
(EMC) dengan lipase Aspergillus niger dan aminopeptidase ...
Upload
brendan-oconnorCategory
view
217download
2
Eur. J. Biochem. 154, 329 -335 (1986) (0 FEBS 1986
Post-proline dipeptidyl-aminopeptidase from synaptosomal membranes of guinea-pig brain A possible role for this activity in the hydrolysis of His-ProNH2, arising from the action of synaptosomal membrane pyroglutamate aminopeptidase on thyroliberin
Brendan O’CONNOR ’ and Gerard O’CUINN
Department of Biochemistry, University College, Galway Department of Life Sciences, Regional Technical College, Galway
(Received June 18/September 3, 1985) - EJB 85 0662
In this paper we report that while 55% of the total post-proline dipeptidyl-aminopeptidase activity in guinea- pig brain is associated with the soluble fraction of the cells, the remaining activity is widely distributed throughout the particulate fractions. A significant portion of this particulate activity is, however, associated with a synaptosomal membrane fraction. The specific activity of this enzyme rose as the synaptosomal membrane fraction was prepared from a synaptosomal fraction and had previously risen at the synaptosomal fraction was prepared from a postmitochondrial pellet. The synaptosomal membrane post-proline dipeptidyl-aminopeptidase was released from the membrane by treatment with Triton X-3 00 and partially purified by chromatography on Sephadex G-200. By contrast with the soluble enzyme the partially purified solubilised synaptosomal membrane post-proline dipeptidyl-aminopeptidase was not inhibited by 1 .0 mM p-chloromercuribenzoate, 1 .0 mM N-ethylmaleimide or 0.5 mM puromycin but was inhibited by 0.5 mM bacitracin. The partially purified solubilised enzyme was capable of releasing His-Pro from His-Pro-Val, His-Pro-Leu, His-Pro-Phe and His-Pro-Tyr and of releasing Gly-Pro from Gly-Pro-Ala but could not release Arg-Pro from Arg-Pro-Pro or from Arg-Pro-Pro-Gly- Phe-Ser-Pro-Phe-Arg (bradykinin). It was also unable to release Pro-Pro from Pro-Pro-Gly or Glp-Pro from Glp-Pro-Ser-Lys-Asp-Ala-Phe-Ile-Gly-Leu-MetNH2 (eledoisin). Using [Pr~-~H]thyroliberin we show that the membrane-bound enzyme converts His-ProNH,, produced by the action of the synaptosomal membrane pyroglutamate aminopeptidase, to His-Pro thus competing with the spontaneous cyclisation of His-ProNH, to His-Pro diketopiperazine. Purified preparations of synaptosomal membrane pyroglutamate aminopeptidase were used to generate His-ProNHz, which could then be converted to His-Pro by the presence of the partially purified synaptosomal membrane post-proline dipeptidyl-aminopeptidase. This preparation was free of contaminating post-proline cleaving endopeptidase, carboxypeptidase P, aminopeptidase P, prolyl carboxypeptidase or proline dipeptidase.
In addition to its well-defined neuroendocrine role as a releasing hormone, thyroliberin (Glp-His-ProNH,) has been suggested to play a neurotransmitter or neuromodulator role in brain [l]. This suggestion has been supported by the recent finding of a thyroliberin-hydrolysing pyroglutamate ainino- peptidase of narrow specificity associated with synaptosomal
Correspondence to G. O’Cuinn, Department of Life Sciences, Regional Technical College, Galway, Ireland
Abbreviu/ions. Glp, pyroglutamic acid, 5-pyrrolidone-2-carbox- ylic acid; Cbz-Gly-Pro-NH-Mec, N-benzyloxycarbonylglycylprolyl- 7-amido-4-methylcoumarin; Gly-Pro-NH-Mec, glycylprolyl-7- amido-4-methylcoumarin.
Enzjmes (IUPAC Recommendations. 1984). Post-proline cleav- ing cndopeptidase (EC 3.4.22.26); post-proline dipeptidyl-amino- pcptidase (EC 3.4.14. -); pyroglutamate aminopeptidase (EC 3.4.19.3); aminopeptidase P (EC 3.4.1 1.9); proline iminopeptidase (EC 3.4.1 1 . S ) ; carboxypeptidase P (EC 3.4. - . -); prolyl car- boxypeptidase (EC 3.4.16.2); prolyl dipeptidase, prolinase (EC 3.4.13.8); proline dipeptidase (EC 3.4.1 3.9); 5’-ribonucleotidase (EC 3.1.3.5); succinate dehydrogenase (EC 1.3.99.1); acid phosphatase (EC 3.1.3.2; NADH: cytochrome c reductase (EC 1.6.99.3); ouabain N a + / K + ATPasc (EC 3.6.1.3).
membranes [2]. This activity inactivates thyroliberin by hydrolysing it to pyroglutamate and His-ProNH,. This latter metabolite cyclises spontaneously and non-enzymatically to His-Pro diketopiperazine, a compound to which is attributed endocrine activity [3] as well as the ability to enhance excitato- ry responses of cortical neurones to acetylcholine [4]. The conversion by of synaptosomal membrane pyroglutamate aminopeptidase of a neuronally active peptide, thyroliberin, to another neuronally active peptide, His-Pro-diketopipera- zine, represents an instance of biotransformation of a neuro- peptide by enzyme activity [ S ] . The presence on synaptosomal membranes of a post-proline dipeptidyl-aminopeptidase capable of deamidating His-ProNH, would be of importance, therefore, in regulating the production of His-Pro diketopiperazine by competing with the cyclisation of His- ProNHz.
Post-proline dipeptidyl-aminopeptidase was first de- scribed in the microsomal fraction of rat liver [6]. Sub- sequently this activity was shown to be present in microvillar membranes of porcine small intestine [7] and kidney [S, 91 and in the microsomal fraction of porcine liver [lo], rat liver [ll], lamb kidney [12, 131 and human submaxillary gland [14, 151.

