
Patent ductus arteriosus in mice with smooth muscle-specific Jag1 ...
9
4191 RESEARCH ARTICLE INTRODUCTION The evolutionarily conserved Notch signaling pathway plays a major role in vascular development in mammals and other vertebrates (Gridley, 2010; Hofmann and Iruela-Arispe, 2007; Phng and Gerhardt, 2009; Roca and Adams, 2007). Among the developmental decisions regulated by the Notch pathway during embryonic vascular development are arterial-venous specification, endothelial tip cell differentiation, and vascular smooth muscle cell differentiation. Whereas most Notch signaling pathway components are expressed in endothelial cells, Jag1 (which encodes a ligand for Notch pathway receptors) is unusual in that it is expressed in both endothelial cells and vascular smooth muscle cells (Villa et al., 2001). Mice homozygous for a Jag1 null mutation die from hemorrhage early during embryogenesis, exhibiting defects in remodeling of the embryonic and yolk sac vasculature (Xue et al., 1999). Endothelial cell-specific deletion of Jag1 with a Cre deleter that is highly expressed during embryogenesis also results in early embryonic lethality owing to vascular defects resulting from the defective development of vascular smooth muscle (High et al., 2008). Similarly, endothelial cell-specific deletion of Jag1 using an inducible Cre deleter line results in reduced coverage of retinal arteries by vascular smooth muscle cells (Benedito et al., 2009). However, it has not been established whether Jag1 plays an essential, cell-autonomous role in the vascular smooth muscle cell lineage. We report here the phenotype of embryos and mice with smooth muscle-specific Jag1 deletion. These mice die in the early postnatal period from patent ductus arteriosus (PDA), a defect of the outflow tract of the heart. The ductus arteriosus is an arterial blood vessel that connects the pulmonary artery and the descending aorta during fetal life. After birth, the ductus arteriosus is normally rapidly and permanently occluded, separating the pulmonary and systemic circulations to establish the normal adult circulatory pattern. Failure of the ductus arteriosus to close after birth is termed PDA and is one of the most common human congenital heart defects. PDA patients are at increased risk of pulmonary and cardiac problems such as pulmonary hemorrhage, congestive heart failure, chronic lung disease, sepsis and necrotizing enterocolitis (Clyman, 2006; Forsey et al., 2009; Schneider and Moore, 2006). Mice with smooth muscle-specific deletion of Jag1 exhibit defects in contractile smooth muscle cell differentiation in the vascular wall of the ductus arteriosus and adjacent descending aorta. These defects appear to arise through an inability to propagate the JAG1-Notch signal by lateral induction throughout the vascular wall of these vessels. Our novel model for this common congenital heart defect provides new insights into the genetic programs that underlie ductus arteriosus development and closure. MATERIALS AND METHODS Mice We described previously the Jag1 null allele (Xue et al., 1999) and Jag1 conditional allele (Jag1 flox ) (Kiernan et al., 2006). SM22a-Cre (official name Tagln-Cre) (Holtwick et al., 2002) and Tek-Cre (also known as Tie2- Cre) (Koni et al., 2001) mice were obtained from the Jackson Laboratory. Animal protocols were approved by the Jackson Laboratory Animal Care and Use Committee. Intracardiac injections To better visualize the outflow tract and ductus arteriosus, hearts of SM22a- Cre/+; Jag1 flox/– mutant and control littermate mice were injected with Microfil silicone rubber injection compound (MV-122; Flow Tech). Pups were isolated at E18.5 by caesarean section, and were euthanized 7 hours post-surgery. The chest cavities were opened and fixed in 10% neutral buffered formalin. After fixing for 1 hour, neonates were rinsed and Microfil compound was injected into the left ventricle using a 27 gauge needle. Development 137, 4191-4199 (2010) doi:10.1242/dev.052043 © 2010. Published by The Company of Biologists Ltd The Jackson Laboratory, Bar Harbor, ME 04609, USA. *These authors contributed equally to this work † Present address: Center for Molecular Medicine, Maine Medical Center Research Institute, Scarborough, ME 04074, USA ‡ Author for correspondence ([email protected]) Accepted 18 October 2010 SUMMARY The ductus arteriosus is an arterial vessel that shunts blood flow away from the lungs during fetal life, but normally occludes after birth to establish the adult circulation pattern. Failure of the ductus arteriosus to close after birth is termed patent ductus arteriosus and is one of the most common congenital heart defects. Mice with smooth muscle cell-specific deletion of Jag1, which encodes a Notch ligand, die postnatally from patent ductus arteriosus. These mice exhibit defects in contractile smooth muscle cell differentiation in the vascular wall of the ductus arteriosus and adjacent descending aorta. These defects arise through an inability to propagate the JAG1-Notch signal via lateral induction throughout the width of the vascular wall. Both heterotypic endothelial smooth muscle cell interactions and homotypic vascular smooth muscle cell interactions are required for normal patterning and differentiation of the ductus arteriosus and adjacent descending aorta. This new model for a common congenital heart defect provides novel insights into the genetic programs that underlie ductus arteriosus development and closure. KEY WORDS: Congenital heart defects, Notch signaling pathway, Vascular smooth muscle differentiation, Mouse Patent ductus arteriosus in mice with smooth muscle-specific Jag1 deletion Xuesong Feng*, Luke T. Krebs* and Thomas Gridley †,‡ DEVELOPMENT
Transcript of Patent ductus arteriosus in mice with smooth muscle-specific Jag1 ...

4191RESEARCH ARTICLE
INTRODUCTIONThe evolutionarily conserved Notch signaling pathway plays a majorrole in vascular development in mammals and other vertebrates(Gridley, 2010; Hofmann and Iruela-Arispe, 2007; Phng andGerhardt, 2009; Roca and Adams, 2007). Among the developmentaldecisions regulated by the Notch pathway during embryonic vasculardevelopment are arterial-venous specification, endothelial tip celldifferentiation, and vascular smooth muscle cell differentiation.Whereas most Notch signaling pathway components are expressedin endothelial cells, Jag1 (which encodes a ligand for Notch pathwayreceptors) is unusual in that it is expressed in both endothelial cellsand vascular smooth muscle cells (Villa et al., 2001). Micehomozygous for a Jag1 null mutation die from hemorrhage earlyduring embryogenesis, exhibiting defects in remodeling of theembryonic and yolk sac vasculature (Xue et al., 1999). Endothelialcell-specific deletion of Jag1 with a Cre deleter that is highlyexpressed during embryogenesis also results in early embryoniclethality owing to vascular defects resulting from the defectivedevelopment of vascular smooth muscle (High et al., 2008).Similarly, endothelial cell-specific deletion of Jag1 using aninducible Cre deleter line results in reduced coverage of retinalarteries by vascular smooth muscle cells (Benedito et al., 2009).However, it has not been established whether Jag1 plays an essential,cell-autonomous role in the vascular smooth muscle cell lineage.
We report here the phenotype of embryos and mice withsmooth muscle-specific Jag1 deletion. These mice die in theearly postnatal period from patent ductus arteriosus (PDA), adefect of the outflow tract of the heart. The ductus arteriosus is
an arterial blood vessel that connects the pulmonary artery andthe descending aorta during fetal life. After birth, the ductusarteriosus is normally rapidly and permanently occluded,separating the pulmonary and systemic circulations to establishthe normal adult circulatory pattern. Failure of the ductusarteriosus to close after birth is termed PDA and is one of themost common human congenital heart defects. PDA patients areat increased risk of pulmonary and cardiac problems such aspulmonary hemorrhage, congestive heart failure, chronic lungdisease, sepsis and necrotizing enterocolitis (Clyman, 2006;Forsey et al., 2009; Schneider and Moore, 2006). Mice withsmooth muscle-specific deletion of Jag1 exhibit defects incontractile smooth muscle cell differentiation in the vascularwall of the ductus arteriosus and adjacent descending aorta.These defects appear to arise through an inability to propagatethe JAG1-Notch signal by lateral induction throughout thevascular wall of these vessels. Our novel model for this commoncongenital heart defect provides new insights into the geneticprograms that underlie ductus arteriosus development andclosure.
