
Oxygen Concentration Within the Nitrogen-Fixing Root ... · lated to oxygen concentration in the...
6
Oxygen Concentration Within the Nitrogen-Fixing Root Nodules of Myrica gale L. Author(s): John D. Tjepkema Source: American Journal of Botany, Vol. 70, No. 1 (Jan., 1983), pp. 59-63 Published by: Botanical Society of America Stable URL: http://www.jstor.org/stable/2443204 . Accessed: 23/08/2011 16:00 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. Botanical Society of America is collaborating with JSTOR to digitize, preserve and extend access to American Journal of Botany. http://www.jstor.org
Transcript of Oxygen Concentration Within the Nitrogen-Fixing Root ... · lated to oxygen concentration in the...

Oxygen Concentration Within the Nitrogen-Fixing Root Nodules of Myrica gale L.Author(s): John D. TjepkemaSource: American Journal of Botany, Vol. 70, No. 1 (Jan., 1983), pp. 59-63Published by: Botanical Society of AmericaStable URL: http://www.jstor.org/stable/2443204 .Accessed: 23/08/2011 16:00
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
Botanical Society of America is collaborating with JSTOR to digitize, preserve and extend access to AmericanJournal of Botany.
http://www.jstor.org

Amer. J. Bot. 70(1): 59-63. 1983.
OXYGEN CONCENTRATION WITHIN THE NITROGEN-FIXING ROOT NODULES OF MYRICA GALE L.i
JOHN D. TJEPKEMA2
Harvard Forest, Harvard University, Petersham, Massachusetts 01366
ABSTRACT
Microelectrodes were used to study the oxygen concentration within Myrica gale L. nodules. Low oxygen concentrations were found only in the region of the mature, nitrogen-fixing en- dophyte, and appeared to correspond to clusters of infected host cells. The oxygen concentration in the remainder of the nodule was much higher. Interconnected intercellular air spaces were demonstrated by infiltration with India ink. Infiltration of the spaces with water greatly reduced oxygen concentration throughout the nodule, indicating that they function in supplying oxygen to the infected cells and remainder of the nodule. These results differ from those found previously for soybean nodules and provide evidence that legume and actinorhizal nodules have different mechanisms for protecting nitrogenase from oxygen.
PURIFIED NITROGENASE is rapidly denatured when exposed to oxygen, and thus requires protection from oxygen when present in or- ganisms living in an aerobic environment. In the Rhizobium-legume symbiosis, there is evi- dence that the inner layer of the nodule cortex lacks intercellular air spaces and thus restricts diffusion of oxygen to the bacteroid-containing tissue in the center of the nodule (Tjepkema and Yocum, 1974). Measurements with an oxygen microelectrode show a sharp decrease in oxygen concentration at the nodule cortex and a low value in the nodule center. Thus, the nodule structure helps to provide the low oxygen concentration required for nitrogen fix- ation by the rhizobia.
Little is known about oxygen relations in actinorhizal (actinomycete-induced) root nod- ules. In Alnus nodules, lenticels allow oxygen to enter the nodules freely, and well-developed intercellular air spaces connect the lenticels di- rectly to the cells containing the actinomycetal endophyte (Wheeler, Gordon and Ching, 1979; Tjepkema, 1979). Thus, there is no cortical barrier to oxygen as in legumes and the Po2 in the air spaces adjacent to the infected cells is probably close to atmospheric Po2. However, oyxgen concentration within the nodule has not been measured.
I Received for publication 25 September 198 1; revision accepted 21 January 1982.
I thank C. S. Yocum for the oxygen microelectrodes and A. M. Berry and C. R. Schwintzer for the section of resin- embedded nodule. This study was supported by NSF Grant DEB 77-02249, USDA Grant 78-59-2252-0-1-055-1, and the Maria Moors Cabot Foundation of Harvard Univer- sity.
Present address: Department of Botany and Plant Pa- thology, University of Maine, Orono, Maine 04469.
