Phylogenetic analysis of the VP1 gene of Enterovirus 71 in Guangzhou during the high occurrence...
5
Phylogenetic analysis of the VP1 gene of Enterovirus 71 in Guangzhou during the high occurrence period of 2008 Ying-xian Yin • Zhi-ying Ou • Yi Xu • Rong Zhou • Hui-min Xia Received: 22 September 2013 / Accepted: 27 January 2014 / Published online: 11 February 2014 Ó Springer Science+Business Media New York 2014 Abstract An outbreak of hand, foot, and mouth disease (HFMD) in Guangzhou in 2008 affected over 10,000 children and resulted in high hospital admission rates. To investigate the molecular epidemiological pattern of EV71 infections in Guangzhou, throat swab samples were col- lected from 102 children clinically diagnosed with HFMD from May to July of 2008 in Guangzhou. Partial VP1 (virus protein 1) fragments of Enterovirus 71 (EV71) isolates were sequenced, and used alongside EV71 sequences entered in GenBank to construct a phylogenetic tree using MEGA5.0. Blast and phylogenetic analyses showed that all 21 sequences belonged to subgenogroup C4 of EV71. In early May, diverse strains were circulating in Guangzhou, but by July, only a small number of these strains could be detected. These results could indicate that geographic and climatic features may affect the epidemic characteristics of EV71, and that some C4 strains might retain their infec- tivity at higher temperatures. Keywords Enterovirus 71 VP1 gene Phylogenetic analysis Molecular epidemiology Introduction Human enterovirus 71 (EV71) is a major causative agent of hand, foot, and mouth disease (HFMD), and compared with other types of Coxsackie virus A16, more frequently causes complications of the central nervous system. The rapid identification and classification of enteroviruses is needed for monitoring outbreaks of HFMD. Molecular diagnostic methods based on reverse-transcriptase polymerase chain reaction (RT-PCR) and sequencing techniques have become the new gold standard for HFMD diagnosis, offering greater sensitivity and specificity than traditional methods [1–4]. The genomic RNA of Enteroviruses comprises a single open reading frame (ORF) encoding a polyprotein (pro- ducing P1, P2, and P3) flanked by nontranslated regions (NTR) at the 5 0 and 3 0 ends. The 5 0 -NTR has been the most frequent target for RT-PCR detection, and for enterovirus classification [5]. Further typing of individual serotypes into serogroups and subgroups based on the 5 0 -NTR is problematic as it is highly conserved within serotypes. VP1 offers an attractive alternative to the 5 0 -NTR as it contains major antigenic epitopes, and has been found to be highly variable among enteroviruses. In addition, high correla- tions between VP1 nucleotide sequences and serotyping, as determined by neutralization tests, have been documented, indicating that the VP1 gene could be used as a determinant for typing of human enteroviruses. This molecular dis- tinction between serotypes is very useful for the typing and classification of enterovirus isolates in the clinical labora- tory [6, 7]. EV71 has been reported in many countries and regions, particularly the Western Pacific Region [8–11]. During the first six months of 2008, outbreaks of HFMD were reported across several provinces in China. The initial cases of Electronic supplementary material The online version of this article (doi:10.1007/s11262-014-1046-z) contains supplementary material, which is available to authorized users. Y. Yin (&) Z. Ou Y. Xu H. Xia Institute of Pediatrics, Guangzhou Women & Children’s Medical Center, Guangzhou 510623, China e-mail: [email protected] R. Zhou Guangzhou Institute of Respiratory Diseases, Guangzhou 510230, China 123 Virus Genes (2014) 48:538–542 DOI 10.1007/s11262-014-1046-z
Transcript of Phylogenetic analysis of the VP1 gene of Enterovirus 71 in Guangzhou during the high occurrence...