MATERIALS AND METHODSMiceWe described previously the Jag1 null allele (Xue et al., 1999) and Jag1conditional allele (Jag1flox) (Kiernan et al., 2006). SM22a-Cre (officialname Tagln-Cre) (Holtwick et al., 2002) and Tek-Cre (also known as Tie2-Cre) (Koni et al., 2001) mice were obtained from the Jackson Laboratory.Animal protocols were approved by the Jackson Laboratory Animal Careand Use Committee.
Intracardiac injectionsTo better visualize the outflow tract and ductus arteriosus, hearts of SM22a-Cre/+; Jag1flox/– mutant and control littermate mice were injected withMicrofil silicone rubber injection compound (MV-122; Flow Tech). Pupswere isolated at E18.5 by caesarean section, and were euthanized 7 hourspost-surgery. The chest cavities were opened and fixed in 10% neutralbuffered formalin. After fixing for 1 hour, neonates were rinsed andMicrofil compound was injected into the left ventricle using a 27 gaugeneedle.
Development 137, 4191-4199 (2010) doi:10.1242/dev.052043© 2010. Published by The Company of Biologists Ltd
The Jackson Laboratory, Bar Harbor, ME 04609, USA.
*These authors contributed equally to this work†Present address: Center for Molecular Medicine, Maine Medical Center ResearchInstitute, Scarborough, ME 04074, USA‡Author for correspondence ([email protected])
Accepted 18 October 2010
SUMMARYThe ductus arteriosus is an arterial vessel that shunts blood flow away from the lungs during fetal life, but normally occludesafter birth to establish the adult circulation pattern. Failure of the ductus arteriosus to close after birth is termed patent ductusarteriosus and is one of the most common congenital heart defects. Mice with smooth muscle cell-specific deletion of Jag1, whichencodes a Notch ligand, die postnatally from patent ductus arteriosus. These mice exhibit defects in contractile smooth musclecell differentiation in the vascular wall of the ductus arteriosus and adjacent descending aorta. These defects arise through aninability to propagate the JAG1-Notch signal via lateral induction throughout the width of the vascular wall. Both heterotypicendothelial smooth muscle cell interactions and homotypic vascular smooth muscle cell interactions are required for normalpatterning and differentiation of the ductus arteriosus and adjacent descending aorta. This new model for a common congenitalheart defect provides novel insights into the genetic programs that underlie ductus arteriosus development and closure.
KEY WORDS: Congenital heart defects, Notch signaling pathway, Vascular smooth muscle differentiation, Mouse
Patent ductus arteriosus in mice with smooth muscle-specificJag1 deletionXuesong Feng*, Luke T. Krebs* and Thomas Gridley†,‡
DEVELO
PMENT

4192
Indomethacin treatmentTo determine whether postnatal indomethacin administration could rescueclosure of the ductus arteriosus of SM22a-Cre/+; Jag1flox/– neonatal mice,newly born pups were injected subcutaneously with indomethacin (6 mg/kgbody weight) within 12 hours of birth. Pups were euthanized 6 hours afterinjection, the chest cavities were opened and closure of the ductusarteriosus was scored visually. In some treated mice, outflow tracts werevisualized by Microfil injection.
Histology and immunofluorescenceEmbryos were fixed in Dent’s (20% DMSO, 80% methanol) and/or 4%paraformaldehyde. Chest cavities were embedded in paraffin, sectioned andstained with Hematoxylin and Eosin. For immunohistochemistry, the sectionswere de-waxed in a standard xylene and ethanol series, then rehydrated withphosphate-buffered saline (PBS). An antigen-retrieval step was performed inboiling 10 mM sodium citrate (pH 6.0) for 10 minutes for antibodies exceptanti-PECAM1 (0.01% trypsin for 15 minutes at 37°C) and anti-phospho-histone H3 (10 mg/ml proteinase K for 5 minutes). Slides were then blockedin 5% goat serum and 2% BSA in PBST (PBS + 0.1% Tween-20) for 2 hoursat room temperature before being incubated with diluted primary antibodiesat 4°C overnight. Tyramide-amplified immunofluorescent staining for theNOTCH1 Val1744 epitope was performed as described (Del Monte et al.,2007). Primary antibodies used in this study include: anti-PECAM1 (BDBiosciences Pharmingen; 1:100; only works for Dent’s-fixed embryos);polyclonal anti-smooth muscle 22a protein (Abcam; 1:400); polyclonal anti-smooth muscle alpha actin (Abcam; 1:200); polyclonal anti-smooth musclemyosin heavy chain (Biomedical Technologies; 1:200); anti-phospho-histoneH3 (Upstate Biotechnology; 1:100); polyclonal anti-JAG1 (Santa Cruz, H-114; 1:50); polyclonal anti-cleaved NOTCH1 (Val1744; Cell Signaling); andpolyclonal anti-NOTCH3 (Abcam ab23426; 1:250). Alexa Fluor fluorescentsecondary antibodies (Molecular Probes), which included goat anti-rabbitAlexa Fluor 488 and 555, goat anti-rat Alexa Fluor 488 and 555, wereapplied for 2 hours at room temperature or at 4°C overnight. All slides weremounted using Vectashield mounting medium with DAPI (VectorLaboratories).
MicroscopyFor transmission electron microscopy, chest cavities of E18.5 and P0 micewere fixed in 2% glutaraldehyde with 0.1 M sodium cacodylate (pH 7.2)overnight at 4°C. The ductus arteriosus was carefully dissected out andembedded in resin at 70°C for 48 hours. Sections (90 nm) were collected ona 300-mesh copper grid and stained with 2% uranyl acetate and Reynold’slead, then viewed on a JEOL 1230 JM transmission electron microscope. Forlight microscopy, histological and immunofluorescent images were takenwith a Zeiss Axioplan microscope or a Leica SP5 confocal microscope.
Quantitative (q) RT-PCRThe ductus arteriosus and ascending aorta from P0 neonates and E18.5embryos were microdissected and immersed in RNAlater (Ambion).Genotypes were identified by allele-specific PCR. We combined two tofive ductus arteriosi or ascending aortas of the same genotype to increasethe RNA extraction yield. RNA was prepared using the Qiagen MinimRNA Extraction Kit. RNA (100 ng of each sample) was reversetranscribed with random hexamers (Ambion). qRT-PCR was performedwith the Super SYBR Master Mix Kit (Applied Biosystems). For eachgene tested we performed three experimental repeats and three biologicalrepeats. Expression levels were normalized to Gapdh expression. The studywas performed on the ABI 7500 platform using SDS software. The primersused were designed by Primer Bank (Spandidos et al., 2008) (forward andreverse; 5� to 3�): smooth muscle 22a, CAACAAGGGTCCATCCTACGGand ATCTGGGCGGCCTACATCA; smoothelin, ATGGCAGACG -AGGCTTTAGC and AGTGTAGCCAGTTCTCCTTGTT; smooth muscleactin a2, GTCCCAGACATCAGGGAGTAA and TCGGATACT -TCAGCGTCAGGA; smooth muscle actin 2, CCGCCCTA -GACATCAGGGT and TCTTCTGGTGCTACTCGAAGC; smooth musclemyosin heavy chain, AAGCTGCGGCTAGAGGTCA and CCCTCCCT -TTGATGGCTGAG; calponin, AGAACCGGCTCCTGTCCAA andGTGTGCATAGGATAACCCCATC; Gapdh, AGGTCGGTGTGAA -CGGATTTG and TGTAGACCATGTAGTTGAGGTCA.