In the present study, another actinorhizal plant, Myrica gale, was examined. Myrica gale fixes nitrogen actively (Schwintzer, 1979) and the literature concerning its root nodules has been recently summarized (Schwintzer, Berry and Disney, 1982). The internal oxygen con- centration was measured with oxygen elec- trodes, and the distribution of intercellular air spaces was traced with India ink and examined in thin, resin-embedded sections. Both high and low oxygen concentrations were found in the region of endophyte-containing cells. The low oxygen concentrations apparently corre- spond to clusters of infected cells. As in Alnus nodules, there was no cortical barrier to oxygen diffusion.
MATERIALS AND METHODS-Seeds of Myrica gale L. were germinated in sand after cold treat- ment. The seedlings were transferred to aero- ponics tanks (Zobel, Del Tredici and Torrey, 1976) containing nitrogen-free nutrient solu- tion and then inoculated with suspensions of ground nodules. The studies were made 3 months after nodule initiation.
Nodules were vacuum infiltrated with India ink by placing them under vacuum for 5 min before immersion in ink followed by release of the vacuum. Prior to infiltration, the larger particles in the ink (Higgins India ink, Faber Castell Corp.) were removed by centrifugation at approximately 12,000 g for 30 min.
Thin sections were obtained from nodule lobes fixed in 3% glutaraldehyde and embed- ded in Spurr's resin as described by Schwintzer, Berry and Disney (1982). Sections (1 to 2 ,um) were stained in 0.9% aqueous KMnO4.
The acetylene reduction assay was used to measure nitrogenase activity in nodule seg-
59
60 AMERICAN JOURNAL OF BOTANY [Vol. 70
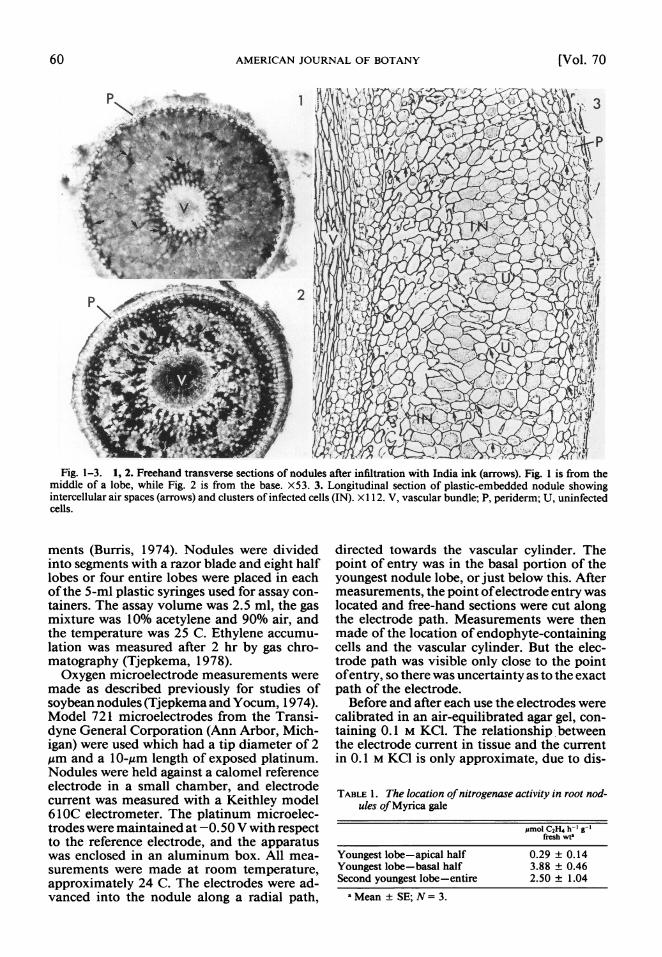
Fig. 1-3. 1, 2. Freehand transverse sections of nodules after infiltration with India ink (arrows). Fig. 1 is from the middle of a lobe, while Fig. 2 is from the base. X53. 3. Longitudinal section of plastic-embedded nodule showing intercellular air spaces (arrows) and clusters of infected cells (IN). X 1 12. V, vascular bundle; P, periderm; U, uninfected cells.