Phylogenetic analysis of the VP1 gene of Enterovirus 71in Guangzhou during the high occurrence period of 2008
Ying-xian Yin • Zhi-ying Ou • Yi Xu •
Rong Zhou • Hui-min Xia
Received: 22 September 2013 / Accepted: 27 January 2014 / Published online: 11 February 2014
� Springer Science+Business Media New York 2014
Abstract An outbreak of hand, foot, and mouth disease
(HFMD) in Guangzhou in 2008 affected over 10,000
children and resulted in high hospital admission rates. To
investigate the molecular epidemiological pattern of EV71
infections in Guangzhou, throat swab samples were col-
lected from 102 children clinically diagnosed with HFMD
from May to July of 2008 in Guangzhou. Partial VP1 (virus
protein 1) fragments of Enterovirus 71 (EV71) isolates
were sequenced, and used alongside EV71 sequences
entered in GenBank to construct a phylogenetic tree using
MEGA5.0. Blast and phylogenetic analyses showed that all
21 sequences belonged to subgenogroup C4 of EV71. In
early May, diverse strains were circulating in Guangzhou,
but by July, only a small number of these strains could be
detected. These results could indicate that geographic and
climatic features may affect the epidemic characteristics of
EV71, and that some C4 strains might retain their infec-
tivity at higher temperatures.
Keywords Enterovirus 71 � VP1 gene � Phylogenetic
analysis � Molecular epidemiology
Introduction
Human enterovirus 71 (EV71) is a major causative agent of
hand, foot, and mouth disease (HFMD), and compared with
other types of Coxsackie virus A16, more frequently causes
complications of the central nervous system. The rapid
identification and classification of enteroviruses is needed
for monitoring outbreaks of HFMD. Molecular diagnostic
methods based on reverse-transcriptase polymerase chain
reaction (RT-PCR) and sequencing techniques have
become the new gold standard for HFMD diagnosis,
offering greater sensitivity and specificity than traditional
methods [1–4].
The genomic RNA of Enteroviruses comprises a single
open reading frame (ORF) encoding a polyprotein (pro-
ducing P1, P2, and P3) flanked by nontranslated regions
(NTR) at the 50 and 30 ends. The 50-NTR has been the most
frequent target for RT-PCR detection, and for enterovirus
classification [5]. Further typing of individual serotypes
into serogroups and subgroups based on the 50-NTR is
problematic as it is highly conserved within serotypes. VP1
offers an attractive alternative to the 50-NTR as it contains
major antigenic epitopes, and has been found to be highly
variable among enteroviruses. In addition, high correla-
tions between VP1 nucleotide sequences and serotyping, as
determined by neutralization tests, have been documented,
indicating that the VP1 gene could be used as a determinant
for typing of human enteroviruses. This molecular dis-
tinction between serotypes is very useful for the typing and
classification of enterovirus isolates in the clinical labora-
tory [6, 7].
EV71 has been reported in many countries and regions,
particularly the Western Pacific Region [8–11]. During the
first six months of 2008, outbreaks of HFMD were reported
across several provinces in China. The initial cases of
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11262-014-1046-z) contains supplementarymaterial, which is available to authorized users.
Y. Yin (&) � Z. Ou � Y. Xu � H. Xia
Institute of Pediatrics, Guangzhou Women & Children’s Medical
Center, Guangzhou 510623, China
e-mail: [email protected]
R. Zhou
Guangzhou Institute of Respiratory Diseases,
Guangzhou 510230, China
123
Virus Genes (2014) 48:538–542
DOI 10.1007/s11262-014-1046-z

HFMD caused by EV71 infection were reported in Fuyang,
Anhui Province, East China. This was followed by a high
occurrence of HFMD in Guangzhou, Guangdong Province,
South China. In the current report, we studied the molec-
ular epidemiology of EV71 infection among young chil-
dren in Guangzhou during the high occurrence period of
2008, via analyzing samples from 102 pediatric patients at
the Guangzhou Children’s Hospital with clinical HFMD,
and who were suspected as enterovirus infection between
May 2008 and July 2008.
VP1 sequences of 50 EV71 strains obtained from Gen-
Bank were included in this analysis, allowing the genera-
tion of dendrograms containing 71 EV71 strains. These
strains were isolated from the United States, Japan, Tai-
wan, Malaysia, Singapore, mainland China, Bulgaria,
Hungary, and the United Kingdom. Homology searches
were carried out using the BLAST program [http://www.
ncbi.nlm.nih.gov/BLAST]. Multiple-sequence alignments
were performed with CLUSTALW [http://www.ebi.ac.uk/
Tools/clustalw2/index.html] [12].
