
PHYLOGENETIC AND BIOGEOGRAPHIC RELATIONSHIPS OF …€¦ · 3 Couper (1960) and Lee, Lee & Mortimer...
59
PHYLOGENETIC AND BIOGEOGRAPHIC RELATIONSHIPS OF CHEIRODENDRON NUTT. EX. SEEMAN (ARALIACEAE) A THESIS SUBMITTED TO THE GRADUATE DIVISON OF THE UNIVERSITY OF HAWAIʻI AT MĀNOA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF MASTER OF SCIENCE IN BOTANY AUGUST 2014 By Chelsea Osaki Thesis Committee: Clifford Morden, Chairperson Donald Drake David Lorence
Transcript of PHYLOGENETIC AND BIOGEOGRAPHIC RELATIONSHIPS OF …€¦ · 3 Couper (1960) and Lee, Lee & Mortimer...

PHYLOGENETIC AND BIOGEOGRAPHIC RELATIONSHIPS OF CHEIRODENDRON
NUTT. EX. SEEMAN (ARALIACEAE)
A THESIS SUBMITTED TO THE GRADUATE DIVISON OF THE
UNIVERSITY OF HAWAIʻI AT MĀNOA IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF
MASTER OF SCIENCE
IN
BOTANY
AUGUST 2014
By
Chelsea Osaki
Thesis Committee:
Clifford Morden, Chairperson
Donald Drake
David Lorence

ii
ACKNOWLEDGEMENTS
I would like to acknowledge the following people whom I am grateful for in helping
me to complete this project: Dr. Clifford Morden for his patience and kindness with the
project and throughout my time as a graduate student; Dr. Donald Drake for his insightful
comments on my thesis; and Dr. David Lorence for his knowledge of the Marquesas Islands.
I would also like to thank Joel Lau, whose ideas about the evolution and speciation of
Cheirodendron have served as the foundations of this project. To Mitsuko Yorkston, who has
taught me every lab technique I know. Thank you to Dr. Anthony Mitchell for his comments
and suggestions for primers and Dr. Ines Schönberger from Allan Herbarium (CHR) for
providing outgroup samples.
There were many people who have been involved in field work, collecting specimen,
sorting herbarium samples, performing DNA extractions, PCRs, and analyses, whose help I
greatly appreciate: Dr. Gregory Plunkett, Dr. Pei-Luen Lu, Dr. Richard Pender, Nipuni
Sirimalwatta, Adam Williams, Seana Walsh, Dylan Morden, Wendy Kishida, Steve Perlman,
Dr. Timothy Gallaher, Vianca Cao, Jesse Adams, Peter Wiggin, Bao Ying Chen, April
Cascasan, Dylan Davis, Jacy Miyaki, Gavin Osaki, Erin Fujimoto, George Akau, Matthew
Campbell, Isaiah Smith, Dr. Daniel Rubinoff, Dr. Michael Thomas, and Robert Tamayo.
Finally, I would like to thank the Botany Department at the University of Hawaiʻi at
Mānoa for providing me with opportunities to do my research; National Tropical Botanical
Garden for excellent hospitality during my stay on Kauaʻi; and finally, the Kōkeʻe Resource
Conservation Program for providing me with the opportunity to learn about conservation,
native Hawaiian plants and the beautiful island of Kauaʻi.

iii
TABLE OF CONTENTS
ACKNOWLEDGEMENTS ...................................................................................................... ii
LIST OF TABLES .................................................................................................................... v
LIST OF FIGURES ................................................................................................................. vi
CHAPTER 1. LITERATURE REVIEW AND THESIS PROPOSAL ................................... 1
INTRODUCTION ................................................................................................................. 1
Family Araliaceae ............................................................................................................. 2
Taxonomic history of the genus Cheirodendron................................................................ 3
Traditional uses ................................................................................................................. 5
PROPOSED RESEARCH AND HYPOTHESES ................................................................ 6
MATERIALS AND METHODS .......................................................................................... 7
Taxon sampling, outgroup sampling and DNA extraction ................................................ 7
DNA sequencing and analysis ........................................................................................... 8
Preliminary data ................................................................................................................ 9
Future directions ............................................................................................................... 9
CHAPTER 2. PHYLOGENETIC AND BIOGEOGRPAHIC RELATIONSHIPS OF
CHEIRODENDRON NUTT. EX. SEEM. (ARALIACEAE) ................................................. 13
ABSTRACT ........................................................................................................................ 13
INTRODUCTION ............................................................................................................... 14
MATERIALS AND METHODS ........................................................................................ 19

iv
Sampling and DNA extraction ......................................................................................... 19
DNA amplification and DNA sequencing ........................................................................ 20
Primer screening and analysis ........................................................................................ 23
RESULTS............................................................................................................................ 27
DISCUSSION ..................................................................................................................... 33
Relationships among taxa ................................................................................................ 34
Current taxonomy vs. phylogenetic relationships ........................................................... 37
Biogeographic relationships............................................................................................ 39
Future directions ............................................................................................................. 40
CHAPTER 3. SYNTHESIS- HYPOTHESES REVISITED ................................................. 42
LITERATURE CITED ........................................................................................................... 44

v
LIST OF TABLES
Table Page
1.1 Comparison of revisions of Cheirodendron………………………………………...10-11
2.1 Voucher information and locality of specimen used in this study………………….21-22
2.2 List of primers and references………………………………………………………24-26

vi
LIST OF FIGURES
Figure Page
1.1 Infrafamilial phylogenetic relationships of Araliaceae………………………………12
2.1 ETS phylogeny of Cheirodendron…………………………………………………...29
2.2 ITS phylogeny of Cheirodendron………………………………………………….…30
2.3 ndhF-rpl32 phylogeny of Cheirodendron…………………………………………....31
2.4 Combined phylogeny comparing MP, ML, and BI methods………………………..32

1
CHAPTER 1. LITERATURE REVIEW AND THESIS PROPOSAL
INTRODUCTION
Understanding infrageneneric relationships are crucial in providing a basis in analysis
for other studies such as biogeography, ecology, macroevolution, systematics, and
conservation biology (Sites & Marshall 2004). One way to determine these relationships is
through phylogenetics. Understanding phylogeny, or how species are related to one another,
provides a comprehensive interpretation of evolutionary processes, including speciation
(Harrison 1998). The study of molecular phylogenetics and phylogenetic trees has provided a
direct record of the speciation events that have led to the extant species we see today
(Barraclough & Nee 2001).
Phylogenetic analysis has been a useful tool in elucidating the microevolutionary and
macroevolutionary relationships in a number of lineages, including the family Araliaceae.
Progress has been made in resolving the placement of Araliaceae within the order Apiales
(Henwood & Hart 2001, Plunkett & Lowry 2001, Plunkett et al. 2004) as well as
understanding relationships within and among closely related genera (e.g. Wen & Zimmer
1996, Mitchell & Wagstaff 1997, Costello & Motley 2001, Eibl, Plunkett & Lowry 2001).
Although studies have been done on some of the more horticulturally (Hedera L., ivy) or
ethnobotanically (Panax L., ginseng) important genera, little attention has been given to
other genera within Araliaceae, particularly Cheirodendron Nutt. ex Seem.
The phylogenetic relationships among species of Cheirodendron are currently
unknown. To date, only a few taxonomical studies have assessed species relationships.
However, speciation is not always accompanied by clear morphological differentiation

2
(Kenfack 2011). The importance of phylogenetics in discovering species relationships of
Cheirodendron will sort out the taxonomy of the genus. My proposed research aims to
uncover species relationships by sequencing various regions of both chloroplast and nuclear
DNA. Using a molecular approach, genetic differences will be assessed, rather than
morphological characters which can be influenced by the environment and highly plastic.
Family Araliaceae
The Araliaceae consists of about 50 genera (Liu et al. 2012) and 1500 species that are
widely distributed in the tropics and subtropics of Asia, the Pacific Islands, and the
Neotropics (Wen et al. 2001, Yi et al. 2004). Most members of Araliaceae are woody with
variable leaf morphologies (simple, palmately compound and pinnately compound), but
maintain conserved floral characteristics (5-merous flowers with inferior ovaries in a
compound umbel) (Wen et al. 2001, Yi et al. 2004, Liu et al. 2012). Some well-known
species in Araliaceae include medicinal herbs such as Panax ginseng C. A. Meyer (ginseng),
and ornamentals including Hedera helix L. (English ivy) and Schefflera actinophylla (Endl.)
Harms (umbrella tree) (Liu et al. 2012).
Molecular studies of Araliaceae agree on a phylogeny with multiple polytomies and
poorly resolved basal lineages (Figure 1.1) (Mitchell & Wagstaff 1997, Plunkett, Wen &
Lowry 2004). Poorly resolved genera that are basal in Araliaceae include Schefflera J. R.
Forster & G. Forster, Cheirodendron Nutt. ex. Seem., Raukaua Seem., Cussonia Thunb.,
Osmoxylon Miq., and Hydrocotyle L. (Plunkett, Wen & Lowry 2004).
Implications about the origins of these taxa and explanations about the unresolved
basal polytomies suggest that there was a rapid diversification and radiation in Gondwana
(Mitchell & Wagstaff 2000, Plunkett, Wen & Lowry 2004). These findings corroborate with

3
Couper (1960) and Lee, Lee & Mortimer (2001) who place the origin of Araliaceae in New
Zealand during the Lower Eocene. These studies also suggest that the closest relative of
Cheirodendron is Raukaua, a genus of six species with a “Gondwanan distribution,” endemic
to New Zealand, Chile, Argentina, and Tasmania (Mitchell, Frodin & Heads 1997).
Taxonomic history of the genus Cheirodendron
The taxonomic history of Cheirodendron is convoluted. The first collections of
Cheirodendron were made by Gaudichaud, who named the plant Aralia trigyna in 1830
(Herat 1981). In his description, he noted the glabrous and opposite leaves of the plant and
its triangular fruit (Herat 1981). De Candolle (1830) placed Gaudichaud’s Aralia trigyna into
the genus Panax as P. gaudichaudii DC. Soon after, with collections made on Captain
Beechey’s voyage around the Pacific, Hooker & Arnott (1832) added two new species to De
Candolle’s Panax: P. ovatum and P. platyphyllum.
Later, well-known botanist at the time Asa Gray (1854) changed the genus to Hedera.
However, when the revision of Hederaceae occurred, Berthold Seemann took up the name
Cheirodendron from Nuttall’s manuscripts and published it as a new genus in his Revision of
the Natural Order Hederaceae (Seemann 1868). In his classification, he grouped five species
with ranges around the Pacific: C. gaudichaudii (DC) Seem. and C. platyphyllum (Hook. &
Arnott) Seem. of the Hawaiian Islands; C. laetivirens (Gay) Seem. and C. valdiviense (Gay)
Seem. of Chile; and C. samoense (A. Gray) Seem. of the Samoan Islands (Seemann 1868).
However, since then, the Chilean and Samoan species were removed from Cheirodendron,
leaving only two species (C. gaudichaudii and C. platyphyllum) in the genus.
Looking closer at Seeman’s Hawaiian species, Hillebrand (1888) was the first to
describe different forms among the Hawaiian species. Hillebrand (1888) saw five forms in C.

