
POLYSACCHARIDES FROM Turbinaria conoides AND...
90
THESIS SUBMITTED FOR THE DEGREE OF DOCTOR OF PHILOSOPHY (SCIENCE) IN CHEMISTRY OF THE UNIVERSITY OF BURDWAN 2013 POLYSACCHARIDES FROM Turbinaria conoides AND Adhatoda vasica : STRUCTURAL FEATURES AND BIOLOGICAL ACTIVITIES Nabanita Chattopadhyay, M. Sc. NATURAL PRODUCTS LABORATORY DEPARTMENT OF CHEMISTRY THE UNIVERSITY OF BURDWAN WEST BENGAL, INDIA
Transcript of POLYSACCHARIDES FROM Turbinaria conoides AND...

THESIS SUBMITTED FOR THE DEGREE OF
DOCTOR OF PHILOSOPHY (SCIENCE) IN CHEMISTRY
OF THE UNIVERSITY OF BURDWAN
2013
POLYSACCHARIDES FROM
Turbinaria conoides AND Adhatoda vasica :
STRUCTURAL FEATURES AND BIOLOGICAL
ACTIVITIES
Nabanita Chattopadhyay, M. Sc.
NATURAL PRODUCTS LABORATORY
DEPARTMENT OF CHEMISTRY
THE UNIVERSITY OF BURDWAN
WEST BENGAL, INDIA

Dedicated to my beloved parentsDedicated to my beloved parentsDedicated to my beloved parentsDedicated to my beloved parents

THE UNIVERSITY OF BURDWAN
TO WHOM IT MAY CONCERN This is to certify that the thesis “POLYSACCHARIDES FROM Turbinaria
conoides AND Adhatoda vasica: STRUCTURAL FEATURES AND
BIOLOGICAL ACTIVITIES” is the result of work done by Nabanita
Chattopadhyay, M.Sc., who has registered her name in The University of
Burdwan on 19-03-2007 for the award of Doctor of Philosophy (Science) in
Chemistry under my supervision and Guidance. This work neither in part nor
whole has been submitted for any degree by Nabanita Chattopadhyay or other. It is
also certified that she has successfully completed Ph.D. course work framed by the
Department of Chemistry, following the guidelines of The University of Burdwan,
and that she has delivered one seminar lecture on 02-08-2012 in our department
regarding fulfillment of all the requirements for submitting the thesis for Ph.D.
degree under new regulation, 2009 of this University.
(Dr. Bimalendu Ray)
Dr. Bimalendu Ray Associate Professor
****
Department of Chemistry
Golapbag, Burdwan 713104, India
Tel: +91-342-2533913 (O)
+91-342-2657709 (R)
Fax: +91-342-2530452(O)
E-mail: [email protected]
Dated…………………..

ACKNOWLEDGEMENT
My journey at the University of Burdwan would not have become fruitful without the advice
and assistance of some special persons. Let me at the outset express my heartfelt gratitude to my
revered teacher and supervisor Dr. Bimalendu Ray, Associate Professor, Department of
Chemistry, The University of Burdwan, for his continuous supportive guidance, untiring help
and encouragement during the course of my work. I am very much thankful to him for his active
participation in all of my publications related to this thesis.
I would like to express my sincere gratitude to Prof. Pradyot Ghosal, Prof. Subrata Laskar,
and Dr. A. K. Ghosh, the present head of the Department of Chemistry, The University of
Burdwan, for their generous help, constant encouragement.
I record my indebtedness to Prof. Gabriella Nosál’ova, Department of Pharmacology,
Jessenius Faculty of Medicine, Comenius University, Martin, Slovakia, who helped me with
Bioassay. I am thankful to all the faculty members, staffs, and research scholars of the
Department of Chemistry, The University of Burdwan, for their kind cooperation in academic,
official and laboratory works related to this thesis. Valuable help rendered by lab-mates Dr.
Utpal Adhikari, Dr. Kausik Chattopadhyay, Dr. Pinaki Mandal, Dr. Tuhin Ghosh, Mr. Sudipta
Saha, Mr. Udipta Ranjan Chatterjee, Mr. Shruti Sourav Bandyopadhyay, Mr. Sujay Majee, Mrs.
Paramita Karmakar, Ms.Sharmistha Sinha, Ms. Debjani Ghosh, Ms. Kanika Ghosh deserve
special mention.
My sincere thanks are cordially extended to Mrs. Tapasi Ray for her enthusiastic
encouragement and cooperation. The infrastructural facilities from The University of Burdwan
are gratefully acknowledged.
Finally, my special acknowledgements are owed to my parents, my sister, brother-in-law,
brother, and last but not the least my husband for their help, constant inspiration, encouragement
in course of my research activity.
Department of Chemistry,
The University of Burdwan,
Burdwan, 713104, India. (Nabanita Chattopadhyay)

CONTENTS
1. POLYSACCHARIDES FROM NATURAL SOURCES: BIOLOGICAL ROLES, INDUSTRIAL USES AND PHARMACOLOGICAL ACTIVITIES
1-1. INTRODUCTION
1-2. BIOLOGICAL ROLES
1-3. INDUSTRIAL USES
1-4. PHARMACOLOGICAL ACTIVITIES
1-4.1. Anticoagulant activity
• Structure-activity relationships
• Mechanism of action
1-4.2. Anticomplementary activity
• Structure-activity relationships
1-4.3. Antioxidative activity
• Structure-activity relationships
1-4.4. Antitumor activity
• Structure-activity relationships
• Mechanism of action
1-4.5. Antitussive activity
• Structure-activity relationships
• Mechanism of action
1-4.6. Antiviral activity
• Structure-activity relationships
• Mechanism of action
1-4.7. Potential clinical applications
10
11
12
14
15
16
18
19
19
20
21
22
9
8
7
6
5
4
3
3
2

AIMS
2. STRUCTURAL FEATURES AND ANTIOXIDATIVE ACTIVITIES OF CARBOHYDRATE POLYMERS FROM THE BROWN SEAWEED Turbinaria conoides
2-1. INTRODUCTION
2-2. CHEMICAL CHARACTERIZATION OF POLYSACCHARIDES FROM Turbinaria conoides
2-2.1. Preparation of depigmented algal power and sugar compositional
analysis
2-2.2. Isolation and sugar composition of polysaccharide fractions
2-2.3. Purification of the fucoidans by chromatography
2-2.4. Molecular mass
2-2.5. Linkage analysis
2-2.6. NMR spectroscopy
2-2.7. Structural features of the glucan and alginic acid rich fractions
2-3. PHARMACOLOGICAL ACTIVITIES OF POLYSACCHARIDES FROM Turbinaria conoides
2-3.1. Antioxidative activity
• Ferric ion reducing/antioxidant power (FRAP) assay
• Scavenging effect on 1, 1-diphenyl-2-picrylhydrazyl (DPPH) radicals
2-4. CONCLUSIONS
24
27
28
28
30
32
33
33
36
37
38
38
38
39
42

3. STRUCTURAL FEATURES AND ANTITUSSIVE ACTIVITIES OF A CARBOHYDRATE POLYMER FROM Adhatoda vasica
3-1. INTRODUCTION
3-2. CHEMICAL CHARACTERIZATION OF THE PECTIC ARABINOGALACTAN FROM Adhatoda vasica
3-2.1. Isolation and chemical composition
3-2.2. Size exclusion chromatography (SEC)
3-2.3. Linkage analysis
3-3. ANTITUSSIVE ACTIVITIES OF THE PECTIC ARABINOGALACTAN FROM Adhatoda vasica
3-3.1. Assessment on chemically induced cough and airways defense reflexes
3-3.2. Mechanism of action
3-4. CONCLUSIONS
4. MATERIALS AND METHODS
4-1. PLANT MATERIALS AND EXTRACTION OF POLYSACCHARIDES
4-1.1. Isolation of sulfated fucan, alginic acid and glucan from Turbinaria conoides by sequential extraction with inorganic solvents
• Plant material and preliminary treatments
• Preparation of depigmented algal powder (DAP)
• Extraction with acid
• Extraction with alkali
• Extraction with water
4-1.2. Isolation of polysaccharides from Adhatoda vasica
• Plant material and preliminary treatments
• Extraction of polysaccharides
44
46
46
47
48
50
50
51
52
55
55
55
55
55
55
55
56
56
56

• Isolation of arabinogalactan protein (AGP) with β-glucosyl Yariv reagent
4-2. ANALYTICAL METHODS
4-2.1. General 4-2.2. Sugar analysis
4-2.3. Protein estimation
4-2.4. Sulfate estimation
• Turbidometric method
• Spectroscopic method
4-2.5. Desulfation 4-2.6. Linkage analysis
4-2.7. Chromatography
• Thin layer chromatography (TLC)
• Size exclusion chromatography (SEC)
• Anion Exchange Chromatography (AEC)
• Gas chromatography (GC)
4-2.8. Spectroscopy
• NMR Spectroscopy
• UV-VIS spectroscopy
• Infra-Red spectroscopy
• Mass spectrometry
4-3. BIOASSAY
4-3.1. FRAP assay
4-3.2. Scavenging capability for 1,1-diphenyl-2-picrylhydrazyl
(DPPH) radicals
56
56
56
57
57
58
58
58
58
58
59
59
59
60
60
60
60
61
61
61
61
61
61

4-3.3. Antitussive activity of the arabinogalactan from A. vasica
• Animals
• Assessment of chemically induced cough and airways defense reflexes
• Statistics
REFERENCES SUMMARY LIST OF PUBLICATIONS
62
62
62
62
64
80
81

1
POLYSACCHARIDES FROM NATURAL SOURCES: BIOLOGICAL ROLES,
INDUSTRIAL USES AND PHARMACOLOGICAL
ACTIVITIES

0
100
200
1-1. INTRODUCTION
Carbohydrates are the fuel of life, being the main source of energy for living organisms and
the central pathway of energy sto
through which the energy of the sun is harnessed and converted into a
by living organisms. According to rough estimates, carbohydrate
annually regrowing biomass of about 200 billion tons
man, the rest decays and recycles along natural
Polysaccharides, proposed as the first biopolymers to have formed on Earth (Tolstoguzov,
2004), are a major group
macromolecule can be defined in terms of the composition
configuration and sequence of its constituent sugars as well as the presence of any non
residues and their positions. Polysaccharides also have secondary, tertiary and quat
structures, which depend upon the repeating sequence in primary structure, the confor
glycosidic linkages and aggregation of polymer chain by various non
two decades the mechanism behind the biological
uses and therapeutic applications
documenting the developments
Fig. 1.1. Carbohydrate is a primary component of renewable biomass
Bil
lio
n t
on
s
Carbohydrate
Non-Carbohydrate
Carbohydrates are the fuel of life, being the main source of energy for living organisms and
the central pathway of energy storage and supply for most cells. They are the major products
through which the energy of the sun is harnessed and converted into a form that can be utilized
by living organisms. According to rough estimates, carbohydrates represent roughly 95% of the
annually regrowing biomass of about 200 billion tons (Fig. 1.1); of these only 3% are used by
the rest decays and recycles along natural pathways (Lichtenthaler &
proposed as the first biopolymers to have formed on Earth (Tolstoguzov,
2004), are a major group in carbohydrate chemistry. The primary structure of this
macromolecule can be defined in terms of the composition, glycosidic linkage, anomeric
and sequence of its constituent sugars as well as the presence of any non
. Polysaccharides also have secondary, tertiary and quat
upon the repeating sequence in primary structure, the confor
linkages and aggregation of polymer chain by various non-covalent bonds.
mechanism behind the biological roles of carbohydrate polymers,
and therapeutic applications have increased tremendously. This
the developments of these macromolecules in these fields.
1. Carbohydrate is a primary component of renewable biomass
2
Carbohydrates are the fuel of life, being the main source of energy for living organisms and
They are the major products
form that can be utilized
represent roughly 95% of the
; of these only 3% are used by
Mondel, 1997).
proposed as the first biopolymers to have formed on Earth (Tolstoguzov,
The primary structure of this
, glycosidic linkage, anomeric
and sequence of its constituent sugars as well as the presence of any non-sugar
. Polysaccharides also have secondary, tertiary and quaternary
upon the repeating sequence in primary structure, the conformation of
covalent bonds. In the last
polymers, their industrial
This chapter aims at
1. Carbohydrate is a primary component of renewable biomass.

3
1-2. BIOLOGICAL ROLES
Carbohydrate polymers occur in almost all living organism and serve diverse functions in the
living material in which they are endogenous. The function of cellulose, the most abundant
naturally occurring substance, as structural support in plant is well established. But, in animals,
they rarely serve such purposes as structural support. However, the special physical texture and
the hydrophilic character are responsible for their multi-various roles. Cell walls of many
bacteria contain polysaccharides, which are responsible for their protective coatings and
serologic specificity (Holst, 1999). Some bacterial polysaccharides are highly antigenic having
endotoxin properties. Other natural macromolecules, which are not composed entirely of sugar
units, contain blocks of monosaccharide units as part of the molecular structure, and contribute
extensively to the production and maintenance of living tissues of animals. The blood-group
polysaccharide constitutes a group of glycoproteins in which arrangement of monosaccharide
residues in carbohydrate subunits controls the blood-group specificity to the overall molecule
(Feizi, 2000). Immunoglobulins are a group of glycoprotein that has antibody activity (van de
Perre, 2003). Transferrin is a glycoprotein, which forms complexes with iron and is responsible
for transporting iron from the storage form in tissues, especially in liver to the metabolically
functioning iron in hemoglobin (Kawabata et al., 2000). Glycosaminoglycans are amongst the
essential building blocks of the macromolecular framework of connective and other tissues
(Taylor & Gallo, 2006). Hyaluronic acid appears to act, on account of its viscosity in solution, as
a lubricant, shock-absorbing gel in limb joints (Kogan et al., 2007).
1-3. INDUSTRIAL USES
Interest in polysaccharides, however, is not purely because of its biological activities but
rather is prompted by their great utility as a raw material in many industries such as food,
pharmaceutical, etc. Water-soluble polysaccharides have enormous use in food industries. Their
non-toxicity, wide availability, and low cost make them suitable for food industries. Also, these
macromolecules are chosen for their added values, as for example for their low calorie intake.
Importantly, they are mostly used as additive to improve or control food properties. Notably,
their applications largely depend on the properties, they impart to solution and gels. The different
families of polysaccharides and their uses in food industry are as follows: (i) anionic exudates

4
polysaccharides such as gum arabic used as an emulsifying agent in the precandy jellies such as
jujubes, fruit gums, fruit pastille, gum drops and cough drops & stabilizing agent as angel kisses,
marshmallows, soft caramels, nougats and meringues (Kubal & Gralen, 1948; Meer, 1980), gum
tragacanth and karaya mainly used as thickener and emulsifier in sauces, salad dressings and
confectionery lozenges (Whistler, 1993), (ii) anionic seaweed polysaccharides including agars
used as gelling agent in wine and fruit juice (Tseng, 1946; Selby & Whistler, 1993), alginates as
gelling agent in making jam, jellies, fruit fillings etc & as stabilizing agent in beer, pulp, fruit
drinks without forming haze and low fat substitutes and carrageenans used as stabilizing agent in
dairy products such as milk chocolate, yogurts and egg nog mixes, ice-cream etc (Painter, 1983;
Piculell, 1995), and also in non-dairy food product, such as instant products, jellies, pet foods,
sauces (Therkelsen,1993; Imeson, 2000), (iii) microbial polysaccharides such as gellan gum &
xanthan gum (extensively used in the food industry as stabilizing, thickening, gelling agent), (iv)
non-ionic seed polysaccharides including guar gum (an economical thickener and stabilizer in ice
creams, sherbets and related products, breads, cakes and donuts & other dairy products),
tamarind seed gum, locust bean gum and (v) pectin, mainly used as gelling agent but also as
stabilizing, thickening and suspending agent (May, 1990; Rolin & DeVries, 1990; Pilnik, 1992).
Polysaccharides commonly used in non food industry are (i) anionic exudates polysaccharides
such as gum tragacanth and karaya (Whistler, 1993), (ii) anionic seaweed polysaccharides such
as agars (Tseng, 1946; Selby & Whistler, 1993) and carrageenans (Painter, 1983; Therkelsen,
1993; Piculell, 1995; Imeson, 2000), (iii) microbial polysaccharides such as dextran (DeBelder,
1993), and xanthan.
The industrial applications of polysaccharides depend on their unique properties, often at a
cost, much lower than the synthetic polymers. They can produce gels of different strength and
stability under controlled condition. Also, they can act as good emulsifier by their interfacial
binding properties. Polysaccharides are useful to supply required consistency by controlling the
moisture. They are important as additives in relatively low proportions, as for example as
thickening, stabilizing agents.
1-4. PHARMACOLOGICAL ACTIVITIES
Recognition of the pharmacological properties of myriad polysaccharides has fueled the
current focus on the search for new drug candidates. Besides their well-attested anticoagulant

5
and antithrombotic activities, they can act on complement systems, have tumoricidal, antitussive
and antioxidative properties, and can protect cells from viral infection (Yamada & Knutsen,
1996; Franz et al., 2000; Paulsen & Samuelsen, 2001; Inngjerdingen et al., 2006; Balzarini &
Van Damme, 2007; Mantovani et al., 2008; Ghosh et al., 2009a; Baek et al., 2010; Sinha et al.,
2011a, 2011b; Bandyopadhyay et al., 2012; Thakur et al., 2012).
The next part of this chapter, which summarizes experimental evidences indicating that
polysaccharides might play increasingly important roles in the prevention of diseases in the near
future, is divided into six parts according to their pharmacological activities. Further
classification within each activity is made by structural type.
1-4.1. Anticoagulant activity
Heparin is a heterogeneous group of straight-chain anionic mucopolysaccharides, called
glycosaminoglycans, having anticoagulant properties (Linhardt & Toida, 1997; Gunay &
Linhardt, 1999; Alban & Franz, 2001; Lee et al., 2008; Fan et al., 2011). This linear
polysaccharide consists of simple disaccharides repeats of alternating unit of (1→4)-linked
uronic acid and glucosamine, made complex by variation of substitutions with O- and N- sulfo
and N- acetyl groups, as well as by epimerization of the uronic acid (Mulloy & Linhardt, 2001;
Feyerabend et al., 2006). This compound has very much higher specific activity as an
anticoagulant than other sulfated polysaccharides and this potency depends on a specific
pentasaccharide sequence (Berteau & Mulloy, 2003; Mulloy, 2005). However, because of
several side effects (Pereira et al., 2002; Athukorala et al., 2007) and limited availability of
heparin, there is considerable interest to obtain safer anticoagulants (heparinoids) from other
natural sources (Alban et al., 2002). In fact, the first report of the heparinoid activity of high-
molecular-weight polysaccharides appeared almost 55 years ago (Springer et al., 1957).
Recently, a number of sulfated polysaccharides from marine sources have emerged as an
important class of compound having anticoagulant effects. Among them, fucoidans are well
known for their heparinoid activity (Nishino & Nagumo, 1991, 1992; Chauvet et al., 1999;
Chevolot et al., 1999; Pereira et al., 1999; Berteau & Mulloy, 2003; Silva et al., 2005; Cumashi
et al., 2007; Chandia & Matsuhiro, 2008). Based on structural features fucoidans are of three
major types: (i) a polysaccharide based on L-fucose with mainly α-(1→3)-glycosidic bonds and
sulfate groups at position 4 (Patankar et al., 1993) (Fig. 1.2), (ii) fucoidan possessing large
proportions of both α-(1→3)-and α-(1→4)-glycosidic bonds (Chevolot et al., 1999, 2001;

6
Daniel et al., 1999, 2001) and (iii) macromolecule is composed primarily of α-(1→2)- and α-
(1→3)-linked fucopyranosyl residues with sulfate groups at position 4 and 2 (Karmakar et al.,
2009).
Besides fucoidan, sulfated galactan (Farias et al., 2000; Matsubara et al., 2001; Farias et al.,
2008; Glauser et al., 2009) and several other sulfated heteropolysaccharides having anticoagulant
activity has also been indicated (Rojers et al., 1990; Guven et al., 1991; Maeda et al., 1991;
Potin et al., 1992). Notably, the anticoagulant activity of some of these compounds such as
rhamnan sulfate, a macromolecule isolated from Monostroma spp., is six folds higher than that
of standard heparin (Maeda et al., 1991). Furthermore structurally well defined sulfated
polysaccharides produced by chemical sulfation also found to have antithrombotics properties
(Alban & Franz, 2001; Alban et al., 2002).
Structure-activity relationships. First, it has been observed that for polysaccharides with the
same structure, the anticoagulant activity depends on the ratio of sulfate group to total sugar
residues of the polysaccharides (Nishino & Nagumo, 1991, 1992). The higher the ratio is, the
OH3C
O
O
O
OR1
R2O
OR3
H3C
O
OH3C
OOR1
R2O
R2O
OH3C
OOR1
R2O
OH3COR1
R2O
R1 = H or SO3- or COCH3
R2 = H or SO3-
R3 = OH3C
HO
O
OR2
R2O
Fig. 1.2. The quasi-repeat unit identified in 1→3-linked fucoidan from Chorda filum (Chizhov et al.,
1999). Other substituents, such as O-acetyl, and branches are present in all these fucoidans and add
considerably to their heterogeneity.

7
better are the chances of exhibiting higher anticoagulant activity. Nishino and colleagues have
found that higher the content of fucose and sulfate groups present, higher is the anticoagulant
activity in native fucoidans from Ecklonia kurome (Nishino et al., 1989; Nishino & Nagumo,
1991, 1992). But it is always not a rule. For example, it has been observed that although the ratio
(0.75) of sulfate to sugars of a fucoidan fraction isolated from commercial Fucus vesiculosus was
higher than those (0.31-0.68) of other fractions, but the former polysaccharide preparation was,
nevertheless, inactive (Nishino et al., 1994). These results clearly demonstrate that not only the
sulfate group but also structural features are responsible for the anticoagulant activity of sulfated
polysaccharides. Second, the location of sulfate group and/or the glycosidic linkage position
affect the activity, as indicated by the comparison between the inactive 3-sulfated 4-linked and
the active 2-sulfated 3-linked α-L-galactans (Mourao, 2004). Third, the occurrence of 2, 4-di-
sulfated units has an amplifying effect on the antithrombin-mediated anticoagulant activity of 3-
linked α-L-fucoidan. This is not merely a consequence of increased charge density. The
anticoagulant activity increases ~38-fold from a 2-sulfated 3-linked α-L-fucoidan to a 2, 4-
disulfated α-L-fucoidan, even though their sulfate content increases only ~1.8-fold (Pomin &
Mourao, 2008). Fourth, the anticoagulant activity of a particular family of polysaccharide also
depends upon their molecular mass. The native fucoidan (MW 320,000 Da) from Lessonia
vadosa have good anticoagulant activity, whereas the radical depolymerized fraction (MW
32,000 Da) exhibits weak anticoagulant activity (Chandia & Matsuhiro, 2008). Finally, the
structures of sulfated polysaccharides vary based on their algal source species to species and give
rise to variation in the degree of anticoagulation action (Chevolot et al., 1999; Pereira et al.,
1999; Boisson-Vidal et al., 2000; Pomin et al., 2005).
Mechanism of action. The biological activities of heparin are due to its binding to cellular
proteins modulating their activities. This interaction is often very specific (Gunay & Linhardt,
1999; Mulloy, 2005), e.g., heparin’s anticoagulant activity mainly results from binding
antithrombin III (ATIII, a serine protease inhibitor of thrombin and other coagulation proteases)
at a discrete pentasaccharide sequence (Fig. 1.3). Sulfated polysaccharides contain several types
of functional groups such as sulfate esters, carboxyls, hydroxyls and, therefore, able to interact
with cellular proteins via: (i) hydrogen bond, (ii) electrostatic interaction, (iii) hydrophobic
interactions and (iv) van deer Waals forces (Hileman et al., 1998; Quiocho & Vyas, 1999).
Electrostatic interaction can occur between locally positively charged patches on the protein

8
(under normal physiological pH) and sulfate groups of heparinoids (reviewed by Gunay &
Linhardt, 1999). The anionic substituents (sulfate and carboxylate groups) in the saccharide
backbone of heparin and some heparinoids have different spatial arrangements, which stem from
a various degree of chirality, different regioisomers, conformers and secondary structure in
carbohydrate backbone. Ease in re-orientation of charged groups, owing to the flexible
saccharide backbone, further assists in such interactions. Heparinoids can also form hydrogen
bonds with the polar amino acids of binding proteins. The residues involved in ionic and
hydrogen bonding interactions are usually spatially arranged either on the surface or on the
binding pocket of the proteins. Substantial hydrophobic contributions to binding may result in
the interaction of heparinoids having hydrophobic character (i.e. fucoidan) making these
effectively potent agents (Gunay & Linhardt, 1999).
1-4.2. Anticomplementary activity
The complement system is part of the innate immune system and consists of a group of serum
proteins which are activated in a cascade mechanism (Samuelsen et al., 1996). This system is
important in initiating inflammation, and its activation might result in the cellular co-operation,
immunopotentiation and regulation of cyclical antibody production. This activation may be
beneficial to the host, but it may also be harmful by damaging host cells and tissues. Many
naturally occurring polysaccharides from plant, bacteria, lichen, fungi have effects on the human
immune system. These compounds include pectin, pectic arabinogalactan (AG), and other acidic
heteroglycans. In addition, several neutral polysaccharides such as glucan (Tomoda et al., 1994a,
1994b, Olafsdottir et al., 1999a), arabinan (Yamada et al., 1989b), arabinoglucan (Yamada et
al., 1989b), galactan (Yamada et al., 1989a), rhamnopyranosylgalactofuranan (Olafsdottir et
al.,1999b), galactomannan (Latge et al., 1994), arabinoxylan (Hromadkova et al., 2012), and
other hetero polysaccharides (Tomoda et al., 1994b; Zhao et al., 1994) with anticomplementary
O
CH2SO3
OH
O
O
O
(Ac or SO3) HN
CO2
OH
OH
O
O
CH2OSO3
OSO3
NHSO3
O O
O
CO2OH
OSO3
O
CH2OSO3
OH
NHSO3
O
Fig. 1.3. Discrete pentasaccharide sequence of Heparin (Mulloy & Linhardt, 2001).

