
Parasite abundance contributes to condition-dependent dispersal in a wild population of large...
5
1121 Parasite abundance contributes to condition-dependent dispersal in a wild population of large herbivore Lucie Debeffe, Nicolas Morellet, Hélène Verheyden-Tixier, Hervé Hoste, Jean-Michel Gaillard, Bruno Cargnelutti, Denis Picot, Julie Sevila and A. J. Mark Hewison Lucie Debeffe (lucie.debeff[email protected]), N. Morellet, H. Verheyden-Tixier, B. Cargnelutti, D. Picot, J. Sevila and A. J. M. Hewison, INRA, UR35 CEFS, B.P-52627, FR-31326 Castanet-Tolosan, France. – LD and J.-M. Gaillard, Univ. Claude Bernard Lyon1, LBBE UMR5558, FR-69622 Villeurbanne, France. – H. Hoste and JS, INRA, UMR1225 IHAP, FR-31076 Toulouse, France and,Univ. de Toulouse, ENVT UMR1225, FR-31076 Toulouse, France. Parasite abundance has been shown to have major consequences for host fitness components such as survival and reproduction. However, although natal dispersal is a key life history trait, whether an individual’s decision to disperse or not is influenced by the abundance of parasites it carries remains mostly unknown. Current and opposing hypotheses suggest that infected individuals should either be philopatric to avoid the energetic costs of dispersal (condition dependence) or disperse to escape from heavily parasitised habitats. From intensive monitoring of a roe deer population inhabiting a multi-use and spatially heterogeneous agricultural landscape, we evaluated the link between an individual’s parasite abundance and its propensity to disperse, while accounting for confounding effects of body mass. Dispersal propensity generally decreased with both increasing nematode abundance and with decreasing body mass. Within the dispersing segment of the population, individuals with high nematode abundance left their natal home range later in the season than less parasitised deer. ese results clearly show that parasite abundance is an important component of condition- dependent dispersal in large herbivores. However, unexpectedly, three individuals that were both heavily parasitised and of low body mass dispersed. We suggest that this ‘leave it’ response to high parasite levels in the natal habitat could represent a last ditch attempt to improve reproductive prospects, constituting a form of emergency life history strategy. Oikos 123: 1121–1125, 2014 doi: 10.1111/oik.01396 © 2014 e Authors. Oikos © 2014 Nordic Society Oikos Subject Editor: Dries Bonte. Accepted 4 February 2014 Parasites impact their host in a variety of ways, often inducing immune responses and/or behavioural modifica- tions. For instance, locomotion and activity of infested individuals are reduced in juvenile toads Bufo bufo (Cameron et al. 1993). In addition, parasites also extract resources from their host and, consequently, may nega- tively impact their fitness (Poulin 2007). High parasite abundances lead to reduced fecundity in mountain hares Lepus timidus (Newey and irgood 2004) and reindeer Rangifer tarandus (Stien et al. 2002), lowered survival in cliff swallows Hirundo pyrrhonota (Brown and Brown 1986) and lowered growth rate and reproductive success (i.e. presence of young after emergence) in yellow bellied marmots Marmota flaviventris (Van Vuren 1996). How- ever, while natal dispersal is a key life history trait for individual fitness (Clobert et al. 2001) which could potentially be affected by parasitism (Heeb et al. 1999, Boulinier et al. 2001), how parasite abundance influences the propensity of individuals to disperse remains virtually unknown. e few published studies have focused on ectoparasites (Brown and Brown 1992, Sorci et al. 1994, Van Vuren 1996, Heeb et al. 1999), while the only study to date on endoparasites failed to find a link between parasite abundance and natal dispersal in Drosophila melanogaster (Jaenike et al. 1995). Parasites have been hypothesised to influence natal dispersal in opposing ways. First, highly parasitised indivi- duals might be more likely to be philopatric or to disperse over shorter distances than healthy individuals (Dufty and Belthoff 2001) so as to minimise dispersal costs (Bonte et al. 2012) (i.e. condition-dependent dispersal). at is, if highly parasitised individuals are unable to cover the energetic costs associated with natal dispersal (Dufty and Belthoff 2001), they should remain philopatric. Fur- themore, it may be less costly from a life history perspective for heavily parasitised individuals to remain philopatric if there is local adaptation to a given parasite community (Kaltz and Shykoff 1998, Heeb et al. 1999, Møller and Szep 2011) and/or if the mosaic of parasite strains potentially encountered during dispersal imposes a supplementary cost (Møller and Szep 2011). ese kinds of effects of parasitism on natal dispersal behaviour have been reported in a great tit Parus major population, where recruits from infested broods had a lower probability of dispersing, and when they did disperse, they did so over shorter distances (Heeb et al. 1999). Alternatively, if natal dispersal is a way
Transcript of Parasite abundance contributes to condition-dependent dispersal in a wild population of large...