ments (Burris, 1974). Nodules were divided into segments with a razor blade and eight half lobes or four entire lobes were placed in each of the 5-ml plastic syringes used for assay con- tainers. The assay volume was 2.5 ml, the gas mixture was 10% acetylene and 90% air, and the temperature was 25 C. Ethylene accumu- lation was measured after 2 hr by gas chro- matography (Tjepkema, 1978).
Oxygen microelectrode measurements were made as described previously for studies of soybean nodules (Tjepkema and Yocum, 1974). Model 721 microelectrodes from the Transi- dyne General Corporation (Ann Arbor, Mich- igan) were used which had a tip diameter of 2 ,um and a 10-,gm length of exposed platinum. Nodules were held against a calomel reference electrode in a small chamber, and electrode current was measured with a Keithley model 61 OC electrometer. The platinum microelec- trodes were maintained at -0. 50 V with respect to the reference electrode, and the apparatus was enclosed in an aluminum box. All mea- surements were made at room temperature, approximately 24 C. The electrodes were ad- vanced into the nodule along a radial path,
directed towards the vascular cylinder. The point of entry was in the basal portion of the youngest nodule lobe, or just below this. After measurements, the point of electrode entry was located and free-hand sections were cut along the electrode path. Measurements were then made of the location of endophyte-containing cells and the vascular cylinder. But the elec- trode path was visible only close to the point of entry, so there was uncertainty as to the exact path of the electrode.
Before and after each use the electrodes were calibrated in an air-equilibrated agar gel, con- taining 0.1 M KCI. The relationship between the electrode current in tissue and the current in 0.1 M KCI is only approximate, due to dis-
TABLE 1. The location of nitrogenase activity in root nod- ules of Myrica gale
IsmoI C2H4 h-' g-I fresh wt'
Youngest lobe-apical half 0.29 ? 0.14 Youngest lobe-basal half 3.88 ? 0.46 Second youngest lobe-entire 2.50 ? 1.04
aMean ? SE; N= 3.
January, 1983] TJEPKEMA-OXYGEN IN MYRICA NODULES 61
100 ~ ~~~45
100
H 10 |
0
O ~~~~INF ~-V-- F*INF1 IN F Y V s *IN l 10 61 7
10 1Ad0>
0.11 . * , . * _* 0 0.2 0.4 0.6 0.8 0 0.2 0.4 0.6 0.8 1.0
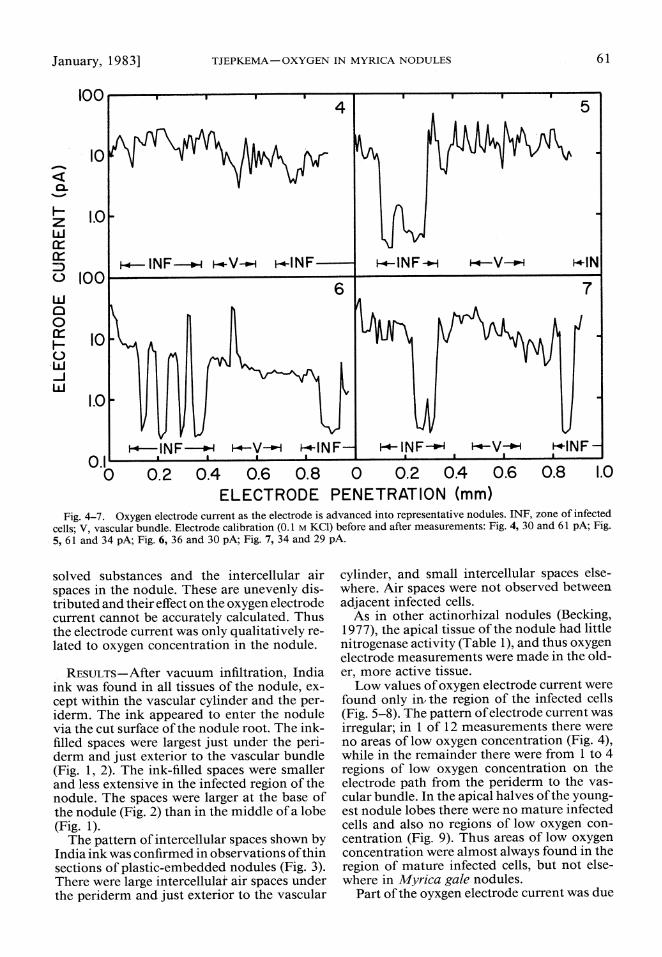
ELECTRODE PENETRATION (mm) Fig. 4-7. Oxygen electrode current as the electrode is advanced into representative nodules. INF, zone of infected
cells; V, vascular bundle. Electrode calibration (0.1 M KCI) before and after measurements: Fig. 4, 30 and 61 pA; Fig. 5, 61 and 34 pA; Fig. 6, 36 and 30 pA; Fig. 7, 34 and 29 pA.
solved substances and the intercellular air spaces in the nodule. These are unevenly dis- tributed and their effect on the oxygen electrode current cannot be accurately calculated. Thus the electrode current was only qualitatively re- lated to oxygen concentration in the nodule.