330
Soluble post-proline dipeptidyl-aminopeptidase has been de- scribed in human serum and in cerebrospinal fluid [16] and a soluble cellular post-proline dipeptidyl-amino-peptidase has been observed in the 100000 x g supernatant fraction of rat brain [17]. Recent reports have indicated that soluble post- proline dipeptidyl-aminopeptidase could participate in the hydrolysis of thyroliberin in vitro to its constituent amino acids in concert with soluble pyroglutamate aminopeptidase, soluble post-proline cleaving endopeptidase and soluble pro- line dipeptidase activities of rat adenohypophysis [18] and of guinea-pig brain [19].
Here we report on the subcellular distribution of post- proline-dipeptidyl-aminopeptidase in guinea-pig brain and a possible role of synaptosomal membrane post-proline dipeptidyl-aminopeptidase in the metabolism of His-ProNH, produced by synaptosomal membrane pyroglutamate aminopeptidase.
MATERIALS AND METHODS
Materials
Post-proline cleaving endopeptidase. Post-proline cleaving endopeptidase was measured by a inodification of the method. of Yoshimoto et al. [21]. 10-p1 samples were incubated with 490 p1 Cbz-Gly-Pro-NH-Mec (0.2 mM in 100 mM potassiur. phosphate pH 7.5 containing 2 mM dithiothreitol, 2 mM EDTA and 5% dimethylformamide) at 37 T for 20 min. The reaction was terminated by addition of 1 in1 1.5 M acetic acid and 7-amino-4-methylcoumarin released was determined as above.
Proline dipeptidase. Proline dipeptidase activity was deter- mined using His-Pro as substrate 1221.
Prolyl carhoxypeptidase and carhoxypeptidase P. Proly'. carboxypeptidase and carboxypeptidase P activities were mea- sured by incubating 50 p1 sample with 450 p1 Cbz-Pro-Phc (2 mM in 75 mM sodium borate pH 7.4 containing 5% dimethylformamide) for 20 min at 30, C and the phenyl- alanine released was measured by the L-amino acidlper- oxidaselp-hydroxyphenylacetic acid system of Nicholson and Peters [23].
Aminopeptidase P. Aminopeptidase P activity was deter- mined by incubating 50 p1 sample with 450 p1 His-Pro-Va' (2 mM in 75 mM sodium borate pH 7.4) for 10 min at 30 C
Nicolson and Peters [231. Prolinusr and prolyl iminopeptidase. Prolinase and proly:
iminopeptidase activities were jointly assayed by incubating
uL-Dithiothreitol, 2-iodoacetamide, p-chloromercuriben-
oline-5-sulphonic acid, benzamidine, bacitracin (B01 25), pu- romycin, phenylmethylsulphonyl fluoride, ninhydrin, bovine Serum albumin, cytochrolne (type V, bovine heart) NADH
zoate, N-ethylmaleimide, 1 ,I O-phenanthroline, g-hydroxyquin- and the histidine released was determined by the method Of
(disodium salt) rotenone, ~-amino-acid oxidase (type VI, with 450 P1 Pro-NH-Mec (0.2 mM in 75 mM Crotalus atrox), peroxidase (type 11, horseradish), p-hydroxy- at 37 and the 7-amino-
phosphate (disodium 2-~-iodophenol)-3-(p-nitrophe- Ouahuin-sensitive N a f / K f ATPase. Ouabain-sensitive
50 P1 'Odium borate pH 7.5) for 30
phenylacetic acid, 7-amino-4-methylcoumarin, adenosine tri-
nyl)-5-phenyltetrazolium chloride, thyroliberin, pyroglutamic N a + / K + acid, proline, prolineamide, histidine and leucine were tained from Sigma Chemical Co. (Poole, Dorset, England); EDTA, ~~i~~~ X-1 00, p p o , popop, dipotassium hydrogen phophate, potassium dihydrogen phosphate and Folin- Ciocalteau phenol reagent were obtained from BDH (Poole, phate was by the method King [251. D ~ ~ ~ ~ ~ , England); Sephadex G-200 and blue dextran were obtained from Pharmacia Fine Chemicals AB (Uppsala, Sweden); Whatman no. 1 and no. 3 chromatography paper
H ~ ~ - P ~ ~ diketopiperazinc, ~ l ~ - ~ i ~ and G ~ ~ - H ~ ~ - T ~ ~ , were from U,C,B, ~ i ~ ~ ~ ~ d ~ ~ ~ ~ ( ~ ~ ~ ~ ~ ~ l ~ , ~ ~ l ~ i ~ ~ ) ; acid thyro- liberin, luliberin, eledoisin, Cbz-Gly-Pro-NH-Mec, Gly-Pro- N H - M ~ ~ , p r o - ~ ~ - ~ e c , ~ i ~ - p ~ ~ , ~ i ~ - p ~ ~ - ~ ~ l , ~ i ~ - p ~ ~ - ~ ~ ~ , His-Pro-Phe, His-Pro-Leu, Gly-Pro-Ala, Pro-Pro-Gly, Pro-Pro, bradykinin and Cbz-Pro-Phe were obtained from Bachem Feincheinikalien (Bubendorf, Switzerland); [Pro- 2,3,4,5-'HH]thyroliberin (102 Ci/mmol) was obtained from New England Nuclear (Dreieich, FRG) guinea-pigs (Dun- can-Hartley strain) were obtained from Biological Laborato- ries Ballina Ltd (Ballina, Ireland).
4-methylcoumarin released was measured as described above.
was measured by the method of et al. [241 and liberated phosphate was estimated by the method
5'-Rihonucleotidasr. This enzyme was measured by the method of Michell and Hawthorne [26] and released phos-
Succinate &hydrogenuse. Succinate dehydrogenase was assayed according to the method of Pennington 1271 as modi- fied by Porteous and [281.
ductase activity was assayed according to the method of' Takesue and Omura 1291 as modified by Jarasch and Franke 13'].
