
Perspectives on Pink Salmon and Sea Lice: Scientific Evidence
18
Downloaded By: [Canadian Research Knowledge Network] At: 22:02 4 May 2008 Reviews in Fisheries Science, 16(4):403–412, 2008 Copyright C Taylor and Francis Group, LLC ISSN: 1064-1262 print DOI: 10.1080/10641260801937131 Perspectives on Pink Salmon and Sea Lice: Scientific Evidence Fails to Support the Extinction Hypothesis KENNETH M. BROOKS 1 and SIMON R. M. JONES 2 1 Aquatic Environmental Sciences, Port Townsend, Washington, USA 2 Pacific Biological Station, Fisheries and Oceans Canada, Nanaimo, British Columbia, Canada Using a Ricker (1975) model and escapement data for a subset of Broughton Archipelago, British Columbia watersheds, Krkoˇ sek et al. (2007a) predicted that sea lice infections originating on Atlantic salmon (Salmo salar)farms will cause the extinction of pink salmon (Oncorhynchus gorbuscha) in the archipelago by 2010. The purpose of this article is to examine this issue in the context of all of the escapement data available for the archipelago and to review additional scientific reports and information not discussed by Krkoˇ sek et al. (2007a). Additional research during the last five years is not consistent with the Krkoˇ sek et al. (2007a) conclusion that sea lice routinely cause in excess of 80% mortality of fry. Rather, the literature reviewed herein indicates that pink salmon fry mount an effective immune response at sizes as small as 0.7 g, resulting in the rapid shedding of lice within two weeks. Pink salmon returns are shown to be highly variable throughout the Northeast Pacific in areas without salmon farms. Following periods of high abundance, pink salmon populations typically fall to low levels, and they may remain depressed for several generations. However, in most cases, the populations then gradually increase to begin the cycle anew. An examination of returns to all of the documented Broughton Archipelago watersheds indicates that following exceptionally high returns in 2000 and 2001, the populations declined to very low numbers in 2002 and 2003. Contrary to the conclusions reached by Krkoˇ sek et al. (2007a), Broughton pink salmon returns have steadily increased since then, with no indication that they are threatened with extinction. Other unsubstantiated assumptions used in Krkoˇ sek et al. (2007a) are also discussed in light of additional scientific reports and theoretical considerations. Keywords Atlantic salmon, aquaculture, netpens, pink salmon fry, sea lice INTRODUCTION Following the release in 2002 of the Pacific Fisheries Re- source Conservation Council Advisory document (PFRCC, 2002), intensive research on sea lice and their interaction with wild and farmed fish in the Broughton Archipelago of British Columbia has been underway. A series of studies claiming that salmon farms are the cause of sea lice (Lepeophtheirus salmonis and Caligus clemensi) infections on pink salmon (Oncorhynchus gorbuscha) fry in the archipelago have been published (Morton et al., 2004; Krkoˇ sek et al., 2005, 2006, 2007b). Krkoˇ sek et al. (2007a) now claim that sea lice originating on salmon farms Address correspondence to Kenneth M. Brooks, Aquatic Environmen- tal Sciences, 644 Old Eaglemount Road, Port Townsend, Washington, USA. E-mail: [email protected] will result in the extinction of Broughton pink salmon stocks in the near future. This review, which is intended to expand the discussion by describing other peer-reviewed papers and public information important to understanding the interaction between pink salmon, farmed fish, and sea lice, includes the following considerations: 1. The variability of pink salmon returns throughout the North Pacific with particular emphasis on the Broughton Archipelago. 2. The general absence of mortality among juvenile pink salmon following controlled laboratory exposure to Lepeophtheirus salmonis. 3. The absence of a cause-and-effect relationship between sea lice infecting pink salmon fry and larval lice released from salmon farms. 4. A reevaluation of trends in pink salmon abundance. 403
Transcript of Perspectives on Pink Salmon and Sea Lice: Scientific Evidence

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 2
2:0
2 4
May 2
008
Reviews in Fisheries Science, 16(4):403–412, 2008Copyright C©© Taylor and Francis Group, LLCISSN: 1064-1262 printDOI: 10.1080/10641260801937131
Perspectives on Pink Salmon and SeaLice: Scientific Evidence Fails toSupport the Extinction Hypothesis
KENNETH M. BROOKS1 and SIMON R. M. JONES2
1Aquatic Environmental Sciences, Port Townsend, Washington, USA2Pacific Biological Station, Fisheries and Oceans Canada, Nanaimo, British Columbia, Canada
Using a Ricker (1975) model and escapement data for a subset of Broughton Archipelago, British Columbia watersheds,Krkosek et al. (2007a) predicted that sea lice infections originating on Atlantic salmon (Salmo salar)farms will cause theextinction of pink salmon (Oncorhynchus gorbuscha) in the archipelago by 2010. The purpose of this article is to examinethis issue in the context of all of the escapement data available for the archipelago and to review additional scientific reportsand information not discussed by Krkosek et al. (2007a). Additional research during the last five years is not consistent withthe Krkosek et al. (2007a) conclusion that sea lice routinely cause in excess of 80% mortality of fry. Rather, the literaturereviewed herein indicates that pink salmon fry mount an effective immune response at sizes as small as 0.7 g, resulting in therapid shedding of lice within two weeks. Pink salmon returns are shown to be highly variable throughout the Northeast Pacificin areas without salmon farms. Following periods of high abundance, pink salmon populations typically fall to low levels,and they may remain depressed for several generations. However, in most cases, the populations then gradually increase tobegin the cycle anew. An examination of returns to all of the documented Broughton Archipelago watersheds indicates thatfollowing exceptionally high returns in 2000 and 2001, the populations declined to very low numbers in 2002 and 2003.Contrary to the conclusions reached by Krkosek et al. (2007a), Broughton pink salmon returns have steadily increased sincethen, with no indication that they are threatened with extinction. Other unsubstantiated assumptions used in Krkosek et al.(2007a) are also discussed in light of additional scientific reports and theoretical considerations.
Keywords Atlantic salmon, aquaculture, netpens, pink salmon fry, sea lice
INTRODUCTION
Following the release in 2002 of the Pacific Fisheries Re-source Conservation Council Advisory document (PFRCC,2002), intensive research on sea lice and their interaction withwild and farmed fish in the Broughton Archipelago of BritishColumbia has been underway. A series of studies claiming thatsalmon farms are the cause of sea lice (Lepeophtheirus salmonisand Caligus clemensi) infections on pink salmon (Oncorhynchusgorbuscha) fry in the archipelago have been published (Mortonet al., 2004; Krkosek et al., 2005, 2006, 2007b). Krkosek et al.(2007a) now claim that sea lice originating on salmon farms
Address correspondence to Kenneth M. Brooks, Aquatic Environmen-tal Sciences, 644 Old Eaglemount Road, Port Townsend, Washington, USA.E-mail: [email protected]
will result in the extinction of Broughton pink salmon stocksin the near future. This review, which is intended to expand thediscussion by describing other peer-reviewed papers and publicinformation important to understanding the interaction betweenpink salmon, farmed fish, and sea lice, includes the followingconsiderations:1. The variability of pink salmon returns throughout the
North Pacific with particular emphasis on the BroughtonArchipelago.
2. The general absence of mortality among juvenile pink salmonfollowing controlled laboratory exposure to Lepeophtheirussalmonis.
3. The absence of a cause-and-effect relationship between sealice infecting pink salmon fry and larval lice released fromsalmon farms.
4. A reevaluation of trends in pink salmon abundance.
403
Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 2
2:0
2 4
May 2
008
404 K. M. BROOKS AND S. R. M. JONES
KAKWEIKEN ODD-YEAR PINKS
0
100,000
200,000
300,000
400,000
500,000
600,000
700,000
800,000
900,000
1953 1957 1961 1965 1969 1973 1977 1981 1985 1989 1993 1997 2001
YEAR
ES
TIM
AT
E
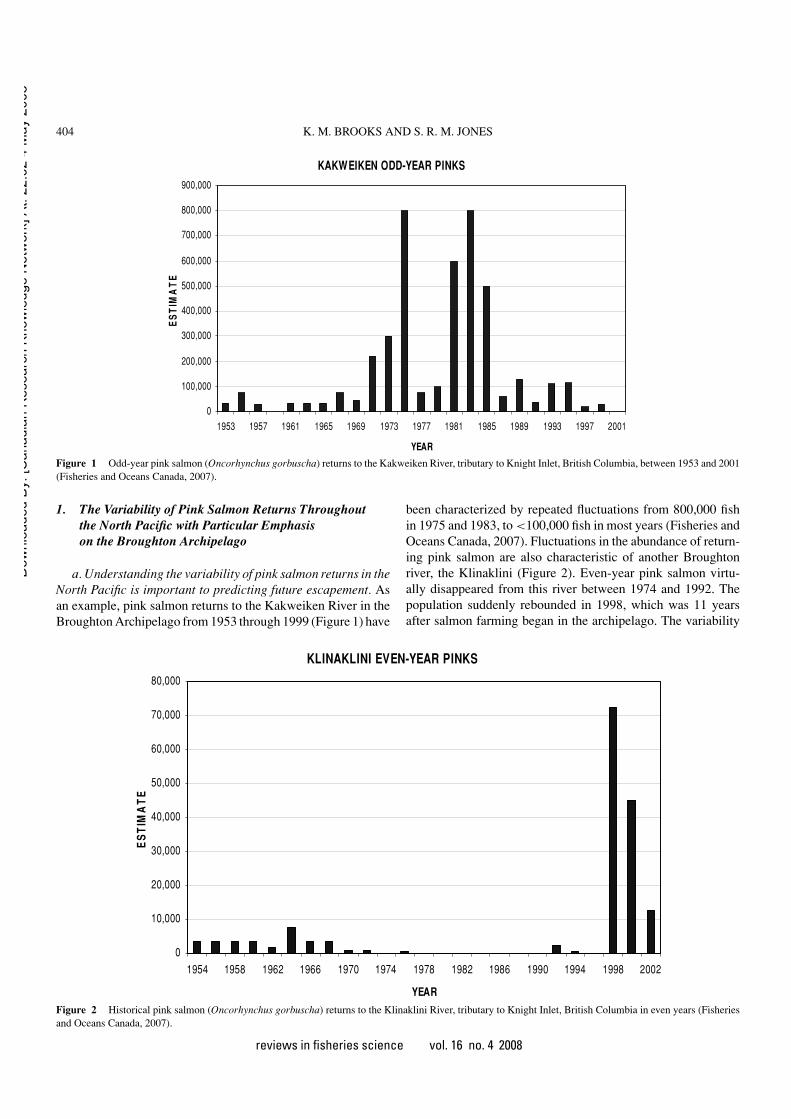
Figure 1 Odd-year pink salmon (Oncorhynchus gorbuscha) returns to the Kakweiken River, tributary to Knight Inlet, British Columbia, between 1953 and 2001(Fisheries and Oceans Canada, 2007).
