
Prevalence and mechanisms of aminoglycoside resistance: A ten-year study
8
Prevalence and Mechanisms of Aminoglycoside Resistance A Ten-Year Study IAN PHILLIPS, M.D. ANNA KING, A.A.I.M.L.T. KEVIN SHANNON, Ph.D. London, United Kingdom Aminoglycoside resistance was monitored at St. Thomas’ Hospital from 1975 to 1984. Gentamicin resistance had appeared in a number of species by 1975, but remained rare (less than 1 percent of iso- lates) in Escherichia coli throughout the study period. Gentamicin- resistant Klebsielleae had become fairly common (8 percent of iso- lates) by 1977, but little change has subsequently occurred in their frequency of isolation. Serratia species are not frequently isolated; gentamicin resistance in these organisms was not observed until 1979. Since then, 10 to 20 percent of isolates have been found to be resistant. Except for Providencia, most isolates of which were gen- tamicin-resistant, less than 5 percent of the Enterobacteriaceae iso- lated were found to be resistant to gentamicin during the lo-year period. Throughout the study, approximately 5 percent of the Pseu- domonas aeruginosa isolates were resistant to gentamicin. Less than 5 percent of the isolates of Acinetobacter were resistant to gentamicin before 1979, at which time 40 percent were found to be resistant; subsequently, gentamicin resistance among these orga- nisms has become somewhat less common. On the whole, tobramy- tin resistance has mirrored gentamicin resistance. However, before 1979, most gentamicin-resistant Klebsielleae isolates had retained susceptibility to tobramycin, as had most gentamicin-resistant iso- lates of Acinetobacter and P. aeruginosa. Amikacin resistance has remained very unusual in all organisms, apart from non-aeruginosa Pseudomonas species. Until 1977, nearly all the resistance among Enterobacteriaceae was attributable to AAC(3)-I, except for that caused by AAC(2’) production in Providencia and the non-enzy- matic resistance observed in E. coli. However, more recently, AAC(3)-II and AAD(2”) have become the most common mechanisms of resistance. The resistance of most gentamicin-resistant isolates of P. aeruginosa from 1974 to 1977 was attributable to non-enzy- matic mechanisms; subsequently, such resistance was more often caused by AAC(3)-I, AAC(G’), or AAD(2”). Gentamicin resistance first appeared in Staphylococcus aureus in 1978, after which about 1 to 2 percent of the isolates from hospitalized patients were found to be resistant, mostly because of production of AAC(6’) and APH(2”). From the United Medical and Dental Schools of Guy’s and St. Thomas’ Hospitals, London, United Kingdom. Requests for reprints should be ad- dressed to Dr. Ian Phillips, St. Thomas’ Hospital Medical School, Department of Microbiology, Lon- don SE1 7EH, United Kingdom. At St. Thomas’ Hospital, since the late 196Os, gentamicin has been the aminoglycoside of choice for use with suitable beta-lactams in the treat- ment of infections caused by gram-negative organisms, Resistance among the Enterobacteriaceae and Pseudomonas species was rare when gentamicin was introduced, but reports of resistance among strains began to be published in the early 1970s. Much of this resistance was 48 June 30, 1988 The American Journal of Medicine Volume 80 (suppl 8B)
-
Upload
ian-phillips -
Category
Documents
-
view
212 -
download
0
Transcript of Prevalence and mechanisms of aminoglycoside resistance: A ten-year study

Prevalence and Mechanisms of Aminoglycoside Resistance
A Ten-Year Study
IAN PHILLIPS, M.D. ANNA KING, A.A.I.M.L.T. KEVIN SHANNON, Ph.D.
London, United Kingdom
Aminoglycoside resistance was monitored at St. Thomas’ Hospital from 1975 to 1984. Gentamicin resistance had appeared in a number of species by 1975, but remained rare (less than 1 percent of iso- lates) in Escherichia coli throughout the study period. Gentamicin- resistant Klebsielleae had become fairly common (8 percent of iso- lates) by 1977, but little change has subsequently occurred in their frequency of isolation. Serratia species are not frequently isolated; gentamicin resistance in these organisms was not observed until 1979. Since then, 10 to 20 percent of isolates have been found to be resistant. Except for Providencia, most isolates of which were gen- tamicin-resistant, less than 5 percent of the Enterobacteriaceae iso- lated were found to be resistant to gentamicin during the lo-year period. Throughout the study, approximately 5 percent of the Pseu- domonas aeruginosa isolates were resistant to gentamicin. Less than 5 percent of the isolates of Acinetobacter were resistant to gentamicin before 1979, at which time 40 percent were found to be resistant; subsequently, gentamicin resistance among these orga- nisms has become somewhat less common. On the whole, tobramy- tin resistance has mirrored gentamicin resistance. However, before 1979, most gentamicin-resistant Klebsielleae isolates had retained susceptibility to tobramycin, as had most gentamicin-resistant iso- lates of Acinetobacter and P. aeruginosa. Amikacin resistance has remained very unusual in all organisms, apart from non-aeruginosa Pseudomonas species. Until 1977, nearly all the resistance among Enterobacteriaceae was attributable to AAC(3)-I, except for that caused by AAC(2’) production in Providencia and the non-enzy- matic resistance observed in E. coli. However, more recently, AAC(3)-II and AAD(2”) have become the most common mechanisms of resistance. The resistance of most gentamicin-resistant isolates of P. aeruginosa from 1974 to 1977 was attributable to non-enzy- matic mechanisms; subsequently, such resistance was more often caused by AAC(3)-I, AAC(G’), or AAD(2”). Gentamicin resistance first appeared in Staphylococcus aureus in 1978, after which about 1 to 2 percent of the isolates from hospitalized patients were found to be resistant, mostly because of production of AAC(6’) and APH(2”).