9
activity have been isolated from medicinal plants or released from pectic polysaccharides during
isolation. Finally, carbohydrate polymers that are not composed entirely of monosaccharide
residues can also activate complement system. For example, the highly branched protein-bound
polysaccharide (~ 1000-2000 kDa), containing terminal, 1,3-, 1,4-, 1,2,6-, 1,3,6-linked glucose;
1,6-, 1,2,6-, 1,3,4-, 1,4,6-linked galactose; 1,5-, 1,3,5-linked arabinose, terminal and 1,2,5-linked
rhamnose residues and 6.55% of protein from Eucommia showed potent anticomplementary
activity (Zhu et al., 2009).
Structure-activity relationships. Because of great chemical variety, it is very difficult to
correlate between structure and activity. However, on the basis of accumulated data the
following conclusions could be made for pectic polysaccharides: (i) the “smooth” region i.e., the
region which consists of polygalacturonide moiety (Voragen et al., 1995) have none or only
weak activities, (ii) esterification, acetylation or uronic acid content of the polymers do not affect
activity (Kiyohara et al., 1989, 1993; Yamada et al., 1989a, 1992; Hirano et al., 1994; Tomoda et
al., 1994a; Samuelsen et al., 1996, 1998; Shin et al.,1998; Yu et al., 1998; Sakurai et al., 1999),
(iii) the hairy region (Voragen et al., 1995) of the pectin in particular the carbohydrate side
chains that attached to the hairy region, might be essential for the expression of this activity
(Kiyohara et al., 1989, 1993; Yamada et al., 1992; Samuelsen et al., 1996, 1998), (iv)
polyanionic smooth regions in the pectin might modulate the mode of complement activation
induced by the hairy region (Kiyohara et al., 1989), (v) a high degree of branching may be
important for the activity and arabinose attached terminally to the galactan side chains reduces
the activity, while arabinose side chain attached directly to C3 of the galacturonic acid residues
increases the activity (Samuelsen et al., 1996), (vi) molecular weight also plays an important role
in exhibiting this activity. For example, an arabinogalactan from Atractylodes lancea DC having
molecular weight (74,000) showed a strong reactivity, whereas other arabinogalactans
(molecular weight 3,100 & 16,000) from the same source did not show the reactivity (Yu et al.,
1998). A careful study of the monomerization and dimerization of the rhamnogalacturonan II
(RG II, a type of pectic polysaccharide having very complex structure) isolated from the leaves
of Panax ginseng clearly indicated that the RG II dimers cross-linked by borate diesters strongly
contribute to the activity (Shin et al., 1998). The higher the proportion of dimer the higher is the
activity. Among branched α-D-glucan, polymers with higher degree of branching have higher
potency (Tomoda et al., 1994a; 1994b), but for linear glucan the presence of α-(1→3)-linkages

10
is essential (Olafsdottir et al., 1999a). Besides, the higher the molecular mass of the
macromolecule, the better is its potency (Olafsdottir et al., 1999a).
These facts clearly demonstrate that besides primary structure some other features are
responsible for the biological activity of pectin. An attractive concept is that polymers having
comb like conformation exhibit anticomplementary activity. Hairy regions of the pectic
substances are characterized by a linear backbone with side chain and hence possess a comb-type
structure. Arabinogalactan, a branched polymer, obtained from various sources showed
anticomplementary activity (Wagner & Jordan, 1988; Kiyohara et al., 1993; Samuelsen et al.,
1998; Yu et al., 1998). So it seems that a comb-like structure with a linear backbone and
branched side-chains may be prerequisite for this activity.
1-4.3. Antioxidative activity
A biological antioxidant has been defined as “any substance that, when present at low
concentrations compared to those of an oxidizable substrate, significantly delays or prevents
oxidation of that substrate” (Halliwell & Gutteridge, 1995). Although oxidation reactions are
crucial for life, they can also be damaging and can cause many diseases like cancer (Paz-Elizur
et al., 2008), liver disease (Preedy et al., 1998), Alzheimer’s disease (Moreira et al., 2005), aging
(Liu & Mori, 2006), arthritis (Colak, 2008), inflammation (Mukherjee et al., 2007), diabetes
(Naito et al., 2006; Jain, 2006), Parkinson’s disease (Beal, 2003; Chaturvedi et al., 2008),
atherosclerosis (Heinecke, 1997), and AIDS (Sepulveda & Watson, 2002). Natural compounds
offer interesting pharmacological properties for their use as antioxidative agent. In recent years, a
number of polysaccharides containing fractions isolated from various sources as for example,
marine algae (Ruperez et al., 2002; Rocha de Souza et al., 2007; Wang et al., 2008, 2009a),
plants (Aguirre et al., 2009; Yang et al., 2011), and even some enzymatic extracts (Je et al.,
2009) possess antioxidative activity. Some of them, in particular the sulfated polysaccharides
from marine algae such as fucoidan (Zhao et al., 2005; Rocha de Souza et al., 2007; Wang et al.,
2008, 2009b; Wang et al., 2010), sulfated galactan (Barahona et al., 2011), sulfated
polysaccharide fractions containing galactose and xylose residues as constituent sugar, and
rhamnose-rich polysaccharide fractions showed considerable antioxidative properties (Hu et al.,
2010; Yang et al., 2011).
Polysaccharides from higher plants such as arabinogalactan (Fig. 1.4) also showed
antioxidative activity (Chatterjee et al., 2011; Sinha et al., 2011a). The highly branched

11
arabinogalactan isolated from Indian medicinal plant Eugenia jambolana is esterified with
phenolic acid and possess potent dose dependent antioxidative activity (Bandyopadhyay et al.,
2012).
In addition, chemically modified macromolecules such as acetylated and benzoylated ulvans
from Ulva pertusa (Chlorophyta) also possess antioxidative property. Potency of some of these
compounds is higher than the others (Qi et al., 2006).
Structure-activity relationships. The antioxidative property of polysaccharides depends upon a
number of parameters including molecular mass (Qi et al., 2005). In case of sulfated
polysaccharides, potency depends upon their sulfate content (Qi et al., 2005; Hu et al., 2010).
For example, sulfated polysaccharides from Undaria pinnitafida had stronger antioxidant
abilities than their de-sulfated derivatives (Wang et al., 2008). Recent study showed that the
position of sulfate group is another important parameter. The increased hydrophobic character of
Fig. 1.4. A model showing the structural features of an arabinogalactan from the Indian medicinal plant Eugenia jambolana (Bandyopadhyay et al., 2012).
12
the macromolecule also leads to higher potency (Qi et al., 2006). Finally, the structure of
polysaccharide also plays an important role in antioxidative property.
1-4.4. Antitumor activity
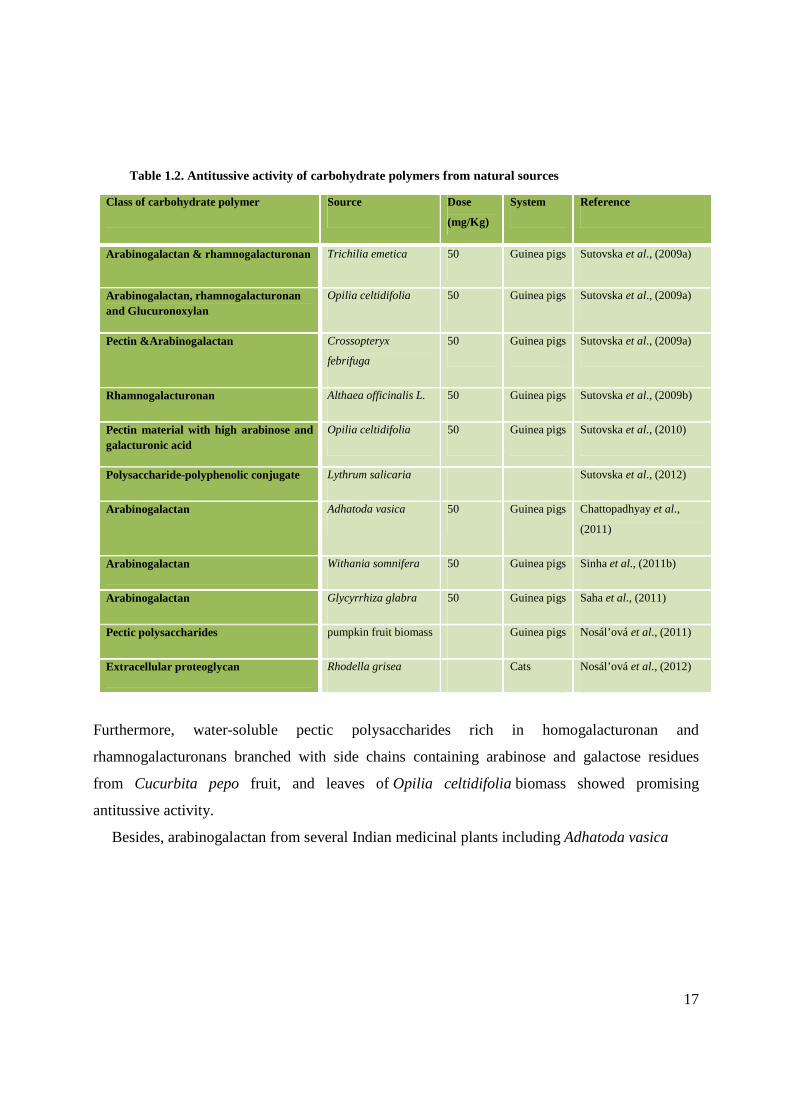
Polysaccharides also possess antitumor activity (Table 1.1). These macromolecules usually
have low toxicity and few side effects, which is essential for immunotherapy against cancer.
Class of carbohydrate polymer
Sources Type of source Reference
ββββ-D glucan Lentinus edodes Fungi Chihara et al., 1970; Ooi & Liu, 2000
ββββ-D glucan Schizophyllum commune Fungi Ooi & Liu, 2000
ββββ-D glucan Ganoderma lucidum Fungi Boh et al., 2007; Chan et al., 2009
ββββ-glucan Thamnolia vermicularis Lichen Olafsdottir et al., 2003
Protein-bound β-glucan Coriolus versicolor Fungi Ooi & Liu, 2000, Hobbs, 2004
Glucomannan Agaricus blazei Fungi Mizuno et al., 1999; Tsuchida et al., 2001
Galactomannans coffee Simões et al., 2010
Glucurono-gluco-galactomannan
Aureobasidium pullulans Bacteria Kataoka-Shirasugi et al., 1994
Fucogalactomannan Grifola frondosa Fungi Zhuang et al., 1994
Acidic glycoprotein Flammulina velutipes (W. Curt.: Fr.) Singer
Fungi Maruyama & Ikekawa, 2007
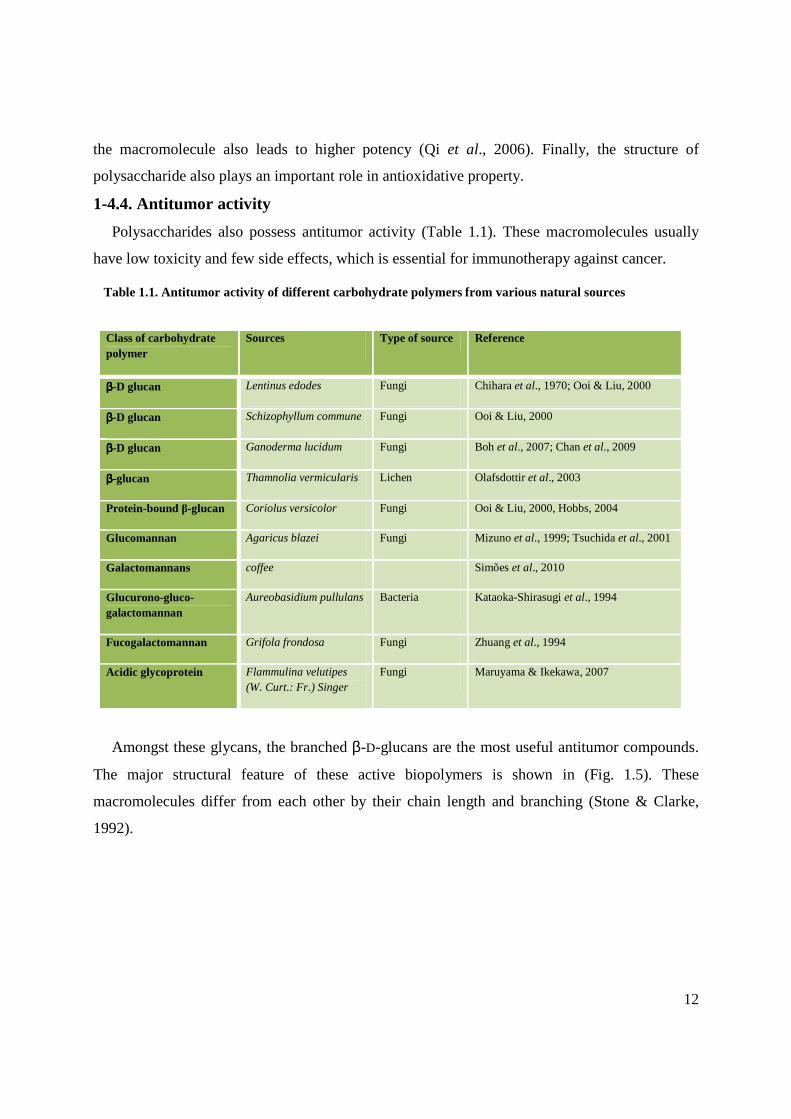
Amongst these glycans, the branched β-D-glucans are the most useful antitumor compounds.
The major structural feature of these active biopolymers is shown in (Fig. 1.5). These
macromolecules differ from each other by their chain length and branching (Stone & Clarke,
1992).
Table 1.1. Antitumor activity of different carbohydrate polymers from various natural sources

13
Fig. 1.5. Molecular core of β-D glucan.
OHO
OOH
O
OH
OHO
OH
OHO
OHO
OH
O*
O
HO
HO
HO
O
OH
n
However, the branches in the glycosidic chain are highly variable and the two main branching
are (1→4) or (1→6) glycosidic chains. Some naturally occurring β-glucans mainly Lentinans,
Schizophyllan, PSK (Krestin) are of particular clinical interest. Lentinan, produced from Shiitake
mushroom, Lentinus edodes, is a β-(1→3)-D-glucan having two β-(1→6)-glucopyranoside
branches for every five β-(1→3)-glucopyranoside linear linkages (Sasaki & Takasuka, 1976;
Saito et al., 1977, 1979). Schizophyllan from the mushroom of Schizophyllum commune consists
of β-(1→3)-D-glucan having one β-(1→6)-D-glucopyranoside branching for every three β-
(1→3)-D-glucopyranoside in the main chain. On the otherhand PSK has a basic β-glucan
structure with β-(1→6)-glucopyranosyl side chains every fourth glucose unit (Fisher & Yang,
2002). The PSK branched structure includes (1→3), or (1→4) and (1→6) bonds, and branches at
3- and 6-positions in a proportion of one per several residual groups of 1→4 bonds (Wasser,
2002; Boh et al., 2007).
Among acidic polysaccharides of natural origin the alginate (Ye et al., 2008), carrageenan
(Zhou et al., 2004), fucoidan (Usui, 1980; Maruyama et al., 2006; Synytsya et al., 2010), pectin
(Edenharder et al., 1995; Hensel & Meier, 1999; Inngjerdingen et al., 2007) etc. have been
examined for their antitumor activities. Alginic acid is one of the best-studied polymers with
pronounced activities against various tumors. It is a linear, high molecular weight glycouronan in
which three distinct types of segment are recognized (Painter, 1983; Kennedy & White, 1988).
These are blocks of β-(1→4)-D-mannuronic acid residues (MM block) and its epimer, α- (1→4)-
L-guluronic acid residues (GG block), and blocks which contain approximately equal quantities
of both types of uronic acid residue which may be randomly distributed or show some alternating

14
sequence. Other studies have explored the antitumor properties of red, brown and green algal
polysaccharides (Zhuang et al., 1995). For example, carrageenan, a family of sulfated galactan,
has shown to be active angiogenesis inhibitors.
Structure-activity relationships. Molecular weight, degree of branching, number of substituents,
water solubility and solution conformation significantly affect the biological activities of β-
glucans (Adachi et al., 2002). For example, the molecular weight and a triple-helical
conformation are known to be important factors for the immune-stimulating activity of lentinan.
The triple-helical lentinan with a moderate molecular weight (5.0x105~15.0x105) exhibits higher
anticancer activities against the growth of cancer cell such as sarcoma 180 than those with too
low or too high molecular weight (Zhang et al., 2005). In aqueous solution, β-glucans undergo
conformational change into triple helix, single helix or random coils. The immune functions of β-
glucans are apparently dependent on their conformational complexity (Bohn & BeMiller, 1995).
As for example, the triple-helical lentinan exhibits distinct antitumor activities but whenever the
triple helix has been broken into single random coils, its antitumor activity decreases
significantly or disappears. Therefore antitumor activity is related to triple helix conformation,
moderate molecular weight and its combined protein (Zhang et al., 2005; Unursaikhan et al.,
2006). However, the highly branched MD-fraction from G. frondosa (MW 1 000 000– 1 200 000
dalton) exerts a high antitumor activity (Nanba et al., 1987; Kodama et al., 2003). Higher
antitumor activity seems to be correlated with higher molecular weight, lower level of branching
and greater water solubility of β-glucans (Zjawiony, 2004).
Less is known about the structure-function relationship of acidic polysaccharides, although
the polyanionic properties of these macromolecules seem to be related to their antitumor activity.
Moreover, amongst alginates having very similar molecular weight (mol. wt. Range: 2.25-2.55 x
105) polymers with a higher content of MM-block showed higher antitumor activity than those
with a lower content despite they have the same poly-anionic properties (Fujihara & Nagumo,
1993). These facts imply that besides polyanionic properties some other features are responsible
for the biological activity of alginic acid. In living system the biospecific molecular recognition
is known to be based on the lock and key concept as proposed by Emil Fischer. On the basis of
this concept bioactive polysaccharides probably possess biospecific “keys” along polymer
backbone. Selected groups on the polysaccharide backbone represent keys, which are
macromolecular chains looped into potential cell surface receptors. Conformational flexibility of

15
Fig. 1.6. Block of β-(1→4)-linked L-mannuronic acid (M) residues.
OO
H
OH
HH
HOO
H
H
OHO
OO
H
OH
HH
HO
H
H
OHO
n
O
HOH
H
H OH
H
H
OO
HOH
H
H OH
H
H
O
OH
O
O
OH
O
n
the carbohydrate polymers facilitates an optimal adaptation to the extra cellular protein. The
molecular architecture of the D-mannuronan (Fig. 1.6) block is completely different from that of
L-guluronan block (Fig. 1.7), the former being twisted, whereas the latter being rigid and buckled
(Kennedy & White, 1988).
Now, in alginic acids of different origin the length of homoglycuronan segments are different
and therefore their three dimensional structure are also different. Alginates having higher
contents of MM-block have the right degree of conformational flexibility necessary for the
polymer-cell surface receptor interaction. Therefore, conformational factors related to the
distribution and sequence of the carboxyl groups on the alginate backbone may be responsible
for their variable biological activity.
Mechanism of action. The molecular mechanism of the inhibition and promotion processes in
carcinogenesis by plant polysaccharides is poorly understood. Polysaccharides or
polysaccharide-protein complex, having antitumor activity, can stimulate effector cells like
Fig. 1.7. Block of α- (1→4)-linked L-guluronic acid (G) residues.

16
macrophases, T-lymphocytes and natural killer (NK) cells to secrete cytokines like TNF-α, IFN-
γ, IL-1β, etc. These are antiproliferative and induce apoptosis and differentiation in tumor cell.
Research data showed that schizophyllan from S. commune can bind to mRNA poly (A) tail and
thus exert antitumor activity (Karinaga et al., 2004). In animal studies, after oral administration,
the specific backbone 1→3 linear β-glycosidic chain of β-glucans cannot be digested. Most β-
glucans enter the proximal small intestine and some are captured by the macrophages. They are
internalized and fragmented within the cells, then transported by the macrophages to the marrow
and endothelial reticular system. The small β-glucans fragments are eventually released by the
macrophages and taken up by other immune cells like the circulating granulocytes, monocytes
and dendritic cells via the complement receptor (CR)-3 leading to various immune responses
(Zhou & Gao, 2002). The immune response will then be turned on; one of the actions is the
phagocytosis of the monoclonal antibody tagged tumor cells. However, β-glucans of different
sizes and branching patterns may have significantly variable immune potency.
1-4.5. Antitussive activity
Recent studies showed that various naturally occurring carbohydrate polymers from medicinal
plants (Table 1.2) affect citric acid-induced cough reflex and reactivity of airways smooth
muscle in vivo conditions. For example, the polysaccharide materials from the leaves of popular
Malian medicinal plants Trichilia emetic (TE) and Opilia celtidifolia (OC), and fruits of
Crossopteryx febrifuga (CF) were able to suppress experimentally induced cough reflex in
guinea pigs. These crude polysaccharides are made up of arabinogalactan (~54%) and
rhamnogalacturonan (~30%) in T. emetic leaves, arabinogalactan (~60%), rhamnogalacturonan
(~14%) and glucuronoxylan (~14%) in O. celtidifolia leaves, and pectic polysaccharides (~75%)
together with arabinogalactan (~17%) in C. febrifuga fruits (Sutovska et al., 2009a). The fruits
extract of C. febrifuga was not active in the dose of 50 mg/kg b.w., however 10–20 fold higher
doses suppressed cough attack and also showed bronchodilatory properties. In addition, the
rhamnogalacturonan from the roots of Althaea officinalis L. showed potent biological effects on
the citric acid-induced cough reflex and reactivity of airways smooth muscle in vitro and in vivo
conditions. It possessed dose-dependent cough suppression effect comparable with opioid
agonist codeine (Sutovska et al., 2009b).
17
Class of carbohydrate polymer Source Dose
(mg/Kg)
System Reference
Arabinogalactan & rhamnogalacturonan Trichilia emetica 50 Guinea pigs Sutovska et al., (2009a)
Arabinogalactan, rhamnogalacturonan and Glucuronoxylan
Opilia celtidifolia 50 Guinea pigs Sutovska et al., (2009a)
Pectin &Arabinogalactan Crossopteryx
febrifuga
50 Guinea pigs Sutovska et al., (2009a)
Rhamnogalacturonan Althaea officinalis L. 50 Guinea pigs Sutovska et al., (2009b)
Pectin material with high arabinose and galacturonic acid
Opilia celtidifolia 50 Guinea pigs Sutovska et al., (2010)
Polysaccharide-polyphenolic conjugate Lythrum salicaria Sutovska et al., (2012)
Arabinogalactan Adhatoda vasica 50 Guinea pigs Chattopadhyay et al.,
(2011)
Arabinogalactan Withania somnifera 50 Guinea pigs Sinha et al., (2011b)
Arabinogalactan Glycyrrhiza glabra 50 Guinea pigs Saha et al., (2011)
Pectic polysaccharides pumpkin fruit biomass Guinea pigs Nosál’ová et al., (2011)
Extracellular proteoglycan Rhodella grisea Cats Nosál’ová et al., (2012)
Furthermore, water-soluble pectic polysaccharides rich in homogalacturonan and
rhamnogalacturonans branched with side chains containing arabinose and galactose residues
from Cucurbita pepo fruit, and leaves of Opilia celtidifolia biomass showed promising
antitussive activity.
Besides, arabinogalactan from several Indian medicinal plants including Adhatoda vasica
Table 1.2. Antitussive activity of carbohydrate polymers from natural sources