1121
Parasite abundance contributes to condition-dependent dispersal in a wild population of large herbivore
Lucie Debeffe , Nicolas Morellet , H é l è ne Verheyden-Tixier , Herv é Hoste , Jean-Michel Gaillard , Bruno Cargnelutti , Denis Picot , Julie Sevila and A. J. Mark Hewison
Lucie Debeff e (lucie.debeff [email protected]), N. Morellet, H. Verheyden-Tixier, B. Cargnelutti, D. Picot, J. Sevila and A. J. M. Hewison, INRA, UR35 CEFS, B.P-52627, FR-31326 Castanet-Tolosan, France. – LD and J.-M. Gaillard, Univ. Claude Bernard Lyon1, LBBE UMR5558, FR-69622 Villeurbanne, France. – H. Hoste and JS, INRA, UMR1225 IHAP, FR-31076 Toulouse, France and, Univ. de Toulouse, ENVT UMR1225, FR-31076 Toulouse, France.
Parasite abundance has been shown to have major consequences for host fi tness components such as survival and reproduction. However, although natal dispersal is a key life history trait, whether an individual ’ s decision to disperse or not is infl uenced by the abundance of parasites it carries remains mostly unknown. Current and opposing hypotheses suggest that infected individuals should either be philopatric to avoid the energetic costs of dispersal (condition dependence) or disperse to escape from heavily parasitised habitats. From intensive monitoring of a roe deer population inhabiting a multi-use and spatially heterogeneous agricultural landscape, we evaluated the link between an individual ’ s parasite abundance and its propensity to disperse, while accounting for confounding eff ects of body mass. Dispersal propensity generally decreased with both increasing nematode abundance and with decreasing body mass. Within the dispersing segment of the population, individuals with high nematode abundance left their natal home range later in the season than less parasitised deer. Th ese results clearly show that parasite abundance is an important component of condition-dependent dispersal in large herbivores. However, unexpectedly, three individuals that were both heavily parasitised and of low body mass dispersed. We suggest that this ‘ leave it ’ response to high parasite levels in the natal habitat could represent a last ditch attempt to improve reproductive prospects, constituting a form of emergency life history strategy.
Oikos 123: 1121–1125, 2014 doi: 10.1111/oik.01396
© 2014 Th e Authors. Oikos © 2014 Nordic Society Oikos Subject Editor: Dries Bonte. Accepted 4 February 2014
Parasites impact their host in a variety of ways, often inducing immune responses and/or behavioural modifi ca-tions. For instance, locomotion and activity of infested individuals are reduced in juvenile toads Bufo bufo (Cameron et al. 1993). In addition, parasites also extract resources from their host and, consequently, may nega-tively impact their fi tness (Poulin 2007). High parasite abundances lead to reduced fecundity in mountain hares Lepus timidus (Newey and Th irgood 2004) and reindeer Rangifer tarandus (Stien et al. 2002), lowered survival in cliff swallows Hirundo pyrrhonota (Brown and Brown 1986) and lowered growth rate and reproductive success (i.e. presence of young after emergence) in yellow bellied marmots Marmota fl aviventris (Van Vuren 1996). How-ever, while natal dispersal is a key life history trait for individual fi tness (Clobert et al. 2001) which could potentially be aff ected by parasitism (Heeb et al. 1999, Boulinier et al. 2001), how parasite abundance infl uences the propensity of individuals to disperse remains virtually unknown. Th e few published studies have focused on ectoparasites (Brown and Brown 1992, Sorci et al. 1994, Van Vuren 1996, Heeb et al. 1999), while the only study to date on endoparasites failed to fi nd a link between
parasite abundance and natal dispersal in Drosophila melanogaster (Jaenike et al. 1995).