4
gaudichaudii, distinguished as: α, β, γ, δ, and ε. However, Heller (1897) recognized that C.
gaudichaud was the same taxon as Gaudichaud’s Aralia trigyna which had priority and
created the new combination Cheirodendron trigynum (Gaud.) Heller (Sherff 1954).
Cheirodendron fauriei Hochreutiner was described by Hochreutiner (1925) and C. dominii
Krajina, found only on Kauai, was described by Krajina (1931). Sherff (1954) later
recognized 14 subspecies of C. trigynum. (Table 1.1).
A Marquesan species, C. marquense Brown, was described in 1935, but was later
found to have been previously described in 1864 as Aralia bastardiana Decaisne (Frodin
1990). Frodin (1990) made a new combination: Cheirodendron bastardianum (Decne.)
Frodin.
Thus far, there have been three treatments classifying all species and subspecies of
Cheirodendron (Table 1.1). Sherff (1954) examined leaflet shape, carpel number, fruit size
and number of flowers per umbellule. He recognized six species, along with associated
variations and forms within the Hawaiian Islands. Although he takes note of a species in the
Marquesas, it was not included in his revision.
Herat examined the morphological and anatomical traits for many different
characters. These include traits involving the petiole and petiolule, lamina, young stem,
wood, pollen, and fruit characteristics (Herat 1981). In his taxonomic delineation, he
recognized six species of Cheirodendron in Hawaii and one species in the Marquesas (Herat
1981) (Table 1.1). His treatment greatly reduced the number of subspecific categories that
Sherff (1954) recognized.
The most recent revision of Cheirodendron was by Lowry (1990) as published in the
Manual of Flowering Plants of Hawaii. Lowry (1990) recognized five Hawaiian species and

5
one Marquesan species, making C. keakuense Kraj. var. forbesii Sherff its own species, C.
forbesii (Sherff) Lowry, and lumping C. helleri Sherff with C. trigynum (Gaud.) A. Heller.
This current taxonomy recognizes C. dominii, C. forbesii, C. fauriei, C. platyphyllum and C.
trigynum as the Hawaiian taxa and C. bastardianum as the Marquesan taxon (Lowry 1990)
(Table 1.1).
In his revision, Lowry suggests that a number of taxa in Sherff’s (1954) classification
may warrant proper species recognition. Although not noted in Table 1.1, Sherff (1954)
recognized slight population differences in distinct island locations (e.g. Northwest Kauai,
Northernmost tip of Hawaii, etc.) where each subspecies was found. Lowry’s suspicions, as
well as suspicions of others who have worked in the field, lead me to believe that these
differences represent genetically distinct taxa. However, more evidence must be presented, in
which this research intends to do.
Traditional uses
This leaf structure is indicative of the name, Cheirodendron, which is derived from
the Greek word cheiros meaning “hand” and dendron meaning “tree,” referring to its five
leaflets fixed at a common point (Rock 1913). In Hawaiian, Cheirodendron (spp.) is called
ʻōlapa or lapalapa which may have gotten its name from the similarities between the plant’s
fluttering leaves in the slightest of wind and the movements of the skirt of a hula dancer of
rank ʽōlapa (Lowry 1990). In the Marquesan language, Cheirodendron is called pimata
(Wagner & Lorence 2002).
Cheirodendron spp. was used traditionally in Hawaii for lei-making (leaves), dye-
making (fruit), and weaponry such as spears (branches) (Abbott 1992). The leaves and bark
of Cheirodendron spp. are recorded to have a carrot-like scent when crushed and, when

6
extracted, provide a fragrance for traditional materials, such as kapa (Abbott 1992, Wagner
& Lorence 2002). Branches would also be slathered with gum from sticky Pisonia L. spp.
seeds to ensnare birds (Abbott 1992). Traditional uses of C. bastardianum in the Marquesas
Islands included utilizing the leaves in lei-making (Wagner & Lorence 2002).
PROPOSED RESEARCH AND HYPOTHESES
The taxonomical work done by Sherff (1954), Herat (1981), and Lowry (1990) are
helpful in discerning species relationships in Cheirodendron. However, genetic data were
unavailable to clarify relationships among the populations and or species. My research will
help to discern these relationships by comparing regions in the nuclear and chloroplast
genomes. Phylogenies based on these regions will show if the current taxonomic
classification corresponds or may show distinct genetic taxa that warrant species recognition.
Discovering the species relationships among Cheirodendron will also elucidate
biogeographic implications regarding the source of the species in Hawaii and Marquesas, its
closest relatives, and biogeographic patterns of dispersal within Hawaii. Three specific
hypotheses have been established:
Hypothesis one: Phylogenetic analysis does not support the current taxonomic
classification based on morphological data.
Based on herbaria specimen and personal field observations, vast morphological
differentiation within the species, especially C. trigynum, causes me to believe that some taxa
were left out of the classification.
Hypothesis two: Cheirodendron is monophyletic.
Mitchell et al. (2012) evaluated the phylogenetic relationships of Raukaua, a
paraphyletic group whose New Zealand species share a sister relationship with

7
Cheirodendron. However, this study only included a single species, C. trigynum, to represent
the genus Cheirodendron (Mitchell et al. 2012). The proposed study will evaluate the
monophyly of Cheirodendron by including all taxa in analyses.
Hypothesis three: The Pacific biogeography of Cheirodendron involves an Austral
origin with long distance dispersal to Hawaii and then to the Marquesas.
Fosberg (1948) suggested the origin of Cheirodendron to be from a single
introduction from an Austral origin. This claim was later supported with phylogenetic
analyses evaluating generic relationships within Araliaceae (Wen et al. 2001, Plunkett, Wen
& Lowry 2004) and studies of Raukaua (Mitchell & Wagstaff 1997, Mitchell & Wagstaff
2000, Mitchell et al. 2012), the genus whose shares a sister relationship to Cheirodendron.
With precedence of other groups with Pacific distributions, Gillett (1972) suggested
that Cheirodendron is likely to have spread from an Austral origin to Hawaii, and then to the
Marquesas. An in depth morphological assessment of the group suggested the same
biogeographic pattern, as the leaf morphology of the Marquesan species, C. bastardianum,
resembles an intermediate form between two Hawaiian species, C. platyphyllum and C.
trigynum (Herat 1981). In addition, the fruit anatomy of the Marquesan species resembles C.
platyphyllum (Herat 1981).
MATERIALS AND METHODS
Taxon sampling, outgroup sampling and DNA extraction
To ensure proper taxon sampling, collection will include all taxa mentioned in the
current revision (Lowry 1990), including samples of the same taxon from different islands. If
personal collection is not possible, collection by collaborators or herbarium specimen will
supplement my personal collection. For all collections made, a voucher will be collected for

8
reference, along with a tissue sample that will be extracted of DNA. Outgroup taxa chosen
for this study will be Raukaua anomalus (Hook.) A. D. Mitch., Frodin & Heads, R. edgerleyi
(Hook. f.) Seem., R. simplex (G. Forst.) A. D. Mitch., Frodin & Heads and Schefflera digitata
J. R. & G. Forst.; all shown to be monophyletic with Cheirodendron in a previous study
(Mitchell et al. 2012). Total genomic DNA will be extracted using a modified CTAB
protocol (Doyle & Doyle 1987) and purified with phenol and chloroform.
DNA sequencing and analysis
Primers will be chosen on the basis of having enough variability. Polymerase chain
reaction will be performed to amplify DNA regions of interest. PCR products will be
visualized on 1% agarose gel stained with EtBr to confirm amplification before being
cleaned with ExoSAP –IT (USB, Cleveland, Ohio, USA). Cleaned PCR products will be sent
to the Greenwood Molecular Biology Facility at the University of Hawaii at Manoa. PCR
products will be sequenced in both the forward and reverse directions to confirm sequences,
and completed sequences will be edited and assembled in Sequencher 3.0 (Gene Codes
Corporation, Ann Arbor, Michigan, USA). Sequences will be aligned using ClustalW in
MEGA version 6 (Tamura et al. 2013) and trimmed to be the same number of base pairs.
Molecular evolutionary analyses using maximum parsimony (MP) will be conducted
using MEGA version 6 (Tamura et al. 2013). Maximum likelihood (ML) analyses will be
carried out on a web-based server, molecularevoution.org, using GARLI version 2.0
(Bazinet, Zwickl & Cummings 2014). Bayesian inference will be carried out using MrBayes
version 3 (Hueselenbeck & Ronquist 2001). If there is congruence among gene regions, a
combined dataset will be analyzed under the phylogenetic methods mentioned above.

9
Preliminary data
Thus far, primer screening for variable regions has been ongoing. The problem has
been finding regions that have enough variability to resolve a phylogenetic tree. At present,
both nuclear and chloroplast regions have been sequenced, including gene, spacer and intron
regions of the genome. Nuclear regions sequenced include Phytochrome C (gene), Nitrate
reductase (intron) and the internal transcribed spacer (ITS) (spacer). Chloroplast regions
include atpB-rbcL (spacer), rbcL (gene), trnL-trnF (spacer), and ndhF-rpl32 (spacer). All
regions tested to date have resulted in little to no variability
Future directions
Regions that show more genetic variation need to be found. Since regions used in
many other phylogenetic studies (ie: trnL-trnF intergenic spacer) do not show variation in
Cheirodendron, faster-evolving regions need to be sequenced. Other chloroplast regions will
be tested as described by Shaw et al. (2005, 2007). Nuclear regions described in Zimmer &
Wen (2012) can also be looked into. Good samples that are representative of the different
taxa around Hawaii need to be analyzed. Thorough collection around all the main Hawaiian
Islands needs to be done.

Table 1.1. Comparison of revisions of Cheirodendron. For location, K=Kauai, O=Oahu, Mo=Molokai, L=Lanai, M=Maui, H=Hawaii,
Marq=Marquesas Islands
Sherff
(1954)
Herat
(1981)
Lowry
(1990)
Species Subspecies Form Location Species Subspecies Location Species Subspecies Location
platyphyllum O platyphyllum platyphyllum O platyphyllum platyphyllum O
kauaiense keakuense K kauaiense K kauaiense K
forbesii K keakuense keakuense K forbesii K
dominii K forbesii K dominii K
trigynum subcordatum H dominii K trigynum trigynum O, Mo,
L, M, H
fosbergii O trigynum trigynum H helleri K
mauiense M acuminatum H fauriei K
oblongum latius M fosbergii O bastardianum Marq
molokaiense angustius Mo, M hillebrandii O, L, Mo
osteostigma Mo mauiense Mo, M
halawanum O, M, L fauriei K
hillebrandii O helleri K
confertiflorum M marquesense Marq
rockii L
skottsbergii L, M
ilicoides H
acuminatum H
degeneri pauciflorum H
fauriei macdanielsii K
10
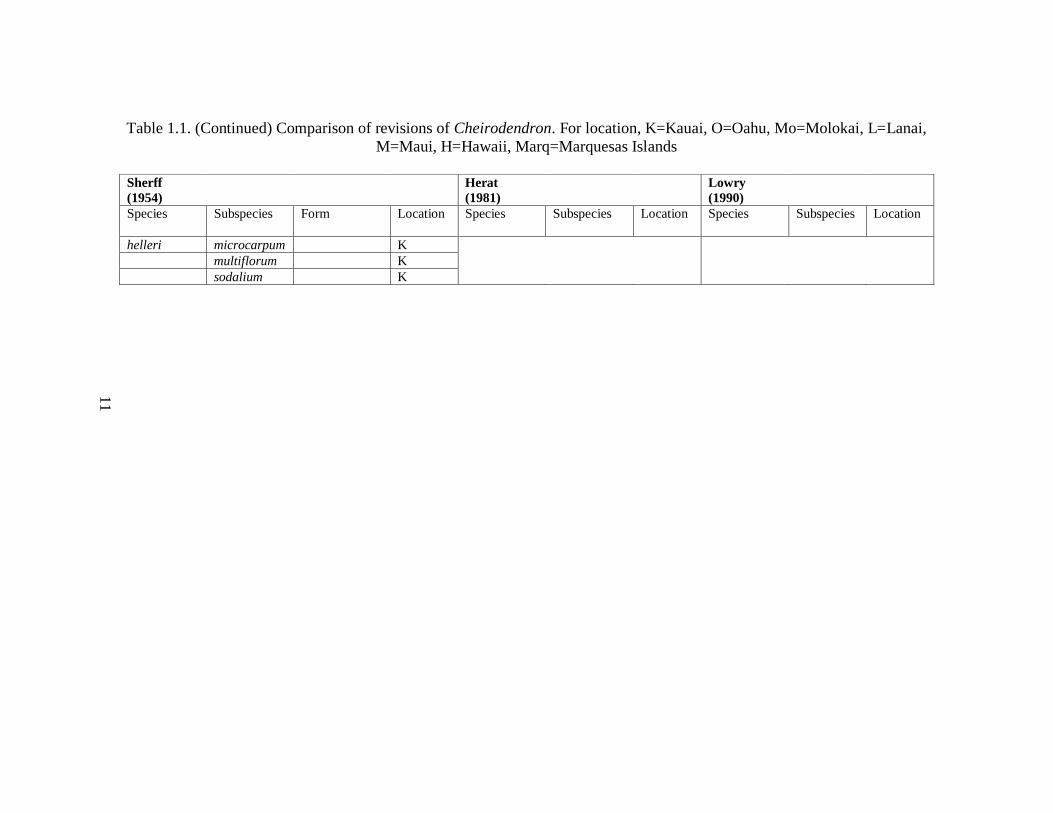
Table 1.1. (Continued) Comparison of revisions of Cheirodendron. For location, K=Kauai, O=Oahu, Mo=Molokai, L=Lanai,
M=Maui, H=Hawaii, Marq=Marquesas Islands
Sherff
(1954)
Herat
(1981)
Lowry
(1990)
Species
Subspecies Form Location Species Subspecies Location Species Subspecies Location
helleri microcarpum K
multiflorum K
sodalium K
11

12
Figure 1.1. Infrafamilial phylogenetic relationships of Araliaceae. Consensus from maximum
parsimony analysis of combined ITS and trnL-trnF data. (from Plunkett, Wen & Lowry
2004).