1. The Variability of Pink Salmon Returns Throughoutthe North Pacific with Particular Emphasison the Broughton Archipelago
a. Understanding the variability of pink salmon returns in theNorth Pacific is important to predicting future escapement. Asan example, pink salmon returns to the Kakweiken River in theBroughton Archipelago from 1953 through 1999 (Figure 1) have
KLINAKLINI EVEN-YEAR PINKS
0
10,000
20,000
30,000
40,000
50,000
60,000
70,000
80,000
1954 1958 1962 1966 1970 1974 1978 1982 1986 1990 1994 1998 2002
YEAR
ES
TIM
AT
E
Figure 2 Historical pink salmon (Oncorhynchus gorbuscha) returns to the Klinaklini River, tributary to Knight Inlet, British Columbia in even years (Fisheriesand Oceans Canada, 2007).
been characterized by repeated fluctuations from 800,000 fishin 1975 and 1983, to <100,000 fish in most years (Fisheries andOceans Canada, 2007). Fluctuations in the abundance of return-ing pink salmon are also characteristic of another Broughtonriver, the Klinaklini (Figure 2). Even-year pink salmon virtu-ally disappeared from this river between 1974 and 1992. Thepopulation suddenly rebounded in 1998, which was 11 yearsafter salmon farming began in the archipelago. The variability
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 2
2:0
2 4
May 2
008
SEA LICE EXTINCTION HYPOTHESIS 405
Number of pink salmon returning to the Duckabush Rivertributary to Hood Canal in Washington State
-
20,000
40,000
60,000
80,000
100,000
120,000
1959
1961
1963
1965
1967
1969
1971
1973
1975
1977
1979
1981
1983
1985
1987
1989
1991
1993
Year of return
Num
ber
of p
ink
salm
on
Figure 3 Historic pink salmon returns to the Duckabush River, tributary to Hood Canal in Washington State, USA. Only odd-year runs are present in this river(National Marine Fisheries Service, 1996).
in returns of pink salmon fry is not unique to the BroughtonArchipelago. Figure 3 describes pink salmon returns to theDuckabush River tributary to Hood Canal in Washington State,where there have never been any salmon farms. Returns therehave varied between 100,000 in 1963 and a very few thousandin 1975, 1981, and 1993. The point in this discussion is thatextreme fluctuations in pink salmon returns are common, mak-ing assessments difficult. This point was emphasized by W. R.Heard (NOAA Fisheries, Auke Bay, Alaska, personal communi-cation) who noted that there was a “coast-wide collapse (at leastin much of North America) of pink salmon returns in 2006. The2006 forecast was for a harvest of around 40 million pinks (inAlaska), while the actual harvest was only 11 million due to theunusual low returns.” Haeseker et al. (2005), Heard (1991), andHard et al. (1996) provide reviews describing the extreme vari-
a) Even year returns b) Odd year returns
Glendale Creek odd year returns
0
200000
400000
600000
800000
1000000
1200000
1400000
1600000
1953195
5195
7195
9196
1196
3196
5196
7196
9197
1197
3197
51977
1979198
1198
3198
5198
7198
9199
1199
3199
5199
7199
92001
2003200
5
Gle
ndal
e od
d ye
ar re
turn
s
Glendale Creek even year returns
0
100,000
200,000
300,000
400,000
500,000
600,000
700,000
800,000
1954195
6195
8196
0196
2196
4196
6196
8197
0197
2197
41976
1978198
01982
1984198
6198
8199
0199
2199
4199
6199
8200
0200
2200
4200
6
Pink
salm
on r
etur
ns
Figure 4 (a) Even-year and (b) odd-year pink salmon returns to Glendale Creek, the major pink salmon-producing watershed in the Broughton Archipelago(Fisheries and Oceans Canada 2007).
ability in pink salmon returns in the Northeast Pacific generally,and the difficulty in forecasting future returns.
b. Exclusion of Glendale Creek data is inappropriate Krkoseket al. (2007a) excluded Glendale Creek escapement data be-cause the watershed includes a spawning channel. However, theydid include escapement from the Kakweiken River, which alsohas a spawning channel. This is important because Glendaleis the major pink salmon-producing system in the archipelago.Between 1999 and 2005, it produced 89% of the Broughtonpink salmon returns in odd years and 39% of the returns ineven years. In addition, Krkosek et al. (2007a) deals withmarine survival, not freshwater survival, and recruits fromGlendale affect marine population dynamics in all circum-stances. Figure 4 describes the historic variability of pink salmonreturns to Glendale Creek. The main difference in Glendale
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 2
2:0
2 4
May 2
008
406 K. M. BROOKS AND S. R. M. JONES
returns has been their increase after the initiation of salmonaquaculture in 1987 and the exceptionally high returns in 1992,2000, 2001, and 2004. Returns since the sharp 2000 decline havebeen within the range of values seen before aquaculture beganin 1987. Glendale returns are important to understanding pinksalmon returns to the Broughton, and exclusion of these databy Krkosek et al. (2007a) is not justified by the presence of aspawning channel.
2. Claims that Pink Salmon Fry Will Suffer up to 97%Mortality Following Infection with Sea Lice Are NotSubstantiated
Krkosek et al.’s (2007) claim that sea lice-induced mortalitywas commonly 80% in fry and ranged from 16% to over 97%.Their assessment is based, in part, on earlier studies (Mortonand Routledge, 2005; Krkosek et al., 2006), in which mortal-ity among captive, naturally infected pink and chum salmonled the authors to conclude that “Farm-origin lice induced 9–95 percent mortality in juvenile pink and chum salmon.” Thelatter studies used wild-caught fish of unknown size and failedto control for alternative sources of mortality. In contrast, noevidence of mortality was observed among naıve pink salmonjuveniles in controlled laboratory trials following exposure toL. salmonis copepodids (Jones et al., 2006b, 2007; Webster etal., 2007). Furthermore, patterns of gene expression showed thatpink salmon mount an effective defense that accelerates the re-jection of lice from their skin (Jones et al., 2007). Recently, thisinnate resistance to sea lice was found to first develop in pinksalmon between 0.3 g and 0.7 g and was associated with changesin skin development, including the formation of scales (Joneset al., in press, a ). In another study, the innate resistance to L.salmonis was retained in 3-g to 13-g pink salmon despite feeddeprivation (Jones et al., in press, b ). These observations areconsistent with numerous earlier studies demonstrating a gen-eral resistance to sea lice infections among species of Pacificsalmon (Oncorhynchus spp.) (Johnson, 1993; Johnson and Al-bright, 1992a, 1992b; Fast et al.,2002a, 2002b). More researchis required to characterize the risk associated with L. salmonisinfection before migrating, post-emergent pink salmon reach 0.7g (Jones et al.,in press, a ). A significant reduction in the riskof death associated with sea lice due to innate resistance has anegative influence on the capacity of the model used by Krkoseket al. (2007a) to predict population extinction.
3. No Cause-and-Effect Relationship Has BeenDemonstrated Between Sea Lice Infecting Pink SalmonFry and Larval Lice Released from Salmon Farms
Krkosek et al. (2007a) assert that larval lice released fromcultured Atlantic salmon are responsible for increased infec-tions on pink salmon fry collected near the farms. This assertionis based in part on correlation analysis and in part on an as-
sumption that there are no other significant sources of sea licein the Archipelago. These assumptions ignore several importantfactors that are well documented in the literature.
a. The “Fallow Route” Described in Krkosek et al. (2007a)Was Not Entirely Fallow in 2003. No empirical evidence of a“primary migration route” has been obtained in the Broughton,and the migration of pink salmon fry is likely dependent on anumber of fry density-dependent factors. Atlantic salmon pro-duction in the Broughton Archipelago between 1998 and 2005varied between 14,323,634 and 20,840,867 kg, with a mean and95% confidence interval of 17,360,469 ± 1,836,984. Produc-tion in 2003 (BC PSF, 2008) was 16,438,333 kg, which was notsignificantly different from the mean for the eight-year period(single sample t-test, t = 1.19, p = 0.28). A portion of thepink salmon fry migrating westward down Knight Inlet fromGlendale Creek and the Klinakini River encounter the DoctorIslet, Sargeaunt Pass, and Humphrey Rocks salmon farms as theyturn north into Tribune Channel, where they join fry from theKakweiken River (Figure 5). Doctor Islets produced 1,929,554kg of marketable salmon in 2003, and the farm was not fal-low until after May 19, 2003. Sargeaunt Pass and HumphreyRock, located in the same area, were stocked in March andApril of 2003. The presence of these stocked farms and thenormal production levels in the Broughton Archipelago during2003 jeopardizes a claim that the Tribune Channel migrationpath was fallow or that farm-derived larval lice were reduced in2003 compared with other years. In 2003, when Krkosek et al.(2007a) saw significant reductions in sea lice infections, nau-plii released from these three farms were competent to infectwild fish from Kumlah Island to the vicinity of the Glacier Fallsfarm. These fry continued migrating around Tribune Channel,including the area reported by Krkosek et al. (2007a). There-fore, reductions in sea lice infections on pink salmon fry oc-curred in the Tribune Channel despite the continued operationof salmon farms and consistent production of farmed salmonin the archipelago. The year 2003 is emphasized because thesefry migrated to sea through the archipelago during a year whenAtlantic salmon production was 16,438,333 kg, and they expe-rienced exceptional marine survival with good returns (950,288pink salmon) in 2004 (Beamish et al., 2006). Atlantic salmonproduction in the Broughton Archipelago was actually lower in2000 (15,575,808 kg), when the fish returning in the year of the2002 decline migrated to sea.
b. There is no evidence that a “pre-infestation period” actu-ally existed. Comparison of “pre-infestation” growth rates with“fallow growth rates” or “exposed population” growth rates byKrkosek et al. (2007a) is not valid, because there are no inven-tories of sea lice on other species of fish or on salmon fry in theBroughton prior to 2001, and thus, there is no basis for assumingthat a “pre-infestation period” actually existed.
c. Farmed salmon are not the only source of either. C.clemensi or L. salmonis in the Broughton Archipelago. Caligusclemensi has many wild hosts, and farmed salmon are not the
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 2
2:0
2 4
May 2
008
SEA LICE EXTINCTION HYPOTHESIS 407
Figure 5 Broughton Archipelago showing two possible migration paths taken by pink salmon fry (dashed lines) and the location of three active salmon farms(Doctor Islets, Sargeaunt Pass, and Humphrey Rocks), together with three inactive farms (Glacier Falls, Burdwood, and Wicklow).