Acid phosphutase. Acid phosphatase activity was assayed according to the method described by Beaufay et al. [3 13.
S~~hcell~~lar,fi.nc.tion~~tion procedures
Freshly isolated guinea-pig brains were homogenised in four volumes of ice-cold sucrose containing 100 mM imidazole/HCl pH 7.4 using a Potter-Elvehjem homogeniser of clearance 0.075-0.125 mm. The tissue was disrupted by
King [251.
was obtained from whatman (Maidstone, ~ ~ ~ t , ~ ~ ~ l ~ ~ d ) ; N A D H : cytochrome C reductuse. NADH cytochroine C re-
Determination of enyzme activities
Post-proline dip~ptid!~l-arninOpeptiduse. Post-proline di- peptidyl-aminopeptidase was measured by a modification of the method of Kato et al. 1201. 50 pl sample was incubated with 450 p1 Gly-Pro-NH-Mec (0.2 mM in 100 mM potas- sium phosphate, pH 7.5) for 30 min at 37 'C. The reaction was terminated by addition of 1 ml 1.5 M acetic acid and the 7-amino-4 inethylcoumarin released was measured in a Perkin-Elmer flouresccnce spectrometer (model MPF-3) using excitation and emission wavelengths of 370 nm and 440 nm respectively. Non-enzymatic degradation of Gly-Pro-NH- Mec was accounted for by control incubations.
twelve up-and-down strokes of the pestle. The subsequent subcellular fractionation steps were based on the methods of Marchbanks 1321 and of Whittaker and Barker 1331. Aliquots of each Craction generated were removed and stored at - 20 C until further assay.
The crude homogenate was centrifuged at 1000 x g for 10 min. The pellet obtained was washed with 0.32 M sucrose containing 100 mM imidazole/HCl pH 7.4. Following re- centrifugation this pellet was resuspended in 0.32 M sucrose containing 100 mM imidazole/HCl pH 7.4 (fraction PI ) while the supernatants from both centrifugations were combined (fraction S,) and centrifuged at 30000 x g for 30 min. After the pellet was washed as before, i t was resuspended in 0.32 M

331
sucrose containing 100 mM imidazole/HCl pH 7.4 (frac- tion P2) while the supernatants from both centrifugations were combined (fraction S,) and centrifuged at 100000 x g for 60 min. The pellet was recovered, washed with 0.32 M sucrose containing 100 mM imidazole/HCl pH 7.4, and recentrifuged at 100000 x g for 60 min. The pellet was recovered and re- suspended in 0.32 M sucrose containing 100 mM imidazole/ HCI pH 7.4 (fraction P3) while the supernatants from both centrifugations were combined (fraction S3).
Aliquots of fraction P2 were placed on discontinuous gradients consisting of 9 ml each of 1.6 M, 1.2 M and 0.8 M sucrose. Following centrifugation at 135 000 x g for 75 min three bands of material were observed on the gradient, one on top of 0.8 M sucrose, one in the 1.2 M sucrose band and one in the 1.6 M sucrose band. Each fraction was diluted with equal volumes of water, centrifuged at 100000 x g for 60 rnin and resuspended in 0.32 M sucrose containing 100 mM imidazole/HCl pH 7.4 to give fi-actions A, B and C respective- ly. In addition some material was found to have travelled through the 1.6 M sucrose and to have pelleted at the bottom of the tube. This material was resuspended in 0.32 M sucrose containing 100 mM imidazole/HCl pH 7.4 (fraction D).
Preparat ion q f synap tosonml membrane, frac tion
Fraction B (prepared as above) was pelleted by centrifuga- tion and resuspended in water, subjected to gentle hand ho- mogenisation and placed over a second discontinuous sucrose gradient consisting of 8 ml 1.6 M sucrose, 8 ml 1.2 M sucrose, 6 ml 0.6 M sucrose and 5 ml 0.5 M sucrose. Following cen- trifugation at 135000 x g for 75 min, four fractions were drawn off from the top of the gradient K (2.5 ml), L (9 ml), M (8 ml) and N (8 ml). The particulate matter in fractions L, M and N was pelleted by centrifugation at lOO00Oxg for 60 min and resuspended in 8 ml of either 0.32 M sucrose (for enzyme marker studies) or 100 mM potassium phosphate pH 7.4.).
Release of post-proline d~eptidyl-aminopeptida,se fiom synaptosomal membranes by rreatment with Triton X-100
To fraction M, resuspended in 100 mM potassium phosphate pH 7.4, was added an equal volume of 2 M NaCl in 100 mM potassium phosphate pH 7.4 and the two were mixed gently together by repeated inversion for 10 min. Then the sample was centrifuged at I00000 x g for 70 min. The pellet was resuspended in 100 mM potassium phosphate pH 7.4 containing 0.2 M NaCl and to this suspension was added an equal volume of 2% Triton X-I00 in 100mM potassium phosphate pH 7.4. The suspension was subjected to gentle hand homogenisation on ice for 5 min then centrifuged at 100000 x g for 70 min and the pellet was re- suspended in 100mM potassium phosphate pH 7.4 containing 0.1 M NaCl. Both supernatant and resuspended pellet were retained for determination of post-proline dipeptidyl-aminopeptidase.
X-100. The applied sample was eluted from the column at a flow rate of 5.4ml h-'. 1-ml fractions were collected and assayed for post-proline dipeptidyl-aminopeptidase activity as described above.
