
pbio 33 406 Mp 590 Allen Press GALLEY File # 06TQ et al. 2007.pdf · FOSSIL FOOD WEBS IN LAKE...
20
PALEOBIOLOGY Wednesday Aug 29 2007 01:41 PM Allen Press • DTProN System GALLEY pbio 33_406 Mp_590 File # 06TQ 2007 The Paleontological Society. All rights reserved. 0094-8373/07/3304-0000/$1.00 Paleobiology, 33(4), 2007, pp. 590–609 Stable isotopic evidence for fossil food webs in Eocene Lake Messel Maia K. Schweizer, Andrew Steele, Jan K. W. Toporski, and Marilyn L. Fogel Abstract.—Carbon and nitrogen stable isotopic compositions of fossil materials from Lake Messel (47 Ma) in Germany are used to investigate Eocene ecosystem dynamics. Autolithified soft tissues of terrestrial and aquatic vertebrate organisms, as well as plant compression fossils, contain organic material (20–50 wt% C, 1–6 wt% N), which appears to retain precursor compositions. Stable iso- topic compositions ( 13 C and 15 N) of Messel fossils are similar to those reported for components in modern lacustrine ecosystems. These data show trophically sensible enrichments relative to food sources, reflect multiple feeding strategies for each organism (e.g., omnivory, planctivory, pisci- vory), and differentiate between benthic and pelagic organic carbon sources. These chemical data broadly confirm existing Messel food web models based on coprolite and gut content analyses. 13 C values for the lacustrine shale range from 30.3 to 26.3‰, pointing to mixed terrestrial and aquatic origins for primary producers in the food web. 13 C values for primary consumers such as insects overlap with those for primary producers but are comparatively enriched in 15 N. Secondary and higher consumers (fish, crocodiles, and frogs) are associated with even more positive 15 N values and show a more constrained range of 13 C values. Omnivory appears widespread in both low and high trophic level consumers. Hence, the stable isotopic compositions of Messel fossils are complex and overlap, and must be combined with paleontological investigations in order to be conclusive. This study represents the first comprehensive isotopic reconstruction, featuring tens of components, of an ecosystem of Eocene age. A thorough understanding of trophic structure in Eo- cene Lake Messel contributes to the global databank of ecological history. Maia K. Schweizer,* Andrew Steele, Jan K. W. Toporski, † and Marilyn L. Fogel. Geophysical Laboratory, Carnegie Institution of Washington, Washington, D.C. 20015. E-mail: [email protected] *Present address: Department of Earth Sciences, Oxford University, Oxford, United Kingdom OX1 3PR †Present address: Department of Mineralogy, Institut fu ¨r Geowissenshaften der Christian-Albrechts- Universita ¨t zu Kiel, Kiel, Germany 24118 Accepted: 13 June 2007 Introduction Our understanding of modern ecosystems would be incomplete without investigations of fossil ecosystems, which may reveal how or- ganisms adapt to long-term environmental changes. However, of the diversity of tools available to modern ecologists—including sta- ble isotope tracers (e.g., Hecky and Hesslein 1995), biomanipulation experiments (e.g., Mehner et al. 2002), and modeling based on thousands of individual observations (Dunne et al. 2004)—only a few can be applied to an- cient ecosystems. The diversity of organisms constituting fossil ecosystems is winnowed by taphonomic processes, and chemical signa- tures are often not observable because even exquisite fossillagersta ¨tten such as the Bur- gess shale and Solnhofen limestone seldom preserve significant quantities of organic mat- ter. Food web analyses of ancient ecosystems therefore rely in large part on paleontological evidence (e.g., Baszio and Richter 2001; Rich- ter and Wedmann 2005), supplemented with limited carbon isotope data. Paleontological food web reconstructions are based on near- est-living-relative comparisons as well as cop- rolite and gut content analyses. They therefore inevitably oversimplify trophic chains because it is difficult to differentiate between whole and partial trophic levels on the basis of iso- lated remains. Fossil organisms are typically represented only by their resistant hard parts, which usually contain less than 25 wt% or- ganic carbon in the form of collagen (Gearing 1991; Sillen and Lee-Thorp 1994). 13 C mea- sured from the organic component of fossil bones and teeth (Ostrom et al. 1993; Ostrom et al. 1994; Sillen and Lee-Thorp 1994; Boch- erens et al. 1995; Clementz and Koch 2001) has previously been used to reconstruct isolated predator-prey relationships and grazing hab-
Transcript of pbio 33 406 Mp 590 Allen Press GALLEY File # 06TQ et al. 2007.pdf · FOSSIL FOOD WEBS IN LAKE...

PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_590File # 06TQ
� 2007 The Paleontological Society. All rights reserved. 0094-8373/07/3304-0000/$1.00
Paleobiology, 33(4), 2007, pp. 590–609
Stable isotopic evidence for fossil food webs in EoceneLake Messel
Maia K. Schweizer, Andrew Steele, Jan K. W. Toporski, and Marilyn L. Fogel
Abstract.—Carbon and nitrogen stable isotopic compositions of fossil materials from Lake Messel(47 Ma) in Germany are used to investigate Eocene ecosystem dynamics. Autolithified soft tissuesof terrestrial and aquatic vertebrate organisms, as well as plant compression fossils, contain organicmaterial (20–50 wt% C, 1–6 wt% N), which appears to retain precursor compositions. Stable iso-topic compositions (�13C and �15N) of Messel fossils are similar to those reported for componentsin modern lacustrine ecosystems. These data show trophically sensible enrichments relative to foodsources, reflect multiple feeding strategies for each organism (e.g., omnivory, planctivory, pisci-vory), and differentiate between benthic and pelagic organic carbon sources. These chemical databroadly confirm existing Messel food web models based on coprolite and gut content analyses. �13Cvalues for the lacustrine shale range from �30.3 to �26.3‰, pointing to mixed terrestrial andaquatic origins for primary producers in the food web. �13C values for primary consumers such asinsects overlap with those for primary producers but are comparatively enriched in 15N. Secondaryand higher consumers (fish, crocodiles, and frogs) are associated with even more positive �15Nvalues and show a more constrained range of �13C values. Omnivory appears widespread in bothlow and high trophic level consumers. Hence, the stable isotopic compositions of Messel fossils arecomplex and overlap, and must be combined with paleontological investigations in order to beconclusive. This study represents the first comprehensive isotopic reconstruction, featuring tens ofcomponents, of an ecosystem of Eocene age. A thorough understanding of trophic structure in Eo-cene Lake Messel contributes to the global databank of ecological history.
Maia K. Schweizer,* Andrew Steele, Jan K. W. Toporski,† and Marilyn L. Fogel. Geophysical Laboratory,Carnegie Institution of Washington, Washington, D.C. 20015. E-mail: [email protected]
*Present address: Department of Earth Sciences, Oxford University, Oxford, United Kingdom OX1 3PR†Present address: Department of Mineralogy, Institut fur Geowissenshaften der Christian-Albrechts-
Universitat zu Kiel, Kiel, Germany 24118
Accepted: 13 June 2007
Introduction
Our understanding of modern ecosystemswould be incomplete without investigations offossil ecosystems, which may reveal how or-ganisms adapt to long-term environmentalchanges. However, of the diversity of toolsavailable to modern ecologists—including sta-ble isotope tracers (e.g., Hecky and Hesslein1995), biomanipulation experiments (e.g.,Mehner et al. 2002), and modeling based onthousands of individual observations (Dunneet al. 2004)—only a few can be applied to an-cient ecosystems. The diversity of organismsconstituting fossil ecosystems is winnowed bytaphonomic processes, and chemical signa-tures are often not observable because evenexquisite fossillagerstatten such as the Bur-gess shale and Solnhofen limestone seldompreserve significant quantities of organic mat-ter.
Food web analyses of ancient ecosystems
therefore rely in large part on paleontologicalevidence (e.g., Baszio and Richter 2001; Rich-ter and Wedmann 2005), supplemented withlimited carbon isotope data. Paleontologicalfood web reconstructions are based on near-est-living-relative comparisons as well as cop-rolite and gut content analyses. They thereforeinevitably oversimplify trophic chains becauseit is difficult to differentiate between wholeand partial trophic levels on the basis of iso-lated remains. Fossil organisms are typicallyrepresented only by their resistant hard parts,which usually contain less than 25 wt% or-ganic carbon in the form of collagen (Gearing1991; Sillen and Lee-Thorp 1994). �13C mea-sured from the organic component of fossilbones and teeth (Ostrom et al. 1993; Ostromet al. 1994; Sillen and Lee-Thorp 1994; Boch-erens et al. 1995; Clementz and Koch 2001) haspreviously been used to reconstruct isolatedpredator-prey relationships and grazing hab-

591FOSSIL FOOD WEBS IN LAKE MESSEL
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_591File # 06TQ
its of fossil organisms (MacFadden et al. 1994,1999; Wang et al. 1994; Sponheimer and Lee-Thorp 1999). Nitrogen is even less likely to bepreserved than carbon, yet is crucial to stableisotopic food web reconstructions.
Eocene Lake Messel in Germany allows usto overcome these obstacles. A complete la-custrine ecosystem and a wide range of ter-restrial organisms are exquisitely preserved inthe Messel Fossillagerstatte (e.g., Storch andHabersetzer 1988; Lutz 1990; Richter 1992;Schaal and Ziegler 1992), with soft-body ma-terials and organic matter intact. The Messeldeposit contains one of the most diverse Eo-cene biotas in the world. The present studyseeks to evaluate paleontological models(Baszio and Richter 2001; Richter and Baszio2001) of the ecosystem at Lake Messel by us-ing stable isotope compositions of carbon andnitrogen preserved in fossil vertebrate soft tis-sues, invertebrates, and leaves. Isotope dataobtained from a similarly preserved Oligo-cene ecosystem demonstrate that relative iso-tope enrichments between organisms may bepreserved over geologic time scales, and thatit is therefore possible to reconstruct trophicrelationships on the basis of the stable isotopiccompositions of autolithified soft tissues(Schweizer et al. 2006). If stable isotopic com-positions of fossil materials are preserved, dothey concur with paleontological data? Thedata presented here represent a novel ap-proach to decoding ancient ecosystems by ap-plying modern techniques to fossil materials.
The Eocene was a time of fluctuating pa-leoenvironment (Janis 1993; Torricelli et al.2006), biome evolution (Blondel 2001; Jacobs2004; Jacobs and Herendeen 2004) and high-latitude species migrations and turnover facil-itated by exceptionally warm temperatures(McKenna 1980, 1983; Hartenberger 1987; Le-gendre and Hartenberger 1992; Janis 1993; Pe-ters and Sloan 2000; Blondel 2001) and minorlatitudinal temperature gradients (Andreas-son and Schmitz 1996, 2000; Janis 1993). Inter-estingly, the Messel shale was deposited at thethermal and mammalian species-diversitymaximum of the early middle Eocene (47 Ma),just prior to a major global cooling event thatpersisted through the remainder of the Eoceneand Oligocene (Haq et al. 1987; Zachos et al.
1993; Blondel 2001). This cooling caused a re-placement of the tropical forest biome withopen forested grasslands and a subsequentturnover of mammals and lower vertebratesadapted to new feeding habits (Janis 1993; Za-chos et al. 1993; Blondel 2001). In other words,the Messel shale offers a rare opportunity totake a census of ecosystems that are pre-gla-cial and hyper-greenhouse in climate.
The fossil record of lakes is persistent andreasonably well known, despite the relativelyephemeral nature of individual lakes. Pale-ontological reconstructions have traced theevolution of modern-type lake ecosystemsfrom the Paleozoic to today through the ap-pearance of important organisms such as in-sects and aquatic macrophytes, changing tec-tonic settings, and substantial terrestrially de-rived organic input (Gray 1988; Buatois andMangano 1993; Cohen 2003). Characteristictop-down trophic cascades, where popula-tions of primary producers and consumers aredetermined by higher predators, were estab-lished in lakes by the Mesozoic and appear tohave remained dominant throughout the Ter-tiary with some fluctuations due to the evo-lution and increasing input of terrestrial andaquatic macrophytes (Gray 1988; Cohen 2003).Reconstructions of specific Eocene lakes(Buchheim 1994; Murray 2001; Melchor et al.2002; Kvacek et al. 2004; Sasche 2005) as wellas widespread lake and terrestrial ecosystemdynamics (Barton and Wilson 1999; Janis1993; Wilf et al. 2003; Williams et al. 2003; Wil-son 1987) have shown that many of the sameintertwining factors important to modernecology, such as temperature and precipita-tion variation, species richness, and ease ofmigration controlled Tertiary ecosystems.Specifically, fossil lacustrine food webs appearto have been determined by lake surface area,morphology, longevity, productivity, and sta-bility (Cohen 2003; Gierlowski-Kordesch andPark 2004). Additionally, fossil and modernlakes exhibit major offshore versus nearshorevariation in organism assemblages, due tohabitat preferences of aquatic organisms andsorting during transportation of allochthon-ous terrestrial organisms (Roth and Dilcher1978; Wilson 1980, 1988).
Site Description. The fossiliferous Messel