only source of larval L. salmonis in the archipelago. The authorsignore the work of Jones et al. (2006a), who reported significantabundance and intensity of both C. clemensi and L. salmonison abundant populations of three-spine sticklebacks (Gasteros-teus aculeatus) in the Broughton, and the work of Trudel et al.(2006), who reported that approximately 25% of 284 juvenilepink salmon examined in the Eastern Bering Sea were infectedwith one to six sea lice, with a mean intensity of 1.5 lice perinfected fish. Trudel et al. (2006) concluded that, “This studydemonstrated that salmon infested with lice remained in coastalwaters throughout the year. We suggest that lice on salmon thatoverwinter in coastal waters will contribute to the infestation ofsalmon smolts migrating to sea in the spring through the releaseof lice nauplii in the water column.” The point in this discus-sion is that we do not know what the relative contributions of L.salmonis or C. clemensi larvae are from farmed salmon in com-parison with wild sources, and it is misleading to assume that sealice infections are associated primarily with nauplii released atsalmon farms. Repeated attempts to collect significant numbersof nauplii or infective copepodids at or near salmon farms havefailed. Salmon farms may contribute 99% of the larvae or theymay contribute 1%, and the answer eludes us. What is clear isthat sea lice are found on juvenile salmon wherever we look inthe Northeast Pacific, and there are numerous wild sources ofthese lice in addition to those contributed by salmon farms.
d. The effects of salinity and temperature on development andsurvival of sea lice larvae are poorly documented. Krokosek etal. (2005, 2006, 2007), Krokosek and Lewis (2005), and Morton
et al. (2004, 2005) have repeatedly asserted that their observa-tions of increased sea lice abundance on pink salmon fry inthe vicinity of salmon farms, in comparison with their controlsites, indicates that the farms are the source of the infectinglice. Their control samples have been collected in areas of lowsalinity, and they contend that L. salmonis larvae are not af-fected by salinities as low as 15 PSU (Morton et al., 2005).That assertion is contrary to our current understanding of thelife history of sea lice and hydrodynamics in the BroughtonArchipelago (Brooks, 2005; Brooks and Stucchi, 2006). Sci-ence proceeds as other researchers attempt to duplicate the re-sults of published work. Consistent with the seminal work ofJohnson and Albright (1991), who observed almost no develop-ment to the L. salmonis copepodid stage at salinities <30 PSU,Bricknell et al. (2006) found that survival of free-swimmingcopepodids was “severely compromised” at salinity <29 PSU,and Tucker et al. (2000) found reduced settlement of copepo-dids at 24 PSU in comparison with 34 PSU. Winter salinityin the Broughton approaches 30 PSU. However, salinity fallsrapidly after April and is typically in the range of 15–25 PSUduring late spring and summer in the eastern portions of thearchipelago, where these studies have been undertaken. This isanother facet of the epizootiology of sea lice that Krkosek et al.(2007a) fail to address in their model, making their control dataquestionable.
e. The dispersion of sea lice between hatching and molting tothe infective copepodid stage are not considered in the models ofKrkosek et al. (2007a). In response to the assertions of Krkosek
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 2
2:0
2 4
May 2
008
408 K. M. BROOKS AND S. R. M. JONES
and Lewis (2005) that increased infection of pink salmon fry inthe vicinity of salmon farms was caused by sea lice larvae origi-nating on salmon farms, Brooks (2005) and Brooks and Stucchi(2006) used a Broughton specific hydrodynamic model (Fore-man et al.,2006) to show that larvae would be advected at least10–12 km from the farms during development to the infectivestage and that, in some cases, they were likely advected out ofthe estuary. The model used in their study did not include theinfluence of wind on resting current vectors. However, an anal-ysis of empirical current data that does include the effects ofwind from 15 farms in the archipelago indicated that maximumcurrent speeds at every farm were between 50 and 75 cm/sec,and that resting currents vectors integrated over full lunar cyclescarried imbedded particles toward Queen Charlotte Strait at amean speed of 1.99 cm/sec or 6.8–10.3 km during developmentto the copepodid stage. Gillibrand and Willis (2007) have de-veloped a model to predict the dispersion of larvae from salmonfarms or other sources of nauplii. Their model includes sea licebehavior, mortality as a function of salinity, development time asa function of temperature, and wind, river discharge, and tidalcurrents. Consistent with Brooks (2005), their model predictsthat larvae will molt to an infective stage at distances of 7–12km from the point where they hatched. Gillibrand and Willis(2007) note that their predictions are consistent with field ob-servations of copepodids. None of this contradictory evidence isdiscussed by Krkosek et al. (2007a), nor do they describe howit affects their model. Basic considerations of sea lice life his-tory and currents suggest a very small likelihood that sea licelarvae are retained in the area where they hatch for four to sixdays while they develop to an infective stage. This conclusionhas now been reached by several other researchers in variousparts of the world. Krokosek and colleagues have refused to ac-knowledge criticisms of their model (Brooks, 2005; Gillibrandand Willis, 2007) or to demonstrate any cause-and-effect rela-tionship between their identified zones of infection and sea liceoriginating on salmon farms.
f. There is no consideration by Krkosek et al. (2007a) thatsalmon farms have been using prophylactic applications ofemamectin benzoate to manage sea lice infestations since 2004.These prophylactic treatments have resulted in farm lice infes-tation decreasing during the migration season. Krkosek et al.(2007a) assume a steady state lice dispersion model. This as-sumption cannot be validated and the model should have beenrelaxed to accommodate the temporal dynamics of sea lice popu-lations available for salmon farms in the Broughton. As a result,Krkosek et al. (2007a) have overestimated the production of sealice nauplii on farms.
4. Analysis of the Entire Pink Salmon Database SupportsConclusions Opposite to Those Reached by Krkosek etal. (2007)
a. Misleading regression analysis. Krkosek et al. (2007a)used linear regression to fit a log-transformed subset of normal-
ized escapement data to the Ricker model. Populations of pinksalmon were grouped by assuming they had experienced vari-ous levels of sea lice exposure. Even a cursory examination ofthe data in Figure 3 of their study suggests that a non-linear re-gression would have been more appropriate, and that a properlyfitted response would asymptotically approach values that arenear zero but slightly negative for all of the modeled popula-tions. In addition, the authors did not provide coefficients of de-termination for the regressions. The scatter in Figure 3 suggeststhat the linear solution explained very little of the variabilityin the database. In addition, the data in Figure S1, describingescapement since 1970, should have been analyzed using an ap-propriate regression model to determine if the coefficients ontime were significantly positive or negative or not significantlydifferent from zero.
b. Incomplete model. Krkosek et al. (2007a) evaluated pinksalmon population dynamics over time using the Ricker (1975)model. In their model ni (t) = ni (t – 2) exp[r – bni (t – 2)],where ni is the number of pink salmon in cohort i , t is thetime at which the estimate is made, (t – 2) is the spawning co-hort producing the generation in question, r is the populationgrowth rate, and bni (t – 2) represents losses in cohort n dueto density-dependent mortality. To be biologically meaningful,the density-dependent mortality should have been expressed as!b j ni (t – 2). There are numerous factors affecting freshwaterand marine survival of pink salmon (Heard,1991; Friedland etal., 2003). Freshwater survival includes random mortality ef-fects and density-dependent effects (number of spawners, etc.).Mortality factors affecting pink salmon fry during their near-shore marine residence include both fresh and marine waterquality (bwq ), predation, and several density-dependent factorssuch as food availability (b f ), horizontally transmitted diseaseagents (bd ), and random disease effects, such as sea lice infec-tions (bsl). Collectively, these factors contribute to the 55–77%mortality expected during the early marine life of pink salmon(Heard, 1991). A more appropriate model would be of the formni (t) = ni (t – 2) exp[r – !b j ni (t – 2)], where the b j includeat least the density-dependent factors cited above. There is nodoubt that the historic return of 3,600,000 pink salmon to theBroughton Archipelago in 2000 collapsed in 2002. Since everysystem has a carrying capacity, it is reasonable to hypothesizethat the unprecedented return of 3.6 million pink salmon to theBroughton Archipelago in 2000 exceeded the available spawn-ing habitat in freshwater and the carrying capacity of the ma-rine environment during the 2001 out-migration of fry. Bugayev(2002) noted a similar collapse in an area of Russia, where therewere no salmon farms, and concluded that, “Due to overflowof the spawning grounds, almost the whole generation of pinksalmon of the Western Kamchatka of 1983 died.” The conclusionby Krkosek et al. (2007a) that sea lice infections derived fromfarmed salmon caused the reduced population growth rates (r ),observed during the “exposed period,” is not substantiated in thestudy by any cause-and-effect relationship, and there could havebeen many causes of the low returns in 2002. A serious error
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 2
2:0
2 4
May 2
008
SEA LICE EXTINCTION HYPOTHESIS 409
Pink salmon returns to the Broughton Archipelago between 1953 and 2006
0
500,000
1,000,000
1,500,000
2,000,000
2,500,000
3,000,000
3,500,000
4,000,000
Yea
r19
5419
5619
5819
6019
6219
6419
6619
6819
7019
7219
7419
7619
7819
8019
8219
8419
8619
8819
9019
9219
9419
9619
9820
0020
0220
04
Num
ber
of r
etur
ning
pin
k sa
lmon
Odd Even
Period of salmon farming
Previous even year declines
Previous odd year declines
Figure 6 Even- and odd-year pink salmon returns to Broughton Archipelago watersheds between 1953 and 2006. Arrows indicate periodic declines that, whenanalyzed over short periods of time, could be used to predict the extinction of pink salmon stocks (Fisheries and Oceans Canada, 2003).
of omission is failure to include and discuss these other causesof mortality, resulting in an incomplete model and misleadingresults.
c. Selective use of data. Figure 6 describes pink salmon re-turns to Broughton Archipelago watersheds between 1953 and2006. Salmon farming began in 1987, and pink salmon returnsincreased during the next 13 years. The solid arrows indicate his-toric periods of even-year pink salmon declines, and the dashedarrows indicate odd-year declines. An analysis that is restrictedto any of these periods, for instance, between 1976 and 1984 orbetween 1983 and 1991, could be interpreted as evidence thatthe stocks were headed for extinction. However, in each case,pink salmon returns to the Broughton Archipelago bottomed andthen increased to at least their long-term average number, justas they are now increasing since the low returns of 2002 and2003.
d. Incomplete analysis. Figure 7 describes total returns tothe 11 Broughton watersheds included in the Fisheries andOceans Canada escapement database for Area 12 (BroughtonArchipelago) from 1999 (before the 2002 decline) through 2006(Fisheries and Oceans Canada, 2007). For this analysis, theindependent variable year was converted to years since 1998and log10(N+ 1) transformed to normalize the residuals. Thedata was then analyzed using non-linear regression (Statistica,Version 6). A linear model, such as that used in Krkosek etal. (2007a), is included in Figure 7. Note that the non-linearpolynomial has a coefficient of determination of 0.55. Krkoseket al. (2007a) did not provide details describing their regres-sion analyses, but the linear regression for this un-normalizedbut log-transformed data resulted in r2 = 0.29, and the co-
efficient on time was not significant (p = 0.18). Of greaterimportance in Figure 7 is the shape of the curves. The lin-ear regression approach, similar to that used by Krkosek etal. (2007a), suggests that pink salmon populations are declin-ing, and the trend would predict their eventual extinction. Thepolynomial fit suggests a very different scenario, in whichthe unprecedented returns in 2000 and 2001 were followedby significant reductions in 2002 and 2003. However, as hasbeen seen repeatedly in pink salmon populations, the trendsince 2003 has been positive, with increasing returns suggestingthat Broughton populations are recovering from the unknowncauses of the 2002 and 2003 lows. A scholarly manuscriptwould have discussed these alternate analyses, and the authorswould have justified their conclusions in light of all of theevidence.
a. Current trends in pink salmon returns. The Broughtondata for both odd and even years between 2002 and 2006 issummarized in Figure 8. Consistent with total returns, data forthe highest-producing pink salmon watershed in the archipelagodemonstrates a trend to increasing returns after 2002. The excep-tionally high return in 2004 resulted in a poor fit, with r2
a = 0.04and a non-significant coefficient on the independent variableyear. If the exceptionally high returns in 2004 are excluded asan outlier (Beamish et al., 2006), a significant (p < 0.05) pos-itive trend is demonstrated with a predicted increase of 49,818pink salmon each year and a coefficient of determination of r2
a= 0.98. Marine survival of pink salmon returning to Broughtonwatersheds has been 1.0–23% (2004 = 23%; 2005 = 3.4%;2006 = 1.0%, and 2007 = 2.6%). Pink salmon marine survivalfor Fraser River pink salmon is typically 1.2%, and it aver-ages 2–3% coastwise (R. Beamish, personal communication).