13
CHAPTER 2. PHYLOGENETIC AND BIOGEOGRPAHIC RELATIONSHIPS OF
CHEIRODENDRON NUTT. EX. SEEM. (ARALIACEAE)
ABSTRACT
Cheirodendron is a genus of six arboreal species in the family Araliaceae, distributed
in the Hawaiian and Marquesas Islands. Previous and current revisions were assessed using
morphological characteristics, resulting in the present taxonomy that consists of five species
and two subspecies in Hawaii and one species in the Marquesas. For the first time, molecular
phylogenetic analyses were carried out to determine species and biogeographic relationships
using sequences from the internal and external transcribed spacer regions of nuclear
ribosomal DNA, and the ndhF-rpl32 chloroplast spacer region. The results suggest that
Cheirodendron is a monophyletic group with Marquesan species C. bastardianum sister to
the Hawaiian Cheirodendron. Within the Hawaiian Cheirodendron, a clade of non-Kauai
taxa was well-supported and a clade of Kauai taxa was weakly supported. However, species
relationships within the Hawaiian clades were unresolved. Phylogenetic differences among
subspecies suggest the recognition of two previously-recognized species, C. helleri Sherff
and C. kauaiense Kraj. Results suggest two possible biogeographic patterns of
Cheirodendron in the Pacific: (1) a stepping stone pattern of dispersal from New Zealand to
Marquesas, and Marquesas to Hawaii or (2) a simultaneous colonization of both Hawaii and
Marquesas. Understanding species relationships and the biogeography of Cheirodendron
adds to our knowledge of the evolution and speciation of Pacific island groups. Long
distance dispersal, along with in situ speciation on island archipelagos provides interesting
evolutionary and biogeographic patterns to be discussed.

14
INTRODUCTION
Cheirodendron Nutt. ex. Seeman is a genus of six arboreal species in the family
Araliaceae. Araliaceae is composed of about 50 genera and 1500 species widely distributed
in the tropics and subtropics of Asia, the Pacific Islands, and the Neotropics (Wen et al. 2001,
Yi et al. 2004). Most members of Araliaceae are woody with variable leaf morphologies, but
maintain conserved floral characteristics (Wen et al. 2001, Yi et al. 2004, Liu et al. 2012).
Some well-known species in Araliaceae include medicinal herbs such as ginseng (Panax
ginseng C. Meyer), and ornamentals including English ivy (Hedera helix L.) and the
umbrella tree (Schefflera actinophylla (Endl. Harms) (Liu et al. 2012).
Cheirodendron has a tropical distribution in the Pacific Ocean with species occurring
in the Hawaiian Islands and the Marquesas Islands. Mature individuals stand at 2-15 meters
and are one of the dominant canopy constituents in mesic forests, wet forests and bogs from
600-1500 meters in elevation (Lowry 1990). All species have compound leaves, with 3-5(-7)
leaflets with margins toothed to entire (Lowry 1990). Flowers are perfect and arranged in
oppositely branched umbels, with carpel numbers varying from two to five (Frodin 1990,
Lowry 1990). When broken or crushed, plant parts emit a strong carrot-like odor (Frodin
1990, Lowry 1990). Fruits are small fleshy drupes that, when ripe, exude a deep purple liquid
which traditional Hawaiians used for dyes.
The current taxonomy recognizes six species distinguishable by morphological
characteristics of leaflet shape and carpel number (Lowry 1990). Cheirodendron trigynum
(Gaud.) A. Heller is the most common species and is known from all the high Hawaiian
Islands except Kahoolawe. It has been reported from Niihau (Hooker & Arnott 1832), but has
since been extirpated (St. John 1959). This species is distinguished from the others by

15
having leaflets longer than wide, ovate to elliptic; margins entire or serrate-crenate and teeth
curved upward and inward (Lowry 1990). The species maintains two morphological forms
recognized as distinct subspecies, C. trigynum ssp. trigynum and ssp. helleri (Sherff) Lowry.
Subspecies helleri is only found on the island of Kauai and flowers have two (occasionally 3)
styles and carpels, while ssp. trigynum is only present on the younger islands (Oahu, Maui,
Lanai, Molokai, and Hawaii) and has three to four (rarely 2) styles and carpels.
Cheirodendron platyphyllum (Hook. & Arnott) Seem. is characterized by broadly
ovate to depressed ovate leaflets that are wider than they are long. This species is known
from Kauai and Oahu, where two subspecies are present, differing in carpel number and
leaflet shape. Subspecies kauaiense (Kraj) Lowry from Kauai has two carpels and rounded
leaflet apices, while subspecies platyphyllum from Oahu maintains five (rarely 4) carpels and
acuminate leaflet apices (Lowry 1990). Three other species, C. dominii Kraj., C. fauriei
Hochr. and C. forbesii (Sherff ) Lowry are restricted to Kauai each having differences in the
degree of dentations on their leaflet margins, carpel numbers and locality. The characteristics
of C. dominii include three broadly ovate leaflets longer than wide, margins caudate-dentate
and spiny in appearance, and three to four (sometimes 2 or 5) styles and carpels (Lowry
1990). This taxon is also quite rare, occurring only in wet forest at 1525-1550 m in elevation
near the slopes of Mount Waialeale (Lowry 1990). Similarly, C. fauriei has leaflets with the
same spiny appearance as C. dominii; however they are more orbicular in shape. The ovary
contains two (sometimes 3) carpels and is widely distributed in diverse mesic to wet forests
at 650-1250 m in elevation (Lowry 1990). Cheirodendron forbesii is distinguished by its
leaflets that are nearly twice as long as they are wide, margins entire, and five (sometimes 4)
carpels (Lowry 1990). Cheirodendron bastardianum is the only species that occurs outside

16
Hawaii and it shares similar morphological characteristics with the Hawaiian taxa (Frodin
1990). It is distinguished from Hawaiian Cheirodendron by morphological characteristics
including leaves as long as or slightly longer than broad and rounded, truncate or slightly
cordate at the base, longer pedicels, and smaller fruit (Brown 1935, Wagner & Lorence
2002).
Previous studies and revisions of Cheirodendron (Sherff 1954, Herat 1981, Lowry
1990) focus only on morphological distinctions of the group. However, these defining
features often overlap. Speciation is not always accompanied by clear morphological
differentiation (Kenfack 2011), and it is important to also look at genetic relationships in an
evolutionary perspective. Studies that focus on infrageneric relationships and species
delimitation are important because species are the basic fundamental unit in other disciplines,
including biogeography, ecology, macroevolution, systematics and conservation (Sites &
Marshall 2004).
To date, no molecular work has been done to assess species relationships and the
biogeographic relationships within Cheirodendron. However, several recent studies have
focused on generic relationships within the family Araliaceae. Plunkett, Wen & Lowry
(2004) showed that there were three major clades within the family (Asian palmate group,
Polyscias J. R. Forst. & G. Forst.-Pseudopanax K. Koch group, and Aralia L. group),
although these clades along with numerous other taxa form an unresolved polytomy. These
unresolved groups were attributed to rapid diversification during the Gondwana break up,
which led to the widespread distribution of these groups (Plunkett, Wen & Lowry 2004).
Cheirodendron, along with a close relative, Raukaua Seem., are placed within this polytomy
(Plunkett, Wen & Lowry 2004). Much attention has been given to the biogeographic

17
implications of Raukaua because of its “Gondwanan distribution” in Chile, Argentina, New
Zealand and Tasmania (Mitchell & Wagstaff 1997, Mitchell & Wagstaff 2000). Recently,
Mitchell et al. (2012) demonstrated that Raukaua is paraphyletic based on chloroplast and
nuclear DNA sequence analysis because of the placement of Cephalaralia cephalobotrys (F.
Muell.) Harms, Motherwellia haplosciadea F. Muell., Cheirodendron trigynum (Gaud.) A.
Heller and Schefflera digitata J. R. Forst. intermixed within the Raukaua clade.
Raukaua is a genus of six species with distribution of extant taxa in South America,
New Zealand and Tasmania. The Raukaua taxa closest to Cheirodendron are those from New
Zealand [R. anomalus (Hook.) A. D. Mitch., Frodin & Heads, R. simplex (G. Forst.) A. D.
Mitch., Frodin & Heads, R. edgerleyi (Hook. f.) Seem.], which form a monophyletic clade in
previous analyses (Mitchell & Wagstaff 1997, Mitchell & Wagstaff 2000, Mitchell et al.
2012). Cheirodendron and New Zealand Raukaua share several synapomorphies including
stipules or ligules reduced or absent, coriaceous adult leaf texture, pedicel articulating below
the flower, wood intervessel pitting scalariform or opposite (Mitchell & Wagstaff 2000), fruit
with laterally compressed endocarps, paniculate inflorescences with opposite umbellules,
pentamorous flowers with 2-5 carpels, and palmately compound leaves (in Raukaua mostly
in juvenile leaves) (Lowry, Plunkett & Wen 2004). This supports the idea that
Cheirodendron is a recent relative of an ancestor that probably originated in New Zealand
(Fosberg 1948) and that the current distribution of Cheirodendron is due to a long distance
dispersal event (Mitchell & Wagstaff 2000).
The biogeographic patterns of Cheirodendron across the Pacific have not been well-
studied. However, molecular phylogenetics has been a useful tool in discerning species
relationships and the biogeographic patterns in many Pacific taxa, such as the Hawaiian

18
Silverswords (Carlquist, Baldwin & Carr 2003), Cyrtandra J. R. Forst. & G. Forst.
(Gesneriaceae) (Cronk et al. 2005), Plantago L. (Plantaginaceae) (Dunbar-Co, Wieczorek &
Morden 2008), Astelia s.l. (Asteliaceae) (Birch & Keeley 2013), amonfelsenstieng many
others. The evolution and speciation of these groups is especially interesting because the
islands that they are native to lie miles from the nearest continental land mass and are
resultant from a single, or sometimes, multiple events of long-distance dispersal (Wagner &
Funk 1995).
There has been one study assessing the biogeographic patterns of Cheirodendron.
Based on morphological analyses, Herat (1981) suggests an Austral origin for Cheirodendron
with dispersal overseas to Hawaii, and subsequent dispersal to the Marquesas. This “stepping
stone” dispersal pattern from island to island in the Pacific is well-known among other
indigenous groups such as Metrosideros Banks ex Gaertn. (Myrtaceae) (Wright et al. 2001),
Astelia (Asteliaceae) (Birch & Keeley 2013), Coprosma J. R. Forst. & G. Forst. (Rubiaceae)
(Cantley et al. 2014) and many others. What is also interesting about Herat’s (1981) findings
is that he concludes a Hawaiian species to be the progenitor of the single Marquesan species,
Cheirodendron bastardianum, a claim that Gillett (1972) also made, although his ideas were
merely speculation on the basis that there were similarities between Hawaiian and Marquesan
floras. Hawaii, being one of the Pacific archipelagos most distant from any continental
landmasses, has often been assumed to be a sink for dispersal, rather than a source (Carlquist
1974). However, recent molecular analyses have found that Hawaiian groups such as
Melicope J. R. Forst. & G. Forst. (Rutaceae) (Harbaugh et al. 2009a) and Santalum L.
(Santalaceae) (Harbaugh & Baldwin 2007) have been the source of colonization on other
Pacific archipelagos.

