
Physiology and Genetics of Acidithiobacillus species
73
1 Physiology and Genetics of Acidithiobacillus species: Applications for Biomining by Olena I. Rzhepishevska ISBN 978-91-7264-509-7 Umeå University Umeå 2008
Transcript of Physiology and Genetics of Acidithiobacillus species

1
Physiology and Genetics of Acidithiobacillus species:
Applications for Biomining
by
Olena I. Rzhepishevska
ISBN 978-91-7264-509-7
Umeå University
Umeå 2008

2

3
TABLE OF CONTENTS
ABSTRACT 5
ABBREVIATIONS AND CHEMICAL COMPOUNDS 6
PAPERS IN THIS THESIS 7
1. INTRODUCTION 9
1.1 General characteristics of the Acidithiobacillus genus 9
1.1.1 Classification 9
1.1.2 Natural habitats and growth requirements 9
1.1.3 Iron and RISCs in natural and mining environments 10
1.1.4 The sulphur cycle and acidithiobacilli 11
1.1.5 Genetic manipulations in Acidithiobacillus spp. 13
1.1.6 Importance in industry and other applications 14
1.2 Acidithiobacillus spp. RISC and iron oxidation 15
1.2.1 RISC oxidation by Acidithiobacillus spp. 15
1.2.2 A. ferrooxidans iron oxidation and regulation 23
1.3 Mineral sulphide oxidation 26
1.4 Acid mine and rock drainage 30
1.5 Biomining 36
1.5.1 Biomining as an industrial process 36
1.5.2 Acidophilic microorganisms in industrial bioleaching 38
2. AIMS OF THE STUDY 41
3. RESULTS AND DISCUSSION 43
4. CONCLUSIONS 51
5. ACKNOWLEDGMENTS 53
6. REFERENCES 55
7. PAPERS 75

4

5
ABSTRACT
Bacteria from the genus Acidithiobacillus are often associated with biomining
and acid mine drainage. Biomining utilises acidophilic, sulphur and iron
oxidising microorganisms for recovery of metals from sulphidic low grade ores
and concentrates. Acid mine drainage results in acidification and contamination
with metals of soil and water emanating from the dissolution of metal sulphides
from deposits and mine waste storage. Acidophilic microorganisms play a
central role in these processes by catalysing aerobic oxidation of sulphides.
Acceleration of mineral solubilisation is a positive aspect in biomining whereas,
in acid mine drainage it is undesirable and accordingly, microbial iron and
sulphur oxidation is promoted in the first case and measures are taken to inhibit
it in the second case. In this thesis, several approaches were taken in order to
understand and increase oxidation efficiency in biomining and to gain an insight
into the biochemical reactions taking place in these environments. A laboratory
scale bioreactor was designed and tested allowing simulation of bioleaching in
heaps of mine tailings at different aeration, irrigation and particle size conditions
(Paper I). A new psychrotolerant strain of Acidithiobacillus ferrooxidans was
characterised that has an application in boreal heap bioleaching. Iron, reduced
inorganic sulphur compound oxidation and bioleaching of various ores by this
strain was studied as well as gene expression during oxidation of tetrathionate
and/or ferrous iron (Papers III & IV). Expression and regulation of a
tetrathionate hydrolase from Acidithiobacillus caldus, a key enzyme in reduced
inorganic sulphur compound metabolism of this bacterium was investigated and
the presence of this enzyme in a bioleaching mixed culture was shown. The gene
cluster that harbours the gene coding for tetrathionate hydrolase (tetH) was
described for the first time (Paper II).

6
ABBREVIATIONS AND CHEMICAL COMPOUNDS
AMD - acid mine drainage
ARD - acid rock drainage
DGGE - denaturing gradient gel electrophoresis
EPS- extracellular polymeric substance
FISH - fluorescent in situ hybridisation
GSH - glutathione
GSSG - GSH disulphide
GSSH - glutathione persulphide
GSSO3- - glutathione S- sulphonate
HiPIP - high potential iron-sulphur protein
H2S - sulphide
PQQ - pyrrolo-quinoline quinone
RISCs - reduced inorganic sulphur compounds
SFORase - sulphide:Fe(III) oxidoreductase
SORa - sulphur oxygenase reductase
SQR - sulphide quinone oxidoreductase
StOR - sulphite:cytochrome c oxidoreductase
Sn2- - polysulphides
S0 - elemental sulphur
S2O32- - thiosulphate
S4O62- - tetrathionate
S5O62- - pentathionate
SO32- - sulphite
SRB - sulphate reducing bacteria
TQO - thiosulphate:quinone oxidoreductase
TQR - thiosulphate quinone oxidoreductase

7
PAPERS IN THIS THESIS
I
Olena I. Rzhepishevska, E. Börje Lindström, Olli H. Tuovinen & Mark Dopson
(2005) Bioleaching of sulfidic tailing samples with a novel, vacuum-positive
pressure driven bioreactor. Biotechnol Bioengin 92(5):559-567
II
Olena I. Rzhepishevska, Jorge Valdes, Liucija Marcinkeviciene, Camelia Algora
Gallardo, Rolandas Meskys, Violaine Bonnefoy, David S. Holmes & Mark
Dopson (2007) Regulation of a novel Acidithiobacillus caldus gene cluster
involved in metabolism of reduced inorganic sulfur compounds. Appl
Environ Microbiol 73(22):7367-7372
III
Daniel Kupka, Olena I. Rzhepishevska, Mark Dopson, E. Börje Lindström, Olia
V. Karnachuk & Olli H. Tuovinen (2007) Bacterial oxidation of ferrous iron
at low temperatures. Biotechnol Bioengin 97(6):1470-1478.
IV
Olena I. Rzhepishevska & Mark Dopson (2008) Sulphur Accumulation
During Low Temperature Bioleaching (Manuscript in preparation)
Papers in this thesis have been reprinted with permission from Wiley (papers I
& III) and ASM (paper II)

8

9
1. INTRODUCTION
1.1 General characteristics of the Acidithiobacillus genus
1.1.1 Classification
The Acidithiobacillus genus belongs to the γ-proteobacteria and comprises four
species: Acidithiobacillus albertensis, Acidithiobacillus caldus,
Acidithiobacillus ferrooxidans and Acidithiobacillus thiooxidans (Kelly &
Wood, 2000). The genus was formed when the former Thiobacillus genus was
split into the genera Acidithiobacillus, Halothiobacillus and Thermithiobacillus.
The members of the genus are acidophilic Gram-negative rods, motile by one or
more flagella and comprise both mesophiles and moderate thermophiles. All
four species are autotrophs capable of growth utilising inorganic compounds
such as reduced inorganic sulphur compounds (RISCs) as sole energy substrate,
whereas A. ferrooxidans is the only representative of the genus that can also
oxidise Fe2+. A. ferrooxidans is represented by a number of strains with
pronounced genetic variation (Bergamo et al., 2004; Karavaiko et al., 2003;
Kelly & Wood, 2000; Paulino et al., 2001) which are predicted by Kelly and
Wood (2000) to be reclassified into separate species. A. albertensis was thought
to be lost from culture and its 16S rRNA sequence was previously unavailable,
but in 2002 the 16S rRNA gene sequence of DSM strain 14366 was published
(accession number AJ459804).
1.1.2 Natural habitats and growth requirements
Acidithiobacilli use various RISCs or Fe2+ as electron donor and therefore, are
often found in metal sulphide deposits all over the world (Karavaiko et al.,
2003), fresh waters associated with sulphide deposits (Gonzalez-Toril et al.,
2003) and seawater (Kamimura et al., 2003). A flexible chemolithoautotrophic
metabolism allows A. ferrooxidans and A. caldus to grow on hydrogen as the

10
electron donor which could potentially broaden their ecological niche (Drobner
et al., 1990; Hallberg & Lindström, 1994). The final electron acceptor is usually
oxygen (Kelly & Wood, 2000) however, Fe3+ and S0 were reported to be
alternative electron acceptors for A. ferrooxidans under anaerobic conditions
(Ohmura et al., 2002; Pronk et al., 1991a; Sugio et al., 1992a). Presence of
Calvin cycle enzymes (Appia-Ayme et al., 2006; Kusano et al., 1991) and hence
the ability to fix CO2 makes acidithiobacilli independent of an organic carbon
source and allows them to thrive in mine waste ecosystems where the organic
carbon concentration is extremely low (Dold et al., 2005). Despite the reports
that organic compounds are toxic to A. ferrooxidans (Tuttle & Dugan, 1976;
Tuttle et al., 1977), A. caldus was reported to reach higher growth yeilds with
addition of yeast extract or glucose and A. ferrooxidans was able to utilise
glucose and formate (Hallberg & Lindström, 1994; Pronk et al., 1991b; Tabita
& Lundgren, 1971). All members of the genus are acidophiles with optimum pH
from 2 to 4 but the upper limit for A. thiooxidans reaches 5.5. All of the
acidithiobacilli are mesophiles with the exception of A. caldus which is a
moderate thermophile with an optimum at 45°C (Kelly & Wood, 2000). When
cultivated in laboratory conditions, acidithiobacilli generally grow much slower
and give much lower yields compared to heterotrophs such as Escherichia coli
(Eccleston & Kelly, 1978; Kamimura et al., 2003; Leduc et al., 1993).
1.1.3 Iron and RISCs in natural and mining environments
Sulphur is one of the most abundant elements in minerals after oxygen and
silicon; together with all its known compounds it constitutes 0.03 % of the
Earth’s crust. Native sulphur (S0) is mainly found in volcanic, oil-bearing or
sedimentary deposits. The biggest sedimentary deposits are located in Texas and
Louisiana (USA) and Sicily (Italy). In the form of sulphate, sulphur is abundant
in oceans and in the soil. Sulphur reacts with all metals except gold and
platinum forming sulphides. The most known and exploited ores that contain

11
sulphides are chalcocite (Cu2S), pyrite (FeS2), chalcopyrite (CuFeS2), galena
(PbS), cinnabar (HgS), sphalerite (ZnS), argenite (Ag2S) and arsenopyrite
(FeAsS) (Weller, 1960; In Encyclopaedia Britanica;
http://search.eb.com/eb/article-9070248). Pyrite is the most widespread metal
sulphide in the world and is often used for sulphur production and rich pyrite
deposits are found in Spain, USA, Canada, Norway, Russia and Portugal.
Galena is often found in association with sphalerite and argenite in USA,
Australia and England, while chalcopyrite is present in almost every sulphide
deposit.
The estimated amount of iron in the Earth’s crust is 5 %; it occurs in
hundreds of different compounds but the most common source of Fe2+ for iron
oxidising acidophiles such as A. ferrooxidans are iron sulphides (Mottana, 1983;
Weller, 1960). The interests of sulphur/iron oxidising microorganisms and
humans collide at sulphide deposits as mining activities exposes sulphide
sources buried under the ground and creates conditions (i.e. exposure to air and
water) for the existence of acidithiobacilli and other acidophilic microorganisms.
1.1.4 The sulphur cycle and acidithiobacilli
Sulphur is one of the key elements in proteins that are present in every living
cell and at the same time sulphur compounds can be used as electron donors or
electron acceptors by various microorganisms. Such a tight dependence of all
forms of life on sulphur creates its turnover in nature, termed the sulphur cycle.
In this process, sulphur atoms are present at different oxidation states and are
incorporated into amino acids of proteins as well as mineralised to inorganic
compounds. The main reactions of the sulphur cycle are assimilatory sulphate
reduction, mineralisation (desulphurisation), dissimilatory oxidation of hydrogen
sulphide and sulphur, and dissimilatory sulphate and sulphur reduction.
A number of groups of microorganisms are involved in dissimilatory
oxidation of RISCs. Among those are the acidithiobacilli, green and purple

12
phototrophic bacteria (e.g. Clorobium and Allochromatium), some thermophilic
Archaea such as Acidianus and Sulfolobus, the facultatively lithoautotrphic
neutrophile Parracoccus pantotrophus and bacteria that live in association with
oceanic sulphurous hot springs from the genera Beggiatoa and Thioploca
(Friedrich et al., 2005; Madigan & Martinko, 2006). Acidithiobacillus spp. are
active in several steps of the sulphur cycle (Fig. 1) but their most important role
in sulphur mobilisation is to catalyse metal sulphide oxidation at low pH (see
section 1.2 for details), thereby releasing sulphate into solution (Schrenk et al.,
1998).
Fig. 1 The sulphur cycle - modified from Robertson & Kuenen (1992). Steps that include specific activities by acidithiobacilli are underlined. Oxidation steps are solid lines and dashed lines are reductive.
Sn2-
S4O62-, S2O3
2-
S0
H2SS2- -Metalsulphides
R-SH
S0
SO32-
Organic compounds
Assimilatory sulphatereduction Mineralization
Dissimilatorysulphur reduction
Dissimilatory sulphate reduction
Biological aerobic andanaerobic oxidation;
spontaneous oxidation
Biological aerobic andanaerobic oxidation
Biological aerobic oxidation
SO42-
Sn2-
S4O62-, S2O3
2-
S0
H2SS2- -Metalsulphides
R-SH
S0
SO32-
Organic compounds
Assimilatory sulphatereduction Mineralization
Dissimilatorysulphur reduction
Dissimilatory sulphate reduction
Biological aerobic andanaerobic oxidation;
spontaneous oxidation
Biological aerobic andanaerobic oxidation
Biological aerobic oxidation
Sn2-
S4O62-, S2O3
2-
S0
H2SS2- -Metalsulphides
R-SH
S0
SO32-
Organic compounds
Assimilatory sulphatereduction Mineralization
Dissimilatorysulphur reduction
Dissimilatory sulphate reduction
Biological aerobic andanaerobic oxidation;
spontaneous oxidation
Biological aerobic andanaerobic oxidation
Biological aerobic oxidation
SO42-SO42-

13
Assimilatory sulphate reduction occurs in all animals, plants and all
microorganisms. Desulphurisation is also a general process which involves
decomposition of proteins through decay and excretion. Dissimilatory sulphate
and sulphur reduction is carried out by bacteria generally termed sulphate
reducers (e.g. the genera Desulfovibrio and Desulfotomaculum) and
dissimilatory sulphur-reducing bacteria (Desulfuromonas). Sulphate reducers
use sulphate as an anaerobic terminal electron acceptor, while dissimilatory
sulphur-reducing bacteria couple anaerobic oxidation of organic compounds to
reduction of S0 to hydrogen sulphide. Certain facultatively aerobic heterotrophs
like Pseudomonas spp. and Salmonella spp. are able to reduce sulphur and other
sulphur compounds (Madigan & Martinko, 2006; Robertson & Kuenen, 1992).
1.1.5 Genetic manipulations in Acidithiobacillus spp.
Genetic manipulations such as gene mutagenesis and cloning, mutation
complementation and protein expression provide excellent approaches to study
gene and protein function in different organisms. Unfortunately, many of these
techniques are inefficient in Acidithiobacillus spp. mainly due to that the outer
membrane and periplasmic space of these bacteria are adapted to acidophilic
conditions, while most of the methods generally used in molecular biology work
optimally at neutral pH (Holmes & Bonnefoy, 2007). One of the main problems
is the absence of an efficient method to import DNA into acidithiobacilli cells.
Though plasmid transfer through conjugation with E. coli was achieved for all
species except A. albertensis, the frequency of transconjugates was only 10-5-10-
7 transconjugates per recipient (Jin et al., 1992; Liu et al., 2007; Peng et al.,
1994; van Zyl et al., 2003). Electroporation was also possible in A. ferrooxidans
but it was apparently only successful for certain strains (Kusano et al., 1992).
Heterologous protein expression in E. coli or other well studied
microorganisms is one way to confirm its function and an efficient option to
produce sufficient protein for further studies. Indeed, some A. ferrooxidans and

14
A. caldus cytoplasmic and inner membrane proteins were successfully expressed
in E. coli (de Groot et al., 2003; Zeng et al., 2007) but outer membrane and
periplasmic proteins often fail to fold correctly at neutral pH (Bengrine et al.,
1998; Bruscella et al., 2005). Expression of heterogeneous phosphofructokinase
in A. thiooxidans was also reported and resulted in glucose consumption by this
chemolithothroph (Tian et al., 2003).
A few mutants achieved both by direct and random mutagenesis have been
reported for A. ferrooxidans. One of them was due to insertion of ISAfe1 (an
insertion sequence element) and the other was constructed using a suicide
plasmid with a selectable marker (Holmes et al., 2001; Liu et al., 2000).
Recently, a successful mutant construction protocol involving a suicide plasmid
was developed for A. caldus (D. Rawlings personal communication) however;
this protocol is un-published and not available for general use. As a result,
molecular genetic studies in acidithiobacilli is a challenging and time consuming
task specially considering their long generation times, low yields in liquid media
and poor growth on solid media.
1.1.6 Importance in industry and other applications
The importance of acidithiobacilli in industry developed as a function of their
ability to catalyse solubilisation of metal sulphides and to produce sulphuric acid
(see chapter 1.4 for details). In some cases their presence is favourable for
industrial processes and sometimes it is undesirable. Acidithiobacilli are
employed for efficient extraction of many valuable metals from sulphidic
minerals, an approach generally termed ‘biomining’ (Rawlings, 2002; Rawlings,
2005). On the other hand, their activity often causes an environmentally
hazardous process called acid mine drainage (AMD) and acid rock drainage
(ARD; for more information see chapter 1.3) (Canovas et al., 2007; Lefebvre et
al., 2001a; Lefebvre et al., 2001b; Olias et al., 2004; Vigneault et al., 2001).
Moreover, A. thiooxidans and A. ferrooxidans speed up concrete corrosion