The alignment outputs were converted to MEGA format,
and phylogenetic analyses conducted using MEGA version
5.0 [13]. Dendrograms were constructed using the neighbor-
joining (NJ) and maximum parsimony (MP) algorithms in
the MEGA program. Statistical significance of phylogenies
was estimated by bootstrap analysis with 1,000 pseudore-
plicates. Nucleotide sequences GZCH01–GZCH21 newly
obtained in this study have been submitted to GenBank and
accession numbers assigned from FJ377563–FJ377583
(Table S1).
Total RNA was extracted from soaking solution of
throat swabs using TRIZOL (Invitrogen). RT-PCR was
successfully performed using EV71 specific primers (50-TCT GTC ACC CTT GTA ATA CCA TGG AT-30, 50-TAR GCR CTC GCR GGT GAC ATG AA-30; R = A, G),
generating a DNA product of 834 bp, visualized on a 2 %
agarose gel. The 834-bp fragment was observed for 21 of
the 102 (20.6 %) specimens obtained from clinically
diagnosed HFMD cases (Fig. S1). Of these 21 samples, 13
were collected in May, two in June, and four in July (Table
S1). BLAST-n was used to align these 21 sequences with
EV71 sequences extracted from GenBank. The alignment
data showed that all 21 sequences belonged to EV71. With
the exception of GZCH11, which was almost similar to
EV71/GDFS/3/2008, all the remaining sequences shared
the greatest sequence homology with isolates from Fuyang,
Anhui Province. The phylogenetic relationship—between
42 EV71 strains, including worldwide reference strains,
and the 21 clinical samples used in this study—was
deduced by the NJ method based on the partial VP1 coding
region (nucleotide position 2,438–2,947). The worldwide
reference strains were representative of most strains of
genogroups A, B, and C. BrCr was the single prototype of
genogroup A. Strains belonging to genogroups B and C had
been in circulation in recent years in various parts of the
world. Phylogenetic analysis of the 21 clinical samples and
typical EV71 isolates was based on the alignment of partial
VP1 gene sequences (510 nucleotides), and the construc-
tion of phylogenetic trees using the NJ method. CA16-G10
was used as the outgroup. The phylogenetic tree shows that
all 21 sequences belong to subgenogroup C4 (Fig. S2). The
high bootstrap value (99 %) for the 21 clinical samples
with Shzh04-12 and Shzh04-38 supports that they should
be clustered into the C4 subgenogroup. Major lineage of
genogroup and subgenogroup in this tree is apparent with
high bootstrap values, which is in accordance with pub-
lished data [3, 14, 15].
To further reveal the phylogenetic relatedness among
these 21 clinical samples and other isolates of subgeno-
group C4, particularly those from mainland China, another
tree was constructed using NJ and MP methods (Fig. 1). In
this analysis, EV71 shzh98 (AF302996), the first isolate
reported in mainland China and belonging to subgenogroup
C4, was used as the outgroup. The two trees have similar
topologic structure. GZCH11 has a high bootstrap value
with EV71/GDFS/3/2008 (99 %), an isolate from Guang-
dong Province during the same period of 2008. The
remaining 20 clinical samples clustered into a clade with
other isolates from mainland China, supported by a boot-
strap value of 70 %. In the current study, phylogenetic
analysis demonstrated that epidemic strains of EV71 iso-
lated in Guangzhou could be grouped into subgenogroup
C4. GZCH11 (FJ377573) was most closely related to
EV71/GDFS/3/2008 and an outlier of the C4 subgeno-
group, but the high bootstrap value (91 %) strongly sug-
gests that it should be included in the C4 subgenogroup
(Fig. 1). GZCH12, GZCH14, GZCH09, GZCH03,
GZCH01, GZCH08, and GZCH19 form a single clade with
an 83 % bootstrap. GZCH13, GZCH15, and GZCH16
cluster as a single clade with GZCH04, and are more
closely related to strains Zhuhai-171 and Zhuhai-152 than
to other strains.