19
The mode of seed dispersal in Cheirodendron across the Pacific is thought to be from
ingestion by birds (Herat 1981) as with many other groups that have Pacific distributions and
fleshy fruits (eg: Howarth & Baum 2005, Harbaugh et al. 2009a, Cantley et al. 2014). The
importance of birds in carrying seeds over long distances is apparent, as a number of
Hawaiian taxa were brought to the islands by birds (Price & Wagner 2004).
The current study used molecular phylogenetics as a tool to determine species
relationships and the biogeography of Cheirodendron by addressing the following questions:
(1) What are the species relationships in Cheirodendron? (2) Does the molecular phylogeny
support the current taxonomy of five species distributed within the Hawaiian Islands? and (3)
Does the molecular data support a colonization of Cheirodendron to the Marquesas Islands
via Hawaiian Islands as Gillett (1972) and Herat (1981) suggest or was there a stepping stone
colonization across the Pacific from the Marquesas Islands to Hawaii? The following study
includes molecular sequence data of two nuclear regions (ETS and ITS) and one chloroplast
region (ndhF-rpl32) to construct a phylogenetic tree for Cheirodendron.
MATERIALS AND METHODS
Sampling and DNA extraction
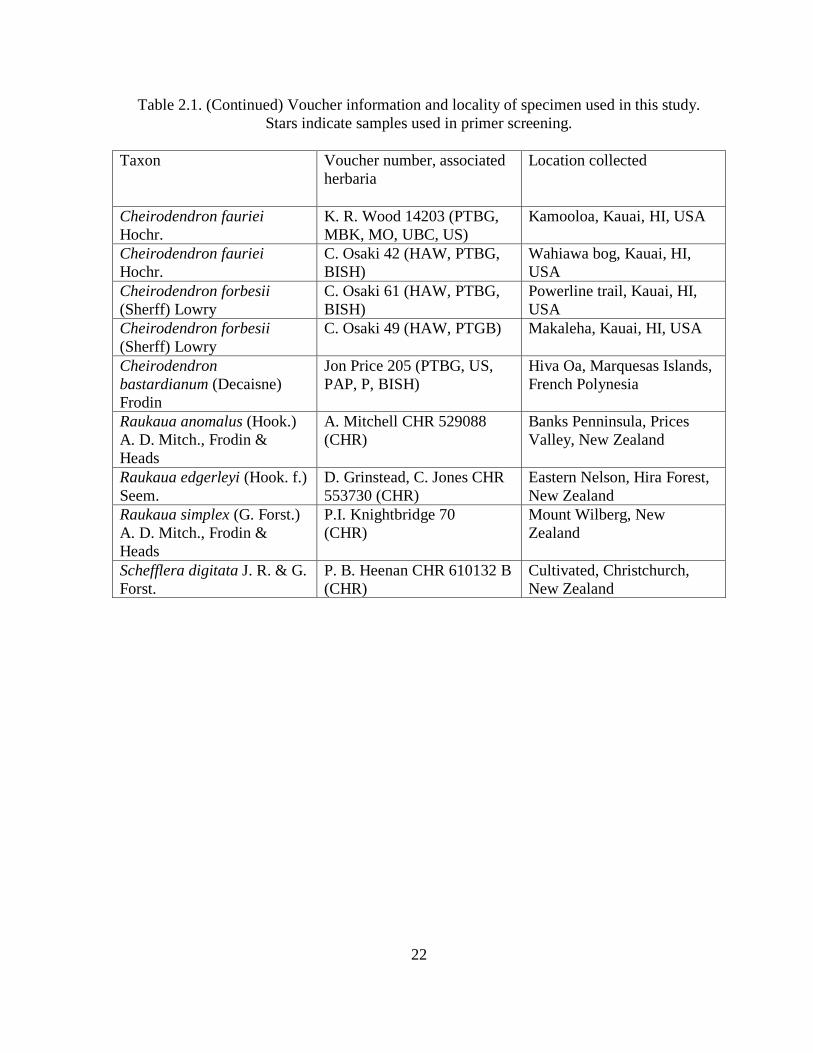
Leaf tissue of all currently recognized taxa (Lowry 1990) was sampled. A total of 22
samples were analyzed, including at least two samples from each taxon in Cheirodendron,
except C. bastardianum, in order to account for possible population differences (Table 2.1).
Although C. bastardianum persists on multiple islands in the Marquesas, only one sample
was available for study from Hiva Oa. However, C. bastardianum has been found to be
morphologically consistent among islands (Steve Perlman, National Tropical Botanical

20
Garden, personal communication). Outgroup taxa chosen for this study were Raukaua
anomalus, R. edgerleyi, R. simplex and Schefflera digitata; all shown to be monophyletic
with Cheirodendron (Mitchell et al. 2012).
Total genomic DNA was extracted from fresh or silica-preserved leaves or herbarium
specimens using a modified CTAB protocol (Morden, Caraway & Motley 1996) and purified
with phenol/chloroform. DNA from Hawaiian taxa were accessioned into the Hawaiian Plant
DNA library and stored at -20°C for future use (Morden, Caraway & Motley 1996).
Vouchers of each individual collected were deposited into the Joseph Rock Herbarium
(HAW).
DNA amplification and DNA sequencing
Each PCR reaction contained a total of 25 µL of 1X GoTaq Flexi Buffer (Promega,
Madison, Wisconsin, USA), 2 mM MgCl2 (Promega), 0.25 µg/µL bovine serum albumin
(Amresco, Solon, Ohio, USA), 0.1 µmol/L of each primer (forward and reverse, see Table
2.2), 0.2 mM each dNTP, 1U GoTaq Flexi DNA polymerase (Promega) and 1 µL of DNA
diluted to 20 ng/µL. PCR was performed using a DNA Thermocycler (MJ Research, St.
Bruno, Quebec, Canada) programed for an initial denaturation at 95°C for 2 min, followed by
30 cycles of 93°C for 1 min, 45-55°C for 1 min, and 72°C for 2 min, followed by an
extended elongation on the final step of 72°C for 3 min. When necessary, PCR products were
cloned using the StrataClone PCR Cloning Kit with associated protocols (Agilent
Technologies, La Jolla, California, USA).
PCR products were visualized on 1% agarose gel stained with EtBr to confirm
amplification before being cleaned with ExoSAP –IT (USB, Cleveland, Ohio, USA).

21
Table 2.1. Voucher information and locality of specimen used in this study. Stars indicate
samples used in primer screening.
Taxon
Voucher number, associated
herbaria
Location collected
Cheirodendron trigynum
(Gaud.) A. Heller ssp.
helleri (Sherff) Lowry
K. R. Wood 14168 (PTBG,
BISH, MBK)
Laau, Kauai, HI, USA
Cheirodendron trigynum
(Gaud.) A. Heller ssp.
helleri (Sherff) Lowry
K. R. Wood 15220 (PTBG,
BISH, MO)
Waiakoali, Kauai, HI, USA
* Cheirodendron trigynum
(Gaud.) A. Heller ssp.
helleri (Sherff) Lowry
C. Osaki 52 (HAW, PTBG,
BISH)
Kokee, Kauai, HI, USA
* Cheirodendron trigynum
(Gaud.) A. Heller ssp.
trigynum
C. Morden (HAW) Puulalaau, Hawaii, HI, USA
Cheirodendron trigynum
(Gaud.) A. Heller ssp.
trigynum
C. Osaki 72 (HAW) Manana trail, Oahu, HI, USA
Cheirodendron trigynum
(Gaud.) A. Heller ssp.
trigynum
S. Walsh (HAW) Munroe trail, Lanai, HI, USA
Cheirodendron trigynum
(Gaud.) A. Heller ssp.
trigynum
S. Walsh (HAW) Waihee trail, Maui, HI, USA
* Cheirodendron
platyphyllum (Hook. &
Arnott) Seem. ssp.
kauaiense (Kraj.) Lowry
C. Osaki 56 (HAW, PTBG,
BISH)
Circle Bog, Kauai, HI, USA
Cheirodendron
platyphyllum (Hook. &
Arnott) Seem. ssp.
kauaiense (Kraj.) Lowry
K. R. Wood 14170 (PTBG,
BISH, K, MBK)
Laau, Kauai, HI, USA
Cheirodendron
platyphyllum (Hook. &
Arnott) Seem. ssp.
platyphyllum
C. Osaki 32 (HAW) Konahuanui, Oahu, HI, USA
*Cheirodendron dominii
Kraj.
K. R. Wood 12284 (PTBG,
BISH, UC)
Waialeale, Kauai, HI, USA
Cheirodendron dominii
Kraj.
C. Osaki 70 (HAW, PTBG,
BISH)
Kilohana vista, Kauai, HI,
USA
Cheirodendron fauriei
Hochr.
K. R. Wood 15185 (PTBG,
BISH, UC, US)
Kalalau rim, Kauai, HI, USA
22
Table 2.1. (Continued) Voucher information and locality of specimen used in this study.
Stars indicate samples used in primer screening.
Taxon
Voucher number, associated
herbaria
Location collected
Cheirodendron fauriei
Hochr.
K. R. Wood 14203 (PTBG,
MBK, MO, UBC, US)
Kamooloa, Kauai, HI, USA
Cheirodendron fauriei
Hochr.
C. Osaki 42 (HAW, PTBG,
BISH)
Wahiawa bog, Kauai, HI,
USA
Cheirodendron forbesii
(Sherff) Lowry
C. Osaki 61 (HAW, PTBG,
BISH)
Powerline trail, Kauai, HI,
USA
Cheirodendron forbesii
(Sherff) Lowry
C. Osaki 49 (HAW, PTGB) Makaleha, Kauai, HI, USA
Cheirodendron
bastardianum (Decaisne)
Frodin
Jon Price 205 (PTBG, US,
PAP, P, BISH)
Hiva Oa, Marquesas Islands,
French Polynesia
Raukaua anomalus (Hook.)
A. D. Mitch., Frodin &
Heads
A. Mitchell CHR 529088
(CHR)
Banks Penninsula, Prices
Valley, New Zealand
Raukaua edgerleyi (Hook. f.)
Seem.
D. Grinstead, C. Jones CHR
553730 (CHR)
Eastern Nelson, Hira Forest,
New Zealand
Raukaua simplex (G. Forst.)
A. D. Mitch., Frodin &
Heads
P.I. Knightbridge 70
(CHR)
Mount Wilberg, New
Zealand
Schefflera digitata J. R. & G.
Forst.
P. B. Heenan CHR 610132 B
(CHR)
Cultivated, Christchurch,
New Zealand