Parasites have been hypothesised to infl uence natal dispersal in opposing ways. First, highly parasitised indivi-duals might be more likely to be philopatric or to disperse over shorter distances than healthy individuals (Dufty and Belthoff 2001) so as to minimise dispersal costs (Bonte et al. 2012) (i.e. condition-dependent dispersal). Th at is, if highly parasitised individuals are unable to cover the energetic costs associated with natal dispersal (Dufty and Belthoff 2001), they should remain philopatric. Fur-themore, it may be less costly from a life history perspective for heavily parasitised individuals to remain philopatric if there is local adaptation to a given parasite community (Kaltz and Shykoff 1998, Heeb et al. 1999, M ø ller and Szep 2011) and/or if the mosaic of parasite strains potentially encountered during dispersal imposes a supplementary cost (M ø ller and Szep 2011). Th ese kinds of eff ects of parasitism on natal dispersal behaviour have been reported in a great tit Parus major population, where recruits from infested broods had a lower probability of dispersing, and when they did disperse, they did so over shorter distances (Heeb et al. 1999). Alternatively, if natal dispersal is a way

1122
for individuals to escape from highly parasitised environ-ments (Boulinier et al. 2001), dispersal propensity and distance should increase with parasite abundance (Brown and Brown 1992, Van Vuren 1996, M ø ller et al. 2004). For instance, dispersing yellow bellied marmots had higher ectoparasite loads than their philopatric counterparts (Van Vuren 1996), while cliff swallows with low parasite levels had a higher probability of returning to their natal site and being recruited locally (Brown and Brown 1992).
We aimed to assess the link between endoparasite abundance, namely gastrointestinal nematodes, and natal dispersal behaviour in a roe deer, Capreolus capreolus , population inhabiting a multi-use, spatially heteroge-neous, landscape. Nematode abundance is known to impact body condition in large herbivores (Stien et al. 2002, Body et al. 2011). Hence, because we have previously shown that dispersal behaviour in roe deer is condition-dependent such that both dispersal propensity and distance increase with increasing mass (Debeff e et al. 2012), we predicted that both dispersal propensity and distance should decrease with increasing nematode abundance. Indeed, condition dependence is a multi-factorial construct (Ims and Hjermann 2001), potentially involving several components including both body mass and parasitism. Because body mass and parasite abundance are negatively related (Body et al. 2011), in order to disentangle the eff ects of these two factors on dispersal behaviour, we fi rst controlled for the body mass – parasite abundance rela-tionship. We were thus subsequently able to distinguish between the eff ects of the body mass component and the parasitism component of condition-dependent dispersal. In addition, because parasitism directly aff ects individual body condition by increasing energetic expenditure, parasite abundance may infl uence the timing of dispersal (Dufty and Belthoff 2001). Hence, as individuals require a certain amount of energy in order to disperse, we expected highly parasitised individuals, presumably in lower condi-tion, to delay dispersal during a given season until they gained suffi cient condition.
Methods
Study site and sampling
Th e data were collected in a heterogeneous agricultural land-scape (43 ° 27 ′ N, 0 ° 85 ′ E) covering around 10 000 ha in southwest France. We identifi ed three sectors of contrasting landscape structure based on the proportion of woodland: a closed sector of two forest blocks, an open landscape with a few fragmented woodlots, and an intermediate sector with inter-connected woodland fragments (Morellet et al. 2011). From 2007 to 2013, we captured 67 juvenile roe deer (less than one year old) during winter, recorded their body mass and sampled faeces directly from the rec-tum, before fi tting them with a GPS collar and releasing them on site.
Assessing natal dispersal
Natal dispersal corresponds to permanent emigration from the natal range to a distinct adult range such that
pre-dispersal locations do not overlap post-dispersal loca-tions (Kenward et al. 2002). As described in Debeff e et al. (2012), to discriminate dispersers from philopatric animals, we examined the degree of range overlap and used the range stability index of Roshier and Reid (2003). Dispersal behaviour was monitored for a total of 67 individuals, of which, 31 dispersed while 36 remained philo-patric. For the 31 individuals that dispersed, the dispersal date corresponded to the last date an individual was located inside its natal range, while dispersal distance was calculated as the Euclidean distance between the pre-dispersal range and the most distant location of the movement trajectory.
