
OUT OF THE QUAGMIRE OF PLANT DEFENSE HYPOTHESES · Secondary metabolites are found mostly in...
33
Volume 78, No. 1 March 2003 THE QUARTERLY REVIEW OF BIOLOGY 23 The Quarterly Review of Biology, March 2003, Vol. 78, No. 1 Copyright 2003 by The University of Chicago. All rights reserved. 0033-5770/2003/7801-0002$15.00 OUT OF THE QUAGMIRE OF PLANT DEFENSE HYPOTHESES Nancy Stamp Department of Biological Sciences, Binghamton University Binghamton, New York 13902 USA e-mail: nstamp@binghamton.edu keywords plant defense, optimal defense hypothesis, carbon:nutrient balance hypothesis, growth-differentiation balance hypothesis, growth rate hypothesis abstract Several hypotheses, mainly Optimal Defense (OD), Carbon:Nutrient Balance (CNB), Growth Rate (GR), and Growth-Differentiation Balance (GDB), have individually served as frameworks for inves- tigating the patterns of plant defense against herbivores, in particular the pattern of constitutive defense. The predictions and tests of these hypotheses have been problematic for a variety of reasons and have led to considerable confusion about the state of the “theory of plant defense.” The primary contribution of the OD hypothesis is that it has served as the main framework for investigation of genotypic expression of plant defense, with the emphasis on allocation cost of defense. The primary contribution of the CNB hypothesis is that it has served as the main framework for investigation of how resources affect phenotypic expression of plant defense, often with studies concerned about allo- cation cost of defense. The primary contribution of the GR hypothesis is that it explains how intrinsic growth rate of plants shaped evolutionarily by resource availability affects defensive patterns. The primary contribution of the expanded GDB hypothesis is that it recognizes the constant physiological tradeoff between growth and differentiation at the cellular and tissue levels relative to the selective pressures of resource availability, including explicitly taking into account plant tolerance of damage by enemies. A clearer understanding of these hypotheses and what we have learned from investigations that use them can facilitate development of well-designed experiments that address the gaps in our knowledge of plant defense. P LANTS PRODUCE TISSUE from raw materials and then consumers (patho- gens and herbivores) harvest some of that for themselves. It appears that plants have evolved a variety of mechanical and chemical defenses primarily to ward off these consumers. In turn the consumers have evolved countermeasures, which then would act as selective pressure on plants for further defense. It has been the quest to understand the levels of defense and the array of defenses in plants that has given rise to the body of knowledge known as the “theory of plant defense.” More specifically, a central goal in plant-herbivore interactions has been to explain and predict phenotypic, genetic, and geographic variation in plant defense. The research on plant defense has been guided by a series of hypotheses that initially seemed to hold great promise for developing a gen- eral theory of plant defense, in particular one that would explain why most plants seem to be so well defended. By the 1950s, knowledge about the
Transcript of OUT OF THE QUAGMIRE OF PLANT DEFENSE HYPOTHESES · Secondary metabolites are found mostly in...

Volume 78, No. 1 March 2003THE QUARTERLY REVIEW OF BIOLOGY
23
The Quarterly Review of Biology, March 2003, Vol. 78, No. 1
Copyright � 2003 by The University of Chicago. All rights reserved.
0033-5770/2003/7801-0002$15.00
OUT OF THE QUAGMIRE OF PLANT DEFENSE HYPOTHESES
Nancy Stamp
Department of Biological Sciences, Binghamton University
Binghamton, New York 13902 USA
e-mail: [email protected]
keywordsplant defense, optimal defense hypothesis, carbon:nutrient balance hypothesis,
growth-differentiation balance hypothesis, growth rate hypothesis
abstractSeveral hypotheses, mainly Optimal Defense (OD), Carbon:Nutrient Balance (CNB), Growth Rate
(GR), and Growth-Differentiation Balance (GDB), have individually served as frameworks for inves-tigating the patterns of plant defense against herbivores, in particular the pattern of constitutivedefense. The predictions and tests of these hypotheses have been problematic for a variety of reasonsand have led to considerable confusion about the state of the “theory of plant defense.” The primarycontribution of the OD hypothesis is that it has served as the main framework for investigation ofgenotypic expression of plant defense, with the emphasis on allocation cost of defense. The primarycontribution of the CNB hypothesis is that it has served as the main framework for investigation ofhow resources affect phenotypic expression of plant defense, often with studies concerned about allo-cation cost of defense. The primary contribution of the GR hypothesis is that it explains how intrinsicgrowth rate of plants shaped evolutionarily by resource availability affects defensive patterns. Theprimary contribution of the expanded GDB hypothesis is that it recognizes the constant physiologicaltradeoff between growth and differentiation at the cellular and tissue levels relative to the selectivepressures of resource availability, including explicitly taking into account plant tolerance of damageby enemies. A clearer understanding of these hypotheses and what we have learned from investigationsthat use them can facilitate development of well-designed experiments that address the gaps in ourknowledge of plant defense.
PLANTS PRODUCE TISSUE from rawmaterials and then consumers (patho-
gens and herbivores) harvest some of that forthemselves. It appears that plants have evolveda variety of mechanical and chemical defensesprimarily to ward off these consumers. In turnthe consumers have evolved countermeasures,which then would act as selective pressure onplants for further defense.
It has been the quest to understand thelevels of defense and the array of defenses inplants that has given rise to the body of
knowledge known as the “theory of plantdefense.” More specifically, a central goal inplant-herbivore interactions has been toexplain and predict phenotypic, genetic, andgeographic variation in plant defense. Theresearch on plant defense has been guidedby a series of hypotheses that initially seemedto hold great promise for developing a gen-eral theory of plant defense, in particular onethat would explain why most plants seem tobe so well defended.
By the 1950s, knowledge about the