592 MAIA K. SCHWEIZER ET AL.
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_592File # 06TQ
oil shale (�8 m thick) was deposited at the an-oxic benthic layer of a large maar lake (Fran-zen 1990; Harms 2002; Robinson et al. 1989;Rolf et al. 2005; Wonik 2005). The lake basinwas formed when rising magma came intocontact with groundwater, explosively vapor-izing the water and leaving behind a steep-sided crater during a period of local volca-nism (Muller-Sohnius et al. 1989; Pirrung etal. 2003) at approximately 47 Ma (Mertz et al.2004; Mertz and Renne 2005) in the middleEocene. Traces of photosynthetic archaebac-teria (Meyers 1997) indicate that even part ofthe photic zone was anoxic. The lake wastherefore strongly stratified. In the Eocene,central Europe was 10� closer to the equator(Schaarschmidt 1992) and mean annual tem-perature ranged from 20� to 30�C (Andreassonand Schmitz 2000; Mai 1981; Markwick 1996,2002; Matthes 1968; Schaarschmidt 1992; Sit-tler 1968; Thiele-Pfeiffer 1988; Wilde 1989)though seasonal variation was similar in mag-nitude to variation today (Andreasson andSchmitz 2000; Schmitz and Andreasson 2001).Lake Messel was surrounded by a dense par-atropical forest (Kohler 1997; Thiele-Pfeiffer1988; Wilde 1989; Lenz et al. 2005) from whichplentiful vegetation and vertebrate carcasseswere carried into the lake. Leaves (Collinson1986), pollen and spores (Schaarschmidt1992), and seeds and fruit are found at Messelthat belong to club mosses, ferns, gymno-sperms, and over 200 species of angiospermsdominated by walnuts and laurels. Palynolog-ical analysis reveals that floral and faunalcomposition varied during the deposition ofthe Messel oil shale because of water table var-iations, increasing paleotemperatures, andMilankovitch cycles (Lenz et al. 2005). Thecontent of coprolites suggests that the aquaticfood web was stable throughout the deposi-tion of the fossiliferous shale, representingsome 50 to 100 Kyr (Goth 1990; Richter andBaszio 2001), though there is evidence for dif-ferent food web compositions lower in theshale sequence (Richter and Wedmann 2005).
Nearest-living-relative comparisons (Schaaland Ziegler 1992) and gut and coprolite con-tent analyses (Schmitz 1991; Baszio and Rich-ter 2001; Richter and Baszio 2001) illuminatepossible omnivorous, insectivorous, carnivo-
rous, and folivorous diets (Richter 1987; Rich-ter and Baszio 2001) for Messel organisms. Apaleontological reconstruction synthesized byRichter and Baszio (2001) describes a six-ti-ered, linear trophic chain. Aquatic primaryproducers included pelagic Tetraedron and Bo-tryococcus (Goth 1990), benthic and floating al-gae, dinoflagellates, monocellular green algaein blooms, and water lilies (Franzen and Mi-chaelis 1988; Hayes et al. 1989; Robinson et al.1989; Goth 1990; Meyers 1997; Nix and Feist-Burkhardt 2003). These were consumed byCladocera, Culicidae, ostracods, caddis flies(Trichoptera), mayflies (Ephemeroptera), andfreshwater sponges. Secondary consumerssuch as water bugs fed on small Crustacea. Ju-venile fish and smaller fish species includingThaumaturus preyed upon such Arthropodaand constituted the tertiary-level consumers.The fourth level of consumers was composedof large predatory fish that preyed on juve-niles, smaller fish species, and large insects(Micklich 1992). Crocodiles, turtles, and pi-scivorous mammals constituted the fifth andhighest level of consumers.
Stable Isotopes as Ecological Tracers. Stableisotopes of carbon and nitrogen (�13C and�15N, respectively) are powerful tools for trac-ing nutrient flow within ecosystems. The ni-trogen isotopic composition of organisms ispassed from primary producers to prey topredator with a fractionation of �1‰ to �5‰�15N (average �3.4‰) per trophic step(DeNiro and Epstein 1981; Minagawa andWada 1984; Harrigan et al. 1989; Vander Zan-den and Rasmussen 2001) whereas �13C re-flects the source carbon signature with a min-imal fractionation of less than �1‰ (DeNiroand Epstein 1978; Fry and Sherr 1984; VanderZanden and Rasmussen 2001; McCutchan etal. 2003). Stable isotope analyses of ecosystemcomponents allow the tracing of multiple car-bon and nitrogen sources and may thereforeyield a nuanced food web instead of a simpli-fied food chain (Cohen et al. 1990; Pimm et al.1991; Kling et al. 1992; Vander Zanden et al.1999). For example, consumer isotopic com-positions differentiate between benthic algaeand pelagic phytoplankton carbon sources(Hecky and Hesslein 1995).
Heterotrophic bacterial reworking of pri-

593FOSSIL FOOD WEBS IN LAKE MESSEL
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_593File # 06TQ
mary organic matter in Eocene Lake Messelwas undoubtedly widespread (Freeman et al.1990) and its effect on the stable isotopic com-position of fossil materials must be consid-ered. Unlike autotrophic microorganisms,which often produce substantial isotopic frac-tionations between source carbon (e.g., CO2)and nitrogen (e.g., N2) and bacterial biomass(Estep et al. 1978; Macko et al. 1982; Whiticaret al. 1986), heterotrophic bacteria such asthose that consumed and replaced Messel ver-tebrate tissues generally incorporate �13C and�15N signatures from the substrate withoutconsiderable fractionation (DeNiro and Ep-stein 1978; Coffin et al. 1989; Coffin et al.1990). Even if replacement is not complete,�13C of the bacterial biomass typically fallswithin 1.0 to 1.5‰ (average ��13C �1.4‰)of the substrate (Hullar et al. 1996). Localizedvariations in stable isotopic composition of in-dividual biomarkers or diagenetic reactionsites tend to balance out and do not substan-tially affect bulk isotopic compositions of a di-verse microbial community (Macko and Estep1984; Blair et al. 1985; Hayes et al. 1989; Coffinet al. 1990; Boschker et al. 1999; Panacost andSinninghe Damste 2003). Although heteroge-neous preservation and microbial processingmay elevate C/N ratios far above what wouldbe expected for living organisms (DeNiro1985), stable isotopic compositions of C and Ncomponents tend to be preserved (Masters1987).
Estimation of trophic position based on�15N values of fossil materials is particularlysensitive to microbial processes because asmall amount of fractionation by microbescould significantly affect apparent trophic lev-el. Other microbial processes such as denitri-fication can greatly influence 15N. Additional-ly, isotopic trophic shifts between primaryproducers, primary consumers, and predatorscan vary. For example, consumers of inverte-brates have a lower than average trophic shiftrelative to their diet (�1.4 � 0.21‰ �15N) thando consumers of other high-protein diets(�3.3 � 0.26‰). Primary consumers feedingon plants and algae exhibit �2.2 � 0.30‰ en-richment in 15N per trophic step (McCutchanet al. 2003). Furthermore, stable isotope com-positions of organism tissues are more likely
to reflect the integration of organic carbon andnitrogen sources rather than the ingested ma-terials themselves (Hecky and Hesslein 1995).These uncertainties require a careful evalua-tion of isotopic data for both modern and an-cient food webs.
Taphonomy of Messel Organisms. Messelleaves and seeds are preserved as compres-sion or impression fossils (Wilde 1989; Man-chester et al. 1994). Beetles with chitinous cu-ticles are the most common insects preserved(Lutz 1992; Wuttke 1992b), while feathers aretypically preserved as replicas made bystrings of autolithified rod-shaped bacteria,arranged end to end (Davis and Briggs 1995).
Vertebrate carcasses were autolithified bysulfate-reducing and denitrifying bacteriasoon after deposition on the bottom of the lake(Wuttke 1983, 1992b,c). By this process, bac-teria inhabiting the mostly anoxic bottom wa-ters immediately colonized the carcasses andbegan releasing metabolic products such asCO2 and H2S into sediment pore waters,which already contained CO2 and iron sup-plied by weathering of nearby Permian strata(Richter 1992). Siderite reached saturation andprecipitated directly onto cell surfaces orwithin the microorganisms themselves untilthe rapidly thickening crust exterminated thebacteria (Toporski et al. 2002).
Messel shale contains a significant amountof low-maturity organic carbon. Kerogen andother materials from Messel are known to con-tain virtually unaltered biolipids (Mycke andMichaelis 1986; Mycke et al. 1987; Robinson etal. 1989) and original albumin (Richter andStorch 1980; von Koenigswald et al. 1981). Re-ducing conditions in the hypolimnion causedby high productivity in the surface waters(Matthes 1966) prevented thorough decom-position of organic matter, and mild burialconditions (�40�C and 200 m depth [Hayes etal. 1987; Kimble et al. 1974; Matthes 1968])preserved the organic matter with little alter-ation. Additionally, autolysis following thedeath of the organisms may have been inhib-ited by humic acids associated with organicmolecules in runoff (Hummel and Herrmann1994), by volcanic gases, or by iron from vol-canic ash (Ferris et al. 1988; Carson 1991). Finevolcanic ash particles and microbial mats fur-

594 MAIA K. SCHWEIZER ET AL.
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_594File # 06TQ
TABLE 1. Common vertebrate, invertebrate, and plant fossils analyzed from the Messel Fossillagerstatte.
Taxon Description Size
Thaumaturus intermedius Small modern fish 15 to 50 mm longCyclurus kehreri Medium-sized bowfin 10 to 60 cm longAtractosteus strausi Large-sized plated garfish 15 to 90� cm longFormicium giganteum Giant ant 10 to 30 mm longElateridae (family) Click beetles Widely variedBuprestidae (family) Jewel beetles Widely variedScarabaeidae (family) Dung beetles Widely variedCurculionidae (family) Weevils 2 to 10 mm longTrichoptera (order) Caddis fly 4 to 12 mm longBlattodea (order) Cockroach 5 to 25 mm longEphemeroptera (order) Mayfly 5 to 20 mm longNymphaeaceae (family) Water lily Up to 10 cm wide
ther preserved the structure of organisms byphysically isolating them and protecting themfrom disruption by turbulence (Zimmerle1993; Harding and Chant 2000; Meyer et al.2001).
Methods
Material Collection. Material from the Mes-sel Fossillagerstatte was collected in July andSeptember 2003 from the Messel pit, located20 km southeast of Frankfurt. Fossils were ex-cavated from a 1-m-thick exposure of shale,the bottom of which is approximately 80 cmabove the horizon (Franzen et al. 1982). Themajority of fossils were subsampled in thefield. A hand-held digital microscope with a100� lens (Scalar DG2) was used to investi-gate the quality of preservation prior to sam-pling in instances where the amount of ma-terial preserved was unclear. Several milli-grams of macroscopically representative ma-terial were loosened with cleaned andsterilized micropicks and placed in aluminumfoil bags, which had previously been cleanedby heating them to 500�C in a muffle furnace.Shale was collected from within a few milli-meters of each fossil to provide a comparisonwith the fossil material. All samples werequickly refrigerated to restrict fungal growthon the organic-rich material, and remained re-frigerated during transport. Upon reachingthe laboratory, the fossil material was kept fro-zen at �20�C.
Seeds were pried whole from the shale, andfoliar material could be peeled from the shalewith little or no sediment contamination. Bac-terial biofilms preserving soft tissues in ver-
tebrate fossils were scraped from the shaleand from bone surfaces as a fine powder. Thedetailed morphological preservation of thespecimens allowed different tissue types to bedistinguished and extracted. Sample materialfrom vertebrate fossils was classified as pure‘‘soft tissues’’ consisting of muscle and inter-nal organs or occasionally as a specific mor-phological feature (such as an eye). Theseterms are based on anatomy and are usedthroughout this paper for simplification,though it should not be forgotten that the orig-inal organic material of the soft tissues hasbeen preserved by bacterial biofilms. A selec-tion of the fossils we sampled are listed anddescribed in Table 1.
Trophic webs for the terrestrial and aquaticecosystems at Messel were reconstructedbased primarily from relationships betweenstable isotopic compositions of fossil organ-isms and secondarily from paleontologicaldata from previous studies.
Stable Isotope Analysis. An aliquot between400 and 2000 �g of each sample was weighedinto a 3.5 � 5 mm tin capsule. The tin capsuleswere sealed and introduced into an elementalanalyzer where they were combusted and re-duced prior to analysis of stable isotope ratiosusing continuous-flow, stable isotope ratiomass spectrometry, as previously describedfor similar samples (Schweizer et al. 2006).Both N2 and CO2 samples were analyzed rel-ative to gas standards and the results are pre-sented in standard notation, defined:
h lX / Xsample sampleh� X � 1 � 1000.h l� �X / Xstd std