The. CffiJct of inhibitors on Triton-X- 100-solubilisedpost-proline dipeptidyl-arviinopeptiduse activity
50-kd aliquots of enzyme sample were incubated at 37°C for 15 min with 50 p1 of each inhibitor sample to be tested, 400 p1 0.2 mM Gly-Pro-NH-Mec was added and incubation was continued at 37°C for a further 30min. Release of 7 amino-4-methylcoumarin was determined as before. Ap- propriate controls and blanks were simultaneously set up.
Substrate specificity of the Triton-X- 100-solubilised post -pr d i n e dipep t idy I-am inopep t idase
50 111 Triton-X-1 00-solubilised post-proline dipeptidyl- aminopeptidase was incubated with 50 pl peptide samples, each at 2 mM in 300 mM potassium phosphate pH 7.4, for 12 h at 37'C. 15 pl was taken from each incubation and spotted onto prepared silica gel G-60 plates (20 x 20 cm) (Merck 5721), which were then developed in chloroform/ methano1/33(Y0 ammonia (60 : 26 : 5). Appropriate standard amino acids and peptides were cochromatographed. The products and standards were visualised with ninhydrin (0.2% in acetone) and with Pauly's reagent. Alternatively 15-pl samples of each incubate were spotted onto Whatman no. 3 paper (57 x 46 cm) and subjected to electrophoresis at 35 V cm- I for 60 min using the buffer system pyridine/acetic acid/ water (1:10:89, v/v/v) pH 3.5. Standard amino acids and peptides were also applied to Whatman no. 3 paper and sub- jected to electrophoresis as above. Products and standards were visualised by chlorination or with Pauly's reagent.
The ability of synapto.somal-mernbrane-boun~~ post-proline dipeptidyl-uminopeptidase and of' the partially purified Triton-X- 100-solubilised post-prolinc d i p q t id.yl-aminopeptidase to hy drolyse His- Pro N H2
20 pl (Pro-3H]thyroliberin (4 pCii0.4 nmol) was incubated with 10 p1 synaptosomal membrane fraction and 10 p1 100 mM potassium phosphate pH 7.6 or 10 p1 100 mM potassium phosphate pH 7.6 containing 4 mM bacitracin or 4 mM N-ethylmaleimide. Alternatively 20 pl [ P ~ O - ~ H I - thyroliberin (5 pCijO.5 nmol) was incubated with 10 p1 purified papain-solubilised synaptosomal pyroglutamate aminopeptidase with or without 10 pl partially purified Triton-X-100-solubilised synaptosomal post-proline dipep- tidyl-aminopeptidase. In some instances 10 pl 5 mM bacitracin was added to the incubation mixture. The final volume was brought to 50 p1 by addition of 100mM potassium phosphate pH 7.4 in instances where some of the above additions above were omitted. After incubation for 8 h at 37 T, 15-pl aliquots were withdrawn from each incubation and applied to preprepared silica gel plates, which were devel- oped using a chloroformimethanol/33% ammonia (60: 26: 5. vjv/v) solvent system. The metabolites containing [3H]proline were located using a Panax thin-layer plate radioscanner. Partial purification ofthe Triton-X-100-solubilisedpast-proline
dipeptidyl-aminopepti~a,~e by gel filtration - _ . _ _ . _ I Preparation of papain-solubilised synaptosomal pyroglutnmute aminopept idase 3.0 ml supernatant, from Triton X-100 treatment of frac-
tion M, was applied to a Sephadex G-200 column (38 x 1.9 cm) previously equilibrated with 100 mM potassium phosphate pH 7.4 containing 0.1 M NaCl and 1% Triton
Pyroglutamate aminopeptidase was solubilised from sy- naptosomal membranes by papain treatment and purified by
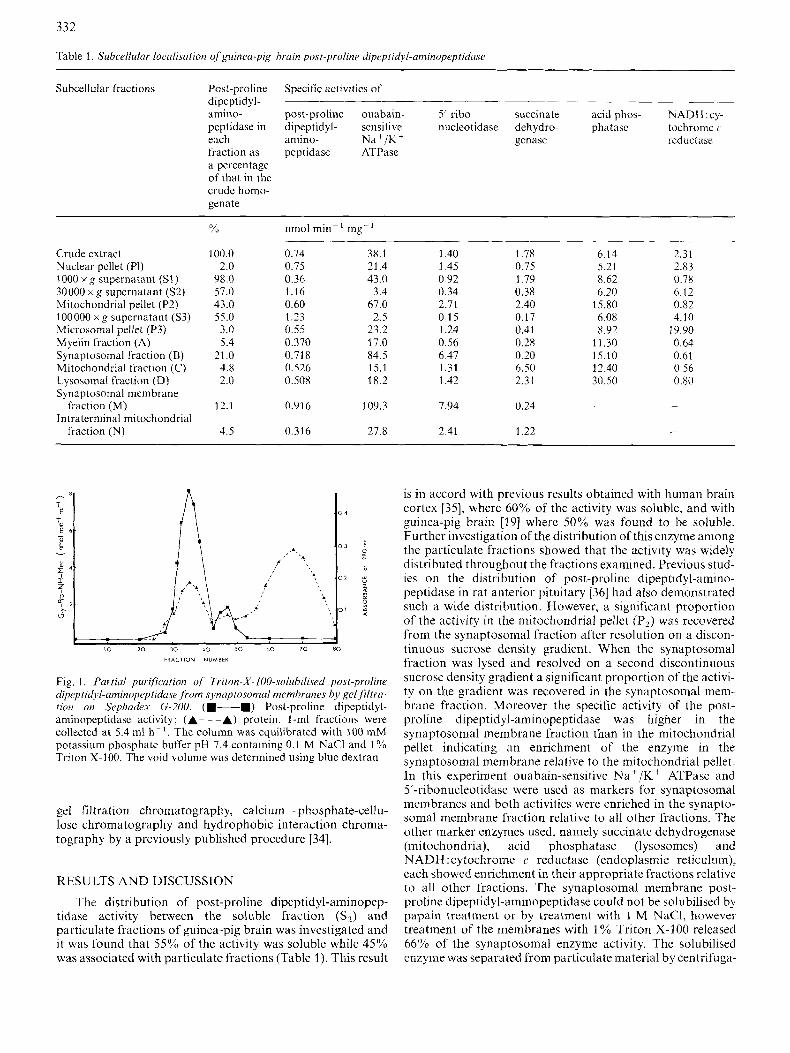
332
Table 1 . Subcellular localisation qf guineu-pig brain post-proiine dipeptidyl-aminopepti~a~~i,
Subcellular fractions Post-proline Specific activities of _ _ ~ _ _ _ ~ _ dipcptidyl-
amino- post-proline Oudbain- 5'-ribo- succinate acid phos- NADI I :cy- nucleotidase dehydro- phatasc tochromc c peptidase in dipeptidyl- sensitive
each amino- Nat /Kt genasc reductase fraction as peptidase ATPase a percentage of that in the crude homo- genate
Crude extract Nuclear pellet (PI) lOO0xg supernatant (Sl) 30000 x g supernatant (S2) Mitochondria1 pellet (P2) I00000 x g supernatant (S3) Microsomal pellet (P3) Myelin fraction (A) Synaptosomal fraction (B) Mitochondria1 fraction (C) Lysosomal fraction (D) Synaptosomal membrane