24 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
amount, array, and biological activity of “sec-ondary” metabolites in plants suggested thatthese compounds served as defense againstpathogens and herbivores (Dethier 1954;Fraenkel 1959). Because these metabolitesdid not appear to have a direct role in pri-mary metabolism, they were labeled “second-ary” (Whittaker and Feeny 1971). Secondarymetabolites are found mostly in plants, fungi,and microorganisms, specifically in organ-isms that lack an immune system (Williams etal. 1989). Allelochemicals, by which an organ-ism of one species affects an organism ofanother species (Whittaker 1975), are a sub-set of secondary metabolites. Some secondarymetabolites have other functions (e.g., ultra-violet protection, drought tolerance).
Further groundwork was laid with Ehrlichand Raven’s (1964) correlation of related but-terfly species using plants that were taxonom-ically and/or chemically related. To explainthe diversification of terrestrial plants andinsect herbivores, they proposed a “coevolu-tion” model based on macroevolutionary pat-terns in plant defensive chemistry and hostplant affiliations of insect herbivores. Theidea is that a plant group develops a newchemical defense that reduces attack by her-bivores and so eventually allows diversifica-tion as the plants proliferate and move intonew niches. At some point this is followed byevolution of counteradaptations by an insectgroup, which also eventually allows diversifi-cation as the insects are able to exploit thisdiverse plant group. In theory, this process ofsuccessive adaptive radiations or repeatedstarbursts of speciation yields reciprocal spe-ciation in mutant host plant and insect her-bivore lineages, but it does not produce par-allel cladogenesis at the level of matchedspecies (Thompson 1999). It is not a lock-stepprocess, with every speciation matched oneon one. Importantly, because the plant line-age undergoes diversification without signifi-cant interaction with herbivores, the eventualcolonization by herbivores is just as likely tostart with a derived plant species as the ances-tral plant species (Thompson 1999). To dis-tinguish it as a special form of coevolution,this model has been called escape-and-radiatecoevolution (Thompson 1989). The best sup-
port for this model is the consistently highdiversification rates in the multiple, indepen-dent plant lineages that developed secretorycanals containing latex or resin (Farrell andMitter 1998).
Ehrlich and Raven’s (1964) model can alsobe classified as diffuse coevolution (Futuymaand Keese 1992). The term diffuse coevolu-tion ( Janzen 1980; Fox 1981) or guild coevo-lution (Thompson 1989) refers to “the evo-lution of a particular trait in one or morespecies in response to a trait or suite of traitsin several other species” (Futuyma and Slat-kin 1983:2). Application of the term rangesfrom the Ehrlich and Raven (1964) scenariodescribed above to the view offered by Fox(1981), with apparent plants (such as woodyspecies) and their large set of herbivoresinvolved in gradual and continuous adjust-ments rather than starburst (or even step-wise) changes. Although diffuse coevolutionhas been suggested as a potentially largeinfluence on the diversity of terrestrial plantsand insect herbivores (Futuyma and Keese1992), there is not much concrete evidenceand little in the way of rigorous tests (Farrelland Mitter 1998), and so it has been difficultto characterize. Central to this problem is thatthe concept can be applied too broadly—e.g.,to a community of plants and herbivores—and so becomes impossible to analyze(Thompson 1989).
In contrast to diffuse coevolution, pairwise(or specific reciprocal) coevolution refers totwo interacting species exerting selectivepressure on each other, resulting in stepwiseevolution, as first one then the other evolvesa response to specific changes in the other( Janzen 1980; Fox 1981). Pairwise coevolu-tion is probably rare (Futuyma and Keese1992). The strongest potential cases are thosein which the host plant species are primarilyattacked by specialized insect herbivores(Futuyma 1983; Farrell and Mitter 1998).Such a situation may occur in unapparentplants, typically herbaceous species, whichhave fewer herbivores as a consequence oftheir small size, relative structural simplicity,and ephemeral nature temporally and spa-tially (Fox 1981). The phylogenic congru-ence of the longhorned beetle genus Tetraopes

March 2003 25PLANT DEFENSE HYPOTHESES
and the milkweed genus Asclepias may be anexample of stepwise or reciprocal diversifica-tion (Farrell and Mitter 1994, 1998). By itself,however, comparison of branching patternsof interacting lineages can show parallel clad-eogenesis, but it does not indicate whetherthe lineages have coevolved (Thompson1999). Examination of genetic variability in aplant species’ resistance to a particular her-bivore species and in the herbivore’s ability tocounter that may provide support for stepwiseevolution. For example, Berenbaum and Zan-gerl (1998) found chemical phenotypematching between wild parsnip introducedinto North America and its principal herbi-vore, the parsnip webworm, and for at leastone furanocoumarin trait interaction seemsto have contributed to escalation. The kindsof data needed to test the ideas of stepwiseand diffuse coevolution have been outlined,but such data are not easily obtained(Futuyma and Keese 1992; Farrell and Mitter1993, 1994, 1998; Thompson 1999).
Whether viewed as pairwise or diffusecoevolution, the idea has been that plantsand insect herbivores are engaged in an evo-lutionary arms race (Feeny 1975). However,the arms race view (Dawkins and Krebs 1979)seems too simplistic now. Besides a lack of evi-dence that is strongly supportive of pairwiseand diffuse coevolution as the dominantexplanation for the patterns of plant defense(Mitter and Brooks 1983; Farrell and Mitter1994, 1998), mathematical models suggestthat in general the arms race analogy foradaptive responses by predator and prey toone another is a poor one (Abrams 1986;Seger 1992). In terms of natural history, it isalso a questionable generalization. Host plantuse by herbivores is shaped by predators andclimatic conditions as well as host plant attrib-utes (Fox 1981; Janzen 1985).
In sum, the concepts of coevolution andthe arms race analogy have served as a usefulframework for study of the evolutionary rela-tionship between groups of host plants andtheir specialized herbivores, but paralleldiversification due to pairwise coevolution is“most certainly the exception rather than therule” (Farrell and Mitter 1994:67). Further-more, the limited evidence for and the diffi-
culty in testing ideas about diffuse coevolu-tion favor a conservative conclusion about thedegree to which it has influenced diversifica-tion. So while plant phylogeny and evolution-ary history of interactions with herbivores cer-tainly play a role in the pattern of plantdefense, the coevolutionary ideas aboutdiversification do not provide a complete oradequate explanation for the defensive pat-tern of most plants and, thus, the plants’ rela-tionships with their enemies. Clearly plantshave evolved an array of chemical and physi-cal defenses against a diverse set of enemiesand under variable abiotic conditions. In thedevelopment of a theory of plant defense, weseek explanations that take all of that intoaccount.
By the mid-1970s, the Optimal DefenseHypothesis, as outlined by Feeny (1975, 1976)and Rhoades and Cates (1976; Rhoades1979), was taking shape. This framework,which will be described later, was very appeal-ing and inspired many tests of the hypothesis.As a result, by the mid-1980s, the cumulativedata, not all of which fit the predictions of thehypothesis, indicated the need for a new syn-thesis. Coley, Bryant, and Chapin’s (1985)Growth Rate (or Resource Availability)Hypothesis provided that. At about the sametime, the Carbon:Nutrient Balance Hypothe-sis was developed (Bryant et al. 1983; Tuomiet al. 1988). These syntheses stimulated muchof the recent work on plant defense. By the1990s, however, data gathered to test thesevarious hypotheses suggested the need for yetanother synthesis. Herms and Mattson (1992)stepped in with an ecological and evolution-ary expansion of Loomis’s (1932, 1953)Growth-Differentiation Balance Hypothesis.In the last decade, all of these hypotheseshave served as a framework for research onplant defense. That is, all have been cited as thetheoretical basis for recently published studies. Fur-thermore, often two or more are cited in astudy, even though the hypotheses addressdifferent issues.
At this point in time, there is considerableconfusion about the hypotheses and, conse-quently, about the state of the theory of plantdefense. There are several reasons for this.The diversity of secondary metabolites and