595FOSSIL FOOD WEBS IN LAKE MESSEL
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_595File # 06TQ
TABLE 2. Summary of average isotope values, and C and N content (�1 SD) of the most common sample types.
Material (n) �13C �15N %C (wt) %N (wt) C/N (wt)
Shale (159) �28.54 � 2.71 6.76 � 1.06 20.94 � 4.02 0.62 � 0.10 33.77Terrestrial leaves (63) �26.81 � 0.83 7.04 � 0.89 38.84 � 6.42 1.46 � 0.38 26.60Terrestrial seeds (4) �25.81 � 1.17 7.00 � 0.81 32.90 � 4.05 1.12 � 0.19 29.38Wood (14) �26.21 � 1.02 7.12 � 0.50 36.60 � 5.28 1.58 � 0.35 23.16Palm fruits (8) �25.30 � 0.51 6.76 � 0.77 33.05 � 4.02 1.42 � 0.23 23.27Water lily leaves (16) �27.03 � 0.75 5.78 � 1.23 43.26 � 6.55 0.95 � 0.33 45.54Beetles (20) �26.69 � 1.63 6.44 � 1.43 33.16 � 5.90 1.64 � 0.43 20.22Caddis fly larvae (8) �28.90 � 1.39 7.03 � 1.63 3.32 � 2.77 0.12 � 0.09 27.67Other insects (9) �26.87 � 1.58 7.44 � 1.49 32.63 � 8.86 1.53 � 0.53 21.33Thaumaturus soft tissue (14) �26.95 � 1.55 8.07 � 1.87 28.86 � 7.97 1.77 � 0.54 16.31Cyclurus soft tissue (4) �26.99 � 0.41 11.78 � 2.19 35.57 � 7.94 3.76 � 0.98 9.46Atractosteus soft tissue (3) �26.53 � 1.54 8.57 � 0.71 26.99 � 13.83 1.71 � 1.02 15.78Cyclurus hard parts (25) �19.51 � 3.58 6.39 � 2.26 4.38 � 2.54 0.13 � 0.08 33.69
Precision was calculated from triplicatemeasurements of the same acetanilide(C8H9NO) standard at the beginning of eachrun of 50 samples, as well as at ten-sample in-tervals during the runs: �15N � 0.24‰ (%N byweight �0.80) and �13C � 0.33‰ (%C byweight �3.30).
Testing of Isotope Measurement Validity. Weused two bulk isotopic tests to evaluate thequality of C and N isotopes as biosignaturesin Messel materials: first, a test for significantdiagenetic alteration, which might be expect-ed to produce uniformly or randomly distrib-uted stable isotope signatures, instead of clus-tered data; and second, a test for stable iso-topic compositions of the shale matrix, whichshould be unrelated to the composition of em-bedded fossil material.
Shale was sampled several centimetersaway from each fossil specimen, within ap-proximately the same lamina. This revealedno clear correlation between shale and fossilisotopic variation. Additionally, for severalvertebrate fossils, shale was sampled at incre-ments of 2 to 5 mm from the fossil, up to 5 cmaway to determine the extent of carbon and ni-trogen exchange between the fossil materialand the shale. There was, again, no significantcorrelation between �13C, �15N, wt% C, or wt%N and distance from the fossil. Both tests in-dicate that carbon and nitrogen have not dif-fused outward from the fossil into the sur-rounding sediments sufficiently to affect thestable isotopic signatures. Plant fossils, bee-tles, and vertebrate soft tissues were found tobe characterized by relatively high C (27 to 43
wt%) and N (1 to 4 wt%) content (Table 2).Caddis fly larva cases, which were originallycomposed of silk from the insect and clingingsedimentary particles, have distinctly lower Cand N contents, as do fish hard parts such asscales and bones (typically 3% to 8% C and0.1% to 0.3% N). These patterns are consistentwith good-quality preservation of the originalprecursor materials.
Results
Primary Producers. Stable isotope ratios forshale and each of the primary producer cate-gories (fruits, seeds, terrestrial and aquaticleaves, and wood) cluster together on a �13Cand �15N bi-plot (Fig. 1). Messel shale and leaf�15N values are statistically similar (average��15N 0.28‰, single-factor ANOVA with p 0.05) and elevated with respect to modernatmospheric nitrogen values (�15Natm � 0‰).Water lilies were abundant primary producersin the aquatic food web of the Messel lake andare depleted in 15N compared to terrestrialleaves (average ��15N �1.26‰, ANOVAwith p K 0.05), although average �13C valuesof water lily and terrestrial leaves are statisti-cally indistinguishable (��13C 0.22‰, AN-OVA with p 0.32). The �13C values of shaleare depleted in 13C relative to terrestrial andwater lily leaves (��13C 1.73‰, single-factorANOVA tests with p K 0.005) (Table 2). Thesecompositional differences between fossilleaves, which are particularly sensitive to con-tamination and diagenetic alteration owing totheir large surface area-to-volume ratio, andthe shale matrix seems to confirm that isotopic

596 MAIA K. SCHWEIZER ET AL.
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_596File # 06TQ
FIGURE 1. Fossil primary producer and background (shale) stable isotope measurements. Error bars represent onestandard deviation from mean for the symbol grouping. Individual shale measurements (n 159) have been omit-ted for clarity.
exchange between fossils and substrate wasnot so extensive that it obscures distinct iso-topic ranges.
Consumers. All varieties of fossil insectsfrom Messel share a similar �15N range, fromapproximately �4‰ to �10‰, whereas �13Cvalues differ between taxonomic groups (Fig.2). Messel beetle �13C values range from ap-proximately �24‰ to �29‰, which may re-flect a number of carbon sources. Note that in-vertebrates sampled were amalgamated onthe basis of gross morphology. This limits thetaxonomic resolution of trophic data; for ex-ample, the grouping denoted ‘‘Beetles’’ (Fig.2) may include detritivorous, herbivorous,omnivorous, and carnivorous specimens fromvarious taxa (Table 1). However, because thematerial sampled was often fragmented, aspecific taxonomic identification was rarelypossible. Caddis fly larvae are aquatic primaryconsumers and their larval cases are generallymore depleted in 13C than the exoskeletons ofbeetles and other terrestrial insects (by 2.21‰on average, single-factor ANOVA with p �
0.005), though their �15N values are statistical-ly similar (p 0.05).
The fish population in Lake Messel wasdominated by three species (Table 1). Thau-maturus, the smallest of the three, has the wid-est range of stable isotope compositions (Fig.3; �13C �23‰ to �29‰, �15N �5‰ to10.5‰). Thaumaturus fossils were sampled asfollows: eye material, often the most obviousand greatest quantity of soft tissue material ina specimen; and muscle and soft tissuesscraped from the body of the fish, usuallyfrom between the delicate ribs. The stable iso-tope compositions of eye material overlap sig-nificantly with muscle compositions (�13C and�15N, single-factor ANOVAs with p k 0.05).On the basis of this observation, we placed eyematerial with muscle in the ‘‘soft tissue’’ cat-egory for Atractosteus strausi, which is foundmuch less frequently in the shale, to providesufficient numbers of samples for robust dataanalysis.
Bones and scales from fish and higher con-sumers were analyzed (Table 2), though their

597FOSSIL FOOD WEBS IN LAKE MESSEL
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_597File # 06TQ
FIGURE 2. Summary of invertebrate stable isotope compositions. Caddis flies represent the aquatic food web, asdo most beetles. Cockroaches and giant ants represent the terrestrial ecosystem. Error bars represent one standarddeviation from mean.
isotopic compositions cannot be directly com-pared to fossilized soft tissue values. Soft tis-sues are typically 35 to 45 wt% C and 1.5 to2.5 wt% N, while bones are 5 to 15 wt% C and0.05 to 0.2 wt% N. The length of Thaumaturusspecimens used in this study ranged from 15to 50 mm. �15N (i.e., degree of carnivory) doesnot vary strongly with Thaumaturus length(slope of linear regression: 0.013‰ �15N permm length, r2 0.0079).
Stable isotopic compositions of Cyclurus keh-reri are strongly enriched in 15N (Fig. 3; �13C �27‰ to �28‰, �15N �10‰ to 15‰). Thisspecies is significantly larger than Thaumatu-rus, allowing for greater distinction betweensample materials from a single specimen (e.g.,gut contents). As with Thaumaturus, there ap-pears to be no relationship between Cyclurusfish length and �15N. However, in the case ofCyclurus, the limited size range of fish sam-pled may be insufficient to reveal such a pat-tern. Because soft tissues are often difficult toisolate beneath Cyclurus scales, we were un-
able to sample soft tissue of both a 45-cm adultand a 12-cm juvenile.
Atractosteus strausi stable nitrogen isotopevalues mostly fall between Thaumaturus andCyclurus values (Fig. 3). Soft tissue �13C and�15N range from �25‰ to �28‰ and 7.5‰to 9‰, respectively. Several higher vertebrateswere sampled and analyzed (Table 3). Stableisotopic compositions of the ribs from a 70-cm-long pipe snake skeleton are comparableto values from hard parts of other Messel fos-sil organisms, such as fish (Table 2). The or-ganic-rich crocodile gut material yielded sta-ble isotope ratios and elemental compositionscharacteristic of soft tissue (Table 3).
Discussion
The Aquatic Food Web. The data presentedabove represent the first isotopic investigationof fossilized soft tissue material of Eocene age,to our knowledge. The isotopic compositionsof Messel fossil organisms will be discussedhere in order of increasing trophic position,
598 MAIA K. SCHWEIZER ET AL.
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_598File # 06TQ
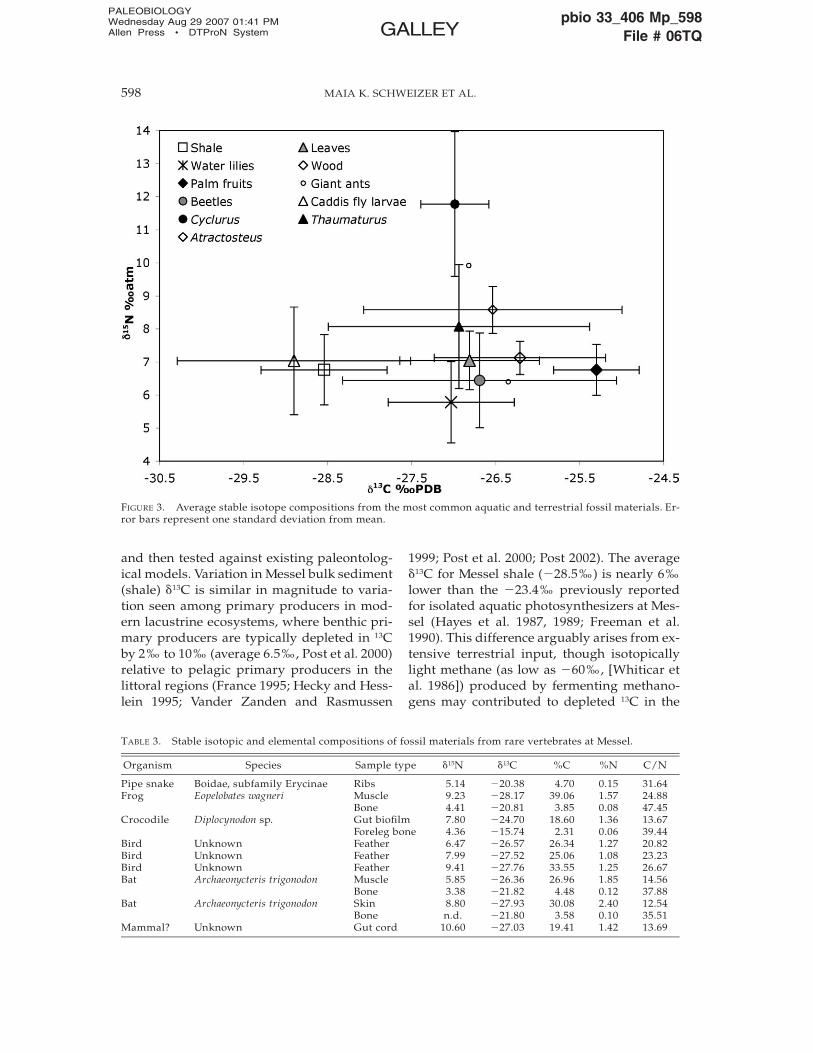
FIGURE 3. Average stable isotope compositions from the most common aquatic and terrestrial fossil materials. Er-ror bars represent one standard deviation from mean.
TABLE 3. Stable isotopic and elemental compositions of fossil materials from rare vertebrates at Messel.
Organism Species Sample type �15N �13C %C %N C/N
Pipe snake Boidae, subfamily Erycinae Ribs 5.14 �20.38 4.70 0.15 31.64Frog Eopelobates wagneri Muscle 9.23 �28.17 39.06 1.57 24.88
Bone 4.41 �20.81 3.85 0.08 47.45Crocodile Diplocynodon sp. Gut biofilm 7.80 �24.70 18.60 1.36 13.67
Foreleg bone 4.36 �15.74 2.31 0.06 39.44Bird Unknown Feather 6.47 �26.57 26.34 1.27 20.82Bird Unknown Feather 7.99 �27.52 25.06 1.08 23.23Bird Unknown Feather 9.41 �27.76 33.55 1.25 26.67Bat Archaeonycteris trigonodon Muscle 5.85 �26.36 26.96 1.85 14.56
Bone 3.38 �21.82 4.48 0.12 37.88Bat Archaeonycteris trigonodon Skin 8.80 �27.93 30.08 2.40 12.54
Bone n.d. �21.80 3.58 0.10 35.51Mammal? Unknown Gut cord 10.60 �27.03 19.41 1.42 13.69
and then tested against existing paleontolog-ical models. Variation in Messel bulk sediment(shale) �13C is similar in magnitude to varia-tion seen among primary producers in mod-ern lacustrine ecosystems, where benthic pri-mary producers are typically depleted in 13Cby 2‰ to 10‰ (average 6.5‰, Post et al. 2000)relative to pelagic primary producers in thelittoral regions (France 1995; Hecky and Hess-lein 1995; Vander Zanden and Rasmussen
1999; Post et al. 2000; Post 2002). The average�13C for Messel shale (�28.5‰) is nearly 6‰lower than the �23.4‰ previously reportedfor isolated aquatic photosynthesizers at Mes-sel (Hayes et al. 1987, 1989; Freeman et al.1990). This difference arguably arises from ex-tensive terrestrial input, though isotopicallylight methane (as low as �60‰, [Whiticar etal. 1986]) produced by fermenting methano-gens may contributed to depleted 13C in the