From the United Medical and Dental Schools of Guy’s and St. Thomas’ Hospitals, London, United Kingdom. Requests for reprints should be ad- dressed to Dr. Ian Phillips, St. Thomas’ Hospital Medical School, Department of Microbiology, Lon- don SE1 7EH, United Kingdom.
At St. Thomas’ Hospital, since the late 196Os, gentamicin has been the aminoglycoside of choice for use with suitable beta-lactams in the treat- ment of infections caused by gram-negative organisms, Resistance among the Enterobacteriaceae and Pseudomonas species was rare when gentamicin was introduced, but reports of resistance among strains began to be published in the early 1970s. Much of this resistance was
48 June 30, 1988 The American Journal of Medicine Volume 80 (suppl 8B)
SYMPOSIUM ON AMINOGLYCOSIDE THERAPY-PHILLIPS ET AL
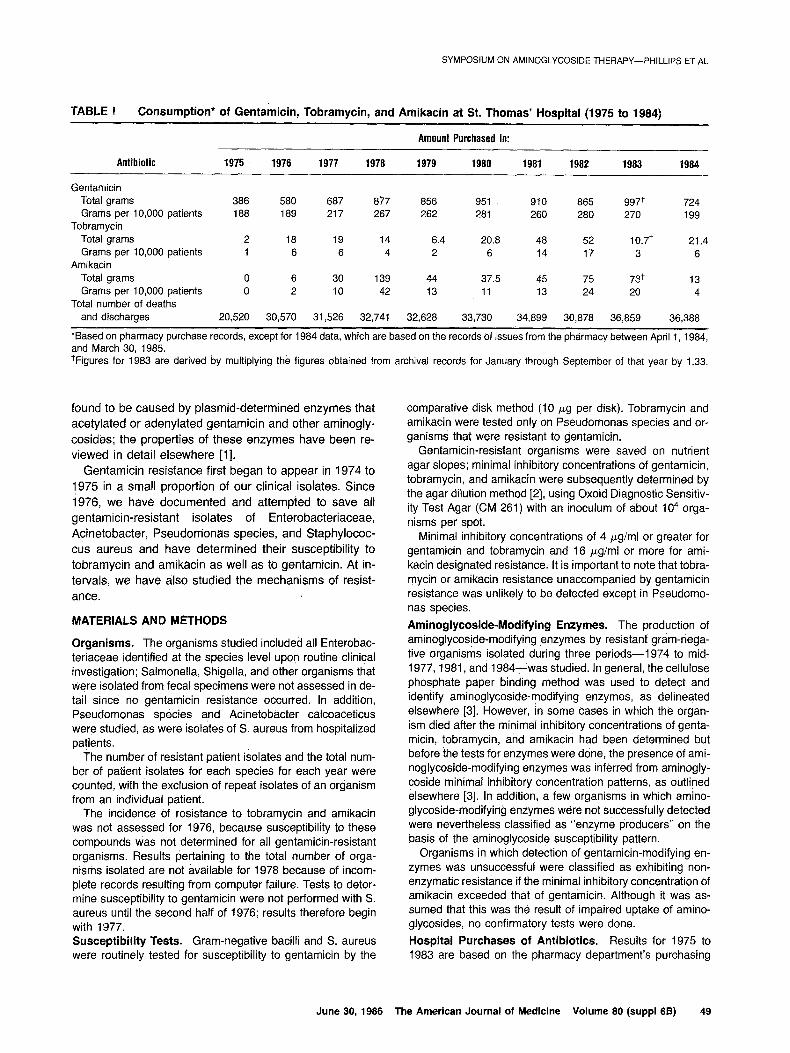
TABLE l Consumption* of Gentamicin, Tobramycin, and Amikacin at St. Thomas’ Hospital (1975 to 1984)
Amount Purchased in:
Antibiotic 1975 1976 1977 1978 1979 1980 1981 1982 1983 1984
Gentamicin Total grams 386 580 687 877 856 951 910 865 997+ 724 Grams 10,000 per patients 188 189 217 267 262 28i 260 280 270 199
Tobramycin Total grams 2 18 19 14 6.4 20.8 48 52 10.7+ 21.4 Grams 10,000 per patients 1 6 6 4 2 6 14 17 3 6
Amikacin Total grams 0 6 30 139 44 37.5 45 75 73+ 13 Grams 10,000 per patients 0 2 IO 42 13 11 13 24 20 4
Total number of deaths and discharges 20,520 30,570 31,526 32,741 32,628 33,730 34,899 30,878 36,859 36,388
*Based on pharmacy purchase records, except for 1984 data, which are based on the records of issues from the pharmacy between April 1, 1984, and March 30, 1985. +Figures for 1983 are derived by multiplying the figures obtained from archival records for January through September of that year by 1.33.
found to be caused by plasmid-determined enzymes that acetylated or adenylated gentamicin and other aminogly- cosides; the properties of these enzymes have been re- viewed in detail elsewhere [l].
comparative disk method (10 pg per disk). Tobramycin and amikacin were tested only on Pseudomonas species and or- ganisms that were resistant to gentamicin.
Gentamicin resistance first began to appear in 1974 to 1975 in a small proportion of our clinical isolates. Since 1976, we have documented and attempted to save all gentamicin-resistant isolates of Enterobacteriaceae, Acinetobacter, Pseudonionas species, and Staphylococ- cus aureus and have determined their susceptibility to tobramycin and amikacin as well as to gentamicin. At in- tervals, we have also studied the mechanisms of resist- ance.