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 2
2:0
2 4
May 2
008
410 K. M. BROOKS AND S. R. M. JONES
Figure 7 Log10 (Broughton pink salmon escapement) since from 1999 through 2006 as a function of time. Values have been Log10 transformed. The coefficientof determination for the polynomial is R2
a = 0.55, whereas R2a for the linear regression is 0.29. However, other than the constant, none of the regression coefficients
were significant, suggesting no significant trends.
Figure 8 Linear regression describing trends in pink salmon returns to Broughton watersheds from 2002 through 2006. Note that the exceptionally high 2004returns are included in this figure.
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 2
2:0
2 4
May 2
008
SEA LICE EXTINCTION HYPOTHESIS 411
Therefore, survival has been equal to or better than observed forthe Fraser River and the rest of the British Columbia coast dur-ing the last four years. It has been six years (2002–2007) sinceBroughton Archipelago pink salmon stocks declined sharply in2002. By examining a subset of the pink salmon watersheds inthe archipelago, Krkosek et al. (2007a) claim that louse-inducedmortality of pink salmon is commonly over 80%, and they ex-pect a 99% collapse in pink salmon population abundance infour salmon generations (8 years). An examination of Figures4, 6, 7, and 8 suggests that current returns are within the historicvariability of the archipelago, and that since 2002 the populationgrowth trends are positive, not negative as suggested in Krkoseket al. (2007a).
CONCLUSIONS
By selective use of data, questionable analytical proce-dures, and several unsubstantiated assumptions presented as fact,Krkosek et al. (2007a) predict the extinction of pink salmonstocks in the Broughton Archipelago within four generations (8years). The authors failed to acknowledge and review the workof numerous scientists from around the world whose results donot necessarily support their conclusions. They have failed topresent alternative hypotheses and analytical approaches or todiscuss how these might influence their conclusions. Researchto this date indicates that pink salmon fry mount an effectiveresponse to sea lice infections. Pink salmon escapements typ-ically fluctuate dramatically throughout the Northeast Pacific.Stocks of these fish reached unprecedented levels in 2000 anddeclined sharply in 2002 for unknown reasons. When all of theBroughton’s watersheds are considered, pink salmon stocks areseen to have steadily increased over the last five years, with noindication that they are headed for extinction. The purpose ofthis article is not to deny that salmon farms may contribute sealice to the marine environment. The fact is that, at this time,research has not determined the relative contribution from wildand farmed sources of lice. Rather, this discussion is intendedto provide additional information, giving readers a broader per-spective of these issues.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the critical commentsand endorsement of this manuscript by the following scien-tists working in this field: Lars Asplin, Ph.D., Øivind Bergh,Ph.D., Karin Boxaspen, Ph.D., Ian Bricknell, Ph.D., James Bron,Ph.D., Walton W. Dickhoff, Ph.D., Ralph A. Elston, Ph.D.,William T. Fairgrieve, Ph.D., Mark D. Fast, Ph.D., Ronald W.Hardy, Ph.D., Lee Harrell, DVM, William R. Heard, M.Sc.,Robert N. Iwamoto, Ph.D., Stewart C. Johnson, Ph.D., Con-rad V. Mahnken, Ph.D., R. Scott McKinley, Ph.D., Colin Nash,Ph.D., and William Waknitz, M.Sc.
REFERENCES
Beamish, R. J., S. Jones, C. Neville, R. Sweeting, G. Karreman, S.Saksida, and E. Gordon. Exceptional production of pink salmon in2003/2004 indicates that farmed salmon and wild Pacific salmon cancoexist successfully in a marine ecosystem on the pacific coast ofCanada. ICES J. Mar. Sci., 63: 1326–1337 (2006).
Bricknell, I. R., S. J. Dalesman, B. O’Shea, C. C. Pert, and A. J. Mor-due Luntz. Effect of environmental salinity on sea lice Lepeoph-theirus salmonis settlement success. Dis. Aquat. Org., 71: 202–212(2006).
BC PSF. Proprietary data provided by the British Columbia PacificSalmon Forum with permission from Marine Harvest Canada andMainstream Canada (2008).
Brooks, K. M. The affects of water temperature, salinity, and currentson the survival and distribution of the infective copepodid stage ofsea lice (Lepeophtheirus salmonis) originating on Atlantic salmonfarms in the Broughton Archipelago of British Columbia, Canada.Rev. Fish. Sci., 13: 177–204 (2005).
Brooks, K. M. and D. J. Stucchi. The effects of water temperature,salinity, and currents on the survival and distribution of the infectivecopepodid stage of the salmon louse (Lepeophtheirus salmonis) orig-inating on Atlantic salmon farms in the Broughton Archipelago ofBritish Columbia, Canada (Brooks, 2005). A response to the rebuttalof Krkosek et al. (2005a). Rev. Fish. Sci., 14: 13–23 (2006).
Bugayev, V. F. On pink salmon (Oncorhynchus gorbuscha) abundanceinfluence on Asian sockeye salmon (Oncorhynchus nerka) abun-dance. NPAFC Doc. 628, 11 pp. (2002).
Fast, M. D., N. W. Ross, A. Mustafa, D. E. Sims, S. C. Johnson, G. A.Conboy, D. J. Speare, G. Johnson, and J. F. Burka. Susceptibility ofrainbow trout Oncorhynchus mykiss, Atlantic salmon Salmo salar,and coho salmon Oncorhynchus kisutch to experimental infectionwith sea lice Lepeophtheirus salmonis. Dis. Aquat. Org., 52: 57–68(2002a).
Fast, M. D., D. E. Sims, J. F. Burka, A. Mustafa, and N. W. Ross.Skin morphology and humoral non-specific defense parameters ofmucus and plasma in rainbow trout, coho and Atlantic salmon. Comp.Biochem. Physiol. A., 132: 645–657 (2002b).
Fisheries and Oceans Canada (DFO). Sea Lice Monitoring Program.http://www-sci.pac.dfo-mpo.gc.ca/mehsd/sea lice/marine reports/(2003).
Fisheries and Oceans Canada (DFO). Area 12 Mainland Inlet PinkSalmon Update Bulletin #7, February 2, 2007. http://www-sci.pac.dfo-mpo.gc.ca/mehsd/sea lice/2006/2006 intro e.htm (2007).
Foreman, M. G. G., D. J. Stucchi, Y. Zhang, and A. M. Baptista. Estu-arine and tidal currents in the Broughton Archipelago. AtmosphericOceans, 44: 47–63 (2006).
Friedland, K. D., D. G. Reddin, J. R. McMenemy, and K. F. Drinkwa-ter. Multidecadal trends in North American Atlantic salmon (Salmosalar) stocks and climate trends relevant to juvenile survival. Can.J. Fish. Aquat. Sci., 60: 562–583 (2003).
Gillibrand, P. A., and K. J. Willis. Dispersal of sea louse larvae fromsalmon farms: Modeling the influence of environmental conditionsand larval behavior. Aquat. Biol., 1: 63–75 (2007).
Haeseker, S. L., R. M. Peterman, and S. Zhenming. Retrospective eval-uation of preseason forecasting models for pink salmon. N. Am. J.Fish. Manage., 25: 897–918 (2005).
Hard, J. J., R. G. Kope, W. S. Grant, F. W. Waknitz, L. T. Parker, and R. S.Waples. Status Review of Pink Salmon from Washington, Oregon, and
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 2
2:0
2 4
May 2
008
412 K. M. BROOKS AND S. R. M. JONES
California. U.S. Department of Commerce, NOAA Tech. Memo.NMFS-NWFSC-25, 131 pp. (1996).
Heard, W. R. Life history of pink salmon (Oncorhynchus gorbuscha).In: Pacific Salmon Life Histories. (C. Groot and L. Margolis, Eds.).Vancouver: UBC Press. pp 121–230 (1991).
Johnson, S. C. A comparison of development and growth rates ofLepeophtheirus salmonis (copepoda: caligidae) on naıve Atlantic(Salmo salar) and chinook (Oncorhynchus tshawytscha) salmon.In: Pathogens of Wild and Farmed Fish Sea Lice. (G. A. Boxhalland D. Defaye, Eds.). Chichester, UK: Ellis Horwood, pp. 68–80(1993).
Johnson, S. C., and L. J. Albright. Development, growth, and survivalof Lepeophtheirus salmonis (copepoda: caligidae) under laboratoryconditions. J. Mar. Biol. Assoc. UK, 71: 425–436 (1991).
Johnson, S. C., and L. J. Albright. Comparative susceptibility andhistopathology of the response of naıve Atlantic, chinook, and cohosalmon to experimental infection with Lepeophtheirus salmonis(copepoda: caligidae). Dis. Aquat. Org., 14: 179–193 (1992a).
Johnson, S. C., and L. J. Albright. Effects of cortisol implants on thesusceptibility and histopathology of the responses of naıve cohosalmon Oncorhynchus kisutch to experimental infection with Lep-eophtheirus salmonis (copepoda: caligidae). Dis. Aquat. Org., 14:195–205 (1992b).
Jones, S. R. M., G. Prosperi-Porta, E. Kim, P. Callow, and N. B.Hargreaves. The occurrence of Lepeophtheirus salmonis and C.clemensi (copepoda: caligidae) on three-spine stickleback Gasteros-teus aculeatus in coastal British Columbia. J. Parasitol., 92: 473–480(2006a).
Jones, S. R. M., E. Kim, and S. Dawe. Experimental infections withLepeophtheirus salmonis (Kroyer) on three-spine sticklebacks, Gas-terosteus aculeatus L., and juvenile Pacific salmon, Oncorhynchusspp. J. Fish Dis., 29: 489–495 (2006b).
Jones, S. R. M., M. D. Fast, S. C. Johnson, and D. B. Groman. Dif-ferential rejection of Lepeophtheirus salmonis by pink and chumsalmon: Disease consequences and expression of proinflammatorygenes. Dis. Aquat. Org., 75: 229–238 (2007).