599FOSSIL FOOD WEBS IN LAKE MESSEL
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_599File # 06TQ
sediment (Chappe et al. 1982; Freeman et al.1990; Goossens et al. 1985; Michaelis and Al-brecht 1979). Two dramatically lighter shalevalues from this study (�13C �36.01‰, and�60.28‰) with typical �15N, %C, and %Nmay correspond to localized high concentra-tions of methanogenic activity.
Messel shale bulk sediment is enriched in15N relative to nitrogen fixed by terrestrialvegetation or aquatic primary producers,which typically has �15N values between�1‰, and �2‰ (Kohl and Shearer 1980). Ac-tive recycling of nitrogen by ammonia-oxidiz-ing bacteria in anaerobic regions and sedi-ment pore waters is here regarded as the likelycause for this enrichment (see Freeman et al.1990). The �15N of the primary producers maytherefore have been lower than the bulk shalemeasurement, perhaps within the range ofwater lily values (Fig. 1), which are isotopi-cally lighter because water lilies fixed nitrogendirectly from the atmosphere. Average shale�15N is also likely to represent a mixture ofbenthic and pelagic contributions.
The carbon isotope fractionations betweendifferent terrestrial plant materials (e.g., be-tween leaves and wood) at Messel are similarto those observed in Oligocene Enspel fossilmaterial (Schweizer et al. 2006). �15N of otherterrestrial plant materials (i.e., wood, seeds,and palm fruits) overlap significantly or en-tirely with leaf values (Fig. 1; single-factorANOVAs with p 0.05).
The isotopic compositions reported here, aswell as observations of fossil caddis fly larvaeattached to water lily stems and leaves (Lutz1991, 1992), indicate that Messel caddis fly lar-vae consumed water lily parts. Specimenswith lower �13C values likely incorporatedmore terrestrial detritus in their diets, as mod-ern caddis flies do (Hutchens et al. 1997). Cad-dis fly �13C values are similar to cockroach(terrestrial detritivore) compositions. Otheraquatic insects are greatly underrepresentedin the shale, either because insect carcasseswere eaten before they could sink or becauseof a genuinely undersized insect population(e.g., Rietschel 1988).
Most Thaumaturus fish �15N values are ap-proximately one trophic step more positivethan the broad cluster of insect values (Table
2), judging from an average enrichment of1.4‰ in modern organisms with invertebratediets (McCutchan et al. 2003). Specimens withoutlying stable isotopic compositions mayhave incorporated more algae, carcasses ofother vertebrates, juvenile fish, or all of theseinto their diets. Unlike many modern pisciv-orous fishes, Thaumaturus does not exhibit in-creasing �15N values with specimen length,which would point to the growing importanceof piscivory to maturing fish diets (Fig. 3). Themany small coprolites containing insects andminute bones described from Messel (Schmitz1991; Micklich 1992; Baszio and Richter 2001)therefore confirm that Thaumaturus was a life-long opportunistic feeder that did not passthrough a insectivore-to-piscivore transition.
Three Atractosteus fish soft tissue samplesshow wide variation in �13C values but reason-ably uniform �15N values. These �15N are tro-phically sensible for a diet of mostly insectsand some Thaumaturus, given an enrichmentof between 1.4‰ and 3.3‰ 15N for modern or-ganisms that consume invertebrates and otheranimals, respectively (McCutchan et al. 2003).Atractosteus specimens may also have con-sumed sufficient primary producers duringalgal blooms (Richter and Baszio 2001) to en-rich their �13C values to just under �25‰ (Fig.3). It should be noted that the Atractosteusspecimens analyzed in this study are at thelower end of the size range for the species, andthus are likely to reflect juvenile diets. Atrac-tosteus could therefore have matured to a com-pletely piscivorous diet with time, as has beensuggested by paleontological reconstructions(Micklich 2002; Richter and Wedmann 2005).
Cyclurus fish specimens (Table 1) have thehighest �15N values of any organism analyzed.Three Cyclurus soft tissue �15N measurementsfell between �10‰ and 12‰, one trophicshift above insects and some Thaumaturus andAtractosteus values. Though medium to largecoprolites at Messel are difficult to assign to asource organism owing to the plethora of can-didates, nearest-living-relative comparisonssuggest that Cyclurus was a formidable pred-ator (Micklich 1992; Richter and Baszio 2001).Stable isotopic data confirm this hypothesis.The highest Cyclurus �15N value of �15‰ isconsistent with a completely carnivorous diet.

600 MAIA K. SCHWEIZER ET AL.
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_600File # 06TQ
FIGURE 4. Flowchart constructed from stable isotopicevidence for trophic structure of the Eocene aquatic eco-system of Lake Messel. Additional trophic levels, suchas primary consumers (e.g., microscopic zooplankton)or additional carnivorous organisms may exist but wereeither not available for study or not recorded in the sta-ble isotopic measurements. Solid arrows represent re-lationships that are recorded in stable isotopic compo-sitions; dashed arrows represent likely relationships ac-cording to paleontological and nearest-living-relativecomparisons that were not tested using stable isotopesin the present study.
Cyclurus �13C values are also much less vari-able than those measured for other organisms.This channeling phenomenon is frequentlyobserved in stable isotopic compositions ofthe highest trophic element of modern ecosys-tems, presumably because these predators re-flect the mean �13C value of the scattered preyin all lower trophic levels (Post 2002).
Black biofilm material sampled from the 40-cm-long crocodile may preserve either the in-ternal organs of the crocodile or partially di-gested flesh from its last meal. If the former isthe case, then the biofilm stable isotopic com-position (Table 3) indicates a nitrogen sourcerelatively depleted in 15N, such as terrestrialinvertebrates. If the biofilm represents par-tially digested flesh in the crocodile’s gut, thenthe stable isotopic composition of this mate-rial reflects the actual composition of the diet.In this case, the crocodile, similar to alligatorsand modern caimans, probably consumed fishand, occasionally, invertebrates (Berg 1966;Keller and Schaal 1992b).
Variations among the stable isotope com-positions of individuals belonging to one spe-cies may result from a number of factors. First,the ecosystem in and surrounding Lake Mes-sel was complex enough to provide a numberof nutrient sources for each organism. Variedeating habits can rapidly and significantly af-fect an organism’s soft tissue stable isotopiccomposition, for example by up to 2‰ �15N inseven days (Adams and Sterner 2000) and byn‰ �13C in n days (e.g., 3‰ in three days; Ol-ive et al. 2003). Additionally, as discussedabove, the diet-to-tissue nitrogen isotope frac-tionation is not constant between organismswith different diets (McCutchan et al. 2003;Pearson et al. 2003; Robbins et al. 2005). Thevariability in trophic step fractionation (3‰ ormore) is itself at least one trophic step. Fur-thermore, different soft tissues from the sameorganism may have slightly different stableisotope compositions. For example, lipid syn-thesis discriminates against 13C, producingdepleted fatty tissues (Pinnegar and Polunin1999), whereas muscle, heart tissue, and livertissue from the same fish typically fall within�1‰ �15N and �2‰ �13C (Bocherens et al.1996). These uncertainties constrain the po-tential accuracy of stable isotopic studies of
ancient ecosystems, particularly given that thebehavior of the organisms cannot be observeddirectly. However, the present data provide atrophic ranking based on �15N and a broaddifferentiation between benthic and pelagichabitats.
Testing the Paleontological Model. A flow-chart showing the path of carbon and nitrogensources through the Messel food web based onstable isotope data (Fig. 4) can be comparedwith the existing paleontological model (Rich-ter and Baszio 2001). Stable isotope data showthat Messel organisms engaged in complexand varied feeding habits beyond those indi-cated by their coprolites. Some trophic levelcomponents as described in the Richter andBaszio’s linear trophic chain might be reclas-sified as partial trophic levels in light of thestable isotope data. For example, Atractosteus,with its combined diet of insects, smaller fish,and even algae (Fig. 4), might belong to thesecond or third trophic level instead of thefourth, depending on the importance of eachof these components to its overall diet. Thestable isotope reconstruction is broadly com-patible with the paleontological model, in that

