
Paavo V: Komi and Albert Gollhofer - humankinetics.com€¦ · Responses to the Target Article 45 1...
10
Responses to the Target Article 45 1 Ingen Schenau, G.J. van, Bobbert, M.F., & de Haan, A. (1997). Does elastic energy enhance work and efficiency in the stretch-shortening cycle? Journal of Applied Biomechanics, 13, 389- 415. A.L. Hof is with the University of Groningen, Laboratory of Medical Physiology, Bloemsingel 10, NL-9712 KZ Groningen, The Netherlands. Stretch Reflexes Can Have an Important Role in Force Enhancement During SSC Exercise Paavo V: Komi and Albert Gollhofer The stretch-shortening cycle (SSC) of muscle function has a well-recognized purpose: enhancement of performance of the final phase (concentric action) when compared to the isolated concentric action (e.g., Komi, 1984). This can be demonstrated in isolated muscle preparations with constant electrical stimulation (e.g., Cavagna, Dusman, & Margaria, 1968), in animal locomotion with natural and variable muscle activation (e.g., Gregor et al., 1988), and in maximal effort conditions of human SSC actions (Cavagna et al., 1968; Komi, 1983). Ingen Schenau et al. seem to agree with this basic concept. However, in discussing the mechanisms involved in the performance potentiation,the authors use coun- termovement jump (CMJ) as their basic model of SSC. In our understanding, an effective SSC requires three fundamental conditions: a well-timed preactivation of the muscle(s) before the eccentric phase, a short and fast eccentric phase, and immediate transition (short delay) between stretch (eccentric) and shortening (concentric phase). While the CMJ can easily be demonstrated to produce higher jumping height as compared to the squatting jump (SJ) (Asmussen & Bonde-Petersen, 1974; Cavagna, Komarek, Citterio, & Margaria, 1971; Komi & Bosco, 1978), it does not meet well all the criteria for an efficient SSC. The purpose of this commentary is not to try to comment on all the aspects of performance enhancement as discussed in the target article. Instead, we have chosen to focus on the argument that stretch reflexes can have considerable importance in stiffness regulation of the muscle, which would consequently enhance force and power during SSCs. Thus, we do not agree with the target article that stretch reflexes have limited pos- sibilities to operate in SSCs. We will not discuss H-reflex-as mentioned in the target article-because H-reflex is not a naturally occurring phenomenon and has little in com- mon with true stretch reflexes. For many neurophysiological and mechanical aspects, it is evident that the CMJ is not a suitable model to elaborate on the specificity of SSCs. In- stead, one has to look for more "normal" activities such as running and hopping, where the conditions of preactivation, faster stretch, and short transition time are well met. In addition, the forces measured in the muscle tendon complex during these activities can present a typical "bouncing ball" type form (Figure 1). Possibilities for Stretch Reflexes to Operate During the SSC There is no doubt that stretch reflexes play an important role in stiffnessregulation. Hoffer and Andreassen (1981) demonstrated well that when reflexes are intact, muscle stiffness is greater per same operating force than in an areflexive muscle. Thus, stretch reflexes may make a net contribution to muscle stiffness already during the eccentric part of SSC.
Transcript of Paavo V: Komi and Albert Gollhofer - humankinetics.com€¦ · Responses to the Target Article 45 1...

Responses to the Target Article 45 1
Ingen Schenau, G.J. van, Bobbert, M.F., & de Haan, A. (1997). Does elastic energy enhance work and efficiency in the stretch-shortening cycle? Journal of Applied Biomechanics, 13, 389- 415.
A.L. Hof is with the University of Groningen, Laboratory of Medical Physiology, Bloemsingel 10, NL-9712 KZ Groningen, The Netherlands.