RESULTSMouse embryos with endothelial cell-specificdeletion of the Jag1 gene phenocopy Jag1 nullmutantsTo assess whether Jag1 function is required in both endothelial andvascular smooth muscle cells, we performed tissue-specific Jag1gene deletions. Endothelial-specific Jag1 deletion has beenreported previously (High et al., 2008) using an independentlygenerated Jag1 conditional allele. Endothelial cell-specific deletionof our Jag1 conditional allele (Kiernan et al., 2006) yielded resultsidentical to those published previously (High et al., 2008).Endothelial cell-specific Jag1 mutant embryos phenocopiedconstitutive Jag1 null mutant embryos (Xue et al., 1999).Endothelial cell-specific Jag1 mutant embryos died atapproximately embryonic day (E) 10.5, exhibiting yolk sacvascular remodeling defects (see Fig. S1 in the supplementarymaterial), hemorrhages (see Fig. S1 in the supplementary material)and non-cell-autonomous defects in vascular smooth muscle celldifferentiation (see Fig. S2 in the supplementary material). Theseresults demonstrate that the early lethality of Jag1 null embryos isdue to Jag1 function in endothelial cells, and that Jag1 expressionin endothelial cells is required for the differentiation of adjacentmural cells into vascular smooth muscle.
Mice with smooth muscle-specific deletion of Jag1die postnatally due to patent ductus arteriosusTo assess whether Jag1 function is also required in vascular smoothmuscle cells, we performed smooth muscle-specific deletion usingSM22a-Cre (also known as Tagln-Cre) mice (Holtwick et al.,2002). SM22a-Cre/+; Jag1flox/– (hereafter referred to as Jag1-SMcko, for Jag1 smooth muscle conditional knockout) mice weregenerated by crossing SM22a-Cre/+; Jag1+/– males withJag1flox/flox female mice. Unlike embryos with endothelial cell-specific Jag1 deletion, Jag1-SMcko mice were found alive and inthe expected Mendelian ratios at postnatal day (P) 0, the day ofbirth. However, ~50% of the Jag1-SMcko neonates died by P1, andno Jag1-SMcko mice survived past P2 (Table 1). Jag1-SMcko micein the early postnatal period could be identified by their cyanoticappearance, which suggested the possibility of cardiovasculardefects. We therefore assessed the morphology of the heart andcardiac outflow tract in Jag1-SMcko and control littermate micefrom E18.5 through P2. This analysis revealed that 95% (53/56) ofJag1-SMcko mice born exhibited PDA (Fig. 1B,C,G; Table 2; seeFig. S3 in the supplementary material). In addition to PDA, asubset of Jag1-SMcko mice also exhibited another outflow tractdefect termed retroesophageal right subclavian artery (Fig. 1C;Table 2).
Jag1-SMcko mice exhibit defects in vascularsmooth muscle differentiation in the ductusarteriosus and the adjacent descending aortaDuctus arteriosus closure postnatally requires differentiatedvascular smooth muscle cells to contract in response to a drop incirculating prostaglandin levels and a rise in oxygen tension thatoccur after birth. We assessed vascular smooth muscle celldifferentiation in the walls of the outflow tract vessels byimmunofluorescent staining for several smooth muscle contractileproteins. Using an antibody against smooth muscle 22a protein(SM22a) at P0 we found that, in contrast to the control littermateductus arteriosus, which has multiple layers of differentiatedsmooth muscle cells occupying almost the entire width of theductus arteriosus wall (Fig. 1E,F), the Jag1-SMcko ductus
RESEARCH ARTICLE Development 137 (24)
DEVELO
PMENT

arteriosus has a much thinner region of SM22a-expressing cellsadjacent to the endothelial cell layer (Fig. 1H,I). Reduced numbersof SM22a-expressing cells were also observed in the medial wallof the connecting descending aorta (Fig. 1H,I). Analyzingadditional contractile smooth muscle cell markers, such as smoothmuscle a-actin (SMA; also known as ACTA2) and smooth musclemyosin heavy chain (SMMHC; also known as MYH11), defects invascular smooth muscle cell differentiation in the ductus arteriosusand descending aorta of Jag1-SMcko embryos could be observedat E17.5 (Fig. 2A-C versus 2A�-C�), when the ductus arteriosus inboth the Jag1-SMcko and littermate control embryos is still patent.Surprisingly, we found that vascular smooth muscle celldifferentiation defects in Jag1-SMcko mice were restricted to theregion of the ductus arteriosus and the adjacent descending aorta.Both the ascending limb of the aorta (Fig. 2D�-F�) and thepulmonary artery trunk (Fig. 2G�-I�) of the mutant embryosexhibited normal differentiation of contractile vascular smoothmuscle cells throughout the medial wall.
Additional assays support our finding of defects in vascularsmooth muscle cell differentiation in the outer layers of the medialwall of the ductus arteriosus and descending aorta of Jag1-SMckomice. We assessed expression levels of genes involved incontractile smooth muscle cell differentiation by quantitative real-time RT-PCR of RNA isolated at P0 from the ductus arteriosus(which displays the differentiation defect) and ascending aorta(which does not display the defect) of Jag1-SMcko and controllittermate mice. The expression of all these contractile smoothmuscle cell marker genes was downregulated in the ductusarteriosus of Jag1-SMcko compared with wild-type littermates (Fig.3). However, in RNA isolated from the ascending aorta, only theSMMHC gene (Myh11) exhibited substantial downregulation inJag1-SMcko mutants. Ultrastructural analysis further revealed thatwhereas the control ductus arteriosus medial wall is composed ofcompact, well-organized smooth muscle cell layers throughout thewidth of the vessel wall (Fig. 4A,C), the Jag1-SMcko ductusarteriosus is composed of more loosely organized cells surroundedby thicker layers of extracellular matrix (Fig. 4B,D). Takentogether, these data demonstrate that PDA in Jag1-SMcko mice iscaused by defects in vascular smooth muscle cell differentiation inthe ductus arteriosus and descending aorta.