601FOSSIL FOOD WEBS IN LAKE MESSEL
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_601File # 06TQ
none of the trophic links are reversed. Al-though Richter and Baszio incorporated cer-tain organisms for which there are no stableisotope data (e.g., sponges, crustaceans, andhigher consumers such as piscivorous mam-mals), the isotope data presented here providean independent and nuanced reconstructionof feeding strategies, even accommodatingchanges in diet with time or maturation. Con-versely, in some cases the isotopic data rep-resent averages for an entire clade of organism(e.g., ‘‘other insects’’), whereas paleontologi-cal observations (e.g., of gut contents) may betied to specific species and even individuals.The resulting flowchart (Fig. 4) presents pos-sible trophic connections suggested by the iso-topic data, but departs from convention in thatit does not take into account paleontologicalinformation unless specifically noted. Isotopicreconstructions such as the one presented hereare therefore most effective when used in con-junction with more traditional techniques.
The Terrestrial Food Web. This study repre-sents the first formal reconstruction of the ter-restrial food web at Messel. However, terres-trial vertebrates, including obligate carnivoressuch as Messelogale kessleri, are extremely rareand were not available for sampling. Thus theuppermost trophic level (top consumers), andpresumably several lower levels (other omniv-orous, detritivorous, and carnivorous verte-brates), are not represented by these data. Theplacement of each component of the terrestrialecosystem will be discussed in order of in-creasing trophic level.
Messel leaf �13C values fall within the�36‰ to �22‰ range typical for modern C3
vegetation (Farquhar et al. 1989). Variationamong the fossil stable isotope measurementsis similar in magnitude to variation amongmodern primary producers from a single lo-cation (Smith and Epstein 1971; Leavitt andLong 1986). These observations are consistentwith evidence for comparable levels of atmo-spheric CO2 in the middle Eocene and today(Royer et al. 2001) and also with previous sta-ble isotopic measurements of Eocene plantfossils, for example �13C �27.5‰ fromNorth America (Hayes et al. 1987).
Messel leaf �15N is enriched above the rangeof expected values (0‰ to �6‰) based on the
fixation of atmospheric nitrogen (Delwiche etal. 1979; Kohl and Shearer 1980; Shearer andKohl 1989). However, nitrogen is typically inexcess as a nutrient in tropical forests (Kelleret al. 1986; Vitousek and Sanford 1986; Vogt etal. 1986; Niell et al. 1995), which allows plantsto lose nitrogen through a more ‘‘open’’ nitro-gen cycle (Martinelli et al. 1999). As a result offractionating pathways along the cycle, com-bined with the greater turnover of nitrogen,plants thereby become enriched in 15N. Aver-age foliar �15N under these conditions hasbeen estimated at 4.7 � 2.1‰, with some spe-cies approaching �10‰ (Martinelli et al.1999), similar to Messel values. Water lilyleaves are therefore not similarly enriched, be-cause the plants did not live in terrestrial soils.The average wt% N is similar between mod-ern tropical leaves (1.9 � 0.8 [Martinelli et al.1999]) and Messel fossil leaves (Table 2). Likemodern plant components (Evans et al. 1996),Messel wood and leaves share similar 15Ncompositions (Fig. 1).
Messel beetles with high �15N values typi-cally belong to terrestrial taxa (Table 1) whoseextant relatives consume dung, leaves, detri-tus, pollen, and other insects (Lutz 1990,1992). Beetles with lower �15N and higher �13C(�24‰ to �26‰) probably consumed com-ponents of the aquatic ecosystem such as cad-dis fly larvae and water lily leaves with theirassociated algae.
Messel giant ants have high �15N values con-sistent with their active hunting strategies, ashypothesized by Lutz (1986, 1992). Depend-ing on the species, they may also have con-sumed leaves, fungi, carcasses of other in-sects, or all of the above (Lutz 1992). Insect sta-ble isotopic compositions change to reflectvarying diet on a short time scale (Olive et al.2003), so the highest giant ant �15N value(�9.92‰, Fig. 2) may reflect consumption ofa vertebrate or invertebrate carcass.
The muscle �15N of a 6-cm-long terrestrialfrog (Wuttke 1992a) also implies a high tro-phic level (Table 3). The sole known Eopelobatesspecimen with preserved gut material hadconsumed a small reptile before its death(Wuttke 1992a). This paleontological datum isconsistent with its enrichment in 15N, whichindicates a tertiary consumer status (i.e., pri-

602 MAIA K. SCHWEIZER ET AL.
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_602File # 06TQ
FIGURE 5. Flowchart based on stable isotopic evidence for trophic structure of the terrestrial ecosystem surround-ing Lake Messel. Additional trophic components might include primitive horses and other such rare finds. Solidarrows represent relationships that are reflected in stable isotopic compositions; dashed arrows represent likelyrelationships according to paleontological and nearest-living-relative comparisons that were not tested using stableisotopes.
mary producer → insect → small lizard, am-phibian, or mammal → Eopelobates).
Soft tissue isotopic compositions of two 8-cm (anterioposterior body length) bats span awhole trophic level and are consistent with asecondary or tertiary consumer’s variable diet(Table 3). Messel Archaeonycteris bats arethought to have specialized in hunting insectsin the midlevel trees of the tropical forest sur-rounding Lake Messel (Habersetzer et al.1992), and their isotopic compositions pre-sumably reflect the diets of the beetles andother insects they consumed (Table 2).
Nitrogen isotope ratios measured for threefeathers (Table 3) span at least one trophic lev-el and are consistent with any number of feed-ing strategies used by these birds. With an en-richment of 2.5‰ to 3.6‰ in �13C and 3.7 ‰to 5.6‰ in �15N relative to diet (Peters 1992),two feathers can be traced back to a diet ofbeetles and other insects (Table 3). The highestfeather �15N (�9.41‰) may have belonged to
a bird of prey, which fed on frogs or bats aswell as insects. The avian fauna of Messel in-cluded birds of prey, ibises, fowl, cranes, rails,owls, woodpeckers, and even flamingoes andostriches (Mizutani et al. 1992; Mayr 2005).
An isolated section of intestine, probablyfrom a rare carnivorous terrestrial mammal(Schaal and Ziegler 1992), has the highest ter-restrial �15N value in this study (�10.60‰).Given this value, the unknown organism mayhave hunted or scavenged frogs, birds, in-sects, or bats. Like the aquatic food web, theterrestrial ecosystem is characterized by inter-relationships and mixed feeding strategies(Fig. 5).
Messel Ecosystem Dynamics. The stable iso-topic data record two nuanced, intertwiningecosystems associated with the Eocene LakeMessel: one in the lake itself and one inhabit-ing the surrounding rainforest. The trophicstructure of this 47-Ma environment sharesmany characteristics with modern environ-

603FOSSIL FOOD WEBS IN LAKE MESSEL
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_603File # 06TQ
ments, including trophic isotopic enrich-ments, flexible, adaptive feeding strategies,and fractionation between tissues from a sin-gle organism. Paleontological studies confirmthat many Messel fossil organisms were high-ly specialized and adapted to their paratrop-ical lacustrine environment (e.g., Habersetzerand Storch 1987). Our stable isotopic data sup-port paleontological evidence for symbioticand parasitic relationships, for example, be-tween caddis fly larvae and water lily leaves,and flowering plants and pollinating beetles(Schaarschmidt 1992).
Network structure parameters (see Dunneet al. 2002, 2004) extrapolated from the twoflowcharts depicting generalized trophic re-lationships (Figs. 4, 5) are also comparable tothose calculated for modern ecosystems. Theconnectance values (trophic links per species2)of the terrestrial and aquatic food webs areL/S2 0.22 and 0.23, respectively. These val-ues fall within the upper end of the range re-ported for diverse modern ecosystems (ap-proximately 0.05 to 0.3 [Dunne et al. 2004]).Link densities (trophic links per species) inMessel terrestrial (L/S 2) and aquatic (L/S 1.85) ecosystems are also within the rangeof modern networks, although at the lowerend of the range. These parameters vary wide-ly between modern lacustrine ecosystems(Williams and Martinez 2000, 2004; Dunne etal. 2002), and so it is possible that the valuescalculated for Messel’s ecosystems accuratelyreflect their network structures. However, theabsence of data for some species and entiregroups no doubt skews the connectance up-ward because our trophic ‘‘species’’ in factrepresent generalized groups encompassingsome degree of variation, and some trophicspecies are not represented at all. The degreeof omnivory (i.e., fraction of organisms feed-ing from more than one trophic level) ob-served in Messel food webs (�0.3 for bothfood webs) is also similar to modern values forterrestrial and lacustrine ecosystems (Dunneet al. 2002, 2004).
However, some puzzles remain. For exam-ple, Messel appears to have fewer trophic lev-els than expected for a lake of its size. In mod-ern lacustrine ecosystems, the maximum tro-phic level depends more on lake volume than
nutrient availability or primary productionlevels (Post et al. 2000). According to best-fitdata on highest-trophic-level organisms in 25North American lakes (Post et al. 2000), a lakeof Messel’s volume (�107 m3) should have amaximum trophic level between 3.7 and 3.9,where
maximum trophic level15 15� N � � Nhighest organism primary producers
.3.4‰ (avg. enrichment)
For Messel,
15 15� N � � N 11.78‰ � 5.78‰Cyclurus water-lily
6.00‰,
corresponding to approximately two trophicsteps of �3.4‰ each (Vander Zanden and Ras-mussen 2001). There are several possible ex-planations for this discrepancy. As discussedabove, trophic fractionation between organ-isms is determined by diet, and there may besubstantial deviations from the canonical�3.4‰ enrichment. For example, the majorityof Messel’s organisms incorporated varyingproportions of insects into their diets (Figs. 4,5), where the fractionation between consumerand insect is typically �1.4‰ �15N (Mc-Cutchan et al. 2003). Additionally, extremelyrare aquatic organisms such as turtles, croco-diles, and the less common fish species, whichwere not available for sampling, are allthought to be mostly or purely piscivorous,and may therefore constitute another partialor whole trophic level.
Finally, the elevated �15N values for primaryproducers at Messel may have condensed theapparent fractionation between bottom andtop trophic levels. These high values may bedue to original microorganism compositionsor post-deposition alteration by continued ni-trogen cycling. If the former is correct, then wewould expect a food web with overall elevated�15N but sensible trophic separations, whereasif the latter is correct (and the sedimentary ni-trogen cycling affected primary producer val-ues but not fossil macroorganism values) wewould expect only the bottom trophic levels tobe condensed while the top trophic levelsshow typical fractionations. The data (Fig. 1)suggest that the latter is the case. The stable

604 MAIA K. SCHWEIZER ET AL.
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_604File # 06TQ
isotope data exhibit a sensible trophic rankingbased on �15N, although the lowest levels,composed of shale, terrestrial plants, insects,and even some Thaumaturus, overlap signifi-cantly. Despite these limitations, however, thestable isotopic compositions here reported for47-Ma fossil organisms seem to broadly re-flect trophic position and provide a tantaliz-ing example of what stable isotopic recon-structions of ancient food webs may reveal.
Conclusions
Messel fossils contain sufficiently well pre-served carbon and nitrogen compounds to al-low stable isotopic decoding of ecosystemstructure. A diverse aquatic ecosystem inhab-ited the lake, including primary producers(water lilies and algae), invertebrate consum-ers (insects and their larvae), and vertebrates(fish, turtles, and crocodiles). The stable iso-topic compositions of many of these compo-nents show trophically sensible enrichmentsrelative to food sources, enabling the recon-struction of a flow-chart food web that takesinto account multiple feeding strategies foreach organism. This food web, although insome cases based on average values for a largegroup of organisms (e.g., insects), is broadlycompatible with existing paleontologicalmodels based on coprolites and gut contents.These data highlight the potential of stableisotopic studies to unravel ancient food websand their ecosystems.
Trophic structure relates to aspects of eco-system dynamics, including species diversity,biomass, biogeochemistry, and stability, all ofwhich are critical to a thorough understand-ing of ecosystems in the past. The Messel floraand fauna share genera with other Eocene fos-sil deposits around the world, including Di-plocynodon crocodiles and avian communitiesin Europe and North America (Peters 1991;Keller and Schaal 1992a). The biota is also sim-ilar to extant organisms in subtropical re-gions, including India (Habersetzer andStorch 1987) and South America (Storch andSchaarschmidt 1992). Indeed, modern South-east Asian lakes have been proposed as themost appropriate modern analogues to Messelon the basis of environmental parameters (Ra-benstein et al. 2004). Stable isotopic studies of
the same, or related, species in these other lo-cations would yield insights into their behav-ioral adaptations to distant and different en-vironments to complement the paleontologi-cal studies of evolutionary morphological fea-tures. The apparent fidelity of stable isotopicsignatures at Messel raises the possibility offurther investigations into older fossillager-statten, especially those with soft tissue pres-ervation such as the Ordovician Soom shale ofSouth Africa or any of several Early Cambriansites.
Although carbon and nitrogen stable iso-tope compositions of Messel fossils broadlyreflect those of the living organisms, basingan ecosystem reconstruction solely on isotopicdata is not possible even at this exceptionallypreserved site. Paleontological clues such asgut contents, coprolites, plant-insect associa-tions, functional morphology, and nearest liv-ing relatives are essential for decoding thecomplex and overlapping isotopic values justas observations of organism behavior are sup-plemental to modern isotope studies.
Acknowledgments
The authors thank D. Erwin, C. Labandeira,P. Koch, and an anonymous reviewer for athorough revision; P. Morrill, M. Brasier, G.Keller, and D. Sigman for helpful comments;B. Behr, head of excavation operations at GrubeMessel, for identifying fossils; and F.-J. Harms,M. Felder, and M. Wuttke for helpful discus-sions and permissions at Grube Messel. Field-work and analysis was funded by M.L.F.’s Na-tional Science Foundation Biocomplexitygrant 0221838 and the Carnegie NASA Astro-biology Institute grant (PI Sean Solomon).
Literature CitedAdams, T. S., and R. W. Sterner. 2000. The effect of dietary ni-
trogen content on trophic level 15N enrichment. Limnologyand Oceanography 45:601–607.
Andreasson, F. P., and B. Schmitz. 1996. Winter and summertemperatures of the early middle Eocene of France from Tur-ritella d18O profiles. Geology 24:1067–1070.
———. 2000. Temperature seasonality in the early middle Eo-cene North Atlantic region: evidence from stable isotope pro-files of marine gastropod shells. Geological Society of Amer-ica Bulletin 112:628–640.
Barton, D. G., and M. V. Wilson. 1999. Microstratigraphic studyof meristic variation in an Eocene fish from a 10000-yearvarved interval at Horsefly, British Columbia. Canadian Jour-nal of Earth Sciences 36:2059–2072.

