genomes, including replication-associated (Rep, red arrow) and capsid (Cp, 476
grey arrow) protein coding regions, and the nonanucleotide motif marking the putative origin of 477
replication within a predicted stem-loop structure. Lower case letters within the nonanucleotide 478
motif indicate nucleotides that were observed in a single genome sequence and ‘n’ represents any 479
nucleotide. Panel B depicts conserved motifs found within the Rep through amino acid sequence 480
logos generated in WebLogo 3. The endonuclease domain sequence logo, including rolling circle 481
replication (RCR) motifs I through III, is shown above the superfamily 3 (SF3) helicase domain 482
logo, which includes Walker-A, Walker-B, Motif C, and arginine (R) finger motifs. Panel C shows 483
pairwise identities calculated using complete genomes (left) or predicted amino acid sequences 484
(right) for the Rep (dark grey) and Cp (light grey). The information for each panel was extracted 485
from the nine genome sequences representing members of the circularisviruses, namely dragonfly 486
circularisvirus (DfCir), two isolates of spinybacked orbweaver circular virus 1 (SpOrbCV-1A and 487
SpOrbCV-1B), golden silk orbweaver associated circular virus 1 (GoSOrbACV-1), longjawed 488
orbweaver circular virus 1 (LjOrbCV-1), cybaeus spider associated circular virus 1 (CySACV-1), 489
pacific flying fox feces associated circular DNA virus 6 (PfffACV-6), and panicum ecklonii-490
associated virus (PeaV). 491
492
Figure 2. Schematic representation of methodological framework used to elucidate the single- or 493
double-stranded nature of spinybacked orbweaver circular virus 1 DNA found within virions and 494
spinybacked orbweaver tissues. Note that S1 nuclease degrades ssDNA templates (dotted line) 495
non-specifically (left panel), whereas EcoRI restriction endonuclease nicks dsDNA at a specific 496
site (shown as a gap) (right panel). PCR was used to confirm endonuclease treatment results. Grey 497
arrows represent primer positions. PCR results for DNA from virions are highlighted in blue, 498
whereas results for DNA from tissues are highlighted in orange. Note that S1 nuclease treated 499
DNA from spinybacked orbweaver tissues was then treated with EcoRI to confirm its double-500

stranded nature (indicated by the orange spider). An example gel image is provided in 501
Supplemental Figure S2. 502
Figure 3. Images of spinybacked orbweaver life stages investigated during this study (A) and 503
percentage of specimens from each life stage that were positive for spinybacked orbweaver circular 504
virus (SpOrbCV) 1 and 2 (B). Each specimen category on the x-axis specifies if the data reflects 505
the prevalence for specimens collected during 2017, 2018 or both years and the number (N) of 506
individuals processed. Prevalence in adult specimens based on whole body (WB) and leg (L) 507
samples are distinguished. 508
Figure 4. Transmission electron microscope images of virus-like particles (VLPs) found in leg 509
tissues from spiders that tested positive for spinybacked orbweaver circular virus 1. VLPs had an 510
average diameter of 25 nm and each image originates from a different specimen. 511
512
513
Supplemental Table and Figure Legends 514
Table S1. List of spider species tested for spinybacked orbweaver circular viruses (SpOrbCV). 515 516
Figure S1. Transmission electron microscope images showing a second type of virus-like particles 517
(VLPs) observed within the leg tissues of a spinybacked orbweaver. Observed VLPs averaged ~66 518
nm in diameter. 519
520
Figure S2. Example gel image showing results for PCR assays targeting DNA extracted from 521
virions (top) or spider tissues (bottom) after treatment with S1 nuclease or EcoRI restriction 522
enzyme. Samples include an untreated positive control (U), PCR products from virion or tissue 523
samples (S), and a negative control (N). 524