Gentamicin-resistant organisms were saved on nutrient agar slopes; minimal inhibitory concentrations of gentamicin, tobramycin, and amikacin were subsequently determined by the agar dilution method [2], using Oxoid Diagnostic Sensitiv- ity Test Agar (CM 261) with an inoculum of about lo4 orga- nisms per spot.
Minimal inhibitory concentrations of 4 pgiml or greater for gentamicin and tobramycin and 16 pgiml or more for ami- kacin designated resistance. It is important to note that tobra- mycin or amikacin resistance unaccompanied by gentamicin resistance was unlikely to be detected except in Pseudomo- nas species.
MATERIALS AND METHODS
Organisms. The organisms studied included all Enterobac- teriaceae identified at the species level upon routine clinical investigation; Salmonella, Shigella, and other organisms that were isolated from fecal specimens were no2 assessed in de- tail since no gentamicin resistance occurred. In addition, Pseudomonas species and Acinetobacter calcoaceticus were studied, as were isolates of S. aureus from hospitalized patients.
The number of resistant patient isolates and the total num- ber of patient isolates for each species for each year were counted, with the exclusion of repeat isolates of an organism from an individual patient.
The incidence of resistance to tobramycin and amikacin was not assessed for 1976, because susceptibility to these compounds was not determined for all gentamicin-resistant organisms. Results pertaining to the total number of orga- nisms isolated are not available for 1978 because of incom- plete records resulting from computer failure. Tests to deter- mine susceptibility to gentamicin were not performed with S. aureus until the second half of 1976; results therefore begin with 1977.
Aminoglycoside-Modifying Enzymes. The production of aminoglycoside-modifying enzymes by resistant gram-nega- tive organisms isolated during three periods-1974 to mid- 1977, 1981, and 1984-was studied. In general, the cellulose phosphate paper binding method was used to detect and identify aminoglycoside-modifying enzymes, as delineated elsewhere [3]. However, in some cases in which the organ- ism died after the minimal inhibitory concentrations of genta- micin, tobramycin, and amikacin had been determined but before the tests for enzymes were done, the presence of ami- noglycoside-modifying enzymes was inferred from aminogly- coside minimal inhibitory concentration patterns, as outlined elsewhere [3]. In addition, a few organisms in which amino- glycoside-modifying enzymes w&e not successfully detected were nevertheless classified as “enzyme firoducers” on the basis of the aminoglycoside susceptibility pattern.
Organisms in which detection of gentamicin-modifying en- zymes was unsuccessful were classified as exhibiting non- enzymatic resistance if the minimal inhibitory concentration of amikacin exceeded that of gentamicin. Although it was as- sumed that this was the result of impaired uptake of amino- glycosides, no confirmatory tests were done.
Susceptibility Tests. Gram-negative bacilli and S. aureus Hospital Purchases of Antibidtics. Results for 1975 to were routinely tested for susceptibility to gentamicin by the 1983 are based on the pharmacy department’s purchasing
June 30, 1986 The American Journal of Medicine Volume 80 (suppl 6B) 49
SYMPOSIUM ON AMINOGLYCOSIDE THERAPY-PHILLIPS ET AL
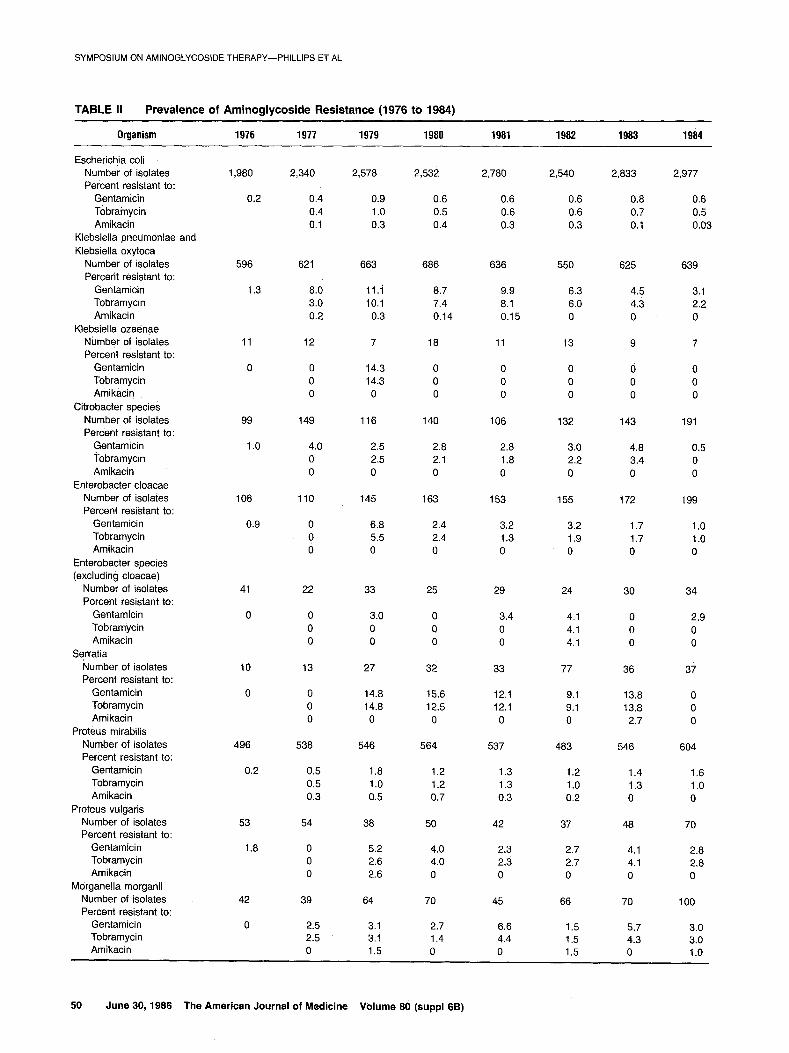
TABLE II Prevalence of Aminoglycoside Resistance (1976 to 1984)