26 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
their functions (which includes chemicalsthat negatively affect herbivores), the profu-sion of hypotheses that are not mutuallyexclusive, the difficulty in testing the hypoth-eses, and seemingly contradictory resultshave contributed to a perception that thereis no tangible theory of plant defense. Fur-thermore, the nature of these hypothesesnecessitates testing subhypotheses, whichsometimes have not been appropriately con-structed and/or tested adequately, and theresults have added to the confusion. Not sur-prisingly then, it has been suggested that anall-encompassing theory may be biologicallyunrealistic (Berenbaum 1995). Such confu-sion is typical of certain stages in the devel-opment of scientific theory, however (Loehle1987). Young and consequently immaturetheory lacks precise statements, which makeit difficult to resolve the questions posed(Loehle 1987,1988). Progress in developingtheory will be slow when time-scales for thesystem are long, little history of the system isleft, and the system is complex (Loehle 1987).
It is unclear whether the current disarrayreflects one of those immature stages orwhether it signals that these hypotheses areunsuitable, and moreover that an all-encompassing theory of plant defense isimpossible. Before discarding the hypothe-ses or accepting the conclusion that an all-encompassing theory is unobtainable, weneed to consider the degree to which thehypotheses have been tested adequately anddetermine exactly what we do and do notknow about plant defense as a result of theseframeworks.
Four hypotheses of plant defense will beexamined in detail here: 1) Optimal Defense(OD), 2) Carbon:Nutrient Balance (CNB),3) Growth Rate (GR), and 4) Growth-Differ-entiation Balance (GDB). These plant defensehypotheses were chosen because they are theones that are cited most often and also are rou-tinely misinterpreted. Each of these hypothe-ses has assumptions and predictions, but onlyto some degree have these hypotheses actuallybeen tested. In fact, the major assumptions,predictions, tests, and results have not beenspelled out clearly, and so it has been difficultto evaluate what we have and have not learnedfrom testing these hypotheses.
Hypotheses of Plant DefenseAgainst Herbivores
the optimal defense hypothesesThe Optimal Defense (OD) Hypotheses
address how defensive needs of a plant con-tribute to the evolution of secondary metab-olites, with costs of defense paid to maximizeplant fitness (McKey 1974, 1979; Rhoades1979). The basic hypothesis (Hypothesis 1)states that, “Organisms evolve and allocatedefenses in a way that maximizes individualinclusive fitness” (Rhoades 1979:12). Inessence, this hypothesis states that any defen-sive pattern is possible if it is adaptive (McKey1974, 1979; Rhoades 1979). However, it is dif-ficult to test this hypothesis without more for-malization of its language because neither theconcepts nor predictions are obvious (Fager-strom et al. 1987). Thus, while the conclu-sions have been that “any defensive pattern ispossible if it is adaptive,” in practice thehypothesis is not really falsifiable. EvenRhoades (1979) said that this hypothesis isprobably impossible to test directly.
The second hypothesis (Hypothesis 2) statesthat “Defenses are costly, in terms of fitness”because they divert resources from “otherneeds,” which mainly has been interpreted asgrowth and reproduction (Rhoades 1979:13).Although measuring cost of defense to fitnessremains an issue, the derivative hypotheses aremore testable than the basic OD hypothesis.The subhypotheses are: a) “Organisms evolvedefenses in direct proportion to their risk frompredators and in inverse proportion to the costof defense” (i.e., the Plant Apparency Hypoth-esis); b) within an organism, defenses are allo-cated in proportion to risk of the plant partand value of it to plant fitness, and in inverseproportion to cost of defense; c) defense isdecreased when enemies are absent andincreased when they are present (i.e., alloca-tion pattern of constitutive and inducibledefenses); and d) there is a tradeoff betweendefense and other plant functions (growthand reproduction) such that stressed individ-uals are less defended (Rhoades 1979:13).
Subhypotheses 2a and 2b refer to genotypicexpression of defense, whereas subhypoth-eses 2c and 2d refer to phenotypic expressionof defense. Cost of defense is an issue in all

March 2003 27PLANT DEFENSE HYPOTHESES
four subhypotheses. Examination of cost instudies of genotypic expression of defenseaddresses genetic constraints and microevo-lutionary responses to selection. In contrast,examination of cost in studies of phenotypicexpression of defense focuses on allocationpatterns and cost of expressing particularphenotypes.
Assumptions
The OD hypotheses assume that: 1) thereis genetic variation in secondary metabolitesfor selection to act upon; 2) herbivory is theprimary selective force for production of sec-ondary metabolites; and 3) defenses reduceherbivory.
In general, extensive data support theassumptions. For instance, studies haveshown intraspecific genetic variation inamount and type of secondary metabolites(Dirzo and Harper 1982; Zangerl and Ber-enbaum 1990; Vrieling et al. 1993; van Damand Vrieling 1994; Mauricio 1998). There isconsiderable circumstantial evidence thatherbivory is a major selective force on plants.In grasslands about 33% of the production isconsumed by herbivores, and over 60% of theseaweed production is consumed by herbi-vores (Hay 1991). Herbivory can adverselyaffect plant growth and reproduction (e.g.,Marquis 1984; Wisdom et al. 1989; Strauss1991; Karban and Strauss 1993). In general,loss to herbivores exceeds the allocation toreproduction (Mooney 1972). Of course,most plants can tolerate some herbivory with-out a reduction in fitness, and some speciesor populations can tolerate a great deal ofherbivory (McNaughton 1983; Paige andWhitham 1987). Nonetheless, a vast array ofsecondary metabolites have evolved in plants,most of which seem primarily involved inplant defense against herbivores (Berenbaum1995). Palatability is mainly determined byplant defenses. Furthermore, ingested plantdefenses can be toxic to herbivores. Forexample, terpenes can inhibit ATP forma-tion, alkylate nucleophiles, disrupt moltinghormonal activity, bind with protein and ster-ols in the gut, and disrupt the nervous system(Langenheim 1994). Not surprisingly, thereis direct evidence indicating that herbivorescan act as selective agents for plant traits that
reduce herbivory (Simms and Rausher 1989;Mauricio and Rausher 1997).
Hypothesis 2 and its Subhypotheses
Allocation Cost of Genotypic Defense(Hypothesis 2)
The major prediction is that a cost ofdefense to fitness results from diversion ofresources from other needs (“allocationcost”). Accordingly, when enemies are absent,less well-defended individuals should havehigher fitness than better defended individ-uals.
As expected, there are costs to biosynthesisof secondary metabolites (Mooney et al.1983; Baas 1989; Gershenzon 1994a,b). Met-abolic cost depends on substrate and cofactorsources, and defenses have additional costssuch as transport and vacuole or resin ductconstruction (Gershenzon 1994a,b). In addi-tion, turnover and recycling affect cost. Largemolecule nonnitrogen-containing defenses,such as lignin, are not recycled (Mooney etal. 1983). Other defenses, such as terpenes,can be degraded and the products recycledinto primary metabolism (Gershenzon1994a,b). But turnover of defenses does notnecessarily mean that limited nutrients, suchas nitrogen, are recovered by the plant (Bald-win and Ohnmeiss 1994).
The cost of defense can be subject to evo-lution, such that natural selection operatingon variation in cost among individuals couldeliminate individuals with high fitness cost ofdefense (Simms 1992; Karban 1993). Exam-ples are the proposed cost-reducing adapta-tions for terpenes: sharing biosyntheticenzymes among multiple pathways, minimi-zation of enzyme turnover, use of a singleenzyme to produce a mixture of products,catabolism of products no longer needed,and use of products for more than one func-tion (Gershenzon 1994a).
Clearly there can be tradeoffs in allocationbetween defense and growth or reproduction(Vrieling and van Wijk 1994; Bergelson andPurrington 1996; Strauss et al. 2002). Forexample, an increase across genotypes inmyrosinase (which acts on glucosinolates toform isothiocyanates) was correlated withincreased resistance to herbivores but at the