A possible explanation for the regional restriction of the vascularsmooth muscle cell differentiation defect in Jag1-SMcko mice isthat SM22a-Cre-mediated recombination is more efficient in theductus arteriosus and its adjacent descending aorta than it is in thepulmonary trunk and ascending aorta. Cre-mediated deletion of ourJag1flox allele generates a nonfunctional, but still immunoreactive,JAG1 protein that exhibits altered cellular trafficking (Kiernan etal., 2006) (Fig. 5). Detection of this nonfunctional JAG1 protein byimmunofluorescence enabled us to assess Cre-mediated Jag1deletion in the outflow tract of Jag1-SMcko embryos (Fig. 5).Whereas the wild-type JAG1 protein localized primarily around the
cell surface (Fig. 5A), the nonfunctional JAG1 protein generatedafter deletion of the Jag1flox allele appeared to localize to thecytoplasm (Fig. 5B). Using this assay, we compared Jag1 genedeletion in Jag1-SMcko and control littermate embryos at E13.5.This analysis revealed equivalent Jag1 gene deletion in theembryonic ascending aorta (Fig. 5H), which does not exhibit thedifferentiation defect, and embryonic ductus arteriosus anddescending aorta (Fig. 5F). These data demonstrate that regional
4193RESEARCH ARTICLEJag1 in vascular development
Fig. 1. Jag1-SMcko mice exhibit patent ductus arteriosus.(A-C)Outflow tracts of P0 mice. In the control littermate (CT) outflowtract (A), the ductus arteriosus (arrowhead) has closed. The ductusarteriosus in Jag1-SMcko mice (B,C, arrowhead) is not closed and bloodcan be observed flowing through the still-patent ductus. The Jag1-SMcko mouse shown in B represents the most common Jag1-SMckomutant phenotype. These mice have a normally patterned outflowtract, but exhibit patent ductus arteriosus (PDA). The mouse shown in Crepresents a subset of Jag1-SMcko mice that displays both PDA andretroesophageal right subclavian artery (arrow). (D,G)Histologicalanalysis confirmed PDA in P0 Jag1-SMcko (G) versus control (D) mice.(E,F,H,I) Fluorescent immunostaining using markers for smooth musclecells (orange, anti-SM22a antibody) and endothelial cells (green, anti-PECAM1 antibody) revealed reduced vascular smooth muscle celldifferentiation in the ductus arteriosus and adjacent descending aortaof Jag1-SMcko (H,I) versus control (E,F) mice. F and I are highermagnification views of the descending aorta vessel wall shown in E andH, respectively. All sections are counterstained with DAPI to highlightcell nuclei (blue). aAo, ascending aorta; dAo, descending aorta; DA,ductus arteriosus; PT, pulmonary artery trunk; RCA, right commoncarotid artery; LCA, left common carotid artery; RSA, right subclavianartery. Scale bars: 1 mm in A-C; 200m in D,E,G,H.
Table 1. Survival of Jag1-SMcko embryos and miceAge Jag1-SMcko All other genotypes
E18.5 3 (3) 19P0 24 (24) 76P1 29 (15) 80P2 0 29Weaning 0 52
Shown is the number of littermates of Jag1-SMcko (with the number surviving inparentheses) versus all other genotypes.
Table 2. Phenotypes of Jag1-SMcko embryos and micePhenotype Jag1-SMcko All other genotypes
PDA 53/56 (95%) 7/204 (3.4%)RRSA 14/56 (25%) 1/204 (0.5%)
Shown is the occurrence of patent ductus arteriosus (PDA) and retroesophagealright subclavian artery (RRSA) among total littermates of Jag1-SMcko versus all othergenotypes. D
EVELO
PMENT

4194
restriction of the vascular smooth muscle cell differentiation defectin Jag1-SMcko embryos is not caused by regional differences inJag1 gene deletion.
JAG1 regulates its own expression via lateralinductionPrevious studies have shown that Jag1 is highly expressed in bothendothelial cells and smooth muscle cells of large arteries at earlyembryonic stages in mice (High et al., 2008; Loomes et al., 1999).However, Jag1 expression in the outflow tract at later embryonicstages has not been closely examined. We assessed JAG1 proteinexpression by immunofluorescence from E13.5 through E16.5. AtE13.5, JAG1 was expressed at high levels across the full width of thevessel wall of the ductus arteriosus and descending aorta (Fig. 6A,J).By contrast, in the ascending aorta and pulmonary artery trunk, onlythe layer of smooth muscle cells adjacent to the endothelial layerdisplayed a similarly high level of JAG1 expression (Fig. 6A,J versus6D,G). The expression pattern seen at E13.5 was maintained atstages as late as E15.5 (see Fig. S4 in the supplementary material),although the high levels of JAG1 expression observed in the ductusarteriosus and descending aorta at E13.5 gradually declined. ByE16.5, JAG1 expression had decreased to a low level in all parts ofthe outflow tract.
We utilized the altered cellular localization of the mutant JAG1protein to assess the consequences of inactivation of JAG1 functionin Jag1-SMcko embryos. Expression of mutant JAG1 protein inJag1-SMcko ductus arteriosus and descending aorta was restricted tovascular smooth muscle cells in the region of the medial wall
RESEARCH ARTICLE Development 137 (24)
Fig. 3. Transcription of contractile smooth muscle cell markergenes is reduced in Jag1-SMcko mice. Relative gene expressionlevels were examined in ductus arteriosus (top) and ascending aorta(bottom) in P0 wild-type (WT) and Jag1-SMcko (CKO) mice. Genesincluded encode smooth muscle 22a (SM22), smooth muscle actin a2(SMA2a), smooth muscle actin 2 (SMA2g), smooth muscle myosinheavy chain (SMMHC), smoothelin (SML) and calponin 2 (CALP).P<0.05 for control versus mutant DA comparisons (Student’s t-test);aAo comparisons (except the SMMHC comparison) were notstatistically significant. Error bars indicate standard deviation of themean.
Fig. 2. Jag1-SMcko mouse embryos exhibit regionally restrictedvascular smooth muscle cell differentiation defects.(A-I�) Immunofluorescent staining of outflow tract vessels in E17.5 controllittermate (A-I) and Jag1-SMcko (A�-I�) embryos stained with antibodies(red) against smooth muscle 22a (SM22), smooth muscle myosin heavychain (SMMHC) and smooth muscle actin (SMA). All sections werecounterstained with DAPI (blue). In the control (A-C), expression of thesecontractile smooth muscle cell proteins was observed throughout thewidth of the medial wall of the ductus arteriosus (DA) and descendingaorta (dAo). However, in the Jag1-SMcko ductus arteriosus anddescending aorta (A�-C�), expression of all three contractile smooth musclecell proteins was observed only in the region adjacent to the endothelialcell layer lining the lumens of the ductus arteriosus and descending aorta.By contrast, in both the ascending aorta (aAo) (D-F versus D�-F�) andpulmonary artery (PA) (G-I versus G�-I�), no differences in expression ofthese contractile smooth muscle cell proteins were observed between thecontrol littermates and Jag1-SMcko embryos. Asterisks (A-C�) mark theentry point of the ductus arteriosus (left side) with the descending aorta(right side). Scale bars: 100m for A-F�; 50m for G-I�.
DEVELO
PMENT
adjacent to the endothelium (Fig. 6B,K). This suggests thatexpression of functional JAG1 protein at the surface of differentiatedvascular smooth muscle cells adjacent to the endothelium is requiredfor JAG1 expression in adjacent smooth muscle cells located moredistally in the ductus arteriosus and descending aorta medial wall.
To confirm that the mutant JAG1 protein is capable of beingexpressed throughout the medial wall of the ductus arteriosus anddescending aorta, we analyzed JAG1 expression in SM22a-Cre/+;Jag1flox/+ embryos. These embryos contain the SM22a-Cre andJag1flox alleles, but also have the wild-type Jag1 allele. SM22a-Cre/+; Jag1flox/+ embryos and mice therefore express both thewild-type JAG1 protein and the mutant JAG1 protein that exhibitsaltered cellular localization. Since JAG1-mediated Notch signalingis active in SM22a-Cre/+; Jag1flox/+ embryos, which exhibit nomutant phenotype, we should observe the superimposition of thewild-type JAG1 and mutant JAG1 expression patterns. Analysis ofE13.5 SM22a-Cre/+; Jag1flox/+ embryos revealed that both thewild-type and mutant JAG1 proteins were expressed throughout theentire medial wall in SM22a-Cre/+; Jag1flox/+ embryos (Fig.6F,I,L,O). Taken together, these results indicate that expression offunctional JAG1 protein at the cell surface of smooth muscle cellsadjacent to the endothelial cell layer is required for expression ofJAG1 protein in smooth muscle cell layers located more distally,which is a hallmark of Notch signaling via lateral induction(Eddison et al., 2000; Lewis, 1998; Saravanamuthu et al., 2009).