Estimating endoparasite abundance
All faeces were collected during winter, between early January and mid February, so that confounding seasonal variations in parasite abundance were minimised. As a result, sampling for parasites occurred two to three months before the dispersal period, so that parasite exposure was a priori equivalent for individuals that subsequently were either philopatric or dispersed. From one gram of fresh faeces, we identifi ed the eggs of gastrointestinal parasites: Strongylida , Trichocephalida , Ascaridida , three diff erent orders of worms, were pooled together as ‘ nematodes ’ . We estimated parasite abundance, defi ned as the number of individuals of a particular parasite in a single host regardless of whether or not the host is infected (Bush et al. 1997), as the abundance of these nematodes. Nematode abundance was measured using faecal egg counts (R ó zsa et al. 2000) which were log-transformed as ln (x � 25) to achieve normality. When parasite abundance was estimated to be less than 100 eggs per gram of faeces (corresponding to less that two eggs on the slide), that individual was con-sidered to be non-infested. Faecal egg counts provide a reli-able estimate of gastrointestinal parasite abundances (Raynaud 1970), and this method has been validated on another roe deer population for Trichuris using 244 roe deer shot during the hunting season (linear regression between faecal egg counts and number of adult worms found at necroscopy: n � 244, r ² � 0.811, p � 0.001) (see Appendix in Body et al. 2011). Since parasite abun-dance is negatively correlated with body mass in roe deer (Body et al. 2011), for subsequent analyses, we generated a standardised measure of nematode abundance as the resid-uals from the linear regression between nematode abun-dance and body mass (McCullagh and Nelder 1989), allowing us to discriminate between the eff ects of body mass per se and the eff ects of parasite abundance on the thee dispersal metrics (propensity, date and distance).
Data analysis
We assessed the infl uence of parasite abundance on the three dispersal metrics using generalized linear mixed models (GLMM) in R software ver. 3.0.0. We included body mass as a fi xed eff ect in our models, while landscape sector was included as a random factor to account for observed variation in dispersal among sectors (Debeff e et al. 2012). Both body mass and the standardised abundance of nematodes were included in the models to
1123