15
processes (Maeda et al., 1999; Sand, 1987) and A. thiooxidans is reported to
dissolve human urinary stones in in vivo experiments (Nishio et al., 2003).
Lately, promising experiments were carried out showing the possibility of A.
ferrooxidans usage in a microbial fuel cell running on Fe2+ (Ter Heijne et al.,
2007).
1.2 Acidithiobacillus spp. RISC and iron oxidation
1.2.1 RISC oxidation by Acidithiobacillus spp.
Chemolithotrophic RISC oxidation occurs in various groups of microorganisms
and several types of enzyme systems are known that can carry out this process.
The well studied Sox complex from α-proteobacterium P. pantotrophus is
important for oxidation of thiosulphate to sulphate and also occurs in many other
bacterial species (Friedrich et al., 2005). RISC oxidation by Archaea has been
studied in Acidianus ambivalens where sulphur oxygenase reductase (SORa)
and thiosulphate:quinone oxidoreductase (TQO) have been discovered. SORa
catalyses the disproportionation of sulphur to sulphite, thiosulphate and
hydrogen sulphide and TQO couples the reduction of caldariella quinone to
thiosulphate oxidation with tetrathionate as a product (Muller et al., 2004; Urich
et al., 2004). Interestingly, neither sox nor sor genes have been found in the A.
ferrooxidansT genome (ATCC 23270), but instead acidithiobacilli have their
own unique enzymes dedicated to RISC oxidation. Though many of these
enzymes have been purified and characterised, the whole picture is far from
being clear. Another unsolved question is how S0 is transported into the
periplasm of acidithiobacilli. It is proposed that it is mobilised by thiol groups of
putative outer-membrane proteins and transported to the periplasm in form of
persulphide sulphur (Rohwerder & Sand, 2003; Silver & Lundgren, 1968b).
Acidithiobacillus thiooxidans. A. thiooxidans was the first bacterium of the
genus to be discovered (Kelly & Wood, 2000; Waksman, 1922). It was shown

16
not to oxidise Fe2+ but was able to metabolise various RISCs such as S0,
thiosulphate (S2O32-), tetrathionate (S4O6
2-) and sulphite (SO32-) with different pH
optimums for certain compounds (Masau et al., 2001; Suzuki, 1965a; Suzuki,
1965b; Suzuki et al., 1992). Thus, the pH optimum for sulphur oxidation is 2.3
but pH 5 for tetrathionate, thiosulphate and sulphite (Masau et al., 2001). The
enzyme responsible for oxidation of S0 was purified in 1965 (Suzuki, 1965b)
and participation of glutathione (GSH) in the reaction was reported even earlier
(Suzuki & Werkman, 1959). Rohwerder and Sand (2003) reported that GSH
activates sulphur through a non-enzymatic reaction forming glutathione
persulphide (GSSH), which in its turn yields sulphite in an enzymatic reaction
catalysed by sulphur dioxygenase. Sulphite was further oxidised in a non-
enzymatic reaction to sulphate, thiosulphate and glutathione S- sulphonate
(GSSO3-). Oxidation of H2S occurred only in the presence of GSH disulphide
(GSSG).
Oxidation of thiosulphate by A. thiooxidans follows two different pathways
depending on the pH (Masau et al., 2001). At pH 5, the intermediate products
are sulphite and sulphur and at pH 2.3 they are tetrathionate and sulphur. In both
cases sulphur and sulphite are finally oxidised to sulphate. Thiosulphate
dehydrogenase containing cytochrome c was purified from A. thiooxidans JCM
7814 (Nakamura et al., 2001) and tetrathionate decomposing enzyme was
purified from A. thiooxidans ON 107 (Tano et al., 1996). The main product of
tetrathionate metabolism in A. thiooxidans ON 107 was thiosulphate. Sulphite
oxidation in A. thiooxidans JCM 7814 was studied by Nakamura et al.
(Nakamura et al., 1992; Nakamura et al., 1995) and proven to be catalysed by an
adenosine 5-monophosphate independent membrane bound sulphite:cytochrome
c oxidoreductase (sulphite dehydrogenase). Sugio et al. (2006) purified an aa3-
type ubiquinol oxidase and demonstrated the existence of sulphite:ubiquinone
oxidoreductase activity in A. thiooxidans BN1-3. These findings allowed them

17
to propose aa3- type ubiquinol oxidase as a terminal oxidase in sulphite and
consequently the A. thiooxidans sulphur oxidation pathway.
Acidithiobacillus caldus. Only three studies have been dedicated to sulphur
metabolism in A. caldus (Bugaytsova & Lindström, 2004; Dopson et al., 2002;
Hallberg et al., 1996). It was shown that A. caldus could metabolise S2-, S0,
S2O32-, S4O6
2- and SO32- and an increase in phosphorylation potential was
observed during oxidation of all these compounds (Dopson et al., 2002).
Fig. 2 Oxidation of thiosulphate (■) to tetrathionate (●) by A. caldus KU in the
absence (A) and in the presence (B) of 200 μM 2,4-dinitrophenol. Re-printed from Hallberg et al. (1996).
Con
cent
ratio
n (m
M)
0 4 8 12 16 20 24minutes
Con
cent
ratio
n (m
M)
0 4 8 12 16 20 24minutes

18
No substrate level phosphorylation was detected and it was concluded that all
ATP is synthesised by membrane bound ATPase driven by a proton gradient.
Thiosulphate was oxidised to tetrathionate by A. caldus cells and in presence of
2,4-dinitrophenol tetrathionate was accumulated (Fig. 2). Inhibition of oxygen
consumption by cyanide had a biphasic character that suggested the presence of
more than one terminal oxidase. It was proposed that thiosulphate is oxidised to
tetrathionate in the periplasm which agreed with the localisation of thiosulfate
oxidising enzyme in other acidithiobacilli. The products of tetrathionate
hydrolysis were thiosulphate, pentathionate and sulphate (Bugaytsova &
Lindström, 2004). Based on inhibition by various uncouplers, it was suggested
that S2- and S0 oxidation yields sulphite and enzymes for these reactions were
located in the cytoplasm (Hallberg et al., 1996) which disagrees with proposed
localisation of sulphur dioxygenase in other acidithiobacilli (Rohwerder & Sand,
2003). Based on subsequent data (Bugaytsova & Lindström, 2004) it is probable
that the inhibition of S2- and S0 oxidation by uncouplers is due to their uptake to
the periplasm rather than the cytoplasm and data in the Hallberg et al. (1996)
study was misinterpreted.
Acidithiobacillus albertensis. A. albertensis is the least studied species of
all acidithiobacilli. DSM 14366 and BY-05 are the only two strains known and
both were demonstrated to metabolise S0, S2O32-, S4O6
2- and ZnS (Kelly, 1989;
Xia et al., 2007). The details of S0 oxidation in this bacterium are unknown. A.
albertenis was shown to colonise elemental sulphur particles by micro colonies
and it was suggested that glycocalyx plays an important role in the attachment
(Bryant et al., 1984).
Acidithiobacillus ferrooxidans. A. ferrooxidans is the best studied
representative of all acidithiobacilli. A. ferrooxidans can oxidise RISCs both
aerobically and anaerobically and all known aerobic pathways of RISC
oxidation were summarised (Fig. 3) by Holmes and Bonnefoy (2007). The S0
oxidising protein was purified by Silver and Lundgren in 1968 but the actual
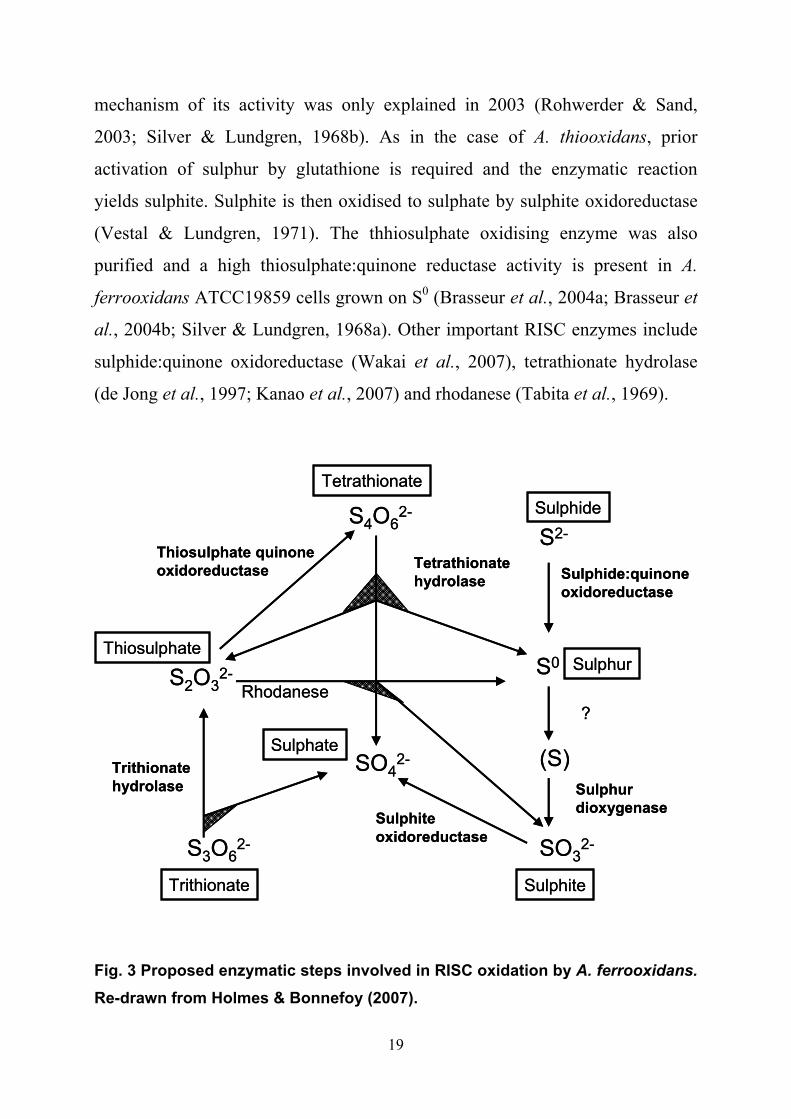
19
mechanism of its activity was only explained in 2003 (Rohwerder & Sand,
2003; Silver & Lundgren, 1968b). As in the case of A. thiooxidans, prior
activation of sulphur by glutathione is required and the enzymatic reaction
yields sulphite. Sulphite is then oxidised to sulphate by sulphite oxidoreductase
(Vestal & Lundgren, 1971). The thhiosulphate oxidising enzyme was also
purified and a high thiosulphate:quinone reductase activity is present in A.
ferrooxidans ATCC19859 cells grown on S0 (Brasseur et al., 2004a; Brasseur et
al., 2004b; Silver & Lundgren, 1968a). Other important RISC enzymes include
sulphide:quinone oxidoreductase (Wakai et al., 2007), tetrathionate hydrolase
(de Jong et al., 1997; Kanao et al., 2007) and rhodanese (Tabita et al., 1969).
Fig. 3 Proposed enzymatic steps involved in RISC oxidation by A. ferrooxidans.
Re-drawn from Holmes & Bonnefoy (2007).
TetrathionateSulphide
Sulphate
Trithionate Sulphite
SulphurThiosulphate
S4O62-
S2O32-
SO42-
SO32-S3O6
2-
S0
S2-
(S)
Sulphide:quinoneoxidoreductase
?
Sulphurdioxygenase
Tetrathionatehydrolase
Sulphiteoxidoreductase
Trithionatehydrolase
Rhodanese
Thiosulphate quinoneoxidoreductase
TetrathionateSulphide
Sulphate
Trithionate Sulphite
SulphurThiosulphate
S4O62-
S2O32-
SO42-
SO32-S3O6
2-
S0
S2-
(S)
Sulphide:quinoneoxidoreductase
?
Sulphurdioxygenase
Tetrathionatehydrolase
Sulphiteoxidoreductase
Trithionatehydrolase
Rhodanese
Thiosulphate quinoneoxidoreductase

20
A. ferrooxidans has a number of terminal oxidases and electron flow from
various substrates to oxygen is quite complicated. The most recent findings
suggest that electrons arising from growth on RISCs enter the respiratory chain
through sulphide quinone oxidoreductase (SQR), TQO or sulphite
oxidoreductase (StOR) and are transferred to bd- or bo3-type oxidases. Those
electrons which are directed to the bo3-type oxidase pass through a bc1-complex
(Fig. 4).
bd quinol oxidase
SQR TQR
CycA2( c4 )
bo3 quinoloxidaseUQ
Hip
bc1 complex(PetABC2)
S2O32- S4O6
2-H2S S0
SO SOR?
GSSG SO32- SO4
2-
O2 H2O O2 H2O
Operon petII Operon cyo Operon cyd Operon doxII sqr3
cycA2,petA2,petC, sdrA2, petB2, hip
cyoB, cyoC, cyoA,cyoD
cydB, cydA doxDA, rod
bd quinol oxidase
SQR TQR
CycA2( c4 )
bo3 quinoloxidaseUQ
HipHip
bc1 complex(PetABC2)
S2O32- S4O6
2-H2S S0
SO SOR??
GSSG SO32- SO4
2-
O2 H2O O2 H2O
Operon petII Operon cyo Operon cyd Operon doxII sqr3
cycA2,petA2,petC, sdrA2, petB2, hip
cyoB, cyoC, cyoA,cyoD
cydB, cydA doxDA, rod
Fig. 4 Model proposed for RISC energetic metabolism of A. ferrooxidans. The transcriptional units and the corresponding redox proteins located in the inner membrane are presented in the same color. Operons cyo, cyd and doxII are up regulated on S0; operons petII and sqr3 are expressed to the same or higher level on S0 comparing to Fe2+. TQR: thiosulphate quinol reductase; SQR: sulfide quinone reductase; Hip: High Potential Iron-sulphur Protein. Re-drawn from Quatrini et al. (2006).

21
Electrons resulting from sulphide and thiosulphate oxidation join the
respiratory chain at the level of the quinol pool and those coming from sulphite
oxidation enter at the level of cytochrome c4 or high potential iron-sulphur
protein (HiPIP) protein (Fig. 4) (Holmes & Bonnefoy, 2007; Quatrini et al.,
2006). Unusually, the electron transport chain in A. ferrooxidans uses two
differentially expressed bc1 complexes (Appia-Ayme et al., 1999; Brasseur et
al., 2002; Bruscella et al., 2007). The first is coded by the petI operon and is
expressed during growth on Fe2+. The other is coded by the petII operon and
dominates in S0 grown A. ferrooxidans. It was shown that petII coded bc1
complex supports downhill electron transport to bo3-type oxidase, but the bc1
coded by petI works in the opposite direction and supplies electrons for NAD(H)
reduction (for more details, see chapter 1.2.2). This theory was built using data
obtained by microarray analysis of differential gene expression in A.
ferrooxidans grown on Fe2+ and S0 and verified by quantitative PCR and
supported by measurements of enzyme activity.
Higher transcription levels were found for genes from petII, cyo, cyd and
doxII operons from S0 grown A. ferrooxidansT ATCC 23270 compared to cells
grown on Fe2+ (Brasseur et al., 2004b; Bruscella et al., 2007; Quatrini et al.,
2006). Surprisingly, similar experiments in A. ferrooxidans strain NASF-1
showed the same level of cydA gene expression both in S0 and iron grown cells
(Wakai et al., 2004). Moreover, coxB which was down regulated in S0 grown A.
ferrooxidans ATCC 23270 (Quatrini et al., 2006) was equally expressed in A.
ferrooxidans NASF-1. The same authors reported that a thiosulphate:ubiquinone
oxidoreductase activity was not detected in NASF-1 strain which is
contradictory to findings in A. ferrooxidans BRGM strain (Brasseur et al., 2002;
Wakai et al., 2004). These contradictions might indicate that there could be
other pathways of electron transport during aerobic RISC oxidation in different
strains within the A. ferrooxidans species.

22
A. ferrooxidansT was shown to grow in anaerobic conditions using S0 as
electron donor and Fe3+ as electron acceptor (Pronk et al., 1992). Strain 21834
was capable of glycine uptake under the same conditions which was seen as a
proof of energy transduction during Fe3+ respiration because glycine was not
taken up by starving cells (Pronk et al., 1991a). Interestingly, A. ferrooxidans
strain JCM 7811 expressed a specific cytochrome when grown on sulphur and
respiring Fe3+. The cytochrome had a molecular mass of 28-kD, was stable at
acidic pH and able to reduce Fe3+ (Ohmura et al., 2002).
The observation that S0 reduction coupled to Fe3+ oxidation takes place
even in the presence of oxygen gave rise to a new theory proposing an
alternative mechanism for S0 oxidation in A. ferrooxidans strain AP19-3 under
aerobic conditions. A soluble sulphur:Fe(III) oxidoreductase and a membrane
bound sulfite:Fe(III) oxidoreductase were purified from strain AP19-3 (Sugio et
al., 1992a; Sugio et al., 1992b). The first enzyme requires GSH and oxidises S0
to sulphite and the second one oxidises sulphite to sulphate. It is suggested that
Fe2+ resulting from hydrogen sulphide:Fe(III) oxidoreductase (SFORase) and
sulphite:Fe(III) oxidoreductase reactions is further oxidised via iron oxidase
(Sugio et al., 1985; Sugio et al., 1988b). Sugio et al. (1989) proposed hydrogen
sulphide to be the actual substrate for sulphur:Fe(III) oxidoreductase using the
argument that it was oxidising hydrogen sulphide in the presence of 8mM GSH.
However, contrary to this, Rohwerder and Sand (2003) argue that GSH at such a
high concentration forms GSSG which reacts with H2S producing GSSH, which
in its turn is the real substrate for the oxidoreductase. Nevertheless, the enzyme
is often referred to as a SFORase. SFORase oxidises S0 using not only Fe3+ but
also Mo6+ and Cu2+ as electron acceptors (Sugio et al., 1988a; Sugio et al.,
1990b). SFORase and iron oxidase activities were high even in the type strain
grown for many generations on S0 and SFORase activities comparable to that of
strain AP19-3 were found in eight different strains of A. ferrooxidans and five
strains of Leptospirillum ferrooxidans (Sugio et al., 1992b; Sugio et al., 2007).