Cleaved NOTCH1 protein is reduced in Jag1-SMckoembryos at E13.5We assessed expression of cleaved NOTCH1 by utilizing the anti-Val1744 antibody, which recognizes an epitope generated bygamma secretase cleavage of the NOTCH1 protein (Del Monte etal., 2007; Schroeter et al., 1998). At E13.5, at the level of theconnection between the ductus arteriosus and descending branch ofthe dorsal aorta, cleaved NOTCH1 protein was observedthroughout the developing medial wall in the control littermate(Fig. 7A), whereas in the Jag1-SMcko mutant cleaved NOTCH1was observed in only a few cells adjacent to the vascular
4195RESEARCH ARTICLEJag1 in vascular development
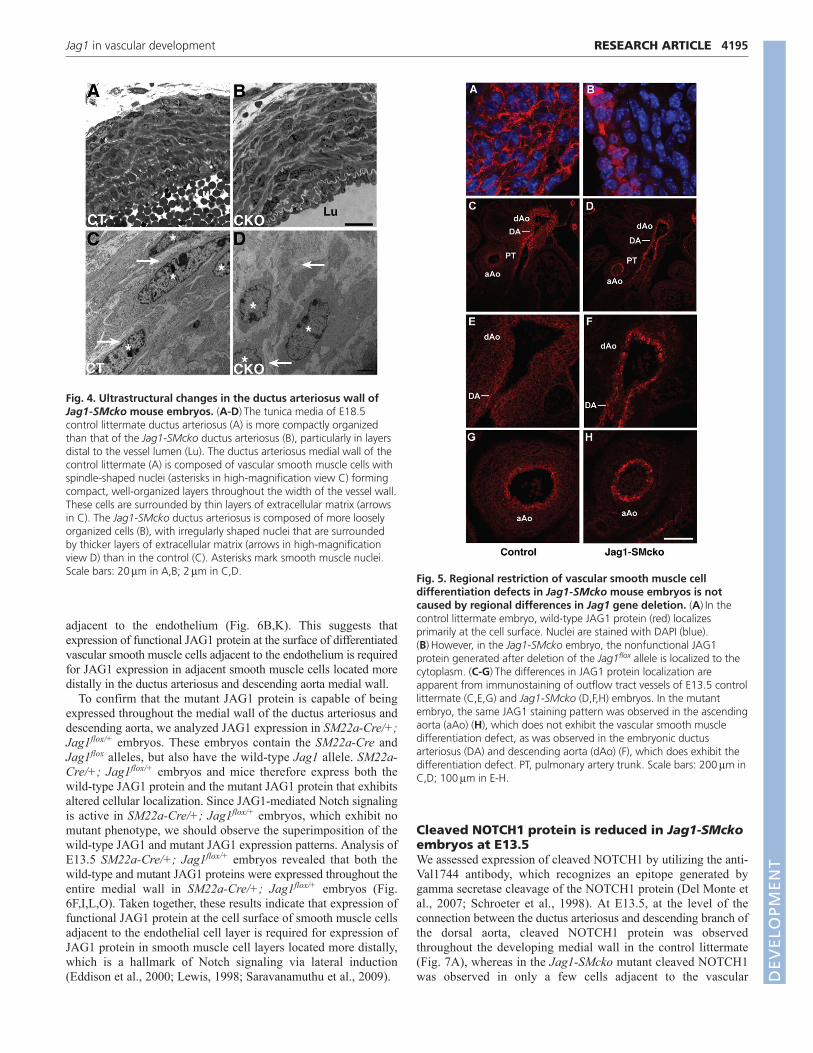
Fig. 4. Ultrastructural changes in the ductus arteriosus wall ofJag1-SMcko mouse embryos. (A-D)The tunica media of E18.5control littermate ductus arteriosus (A) is more compactly organizedthan that of the Jag1-SMcko ductus arteriosus (B), particularly in layersdistal to the vessel lumen (Lu). The ductus arteriosus medial wall of thecontrol littermate (A) is composed of vascular smooth muscle cells withspindle-shaped nuclei (asterisks in high-magnification view C) formingcompact, well-organized layers throughout the width of the vessel wall.These cells are surrounded by thin layers of extracellular matrix (arrowsin C). The Jag1-SMcko ductus arteriosus is composed of more looselyorganized cells (B), with irregularly shaped nuclei that are surroundedby thicker layers of extracellular matrix (arrows in high-magnificationview D) than in the control (C). Asterisks mark smooth muscle nuclei.Scale bars: 20m in A,B; 2m in C,D.
Fig. 5. Regional restriction of vascular smooth muscle celldifferentiation defects in Jag1-SMcko mouse embryos is notcaused by regional differences in Jag1 gene deletion. (A)In thecontrol littermate embryo, wild-type JAG1 protein (red) localizesprimarily at the cell surface. Nuclei are stained with DAPI (blue).(B)However, in the Jag1-SMcko embryo, the nonfunctional JAG1protein generated after deletion of the Jag1flox allele is localized to thecytoplasm. (C-G)The differences in JAG1 protein localization areapparent from immunostaining of outflow tract vessels of E13.5 controllittermate (C,E,G) and Jag1-SMcko (D,F,H) embryos. In the mutantembryo, the same JAG1 staining pattern was observed in the ascendingaorta (aAo) (H), which does not exhibit the vascular smooth muscledifferentiation defect, as was observed in the embryonic ductusarteriosus (DA) and descending aorta (dAo) (F), which does exhibit thedifferentiation defect. PT, pulmonary artery trunk. Scale bars: 200m inC,D; 100m in E-H.
DEVELO
PMENT

4196
endothelium (Fig. 7B). By E17.5, levels of cleaved NOTCH1 werereduced in the medial wall of both control and mutant embryos(Fig. 7C,D). However, cleaved NOTCH1 was still observed in bothcontrols and Jag1-SMcko mutants at E17.5 in other tissues, such asin the hair follicles in the skin (Fig. 7E,F). Analysis of NOTCH3
expression revealed that in both controls and Jag1-SMcko mutants,expression was high throughout the medial wall of the ductusarteriosus and descending dorsal aorta at both E13.5 and E17.5.(see Fig. S5 in the supplementary material).
Postnatal indomethacin injection rescues ductusarteriosus closure in a subset of Jag1-SMckoneonatesProstaglandins are key regulators of ductus arteriosus patency. Cyclo-oxygenases 1 and 2 (COX1 and 2; PTGS1 and 2; formally known asprostaglandin H synthases 1 and 2) are rate-limiting enzymesinvolved in the conversion of arachidonic acid to prostaglandins.Non-steroidal anti-inflammatory drugs such as indomethacin andibuprofen inhibit the formation of prostanoids by the COX enzymes(Mitchell and Warner, 2006). In premature infants exhibiting PDA,COX inhibitors are used clinically to induce ductus arteriosus closureby reducing prostaglandin levels, thereby inducing constriction of theductus arteriosus and closure of its lumen.
We assessed whether indomethacin administration to naturallyborn neonatal mice could rescue the PDA of Jag1-SMcko neonates.Indomethacin administered by subcutaneous injection within 12hours of birth rescued ductus arteriosus closure in ~50% of theJag1-SMcko mutants (Table 3; see Fig. S6 in the supplementary
RESEARCH ARTICLE Development 137 (24)
Fig. 6. JAG1 protein is highly expressed in the ductus arteriosusand descending aorta and is required for its own expression.Transverse sections of E13.5 mouse embryos through the level at whichthe ductus arteriosus (DA) connects to the descending aorta (dAo).Sections are immunostained with JAG1 antibody (red); A-L are alsocounterstained with DAPI (blue) to highlight nuclei. (A-C)Low-magnification views showing the JAG1 expression pattern in the ductusarteriosus and descending aorta, ascending aorta (aAo) and pulmonarytrunk (PT). (D-L)High-magnification views of the medial walls of theascending aorta (D-F), pulmonary trunk (G-I) and descending aorta (J-L).(M-O)JAG1 immunofluorescent staining reveals the superimposition ofthe wild-type JAG1 and mutant JAG1 expression patterns throughoutthe entire medial wall in the descending aorta of SM22a-Cre/+;Jag1flox/+ embryos (O). Scale bars: 200m in A-C; 40m in D-L; 25min M-O.