Jones, S., E. Kim, and W. Bennett. Early development of resistance tothe salmon louse Lepeophtheirus salmonis (Krøyer) in juvenile pinksalmon Oncorhynchus gorbuscha (Walbaum). J. Fish Dis. (in press,a).
Jones, S. R. M., M. D. Fast, and S. C. Johnson. Influence of reduced feedration on Lepeophtheirus salmonis infestation and gene expressionin juvenile pink salmon. J. Aquat. Anim. Health (in press b).
Krkosek, M., M. A. Lewis, and J. P. Volpe. Transmission dynamics ofparasitic sea lice from farm to wild salmon. Proc. R. Soc. B., 272:689–696 (2005).
Krkosek, M., M. A. Lewis, A. Morton, L. N. Frazer, and J. P. Volpe.Epizootics of wild fish induced by farm fish. Proc. Natl. Acad. Sci.USA, 103: 15506–15510 (2006).
Krkosek, M., J. S. Ford, A. Morton, S. Lele, R. A. Myers, and M. A.Lewis. Declining wild salmon populations in relation to parasitesfrom farm salmon. Science, 318: 1772– 1775 (2007a).
Krkosek, M., A. Gottesfeld, B. Proctor, D. Rolston, C. Carr-Harris, andM. A. Lewis. Effects of host migration, diversity and aquaculture onsea lice threats to Pacific salmon populations. Proc. R. Soc. B., 274:3141–3149 (2007b).
Morton, A., and R. Routledge. Mortality rates for juvenile pink On-corhynchus gorbuscha and chum O. keta salmon infested with sealice, Lepeohtheirus salmonis, in the Broughton Archipelago. AlaskaFish. Res. Bull. 11: 146–152 (2005).
Morton, A., R. Routledge, and R. Williams. Temporal patterns of sealouse infestation on wild Pacific salmon in relation to the fallowingof Atlantic salmon farms. N. Am. J. Fish. Manage. 25: 811–821(2005).
Morton, A., R. Routledge, C. Peet, and A. Ladwig. Sea lice (Lepeoph-theirus salmonis) infection rates on juvenile pink (Oncorhynchusgorbuscha) and chum (Oncorhynchus keta) salmon in the nearshoremarine environment of British Columbia, Canada. Can. J. Fish.Aquat. Sci., 61: 147–157 (2004).
National Marine Fisheries Service (NMFS). Status Review ofPink Salmon from Washington, Oregon, and California.NOAA Technical Memorandum NMFS-NWFSC-25, 131pp. (1996).
PFRCC. Vancouver, BC: Pacific Fisheries Resource ConservationCouncil (2002).
Ricker, W. E. Computation and interpretation of biological statis-tics of fish populations. J. Fish. Res. Board Can. Bull., 191: 381pp. (1975).
Trudel, M., S. R. M. Jones, M. E. Thiess, J. F. T. Morris, D. W. Welch,R. M. Sweeting, J. H. Moss, B. L. Wing, E. V. Farley, Jr., J. M.Murphy, R. E. Baldwin, and K. C. Jacobson. Infestations of motilesalmon lice (Lepeophtheirus salmonis) on Pacific salmon along thewest coast of North America. Am. Fish. Soc. Symp. Ser., 57: 157(2006).
Tucker, C. S., C. Sommerville, and R. Wootten. The effect of temper-ature and salinity on the settlement and survival of copepodids ofLepeophtheirus salmonis (Kroyer, 1837) on Atlantic salmon, Salmosalar L. J. Fish Dis., 23: 309–320 (2000).
Webster, S. J., L. M. Dill, and K. Butterworth. The effect of sea liceinfestation on the salinity preference and energetic expenditure of ju-venile pink salmon (Oncorhynchus gorbuscha). Can. J. Fish. Aquat.Sci., 64: 672–680 (2007).
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 1
6:5
6 2
4 A
pril 2008
Reviews in Fisheries Science, 16(4):413–420, 2008Copyright C©© Taylor and Francis Group, LLCISSN: 1064-1262 printDOI: 10.1080/10641260802013692
Sea Lice and Pink Salmon Declines:A Response to Brooks andJones (2008)
MARTIN KRKOSEK,1,2 JENNIFER S. FORD,3 ALEXANDRA MORTON,4SUBHASH LELE,1 and MARK A. LEWIS1,2
1Centre for Mathematical Biology, Department of Mathematical and Statistical Sciences, University of Alberta, Edmonton,AB, Canada2Department of Biological Sciences, University of Alberta, Edmonton, AB, Canada3Biology Department, Dalhousie University, Halifax, NS, Canada4Salmon Coast Field Station, Simoom Sound, BC, Canada
In this article, we respond to concerns raised by Brooks and Jones (2008) about recent advances in sea lice and salmonpopulation dynamics in the Broughton Archipelago, British Columbia. We show that the assessment by Brooks and Jones(2008) is thoroughly mistaken and that their conclusions are based on a combination of obfuscation, misrepresentation,and fundamental misunderstandings. The extinction hypothesis is not actually a hypothesis at all, but rather an inevitableconsequence of sustained population decline. Local extinction of Broughton Archipelago pink salmon can be prevented ifpopulation declines are turned around, and the data and models suggest this can be achieved if the infestations are stopped.We have organized our responses in an itemized manner according to the headings and subheadings in Brooks and Jones(2008).
Keywords aquaculture, conservation, population dynamics, transmission, salmon, sea lice
1. THE VARIABILITY OF PINK SALMON RETURNSTHROUGHOUT THE NORTH PACIFIC WITHPARTICULAR EMPHASIS ON THE BROUGHTONARCHIPELAGO
a. Understanding the Variability of Pink Salmon Returnsin the North Pacific is Important to PredictingFuture Escapement
Brooks and Jones (2008) emphasize that pink salmon popu-lation dynamics are naturally variable. We fully agree and pointout that the variation is due to many factors—both biotic andenvironmental—and that the variability is clearly representedin both Figures 2 and 3 of Krkosek et al. (2007a). We do notclaim that sea lice are the only factors that affect salmon pop-ulation dynamics; rather, in Krkosek et al. (2007a), we have
Address correspondence to Martin Krkosek, Centre for Mathematical Biol-ogy, Department of Mathematical and Statistical Sciences, 632 Central Aca-demic Building (CAB), University of Alberta, Edmonton, AB, T6G 2G1,Canada. E-mail: [email protected]
shown that sea lice are one key factor that affects the popu-lation dynamics of pink salmon. To accommodate the naturalvariability of pink salmon population dynamics in the analy-sis, we used the stochastic Ricker model (Dennis and Taper,1994), which is a nonlinear stochastic model that explicitly mod-els density-dependent mortality and environmental variation inpink salmon population dynamics. This quantitative frameworkis well known and well established for analyzing the populationdynamics of fish (Myers et al., 1999; Hilborn and Walters, 2001)as well as many other species (Brook and Bradshaw, 2006). Tocontrol for other factors that affect pink salmon population dy-namics and isolate the effects of sea lice on pink salmon pop-ulation dynamics, we used a comparative analysis of exposedpre-infestation, exposed infested, and unexposed populations.The exposed pre-infestation populations and exposed infestedpopulations are in the vicinity of salmon farms prior to and af-ter the sea lice infestations, which were first observed in 2001.The unexposed populations are far from salmon farms, with noknown sea lice infestations. The exposed pre-infestation andunexposed populations share the many factors that affect pink
413

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 1
6:5
6 2
4 A
pril 2008
414 M. KRKOSEK ET AL.
salmon population dynamics, evidenced by their similar pop-ulation growth rates (Krkosek et al., 2007a) and synchronouspopulation dynamics (Pyper et al., 2001). The exposed infestedpopulations have a markedly depressed growth rate, which is iso-lated to the exposed infested populations. This statistical design,similar to the matched case-control study design in epidemiol-ogy (Rothman, 1986), indicates sea lice are the factor drivingthe difference in population growth rates.
Brooks and Jones (2008) present pink salmon escapementdata from the Klinaklini River as an example of natural vari-ation in pink salmon population dynamics. Their assessmentsuggests pink salmon disappeared from this river in even yearsbetween 1974 and 1990 and then abruptly returned to abun-dances in the tens of thousands in 1998 and subsequent years. Wechecked the validity of their assessment against information heldin the Fisheries and Oceans Canada Pacific salmon escapementdatabase using the Mapster program (http://www-heb.pac.dfo-mpo.gc.ca/maps/maps-data e.htm). Most of the data portrayedas showing zero abundance between 1974 and 1990 in Figure 2of Brooks and Jones (2008) correspond to entries of “no data”or “not inspected” in the Pacific salmon escapement database,rather than “zero abundance.” There is also a problem with theirinterpretation of the increased abundance in this river in 1998.The rise in pink salmon escapement estimates in the Kliniklinifollowing 1998 coincides with a change in escapement estima-tion from overhead flights and river walks (1996 and earlier) tomark-recapture programs that used a fishwheel to capture fish(1998 and afterward). The Klinaklini is a large glacier-fed andheavily-silted river, making visibility of fish and escapementestimates based on overhead flights very difficult. A fishwheel-based mark-recapture approach would be enhanced by silted wa-ter because the low visibility will limit the salmon’s vision duringcapture. The data from the two escapement methods have notbeen calibrated (Pieter van Will, Fisheries and Oceans Canada,personal communication). These limitations of the KlinakliniRiver data suggest that the extinction-colonization dynamics thatBrooks and Jones (2008) described did not actually occur andare rather the spurious result of a poor quality data set. The Kli-naklini data were not used in the analysis in our Science report(Krkosek et al., 2007a).
b. Exclusion of Glendale Creek Data is Inappropriate
Brooks and Jones (2008) expressed concern about Krkoseket al. (2007a) excluding the Glendale populations but includingthe Kakweiken populations. In Krkosek et al. (2007a), it was sta-tistically necessary to exclude rivers that recently had spawningchannels constructed. As stated in the paper, this was done sys-tematically for rivers in the Broughton as well as the unexposedarea to the north. The objective of spawning channel construc-tion is to improve spawning habitat quantity and spawning gravelquality. If the spawning channels are effective, the increased pro-ductivity due to spawning channels prevents evaluation of recenttrends relative to historical abundance, which is the basis of anal-
ysis in the paper. The Kakweiken was included in the analysisbecause its spawning channel has not been successfully utilized(usually less than 7% and frequently 0% of the run use it; GlenNeidrauer, Fisheries and Oceans Canada, personal communica-tion). In contrast, the Glendale spawning channel has been wellutilized (typically 70,000 fish use it; Pieter van Will, Fisheriesand Oceans Canada, personal communication) and has greatlyincreased the escapement in this system since its construction. Itis straightforward to reanalyze the data with Glendale includedand Kakweiken excluded, despite the biases. The populationgrowth rate, r , with 95% confidence intervals for Broughtonpink salmon populations during the sea lice infestations are:
With Glendale
r = −1.002, 95%CI : −1.52 to − 0.52
Without Glendale and Kakweiken
r = −1.23, 95%CI : −1.80 to − 0.62
The population growth rate for Broughton pink salmon popula-tions during the infestations as reported in our study are:
r = −1.17, 95% , CI : −1.71 to − 0.59
All of these analyses show that Broughton pink salmon popu-lations were depressed and rapidly declining during the sea liceinfestations.