28 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
cost of lower seed production (Mitchell-Oldset al. 1996). Tests are difficult when thegenetic backgrounds of the susceptible andresistant genotypes differ in more ways thanjust a resistant gene. Through genetic engi-neering it is possible to conduct a more con-trolled experiment. For example, a resistantgene transplanted into a herbaceous plantreduced lifetime seed production by 34%,illustrating that the cost of resistance to ene-mies can be substantial (Bergelson et al.1996).
But total cost of resistance is likely to reflectother mechanisms than just “allocation costs”(costs via diversion of energy and nutrients),in particular ecological costs (Purrington2000; Strauss et al. 2002). Ecological costs ofresistance include deterrence of mutualists(e.g., pollinators, mycorrhizal fungi), reduc-tion in competitive ability, traits that deterone enemy but attract another, and traits thatconfer resistance against one enemy but con-strain resistance to another.
Furthermore, evaluating the tradeoffbetween growth-reproduction and defense iscomplicated by the “third party” tradeoff withtolerance. As opposed to resistance (traitsthat reduce the amount of damage), toler-ance refers to traits that reduce the impact ofdamage on plant fitness (Stowe et al. 2000).Tolerance to tissue damage and loss is anemergent property, reflecting intrinsicgrowth rate, storage capacity, allocation pat-tern, flexible photosynthetic rate, flexiblenutrient uptake, and developmental plasticity(Rosenthal and Kotanen 1994). Just as sec-ondary metabolites are a product of variousselective pressures besides herbivory, so too istolerance. Herbivory is probably the greatestselection pressure for tolerance (or basicallyregrowth capacity), however. Plants exhibitgenetic variation for tolerance to herbivory(Fineblum and Rausher 1995; Mauricio et al.1997), and tolerance can counter the poten-tial negative impact of herbivory on repro-duction (Maschinski and Whitham 1989).
Plants appear to have one of three strate-gies: well-developed defense and poor toler-ance, well-developed tolerance and poordefense, or an intermediate of both (van derMeijden et al. 1988). For instance, damagehad no effect on alkaloid concentration of a
plant species that typically has little herbivorybut maintains a high constitutive level ofdefense, whereas damage resulted in loweralkaloid concentration in a second speciesthat suffers high herbivory but exhibits sub-stantial regrowth, and damage increased alka-loids in a third plant species that has little her-bivory but also slow regrowth (van Dam et al.1993). Some evidence indicates that defenseand tolerance covary negatively (Bilbroughand Richards 1993; Fineblum and Rausher1995), but defense and tolerance can co-occur together (Rosenthal and Kotanen1994; Mauricio et al. 1997).
We might expect a fitness cost to tolerancein the absence of herbivory. An example of afitness cost to tolerance in the absence of her-bivory is grazing-tolerant plants being com-petitively inferior to grazing-intolerant plantswhen herbivores are absent (Painter 1987).Costs of tolerance could arise directly—e.g.,allocation costs, opportunity costs (small costsearly in life that are compounded as plantsage)—and indirectly (i.e., ecological costs,such as altering the soil microbial relation-ship with the plant).
The greater an allocation to tolerance (“athird party”), the more likely that any tradeoffbetween growth and tolerance, or betweendefense and tolerance, will be undetected(Mole 1994). Paying attention to such thirdparty tradeoffs is especially importantbecause allelochemicals often may requirerelatively small allocations of resources; thus,due to third party tradeoffs, measuring costsof defense is quite difficult (Mole 1994).
It may be especially difficult to demonstratecost of defense to fitness for plants with thestrategy of “well-developed tolerance andpoor defense” and plants with the strategy of“intermediate tolerance and defense”because cost of tolerance may obscure fitnesscosts of defense. For example, if the scale forcost of tolerance is greater than that fordefense, then an increase in defense willdecrease the cost of tolerance disproportion-ately and, thus, decrease the total cost of tol-erance and defense (Simms and Triplett1994). This could yield a positive genetic cor-relation between defense level and fitness inthe absence of herbivory; i.e., there would bethe appearance of no cost of defense.

March 2003 29PLANT DEFENSE HYPOTHESES
In sum, defenses do have a cost in terms ofconstruction and maintenance, but there islikely to have been selection to reduce thatcost. Tradeoffs in allocation to different func-tions (e.g., growth-reproduction versusdefense) can occur, but other costs, such asecological costs, may be as important or moreimportant. The sum of defenses in theabsence of herbivory may show a cost to fit-ness, but detection is complicated by the thirdparty tradeoff with tolerance. Consequently,when no cost of resistance is found, we do notknow if there is no cost, or whether the inves-tigation is incomplete (which it most certainlywill be). Thus, no study has emphatically con-cluded that there is no cost of resistance forthe focal plant(s). It seems that a more usefulapproach is to focus on why certain traits costas much as they do, effect of environmentalconditions on the magnitude of cost, mech-anisms and implications of cost between resis-tance and tolerance, and the sum benefit andcost of a trait that has negative effects on someenemies but positive effects on others (Pur-rington 2000; Hamilton et al. 2001).
Plant Apparency (Subhypothesis 2a)The Plant Apparency Hypothesis (Feeny
1975, 1976; Rhoades and Cates 1976) is basedon the optimal defense idea (Rhoades 1979).It predicts that apparent plants (e.g., oaktrees) are easily found by herbivores and,therefore, these plants have a large invest-ment in broadly effective defenses. These dos-age dependent or “quantitative” defenses(Feeny 1976) have the characteristic of inter-fering with nutrient acquisition by herbivoresand so are called “digestibility reducing”defenses (Rhoades and Cates 1976). Theinvestment is predicted to be large becausethe molecules are large, and it may take arelatively high concentration to have a nega-tive impact. Tannins are an example of aquantitative defense. These quantitativedefenses slow the growth rate of herbivores,and so are predicted to subject them tohigher rates of predation and parasitism(Feeny 1976). Even herbivores that naturallyfeed on such plant material are subject to thenegative effect of quantitative defenses ongrowth. In contrast, unapparent plants (e.g.,crucifers) have an unpredictable distribution,
and so they are expected to often escape spe-cialist herbivores. Consequently, these plantsare predicted to invest in less costly defensesthat are useful against nonadapted general-ists. These “qualitative” defenses are expectedto be less costly because they are relativelysmall molecules and toxic at low dosage(Feeny 1976). Glucosinolates, cyanogenic gly-cosides, and alkaloids are examples of quali-tative defenses.
While certainly an appropriate and usefulhypothesis at the time, evidence gathereddoes not fully support the Plant ApparencyHypothesis (Fox 1981; Futuyma 1983; Coleyet al. 1985; Waterman and Mole 1989). Insupport of the hypothesis, there are correla-tions between apparency of plants and moregeneralized defense (Berenbaum 1981, 1983;Coley 1983), but there are also many exam-ples that contradict the predictions. The arse-nal of defenses of plants that fit the “unap-parent” lifestyle may include quantitativedefenses, and “apparent” plants may havequalitative defenses (Futuyma 1976).Although described as a quantitative defense,tannins are not all-purpose digestibilityreducers (Martin et al. 1987) and can act astoxins (Steinly and Berenbaum 1985). Clearlymany herbivore species have counteradapta-tions to tannin-rich food (Berenbaum 1980;Bernays et al. 1980; Schultz and Lechowicz1986; Martin et al. 1987). Furthermore, formany insects the deterrent effect of tannin-containing food may reflect associated quali-ties (e.g., low nitrogen, low water, toughness)that render the food poor for herbivores(Bernays 1981). Then there are the putative“qualitative” defenses, such as terpenes. Forsome plant species, terpenes seem to be qual-itative defenses, but for other species theyseem to be quantitative defenses (Fox 1981).In addition, even at low concentrations, qual-itative defenses may be costly, due to nitrogeninvestment (Baldwin et al. 1990) and rapidturnover rate.
In sum, the Plant Apparency Hypothesiswas a useful framework when it was proposed,and it guided a number of studies that madesignificant contributions to the currentunderstanding of plant defense, but the dif-ficulty in trying to measure apparency (Feeny1991) and the mixed support for the hypo-