Stretch Reflexes Can Have an Important Role in Force Enhancement During SSC Exercise
Paavo V: Komi and Albert Gollhofer
The stretch-shortening cycle (SSC) of muscle function has a well-recognized purpose: enhancement of performance of the final phase (concentric action) when compared to the isolated concentric action (e.g., Komi, 1984). This can be demonstrated in isolated muscle preparations with constant electrical stimulation (e.g., Cavagna, Dusman, & Margaria, 1968), in animal locomotion with natural and variable muscle activation (e.g., Gregor et al., 1988), and in maximal effort conditions of human SSC actions (Cavagna et al., 1968; Komi, 1983). Ingen Schenau et al. seem to agree with this basic concept. However, in discussing the mechanisms involved in the performance potentiation, the authors use coun- termovement jump (CMJ) as their basic model of SSC. In our understanding, an effective SSC requires three fundamental conditions: a well-timed preactivation of the muscle(s) before the eccentric phase, a short and fast eccentric phase, and immediate transition (short delay) between stretch (eccentric) and shortening (concentric phase). While the CMJ can easily be demonstrated to produce higher jumping height as compared to the squatting jump (SJ) (Asmussen & Bonde-Petersen, 1974; Cavagna, Komarek, Citterio, & Margaria, 1971; Komi & Bosco, 1978), it does not meet well all the criteria for an efficient SSC.
The purpose of this commentary is not to try to comment on all the aspects of performance enhancement as discussed in the target article. Instead, we have chosen to focus on the argument that stretch reflexes can have considerable importance in stiffness regulation of the muscle, which would consequently enhance force and power during SSCs. Thus, we do not agree with the target article that stretch reflexes have limited pos- sibilities to operate in SSCs. We will not discuss H-reflex-as mentioned in the target article-because H-reflex is not a naturally occurring phenomenon and has little in com- mon with true stretch reflexes. For many neurophysiological and mechanical aspects, it is evident that the CMJ is not a suitable model to elaborate on the specificity of SSCs. In- stead, one has to look for more "normal" activities such as running and hopping, where the conditions of preactivation, faster stretch, and short transition time are well met. In addition, the forces measured in the muscle tendon complex during these activities can present a typical "bouncing ball" type form (Figure 1).
Possibilities for Stretch Reflexes to Operate During the SSC
There is no doubt that stretch reflexes play an important role in stiffness regulation. Hoffer and Andreassen (1981) demonstrated well that when reflexes are intact, muscle stiffness is greater per same operating force than in an areflexive muscle. Thus, stretch reflexes may make a net contribution to muscle stiffness already during the eccentric part of SSC.

452 Responses to the Target Article
anterior
M. gastrocnemius
M. sdeus
Segment length (A%)
Vertical force
Achilla tendon force
Horizontal force
Figure 1 -A representative record of the Achilles tendon force and segmental length changes of the gastrocnemius and soleus muscles together with Fz and Fy ground reaction forces and selected EMG activities when the subject was running along a long force platform. The vertical line indicates the beginning of the ground contact as well as of the stretch-shortening cycle (SSC) of the muscles. The upward and downward deflections of the segmental length changes signify, respectively, stretching (eccentric) and shortening (concentric) phases of the cycle. From "Stretch-Shortening Cycle" by P.V. Komi, 1992, in P.V. Komi (Ed.), Strength and Power in Sport (p. 171) Oxford, UK: Blackwell Scientific.