18
(Chattopadhyay et al., 2011), Withiania somnifera (Sinha et al., 2011b) and Glycyrrhiza glabra
(Saha et al., 2011) showed in vivo antitussive activities. This polysaccharide is consisted mainly
of (1,3)/-(1,6)/-(1,3,6) linked galactopyranosyl and (1,5)/-(1,3,5) linked arabinofuranosyl
residues.
Other natural macromolecules, which are not composed entirely of sugar units, contain blocks
of monosaccharide units as part of the molecular structure, and contribute extensively to their
antitussive activities. For example, a low molecular mass arabinogalactan-protein (AGP) from
the instant coffee powder of Coffea arabica beans, possess prominent antitussive (in vivo)
activity in a dose dependant way (Nosál’ová et al., 2011).
The mucilagineous extracellular proteoglycan (EPG) from culture medium of red alga
Rhodella grisea, which contained xylose and its 3-O-and 4-O-methyl-derivates (55%),
glucuronic acids (17%), rhamnose (14%), galactose (8%), glucose (4%) & contained protein
(13%) & minor amounts of other sugars (∼2%) also showed a cough suppressing effect on
laryngopharyngeal type of cough (Nosáľová et al., 2012).
The high molecular mass polysaccharide-polyphenolic conjugate (rhamnogalacturonan
associated with arabinogalactan in Lythrum conjugate) from flowering parts of Lythrum
salicaria that contains 74% of carbohydrates and 17% of phenolics showed antitussive activity
(Sutovska et al., 2012). The polyphenolic–polysaccharide–protein complex (molar mass
11.2 kDa) from flowers of Solidago canadensis L. composed of carbohydrates (43%), protein
(27%), phenolics (12%), uronic acids (10%) and inorganic material (8%) showed antitussive
activity (Sutovska et al., 2013).
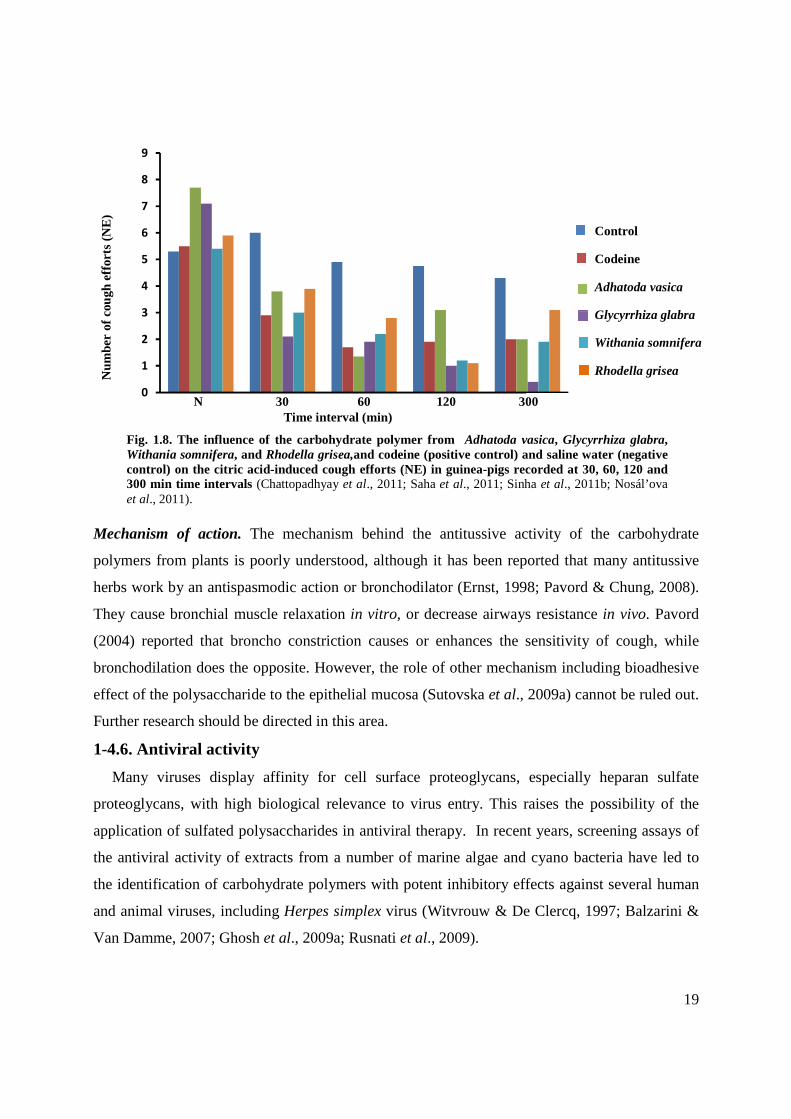
Structure-activity relationships. The antitussive properties of polysaccharide from various
sources are different (Fig.1.8), but because of limited data available the relationship between the
structure of polysaccharides and their antitussive activity is not yet clear (Sutovska et al., 2010;
Nosáľová et al., 2011).
19
Mechanism of action. The mechanism behind the antitussive activity of the carbohydrate
polymers from plants is poorly understood, although it has been reported that many antitussive
herbs work by an antispasmodic action or bronchodilator (Ernst, 1998; Pavord & Chung, 2008).
They cause bronchial muscle relaxation in vitro, or decrease airways resistance in vivo. Pavord
(2004) reported that broncho constriction causes or enhances the sensitivity of cough, while
bronchodilation does the opposite. However, the role of other mechanism including bioadhesive
effect of the polysaccharide to the epithelial mucosa (Sutovska et al., 2009a) cannot be ruled out.
Further research should be directed in this area.
1-4.6. Antiviral activity
Many viruses display affinity for cell surface proteoglycans, especially heparan sulfate
proteoglycans, with high biological relevance to virus entry. This raises the possibility of the
application of sulfated polysaccharides in antiviral therapy. In recent years, screening assays of
the antiviral activity of extracts from a number of marine algae and cyano bacteria have led to
the identification of carbohydrate polymers with potent inhibitory effects against several human
and animal viruses, including Herpes simplex virus (Witvrouw & De Clercq, 1997; Balzarini &
Van Damme, 2007; Ghosh et al., 2009a; Rusnati et al., 2009).
0
1
2
3
4
5
6
7
8
9
Num
ber
of c
ough
effo
rts
(NE
)
Control
Codeine
Adhatoda vasica
Glycyrrhiza glabra
Withania somnifera
Rhodella grisea
Fig. 1.8. The influence of the carbohydrate polymer from Adhatoda vasica, Glycyrrhiza glabra, Withania somnifera, and Rhodella grisea,and codeine (positive control) and saline water (negative control) on the citric acid-induced cough efforts (NE) in guinea-pigs recorded at 30, 60, 120 and 300 min time intervals (Chattopadhyay et al., 2011; Saha et al., 2011; Sinha et al., 2011b; Nosál’ova et al., 2011).
N 30 60 120 300 Time interval (min)
20
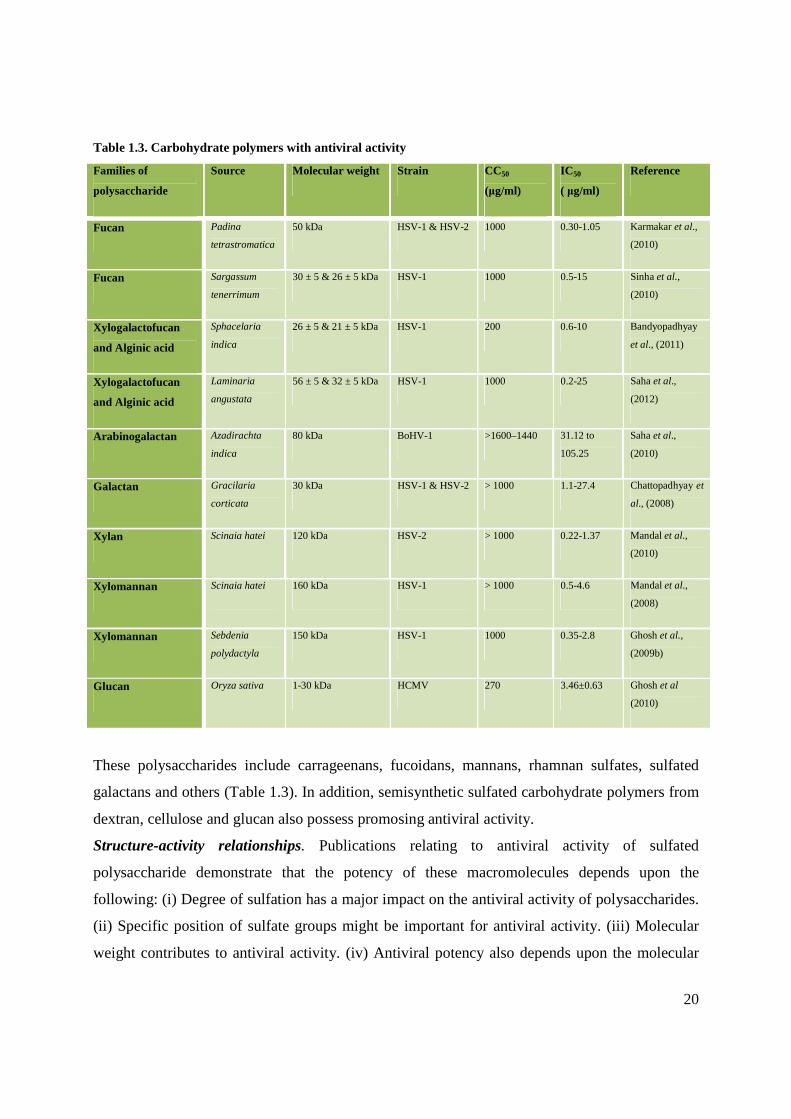
Table 1.3. Carbohydrate polymers with antiviral activity
Families of
polysaccharide
Source Molecular weight Strain CC50
(µg/ml)
IC 50
( µg/ml)
Reference
Fucan Padina
tetrastromatica
50 kDa HSV-1 & HSV-2 1000 0.30-1.05 Karmakar et al.,
(2010)
Fucan Sargassum
tenerrimum
30 ± 5 & 26 ± 5 kDa HSV-1 1000 0.5-15 Sinha et al.,
(2010)
Xylogalactofucan
and Alginic acid
Sphacelaria
indica
26 ± 5 & 21 ± 5 kDa HSV-1 200 0.6-10 Bandyopadhyay
et al., (2011)
Xylogalactofucan
and Alginic acid
Laminaria
angustata
56 ± 5 & 32 ± 5 kDa HSV-1 1000 0.2-25 Saha et al.,
(2012)
Arabinogalactan Azadirachta
indica
80 kDa BoHV-1 >1600–1440 31.12 to
105.25
Saha et al.,
(2010)
Galactan Gracilaria
corticata
30 kDa HSV-1 & HSV-2 > 1000 1.1-27.4 Chattopadhyay et
al., (2008)
Xylan Scinaia hatei 120 kDa HSV-2 > 1000 0.22-1.37 Mandal et al.,
(2010)
Xylomannan Scinaia hatei 160 kDa HSV-1 > 1000 0.5-4.6 Mandal et al.,
(2008)
Xylomannan Sebdenia
polydactyla
150 kDa HSV-1 1000 0.35-2.8 Ghosh et al.,
(2009b)
Glucan Oryza sativa 1-30 kDa HCMV 270 3.46±0.63 Ghosh et al
(2010)
These polysaccharides include carrageenans, fucoidans, mannans, rhamnan sulfates, sulfated
galactans and others (Table 1.3). In addition, semisynthetic sulfated carbohydrate polymers from
dextran, cellulose and glucan also possess promosing antiviral activity.
Structure-activity relationships. Publications relating to antiviral activity of sulfated
polysaccharide demonstrate that the potency of these macromolecules depends upon the
following: (i) Degree of sulfation has a major impact on the antiviral activity of polysaccharides.
(ii) Specific position of sulfate groups might be important for antiviral activity. (iii) Molecular
weight contributes to antiviral activity. (iv) Antiviral potency also depends upon the molecular

21
structure of the polysaccharide. (v) Low-molecular weight compounds inhibit cell-to-cell spread
of viruses more efficiently than high molecular weight compounds (Ekblad et al., 2010).
Mechanism of action. The entry of HSV into host cells is a complex process initiated by the
specific interaction between host-cell-surface receptors and viral envelope glycoproteins
(Schneider-Schaulies, 2000; Spear, 2004; Kleymann, 2005; Olofsson & Bergstrom, 2005). In the
case of HSV-1 and HSV-2, attachment to HS seems to be primarily mediated through
glycoprotein C (gC) although glycoprotein B (gB) may contribute to this function (Herold et al.,
1991, 1994; Cheshenko & Herold 2002). Clusters of basic and hydrophobic amino acids located
between residues 129 and 160 of gC1 (Trybala et al., 1994; Mardberg et al., 2001, 2002) as well
as the mucin-like region (amino acids 33–123) of this protein (Tal-Singer et al., 1995) were
identified as important for HSV-1 attachment to cell-surface HS/CS. HSV-1 gB1 consists of 904
amino acids and approximately 85% of the sequence is homologous to its HSV-2 counterpart
(Heldwein et al., 2006). Most of the variability between gB1 and gB2 is seen in a lysine-rich
region (amino acids 68–76), which is also responsible for binding to HS (Laquerre et al., 1998).
gB1 occurs as a trimer with each of the monomers divided in five distinctive domains: I-base, II-
middle, III-core, IV-crown, and V-arm (Heldwein et al., 2006). The results from a more recent
study suggest that specific hydrophobic/aromatic amino acids from domain I are important for
the fusogenic activity of gB (Hannah et al., 2007). An attractive concept is that sulfated
polysaccharides act as antiviral agents in cell culture due to the fact that these charged polymers
may mimic HS chains on cell-surface proteoglycans and thus block viral attachment by
competitive inhibition.
A novel approach to inhibit HSV-1 infection by targeting the gD-mediated membrane fusion
step has been described (Copeland et al., 2008). The antiherpetic properties of sulfated
polysaccharides may depend not only on their charge density but also on the characteristics of
their uncharged portions which may be involved in hydrophobic and hydrogen bonding
interactions. Mardberg and co-workers (2001) reported that hydrophobic interactions, in addition
to electrostatic forces, are decisive for the CS as well as HS binding to viral glycoprotein gC.
The interaction of the methyl groups of fucoidan with the hydrophobic pocket of HSV-1 gC
seems to be important in the binding of the polysaccharide to the viral glycoprotein. Finally, in
addition to the polysaccharide-mediated antiviral effects directed to the cell surface (viral
receptor binding, entry, fusion); a second type of effects may play a role, i.e., the induction of

22
intracellular events contributing to the antiviral activity of sulfated polysaccharides. As the
binding of a number of known polysaccharides to cell-surface receptors can induce intracellular
signaling pathways, this second type of effects should be additionally taken into consideration.
As an example, the anticytomegalo viral effect of spirulan-like polysaccharides was
demonstrated to be composed of these two antiviral activities, i.e., an inhibition of HCMV entry
on the one side in addition to the induction of intracellular antiHCMV effects on the other side
(Rechter et al., 2006).The replication efficiency of most viruses is dependent on specific
intracellular signaling pathways, the inhibition or the induction of particular signaling by
surface-binding polysaccharides can provide a significant part of the overall antiviral activity.
One explanation for such intracellularly produced activity is the stimulatory effect of sulfated
polysaccharides onto interferon production with the consequence of a broad antiviral effect.
1-4.7. Potential clinical applications
During the past decades many significant developments in the utilization of carbohydrate
polymers as drug have been put forward. This, however, is not surprising since it is known that
many of these biopolymers play an essential role in key biological processes. Some of the new
cases of polysaccharide applications have been approved by the scientific world; others are still a
matter of controversies and hence are not being accepted for clinical approval (Witezak, 1995).
Many fungal and plant derived bioactive polysaccharides with a broad range of
immunomodulatory activities are found in traditional medicine. Some such polysaccharides have
been developed into drugs and show clinical efficacy in controlled trials while the majority of
such compounds remain as nutraceuticals with only preliminary research.
Three polysaccharide based carcinostatic (immunotherapeutic) agents, Krestin, Lentinan and
Sonifilan, have already been developed from mushroom (Borchers et al., 2004; Zhang et al.,
2011; Maehara et al., 2012). These are used currently in the treatment of cancer of the digestive
organs, lung and breast, as well as cancer of the stomach and cervical cancer, ovarian cancer
(Fujimoto et al., 2006), gastric or colorectal cancer, (Maehara et al., 2012) respectively. The
potential usefulness of bioactive polysaccharides in the treatment of diseases has been
demonstrated in preclinical and clinical studies on β-glucan. Such polysaccharides are generally
nontoxic and possess other bioactivities such as inducing differentiation, stimulating
hematopoiesis, antimetastasis, and antiangiogenesis, which make them ideal adjuvants in modern

23
cancer therapy. Again, other clinical experiments have showed the potential application of
lentinan on anti-HIV activity (Gordon et al., 1995, 1998).
For more than 70 years, heparin is a drug of choice in the prevention and treatement of
thromboembolic disorders (Linhardt et al., 1997; Gunay & Linhardt, 1999; Alban & Franz,
2001; Lee et al., 2008; Fan et al., 2011). In addition to heparin, partially depolymerized forms
called low molecular weight (LMW) heparins and a synthetic heparin pentasaccharide are
currently in clinical use (Murugesan et al., 2008). The subcutaneous injection of the LMW
heparins for the treatment of deep venous thrombosis has made anticoagulant therapy more
effective and easier to administer, making it possible to treat the patients without hospitalization
and at no increased risk of recurrent thromoembolism or bleeding complications (Rydberg et al.,
1999).
Although many polysaccharides and their functions have been described in this chapter, the
structure–function relationship remains to be elucidated. A great deal of fascinating chemistry
remains to be discovered. Elucidation of this interrelationship is the main objective of this work.
This research work has been presented into three parts. The first part provides an interesting
story about the structural features and antioxidant capacity of polysaccharides from the brown
seaweed Turbinaria conoides. The second part is devoted to structural features and antitussive
activity of water extracted polysaccharide from Adhatoda vasica. The final chapter describes the
materials and methods used in this study.

24
The goals of this research are to identify the polysaccharides present in Indian samples of
Turbinaria conoides and Adhatoda vasica and to study their biological activities.
The strategy adopted to achieve these goals involves:
� Isolation of the high molecular weight compounds.
� Their purification.
� Screening of the biological activities of the crude extracts as well as that of pure
compounds.
� Finally, determination of the structural features of the biologically active high
molecular weight compounds.
AIMS

RESULTS AND DISCUSSIONS

26
2
STRUCTURAL FEATURES AND ANTIOXIDATIVE
ACTIVITIES OF CARBOHYDRATE
POLYMERS FROM THE BROWN SEAWEED Turbinaria conoides

27
2-1. INTRODUCTION
Reactive oxygen species (ROS) are highly reactive molecules that are constantly produced by
enzymatic reactions in cells. In normal physiological conditions, ROS are produced at low levels,
which are necessary for maintaining normal cell functions and the endogenous antioxidative
defense systems of the body have the capacity to avert any harmful effects. However, free
radicals can escape from cellular defense system, leading to modification of DNA, proteins,
lipids and small cellular molecules are associated with a number of pathological processes,
including atherosclerosis, cancer and rheumatoid arthritis (Halliwell & Gutteridge, 1984).
Therefore, antioxidants are important for bodily protection against oxidative stress. Lipid
oxidation by reactive oxygen species (ROS) such as super oxide anion, hydroxyl radicals and
hydrogen peroxide also causes a decrease in nutritional value of lipids, in their safety and
appearance. In addition, it is the predominant cause of qualitative decay of foods, which leads to
rancidity, toxicity and destruction of biochemical components important in physiologic
metabolism. Recently, there is a considerable interest in the food industry and in the preventive
medicine for the development of antioxidants from natural sources, such as marine flora and
fauna, bacteria, fungi and higher plants. Among them, marine algae represent one of the richest
sources of bioactive compounds, and algae-derived products are increasingly used in medical
and biochemical research (Mayer & Lehmann, 2000). One particularly interesting feature of
marine algae is their richness in Sulfated polysaccharides, the uses of which span from food,
cosmetic and pharmaceutical industries to microbiology and biotechnology (Renn, 1997). These
macromolecules have been proven to show a wide range of biological activities important to
human health, for example, antiviral, antitumoral, antiinflammatory and anticoagulant activity
(Cumashi et al., 2007; Pomin & Mourao, 2008; Ghosh et al., 2009a).
In recent years, several classes of sulfated polysaccharides have been demonstrated to show
antioxidative activity, too. The compounds tested included glucan, alginic acid, fucoidan and
other unidentified macromolecules present in the extracts (Ruperez et al., 2002; Rocha de Souza
et al., 2007; Wang et al., 2008). Indian coastal area is inhabited by variety of marine algae which
are not yet to be fully explored and exploited for socio-economic development of the nation
(Wealth of India, 1985). Turbinaria conoides (Fucales, Sargassaceae) is one such seaweed,
which is widely distributed in the coastal areas of Andaman and Nicobar island, Laccadives,
Maharastra, Kerala, Tamilnadu, Gujrat, Andhra Pradesh (Sahoo, 2010). This brown alga contains

28
phytochemicals of great interest to researchers including highly cytotoxic hydroperoxysterol 24-
hydroperoxy-24-vinylcholesterol and fucosterol (Sheu & Sung, 1991), 24 ξ-hydroperoxy-24-
vinylcholesterol; 29-hydroperoxystigmasta-5,24(28)- dien-3β-ol; 24-ethylcholesta-4,24(28)-dien-
3-one; 24 ξ-hydroperoxy-24-ethyl cholesta-4, 28(29)-dien-3-one; 24-ethyl cholesta-4,24(28)-
dien-3,6-dione; 24 ξ-hydro peroxy-24-ethyl cholesta-4,28(29)-dien-3,6-dione; 6β-hydroxy-24-
ethyl cholesta-4,24(28)-dien-3-one; 24 ξ-hydroperoxy-6 β-hydroxy-24-ethylcholesta-4,28(29)-
dien-3-one (Sheu et al., 1999). The presence of two new antifungal steroids namely, 3,6,17-
trihydroxy-stigmasta-4,7,24(28)-triene and 14,15,18,20-diepoxy turbinarin have also been
reported (Kumar et al., 2010, 2011). Despite the general interest in phytoconstituents it remains
ironic that research on the chemical and biological aspects of the high molecular weight
bioactive has been neglected. Notably, it is intriguing to observe that polysaccharide from plant
extracts exhibit a large range of pharmacological activities (Inngjerdingen et al., 2006;
Mantovani et al., 2008; Ghosh et al., 2009a; Baek et al., 2010; Thakur et al., 2012).
Hence the central goal of this study was to investigate the structural features of the different
classes of polysaccharides present in T.conoides. We have also evaluated in vitro the
antioxidative activity of fucoidan, glucan and alginic acid isolated from this brown seaweed.
These polysaccharides may represent a new approach for inhibiting the harm caused by
excessive free radicals.
2-2. CHEMICAL CHARACTERIZATION OF POLYSACCHARIDES
FROM Turbinaria conoides
2-2.1. Preparation of depigmented algal power and sugar compositional analysis
The major goal of this study was to develop antioxidative drug candidate from Turbinaria
conoides. In order to study the chemical structures and antioxidative properties of polymers
present in T. conoides, the dried seaweed was sequentially extracted in a Soxhlet apparatus with
petroleum ether and acetone to leave a depigmented algal powder named as DAP (Fig. 2.1).
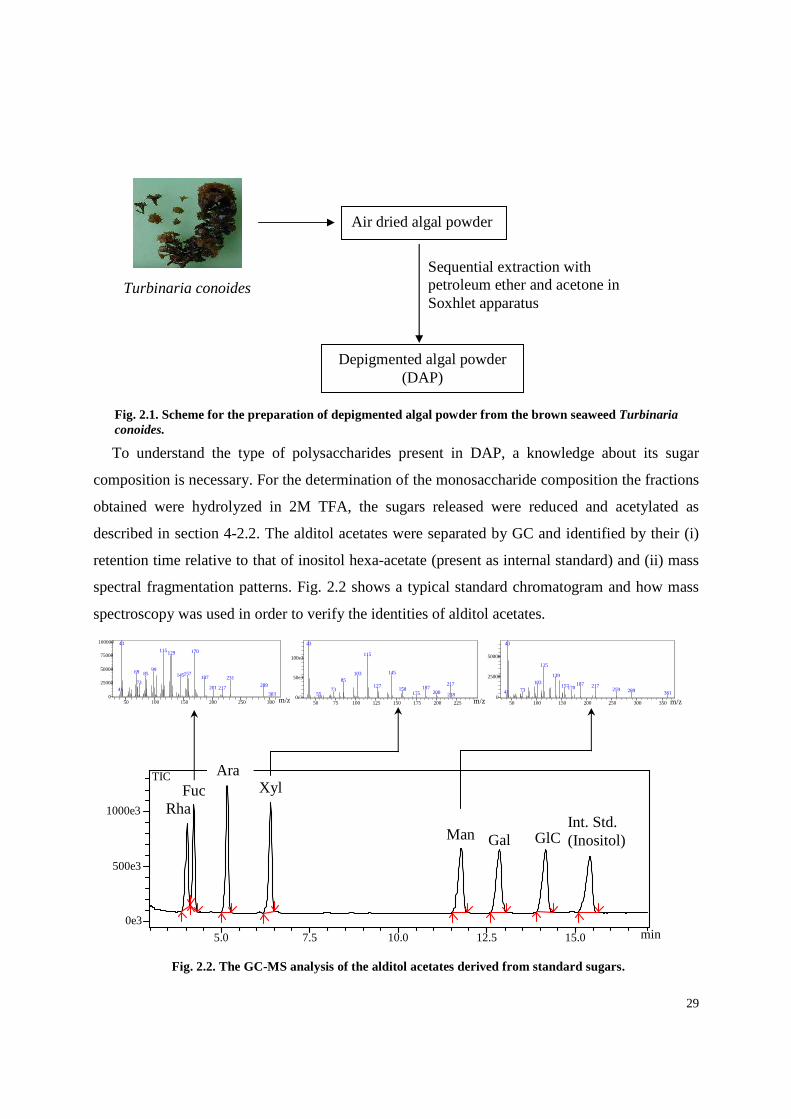
29
To understand the type of polysaccharides present in DAP, a knowledge about its sugar
composition is necessary. For the determination of the monosaccharide composition the fractions
obtained were hydrolyzed in 2M TFA, the sugars released were reduced and acetylated as
described in section 4-2.2. The alditol acetates were separated by GC and identified by their (i)
retention time relative to that of inositol hexa-acetate (present as internal standard) and (ii) mass
spectral fragmentation patterns. Fig. 2.2 shows a typical standard chromatogram and how mass
spectroscopy was used in order to verify the identities of alditol acetates.
Fig. 2.2. The GC-MS analysis of the alditol acetates derived from standard sugars.
min
m/z m/z m/z
Gal GlC Man Int. Std. (Inositol)
Xyl Fuc Rha
50 100 150 200 250 3000
25000
50000
75000
100000 43115 170129
9969 85 157145 187 23173 28920121741
303
50 75 100 125 150 175 200 2250e3
50e3
100e3
43
115
14510385
217127 18715873 20017555 218
50 100 150 200 250 300 3500
25000
50000
43
115
139103 187 217157170 25973 28941 361
5.0 7.5 10.0 12.5 15.0
0e3
500e3
1000e3
TIC Ara
Fig. 2.1. Scheme for the preparation of depigmented algal powder from the brown seaweed Turbinaria conoides.
Depigmented algal powder (DAP)
Sequential extraction with petroleum ether and acetone in Soxhlet apparatus
Air dried algal powder
Turbinaria conoides

30
GC analysis of the alditol acetates derived from DAP (Fig. 2.3) showed the presence of fucose
(56%), xylose (11%), galactose (15%), glucose (12%) and small amount of mannose (Table 2.1).
2-2.2. Isolation and sugar composition of polysaccharide fractions
Sugar composition of depigmented algal powder (DAP) revealed that, it contained 35% 2 M
TFA hydrolysable sugars of which about seventh were uronic acids. The main neutral sugar was
fucose and it also contained other sugar including glucose, galactose
5.0 7.5 10.0 12.5 15.0
0.0e6
5.0e6
10.0e6
15.0e6
20.0e6 T IC
Fig. 2.3. GC analysis of alditol acetates derived from DAP of Turbinaria conoides.
Fu
cose
Xyl
ose
Man
nose
Gal
acto
se
Glu
cose
Int.
Std
. (In
osi
tol)
min
SEC
Algal Powder (DAP)
0.1 M HCl, 30-350C
A
AF1
AF2
AF3 Residue
3% Aqueous Na2CO3 Extract B
2% CaCl2
Residue
INS
C
2% CaCl2
Precipitate (C2)
Solution (C1)
AEC
Water, pH~6.5, 800C
C1F1 C1F2
Fig. 2.4. Scheme for the extraction and purification of polysaccharides from the marine brown alga T. conoides.

31
and xylose (Table 2.1). The DAP was fractionated by the sequential extraction procedure as
shown in Fig. 2.4, was based on the different solubility, molecular mass and charge distribution
of polysaccharides from brown seaweeds. Glucans are soluble in warm waters and therefore
extracted at 80°C. Fucoidans were extracted with diluted hydrochloric acid, whereas alginates
were extracted with sodium carbonate. Alginates form insoluble precipitates with bivalent
calcium and at acidic pH, and are soluble in solution between pH 6 and 9. Procedures described
in the materials and methods (Section 4-1.1.).
GC and GC-MS analysis of the alditol acetates derived from A (Fig. 2.5) revealed the
presence of fucose as the major monosaccharide together with smaller amounts of xylose and
galactose (Table 2.1). No amino sugars were detected during GC-MS analysis of the derived
alditol acetates.
50 100 150 200 250 3000
25000
50000
75000
100000 43115 170129
9969 85 157145 187 23173 289201 21741
303
5.0 7.5 10.0 12.50.0e6
1.0e6
2.0e6
3.0e6
4.0e6 TIC
Gal
acto
se
Int.
Std
. (I
nos
itol)
Fu
cose
Xyl
ose
m/z
min
Fig. 2.5. GC-MS analysis of the alditol acetate derived from the fraction A of T. conoides.