23
Both these facts were interpreted by the authors as evidence favouring a wide
distribution of the alternative pathway for S0 oxidation in iron oxidising
microorganisms.
Sulphur oxidation is strongly inhibited by Fe2+ in strain AP19-3 due to
inhibition of sulphite:Fe(III) oxidoreductase by this ion. As a result, sulphite
accumulates in the cells and damages SFORase and iron oxidase which halts cell
growth (Sugio et al., 1990a). Such sensitivity of strain AP19-3 to sulphite
suggests that this strain might lack sulphite oxidoreductase.
1.2.2 A. ferrooxidans iron oxidation and regulation
A. ferrooxidans is the only species of acidithiobacilli able to oxidise Fe2+ and
consequently these bacteria have both RISCs and Fe2+ for energy metabolism at
their disposal. Identification of the primary protein that oxidises Fe2+ to Fe3+ in
A. ferrooxidans has a long history. Among others, high potential iron-sulphur
protein (HiPIP) from Fe-1 and BMGR strains, the 26 kDa cytochrome c4 (also
known as Cyc1 or CYC41) and outer membrane cytochrome Cyc2 have all been
suggested to serve as the iron oxidase (Appia-Ayme et al., 1999; Cavazza et al.,
1995; Giudici-Orticoni et al., 2000; Yamanaka & Fukumori, 1995). Both latter
authors agreed that electrons from Fe2+ were transferred by the iron oxidase to
rusticyanin, a blue copper protein which constitutes up to 5 % of the total
protein in iron grown A. ferrooxidans. Later it was demonstrated that the iro
gene (coding for HiPIP in Fe-1 and BMGR strains) had hip gene as the only
counterpart in three other strains. Hip shared 51% similarity with Iro and was up
regulated in S0 grown cells which suggests that it did not act as an iron oxidase
(Bruscella et al., 2005). At the same time, 26 kDa cytochrome c4 was shown by
spectroscopic analysis and site specific mutagenesis to mediate electron
transport between rusticyanin and cytochrome oxidase (Malarte et al., 2005).

24
RusticyaninCyc1(c4)
CycA1(c4)
Cyc2ORF
bc1 complex(PetABC1)
Cytochromeoxidase UQ
O2 H2O
NAD
HI
NAD(P) NAD(P)H
Fe(II) Fe(III)
Operon rus Operon petI
cyc2, orf, coxA, coxD, cyc1,coxB, coxC, rus
cycA1, petA1, petC1,sdrA1, petB1
OM
IM
Periplasm
Cytoplasm
RusticyaninCyc1(c4)
CycA1(c4)
Cyc2ORF
bc1 complex(PetABC1)
Cytochromeoxidase UQ
O2 H2O
NAD
HI
NAD
HI
NAD(P) NAD(P)H
Fe(II) Fe(III)
Operon rus Operon petI
cyc2, orf, coxA, coxD, cyc1,coxB, coxC, rus
cycA1, petA1, petC1,sdrA1, petB1
OM
IM
Periplasm
Cytoplasm Fig. 5 Model proposed for direct and reverse electron transport in A.
ferrooxidans grown on iron. Figure re-drawn from Quatrini et al. (2006).
However, evidence favouring cytochrome Cyc2 as being a potential
iron:rusticyanin oxidoreductase resulted in the model in Fig. 5 (Quatrini et al.,
2006; Yarzabal et al., 2002; Yarzabal et al., 2004). Cyc2 oxidises Fe2+ and
reduces rusticyanin which is the branching point in this electron transport chain.
From rusticyanin, electrons are directed either to cytochrome oxidase via Cyc1
or to the bc1 complex via CycA1. Cyc1 and cytochrome oxidase represent the
“downhill” electron pathway resulting in ATP production, while electron flow
through CycA1 and bc1 is “uphill” i.e. an energy consuming pathway that results
in NAD(P)H production. The requirement for A. ferrooxidans cultured on Fe2+
to expend energy on uphill or reverse electron transport was proposed as early as

25
1982 (Ingledew, 1982). The requirement for uphill electron transport results
from that the redox potential of the couple Fe2+/Fe3+ at pH 2 is more positive
than for NAD(P)H/NAD(P)+ at pH 6.5 (Ingledew, 1982). NAD(P)H is necessary
in the Calvin cycle for CO2 fixation and carbohydrate synthesis (Fig. 6). As
already mentioned, A. ferrooxidans has two bc1 complexes, one of which is
upregulated in S0 grown cells and the other one is upregulated in iron grown
bacteria. The latter bc1 complex is coded by the pet1 operon which also contains
cycA1 and sdrA1. Gene sdrA1 is similar to ribitol/glucose oxidoreductases.
Oxidation of ribitol yields ribulose which can be further converted to ribulose-5-
P and directed to the Calvin cycle. Thus, the suggested role of SdrA1 is to
trigger the Calvin cycle to utilise NAD(P)H produced by reverse electron
transport (Levican et al., 2002). It is also noteworthy that two isotypes of
rusticyanin are found in several strains of A. ferrooxidans. They differ in signal
-0.2
0
0.2
0.4
0.6
0.8
1.0
-0.4
O2
Fe 2+/3+
NAD(P)+
∆μH+ ATP/ADP
Calvin cycleVolt
-0.2
0
0.2
0.4
0.6
0.8
1.0
-0.4
O2
Fe 2+/3+
NAD(P)+
∆μH+ ATP/ADP
Calvin cycle
-0.2
0
0.2
0.4
0.6
0.8
1.0
-0.4
-0.2
0
0.2
0.4
0.6
0.8
1.0
-0.4
O2
Fe 2+/3+
NAD(P)+
∆μH+ ATP/ADP∆μH+ ATP/ADP
Calvin cycleVolt
Fig. 6 Redox and metabolic relationships in A. ferrooxidans. The ∆μH+ (H+ -electrochemical gradient) is used to drive ATP synthesis or reverse electron transport. Re-drawn from Ingledew (1982).

26
peptide sequence and DNA context of their genes (Sasaki et al., 2003) and it is
probable that these strains would not exactly fit to the model in Fig. 5.
Recently, it was shown by several authors that expression of different
protein complexes involved in oxidation of RISCs and Fe2+ is regulated
depending on the availability of the substrate (Bruscella et al., 2007; Quatrini et
al., 2006; Ramirez et al., 2004; Yarzabal et al., 2004). Transcription of petII,
cyo, cyd and doxII operons is upregulated during cultivation on S0, while the rus
and petI operons are upregulated in iron grown bacteria. Intriguingly, closer
investigation of this process revealed transient expression of rus and petII
operons on S0 and iron, respectively. Genes from the rus operon are always
expressed in the early exponential phase on S0 but by the mid-exponential phase
mRNA and corresponding proteins levels were drastically decreased (Yarzabal
et al., 2004). This observation probably explains the conflicting reports on
rusticyanin expression during A. ferrooxidans cultivation on S0 (Ramirez et al.,
2004; Suzuki et al., 1990; Yarzabal et al., 2003). The rus operon is regulated
from three different promoters and post transcriptional processing of its mRNA
is proposed. A similar pattern is observed for petII promoter which is
upregulated and suggested to be involved in direct electron transport in S0 grown
A. ferrooxidans, but transient expression of the operon was observed at the early
stages of growth on iron (Bruscella et al., 2007). The regulatory proteins
involved in this process have not been identified though existence of Fur
regulated genes in A. ferrooxidans was shown (Quatrini et al., 2005) but the
binding site for this transcription factor has not been found for rus, petI or petII.
1.3 Mineral sulphide oxidation
It is generally accepted that acidithiobacilli are involved in release of metals
from metal sulphides (Colmer & Hinkle, 1947; Singer & Stumm, 1970).
However, a debate on the nature of the metal sulphide oxidation has occurred

27
since Silverman and Ehrlich proposed the direct and indirect mechanisms
(Silverman & Ehrlich, 1964; Silverman, 1967). The indirect mechanism implies
that the actual leaching agent is Fe3+ ion which extracts electrons from the
mineral while being reduced to Fe2+. The role of the bacteria is to oxidise Fe2+
into Fe3+ thereby, regenerating the primary oxidant. The direct mechanism
suggests that the microorganisms themselves enzymatically oxidise the mineral
whilst in direct contact. A few workers still support this theory (Lilova &
Karamanev, 2005) though, no enzymes are reported which could be involved in
direct metal sulphide oxidation. On the other hand, a study of the complex
chemical reactions behind the indirect mechanism resulted in an integral model
for bioleaching (Sand et al., 2001; Schippers & Sand, 1999).
An important pre-requisite for this model was a study showing that there
are two types of sulphides and accordingly, two types of possible indirect
leaching mechanisms (Tributsch & Bennett, 1981). Sulphides with valence
bands derived from metal electron orbitals only have their electrons extracted by
the attack of Fe3+ (e.g. MoS2, FeS2 and WS2) while sulphides with valence bands
derived from both metal and sulphur electron orbitals could also be oxidised via
proton attack (e.g. CdS and ZnS). The valence band in the band theory of solids
is an energy level occupied by valence electrons (i.e. electrons from the outer
shell of an atom) and valence electrons are thought to determine the chemical
properties of atoms in reactions (Atkins & de Paula, 2006). Metal sulphides
from the first group (MoS2, FeS2 and WS2) are oxidised via the indirect
thiosulphate pathway (Fig. 7) that results in the production of thiosulphate and
Fe2+ as the primary leaching products (Schippers & Sand, 1999). Thiosulphate is
further oxidised in a cyclic manner to sulphate with tetrathionate, disulphane-
monosulphonic acid and trithionate as intermediate products (Schippers et al.,
1996). It is speculated that the oxidation of these sulphur compounds is mainly
of a chemical character but participation of such enzymes as
tetrathionatehydrolase of acidithiobacilli is also suggested (Sand et al., 2001).
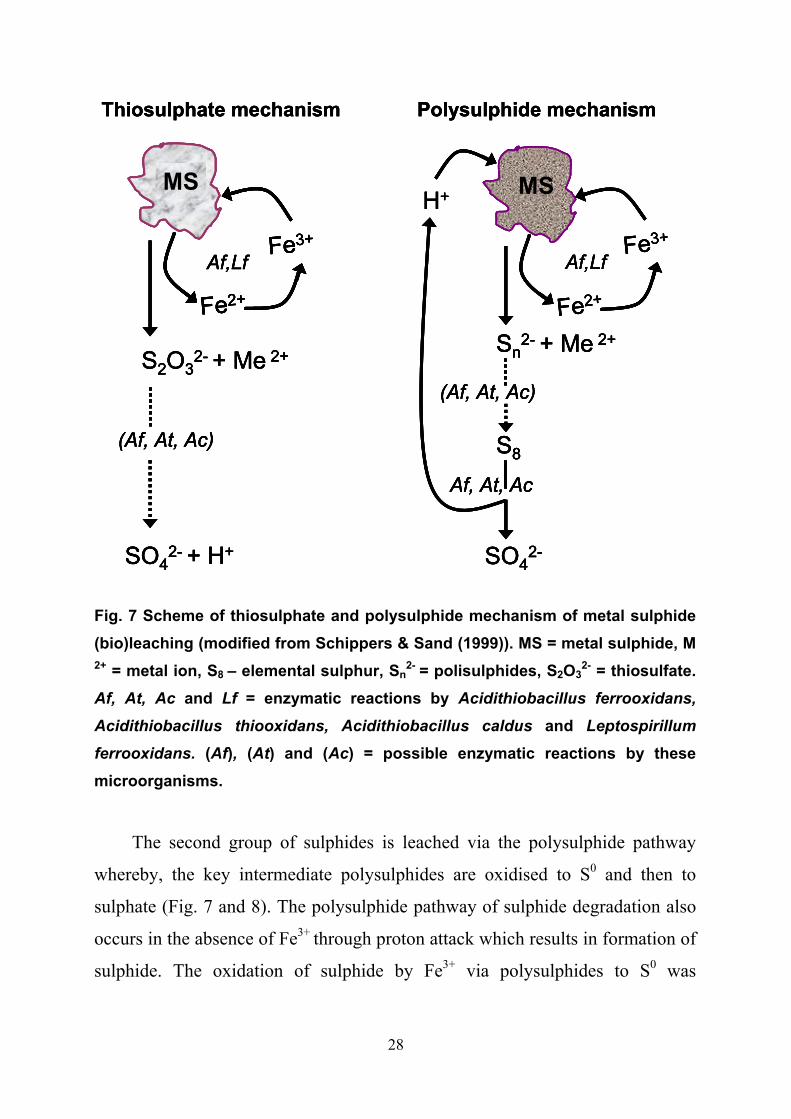
28
MS
Fe3+
Fe2+
MS
Fe3+
Fe2+
SO42- + H+
(Af, At, Ac)
S2O32- + Me 2+
Af,Lf
H+
SO42-
S8
Sn2- + Me 2+
Af, At, Ac
(Af, At, Ac)
Af,Lf
Thiosulphate mechanism Polysulphide mechanism
MS
Fe3+
Fe2+
MSMS
Fe3+
Fe2+
Fe3+
Fe2+
MS
Fe3+
Fe2+
SO42- + H+
(Af, At, Ac)
S2O32- + Me 2+
Af,Lf
H+
SO42-
S8
Sn2- + Me 2+
Af, At, Ac
(Af, At, Ac)
Af,Lf
Thiosulphate mechanism Polysulphide mechanism
MS
Fe3+
Fe2+
SO42- + H+
(Af, At, Ac)
S2O32- + Me 2+
Af,Lf
MSMS
Fe3+
Fe2+
Fe3+
Fe2+
SO42- + H+
(Af, At, Ac)
S2O32- + Me 2+
Af,Lf
H+
SO42-
S8
Sn2- + Me 2+
Af, At, Ac
(Af, At, Ac)
Af,Lf
H+
SO42-
S8
Sn2- + Me 2+
Af, At, Ac
(Af, At, Ac)
Af,Lf
Thiosulphate mechanism Polysulphide mechanism
Fig. 7 Scheme of thiosulphate and polysulphide mechanism of metal sulphide (bio)leaching (modified from Schippers & Sand (1999)). MS = metal sulphide, M 2+ = metal ion, S8
– elemental sulphur, Sn2- = polisulphides, S2O3
2- = thiosulfate. Af, At, Ac and Lf = enzymatic reactions by Acidithiobacillus ferrooxidans,
Acidithiobacillus thiooxidans, Acidithiobacillus caldus and Leptospirillum
ferrooxidans. (Af), (At) and (Ac) = possible enzymatic reactions by these microorganisms.
The second group of sulphides is leached via the polysulphide pathway
whereby, the key intermediate polysulphides are oxidised to S0 and then to
sulphate (Fig. 7 and 8). The polysulphide pathway of sulphide degradation also
occurs in the absence of Fe3+ through proton attack which results in formation of
sulphide. The oxidation of sulphide by Fe3+ via polysulphides to S0 was

29
MS2H+ M2+
Fe3+ Fe2+
[H2S*+ H2Sn]HS* S8
H+ H+
Fe3+ Fe2+
MS2H+ M2+
Fe3+ Fe2+
[H2S*+ H2Sn]HS* S8
H+ H+
Fe3+ Fe2+
Fig. 8 Simplified scheme from the polysulphide mechanism. Re-drawn from Schippers & Sand (1999).
demonstrated (Steudel, 1996) but no information is available on the involvement
of microorganisms in this step. Additionally, Tributsch (2001) claims that
polysulphides may also cause dissolution of metal sulphides.
It was proposed to distinguish contact and non-contact leaching within
indirect oxidation of metal sulphides (Tributsch, 2001) as A. ferrooxidans is able
to attach to mineral surfaces and create dents and pits (Escobar et al., 1997;
Gehrke et al., 1998; Mangold et al., 2008; Sand & Gehrke, 2006). Attachment
occurred with the help of an extracellular polymeric substance (EPS) layer
which is proposed to facilitate mineral dissolution due to increased Fe3+
concentration. The iron oxidising bacterium L. ferrooxidans has also been
shown to attach to pyrite and create a characteristic hole pattern on the mineral
surface (Rojas-Chapana & Tributsch, 2004). Thus, leaching by Fe3+ ions
concentrated within EPS of the attached bacteria is referred to as contact indirect
bioleaching. Leaching catalysed by free floating microorganisms is termed non-
contact indirect bioleaching.