30 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
thesis have prevented further profitableresearch along those lines.
Optimal Defense Within Plant(Subhypothesis 2b)
Another subhypothesis of the OD hypoth-esis is that, within a plant, defenses are allo-cated in direct proportion to the risk of theparticular tissue to herbivory and the value ofthat tissue in terms of loss of fitness (McKey1974, 1979; Rhoades 1979).
In general, data support the idea thatdefenses are allocated in proportion to risk oftissue to herbivory (Bryant et al. 1983; Zan-gerl and Bazzaz 1992; Baldwin and Schmelz1994; Baldwin and Karb 1995; van Dam et al.1995; Wallace and Eigenbrode 2002). Forexample, the reproductive parts of wild pars-nip, which had an estimated high probabilityof being attacked and greatest value to fitness,had the highest constitutive level of a toxicfuranocoumarin, and the furanocoumarinwas not inducible there (Zangerl and Rut-ledge 1996). Roots of wild parsnip, whichwere the least likely plant part to be attacked,had the lowest constitutive level of the fur-anocoumarin but were highly inducible.Leaves, which had a high probability ofattack, had intermediate constitutive andinducible levels.
Examining this hypothesis requires mea-surement of three factors: 1) value of plantpart, 2) benefit of defense, and 3) probabilityof attack (Zangerl and Bazzaz 1992; Hamiltonet al. 2001). Value of plant part can and hasbeen measured, but usually studies just inferdifferent values of plant parts based on thegeneralization that reproductive parts carryhigher value to plant fitness than nonrepro-ductive parts (Zangerl and Bazzaz 1992; Ham-ilton et al. 2001). To determine benefit ofdefense would require having plants with andwithout defense exposed to herbivores (Ham-ilton et al. 2001), which is not feasible (untilgenetic manipulation can eliminate a plant’sdefense system). Instead researchers haveassumed that a correlation between fitnessand defense of plants exposed to herbivoresis evidence for a benefit of defense propor-tional to defense level (Hamilton et al. 2001).But that is problematic. For example, syner-gerism among defenses can be high (Witts-
tock and Gershenzon 2002), and so benefitof defense and defense level may not corre-late. In addition, to conduct a test, the samekind of defense must occur throughout theplant, so that cost of defense and effectivenessof defense only vary with level of defense(Zangerl and Rutledge 1996). Consequently,it may only be possible to conduct tests withsystems in which the defensive array is rela-tively simple (e.g., wild parsnip). The few teststhat have been done were with herbaceoustemperate species (Hamilton et al. 2001).Estimating probability of attack is seldomdone (but see Zangerl and Rutledge 1996)and is difficult to do with any confidence(Hamilton et al. 2001). Importantly, severalpopulations with different levels of herbivoryshould be measured.
So while the logic of the Optimal DefenseWithin Plant Hypothesis is sound, which isbased on generalities about within-plant dis-tributional patterns of secondary metabolitesand plant part function, in truth the empiri-cal evidence is limited due to what we canrealistically measure. Nonetheless, at this pointin time, it seems reasonable to accept the ideathat defense allocation among plant parts ofherbaceous species reflects cost:benefit pat-terns in plant fitness.
Inducible Defense (Subhypothesis 2c)Another subhypothesis of the OD hypoth-
esis describes inducible defenses. Inducibledefenses refer to defenses produced inresponse to damage and so defense levels riseabove the baseline or constitutive level.Because defense is predicted to be a costlycommitment, defense should be reducedwhen herbivores are absent and increasedwhen plants are subject to attack (Rhoades1979). Furthermore, because defense is pre-dicted to be costly, the relaxation time ofinduced defenses is expected to be short.“Relaxation time” refers to how long it takesthe level of induced defense to drop to thebaseline or previous constitutive defensivelevel. Thus, the “defense view” of induction isthat changes in phytochemistry in responseto herbivory have been shaped over evolu-tionary time as a defense (Rhoades 1979;Haukioja and Neuvonen 1985).
Rapid induction of specific defenses in

March 2003 31PLANT DEFENSE HYPOTHESES
response to herbivory (within hours, followedby relaxation over days or weeks) can beexplained by the OD induction prediction.An example is the production of proteinaseinhibitors in response to a wound-signalingmechanism within hours of damage intomato leaves (Green and Ryan 1972; Broad-way et al. 1986). Herbivory triggers release ofsystemin, which activates the jasmonic acidpathway, which in turn increases products of15 or more genes, including proteinase inhib-itor (Bergey et al. 1996). Interference in thisprocess (via mutant genes) results in less pro-teinase inhibitor (Orozco-Cardenas et al.1993) and more feeding and growth by insectherbivores (Howe et al. 1996). Other studieshave shown that induced plant responses toherbivory or jasmonic acid provided higherlevels of defense that resulted in greater seedproduction compared to controls (Agrawal1998; Baldwin 1998).
Induced resistance can result in an alloca-tion cost. Seed production in tobacco plantswas not reduced in damaged plants in whichalkaloid production was inhibited by auxin,but it was reduced in plants that increasedalkaloid production in response to damage(Baldwin et al. 1990). In addition, even in theabsence of enemies, inducible defenses mayhave allocation costs and opportunity costs,via the requirement of maintaining wound-detection pathways and defense precursors(Cipollini 1998; DeWitt et al. 1998; Purring-ton 2000). Induced resistance may also haveecological costs (Strauss et al. 2002).
A key issue in testing hypotheses aboutinduction is an estimate of probability ofattack (Karban and Baldwin 1997). Plantswith a low probability of attack are expectedto exhibit greater inducibility than those witha high probability of attack because the latterwould benefit more by high levels of consti-tutive defense. But probability of attack is dif-ficult to assess and can result in misinterpre-tation if reciprocal transplanting is not done.For instance, based on the OD induction sub-hypothesis, the expectation is that popula-tions subject to high herbivory would exhibithigh levels of constitutive defense and low lev-els of inducibility, whereas populations sub-ject to low herbivory would exhibit low levelsof constitutive defense and be highly induc-
ible. Comparison of two populations of wildparsnip indicated that one was subject to fre-quent herbivory (42%) in contrast to theother (7%), but a transplant experimentshowed that these differences reflected thesites (or habitats), since those levels of her-bivory occurred at the sites regardless of thepopulation source (Zangerl and Berenbaum1990). Furthermore, the two populations didnot differ in inducible levels and only in twoof four constitutive defense levels (and appar-ently those differences were not biologicallysignificant, as herbivory levels reflected siterather than population differences).
A prediction of the OD induction subhy-pothesis is that among species there shouldbe a negative correlation between the level ofconstitutive defense and level of inducibility.Some data support that prediction, but inmany other cases the data indicate either apositive correlation or no correlation(reviewed by Karban and Baldwin 1997).Even between populations, there may be nocorrelation among constitutive levels,induced levels, and level of herbivory (Zan-gerl and Berenbaum 1990). One explanationfor a lack of negative correlation betweenconstitutive and inducible defense levels isthat perhaps plants do not take on a partic-ular phenotype matched to a particular envi-ronment (e.g., increased defense in responseto increased herbivory). In contrast, plantsmay respond to attack by simply changingphenotypes (levels of defenses within thedefensive array change, which changes foodquality) and, thus, plants present a “movingtarget” to enemies (Adler and Karban 1994).In the only clear test of this idea, natural pop-ulations of a postfire annual exhibited defen-sive changes in phenotype that matched thepredictions of the OD Inducible DefenseHypothesis rather than the Moving TargetHypothesis (Baldwin 1998). Jasmonate-induced defenses reduced seed production(so had a cost to fitness), but having an induc-ible defense allowed the plants to avoid thatcost when plants were not attacked (i.e.,defenses were not induced) (Baldwin 1998).Interestingly, as the postfire environmentaged and, in turn, herbivory increased, thenature of the induced defense benefitschanged from that of greater seed produc-