23
Cleaned PCR products were sent to the Greenwood Molecular Biology Facility at the
University of Hawaii at Manoa. PCR products were sequenced in both the forward and
reverse directions to confirm sequences, and completed sequences were edited and assembled
in Sequencher 3.0 (Gene Codes Corporation, Ann Arbor, Michigan, USA). Sequences were
aligned using ClustalW in MEGA version 6 (Tamura et al. 2013) and trimmed to be the same
number of base pairs.
Primer screening and analysis
Before choosing which regions to analyze, primers were screened based on their
ability to amplify and the number of indels among individuals of the genus. To identify
variable gene regions for analysis, four samples representing 3 species from different
locations were used: C. trigynum ssp. helleri (Kauai), C. trigynum ssp. trigynum (Hawaii), C.
dominii (Kauai) and C. platyphyllum ssp. kauaiense (Kauai) (See Table 2.1, taxa used for
screening indicated with a star *). In total, 15 chloroplast and 9 nuclear regions were
examined (Table 2.2). However, only three regions had enough variation where all samples
amplified, and these were then sequenced for all individuals. ITS sequences for outgroup
taxa (Schefflera digitata, Raukaua simplex, R. anomalus, and R. edgerleyi) were taken from
GenBank identified by the following respective accession numbers: JX106299.1, U63180.2,
U63164.1, U63171.1.
Before performing phylogenetic analysis, JModel Test 2 (Darriba et al. 2012,
Guindon & Gascuel 2003) was used to determine the best parameters for each region
separately and combined according to the Akaike information criterion (AIC).
Molecular evolutionary analyses using maximum parsimony (MP) were conducted
using MEGA version 6 (Tamura et al. 2013). All characters were equally weighted and gaps

Table 2.2. List of primers and references.
Locus Genome
origin, type of
DNA
Primer names, sequences (5’-3’) and reference Approximate
length of
region (base
pairs)
Number
of indels
Percent
variability
(%)
Internal
transcribed
spacer (ITS)
Nuclear,
spacer
ITS 5: GGAAGTAAAAGTCGTAACAAGG
ITS 4: TCCTCCGCTTATTGATATGC
(Baldwin 1992)
600 9 1.5
Phytochrome C
(PHYC)
Nuclear, gene Phy C F: GAYTTRGARCCWGTDAAYC
Phy C R: GRATKGCATCCATYTCMAYRTC
(Matthews & Donoghue 1999)
600 1 0.2
Nitrate
reductase (NIA)
Nuclear, gene NIA F5: GCTGAACTTGCTAACGCTGA
NIA i2R: CCATGTCTCTCCTCCATCCA
(Levin, Blanton & Miller 2009)
600 5 0.8
Nitrate
reductase (NIA)
Nuclear,
intron
NIA i3 F:
AARTAYTGGTGYTGGTGYTTYTGGTC
NIA i3 R: GAACCARCARTTGTTCATCATDCC
(Howarth & Baum 2002)
1300 22
1.7
Alcohol
dehydrogenase
(Adh)
Nuclear, gene Adhx2-1 F: CTTCACTGCTTTATGTCACACT
Adhx8-1 R: GGACGCTCCCTGTACTCC
(Small & Wendel 2000)
Did not
amplify
- -
Phytochrome A
(PHYA)
Nuclear, gene Phy A F: CCYTAYGARGRNCCYATGACWGC
Phy A R: GDATDGCRTCCATYTCRTAGTC
(Matthews & Donoghue 1999)
Did not
amplify
- -
5S-Non-
transcribed
spacer
Nuclear,
spacer
5SFUL: TTAGTGCTGGTATGATCGCA
5SR: CACCGGATCCCATCAGAACT
(Udovicic, McFadden & Ladiges 1995)
Did not
amplify
- -
External
transcribed
spacer (ETS)
Nuclear,
spacer
ETS 18S: GAGCCATTCGCAGTTTCACAG
(Wright et al. 2001)
jkETS 9: CGT WMA GGY GYA TGA GTG GT
(Mitchell, Heenan & Patterson 2009)
500 4
0.8
24

Table 2.2. (Continued) List of primers and references.
Locus Genome
origin, type of
DNA
Primer names, sequences (5’-3’) and reference Approximate
length of
region (base
pairs)
Number
of indels
Percent
variability
(%)
Xanthine
dehydrogenase
(Xdh)
Nuclear, gene X502F: TGTGATGTCGATGTATGC
X1599R: G(AT)GAGAGAAA(CT)TGGAGCAAC
(Gorniak, Paun & Chase 2010)
1000 3 0.3
trnL-trnF Chloroplast,
spacer
E: GGTTCAAGTCCCTCTATCCC
F: ATTTGAACTGGTGACACGAG
(Taberlet et al. 1991)
400 1 0.3
trnL Chlroplast,
intron
C: CGAAATCGGTAGACGCTACG
D: GGGGATAGAGGGACTTGAAC
(Taberlet et al. 1991)
535 2 0.4
psbA-trnH Chloroplast,
spacer
psbA: GTTATGCATGAACGTAATGCTC
trnH: CGCGCATGGTGGATTCACAATCC
(Shaw et al. 2005)
450 2 0.4
ndhA Chloroplast,
intron
ndhAx1:
GCYCAATCWATTAGTTATGAAATACC
ndhAx2: GGTTGACGCCAMARATTCCA
(Shaw et al. 2007)
Did not
amplify
- -
atpB-rbcL Chloroplast,
spacer
atpB: GAAGTAGTAGGATTGATTCTC
rbcL: ATGTCAACAGGTACATGGTC
(Manen, Natali & Ehrendorfer 1994)
800 0 0
rbcL Chloroplast,
gene
ESRBCLF:
ATGTCACCACAAACGGAGACTAAAGC
ESRBCL1361R:
TCAGGACTCCACTTACTAGCTTCACG
(Schuettpelz & Pryer 2007)
1200 0 0
ndhF-rpl32 Chloroplast,
spacer
ndhF: GAAAGGTATKATCCAYGMATATT
rpl32r: CCAATATCCCTTYYTTTTCCAA
(Shaw et al. 2007)
1200 8 0.7
25

Table 2.2. (Continued) List of primers and references.
Locus Genome
origin, type of
DNA
Primer names, sequences (5’-3’) and reference Approximate
length of
region (base
pairs)
Number
of indels
Percent
variability
(%)
trnQ-rps16 Chloroplast,
spacer
trnQ: GCGTGGCCAAGYGGTAAGGC
rps16-1: GTTGCTTTYTACCACATCGTTT
(Shaw et al. 2007)
2000 21 1
rpl16 Chloroplast,
gene
rpL16F71:GCTATGCTTAGTGTGTGACTCGTTG
rpl16R1516: CCCTTCATTCTTCCTCTATGTTG
(Shaw et al 2005)
Did not
amplify
- -
trnS-trnG Chloroplast,
spacer
trnS: AACTCGTACAACGGATTAGCAATC
trnG: GAATCGAACCCGCATCGTTAG
(Shaw et al. 2007)
1000 4
0.4
ndhF Chloroplast,
portion of
gene
536F: TTGTAACTAATCGTGTAGGGGA
1603R:
GCATAGTATTGTCCGATTCAT(A/G)AGG
(Olmstead & Sweere 1994)
800 1 0.1
ndhC-trnV Chloroplast,
spacer
ndhC:
TATTATTAGAAATGYCCARAAAATATCA
TATTC
trnVx2: GTCTACGGTTCGARTCCGTA
(Shaw et al. 2007)
1000 4 0.4
trnT-trnL Chloroplast,
spacer
trnT: CAAATGCGATGCTCTAACCT
trnL: GGGGATAGAGGGACTTGAAC
(Shaw et al. 2005)
800 4 0.5
rpl32-trnL Chloroplast,
spacer
rpl32F: CAGTTCCAAAAAAACGTACTTC
trnL: CTGCTTCCTAAGAGCAGCGT
(Shaw et al. 2007)
950
5
0.5
petL-psbE Chloroplast,
spacer
petL: AGTAGAAAACCGAAATAACTAGTTA
psbE: TATCGAATACTGGTAATAATATCAGC
(Shaw et al. 2007)
Did not
amplify
- -
26

27
or missing data were treated as complete deletion. Heuristic searches were conducted using
the tree-bisection reconnection method. A consensus of equally parsimonious trees was
constructed where branches of less than 50% consensus were collapsed into a polytomy. A
total of 1000 bootstrap replicates were carried out to construct the tree. Maximum likelihood
(ML) analyses were carried out on a web-based server, molecularevoution.org, using GARLI
version 2.0 (Bazinet, Zwickl & Cummings 2014). An optimal tree was inferred from 2000
non-parametric bootstrap replicates. If the best-fit model identified in JModel Test was not
available, the general time-reversible (GTR) model was chosen. Bootstrap resampling in both
MP and ML (reported as percentages below) were used to estimate the robustness of the
nodes (Felsenstein 1985). Bayesian inference (BI) was carried out using MrBayes version 3
(Hueselenbeck & Ronquist 2001). The GTR+I+G substitution model and default priors were
used in analysis. Markov chain Monte Carlo (MCMC) sampling was done with two
replicates of four chains (three cold, one hot). 10,000,000 generations were run with a subset
sampling every 1000 generations. A burn-in period excluding the first 25% of trees was set.
Posterior probabilities (reported below as BI) for tree nodes give an overestimate of branch
support (Suzuki, Glazco & Nei 2002) and were interpreted with caution. FigTree version 1
(Rambaut 2012) was used to view trees. Adobe Illustrator CS version 3 (Adobe Systems
Incorporated, San Jose, California, USA) was used to format and edit final trees.
RESULTS
In total, 15 chloroplast and 9 nuclear primer pairs were used to screen for possible
regions for phylogenetic analysis (Table 2.2). Assessment of the percentage of indels within
the region and the ability to amplify resulted in choosing two nuclear and one chloroplast

28
region for phylogenetic analysis: a portion of the external transcribed spacer (ETS), the
internal transcribed spacer (ITS) and ndhF-rpl32 (Table 2.2). There was no genetic variation
among the Hawaiian Cheirodendron species for the other 21 regions examined. When
aligned and trimmed, the total combined dataset included a total of 2365 characters including
the nuclear regions ETS (510 bp), ITS (641 bp) and the chloroplast region ndhF-rpl32
intergenic spacer (1214 bp). JModel Test (Darriba et al. 2012, Guindon & Gascuel 2003)
estimated the best fit model for each region to be HKY (ETS), TPM1uf+I (ITS), TVM+I
(ndhF-rpl32), and TPMuf+I+Γ (combined).
Comparison of each gene region under different phylogenetic methods (parsimony,
maximum likelihood and Bayesian inference) produced similar topologies. Among trees
constructed from the ETS region, maximum likelihood and Bayesian phylogenies showed
weak support for C. bastardianum and C. forbesii as basal to the rest of Cheirodendron
(Figure 2.1) although the relationship among all Cheirodendron spp. is collapsed into a
polytomy in the parsimony analysis. Among ITS trees (Figure 2.2), parsimony, maximum
likelihood and Bayesian analyses agree on an unresolved, but well-supported monophyletic
Cheirodendron clade. Within this larger clade, three subclades were shared among all
analyses: C. forbesii (from Kauai) and C. trigynum ssp. trigynum (from Hawaii) (MP: 75%,
ML: 57%, BI: 0.86), C. trigynum ssp. trigynum (from Maui) and C. trigynum ssp. trigynum
(from Lanai) (MP: 100%, ML: 55%, BI: 0.94), and two of the three collections of C. fauriei
(from Kauai) (MP: 100%, ML: 72%, BI: 0.99). Parsimony and Bayesian inference showed an
additional relationship between two collections of C. trigynum ssp. helleri (MP: 100%, BI:
0.74) that was not inferred in maximum likelihood analysis. Phylogenetic analyses of the
chloroplast region ndhF-rpl32 were congruent (Figure 2.3). In all analyses, C. bastardianum

29
Figure 2.1. ETS phylogeny of Cheirodendron. Bootstrap values and posterior probabilities
are listed near each node as MP (%), ML (%), BI (probability), respectively. Collapsed
branches are denoted as * for any particular analysis. If analysis does not support a particular
relationship, a dash (-) is indicated. The location of where each taxon was collected is in
parentheses next to each name. K=Kauai; O=Oahu; M=Maui; L=Lanai; H=Hawaii;
Marq=Marquesas; NZ=New Zealand.