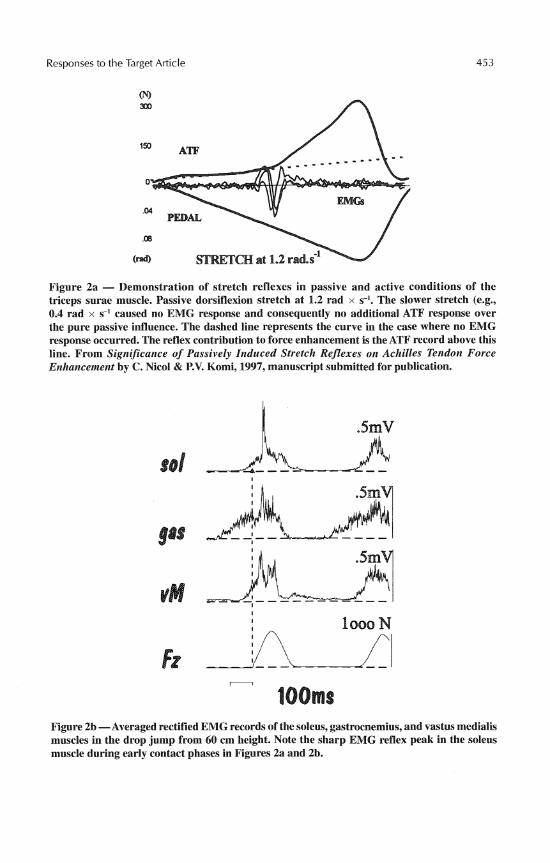
This could be a logical consequence of how muscle spindles and Golgi tendon organs (GTO) operate in the control of muscle length and tension (Houk & Rymer, 1981). It is difficult to imagine that proprioceptive reflexes, the existence of which has been known for centuries, would not play any significant role in human locomotion including SSCs. It is, however, true that in normal movements with high EMG activity, the magnitude and net contribution of reflex regulation of muscle force are methodologically difficult to assess. The task becomes much easier when one studies relatively slow (1.2-1.9 rad x s-I) passive dorsiflexions, where the stretch-induced reflex EMG has been reported to en- hance Achilles tendon force (ATF) by 200-500% over the pure passive stretch without reflex EMG potential (Gollhofer, Komi, Voigt, & Nicol, 1995; Nicol & Komi, 1997; Nicol, Komi, Belli, Huttunen, & Partio, 1995; see also Figure 2a). In these conditions, the time delay between EMG and force enhancement varies between 13 and 15 ms.
In normal hopping and running, even when performed submaximally, the reflex contribution to total EMG can be substantial due to a great number of motor units receiving Ia afferent stimuli from the condition of relatively high stretch veloci- ties (10-12 rad x s-' in the ankle joint). In this regard, hopping and running differ greatly from the CMJ or SJ. While the angular displacement of the ankle joint is clear and fast during hopping, CMJ and SJ may even demonstrate no stretching phase at all (see Figure 3 in Fukashiro, Komi, J h i n e n , & Miyashita, 1993). The situation would not change considerably if one considers the possible consequences of the two-joint muscle behavior.
Responses to the Target Article
ls, ATF
.08
(na) STRETCH at 1.2 ra
Figure 2a - Demonstration of stretch reflexes in passive and active conditions of the triceps surae muscle. Passive dorsiflexion stretch at 1.2 rad x s-I. The slower stretch (e.g., 0.4 rad x s-' caused no EMG response and consequently no additional ATF response over the pure passive influence. The dashed line represents the curve in the case where no EMG response occurred. The reflex contribution to force enhancement is the ATF record above this line. From Significance of Passively Induced Stretch Reflexes on Achilles Tendon Force Enhancement by C. Nicol & P.V. Komi, 1997, manuscript submitted for publication.
Figure 2b -Averaged rectified EMG records of the soleus, gastrocnemius, and vastus medialis muscles in the drop jump from 60 cm height. Note the sharp EMG reflex peak in the soleus muscle during early contact phases in Figures 2a and 2b.

Responses to the Target Article
SOL ENIG
Figure 2c - Rectified and smoothed EMG response from the soleus muscle in short contact time hopping. From Modulation of Short Latency Refexes During Human Hopping by M. Voigt, P. Dyhre-Poulsen, & E.B. Simonsen, 1997, manuscript submitted for publication.