32
Table 2.1. Yield and chemical composition (mol %) of fractions obtained from T. conoides
*percent weight of fraction dry weight.†mol percent of anhydro sugar - = not determined. nd = not detected. Tr = trace. TS = Total sugar
2-2.3. Purification of the fucoidans by chromatography
Anion exchange chromatography
on a DEAE Sepharose column
separated fraction A of T.
conoides into three sub-fractions
(AF1, AF2, and AF3) (Fig. 2.4).
AF1, which accounted for 11% of
the total sugars recovered from
the anion exchanger, was the
minor component of A. It
consisted mostly of fucose
together with smaller amount of
galactose, xylose and glucose (Table 2.1). The second fraction AF2, eluted at the beginning of
the salt gradient, is the second largest fraction. AF3 was the major fraction, amounting to 51% of
the total polymers recovered from the column. Fucose, accounted for more than 54% of the total
sugars of AF3 together with smaller amount of galactose and xylose units (Table 2.1). Thin layer
chromatographic analysis of the monosaccharides present in the hydrolysate indicates the
presence of an uronic acid with an Rf value
similar to that of glucuronic acid. GC analysis of
the TMS derivatives of the derived methyl
glycosides confirmed this result. The presence of
glucuronic acid has already been reported in
fucoidan from brown seaweeds (Adhikari et al.,
2006; Mandal et al., 2007).Therefore, AF3 is
essentially a fucoidan that might contain a high
number of sulfate groups, as indicated by its late
elution. Indeed, the high charge density of this
polysaccharide was confirmed by its sulfate
content (4%, w/w). The FT-IR spectrum
Fractions DAP A AF3 AF3D B C1 C1F2
Yield 100 8.8 nd nd 22.6 6.2 nd
TS* 35 36 37 56 46 41 58
Sulfate* nd 3 4 Tr 1 2 Tr
Protein* nd 1.3 nd nd 2.4 2.1 nd
Fucose† 56 57 54 52 Tr 59 6
Xylose† 11 15 18 19 Tr 12 5
Mannose† 7 Tr Tr - Tr Tr Tr
Galactose† 15 23 28 29 Tr 18 -
Glucose† 12 5 - - - 11 89
Fig. 2.6. FT-IR spectrum of fucoidan (AF3) obtained from T. conoides.
Sulfate band 1252 cm-1

33
0
0.5
1
1.5
2
2.5
3
00.0
80.1
50.23 0.3
0.380.45
0.53 0.6 0.68
0.75
0.83 0.9
0.98
Fig. 2.7. Elution profile of the fucoidan (AF3) of T. conoides on Sephacryl S-200 column with 500 mM sodium acetate buffer (pH 5.0) at 20 mL/h. Elution of polysaccharide was expressed as a function of the partition coefficient Kav [Kav = (Ve - V0)/(Vt - V0) with Vt and V0 being the total and void volume of the column determined as the elution volume of glucose and blue dextran, respectively, and Ve is the elution volume of the sample].
Kav
Su
gar
Co
nce
ntr
atio
n (µg
/mL)
(Fig. 2.6) of AF3 showed an absorption band at 1252 cm-1 related to a >S=O stretching vibration
of the sulfate group. An additional sulfate absorption band at 848 cm-1 (C–O–S, secondary axial
sulfate) indicated that the sulfate group is located at position 4 of the fucopyranosyl residue
(Patankar et al., 1993).
Solvolytic desulfation of the purified (AF3) fucoidan (as pyridinium salt) produces a
desulfated derivative (AF3D). Preliminary experiments showed a higher recovery with this
method compared to methanol-HCl and auto-desulfation methods. In the IR spectrum of
desulfated derivative (AF3D) absorbances at 1252 cm−1 and 848 cm−1 become weak,
demonstrating that they were associated with sulfate groups. This purified sulfated fucan had
negative specific rotation [α]D -109o (c 0.2, H2O), revealing that fucose in AF3 belongs to the L-
series, like other sulfated fucans from brown seaweeds (Kariya et al., 2004; Adhikari et al.,
2006; Mandal et al., 2007). AF3 was subjected to further chemical analysis.
2-2.4. Molecular mass
AF3 was subjected to
further chemical analysis. First,
the apparent molecular mass
was determined by size
exclusion chromatography. The
elution profile of this
macromolecule on Sephacryl
S-200 suggests that this
polymer is homogeneous (Fig.
2.7). Based on calibration with
standard dextrans, the apparent
molecular mass of AF3 would
be 50 kDa.
2-2.5. Linkage analysis
Table 2.2 shows that the desulfated fucoidan yielded partially methylated alditol acetates
(PMAA) corresponding to (1→2)-Xylp (xylopyranose), (1→3)-linked Fucp, and (1→3)- and
(1→2,3)-linked Galp residues, consistent with the presence of fucoidan. The identity of each

34
PMAA was confirmed by their relative retention time and mass spectral fragmentation pattern.
Fig. 2.8 shows EI-MS fragmentation pattern of selected PMAA. The presence of large quantities
of T-Xylp (terminal xylopyranose) and T-Fucp residues suggest that this polymer is highly
branched with 29 terminals for every 71 residues in the main chain. Interestingly, 33–34% of the
total fucose residues are terminal, 27–28% are (1→3)-linked, 21–22% are branched points
(Table 2.2). Small amount of (1→4)- and (1→2)-linked Fucp residues were also present. So far,
fucose residues in algal fucoidans are either (1→3)- or (1→3)- and (1→4)-linked (Cumashi et
al., 2007; Pomin & Mourao, 2008). Linkage analysis of the native fucan sulfate AF3 yielded a
variety of monomethylated, dimethylated and trimethylated products (Table 2.2) indicating that
the structure of this polymer is highly complex. The amount of sulfate groups in the native
Methylation
products†
Deduced linkage Peak area* AF3 AF3D
2,3,4-Xyl Xylp(1→ 11 12
3,4-Xyl →2)Xylp(1→ 7 9
2,3,4-Fuc Fucp(1→ 8 17
2,4-Fuc →3)Fucp(1→ 3 14
2,3-Fuc →4)Fucp(1→ 3 3
3,4-Fuc →2)Fucp(1→ 5 6
2-Fuc →3,4)Fucp(1→ 17 5
3-Fuc
Fuc
→2,4)Fucp(1→
→2,3,4)Fucp(1→
2
16
6
-
3,4,6-Gal →2)Galp(1→ 6 7
2,4,6-Gal
4,6-Gal
2-Gal
→3)Galp(1→
→2,3)Galp(1→
→3,4,6)Galp(1→
10
6
6
10
6
5
polysaccharide as calculated from methylation was not in good agreement with the
experimentally determined sulfate. But, methylation of sulfated polysaccharides does not always
yield reliable proportions of methylated alditol acetates (Pereira et al., 1999; Adhikari et al.,
2006). 2-O-methyl fucitol, and unmethylated fucitol were amongst the abundant products of
methylation analysis of the native polymer. The increase in the proportions of 2,4-di-O-methyl
*Percentage of total area of the identified peaks. †2,3,4-Fuc denotes 1,5-di-O- acetyl-2,3,4- tri-O-methylfucitol, etc. - = not detected.
Table 2.2. Partially methylated alditol acetates derived from the fucoidan (AF3) of T. conoides and its desulfated derivative (AF3D)
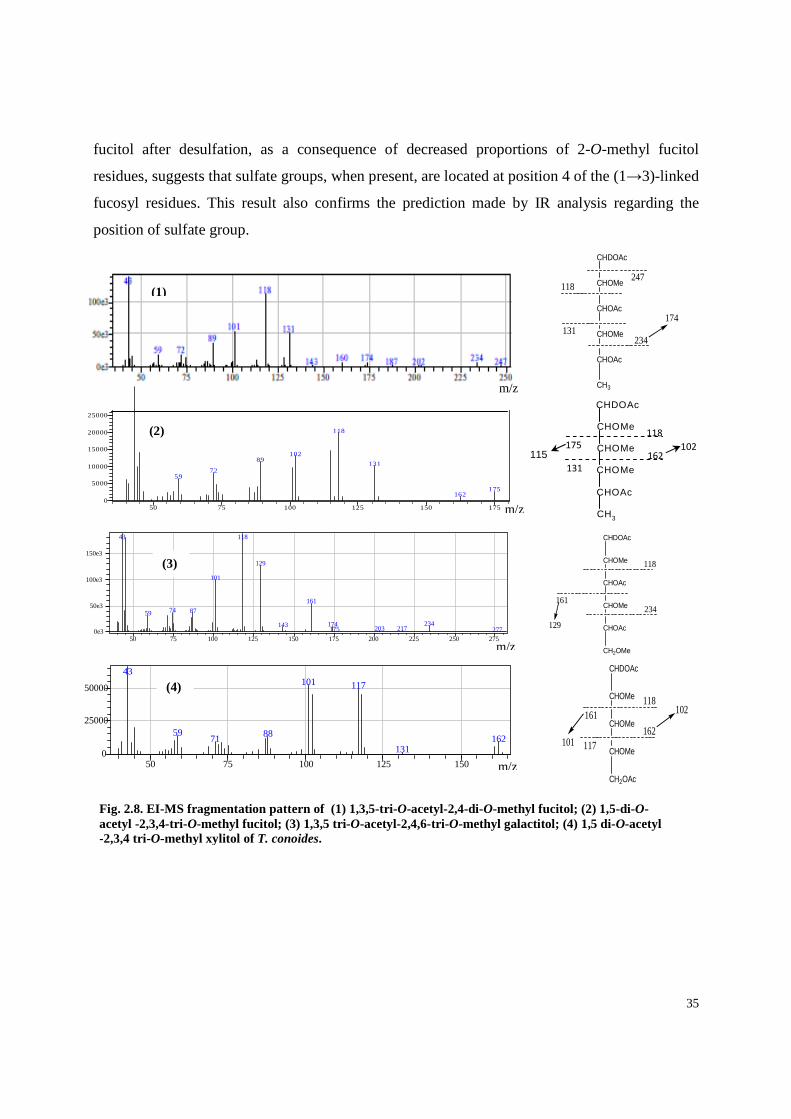
35
fucitol after desulfation, as a consequence of decreased proportions of 2-O-methyl fucitol
residues, suggests that sulfate groups, when present, are located at position 4 of the (1→3)-linked
fucosyl residues. This result also confirms the prediction made by IR analysis regarding the
position of sulfate group.
CHDOAc
CHOMe
CHOAc
CHOMe
CHOAc
CH2OMe
118
161
129
234
102
CHOAc
CHDOAc
CHOMe
CH3
CHOMe
CHOMe
118
131 162 175 115
m/z 50 75 100 125 150 1750
5000
10000
15000
20000
25000
118
10289
13172
59
175162
CHDOAc
CHOMe
CHOMe
CHOMe
CH2OAc
118161
117162
102
101
(2)
m/z
Fig. 2.8. EI-MS fragmentation pattern of (1) 1,3,5-tri -O-acetyl-2,4-di-O-methyl fucitol; (2) 1,5-di-O-acetyl -2,3,4-tri-O-methyl fucitol; (3) 1,3,5 tri-O-acetyl-2,4,6-tri-O-methyl galactitol; (4) 1,5 di-O-acetyl -2,3,4 tri-O-methyl xylitol of T. conoides.
50 75 100 125 150 175 200 225 250 2750e3
50e3
100e3
150e3
43 118
129
101
16174 8759
234174143 203 217175 277
m/z
(1)
CHDOAc
CHOMe
CHOAc
CHOMe
CHOAc
CH3
247118
131234
174
m/z
(3)
50 75 100 125 1500
25000
50000
43101 117
59 8871 162131
(4)
m/z
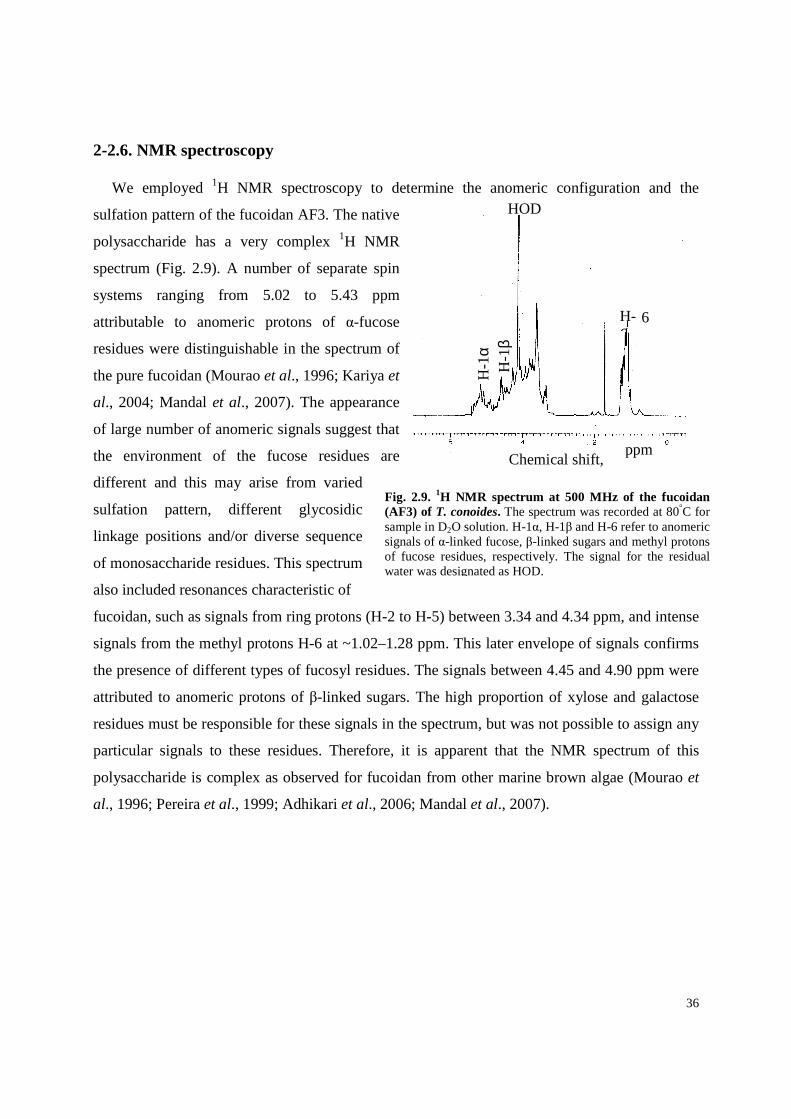
36
HOD
H-6
H-1
α H
-1β
Chemical shift,
Fig. 2.9. 1H NMR spectrum at 500 MHz of the fucoidan (AF3) of T. conoides. The spectrum was recorded at 80°C for sample in D2O solution. H-1α, H-1β and H-6 refer to anomeric signals of α-linked fucose, β-linked sugars and methyl protons of fucose residues, respectively. The signal for the residual water was designated as HOD.
6
2-2.6. NMR spectroscopy
We employed 1H NMR spectroscopy to determine the anomeric configuration and the
sulfation pattern of the fucoidan AF3. The native
polysaccharide has a very complex 1H NMR
spectrum (Fig. 2.9). A number of separate spin
systems ranging from 5.02 to 5.43 ppm
attributable to anomeric protons of α-fucose
residues were distinguishable in the spectrum of
the pure fucoidan (Mourao et al., 1996; Kariya et
al., 2004; Mandal et al., 2007). The appearance
of large number of anomeric signals suggest that
the environment of the fucose residues are
different and this may arise from varied
sulfation pattern, different glycosidic
linkage positions and/or diverse sequence
of monosaccharide residues. This spectrum
also included resonances characteristic of
fucoidan, such as signals from ring protons (H-2 to H-5) between 3.34 and 4.34 ppm, and intense
signals from the methyl protons H-6 at ~1.02–1.28 ppm. This later envelope of signals confirms
the presence of different types of fucosyl residues. The signals between 4.45 and 4.90 ppm were
attributed to anomeric protons of β-linked sugars. The high proportion of xylose and galactose
residues must be responsible for these signals in the spectrum, but was not possible to assign any
particular signals to these residues. Therefore, it is apparent that the NMR spectrum of this
polysaccharide is complex as observed for fucoidan from other marine brown algae (Mourao et
al., 1996; Pereira et al., 1999; Adhikari et al., 2006; Mandal et al., 2007).
ppm
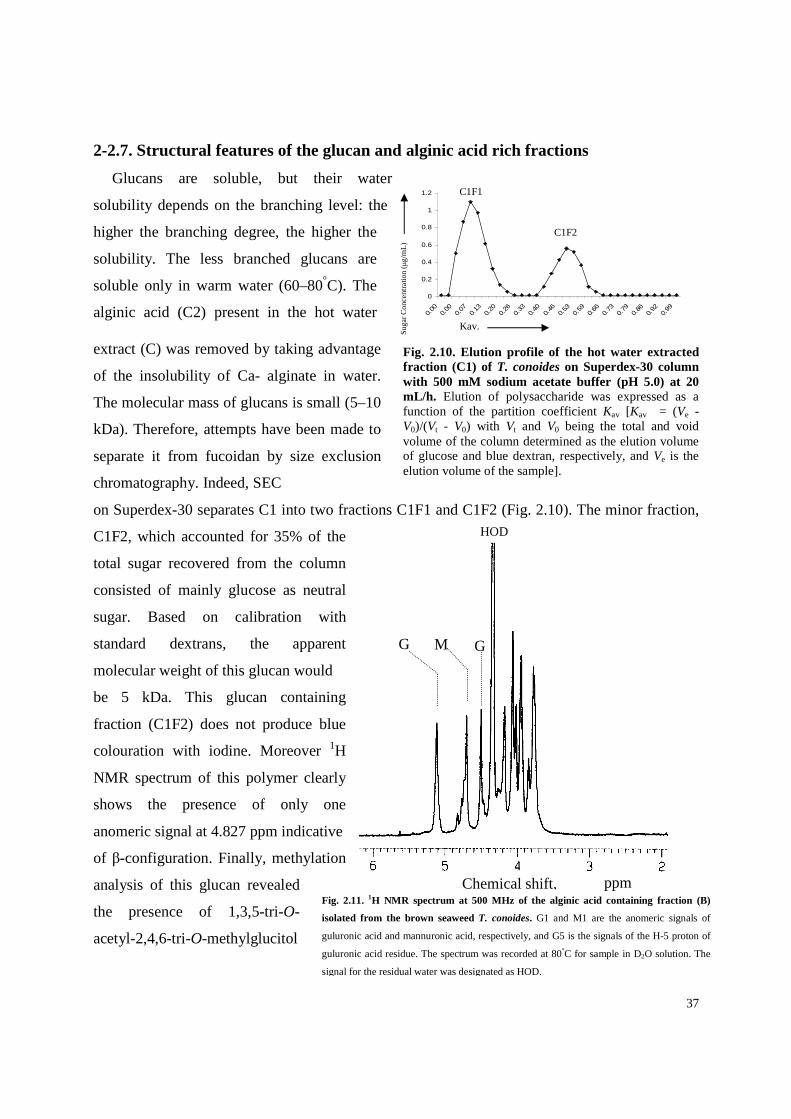
37
p
G M G
HOD
Chemical shift, ppm Fig. 2.11. 1H NMR spectrum at 500 MHz of the alginic acid containing fraction (B)
isolated from the brown seaweed T. conoides. G1 and M1 are the anomeric signals of
guluronic acid and mannuronic acid, respectively, and G5 is the signals of the H-5 proton of
guluronic acid residue. The spectrum was recorded at 80°C for sample in D2O solution. The
signal for the residual water was designated as HOD.
HOD
0
0.2
0.4
0.6
0.8
1
1.2
0.00
0.00
0.07
0.13
0.20
0.26
0.33
0.40
0.46
0.53
0.59
0.66
0.73
0.79
0.86
0.92
0.99
Sug
ar
Co
nce
ntra
tion
(µg/m
L)
Kav.
Fig. 2.10. Elution profile of the hot water extracted fraction (C1) of T. conoides on Superdex-30 column with 500 mM sodium acetate buffer (pH 5.0) at 20 mL/h. Elution of polysaccharide was expressed as a function of the partition coefficient Kav [Kav = (Ve - V0)/(Vt - V0) with Vt and V0 being the total and void volume of the column determined as the elution volume of glucose and blue dextran, respectively, and Ve is the elution volume of the sample].
2-2.7. Structural features of the glucan and alginic acid rich fractions
Glucans are soluble, but their water
solubility depends on the branching level: the
higher the branching degree, the higher the
solubility. The less branched glucans are
soluble only in warm water (60–80°C). The
alginic acid (C2) present in the hot water
extract (C) was removed by taking advantage
of the insolubility of Ca- alginate in water.
The molecular mass of glucans is small (5–10
kDa). Therefore, attempts have been made to
separate it from fucoidan by size exclusion
chromatography. Indeed, SEC
on Superdex-30 separates C1 into two fractions C1F1 and C1F2 (Fig. 2.10). The minor fraction,
C1F2, which accounted for 35% of the
total sugar recovered from the column
consisted of mainly glucose as neutral
sugar. Based on calibration with
standard dextrans, the apparent
molecular weight of this glucan would
be 5 kDa. This glucan containing
fraction (C1F2) does not produce blue
colouration with iodine. Moreover 1H
NMR spectrum of this polymer clearly
shows the presence of only one
anomeric signal at 4.827 ppm indicative
of β-configuration. Finally, methylation
analysis of this glucan revealed
the presence of 1,3,5-tri-O-
acetyl-2,4,6-tri-O-methylglucitol
C1F1
C1F2
ppm

38
(98%) and 1,5-di-O-acetyl-2,3,4,6-tera-O-methyl glucitol (2%) residues. On the basis of the
glycosidic linkage and 1H NMR analysis it may be concluded that the glucan of T. conoides is a
linear polysaccharide and contained β-(1→3)-linked glucopyranosyl residues. Sodium alginates
form insoluble precipitates at acid pH and with calcium salts, but they are soluble in solution
between pH 6 and 9. Therefore, these macromolecules were extracted with Na2CO3. The
fractional product (B) was analysed using FT-IR and 1H NMR spectroscopy. The FT-IR
spectrum of this fraction contains band at 3400 (OH stretching), 2925 (CH stretching), 1675 and
1420 (COO stretching) cm-1 characteristic of alginate. Moreover two bands at approximately
1100 and 1025 cm-1 responsible for mannuronic (M) and guluronic (G) units, respectively, were
also observed. To evaluate the content of G, M and G–G linkage, we have investigated the 1H
NMR spectrum (Fig. 2.11) of sodium alginate using procedures as described by Grasdalen, 1983
and Matsushima, 2005. The relative areas of anomeric protons G-1 (H-1 of G) and M-1 (H-1 of
M) correspond to the mole fractions of G and M, respectively. The peak areas of G-5 (H-5
resonances of G) give the distribution of G in G-block and GM-block, and the algebraic sum of
their intensities accounts for the total G content, i.e., the relative area of G-5 is equal to that of G-
1. From the measurement of peak areas of A (δH 5.13 (G-1)), B (δH 4.72–4.84 (G-5 in GM-
block and M-1)) and C (δH 4.45–4.52 (G-5 in G-block)), the guluronic acid content (G%) and
G–G diad frequency (GG%) were calculated using Eqs. (1) and (2), and the calculated values are
46.19 and 42.12, respectively
2-3. PHARMACOLOGICAL ACTIVITIES OF POLYSACCHARIDES
FROM Turbinaria conoides
2-3.1. Antioxidative activity
Ferric ion reducing/antioxidant power (FRAP) assay. In recent years many different methods
are being used to evaluate antioxidative capacity of foods and biological samples (Huang et al.,
(2) (area of B) + (area of C)
GG% (%) = area of C X 100
(1) (area of B) + (area of C)
G% (%) = area of A X 100

39
Table 2.3. Ferric ion reducing abilitya of polysaccharide containing fractions from T. conoides
4 min 30 min
Fractions µµµµmol
Trolox/g
µµµµmol
Fe(II)/g
µµµµmol
Fe(II)/g
µµµµmol
Trolox/g
AF3b 165 79 190 96
Bb 92 53 137 85
C1F2 b 72 38 82 43
aResults are expressed as equivalent of Fe(II) or Trolox per gram dry weight of fractions in aqueous solution. Each value, which is given to its nearest whole number, was the mean of 2 replicates. bFor a description of fractions obtained see “Materials and methods”.
2005). In some of these protocols, antioxidative assays were performed in alcoholic solutions,
but in this condition polysaccharide
would precipitate. Therefore,
the antioxidative capacity of
sulfated polysaccharides (AF3, B
and C1F2) from Turbinaria
was determined by the FRAP
assay. This assay in which a ferric
salt Fe(III) (TPTZ)2Cl3
(TPTZ=2,4,6-tripyridyl-s-triazine)
is used as oxidant (Benzie &
Strain, 1996) takes advantage of
electron transfer reactions (Huang et al., 2005). FRAP values increased considerably from 4 to
30 min, as it has been described for other vegetable and seaweed samples (Ruperez et al., 2002).
Regarding antioxidative capacity of the polysaccharides of present study it is clear that the
fucoidan (AF3) showed the highest reducing power at 4 and 30 min (165 and 190 mol Fe(II) per
g sample dry weight, respectively). This is followed by alginic acid and glucan (Table 2.3).
Results are expressed as mol Fe(II) per g sample dry weight. For comparison of potencies, values
are also calculated as mol Trolox/g sample dry weight from regression equations as described by
Pulido, 2000 of Trolox at 4 and 30 min of reaction with the FRAP reagent.
Scavenging effect on1,1-diphenyl-2-picrylhydrazyl (DPPH) radicals. For further insight into
the activation mechanism, we examined whether the protective effect was associated with DPPH
radicals. The proton radical scavenging action is known to be one of the various mechanisms

40
Fig. 2.12. Scavenging effect of polysaccharides isolated from the brown seaweed T. conoides on DPPH radicals. BHA: Butylated hydroxyanisole. AF3: The fucoidan rich fraction. B: The alginic acid containing fraction. C1F2: The glucan fraction.
0 1 2 3 4 5
0
20
40
60
80
100
BHA AF3 B C2F3
Sca
veng
ing
effe
cts
(%)
Concentration (mg/ml)
C1F2
for antioxidation. DPPH is one of the compounds that possess a proton free radical and shows a
characteristic absorption at 517 nm (purple) (Matsukawa et al., 1997). When DPPH encounters
proton radical scavengers, its purple colour would fade rapidly (Yamaguchi et al., 1998). An
excellent scavenging capability on DPPH radicals at a dosage of 1 mg/ml (61%) was found with
the fucoidan fraction AF3 as compared with the control BHA (100%) regarding the low dosage
ranges used (Fig. 2.12). More significant and effective radical scavenging capability (90%) was
also found with the fraction AF3 at a
higher dosage (5 mg/ml).
Comparable results with the fraction
AF3 were found to be 62% at the
dosages of 5.0 mg/ml for the alginate
rich fraction B (Fig. 2.12). In
contrast, the glucan fraction C1F2
demonstrated least scavenging
capability (50%) among three
fractions at the concentration of 5.0
mg/ml. Moreover, the marked
inhibitory effect of these
polysaccharides on DPPH radicals was
found to be concentration dependent (Fig. 2.12), though the activities were low. These results
reveal that the polysaccharides of present study are potent scavenger and their antioxidative
activity may be attributed to their proton-donating ability (Shimada et al., 1992). Probably, these
fractions might contain a higher amount of reductone which could react with radicals to stabilise
and terminate radical chain reaction. Whether the structures can be related to scavenging
capacity for DPPH radicals remains to be determined in future studies. However, our preliminary
results suggest that different bioactivities of these fractions apparently may be, in some respects,
linked to their different molecular structures. Most of earlier data on antioxidants of foods and
biological samples were on phenolic compounds and many researchers have been reported
positive correlation between free radical scavenging activity and total phenolic compound. But
this correlation between structure and activity is not always valid. For example, selected
enzymatic extracts of seaweeds (Ecklonia cava and Sargassum coreanum) did not possess

41
antioxidative activity, although they contained as much phenolic compounds as the other extracts
of E. cava. Feruloyl oligosaccharides showed a higher antioxidative capacity than free ferulic
acid (Yuan et al., 2005). Moreover, the former showed greater antioxidative capacity in vivo than
in vitro when compared with vitamin C (Ou et al., 2007). Therefore, it is believed that other
materials in seaweed extracts, such as small molecular weight polysaccharides, pigments,
proteins or peptides, may influence the activity. Indeed, recent data showed that a number of
polysaccharide containing fractions isolated from various sources such as medicinal mushrooms
(Shimada et al., 1992; Jiang et al., 2008), higher plants (Aguirre et al., 2009), marine algae
(Ruperez et al., 2002; Rocha de Souza, 2007; Wang et al., 2008, 2009a) and even some
enzymatic extracts (Je et al., 2009) possess antioxidative activity. Our results are in agreement
with these studies. In addition, the polysaccharides used in this study were purified and,
therefore, conclusively prove their antioxidative potency. Literature data also shows that some
polysaccharides have higher antioxidative potency than the others. For example, the
antioxidative potential of sulfated polysaccharides from the brown seaweed Fucus vesiculosus
was higher than that of agar-like sulfated galactans from the red seaweed Nori (Ruperez, 2001).
In a more recent study this group (Ruperez et al., 2002) found that antioxidative potency of
fucoidan containing fraction from the brown alga Fucus vesiculosus was higher than that of
glucan and alginic acid containing fractions. The glucan (C1F2) and alginate (B) rich fractions of
present study exhibited lower antioxidative potentials than the fucoidan containing fraction AF3
(Table 2.3). This result is in agreement with that of (Ruperez et al., 2002) except that the
antioxidative potency of the purified polysaccharides of present study is lower. Although the
antioxidative capacity of polysaccharide has been proven, the relationships between structure and
antioxidative capacity have not yet been elucidated. This is primarily due to the interplay of two
important factors. Firstly, the huge structural diversity of these polysaccharides has given a
major hindrance in the structure–activity relationship establishment. However, on the basis of the
accumulated data several common structural motifs emerge that are important for activity.
Secondly, many of the hitherto used sulfated polysaccharides contained a number of other
molecules. These later molecules may have their own activities, and or at least dilute the efficacy
of the sulfated polymer itself. However, the polysaccharides described in this work are purified
and therefore, conclusively proves their antioxidative activities.