evaluate the contribution of these two components of condition-dependent dispersal. We also included the body mass x nematode abundance interaction in the full model to test for consistency in the eff ects of parasite abundance on dispersal behaviour across individuals of diff ering body mass. To simplify interpretation, individuals were assigned to one of three body mass categories: heavy individuals, from 17.9 to 20 kg (20% of the data), average individuals, from 15 to 17.9 kg (60% of the data) and light individuals, from 11.5 to 15 kg (20% of the data). Body mass of male and female juveniles did not diff er (mean � SD: 16.74 � 1.85 kg and 16.12 � 1.68 kg for males and females respectively, n � 67, r 2 � 0.03, F 1,65 � 2.04, p � 0.16), hence we did not correct individual body mass for any sex-specifi c diff erence. We fi tted fi ve candidate models for each metric of dispersal behaviour: dispersal propensity, distance and departure date. We selected the models with the lowest AICc value (Akaike information criterion corrected for small sample size, Burnham and Anderson 1998). Analyses using body mass as a continuous variable did not change the results or our interpretations and are provided in the electronic supplementary materials.
Results
Dispersal propensity
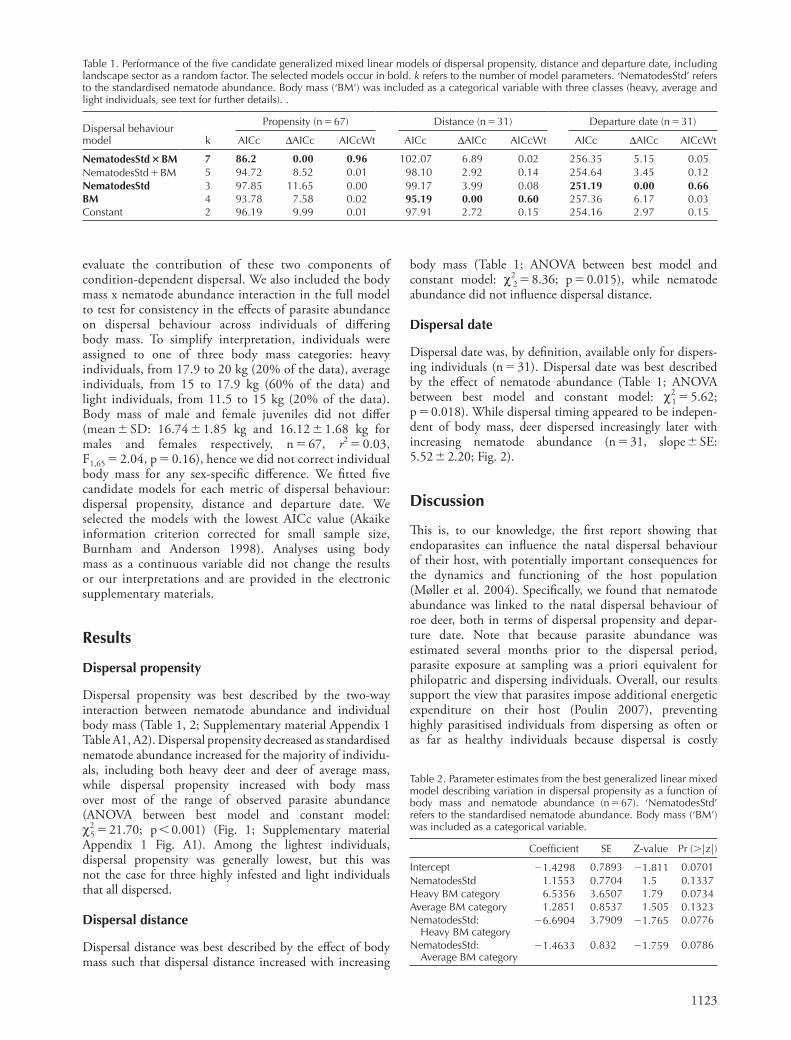
Dispersal propensity was best described by the two-way interaction between nematode abundance and individual body mass (Table 1, 2; Supplementary material Appendix 1 Table A1, A2). Dispersal propensity decreased as standardised nematode abundance increased for the majority of individu-als, including both heavy deer and deer of average mass, while dispersal propensity increased with body mass over most of the range of observed parasite abundance (ANOVA between best model and constant model: χ 2 5 � 21.70; p � 0.001) (Fig. 1; Supplementary material Appendix 1 Fig. A1). Among the lightest individuals, dispersal propensity was generally lowest, but this was not the case for three highly infested and light individuals that all dispersed.
Dispersal distance
Dispersal distance was best described by the eff ect of body mass such that dispersal distance increased with increasing
body mass (Table 1; ANOVA between best model and constant model: χ 2 2 � 8.36; p � 0.015), while nematode abundance did not infl uence dispersal distance.
Dispersal date
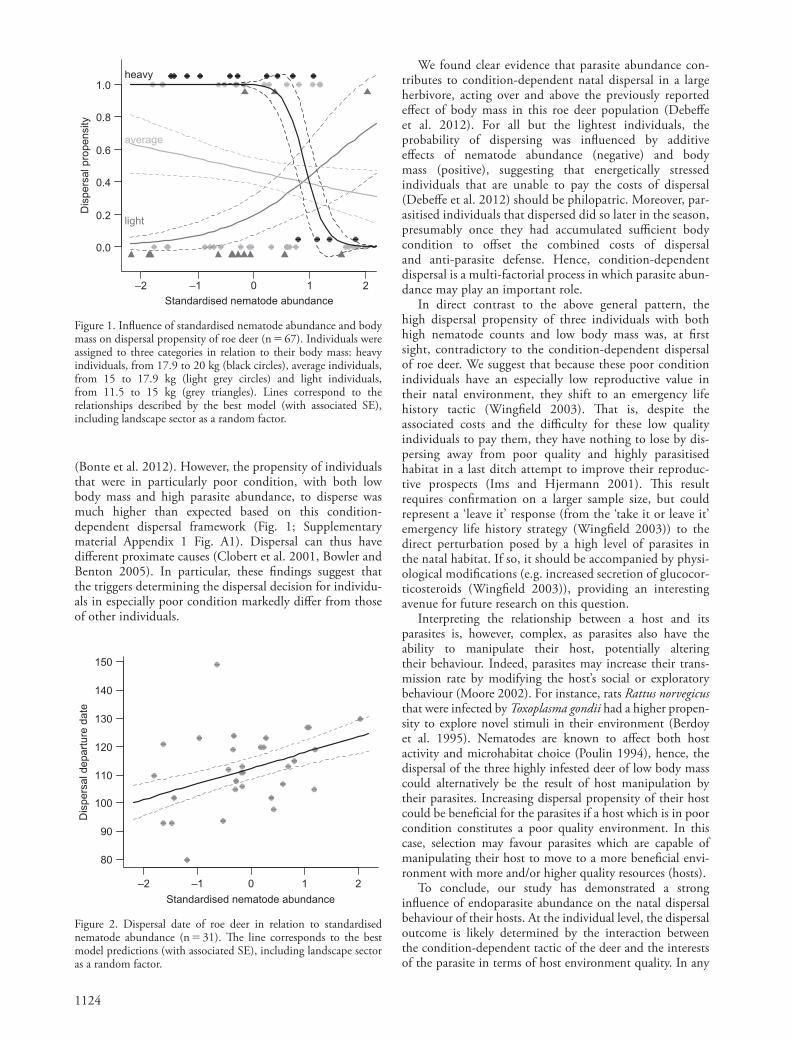
Dispersal date was, by defi nition, available only for dispers-ing individuals (n � 31). Dispersal date was best described by the eff ect of nematode abundance (Table 1; ANOVA between best model and constant model: χ 2 1 � 5.62; p � 0.018). While dispersal timing appeared to be indepen-dent of body mass, deer dispersed increasingly later with increasing nematode abundance (n � 31, slope � SE: 5.52 � 2.20; Fig. 2).