605FOSSIL FOOD WEBS IN LAKE MESSEL
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_605File # 06TQ
Baszio, S., and G. Richter. 2001. First proof of planctivory/in-sectivory in a fossil fish: Thaumaturus intermedius from the Eo-cene Lake Messel (FRG). Palaeogeography, Palaeoclimatolo-gy, Palaeoecology 173:75–85.
Berg, D. E. 1966. Die Krokodile, insbesondere Asiatosuchus undaff. Sebecus? aus den Eozan von Messel bei Darmstadt/Hes-sen. Abhandlungen des Hessischen Landesamt Bodenfor-schung 52:1–105.
Blair, N., A. Leu, E. Nunoz, J. Olsen, E. Kwong, and D. Des-Marais. 1985. Carbon isotopic fractionation in heterotrophicmicrobial metabolism. Applied and Environmental Microbi-ology 50:996–1001.
Blondel, C. 2001. The Eocene-Oligocene ungulates from WesternEurope and their environment. Palaeogeography, Palaeocli-matology, Palaeoecology 168:125–139.
Bocherens, H., M. L. Fogel, N. Tuross, and M. Zeder. 1995. Tro-phic structure and climate information from isotopic signa-tures in Pleistocene cave fauna of southern England. Journalof Archaeological Science 22:327–340.
Bocherens, H., G. Pacaud, P. A. Lazarev, and A. Mariotti. 1996.Stable isotope abundances (13C, 15N) in collagen and soft tis-sues from Pleistocene mammals from the Yakutia: implica-tions for the paleobiology of the Mammoth Steppe. Palaeo-geography, Palaeoclimatology, Palaeoecology 126:31–44.
Boschker, H. T. S., J. F. C. de Brouwer, and T. E. Cappenberg.1999. The contribution of macrophyte-derived organic matterto microbial biomass in salt-marsh sediments: stable carbonisotope analysis of microbial biomarkers. Limnology andOceanography 44:309–319.
Buatois, L. A., and M. G. Mangano. 1993. Ecospace utilization,paleoenvironmental trends, and the evolution of early non-marine biotas. Geology 21:595–598.
Buchheim, H. P. 1994. Eocene Fossil Lake, Green River Forma-tion, Wyoming: a history of fluctuating salinity. Pp. 239–247in R. W. Renaut and W. M. Last, eds. Sedimentology and geo-chemistry of modern and ancient saline lakes. Society for Sed-imentary Geology, Tulsa, Okla.
Carson, G. A. 1991. Silicification of fossils. Pp. 455–499 in P. A.Allison and D. E. G. Briggs, eds. Taphonomy: releasing datalocked in the fossil record. Plenum, New York.
Chappe, B., P. Albrecht, and W. Michaelis. 1982. Polar lipids ofArchaebacteria in sediments and petroleums. Science 217:65–66.
Clementz, M. T., and P. L. Koch. 2001. Differentiating aquaticmammal habitats and foraging ecology with stable isotopesin tooth enamel. Oecologia 129:461–472.
Coffin, R. B., B. Fry, B. J. Peterson, and R. T. Wright. 1989. Carbonisotope composition of estuarine bacteria. Limnology andOceanography 34:1305–1310.
Coffin, R. B., D. J. Velinsky, R. Devereux, W. A. Price, and L. A.Cifuentes. 1990. Stable carbon isotope analysis of nucleic ac-ids to trace sources of dissolved substrates used by estuarinebacteria. Applied and Environmental Microbiology 56:2012–2020.
Cohen, A. S. 2003. Paleolimnology: the history and evolution oflake systems. Oxford University Press, Oxford.
Cohen, J. E., F. Briand, and C. M. Newman. 1990. Communityfood webs: data and theory. Springer, New York.
Collinson, M. E. 1986. Fruchte und Samen aus dem MesselerOlscheifer. Courier Forschungsinstitut Senckenberg 85:217–220.
Davis, P. G., and D. E. G. Briggs. 1995. Fossilization of feathers.Geology 23:783–786.
Delwiche, C. C., P. J. Zinke, C. M. Johnson, and R. A. Virginia.1979. Nitrogen isotope distribution as a presumptive indica-tor of nitrogen fixation. Botanical Gazette 140:65–69.
DeNiro, M. J. 1985. Postmortem preservation and alteration of
in vivo bone collagen isotope ratios in relation to palaeodie-tary reconstruction. Nature 317:806–809.
DeNiro, M. J., and S. Epstein. 1978. Influence of diet on the dis-tribution of carbon isotopes in animals. Geochimica et Cos-mochimica Acta 42:495–506.
———. 1981. Influence of diet on the distribution of nitrogenisotopes in animals. Geochimica et Cosmochimica Acta 45:341–351.
Dunne, J. A., R. J. Williams, and N. D. Martinez. 2002. Networkstructure and biodiversity loss in food webs: robustness in-creases with connectance. Ecology Letters 5:558–567.
———. 2004. Network structure and robustness of marine foodwebs. Marine Ecology Progress Series 273:291–302.
Estep, M. F., F. R. Tabita, P. L. Parker, and C. Van Baalen. 1978.Carbon isotope fractionation by ribulose-1.5-bisphosphatecarboxylase from various organisms. Plant Physiology 61:680–687.
Evans, R. D., A. J. Bloom, S. S. Sukrapanna, and J. R. Ehleringer.1996. Nitrogen isotope composition of tomato (Lycopersicon es-culentum Mill. cv T-5) grown under ammonium or nitrate nu-trition. Plant Cell Environment 19:1317–1323.
Farquhar, G. D., J. R. Ehleringer, and K. T. Hubick. 1989. Carbonisotope discrimination and photosynthesis. Annual Review ofPlant Physiology and Plant Molecular Biology 40:503–537.
Ferris, F. G., W. S. Fyfe, and T. J. Beveridge. 1988. Metallic ionbinding by Bacillus substilis: implications for the fossilizationof microorganisms. Geology 16:149–152.
France, R. L. 1995. Differentiation between littoral and pelagicfood webs in lakes using carbon isotopes. Limnology andOceanography 40:1310–1313.
Franzen, J. L. 1990. Grube Messel. Pp. 289–294 in D. E. G. Briggsand P. R. Crowther, eds. Paleobiology: a synthesis. BlackwellScientific, Oxford.
Franzen, J. L., and W. Michaelis. 1988. Eocene Lake Messel. Cou-rier Forschungsinstitut Senckenberg 107:1–452.
Franzen, J. L., J. Weber, and M. Wuttke. 1982. Senckenberg-Gra-bungen in der Grube Messel bei Darmstadt. 3. Eergenmosse1979–1981. Courier Forschungsinstitut Senckenberg 54:1–118.
Freeman, K. H., J. M. Hayes, J. Trendel, and P. Albrecht. 1990.Evidence from carbon isotope measurements for diverse or-igins of sedimentary hydrocarbons. Nature 343:254–256.
Fry, B., and E. B. Sherr. 1984. �13C measurements as indicatorsof carbon flow in marine and freshwater ecosystems. Contri-butions to Marine Science 27:13–47.
Gearing, J. N. 1991. The study of diet and trophic relationshipsthrough natural abundance 13C. Pp. 201–218 in D. Colemanand B. Fry, eds. Carbon isotope techniques. Academic Press,San Diego.
Gierlowski-Kordesch, E. H., and L. E. Park. 2004. Comparingspecies diversity in the modern and fossil record of lakes.Journal of Geology 112:703–717.
Goossens, H., W. I. C. Rijpstra, R. R. Duren, J. W. de Leeuw, andP. A. Schenck. 1985. Bacterial contribution to sedimentary or-ganic matter; a comparative study of lipid moieties in bacteriaand Recent sediments. Organic Geochemistry 10:683–696.
Goth, K. 1990. Der Messeler Olschiefer—ein Algenlaminit. Cou-rier Forschungsinstitut Senckenberg 131:1–143.
Gray, J. 1988. Evolution of the freshwater ecosystem: the fossilrecord. Palaeogeography, Palaeoclimatology, Palaeoecology62:1–214.
Habersetzer, J., and G. Storch. 1987. Klassifikation und funkti-onelle Flugelmorphologie palaogener Fledermause (Mam-malia, Chiroptera). Courier Forschungsinstitut Senckenberg91:117–150.
Habersetzer, J., G. Richter, and G. Storch. 1992. Bats: alreadyhighly specialized insect predators. Pp. 181–191 in Schaal andZiegler 1992.