2. CLAIMS THAT PINK SALMON FRY WILL SUFFERUP TO 97% MORTALITY FOLLOWING INFECTIONWITH SEA LICE ARE NOT SUBSTANTIATED
The estimates of annual pink salmon mortality due to salmonlice in Krkosek et al. (2007a) are not based on previous ex-perimental work as Brooks and Jones (2008) claim. Rather,the mortality estimates in Krkosek et al. (2007a) are based ona direct analysis of pink salmon escapement data and annualaverage salmon lice abundances. The estimates for mortalityranged from 16% to over 97%, and were commonly over 80%.However, the pathogenicity of motile L. salmonis on juvenilepink salmon, estimated in Krkosek et al. (2007a) from escape-ment data, gives similar values to those estimated independentlyfrom survival experiments of infected juvenile salmon held inocean enclosures (Krkosek et al., 2006). The reduction in wildsalmon survival from these two studies is also similar to the ob-served decline in wild pink salmon in Ford and Myers (2008)meta-analysis. The studies by Jones et al. (2006a, 2007) eval-uated the effect of salmon lice on the survival of juvenile pinksalmon at sizes over 10 times larger than the juvenile salmoninfested each spring in the Broughton Archipelago (Morton andWilliams, 2003; Morton et al., 2004, 2005). Jones et al. (2006a,2007) used juvenile salmon that weighed over 10 g and werefully scaled, whereas the juvenile pink salmon studied in the
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 1
6:5
6 2
4 A
pril 2008
SEA LICE AND PINK SALMON DECLINES 415
Broughton Archipelago weigh less than 1 g and do not havescales (Morton and Williams, 2003; Morton et al., 2004, 2005).The effect of salmon lice on salmon survival is host size de-pendent (Pike and Wadsworth, 2000; Boxaspen, 2006), and sothe studies by Jones et al. (2006a, 2007) likely underestimatethe impact of sea lice on juvenile salmon survival at the sizerelevant to the infestations in the Broughton Archipelago. Thestudy by Webster et al. (2007) tested for the effect of salmonlice on juvenile pink salmon behavior, not survival. Webster etal. (2007) examined the juvenile pink salmon after 14 days andfound most of the lice were gone before the lice reached theirpathogenic motile stages. Studies that have reared salmon liceto motile stages on juvenile pink salmon weighing less than 1g have observed high mortality of the infected juvenile pinksalmon (Morton and Routledge, 2005; Krkosek et al., 2006). Inaddition, all experimental work on pink salmon mortality fromsea lice does not reflect the sustained rate of new infections char-acteristic of the Broughton Archipelago as the fish migrate pastmultiple farms.
3. NO CAUSE AND EFFECT RELATIONSHIP HASBEEN DEMONSTRATED BETWEEN SEA LICEINFECTING PINK SALMON FRY AND LARVALLICE RELEASED FROM SALMON FARMS
Many studies have demonstrated spatial or temporal asso-ciations of sea lice infestation of wild juvenile Pacific salmonwith salmon farming operations (Morton and Williams, 2003;Morton et al., 2004, 2005, 2007; Krkosek et al., 2005, 2006).Some of these studies have shown excellent agreement be-tween models of sea lice dispersion and infection with fielddata of sea lice infecting juvenile pink and chum salmon mi-grating past salmon farms (Krkosek et al., 2005, 2006). Thesestudies have allowed for significant sources of lice other thansalmon farms. In fact, two of these studies statistically de-tected and quantified the transmission of sea lice from non-farm sources to wild juvenile pink and chum salmon (Krkoseket al., 2005, 2006). The estimated transmission from non-farmhosts has been consistently very low and overwhelmed by trans-mission from farm salmon (Krkosek et al., 2005, 2006; Orr,2007).
a. The “Fallow Route” Described in Krkosek et al. (2007)was Not Entirely Fallow in 2003
The Broughton Archipelago is a system of linear inlets andchannels. Because the juvenile pink salmon migrate from theirnatal stream to the open ocean, they have to migrate down theinlets and channels. Primary migration routes in a system likethis do not require empirical evidence. Rather the migrationroutes are obviously based on the topography of the Archipelago.Krkosek et al. (2007a) clearly identified the fallow route as Tri-bune Channel and Fife Sound. The Doctor Islet farm, which was
stocked with adult salmon in the spring of 2003, is not locatedin Tribune Channel, but rather in Knight Inlet. Krkosek et al.(2007a) clearly identified that some of the lice observed on thejuvenile pink salmon in 2003 could have originated from salmonfarms located outside this fallowed migration corridor, and thisis known based on previous work documenting the spread of licefrom the Doctor Islet farm (Krkosek et al., 2005). It is true thatSergeants Pass and Humphrey Rock salmon farms were stockedwith smolts later in the spring of 2003. These farms could nothave been a source of lice because the stocked smolts enter theocean without lice, and there was insufficient time for sea lice tocolonize the smolts, reproduce, and become a significant sourceof lice to infect the wild juvenile salmon migrating past thesefarms.
b. There is No Evidence that a “Pre-Infestation Period”Actually Existed
There is a strong basis for assuming an L. salmonis pre-infestation period existed. In areas without salmon farms, theprevalence of salmon lice on juvenile pink salmon during theirfirst 1–3 months of marine life is less then 5% (Wertheimer et al.,2003; Morton et al., 2004; Krkosek et al., 2007b; Peet, 2007).The low prevalence of salmon lice on juvenile pink salmon inthese areas during this time is because the vast majority of theadult wild Pacific salmon that carry the parasite are located off-shore when the juvenile salmon enter sea (Groot and Margolis,1991; Krkosek et al., 2007b). In this way, salmon migrationprotects juvenile pink salmon from salmon lice during earlymarine life (Krkosek et al., 2007b). Salmon lice infestations ofjuvenile pink salmon have only been observed in areas withsalmon farms, and the primary source of salmon lice has beenconsistently identified to be salmon farms (Morton and Williams,2003; Morton et al., 2004, 2005, 2007; Krkosek et al., 2005).Salmon lice infestations of juvenile pink salmon are conspic-uous, and because the Broughton Archipelago is inhabited bybiologists, fishermen, and First Nations people, it is unlikelythat the infestations occurred before 2001 without notice. Thesudden occurrence of the infestations could be explained byfarmed salmon regional density in the Broughton Archipelagoexceeding a host density threshold that previously suppressedoutbreaks. Such thresholds are common in epidemiology (Mayand Anderson, 1991; Grenfell and Dobson, 1995) and are thebasis for culling as a means for disease control in domestic andwild animals. Such thresholds, however, may be difficult to pre-dict and identify (Lloyd-Smith et al., 2005).
c. Farmed Salmon are Not the Only Source of Either C.clemensi or L. salmonis in the Broughton Archipelago
The infestations associated with the rapid decline inBroughton Archipelago pink salmon populations are L. salmo-nis, not C. clemensi (Morton and Williams, 2003; Morton et al.,2004, 2005; Krkosek et al., 2006). Much previous work by
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 1
6:5
6 2
4 A
pril 2008
416 M. KRKOSEK ET AL.
some of the authors has examined the sources of sea lice infest-ing juvenile salmon in the Broughton Archipelago (Morton andWilliams, 2003; Morton et al., 2004, 2005; Krkosek et al., 2005,2006). None of this work has ignored the presence of alternatehosts for sea lice in the Broughton Archipelago. In fact, someof the studies have specifically tested for the presence of naturalorigin sea lice in the Broughton Archipelago and quantified howmany lice are coming from these natural hosts (Krkosek et al.,2005, 2006). These analyses have indicated that the vast major-ity of sea lice infecting wild juvenile salmon in the BroughtonArchipelago have originated from farm salmon. It is simply nottrue that sea lice commonly infest juvenile pink salmon every-where in the Northeast Pacific. The studies examining salmonlice infections on juvenile pink salmon during their first 1–3 months of marine life in areas without salmon farms in theNortheast Pacific have found salmon lice on less than 5% of thejuvenile pink salmon when the salmon are less than 80 mm forklength (Wertheimer et al., 2003; Morton et al., 2004; Krkoseket al., 2007b; Peet, 2007). Three-spine stickleback cannot be theorigin of salmon lice infestations because stickleback are dis-tributed throughout the North Pacific coastal waters, whereas L.salmonis infestations of wild juvenile Pacific salmon occurredonly in areas with salmon farms (Morton and Williams, 2003;Morton et al., 2004, 2005, 2007; Krkosek et al., 2005, 2006),and infestations of juvenile salmon near salmon farms are dom-inated by larval copepodid and chalimus lice, whereas salmonlice do not survive to reproductive age on stickleback (Joneset al., 2006a, 2006b).
d. The Effects of Salinity and Temperature on Developmentand Survival of Sea Lice Larvae are Poorly Documented
In our Science report (Krkosek et al., 2007a), we did notinvestigate the effects of salinity and temperature on sea licelarvae as Brooks and Jones (2008) suggest. Rather, we inves-tigated the relationship between salmon returns and measuredlevels of sea louse infection on juvenile pink salmon. However,it is known that abiotic factors such as temperature and salin-ity affect salmon lice survival and developmental rates (Stien etal., 2005; Bricknell et al., 2006). In our earlier work examin-ing salmon lice infestations of wild juvenile salmon (Krkoseket al. 2005, 2006), the louse developmental rates estimated atthe observed ocean temperatures were related to those expectedfrom experimental data (Stien et al., 2005) by the average ju-venile salmon migration speed, ∼1 km per day (Krkosek etal., 2006). In other work, the effects of salinity on sea liceabundance on wild juvenile Pacific salmon were insignificant(Morton et al., 2004, 2007), possibly because of louse behavior.Salinity is vertically distributed in the water column and larvallice have a diel vertical migration (Heuch et al., 1995) and canselect locations suitable for their survival and/or transmission(Heuch, 1995). The behavior of lice can combine with thesephysical variables including tides, currents, and wind to gener-ate spatial distributions of nauplii and copepodids from a point
source of release that take a variety of forms, including thoseobserved in the Broughton Archipelago (Gillibrand and Kate,2007).