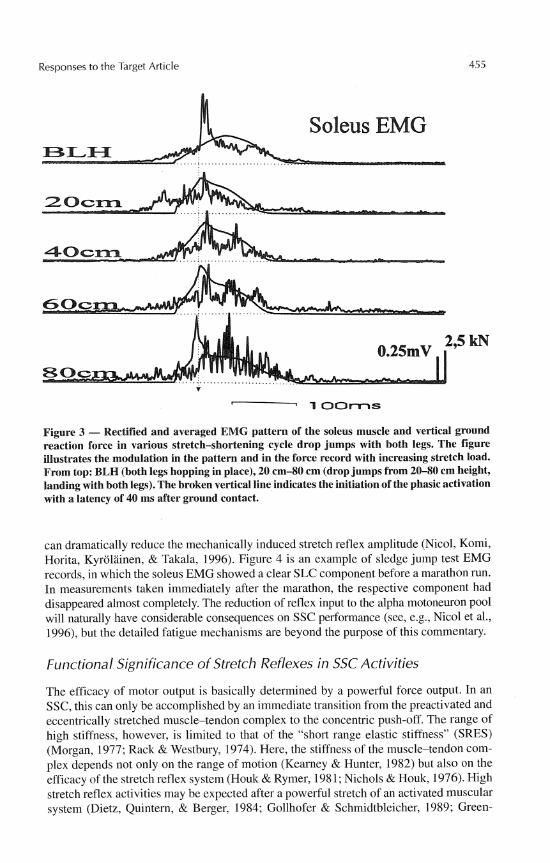
In hopping and running, the short latency stretch reflex component (SLC) can be quite easily observed, especially in the soleus muscle. Figure 2 illustrates records ob- tained in the conditions where this component appears clearly in the EMG patterns when averaged over several trials. Figures 2b (Gollhofer, Strojnik, Rapp, & Schweizer, 1992) and 2c (Voigt, Dyhre-Poulsen, & Simonsen, 1997) are comparable two-leg hops with short contact times. Voigt et al. (1997) measured in these conditions both the origin-to- insertion muscle lengthening and the muscle fiber lengthening. Both measurements showed high stretch velocities in the early contact phase, which led the authors to conclude that the conditions were sufficient for muscle spindle afferent activation. The SLC is sensitive to loading conditions, as shown in Figure 3, where the stretch loads vary from the pre- ferred submaximal hopping (the records on the top) to drop jumps. In the highest drop jump condition (80 cm) the SLC component becomes less clear, suggesting decreased facilitation from the muscle spindles andlor increased inhibitory drive from various sources (e.g., GTO, voluntary protection mechanisms, etc.). In cases where drop jumps have been performed from excessive heights (e.g., from 140 cm, Kyrolainen & Komi, 1995), the subjects have had to sustain extreme powers during contact. In these situations, the re- duced reflex activation may functionally serve as a protection strategy to prevent muscle and/or tendon injury.
It has been shown during passive dorsiflexion tests that the SLC and the medium latency component (MLC) can be dramatically reduced if the measurements are made during an ischemic blockade of the lower limb (e.g., Fellows, Domges, Topper, Thilmann, & Nith, 1993). This method was applied earlier to conditions of fast running (Dietz, Schmidtbleicher, & Noth, 1979), in which the control runs before ischemia demonstrated that the gastrocnemius EMG had a clear SLC component during contact. The average peak EMG was almost three times higher than that measured during a maximal, voluntary, isometric plantar flexion test. During sufficient ischemic blockade, gastrocnemius EMG during contact was dramatically reduced in the fast running test with the same velocity, but there was no change in preactivation. These results emphasize the potential role of Ia afferent input in SSC-type activities such as running. The ischemic blockade is used to isolate the Ia afferent information acting on spinal pathways (Fellows et al., 1993).
Variations in Ia afferent input during SSC have been assessed also in fatigue experi- ments. The moderate SSC fatigue could result in slight potentiation of the SLC compo- nent (Gollhofer, Komi, Fujitsuka, & Miyashita, 1987), but an exhaustive SSC exercise
Responses to the Target Article
Soleus EMG
1 OOms
Figure 3 - Rectified and averaged EMG pattern of the soleus muscle and vertical ground reaction force in various stretch-shortening cycle drop jumps with both legs. The figure illustrates the modulation in the pattern and in the force record with increasing stretch load. From top: BLH (both legs hopping in place), 20 cm-80 cm (drop jumps from 20-80 cm height, landing with both legs). The broken vertical line indicates the initiation of the phasic activation with a latency of 40 ms after ground contact.
can dramatically reduce the mechanically induced stretch reflex amplitude (Nicol, Komi, Horita, Kyrolainen, & Takala, 1996). Figure 4 is an example of sledge jump test EMG records, in which the soleus EMG showed a clear SLC component before a marathon run. In measurements taken immediately after the marathon, the respective component had disappeared almost completely. The reduction of reflex input to the alpha motoneuron pool will naturally have considerable consequences on SSC performance (see, e.g., Nicol et al., 1996), but the detailed fatigue mechanisms are beyond the purpose of this commentary.