606 MAIA K. SCHWEIZER ET AL.
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_606File # 06TQ
Haq, B. U., J. Hardenbol, and P. R. Vail. 1987. Chronology of fluc-tuating sea levels since the Triassic. Science 235:1156–1167.
Harding, I. C., and L. S. Chant. 2000. Self-sedimented diatommats as agents of exceptional fossil preservation in the Oli-gocene Florissant Lake Beds, Colorado, United States. Geol-ogy 28:195–198.
Harms, F.-J. 2002. Steine erzahlen Geschichte(n): Ursachen furdie Entstehung des Messel-Sees gefunden. Natur und Muse-um 132:1–4.
Harrigan, P., J. C. Zieman, and S. A. Macko. 1989. The base ofnutritional support for the gray snapper (Lutjanus griseus): anevaluation based on a combined stomach content and stableisotope analysis. Bulletin of Marine Science 44:65–77.
Hartenberger, J. L. 1987. Modalites des extinctions et appari-tions chez les mammiferes du Paleogene d’Europe. Memoiresde la Societe Geologique de France 150:133–143.
Hayes, J. M., R. Takigiku, R. Ocampo, H. J. Callot, and P. Al-brecht. 1987. Isotopic compositions and probably origins oforganic molecules in the Eocene Messel Shale. Nature 329:48–51.
Hayes, J. M., K. H. Freeman, B. N. Popp, and C. H. Hoham. 1989.Compound-specific isotopic analyses: a novel tool for recon-struction of ancient biogeochemical processes. Organic Geo-chemistry 16:1115–1128.
Hecky, R. E., and R. H. Hesslein. 1995. Contributions of benthicalgae to lake food webs as revealed by stable isotope analysis.Journal of the North American Benthological Society 14:631–653.
Hullar, M. A. J., B. Fry, B. J. Peterson, and R. T. Wright. 1996.Microbial utilization of estuarine dissolved organic carbon: astable isotope tracer approach tested by mass balance. Ap-plied and Environmental Microbiology 62:2489–2493.
Hummel, S., and B. Herrmann. 1994. General aspects of samplepreparation. Pp. 59–68 in S. Hummel and B. Herrmann, eds.Ancient DNA: recovery and analysis of genetic material frompaleontological, archaeological, museum, medical, and foren-sic specimens. Springer, New York.
Hutchens, J. J., E. F. Benfield, and J. R. Webster. 1997. Diet andgrowth of a leaf-shredding caddisfly in southern Appalachianstreams of contrasting disturbance history. Hydrobiologia364:193–201.
Jacobs, B. F. 2004. Palaeobotanical studies from tropical Africa:relevance to the evolution of forest, woodland and savannahbiomes. Philosophical Transactions of the Royal Society ofLondon B 359:1573–1583.
Jacobs, B. F., and P. S. Herendeen. 2004. Eocene dry climate andwoodland vegetation in tropical Africa reconstructed fromfossil leaves from northern Tanzania. Palaeogeography, Pa-laeoclimatology, Palaeoecology 213:115–123.
Janis, C. M. 1993. Tertiary mammal evolution in the context ofchanging climates, vegetation, and tectonic events. AnnualReview of Ecology and Systematics 24:467–500.
Keller, M., and S. Schaal. 1992a. Lizards: reptiles en route to suc-cess. Pp. 121–133 in Schaal and Ziegler 1992.
Keller, M., W. A. Kaplan, and S. C. Wofsy. 1986. Emissions ofN20, CH4, and CO2 from tropical soils. Journal of GeophysicalResearch 91:1791–1802.
———. 1992b. Crocodiles: large ancient reptiles. Pp. 109–118 inSchaal and Ziegler 1992.
Kimble, B. J., J. R. Maxwell, G. Eglinton, P. Albrecht, A. Ensmin-ger, P. Arpino, and G. Ourisson. 1974. Tri- and tetraterpenoidhydrocarbons in the Messel oil shale. Geochimica et Cosmo-chimica Acta 28:1165–1181.
Kling, G. W., B. Fry, and W. J. O’Brien. 1992. Stable isotopes andplanktonic trophic structure in arctic lakes. Ecology 73:561–566.
Kohl, D. H., and G. Shearer. 1980. Isotopic fractionation associ-
ated with symbiotic N2 fixation and uptake of NO3� by plants.
Plant Physiology 66:51–56.Kvacek, Z., M. Bohme, Z. Dvorak, M. Konzalova, K. Mach, J.
Prokop, and M. Rajchl. 2004. Early Miocene freshwater andswamp ecosystems of the Most Basin (northern Bohemia)with particular reference to the Bilina Mine section. Journalof the Czech Geological Society 49:1–40.
Kohler, J. 1997. Die Fossillagerstatte Enspel: vegetation, vege-tationsdynamik, und klima im Oberoligozan. Ph.D. thesis.Universitat Tubingen, Tubingen.
Leavitt, S. W., and A. Long. 1986. Stable-carbon isotope vari-ability in tree foliage and wood. Ecology 67:1002–1010.
Legendre, S., and J. L. Hartenberger. 1992. Evolution of mam-malian faunas in Europe during the Eocene and Oligocene.Pp. 516–528 in D. R. Prothero and W. A. Berggren, eds. Eo-cene-Oligocene climatic and biotic evolution. Princeton Uni-versity Press, Princeton, N.J.
Lenz, O. K., V. Wilde, W. Riegel, and F. Schaarschmidt. 2005. Cli-mate and vegetation dynamics in the pit in Messel—a reap-praisal of paleontological data from 1980. Courier Forschung-sinstitut Senckenberg 255:81–101.
Lutz, H. 1986. Eine neue Unterfamilie der Formicidae (Insecta:Hymenoptera) aus dem mittel-eozanen Olschiefer der ‘GrubeMessel’ bei Darmstadt (Deutschland, S.-Hessen). Senckenber-giana Lethaea 67:177–218.
———. 1990. Systematische und Palokologische Untersuchun-gen an Insekten aus dem Mittel-eozan der Grube Messel beiDarmstadt. Courier Forschungsinstitut Senckenberg 124:1–165.
———. 1991. Autochthone aquatische Arthropoda aus dem Mit-tel-Eozan der Funstatte Messel (Insecta: Heteroptera; Cole-optera; cf. Diptera-Nematocera; Crustacea: Cladocera). Cou-rier Forschungsinstitut Senckenberg 139:119–125.
———. 1992. Giant ants and other rarities: the insect fauna. Pp.55–67 in Schaal and Ziegler 1992.
MacFadden, B. J., Y. Wang, T. E. Cerling, and F. Anaya. 1994.South American fossil mammals and carbon isotopes: a 25million-year sequence from the Bolivian Andes. Palaeogeog-raphy, Palaeoclimatology, Palaeoecology 103:257–268.
MacFadden, B. J., N. Solounias, and T. E. Cerling. 1999. Ancientdiets, ecology, and extinction of 5-million-year-old horsesfrom Florida. Science 283:824–827.
Macko, S. A., and M. F. Estep. 1984. Microbial alteration of stablenitrogen and carbon isotopic compositions of organic matter.Organic Geochemistry 6:787–790.
Macko, S. A., M. F. Estep, and T. C. Hoering. 1982. Nitrogen iso-tope fractionation by blue-green algae cultured in molecularnitrogen and nitrate. Carnegie Institution of WashingtonYearbook 81:413–417.
Mai, D. H. 1981. Entwicklung und klimatische Differenzierungder Laubwaldflora Mitteleuropas im Tertiar. Flora 171:525–582.
Manchester, S. R., M. E. Collinson, and K. Goth. 1994. Fruits ofthe Juglandaceae from the Eocene of Messel, Germany, andimplications for early Tertiary phytogeographic exchange be-tween Europe and western North America. International Jour-nal of Plant Science 155:388–394.
Markwick, P. J. 1996. Late Cretaceous to Pleistocene climates:nature of the transition from a ‘hot-house’ to an ‘ice-house’world. Ph.D. dissertation. University of Chicago, Chicago.
———. 2002. Integrating the present and past records of climate,biodiversity and biogeography: implications for palaeoecol-ogy and palaeoclimatology. In J. A. Crame and A. W. Owen,eds. Palaeobiogeography and biodiversity change: a compar-ison of the Ordovician and Mesozoic-Cenozoic radiations.Geological Society of London Special Publication 194:179–199.
Martinelli, L. A., M. C. Piccolo, A. R. Townsend, P. M. Vitousek,

607FOSSIL FOOD WEBS IN LAKE MESSEL
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_607File # 06TQ
E. Cuevas, W. McDowell, G. P. Robertson, O. C. Santos, and K.Tresedar. 1999. Nitrogen stable isotopic composition of leavesand soil: tropical versus temperate forests. Biogeochemistry46:45–65.
Masters, P. M. 1987. Preferential preservation of noncollagenousprotein during bone diagenesis: implications for chronomet-ric and stable isotopic measurements. Geochimica et Cos-mochimica Acta 51:3209–3214.
Matthes, G. 1966. Zur geologie des Olschiefervorkommens vonMessel bei Darmstadt. Abhandlungen des Hessischen Lan-desamtes fur Bodenforschung 51:1–87.
———. 1968. Les couches eocenes dans la region du fosse rhen-an septentrional. Memoires de la Bureau de Recherche Geo-logiques et Minieres 58:327–337.
Mayr, G. 2005. The Paleogene fossil record of birds in Europe.Biological Review 80:515–542.
McCutchan, J. H., W. M. Lewis, C. Kendall, and C. C. McGrath.2003. Variation in trophic shift for stable isotope ratios of car-bon, nitrogen and sulfur. Oikos 102:378–390.
McKenna, M. C. 1980. Eocene paleolatitude, climate, and mam-mals of Ellesmere Island. Palaeogeography, Palaeoclimatolo-gy, Palaeoecology 30:349–362.
———. 1983. Holarctic landmass rearrangement, cosmic events,and Cenozoic terrestrial organisms. Annals of the MissouriBotanical Garden 70:459–489.
Mehner, T., J. Benndorf, P. Kasprzak, and R. Koschel. 2002. Bio-manipulations of lake ecosystems: successful applicationsand expanding complexity in the underlying science. Fresh-water Biology 47:2453–2465.
Melchor, R. N., J. F. Genise, and S. E. Miquel. 2002. Ichnology,sedimentology, and paleontology of Eocene calcareous paleo-sols from a palustrine sequence, Argentina. Palaios 17:16–35.
Mertz, D. F., and P. R. Renne. 2005. A numerical age for the Mes-sel fossil deposit (UNESCO World Heritage Site) derivedfrom 40Ar/39Ar dating on a basaltic rock fragment. CourierForschungsinstitut Senckenberg 255:67–75.
Mertz, D. F., F.-J. Harms, G. Gabriel, and M. Felder. 2004. Ar-beitstreffen in der Forschungsstation Grube Messel mit neuenErgebnissen aus der Messel-Forschung. Natur und Museum134:289–290.
Meyer, H. W., K. Rielly, S. Maguire, N. R. O’Brien, and A. M.Ross. 2001. Fossilization of Tertiary insects and plants bypolysaccharide film. Geological Society of America Abstractswith Programs 33:63.
Meyers, P. A. 1997. Organic geochemical proxies of paleocean-ographic, paleolimnologic, and paleoclimatic processes. Or-ganic Geochemistry 27:213–250.
Michaelis, W., and P. Albrecht. 1979. Molecular fossils of Ar-chaebacteria in kerogen. Naturwissenschaften 66:420–422.
Micklich, N. 1992. Ancient knights-in-armour and modern can-nibals. Pp. 69–91 in Schaal and Ziegler 1992.
———. 2002. The fish fauna of Messel Pit: a nursery school? Cou-rier Forschungsinstitut Senckenberg 237:97–127.
Minagawa, M., and E. Wada. 1984. Stepwise enrichment of 15Nalong food chains: further evidence and the relation between�15N and animal age. Geochimica et Cosmochimica Acta 48:1135–1140.
Mizutani, H., M. Fukuda, and Y. Kabaya. 1992. 13C and 15N en-richment factors of feathers of 11 species of adult birds. Ecol-ogy 73:1391–1395.
Muller-Sohnius, D., P. Horn, and H. G. Hukkenholz. 1989. Ka-lium-Argon Datierungen an tertiaren Vulkaniten der Hoch-eifel (BRD). Chemie Erde 49:119–136.
Murray, A. M. 2001. The oldest fossil cichlids (Teleostei: Perci-formes): indication of a 45 million-year-old species flock. Pro-ceedings of the Royal Society of London B 268:679–684.
Mycke, B., and W. Michaelis. 1986. Molecular fossils from chem-
ical degradation of macromolecular organic matter. OrganicGeochemistry 10:847–858.
Mycke, B., F. Narjes, and W. Michaelis. 1987. Bacteriohopanete-trol from chemical degradation of an oil shale kerogen. Nature326:179–181.
Niell, C., M. C. Piccolo, P. A. Steudler, J. M. Melillo, B. J. Feigl,and C. C. Cerri. 1995. Nitrogen dynamics in soils of forestsand active pastures in the western Brazilian Amazon Basin.Soil Biology and Biochemistry 27:1167–1175.
Nix, T., and S. Feist-Burkhardt. 2003. New methods applied tothe microstructure analysis of Messel oil shale: confocal laserscanning microscopy (CLSM) and environmental scanningelectron microscopy (ESEM). Geology Magazine 140:469–478.
Olive, L. W., J. K. Pinnegar, N. V. C. Polunin, G. Richards, andR. Welch. 2003. Isotope trophic-step fractionation: a dynamicequilibrium model. Journal of Animal Ecology 72:608–617.
Ostrom, P. H., S. A. Macko, M. H. Engel, and D. A. Russell. 1993.Assessment of trophic structure of Cretaceous communitiesbased on stable nitrogen isotope analyses. Geology 21:491–494.
Ostrom, P. H., J.-P. Zonneveld, and L. L. Robbins. 1994. Organicgeochemistry of hard parts: assessment of isotopic variabilityand indigeneity. Palaeogeography, Palaeoclimatology, Pa-laeoecology 107:201–212.
Panacost, R. D., and J. S. Sinninghe Damste. 2003. Carbon iso-topic compositions of prokaryotic lipids as tracers of carboncycling in diverse settings. Chemical Geology 195:29–58.
Pearson, S. F., D. J. Levey, C. H. Greenberg, and C. M. del Rio.2003. Effects of elemental composition on the incorporation ofdietary nitrogen and carbon isotopic signatures in an omniv-orous songbird. Oecologia 135:516–523.
Peters, D. S. 1991. Zoogeographical relationships of the Eoceneavifauna from Messel (Germany). Acta XX Congressus Inter-nationalis Ornithologici 1:572–577.
———. 1992. Messel birds: a land-based assemblage. Pp. 137–151 in Schaal and Ziegler 1992.
Peters, R. B., and L. C. Sloan. 2000. High concentrations ofgreenhouse gases and polar stratospheric clouds: a possiblesolution to high-latitude faunal migration at the latest Paleo-cene thermal maximum. Geology 28:979–982.
Pimm, S. L., J. H. Lawton, and J. E. Cohen. 1991. Food web pat-terns and their consequences. Nature 350:669–674.
Pinnegar, J. K., and N. V. C. Polunin. 1999. Differential fraction-ation of �13C and �15N among fish tissues: implications for thestudy of trophic interactions. Functional Ecology 12:225–231.
Pirrung, M., C. Fischer, G. Buchel, R. Gaupp, H. Lutz, and F. O.Neuffer. 2003. Lithofacies succession of maar crater depositsin the Eifel area (Germany). Terra Nova 15:125–132.
Post, D. M. 2002. Using stable isotopes to estimate trophic po-sition: models, methods, and assumptions. Ecology 83:703–718.
Post, D. M., M. L. Pace, and N. G. Hairston. 2000. Ecosystem sizedetermines food-chain length in lakes. Nature 405:1047–1049.
Rabenstein, R., R. Usman, and S. Schaal. 2004. Looking at mod-ern lakes as models for Eocene habitats from Messel. CourierForschungsinstitut Senckenberg 252:115–138.
Richter, G. 1987. Untersuchungen zur Ernahrung eozaner Sau-ger aus der Fossilfundstatte Messel bei Darmstadt. CourierForschungsinstitut Senckenberg 91:1–33.
———. 1992. Fossilized gut contents: analysis and interpreta-tion. Pp. 285–289 in Schaal and Ziegler 1992.
Richter, G., and S. Baszio. 2001. Traces of a limnic food web inthe Eocene Lake Messel—a preliminary report based on fishcoprolite analyses. Palaeogeography, Palaeoclimatology, Pa-laeoecology 166:345–368.
Richter, G., and G. Storch. 1980. Beitrage zur Ernahrungsbiolo-gie eozaner Fledermause aus der ‘Grube Messel.’ Natur undMuseum 110:353–367.