Table 1. Primers used for PCR assays performed during this study.
1Genome positions for each primer used to target spinybacked orbweavers circular viruses (SpOrbCV) and bacteriophage phiX174 (GenBank accession number NC_001422). 2Product sizes are listed in kilobases (kb) or base pairs (bp). 3Annealing temperatures highlighted with an asterisk (*) were incrementally decreased by 0.1˚C every cycle.
Targeted virus Purpose Primer pair and sequences (5’ – 3’) Primer
positions1 Product length2
Annealing temp (˚C)3
SpOrbCV-1 Complete genome recovery (inverse PCR)
SpOrbCV1_F1: CACCACCTCCTGATTTAG SpOrbCV1_R1: CATTTAAGGATGTTTTGCTT
377 – 394 395 – 414 2 kb 42
SpOrbCV-1 Sample screening for prevalence estimates
SpOrbCV1_F2: ACGGATGAACAAGCCTGGAT SpOrbCV1_R2: ACCGTCGAAGAAGAAGAAACG
455 – 474 1038 – 1058 1.2 kb 50
SpOrbCV-1 Genome characterization assays
SpOrbCV1_F3: ATCCAGGCTTGTTCATCCGT SpOrbCV1_R3: CGTTTCTTCTTCTTCGACGGT
455 – 474 1038 – 1058 600 bp 52
SpOrbCV-1 Nested PCR for genome characterization assays
SpOrbCV1_F3n1: TCTGTTAAGTTTCGAGTTGAGGA SpOrbCV1_R3n1: AGCCAAACACAATGCGAAGG
496 – 518 673 – 692 200 bp 52
SpOrbCV-1 Nested PCR for genome characterization assays
SpOrbCV1_F3n2: GTGGAACCTTCGCATTGTGT SpOrbCV1_R3n2: CTGGCGGGGAGAAAATGAAC
667 – 686 875 – 894 230 bp 52
SpOrbCV-2 Sample screening for prevalence estimates
SpOrbCV2_F1: ATCTTATCTGGGGCCGATGG SpOrbCV2_R1: AACATTTCTGCGGCCTCTTG
24 – 43 639 – 658 1.1 kb 54
phiX174 S1 nuclease experiment phiX174_F1: TCAGATATGGACCTTGCTGCT phiX174_R1: TCCTTGACGAACGTGCCAA
5034 – 5054 254 – 272 625 bp 54*
phiX174 S1 nuclease experiment phiX174_F3: ACGCCGTTCAACCAGATATTG phiX174_R3: GGTTGAACAGCATCGGACTC
5226 – 5246 352 – 371 530 bp 53*

Table 2. Summary of viral isolates representing circularisviruses.
Virus1 Accession No.2 Isolate
Isolation source Reference Source Scientific name Sample type
Dragonfly circularisvirus JX185415 TO-DF3E-2010 Dragonfly Pantala flavescens Abdomen [27]
Cybaeus spider ACV-1 MH545522 BC_I1644C_F12 Spider Cybaeus signifer Whole specimen [2]
Golden silk orbweaver ACV-1 MH545520 PR_I0960_F8 Spider Nephila clavipes Whole specimen [2]
Longjawed orbweaver CV-1 MH545521 BC_I1601_F12 Spider Leucauge argyra Whole specimen [2]
Spinybacked orbweaver CV-1 MH545518 FL_I0831_I69-H8 Spider Gasteracantha cancriformis Whole specimen [2]
Spinybacked orbweaver CV-1 MH545519 FL_I1594-I104_E11 Spider Gasteracantha cancriformis Whole specimen [2]
Pacific flying fox faeces ACV-6 KT732823 Tbat_H_77994 Bat Pteropus tonganus Feces [66]
Panicum ecklonii AV MH425571 2-82-I Grass Panicum ecklonii Leaf [41]
Odonata ACV-13 KM598396 US-1591LM1-12 Dragonfly Libellula quadrimaculata Abdomen [46] 1Associated circular virus (ACV); circular virus (CV); associated virus (AV) 2GenBank accession number