30
1.4 Acid mine and rock drainage
AMD or ARD are terms used to describe drastic acidification (down to pH –3.6)
of surface and ground waters by products of metal sulphide (bio)leaching
(Nordstrom et al., 2000). Acidification is accompanied by high concentrations
of iron and other metals such as copper, zinc, lead, chromium and arsenic.
Opencast and underground abandoned mines as well as mining industry waste
(tailings) are the source of the AMD/ARD. The mining industry is widespread
over the world and hence AMD is a common phenomenon and the involvement
of microorganisms in AMD was proved long ago (Colmer & Hinkle, 1947).
Leaching of metal sulphides exposed to water and oxygen occurs via a purely
chemical mechanism but the presence of S0 and iron oxidising microorganisms
increases the chemical reaction rate by more than 106 fold (Singer & Stumm,
1970). Several factors such as temperature, age, chemical composition of the
rocks, hydrological regime of the site and mining technique influences AMD
development. Consequently, every AMD is unique but nevertheless, some
general trends in chemical and microbial characteristics can be found. An
optimal combination of AMD forming factors gave one of the biggest and most
studied AMDs in the world at Iron Mountain, Shasta County, northwest
California. The Iron mountain mining area comprises several mines that were
operated from the 1860s to 1962 when Ag, Au, Cu, Fe, Zn and pyrite were
recovered. The brittle rhyolite bedrock of the site creates a fracture-flow type
hydrologic conditions that has little buffering capacity. The massive deposit (95
% pyrite) located at, or above the water table was excavated by tunnels and
shafts where airflow generated by heat production from pyrite oxidation
produces a subsurface temperature of 50ºC (Nordstrom & Alpers, 1999;
Nordstrom et al., 2000). High availability of oxygen and moisture as well as a
mild to high temperature range benefits a wide variety of iron and S0 oxidisers
(Bond et al., 2000; Edwards et al., 1999a; Edwards et al., 1999b; Schrenk et al.,

31
1998). In 1983 Iron Mountain was ranked as the third largest polluter in the
State of California on National Priorities List for Environmental Protection
Agency Superfund (Iron Mountain Mine Study Case,
http://www.epa.gov/aml/tech/imm.pdf, accessed January 13, 2008).
Approximately 2,500 tons of pyrite were annually weathered from the
Richmond mine before large scale remediation efforts were initiated in the late
1980s (Nordstrom & Alpers, 1999). The AMD contained very high levels of
metals and approximately 300 tons of dissolved Cu, Cd and Zn were released
into the Sacramento river. Sudden increases in AMD release into the
Sacramento river caused massive fish deaths and > 47,000 trout died in a single
week in 1967 (Nordstrom & Alpers, 1999).
AMD is gravely hazardous for most living organisms due to its high acidity
and presence of heavy metals producing symptoms such as gill lesions in fish
(Henry et al., 1999; Henry et al., 2001). Also, sheep refuse to drink synthetic
AMD water even without being given an alternative due to dissolved iron rather
than the pH (Horvath, 1985). AMD waters are lethal to seaweeds and
invertebrates in laboratory and field conditions (Marsden & DeWreede, 2000;
O'Halloran et al., 2008). Damage caused by heavy metals is detected in rodent
liver, kidney and testis caught near the metal processing plants (Damek-Poprawa
& Sawicka-Kapusta, 2003). Cd, Cu and Zn were shown to be lethal in high
doses and decrease the survival in low doses to various vertebrates and
invertebrates (Baker et al., 1998; Gheorghiu et al., 2007; Handy, 2003; Jianhua
et al., 2006). Heterotrophic bacteria commonly occurring in non-acidic streams
could not survive in streams polluted with AMD at pH <3 (Tuttle et al., 1968;
Walton & Johnson, 1992). The environmental pollution caused by AMD is long-
lasting and some areas have suffered its impact for hundreds of years (Demchak
et al., 2004; Younger, 1997). Species number and abundance in marine and
fresh waters remain decreased for several years after remediation or even total
neutralisation of the AMD source (Simon et al., 2006; Zis et al., 2004).

32
AMD is extremely toxic for neutrophilic organisms though it creates
suitable conditions for a variety of acidophilic bacteria, archaea and algae that
often possess genetic determinants for metal resistance (Dopson et al., 2001).
An interesting characteristic of such biogeosystems is a gradual shift in pH,
metal concentration, dissolved oxygen and organic carbon depending on the
distance from the AMD source which is reflected in the changing composition
of microbial communities. Moreover, sediment layers from different depths of
AMD streams are populated by defined groups of bacteria, algae and archaea
(Johnson et al., 2001; Rowe et al., 2007). Microbial communities analysed at
different sites of Iron Mountain AMD (pH 0.77 to 1.31) were dominated by
“Ferroplasma” spp., Leptospirillum group I, II and III, Sulfobacillus spp. and
Acidimicrobium spp.. A. ferrooxidans was not found suggesting its lack of
involvement in pyrite dissolution at Iron Mountain (Baker & Banfield, 2003;
Bond et al., 2000; Schrenk et al., 1998). On the other hand, data was obtained
showing A. ferrooxidans to be important in colonisation of pyrite at
circumneutral pH. When attached to pyrite bacteria trigger leaching reactions
and push the pH down preparing conditions suitable for other acidophiles
(Mielke et al., 2003). A. ferrooxidans dominates in oxygen-depleted mine water,
Huelva Spain (pH 2.67) which was supposedly conditioned by facultatively
anaerobic metabolism of A. ferrooxidans (Rowe et al., 2007). Studies of
microbial communities of Tinto river (pH 2.1 to 4.7) showed A. ferrooxidans to
be one of the most available species both in aerobic and anaerobic conditions
(Gonzalez-Toril et al., 2003). In oxic wetlands receiving acid coal mine drainage
waters (influent pH 2 to 3.9) A. ferrooxidans together with A. thiooxidans
accounted for 34 % of the bacterial population detected by fluorescent in situ
hybridisation (FISH) (Nicomrat et al., 2006a; Nicomrat et al., 2006b). A.
ferrooxidans is often found in numbers lower than L. ferrooxidans in AMD and
bioleaching set-ups (Rawlings et al., 1999). However, Johnson et al. (2001)
reports that an isolate with 98 % similarity to A. ferrooxidans dominated

33
Leptospirillum – like bacteria in all water samples taken at King’s Mine,
Norway (pH 2.75 to 3.71). Although L. ferrooxidans is more tolerant to Fe3+
than A. ferrooxidans the latter has the advantage of being more resistant to low
temperatures (Hallmann et al., 1992; Leduc et al., 1993; Rawlings et al., 1999).
Thus, the presence of a dominant A. ferrooxidans at the subarctic climate zone
where King’s Mine is located with temperatures remaining below zero for more
than half a year is consistent with other studies. Interestingly, low temperature
(7°C) enrichment cultures from the black schist Talvivaara deposit, Finland
demonstrated the presence of A. ferrooxidans, A. thiooxidans and A. albertensis
while at higher temperatures L. ferrooxidans and A. caldus were prevailing
(Puhakka et al., 2007). It appears that A. ferrooxidans plays multiple roles in
ecosystems based on iron and sulphur compounds as the main source of energy
and its colonisation on pyrite decreases the pH in the initial step of leaching. It
oxidises Fe2+ to Fe3+ under aerobic conditions and reducing Fe3+ by oxidation of
RISCs under anaerobic conditions supporting iron cycling in AMD waters and
remediation wetlands. Finally, it is likely that A. ferrooxidans is the main
microorganism catalysing pyrite oxidation in boreal climates.
Chemolithotrophic bacteria and archaea are not the only microorganisms
found in AMD environments and algae such as Chlamydomonas sp., Euglena cf.
mutabilis, Chlorella sp. as well as several others species were isolated from the
Tinto river (Aguilera et al., 2006; Amaral Zettler et al., 2003; Rowe et al.,
2007). No photosynthetic algae were found in eukaryotic Iron Mountain
subsurface biofilms which is attributed to the absence of a phototrophic niche
rather than the extremely low pH or high temperature. Curiously, the 16S rRNA
gene sequences of Dodhideomycetes and Eurotiomycetes found in this
ecosystem were highly similar to neutrophilic lineages which implies that either
colonisation of AMD by these fungi occurred relatively recently or that AMD
conditions are quite acceptable for these species and do not exert significant
evolutionary pressure (Baker et al., 2004). Twenty-seven yeast species from

34
different AMD sampling points have been isolated from the Iberian Pyrite Belt
region, Spain and Portugal (Gadanho et al., 2006). Many of these species
belonged to Candida and Cryptococcus genera whereas, 48 % probably
constituted new taxa. The authors note that the composition of yeast consortia is
similar in related environmental stress conditions (pH and salt concentration
etc.) and did not depend on geographical location (Gadanho et al., 2006).
Similarly, principal-component analysis of 16S rRNA and gyrB genes from
AMD microbial communities of Dexing copper mine, Jiangxi, China suggests
that sites with similar geochemical characteristics harbour communities of a
close structure (Yin et al., 2008). Heterotrophic, acidophilic bacteria have been
isolated and characterised from AMD sites. Despite their ability to oxidise Fe2+
their contribution to the total iron oxidation is considered to be negligible.
Nevertheless, under certain conditions these bacteria could be responsible for
the larger part of the biomass production (Johnson et al., 1992; Wichlacz & Unz,
1981).
Many approaches have emerged to prevent and/or mitigate the problem of
AMD that have been divided into “source control” and “migration control”
measures or in other words prevention and remediation (Johnson & Hallberg,
2005). The “migration control” group comprises both active and passive
methods while “source control” alternatives include:-
flooding or sealing of a mine
land or water covering of mine tailings / rock capping
mineral waste blending with acid consuming material
bioshrouding / biological source treatment
application of biocidic agents
solidification of tailings
chemical coating

35
All these methods have different degrees of efficiency, certain drawbacks and
should be applied carefully taking into concern the geochemical characteristics
of the site and nature of AMD source (Anonymous, 1994; Gatzweiler et al.,
2001). For example, efflorescent salts that accumulate at the site could produce
even heavier AMD if dissolved, which makes a widely-used method of mine
flooding undesirable (Nordstrom & Alpers, 1999). Tailings covered with water
release heavy metals from the surface layer even under anoxic conditions and
major spills from malfunctioning and corroded tailings dams cause serious
damage to environment and agriculture (Bjelkevik, 2005; Olias et al., 2006;
Vigneault et al., 2001).
Biological source treatment and bioshrouding are recently proposed
methods for source treatment (Jin et al., 2007; Johnson et al., 2008). Biological
source treatment is a down-hole injection of sulphate reducing bacteria (SRB)
and organic substrate in electromagnetically determined underground AMD “hot
spots”. Microbial sulphate reduction occurs naturally in AMD sites or at
locations receiving acid mine flows (Herlihy & Mills, 1985; Tuttle et al., 1969a)
and it has been suggested to be applied for amendment of AMD (Tuttle et al.,
1969b). SRB are used to mitigate AMD in bioreactors and in situ providing
efficient metal removal (Dvorak et al., 1992; Koschorreck et al., 2007; Neculita
et al., 2007). Bioshrouding utilises inoculation of freshly ground tailings with
heterotrophic, Fe3+ reducing acidophiles from the Acidiphilium and Acidocella
genera. Bacteria form a biofilm on the tailings surface and reduce Fe3+, the
primary oxidant of AMD (Johnson et al., 2008). Measures of AMD remediation
are summarised in Fig. 9.

36
Biological
Abiotic
Aerobic wetlands
Compost reactors wetlands
Permeable reactive barriers
Packed bed iron-oxidationbioreactor
Active
Passive
Off-line sulphidogenic bioreactors
Active systems: aeration and lime additionPassive systems: e.g. anoxic limestone drains
RE
ME
DIA
TIO
N
Biological
Abiotic
Aerobic wetlands
Compost reactors wetlands
Permeable reactive barriers
Packed bed iron-oxidationbioreactor
Active
Passive
Off-line sulphidogenic bioreactors
Active systems: aeration and lime additionPassive systems: e.g. anoxic limestone drains
RE
ME
DIA
TIO
N
Fig. 9 Biological and abiotic strategies for remediation of acid mine drainage
waters. Adapted from Johnson & Hallberg (2005).
1.5 Biomining
1.5.1 Biomining as an industrial process
The ability of microorganisms to oxidise Fe2+ and S0 is widely used for metal
recovery from sulphidic ores, a process known as biomining. The term
‘bioleaching’ refers to recovery of the metal of interest when it is chemically
bound by sulphide (e.g. copper in chalcopyrite (CuFeS2)) and ‘biooxidation’ is
the biomining of a metal of interest which is not chemically bound in the
sulphide (e.g. gold particles embedded in arsenopyrite). Biomining is an
increasingly developing industry (McCready, 1987; Songrong et al., 2002;
Torma, 1985) and the following commercial-scale leaching techniques have
been developed: in situ leaching, dump leaching, heap leaching, vat leaching
and reactor leaching (Rossi, 1990). In situ leaching is a little utilised approach,
limited mainly to bioleaching of copper and uranium. The low grade deposit is
blasted and bioleached at place making the capital and operational costs low but

37
significantly increasing the negative environmental impact of this technology
(Rossi, 1990; Torma, 1985; Tuovinen, 1985). Dump and heap bioleaching are
similar techniques exploiting metal recovery from the leachate collected in the
base of a dump (heap) of crushed rock. However, a dump is an unorganised pile
of mine waste or run-off-mine ore while a heap is an irrigated and aerated
construction with drainage layer and watertight lining at the bottom (Fig. 10)
(Groudev et al., 2001; Logan et al., 2007; Petersen & Dixon, 2007). The
drawback of even highly engineered heaps is that bioleaching takes months and
a large percentage of the target metals remains unleached (Petersen & Dixon,
2007). GEOCOAT® is a new technology involving heap construction of a
specially prepared metal sulphide source that promises substantially increased
oxidation time and higher metal output. The source for the heap construction is
Irrigation
Aeration
Solution collectionDrainage layer
Heap packing (ore)
Irrigation
Aeration
Solution collectionDrainage layer
Heap packing (ore)
Fig. 10 A construction of an engineered heap (up arrows designate air flow and
down arrows the liquid percolation). Adapted from Petersen & Dixon (2007).

38
particles of a bare rock 6 to 25mm in diameter with 0.5 mm thick coating of a
fine milled ore (Harvey & Bath, 2007). In contrast, vat leaching is carried out in
a series of concrete vats (tanks) filled with crushed ore where the leach solution
is fed from the bottom of the vat and overflows to the next vat (Rossi, 1990).
Additionally, bioleaching of high-value ores and concentrates is economically
successful in stirred, aerated and temperature controlled reactors and constitute a
highly developed biomining technology (Do Carmo et al., 2001; Rawlings,
2002; Van Aswegen et al., 2007). Stirred bioreactors with different kinds of
impellers are mainly used in these processes but new designs like “aerated
through bioreactor” or “low energy bioreactor” are being currently being
developed (Rossi, 2001).
1.5.2 Acidophilic microorganisms in industrial bioleaching
For many decades A. ferrooxidans was considered to be the primary
microorganism responsible for catalysis of metal sulphide oxidation due to their
selection in culture-dependent analyses of bioleaching samples (Leveille et al.,
2001; Mishra et al., 1983; Schippers et al., 1995). However, the development of
culture independent microbial molecular techniques identified that other species
were dominant in many bioleaching environments, especially bioreactors
treating mineral concentrates (Hugenholtz et al., 1998; Johnson, 2001; Tan et
al., 2007). Biodiversity in industrial bioleaching processes can be divided
according to their growth temperature ranges into mesophiles at approximately
30°C, moderate thermophiles around 45°C and thermophiles at approximately
70°C (Norris, 2007; Rawlings, 2002). Typical mesophilic bioleaching
microorganisms include the iron and S0 oxidisers A. ferrooxidans, “Sulfobacillus
monseratensis” and Thiomonas intermedia; the iron oxidiser L. ferrooxidans;
the sulphur oxidiser A. thiooxidans; and heterotrophic Acidiphilium spp.
(Battaglia-Brunet et al., 2006; Norris et al., 2000; Rawlings & Johnson, 2007;

39
Yahya & Johnson, 2002). Recently, L. ferrooxidans was reclassified and the
new species Leptospirillum ferriphilum and Leptospirillum ferrodiazotrophum
were formed. L. ferriphilum has a higher optimal growth temperature than L.
ferrooxidans and L. ferrodiazotrophum was proposed as the main nitrogen fixer
in an acidophilic AMD microbial community (Coram & Rawlings, 2002; Tyson
et al., 2005). Consequently, L. ferriphilum is found in moderately thermophilic
consortia together with A. caldus, Acidimicrobium ferrooxidans, Ferroplasma
acidiphilum and Sulfobacillus spp. (Dopson & Lindström, 1999; Karavaiko et
al., 1987; Norris, 2007). Thermophilic stirred tank bioleaching consortia are
dominated by archaea of Sulfolobus, Metallosphaera, Stygiolobus and Acidianus
genera (Dopson et al., 2006; Mikkelsen et al., 2006). Mesophiles and moderate
thermophiles are often isolated from heap leaching but thermophilic archaea of
Thermoplasma and Sulfolobus genera are also present due to high temperatures
measured in the deeper layers of a heap (Puhakka et al., 2007a; Rawlings &
Johnson, 2007; Xie et al., 2007).
The microbiology of biomining has several peculiar characteristics that are
worth mentioning. The catalysis of mineral oxidation is carried out by mixed
consortia in non sterile conditions (Rawlings, 2002; Rossi, 1990). It was noticed
that adaptation of the culture to specific mineral, temperature and other factors
is crucial for efficient bioleaching/biooxidation and though stirred tanks and
heaps are supplied with the initial inoculum the microbial consortium
composition, as a rule, changes during leaching due to the presence of
indigenous microorganisms in the mineral and selective pressure of the system
(Morin, 2007; Rawlings, 2007; Plumb et al., 2007). In stirred tanks where
temperature, aeration and pH are maintained at the same level only a few
species were found to be dominating (Mikkelsen et al., 2006). In heap
bioleaching a temperature gradient across a heap creates multiple niches for
survival of species with different growth temperature requirements (Plumb et
al., 2007; Rawlings, 2007). Though bioleaching in stirred tanks and heaps are

40
dominated primarily by autotrophic microorganisms, heterotrophic and
facultatively heterotrophic acidophiles and iron oxidisers also contribute to
mineral sollubilisation. The relationships of autotrophs and heterotrophs in such
systems are reciprocal; heterotrophs remove organic substances which are toxic
to certain autotrophs (e.g. Leptospirillum spp.) and autotrophs provide necessary
organic compounds to heterotrophic iron oxidisers (Okibe & Johnson, 2004).