Discussion
Th is is, to our knowledge, the fi rst report showing that endoparasites can infl uence the natal dispersal behaviour of their host, with potentially important consequences for the dynamics and functioning of the host population (M ø ller et al. 2004). Specifi cally, we found that nematode abundance was linked to the natal dispersal behaviour of roe deer, both in terms of dispersal propensity and depar-ture date. Note that because parasite abundance was estimated several months prior to the dispersal period, parasite exposure at sampling was a priori equivalent for philopatric and dispersing individuals. Overall, our results support the view that parasites impose additional energetic expenditure on their host (Poulin 2007), preventing highly parasitised individuals from dispersing as often or as far as healthy individuals because dispersal is costly
Table 2. Parameter estimates from the best generalized linear mixed model describing variation in dispersal propensity as a function of body mass and nematode abundance (n � 67). ‘ NematodesStd ’ refers to the standardised nematode abundance. Body mass ( ‘ BM ’ ) was included as a categorical variable.
Coeffi cient SE Z-value Pr ( � |z|)
Intercept � 1.4298 0.7893 � 1.811 0.0701NematodesStd 1.1553 0.7704 1.5 0.1337Heavy BM category 6.5356 3.6507 1.79 0.0734Average BM category 1.2851 0.8537 1.505 0.1323NematodesStd:
Heavy BM category � 6.6904 3.7909 � 1.765 0.0776
NematodesStd: Average BM category
� 1.4633 0.832 � 1.759 0.0786
Table 1. Performance of the fi ve candidate generalized mixed linear models of dispersal propensity, distance and departure date, including landscape sector as a random factor. The selected models occur in bold. k refers to the number of model parameters. ‘ NematodesStd ’ refers to the standardised nematode abundance. Body mass ( ‘ BM ’ ) was included as a categorical variable with three classes (heavy, average and light individuals, see text for further details). .
Dispersal behaviourmodel
Propensity (n � 67) Distance (n � 31) Departure date (n � 31)
k AICc Δ AICc AICcWt AICc Δ AICc AICcWt AICc Δ AICc AICcWt
NematodesStd � BM 7 86.2 0.00 0.96 102.07 6.89 0.02 256.35 5.15 0.05NematodesStd � BM 5 94.72 8.52 0.01 98.10 2.92 0.14 254.64 3.45 0.12 NematodesStd 3 97.85 11.65 0.00 99.17 3.99 0.08 251.19 0.00 0.66 BM 4 93.78 7.58 0.02 95.19 0.00 0.60 257.36 6.17 0.03Constant 2 96.19 9.99 0.01 97.91 2.72 0.15 254.16 2.97 0.15
1124
We found clear evidence that parasite abundance con-tributes to condition-dependent natal dispersal in a large herbivore, acting over and above the previously reported eff ect of body mass in this roe deer population (Debeff e et al. 2012). For all but the lightest individuals, the probability of dispersing was infl uenced by additive eff ects of nematode abundance (negative) and body mass (positive), suggesting that energetically stressed individuals that are unable to pay the costs of dispersal (Debeff e et al. 2012) should be philopatric. Moreover, par-asitised individuals that dispersed did so later in the season, presumably once they had accumulated suffi cient body condition to off set the combined costs of dispersal and anti-parasite defense. Hence, condition-dependent dispersal is a multi-factorial process in which parasite abun-dance may play an important role.
In direct contrast to the above general pattern, the high dispersal propensity of three individuals with both high nematode counts and low body mass was, at fi rst sight, contradictory to the condition-dependent dispersal of roe deer. We suggest that because these poor condition individuals have an especially low reproductive value in their natal environment, they shift to an emergency life history tactic (Wingfi eld 2003). Th at is, despite the associated costs and the diffi culty for these low quality individuals to pay them, they have nothing to lose by dis-persing away from poor quality and highly parasitised habitat in a last ditch attempt to improve their reproduc-tive prospects (Ims and Hjermann 2001). Th is result requires confi rmation on a larger sample size, but could represent a ‘ leave it ’ response (from the ‘ take it or leave it ’ emergency life history strategy (Wingfi eld 2003)) to the direct perturbation posed by a high level of parasites in the natal habitat. If so, it should be accompanied by physi-ological modifi cations (e.g. increased secretion of glucocor-ticosteroids (Wingfi eld 2003)), providing an interesting avenue for future research on this question.