30
Figure 2.2. ITS phylogeny of Cheirodendron. Bootstrap values and posterior probabilities are
listed near each node as MP (%), ML (%), BI (probability), respectively. Collapsed branches
are denoted as * for any particular analysis. If analysis does not support a particular
relationship, a dash (-) is indicated. The location of where each taxon was collected is in
parentheses next to each name. K=Kauai; O=Oahu; M=Maui; L=Lanai; H=Hawaii;
Marq=Marquesas; NZ=New Zealand.

31
Figure 2.3. ndhF-rpl32 phylogeny of Cheirodendron. Bootstrap values and posterior
probabilities are listed near each node as MP (%), ML (%), BI (probability), respectively.
Collapsed branches are denoted as * for any particular analysis. If analysis does not support a
particular relationship, a dash (-) is indicated. The location of where each taxon was collected
is in parentheses next to each name. K=Kauai; O=Oahu; M=Maui; L=Lanai; H=Hawaii;
Marq=Marquesas; NZ=New Zealand.

32
Figure 2.4. Combined phylogeny comparing MP, ML, and BI methods. Bootstrap values and
posterior probabilities are listed near each node as MP (%), ML (%), BI (probability),
respectively. Collapsed branches are denoted as * for any particular analysis. If analysis does
not support a particular relationship, a dash (-) is indicated. The location of where each taxon
was collected is in parentheses next to each name. K=Kauai; O=Oahu; M=Maui; L=Lanai;
H=Hawaii; Marq=Marquesas; NZ=New Zealand.

33
was closely associated with the outgroup species of Raukaua. The remaining species of
Cheirodendron form two clades, one consisting of the Kauai taxa and the other of non-Kauai
taxa. The Kauai clade formed a polytomy that was strongly supported in MP analysis but
weakly-supported in maximum likelihood and Bayesian inference (MP: 100%, ML: 65%, BI:
0.52). The non-Kauai clade was strongly supported in all three analyses (MP: 100%, ML:
100%, BI: 1).
Though individual genetic regions showed some inconsistencies, the combined
analysis shows strong support for Cheirodendron as a monophyletic lineage, with C.
bastardianum as sister to the Hawaiian species, and the Hawaiian species consisting of two
clades representing Kauai and non-Kauai taxa. Bayesian analysis (Figure 2.4) shows branch
support for C. bastardianum as sister to the rest of the Hawaiian species with a posterior
probability of 1.0 and strong support for a Hawaiian clade (BI: 1). Within the Hawaiian
group, the unresolved Kauai clade shows low support in Bayesian analysis (0.52), which is
reflected in the ML and MP tree that shows all the Hawaiian taxa in a single large clade with
the non-Kauai clade included as a strongly supported subclade within. The MP analysis was
similar to the ML with the exception that the two C. forbesii collections group together and
were sister to the remainder of the Hawaiian Cheirodendron (MP: 100%).
DISCUSSION
Results from this study indicate that Cheirodendron is a monophyletic group that
shares a common ancestor with Schefflera digitata and a sister relationship to New Zealand
Raukaua. Though Raukaua is paraphyletic with the placement of Cheirodendron trigynum,
Schefflera digitata, Motherwellia haplosciadea and Cephalaria cephalobotrys intermixed

34
within the clade (Mitchell et al. 2012), New Zealand Raukaua and Cheirodendron share
many similarities including laterally compressed endocarp fruits, paniculate inflorescences
with opposite umbellules, pentamorous flowers with 2-5 carpels and an articulated pedicel,
and palmately compound leaves (mostly in juvenile leaves in Raukaua) (Lowry, Plunkett &
Wen 2004). The current study also demonstrates a close relationship between Cheirodendron
and Raukaua. In particular, the chloroplast phylogeny has C. bastardianum nested within
(BI) or sister to (MP and ML) Raukaua. Although analysis of the chloroplast region resulted
in Cheirodendron being paraphyletic, combined analysis showed strong support for
monophyly of Cheirodendron and for C. bastardianum as the sister group to the Hawaiian
clade. Consistent with this relationship, C. bastardianum shares many morphological
characteristics with Hawaiian Cheirodendron, rather than Raukaua (Brown 1935) and looks
almost identical to C. trigynum (Frodin 1990).
Relationships among taxa
Phylogenetic analysis revealed Cheirodendron as monophyletic, with C.
bastardianum (from Marquesas) sister to the Hawaiian species. Within the Hawaiian species,
two clades are supported, a Kauai taxa clade and a non-Kauai taxa clade. Both the Kauai
clade and non-Kauai clade are unresolved, representing little or no gene sequence divergence
among these taxa. Thus, fine-tuned phylogenetic relationships cannot be inferred at this time.
Further study at the population level might be of help in sorting out infraspecific
relationships. However, within the scope of this project, this is not possible.
Short-branching polytomies within the Kauai clade and non-Kauai clade agree with a
similar pattern of evolution found in other Pacific Island groups. Many Hawaiian radiations

35
exhibit great degrees of morphological diversity with little genetic variation (Wagner & Funk
1995). For example, the Hawaiian Cyrtandra (Gesneriaceae) is composed of 58 species, all
resultant from a single ancestor and exhibits little infraspecific genetic variation (Cronk et al.
2005). Hawaiian Bidens L. (Asteraceae) is one of the most species-rich radiations within the
genus, but has little differentiation in both nuclear and chloroplast regions (Knope et al.
2012). Many other examples, including the Hawaiian mints (Lindqvist et al. 2003),
Silverswords (Carlquist, Baldwin & Carr 2003), Lobeliads (Givnish et al. 2009) and
Coprosma (Cantley et al. 2014) have demonstrated great morphological diversity with little
genetic differentiation.
The lack of genetic divergence within Kauai taxa and non-Kauai taxa, despite
morphological differentiation, could be explained by a recent and rapid radiation of the
group. Previous analyses have estimated the divergence rates of angiosperms to range from
0.078 to 0.091 net speciation events per million years (Magallon & Castillo 2009) which is
considerably slower than other Hawaiian groups. Hawaiian Bidens was estimated to have a
diversification rate of 0.3-2.3 species per million years (Knope et al. 2012). Baldwin &
Sanderson (1998) estimate the diversification rate of the Hawaiian Silverswords to be 0.56 ±
0.17 species per million years. Rapid speciation rates seen in other groups within Hawaii
could be mirrored in Cheirodendron, although no analyses have estimated the age and rate of
diversification of the genus. Further study using a molecular clock might give some insight
as to the time frame and diversification rate of Cheirodendron.
Unresolved species relationships and poor branch support could suggest that
Cheirodendron is composed of only a few species with a great deal of morphological
variation and that phenotypic plasticity might be the cause of variation in form. Phenotypic

36
plasticity is described as “the ability of an individual to express different features under
different environmental conditions” (Travis 2009). To some extent, different morphologies
might be the result of phenotypic plasticity in Cheirodendron, especially in C. trigynum,
which exhibits local population differences (see Sherff 1954). However, phenotypic plasticity
does not explain the morphological differences in taxa on Kauai that persist adjacent to one
another. On Kauai, C. trigynum ssp. helleri can be found in lowland bogs to mesic forests
(440-1250 m) (Lowry 1990), overlapping with most Kauai species, including C. fauriei,
which also occurs in low-elevation bogs (Lowry 1990). Although these two species can be
found in the same ecological conditions, they still maintain separate morphologies. If
plasticity relies heavily on environmental pressure to produce phenotypic differences,
different morphological taxa would not be present next to each other.
The study by Knope et al. (2012) on Hawaiian Bidens revealed that these species
consisted of closely related individuals with a great degree of morphological differences
among them. Common garden experiments showed that these differences were not plastic
traits, and forms held true to their phenotypes (Knope et al. 2012). Common garden
experiments to rule out phenotypic plasticity have not been done with Cheirodendron before,
thus phenotypic plasticity cannot be ruled out. However, it is more likely that the consistent
differences found among individuals in similar or the same habitats are genetically based
rather than a plastic response to ecological pressures, although genetic differences among
them were not detected with the methods used in this study.

37
Current taxonomy vs. phylogenetic relationships
Phylogenetic analysis suggests that two taxa of Cheirodendron presently regarded as
subspecies (Lowry 1990) are to be recognized as distinct species. Cheirodendron trigynum
ssp. helleri and C. platyphyllum ssp. kauaiense (both in the Kauai clade) are polyphyletic
relative to their respective species (in the non-Kauai clade). This strongly supports the
reclassification of subspecific taxa presently recognized in either C. trigynum or C.
platyphyllum that they do not belong to the same species. As such, C. trigynum ssp. helleri
(Sherff) Lowry is now recognized as C. helleri Sherff and C. platyphyllum ssp. kauaiense
(Kraj.) Lowry is now recognized as C. kauaiense Kraj.
As far as the assessments of Sherff (1954), Herat (1981) and Lowry (1990), each
hinted that the vast morphological diversity present within the Hawaiian Cheirodendron
could represent more taxa. In his revision, Lowry (1990) takes note of the many different
varieties seen by Sherff, stating that “some of these [populations] may represent genetically-
based differences and could warrant taxonomic recognition.” This statement is supported for
some taxa in the current study (see above), but is not supported for other species differences
as Lowry (1990) suggests due to the lack of resolution within the Kauai and non-Kauai
clades.
Disagreement between the molecular phylogeny and taxonomy could be caused by
certain traits being convergent within Cheirodendron, the result masking the true
evolutionary relationships. Convergent evolution is the process whereby similar characters
arise independently in different lineages rather than as a result of common ancestry (Grande
& Rieppel 1994). These certain characters are said to be homoplasic, and can result from
adaptive responses to similar selection pressures (Wake, Wake & Specht 2011). Homoplasy

38
and convergent evolution present a problem in phylogenetics because it gives a false sense of
what the true relationships are. For example, thorns, spines and prickles are evolutionarily
and ontogenetically different structures, though they arose from the same selective pressure
to serve the same purpose—defense. If, then, this characteristic (having sharp protrusions for
protection) was used to infer phylogentic relationships, Rosaceae (with prickles), Cactaceae
(with spines) and Rutaceae (with thorns), would infer a close relationship, however, this is
definitely not the case (see Soltis et al. 2005).
Convergence in morphology could also be occurring in Cheirodendron. If the
characters used to classify species were homoplasic due to convergent evolution, species
delineations would not agree with phylogenetic relationships. Lowry’s (1990) classification
of C. trigynum with two subspecies was based on indistinguishable characteristics in leaf
morphology. Lowry’s (1990) ssp. helleri and ssp. trigynum are readily distinguished from
each other based on carpel number, with ssp. helleri having two carpels and ssp. trigynum
usually having three or occasionally four, although ssp. helleri is found to have three carpels
in some individuals. This gives additional credibility to the separation of these as distinct
species as noted above.
The question of why certain characters such as leaf morphology converged in
different taxa is an interesting question that cannot be answered within the scope of this
project. All C. trigynum share the same leaflet morphology (the most commonly recognized
features being leaflets usually 3 to 5 and longer than wide, a large central leaflet, and margins
thickened), and occur in similar habitats on their respective islands. Perhaps convergence on
this particular leaf morphology has provided some ecological advantage to allow this form to
occur in different lineages.