Intraterminal mitochondria1 fraction (M)
fraction (N)
100.0 2.0
98.0 57.0 43.0 55.0 3.0 5.4
21 .o 4.8 2.0
12.1
4.5
nmol min-
0.74 0.75 0.36 1.16 0.60 1.23 0.55 0.370 0.718 0.526 0.508
_ _ ~
0.916
0.316
I mg-'
38.1 21.4 43.0
3.4 67.0 2.5
23.2 17.0 84.5 15.1 18.2
109.3
27.8
~~
1.40 1.45 0.92 0.34 2.71 0.1 5 1.24 0 56 6 47 1.31 1.42
7.94
2.41
1.78 0.75 1.79 0.38 2.40 0.17 0.41 0.28 0.20 6.50 2.31
0.24
1.22
6.14 5.21 8.62 6.20
15.80 6.08 8.92
11.30 15.10 12.40 30.50
_ _ _ - 2.31 2.83 0.78 6.12 0.82 4.10
19.90 0.64 0.61 0.56 0.80
$ P
FRACTION NUMBER
Fig. 1 , Partial purification of Tritot~-X-100-.~oiuhilised post-proline d~rpti~i,l-aminopeptidase from synaptosomal membranes by gelfl'ltra- /ion on Srphadex G-200. (m-m) Post-proline dipcptidyl- aminopeptidasc activity; (A- - -A) protein. I-ml fractions were collected at 5.4 ml h - '. The column was equilibrated with 100 mM potassium phosphatc buffer pH 7.4 containing 0.1 M NaCl and 1 YO Triton X-100. The void volume was determined using bluc dextran
gel filtration chromatography, calcium - phosphate-cellu- lose chromatography and hydrophobic interaction chroma- tography by a previously published procedure [34].
RESULTS AND DISCUSSION
The distribution of post-proline dipeptidyl-aminopep- tidase activity between the soluble fraction (S,) and particulate fractions of guinea-pig brain was investigated and it was found that 55% of the activity was soluble while 45% was associated with particulate fractions (Table 1). This result
is in accord with previous results obtained with human brain cortex [35], where 60% of the activity was soluble, and with guinea-pig brain [I91 where 50% was found to be soluble. Furthcr investigation of the distribution of this enzyme among the particulate fractions showed that the activity was widely distributed throughout the fractions examined. Previous stud- ies on the distribution of post-prolinc dipeptidyl-amino- peplidase in rat anterior pituitary [36] had also demonstrated such a wide distribution. However, a significant proportion of the activity in the mitochondria1 pellet (P,) was recovered from the synaptosomal fraction after resolution on a discon- tinuous sucrose density gradient. When the synaptosomal fraction was lysed and resolved 011 a second discontinuous sucrose density gradient a significant proportion of the activi- ty on the gradient was recovered in the synaptosomal meni- brane fraction. Moreover the specific activity of the post- proline dipeptidyl-aminopeptidase was higher in the synaptosoinal membrane fraction than in the mitochondrial pellct indicating an enrichment of the enzyme in the synaptosomal membrane relative to the mitochondrial pellet. In this experiment ouabain-sensitive Na'/Kt ATPase and 5'-ribonucleotidase were used as markers for synaptosomal membranes and both activities were enriched in the synapto- soma1 membrane fraction relative to all other fractions. The other marker enzymes used, namely succinate dehydrogenase (mitochondria), acid phosphatase (lysosomes) and NADH :cytochrome c reductase (endoplasmic reticulum), each showed enrichment in their appropriate fractions relative to all other fractions. The synaptosomal membrane post- proline dipeptidyl-aminopeptidase could not be solubilised by papain treatment or by treatment with I M NaCl, however treatment of the membranes with 1% Triton X-100 released 66Yo of the synaptosomal enzyme activity. The solubilised enzyme was separated from particulate material by centrifuga-

333
HIS-PRO HIS : PRO
F n
I T H Y R O L I B E R I N
I ORIGIN 1
HIS- PRO H I S : PRO