41
2. AIMS OF THE STUDY
Heap leaching is a developing industry and there is a need for experimental
studies of ore arrangement and management in the heap as well as analysis of
microbial activity during heap bioleaching (Petersen & Dixon, 2007).
The first aim of this study was to test whether the novel vacuum-positive
pressure driven bioreactor was suitable for modelling of heap bioleaching.
Heap leaching in arctic and sub arctic environments has a big potential (Puhakka
et al., 2007) though new technologies must be developed for bioleaching in cold
temperatures.
The second aim of this study was to characterise a new psychrotolerant
strain of A. ferrooxidans which dominates heap leaching of black schist at
low temperatures (Finland).
Occurrence of polythionates (tetrathionate and thiosulphate etc.) in cyanidation
slurry increases cyanide consumption in industrial bioleaching (Van Aswegen et
al., 2007).
The third aim of this study was to investigate regulation of expression of
tetrathionate hydrolase from A. caldus, a candidate microorganism to use
in pre-cyanidation treatment.

42

43
3. RESULTS AND DISCUSSION
PAPER I
A novel reactor design was presented that was used for bioleaching with an
acidophilic mixed culture and its performance compared to bioleaching in a
stirred tank reactor inoculated with the same culture. The new reactor utilised
cycles of positive and negative pressure applied to a system of two connected
vessels (Fig. 11).
Fig. 11 Design of the novel reactor.
The design allowed the leachate to circulate between the vessels and drain
through a layer of finely milled mineral (tailings) placed on the filter in vessel B
and thus, simulate heap bioleaching. Resting periods between phases when
positive or negative pressure was applied helped to lower energy consumption.
Compressed air pump
Ground
Spray nozzle
Tube a
One-way valve for aeration
Filter cloth (felt liner)
Tube b
Sensor controlling upper level of liquid
One-way valve Vessel B
Vessel A

44
The average leaching rate in the new bioreactor was 0.17 g Fe/l·day, while in the
conventional stirred reactor it was 0.50 g Fe/l·day. The dissolved oxygen
concentration in vessel A fluctuated from 8.4 to 9.4 mg/l depending on the phase
of the cycle whereas; typical dissolved oxygen concentrations in commercial
leaching bioreactors are between 1 to 2 mg/l (Van Aswegen et al., 2007 and
Anthony Pinches, personal communication). The bioleaching proceeded for 83
days and the changes in microbial composition of the mixed culture was
analysed by denaturing gradient gel electrophoresis (DGGE; Paper I, Fig. 4). By
cloning and sequencing the 16S rRNA gene, clones related to A. thiooxidans,
Leptospirillum OS7, L. ferriphilum and Sulfobacillus Y0017 were identified
from both reactor types. Additionally, an archaeon similar to F. acidiphilum was
identified from the novel bioreactor.
Although many microorganisms were observed in the leachate (liquid
media) no biofilm was found on the tailings (vessel B). During the resting phase
of the cycle the leachate percolated through the vessel and thereby, wetted and
aerated the tailings. It is possible that the leachate drained through the tailings
too quickly and over drying of the mineral particles occurred during the resting
and negative pressure phases (95 s) that prevented biofilm formation. Recent
data on the importance of biofilms in bioleaching (contact indirect bioleaching)
suggests that the results obtained from the novel bioreactor may be improved if
biofilm growth could be promoted (Sand & Gehrke, 2006). To obtain a biofilm
on the tailings, the resting time could be shortened or vessel B could be packed
with a thicker layer of tailings to lower the percolation rate and facilitate
bacterial attachment. By FISH, most of the active Leptospirillum spp. and
Sulfobacillus spp. were found at the bottom of vessel B (Fig. 12). Fe3+ and
protons generated by these bacteria oxidised the tailings during leachate
circulation and thus, leaching was due to non-contact in-direct bioleaching. The
bioreactor is suitable for modelling of heap bioleaching and microbial
community studies in AMD environments.
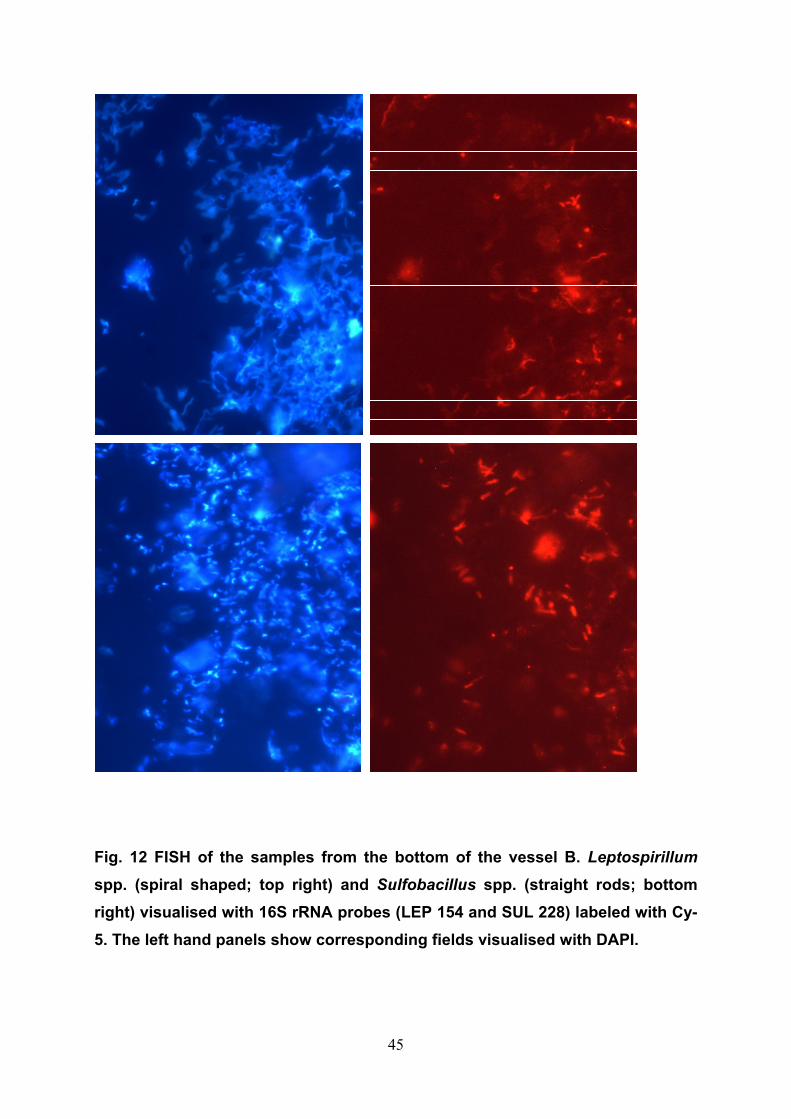
45
Fig. 12 FISH of the samples from the bottom of the vessel B. Leptospirillum spp. (spiral shaped; top right) and Sulfobacillus spp. (straight rods; bottom right) visualised with 16S rRNA probes (LEP 154 and SUL 228) labeled with Cy-5. The left hand panels show corresponding fields visualised with DAPI.

46
PAPER II
Knowledge of RISC metabolism and its regulation in acidophilic
microorganisms might be useful for industrial bioleaching processes.
Cyanidation is a common hydrometallurgical gold recovery process based on the
reaction of gold with cyanide whereby, gold is converted into the water soluble
aurocyanide complex that is collected by adsorption on carbon particles
(International Cyanide Management Institute
http://www.cyanidecode.org/cyanide_use.php; accessed 25.01.2008). Pre-
treatment is necessary to extract gold from sulphide ores (typically arsenopyrite)
in order to dissolve the mineral and liberate the gold. Polysulphides and
polythionates (e.g. tetrathionate and thiosulphate) produced during mineral
sulphide oxidation compete for cyanide in the cyanidation step according to the
following reactions:
S4O62- + 3CN- + H2O → S2O3
2- +2HCN +SCN- (Kelly et al., 1969)
SxO32- + CN- → SO3
2- +SCN- (Van Aswegen et al., 2007)
Therefore, RISCs present in the slurry consume cyanide required for successful
gold recovery and one solution to decrease its consumption is oxidation of the
residual RISCs by acidophilic sulphur oxidisers. Reductions in cyanide
consumption will not only reduce costs but also lower nitrate (the final SCN-
breakdown product) release into the environment that can result in
eutrophication of water bodies. Higher temperatures during pre-cyanidation
treatment ensures more efficient RISC oxidation (Van Aswegen et al., 2007)
and therefore, use of the moderately thermophilic sulphur oxidiser A. caldus in
this step might be of interest. Tetrathionate hydrolase (TetH) has been purified
and described as the enzyme responsible for tetrathionate decomposition

47
ISac1 rsrR rsrS tetH doxD
1000 bp
P0 P1
ISac1 rsrR rsrS tetH doxD
1000 bp1000 bp
P0 P1
Fig. 13 Schematic representation of tetH gene cluster.
(Bugaytsova & Lindström, 2004). In this study, we investigated A. caldus TetH
regulation and characterised the first gene cluster containing tetH (Fig. 13).
Expression of the enzyme was compared during A. caldus cultivation on
tetrathionate and S0 by Western-blot analysis (Paper II, Fig. 3) and Q-PCR. TetH
expression during bioleaching was demonstrated in the Western blot confirming
that investigations of TetH are relevant to bioleaching environments. It was
demonstrated that TetH expression was transcriptionally regulated and was
233.5 ± 134 fold higher in tetrathionate grown cells compared those grown on
S0. Genes coding for a putative transposase, a two component regulatory system
(RsrRS) and DoxD were co-transcribed with tetH. P0 and P1 putative promoters
(Fig. 13) were identified in the cluster and shown to increase transcription of β-
galactosidase when cloned up-stream of the lac operon of pRW2 plasmid in E.
coli. The transcription start site of the P1 promoter was mapped by primer
extension (Paper II, Fig.4).
TetH was similar to pyrrolo-quinoline quinine (PQQ) binding
dehydrogenases ((Paper II, Fig. 2) and the presence of a quinone moiety was
suggested for this protein based on the positive staining with NBT-glycinate.
Previously, it was shown that tetrathionate oxidation is inhibited by electron
transport uncouplers (Hallberg et al., 1996). This data taken together, led us to
propose that TetH was feeding electrons from tetrathionate to the electron
transport chain. Protein crystallisation experiments are on-going to elucidate the

48
A BA B
Fig. 14 Crystals of TetH in 3M MgSO4 (A) and 1.5M MgSO4, 2.5M NaCl and zwittergent 3-12 (B).
mechanism of tetrathionate oxidation reaction through analysis of its crystal
structure. Initial crystals have been obtained (Fig. 14) but further optimisation of
the crystallisation conditions is needed.
PAPERS III AND IV
Papers III and IV are tightly connected. In paper III we characterise A.
ferrooxidans strain SS3 as the first psychrotolerant acidophilic
chemolithotrophic iron oxidiser. In paper IV we investigate RISC oxidation
during cold temperature leaching by A. ferrooxidans strain SS3 and T7 mix
psychrotolerant cultures. Both studies were aimed at understanding and
improving bioleaching processes in boreal climates. We show that strain SS3
was able to grow at 5°C by oxidising Fe2+ (Fig.15) with the rate constant ranging
from 0.0162 to 0.0104 h-1 (generation time of 43-58 h) depending on the initial
pH. A mesophilic A. ferrooxidans reference strain was substantially less
efficient under the same conditions (Paper III, Fig. 2).

49
Fig. 15 Iron oxidation at 5°C. A. ferrooxidans SS3 at the initial pH 2.8 (■) and 2.0 (∆); mesophilic A. ferrooxidans reference strain at initial pH2.0 (◊); abiotic control (Χ). From paper III.
Tetrathionate was shown to support the growth of strain SS3 at both 5° and
30°C (Paper III, Fig. 5). 16S rRNA gene phylogeny showed strains SS3, SK3
and SK5 to be similar to several A. ferrooxidans isolates and clones acquired
from arctic and sub-arctic environments (Paper III, Fig. 1). Also, several of these
isolates are psychrotolerant suggesting that they may constitute a new species
and experiments to test this hypothesis are ongoing. Bioleaching of different
sulphidic ores at 5°C with strain SS3 showed that it was 5 to 6 times more
efficient than the mesophilic A. ferrooxidansT (Dopson et al., 2007). Chemical
analysis of leachate and solids of chalcopyrite and complex Talvivaara ore
showed a transient formation of S0 in the reactors (Paper IV, Fig. 1).
Considering that accumulation of S0 was accompanied by a halt in metal
solubilisation we suspected that the S0 was responsible for mineral passivation.
4000 200
Oxi
dize
d iro
n (m
M)
Hours4000 200
Oxi
dize
d iro
n (m
M)
Hours4000 200
Oxi
dize
d iro
n (m
M)
Hours

50
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.40
Gene expression relative to rrs
0 2 4 6 8 10 12 14 160
10
20
30
40
50
60
70
80
90
100
Sub
stra
te (m
M)
Culture volumes
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.40
Gene expression relative to rrs
0 2 4 6 8 10 12 14 160
10
20
30
40
50
60
70
80
90
100
Sub
stra
te (m
M)
Culture volumes
Fig. 16 Expression of A. ferrooxidans SS3 cyoB (black), petA (light grey) and petB (dark grey) genes in tetrathionate and Fe2+ limited continuous culture (all values are expressed as n-fold relative to rrs gene (5 x 10-4)). Lines represent theoretical changes in tetrathionate (dashed) and Fe2+ (solid) concentrations.
Almost no S0 was detected during pyrite leaching as its oxidation occurs via the
thiosulphate mechanism (Schippers & Sand, 1999). Expression of the A.
ferrooxidans SS3 genes involved in S0 and Fe2+ oxidation (Fig. 16) had the same
pattern as for the mesophilic A. ferrooxidansT such that cyoB was up regulated
on the RISC compound tetrathionate and petA and petB were up regulated on
Fe2+ (Quatrini et al., 2006). When bacteria were cultivated on both substrates,
intermediate levels of the expression for all three genes were measured (Paper
IV, Fig. 2). The results suggest that transient oxidation of RISCs in strain SS3
can occur alongside with Fe2+ oxidation.

51
4. CONCLUSIONS
A novel bioreactor performed efficiently during the conditions tested for
simulating heap bioleaching though it was not as efficient as a stirred reactor.
Oxidation of iron and catalysis of mineral dissolution by a novel
psychrotolerant A. ferrooxidans strain SS3 was more efficient compared to A.
ferrooxidansT at low temperatures.
S0 was transiently accumulated in low temperature bioleaching of
chalcopyrite and chalcopyrite containing ore with psychrotolerant pure and
mixed cultures potentially causing passivation of the mineral surface.
Expression of the genes proposed to be responsible for iron and S0
oxidation in A. ferrooxidansT had a similar pattern in strain SS3 suggesting that
iron and RISC oxidation in both strains occurs through similar pathways.
During cultivation on mixed substrate (Fe2+ and tetrathionate) a transient
expression of corresponding genes proposed to be involved in Fe2+ and
tetrathionate oxidation was observed.
Expression of TetH was transcriptionally regulated in a substrate
dependent manner. It was proposed that this enzyme belongs to a group of
acidophilic dehydrogenases.