Fig. 7. Cleaved NOTCH1 protein expression is reduced in Jag1-SMcko mouse embryos at E13.5. (A-F)Immunofluorescent stainingwith anti-cleaved NOTCH1 antibody (Val1744 epitope; red) of a controllittermate (A,C,E) and Jag1-SMcko mutant (B,D,F) at E13.5 (A,B) andE17.5 (C-F). At the level of the connection between the ductusarteriosus (left) and descending branch of the dorsal aorta (right) (A-D),cleaved NOTCH1 protein was observed throughout the developingmedial wall in the control littermate at E13.5 (A), whereas in the Jag1-SMcko mutant (B) expression was only observed in a few cells adjacentto the vascular endothelium. By E17.5, cleaved NOTCH1 protein wasdownregulated in the medial wall of both control and mutant embryos(C,D), whereas nuclear cleaved NOTCH1 protein was still observed atthis stage in hair follicles in the skin (E,F, arrowheads). Scale bars:100m.
DEVELO
PMENT

material). These data suggest that the ductus arteriosus of someJag1-SMcko mutants retains sufficient smooth muscle to constrictin response to the drop in prostaglandin levels caused by postnatalindomethacin administration.
DISCUSSIONOne of the primary physiological signals regulating patency of theductus arteriosus is the level of circulating prostaglandins (Clyman,2006; Forsey et al., 2009; Schneider and Moore, 2006). The mostimportant of these is prostaglandin E2 (PGE2), which signals throughthe G protein-coupled receptor EP4 (also known as PTGER4).Targeted deletion of several genes encoding components of theprostaglandin signaling pathway, including the receptor EP4(Nguyen et al., 1997; Segi et al., 1998), 15-hydroxy prostaglandindehydrogenase (HPGD) (Coggins et al., 2002), the prostaglandintransporter PGT (also known as SLCO2A1) (Chang et al., 2010) andCOX1 and 2 (Loftin et al., 2001), lead to PDA in neonatal mice.However, several recent studies have demonstrated that, in both miceand humans, defects in smooth muscle cell differentiation or functionin the ductus arteriosus wall can cause PDA. Smooth muscle cellsdiffer from most other differentiated cell lineages in adult mammalsin that they are not irreversibly, terminally differentiated. Smoothmuscle cells, unlike skeletal muscle cells and cardiomyocytes, retainthe ability to reversibly alter their phenotype in response to variousenvironmental, physiological and genetic cues (McDonald andOwens, 2007; Owens et al., 2004). This property has been termedphenotypic switching or smooth muscle cell plasticity. Contractilesmooth muscle cells express high levels of contractile proteinsinvolved in establishing and maintaining myofilament structure andfunction, including SM22a, SMA and SMMHC. By contrast,synthetic smooth muscle cells express lower levels of contractilemuscle proteins and have higher rates of proliferation, migration andproduction of extracellular matrix components.
In humans, mutations in both the smooth muscle myosin heavychain (MYH11) and the smooth muscle a-actin (ACTA2) genes arefound in inherited forms of thoracic aortic aneurysm and dissectionwith PDA (Guo et al., 2007; Zhu et al., 2006). In mice, neural crestcell-specific deletion of the myocardin gene (Myocd) results in deathof the mutant neonates from PDA (Huang et al., 2008). The Myocdconditional mutant mice exhibit reduced expression of smoothmuscle-specific contractile proteins and display ultrastructuralfeatures indicating that vascular smooth muscle in the ductusarteriosus medial wall has acquired a synthetic, rather than acontractile, phenotype (Huang et al., 2008). Our analyses of Jag1-SMcko embryos and mice indicate that most smooth muscle cells inthe medial wall of the ductus arteriosus and descending aorta fail toexpress contractile smooth muscle cell proteins. In the Jag1-SMckomutants, the differentiation of contractile vascular smooth musclecells is confined to the region adjacent to the endothelial cell layer.
Our studies of both Jag1-SMcko mutants and embryos withendothelial cell-specific Jag1 deletion indicate that Jag1-mediatedNotch signaling induces Jag1 expression in the signal-receiving
cells, which is a hallmark of Notch signaling via lateral induction(Eddison et al., 2000; Lewis, 1998; Saravanamuthu et al., 2009).Several other studies have also demonstrated that expression ofJag1 is induced by Notch signal reception (Ascano et al., 2003;Daudet et al., 2007; Eddison et al., 2000; Luo et al., 1997; Ross andKadesch, 2004; Saravanamuthu et al., 2009), although some ofthese studies indicate that Jag1 might not be a direct Notch targetgene (Ross and Kadesch, 2004). Significantly, a recent studydemonstrated that both the JAG1 and NOTCH3 proteins wereinduced in vascular smooth muscle cells upon co-culture withendothelial cells (Liu et al., 2009).
Requirements for the NOTCH1 and NOTCH3 receptors havebeen studied previously in an in vivo vascular injury model (Li etal., 2009). Subsequent to vascular injury, smooth muscle cells inthe vascular wall in the injured area proliferate and form athickened layer of smooth muscle cells termed the neointima.Notch1+/– mice and mice with a heterozygous deletion of Notch1in smooth muscle cells exhibit a 70% decrease in neointimalformation after carotid artery ligation. However, neointimalformation is unaffected after carotid artery ligation in Notch3–/–
mice (Li et al., 2009). These data indicate that the NOTCH1receptor plays a dominant role in vascular smooth muscle cellsduring the response to vascular injury. Whether NOTCH1 plays asimilar dominant role during embryonic vascular smooth muscledevelopment is not known.
Our analyses, along with those of others (Benedito et al., 2009;High et al., 2008), suggest a model for JAG1-mediated Notchsignaling during development of the vascular wall of the ductusarteriosus and descending aorta (Fig. 8). In the ductus arteriosus
4197RESEARCH ARTICLEJag1 in vascular development
Fig. 8. Model for JAG1-Notch signaling in the ductus arteriosusand descending aorta. In the ductus arteriosus and descending aortaof wild-type mice, endothelial cells expressing JAG1 protein undergoheterotypic interactions with adjacent vascular mural/smooth musclecells expressing Notch receptors (most likely through NOTCH1,although a role for NOTCH3 cannot be excluded). Notch signalreception via lateral induction induces JAG1 expression in the receivingcells, leading to JAG1-mediated Notch signal transmission throughhomotypic interactions with the adjacent layer of smooth muscle cells.We further propose that Notch signal reception by vascular smoothmuscle cells is required for their differentiation into contractile, ratherthan synthetic, smooth muscle cells. In embryos with endothelial cell-specific Jag1 deletion, the mutant endothelial cells are unable to signalto adjacent smooth muscle cells, which consequently differentiate intosynthetic smooth muscle cells. In embryos with smooth muscle cell-specific Jag1 deletion, JAG1-expressing endothelial cells signalheterotypically to the adjacent layer of mutant vascular smooth musclecells. These cells differentiate as contractile smooth muscle andupregulate expression of the mutant JAG1 protein. However, thesemutant smooth muscle cells are unable to propagate the JAG1 signalthrough homotypic cellular interactions to more distal layers of smoothmuscle, which therefore differentiate as synthetic smooth muscle cells.