Organism 1976 1977 1979 1980 1981 1982 1983 1984
Escherichja coli Number of isolates Percent resistant to:
Gentamicin Tobramycin Amikacin
Klebsiella pneumoniae and Klebsiella oxytoca
Number of isolates Percent resistant to:
Gentamicin Tobramycin Amikacin
Klebsiella ozadnae Number of isolates Percent resistant to:
Gentamicin Tobramycin Amikacin
Citrobacter species Number of isolates Percent resistant to:
Gentamicin Tobramycin Amikacin
Enterobacter cloacae Number of isolates Percent resistant to:
Gentamicin Tobramycin Amikacin
Enterobacter species (excluding cloacae)
Number of isolates Percent resistant to:
Gentamicin Tobramycin Amikacin
Serratia Number of isolates Percent resistant to:
Gentamicin Tobramycin Amikacin
Proteus mirabilis Number of isolates Percent resistant to:
Gentamicin Tobramycin Amikacin
Proteus vulgaris Number of isolates Percent resistant to:
Gentamicin Tobramycin Amikacin
Morganella morganii Number of isolates Percent resistant to:
Gentamicin Tobramycin Amikacin
1,980
0.2
2,340 2,578 2,532 2,780 2,540 2,833 2,977
0.4 0.9 0.6 0.6 0.6 0.8 0.6 0.4 1 .o 0.5 0.6 0.6 0.7 0.5 0.1 0.3 0.4 0.3 0.3 0.1 0.03
596 621 663 686 636 550 625 639
1.3 8.0 ii.i 8.7 9.9 6.3 4.5 3.1 3.0 10.1 7.4 8.1 6.0 4.3 2.2 0.2 0.3 0.14 0.15 0 0 0
11 12 7 18 11 13 9 7
0 0 0 0
14.3 14.3
0
0 0 0
0 0 0
0 0 0
0 0 0
0 0 0
99 149 116 140 106 132 143 191
1 .o 4.0 2.5 2.8 2.8 3.0 4.8 0.5 0 2.5 2.1 1.8 2.2 3.4 0 0 0 0 0 0 0 0
108 110 145 163 153 155 172 199
0.9 0 6.8 2.4 3.2 3.2 1.7 1.0 0 5.5 2.4 1.3 1.9 1.7 1.0 0 0 0 0 0 0 0
41 22 33 25 29 24 30 34
0 0 0 0
3.0 0 0
0 0 0
3.4 0 0
4.1 4.1 4.1
0 0 0
2.9 0 0
10 13 27 32 33 77 36 37
0 0 14.8 15.6 12.1 9.1 13.8 0 0 14.8 12.5 12.1 9.1 13.8 0 0 0 0 0 0 2.7 0
496 538 546 564 537 483 546 604
0.2 0.5 1 .a 1.2 1.3 1.2 1.4 1.6 0.5 1.0 1.2 1.3 1 .o 1.3 1.0 0.3 0.5 0.7 0.3 0.2 0 0
53
1.8
54 38 50 42 37 48 70
0 5.2 4.0 2.3 2.7 0 2.6 4.0 2.3 2.7 0 2.6 0 0 0
2.8 2.8 0
42 39 64 70
0 2.5 3.1 2.7 2.5 3.1 1.4 0 1.5 0
45
6.6 4.4 0
66
1.5 1.5 1.5
4.1 4.1 0
70
5.7 4.3 0
100
3.0 3.0 1 .o
50 June 30, 1986 The American Journal of Medicine Volume 80 (suppl 66)

SYMPOSIUM ON AMINOGLYCOSIDE THERAPY-PHILLIPS ET AL
TABLE II Prevalence of Aminoglycoside Resistance (1976 to 1984) (continued)
Organism 1976 1977 1979 1980 1981 1982 1983 1984
Providencia rettgeri Number of isolates Percent resistant to:
Gentamicin Tobramycin Amikacin
Providencia stuartii Number of isolates Percent resistant to:
Gentamicin Tobramycin Amikacin
Acinetobacter calcoaceticus Number of isolates Percent resistant to:
Gentamicin Tobramycin Amikacin
Pseudomonas aeruginosa Number of isolates Percent resistant to:
Gentamicin Tobramycin Amikacin
Staphylococcus aureus Number of isolates Percent resistant to:
Gentamicin
0 0 0 27.2 33.3 28.6 14.3 16.7 0 0 0 27.2 33.3 28.6 14.3 16.7 0 0 0 0 0 0 0 0
8 20 25 29 28 29 34 31
50 85 88 75.8 85.7 82.7 70.5 83.8 50 70 92 75.8 85.7 82.7 70.5 80.6
0 5 0 0 0 0 5.8 2.7
77
2.6
473 407 406 458 444 415
4.0 6.1 4.6 6.1 5.4 8.7 1.4 2.9 1.4 2.1 2.2 2.4 1.2 2.7 1.9 2.4 1.5 2.4
10 8 11 3 7 7 6
66 113 144 122 95 110 128
4.5 38.9 36.1 20.5 26.3 20.0 13.3 0 4.4 4.2 1.6 2.1 10.0 4.7 0 0 4.9 2.5 2.1 3.6 3.1
481
2,305 1,982 2,180 2,338 1,919
1.9 1.8 1.1 2.0 0.6
451
5.7 2.2 1.5
1,705
0.1
3.1 0.8 1.2
1,760
3.3
archives. A computer system was introduced in October 1983; for that year, the archival figures for January to Sep- tember have been multiplied by 1.33. Figures for 1984 are from the computer records.