42
2-4. CONCLUSIONS
In this study, we found that the soluble polysaccharides of T. conoides possess antioxidative
property. The higher molecular mass fucoidan (50 kDa) exhibits stronger activity than the low
molecular mass glucan (5 kDa) and, therefore, this potency probably be directly related with the
molecular mass of these polysaccharides. These differences can as well be attributed to the
differences in structure. The antioxidative activity of these polysaccharides may be attributed to
their proton-donating activity as evidenced through DPPH radical scavenging results. Because
the polysaccharides tested in this study was basically prepared without toxic chemical reagents, it
can be assumed to be potentially useful as a safe antioxidants for food processing industries.
Furthermore, as the isolation of these polysaccharides involves a few inexpensive and easy steps
it will be of an added advantage.

3
STRUCTURAL FEATURES AND ANTITUSSIVE
ACTIVITIES OF A CARBOHYDRATE POLYMER FROM
Adhatoda vasica

44
3-1. INTRODUCTION
Cough is a protective reflex that is vital to remove foreign material and secretions from the
airways (Nasra & Belvisi, 2009). In the physiological conditions in healthy persons this reflex
serves its function appropriately. However, modulation of the cough reflex pathway can lead to
inappropriate coughing and an augmented cough response. Ineffective cough is associated with
respiratory morbidity such as recurrent pneumonia. However, chronic cough can be troublesome.
It impairs the quality of life in adults (French et al., 2002) and significantly worries the parents
of coughing children (Cornford et al., 1993). Therefore, cough is the most common condition for
which patients seek consultation from a doctor (Schappert & Burt, 2006). Codeine and
dextromethorphan are extensively used for the treatment of cough (Chung, 2009). But current
antitussives, such as opiods, have unwanted side-effects (Belvisi & Hele, 2009). Therefore, new
antitussive agents are needed. In this context, natural products scaffolds have been invaluable as
biologically validated platforms for drug development (Ramallo et al., 2011).
Adhatoda vasica, also known as Basak is a shrub belonging to the Acanthaceae family. This
small evergreen shrub has been used in traditional Indian medicine for more than 2000 years
(Wealth of India, 1988; Kapoor, 1990). Leaves of this shrub are the main source of drug
preparation. The juice from the leaves and a decoction or infusion of the leaves and roots, water
extracts or syrups are used as herbal remedy for asthma, bronchitis, and chronic coughs and
breathlessness (Amin & Mehta, 1959; Dhuley, 1999). Basak contains alkaloids of great interest
to researchers including vasicine, vasicinone, deoxyvasicine, vasicol, adhatodinine, vasicinol and
others (Claeson et al., 2000). Other constituents include the vitamins, saponins, flavonoids as
well as steroids, and fatty acids (Wealth of India, 1988). As for example, roots contain a steroid,
called daucosterol, carbohydrates and alkanes (Huq et al., 1967; Jain et al., 1980; Jain & Sharma,
1982). Flowers were shown to harbor a triterpene α-amyrin; flavonoids (astragalin, kaempferol,
quercetin, etc) and alkanes (Huq et al., 1967; Claeson et al., 2000). Two aliphatic
hydroxyketones were also isolated from the aerial parts of A. vasica, namely, 37-
hydroxyhexatetracont-1-en-15-one and 37-hydroxyhentetracontan-19-one (Singh et al., 1991).
Two new pyrroloquinazoline alkaloids [1,2,3,9-tetrahydropyrrolo(2,1-b-quinazolin-9-one-3R-
hydroxy-3(2'-dimethylamino phenyl (desmethoxyaniflorine) and 7-methoxy-3R-hydroxy-1,2,3,9-
tetrahydropyrrolo[2,1-b]-quinazolin-9-one (7-methoxyvasicinone)] were also isolated from A.

45
vasica and their structures were deduced using spectroscopic and X-ray diffraction analyses
(Thappa et al., 1996). Four quinazoline alkaloids anisotine, 3-hydroxy-anisotine, vasicolinone,
vasicoline and one isoquinoline alkaloid 1-phenyl-2-methyl-6,7-dimethoxy-1,2,3,4-
tetrahydroisoquinoline were also isolated from the leaves of A. vasica. Additionally
unsaponifiable fraction of petrol ether extract of leaves were shown to contain higher levels of
beta-sitosterol and tricontane, whereas saponifiable fraction contains higher levels of linolenic,
arachedonic, linoleic, palmitic and oleic acids (Abd El-Megeed Hashem & Ahmed Elsawi,
1998). A new triterpenoid, 3 α-hydroxyoleanane-5-ene was also isolated from the aerial part of
A. vasica (Sultana et al., 2005). Some of these compounds contribute to the observed medicinal
effect of this plant. For example, the alkaloid vasicine showed bronchodilatory activity both in
vitro and in vivo. Vasicinone showed bronchodilatory activity in vitro but bronchoconstrictory
activity in vivo (Atal, 1980). Notably, these compounds cannot be used during pregnancy.
However, there have been no reports on the high molecular weight bioactive compounds present
in A. vasica although plant extracts consisting of carbohydrate polymers exhibit a large range of
pharmacological activities (Inngjerdingen et al., 2006; Cumashi et al., 2007; Sutovska et al.,
2007, 2009; Mantovani et al., 2008; Ghosh et al., 2009a; Baek et al., 2010; Thakur et al., 2012).
The present study reports isolation and chemical characterization of a water extracted
polysaccharide isolated from A. vasica leaf. Using chemical and chromatographic methods we
have been able to deduce structural features of a pectic arabinogalactan. We have also
investigated the antitussive activity of this polysaccharide in terms of number of cough efforts on
the citric acid-induced cough reflex and the reactivity of the airway smooth muscle in vivo
conditions in guinea pigs.

46
Table 3.1. Sugar composition and protein content of the water extracted polysaccharides containing fraction (WE) isolated from A. vasica leaves, the arabinogalactan protein (AGP) and of fractions (F1 – F3) derived by size exclusion chromatography
WE AGP F1 F2 F3 Yielda - - 16 72 12 Total sugarb 49 ndd 51 49 47
Uronicacidb 7.3 ndd 7.6 7.2 6.9
Proteinb Rhamnosec
21 2
ndd 1
19 3
21 2
22 2
Arabinosec 29 32 29 30 30
Xylosec 1 tre 1 1 tre
Mannosec 1 1 1 tre tre
Galactosec 60 66 60 62 61
Glucosec 7 tre 6 5 7
apercent weight of total neutral sugar recovered. bpercent weight of fraction dry weight. cmol percent of neutral sugars. ndd, not determined. tre, trace.
3-2.CHEMICAL CHARACTERIZATION OF THE PECTIC ARABINOGALACTAN FROM Adhatoda vasica
3-2.1. Isolation and chemical composition
The objectives of this research were to analyze the water extracted polysaccharide generated
from the medicinal plant Adhatoda vasica and to study its antitussive activity. In Indian
Ayurvedic system of medicine a decoction of its leaves in water is used as herbal remedy for
chronic coughs and breathlessness (Amin, 1959), therefore fresh leaves of this evergreen shrub
were extracted with water.
The yield of the water extracted polymer (named as WE), after fractional precipitation with
ethanol, was 10mg per gram of fresh leaves (Fig. 3.1). The use of cold water, in principle, may
exclude the extraction of physiologically inactive starch, present in the leaves. Fraction WE
contains 49% (w/w) neutral sugar along with 21% (w/w) protein. The amino acid composition of
protein associated with
fraction WE showed that
glutamic acid / glutamine
(38.5%), alanine (13.3%),
serine (9.4%), and glycine
(6.5%) were the major
constituents. The uronide
content of this fraction is
7.3% (w/w). Thin layer
chromatographic analysis of
the monosaccharides present
in the hydrolysate indicates
Fig. 3.1. Scheme for the isolation and purification of carbohydrate polymers from Adhatoda vasica.
Water, (25-32)°C WE
Size Exclusion Chromatography
Sephacryl S-100
F1
F2
F3

47
Fig. 3.2. Elution profile of the water extracted polysaccharide obtained from Adhatoda vasica leaves on Sephacryl S-100 column with 500 mM sodium acetate buffer (pH 5.5) at 30 ml/h. Collected fractions were analyzed for total sugar content by phenol-sulfuric acid. Elution of polysaccharide was expressed as a function of the partition coefficient Kav [Kav = (Ve-Vo)/(Vt-Vo) with Vt and Vo being the total and void volume of the column determined as the elution volume of potassium hydrogen phthalate and dextran (500 kDa), respectively and Ve is the elution volume of the sample].
F2
F1
F3
the presence of an uronic acid with Rf values similar to that of galacturonic acid. GC analysis of
the TMS derivatives of the derived methyl glycosides confirmed this result, but it also shows the
presence of traces of glucuronic acid. Sugar compositional analysis revealed that fraction WE
consist mainly of arabinose and galactose as the major neutral sugar together with smaller
amount of glucose, rhamnose, mannose and xylose units (Table 3.1). Considering that the water
extracted polymeric fraction (WE) contained galactosyl and arabinosyl residues as the major
sugars, and its protein content is 21%, we have tested its reactivity with β-glucosyl Yariv reagent
that specifically precipitates AGPs. We found that the major part of WE was Yariv soluble.
Sugar compositional analysis of this precipitate (named AGP) shows that it consisted mainly of
galactose residues and, to a lesser extent, arabinose residues, confirming the presence of AGP
(Table 3.1). This material also contained mannose residues probably originating from N-glycans.
3-2.2. Size exclusion chromatography (SEC)
SEC of WE on Sephacryl S-100 yielded three
overlapping subfractions (F1, F2 and F3; Fig.3.2). The
yield and chemical composition of these subfractions
are given in Table 3.1. They had similar
monosaccharide compositions. The only difference
between these three samples, as judged by size
exclusion chromatography, seems to be the molecular
weight. Based on calibration with standard dextrans,
the apparent average molecular mass of
macromolecules present in subfractions F1, F2
and F3 would be 61, 42 and 9 kDa, respectively.
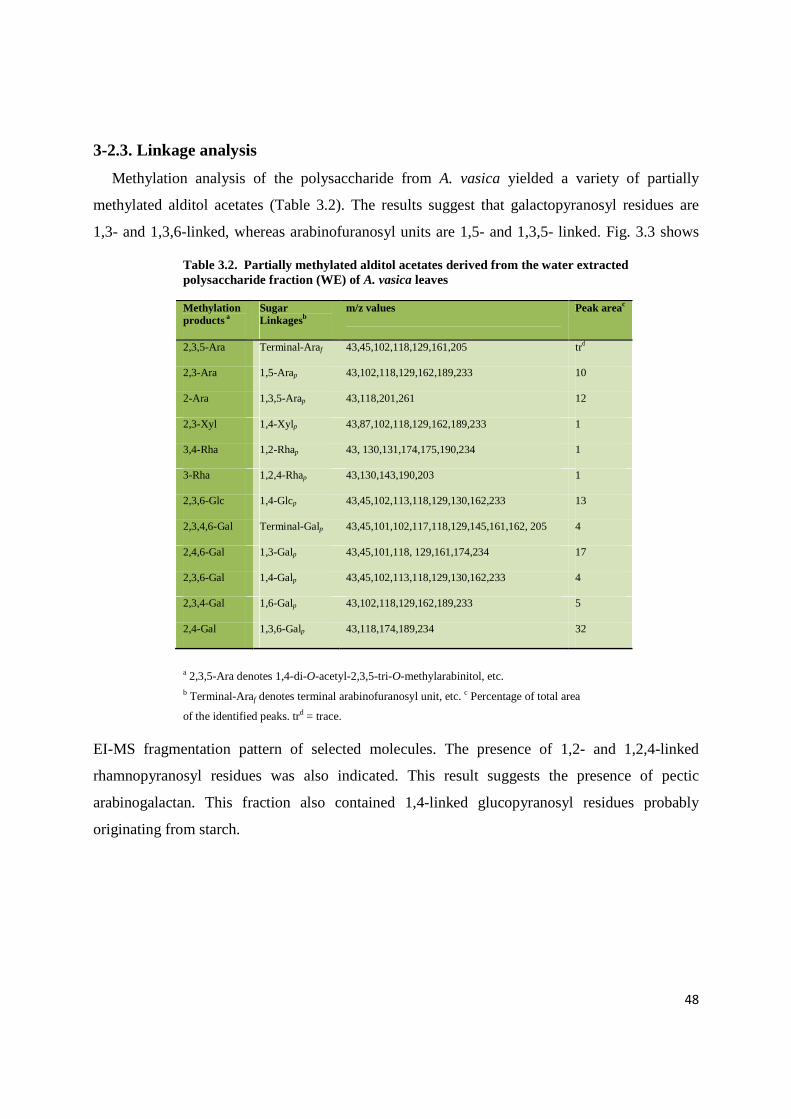
48
Table 3.2. Partially methylated alditol acetates derived from the water extracted polysaccharide fraction (WE) of A. vasica leaves
Methylation products a
Sugar Linkagesb
m/z values Peak areac
2,3,5-Ara Terminal-Araf 43,45,102,118,129,161,205 trd
2,3-Ara 1,5-Arap 43,102,118,129,162,189,233 10
2-Ara 1,3,5-Arap 43,118,201,261 12
2,3-Xyl 1,4-Xylp 43,87,102,118,129,162,189,233 1
3,4-Rha 1,2-Rhap 43, 130,131,174,175,190,234 1
3-Rha 1,2,4-Rhap 43,130,143,190,203 1
2,3,6-Glc 1,4-Glcp 43,45,102,113,118,129,130,162,233 13
2,3,4,6-Gal Terminal-Galp 43,45,101,102,117,118,129,145,161,162, 205 4
2,4,6-Gal 1,3-Galp 43,45,101,118, 129,161,174,234 17
2,3,6-Gal 1,4-Galp 43,45,102,113,118,129,130,162,233 4
2,3,4-Gal 1,6-Galp 43,102,118,129,162,189,233 5
2,4-Gal 1,3,6-Galp 43,118,174,189,234 32
a 2,3,5-Ara denotes 1,4-di-O-acetyl-2,3,5-tri-O-methylarabinitol, etc. b Terminal-Araf denotes terminal arabinofuranosyl unit, etc. c Percentage of total area
of the identified peaks. trd = trace.
3-2.3. Linkage analysis
Methylation analysis of the polysaccharide from A. vasica yielded a variety of partially
methylated alditol acetates (Table 3.2). The results suggest that galactopyranosyl residues are
1,3- and 1,3,6-linked, whereas arabinofuranosyl units are 1,5- and 1,3,5- linked. Fig. 3.3 shows
EI-MS fragmentation pattern of selected molecules. The presence of 1,2- and 1,2,4-linked
rhamnopyranosyl residues was also indicated. This result suggests the presence of pectic
arabinogalactan. This fraction also contained 1,4-linked glucopyranosyl residues probably
originating from starch.
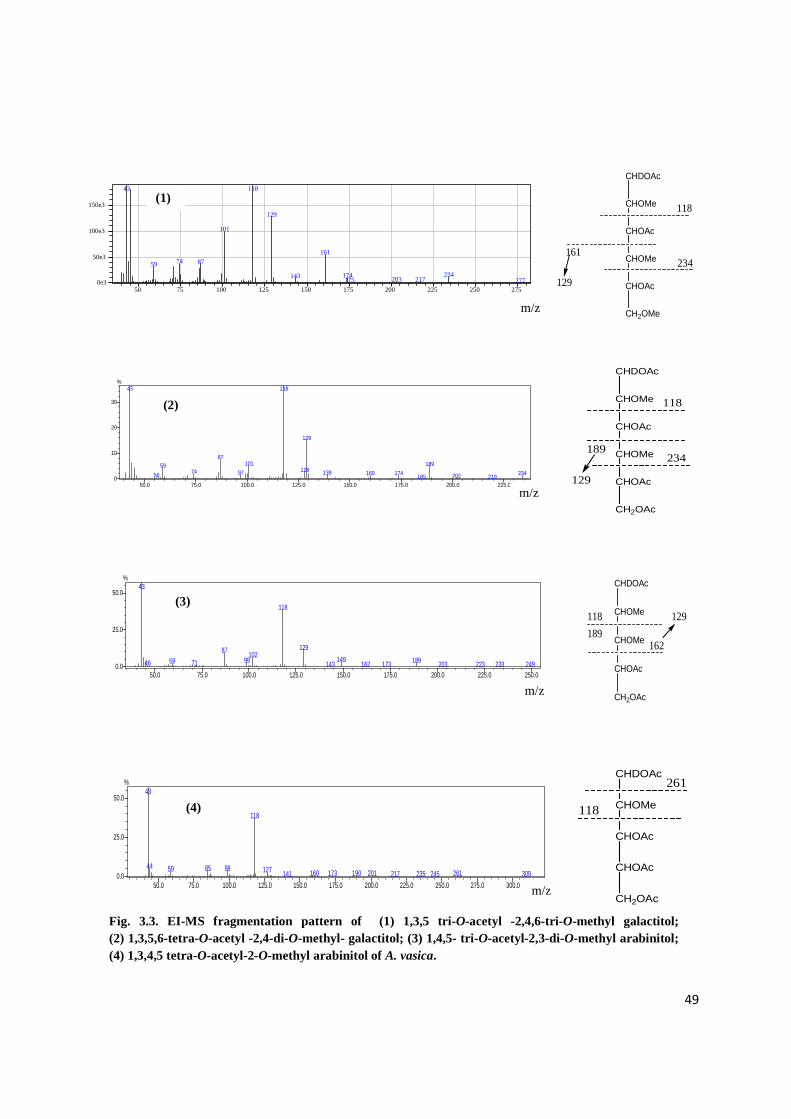
49
50.0 75.0 100.0 125.0 150.0 175.0 200.0 225.00
10
20
30
%43 118
129
87101 18959
12874 97 139 234160 17456 202185 219
CHDOAc
CHOMe
CHOAc
CHOMe
CHOAc
CH2OMe
118
161
129
234
50 75 100 125 150 175 200 225 250 2750e3
50e3
100e3
150e3
43 118
129
101
16174 8759
234174143 203 217175 277
m/z
m/z
CHDOAc
CHOMe
CHOAc
CHOMe
CHOAc
CH2OAc
118
189
129
234
50.0 75.0 100.0 125.0 150.0 175.0 200.0 225.0 250.00.0
25.0
50.0
%43
118
12987102
149 1899959 7146 162 223143 203173 233 249
m/z
50.0 75.0 100.0 125.0 150.0 175.0 200.0 225.0 250.0 275.0 300.00.0
25.0
50.0
%43
118
44 998559 127 261201160 173 190141 217 235 245 309
m/z
CHDOAc
CHOMe
CHOMe
CHOAc
CH2OAc
118
189162
129
CHDOAc
CHOMe
CHOAc
CHOAc
CH2OAc
118
261
Fig. 3.3. EI-MS fragmentation pattern of (1) 1,3,5 tri- O-acetyl -2,4,6-tri-O-methyl galactitol; (2) 1,3,5,6-tetra-O-acetyl -2,4-di-O-methyl- galactitol; (3) 1,4,5- tri-O-acetyl-2,3-di-O-methyl arabinitol; (4) 1,3,4,5 tetra-O-acetyl-2-O-methyl arabinitol of A. vasica.
(1)
(2)
(3)
(4)

50
3-3. ANTITUSSIVE ACTIVITY OF THE PECTIC ARABINOGALA CTAN FROM Adhatoda vasica
3-3.1. Assessment on chemically induced cough and airways defense reflexes
The effect of water extracted pectic arabinogalactan from A. vasica (WE) was evaluated on
the citric acid-induced cough reflex and reactivity of airways smooth muscle in vivo conditions.
For comparative purposes, codeine was simultaneously assayed as known reference compound.
The results of antitussive test showed that peroral administration of the pectic arabinogalactan in
a dose 50 mg kg−1 body weight brought about a significant decrease in the number of citric acid
induced cough efforts (NE) in adult healthy awaken guinea-pigs (Fig. 3.4). The first statistically
significant result was observed within 30 min after application. Furthermore, this positive effect
was observed during all the study time intervals. Notably, the suppression of cough efforts by the
polymer was quantitatively similar to that of codeine. Airway resistance is a concept used in
respiratory physiology to describe mechanical factors which limit the access of inspired air to the
pulmonary alveoli, and thus determine airflow. It is dictated by inter alia, the diameter of the
airways.
At present the relationship between cough and bronchoconstriction is not known with
**
**
** **
0
1
2
3
4
5
6
7
8
9
N 30 60 120 300 min
Nu
mb
er o
f co
ug
h e
ffo
rts
(NE
) control
codeine
WE
**
***
**
***
**
***
**
***
Fig. 3.4.The influence of the pectic arabinogalactan from Adhatoda vasica (WE), codeine (positive control) and saline water (negative control) on the citric acid-induced cough efforts (NE) in guinea-pigs recorded at 30, 60, 120 and 300 min time intervals. All used substances were applied by peroral route of administration: plant polysaccharides in the dose of 50 mg kg-1, codeine in the dose 10 mg kg-1and saline water in the dose 1 ml kg-1body weight. N, Initial values before application of the polysaccharides and codeine. Statistical significance

51
N 30 60 120 300 min0
5
10
15
20
25
R x
V (cm
H2O
s-1) Contr
Codeine
WE
certainty. Although it is generally accepted that bronchodilating substances can cause cough
suppression. Therefore, we have evaluated the changes of specific airway resistance as indictor
of this activity. Our result suggests that the application of arabinogalactan from A. vasica in the
dose which provoked cough suppressive activities did not significantly change the values of
specific airway resistance (Fig. 3.5). It also showed that after 30 min from application of
polysaccharide sample a slightly decreased value of specific airway resistance is registered.
Notably, in this time interval application of codeine provoked the same result. Recent studies
showed that several naturally occurring polysaccharides possess antitussive activity (Nosál’ova
et al., 2005; Sutovska et al., 2007, 2009a; Saha et al., 2011; Sinha et al., 2011b). The most potent
antitussive activity is observed with polysaccharide containing the highest proportion of uronic
acid constituent (Sutovska et al., 2007), although neutral polysaccharides may also have
significant activity (Nosál’ova et al., 2005). Therefore, the fact that the pectic arabinogalactan
from A. vasica modulates chemically induced cough reflex and specific airway resistance is not
surprising.
3-3.2. Mechanism of action
The mechanism behind the antitussive activity of the pectic arabinogalactan of present study
is not known, but many antitussive herbs probably work through an antispasmodic action or as
bronchodilator (Ernst, 1998; Pavord & Chung, 2008). They cause bronchial muscle relaxation in
Fig. 3.5. The influence of water extracted polysaccharide from the medicinal plant Adhatoda vasica and control agents (vehicle and codeine) on citric acid induced changes of specific airway resistance (R x V) in vivo conditions, registered before any agent application (values labelled as N in graphs) and after that in 30, 60, 120 and 300 min time intervals.

52
vitro, or decrease airways resistance in vivo. The arabinogalactan of present study possesses very
high cough suppressive effect and decreases the values of specific airway resistance in vivo
conditions only slightly. Pavord (2004) reported that bronchoconstriction causes or enhances the
sensitivity of cough, while bronchodilation does the opposite. However, the role of other
mechanism including bioadhesive effect of the polysaccharide to the epithelial mucosa (Sutovska
et al., 2009a) cannot be ruled out.
3-4. CONCLUSIONS
This study represents the first account on the in vivo antitussive activity of the water extracted
polysaccharide and its structural features. Biological investigations indicated that the pectic
arabinogalactan of A. vasica displayed promising activity in the antitussive assays. Moreover, as
the macromolecule tested in this study was basically isolated without toxic chemical reagents, it
can be assumed to be potentially useful as a safe antitussive agent for industries. Furthermore, as
the isolation of these polysaccharides involves few inexpensive and easy steps it will be of an
added advantage. Finally, the biological activity observed in A. vasica provides a scientific basis
for the use of the plant in traditional medicines.