Functional Significance of Stretch Reflexes in SSC Activities
The efficacy of motor output is basically determined by a powerful force output. In an SSC, this can only be accomplished by an immediate transition from the preactivated and eccentrically stretched muscle-tendon complex to the concentric push-off. The range of high stiffness, however, is limited to that of the "short range elastic stiffness" (SRES) (Morgan, 1977; Rack & Westbury, 1974). Here, the stiffness of the muscle-tendon com- plex depends not only on the range of motion (Kearney & Hunter, 1982) but also on the efficacy of the stretch reflex system (Houk & Rymer, 1981; Nichols & Houk, 1976). High stretch reflex activities may be expected after a powerful stretch of an activated muscular system (Dietz, Quintern, & Berger, 1984; Gollhofer & Schmidtbleicher, 1989; Green-

sa8ueq3 uo!sual pw ql8ual aq 01 anqsuas h[q%!q s! ley1 maids loida3a~ e WOJJ h[an!i3aj -ja )sour paprno~d aq ue3 uo!~aap3e nln3snur pa~a88ui pug luapuadap aseqd q8!~
.uog~np3e mln3snur asualu! hq paluana~d aq ue3 ST
Responses to the Target Article 457
in the muscle-tendon system. For several reasons, the stretch reflex system seems to be the most appropriate candidate to generate this intense phasic muscular activation: First, the latencies of the EMG peaks in the stretched muscles are 40-45 ms after ground contact (see also Figure 5). Depending on the muscle and on the subject's size, these latencies match very well with the stretch responses observable after mechanical stimulation (Gollhofer & Rapp, 1993; Nicol & Komi, 1997). Second, instantaneous force-length (Komi, 1992) and EMG-length (Gollhofer et al., 1992) diagrams show that 45 ms after contact, the triceps surae muscle is in the isometric state, which usually lasts 80 ms or less. The latencies as well as the duration fit well with the occurrence of the SLC and MLC (Dietz, 1987; Lee & Tatton, 1982; Marsden, Merton, & Morton, 1976). Third, according to Stein (1982) and Nichols (1987), it is the stretch reflex system that provides high linearity in muscular stiffness. And fourth, the above-mentioned aspects may be partly responsible for the observations that mechanical efficiency in natural SSC exercise is higher than that in pure concentric exercise (e.g., Aura & Komi, 1986; Kyrolainen et al., 1990). Especially the concept of elastic storage favors the existence of reflex activation. Thus, it is evident that high muscular activation during the eccentric phase of SSC is a prerequisite for effi- cient storage of elastic energy. Functionally, the muscle provides tension in the tendon- muscle complex, and the tendon attenuates length changes.
In drop jumps with high stretch loads, neuromuscular inhibition is often observed prior to reflex activation (Figure 4). Prochazka, Schofield, Westerman, and Ziccone (1977) observed similar reflex behavior in cats. The authors discussed these reductions as a strat- egy of the neuromuscular system to reset the motoneurons in order to produce a triggered and synchronized activation of all units, simultaneously with the ground contact (i.e., with the stretching of the extensor muscles).
An electrically stimulated muscle responds to ramp stretches with linear tension increments only if the muscle has an intact reflex system (Nichols, 1987; Nichols & Houk,
Figure 5 - Rectified and averaged EMG pattern of m. gastrocnemius and vertical ground reaction force of a drop jump from 72 cm height. The EMG profile shows a clear reduction in amplitude prior to contact, which is consistent until the middle phase of contact. The phase of reduction is dispersed by a phasic EMG contribution (see the vertical arrow) occurring 43 ms after the ground contact.