Fig. 2. Drgruda (ion of thyroliberin und His-ProNH2 by .synuptosomul mcmhranr preparations and by solubilised preparations qjpyryroglutumure aminopepiiduse und postproline dipeptidyl aminopeptiduse .from .synaptosomul membranes. Each panel (A - F) represents a radioscan of silica gcl thin-layer plates to which 15 p1 appropriate incubation mixture had bccn applied and which had been developed using a chloroform] methanol/33% ammonia (60: 26: 5 , v/v/v) solvent system. The incubation mixtures contained (A) 20 p1 [Pr~-~H]thyroliberin (4 pCi/o.5 mmol), 10 p1 synaptosomal membrane fraction and 10 pI 100 mM potassium phosphate pH 7.6. (B) 20 p1 [Pr~-~H]thyroliberin (4 pCijO.4 nmol), 10 pI synaptosomal membrane fraction and 10 pl of 100 mM potassium phosphate pH 7.6 containing4 mM bacitracin. (C) 20 p1 [Pr~-~H]thyroliberin (4 pCi/0.4 nmol), 10 pI synaptosomal membrane fraction and 10 p1 100 mM potassium phosphate pH 7.6 containing 4 mM N-cthylmaleimide. (D) 20 pI [Pr~-~H]thyroliberin (5 pCi/0.5 nmol), 10 pl purified synaptosomal pyroglutamate aminopeptidasc and 20 p1 100 mM potassium phosphate pH 7.6. (E) 20 p1 [Pm3H]thyroliberin (5 pCi/0.5 nmol), 10 p1 purified synaptosomal pyroglutamate aminopeptidase, 10 p1 of partially purified synaptosomal post-proline dipeptidyl-aminopeptidase and 10 pI 100 mM potassium phosphate pH 7.6. (F) 20 pl [Pro- 'HH]thyroliberin 5 pCij0.5 nmol), 10 p1 purified synaptosomal pyroglutamate aminopeptidase, 10 pI partially purified synaptosomal post- proline dipeptidyl-aminopeptidase and 10 pl 100 mM potassium phosphate pH 7.6 containing 5 mM bacitracin. His:Pro, histidylprolinc di ketopiperazinc
tion at 100000 x g for 75 min and applied to a Sephadex G-200 column, pre-equilibrated with 100 mM potassium phosphate containing 0.1 M NaCl and 1 % Triton X-100. The enzyme eluted as a major peak directly behind the void volume and a minor peak of activity emerging at an elution volume consis- tent with a lower molecular mass (Fig. 1). The major peak was found to have a relative molecular mass in excess of the value of 130000, reported for kidney microvillar membrane monomeric post-proline dipeptidyl-aminopeptidase [9], and in excess of 200000 reported for the soluble enzyme 1191. It is possible that the species eluting from the column in this study represents aggregated forms of the enzyme augmented with Triton X-100 micelles as described in a previous study on a neutral endopeptidase [37]. A pH optimum of 8.0, using 100 mM potassium phosphate buffers, was determined for the major peak of the solubilised enzyme.
The effect of a range of inhibitors on the activity of the synaptosomal post-proline dipeptidyl-aminopeptidase was in- vestigated (Table 2). Whereas the soluble cytoplasmic activity was completely abolished by treatment with N-ethylmaleimide (1 mM), p-chloromercuribenzoate (1 mM) and puromycin (0.5 mM), the solubilised synaptosomal post-proline dipep- tidyl-aminopeptidase was only slightly affected by these rea- gents. On the other hand bacitracin, which displaced a minimal inhibitory effect on the soluble enzyme, almost
completely inhibited the activity solubilised from synapto- somal membranes.
When the post-proline dipeptidyl-aminopeptidase activi- ties, associated with the intraterminal mitochondria1 fraction (N) the lysosomal fraction (D), myelin fraction (A) and the microsomal pellet (P3), were assayed in the presence and ab- sence of 1 mM bacitracin, each activity was almost totally abolished as was the post-proline dipeptidyl-aminopeptidase activity of the synaptosomal fraction (B) and the synapto- somal membrane fraction (M). This result indicates that the post-proline dipeptidyl-aminopeptidase activity associated with synaptosomal membranes is probably closely related to the post-proline dipeptidyl-aminopeptidase activities associ- ated with the other particulate fractions and it is likely that the synaptosomal membrane activity is distributed throughout other particulate fractions. However, it must be remembered that the highest yields of this activity occur in synaptosomal membrane and that the specific activity of this enzyme rises only in the synaptosomal fraction and again in the synaptosomal membrane fraction relative to the mitochon- drial pellet (P?).