Table 3. Indomethacin injection rescues ductus arteriosusclosure in a subset of Jag1-SMcko mutant neonatesPhenotype Jag1-SMcko All other genotypes
Closed DA 4/7 (57%) 25/25 (100%)Patent DA 3/7 (43%) 0/25 (0%)
Indomethacin (6 mg/kg body weight) was injected subcutaneously into pups within12 hours of birth. After 6 hours, pups were euthanized and the ductus arteriosus(DA) scored for patency or closure among total littermates (from four litters) of Jag1-SMcko versus all other genotypes.
DEVELO
PMENT

4198
and descending aorta of wild-type mice, endothelial cellsexpressing JAG1 protein undergo heterotypic interactions withadjacent vascular mural/smooth muscle cells expressing Notchreceptors (most likely through the NOTCH1 receptor, although ourdata cannot exclude a role for the NOTCH3 receptor). Notch signalreception via lateral induction induces JAG1 expression in thereceiving cells, leading to JAG1-mediated Notch signaltransmission through homotypic interactions with the adjacentlayer of smooth muscle cells. We further propose that Notch signalreception by vascular smooth muscle cells is required for theirdifferentiation into contractile, rather than synthetic, smooth musclecells. In embryos with endothelial cell-specific Jag1 deletion, themutant endothelial cells are unable to signal to adjacent smoothmuscle cells, which consequently differentiate into syntheticsmooth muscle cells. In embryos with smooth muscle cell-specificJag1 deletion, JAG1-expressing endothelial cells signalheterotypically to the adjacent layer of mutant vascular smoothmuscle cells. These cells differentiate as contractile smooth muscleand upregulate expression of the mutant JAG1 protein. However,the Jag1-SMcko smooth muscle cells are unable to propagate theJAG1 signal through homotypic cellular interactions to more distallayers of smooth muscle, which therefore differentiate as syntheticsmooth muscle cells.
The mechanism that terminates the propagation of the JAG1-mediated signal in wild-type embryos is not understood. Ourmodel proposes that the Jag1 gene is induced in vascular smoothmuscle cell progenitors adjacent to the innermost layer of cellsexpressing the JAG1 ligand on their surface (Fig. 8). There is notan obvious signal to terminate propagation of this JAG1-mediated signal. Recent studies of NOTCH1-DLL1 signaling inmammalian cells have demonstrated that cis-interactionsresulting in mutual inactivation of NOTCH1 and DLL1 in thesame cell can generate a molecular switch between mutuallyexclusive signal-sending and signal-receiving states (Sprinzak etal., 2010). Whether such a mechanism contributes to terminationof the JAG1-mediated signal during the development of vascularsmooth muscle cells in the ductus arteriosus and descendingaorta remains to be determined.
It is also unclear why the vascular smooth muscle celldifferentiation defect in Jag1-SMcko mice is restricted to theductus arteriosus and descending aorta. Our data indicate thatregional restriction of the vascular smooth muscle celldifferentiation defect is not caused by regional differences inJag1 gene deletion. Jag1 deletion in the ascending aorta of theJag1-SMcko embryos, which does not exhibit the differentiationdefect, was equivalent to that in the embryonic ductus arteriosusand descending aorta, which do exhibit the defect (Fig. 5). Theductus arteriosus is a very specialized blood vessel, with avascular wall composed of highly differentiated and contractilesmooth muscle (Slomp et al., 1997). Smooth muscle of theductus arteriosus activates a unique transcriptional programduring its development (Ivey et al., 2008). This ductusarteriosus-specific transcriptional program might contribute tothe regional restriction of the vascular smooth muscledifferentiation defects exhibited by Jag1-SMcko mice.
The models for PDA exhibited by Jag1-SMcko mice and neuralcrest cell-specific Myocd gene deletion mice (Huang et al., 2008)represent a new paradigm for the etiology of PDA in mammals. Inthese mice, the differentiation of ductal smooth muscle cells alongthe synthetic, rather than the contractile, pathway leads to PDA.Further work will be required to determine whether this mechanismfor the etiology of PDA occurs in humans.
AcknowledgementsWe thank Greg Sousa and Chris Norton for helpful discussions. This work wassupported by grants from the NIH (HD034883) and the March of DimesFoundation (1-FY10-367) to T.G., and by a Center Grant from the NationalCancer Institute (CA034196) to the Jackson Laboratory. Deposited in PMC forrelease after 12 months.
Competing interests statementThe authors declare no competing financial interests.
Supplementary materialSupplementary material for this article is available athttp://dev.biologists.org/lookup/suppl/doi:10.1242/dev.052043/-/DC1
ReferencesAscano, J. M., Beverly, L. J. and Capobianco, A. J. (2003). The C-terminal PDZ-
ligand of JAGGED1 is essential for cellular transformation. J. Biol. Chem. 278,8771-8779.
Benedito, R., Roca, C., Sorensen, I., Adams, S., Gossler, A., Fruttiger, M. andAdams, R. H. (2009). The Notch ligands Dll4 and Jagged1 have opposingeffects on angiogenesis. Cell 137, 1124-1135.
Chang, H. Y., Locker, J., Lu, R. and Schuster, V. L. (2010). Failure of postnatalductus arteriosus closure in prostaglandin transporter-deficient mice. Circulation121, 529-536.
Clyman, R. I. (2006). Mechanisms regulating the ductus arteriosus. Biol. Neonate89, 330-335.
Coggins, K. G., Latour, A., Nguyen, M. S., Audoly, L., Coffman, T. M. andKoller, B. H. (2002). Metabolism of PGE2 by prostaglandin dehydrogenase isessential for remodeling the ductus arteriosus. Nat. Med. 8, 91-92.
Daudet, N., Ariza-McNaughton, L. and Lewis, J. (2007). Notch signalling isneeded to maintain, but not to initiate, the formation of prosensory patches inthe chick inner ear. Development 134, 2369-2378.
Del Monte, G., Grego-Bessa, J., Gonzalez-Rajal, A., Bolos, V. and De LaPompa, J. L. (2007). Monitoring Notch1 activity in development: evidence for afeedback regulatory loop. Dev. Dyn. 236, 2594-2614.
Eddison, M., Le Roux, I. and Lewis, J. (2000). Notch signaling in thedevelopment of the inner ear: lessons from Drosophila. Proc. Natl. Acad. Sci.USA 97, 11692-11699.
Forsey, J. T., Elmasry, O. A. and Martin, R. P. (2009). Patent arterial duct.Orphanet. J. Rare Dis. 4, 17.
Gridley, T. (2010). Notch signaling in the vasculature. Curr. Top. Dev. Biol. 92, 277-309.
Guo, D. C., Pannu, H., Tran-Fadulu, V., Papke, C. L., Yu, R. K., Avidan, N.,Bourgeois, S., Estrera, A. L., Safi, H. J., Sparks, E. et al. (2007). Mutations insmooth muscle alpha-actin (ACTA2) lead to thoracic aortic aneurysms anddissections. Nat. Genet. 39, 1488-1493.
High, F. A., Lu, M. M., Pear, W. S., Loomes, K. M., Kaestner, K. H. and Epstein,J. A. (2008). Endothelial expression of the Notch ligand Jagged1 is required forvascular smooth muscle development. Proc. Natl. Acad. Sci. USA 105, 1955-1959.
Hofmann, J. J. and Iruela-Arispe, M. L. (2007). Notch signaling in blood vessels:who is talking to whom about what? Circ. Res. 100, 1556-1568.