458 Responses to the Target Article
1976). This linearity is restricted to small length changes (e.g., Hoffer & Andreassen, 1981; Joyce et al., 1969; Noth, 1985). The question then arises whether these small length changes are relevant for drop jump performances. Various length changes are reported in the literature demonstrating that effective range of SRES in in vitro preparations is 1 to 4% (e.g., Ford et a]., 1978; Huxley & Simmons, 1971). In the intact muscle tendon com- plex in vivo, this value is increased because series elasticity and fiber geometry must be taken into account. This would bring the muscle-tendon lengthening to 6 to 8%. As dem- onstrated by Komi (1992) and Gollhofer e t al. (1992), length changes of the triceps surae in running and in drop jumps are between 6 and 9% during contact. When the measure- ments are made on muscle fiber level, the values are naturally smaller, as shown by Rob- erts, Marsch, Weyand, and Taylor (1997) in turkeys running on level ground.
Thus, there seems to be sufficient support for the argument that the stretch reflex contributes significantly to force generation during the touchdown in SSC activities such as hopping and running. Depending on the types of hopping, for example, the amplitude of the SLC reflex peak may vary considerably. Additionally, the combination of the "prereflex" background activation and the following reflex activation might represent a scenario that supports yield compensation and fast rate of force development (Voigt et al., 1997).
References
Asmussen, E., & Bonde-Petersen, F. (1974). Apparent efficiency and storage of elastic energy in human muscles during exercise. Acta Physiologica Scandinavica, 92,537-545.
Aura, O., & Komi, P.V. (1986). The mechanical efficiency of locomotion in men and women with special emphasis on stretch-shortening cycle exercises. European Journal ofApplied Physi- ology, 55,3743.
Avela, J., & Komi, P.V. (1997). Interaction between muscle stifiess and stretch reflex sensitivity after marathon running. Manuscript submitted for publication.
Cavagna, G.A., Dusman, B., & Margaria, R. (1968). Positive work done by a previously stretched muscle. Journal of Applied Physiology, 24,21-32.
Cavagna, G.A., Komarek, L., Citterio, G., & Margaria, R. (1971). Power output of the previously stretched muscle. In J. Vredenbregt & J. Wartenweiler (Eds.), Medicine and sport, Vol 6: Biomechanics I1 (pp. 159-167). Basel, Switzerland: Karger.
Dietz, V. (1987). Role of peripheral afferents and spinal reflexes in normal and impaired human locomotion. Revue Neurologique, 4,241-254.
Dietz, V., Quintern, J., & Berger,.W. (1984). Corrective reactions to stumbling in man. Functional significance of spinal and transcortical reflexes. Neuroscience Letters, 44, 13 1-135.
Dietz, V., Schmidtbleicher, D., & Noth, J. (1979). Neuronal mechanisms of human locomotion. Journal of Neurophysiology, 42,1212-1222.
Fellows, S., Domges, F., Topper, R., Thilmann, A., & Noth. J. (1993). Changes in the short and long latency stretch reflex components of the triceps surae muscle during ischaemia in man. Jour- nal of Physiology, 472,737-748.
Flitney, F.W., & Hirst, D.G. (1978). Cross-bridge detachment and sarcomere "give" during stretch of active frog's muscle. Journal of Physiology, 276,449-465.
Ford, L.E., Huxley, A.F., & Simmons, R.M. (1978). Tension responses to sudden length change in stimulated frog muscle fibres near slack length. Journal of Physiology, 269,441-515.
Fukashiro, S., Komi, P.V., T i inen , M., & Miyashita, M. (1993). Comparison between the directly measured Achilles tendon force and the tendon force calculated from the joint ankle move- ment during vertical jumps. Journal of Clinical Biomechanics, 8,25-30.
Gollhofer, A., Komi, P.V., Fujitsuka, N., & Miyashita, M. (1987). Fatigue during stretch-shortening cycle exercises: 11. Changes in neuromuscular activation patterns of human skeletal muscle. International Journal of Sports Medicine, 8 ,3841 .

Responses to the Target Article
Gollhofer, A., Komi, P.V., Voigt, M., & Nicol, C. (1995). Mechanical contribution of isolated stretch evoked EMG responses. In K. Hakkinen, K.L. Keskinen, P.V. Komi, &A. Mero (Eds.), Book of abstractsfrom the XVth International Congress of Biomechanics, University of Jyvtiskyla, Finland (pp. 330-331).