32 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
tion in the first year after a burn, comparedto noninduced plants, to greater survival tothe stage of seed production in the secondyear.
Most likely the studies with a lack of nega-tive correlation between constitutive andinducible levels of defense reflect the evolu-tionary and ecological complications of hav-ing a defense system that can handle a widevariety of enemies, but which in turn carriesecological costs (Agrawal and Karban 1999).For example, insect herbivores in the samefeeding guild caused different inducible pat-terns: strong resistance, no resistance, orincreased susceptibility (Agrawal 2000). Forsome plants, induced resistance against somepathogens confers resistance to other patho-gens, but not to arthropod herbivores(reviewed in Agrawal and Karban 1999; Agra-wal 2000). Likewise, induced resistance againstherbivores may not confer resistance againstpathogens. For some plants, in response tosome enemies, the inducible defense pathwaysmay be separate and for others there may besome “cross-talk” among pathways (Agrawaland Karban 1999; Felton and Korth 2000; Heiland Bostock 2002; Kunkel and Brooks 2002).
An alternative explanation for inducedresistance is that it simply reflects physiologi-cal change due to tissue damage rather thanan evolved defense. Strictly speaking, induc-tion refers to a change in phytochemistry inresponse to stress, which can include changesin secondary metabolism (Karban and Bald-win 1997). In many cases, increases in con-centration of secondary metabolites may bean unavoidable consequence of changes inprimary physiology and with no functionalimportance to the plant (Gershenzon 1984).Furthermore, induction does not necessarilyresult in deterring herbivores and improvingplant fitness (Fowler and Lawton 1985; Kar-ban and Myers 1989; Karban and Baldwin1997).
In the case of delayed induction (responsesoccurring in the season(s) after damage), therelaxation time is generally long (e.g., a yearor more) (Rhoades 1979; Tuomi et al. 1984).In general, insect herbivore performance isless on trees with delayed induced responsescompared to controls, but there are excep-tions (reviewed by Karban and Baldwin 1997).
Even though the phenomena of delayed in-duction and slow relaxation time do not fit theOD induction prediction (Rhoades 1979),they actually fit the general statement of theOD hypothesis (i.e., any observed defensivepattern is possible if it is adaptive) (Haukiojaand Neuvonen 1985). This duality in predic-tions illustrates one reason why the basic ODhypothesis has been difficult to test and, con-sequently, has been a source of confusionabout progress with the theory of plantdefense.
Allocation Cost of Phenotypic Defense(Subhypothesis 2d)
This subhypothesis states that because allo-cation to defense results in less allocation ofenergy and nutrients to other needs (e.g.,growth and reproduction), environmentallystressed plants should be less well defendedthan unstressed individuals (Rhoades 1979).Furthermore, under stress, plants are ex-pected to decrease commitment to costlydefenses and increase commitment to lesscostly but accordingly less effective defenses.
It may be that fitness costs are only signifi-cant at certain developmental stages (e.g.,reproduction) or in stressful situations (Bald-win et al. 1990; Briggs and Schultz 1990;Agren and Schemske 1993; Bergelson 1994;Gershenzon 1994b). A meta-analysis thatshowed negative correlation of defense andplant fitness under uncontrolled environ-mental conditions and persistence in thepresence of herbivores suggests that a cost ofdefense to fitness may often occur throughinteraction of plants and various componentsof their environment (Koricheva 2002).Responses to damage are strongly affected bythe source-sink relationships within a plant,and tolerance decreases with nutrient stress(Chapin and McNaughton 1989; Maschinskiand Whitham 1989). In general, stressed indi-viduals have low defense levels (reviewed inHerms and Mattson 1992). But as will beexplained later, the CNB and GDB hypothe-ses have more explicit explanations for pat-tern of defense across resource gradientsthan does the OD view. It is not clear whethercommitment to defense consists of less of alldefenses or less of the more costly and effec-tive ones, as predicted.