EXPERIMENTAL SECTION

54
4
MATERIALS AND
METHODS

55
4-1. PLANT MATERIALS AND EXTRACTION OF POLYSACCHARI DES
4-1.1.Isolation of fucoidan, alginic acid and glucan from Turbinaria conoides by
sequential extraction with inorganic solvents
Plant material and preliminary treatments. Samples of Turbinaria conoides were collected
from Okha coast of Gujarat, India, in August 1995. The collected seaweeds were washed
thoroughly with tap water and dried by forced air circulation, followed by pulverization in a
Waring Blender.
Preparation of depigmented algal powder (DAP). After pulverization in a commercial grinder,
Depigmentation of algal powder (270 g) was done using sequential extraction with petroleum
ether (24 h) and acetone (24 h) as solvent in a Soxhlet apparatus. The unextracted material was
placed in a plastic beaker, and air dried to obtain depigmented algal powder (DAP, 173 g).
Extraction with acid. Extractions of DAP (3 g) with 0.1 M HCl (w/v::1:100) were conducted at
30–35°C for 12 h under constant stirring (twice). Separation of the residue from the extract was
performed by filtration through a glass filter (G-2). The residue was washed with additional 0.1
M HCl and the wash was collected to maximize polysaccharide recovery. The extracts were
combined, dialyzed and lyophilized to give fraction A (yield, 8.8%).
Extraction with alkali. The acid insoluble residue was extracted with 3% Na2CO3 (w/v::1:100) at
45–50°C for 4 h under constant stirring (twice). The extract was carefully acidified with
concentrated HCl to pH~1, the precipitate formed was collected by centrifugation, washed with
0.1 M HCl, suspended in water and dissolved by careful addition of NaOH. The slightly alkaline
solution was dialyzed and diluted with 4.0 M CaCl2 solution to make a final concentration of 2%
CaCl2. The precipitate formed at this stage was isolated by centrifugation, washed with water and
treated with 0.1 M HCl (4 x 50 ml, stirring at room temperature for ~2 h). Then the precipitate
was dissolved in dilute NaOH as above, the solution dialyzed and finally lyophilized to yield
sodium alginate (B, yield, 22.6%).
Extraction with water. The residue left after extraction with alkali was once again extracted with
water at 80°C (twice) and the combined extract (C) dialyzed. Then 4.0 M CaCl2 solution was
added to the extract to make a final concentration of 2% CaCl2, and the suspension kept at 5–9°C
overnight. After separating the precipitate (C2) by centrifugation the supernatant dialyzed,

56
concentrated and lyophilized to give fraction C1 (yield, 6.2%). The resulting insoluble residue
was dialyzed against distilled water and finally lyophilized (INS).
4-1.2.Isolation of polysaccharides from Adhatoda vasica
Plant material and preliminary treatments. Leaves of Adhatoda vasica were collected from the
garden of medicinal plants, The University of Burdwan, West Bengal, India. Collected leaves
(10 g) were washed thoroughly with tap water and then blended with water (800 ml) in a mixer
(Waring Products, Inc., Torrington, CT, USA).
Extraction of polysaccharides. Extraction of polysaccharide was conducted by stirring a
suspension of this paste in water (pH 6.0) at 25–32°C for 12 h. Separation of the residue from the
extract was performed by filtration through a glass filter (G-2). The insoluble material was
extracted twice more under similar condition at a solute to solvent ratio of 1:100 (w/v). The
combined liquid extract was dialyzed extensively against water and lyophilized. The recovered
material was dissolved in water, precipitated by the addition of ethanol (4 volumes) and then
collected by centrifugation (repeated three times). The final pellet was dissolved in water and
lyophilized to yield the water extracted polysaccharide, named WE (100 mg).
Isolation of arabinogalactan protein (AGP) with β-glucosyl Yariv reagent. AGP was isolated
according to Schultz et al., 2000. Briefly to a solution of WE in 1% NaCl (w/v) was added an
equal volume of β-glucosyl Yariv reagent also in 1% NaCl. The mixture was kept at 4°C for 18 h
and then centrifuged. The pellet was washed with 1% NaCl followed by pure methanol (3 times
each), dried and treated with sodium metabisulfite (10%). The resulting solution was dialyzed
and freeze dried to yield the arabinogalactan protein (AGP).
4-2. ANALYTICAL METHODS
4-2.1. General
All the experiments were carried out at least in duplicate, the mean and standard deviation
was directly calculated from the functions present in excel program. Evaporations were executed
under reduced pressure at around 45°C. Dialysis was performed against distilled water with
continuous stirring, toluene being added to inhibit microbial growth. During experiments,
moisture was determined by drying ground material in an air-circulated oven at 110°C for 3
hours. Chemicals used were analytical grade or best available. Except otherwise stated data

57
presented are the mean values of at least three independent analyses. When the standard
deviation was greater than 5%, experiments were repeated.
4-2.2. Sugar analysis
Total sugars were estimated by the phenol-sulphuric acid method (Dubois et al., 1956).
Evaluation was corrected afterwards, depending on sugar composition. Total uronic acids
present, were assayed using m-phenyl phenol color reagent (Ahmed & Labavitch, 1977). For
measurement of individual neutral sugar polysaccharides (10-12 mg) were hydrolyzed with 2 M
trifluoro acetic acid (2 h, 100°C).For the insoluble residues, this hydrolysis was followed by a
treatment with 72% (w/w) H2SO4 for 1 h at room temperature and after that, with 1 M H2SO4 for
2 h at 100°C. Myo-Inositol was used as internal standard, and hydrolytic losses were accounted
for by using an external standard (Chattopadhyay et al., 2007b). Monosaccharides were reduced
with sodium borohydride in anhydrous dimethyl sulfoxide and after reduction, the excess sodium
borohydride was decomposed by the addition of acetic acid. Acetylation was conducted by acetic
anhydride and 1-Methylimidazole was added as catalyst (Blakeney et al., 1983). Derived alditol
acetates were analysed by GC (Blakeney et al., 1983) on columns of 3% SP-2340 on
Supelcoport 100–120 mesh, and DB-225 (JW) and by GC-MS (Ghosh et al., 2009b; Ghosh et
al., 2010) or alternatively, trimethylsilyl (TMS) derivatives of methyl glycosides were analysed
by gas chromatography.
4-2.3. Protein estimation
Proteins present in soluble fractions were estimated by the method of Lowry et al., 1951 using
bovine serum albumin as standard. In the insoluble residues it was obtained by estimated the
total nitrogen and multiplying the value with 6.25.
Alternatively, protein was measured in the soluble material by the micro-Bradford method
using bovine serum albumin as standard, and was detected in some chromatography fractions by
the absorbance at 280 nm. Amino acids were released by hydrolysis with 6 M HCl at 110°C for
22 h in a sealed tube. Protections were done for cysteine, methionine and tyrosine using proper
protecting reagents. The liberated amino acids were analyzed by Pharmacia LKB ALPHA PLUS
amino acid analyzer.

58
4-2.4. Sulfate estimation
Turbidometric method. Samples (90 - 100 mg) were hydrolyzed in of 2 N hydrochloric acid (5
ml) in sealed glass tubes at 100oC (Chattopadhyay et al., 2010). Sulfate was then estimated using
a modified turbidometric barium chloride method (Craigie et al., 1984). The reagent was
prepared by dissolving 600 mg of gelatin in 200 ml of distilled water at 70oC and the solution
was allowed to cool. After 16 h at 4oC the temperature was brought to 30 – 35oC and 2.0 g of
BaCl2 .2H2O was added and then dissolved. AR grade K2SO4 was used here as a standard in the
range of 10 – 50 µg S ml-1. Accurately weighted samples (25-50 mg) were hydrolyzed for 2 h at
100oC with 2N HCl (0.75 ml) in a sealed tube. After that, the contents were transferred and made
to volume in a 10 ml volumetric flask. The humic substances were removed by centrifugation. 2
ml of sample, 18 ml of distilled water and 2 ml of HCl (0.5 N) was mixed in a 50 ml Erlenmeyer
flask. 1 ml of BaCl2-gelatin reagent was added to that and swirled. After 30 minutes swirling
again mixed the contents of the flasks and turbidity was measured at 550 nm against a reagent
blank.
Spectroscopic method. The sulfate content was measured according to the method of Rochas &
coworkers (1986) by considering the ratio of the intensity of the 1252 cm-1 band to the intensity
of the 2920 cm-1 band.
4-2.5. Desulfation
Desulfation of the purified fucoidan (AF3) obtained from T. conoides was carried out
according to the procedure of Falshaw & Furneaux (1998). The pyridinium salt of the sample (1
g) was dissolved in DMSO containing 10% MeOH, and then the solution was kept for 3 h at
100oC. The reaction mixture was cooled, diluted with an equal volume of water, and the pH
adjusted to 9.0 by the addition of sodium hydroxide (0.1 M). The solution was dialyzed against
running water for 20 h and after that lyophilization of the dialyzate gave desulfated sample.
4-2.6. Linkage analysis
Methylation was carried out using the method of Blakeney & Stone (1985) as described by
Adhikari & coworkers (2006). Exactly weighed (1-2mg) of the dried samples along with
magnetic bars were taken in tubes fitted with air tight rubber corks. The nitrogen atmosphere was
maintained within the tubes. The samples were dissolved in DMSO and to this solution, lithium
methyl carbanion were added. The mixture was then stirred in nitrogen atmosphere for 1 hr. The
tubes were cooled under ice bath and ice cold methyl iodide (0.4 ml) was added to it. After that

59
the tubes was left for 1 hr at room temperature. The methylation process was repeated once more
and excess methyl iodide was removed under vacuum. The resulting methylated polymer was
then extracted with chloroform-methanol [2:1 (v/v)]. The permethylated polysaccharide thus
obtained was hydrolyzed with trifluoro acetic acid (2.5 M) at 120°C for 75 min, reduced with
NaBD4(1 M) in NH4OH (2 M) for 3 h at room temperature and acetylated with acetic anhydride
using perchloric acid as a catalyst. The partially methylated alditol acetates (PMAA) were then
analyzed by GC and GC/MS as described by Sinha et al., 2010. The PMAA were identified by
(i) measurement of relative retention times (ii) methoxyl substitution pattern as obtained from
GC-MS and (iii) carbohydrate composition of the non methylated polymers.
4-2.7. Chromatography
Thin layer chromatography (TLC). The sugars released by acid hydrolysis (as described above)
were also analyzed by TLC as described by Ray, 2006. Briefly, TLC was then conducted by
ascending technique using silica gel G impregnated with 0.5M NaH2PO4 as the supporting
material. Small amounts of samples were spotted in the plate for identification. The Rf values of
the test material were compared to that with the standard compounds to get an idea about the
components present and their purity. The color of the spot developed also conveys message
about its components.
The solvent system used for developing TLC was the following: (1) Ethanol: Phenol: Pyridine:
0.1 M Phosphoric acid :: 10 : 2 : 2 : 4 and (2) 2-Propanol: Methanol: Water :: 16 : 1 : 3
Detection: By spraying saturated solution of aniline phthalate and heating the plate at 100°C.
Size exclusion chromatography (SEC). SEC was performed as described by Chattopadhyay et
al., 2007a. In brief, solutions (1-2 ml) of different extracts in 0.5 M sodium acetate buffer (pH
5.0), were loaded to columns (i) (2.6 x 50 cm; Amersham Biosciences AB) of Superdex-30 (for
C1 solution of T. conoides), (ii) (90 cm x 2.6 cm, Amersham Biosciences AB) of Sephacryl S-
200 (for the solution of the fucoidan fraction (AF3) of T. conoides ) and (iii) (90 cm x 2.6 cm,
Amersham Biosciences AB) Sephacryl S-100 (for the solution of water extracted polymer (WE)
solution of A. vasica ) equilibrated with the same buffer. The column was then eluted
ascendingly with the same buffer at 0.5 ml min-1 at a temperature of 30–35°C. Elution of
polysaccharide was expressed as a function of the partition coefficient Kav [Kav = (Ve-V0)/(Vt-
V0) where, Vt and V0 are the total and void volume of the column determined as the elution

60
volume of glucose and blue dextran, respectively and Ve is the elution volume of the sample].
Here, the column was calibrated with standard dextrans.
Anion Exchange Chromatography (AEC). A solution (20 ml) of dilute acid extracted polymer
from T. conoides (fraction A) in 50 mM sodium acetate (pH 5.5) was applied to a column (2.6 x
25 cm) of DEAE-Sepharose FF (AcO-1). After that, the column was eluted (0.6 ml min-1)
successively with 0.05 M (fraction AF1), 0.7 M (fraction AF2) and 2.0 M (fraction AF3) NaOAc
buffer in a stepwise manner. Appropriate fractions were pooled, dialyzed and then lyophilized.
Gas chromatography (GC).GC was conducted on (i) GC-17A, Shimadzu and (ii) HP6890 series
Chromatograph.
The gas chromatograph was equipped with a flame ionization detector, the following columns
(a – d) and helium as gas vector.
(a) WCOT fused silica capillary column (length 25 m, i.d. 0.25 mm) and film thickness 0.4µm
with CP-Sil 5 CP as stationary phase and helium as gas vector, (b) 3 % SP-2340 on Supelcoport
100-120 mesh, (c) DB-225 (JW) and (d) SGE BP 225.
The oven temperature program was:
(a) 210°C isothermal or (b) 2 min at 120°C, 10°C/min to 160°C, and 1.5°C/min to 220°C and
then 20°C/min to 280°C or (c)170°C for 15min, 170-210°C at 5°C/min and 210°C for 15 min.
Gas liquid chromatography–Mass Spectrometry (GC-MS): The GC-MS analysis were conducted
using (a) Shimadzu (Japan) GC 17A coupled to QP 5050A MS and (b) GC (HP 6890) coupled to
an Autospec (Micromass, Manchester, UK) MS.
The mass spectra were recorded with MS instrument at 70 eV.
4-2.8. Spectroscopy
NMR Spectroscopy. The 1H NMR spectra of the polysaccharide containing fractions of T.
conoides were recorded on a Bruker 500 spectrometer (Bruker Biospin AG, Fallanden,
Switzerland) operating at 500 MHz, respectively, for 1H. The samples (~10 mg of each) were
heated at 80°C for 30 min with water (1 ml), centrifuged and the resulting supernatant
lyophilised. The freeze-dried sample was deuterium-exchanged by lyophilisation with D2O and
then examined in 99.9% D2O.

61
Scavenging effect (%) =
UV-VIS spectroscopy. UV-VIS spectra were recorded with Shimadzu UV- 1600A
spectrophotometer.
Infra-Red spectroscopy. IR spectra were obtained on a FT-IR spectrophotometer (JASCO FT/IR
- 420) using KBr discs containing finely ground samples.
Mass spectrometry. EI mass spectra were recorded using Shimadzu QP5050A GC/MS
instrument at 70 eV. The characterization of sugars, including 6-O-methyl galactose, was done
on the basis of previously reported mass spectra (Carpita & Shea, 1988) and relative retention
times as described by Ray & Lahaye, 1995.
4-3. BIOASSAY
4-3.1. FRAP assay
The FRAP assay was conducted according to Benzie & Strain, 1996 as modified by Pulido et
al., 2000. Briefly, the oxidant was prepared by mixing 2.5 ml of a 10 mM TPTZ [2,4,6-tri(2-
pyridyl-5-triazine) Fluka Chemicals, Madrid, Spain] solution in 40 mM HCl with 25 ml of 0.3 M
acetate buffer (pH 3.6) and 2.5 ml of 20 mM FeCl3.6H2O. The final solution has Fe(III) of 1.67
mM and TPTZ of 0.83 mM. To measure FRAP value, 900 µl of freshly prepared FRAP reagent
was warmed to 37°C and a reagent blank reading was taken at 593 nm; then 30 µl of test sample
and 90µl of distilled water were added. Absorbance readings were taken after 0.5 s and every 15
s until 30 min using a Shimadzu UV-1601 (PC) Spectrophotometer. The change of absorbance
(DA = A30 min_ A0min) was calculated and related to DA of an Fe(II) standard solution. Aqueous
solutions of known Fe(II) concentrations (100–2000 µM FeSO4.7H2O) were used for calibration.
4-3.2. Scavenging capability for 1,1-diphenyl-2-picrylhydrazyl (DPPH) radicals
The method reported by Shimada et al., 1992 was adopted for measurement of free radical
scavenging capability. To each 4 ml of sample solution, 1 ml of freshly prepared methanolic
DMSO solution of DPPH (0.5 mM) was added, mixed well and then let stand for 30 min at room
temperature in the dark. The absorbance of the resulting was recorded at 517 nm. Butylated
hydroxyanisole (BHA) was used as reference compound. The capability to scavenge the DPPH
radical was calculated using the following equation:
x100

62
4-3.3. Antitussive activity of the arabinogalactan from A. vasica
Animals. Animals used in this experiment were kept in faculty animal house, Department of
Experimental Pharmacology, Slovak Academy of Science, Dobra Voda, Slovakia, with food and
water ad libitum and with a standard air conditioning system. The animals were kept one week in
quarantine before starting the experiment. The experimental protocols were approved by the
Institutional Ethics Committee of the Jessenius Faculty of Medicine, Comenius University in
Martin, Slovakia, registered in Institutional Review Board/Institutional Ethic Board Office (IRB
00005636), complied with Slovakian and European Community regulations for the use of
laboratory animals and follow the criteria of experimental animal’s well fare.
Assessment of chemically induced cough and airways defense reflexes. Awaken guinea-pigs
were individually placed in a body plethysmograph box (HSE type 855, Hugo Sachs Elektronik,
Germany) and restricted so that the head protrudes into the nasal chamber and the neck were
sealed with a soft diaphragm. The cough reflex was induced by aerosol of citric acid in a
concentration 0.3 M. The citric acid aerosol was generated by a jet nebulizer (PARI jet nebulizer,
Paul Ritzau, Pari-Werk GmbH, Germany, output 5 l s−1, particles mass median diameter 1.2µm)
and delivered to the head chamber of the plethysmograph for 3min interval, in which number of
cough efforts was counted. The cough effort was defined as sudden PC-recorded enhancement of
expiratory flow associated with typical cough motion and sound followed by trained observer
(Sutovska et al., 2009a).The reactivity of the airway smooth muscle in vivo conditions was
expressed as values of specific airway resistance calculated according to Pennock et al., 1979 by
time difference between pressure changes in head and chest parts of body plethysmograph during
normal breathing pattern. Both, influence on citric acid-induced cough and specific airway
resistance were registered before any agent application (values labeled as N in graphs) and after
that in 30, 60, 120 and 300 min time intervals. The minimal time interval between two
measurements was 2 h to prevent cough receptors adaptation as well as adaptation of laboratory
animals on kind of irritation. All tested compounds (polysaccharide, codeine and vehicle) were
applied by peroral route of administration, plant polysaccharides in the dose of 50mgkg−1,
codeine in the dose 10 mg kg−1 and saline water (vehicle) in the dose 1ml kg−1 body weight.
Statistics. Student’s t-test was used for the statistical analysis of the obtained results. Data are
presented as mean±standard error of the mean (S.E.M.). p< 0.05 was considered statistically

63
significant. Significance of p < 0.05; p < 0.01 and p < 0.001 is shown by one, two or three
asterisks, respectively.

64
REFERENCES
Abd El-Megeed Hashem, F., & Ahmed Elsawi, S. (1998). Isoquinoline and quinazoline alkaloids of Adhatoda vasica NEES. Pharmaceutical and Pharmacological Letters, 8, 167-169.
Adachi, Y. S., Jinushi, T., Yadomae, T., & Ohno, N. (2002). Th1-oriented immunomodulating activity of gel-forming fungal (1-3)-beta-glucans. International journal of medicinal mushrooms, 4, 95–109.
Adhikari, U., Mateu, C. G., Chattopadhyay, K., Pujol, C. A., Damonte, E. B., & Ray, B. (2006). Structure and antiviral activity of sulphated fucans from Stoechospermum marginatum. Phytochemistry, 67, 2474–2482.
Aguirre, M. J., Isaacs, M., Matsuhiro, B., Mendoza, L., & Zúñiga, E. A. (2009). Characterization of a neutral polysaccharide with antioxidant capacity from red wine. Carbohydrate Research, 344, 1095-1101.
Ahmed, A., & Labavitch, J. M. (1977). A simplified method for accurate determination of cell wall uronide content. Journal of Food Biochemistry, 1, 361–365.
Alban, S., & Franz, G. (2001). Partial synthetic glucan sulfates as potential new antithrombotics: a review. Biomacromolecules, 2, 354-361.
Alban, S., Schauerte, A., & Franz, G. (2002). Anticoagulant sulfated polysaccharides. Part I. Synthesis and structure-activity relationships of new pullulan sulfates. Carbohydrate Polymers, 47, 267–276.
Amin, A. H., & Mehta, D. R. (1959). A bronchodilator alkaloid (vasicinone) from Adhatoda vasica Nees. Nature, 184, 1317.
Atal, C. K., (1980). Chemistry and pharmacology of vasicine – A new oxytocic and abortifacient. Indian Drugs, 15, 15–18.
Athukorala, Y., Lee, K. W., Kim, S. K., & Jeon, Y. J. (2007). Anticoagulant activity of marine green and brown algae collected from Jeju Island in Korea. Bioresource Technology, 98, 1711–1716.
Baek, S. H., Lee, J. G., Park, S. Y., Bae, O. N., Kim, D. H., & Park, J. H. (2010). Pectic polysaccharides from Panax ginseng as the antirotavirus principals in ginseng. Biomacromolecules, 11, 2044–2052.
Balzarini, J., & Van Damme, L. (2007). Microbicide drug candidates to prevent HIV infection. Lancet, 369, 787-797.
Bandyopadhyay, S. S., Ghosh, D., Micard, V., Sinha, S., Chatterjee, U. R., & Ray, B. (2012). Structure, fluorescence quenching and antioxidant activity of a carbohydrate polymer from Eugenia jambolana. International Journal of Biological Macromolecules, 51, 158-164.
Bandyopadhyay, S. S., Navid, M. H., Ghosh, T., Schnitzler, P., & Ray, B. (2011). Structural features and in vitro antiviral activities of sulfated polysaccharides from Sphacelaria indica. Phytochemistry, 72, 276-283.
Barahona, T., Chandía, N. P., Encinas, M. V., Matsuhiro, B., & Zúñiga, E. A. (2011). Antioxidant capacity of sulfated polysaccharides from seaweed: a kinetic approach. Food Hydrocolloids, 25, 529-535.
Beal, M. F. (2003). Mitochondria, oxidative damage, and inflammation in Parkinson's disease. Annals of the New York Academy of Sciences, 991, 120-131.

65
Belvisi, M. G., & Hele, D. J. (2009). Cough sensors III. Opioid and cannabinoid receptors on vagal sensory nerves. In F. Chung, & J. G. Widdicombe (Eds.), Pharmacology and therapeutics of cough. Berlin/Heidelberg: Springer-Verlag, 63-76.
Benzie, I. F. F., & Strain, J. J. (1996). The ferric reducing ability of plasma (FRAP) as a measure of ‘‘antioxidant power”: The FRAP assay. Analytical Biochemistry, 239, 70–76.
Berteau, O., & Mulloy, B. (2003). Sulfated fucans, fresh perspectives: structures, functions, and biological properties of sulfated fucans and an overview of enzymes active toward this class of polysaccharide. Glycobiology, 13, 29R-40R.
Blakeney, A. B., Harris, P., Henry, R. J., & Bruce, A. B. (1983). A simple rapid preparation of alditol acetates for monosaccharide analysis. Carbohydrate Research, 113, 291–299.
Blakeney, A. B., & Stone, B. A. (1985). Methylation of carbohydrates with lithium methylsulphinyl carbanion. Carbohydrate Research, 140, 319-324.
Boh, B., Berovic, M., Zhang, J., & Zhi-Bin, L. (2007). Ganoderma lucidum and its pharmaceutically active compounds. Biotechnology annual review, 13, 265-301.
Bohn, J. B., & BeMiller, J. N. (1995). (1→3)-β-d-Glucans as biological response modifiers: a review of structure-functional activity relationships. Carbohydrate Polymers, 28, 3–14.
Boisson-Vidal, C., Chaubet, F., Chevolot, L., Sinquin, C., Theveniaux, J., Millet, J., Sternberg, C., Mulloy, B., & Fischer, A. M. (2000). Relationship between antithrombotic activities of fucans and their structure. Drug Development Research, 51, 216–224.
Borchers, A. T., Keen, C. L., M. Gershwin, & M. E. (2004). Mushrooms, Tumors, and Immunity: An Update. Experimental Biology and Medicine, 229, 393-406.
Carpita, N. C., & Shea, E. M. (1988). Linkage structure of carbohydrates by gas chromatography mass spectrometry (GC-MS) of partially methylated alditol acetates. In Analysis of Carbohydrates by GC and MS; Bermann, C. J., McGinis, G. D., Eds.; CRC Press: BocaRaton, FL, 157-212.
Chan, G. C., Chan, W. K., & Sze, D. M. (2009). The effects of beta-glucan on human immune and cancer cells. Journal of hematology & oncology, 2, 25.
Chandia, N. P., & Matsuhiro, B. (2008).Characterization of a fucoidan from Lessonia vadosa (Phaeophyta) and its anticoagulant and elicitor properties. International Journal of Biological Macromolecules, 42, 235-240.
Chatterjee, U. R., Bandyopadhyay, S. S., Ghosh, D., Ghosal, P. K., & Ray, B. (2011). In vitro anti-oxidant activity, fluorescence quenching study and structural features of carbohydrate polymers from Phyllanthus emblica. International Journal of Biological Macromolecules, 49, 637– 642.
Chattopadhyay, K., Adhikari, U., Lerouge, P., & Ray, B. (2007a). Polysaccharides from Caulerpa racemosa: Purification and structural features. Carbohydrate Polymers, 68, 407.
Chattopadhyay, K., Mateu, C. G., Mandal, P., Pujol, C. A., Damonte, E. B., & Ray, B. (2007b). Galactan sulfate of Grateloupia indica: Isolation, structural features and anti-viral activity. Phytochemistry, 68, 2428-2435.
Chattopadhyay, K., Ghosh, T., Pujol, C. A., Carlucci, M. J., Damonte, E.B., & Ray, B. (2008). Polysaccharides from Gracilaria corticata: Sulfation, chemical characterization and anti-HSV activities. International Journal of Biological Macromolecules, 43, 346–351.
Chattopadhyay, N., Ghosh, T., Sinha, S., Chattopadhyay, K., Karmakar, P., & Ray, B. (2010). Polysaccharides from Turbinaria conoides: structural features and antioxidant capacity. Food Chemistry, 118, 823–829.
Chattopadhyay, N., Nosál’ová, G., Saha, S., Bandyopadhyay, S. S., Flešková, D., & Ray, B. (2011). Structural features and antitussive activity of water extracted polysaccharide from Adhatoda vasica. Carbohydrate Polymers, 83, 1970-1974.

66
Chaturvedi, R. K., & Beal, M. F. (2008). PPAR: a therapeutic target in Parkinson's disease. Journal of neurochemistry, 106, 506-518.
Chauvet, P., Bienvenu, J. G., Theoret, J. F., Latour, J. G., & Merhi, Y. (1999). Inhibition of platelet-neutrophil interactions by fucoidan reduces adhesion and vasoconstriction after acute arterial injury by angioplasty in pigs. Journal of cardiovascular pharmacology, 34, 597–603.
Cheshenko, N., & Herold, B. C. (2002). Glycoprotein B plays a predominant role in mediating herpes simplex virus type 2 attachment and is required for entry and cell-to-cell spread. The Journal of general virology, 83, 2247–2255.
Chevolot, L., Foucault, A., Chaubet, F., Kervarec, N., Sinquin, C., Fisher, A. M., & Boisson-Vidal, C. (1999). Further data on the structure of brown seaweed fucans: relationships with anticoagulant activity. Carbohydrate Research, 319, 154-165.
Chevolot, L., Mulloy, B., Ratiskol, J., Foucault, A., & Colliec-Jouault, S. (2001). A disaccharide repeat unit is the major structure in fucoidans from two species of brown algae. Carbohydrate Research, 330, 529-535.
Chihara, G., Hamuro, J., Maeda, Y.Y., Arai, Y., & Fukuoka, F. (1970). Fractionation and Purification of the Polysaccharides with Marked Antitumor Activity, Especially Lentinan, from Lentinus edodes (Berk.) Sing. (An Edible Mushroom).Cancer Research, 30, 2776-2781.
Chizhov, A. O., Dell, A., Morris, H. R., Haslam, S. M., McDowell, R. A., Shashkov, A. S., Nifant’ev, N. E., Khatuntseva, E. A., & Usov, A. I. (1999). A study of fucoidan from brown seaweed Chorda filum. Carbohydrate Research, 320, 108–119.
Chung, K. F. (2009). Clinical cough VI: The need for new therapies for cough: Disease specific and symptom-related anti-tussives. Handbook of Experimental Pharmacology, 187, 343–368.
Claeson, U. P., Malmfors, T., Wikman, G., & Bruhn, J. G. (2000). Adhatoda vasica: A critical review of ethnopharmacological and toxicological data. Journal of Ethnopharmacology, 72, 1–20.
Colak, E. (2008). New markers of oxidative damage to macromolecules. Journal of medical Biochemistry, 27, 1-16.
Copeland, R., Balasubramaniam, A, Tiwari, V., Zhang, F., Bridges, A., Linhardt, R. J, Shukla, D., & Liu, J. (2008).Using a 3-O-sulfated heparin octasaccharide to inhibit the entry of herpes simplex virus type 1. Biochemistry, 47, 5774–5783.
Cornford, C. S., Morgan, M., & Ridsdale, L. (1993). Why do mothers consult when their children cough? Family Practice, 10, 193–196.
Craigie, J. S., Wen, Z. C., & van der Meer, J. P. (1984). Interspecific, Intraspecific and Nutritionally-Determined Variations in the Composition of Agars from Gracilaria spp. Botanica Marina, XXVII, 55-61.
Cumashi, A., Ushakova, N. A., Preobrazhenskaya, M. E., D’Incecco, A., Piccoli, A., Totani, L., Tinari, N., et al. (2007). A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology, 17, 541–552.
Daniel, R., Berteau, O., Chevolot, L., Varenne, A., Gareil, P., & Goasdoue, N. (2001). Regioselective desulfation of sulfated L-fucopyranoside by a new sulfoesterase from the marine mollusk Pecten maximus: application to the structural study of algal fucoidan (Ascophyllum nodosum). European journal of biochemistry / FEBS, 268, 5617-5626.
Daniel, R., Berteau, O., Jozefonvicz, J., & Goasdoue, N. (1999). Degradation of algal (Ascophyllum nodosum) fucoidan by an enzymatic activity contained in digestive glands of the marine mollusc Pecten maximus. Carbohydrate Research, 322(3-4), 291–297.