Gollhofer, A., & Rapp, W. (1993). Recovery of stretch reflex responses following mechanical stimu- lation. European Journal of Applied Physiology, 66,415-420.
Gollhofer, A., & Schrnidtbleicher, D. (1989). Stretch reflex responses of the human m. triceps surae following mechanical stimulation. J o u m l of Biomechanics, 22, 1016.
Gollhofer, A., Strojnik, V., Rapp, W., & Schweizer, L. (1992). Behavior of triceps surae muscle- tendon complex in different jump conditions. European Journal of Applied Physiology, 64, 283-29 1.
Greenwood, R., & Hopkins, A. (1976). Landing from an unexpected fall and a voluntary step. Brain Research, 99,375-386.
Greenwood, R., & Hopkins, A. (1977). Muscle responses during sudden falls in man. Journal of Physiology, 254,507-5 18.
Gregor, R.J., Roy, R.R., Whiting, W.C., Lovely, R.G., Hodgson, J.A., & Edgerton, V.R. (1988). Mechanical output of the cat soleus during treadmill locomotion: In vivo vs. in situ character- istics. Journal of Biomechanics, 21(9), 721-732.
Hoffer, J.A., & Andreassen, S. (1981). Regulation of soleus muscle stiffness in premamillary cats. Intrinsic and reflex components. Journal of Neurophysiology, 45,267-285.
Houk, J.C., & Rymer, W.Z. (1981). Neural control of muscle length and tension. In V.B. Brooks (Ed.), Handbook ofphysiology. The nervous system (pp. 257-323). Baltimore: Waverly Press.
Hufschmidt, A., & Schwaller, I. (1987). Short-range elasticity and resting tension of relaxed human lower leg muscles. Journal of Physiology, 393,451-465.
Huxley, A.F., & Simmons, R.M. (1971). Proposed mechanism of force generation in striated muscle. Nature, 233,533-538.
Ingen Schenau, G.J. van, Bobbert, M.F., & Haan, A. de. (1997). Does elastic energy enhance work and efficiency in the stretch-shortening cycle? Journal of Applied Biomechanics, 13,389-415.
Joyce, G.C., Rack, P.M.H., & Westbury, D.R. (1969). The mechanical properties of cat soleus muscle during controlled lengthening and shortening movements. Journal of Physiology, 204, 461- 474.
Kearney, R.E., & Hunter, I.W. (1982). Dynamics of human ankle stiffness. Variation with displace- ment amplitude. Journal of Biomechanics, 15,753-756.
Komi, P.V. (1983). Elastic potentiation of muscles and its influence on sport performance. In W. Baumann (Ed.), Biomechanik und sportliche Leistung (pp. 59-708). Schomdorf, Germany: Verlag Karl Hofrnann.
Komi, P.V. (1984). Physiological and biomechanical correlates of muscle function: Effects of muscle structure and stretch-shortening cycle on force and speed. Exercise and Sport Sciences Re- views, 12, 81-121.
Komi, P.V. (1992) Stretch-shortening cycle. In P.V. Komi (Ed.), Strength and power in sport (pp. 169-179). Oxford, UK: Blackwell Scientific.
Komi, P.V., & Bosco, C. (1978). Utilization of stored elastic energy in leg extensor muscles by men and women. Medicine and Science in Sports, 10(4), 261-265.
Kyrolilinen, H., & Komi, P.V. (1995). Differences in mechanical efficiency in athletes during jump- ing. European Journal ofApplied Physiology, 70,36-44.
Kyrolainen, H., Komi, P.V., Oksanen, P., Hakkinen, K., Cheng, S., &Kim, D.H. (1990). Mechanical efficiency of locomotion in females during different kinds of muscle actions. European Jour- nal of Applied Physiology, 61,446-452.
Lee, R.G., & Tatton, W.G. (1982). Long latency reflexes to imposed displacements of the human wrist. Dependence on duration of movement. Experimental Brain Research, 45,207-216.