March 2003 33PLANT DEFENSE HYPOTHESES
In sum, the results are mixed for supportof the subhypotheses of the OD view. Basi-cally, the observed patterns are more compli-cated than originally envisioned. Of moreinterest now than cost of defense is how plantsevolve and balance an array of defenses com-plementing an array of tolerance traits towithstand a complex of enemies within theframework of optimizing growth and repro-duction. Nonetheless, all of the results areconsistent with the OD statement that plants“evolve and allocate defenses in the way thatmaximizes individual inclusive fitness”(Rhoades 1979:12). This framework and theensuing research led to the development ofother hypotheses of plant defense.
the carbon:nutrient balancehypothesis
The Carbon:Nutrient Balance (CNB)Hypothesis (also called the EnvironmentalConstraint Hypothesis) is a model of how thesupply of carbon and nutrients in the envi-ronment influence the phenotypic expres-sion of defense by plants (Bryant et al. 1983;Tuomi et al. 1988, 1991). Originally it wasdeveloped to explain the influence of soilnutrients and shade on plant defensive chem-istry via the effects on the carbon:nutrientratio of the plant (Bryant et al. 1983). Basically,the hypothesis is that if the carbon:nutrientratio acquired by a plant controls allocationof resources to plant functions, then the phe-notypic expression of that plant’s geneticpotential for defenses will be affected. Thevariation of phenotypic expression in defenseamong genotypes may range from none(complete genetic determination) to substan-tial (great plasticity in response to environ-mental conditions). For example, a specieswith a high constitutive level of defense thattypically grows in nutrient poor soil may bewell defended because: 1) herbivory in suchsites selected for such genotypes, 2) chroniccarbon surplus is shunted into allelochemi-cals, or 3) some combination of these (Tuomiet al. 1988, 1991).
Some of the confusion and controversy inthe literature about the CNB hypothesisstems from conflicting statements in the origi-nal descriptions of the hypothesis. Some state-ments in the descriptions suggest that the
original view was that all defensive productionoccurs after growth requirements are met;e.g., “it implies no reduction in growth to sup-port carbon-based resistance” and “allocationto secondary metabolite production occursonly when growth demands for carbon havebeen met” (Tuomi et al. 1988:59). But otherstatements suggest that the original view wasthat a plant could produce some defenses tomeet a fixed allocation (in conjunction withgrowth) and produce some defenses via flex-ible allocation (altered carbon:nutrient ratioyielding carbon surplus, which is shuntedinto defense). For instance, “Some com-pounds may show a fixed allocation pattern,whereas others may depend more on the con-straints limiting plant primary metabolism”(Tuomi et al. 1991:92). “Thus, a model withboth P1 [proportion of carbon invested indefense when it could be allocated to growth]and P2 [proportion of carbon surplus beyondgrowth requirements allocated to defense]could presumably better describe actual allo-cation of carbon between defense andgrowth. In such a model, active defense allo-cation could determine some backgroundlevel of defense investments (i.e., P1) thatimply costs on growth. This background levelof defense could also be modified by selec-tion . . . Plant carbon/nutrient balance couldthus be a factor that accounts for a part of thephenotypic variation in constitutive levels ofcarbon-based allelochemicals” (p 90).
Since the original descriptions of the CNBhypothesis, however, researchers havefocused on the former view (i.e., allocation todefense comes after growth, therefore no costto defense). Much of the criticism of the CNBhypothesis (Karban and Baldwin 1997; Ham-ilton et al. 2001) reflects the problems withthat interpretation. But the original descrip-tions anticipated some of the criticisms in away that suggests that the latter view (consti-tutive defense � fixed and flexible alloca-tions) is what the authors of the CNB hypoth-esis intended.
Following up on the view that a plant’sdefense accrues from a combination of fixedallocation and flexible allocation, there aretwo scenarios to explain constitutive defenselevels. The first, “baseline plus,” is that plantsmay have a fixed baseline allocation to

34 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
defense (proportional to growth), the car-bon:nutrient balance may be affected by envi-ronmental conditions, carbon surplus(beyond use for growth) may be shunted intodefense (flexible allocation), and thusdefense at any point in time is a combinationof the baseline allocation and flexible allo-cation. The second, “variable plus,” is thatplants may have a fixed allocation to defense,but this allocation is proportional to bothgrowth and carbon surplus; the car-bon:nutrient balance may be affected by envi-ronmental conditions, carbon surplus(beyond use for growth) may be shunted intodefense (flexible allocation), and thusdefense at any point in time is a combinationof the shifting-but-fixed allocation and flexi-ble allocation. Either way, a plant’s responseto changes in the carbon:nutrient ratio canalter the phenotypic expression of the plant’sgenetic potential for defense.
Assumptions
The CNB hypothesis has several assump-tions. It assumes that carbon gain and growthdepend on the mineral nutrient reserves of aplant, that carbon is allocated to growthwhenever the supply of mineral nutrients isadequate, and that carbon accumulatedbeyond the level used for growth is allocatedto defense or storage; data support theseassumptions (Tuomi et al. 1988). The CNBhypothesis also assumes that growth is inhib-ited more by nutrient limitation than is pho-tosynthesis (Bryant et al. 1983; Tuomi et al.1988; Tuomi 1992); data support this idea aswell (reviewed by Bryant et al. 1983; Tuomi etal. 1988; Luxmoore 1991; Herms and Mattson1992). Lastly, the CNB hypothesis assumesthat herbivory is a primary selective force forconstitutive secondary metabolites and thatdefenses reduce herbivory (Bryant et al.1983; Tuomi et al. 1988), but it does notassume that the total amount or general typeof defense (nonnitrogen-containing versusnitrogenous) is selected for by herbivory(Tuomi et al. 1988).
Although a recent review claims that thefundamental assumptions of the CNB hypoth-esis are now known to be “incorrect” (Ham-ilton et al. 2001), the so-called “assumptions”were not presented accurately. It is not an
assumption of the CNB hypothesis that “theability of a plant to synthesise a defence com-pound can be predicted based on the atomscontained in the compound relative to theavailability of these atoms in the plant” (Ham-ilton et al. 2001:89). No such statement orimplication is made in the CNB hypothesispapers (Bryant et al. 1983; Tuomi et al. 1988,1991), nor is such an assumption required.What the reviewers are referring to is a predic-tion based on the CNB hypothesis that, forexample, shade will decrease the car-bon:nutrient ratio and thus cause an increasein internal nitrogen available for defense andstorage. It is also not an assumption “thatgrowth always takes priority over secondarymetabolism in allocation of resources” (Ham-ilton et al. 2001:89); in other words, thatgrowth takes priority over any level of defenseincluding genetically determined baselinedefense. What Tuomi et al. state is that “car-bon is allocated to growth whenever there aresufficient mineral nutrients to construct newcells, and . . . the carbon surplus accumulatedabove the levels required for growth is allo-cated among different carbon-based allelo-chemicals and/or carbohydrate storage”(1988:59). This statement refers to the phe-notypic expression of defense affected by thecarbon:nutrient ratio and not to the level ofdefense in place due to intrinsic geneticdetermination. Statements elsewhere makethis clear (Bryant et al. 1983; Tuomi et al.1988:60–61, 1991:90–92). It is not an assump-tion “that simple mass-action drives rates ofsecondary metabolite production (Reichardtet al. 1991) implies that plants have little abil-ity to control their chemical composition”(Hamilton et al. 2001:89). The CNB hypoth-esis acknowledges that carbon surplus is allo-cated to defense and/or storage (and thusregulated), and that surplus shunted intodefense involves regulation of synthesis path-ways; thus, precursors, substrates, and prod-ucts are under control of the plant (Bryant etal. 1983; Tuomi et al. 1988, 1991). Finally, itis not an assumption of the CNB hypothesisthat “an effect of environment is independentof the action of genes” (Hamilton et al.2001:91). The CNB hypothesis acknowledgesthat plant defense is programmed geneticallyand that plant species exhibit a range of plas-