1960 - 2070 nt
(C/T)A(G/n)T(A/g)(T/c)TAC
)p
C( di s
pa
C
PfffACV-6
PeaV
SpOrbCV-1A
GoSOrbACV-1
OdACV-13
DfCirc
LjOrbCV-1
CySACV-1
SpOrbCV-1B
KT
732823
MH
425571
MH
425518
MH
425519
MH
425522
MH
425520
KM
598396
JX 1
85415
MH
425521
Pairwise identity (%)
100 92 84 76 68 60 52 44 36 28 20
Genome
DfCirc
SpOrbCV-1A
SpOrbCV-1B
LjOrbCV-1
GoSOrbCV-1
OdACV-13
CySACV-1
PfffACV-6
SpO
rbC
V-1
B
SpO
rbC
V-1
A
PeaV
KT
732823
MH
425571
MH
425518
MH
425519
MH
425522
MH
425520
KM
598396
JX 1
85415
MH
425521
PeaV
DfC
irc
LjO
rbC
V-1
GoS
Orb
CV
-1
OdA
CV
-13
CyS
AC
V-1
PfffA
CV
-6
KT 732823
MH 425571
MH 425518
MH 425519
MH 425522
MH 425520
KM 598396
JX 185415
MH 425521
Rep
Cp
A) Genome organization B) Rep conserved motifs
C) Pairwise identities
RCR I RCR II RCR III
Walker-A Walker-B Motif C R-Finger
0
1
2
3
bits
YWVLTCYFTRAVSYF HYKILQGYF YCSKKD
0
1
2
3bits GKPSP G
RICSGKD I
VVHL ILQSDMLIFVQED M
VTSNYF A IVLRQAKSTRARCF

S1 nuclease digest EcoRI restriction digest
PCR PCR
- + -+
? ?

20
40
60
80
100
Adults-WB Adults-L Eggs SpiderlingsN = 282017
N = 1212017-2018
N = 262017-2018
N = 142018
SpOrbCV-1 SpOrbCV-2
Posi
tive s
am
ple
s (%
)
Adult Eggs Spiderlings
A) Investigated spinybacked orbweaver life stages
B) Viral prevalence in specimens representing each life stage
65%
36%
79%
2.5%
74%
0% 0%
79%