Interpreting the relationship between a host and its parasites is, however, complex, as parasites also have the ability to manipulate their host, potentially altering their behaviour. Indeed, parasites may increase their trans-mission rate by modifying the host ’ s social or exploratory behaviour (Moore 2002). For instance, rats Rattus norvegicus that were infected by Toxoplasma gondii had a higher propen-sity to explore novel stimuli in their environment (Berdoy et al. 1995). Nematodes are known to aff ect both host activity and microhabitat choice (Poulin 1994), hence, the dispersal of the three highly infested deer of low body mass could alternatively be the result of host manipulation by their parasites. Increasing dispersal propensity of their host could be benefi cial for the parasites if a host which is in poor condition constitutes a poor quality environment. In this case, selection may favour parasites which are capable of manipulating their host to move to a more benefi cial envi-ronment with more and/or higher quality resources (hosts).
To conclude, our study has demonstrated a strong infl uence of endoparasite abundance on the natal dispersal behaviour of their hosts. At the individual level, the dispersal outcome is likely determined by the interaction between the condition-dependent tactic of the deer and the interests of the parasite in terms of host environment quality. In any
(Bonte et al. 2012). However, the propensity of individuals that were in particularly poor condition, with both low body mass and high parasite abundance, to disperse was much higher than expected based on this condition-dependent dispersal framework (Fig. 1; Supplementary material Appendix 1 Fig. A1). Dispersal can thus have diff erent proximate causes (Clobert et al. 2001, Bowler and Benton 2005). In particular, these fi ndings suggest that the triggers determining the dispersal decision for individu-als in especially poor condition markedly diff er from those of other individuals.
–2 –1 0 1 2
0.0
0.2
0.4
0.6
0.8
1.0
Standardised nematode abundance
Dis
pers
al p
rope
nsity
light
average
heavy
Figure 1. Infl uence of standardised nematode abundance and body mass on dispersal propensity of roe deer (n � 67). Individuals were assigned to three categories in relation to their body mass: heavy individuals, from 17.9 to 20 kg (black circles), average individuals, from 15 to 17.9 kg (light grey circles) and light individuals, from 11.5 to 15 kg (grey triangles). Lines correspond to the relationships described by the best model (with associated SE), including landscape sector as a random factor.
–2 –1 0 1 2
80
90
100
110
120
130
140
150
Standardised nematode abundance
Dis
pers
al d
epar
ture
dat
e
Figure 2. Dispersal date of roe deer in relation to standardised nematode abundance (n � 31). Th e line corresponds to the best model predictions (with associated SE), including landscape sector as a random factor.

1125
Heeb, P. et al. 1999. Ectoparasite infestation and sex-biased local recruitment of hosts. – Nature 400: 63 – 65.
Ims, R. A. and Hjermann, D. O. 2001. Condition-dependent dispersal. – In: Clobert, J. et al. (eds), Dispersal. Oxford Univ. Press, pp. 203 – 216.
Jaenike, J. et al. 1995. Parasite-induced mortality in Mycophagous drosophila . – Ecology 76: 383 – 391.
Kaltz, O. and Shykoff , J. A. 1998. Local adaptation in host – parasite systems. – Heredity 81: 361 – 370.
Kenward, R. E. et al. 2002. From marking to modelling: dispersal study techniques for land vertebrates. – In: Bullock, J. M. et al. (eds), Dispersal ecology. Blackwell, pp. 50 – 71.
McCullagh, P. and Nelder, J. A. 1989. Generalized linear models. – Chapman and Hall.
M ø ller, A. P. and Szep, T. 2011. Th e role of parasites in ecology and evolution of migration and migratory connectivity. – J. Ornithol. 152: 141 – 150.
M ø ller, A. P. et al. 2004. Parasitism, host immune defence and dispersal. – J. Evol. Biol. 17: 603 – 612.
Moore, J. 2002. Parasites and the behaviour of animals. – Oxford Univ. Press.
Morellet, N. et al. 2011. Landscape composition infl uences roe deer habitat selection at both home range and landscape scales. – Landscape Ecol. 26: 999 – 1010.
Newey, S. and Th irgood, S. 2004. Parasite-mediated reduction in fecundity of mountain hares. – Proc. R. Soc. B 271: S413 – S415.