HFMD caused by EV71 has been reported sporadically
in many parts of mainland China since 1998 [15]. Localized
outbreaks of HFMD at geographically distinct locations
occur annually in China, but before 2008, simultaneous
outbreak of HFMD across several provinces in one season
had not been reported. In 2008, isolates of EV71 from
several provinces had been sent to the Center for Disease
Control and Prevention of China for further study. How-
ever, little is understood about the relationship between the
2008 outbreak in Guangzhou and other epidemics across
China. Sequencing of the 50-NTR region of samples of
children clinically diagnosed with HFMD in the early out-
break of 2008, among whom 81.4 %(83/102) were positive
for 50-NTR amplification, showed some samples strongly
Virus Genes (2014) 48:538–542 539
123

identified with Fuyang/17.08/3, an isolate from Fuyang,
Anhui Province (data not shown). To further understand the
phylogenetic relationship of EV71 strains isolated from two
distant regions, the current study used partial VP1 sequen-
ces from 21 samples to construct phylogenetic trees with
reference sequences stored in GenBank.
According to phylogenetic analysis based on VP1
sequences, EV71 is currently divided into three genogroups:
A, B, and C [16]. BrCr prototype strain is the only member of
genogroup A. Most EV71 isolates belong to either geno-
group B or C, and they are further classified into five sub-
genogroups: B1–B5 and C1–C5 [9–11, 14, 17]. C4 first
reported as Shzh98 (AF302996) was isolated in Shenzhen,
China in 1998. It was debated whether the outbreaks in
mainland China during 2008 were linked to the 2004 C4
outbreaks in Taiwan. The two methods used in the
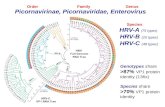
Fig. 1 Dendrograms constructed using the neighbor-joining (a) and
maximum parsimony (b) methods showing the genetic relationships
among 21 clinical EV71 strains collected from Guangzhou Children’s
Hospital, and reference sequences of genogroup C retrieved from
GenBank. The alignments were based on partial VP1 gene sequences.
Branch lengths are proportional to the number of nucleotide
differences. The bootstrap values in 1,000 pseudoreplicates for major
lineages within the tree are shown as percentages. Values less than
70 % were not shown. The marker denotes a measurement of relative
phylogenetic distance. Shzh98 belonging to C4 subgenogroup was
used as the outgroup in this analysis. The date in parenthesis tells only
the date of samples collected, not the date of strains. The different
months of samples collected were colored (green for May, orange for
June, and red for July) (Color figure online)
540 Virus Genes (2014) 48:538–542
123

construction of phylogenetic trees in the current study show
similar topology (Fig. 1). Despite some differences in the
tree twigs, the evolutionary hierarchy is almost identical.
Shzh98 at the tree roots can be considered as the ancestor of
the C4 subgenogroup. GZCH11 and EV71/GDFS/3/2008,
which are near the root, were isolated from two adjacent
regions of Guangdong Province.
It is of interest to note that for nearly ten years, all isolates
from mainland China belonged to subgenogroup C4, and that
their genotypes were not affected by epidemic situations in
adjacent regions including Japan, Taiwan, Singapore, and
Malaysia [15, 18–20]. In addition, neither genogroup A nor
B has been reported in mainland China since 2001. This
contrasts with Taiwan, where there was a more complex
epidemic pattern of EV71, where subgenogroups B1, B4, B5,
C2, C4, and C5 had been simultaneously predominant over
the last 10 years [17, 21–24]. These results indicate that
geographic and climatic features could affect the epidemic
characteristics of EV71, and specific subgenogroups might
be sustained in mainland China.
An interesting phenomenon encountered in this study
was the presence of multiple strains of EV71 cocirculating
during the early stages of outbreak compared with fewer
strains detected in clinical samples during the later stages
of outbreak. Our analysis showed a great diversity of EV71
strains belonging to subgenogroup C4 circulating in Gu-
angzhou in early May. Many of these isolates shared
similar identities to Zhuhai-213, Fuyang22, and Zhe-
jiang08, or with ancestral strains (according to the phylo-
genetic tree) C4, such as Shzh04-12. In contrast, in late
July, only isolates from the same branch of the phyloge-
netic tree as Fuyang/17.08/3 were detected in the Gu-
angzhou area. Isolates GZCH01, GZCH03, GZCH05,
GZCH06, GZCH07, GZCH08, GZCH09, GZCH10,
GZCH11, GZCH14, GZCH18, GZCH19, and GZCH21
were collected in early May 2008, while GZCH02,
GZCH04, GZCH17, and GZCH20 were collected in late
July (Fig. 1). GZCH02, GZCH04, GZCH13, GZCH15,
GZCH16, and GZCH20 clustered in a clade within the C4
subgenogroup, and were phylogenetically related to Zhu-
hai152 and Zhuhai171.