52

53
5. ACKNOWLEDGMENTS
Six years at the Umea University were happy, interesting and intensive time for me and here
comes the list of people who contributed to this feeling. I’m grateful to my supervisors, Börje
Lindström and Mark Dopson; to Börje for taking me in this project and to Mark for
helping me to get out of it. Both of the guys gave me a lot of freedom so I could go to
different conferences and courses, sleep in the morning as much as I like and do whatever
experiments. Mark only insisted that whatever experiments should eventually take a form of
published articles, which is in fact a sober thinking in the modern science. I’d like to thank all
the former and present members of my group: Siv Sääf who for me is an expression of
stability, energy, good mood and perfect technical skills. The group can have a different
supervisor but Siv is always Siv; Zhanna Bugaytsova, a woman of a strong character and an
author of an inspiring article on the tetrathionate hydrolase; Rana Atahr, for her careful
teaching and extremely polite instructions; Stefanita Mangold and Dr. Shabir, who arrived
recently but it feels like we’ve been friends since long ago. I’d like to acknowledge my great
co-workers and teachers Violaine Bonnefoy (for hosting me in her lab and opening my eyes
on French food) and David Holmes (for taking my invitation and coming to Umeå from the
other side of the world with a talk, lots of ideas and a stone egg), remote co-supervisor Olli
Tuovinen for involving me in the psychrotolerant affairs, Jaakko Puhakka for a couple of
creative discussions and Anna Kakksonen for FISHy guidelines which were actually quite
precise, my teachers from Odessa Mechnikov University Ivanitsa V., Elinskaya N. and
Vasilyeva T.. Thanks to Sven Bergström who sorted out my supervisors and sorted out me
when it was needed; to my favourite dealer Johnny with his hit “ethanol for dates, methanol
for blind dates”; “catch me if you can” guy Marek Wilczynski –for all reinstalling and total
upgrading; our administrators Marita and Berit who despite their fragile appearance are real
batmen in the tuff world of papers, numbers and signatures; Gunilla Jäger for eternal fight
with toxic chemicals. Marionella, gracias, por la limpieza y por todo! Thanks to media
department and especially to Agneta for all the important work they do. My best wishes to
the office inhabitants of different periods Arunas, Dafne and Karina. Karrrinita, your
vitality and sense of humor kept me going when I was a big ball with short legs (kept me
rolling I meant). Thanks, to the helpful and jolly neighbours Eleonore, Lisandro, Linda,
Odil, Joanna and Teresa, transcriptionally regulated by Vicky Shingler. Big hug to my
teaching partners from whom I learned a great deal of molecular genetics, e.g. single malt is a

54
good stuff (Jocke), boiled cucumbers taste very good - no kidding (Bo) and that if you are
tiered of sport it is better to do some other kind of sport (Anders). Hello to Chenito,
Changchung and Tiago. It is important to discuss here my most pleasant membership in
THE JOURNAL CLUB. Thanks to BEU and SNW for the leadership, to Annika and Jorgen
for funny presentations, to Carlos for being dangerously good looking, to Klaudia and Anna
for “primer extension intensive course”, to Jurate for being a friend anyway, to Stina for
blodomloppet, to Ingve and Berit for smart advice, to Edmund for having his critical opinion
on everything, to Kaloris for quiet enthusiasm, to Barbro for loud enthusiasm, to Connie for
tricky hair styling, to Soni for a beautiful smile, to Hyun-Sook, Tianyan and others for
questions, suggestions, interesting cakes and tasty papers. Thanks to the guys who stir the
social life and organise non scientific activities in the department: Jenny, Christer, Lelle,
Ignas, Sara, Maria, Pär, Mike and Betty; to Gosia for intelligent jokes and Monika
Vestling for the vivid descriptions of labour and breast feeding; to Kristoffer Brännström
for introduction to HPLC and to Per for corridor attacks. Many thanks to Marija Pinne for
sharing so many things and skiing (snowboarding) trips. Hej to the people from up above,
Mazan, Margaret, Olga, Gautam, Uwe and Tobias Hainzl. Tobias, may live our crystal
venture! Regards to fellows from the other side, Gerhard for his wine and power drive in
everything, his students “lazy bone” Marcus and sunny Marco Sani, Maribel for Spanish
training and crazy squash, Ana Virel for the cute three language cocktaile and MEGA help,
my first lab-mates in Umeå Mantas and Vovka and to Alvaro for unforgettable trip to
Castellon. Thanks to the Russian gang, Elena, Asia, Zhenia and Misha for the beautiful
sounds of the mother toungue; to Darius for birdwatching; to my ex room-mates Lijana and
Aurelija ( we don’t live together anymore but you always have one room each in my heart);
to my just exs, Burkhard and Denis - it was nice to meet you guys; to Frederick, the boy-
friend of Lijana (who used to be my room-mate) and a perfect cook; I thank my sister Anna
and my friends Valentina and Jana who lent me some money for a flight to Umeå otherwise
I’d had to walk here.
My parents should be acknowledged (Mila and Ivan) cause they are my parents☺ and
probably it was them who planted the affection for biology in my head (it was a sheer brain
washing!). Finally thanks to Patrik for his love, for letting me love him and for the best
present I ever got in my life – our son Sasha. And to Patrik’s parents of course for all this
terrific food and babysitting!
To all people who read this I wish good health, true love and a lot of scientific
achivements!

55
6. REFERENCES
Aguilera, A., Manrubia, S. C., Gomez, F., Rodriguez, N. & Amils, R. (2006). Eukaryotic community distribution and its relationship to water physicochemical parameters in an extreme acidic environment, Rio Tinto (southwestern Spain). Appl Environ Microbiol 72, 5325-5330. Amaral Zettler, L. A., Messerli, M. A., Laatsch, A. D., Smith, P. J. & Sogin, M. L. (2003). From genes to genomes: beyond biodiversity in Spain's Rio Tinto. Biol Bull 204, 205-209. Anonymous (1994). Design and evaluation of tailings dams. Washington, DC: U.S. Environmental Protection Agency, Office of Solid Waste Special Waste Branch. Appia-Ayme, C., Guiliani, N., Ratouchniak, J. & Bonnefoy, V. (1999). Characterization of an operon encoding two c-type cytochromes, an aa3-type cytochrome oxidase, and rusticyanin in Thiobacillus ferrooxidans ATCC 33020. Appl Environ Microbiol 65, 4781-4787. Appia-Ayme, C., Quatrini, R., Denis, Y. & other authors (2006). Microarray and bioinformatic analyses suggest models for carbon metabolism in the autotroph Acidithiobacillus ferrooxidans. Hydrometallurgy 83, 273-280. Atkins, P. & de Paula, J. (2006). Elements of physical chemistry: Oxford University press. Baker, B. J. & Banfield, J. F. (2003). Microbial communities in acid mine drainage. FEMS Microbiol Ecol 44, 139-152. Baker, B. J., Lutz, M. A., Dawson, S. C., Bond, P. L. & Banfield, J. F. (2004). Metabolically active eukaryotic communities in extremely acidic mine drainage. Appl Environ Microbiol 70, 6264-6271. Baker, R. T. M., Handy, R. D., Davies, S. J. & Snook, J. C. (1998). Chronic dietary exposure to copper affects growth, tissue lipid peroxidation, and metal composition of the grey mullet, Chelon labrosus. Mar Environ Res 45, 357-365. Battaglia-Brunet, F., Joulian, C., Garrido, F., Dictor, M. C., Morin, D., Coupland, K., Johnson, D. B., Hallberg, K. B. & Baranger, P. (2006). Oxidation of arsenite by Thiomonas strains and characterization of Thiomonas arsenivorans sp nov. Anton Leeuw Int J G 89, 99-108. Bengrine, A., Guiliani, N., Appia-Ayme, C., Jedlicki, E., Holmes, D. S., Chippaux, M. & Bonnefoy, V. (1998). Sequence and expression of the rusticyanin structural gene from Thiobacillus ferrooxidans ATCC33020 strain. BBA-Gene Struct Expr 1443, 99-112.

56
Bergamo, R. F., Novo, M. T., Verissimo, R. V. & other authors (2004). Differentiation of Acidithiobacillus ferrooxidans and Acidithiobacillus thiooxidans strains based on 16S-23S rDNA spacer polymorphism analysis. Res Microbiol 155, 559-567. Bjelkevik, A. (2005). Water cover closure design for tailings dams. Luleå: Luleå University of Technology, Department of Civil and Environmental Engineering, Division of Geotechnology. Bond, P. L., Druschel, G. K. & Banfield, J. F. (2000). Comparison of acid mine drainage microbial communities in physically and geochemically distinct ecosystems. Appl Environ Microb 66, 4962-4971. Brasseur, G., Bruscella, P., Bonnefoy, V. & Lemesle-Meunier, D. (2002). The bc1 complex of the iron-grown acidophilic chemolithotrophic bacterium Acidithiobacillus ferrooxidans functions in the reverse but not in the forward direction. Is there a second bc1 complex? Biochim Biophys Acta 1555, 37-43. Brasseur, G., Levican, G., Bonnefoy, V., Holmes, D., Jedlicki, E. & Lemesle-Meunier, D. (2004a). Apparent redundancy of electron transfer pathways to oxygen in the extremophilic chemolithautotrophic bacterium Acidithiobacillus ferrooxidans. BBA-Bioenergetics 1658, 247-247. Brasseur, G., Levican, G., Bonnefoy, V., Holmes, D., Jedlicki, E. & Lemesle-Meunier, D. (2004b). Apparent redundancy of electron transfer pathways via bc1 complexes and terminal oxidases in the extremophilic chemolithoautotrophic Acidithiobacillus ferrooxidans. BBA-Bioenergetics 1656, 114-126. Bruscella, P., Cassagnaud, L., Ratouchniak, J., Brasseur, G., Lojou, E., Amils, R. & Bonnefoy, V. (2005). The HiPIP from the acidophilic Acidithiobacillus ferrooxidans is correctly processed and translocated in Escherichia coli, in spite of the periplasm pH difference between these two micro-organisms. Microbiology 151, 1421-1431. Bruscella, P., Appia-Ayme, C., Levican, G., Ratouchniak, J., Jedlicki, E., Holmes, D. S. & Bonnefoy, V. (2007). Differential expression of two bc1 complexes in the strict acidophilic chemolithoautotrophic bacterium Acidithiobacillus ferrooxidans suggests a model for their respective roles in iron or sulfur oxidation. Microbiology 153, 102-110. Bryant, R. D., Costerton, J. W. & Laishley, E. J. (1984). The role of Thiobacillus albertis glycocalyx in the adhesion of cells to elemental sulfur. Can J Microbiol 30, 81-90. Bugaytsova, Z. & Lindström, E. B. (2004). Localization, purification and properties of a tetrathionate hydrolase from Acidithiobacillus caldus. Eur J Biochem 271, 272-280.

57
Canovas, C. R., Olias, M., Nieto, J. M., Sarmiento, A. M. & Ceron, J. C. (2007). Hydrogeochemical characteristics of the Tinto and Odiel Rivers (SW Spain). Factors controlling metal contents. Sci Total Environ 373, 363-382. Cavazza, C., Guigliarelli, B., Bertrand, P. & Bruschi, M. (1995). Biochemical and EPR characterization of a high-potential iron-sulfur protein in Thiobacillus ferrooxidans. FEMS Microbiol Lett 130, 193-199. Colmer, A. R. & Hinkle, M. E. (1947). The role of microorganisms in acid mine drainage: A preliminary report. Science 106, 253-256. Coram, N. J. & Rawlings, D. E. (2002). Molecular relationship between two groups of the genus Leptospirillum and the finding that Leptospirillum ferriphilum sp nov dominates South African commercial biooxidation tanks that operate at 40°C. Appl Environ Microb 68, 838-845. Damek-Poprawa, M. & Sawicka-Kapusta, K. (2003). Damage to the liver, kidney, and testis with reference to burden of heavy metals in yellow-necked mice from areas around steelworks and zinc smelters in Poland. Toxicology 186, 1-10. de Groot, P., Deane, S. M. & Rawlings, D. E. (2003). A transposon-located arsenic resistance mechanism from a strain of Acidithiobacillus caldus isolated from commercial, arsenopyrite biooxidation tanks. Hydrometallurgy 71, 115-123. de Jong, G. A. H., Hazeu, W., Bos, P. & Kuenen, J. G. (1997). Polythionate degradation by tetrathionate hydrolase of Thiobacillus ferrooxidans. Microbiology-UK 143, 499-504. Demchak, J., Skousen, J. & McDonald, L. M. (2004). Longevity of acid discharges from underground mines located above the regional water table. J Environ Qual 33, 656-668. Do Carmo, O. A., Lima, V. M. & Guimaraes, S. M. R. (2001). BIOX® Process - the Sao Bento experience. In International Biohydrometallurgy Symposium, pp. 81-86. Edited by V. S. T. Ciminelli & O. Garcia. Ouro Preto: Elsevier. Dold, B., Blowes, D. W., Dickhout, R., Spangenberg, J. E. & Pfeifer, H. R. (2005). Low molecular weight carboxylic acids in oxidizing porphyry copper tailings. Environ Sci Technol 39, 2515-2521. Dopson, M. & Lindström, E. B. (1999). Potential role of Thiobacillus caldus in arsenopyrite bioleaching. Appl Environ Microbiol 65, 36-40. Dopson, M., Lindström, E. B. & Hallberg, K. B. (2001). Chromosomally encoded arsenical resistance of the moderately thermophilic acidophile Acidithiobacillus caldus. Extremophiles 5, 247-255.

58
Dopson, M., Lindström, E. B. & Hallberg, K. B. (2002). ATP generation during reduced inorganic sulfur compound oxidation by Acidithiobacillus caldus is exclusively due to electron transport phosphorylation. Extremophiles 6, 123-129. Dopson, M., Sundkvist, J. E. & Lindström, B. (2006). Toxicity of metal extraction and flotation chemicals to Sulfolobus metallicus and chalcopyrite bioleaching. Hydrometallurgy 81, 205-213. Dopson, M., Halinen, A. K., Rahunen, N., Ozkaya, B., Sahinkaya, E., Kaksonen, A. H., Lindström, E. B. & Puhakka, J. A. (2007). Mineral and iron oxidation at low temperatures by pure and mixed cultures of acidophilic microorganisms. Biotechnol Bioeng 97, 1205-1215. Drobner, E., Huber, H. & Stetter, K. O. (1990). Thiobacillus ferrooxidans, a facultative hydrogen oxidizer. Appl Environ Microb 56, 2922-2923. Dvorak, D. H., Hedin, R. S., Edenborn, H. M. & Mcintire, P. E. (1992). Treatment of metal-contaminated water using bacterial sulfate reduction - results from pilot-scale reactors. Biotechnol Bioengin 40, 609-616. Eccleston, M. & Kelly, D. P. (1978). Oxidation kinetics and chemostat growth kinetics of Thiobacillus ferrooxidans on tetrathionate and thiosulfate. J Bacteriol 134, 718-727. Edwards, K. J., Schrenk, M. O., Hamers, R. & Banfield, J. F. (1998). Microbial oxidation of pyrite: Experiments using microorganisms from an extreme acidic environment. Am Mineral 83, 1444-1453. Edwards, K. J., Gihring, T. M. & Banfield, J. F. (1999a). Seasonal variations in microbial populations and environmental conditions in an extreme acid mine drainage environment. Appl Environ Microb 65, 3627-3632. Edwards, K. J., Goebel, B. M., Rodgers, T. M. & other authors (1999b). Geomicrobiology of pyrite (FeS2) dissolution: Case study at Iron Mountain, California. Geomicrobiol J 16, 155-179. Escobar, B., Huerta, G. & Rubio, J. (1997). Influence of lipopolysaccharides on the attachment of Thiobacillus ferrooxidans to minerals. World J Microb Biotechnol 13, 593-594. Fowler, T. A. & Crundwell, F. K. (1999). Leaching of zinc sulfide by Thiobacillus ferrooxidans: Bacterial oxidation of the sulfur product layer increases the rate of zinc sulfide dissolution at high concentrations of ferrous ions. Appl Environ Microb 65, 5285-5292. Friedrich, C. G., Bardischewsky, F., Rother, D., Quentmeier, A. & Fischer, J. (2005). Prokaryotic sulfur oxidation. Curr Opin Microbiol 8, 253-259.

59
Gadanho, M., Libkind, D. & Sampaio, J. P. (2006). Yeast diversity in the extreme acidic environments of the Iberian Pyrite Belt. Microbial Ecology 52, 552-563. Gatzweiler, R., Jahn, S., Neubert, G. & Paul, M. (2001). Cover design for radioactive and AMD-producing mine waste in the Ronneburg area, Eastern Thuringia. Waste Manage 21, 175-184. Gehrke, T., Telegdi, J., Thierry, D. & Sand, W. (1998). Importance of extracellular polymeric substances from Thiobacillus ferrooxidans for bioleaching. Appl Environ Microbiol 64, 2743-2747. Gheorghiu, C., Cable, J., Marcogliese, D. J. & Scott, M. E. (2007). Effects of waterborne zinc on reproduction, survival and morphometrics of Gyrodactylus turnbulli (Monogenea) on guppies (Poecilia reticulata). Int J Parasitol 37, 375-381. Giudici-Orticoni, M. T., Leroy, G., Nitschke, W. & Bruschi, M. (2000). Characterization of a new dihemic c4-type cytochrome isolated from Thiobacillus ferrooxidans. Biochemistry-USA 39, 7205-7211. Gonzalez-Toril, E., Llobet-Brossa, E., Casamayor, E. O., Amann, R. & Amils, R. (2003). Microbial ecology of an extreme acidic environment, the Tinto River. Appl Environ Microbiol 69, 4853-4865. Groudev, S. N., Spasova, I. I., Komnitsas, K. & Paspaliaris, I. (2001). Microbial heap leaching of copper and gold from a sulphide ore. In International Biohydrometallurgy Symposium. Edited by V. S. T. Ciminelli & O. Garcia. Ouro Preto: Elsevier. Hallberg, K. B. & Lindström, E. B. (1994). Characterization of Thiobacillus caldus sp. nov., a moderately thermophilic acidophile. Microbiology 140, 3451-3456. Hallberg, K. B., Dopson, M. & Lindström, E. B. (1996). Reduced sulfur compound oxidation by Thiobacillus caldus. J Bacteriol 178, 6-11. Hallmann, R., Friedrich, A., Koops, H. P., Pommereningroser, A., Rohde, K., Zenneck, C. & Sand, W. (1992). Physiological characteristics of Thiobacillus ferrooxidans and Leptospirillum ferrooxidans and physicochemical factors influence microbial metal meaching. Geomicrobiol J 10, 193-206. Handy, R. D. (2003). Chronic effects of copper exposure versus endocrine toxicity: Two sides of the same toxicological process? Comp Biochem Phys A 135, 25-38.