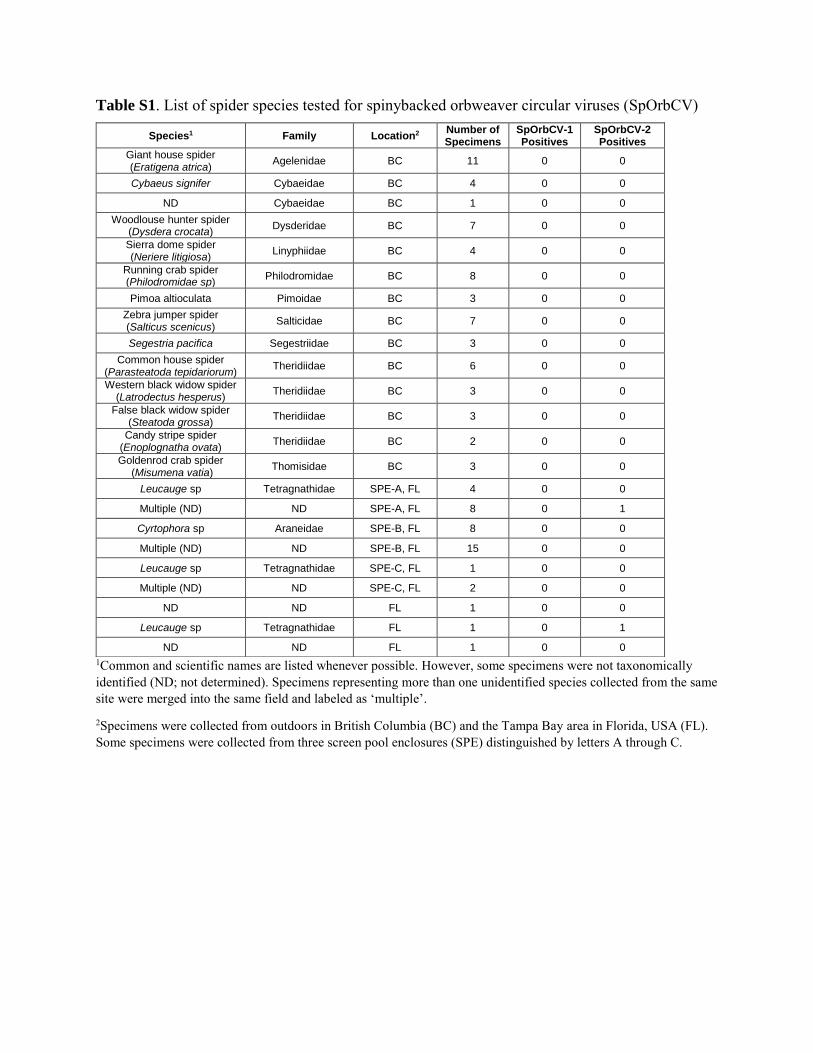
Table S1. List of spider species tested for spinybacked orbweaver circular viruses (SpOrbCV)
1Common and scientific names are listed whenever possible. However, some specimens were not taxonomically identified (ND; not determined). Specimens representing more than one unidentified species collected from the same site were merged into the same field and labeled as ‘multiple’.
2Specimens were collected from outdoors in British Columbia (BC) and the Tampa Bay area in Florida, USA (FL). Some specimens were collected from three screen pool enclosures (SPE) distinguished by letters A through C.
Species1 Family Location2 Number of Specimens
SpOrbCV-1 Positives
SpOrbCV-2 Positives
Giant house spider (Eratigena atrica) Agelenidae BC 11 0 0
Cybaeus signifer Cybaeidae BC 4 0 0
ND Cybaeidae BC 1 0 0 Woodlouse hunter spider
(Dysdera crocata) Dysderidae BC 7 0 0
Sierra dome spider (Neriere litigiosa) Linyphiidae BC 4 0 0
Running crab spider (Philodromidae sp) Philodromidae BC 8 0 0
Pimoa altioculata Pimoidae BC 3 0 0 Zebra jumper spider (Salticus scenicus) Salticidae BC 7 0 0
Segestria pacifica Segestriidae BC 3 0 0 Common house spider
(Parasteatoda tepidariorum) Theridiidae BC 6 0 0
Western black widow spider (Latrodectus hesperus) Theridiidae BC 3 0 0
False black widow spider (Steatoda grossa) Theridiidae BC 3 0 0
Candy stripe spider (Enoplognatha ovata) Theridiidae BC 2 0 0
Goldenrod crab spider (Misumena vatia) Thomisidae BC 3 0 0
Leucauge sp Tetragnathidae SPE-A, FL 4 0 0
Multiple (ND) ND SPE-A, FL 8 0 1
Cyrtophora sp Araneidae SPE-B, FL 8 0 0
Multiple (ND) ND SPE-B, FL 15 0 0
Leucauge sp Tetragnathidae SPE-C, FL 1 0 0
Multiple (ND) ND SPE-C, FL 2 0 0
ND ND FL 1 0 0
Leucauge sp Tetragnathidae FL 1 0 1
ND ND FL 1 0 0

Figure S1. Transmission electron microscope images showing a second type of virus-like particles (VLPs) observed within the leg tissues of a spinybacked orbweaver. Observed VLPs averaged ~66 nm in diameter.
Figure S1

U S6 S7 S8 S9 S10 N S6 S7 S8 S9 S10 N
Figure S2. Example gel image showing results for PCR assays targeting DNA extracted from virions (top) or spider tissues (bottom) after treatment with S1 nuclease or EcoRI restriction enzyme. Samples include an untreated positive control (U), PCR products from virion or tissue samples (S), and a negative control (N).
Figure S2
U S1 S2 S3 S4 S5 N S1 S2 S3 S4 S5 N
U S6 S7 S8 S9 S10 N
Virions
Tissues
S1 nuclease EcoRI
S1 nuclease S1 followed by EcoRI
S6 S7 S8 S9 S10 N


