RESULTS
Aminoglycoside Consumption. As judged by the phar- macy’s purchases, gentamicin was used much more ex- tensively than tobramycin or amikacin (Table I), which were generally given only after consultation with one of the medical staff of the microbiology department. Deaths and discharges from the St. Thomas’ group of hospitals increased by 50 percent between 1975 and 1976, when two outlying hospitals joined the main hospital. Since that time, deaths and discharges have tended to show a small annual increase. Gentamicin use increased between 1976 and 1979, relative to both the numbers of deaths and discharges and to the total number of patient isolates of Enterobacteriaceae, Pseudomonas aeruginosa, and A. calcoaceticus; these figures subsequently showed little change until 1984, when there was a reduction in use. Prevalenke and Mechanisms of Aminoglycoside Re- sistance. The prevalence of resistance to gentamicin, tobramycin, and amikacin among gram-negative bacilli is shown in Table II; the mechanisms of gentamicin resist-
ante for three periods, 1974 to mid-l 977, 1981, and 1984, are shown in Table 111.
Gentamicin resistance was extremely rare in Esche- richia coli in 1976. It had increased somewhat by 1979 but was still less than 1 percent. It remained at this level for the rest of the study period. Tobramycin resistance oc- curred at a rate similar to that of gentamicin resistance. However, amikacin resistance occurred at about one third to two thirds of the frequency of gentamicin resistance throughout most of the study and was even less common in 1983 and 1984. From 1974 to 1977, all but one of the seven tested gentamicin-resistant isolates of E. coli showed non-enzymatic resistance; this mechanism of resistance was still the most common one in 1981, al- though the incidence of resistance caused by it was ex- ceeded by the total incidence caused by enzymatic mech- anisms, mostly AAD(2”). However, in 1984, AAC(3)-II pro- duction was the most common mechanism of gentamicin resistance in E. coli.
Gentamicin-resistant strains of Klebsiella (and Acinetobacter) have caused outbreaks of cross-infection in this hospital. Although more common than in E. coli, gentamicin resistance in Klebsiella pneumoniae (from which we did not distinguish Klebsiella oxytoca) was nev- ertheless rare in 1976. However, it became much more frequent in 1977, when a single strain was responsible for
June 30, 1988 The American Journal of Medicine Volume 80 (suppl 8B) 51

SYMPOSIUM ON AMINOGLYCOSJDE THERAPY-PHILLIPS ET AL
TABLE III Mechanisms of Aminoglycoside Resistance
Mechanism of Resistance (number of strains)
Organism Year AAC(B)-I
Escherichia coli
Klebsiella pneumoniae and Klebsiella oxytoca
Citrobacter species
Enterobacter cloacae
Enterobacter aerogenes
Serratia species
Proteus mirabilis
Proteus vulgaris
Morganella morganii
Providencia rettgeri
Providencia stuartii
Acinetobacter caicoaceticus
Pseudomonas aeruginosa
Pseudomonas species
1974 to 1977 1 0 0 0 0 6 1981 1 2 0 0 4 7 1984 2 8 0 0 1 5
1974 to 1977 10 0 0 0 1 4 1981 10 29 0 0 19 2 1984 5 8 0 0 3 0
1974 to 1977 3 0 0 0 0 1 1981 ? 1 0 0 1 0 1984 1 0 0 0 1 0
1974 to 1977 5 0 0 0 0 0 1981 3 1 0 0 1 0 1984 0 0 0 0 1 0
1974 to 1977 0 0 0 0 0 0 1981 1 0 0 0 0 0 1984 1 0 0 0 0 0
1974 ‘to 1977 3 0 0 2 0 0 1981 Q 0 0 3 1 0 1984 0 0 0 0 0 0
1974 to 1977 1 0 0 0 0 1 1981 0 0 0 0 4 2 1984 1 1 0 0 0 2
1974 to 1977 0 0 0 0 0 0 1981 0 0 0 0 0 0 1984 0 0 0 0 1 1
1974 to 1977 0 0 0 0 0 0 1981 1 1 0 0 1 0 1984 2 0 0 0 0 1
1974 to 1977 0 0 0 0 0 0 1981 0 0 1 0 0 0 1984 0 0 0 0 0 0
1974 to 1977 0 0 17 0 0 0 1981 0 0 24 1 0 0 1984 0 0 26 0 0 0
1974 to 1977 0 0 0 5 0 0 1981 IS 0 0 0 0 1 1984 10 0 0 2 2 0
1974 to 1977 4 0 0 3 2 24 1981 6 0 0 2 4 9 1984 11 0 0 0 1 10
1974 to 1977 1 0 0 1 0 17 1981 0 0 0 0 0 7 1984 1 0 0 1 1 12
AAC(B)-II AAC(2’) AAC(6’) AAD Non-Enzymatic
For organisms isolated in 1984, AAC(3)-I production was inferred from minimal inhibitory concentration profiles (rather than detected) for two isolate; of Acinetobacter calcoaceticus and one of Pseudomonas aeruginosa; similarly inferred were AAD(2”) production for one isolate of Proteus vulgaris and non-enzymatic resistance for three isolates of Pseudomonas species and one of P. aeruginosa.