60
Harvey, J. T. & Bath, M. (2007). The GeoBiotics GEOCOAT® technology - progress and chalanges. In Biomining, pp. 97-112. Edited by D. E. Rawlings & B. D. Johnson. Berlin Heidelberg New York: Springer. Henry, T. B., Irwin, E. R., Grizzle, J. M., Wildhaber, M. L. & Brumbaugh, W. G. (1999). Acute toxicity of an acid mine drainage mixing zone to juvenile bluegill and largemouth bass. T Am Fish Soc 128, 919-928. Henry, T. B., Irwin, E. R., Grizzle, J. M., Brumbaugh, W. G. & Wildhaber, M. L. (2001). Gill lesions and death of bluegill in an acid mine drainage mixing zone. Environ Toxicol Chem 20, 1304-1311. Herlihy, A. T. & Mills, A. L. (1985). Sulfate reduction in freshwater sediments receiving acid mine drainage. Appl Environ Microbiol 49, 179-186. Holmes, D. S., Zhao, H. L., Levican, G., Ratouchniak, J., Bonnefoy, V., Varela, P. & Jedlicki, E. (2001). ISAfe1, an ISL3 family insertion sequence from Acidithiobacillus ferrooxidans ATCC 19859. J Bacteriol 183, 4323-4329. Holmes, D. S. & Bonnefoy, V. (2007). Genetic and bioinformatic insights into iron and sulfur oxidation mechanisms of bioleaching organisms. In Biomining, pp. 281-307. Edited by D. E. Rawlings & B. D. Johnson. Berlin Heidelberg: Springer. Horvath, D. J. (1985). Consumption of simulated acid mine water by sheep. J Anim Sci 61, 474-479. Hugenholtz, P., Goebel, B. M. & Pace, N. R. (1998). Impact of culture-independent studies on the emerging phylogenetic view of bacterial diversity. J Bacteriol 180, 6793-6793. Ingledew, W. J. (1982). Thiobacillus ferrooxidans. The bioenergetics of an acidophilic chemolithotroph. Biochim Biophys Acta 683, 89-117. Jianhua, Z., Lian, X., Shuanlai, Z., Juan, D. & Shuanxi, Y. (2006). DNA lesion and Hprt mutant frequency in rat lymphocytes and V79 Chinese hamster lung cells exposed to cadmium. J Occup Health 48, 93-99. Jin, S., Fallgren, H. P., Morris, M. J. & Cooper, S. J. (2007). Source treatment of acid mine drainage pollution at Backfilled coal mine using remote sencing and biogeochemistry. Water Air Soil Pollut 188, 205-212. Jin, S. M., Yan, W. M. & Wang, Z. N. (1992). Transfer of IncP plasmids to extremely acidophilic Thiobacillus thiooxidans. Appl Environ Microbiol 58, 429-430.

61
Johnson, D. B., Ghauri, M. A. & Said, M. F. (1992). Isolation and characterization of an acidophilic, heterotrophic bacterium capable of oxidizing ferrous iron. Appl Environ Microbiol 58, 1423-1428. Johnson, D. B. (2001). Importance of microbial ecology in the development of new mineral technologies. Hydrometallurgy 59, 147-157. Johnson, D. B., Rolfe, S., Hallberg, K. B. & Iversen, E. (2001). Isolation and phylogenetic characterization of acidophilic microorganisms indigenous to acidic drainage waters at an abandoned Norwegian copper mine. Environ Microbiol 3, 630-637. Johnson, D. B. & Hallberg, K. B. (2005). Acid mine drainage remediation options: A review. Sci Tot Environ 338, 3-14. Johnson, D. B., Yajie, L. & Okibe, N. (2008). "Bioshrouding"- a novel approach for securing reactive mineral tailings. Biotechnol Lett 30, 445-449. Kamimura, K., Higashino, E., Moriya, S. & Sugio, T. (2003). Marine acidophilic sulfur-oxidizing bacterium requiring salts for the oxidation of reduced inorganic sulfur compounds. Extremophiles 7, 95-99. Kanao, T., Kamimura, K. & Sugio, T. (2007). Identification of a gene encoding a tetrathionate hydrolase in Acidithiobacillus ferrooxidans. J Biotechnol 132, 16-22. Karavaiko, G. I., R.S., G., Pivovarova, T. A., I.A., T. & S.N., V. (1987). Thermophilic bacteria of the genus Sulfobacillus. In Biohydrometalurgy, pp. 29-42. Edited by P. R. Norris & D. P. Kelly. Warwick: Antony Rowe Ltd. Karavaiko, G. I., Turova, T. P., Kondrat'eva, T. F., Lysenko, A. M., Kolganova, T. V., Ageeva, S. N., Muntyan, L. N. & Pivovarova, T. A. (2003). Phylogenetic heterogeneity of the species Acidithiobacillus ferrooxidans. Int J Syst Evol Microbiol 53, 113-119. Kelly, D. P., Cambers, L. A. & Trudinger, P. A. (1969). Cyanolysis and spectrophotometric estimation of trithionate in mixture with thiosulfate and tetrathionate. Anal Chem 41, 898-902. Kelly, D. P. & Wood, A. P. (2000). Reclassification of some species of Thiobacillus to the newly designated genera Acidithiobacillus gen. nov., Halothiobacillus gen. nov. and Thermithiobacillus gen. nov. Int J Syst Evol Microbiol 50, 511-516. Kelly, D. P. H. A. H. (1989). Genus Thiobacillus. In Bergey's Manual of Systematic Bacteriology, pp. 1842-1858. Edited by M. P. B. J.T. Staley, N. Pfennig & J.G. Holt. Baltimore: Williams & Wilkins.

62
Koschorreck, M., Bozau, E., Frommichen, R., Geller, W., Herzsprung, P. & Wendt-Potthoff, K. (2007). Processes at the sediment water interface after addition of organic matter and lime to an Acid Mine Pit Lake mesocosm. Environ Sci Technol 41, 1608-1614. Kupka, D., Rzhepishevska, O. I., Dopson, M., Lindström, E. B., Karnachuk, O. V. & Tuovinen, O. H. (2007). Bacterial oxidation of ferrous iron at low temperatures. Biotechnol Bioengin 97, 1470-1478. Kusano, T., Takeshima, T., Inoue, C. & Sugawara, K. (1991). Evidence for 2 sets of structural genes-coding for ribulose bisphosphate carboxylase in Thiobacillus ferrooxidans. J Bacteriol 173, 7313-7323. Kusano, T., Sugawara, K., Inoue, C., Takeshima, T., Numata, M. & Shiratori, T. (1992). Electrotransformation of Thiobacillus ferrooxidans with plasmids containing a mer determinant. J Bacteriol 174, 6617-6623. Leduc, L. G., Trevors, J. T. & Ferroni, G. D. (1993). Thermal characterization of different isolates of Thiobacillus ferrooxidans. FEMS Microbiol Lett 108, 189-193. Lefebvre, R., Hockley, D., Smolensky, J. & Gelinas, P. (2001a). Multiphase transfer processes in waste rock piles producing acid mine drainage 1: Conceptual model and system characterization. J Contam Hydrol 52, 137-164. Lefebvre, R., Hockley, D., Smolensky, J. & Lamontagne, A. (2001b). Multiphase transfer processes in waste rock piles producing acid mine drainage 2: Applications of numerical simulation. J Contam Hydrol 52, 165-186. Leveille, S. A., Leduc, L. G., Ferroni, G. D., Telang, A. J. & Voordouw, G. (2001). Monitoring of bacteria in acid mine environments by reverse sample genome probing. Can J Microbiol 47, 431-442. Levican, G., Bruscella, P., Guacunano, M., Inostroza, C., Bonnefoy, V., Holmes, D. S. & Jedlicki, E. (2002). Characterization of the petI and res operons of Acidithiobacillus ferrooxidans. J Bacteriol 184, 1498-1501. Lilova, K. & Karamanev, D. (2005). Direct oxidation of copper sulfide by a biofilm of Acidithiobacillus ferrooxidans. Hydrometallurgy 80, 147-154. Liu, X., Lin, J., Zhang, Z., Bian, J., Zhao, Q., Liu, Y. & Yan, W. (2007). Construction of conjugative gene transfer system between E. coli and moderately thermophilic, extremely acidophilic Acidithiobacillus caldus MTH-04. J Microbiol Biotechnol 17, 162-167.

63
Liu, Z., Guiliani, N., Appia-Ayme, C., Borne, F., Ratouchniak, J. & Bonnefoy, V. (2000). Construction and characterization of a recA mutant of Thiobacillus ferrooxidans by marker exchange mutagenesis. J Bacteriol 182, 2269-2276. Logan, C. T., Seal, T. & Brierley, J. A. (2007). Whole-ore heap biooxidation of sulfidic gold-bearing ores. In Biomining, pp. 113-137. Edited by D. E. Rawlings & B. D. Johnson. Berlin Heidelberg New York: Springer. Madigan, M. T. & Martinko, J. M. (2006). Brock Biology of Microorganisms. Upper Saddle River: Pearson Education, Inc. Maeda, T., Negishi, A., Komoto, H., Oshima, Y., Kamimura, K. & Sugio, T. (1999). Isolation of iron-oxidizing bacteria from corroded concretes of sewage treatment plants. J Biosci Bioeng 88, 300-305. Malarte, G., Leroy, G., Lojou, E., Abergel, C., Bruschi, M. & Giudici-Orticoni, M. T. (2005). Insight into molecular stability and physiological properties of the diheme cytochrome CYC41 from the acidophilic bacterium Acidithiobacillus ferrooxidans. Biochemistry-US 44, 6471-6481. Mangold, S., Harneit, K., Rohwerder, T., Claus, G. & Sand, W. (2008). Novel combination of atomic force microscopy and epifluorescence microscopy for visualization of leaching bacteria on pyrite. Appl Environ Microbiol 74, 410-415. Marsden, A. D. & DeWreede, R. E. (2000). Marine macroalgal community structure, metal content and reproductive function near an acid mine drainage outflow. Environ Pollut 110, 431-440. Masau, R. J., Oh, J. K. & Suzuki, I. (2001). Mechanism of oxidation of inorganic sulfur compounds by thiosulfate-grown Thiobacillus thiooxidans. Can J Microbiol 47, 348-358. McCready, R. G. L. (1987). Progress in the bacterial leaching of metals in Canada. In Biohydrometallurgy. Edited by P. R. Norris & D. P. Kelly. Warwick: Antony Rowe Ltd. Meldrum, J. L., Jamieson, H. E. & Dyke, L. D. (2001). Oxidation of mine tailings from Rankin Inlet, Nunavut, at subzero temperatures. Can Geotech J 38, 957-966. Mielke, R. E., Pace, D. L., Porter, T. & Southam, G. (2003). A critical stage in the formation of acid mine drainage: Colonization of pyrite by Acidithiobacillus ferrooxidans under pH-neutral conditions. Geobiology 1, 81-90. Mikkelsen, D., Kappler, U., McEwan, A. G. & Sly, L. I. (2006). Archaeal diversity in two thermophilic chalcopyrite bioleaching reactors. Environ Microbiol 8, 2050-2055.

64
Mishra, K. A., Roy, P. & Mahapatra, R. S. S. (1983). Isolation of Thiobacillus ferrooxidans from various habitats and their growth patter on solid medium. Curr Microbiol 8, 147-152. Morin, D. H. R. (2007). Bioleaching of sulfide minerals in continuous stirred tanks. In Microbial processing of metal sulfides. Edited by E. R. Donati & W. Sand. Dordrecht: Springer. Mottana, A., R. Crespi, R., and Liborio, G. (1983). The Macdonald Encyclopedia of Rocks and Minerals. London: Macdonald & Co, London & Sydney. Muller, F. H., Bandeiras, T. M., Urich, T., Teixeira, M., Gomes, C. M. & Kletzin, A. (2004). Coupling of the pathway of sulphur oxidation to dioxygen reduction: characterization of a novel membrane-bound thiosulphate:quinone oxidoreductase. Mol Microbiol 53, 1147-1160. Nakamura, K., Kuribayashi, S., Kurosawa, H. & Amano, Y. (1992). Pathway of sulfite oxidation in Thiobacillus thiooxidans JCM7814. Biosci Biotech Bioch 56, 261-263. Nakamura, K., Yoshikawa, H., Okubo, S., Kurosawa, H. & Amano, Y. (1995). Purification and properties of membrane-bound sulfite dehydrogenase from Thiobacillus thiooxidans JCM7814. Biosci Biotech Bioch 59, 11-15. Nakamura, K., Nakamura, M., Yoshikawa, H. & Amano, Y. (2001). Purification and properties of thiosulfate dehydrogenase from Acidithiobacillus thiooxidans JCM7814. Biosci Biotechnol Biochem 65, 102-108. Neculita, C. M., Zagury, G. J. & Bussiere, B. (2007). Passive treatment of acid mine drainage in bioreactors using sulfate-reducing bacteria: critical review and research needs. J Environ Qual 36, 1-16. Nicomrat, D., Dick, W. A. & Tuovinen, O. H. (2006a). Microbial populations identified by fluorescence in situ hybridization in a constructed wetland treating acid coal mine drainage. J Environ Qual 35, 1329-1337. Nicomrat, D., Dick, W. A. & Tuovinen, O. H. (2006b). Assessment of the microbial community in a constructed wetland that receives acid coal mine drainage. Microbial Ecol 51, 83-89. Nishio, S., Aoki, K. & Yokoyama, M. (2003). A new method using a bacterium for dissolution of urinary stones. Aktuelle Urol 34, 253-255.

65
Nordstrom, D. K. & Alpers, C. N. (1999). Negative pH, efflorescent mineralogy, and consequences for environmental restoration at the Iron Mountain Superfund site, California. Proc Natl Acad Sci USA 96, 3455-3462. Nordstrom, D. K., Alpers, C. N., Ptacek, C. J. & Blowes, D. W. (2000). Negative pH and extremely acidic mine waters from Iron Mountain, California. Environ Sci Technol 34, 254-258. Norris, P. R., Burton, N. P. & Foulis, N. A. M. (2000). Acidophiles in bioreactor mineral processing. Extremophiles 4, 71-76. Norris, P. R. (2007). Acidophile diversity in mineral sulfide oxidation. In Biomining, pp. 199-212. Edited by D. E. Rawlings & B. D. Johnson: Springer. O'Halloran, K., Cavanagh, J. A. & Harding, J. (2008). Response of a New Zealand mayfly (Deleatidium spp.) to acid mine drainage: Implications for mine remediation. Environ Toxicol Chem, 1. Ohmura, N., Sasaki, K., Matsumoto, N. & Saiki, H. (2002). Anaerobic respiration using Fe3+, S0, and H2 in the chemolithoautotrophic bacterium Acidithiobacillus ferrooxidans. J Bacteriol 184, 2081-2087. Okibe, N., Gericke, M., Hallberg, K. B. & Johnson, D. B. (2003). Enumeration and characterization of acidophilic microorganisms isolated from a pilot plant stirred-tank bioleaching operation. Appl Environ Microb 69, 1936-1943. Okibe, N. & Johnson, D. B. (2004). Biooxidation of pyrite by defined mixed cultures of moderately thermophilic acidophiles in pH-controlled bioreactors: Significance of microbial interactions. Biotechnol Bioengin 87, 574-583. Olias, M., Nieto, J. M., Sarmiento, A. M., Ceron, J. C. & Canovas, C. R. (2004). Seasonal water quality variations in a river affected by acid mine drainage: the Odiel River (South West Spain). Sci Total Environ 333, 267-281. Olias, M., Ceron, J. C., Moral, F. & Ruiz, F. (2006). Water quality of the Guadiamar River after the Aznalcollar spill (SW Spain). Chemosphere 62, 213-225. Paulino, L. C., Bergamo, R. F., Garcia, O., Jr., de Mello, M. P., Manfio, G. P. & Ottoboni, L. M. (2001). Molecular characterization of Acidithiobacillus ferrooxidans and Acidithiobacillus thiooxidans strains isolated from mine wastes in Brazil. Anton Van Leeuwen 80, 65-75. Peng, J. B., Yan, W. M. & Bao, X. Z. (1994). Plasmid and transposon transfer to Thiobacillus ferrooxidans. J Bacteriol 176, 2892-2897.

66
Petersen, J. & Dixon, D. G. (2007). Modeling and optimization of heap bioleach processes. In Biomining, pp. 281-307. Edited by D. E. Rawlings & B. D. Johnson. Berlin Heidelberg New York: Springer. Pronk, J. T., Liem, K., Bos, P. & Kuenen, J. G. (1991a). Energy transduction by anaerobic ferric iron respiration in Thiobacillus ferrooxidans. Appl Environ Microbiol 57, 2063-2068. Pronk, J. T., Meijer, W. M., Hazeu, W., van Dijken, J. P., Bos, P. & Kuenen, J. G. (1991b). Growth of Thiobacillus ferrooxidans on formic acid. Appl Environ Microbiol 57, 2057-2062. Pronk, J. T., Debruyn, J. C., Bos, P. & Kuenen, J. G. (1992). Anaerobic growth of Thiobacillus ferrooxidans. Appl Environ Microb 58, 2227-2230. Puhakka, A. J., Kaksonen, A. H. & Riekkola-Vanhanen, M. (2007). Heap leaching of black schist. In Biomining, pp. 139-150. Edited by D. E. Rawlings & B. D. Johnson. Berlin Heidelberg New York: Springer. Quatrini, R., Lefimil, C., Holmes, D. S. & Jedlicki, E. (2005). The ferric iron uptake regulator (Fur) from the extreme acidophile Acidithiobacillus ferrooxidans. Microbiology-UK 151, 2005-2015. Quatrini, R., Appia-Ayme, C., Denis, Y. & other authors (2006). Insights into the iron and sulfur energetic metabolism of Acidithiobacillus ferrooxidans by microarray transcriptome profiling. Hydrometallurgy 83, 263-272. Ramirez, P., Guiliani, N., Valenzuela, L., Beard, S. & Jerez, C. A. (2004). Differential protein expression during growth of Acidithiobacillus ferrooxidans on ferrous iron, sulfur compounds, or metal sulfides. Appl Environ Microb 70, 4491-4498. Rawlings, D. E., Tributsch, H. & Hansford, G. S. (1999). Reasons why 'Leptospirillum'-like species rather than Thiobacillus ferrooxidans are the dominant iron-oxidizing bacteria in many commercial processes for the biooxidation of pyrite and related ores. Microbiology 145, 5-13. Rawlings, D. E. (2002). Heavy metal mining using microbes. Annu Rev Microbiol 56, 65-91. Rawlings, D. E. (2005). Characteristics and adaptability of iron- and sulfur-oxidizing microorganisms used for the recovery of metals from minerals and their concentrates. Microb Cell Fact 4, 13. Rawlings, D. E. & Johnson, D. B. (2007). The microbiology of biomining: development and optimization of mineral-oxidizing microbial consortia. Microbiology 153, 315-324.