Many factors such as temperature, humidity, and
population mobility can affect the epidemiological pattern
of EV71; however, when environmental conditions are
kept the same for all isolates, theoretically, the differential
pattern for different isolates should be predominantly
attributed to genetic factors. Mutations in specific regions
of viral genomic RNA can alter virus sensitivity to tem-
perature [25]. Our previous sequencing results also
showed that no nucleotide mutations in position 251 of
the 3D polymerase region of all isolates obtained were
found. Other mutations besides this region might alter
sensitivity to temperature, while this task is time
consuming and needs a large number of samples to verify.
Based on the phylogenetic tree, it could be surmised that
from May to July, with a gradual rise in temperature in
Guangzhou, the higher temperature is optimal for isolates.
Urashima et al. argued the number of patients with
HFMD will increase if summer and autumn temperatures
increase [26]. An epidemiological study in Guangzhou
showed that each 1 �C rise in temperature corresponded
to an increase of 9�38 % in the weekly number of HFMD
cases [27]. Our findings seem to contradict their conclu-
sions, although those authors argue that their viewpoint is
based on seasonal models. The average temperature in
Guangzhou in June and July was higher than that in May.
Perhaps more importantly, the history climatological data
indicate that there were two temperature peaks greater
than 35 �C in July, one in June, and nil in May (Fig. S3).
In late July (the hottest time in Guangzhou) versus early
May, incidences of HFMD have decreased dramatically
(Table S3). In our study, the number of EV71 cases was
reduced in late July, as was the mean diversity of C4
subgenogroups of EV71(Fig. S3).
Other factors such as relative humidity, atmospheric
pressure, and wind velocity may also affect the epide-
miological pattern of HFMD; nevertheless, the most
likely climatological factor that affects the genetic
diversity should be temperature, because other factors
except temperature only contribute to the viability and the
spread of viruses, and influence of which is fair for all
isolates. Another driving force accelerating the evolution
of virus is host–pathogen interaction. However, this study
is hard to be carried out in very inadequate samples and
population. Moreover, in the first outbreak of EV71
infection, virus–host interaction including immune state is
relatively simpler than other enteroviruses, and so EV71
was utilized to study the temperature–genetic diversity in
our study.
Further studies on the role of temperature in virus sus-
ceptibility for different isolates of subgenogroup C4 will be
undertaken using cell culture and site-directed mutagenesis
of EV71 genes. Viability test in high temperature greater
than 35 �C for different isolates should be performed too.
Moreover, a greater number of isolates should be included
for more extensive epidemiologic studies.
Acknowledgments Financial support was provided by grants from
the Guangzhou Municipal Health Bureau (30307-2080048) and the
Guangdong Natural Science Foundation (s2012010008319).
References
1. S.C. Huang, Y.W. Hsu, H.C. Wang, S.W. Huang, D. Kiang, H.P.
Tsai, S.M. Wang, C.C. Liu, K.H. Lin, I.J. Su, J.R. Wang, Virus
Res. 131, 250–259 (2008)
Virus Genes (2014) 48:538–542 541
123

2. S. Singh, V.T. Chow, K.P. Chan, A.E. Ling, C.L. Poh, J. Virol.
Methods 88, 193–204 (2000)
3. P. McMinn, K. Lindsay, D. Perera, H.M. Chan, K.P. Chan, M.J.
Cardosa, J. Virol. 75, 7732–7738 (2001)
4. T. Vuorinen, R. Vainionpaa, T. Hyypia, Clinical infectious dis-
eases 37, 452–455 (2003)
5. G. Lee, C. Lee, J microbiol. 46, 319–324 (2008)
6. M.S. Oberste, K. Maher, D.R. Kilpatrick, M.R. Flemister, B.A.
Brown, M.A. Pallansch, J. Clin. Microbiol. 37, 1288–1293 (1999)
7. M.S. Oberste, W.A. Nix, K. Maher, M.A. Pallansch, J clin virol.
26, 375–377 (2003)
8. K.H. Lin, K.P. Hwang, G.M. Ke, C.F. Wang, L.Y. Ke, Y.T. Hsu,
Y.C. Tung, P.Y. Chu, B.H. Chen, H.L. Chen, C.L. Kao, J.R.