Holtwick, R., Gotthardt, M., Skryabin, B., Steinmetz, M., Potthast, R.,Zetsche, B., Hammer, R. E., Herz, J. and Kuhn, M. (2002). Smooth muscle-selective deletion of guanylyl cyclase-A prevents the acute but not chroniceffects of ANP on blood pressure. Proc. Natl. Acad. Sci. USA 99, 7142-7147.
Huang, J., Cheng, L., Li, J., Chen, M., Zhou, D., Lu, M. M., Proweller, A.,Epstein, J. A. and Parmacek, M. S. (2008). Myocardin regulates expression ofcontractile genes in smooth muscle cells and is required for closure of the ductusarteriosus in mice. J. Clin. Invest. 118, 515-525.
Ivey, K. N., Sutcliffe, D., Richardson, J., Clyman, R. I., Garcia, J. A. andSrivastava, D. (2008). Transcriptional regulation during development of theductus arteriosus. Circ. Res. 103, 388-395.
Kiernan, A. E., Xu, J. and Gridley, T. (2006). The Notch ligand JAG1 is requiredfor sensory progenitor development in the mammalian inner ear. PLoS Genet. 2,e4.
Koni, P. A., Joshi, S. K., Temann, U. A., Olson, D., Burkly, L. and Flavell, R. A.(2001). Conditional vascular cell adhesion molecule 1 deletion in mice: impairedlymphocyte migration to bone marrow. J. Exp. Med. 193, 741-754.
Lewis, J. (1998). Notch signalling and the control of cell fate choices invertebrates. Semin. Cell Dev. Biol. 9, 583-589.
Li, Y., Takeshita, K., Liu, P. Y., Satoh, M., Oyama, N., Mukai, Y., Chin, M. T.,Krebs, L., Kotlikoff, M. I., Radtke, F. et al. (2009). Smooth muscle Notch1mediates neointimal formation after vascular injury. Circulation 119, 2686-2692.
Liu, H., Kennard, S. and Lilly, B. (2009). NOTCH3 expression is induced in muralcells through an autoregulatory loop that requires endothelial-expressedJAGGED1. Circ. Res. 104, 466-475.
Loftin, C. D., Trivedi, D. B., Tiano, H. F., Clark, J. A., Lee, C. A., Epstein, J. A.,Morham, S. G., Breyer, M. D., Nguyen, M., Hawkins, B. M. et al. (2001).Failure of ductus arteriosus closure and remodeling in neonatal mice deficient in
RESEARCH ARTICLE Development 137 (24)
DEVELO
PMENT

cyclooxygenase-1 and cyclooxygenase-2. Proc. Natl. Acad. Sci. USA 98, 1059-1064.
Loomes, K. M., Underkoffler, L. A., Morabito, J., Gottlieb, S., Piccoli, D. A.,Spinner, N. B., Baldwin, H. S. and Oakey, R. J. (1999). The expression ofJagged1 in the developing mammalian heart correlates with cardiovasculardisease in Alagille syndrome. Hum. Mol. Genet. 8, 2443-2449.
Luo, B., Aster, J. C., Hasserjian, R. P., Kuo, F. and Sklar, J. (1997). Isolation andfunctional analysis of a cDNA for human Jagged2, a gene encoding a ligand forthe Notch1 receptor. Mol. Cell. Biol. 17, 6057-6067.
McDonald, O. G. and Owens, G. K. (2007). Programming smooth muscleplasticity with chromatin dynamics. Circ. Res. 100, 1428-1441.
Mitchell, J. A. and Warner, T. D. (2006). COX isoforms in the cardiovascularsystem: understanding the activities of non-steroidal anti-inflammatory drugs.Nat. Rev. Drug Discov. 5, 75-86.
Nguyen, M., Camenisch, T., Snouwaert, J. N., Hicks, E., Coffman, T. M.,Anderson, P. A., Malouf, N. N. and Koller, B. H. (1997). The prostaglandinreceptor EP4 triggers remodelling of the cardiovascular system at birth. Nature390, 78-81.
Owens, G. K., Kumar, M. S. and Wamhoff, B. R. (2004). Molecular regulationof vascular smooth muscle cell differentiation in development and disease.Physiol. Rev. 84, 767-801.
Phng, L. K. and Gerhardt, H. (2009). Angiogenesis: a team effort coordinated byNotch. Dev. Cell 16, 196-208.
Roca, C. and Adams, R. H. (2007). Regulation of vascular morphogenesis byNotch signaling. Genes Dev. 21, 2511-2524.
Ross, D. A. and Kadesch, T. (2004). Consequences of Notch-mediated inductionof Jagged1. Exp. Cell Res. 296, 173-182.
Saravanamuthu, S. S., Gao, C. Y. and Zelenka, P. S. (2009). Notch signaling isrequired for lateral induction of Jagged1 during FGF-induced lens fiberdifferentiation. Dev. Biol. 332, 166-176.
Schneider, D. J. and Moore, J. W. (2006). Patent ductus arteriosus. Circulation114, 1873-1882.
Schroeter, E. H., Kisslinger, J. A. and Kopan, R. (1998). Notch-1 signallingrequires ligand-induced proteolytic release of intracellular domain. Nature 393,382-386.
Segi, E., Sugimoto, Y., Yamasaki, A., Aze, Y., Oida, H., Nishimura, T., Murata,T., Matsuoka, T., Ushikubi, F., Hirose, M. et al. (1998). Patent ductusarteriosus and neonatal death in prostaglandin receptor EP4-deficient mice.Biochem. Biophys. Res. Commun. 246, 7-12.
Slomp, J., Gittenberger-de Groot, A. C., Glukhova, M. A., Conny vanMunsteren, J., Kockx, M. M., Schwartz, S. M. and Koteliansky, V. E. (1997).Differentiation, dedifferentiation, and apoptosis of smooth muscle cells duringthe development of the human ductus arteriosus. Arterioscler. Thromb. Vasc.Biol. 17, 1003-1009.
Spandidos, A., Wang, X., Wang, H., Dragnev, S., Thurber, T. and Seed, B.(2008). A comprehensive collection of experimentally validated primers forpolymerase chain reaction quantitation of murine transcript abundance. BMCGenomics 9, 633.
Sprinzak, D., Lakhanpal, A., Lebon, L., Santat, L. A., Fontes, M. E.,Anderson, G. A., Garcia-Ojalvo, J. and Elowitz, M. B. (2010). Cis-interactions between Notch and Delta generate mutually exclusive signallingstates. Nature 465, 86-90.
Villa, N., Walker, L., Lindsell, C. E., Gasson, J., Iruela-Arispe, M. L. andWeinmaster, G. (2001). Vascular expression of Notch pathway receptors andligands is restricted to arterial vessels. Mech. Dev. 108, 161-164.
Xue, Y., Gao, X., Lindsell, C. E., Norton, C. R., Chang, B., Hicks, C., Gendron-Maguire, M., Rand, E. B., Weinmaster, G. and Gridley, T. (1999). Embryoniclethality and vascular defects in mice lacking the Notch ligand Jagged1. Hum.Mol. Genet. 8, 723-730.
Zhu, L., Vranckx, R., Khau Van Kien, P., Lalande, A., Boisset, N., Mathieu, F.,Wegman, M., Glancy, L., Gasc, J. M., Brunotte, F. et al. (2006). Mutations inmyosin heavy chain 11 cause a syndrome associating thoracic aorticaneurysm/aortic dissection and patent ductus arteriosus. Nat. Genet. 38, 343-349.
4199RESEARCH ARTICLEJag1 in vascular development
DEVELO
PMENT


