e. The Dispersion of Sea Lice between Hatching andMolting to the Infective Copepodid Stage are NotConsidered in the Models of Krkosek et al. (2007)
In our Science report (Krkosek et al., 2007a), we did notinvestigate sea lice dispersion as Brooks and Jones (2008) sug-gest. Rather, Krkosek et al. (2007a) investigated the effects of L.salmonis infestations on wild pink salmon population dynam-ics. However, in some of our previous work, we have used amodel of sea lice dispersion as part of the analyses of sea lice in-fecting juvenile salmon migrating past salmon farms (Krkoseket al., 2005, 2006). This model tracked sea lice developmentthrough nauplii and then copepodid stages, and was parameter-ized by data on the current speeds measured in the BroughtonArchipelago (Krkosek et al., 2006). This constrained model,which represents the simplest possible model of sea lice disper-sal, explains the data very well and has been spatially, tempo-rally, and taxonomically replicated (Krkosek et al., 2005, 2006).The predictions cited by Brooks and Jones (2008) of sea lice lar-vae being dispersed from the Broughton Archipelago before thelice reach infectious stages (Brooks, 2005) are probably over-estimated because the models overestimate advective flow, donot include the effects of wind on sea surface water flow, andthey do not represent the behavioral ecology of planktonic lar-val sea lice. Inclusion of these factors can give rise to a widerange of dispersion patterns for sea lice larvae, including thoseobserved in the Broughton Archipelago (Gillibrand and Kate,2007). The theoretical oceanographic model of Gillibrand andKate (2007) does not refute the connections between sea liceand salmon farms in the Broughton Archipelago, but rathershows sea lice infestation patterns observed in the BroughtonArchipelago are within the range of those predicted bytheir model.
f. Prophylactics and Sea Lice Dispersion
Our Science report (Krkosek et al., 2007a) tested for andquantified the effects of L. salmonis infestations of wild juve-nile pink salmon on wild pink salmon population dynamics in theBroughton Archipelago. Contrary to Brooks and Jones (2008)interpretation of Krkosek et al. (2007a), we did not model sealice dispersion, assume a steady state dispersion model, estimatethe production of sea lice nauplii on salmon farms, or investigatethe effects of prophylactic treatment on sea lice abundances onfarm salmon. Brooks and Jones (2008) claim the prophylactictreatments have reduced sea lice abundance on salmon farms,which may be the case, but they provide no supporting evi-dence for their claim. The effects of prophylactic use on sealice control on salmon farms and sea lice transmission fromfarm to wild salmon is an important line of future research,
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 1
6:5
6 2
4 A
pril 2008
SEA LICE AND PINK SALMON DECLINES 417
but was not the focus of our Science report (Krkosek et al.,2007a).
4. ANALYSIS OF THE ENTIRE PINK SALMONDATABASE SUPPORTS CONCLUSIONS OPPOSITETO THOSE REACHED BY KRKOSEK ET AL. (2007)
a. Misleading Regression Analysis
In our Science report (Krkosek et al., 2007a), we did not con-duct a regression analysis. Rather, we fit the stochastic Rickermodel to estimates of the number of pink salmon returning an-nually to rivers on the Central Coast of British Columbia. TheRicker model is well established in fisheries science (Hilbornand Walters, 2001) and is the preferred method for estimatingpopulation growth rates (Myers et al., 1999) and testing for den-sity dependence in time series data of population abundances(Dennis and Taper, 1994; Mueter et al., 2002). Brooks and Jones(2008) place high emphasis on the coefficients of determinationin their regression analysis, but these statistics are meaninglessin this context because of the problem of non-independence be-tween xand yvariables. Because of the dependence of ln[n(t+2)/n(t)] on n(t) in the Ricker model, the variation in the data mustbe represented by parametric bootstrapping (Dennis and Taper,1994). Brooks and Jones (2008) advocate using non-linear re-gression to analyze the data, but fail to describe how the variationin the data can be correctly represented in such an analysis. Morefundamentally, they fail to explain the biological significance ofsuch an analysis.
b. Incomplete Model
Brooks and Jones (2008) suggest the stochastic Ricker modelused in Krkosek et al. (2007a) was inadequate to model pinksalmon population dynamics. The stochastic Ricker model isa nonlinear stochastic model that captures density-dependentmortality as well as stochastic environmental variation (Dennisand Taper, 1994). The Ricker model may be the most funda-mental model in fisheries science and is used widely in fish-eries and ecology (Dennis and Taper, 1994; Myers et al., 1999;Hilborn and Walters, 2001; Brook and Bradshaw, 2006). TheRicker model directly models overcompensation - the cycle ofpopulation crash following high brood-year abundance (Kot,2001) and so fully accommodates the high spawner abundancein 2000. The population growth rate, r , is estimated as the inter-cept, and density-dependent mortality is estimated as the slopewhen fitting the stochastic Ricker model (Dennis and Taper,1994), meaning that these two parameters are well distinguished.Figure 3 in Krkosek et al. (2007a) shows that a common slopeparameter (strength of density dependence) fit all the data well,including the data from the Broughton Archipelago during thesea lice infestations. This indicates at least two things. First, thesea lice infestations have affected the population growth rate butnot density-dependent mortality, which is expected from theory
(Krkosek et al., 2007a, 2007b). Second, and importantly, thedensity-dependent parameter estimated from all the data rep-resents well the high spawner density and subsequent collapsethat occurred for Broughton pink salmon populations in 2002.This latter point can be seen by the fact that there are no majoroutliers in the data from the model fit in panel C relative to themodel fits in panels A and B in Figure 3 in Krkosek et al., 2007a.Brooks and Jones (2008) introduce a new alternative model thatsupposedly tracks fresh and marine water quality, predation,and several density-dependent factors such as food availability,horizontally transmitted disease agents, and random disease ef-fects. The model looks interesting, but Brooks and Jones (2008)provide no quantitative basis for its validity, nor any empiricalevidence that it is a suitable alternative model. It is not clearhow the Brooks and Jones (2008) model accounts mechanis-tically for the effects they list, and it is difficult to imagine asuitable dataset that contains sufficient information to estimateall the parameters.
c. Selective Use of Data
Brooks and Jones (2008) combine all the pink salmon popula-tions as though they were one population. The problem with thisapproach is that the functional unit in the pink salmon metapop-ulation structure is the odd- or even-year lineage in each individ-ual river, not the regional summation of many rivers. Figures 2and S1 in Krkosek et al. (2007a) show clearly that there is sub-stantial variation in the data at the individual river level. Thisvariation and the information it contains about the processesunderlying it is lost when simply summing the escapement es-timates. The natural periodic declines in the summed escape-ment data do not provide an alternate explanation to our resultsin Krkosek et al. (2007a). A more careful examination of thevariation in the individual populations of Figure 2 in Krkoseket al. (2007a) shows that the catastrophic event currently affect-ing Broughton pink salmon populations is unprecedented, ratherthan common. Table 1 shows that all the periods of decline insummed escapement estimates identified by Brooks and Jones(2008) do not yield negative growth rates and extinction predic-tions when applying our analytical approach. This is becausetrends in summed escapement data reflect trends in only thelargest populations, and, while the largest populations may havedeclined due to nonlinear density dependence, the smaller pop-ulations did not. Similar obfuscation of ecological processes byanalyzing data at the stock complex level rather than the individ-ual river level is known elsewhere in fisheries, where simulationmodels show that evidence for depensation is lost at the stockcomplex level when it is in fact present at the individual popu-lation level (Frank and Brickman, 2000). With the analysis thatBrooks and Jones (2008) conduct, some populations could goextinct, and that may not be statistically detectable or even evi-dent in the data if other populations were healthy, declining lessrapidly, or equally declining but starting with higher abundance.Because the Glendale populations had recently increased due
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 1
6:5
6 2
4 A
pril 2008
418 M. KRKOSEK ET AL.
to the spawning channel, this hides the trends happening in theother populations. By analyzing the data at a level that permitsexamination of all the variation it contains, it becomes clearthat all the populations, including Glendale, had significantlynegative population growth rates during the sea lice infestations(Krkosek et al., 2007a).
d. Incomplete Analysis
The alternate analysis in this section of Brooks andJones (2008) involves fitting arbitrary polynomials to log-transformed summed escapement and log-transformed timevariables. Brooks and Jones (2008) do not provide a biolog-ical basis for the analysis or discuss the associated scientificinference. It is not clear to us what can be learned about the ef-fects of sea lice on pink salmon population dynamics by fitting acubic function to seven data points on log-transformed summedescapement and log-transformed year axes. The presentation byBrooks and Jones (2008) of the stochastic Ricker model as alinear regression through a plot of log escapement vs log year isnot correct. The correct method for fitting the stochastic Rickermodel and comparing parameter estimates is conducted by lin-ear regression of ln[n(t+ 2)/n(t)] vs n(t) to obtain the pointestimates of the parameters and then parametric bootstrappingto construct the 95% confidence intervals on the parameter esti-mates (Dennis and Taper, 1994).
e. Current Trends in Pink Salmon Returns
Brooks and Jones (2008) suggest that recent trends inBroughton Archipelago pink salmon populations are increasingrather than decreasing (their Figure 8). Their analysis is basedon regionally summed escapement estimates, and we have al-ready explained how this obfuscates the variation and processesin the underlying data. But there are more fundamental flawsin their analysis. Odd- and even-year pink salmon lineages areindependent populations that commonly have different abun-dances, population dynamics, and divergent genetics. It can beappropriate to pool data from odd- and even-year lineages if thedata are first scaled by odd- and even-year mean abundancesto make the data comparable (Krkosek et al., 2007a). The datamust then be analyzed on n(t+ 2) vs n(t) or ln[n(t+ 2)/n(t)]
Table 1 Point estimates of the population growth rate, r , for subsets ofthe Broughton Archipelago pink salmon escapement database suggestedby Brooks and Jones (2008) to be periods of natural decline
Data period Population growth rate, r
Odd years 1959–1969 0.45Odd years 1983–1991 0.39Odd years 2001–2003 Sea lice infestation yearsEven years 1976–1984 0.12Even years 1990–1996 0.64Even years 2000–2006 Sea lice infestation years
vs n(t) axes to represent the autocorrelation in the time seriescorrectly. In their Figure 8, Brooks and Jones (2008) analyze thedata as though the data points are all independent, which they arenot, and the odd- and even-year lineages are not comparable be-cause they have not been scaled appropriately. After excludingthe year following the fallow for outmigrating juveniles (2004),there is only one valid n(t+ 2) and n(t) data pair present in theiranalysis (2003 and 2005). Despite these fundamental flaws, weattempted to verify the analysis in Figure 8 of Brooks and Jones(2008) using summed escapement data from the seven rivers inour Science report (Krkosek et al., 2007a) plus Glendale Creek.We found that the increasing trend claimed by Brooks and Jones(2008) in unscaled summed escapement data in the years 2002–2006 (excluding 2004) is not statistically significant (t = 2.87,df = 2, p = 0.103), and when the data are appropriately scaledthe statistical results are further weakened (t = 1.38, df = 2, p =0.302). This means that Brooks and Jones (2008) conclusion thatBroughton Archipelago pink salmon are recovering is based ona statistically insignificant result from a flawed analysis. A care-ful examination of all the variation in the individual populationsof Figures 2 and 3 in Krkosek et al. (2007a) indicates steep andunprecedented declines in Broughton Archipelago pink salmonpopulations during the sea lice infestations. This is confirmed byexamination of Table 2, which shows that the survival of pinksalmon populations during the infestation years was negativeoverall, with some underlying variation. The underlying vari-ation is expected due to the high stochasticity in the data andmodel. While the populations decline overall, there are tempo-rary exceptions due to the stochastic process (Figure 1).