Poulin, R. 1994. Meta-analysis of parasite-induced behavioural changes. – Anim. Behav. 48: 137 – 146.
Poulin, R. 2007. Evolutionary ecology of parasites. – Princeton Univ. Press.
Raynaud, J. 1970. Etude de l ’ effi cacit é d ’ une technique de coproscopie quantitative pour le diagnostic de routine et le contr ô le des infestations parasitaires des bovins, ovins, é quins et porcins. – Ann. Parasitol. Hum. Compar é e 45: 321 – 342.
Roshier, D. A. and Reid, J. R. W. 2003. On animal distributions in dynamic landscapes. – Ecography 26: 539 – 544.
R ó zsa, L. et al. 2000. Quantifying parasites in samples of hosts. – J. Parasitol. 86: 228 – 232.
Sorci, G. et al. 1994. Maternal parasite load increases sprint speed and philopatry in female off spring of the common lizard. – Am. Nat. 144: 153 – 164.
Stien, A. et al. 2002. Th e impact of gastrointestinal nematodes on wild reindeer: experimental and cross-sectional studies. – J. Anim. Ecol. 71: 937 – 945.
Van Vuren, D. 1996. Ectoparasites, fi tness, and social behaviour of yellow-bellied marmots. – Ethology 102: 686 – 694.
Wingfi eld, J. C. 2003. Control of behavioural strategies for capricious environments. – Anim. Behav. 66: 807 – 815.
case, by modifying their host ’ s natal dispersal behaviour, endoparasites likely have an important impact on the dynamics and functioning of their host populations (M ø ller et al. 2004).
Acknowledgements – Th e study was supported by an INRA grant. We thank the local hunting associations and all volunteers for their assistance during capture and Eric Pardo for carrying out fecal analyses. We thank Atle Mysterud and for comments on a previous version of this manuscript.
References
Berdoy, M. et al. 1995. Parasite-altered behaviour: is the eff ect of Toxoplasma – Gondii on Rattus norvegicus specifi c? – Parasitology 111: 403 – 409.
Body, G. et al. 2011. Population density and phenotypic attributes infl uence the level of nematode parasitism in roe deer. – Oecologia 167: 635 – 646.
Bonte, D. et al. 2012. Costs of dispersal. – Biol. Rev. 87: 290 – 312.
Boulinier, T. et al. 2001. Dispersal and parasitism. – In: Clobert, J. et al. (eds), Dispersal. Oxford Univ. Press, pp. 169 – 179.
Bowler, D. E. and Benton, T. G. 2005. Causes and consequences of animal dispersal strategies: relating individual behaviour to spatial dynamics. – Biol. Rev. 80: 205 – 225.
Brown, C. and Brown, M. 1986. Ectoparasitism as a cost of coloniality in cliff swallows ( Hirundo pyrrhonota ). – Ecology 67: 1206 – 1218.
Brown, C. R. and Brown, M. B. 1992. Ectoparasitism as a cause of natal dispersal in cliff swallows. – Ecology 73: 1718 – 1723.
Burnham, K. P. and Anderson, D. R. 1998. Model selection and inference. – Springer.
Bush, A. O. et al. 1997. Parasitology meets ecology on its own terms: Margolis et al. revisited. – J. Parasitol. 83: 575 – 583.
Cameron, P. G. et al. 1993. Eff ects of body size and parasite infection on the locomotory performance of juvenile toads, Bufo bufo . – Oikos 66: 129 – 136.
Clobert, J. et al. 2001. Dispersal. – Oxford Univ. Press. Debeff e, L. et al. 2012. Condition-dependent natal dispersal in a
large herbivore: heavier animals show a greater propensity to disperse and travel further. – J. Anim. Ecol. 81: 1327 – 1337.
Dufty, A. M. and Belthoff , J. R. 2001. Proximate mechanisms of natal dispersal: the role of body condition and hormones. – In: Clobert, J. et al. (eds), Dispersal. Oxford Univ. Press, pp. 2017 – 229.
Supplementary material (available online as Appendix oik.01396 at � www.oikosjournal.org/readers/appendix � ). Appendix 1 (Table A1: Performance of the fi ve candidate generalized mixed linear models of dispersal metrics, with body mass included as a continuous variable. Table A2: Parameter estimates from the best generalized linear mixed model describing variation in dispersal propensity as a func-tion of body mass and nematode abundance, with body mass included as a continuous variable. Figure A1: Infl uence of the two-way interaction between nematode abundance and individual body mass on dispersal propensity with body mass included as a continuous variable).


