an outbreak of infection and colonization that reached a peak of 11 percent of isolates in 1979; by 1984, this level h&d slowly declined to 3.1 percent. Most of the genta- micin-resistant Klebsielleae remained susceptible to to- bramycin until 1979, after which gentamicin resistance was generally accompanied by tobramycin resistance. Amikacin-resistant Klebsielleae were exceedingly rare throughout the study period; none was isolated after 1981. We isoiated only one gentamicin- and tobramycin-resis- tant strain of Klebsiella ozaenae. AAC(S)-I production, the only mechanism to confer resistance in Enterobacteria-
ceae, to gentamicin but not tobramycin, accounted for the bulk of the gentamicin resistance in Klebsiella in 1974 to 1977. By 1981, however, the picture had changed dra- matically; AAC(3)-II was the most commonly detected enzyme, followed in order by AAD(2”) and AAC(3)-I. Gen- tamic/n-resistant Klebsielleae were rarer in 1984 than in 1981; at this time,‘AAC(3)-I was detected only slightly less frequently than was AAC(3)-II, followed by AAD(2”).
Enterobacter, mostly Enterobacter cloacae, and Citro- batter are not particularly common organisms in this hos- pital, each accountirig for only 100 to 200 isolates per
52 June 30, 1988 The American Journal of Medicine Volume 80 (suppl 68)

year. Throughout most of the study, about 2 to 5 percent of these isolates were resistant to gentamicin; however, the level of resistant isolates dropped to 0.5 to 1 percent in 1984. Most of the gentamicin-resistant strains have also been resistant to tobramycin, at least since 1979, but none was resistant to amikacin. From 1974 to 1977, all these resistant strains produced AAC(3)-I except for one Citrobacter isolate, which exhibited non-enzymatic resist- ance. There was some production of AAC(3)-II and AAD(2”) in 1981, but by 1984, there were only four genta- micin-resistant strains available for study.
tamicin. No gentamicin-resistant strains of Hafnia alvei, Salmonella, or Shigella were isolated, and no Yersinia species were isolated.
Isolates of Serratia were rare. Until recently, our meth- ods were unable to speciate organisms reliably in this genus; we have, therefore, placed all the species into one group. The increase in the number of patient isolates for 1982 is accounted for by an outbreak of infection with an aminoglycoside-susceptible strain in the premature baby unit. Gentamicin resistance was not observed until 1979, after which 9 to 16 percent of isolates were resistant to both gentamicin and tobramycin (except in 1984, when no resistant strains were isolated). We observed only one amikacin-resistant strain during the entire study. AAC(6’) was detected in many of the gentamicin-resistant strains, although in 1981, several strains were observed to pro- duce AAC(3)-I.
Gentamicin resistance was uncommon in A. calco- aceticus until 1978 (results are not shown for that year), when there was an outbreak of infection and colonization with a resistant strain of the anitratus variety that pro- duced AAC(3)-I and, consequently, retained susceptibility to tobramycin and amikacin. Subsequently, there was a gradual decline in the prevalence of gentamicin-resistant Acinetobacter from 38.9 percent of isolates in 1979 to 13.3 percent of isolates in 1984. AAC(3)-I production re- mained the predominant mechanism of resistance; thus, resistance to tobramycin or amikacin was rare. Five iso- lates from 1974 to 1977 produced AAC(6’); they were re- sistant to tobramycin and amikacin but susceptible to gen- tamicin, and were detected only because of a study of the in vitro activity of aminoglycosides on clinical isolates that was being conducted at that time [2].
Gentamicin resistance was more common in Proteus mirabilis than in E. coli, occurring in 1 to 2 percent of iso- lates. Tobramycin resistance was observed somewhat less frequently and amikacin resistance was even rarer. Proteus vulgaris was isolated only about one-tenth as often as P. mirabilis, but gentamicin and tobramycin re- sistance occurred in a somewhat higher proportion of strains; amikacin resistance was detected only once in this species. Most of the resistance in Proteus was caused by AAD(2”).
The prevalence of resistance to gentamicin fluctuated between 1.5 and 6.6 percent for Morganella morganii dur- ing the study period. Most of the gentamicin-resistant iso- lates were also resistant to tobramycin, but few were re- sistant to amikacin. Aminoglycoside resistance was attrib- uted to AAC(B)-I, AAC(3)-II, AAD(2”), and, in one case, a non-enzymatic mechanism.
For P. aeruginosa isolates, the prevalence of resistance to gentamicin ranged from 3 to 9 percent throughout the course of the study, and has declined since 1982. Resist- ance to tobramycin or amikacin was more unusual and was never found in more than 3 percent of isolates. From 1974 to 1977, most of the resistance was due to non- enzymatic mechanisms, with smaller contributions attrib- utable to production of AAC(3)-I, AAC(G’), or AAD(2”). In 1981, however, although non-enzymatic resistance was still the most common mechanism, it had lost its absolute majority; by 1984, AAC(3)-I production had become the most common means of resistance. In contrast to the non- enzymatic resistance observed in Enterobacteriaceae, such resistance among strains of P. aeruginosa often was successfully countered by tobramycin; a typical minimal inhibitory concentration pattern was 4 pg/ml of gentami- tin, 1 pgiml of tobramycin, and 16 to 32 pg/ml of ami- kacin.
Providencia stuartii showed a high incidence of resist- ance to gentamicin and tobramycin throughout the study, but amikacin resistance was rare. Providencia rettgeri, which was resistant to gentamicin less often than was P. stuartii, was isolated less frequently; Providencia al- califaciens was rarer still. The only tested gentamicin- resistant isolate of P. rettgeri produced AAC(2’), as did all the isolates of P. stuartii-even those with gentamicin minimal inhibitory concentrations as Jaw as 2 pg/ml. We suspect that one isolate of P. stuartii, from 1981, produced AAC(6’) in addition to AAC(2’).