67
Robertson, L. A. & Kuenen, J. K. (1992). The colorless sulphur bacteria. In The Prokaryotes, pp. 385-431. Edited by A. Balows, Trueper, H.G., Workin M.D., Hareder, W. and Schleifer K.H.: springer-Verlag. Rohwerder, T. & Sand, W. (2003). The sulfane sulfur of persulfides is the actual substrate of the sulfur-oxidizing enzymes from Acidithiobacillus and Acidiphilium spp. Microbiology 149, 1699-1710. Rojas, J., Giersig, M. & Tributsch, H. (1995). Sulfur colloids as temporary energy reservoirs for Thiobacillus ferrooxidans during pyrite oxidation. Arch Microbiol 163, 352-356. Rojas-Chapana, J. A. & Tributsch, H. (2004). Interfacial activity and leaching patterns of Leptospirillum ferrooxidans on pyrite. FEMS Microbiol Ecol 47, 19-29. Rossi, G. (1990). Biohydrometallurgy. Hamburg New York: McGraw-Hill. Rossi, G. (2001). The design of bioreactors. Hydrometallurgy 59, 217-231. Rowe, O. F., Sanchez-Espana, J., Hallberg, K. B. & Johnson, D. B. (2007). Microbial communities and geochemical dynamics in an extremely acidic, metal-rich stream at an abandoned sulfide mine (Huelva, Spain) underpinned by two functional primary production systems. Environ Microbiol 9, 1761-1771. Sand, W. (1987). Importance of hydrogen sulfide, thiosulfate, and methylmercaptan for growth of thiobacilli during simulation of concrete corrosion. Appl Environ Microbiol 53, 1645-1648. Sand, W., Gehrke, T., Jozsa, P. G. & Schippers, A. (2001). (Bio) chemistry of bacterial leaching - direct vs. indirect bioleaching. Hydrometallurgy 59, 159-175. Sand, W. & Gehrke, T. (2006). Extracellular polymeric substances mediate bioleaching/biocorrosion via interfacial processes involving iron(III) ions and acidophilic bacteria. Res Microbiol 157, 49-56. Sasaki, K., Ida, C., Ando, A., Matsumoto, N., Saiki, H. & Ohmura, N. (2003). Respiratory isozyme, two types of rusticyanin of Acidithiobacillus ferrooxidans. Biosci Biotech Bioch 67, 1039-1047. Schippers, A., Hallmann, R., Wentzien, S. & Sand, W. (1995). Microbial diversity in uranium-mine waste heaps. Appl Environ Microb 61, 2930-2935.

68
Schippers, A., Jozsa, P. G. & Sand, W. (1996). Sulfur chemistry in bacterial leaching of pyrite. Appl Environ Microbiol 62, 3424-3431. Schippers, A. & Sand, W. (1999). Bacterial leaching of metal sulfides proceeds by two indirect mechanisms via thiosulfate or via polysulfides and sulfur. Appl Environ Microbiol 65, 319-321. Schrenk, M. O., Edwards, K. J., Goodman, R. M., Hamers, R. J. & Banfield, J. F. (1998). Distribution of Thiobacillus ferrooxidans and Leptospirillum ferrooxidans: Implications for generation of acid mine drainage. Science 279, 1519-1522. Silver, M. & Lundgren, D. G. (1968a). The thiosulfate-oxidizing enzyme of Ferrobacillus ferrooxidans (Thiobacillus ferrooxidans). Can J Biochem 46, 1215-1220. Silver, M. & Lundgren, D. G. (1968b). Sulfur-oxidizing enzyme of Ferrobacillus ferrooxidans (Thiobacillus ferrooxidans). Can J Biochem 46, 457-461. Silverman, M. P. & Ehrlich, H. L. (1964). Microbial formation and degradation of minerals. Advan Appl Microbiol 6, 153-206. Silverman, M. P. (1967). Mechanism of bacterial pyrite oxidation. J Bacteriol 94, 1046-1051. Simon, M. L., Cherry, D. S., Currie, R. J. & Zipper, C. E. (2006). The ecotoxicological recovery of Ely Creek and tributaries (Lee County, VA) after remediation of acid mine drainage. Environ Monit Assess 123, 109-124. Singer, P. C. & Stumm, W. (1970). Acidic mine drainage: The rate-determining step. Science 167, 1121-1123. Songrong, Y., Jiyuan, X., Guanzhou, Q. & Yuehua, H. (2002). Research and application of bioleaching and biooxidation technologies in China. Miner Engin 15, 361-363. Steudel, R. (1996). Mechanism for the formation of elemental sulfur from aqueous sulfide in chemical and microbiological desulfurization processes. Ind Eng Chem Res 35, 1417-1423. Sugio, T., Domatsu, C., Munakata, O., Tano, T. & Imai, K. (1985). Role of a ferric ion-reducing system in sulfur oxidation of Thiobacillus ferrooxidans. Appl Environ Microbiol 49, 1401-1406. Sugio, T., Tsujita, Y., Katagiri, T., Inagaki, K. & Tano, T. (1988a). Reduction of Mo6+ with elemental sulfur by Thiobacillus ferrooxidans. J Bacteriol 170, 5956-5959.

69
Sugio, T., Wada, K., Mori, M., Inagaki, K. & Tano, T. (1988b). Synthesis of an iron-oxidizing system during growth of Thiobacillus ferrooxidans on sulfur-basal salts medium. Appl Environ Microbiol 54, 150-152. Sugio, T., Katagiri, T., Inagaki, K. & Tano, T. (1989). Actual substrate for elemental sulfur oxidation by sulfur-ferric ion oxidoreductase purified from Thiobacillus ferrooxidans. Biochim Biophys Acta 973, 250-256. Sugio, T., Hirose, T., Oto, A., Inagaki, K. & Tano, T. (1990a). The regulation of sulfur use by ferrous ion in Thiobacillus ferroxidans. Agr Biol Chem Tokyo 54, 2017-2022. Sugio, T., Tsujita, Y., Inagaki, K. & Tano, T. (1990b). Reduction of cupric ions with elemental sulfur by Thiobacillus ferrooxidans. Appl Environ Microbiol 56, 693-696. Sugio, T., Hirose, T., Ye, L. Z. & Tano, T. (1992a). Purification and some properties of sulfite ferric ion oxidoreductase from Thiobacillus ferrooxidans. J Bacteriol 174, 4189-4192. Sugio, T., White, K. J., Shute, E., Choate, D. & Blake, R. C. (1992b). Existence of a hydrogen sulfide: ferric ion oxidoreductase in iron-oxidizing bacteria. Appl Environ Microbiol 58, 431-433. Sugio, T., Hisazumi, T., Kanao, T., Kamimura, K., Takeuchi, F. & Negishi, A. (2006). Existence of aa3-type ubiquinol oxidase as a terminal oxidase in sulfite oxidation of Acidithiobacillus thiooxidans. Biosci Biotech Bioch 70, 1584-1591. Sugio, T., Taha, T. M., Kanao, T. & Takeuchi, F. (2007). Increase in Fe2+-producing activity during growth of Acidithiobacillus ferrooxidans ATCC23270 on sulfur. Biosci Biotechnol Biochem 71, 2663-2669. Suzuki, I. & Werkman, C. H. (1959). Glutathione and sulfur oxidation by Thiobacillus thiooxidans. Proc Natl Acad Sci U S A 45, 239-244. Suzuki, I. (1965a). Incorporation of atmospheric oxygen-18 into thiosulfate by the sulfur-oxidizing enzyme of Thiobacillus thiooxidans. Biochim Biophys Acta 110, 97-101. Suzuki, I. (1965b). Oxidation of elemental sulfur by an enzyme system of Thiobacillus thiooxidans. Biochim Biophys Acta 104, 359-371. Suzuki, I., Takeuchi, T. L., Yuthasastrakosol, T. D. & Oh, J. K. (1990). Ferrous iron and sulfur oxidation and ferric iron reduction activities of Thiobacillus ferrooxidans are affected by growth on ferrous iron, sulfur, or a sulfide ore. Appl Environ Microbiol 56, 1620-1626. Suzuki, I., Chan, C. W. & Takeuchi, T. L. (1992). Oxidation of elemental sulfur to sulfite by Thiobacillus thiooxidans cells. Appl Environ Microbiol 58, 3767-3769.

70
Tabita, R., Silver, M. & Lundgren, D. G. (1969). The rhodanese enzyme of Ferrobacillus ferrooxidans (Thiobacillus ferrooxidans). Can J Biochem 47, 1141-1145. Tabita, R. & Lundgren, D. G. (1971). Utilization of glucose and the effect of organic compounds on the chemolithotroph Thiobacillus ferrooxidans. J Bacteriol 108, 328-333. Tan, G. L., Shu, W. S., Hallberg, K. B., Li, F., Lan, C. Y. & Huang, L. N. (2007). Cultivation-dependent and cultivation-independent characterization of the microbial community in acid mine drainage associated with acidic Pb/Zn mine tailings at Lechang, Guangdong, China. FEMS Microbiol Ecol 59, 118-126. Tano, T., Kitaguchi, H., Harada, M., Nagasawa, T. & Sugio, T. (1996). Purification and some properties of a tetrathionate decomposing enzyme from Thiobacillus thiooxidans. Biosci Biotechnol Biochem 60, 224-227. Ter Heijne, A., Hamelers, H. V. M. & Buisman, C. J. N. (2007). Microbial fuel cell operation with continuous biological ferrous iron oxidation of the catholyte. Environ Sci Technol 41, 4130-4134. Tian, K. L., Lin, J. Q., Liu, X. M., Liu, Y., Zhang, C. K. & Yan, W. M. (2003). Conversion of an obligate autotrophic bacteria to heterotrophic growth: expression of a heterogeneous phosphofructokinase gene in the chemolithotroph Acidithiobacillus thiooxidans. Biotechnol Lett 25, 749-754. Ting, Y. P., Kumar, A. S., Rahman, M. & Chia, B. K. (2000). Innovative use of Thiobacillus ferrooxidans for the biological machining of metals. Acta Biotechnol 20, 87-96. Torma, A. E. (1985). Biohydrometallurgy as en emerging technology. In Biotechnology & Bioengenireeng Symposium, pp. 49-63. Edited by H. L. Ehrlich & D. S. Holmes. New York: John Wiley & Sons. Tributsch, H. & Bennett, J., C. (1981). Semiconductor-electrochemical aspects of bacterial leaching. I. Oxidation of metal sulphides with large energy gaps. J Chem Technol Biotechnol 31, 565-577. Tributsch, H. (2001). Direct versus indirect bioleaching. Hydrometallurgy 59, 177-185. Tuovinen, O. H. (1985). Acid leaching of uranium ore materials with microbial catalysis. In Biotechnology & Bioengenireeng Symposium, pp. 65-72. Edited by H. L. Ehrlich & D. S. Holmes. New York: John Wiley & Sons.

71
Tuttle, J. H., Randles, C. I. & Dugan, P. R. (1968). Activity of microorganisms in acid mine water. I. Influence of acid water on aerobic heterotrophs of a normal stream. J Bacteriol 95, 1495-1503. Tuttle, J. H., Dugan, P. R., Macmillan, C. B. & Randles, C. I. (1969a). Microbial dissimilatory sulfur cycle in acid mine water. J Bacteriol 97, 594-602. Tuttle, J. H., Dugan, P. R. & Randles, C. I. (1969b). Microbial sulfate reduction and its potential utility as an acid mine water pollution abatement procedure. Appl Microbiol 17, 297-302. Tuttle, J. H. & Dugan, P. R. (1976). Inhibition of growth, iron, and sulfur oxidation in Thiobacillus ferrooxidans by simple organic compounds. Can J Microbiol 22, 719-730. Tuttle, J. H., Dugan, P. R. & Apel, W. A. (1977). Leakage of cellular material from Thiobacillus ferrooxidans in the presence of organic acids. Appl Environ Microbiol 33, 459-469. Tyson, G. W., Lo, I., Baker, B. J., Allen, E. E., Hugenholtz, P. & Banfield, J. F. (2005). Genome-directed isolation of the key nitrogen fixer Leptospirillum ferrodiazotrophum sp. nov. from an acidophilic microbial community. Appl Environ Microb 71, 6319-6324. Urich, T., Bandeiras, T. M., Leal, S. S. & other authors (2004). The sulphur oxygenase reductase from Acidianus ambivalens is a multimeric protein containing a low-potential mononuclear non-haem iron centre. Biochem J 381, 137-146. Van Aswegen, P. C., Van Niekerk, J. & Oliver, W. (2007). The BIOXTM Process for the treatment of refractory gold concentrates. In Biomining, pp. 281-307. Edited by D. E. Rawlings & B. D. Johnson. Berlin Heidelberg New York: Springer. van Zyl, L. J., Deane, S. M. & Rawlings, D. E. (2003). Analysis of the mobilization region of the broad-host-range IncQ-like plasmid pTC-F14 and its ability to interact with a related plasmid, pTF-FC2. J Bacteriol 185, 6104-6111. Vestal, J. R. & Lundgren, D. G. (1971). The sulfite oxidase of Thiobacillus ferrooxidans (Ferrobacillus ferrooxidans). Can J Biochem 49, 1125-1130. Vigneault, B., Campbell, P. G., Tessier, A. & De Vitre, R. (2001). Geochemical changes in sulfidic mine tailings stored under a shallow water cover. Water Res 35, 1066-1076. Wakai, S., Kikumoto, M., Kanao, T. & Kamimura, K. (2004). Involvement of sulfide:quinone oxidoreductase in sulfur oxidation of an acidophilic iron-oxidizing bacterium, Acidithiobacillus ferrooxidans NASF-1. Biosci Biotechnol Biochem 68, 2519-2528.

72
Wakai, S., Tsujita, M., Kikumoto, M., Manchur, M. A., Kanao, T. & Kamimura, K. (2007). Purification and characterization of sulfide:quinone oxidoreductase from an acidophilic iron-oxidizing bacterium, Acidithiobacillus ferrooxidans. Biosci Biotechnol Biochem 71, 2735-2742. Waksman, S. A. J., J.S. (1922). Microorganisms concerned in oxidation of sulfur in the soil. II. Thiobacillus thiooxidans, a new sulfur-oxidizing organism isolated from the soil. J Bacteriol 7, 239-256. Walton, K. C. & Johnson, D. B. (1992). Microbiological and chemical characteristics of an acidic stream draining a disused copper mine. Environ Pollut 76, 169-175. Weller, J., M. (1960). Glossary of Geology and Related Sciences. Edited by G. R. Committee. Washington, D.C.: Williams and Heinz Lithograph Div.- Kaufmann Printing Inc. Wichlacz, P. L. & Unz, R. F. (1981). Acidophilic, heterotrophic bacteria of acidic mine waters. Appl Environ Microbiol 41, 1254-1261. Xia, J. L., Peng, A. A., He, H., Yang, Y., Liu, X. D. & Qiu, G. Z. (2007). A new strain Acidithiobacillus albertensis BY-05 for bioleaching of metal sulfides ores. Trans Nonferr Metal Soc 17, 168-175. Xie, X., Xiao, S., He, Z., Liu, J. & Qiu, G. (2007). Microbial populations in acid mineral bioleaching systems of Tong Shankou Copper Mine, China. J Appl Microbiol 103, 1227-1238. Yahya, A. & Johnson, D. B. (2002). Bioleaching of pyrite at low pH and low redox potentials by novel mesophilic Gram-positive bacteria. Hydrometallurgy 63, 181-188. Yamanaka, T. & Fukumori, Y. (1995). Molecular aspects of the electron transfer system which participates in the oxidation of ferrous ion by Thiobacillus ferrooxidans. FEMS Microbiol Rev 17, 401-413. Yarzabal, A., Brasseur, G., Ratouchniak, J., Lund, K., Lemesle-Meunier, D., DeMoss, J. A. & Bonnefoy, V. (2002). The high-molecular-weight cytochrome c Cyc2 of Acidithiobacillus ferrooxidans is an outer membrane protein. J Bacteriol 184, 313-317. Yarzabal, A., Duquesne, K. & Bonnefoy, V. (2003). Rusticyanin gene expression of Acidithiobacillus ferrooxidans ATCC 33020 in sulfur- and in ferrous iron media. Hydrometallurgy 71, 107-114. Yarzabal, A., Appia-Ayme, C., Ratouchniak, J. & Bonnefoy, V. (2004). Regulation of the expression of the Acidithiobacillus ferrooxidans rus operon encoding two cytochromes c, a cytochrome oxidase and rusticyanin. Microbiology 150, 2113-2123.

73
Yin, H., Cao, L., Qiu, G., Wang, D., Kellogg, L., Zhou, J., Liu, X., Dai, Z. & Ding, J. (2008). Molecular diversity of 16S rRNA and gyrB genes in copper mines. Arch Microbiol 189, 101-110. Younger, P. L. (1997). The longevity of minewater pollution: a basis for decision-making. Sci Total Environ 194-195, 457-466. Zeng, J., Zhang, Y. F., Liu, Y. D., Zhang, X. J., Xia, L. X., Liu, J. S. & Qiu, G. Z. (2007). Expression, purification and characterization of a cysteine desulfurase, IscS, from Acidithiobacillus ferrooxidans. Biotechnol Lett 29, 1983-1990. Zis, T., Ronningen, V. & Scrosati, R. (2004). Minor improvement for intertidal seaweeds and invertebrates after acid mine drainage diversion at Britannia Beach, Pacific Canada. Mar Pollut Bull 48, 1040-1047.


