39
Biogeographic relationships
Fosberg (1948) was the first to suggest that the origins of Cheirodendron were from a
single colonization event from the Austral region. Decades later, his ideas were confirmed
with molecular data, supporting that Cheirodendron had an Austral origin and had moved
northward via long-distance dispersal (Wen et al. 2001, Plunkett, Wen & Lowry 2004).
Recent studies have also shown that Raukaua, a paraphyletic group distributed in New
Zealand, Chile, Argentina and Tasmania, is Cheirodendron’s closest relative. The species
sister to Cheirodendron are the New Zealand species of Raukaua: R. edgerleyi, R. anomalus,
and R. simplex (Mitchell et al. 2012). Although there has been much recent information about
the infrafamilial biogeographic relationships within Araliaceae, as well as the infrageneric
biogeographic relationships of Raukaua, there has been no detailed phylogenetic work to
assess these relationships within Cheirodendron until the present study.
Herat (1981) suggested a single colonization event from New Zealand to Hawaii with
a subsequent dispersal event to the Marquesas (based on morphological similarities).
Although much less commonly seen, dispersal events originating from Hawaii have been
confirmed through molecular work (see Harbaugh & Baldwin 2007, Harbaugh et al. 2009a).
However, this pattern of dispersal does not follow traditional ideas about biogeography in the
Pacific. Hennig (1966) proposed the concept of stepping stone biogeography patterns, where
a given organism with a continental origin disperses to the next closest landmass (sometimes
islands), evolutionarily “stepping” from landmass to landmass. This concept is especially
embraced in Pacific biogeography when life on island archipelagos would have occurred via
long distance dispersal along with in situ speciation. Evidence of this is well-supported by

40
many Pacific examples including Metrosideros (Wright et al. 2001), Astelia (Birch & Keeley
2013), and Coprosma (Cantley et al. 2014).
The biogeographic relationships of Cheirodendron inferred from this study suggest a
stepping stone pattern of dispersal from New Zealand to Marquesas, and Marquesas to
Hawaii. As C. bastardianum and the Hawaiian clade have a sister relationship, an alternative
could be that the most recent common ancestor could have colonized both the Marquesas and
Hawaiian Islands simultaneously. However, the previous hypothesis of Herat (1981) that C.
bastardianum is descended from a colonist from the Hawaiian Islands can be discarded.
The biogeographic relationships implications of seven endemic species in the
Hawaiian Islands cannot be fully resolved at this time. Two Hawaiian clades, the Kauai clade
and non-Kauai clade are sister to each other and could represent two simultaneous dispersal
events from Marquesas to Hawaii. The biogeography of Cheirodendron within the Hawaiian
Islands cannot be inferred from this study, although Herat (1981) suggested that
Cheirodendron in Hawaii originated on Kauai, since Kauai appears to be the center of
diversity and has the highest number of species. The lack of resolution within the two
Hawaiian clades suggests a recent and rapid radiation of these species.
Future directions
Although this study has shed light on the evolution and biogeography of
Cheirodendron, there are many questions that still remain. Are the forms of Cheirodendron a
result of phenotypic plasticity? What are the phylogenetic relationships among Hawaiian
species? What are the biogeographic implications of species within the Hawaiian Islands?
Can species relationships be resolved or are there underlying factors that limit our

41
understanding of the group? Perhaps Cheirodendron is part of a species complex, much like
the unresolved species relationships of Metrosideros (see Harbaugh et al. 2009b).
The many challenges in sorting out species relationships have also been opportunities
in deepening our understanding of the evolution of Cheirodendron. New techniques and
methods to help answer these questions such as next generation sequencing, microsatellites,
and restriction site associated DNA (RAD) sequencing may help in sorting out these
relationships. Not only will these answers contribute to our understanding of Cheirodendron,
but will advance our knowledge about the radiation and speciation of other Pacific groups.
Ultimately, this knowledge will provide valuable information for discerning relationships
among all living things.

42
CHAPTER 3. SYNTHESIS- HYPOTHESES REVISITED
The conclusions of this study allow for the acceptance of two hypotheses and
rejection of one hypothesis. The following discusses these conclusions as they relate to each
individual hypothesis:
Hypothesis one: The molecular phylogeny does not support the current taxonomy of
Cheirodendron.
Conclusion: Accept hypothesis
Results of this study found disagreement between the molecular phylogeny and
current taxonomy based on the placement of two Hawaiian subspecies. Based on these
results, it is evident that species with subspecies recognized on different islands are not
closely related and it is necessary to elevate the subspecies C. trigynum ssp. helleri to C.
helleri Sherff and C. platyphyllum ssp. kauaiense to C. kauaiense Kraj. There are seven
Hawaiian species and one Marquesan species that should be recognized, as opposed to the
current classification with five Hawaiian species and one Marquesan species.
Hypothesis two: Cheirodendron is monophyletic.
Conclusion: Accept hypothesis
This study shows evidence that Cheirodendron is a monophyletic lineage, consisting
of C. bastardianum of the Marquesas along with Hawaiian species, C. trigynum, C. helleri,
C. platyphyllum, C. kauaiense, C. forbesii, C. fauriei and C. dominii. Cheirodendron has a
sister relationship to New Zealand species of Raukaua, a paraphyletic genus with species in
New Zealand, Tasmania, Chile and Argentina.

43
Hypothesis three: The Pacific biogeography of Cheirodendron involves an Austral origin
with long distance dispersal to Hawaii and then to the Marquesas.
Conclusion: Reject hypothesis
Phylogenetic analyses show Cheirodendron as a monophyletic clade of Austral
origin. However, C. bastardianum has a sister relationship to the Hawaiian species rather
than being derived from Hawaiian species. This infers that either two dispersal events
simultaneously occurred from an Austral origin to Hawaii and the Marquesas, or dispersal
occurred to Hawaii via Marquesas. There is no supporting data that suggests colonization of
Marquesas via Hawaii.

44
LITERATURE CITED
Abbott, I. A. 1992. Lāʻau Hawaiʻi: Traditional Hawaiian Uses of Plants. Bishop Museum
Press, Honolulu.
Baldwin, B. G. 1992. Phylogenetic utility of the internal transcribed spacers of nuclear
ribosomal DNA in plants: An example from the Compositae. Molecular
Phylogenetics and Evolution 1(1): 3-16.
Baldwin, B. G. & M. J. Sanderson. 1998. Age and rate of diversification of the Hawaiian
silversword alliance (Compositae). Proceedings of the National Academy of Sciences
95(16): 9402-9406.
Barraclough, T. G. & S. Nee. 2001. Phylogenetics and Speciation. Trends in Ecology and
Evolution 16(7): 391-399.
Bazinet, A. L., D. J. Zwickl & M. P. Cummings. 2014. A gateway for phylogenetic analysis
powered by grid computing featuring GARLI 2.0. Systematic Biology. Advance
Access published online April 30, 2014. doi:10.1093/sysbio/syu031
Birch, J. L. & S. C. Keeley. 2013. Dispersal pathways across the pacific: the historical
biogeography of Astelia s.l. (Asteliaceae, Asparagales). Journal of Biogeography 40:
1914–1927.
Brown, F. B. H. 1935. Flora of southeastern Polynesia III. Dicotyledons. Bernice P. Bishop
Museum Bulletin 130: 1–386.
Cantley, J. T., N. G. Swenson, A. Markey & S. C. Keeley. 2014. Biogeographic insights on
Pacific Coprosma (Rubiaceae) indicate two colonizations to the Hawaiian Islands.
Botanical Journal of the Linnaean Society 174(3): 412-424.
Carlquist, S. 1974. Island Biology. Columbia University Press, New York.

45
Carlquist, S., B. G. Baldwin & G. D. Carr. (Eds.) 2003. Tarweeds and silverswords:
evolution of the Madiinae (Asteraceae). Missouri Botanical Garden Press, Saint
Louis.
Costello, A. & T. Motley. 2001. Molecular systematics of Tetraplasandra, Munroidendron
and Reynoldsia sandwicensis (Araliaceae) and the evolution of superior ovaries in
Tetraplasandra. Edinburgh Journal of Botany 58: 229–242.
Couper, R. A. 1960. New Zealand Mesozoic and Cainozoic plant microfossils. New Zealand
Geological Survey Paleontological Bulletin 32: 1-87.
Cronk, Q.C.B., M. Kiehn, W. L. Wagner & J. F. Smith. 2005. Evolution of Cyrtandra
(Gesneriaceae) in the Pacific Ocean: The origin of a supertramp clade. American
Journal of Botany 92(6): 1017-1024.
Darriba, D., G. L. Taboada, R. Doallo & D. Posada. 2012. jModelTest 2: more models, new
heuristics and parallel computing. Nature Methods 9(8): 772.
De Candolle, A. P. 1830. Araliaceae. Prodromus systematis naturalis regut vegetabilis 4:
251-266. Paris.
Doyle, J. J. & J. L. Doyle. 1987. A rapid DNA isolation procedure for small amounts of fresh
leaf tissue. Phytochemistry Bulletin, Botanical Society of America 19: 11–15.
Dunbar-Co, S., A. M. Wieczorek & C. W. Morden. 2008. Molecular phylogeny and adaptive
radiation of the endemic Hawaiian Plantago species (Plantaginaceae). American
Journal of Botany 95(9): 1177-1188.
Eibl J., G. M. Plunkett & P. P. Lowry II. 2001. Phylogenetic relationships in Polyscias sect.
Tieghemopanax (Araliaceae) based on DNA sequence data. Adansonia 3(23): 23–48.

46
Felsenstein, J. 1985. Confidence limits on phylogenies: An approach using the bootstrap.
Evolution 39: 783-791.
Fosberg, F. R. 1948. Derivation of the flora of the Hawaiian Islands. In E. C. Zimmerman
(Ed.), Insects of Hawaii, vol. 1, pp. 107-119. University Press Hawaii, Honolulu.
Frodin, D. G. 1990. Identity of Aralia bastardiana Decaisne. Pacific Science 44(3): 265-276.
Gillett, G. W. 1972. Genetic affinities between Hawaiian and Marquesan Bidens
(Asteraceae). Taxon 21(4): 479-483.
Givnish, T. J., K. C. Millam, A. R. Mast, T. B. Patterson, T.J. Theim, A. L. Hipp, J. M.
Henss, J. F. Smith, K. R. Wood & K. J. Sytsma. 2009. Origin, adaptive radiation and
diversification of the Hawaiian lobeliads (Asterales: Campanulaceae). Proceedings of
the Royal Society B: Biological Sciences 1656: 407–416.
Gorniak, M., O. Paun & M. W. Chase. 2010. Phylogenetic relationships within Orchidaceae
based on a low-copy coding gene, Xdh: Congruence with organellar and nuclear
ribosomal DNA results. Molecular Phylogenetics and Evolution 56: 784-795.
Grande, L. & O. Rieppel. 1994. Interpreting the hierarchy of nature. Academic Press, San
Diego.
Gray, A. 1854. United States Exploring Expedition under the command of Charles Wilkes,
U.S.N. Vol. 15. Botany Phanerogamia. Putnam & Co, New York.
Guindon, S. & O. Gascuel. 2003. A simple, fast and accurate method to estimate large
phylogenies by maximum-likelihood. Systematic Biology 52: 696-704.
Harbaugh, D. T. & B. Baldwin. 2007. Phylogeny and biogeography of the Sandalwoods
(Santalum, Santalaceae): Repeated dispersals throughout the Pacific. American
Journal of Botany 94(6): 1028- 1040.