Gentamicin-resistant isolates of Pseudomonas species other than P. aeruginosa generally showed non-enzy- matic resistance. The resistant isolates generally be- longed to species such as Pseudomonas acidovorans, Pseudomonas cepacia, Pseudomonas maltophilia, or Pseudomonas pseudoalcaligenes; these were almost always aminoglycoside-resistant. Gentamicin-modifying enzymes were detected in a few strains: AAC(3)-I was produced by an isolate of P. pseudoalcaligenes between 1974 and 1977 and by an isolate of P. maltophilia in 1984; AAC(6’) was produced by isolates of P. maltophilia be- tween 1974 and 1977 and in 1984; and AAD(2”) was pro- duced by an isolate of P. maltophilia in 1984. However, the minimal inhibitory concentration patterns suggested that these strains generally also possessed the non- enzymatic mechanism of aminoglycoside resistance.
The few organisms that we were unable to identify more We first isolated a gentamicin-resistant strain of precisely than as “coliforms” were all susceptible to gen- S. aureus in 1976. From 1977 to 1981, 1 to 2 percent of
SYMPOSIUM ON AMINOGLYCOSIDE THERAPY-PHILLIPS ET AL
June 30, 1988 The American Journal of Medicine Volume 80 (suppl 8B) 53

SYMPOSIUM ON AMINOGLYCOSIDE THERAPY-PHILLIPS ET AL
the isolates from hospitalized patients were resistant (Table II). The frequency of their isolation then declined until 1984, when an outbreak of cross-infection with a mul- tiply resistant strain accounted for 3.1 percent of the 3.3 percent resistance rate. Twenty-two strains isolated from 1976 to 1977 were tested for production of aminoglyco- side-modifying enzymes; the gentamicin resistance was attributable to production of APH(2”) and AAC(6’) in 17 strains and to a non-enzymatic mechanism in five strains. Strains that produced APH(2”) and AAC(6’) were resistant to both gentamicin and tobramycin and often showed some reduction in susceptibility to amikacin-in some cases, sufficient to render the organism resistant to the compound. As was the case with resistance in gram- negative bacilli, non-enzymatic resistance was associated with greater resistance to amikacin than to gentamicin or tobramycin.
Except in the case of S. aureus, production of more than one gentamicin-modifying enzyme In any individual isolate was rarely encountered. AAC(3)-I and AAC(2”) were produced by two isolates of E. coli and by one isolate of Citrobacter freundii; AAC(6’) and AAD(2”) were pro- duced by an isolate of P. aeruginosa. We suspected that two isolates of Serratia produced AAC(6’) and AAC(3)-I and that one isolate of P. stuartii produced AAC(2’) and AAC(6’); however, some uncertainty exists when two acetyltransferases are involved. Furthermore, there was the possibility that some strains produced two acetyl- transferases, one of which completely masked the other (AAC(3)-I together with AAC(2’) or AAC(3)-II, for exam- ple).
COMMENTS
In some cases in which the mechanisms of resistance have been studied in a single hospital, one or two en- zymes have been found to account for most of the genta- micin resistance. For example, AAC(3)-I and an enzyme designated as AAC(S)-V accounted for most of the resist- ance in Saragoza, Spain [4]; AAC(S)-I and AAD(2”) were the predominant mechanisms of resistance in Sydney, Australia [5]; in a Boston hospital, AAD(2”) production was predominant [6]. Our strains showed a little more variety: if Providencia, which shall be discussed later, is excluded, three enzymes-AAC(3)-I, AAC(3)-II, and AAD(2”)- accounted for most of the resistance, with smaller contri- butions by AAC(6’) and non-enzymatic mechanisms.
In a large survey of resistant organisms [7] (most from hospitals within the United States) conducted by Bristol Laboratories, AAD(2”) was the most commonly produced enzyme; AAC(3)-I was next in order, followed by other forms of AAC(3). However, Serratia, which accounted for a much higher proportion of the strains in this survey than it did in our study, produced mostly AAC(G’), with or with- out another enzyme. Non-enzymatic resistance was com-
mon in E. coli, P. aeruginosa, and other non-fermenters of glucose. Similar conclusions were drawn by Miller and colleagues [8], although their determination of mecha- nisms of resistance was based on susceptibility patterns. Our findings are consistent with the findings of these stud- ies insofar as non-enzymatic resistance was determined to be common in E. coli and P. aeruginosa and AAC(6’) was determined to be common in the few isolates of Ser- ratia; however, we found AAC(B)-I and AAC(S)-II to be more prevalent than AAD(2”). The difference in the pro- portions of resistance attributable to AAC(3) and AAD(2”) may reflect geographic differences; a later study [9], in- volving institutions in the United States, Chile, Japan, Korea, and Formosa, inferred AAD(2”) production in 82 percent of the strains from the United States and 46 per- cent of those from the Far East, but in none from Chile. In contrast, although AAC(3) production was detected in only 2 percent of the strains from the Far East and 12 percent of the strains from the United States, it was ob- sewed in 96 percent of the strains from Chile.
In only a few groups of organisms were sufficient num- bers of gentamicin-resistant strains isolated to warrant a consideration of the changes in mechanisms of resistance that occurred during the study. However, an analysis of these cases seemed to indicate several different ways in which aminoglycoside resistance can occur and can spread in a hospital.
Virtually all the resistance in P. rettgeri and P. stuartii was attributable to production of AAC(2’); furthermore, this enzyme was not found in any of the other strains. This association between AAC(2’) and Providencia has been noted previously [l]. It appears that, in contrast to the pro- duction of most other aminoglycoside-modifying en- zymes, AAC(2’) production is chromosomally determined [I 0] and thus this microbe seems to be an example of an inherently gentamicin-resistant organism.