RESULTS-After vacuum infiltration, India ink was found in all tissues of the nodule, ex- cept within the vascular cylinder and the per- iderm. The ink appeared to enter the nodule via the cut surface of the nodule root. The ink- filled spaces were largest just under the peri- derm and just exterior to the vascular bundle (Fig. 1, 2). The ink-filled spaces were smaller and less extensive in the infected region of the nodule. The spaces were larger at the base of the nodule (Fig. 2) than in the middle of a lobe (Fig. 1).
The pattern of intercellular spaces shown by India ink was confirmed in observations of thin sections of plastic-embedded nodules (Fig. 3). There were large intercellulat air spaces under the periderm and just exterior to the vascular
cylinder, and small intercellular spaces else- where. Air spaces were not observed between adjacent infected cells.
As in other actinorhizal nodules (Becking, 1977), the apical tissue of the nodule had little nitrogenase activity (Table 1), and thus oxygen electrode measurements were made in the old- er, more active tissue.
Low values of oxygen electrode current were found only in, the region of the infected cells (Fig. 5-8). The pattern of electrode current was irregular; in 1 of 12 measurements there were no areas of low oxygen concentration (Fig. 4), while in the remainder there were from 1 to 4 regions of low oxygen concentration on the electrode path from the periderm to the vas- cular bundle. In the apical halves of the young- est nodule lobes there were no mature infected cells and also no regions of low oxygen con- centration (Fig. 9). Thus areas of low oxygen concentration were almost always found in the region of mature infected cells, but not else- where in Myrica gale nodules.
Part of the oyxgen electrode current was due

62 AMERICAN JOURNAL OF BOTANY [Vol. 70
100
t?? 1.0 21.A
H
w
0
0 10
*0 0.2 0.4 0.6 0.8
ELECTRODE PENETRATION (mm)
Fig. 8. Oxygen electrode current within a nodule in atmospheres of air and then argon. INF, zone of infected cells; V, vascular bundle. Electrode calibration (0. I M KC1: 46 pA before measurements, 31 pA afterwards.
to reduction of substances other than oxygen. When measurements were made with argon flowing through the electrode chamber, the electrode current was about 100 times lower and was probably due to reduction of sub- stances other than oxygen (Fig. 8). The higher electrode current near the nodule exterior dur- ing the measurements in argon was probably due to failure to completely exclude oxygen from the electrode chamber.
Since oxygen diffuses about 300,000 times faster through air than water, it mostly diffuses
100
,,0 02 0.4E 0. 0.
z 1.0V
0.