Marsden, C.D., Merton, P.A., & Morton, H.B. (1976). Stretch reflex and servo action in a variety of human muscles. Journal of Physiology, 259,531-560.
Melvill Jones, G., &Watt, G.D. (1971). Muscular control of landing from unexpected falls in man. Journal of Physiology, 219,729-737.

460 Responses to the Target Article
Morgan, D.L. (1977). Separation of active and passive components of short-range stiffness of muscle. American Journal of Physical Medicine, 232,4549.
Nicol, C., & Komi, P.V. (1997). Significance ofpassively induced stretch reflexes on Achilles tendon force enhancement. Manuscript submitted for publication.
Nicol, C., Komi, P.V., Belli, A., Huttunen, V., & Partio, E. (1995). Reflex contribution of Achilles tendon forces: In-vivo measurements with the optic fibre technique. In K. Htikkinen, K.L. Keskinen, P.V. Komi, &A. Mero (Eds.), Book of abstracts from the XVth International Con- gress of Biomechanics, University of Jyvaskylii, Finland (pp. 674-675).
Nicol, C., Komi, P.V., Horita, T., Kyrolainen, H., & Takala, T.E.S. (1996). Reduced stretch-reflex sensitivity after exhausting stretch-shortening cycle (SSC) exercise. European Journal of Applied Physiology, 72,401-409.
Nichols, T.R. (1974). Soleus muscle stiffness and its reflex control. Unpublished doctoral disserta- tion, Harvard University, Cambridge, MA.
Nichols, T.R. (1987). The regulation of muscle stiffness. Medicine and Science in Sports and Exer- cise, 26,3647.
Nichols, T.R., & Houk, J.C. (1976). Improvement in linearity and regulation of stiffness that results from actions of stretch reflex. Journal of Neurophysiology, 39, 119-142.
Noth, J. (1985). Neurophysiologische Aspekte der Muskelelastizitat [Neurophysiological aspects of muscle elasticity]. Schrijireihe des Bundesinstitutsfur Sportwissenschaft, 56,238-253.
Prochazka, A., Schofield, P., Westerman, R.A., & Ziccone, S.P. (1 977). Reflexes in cat ankle muscles after landing from falls. Journal of Physiology, 272,705-719.
Rack, P.M.H., & Westbury, D.R. (1974). The short range stiffness of active mammalian muscle and its effect on mechanical properties. Journal of Physiology, 240, 331-350.
Roberts, T.J., Marsch, R.L., Weyand, P.G., & Taylor, C.R. (1997). Muscular force in running tur- keys: The economy of minimizing work. Science, 275, 11 13-1 115.
Stein, R.B. (1982). What muscle variable(s) does the nervous system control in limb movements? Behavioral and Brain Sciences, 5,535-577.
Vallbo, A.B. (1981). Basic patterns of muscle spindle discharge in man. In A. Taylor &A. Prochazka (Eds.), Muscle receptors and movement (pp. 263-275). London: Macmillan..
Voigt, M., Dyhre-Poulsen, P., & Simonsen, E.B. (1997). Modulation of short latency reflexes during human hopping. Manuscript submitted for publication.
Paavo K Komi is with the Department of Biology of Physical Activity, University of Jyvaskyla, PO. Box 35, 40351 Jyvaskyla, Finland. Albert Gollhofer is with the Department of Sport Science, University of Stuttgart, Stuttgart, Germany.
Are Efficiency and the Cost of Generating Force Both Relevant Concepts?
Rodger Kram
I focus my response to the stimulating review by Ingen Schenau, Bobbert, and Haan (1997) on the section in Part I1 entitled "Is Efficiency in SSCs a Relevant Concept?My response is organized into three parts: a refutation of some of the arguments presented, a synopsis of recent evidence that is highly relevant, and my perspective on the direction that future research in this area should pursue.
Ingen Schenau et al. state that "it would be hard to explain the high requirement of metabolic energy if running were predominantly driven by isometrically contracting muscles" (p. 403). Their statement is based on a comparison of the metabolic costs of cycling versus running that I believe is misguided. It is well established that forces are


