In contrast, in our hospital, Acinetobacter was usually gentamicin-susceptible; thus, we assumed that the genta- micin resistance resulted from outbreaks of cross-infec- tion and colonization with a single, AAC(3)-l-producing strain [l 11; however, there cannot be complete certainty in the absence of a reliable typing method for this organism.
Gentamicin resistance in Klebsiella was initially associ- ated with AAC(B)-I; it was subsequently primarily associ- ated with AAC(3)-II and AAD(2”). In the first part of the study period (1974 to 1977), most of the gentamicin-resis- tant Klebsielleae were from an outbreak of a strain of cap- sular type 16 [12]; in the two later parts of the period, there were no major incidents of cross-infection, perhaps be- cause of more effective implementation of the infection control policy. There is no obvious reason why AAC(3)-II should have risen to such sudden prominence and then diminished, both proportionately and in absolute terms. Nevertheless, there is some evidence in this hospital that plasmid-determined mechanisms of gentamicin resist-
54 June 30, 1988 The American Journal of Medicine Volume 80 (suppl 8B)

SYMPOSIUM ON AMINOGLYCOSIDE THERAPY-PHILLIPS ET AL
ante tended to appear first in Klebsiella and then shift to other Enterobacteriaceae, since AAC(3)-I, AAC(3)4, and AAD(2”) all appeared first in Klebsiella and were found later in other genera. This exchange has been observed in individual patients in this hospital. Furthermore, Knight and Casewell [13] reported the apparent in vivo transfer of gentamicin resistance from K. pneumoniae to E. coli, E. cloacae, and C. freundii. It will be interesting to discover whether AAC(3)-II production, now the most common re- sistance mechanism in E. coli, diminishes in this species during the next few years, as it recently has in Klebsiella.
In contrast to what has been observed with Klebsiella and E. coli, there has not been an obvious change in the gentamicin-modifying enzymes detected in P. aeruginosa. However, there does seem to have been a shift from non-
enzymatic resistance towards production of modifying enzymes.
The question of why aminoglycoside resistance has become rarer in our hospital since the early 1980s re- mains unanswered. It would be pleasing to propose that this was the result of effective antibiotic and infection con- trol policies. However, it may reflect the result of a de- crease in aminoglycoside selective pressure that accom- panied an increase in the use of cephalosporins for the treatment of infections with gram-negative organisms. It should be remembered that the substantial problems en- countered with multiply resistant strains of S. aureus in the 1950s and 1960s have recurred in recent years. One can- not help wondering whether the gram-negative organisms will be involved in a similar scenario.
REFERENCES
1. Phillips I, Shannon K: Aminoglycoside resistance. Br Med Bull 1984; 40: 28-35.
2. Phillips I, Eykyn S, King A, et al: The in vitro activity of nine aminoglycosides and spectinomycin on clinical isolates of common gram-negative bacteria. J Antimicrob Chemother 1977; 3: 403-410.
3. Shannon KP, Phillips I: Detection of aminoglycoside-modifying strains of bacteria. In: Russell AD, Quesnel LB, eds. Antibiot- ics. Society for Applied Bacteriology technical series no. 18. London: Academic Press, 1983; 183-l 98.
4. Gomez-Lus R, Rubio-Calve MC, Navarro M, et al: Plasmid- determined resistance to aminocyclitols in enterobacteria. Current chemotherapy and immunotherapy. In: Periti P, Grassi GG, eds. Proceedings of the 19th International Con- gress of Chemotherapy. Washington, DC.: American Society of Microbiology, 1982; 218-219.
5. Groot Obbink DJ, George AM, Coombe RG: Aminoglycoside- modifying enzymes associated with hospital isolates of gram- negative rods. J Antimicrob Chemother 1983; 11: 525-533.
6. O’Brien TF, Ross DG, Guzman MA, et al: Dissemination of an antibiotic-resistance plasmid in hospital patient flora. An- timicrob Agents Chemother 1980; 17: 537-543.
7. Price KE, Kresel PA, Farchione LA, et al: Epidemiological stud- ies of aminoglycoside resistance in the U.S.A. J Antimicrob Chemother 1981; 8 (suppl A): 89-105.
8. Miller GH, Sabatelli FJ, Hare RS, Waitz JA: Survey of aminogly- coside resistance patterns. Dev Ind Microbial 1980; 21: 91- 104.
9. Shimizu K, Kumada T, Hsieh W-C, et al: Comparison of amino- giycoside resistance patterns in Japan, Formosa, and Korea, Chile, and the United States. Antimicrob Agents Chemother 1985; 28: 282-288.
10. Hawkey PM: Providencia stuartii: a review of a multiply antibi- otic-resistant bacterium. J Antimicrob Chemother 1984; 13: 209-226.
11. French GL, Casewell MW, Roncoroni AJ, et al: A hospital out- break of antibiotic-resistant Acinetobacter anitratus: epidemi- ology and control. J Hosp Infect 1980; 1: 125-131.
12. Casewell MW, Dalton MT, Webster M, Phillips I: Gentamicin- resistant Klebsiella aerogenes in a urological ward. Lancet 1977; II: 444-446.
13. Knight S, Casewell M: Dissemination of resistance plasmids among gentamicin-resistant enterobacteria from hospital pa- tients Br Med J 1981; 283: 755-756.
June 30, 1986 The American Journal of Medicine Volume 80 (suppl66) 55


