ELECTRODE PENETRATION (mm)
Fig. 8. Oxygen electrode current wi thin alenodue in
adatoperes of ai an.dIN zone of infcted
cel;V aclrbundle. Electrode calibration (0.1 M KC1)L3)A:efr 46p eoemeasurements, 31 pA afterwards.
tordcinoCsbtne terta xgn Whe esrmnswr aewt ro
ei.9 xye lectrode current neastetndue exeteriore dur idvng eit the me iasurementsoi argonueloe was probablyr
fromdte. Electrode chlbamber(. ImK :3 Abfr
feastreernthroughp airthanwatr,dtmotysifue
100
10 w llJ \, ~ INF V
w 0 0
H 1.0 e w w
0.1 . 0 0.2 0.4 0.6 0.8
ELECTRODE PENETRATION (mm)
Fig. 10. Oxygen electrode current within a nodule after infiltration of the intercellular air spaces with water. INF, zone of infected cells, V, vascular bundle. Electrode cali- bration: 23 pA before measurements, 34 pA afterwards.
to the nodule interior via the intercellular air spaces. When these spaces were infiltrated with water, the oxygen concentration in the nodule interior was much lower (Fig. 10).
DISCUSSION-The results show that low oxy- gen concentrations occur only in the region of the mature endophyte in root nodules of My- rica gale. Within this region, areas of both high and low oxygen concentrations were found (Fig. 4-8). The regions of low oxygen concentration varied from about 40 Am to 200 Am in width. They probably corresponded to the locations of individual infected cells or to several ad- jacent infected cells, since the oxygen electrode has a spatial resolution of approximately 30 Am (Tjepkema and Yocum, 1974), and the in- fected cells varied in diameter from about 30 to 40 gim. Uninfected cells were probably not responsible for the areas of low oxygen con- centration since such cells were highly vacu- olate (Fig. 3) and thus not capable of high rates of oxygen uptake. Furthermore, low electrode currents were not observed in the apical halves of nodule lobes, where there were only unin- fected cells or immature infected cells (Fig. 9).
This pattern of oxygen concentration is con- sistent with the distribution of intercellular air spaces in Myrica gale nodules (Fig. 1-3). In- terconnected, ink-filled spaces were observed in all regions of the nodule except within the vascular cylinder. Air spaces provide a low resistance to oxygen diffusion, and one would thus expect cells connected by them to be at similar oxygen concentrations, as was ob- served. Intercellular spaces were lacking be-

January, 1983] TJEPKEMA-OXYGEN IN MYRICA NODULES 63
tween adjacent infected cells (Fig. 3), thus in- creasing the resistance to oxygen diffusion into these cells. Combined with a high rate of res- piration by the infected cells, this would ex- plain the observed regions of low oxygen con- centration.
The intercellular air spaces of the nodule are connected to those of the nodule root, which provides a large surface area for the absorption of oxygen (Tjepkema, 1978; Torrey and Cal- laham, 1978). The resistance to oxygen dif- fusion through the nodule root epidermis and nodule periderm appears to be relatively small (Tjepkema, 1978), so that the Po2 in the in- tercellular air spaces is probably not substan- tially below that at the nodule exterior. This conclusion is supported by the observation of oxygen electrode currents in the nodule center that are almost as large as those observed at the nodule surface or in air-equilibrated agar.
These results are very different from those observed previously for the root nodules of soybeans, a legume. Using the same type of electrode, very low oxygen concentrations were observed throughout the endophyte-contain- ing region of soybean nodules, even though uninfected cells were present (Tjepkema and Yocum, 1974). It was concluded that there is a cell layer in the inner nodule cortex of soy- bean nodules that restricts oxygen diffusion into the region of the endophyte. No such layer is present in the nodules of Myrica gale, and thus high oxygen concentrations as well as low oxygen concentrations occur in the infected region. This suggests that protection of nitrog- enase occurs within the infected cells of Myrica gale, rather than relying on a diffusion barrier that entirely surrounds the infected cells, as is the case in soybean nodules.