608 MAIA K. SCHWEIZER ET AL.
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_608File # 06TQ
Richter, G., and S. Wedmann. 2005. Ecology of the Eocene LakeMessel revealed by analysis of small fish coprolites and sed-iments from a drilling core. Palaeogeography, Palaeoclima-tology, Palaeoecology 223:147–161.
Rietschel, S. 1988. Taphonomic biasing in the Messel fauna andflora. Courier Forschungsinstitut Senckenberg 107:169–182.
Robbins, C. T., L. A. Felicetti, and M. Sponheimer. 2005. The ef-fect of dietary protein quality on nitrogen isotope discrimi-nation in mammals and birds. Oecologia 144:534–540.
Robinson, N., G. Eglinton, P. A. Cranwell, and Y. B. Zeng. 1989.Messel oil shale (western Germany): assessment of deposi-tional palaeoenvironment from the content of biologicalmarker compounds. Chemical Geology 76:152–173.
Rolf, C., R. Pucher, R. Schulz, and T. Wonik. 2005. Magnetic in-vestigations at the kernel of the Messel Pit 2001—first mag-netic knowledge. Courier Forschungsinstitut Senckenberg255:47–55.
Roth, J. L., and D. L. Dilcher. 1978. Some considerations in leafsize and leaf margin analysis of fossil leaves. Courier For-schungsinstitut Senckenberg 30:165–171.
Royer, D. L., S. L. Wing, D. J. Beerling, D. W. Jelley, P. L. Koch,L. J. Hickey, and R. A. Berner. 2001. Paleobotanical evidencefor near present-day levels of atmospheric CO2 during part ofthe Tertiary. Science 292:2310–2313.
Sasche, M. 2005. A remarkable fossiliferous mass flow depositin the Eocene Eckfeld Maar (Germany)—sedimentological, ta-phonomical, and palaeoecological considerations. Facies 51:173–184.
Schaal, S., and W. Ziegler, eds. 1992. Messel—an insight into thehistory of life and of the Earth. Clarendon, Oxford.
Schaarschmidt, F. 1992. The vegetation: fossil plants as witness-es of a warm climate. Pp. 29–52 in Schaal and Ziegler 1992.
Schmitz, B., and F. P. Andreasson. 2001. Air humidity and laked18O during the latest Paleocene-earliest Eocene in Francefrom recent and fossil fresh-water and marine gastropodd18O, d13C, and 87Sr/86Sr. Geological Society of America Bul-letin 113:774–789.
Schmitz, M. 1991. Die Koprolithen mitteleozaner Vertebratenaus der Grube Messel bei Darmstadt. Courier Forschungsin-stitut Senckenberg 137:1–159.
Schweizer, M. K., M. W. Wooller, J. K. W. Toporski, M. L. Fogel,and A. Steele. 2006. Examination of an Oligocene lacustrineecosystem using C and N isotopes. Palaeogeography, Palaeo-climatology, Palaeoecology 230:335–351.
Shearer, G., and D. H. Kohl. 1989. Estimates of N2 fixation in eco-systems: the need for and basis of the 15N natural abundancemethod. Pp. 342–374 in R. W. Rundel, J. R. Ehleringer, and K.A. Nagy, eds. Stable isotopes in ecological research. Springer,New York.
Sillen, A., and J. A. Lee-Thorp. 1994. Trace element and isotopeaspects of predator-prey relationships in terrestrial foodwebs.Palaeogeography, Palaeoclimatology, Palaeoecology 107:243–255.
Sittler, C. 1968. L’analyse pollinique dans l’Est de la France:etude des formations eocenes ou rapportees a l’Eocene et desstratotypes polynologiques de Borken et de Messel. Memoiresde la Bureau de Recherche Geologiques et Minieres 58:165–171.
Smith, B. N., and S. Epstein. 1971. Two categories of 13C/12C ra-tios for higher plants. Plant Physiology 47:380–384.
Sponheimer, M., and J. A. Lee-Thorp. 1999. Reconstructing thediet of the early hominid Australopithecus africans using 13C/12C analysis. Science 238:368–370.
Storch, G., and J. Habersetzer. 1988. Archaeonycteris pollex (Mam-malia, Chiroptera), eine neue Fledermaus aus dem Eozan derGrube Messel bei Darmstadt. Courier ForschungsinstitutSenckenberg 107:263–273.
Storch, G., and F. Schaarschmidt. 1992. The Messel fauna and
flora: a biogeographical puzzle. Pp. 291–297 in Schaal andZiegler 1992.
Thiele-Pfeiffer, H. 1988. Die Mikroflora aus dem mitteleozanOlscheifer von Messel bei Darmstadt. Palaeontographica, Ab-teilung B 211:1–86.
Toporski, J. K. W., A. Steele, F. Westall, R. Avci, D. M. Martill,and D. S. McKay. 2002. Morphological and spectral investi-gation of exceptionally well-preserved bacterial biofilms fromthe Oligocene Enspel formation, Germany. Geochimica etCosmochimica Acta 66:1773–1791.
Torricelli, S., G. Knezaurek, and U. Biffi. 2006. Sequence bio-stratigraphy and paleoenvironmental reconstruction in theEarly Eocene Figols Group of the Tremp-Graus Basin (south-central Pyrenees, Spain). Palaeogeography, Palaeoclimatolo-gy, Palaeoecology 232:1–35.
Vander Zanden, M. J., and J. B. Rasmussen. 1999. Primary con-sumer �15N and �13C and the trophic position of aquatic con-sumers. Ecology 80:406–416.
———. 2001. Variation in �15N and �13C trophic fractionation: im-plications for aquatic food web studies. Limnology andOceanography 46:2061–2066.
Vander Zanden, M. J., B. J. Shuter, N. Lester, and J. B. Rasmus-sen. 1999. Patterns of food chain length in lakes: a stable iso-tope study. American Naturalist 154:406–416.
Vitousek, P. M., and R. L. Sanford. 1986. Nutrient cycling inmoist tropical forest. Annual Review of Ecology and System-atics 17:137–167.
Vogt, K. A., C. C. Grier, and D. J. Vogt. 1986. Production, turn-over, nutrient dynamics of above- and below-ground detritusof world forests. Advances in Ecological Research 15:303–377.
von Koenigswald, W., G. Richter, and G. Storch. 1981. Nachweizvon Hornschuppen bei Eomanis waldi aus der ‘Grube Messel’bei Darmstadt (Mammalia, Pholidota). Senckenbergiana Le-thaea 61:291–298.
Wang, Y., T. E. Cerling, and B. J. MacFadden. 1994. Fossil horsesand carbon isotopes: new evidence for Cenozoic dietary, hab-itat, and ecosystem changes in North America. Palaeogeog-raphy, Palaeoclimatology, Palaeoecology 107:269–279.
Whiticar, M. J., E. Faber, and M. Schoell. 1986. Biogenic methaneformation in marine and freshwater environments: CO2 re-duction vs. acetate fermentation—isotope evidence. Geochim-ica et Cosmochimica Acta 50:693–709.
Wilde, V. 1989. Untersuchungen zur Systematik der Blattresteaus dem Mitteleozan der Grube Messel bei Darmstadt (Hes-sen, Bundesrepublik Deutschland). Courier Forschungsinsti-tut Senckenberg 115:1–213.
Wilf, P., N. R. Cuneo, K. R. Johnson, J. F. Hicks, S. L. Wing, andJ. D. Obradovich. 2003. High plant diversity in Eocene SouthAmerica: evidence from Patagonia. Science 300:122–125.
Williams, C. J., A. H. Johnson, B. A. LePage, D. R. Vann, and T.Sweda. 2003. Reconstruction of Tertiary Metasequoia forests.II. Structure, biomass and productivity in Eocene floodplainforests in the Canadian Arctic. Paleobiology 29:271–292.
Williams, R. J., and N. D. Martinez. 2000. Simple rules yieldcomplex food webs. Nature 404:180–183.
———. 2004. Limits to trophic levels and omnivory in complexfood webs: theory and data. American Naturalist 163:458–468.
Wilson, M. V. 1980. Eocene lake environments: depth and dis-tance-from-shore variation in fish, insect and plant assem-blages. Palaeogeography, Palaeoclimatology, Palaeoecology32:21–44.
———. 1987. Predation as a source of fish fossils in Eocene lakesediments. Palaios 2:497–504.
———. 1988. Reconstruction of ancient lake environments usingboth autochthonous and allochthonous fossils. Palaeogeog-raphy, Palaeoclimatology, Palaeoecology 62:609–623.
Wonik, T. 2005. First results of the measurement in the For-

609FOSSIL FOOD WEBS IN LAKE MESSEL
PALEOBIOLOGYWednesday Aug 29 2007 01:41 PMAllen Press • DTProN System GALLEY
pbio 33_406 Mp_609File # 06TQ
schungsbohrung Messel 2001. Courier ForschungsinstitutSenckenberg 255:11–20.
Wuttke, M. 1983. Weichteilerhaltung durch lithifizierte Mikoor-ganismen bei mittel-eozanen Vertebraten aus dem olschieferder ‘‘Grube Messel’’ bei Darmstadt. Senckenbergiana Lethaea64:503–527.
———. 1992a. Amphibia at Lake Messel: salamanders, toads,and frogs. Pp. 95–98 in Schaal and Ziegler 1992.
———. 1992b. Conservation, dissolution, transformation: on thebehavior of biogenic materials during fossilization. Pp. 263–275 in Schaal and Ziegler 1992.
———. 1992c. Death and burial of the vertebrates. Pp. 263–275in Schaal and Ziegler 1992.
Zachos, J. C., K. C. Lohmann, J. C. G. Walker, and W. W. Sher-wood. 1993. Abrupt climate change and transient climatesduring the Paleogene: a marine perspective. Journal Of Ge-ology 101:191–213.
Zimmerle, W. 1993. Some aspects of Cenozoic maar sedimentsof Europe: the source-rock potential and their exceptionallygood fossil preservation. Pp. 467–476 in J. F. W. Negendankand B. Zolitschka, eds. Paleolimnology of European maarlakes. Springer, Berlin.










![Lignin Dening Ye Oct 29 th 2010 PBIO 691: [Plant Cell Walls]](https://static.fdocuments.us/doc/165x107/56649c7c5503460f9492ffe1/lignin-dening-ye-oct-29-th-2010-pbio-691-plant-cell-walls.jpg)