Wang, H.L. Eng, S.Y. Wang, L.C. Hsu, H.Y. Chen, J. Med. Virol.
78, 254–262 (2006)
9. K. Mizuta, C. Abiko, T. Murata, Y. Matsuzaki, T. Itagaki, K.
Sanjoh, M. Sakamoto, S. Hongo, S. Murayama, K. Hayasaka, J.
Clin. Microbiol. 43, 6171–6175 (2005)
10. H. Shimizu, A. Utama, N. Onnimala, C. Li, Z. Li-Bi, M. Yu-Jie,
Y. Pongsuwanna, T. Miyamura, Pediatrics international 46,
231–235 (2004)
11. P.V. Tu, N.T. Thao, D. Perera, T.K. Huu, N.T. Tien, T.C. Thu-
ong, O.M. How, M.J. Cardosa, P.C. McMinn, Emerg. Infect. Dis.
13, 1733–1741 (2007)
12. J.D. Thompson, D.G. Higgins, T.J. Gibson, Nucleic Acids Res.
22, 4673–4680 (1994)
13. K. Tamura, D. Peterson, N. Peterson, G. Stecher, M. Nei, S.
Kumar, Mol. Biol. Evol. 28, 2731–2739 (2011)
14. M.J. Cardosa, D. Perera, B.A. Brown, D. Cheon, H.M. Chan, K.P.
Chan, H. Cho, P. McMinn, Emerg. Infect. Dis. 9, 461–468 (2003)
15. L. Li, Y. He, H. Yang, J. Zhu, X. Xu, J. Dong, Y. Zhu, Q. Jin, J.
Clin. Microbiol. 43, 3835–3839 (2005)
16. B.A. Brown, M.S. Oberste, J.P. Alexander Jr, M.L. Kennett, M.A.
Pallansch, J. Virol. 73, 9969–9975 (1999)
17. Y.P. Huang, T.L. Lin, C.Y. Kuo, M.W. Lin, C.Y. Yao, H.W.
Liao, L.C. Hsu, C.F. Yang, J.Y. Yang, P.J. Chen, H.S. Wu, Virus
Res. 137, 206–212 (2008)
18. T. Munemura, M. Saikusa, C. Kawakami, H. Shimizu, M. Oseto,
A. Hagiwara, H. Kimura, T. Miyamura, Arch. Virol. 148,
253–263 (2003)
19. M.Y. Liu, W. Liu, J. Luo, Y. Liu, Y. Zhu, H. Berman, J. Wu,
PLoS ONE 6, e25287 (2011)
20. X. Tan, X. Huang, S. Zhu, H. Chen, Q. Yu, H. Wang, X. Huo, J.
Zhou, Y. Wu, D. Yan, Y. Zhang, D. Wang, A. Cui, H. An, W. Xu,
PLoS ONE 6, e25662 (2011)
21. K.T. Chen, H.L. Chang, S.T. Wang, Y.T. Cheng, J.Y. Yang,
Pediatrics 120, e244–e252 (2007)
22. S.H. Kung, S.F. Wang, C.W. Huang, C.C. Hsu, H.F. Liu, J.Y.
Yang, Clin microbiol infect. 13, 782–787 (2007)
23. T.N. Wu, S.F. Tsai, S.F. Li, T.F. Lee, T.M. Huang, M.L. Wang,
K.H. Hsu, C.Y. Shen, Emerg. Infect. Dis. 5, 458–460 (1999)
24. C. Yoke-Fun, S. AbuBakar, BMC Microbiol. 6, 74 (2006)
25. Y.H. Kung, S.W. Huang, P.H. Kuo, D. Kiang, M.S. Ho, C.C. Liu,
C.K. Yu, I.J. Su, J.R. Wang, Virology 396, 1–9 (2010)
26. M. Urashima, N. Shindo, N. Okabe, Jpn J infect dis. 56, 48–53
(2003)
27. Li T., Yang Z., Di B., and Wang M., Epidemiology and infection,
1-10, 2013
542 Virus Genes (2014) 48:538–542
123