67
DeBelder, A. N. (1993). In ‘‘Industrial Gums’’,eds, Whistler R. L., BeMiller J. N., Academic Press, INC, 399.
Dhuley, J. N. (1999). Antitussive effect of A. vasica extract on mechanical and chemical stimulation-induced coughing in animals. Journal of Ethnopharmacology, 67, 361–365.
Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., & Smith, F. (1956).Colorimetric method for determination of sugars and related substances. Analytical Chemistry, 28, 350–366.
Edenharder, R., Leopold, C., & Kries, M. (1995). Modifying actions of solvent extracts from fruit and vegetable residues on 2-amino-3-methylimidazo[4,5-f]quinoline (IQ) and 2-amino-3,4-dimethylimidazo[4,5-f]quinoxaline (MeIQx) induced mutagenesis in Salmonella typhimurium TA 98. Mutation research, 341, 303-318.
Ekblad, M., Adamiak, B., Bergstrom, T., Johnstone, K. D., Karoli, T., Liu, L., Ferro, V., & Trybala, E. (2010). A highly lipophilic sulfated tetrasaccharide glycoside related to muparfostat (PI-88) exhibits virucidal activity against herpes simplex virus. Antiviral Research, 86, 196–203.
Ernst, E. (1998). Complementary therapies for asthma: What patient use? The Journal of Asthma: Official Journal of the Association for the Care of Asthma, 35, 667–671.
Falshaw, R., & Furneaux, R. H. (1998). Structural analysis of carrageenans from the tetrasporic stages of the red algae Gigartina lanceata and Gigartina chapmanii (Gigartinaceae, Rhodophyta). Carbohydrate Research, 307, 325–331.
Fan, L., Jiang, L., Xu, Y., Zhou, Y., Shen, Y., Xie, W., Long, Z., & Zhou, J. (2011). Synthesis and anticoagulant activity of sodium alginate sulfates. Carbohydrate Polymers, 83, 1797–1803.
Farias, E. H. C., Pomin, V. H., Valente, A. P., Nader, H. B., Rocha, H. A. O., & Mourao, P. A. S. (2008). A preponderantly 4-sulfated, 3-linked galactan from the green alga Codium isthmocladum. Glycobiology, 18, 250–259.
Farias, W. R. L., Valente, A. P., Pereira, M. S., & Mourao, P. A. S. (2000).Structure and anticoagulant activity of sulfated galactans. Isolation of a unique sulfated galactan from the red algae Botryocladiaoccidentalis and comparison of its anticoagulant action with that of sulfated galactans from invertebrates. Journal of Biological Chemistry, 275, 29299–29307.
Feizi, T. (2000). Carbohydrate-mediated recognition systems in innate immunity. Immunological Reviews, 173, 79-88.
Feyerabend, T. B., Li, J. P., Lindahl, U., & Rodewald, H. R. (2006). Heparan sulfate C5-epimerase is essential for heparin biosynthesis in mast cells. Nature Chemical Biology, 2,195-196.
Franz, G., Pauper, D., & Alban, S. (2000). In B. S. Paulsen (Ed.), Bioactive carbohydrate polymers, Dordrecht: Kluwer Academic Publishers, 47–58.
French, C. T., Irwin, R. S., & Fletcher, K. E. (2002). Evaluation of a cough-specific quality-of-life questionnaire. Chest, 121, 1123–1131.
Fujihara, M., & Nagumo, T. (1993). An influence of the structure of alginate on the chemotactic activity of macrophages and the antitumor activity. Carbohydrate Research, 243, 211–216.
Fujimoto, K., Tomonaga, M., & Goto, S. (2006). A case of recurrent ovarian cancer successfully treated with adoptive immunotherapy and lentinan. Anticancer Research, 26, 4015-4018.
Ghosh, T., Auerochs, S., Saha, S., Ray, B., & Marschall, M. (2010). Anti-cytomegaloviral activity of sulfated glucans generated from a commercial preparation of rice bran. Antiviral Chemistry & Chemotherapy, 21, 85–95.
Ghosh, T., Chattopadhyay, K., Marschall, M., Karmakar, P., Mandal, P., & Ray, B. (2009a). Focus on antivirally active sulfated polysaccharides: From structure–activity analysis to clinical evaluation. Glycobiology, 19, 2–15.

68
Ghosh, T., Pujol, C. A., Damonte, E. B., Sinha, S., & Ray, B. (2009b). Sulfated xylomannans from the red seaweed Sebdenia polydactyla: structural features, chemical modification and antiviral activity. Antiviral Chemistry & Chemotherapy, 19, 235–242.
Glauser, B. F., Rezende, R. M., Melo, F. R., Pereira, M. S., Francischetti, I. M. B., Monteiro, R. Q., Rezaie, A. R., & Mourao, P. A. S. (2009). Anticoagulant activity of a sulfated galactan: Serpin-independent effect and specific interaction with factor Xa. Journal of Thrombosis and Haemostasis, 102, 1183–1193.
Gordon, M., Bihari, B., Goolsby, E., Gorter, R., Guralnik, M., Mimura, T., et al. (1998). A placebo e controlled trial of immune modulator, lentinan, in HIV positive patients: a Phase I/II trial. Journal of Medicine, 29, 305-330.
Gordon, M., Guralnik, M., Kaneko, Y., Mimura, T., Goodgame, J., & Lang, W. (1995). Further clinical studies of curdlan sulfate (CRDS)-an anti-HIV agent. Journal of Medicine, 26, 97-131.
Grasdalen, H. (1983). High-field 1H NMR spectroscopy of alginate: Sequential structure and linkage conformations. Carbohydrate Research, 118, 255–260.
Gunay, N. S., & Linhardt, R. J. (1999). Heparinoids: Structure, Biological Activities and Therapeutic Applications. Planta Medica, 65, 301-306.
Guven, K. V., Ozsoy, Y., & Ulutin, O. N. (1991). Anticoagulant, fibrinolytic and antiaggregant activity of carrageenans and alginic acid. Botanica Marina, 34, 429-432.
Halliwell, B., & Gutteridge, J. M. (1995). The definition and measurement of antioxidants in biological systems. Free Radical Biology & Medicine, 18, 125-126.
Halliwell, B., & Gutteridge, J. M. C. (1984). Oxygen toxicity, oxygen radicals, transition metals and disease. Biochemical Journal, 219, 1–4.
Hannah, B. P., Heldwein, E. E., Bender, F. C., Cohen, G. H., & Eisenberg, R. J.(2007). Mutational evidence of internal fusion loops in herpes simplex virus glycoprotein B. Journal of virology, 81, 4858–4865.
Heinecke, J. W. (1997). Mechanisms of oxidative damage of low density lipoprotein in human atherosclerosis. Current Opinion in Lipidology, 8, 268-274.
Heldwein, E. E, Lou, H., Bender, F. C, Cohen, G. H. Eisenberg R. J, & Harrison SC. (2006).Crystal structure of glycoprotein B from herpes simplex virus 1. Science.313, 217–220.
Hensel, A., & Meier, K. (1999). Pectins and xyloglucans exhibit antimutagenic activities against nitroaromatic compounds. Planta Medica, 65, 395-399.
Herold, B. C., Visalli, R. J., Susmarski, N., Brandt, C. R., & Spear, P.G. (1994). Glycoprotein C-independent binding of herpes simplex virus to cells requires cell surface heparan sulphate and glycoprotein B. The Journal of general virology,75, 1211–1222.
Herold, B. C., WuDunn, D., Soltys, N., & Spear, P. G. (1991). Glycoprotein C of herpes simplex virus type 1 plays a principal role in the adsorption of virus to cells and in infectivity. Journal of virology, 65, 1090–1098.
Hileman, R. E., Fromm, J. R., Weiler, J. M., & Linhardt, R. J. (1998). Glycosaminoglycan-protein interactions: definition of consensus sites in glycosaminoglycan binding proteins. BioEssays, 20, 156-167.
Hirano, M., Kiyohara, H., & Yamada, H. (1994). Existence of a Rhamnogalacturonan II-like Region in Bioactive Pectins from Medicinal Herbs. Planta Medica, 60, 450-454.
Hobbs, C. R. (2004). Medicinal value of Turkey Tail fungus Trametes versicolor (L.:Fr.) Pilát (Aphyllophoromycetideae). International journal of medicinal mushrooms, 6,195–218.

69
Holst, O. (1999). In ‘‘Endotoxin in Health and Disease’’, eds, Brade H., Morrison D. C., Opal S., Vogel S., Marcel Dekker, Inc., New York, 115.
Hromadkova, Z., Paulsen, B. S., Polovka, M., Kostalova, Z., & Ebringerova, A. (2012). Structural features of two heteroxylan polysaccharide fractions from wheat bran with anti-complementary andantioxidant activities. Carbohydrate Polymers, DOI: http://dx.doi.org/10.1016/ j. carbpol. 2012.05.021.
Hu, T., Liu, D., Chen, Y., Wu, J., & Wang, S. (2010). Antioxidant activity of sulfated polysaccharide fractions extracted from Undaria pinnitafida in vitro. International Journal of Biological Macromolecules, 46, 193-198.
Huang, D., Ou, B., & Prior, R. L. (2005). The Chemistry behind antioxidant capacity assays. Journal of Agricultural and Food Chemistry, 53, 1841–1856.
Huq, M. E., Ikram, M., & Warsi, S. A. (1967). Chemical composition of Adhatoda vasica Linn. II. Pakistan Journal of Scientific and Industrial Research, 10, 224-225.
Imeson, A. P. (2000). In “Handbook of hydrocolloids”, ed, Phillips, G. O.;Williams, P. A, 87-102.
Inngjerdingen, K. T., Coulibaly, A.; Diallo, D., Michaelsen, T. E., & Paulsen, B. S. (2006). A complement fixing polysaccharide from Biophytum petersianum Klotzsch, a medicinal plant from Mali, West Africa. Biomacromolecules, 7, 48–53.
Inngjerdingen, K. T., Kiyohara, H., Matsumoto, T., Petersen, D., Michaelsen, T. E., Diallo, D., Inngjerdingen, M., Yamada, H., & Paulsen, B. S. (2007). An immunomodulating pectic polymer from Glinus oppositifolius. Phytochemistry, 68, 1046-1058.
Jain, M. P., & Sharma, V. K. (1982). Phytochemical Investigation of Roots of AdhatodaVasica. Planta Medica, 46, 250-250.
Jain, M. P., Koul, S. K., Dhar, K. L., & Atal, C. K. (1980). Novel nor-Harmal Alkaloid from Adhatoda-Vasica. Phytochemistry, 19, 1880-1882.
Jain, S. K. (2006). Superoxide dismutase overexpression and cellular oxidative damage in diabetes. A commentary on "Overexpression of mitochondrial superoxide dismutase in mice protects the retina from diabetes-induced oxidative stress". Free radical biology & medicine, 41, 1187-1190.
Je, J. Y., Park, P. J., Kim, E. K., Park, J. S., Yoon, H. D., Kim, K. R., et al., (2009). Antioxidant activity of enzymatic extracts from the brown seaweed Undariapinnatifida by electron spin resonance spectroscopy. LWT – Food Science and Technology, 42, 874–878.
Jiang, Y. H., Jiang, X. L., Wang, P., Mou, H. L., Hu, X. K., & Liu, S. Q. (2008). The antitumor and antioxidative activities of polysaccharides isolated from Isaria farinosa B05. Microbiological Research, 163, 424–430.
Kapoor, L. D. (1990). CRC handbook of ayurvedic medicinal plants. Boca Raton: CRC Press.
Karinaga, R., Mizu, M., Koumoto, K., Anada, T., Shinkai, S., Kimura, T., et al., (2004). First observation by fluorescence polarization of complexation between mRNA and the natural polysaccharide schizophyllan. Chem Biodivers, 1, 634–639.
Kariya, Y., Mulloy, B., Imai, K., Tominaga, A., Kaneko, T., Asari, A., et al., (2004). Isolation and partial characterization of fucan sulphates from the body wall of sea cucumber Stichopus japonicus and their ability to inhibit osteoclastogenesis. Carbohydrate Research, 339, 1339–1346.
Karmakar, P., Ghosh, T., Sinha, S., Saha, S., Mandal, P., Ghosal, P. K., & Ray, B. (2009). Polysaccharides from the brown seaweed Padina tetrastromatica: Characterization of a sulfated fucan. Carbohydrate Polymers, 78, 416–421.

70
Karmakar, P., Pujol, C.A., Damonte, E.B., Ghosh, T., & Ray, B. (2010). Polysaccharides from Padina tetrastromatica: Structural features, chemical modification and antiviral activity. Carbohydrate Polymers 80, 514–521.
Kataoka-Shirasugi, N., Ikuta, J., Kuroshima, A., & Misaki, A. (1994). Antitumor Activities and Immunochemical Properties of the Cell-wall Polysaccharides from Aureobasidium pullulans. Bioscience, biotechnology, and biochemistry, 58, 2145-2151.
Kawabata, H., Germain, R. S., Vuong, P. T., Nakamaki, T., Said, J. W., & Koeffler, H. P. (2000). Transferrin Receptor 2-a Supports Cell Growth Both in Iron-chelated Cultured Cells and in Vivo. Journal of Biological Chemistry, 275, 16618-16625.
Kennedy, J. F., & White, C. A. (1988). In Carbohydrate Chemistry (Kennedy J. F ed) Clarendon Press Oxford, 220.
Kiyohara, H., Cyong, J. C., &Yamada, H. (1989). Relationship between structure and activity of the “ramified” region in anti-complementary pectic polysaccharides from Angelica acutiloba kitagawa. Carbohydrate Research, 193, 201-214.
Kiyohara, H., Yamada, H., Takemoto, N., Kawamura, H., Komatsu, Y., & Oyama, T. (1993).Characterization of in vitro IL-2 production enhancing and anticomplementary pectic polysaccharides from kampo (Japanese herbal) medicine ‘Juzen-Taiho-To’. Phytotherapy Research, 7, 367-375.
Kleymann, G. (2005). Agents and strategies in development for improved management of herpes simplex virus infection and disease. Expert opinion on investigational drugs, 14, 135–161.
Kodama, N., Komuta, K., & Nanba, H. (2003).Effect of Maitake (Grifola frondosa) D-Fraction on the activation of NK cells in cancer patients. Journal of medicinal food, 6, 371-377.
Kogan, G., Šoltés, L., Stern, R., & Gemeiner, P. (2007). Hyaluronic acid: a natural biopolymer with a broad range of biomedical and industrial applications. Biotechnology Letters, 29, 17-25.
Kubal J. V., & Gralen, N. (1948). Industrial Gums: Polysaccharides and Their Derivatives. Journal of Colloid Science, 3, 457-471.
Kumar, S. S., Kumar, Y., Khan, M. S., & Gupta, V. (2010). New antifungal steroids from Turbinaria conoides (J. Agardh) Kutzing. Natural Product Research, 24, 1481-1487.
Kumar, S. S., Kumar, Y., Khan, M. S., Anbu, J., & De Clercq, E. (2011). Antihistaminic and antiviral activities of steroids of Turbinaria conoides. Natural Product Research, 25, 723-729.
Laquerre, S., Argnani, R., Anderson, D. B., Zucchini, S., Manservigi, R., & Glorioso, J. C. (1998). Heparan sulfate proteoglycan binding by herpes simplex virus type 1 glycoproteins B and C, which differ in their contributions to virus attachment, penetration, and cell-to-cell spread. Journal of virology, 72, 6119– 6130.
Latge, J. P., Kobayashi, H., Debeaupuis, J. P., Diaquin, M., Sarfati, J., Wieruszeski, J.M., Parra, E, Bouchara, J. P., & Fournet, B. (1994). Chemical and immunological characterization of the extracellular galactomannan of Aspergillus fumigatus. Infection and Immunity, 62, 5424-5433.
Lee, S. H., Athukorala, Y., Lee, J. S., & Jeon, Y. J. (2008). Simple separation of anticoagulant sulfated galactan from red algae. Journal of Applied Physiology, 20, 1053–1059.
Lichtenthaler, F., & Mondel, S. (1997). Carbohydrates as Organic Raw Materials. Pure and Applied Chemistry, 69, 1853-1866.
Linhardt, R. J., & Toida, T. (1997). Heparin Oligosaccharides: New Analogues Development and Applications, in: Carbohydrate in Drug Designing, (Zbigniew, J. W., Karl, A. N., eds.), Marcel Dekker, Inc., New York, 277-341.
Liu, J., & Mori, A. (2006). Oxidative damage hypothesis of stress-associated aging acceleration:

71
neuroprotective effects of natural and nutritional antioxidants. Research Communications in Biological Psychology, Psychiatry & Neuroscience, 30-31, 103-119.
Lowry, O. H., Rosebrough, N. J., Lewsfarr, A., & Randall, R. J. (1951).Protein measurement with the folin reagent. Journal of Biological Chemistry, 193, 265–275.
Maeda, M., Totomu, U., Harada, N., Sekiguchi, M., & Hiraoka, A. (1991). Heparinoid-active sulphated polysaccharides fromMonostroma nitidum and their distribution in the chlorophyta. Phytochemistry, 30, 3611-3614.
Maehara, Y., Tsujitani, S., Saeki, H., Oki, E., Yoshinaga, K., Emi, Y., Morita, M., Kohnoe, S., Kakeji, Y., Yano, T., & Hideo Baba, H. (2012). Biological mechanism and clinical effect of protein-bound polysaccharide K (KRESTIN): review of development and future perspectives. Surgery Today, 42, 8–28.
Mandal, P., Mateu, C. G., Chattopadhyay, K., Pujol, C. A., Damonte, E. B., & Ray, B. (2007). Structural features and antiviral activity of sulphated fucans from the brown seaweed Cystoseira indica. Antiviral Chemistry and Chemotherapy, 18,153–162.
Mandal, P., Pujol, C. A., Carlucci, M. J., Chattopadhyay, K., Damonte, E. B., & Ray, B. (2008). Sulfated xylomannan of Scinaia hetai: Isolation, structural features and antiviral activity. Phytochemistry, 69, 2193–2199.
Mandal, P., Pujol, C. A., Damonte, E. B., Ghosh, T., & Ray, B. (2010). Xylans from Scinaia hatei: Structural features, sulfation and anti-HSV activity. International Journal of Biological Macromolecules, 46, 173–178.
Mantovani, M. S., Bellini, M. F., Angeli, J. P. F., Oliveira, R. J., Silva, A. F., & Ribeiro, L. R. (2008). beta-Glucans in promoting health: prevention against mutation and cancer. Mutation Research, 658, 154−161.
Mardberg, K., Trybala, E., Glorioso, J. C., & Bergstr¨om, T. (2001).Mutational analysis of the major heparin sulfate-binding domain of herpes simplex virus type 1 glycoprotein C. The Journal of general virology,82, 1941–1950.
Mardberg, K., Trybala, E., Tufaro, F., & Bergstr¨om, T. (2002). Herpes simplex virus glycoprotein C is necessary for efficient infection of chondroitin sulphate expressing gro2C cells. The Journal of general virology, 83, 291–300.
Maruyama, H., & Ikekawa, T. (2007). Immunomodulation and antitumoractivity of a mushroom product, proflamin, isolated from Flammulina velutipes (W. Curt:Fr.) Singer(Agaricomycetideae). International journal of medicinal mushrooms 9,109–122.
Maruyama, H., Tamauchi, H., Iizuka, M., & Nakano, T. (2006). The role of NK cells in antitumor activity of dietary fucoidan from Undaria pinnatifida sporophylls (Mekabu). Planta Medica, 72, 1415-1417.
Matsubara, K., Matsuura, Y., Bacic, A., Liao, M.L., Hori, K., & Miyazawa, K. (2001). Anticoagulant properties of a sulfated galactan preparation from a marine green alga, Codium cylindricum. International Journal of Biological Macromolecules, 28, 395–399.
Matsukawa, R., Dubinsky, Z., Kishimoto, E., Masaki, K., Masuda, Y., & Takeuchi, T. (1997).A comparison of screening methods for antioxidant activity in seaweeds. Journal of Applied Phycology, 9, 29–35.
Matsushima, H., Minoshima, H., Kawanami, H., Ikushima, Y., Nishizawa, M., Kawamukai, A., et al., (2005). Decomposition reaction of alginic acid using subcritical and supercritical water. Industrial and Engineering Chemistry Research, 44, 9626–9630.
May, C. D. (1990). Industrial pectins: Sources, production and applications. Carbohydrate Polymers, 12, 79-99.

72
Mayer, A. M. S., & Lehmann, V. K. B. (2000). Marine pharmacology in 1998: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, anthelmintic, antiplatelet, antiprotozoal and antiviral activities; with actions on the cardiovascular, endocrine, immune and nervous systems: And other miscellaneous mechanisms of action. Pharmacologist, 42, 62–69.
Mazumder, S., Morvan, C., Thakur, S., & Ray, B. (2004).Cell wall polysaccharides from Chalkumra (Benincasa hispida) fruit.Part I. Isolation and characterization of pectins. Journal of Agriculture and Food Chemistry, 52, 2356–2362.
Meer, W. (1980). In ‘‘Handbook of water soluble gums and resins’’, ed, Davidson R. I., McGraw Hill, New York, 81.
Mizuno, M., Minato, K., Ito, H., Kawade, M., Terai, H., & Tsuchida, H. (1999).Anti-tumor polysaccharide from the mycelium of liquid-cultured Agaricus blazei mill. Biochemistry & Molecular Biology International, 47, 707-714.
Moreira, P. I., Smith, M. A., Zhu, X., Honda, K., Lee, H. G., Aliev, G., & Perry, G. (2005). Oxidative damage and Alzheimer's disease: are antioxidant therapies useful? Drug News Perspect, 18, 13-19.
Mourao, P. A. S. (2004). Use of sulfated fucans as anticoagulantand antithrombotic agents: Future perspectives. Current Pharmaceutical Design, 10, 967–981.
Mourao, P. A. S., Pereira, M. S., Pavao, M. S. G., Mulloy, B., Tollefsen, D. M., Mowinckel, M. C., et al., (1996). Structure and anticoagulant activity of a fucosylated chondroitin sulphate from Echinoderm. Journal of Biological Chemistry, 271, 23973–23984.
Mukherjee, A. B., Zhang, Z., & Chilton, B. S. (2007). Uteroglobin: a steroid-inducible immunomodulatory protein that founded the Secretoglobin superfamily. Endocrine reviews, 28, 707-725.
Mulloy, B. (2005). The specificity of interactions between proteins and sulfated polysaccharides. Anais da Academia Brasileira de Ciências, 77, 651-664.
Mulloy, B., & Linhardt, R. J. (2001). Order out of complexity--protein structures that interact with heparin.Current opinion in structural biology, 11, 623-628.
Murugesan, S., Xie, J., & Linhardt, R. J. (2008). Immobilization of Heparin: Approaches and Applications. Current Topics in Medicinal Chemistry,8, 80-100.
Nanba, H., Hamaguchi, A., & Kuroda, H. (1987). The chemical structure of an antitumor polysaccharide in fruit bodies of Grifola frondosa (maitake).Chemical & pharmaceutical bulletin, (Tokyo), 35, 1162-1168.
Nasra, J., & Belvisi, M. G. (2009). Modulation of sensory nerve function and the cough reflex: Understanding disease pathogenesis. Pharmacology & Therapeutics, 124, 354–375.
Nishino, T., & Nagumo, T. (1991). The sulfate-content dependence of the anticoagulant activity of a fucan sulfate from the brown seaweed Ecklonia kurome. Carbohydrate Research, 214, 193-197.
Nishino, T., & Nagumo, T. (1992). Anticoagulant and antithrombin activities of oversulfated fucans. Carbohydrate Research, 229, 355-362.
Nishino, T., Nishioka, C., Ura, H., & Nagumo, T. (1994). Isolation and partial characterization of a novel amino sugar-containing fucan sulfate from commercial Fucus vesiculosus fucoidan. Carbohydrate Research, 255, 213-224.
Nishino, T., Yokoyama, G., Dobashi, K., Fujihara, M., & Nagumo, T. (1989). Isolation, purification, and characterization of fucose-containing sulfated polysaccharides from the brown seaweed Ecklonia kurome and their blood-anticoagulant activities. Carbohydrate Research, 186, 119-129.

73
Nosál’ová, G., Sutovska, M., Mokry´, J., Kardosˇová, A., Capek, P., & Khan, T. H. M. (2005). Efficacy of the herbal substances according to cough reflex. Minerva Biotechnologia, 17, 141–152.
Nosáľová, G., Prisenžňáková, Ľ., Košťálová, Z., Ebringerová, A., & Hromádková. Z., (2011).Suppressive effect of pectic polysaccharides from Cucurbita pepo L. var. Styriaca on citric acid-induced cough reflex in guinea pigs. Fitoterapia, 82, 357-364.
Nosáľová, G., Capek, P., Matáková, T., Nosáľ, S., Flešková, D., & Jureček, Ľ. (2012).Antitussive activity of an extracellular Rhodella grisea proteoglycan on the mechanically induced cough reflex. Carbohydrate Polymers, 87, 752-756.
Olafsdottir, E. S., Ingolfsdottir, K., Barsett, H., Paulsen, B. S., Jurcic, K., &Wagner, H. (1999a). Immunologically active (1→3)-(1→4)-α-D-Glucan from Cetraria islandica. Phytomedicine,6, 33-39.
Olafsdottir, E. S., Omarsdottir, S., Paulsen, B. S., Jurcic, K., & Wagner, H. (1999b). Rhamnopyranosylgalactofuranan, a new immunologically active polysaccharide from Thamnolia subuliformis. Phytomedicine, 6, 273-279.
Olafsdottir, E. S., Omarsdottir, S., Paulsen, B. S., & Wagner, H. (2003). Immunologically active O6-branched (1-->3)-beta-glucan from the lichen Thamnolia vermicularis var. subuliformis. Phytomedicine, 10, 318-324.
Olofsson, S., & Bergstrom, T. (2005). Glycoconjugate glycans as viral receptors. Annals of Medicine.37,154–172.
Ooi, V. E., & Liu, F. (2000). Immunomodulation and anti-cancer activity of polysaccharide-protein complexes.Current Medicinal Chemistry, 7, 715-729.
Ou, S. Y., Jackson, G. M., Jiao, X., Chen, J., Wu, J. Z., & Huang, X. S. (2007). Protection against oxidative stress in diabetic rats by wheat bran feruloyl oligosaccharides. Journal of Agricultural and Food Chemistry, 55, 3191–3195.
Painter, T. J. (1983). In: Aspinall G. O, editor. The polysaccharides, Academic Press Inc, 2, 195.
Patankar, M. S., Oehninger, S., Barnett, T., Williams, R. L., & Clark, G. F. (1993). A revised structure for fucoidan may explain some of its biological activities. The Journal of biological chemistry, 268, 21770-21776.
Paulsen, B., & Samuelsen, A. B. (2001). Plantago Species. Harwood Academic Publishers GmBH, Amsterdam.
Pavord, I. D. (2004). Cough and asthma. Pulmonary Pharmacology & Therapeutics, 17, 399–402.
Pavord, I. M., & Chung, K. F. (2008). Chronic cough: Treatment and management. The Lancet, 371, 1375–1384.
Paz-Elizur, T., Sevilya, Z., Leitner-Dagan, Y., Elinger, D., Roisman, L. C., & Livneh, Z. (2008). DNA repair of oxidative DNA damage in human carcinogenesis: potential application for cancer risk assessment and prevention. Cancer letters, 266, 60-72.
Pennock, B. E., Cox, C. P., Rogers, R. M., Cain, W. A., & Wells, J. H. (1979). A noninvasive technique for measurement of changes in specific airway resistance. Journal of Applied Physiology, 46, 399–406.
Pereira, M. S., Mulloy, B., & Mourao, P. A. S. (1999). Structure and anticoagulant activity of sulfated fucans: Comparison between the regular repetitive and linear fucansfrom echinoderms with the more heterogeneous and branched polymersfrom brown algae. Journal of Biological Chemistry, 274, 7656–7667.
Pereira, M. S., Silva, A. C. E. S. V., Valente, A. P., & Mourao, P. A. S. (2002). A 2-sulfated, 3-linked α-L-galactan is an anticoagulant polysaccharide. Carbohydrate Research, 337, 2231–2238.