47
Harbaugh, D. T., W. L. Wagner, G. J. Allan, & E. A. Zimmer. 2009a. The Hawaiian
Archipelago is a stepping stone for dispersal in the Pacific: an example from the plant
genus Melicope (Rutaceae). Journal of Biogeography 36: 230–241.
Harbaugh, D. T., W. L. Wagner, D. M. Percy, H. F. James, & R. C. Fleischer. 2009b. Genetic
Structure of the Polymorphic Metrosideros (Myrtaceae) Complex in the Hawaiian
Islands Using Nuclear Microsatellite Data. PLoS ONE 4(3): e4698.
Harrison, R. G. 1998. Linking evolutionary pattern and process: the relevance of species
concepts for the study of speciation. In: D. J. Howard, S. H. Berlocher (Eds.), Endless
Forms: Species and Speciation. Oxford University Press, New York.
Heller, A. A. 1897. Observations on the ferns and flowering plants of the Hawaiian Islands.
Minnesota Botanical Studies 58: 760-922.
Hennig, W. 1966. Phylogenetic Systematics. University of Illinois Press, Urbana.
Henwood, M. J. & J. M. Hart. 2001. Towards an understanding of the phylogenetic
relationships of Australian Hydrocotyloideae (Apiaceae). Edinburgh Journal of
Botany 58: 269–289.
Herat, R. M. 1981. A systematic study of the genus Cheirodendron (Araliaceae). Ph.D.
dissertation, University of Hawaii at Manoa, Honolulu, 246 pp.
Hillebrand, W. F. 1888. Flora of the Hawaiian Islands. Hafner Publishing Co. Reprint
edition, New York.
Hochreutiner, B. P. G. 1925. Plantae Hochreutineranae II. Candollea 2: 317-513.
Hooker, W. J. & W. G. Arnott. 1832. The Botany of Captain Beechey's Voyage. London.
Howarth, D. G. & D. A. Baum. 2002. Phylogenetic utility of a nuclear intron from

48
nitrate reductase for the study of closely related plant species. Molecular
Phylogenetics & Evolution 23: 525–528.
Howarth D. G. & D. A. Baum. 2005. Genealogical evidence of homoploid hybrid speciation
in an adaptive radiation of Scaevola (Goodeniaceae) in the Hawaiian Islands.
Evolution 59: 948–961.
Hueselenbeck, J. P. & F. Ronquist. 2001. MrBayes: Bayesian inference of phylogeny.
Biometrics 17: 754–755.
Kenfack, D. 2011. Resurrection in Carapa (Meliaceae): a reassessment of morphological
variation and species boundaries using multivariate methods in a phylogenetic
context. Botanical Journal of the Linnaean Society 165: 186-221.
Knope, M. L., C. W. Morden, V. A. Funk & T. Fukami. 2012. Area and the rapid radiation of
Hawaiian Bidens (Asteraceae). Journal of Biogeography 39(7): 1206-1216.
Krajina, V. 1931. Generis Cheirodendrum species hawaiiensis ex affinitate Cheirodendron
platyphylli. Pres1ia 10: 91-100.
Lee, D. E., W. G. Lee & N. Mortimer. 2001. Where and why have all the flowers gone?
Depletion and turnover in the New Zealand Cenozoic angiosperm flora in relation to
palaeogeography and climate. Australian Journal of Botany 49: 341–356.
Levin, R. A., J. Blanton & J. S. Miller. 2009. Phylogenetic utility of nuclear nitrate reductase:
A multi-locus comparison of nuclear and chloroplast sequence data for inference of
relationships among American Lycieae (Solanaceae). Molecular Phylogenetics &
Evolution 50: 608-617.
Lindqvist, C., T. J. Motley, J. J. Jeffrey & V. A. Albert. 2003. Cladogenesis and reticulation
in the Hawaiian endemic mints (Lamiaceae). Cladistics 19: 480–495.

49
Liu, Z., X. Zeng, D. Yang, G. Chu, Z. Yuan & S. Chen. 2012. Applying DNA barcodes for
identification of plant species in the family Araliaceae. Gene 499: 76-80.
Lowry, P. P. 1990. Araliaceae. In W. L. Wagner, D. R. Herbst & S. H. Sohmer (Eds.)
Manual of the flowering plants of Hawaii, vol. 1, pp. 224-237. University of Hawaii
Press, Honolulu.
Lowry, P. P., G. M. Plunkett & J. Wen. 2004. Generic relationships in Araliaceae: looking
into the crystal ball. South African Journal of Botany 70(3): 382-392.
Magallon, S. & A. Castillo. 2009. Angiosperm diversification through time. American
Journal of Botany 96: 349–365.
Manen, J. F., A. Natali & F. Ehrendorfer. 1994. Phylogeny of Rubiaceae-Rubieae inferred
from the sequence of a cpDNA intergene region. Plant Systematics & Evolution 190:
195-211.
Matthews, S. & M. J. Donoghue. 1999. The root of angiosperm phylogeny inferred from
duplicate phytochrome genes. Science 286(5441): 947-950.
Mitchell, A. D., D. G. Frodin & M. J. Heads. 1997. Reinstatement of Raukaua, a genus of
Araliaceae centered in New Zealand. New Zealand Journal of Botany 35: 309–315.
Mitchell, A. D. & S. J. Wagstaff. 1997. Phylogenetic relationships of Pseudopanax species
(Araliaceae) inferred from parsimony analysis of rDNA sequence data and
morphology. Plant Systematics & Evolution 208: 121-138.
Mitchell A. D. & S. J. Wagstaff. 2000. Phylogeny and biogeography of the Chilean
Pseudopanax laetevirens. New Zealand Journal of Botany 38: 409-414.

50
Mitchell, A. D., P. B. Heenan, & A. M. Paterson. 2009. Phylogenetic relationships of
Geranium species indigenous to New Zealand. New Zealand Journal of Botany 47:
21-31.
Mitchell, A. D., R. Li, J. W. Brown, I. Schonberger & J. Wen. 2012. Ancient divergence and
biogeography of Raukaua (Araliaceae) and close relatives in the southern
hemisphere. Australian Systematic Botany 25: 432-446.
Morden, C. W., V. Caraway & T. J. Motley. 1996. Development of a DNA library for native
Hawaiian plants. Pacific Science 50: 324-335.
Olmstead, R. G. & J. A. Sweere. 1994. Combining data in phylogenetic systematics: An
empirical approach to using three molecular data sets in the Solanaceae. Systematic
Biology 43(4): 467-481.
Plunkett, G. M. & P. P. Lowry II. 2001. Relationships among ‘‘ancient araliads’’ and their
significance for the systematics of Apiales. Molecular Phylogenetics & Evolution 19:
259–276.
Plunkett, G. M., J. Wen & P. P. Lowry II. 2004. Infrafamilial classifications and characters in
Araliaceae: Insights from the phylogenetic analysis of nuclear (ITS) and plastid (trnL-
trnF) sequence data. Plant Systematics & Evolution 245: 1–39.
Plunkett, G. M., G. T. Chandler, P. P. Lowry II, S. M. Pinney & T. S. Sprenkle. 2004. Recent
advances in understanding Apiales and a revised classification. South African Journal
of Botany 70: 371-381.
Price, J. P. & W. L. Wagner. 2004. Speciation in Hawaiian angiosperm lineages: cause,
consequence, and mode. Evolution 58: 2185–2200.

51
Rambaut, A. 2012. FigTree: Tree figure drawing tool version 1.4.0. Institute of Evolutionary
Biology, University of Edinburgh.
Rock, J. F. 1913. The Indigenous Trees of the Hawaiian Islands. Pacific Tropical Botanical
Gardens, Lawai. Reprint edition.
St. John, H. 1959. Botanical novelties on the island of Niihau, Hawaiian Islands. Hawaiian
plant studies 25. Pacific Science 13: 156-190.
Schuettpelz, E. & K. M. Pryer. 2007. Fern phylogeny inferred from 400 leptosporangiate
species and three plastid genes. Taxon 56: 1037-1050.
Seemann, B. 1868. Revision of the natural order of Hederaceae L. Reeve & Co., London.
Shaw, J., E. Lickey, J. T. Beck, S. B. Farmer, W. Liu, J. Miller, K. C. Siripun, C. T. Winder,
E. E. Schilling & R. L. Small. 2005. The tortoise and the hare II: relative utility of 21
noncoding chloroplast DNA sequences for phylogenetic analysis. American Journal
of Botany 92: 142–166.
Shaw, J., E. B. Lickey, E. E. Schilling & R. L. Small. 2007. Comparison of whole chloroplast
genome sequences to choose noncoding regions for phylogenetic studies in
angiosperms: The tortoise and the hare III. American Journal of Botany 94(3): 275-
288.
Sherff, E. E. 1954. Revision of the genus Cheirodendron Nutt. ex Seem. for the Hawaiian
Islands. Fieldiana Botany: 29(1). Chicago Natural History Museum, Chicago, IL.
Sites, J. W. Jr. & J. C. Marshall. 2004. Operational criteria for delimiting species. Annual
Review of Ecology, Evolution & Systematics 35: 199–227.

52
Small, R. L. & J. F. Wendel. 2000. Phylogeny, duplication, and intraspecific variation of Adh
sequences in new world diploid cottons (Gossypium L., Malvaceae). Molecular
Phylogenetics & Evolution 16(1): 73-84.
Soltis, D. E., P. S. Soltis, P. K. Endress & M. W. Chase. 2005. Phylogeny and evolution of
angiosperms. Sinauer Associates, Inc., Massachusetts.
Suzuki, Y., G. V. Glazco & M. Nei. 2002. Overcredibility of molecular phylogenies obtained
by Bayesian phylogenetics. Proceedings of the National Academy of Science 99:
16138-16143.
Taberlet, P., L. Gielley, G. Pautou & J. Bouvet. 1991. Universal primers for amplification of
three non-coding regions of chloroplast DNA. Plant Molecular Biology 17: 1105–
1109.
Tamura, K., G. Stecher, D. Peterson, A. Filipski & S. Kumar. 2013. MEGA6: Molecular
Evolutionary Genetics Analysis version 6.0. Molecular Biology & Evolution 30:
2725-2729.
Travis, J. 2009. Phenotypic Plasticity. In S. A. Levin (Ed.), The Princeton Guide to Ecology,
pp. 65-71. Princeton University Press, Princeton.
Udovicic, F., G. I. McFadden & P. Y. Ladiges. 1995. Phylogeny of Eucalyptus and
Angophora based on 5S rDNA spacer sequence data. Molecular Phylogenetics &
Evolution 4: 247-256.
Wagner, W. L. & V. A. Funk (Eds.). 1995. Hawaiian biogeography: Evolution on a hot spot
archipelago. Smithsonian Institution Press, Washington D. C.
Wagner, W. L. & D. H. Lorence. 2002. Flora of the Marquesas Islands website.
http://botany.si.edu/pacificislandbiodiversity/marquesasflora/index.htm.

53
Wake, D. B., M. H. Wake, & C. D. Specht. 2011. Homoplasy: From Detecting Pattern to
Determining Process and Mechanism of Evolution. Science 331(6020): 1032-1035.
Wen, J., & E. A. Zimmer. 1996. Phylogeny and biogeography of Panax L. (the ginseng
genus, Araliaceae): Inferences from ITS sequences of nuclear ribosomal DNA.
Molecular Phylogenetics & Evolution 6: 167–177.
Wen, J., G. M. Plunkett, A. D. Mitchell & S. J. Wagstaff. 2001. The Evolution of Araliaceae:
A Phylogenetic Analysis Based on ITS Sequences of Nuclear Ribosomal DNA.
Systematic Botany 26(1): 144-167.
Wright, S. D., C. G. Yong, S. R. Wichman, J. W. Dawson, & R. C. Gardner. 2001. Stepping
stones to Hawaii: A trans-equatorial dispersal pathway for Metrosideros (Myrtaceae)
inferred from nrDNA (ITS + ETS). Journal of Biogeography 28(6): 769-774.
Yi, T., P. P. Lowry, G. M. Plunkett & J. Wen. 2004. Chromosomal evolution in Araliaceae
and close relatives. Taxon 53(4): 987-1005.
Zimmer, E. A. & J. Wen. 2012. Using nuclear gene data for plant phylogenetics: Progress
and prospects. Molecular Phylogenetics & Evolution 65: 774-785.

