There are two ways in which nitrogenase in Myrica gale nodules may be protected from oxygen. Figures 5-8 demonstrate that regions of low oxygen concentration exist in nodules and one possibility is that nitrogenase activity is localized in these regions. The second pos- sibility is that the vesicles of the endophyte are analogous to the heterocysts of blue-green algae in providing protection of nitrogenase against oxygen inactivation. The vesicles are believed to be the site of nitrogen fixation and our results demonstrate that this is also true for cultures of the endophyte, Frankia (Tjepkema, Or- merod and Torrey, 1980, 1981). Nitrogen fix- ation by cultures occurs only when vesicles are present and maximum rates of nitrogen fixa- tion occur at atmospheric Po2 (20 kPa). Thus the endophyte itself is able to protect nitrog-
enase from oxygen and does not require a nod- ule structure that provides an environment of low oxygen concentration.
We conclude that the mechanisms for the protection of nitrogenase from oxygen are fun- damentally different in legume and actino- rhizal nodules. In legume nodules, nitrogen fix- ation by Rhizobium can take place only at low oxygen concentrations, and the nodule struc- ture provides such a low oxygen environment. In actinorhizal nodules, nitrogen fixation by Frankia can occur at atmospheric Po2 and no barrier to oxygen diffusion into the nodule is required.
LITERATURE CITED
BECKING, J. H. 1977. Endophyte and association estab- lishment in non-leguminous nitrogen-fixing plants. In W. Newton, J. R. Postgate, and C. Rodriguez-Bar- rueco [eds.], Recent developments in nitrogen fixa- tion, pp. 551-567. Academic Press, New York.
BURRIS, R. H. 1974. Methodology. In A. Quispel [ed.], The biology of nitrogen fixation, pp. 9-33. American Elsevier, New York.
SCHWINTZER, C. R. 1979. Nitrogen fixation by Myrica gale root nodules in a Massachusetts wetland. Oeco- logia 43: 283-294.
5 A. M. BERRY, AND L. D. DISNEY. 1982. Seasonal patterns of root nodule growth, endophyte morphol- ogy, nitrogenase activity, and shoot development in Myrica gale L. Can. J. Bot. 60: 746-757.
TJEPKEMA, J. 1978. The role of oxygen diffusion from the shoots and nodule roots in nitrogen fixation by root nodules of Myrica gale. Can. J. Bot. 56: 1365- 1371.
1979. Oxygen relations in leguminous and ac- tinorhizal nodules. In J. C. Gordon, C. T. Wheeler, and D. A. Perry [eds.], Symbiotic nitrogen fixation in the management of temperate forests, pp. 175-186. Forest Research Laboratory, Oregon State University, Corvallis.
, AND C. S. YOCUM. 1974. Measurement of oxy- gen partial pressure within soybeani nodules by oxygen microelectrodes. Planta 119: 351-360.
, W. ORMEROD, AND J. G. TORREY. 1980. Vesicle formation and acetylene reduction activity in Frankia sp. CPI1 cultured in defined nutrient media. Nature (London) 287: 633-635.
,AND . 1981. Factors affecting vesicle formation and acetylene reduction (nitrogen- ase activity) in Frankia sp. CPI 1. Can. J. Microbiol. 27: 815-823.
TORREY, J. G., AND D. CALLAHAM. 1978. Determinate development ofnodule roots in actinomycete-induced root nodules of Myrica gale. Can. J. Bot. 56: 1357- 1364.
WHEELER, C. T., J. C. GORDON, AND T. M. CHING. 1979. Oxygen relations of the root nodules of Alnus rubra Bong. New Phytol. 82: 449-457.
ZOBEL, R. W., P. DEL TREDICI, AND J. G. TORREY. 1976. Method for growing plants aeroponically. Plant Phys- iol. 57: 344-346.


