Figure 1 Simulation of the stochastic extinction model n (t+ 2) = n(t)·exp(r + Zt (0,v)) described and parameterized in Krkosek et al. (2007a). Pa-rameters are r , the population growth rate (r = –1.17), and v, the variance (v =1.91). The environmental stochasticity term, Z , is a normally distributed randomvariable with mean 0 and variance v, which is drawn each generation. There are100 stochastic simulations shown (grey open circles), all starting at historicalmean abundance n (t = 0) = 1. The solid black line is the decline predicted bythe deterministic model n (t+ 2) = n (t)· exp(r ).
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 1
6:5
6 2
4 A
pril 2008
SEA LICE AND PINK SALMON DECLINES 419
Table 2 Annual survival estimates, ln[n (t)/n (t – 2)] per pink salmonpopulation in the Broughton Archipelago during 2002–2006
2002 2003 2004* 2005 2006
Ahta −4.48 −3.16 3.40 1.05 −2.73Kakweiken −5.03 −1.93 2.46 0.75 −0.55Kingcome −3.79 −1.08 2.63 −1.94 −1.93Wakeman −2.54 1.02 0.86 −3.65 −1.55Viner −0.51 −2.10 1.57 −1.05 −3.02Lull −6.73 −5.31 4.28 3.55 −3.16Ahnuhati −4.11 0.40a2.82 −0.01 −2.54Glendale −3.73 −2.12 3.59 0.32 −1.29Average −3.87 −1.79 2.70 −0.12 −2.10
∗This year corresponds to the pink salmon cohort whose juvenile outmigrationoccurred during the fallow year 2003.
ACKNOWLEDGMENTS
We thank several colleagues for helpful comments. Fund-ing came from the Natural Science and Engineering ResearchCouncil of Canada and a Canada Research Chair.
REFERENCES
Boxaspen, K. A review of the biology and genetics of sea lice. ICES J.Mar. Sci., 63: 1304–1316 (2006).
Bricknell, I. R., S. J. Dalesman, B. O’Shea, C. C. Pert, and A. J.M. Luntz. Effect of environmental salinity on sea lice, Lepeoph-theirus salmonis, settlement success. Dis. Aquat. Org., 71: 201–212 (2006).
Brook, B. W., and C. J. A. Bradshaw. Strength of evidence for densitydependence in abundance time series of 1198 species. Ecology, 87:1445–1451 (2006).
Brooks, K. M. The effects of water temperature, salinity, and currentson the survival and distribution of the infective copepodid stage ofsea lice (Lepeophtheirus salmonis) originating on Atlantic salmonfarms in the Broughton Archipelago of British Columbia, Canada.Rev. Fish. Sci., 13: 177–204 (2005).
Brooks, K. M., and S. R. M. Jones. Perspectives on pink salmon and sealice: Scientific evidence fails to support the extinction hypothesis.Rev. Fish. Sci., 16: 403–412 (2008).
Dennis, B., and M. L. Taper. Density dependence in time series observa-tions of natural populations: Estimation and testing. Ecol. Monogr.,64: 205–224 (1994).
Ford, J. S., and R. A. Myers. A global assessment of salmon aquacultureimpacts on wild salmonids. PLoS Biol., 6: e33 (2008).
Frank, K. T., and D. Brickman. Allee effects and compensatory popu-lation dynamics within a stock complex. Can. J. Fish. Aquat. Sci.,57: 513–517 (2000).
Gillibrand, P. A., and J. W. Kate. Dispersal of sea lice larvae from salmonfarms: A model study of the influence of environmental conditionsand larval behavior. Aquat. Biol., 1: 63–75 (2007).
Grenfell, B. T., and A. P. Dobson. Ecology of Infectious Diseases in Nat-ural Populations. Cambridge: Cambridge University Press (1995).
Groot, C., and L. Margolis. Pacific Salmon Life Histories. Vancouver:UBC Press (1991).
Heuch, P. A. Experimental-evidence for aggregation of salmon lousecopepodids (Lepeophtheirus salmonis) in Step Salinity Gradients. J.Mar. Biol. Asso. UK, 75: 927–939 (1995).
Heuch, P. A., A. Parsons, and K. Boxaspen. Diel vertical migra-tionA possible host-finding mechanism in salmon louse (Lepeoph-theirus salmonis) copepodids. Can. J. Fish. Aquat. Sci., 52: 681–689 (1995).
Hilborn, R., and C. J. Walters. Quantitative Fisheries Stock Assessment.Norwell, MA: Kluwer (2001).
Jones, S., E. Kim, and S. Dawe. Experimental infections with Lepeoph-theirus salmonis (Kroyer) on three-spine sticklebacks, Gasterosteusaculeatus L., and juvenile Pacific salmon, Oncorhynchus spp. J. FishDis., 29: 489–495 (2006a).
Jones, S. R. M., G. Prosperi-Porta, E. Kim, P. Callow, and N. B. Har-greaves. The occurrence of Lepeophtheirus salmonis and Caligusclemensi (Copepoda: Caligidae) on three-spine stickleback Gas-terosteus aculeatus in coastal British Columbia. J. Parasitol., 92:473–480 (2006b).
Jones, S. R. M., M. D. Fast, S. C. Johnson, and D. B. Groman. Differ-ential rejection of salmon lice by pink and chum salmon: Diseaseconsequences and expression of proinflammatory genes. Dis. Aquat.Org., 75: 229–238 (2007).
Kot, M. Elements of Mathematical Ecology. Cambridge: CambridgeUniversity Press (2001).
Krkosek, M., A. Gottesfeld, B. Proctor, D. Rolston, C. Carr-Harris, andM. A. Lewis. Effects of host migration, diversity, and aquacultureon sea lice threats to Pacific salmon populations. Proceedings of theRoyal Society of London, Series B, 274: 3141–3149 (2007b).
Krkosek, M., J. S. Ford, A. Morton, S. Lele, R. A. Myers, and M. A.Lewis. Declining wild salmon populations in relation to parasitesfrom farm salmon. Science, 318: 1772–1775 (2007a).
Krkosek, M., M. A. Lewis, A. Morton, L. N. Frazer, and J. P. Volpe.Epizootics of wild fish induced by farm fish. Proceedings of theNational Academy of Sciences of the USA, 103: 15506–15510(2006).
Krkosek, M., M. A. Lewis, and J. P. Volpe. Transmission dynamics ofparasitic sea lice from farm to wild salmon. Proceedings of the RoyalSociety of London Series B, 272: 689–696 (2005).
Lloyd-Smith, J. O., P. C. Cross, C. J. Briggs, M. Daugherty, W. M. Getz,J. Latto, M. S. Sanchez, A. B. Smith, and A. Swei. Should we expectpopulation thresholds for wildlife disease? Trends Ecol. Evol., 20:511–519 (2005).
May, R. M., and R. M. Anderson. Infectious Diseases of Humans. Ox-ford: Oxford University Press (1991).
Morton, A., and R. Routledge. 2005. Mortality rates for juvenile pinkOncorhynchus gorbuscha and chum O. keta salmon infested with sealice Lepeophtheirus salmonis in the Broughton Archipelago. AlaskaFish. Res. Bull., 11: 146–152.
Morton, A., R. Routledge, C. Peet, and A. Ladwig. Sea lice (Lepeoph-theirus salmonis) infection rates on juvenile pink (Oncorhynchusgorbuscha) and chum (Oncorhynchus keta) salmon in the nearshoremarine environment of British Columbia, Canada. Can. J. Fish.Aquat. Sci., 61: 147–157 (2004).
Morton, A., R. D. Routledge, and R. Williams. Temporal patterns of sealouse infestation on wild Pacific salmon in relation to the fallowingof Atlantic salmon farms. N. Am. J. Fish. Manage., 25: 811–821(2005).
Morton, A. B., and R. Williams. First report of a sea louse, Lepeoph-theirus salmonis, infestation on juvenile pink salmon, Oncorhynchus
reviews in fisheries science vol. 16 no. 4 2008

Dow
nlo
aded B
y: [C
anadia
n R
esearc
h K
now
ledge N
etw
ork
] A
t: 1
6:5
6 2
4 A
pril 2008
420 M. KRKOSEK ET AL.
gorbuscha, in nearshore habitat. Can. Field Nat., 117: 634–641(2003).
Morton, A. M., R. Routledge, and M. Krkosek. Sea lice infestation ofwild juvenile salmon and herring associated with fish farms off theeast central coast of Vancouver Island, BC. N. Am. J. Fish. Manage.,in press (2008).
Mueter, F. J., R. M. Peterman, and B. J. Pyper. Opposite effects ofocean temperature on survival rates of 120 stocks of Pacific salmon(Oncorhynchus spp.) in northern and southern areas. Can. J. Fish.Aquat. Sci., 59: 456–463 (2002).
Myers, R. A., K. G. Bowen, and N. J. Barrowman. Maximum repro-ductive rate of fish at low population sizes. Can. J. Fish. Aquat. Sci.,56: 2404–2419 (1999).
Orr, C. Estimated sea louse egg production from Marine HarvestCanada farmed Atlantic salmon in the Broughton Archipelago,British Columbia, 2003–2004. N. Am. J. Fish. Manage., 27: 187–197 (2007).
Peet, C. R. Interactions between sea lice (Lepeophtheirus salmo-nis and Caligus clemensi) and juvenile chum (Oncorhynchusketa) and pink salmon (Oncorhynchus gorbuscha) in BritishColumbia. M.Sc. Thesis. University of Victoria, Victoria, Canada(2007).
Pike, A. W., and S. L. Wadsworth. Sea lice on salmonids: Their biologyand control. Adv. Parasitol., 44: 233–337 (2000).
Pyper, B. J., F. J. Mueter, R. M. Peterman, D. J. Blackbourn, and C.C. Wood. Spatial covariation in survival rates of Northeast Pacificpink salmon (Oncorhynchus gorbuscha). Can. J. Fish. Aquat. Sci.,58: 1501–1515 (2001).
Ricker, W. E. Stock and recruitment. J. Fish. Res. Board Can., 11:559–623 (1954).
Rothman, K. J. Modern Epidemiology. Boston: Little Brown (1986).Stien, A., P. A. Bjorn, P. A. Heuch, and D. A. Elston. Population dy-
namics of salmon lice Lepeophtheirus salmonis on Atlantic salmonand sea trout. Mar. Ecol. Prog. Ser., 290: 263–275 (2005).
Webster, S. J., L. M. Dill, and K. Butterworth. The effect of sea liceinfestation on the salinity preference and energetic expenditure of ju-venile pink salmon (Oncorhynchus gorbuscha). Can. J. Fish. Aquat.Sci., 64: 672–680 (2007).
Wertheimer, A. C., E. A. Fergusson, R. L. Focht, W. R. Heard, J. A.Orsi, M. V. Sturdevant, and B. L. Wing. Sea lice infection of juve-nile salmon in the marine waters of the northern region of south-eastern Alaska, May–August, 2003. (NPAFC Doc. 706) Auke BayLab., Alaska Fisheries Science Center, National Marine FisheriesServices, NOAA, Juneau (2003).
reviews in fisheries science vol. 16 no. 4 2008


