The possible role of post-proline dipeptidyl-amino- peptidase in thyroliberin breakdown by plasma membranes was investigated by incubating [Pr~-~H]thyroIiberin with either synaptosoinal membrane preparations or with purified

334
Table 2. The effect uf inhibitors on the major peak, f rom Sephadex G-200 chromatography, of post-proline dipeptidyl-aminopepptidase ac- tivity soluhilised,from synuptosomul membranes by Triton X I 0 0 treut- men t In each case the enzyme was preincubated with inhibitor for 15 min at 37°C and the reaction initiated by addition of substrate
Inhibitor Concn Activity in the presence of inhibitor as a percentage of controls incubated in the absencc of inhibitor
mM % 2-Iodoacetate 1 .o 100
p-Chloromercuribenzoate 1 .o 82 Di thiothreitol 2.0 100 E DTA 5.0 93 8-H ydroxyquinoline 1 .0 25 1 ,lo-Phenanthrolinc 1 .o 65 Phcnyl-methylsulphonyl Fluoride" 0.5 98 Benzamidine 1 .o 99 Baci tracin 0.5 4 Puromycin 0.5 83 Bestatin 1 .0 40
N-Ethylmaleimide 1 .O 85
Assayed in the presence of 5% acetone
Table 3. Substrate specificity of' a partially purified Triton-X-100- .solubilised post-prolina dipepti~yl-uminopepti~use Downward arrows indicate sites of cleavage where these are known
Substrates hydrolysed Substrates not hydrolysed
V
V
V
V
V
His-ProNH,
His-Pro-Val
His-Pro-Leu
His-Pro-Phe
His-Pro-Tyr GI y-Pro-Ala Tyr-Pro-Phe Tyr-Pro-Phc-Pro Tyr-Pro-Phe-Pro-Gly-Pro-Ile Arg-Pro-Lys-Pro Arg-Pro-Lys-Pro-Gln-Gln-Phe
Phe-Gly-Leu-MetNH2 V
Gly-Pro-N H-Mec
Leu-NH,
Leu-Leu-Leu
Leu-Pro
His-Pro
His-Pro diketopiperazine Pro-Pro-Gly Pro-Gly-Gly
Tyr-Gly-Gly Phe-Gly-Gly
Leu-Gly-GI y
Arg-Pro-Pro Arg-Pro-Pro-Gly-Phe-Ser-Pro- Phe-Arg < Glp-Pro-Ser-Lys-Asp-Ala-Phc- Ile-Gly-Leu-MetNH, Cbz-Pro-Phe Pro-MCA Cbz-Gly-Pro-MC A
Mct-Gly-GI y
or partially purified preparations of enzymes released from synaptosomal preparations. The results show that thyroliberin is metabolised to His-Pro diketopiperazine, the cyclisation product of His-ProNH,, and His-Pro by enzymes present in the synaptosomal membrane preparation (Fig. 2A). There were two possible explanations for the appearance of His-Pro as a product. First, thyroliberin may be converted to acid thyroliberin by soluble post-proline cleaving endopeptidase, possibly present as a contaminant in synaptosomal membrane preparations, and that subsequently acid thyroliberin may be converted to His-Pro by the pyroglutamate aminopeptidase, which is known to act on this substrate [2, 341. Secondly, thyroliberin may be converted to His-ProNH, by the action of synaptosomal membrane pyroglutamate aminopeptidase. The His-ProNH, produced spontaneously cyclises to His-Pro-diketopiperazine and may also be converted to His-Pro by the action of synaptosomal post-proline dipeptidyl-aminopeptidase. To determine which pathway had provided for the result observed the incubation was repeated in the presence of 1 mM bacitracin, a cyclic peptide which inhibited both synaptosomal post-proline dipeptidyl-aminopeptidase and soluble post-proline cleaving endopeptidase [19]. The results (Fig. 2B) indicate a marked reduction in the production of His-Pro. However, incubation of synaptosomal membrane fractions with [P~o-~HIthyro- liberin in the presence of N-ethylmaleimide (1 mM), a sulphydryl reagent which completely inhibited the action of soluble post-proline cleaving endopeptidase, while only modestly inhibiting synaptosomal post-proline dipeptidyl- aminopeptidase, resulted in metabolism of thyroliberin to His-Pro (Fig. 2C). These results suggest that the post-proline dipeptidyl-aminopeptidase activity of synaptosomal mem- brane mediates the conversion of His-ProNH, to His-Pro by synaptosomal membranes. When [Pr~-~H]thyroliberin was
incubated with purified synaptosomal membrane pyroglut- amate aminopeptidase, His-Pro diketopiperazine, the cyclisa- tion product of His-ProNH,, was the only metabolite ob- served (Fig. 2D). When the incubation was repeated in the presence of partially purified post-proline dipeptidyl- aminopeptidase, His-Pro was observed as a metabolite along with His-Pro diketopiperazine indicating that the post-proline dipeptidyl-aminopeptidase was capable of competing with the cyclisation process to bring about conversion of His-ProNH, to His-Pro (Fig. 2E). The effect of adding 1 mM bacitracin to the incubation of [P~o-~H]thyroliberin with both purified pyroglutamate aminopeptidase and partially purified post- proline dipeptidyl-aminopeptidase was to totally abolish the production of His-Pro (Fig. 2 F). The partially purified post- proline dipeptidyl-aminopeptidase from synaptosomal membranes had no effect on thyroliberin (results not shown).
The partially purified synaptosomal post-proline dipeptidyl-aminopeptidase released His-Pro from His- ProNH, (Fig. 2 E) His-Pro-Val, His-Pro-Phe and His-Pro-Tyr and Gly-Pro from Gly-Pro-Ala but was unable to release Pro- Pro from Pro-Pro-Gly, Arg-Pro from Arg-Pro-Pro or Arg- Pro-Pro-Gly-Phe-Ser-Pro-Phe-Arg (bradykinin) or Glp-Pro from Glp-Pro-Ser-Lys-Asp-Ala-Phe-Ile-Gly-Leu-MetNH2 (eledoisin) (Table 3 ) . These results indicate that, while the partially purified enzyme acted as a post-proline dipeptidyl- aminopeptidase on a number of tripeptides with proline in the central position, it was unable to effect a cleavage where proline was present in the amino-terminal position as well as the central position as in Pro-Pro-Gly. A free amino terminus was required as indicated by the enzymes failure to cleave Glp-Pro from eledoisin. Agreement was also noted with a previous study [38] where post-proline dipeptidyl-amino-

peptidase was unable to act as an iminopeptidase and hydrolyse the Pro-Pro bond of Arg-Pro-Pro and bradykinin. Neglibile activity was observed with Cbz-Gly-Pro-NH-Mec and Cbz-Pro-Phe as substrates, ruling out the involvement of post-proline cleaving endopeptidase or carboxypeptidase P activities in the observed hydrolysis of His-ProNH,. Also no hydrolysis of His-Pro-Val and His-Pro was observed, in- dicating the absence of aminopeptidase P and proline dipeptidase activities in the partially purified post-proline dipeptidyl-aminopeptidase preparation, thus explaining the non-appearance of Pro or ProNH, on the radioscans. Similarly no prolinase or proline iminopeptidase activity was present as indicated by the non-hydrolysis of Pro-NH-Mec.