74
Piculell, L. (1995). In ‘‘Food Polysaccharides and their Applications’’, ed, Stephen A. M., Marcel Dekker, Inc, Newyork, 2005.
Pomin, V. H., & Mourao, P. A. S. (2008). Structure, biology, evolution, and medical importance of sulfated fucans and galactans. Glycobiology, 18, 1016–1027.
Pomin, V. H., Pereira, M. S., Valente, A. P., Tollefsen, D. M., Pavao, M. S., & Mourao, P. A. S. (2005). Selective cleavage and anticoagulant activity of a sulfated fucan: stereospecific removal of a 2-sulfate ester from the polysaccharide by mild acid hydrolysis, preparation of oligosaccharides, and heparin cofactor II-dependent anticoagulant activity. Glycobiology, 15, 369-381.
Potin, P., Patier, P., Floc'h, J. Y., Yvin, J. C., Rochas, C., & Kloareg, B. (1992). Chemical characterization of cell-wall polysaccharides from tank-cultivated and wild plants of Delesseria sanguinea (Hudson) Lamouroux (Ceramiales, delesseriaceae): culture patterns and potent anticoagulant activity. Journal of Applied Phycology, 4, 119-128.
Pulido, R., Bravo, L., & Saura-Calixto, F. (2000).Antioxidant activity of dietary polyphenols as determined by a modified ferric reducing/antioxidant power assay. Journal of Agricultural and Food Chemistry, 48, 3396–3402.
Qi, H., Zhang, Q., Zhao, T., Chen, R., Zhang, H., Niu, X., & Li, Z. (2005). Antioxidant activity of different sulfate content derivatives of polysaccharide extracted from Ulva pertusa (Chlorophyta) in vitro. International Journal of Biological Macromolecules, 37, 195-199.
Qi, H., Zhang, Q., Zhao, T., Hu, R., Zhang, K., & Li, Z. (2006). In vitro antioxidant activity of acetylated and benzoylated derivatives of polysaccharide extracted from Ulva pertusa (Chlorophyta). Bioorganic & medicinal chemistry letters, 16, 2441-2445.
Quiocho F. A., & Vyas, N. K. (1999). In Bioorganic Chemistry: Carbohydrates (Hecht, S. M. edited), Oxford University Press, New York, 441-457.
Ramallo, I. A., Salazar, M. O., Mendez, L., & Furlan, R. L. E. (2011). Chemically Engineered Extracts: Source of Bioactive Compounds. Accounts of Chemical Research, 44, 241-250.
Ray, B. (2006). Polysaccharides from Enteromorpha compressa: isolation, purification and structural features. Carbohydrate Polymers, 66, 408-416.
Ray, B., & Lahaye, M. (1995). Cell-wall polysaccharides from the marine green algae Ulva rigida (Ulvales, Chlorophyta). Chemical structure of ulvan. Carbohydrate Research, 274, 313-318.
Rechter, S., K¨onig, T., Auerochs, S., Thulke, S., Walter, H., D¨ornenburg, H., Walter, C., & Marschall, M. (2006). Antiviral activity of arthrospira-derived spirulan-like substances. Antiviral research,72(3), 197–206.
Renn, D. W. (1997). Biotechnology and the red seaweed polysaccharide industry:Status, needs and prospects. TIBTECH, 15, 9–14.
Rocha de Souza, M. C., Marques, C. T., Dore, C. M. G., Ferreira da Silva, F. R., Rocha, H. A. O., & Leite, E. L. (2007). Antioxidant activities of sulphated polysaccharides from brown and red seaweeds.Journal of Applied Phycology, 19, 153–160.
Rochas, C., Lahaye, M., & Yaphe, W., (1986). Sulfate content of carrageenan and agar determined by infrared spectroscopy. Botanica Marina,29, 335-340.
Rojers, D. J., Jurd, K. M., Blunden, G., Paoletti, S., & Zanetti, F. (1990). Journal of Applied Physiology, 2, 357.
Rolin, C., & De Vries, J. (1990). In ‘‘Pectin, Food Gels’’, ed, P. Harris, Elsevier Applied Science, New York, 401.
Ruperez, P. (2001). Antioxidant activity of sulphated polysaccharides from the Spanish marine seaweed Nori. In Proceedings of the COST 916 European conference on bioactive compounds in plant

75
foods. Tenerife, Canary Islands, Spain: Health Effects and Perspectives for the Food Industry, 114.
Ruperez, P., Ahrazem, O., & Leal, A. (2002). Potential antioxidant capacity ofsulphated polysaccharides from the edible marine brown seaweed Fucus vesiculosus. Journal of Agricultural and Food Chemistry, 50, 840–845.
Rusnati, M., Vicenzi, E., Donalisio, M., Oreste, P., Landolfo, S., & Lembo, D. (2009). Sulfated K5 Escherichia coli polysaccharide derivatives: A novel class of candidate antiviral microbicides. Pharmacology and Therapeutics, 123, 310–322.
Rydberg, E., Westfall, M. J., Nicholas, R. A. (1999). Low Molecular weight heparin in preventing and treating DVT. American Family Physician, 59, 1607-1612.
Saha, S., Galhardi, L. C. F., Yamamoto, K. A., Linhares, R. E. C., Bandyopadhyay, S. S., Sinha, S., Nozawa, C., & Ray, B. (2010). Water-extracted polysaccharides from Azadirachta indica leaves: Structural features, chemical modification and anti-bovine herpesvirus type 1 (BoHV-1) activity. International Journal of Biological Macromolecules, 47, 640–645.
Saha, S., Navid, M. H., Bandyopadhyay, S. S., Schnitzler, P., & Ray, B. (2012). Sulfated Polysaccharides from Laminaria angustata: Structural features and in vitro antiviral activities. Carbohydrate polymers, 87, 123-130.
Saha, S., Nosál’ová, G., Ghosh, D., Flešková, D., Capek, P., & Ray, B. (2011). Structural features and in vivo antitussive activity of the water extracted polymer from Glycyrrhiza glabra. International Journal of Biological Macromolecules,48, 634-638.
Sahoo, D. (2010). Common Seaweeds of India. I. K. International Pvt Ltd, 102.
Saito, H., Ohki, T., & Sasaki, T. (1979). A 13C-nuclear magnetic resonance study of polysaccharide gels. Molecular architecture in the gels consisting of fungal, branched (1 → 3)-β-D-glucans (lentinan and schizophyllan) as manifested by conformational changes induced by sodium hydroxide. Carbohydrate Research, 74, 227-240.
Saito, H., Ohki, T., Takasuka, N., & Sasaki, T. (1977).A 13C-N.M.R.-Spectral study of a gel-forming, branched (1→3)-β-d-glucan, (lentinan) from lentinus edodes, and its acid-degraded fractions.Structure, and dependence of conformation on the molecular weight. Carbohydrate Research, 58, 293–305.
Sakurai, M. H., Matsumoto, T., Kiyohara, H., & Yamada, H. (1999). B-cell proliferation activity of pectic polysaccharide from a medicinal herb, the roots of Bupleurum falcatum L. and its structural requirement. Immunology, 973, 540-547.
Samuelsen, A. B., Paulsen, B. S., Wold, J. K., Knutsen, S. H., & Yamada, H. (1998). Characterization of a biologically active arabinogalactan from the leaves of Plantagomajor L. Carbohydrate Polymers, 35, 145-153.
Samuelsen, A. B., Paulsen, B. S., Wold, J. K., Otsuka, H., Kiyohara, H., Yamada, H., & Knutsen, S. H. (1996). Characterization of a biologically active pectin from Plantago major L. Carbohydrate Polymers,30, 37-44.
Sasaki, T., & Takasuka, N. (1976). Further study of the structure of lentinan, an anti-tumor polysaccharide from Lentinus edodes. Carbohydrate Research, 47, 99-104.
Schappert, S. M., & Burt, C. W. (2006). Ambulatory care visits to physician offices, hospital outpatient departments and emergency departments: United States, 2001–02. Vital and Health Statistics, 13, 1–66.
Schneider-Schaulies, J. (2000). Cellular receptors for viruses: Links to tropism and pathogenesis. The Journal of general virology,81, 1413–1429.

76
Schultz, C. R., Johnson, K. L., Currie, G., & Bacic, A. (2000).The classical arabinogalactan protein gene family of Arabidopsis. The Plant Cell, 12, 1751–1767.
Selby, H. H., & Whistler, R. L. (1993). In ‘‘Industrial Gums’’, ed, Whistler R. L., BeMiller J. N., Academic Press, INC, 87.
Sepulveda, R. T., & Watson, R. R. (2002). Treatment of antioxidant deficiencies in AIDS patients. Nutrition research. (N.Y.), 22, 27-37.
Sheu, J. H., & Sung, P. J. (1991). Isolation of 24-hydroperoxy-24-vinylcholesterol and fucosterol from the brown alga Turbinaria conoides. Journal of the Chinese Chemical Society, 38, 501-503.
Sheu, J. H., Wang, G. H., Sung, P. J., & Duh, C. Y. (1999).New cytotoxic oxygenated fucosterols from the brown alga Turbinaria conoides.Journal of Natural Products, 62, 224-227.
Shimada, K., Fujikawa, K., Yahara, K., & Nakamura, T. (1992). Antioxidative properties of xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. Journal of Agricultural and Food Chemistry, 40, 945–948.
Shin, K. S., Kiyohara, H., Matsumoto, T., & Yamada, H. (1998).Rhamnogalacturonan II dimers cross-linked by borate diesters from the leaves of Panax ginseng C.A. Meyer are responsible for expression of their IL-6 production enhancing activities. Carbohydrate Research, 307, 97-106.
Silva, T. M. A., Alves, L. G., Queiroz, K. C. S., Santos, M. G. L., Marques, C. T., Chavante, S. F., Rocha, H. A. O., & Leite, E. L. (2005). Partial characterization and anticoagulant activity of a heterofucan from the brown seaweed Padina gymnospora. Brazilian Journal of Medical and Biological Research, 38, 523-533.
Simões, J. N., Nunes, F. M., Domingues, M. d. R. M., & Coimbra, M. A. (2010).Structural features of partially acetylated coffee galactomannans presenting immunostimulatory activity.Carbohydrate Polymers, 79, 397–402.
Singh, R. S., Misra, T. N., Pandey, H. S., & Singh, B. P. (1991).Aliphatic Hydroxyketones from Adhatoda-Vasica.Phytochemistry, 30, 3799-3801.
Sinha, S., Astani, A., Ghosh, T., Schnitzler, P., & Ray, B. (2010). Polysaccharides from Sargassum tenerrimum: Structural features, chemical modification and antiviral activity. Phytochemistry, 71, 235-42.
Sinha, S. Bandyopadhyay, S. S., Ghosh, D., Chatterjee, U. R., Saha, S., Ghosal, P. K., & Ray, B. (2011a). Structural Characteristics, Fluorescence Quenching, and Antioxidant Activity of the Arabinogalactan Protein-rich Fraction from Senna (Cassia angustifolia) Leaves. Food Science Biotechnology, 20, 1005-1011.
Sinha, S., Nosal’ova, G., Bandyopadhyaya, S. S., Fleˇskovab, D., & Ray, B., (2011b). In vivo anti-tussive activity and structural features of a polysaccharide fraction from water extracted Withania somnifera. Journal of Ethnopharmacology, 134, 510–513.
Spear, P.G. (2004). Herpes simplex virus: Receptors and ligands for cell entry. Cellular microbiology,6, 401–410.
Springer, G. F., Wurzel, H. A., McNeal, Jr. G. M., Ansell, N. J., & Doughty, M. F. (1957).Isolation of anticoagulant fractions from crude fucoidan. Proceedings of the Society for Experimental Biology and Medicine. 94, 404–409.
Stone, B. A., &Clarke, A. E. (1992). Chemistry and biology of (1,3)-D-glucans. Victoria, Australia: La Trobe University Press; 882.
Sultana, N., Anwar, M. A., Ali, Y., & Afza, N. (2005). Phytochemical studies on Adhatoda vasica. Pakistan Journal of Scientific and Industrial Research, 48, 180-183.

77
Sutovska, M., Nosal’ova, G., Fraˇnova, S., & Kardoˇsova, A. (2007). Isolation, characterization, and antitussive activity of polysaccharides from the flowers of Althaea officinalis L., var Robusta, leaves of Arctium lappa L., var. Herkules, and gum exutade of peach tree (Prunus persica L., Batsch). Bratislava Medical Journal, 108, 93–99.
Sutovska, M., Fraňová, S., Prisežnaková, L., Nosáľová, G., Togola, A., Diallo, D., Paulsen, B. S., & Capek, P., (2009a).Antitussive activity of polysaccharides isolated from the Malian medicinal plants, International Journal of Biological Macromolecules, 44, 236-239.
Sutovska, M., Nosáľová, G., Šutovský, J., Fraňová, S., Prisenžňáková, Ľ., & Capek, P. (2009b). Possible mechanisms of dose-dependent cough suppressive effect of Althaea officinalis rhamnogalacturonan in guinea pigs test system. International Journal of Biological Macromolecules, 45, 27-32.
Sutovska, M., Fraňová, S., Sadloňová, V.,Grønhaug, T. E.,Diallo, D.,Paulsen, B. S., & Capek, P. (2010). The relationship between dose-dependent antitussive and bronchodilatory effects of Opilia celtidifolia polysaccharide and nitric oxide in guinea pigs. International Journal of Biological Macromolecules, 47, 508-513.
Sutovska, M., Capek, P., Fraňová, S., Pawlaczyk, I., & Gancarz, R. (2012).Antitussive and bronchodilatory effects of Lythrum salicariapolysaccharide-polyphenolic conjugate. International Journal of Biological Macromolecules, 51, 794-799.
Sutovska, M., Capek, P., Kocmálová, M., Fraňová, S., Pawlaczyk, I., & Gancarz, R. (2013). Characterization and biological activity of Solidago canadensis complex. International Journal of Biological Macromolecules, 52, 192-197.
Synytsya, A., Kim, W. J., Kim, S. M., Pohl, R., Synytsya, A., Kvasnička, F., Čopíková, J., & Park, Y. I. (2010). Structure and antitumour activity of fucoidan isolated from sporophyll of Korean brown seaweed Undaria pinnatifida. Carbohydrate Polymers, 81, 41-48.
Tal-Singer, R., Peng, C., Ponce de Leon, M., Abrams, W. R., Banfield, B. W., Tufaro, F., Cohen, G. H., & Eisenberg, R. J. (1995). Interaction of herpes simplex virus glycoprotein gC with mammalian cell surface molecules. Journal of virology, 69, 4471– 4483.
Taylor, K. R., & Gallo, R. L. (2006). Glycosaminoglycans and their proteoglycans: host-associated molecular patterns for initiation and modulation of inflammation.The FASEB Journal, 20, 9-22.
Thakur, M., Weng, A., Fuchs, H., Sharma, V., Bhargava, C. S., Chauhan, N. S., Dixit, V. K., & Bhargava.S. (2012). Rasayana properties of Ayurvedic herbs: Are polysaccharides a major contributor. Carbohydrate Polymers, 87, 3-15.
Thappa, R. K., Agarwal, S. G., Dhar, K. L., Gupta, V. K., & Goswami, K. N. (1996). Two pyrroloquinazolines from Adhatoda vasica. Phytochemistry, 42, 1485-1488.
Therkelsen, G. H. (1993). In ‘Industrial Gums: Polysaccharides and Their Derivatives, edited by James N. BeMiller, Roy L. Whistler, Academic Press Inc, 7, 146-180.
Tolstoguzov, V., (2004). Why are polysaccharides necessary? Food Hydrocolloids, 18, 873-877.
Tomoda, M., Ohara, N., Shizimu, N., & Gonda, R. (1994a). Characterization of a novel glucan, which exhibits reticuloendothelial system-potentiating and anti-complementary activities, from the rhizome of Cnidium officinale. Chemical & pharmaceutical bulletin, 42, 630-633.
Tomoda, M., Ohara, N., Shizimu, N., & Gonda, R. (1994b). Characterization of a novel heteroglucan from the rhizome of Cnidium officinale exhibiting high reticuloendothelial system-potentiating and anti-complementary activities. Biological & pharmaceutical bulletin, 17, 973-976.

78
Trybala, E., Bergstr¨om, T., Svennerholm, B., Jeansson, S., Glorioso, J. C., & Olofsson, S. (1994). Localization of a functional site on herpes simplex virus type 1 glycoprotein C involved in binding to cell surface heparan sulphate. The Journal of general virology, 75, 743–752.
Tseng C. K. (1946). In ‘‘Colloid Chemistry’’, J. Alexander, ed, Rheinhold Publishing corp, New York, 6, 630.
Tsuchida, H. M., Taniguchi, Y., Ito, H., Kawade, M., & Akasaka, K. (2001). Glucomannan separated from Agaricus blazei mushroom culture and antitumor agent containing as active ingredient. In J. Patent (Ed.) (Vol. 11–080206).
Unursaikhan, S., Xu, X., Zeng, F., & Zhang, L. (2006). Antitumor activities of O-sulfonated derivatives of (1-->3)-alpha-D-glucan from different Lentinus edodes. Bioscience, biotechnology, and biochemistry, 70, 38-46.
Usui, T. (1980). Isolation of highly purified fucoidan from Eisenia bicyclics and its anticoagulant and antitumor activities. Agricultural and biological chemistry, 44, 1965-1966.
Van de Perre, P. (2003). Transfer of antibody via mother's milk. Vaccine, 21, 3374-3376.
Voragen, A. G. J., Pilnik, W., Thibault, J. F., Axelos, M. A. V., & Renard, C. M. G. C. (1995). In: Food polysaccharides and their applications, Stephen A M. (Eds), Marcel Dekker, Inc., New York, 287-339.
Wagner, H., & Jordan, E. (1988). An immunologically active arabinogalactan from Viscum album ‘berries’. Phytochemistry, 27, 2511-2517.
Wang, B. G., Zhang, W.W., Duan, X. J., & Li, X. M. (2009a). In vitro antioxidative activities of extract and semi-purified fractions of the marine red alga, Rhodomela confervoides (Rhodomelaceae).Food Chemistry, 113, 1101–1105.
Wang, J., Liu, L., Zhang, Q., Zhang, Z., Qi, H., & Li, P. (2009b). Synthesized oversulphated, acetylated and benzoylated derivatives of fucoidan extracted from Laminaria japonica and their potential antioxidant activity in vitro. Food Chemistry, 114, 1285–1290.
Wang, J., Zhang, Q., Zhang, Z., & Li, Z. (2008). Antioxidant activity of sulphated polysaccharide fractions extracted from Laminaria japonica. International Journal of Biological Macromolecules, 42, 127–132.
Wang, J., Zhang, Q., Zhang, Z., Song, H., & Li, P. (2010). Potential antioxidant and anticoagulant capacity of low molecular weight fucoidan fractions extracted from Laminaria japonica. International Journal of Biological Macromolecules, 46, 6-12.
Wasser, S. P. (2002). Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Applied microbiology and biotechnology, 60, 258-274.
Wealth of India, (1988). A dictionary of Indian raw materials: Volume II: B (revised). New Delhi: Publications and Information Directorate. (350 + 90 pp. indexes).
Wealth of India, (1985). Algae, CSIR, New Delhi, Vol-IA, 138.
Whistler, R. (Ed.) (1993). Exudate gums. New York: Academic Press.
Witezak, Z. J. (1995). Carbohydrates as Drug and Potential Therapeutics. Current Medicinal Chemistry, 1, 392-405.
Witvrouw, M., & De Clercq, E. (1997). Sulfated polysaccharides extracted from sea algae as potential antiviral drugs. General Pharmacology, 29, 497–511.
Yamada, H., Ra, K. S., Kiyohara, H., Cyong, J. C., & Otsuka, Y. (1989a). Structural characterization of an anti-complementary pectic polysaccharide from the roots of Bupleurum falcatum L. Carbohydrate Research, 189, 209-226.

79
Yamada, H, Ra, K. S., Kiyohara, H., Cyong, J. C., Yang, H. C., & Otsuka,Y. (1989b). Characterization of anti-complementary neutral polysaccharides from the roots of Bupleurum falcatum. Phytochemistry, 27, 3163-3168.
Yamada, H., Kiyohara, H., Takemoto, N., Zhao, J. F., Kawamura, H., Komatsu, Y., Cyong, J. C., & Hosoya, E. (1992). Mitogenic and Complement Activating Activities of the Herbal Components of Juzen-Taiho-To1. Planta Medica, 58, 166-170.
Yamada, H., & Knutsen, S. H. (1996). Characterization of a biologically active pectin from Plantago major L. Carbohydrate Polymers, 30, 37-44.
Yamaguchi, T., Takamura, H., Matoba, T., & Terao, J. (1998). HPLC method for evaluation of the free radical scavenging activity of foods by using 1, 1,-diphenyl-2-picrylhydrazyl.Bioscience Biotechnology and Biochemistry, 62,1201–1204.
Yang, Y., Liu, D., Wu, J., Chen, Y., & Wang, S. (2011). In vitro antioxidant activities of sulfated polysaccharide fractions extracted from Corallina officinalis. International Journal of Biological Macromolecules, 49, 1031-1037.
Ye, H., Wang, K., Zhou, C., Liu, J., & Zeng, X. (2008). Purification, antitumor and antioxidant activities in vitro of polysaccharides from the brown seaweed Sargassum pallidum. Food Chemistry, 111, 428–432.
Yu, K. W., Kiyohara, H., Matsumoto, T., Yang, H. C., & Yamada, H. (1998). Intestinal Immune System Modulating Polysaccharides from Rhizomes of Atractylodes lancea. Planta Medica, 64, 714-719.
Yuan, X. P., Wang, J., Yao, H. Y., & Chen, F. (2005). Free radical scavenging capacity and inhibitory activity on rat erythrocyte hemolysis of feruloyl oligosaccharides from wheat bran insoluble dietary fiber. LWT – Food Science and Technology, 38, 877–883.
Zhang, L., Li, X., Xu, X., & Zeng, F. (2005). Correlation between antitumor activity, molecular weight, and conformation of lentinan. Carbohydrate Research, 340, 1515-1521.
Zhang, Y., Li, S., Wang, X., Zhang, L., & Cheung, P. C. K. (2011). Advances in lentinan: Isolation, structure, chain conformation and bioactivities. Food Hydrocolloids, 25, 196-206.
Zhao, J. F., Kiyohara, H., Matsumoto, T., & Yamada, H. (1994). Anti-complementary acidic polysaccharides from roots of Lithospermum euchromum. Phytochemistry, 34, 719-724.
Zhao, X., Xue, C., Cai, Y., Wang, D., & Fang, Y. (2005). Study of antioxidant activities of fucoidan from Laminaria japonica. High Technology Letters, 11, 91-94.
Zhou, G., Sun, Y., Xin, H., Zhang, Y., Li, Z., & Xu, Z. (2004). In vivo antitumor and immunomodulation activities of different molecular weight lambda-carrageenans from Chondrus ocellatus. Pharmacological research, 50, 47-53.
Zhou, S., & Gao Y. (2002). The immunomodulating effects of Ganoderma lucidum (Curt.:Fr.) P.Karst (LingZhi, Reishi Mushroom) (Aphylloromycetidae). International journal of medicinal mushrooms,4,1–11.
Zhu, H., Di, H., Zhang, Y., Zhang, J., & Chen, D. (2009). A protein-bound polysaccharide from the stem bark of Eucommia ulmoides and its anti-complementary effect. Carbohydrate Research, 344, 1319-1324.
Zhuang, C. Itoh, H., Mizuno, T., & Ito, H. (1995). Antitumor Active Fucoidan from the Brown Seaweed, Umitoranoo (Sargassum thunbergii). Bioscience, biotechnology, and biochemistry, 59, 563-567.
Zhuang, C., Mizuno, T., Ito, H., Shimura, K., Sumiya,T., Kawade, M., & Inamori, Y. (1994). Fractionation and Antitumor Activity of Polysaccharides from Grifola frondosa Mycelium.Bioscience, biotechnology, and biochemistry, 58, 185-188.
Zjawiony, J. K. (2004). Biologically active compounds from Aphyllophorales (polypore) fungi. Journal of natural products, 67, 300-310.

80
The novel findings presented in this thesis are:
1. Three families of water soluble polysaccharides, namely, sulfated fucan, alginic acid and
glucan can be easily isolated from the marine alga Turbinaria conoides.
2. A 50 kDa sulfated fucan containing inter alia, α-(1→3)-linked fucopyranosyl residues with a
sulfate group at the position 4, has been identified.
3. The alginic acid contains equimolar proportions of guluronic and mannuronic acid residues.
The guluronic acid content (G %) and G–G diad frequency (GG %) of this polymer are 46.2 and
42.1, respectively.
4. A 5 kDa linear glucan containing β-(1→3)-linked glucopyranosyl residue has also been
isolated.
5. These polymers T. conoides from demonstrated dose dependent antioxidative activity. The
fucoidan showed the highest antioxidative activity followed by the alginate and glucan.
6. The decoction of Adhatoda vasica contains a highly branched arabinogalactan.
7. This branched arabinogalactan containing 1,3- and 1,3,6-linked galactopyranosyl & 1,5- and
1,3,5- linked arabinofuranosyl residue, possesses potent in vivo antitussive activity.
8. Peroral administration of this arabinogalactan (50 mg kg−1 body weight) from A. vasica
inhibited the number of coughs induced by citric acid in guinea pigs and slightly decreased the
values of specific airway resistance.
9. Thus, traditional aqueous extraction methods provide polysaccharides, which stimulate an
intense biological response. This could represent an interesting approach in phytotherapeutic
treatments.
SUMMARY

81
LIST OF PUBLICATIONS
Nabanita Chattopadhyay, Tuhin Ghosh, Sharmistha Sinha, Kausik Chattopadhyay, Paramita
Karmakar, Bimalendu Ray (2010). Polysaccharides from Turbinaria conoides: Structural
features and antioxidant capacity. Food Chemistry, 118, 823–829.
Nabanita Chattopadhyay, Gabriella Nosal’ova, Sudipta Saha, Shruti S. Bandyopadhyay, Dana
Fleˇskova, Bimalendu Ray (2011). Structural features and antitussive activity of water extracted
polysaccharide from Adhatoda vasica. Carbohydrate Polymer, 83, 1970-1974.


