If thyroliberin acts as a neurotransmitter or neuro- modulator its activity may be controlled by the synaptosomal membrane pyroglutamate aminopeptidase. His-ProNH2 would be a by-product of this action and the location of a post-proline dipeptidyl-amino-peptidase on the synaptosomal membrane may represent a mechanism for controlling the cyclisation of His-ProNH, to His-Pro diketopiperazine.
We gratefully acknowledge helpful discussion with Dr John Donlon. We thank the Medical Research Council of Ireland for awiirdiI1g 21 grant-in-aid to Gerard O’Cuinn and the Department of Education, Dublin, Ireland for awarding a research maintenance grant to Brendan O’Connor. We also thank Patricia McLaughlin for typing this manuscript and Daniel Collins for Animal Care.
REFERENCES 1 . 2.
3.
4. 5.
6.
7.
8.
9.
Morley, J. E. (1980) Life Sci. 25, 1539-1550. O’Connor, B. & O’Cuinn, G . (1984) Eur. J . Biochem. 144, 271 -
Brabant, G.. Wickings, E. J. & Nieschlag, E. (1981) Acta
Stone, T. W. (1 983) Eur. J. Pharmorol. 92, 11 3 - 11 8. Griffiths, E. & McDermott, J. (1984) Neuroendocrinology 39,
573 - 581. Hopsu-Havu, V. K . & Glenner, G. G. (1966) Histochetnic 7,197 -
201. Svensson, B., Danielsen, M., Staun, M., Jeppesen, L., Noren,
0. & Sjostrom, H. (1978) Eur. J . Biochem. 90,489-498. Kenny. A. J., Booth, A. G., George, S . G., Ingram, J., Kershaw,
D., Wood. E. J. & Young, A. R. (1976) Biochem. J. 155, 169 - 1x2.
Macnair. R. D. C. & Kcnny, A. J. (1979) Biochrm. J . lY7, 379- 395.
278.
Endocrinol. 98, 189 - 194.
10.
11. 12.
13.
14.
15.
16.
17.
18.
19. 20.
21.
22. 23.
24.
25. 26.
27. 28. 29.
30.
31.
32.
33.
34.
35.
36.
37. 38.
335
Fukasawa, K. M., Fukasawa, K., Hiraoka, B. Y. & Harada, M.
Heymann, E. & Mentlein, R. (1978) FEBS Lcti. 91, 360-364. Yoshimoto, T. & Walter, R . (1977) Biochim. Biophys. Acta 485,
Yoshimoto, T., Fischl, M., Orlowski, R. C. & Walter, R. (1978)
Oya, H., Nagatsu, I. & Nagatsu, T. (1972) Biochim. Biophys. Acta
Kqjima, K., Hama, T., Kato, T. & Nagatsu, T. (1980) J .
Kato, T.. Iwase, K., Nagatsu, T., Sakakibara, S . & Fujita, K.
Kato, T., Iwase, K., Nagatsu, T., Sakakibara, S. & Fujita, K.
Bauer, K. & Kleinkauf, H. (1980) Euu. .I. Biochem. 106, 107-
Browne. P. & O’Cuinn, G. (1983) Eur. J . Biochem. 137, 75-87. Kato, T., Nagatsu, T., Kimura. T. & Sakakibara, S. (1978) Bio-
Yoshimoto, T., Ogita, K., Walter, R., Koida, M. & Tsuru, D.
Browne, 1’. & O’Cuinn, G. (1983) J . Biol. Chem. 258,6147-6154. Nicholson, J. A . & Peters, T. J. (1978) Anul. Biochem. 87, 418-
Severson, D. L., Drummond, G. T. & Sulakhe, P. V. (1972) J .
King, E. J. (1932) Biochem. J . 26, 292-297. Michell, R. H. &Hawthorne, J. N. (1965) Biochim. Biophys. Res.
Pennington, R. J. (1961) Biochem. J . 80, 649-654. Portcous, J. W. & Clark, B. (1965) Biochem. J . 96, 159-171. Takesue, J. & Omura, T. (1970) J . Biochem. (Tokyo) 67, 267-
276. Jarasch, E. D. & Franke, W. W. (1974) J . Biol. (:hem. 249, 7245-
7254. Beaufay, H., Amar-Costesec, A., Feytmans, E., Thines-Sempoux,
D., Wibo, M., Robbi, M. & Berthet, J. (1974) J . Cell. B id . 61,
Marchbanks, R . M. (1976) in Practical neurochemistrv (McIllwain, H., ed.) pp. 208 -242 Churchill-Liviugstone, Lon- don.
Whittaker, V. P. & Barker, L. A. (1972) Methods Neurochem. 2, 1-52.
O‘Connor, B. & O’Cuinn, G. (1985) Eur. J . Biochem. 150, 47- 52.
Kato, T., Hama, T. & Nagatsu, T. (1980).1. Neurochem. 34,602- 608.
Horsthemke, B., Leblanc, P., Kordon, C., Wattiaux.De Coninck, S., Wattiaux, R. & Bauer, K. (1984) Eur. J . Biochem. 139,315- 320.
(1981) Biochim. Biophys. Acta. 657, 179-189.
391 -401.
J . B i d . Chem. 253, 3708 - 3716.
258, 591 - 599.
C‘hruinatogr. 189, 233 - 240.
(1979) Experientiii (Basel) 35, 20-21.
(1979) E.xpc>rientia (Busel) 35, 1329-1330.
117.
chitn. Med. I Y , 352 -359.
(1979) Biochim. Biophys. Acta 569, 184-192.
424.
Biol. Chcvn. 247, 2949 -2958.
Cotnmun. 21, 333 -337.
188 - 200.
Fulcher, I . S. & Kenny, A. J . (1983) Biochem. J . 211, 743-753. Kato, T., Nagatsu, T., Fukasawa, K., Harada, M., Nagatsu, I . &
Sakakibara, S. (1978) Biochim. Biophys. Acia 525, 417-422.


