March 2003 35PLANT DEFENSE HYPOTHESES
ticity in expression of levels and array ofdefenses, which is a genetically determinedresponse to the environment (Bryant et al.1983; Tuomi et al. 1988, 1991). Contrary tothe Hamilton et al. (2001) interpretation, theCNB hypothesis has its roots in the ideas ofMooney (1972), Grime (1977, 1979), andChapin (1980) about the effects of environ-mental resources on the evolution of plantfunctions (Bryant et al. 1983). Much of theHamilton et al. (2001) viewpoint aboutassumptions and predictions of the CNBhypothesis seems to be based on the discus-sion of Karban and Baldwin (1997:75–77),which also provides some misleading state-ments (e.g., that the CNB hypothesis presentsinduced metabolites as being essentially wasteproducts).
Predictions
The CNB hypothesis makes specific andtestable predictions about how stressful envi-ronments affect the amount and general typeof plant defense (Bryant et al. 1983). “Gen-eral type of defense” refers to nonnitrogen-containing versus nitrogenous. For genotypeswith little or no phenotypic plasticity in defense, theprediction is that any effects of resource con-ditions on the carbon:nutrient ratio of a plantdo not translate into a change in defense lev-els. For example, woody plants adapted to lowresource situations are expected to have a lowintrinsic growth rate and therefore low capac-ity for compensatory growth after herbivory,which in turn would favor selection for main-tenance of high defense levels (i.e., completegenetic determination of defense) and car-bon surplus into storage rather than defense.For genotypes with phenotypic plasticity in defense,the prediction is that any effects of resourceconditions on the carbon:nutrient ratio cancause a change in the total defense level.More specifically, the predictions are that fer-tilization or shade will decrease the car-bon:nutrient ratio of a plant, reducing theexcess carbon production and, consequently,decreasing nonnitrogen-containing defenseswhile increasing the availability of assimilatednitrogen for defense. That is, high levels ofnitrogenous defenses reflect nitrogen assim-ilated in excess of growth requirements and,vice versa, high levels of nonnitrogen-contain-
ing defenses reflect an accumulation ofexcess carbon production. For high plasticitygenotypes, the CNB hypothesis assumes thatcarbon surplus is “cheap” (Bryant et al. 1983),and so predicts that “secondary compoundsare not costly” (Tuomi et al. 1988:67,1991:89). This refers to the production of sec-ondary metabolites from carbon surplus,however.
Importantly, the amount of defense abovethe baseline genetic level will reflect the car-bon surplus, whereas specific types of allelo-chemicals presumably reflect, in part, selec-tion over time for functions such as defenseagainst pathogens and herbivores. Conse-quently, the amount and array of secondarymetabolites are not expected to map preciselyto function (Tuomi et al. 1988, 1991). Fur-thermore, the CNB hypothesis does not pre-dict which allelochemicals will be produced,but because the limiting nutrient is usuallynitrogen, the carbon:nutrient balance of aplant determines the production and accu-mulation of nonnitrogenous and nitrogenousdefenses (Tuomi et al. 1988).
The CNB hypothesis provides specific pre-dictions about when plants may respond todamage by increasing production of second-ary metabolites (Tuomi et al. 1988). Thenutrient stress view of induction, derivedfrom the CNB hypothesis, is that herbivorycan disrupt the carbon:nutrient balance,which may lead to a nonspecific accumula-tion of nonnitrogen-containing defenses andan associated decline in leaf protein (Bryantet al. 1983; Tuomi et al. 1984). That is, thereis an optimal balance between internalreserves of carbon and mineral nutrients thatpromotes maximal growth (Chapin 1980;Ingestad 1982). Defoliation reduces a plant’sability to gain carbon and may affect carbonand nutrient stores. Consequently, the car-bon:nutrient balance may be altered.
The alteration may or may not result inaccumulation of secondary metabolites(Tuomi et al. 1991). Whether an accumula-tion of nonnitrogen-containing defensesoccurs depends on where carbon is storedand the availability of soil nutrients (Tuomiet al. 1988). For example, evergreen treesstore carbon reserves in their leaves, and thusdamage to leaves (because it removes carbon

36 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
reserves) should result in a lower car-bon:nutrient ratio and, even with high nutri-ent availability, these trees would not have car-bon reserves to increase defenses. In contrast,deciduous trees store carbon reserves instems and roots, and damage to their leavesshould result in a marked and long-termincrease in nonnitrogen-containing defenseswhen nutrients are in short supply, but less sowhen nutrients are more plentiful and thepredamage carbon:nutrient ratio can berestored quickly. In these scenarios, thechange in defense is simply passive deterio-ration of leaf quality due to an altered car-bon:nutrient ratio for the plant. Conse-quently, long-term induction can occur overa few years, followed by a gradual relaxationover a few years, which may reflect the popu-lation dynamics of the herbivores (Tuomi etal. 1991). Of course, whether a plant geno-type exhibits these patterns depends on thedegree of phenotypic plasticity in defense.
Just as there has been confusion about theassumptions of the CNB hypothesis, there hasbeen confusion about the predictions. Anexample of that is the misleading statementthat “the predictions of this theory for a rap-idly induced response involving the putativelynitrogen-intensive metabolite nicotine havebeen unambiguously falsified” (Karban andBaldwin 1997:76–77). This is misleading inthree respects. First, the CNB hypothesis doesnot make predictions about wound-inducedshort-term rapid responses: “The carbon/nutrient balance explanations of inducedresponses are most unlikely in conditionswhere the responses are triggered by chemi-cal cues . . . in wound-induced short-termresponses” (Tuomi et al. 1991:99); “specificdefensive responses are associated with theenzymatic regulation of secondary metabo-lism, whereas the plant carbon/nutrient bal-ance mainly affects the amount of precursorsand substrates available to the synthesis of sec-ondary metabolites” (p 99); and “short-termas well as long-term inducible responses thatspread from attacked to nonattacked parts ofthe plant can less likely be attributed tochanges in the carbon/nutrient balance” (p98). Second, in the study that Karban andBaldwin (1997) referred to, Ohnmeiss andBaldwin state, “[W]e interpret C/N theory to
predict that as the internal C:N ratioincreases there should be a concomitantdecrease in the allocation to nitrogen-basedsecondary metabolites in order for RGR to bemaximized” (1994:996). But actually what theCNB hypothesis predicts is that, relative toconstitutive defense, plant species can havesome combination of fixed and flexible allo-cation, varying between having completefixed allocation to complete flexible alloca-tion. Consequently, data gathered to testOhnmeiss and Baldwin’s (1994) predictioncannot be evaluated without knowing whatthe fixed and flexible defense allocation pat-terns are for the test species. These examplesof misapplication are quite typical of theresearch on the CNB hypothesis.
Evidence
Numerous studies (200�; e.g., 147 cited byKoricheva et al. 1998) have been used to exam-ine the CNB hypothesis. Overall, the resultsare equivocal. For instance, the evidence forthe CNB hypothesis consists of some studiesthat show that fertilization can decrease con-centration of nonnitrogen-containing second-ary metabolites and increase concentration ofnitrogenous secondary metabolites, and thatshade can decrease nonnitrogen-containingdefenses and increase nitrogenous defenses.Just as many studies have been interpreted ascontradictory evidence.
Part of the problem with interpretation ofthe results of these studies is a misunderstand-ing of the hypothesis and, consequently, mis-statement of the predictions. For example, arecent review stated, “In an early test of CNB,Bryant and coworkers determined that a slow-growing tree species adapted to a resource-limited environment did not respond as pre-dicted (Bryant et al. 1987)” (Hamilton et al.2001:88). That statement is a misinterpreta-tion of the data and conclusion of Bryant etal. (1987), however, which in fact supportedthe CNB hypothesis. The slow-growing treespecies typically growing in shaded and lessfertile sites was predicted to show little plas-ticity in production of secondary metabolites,with fertilization and shading treatmentscompared to a fast-growing tree species typi-cally growing in sunnier and more fertilesites. The pattern that Bryant at el. (1987)

March 2003 37PLANT DEFENSE HYPOTHESES
found basically fit the prediction. The misin-terpretation by Hamilton et al. (2001) lies inthe expectation that all plants subject to fer-tilization or shading should exhibit a changein the carbon:nutrient ratio that in turn resultsin less nonnitrogen-containing secondary metabo-lites. That is not what the CNB hypothesis pre-dicts. The hypothesis allows for genotypeswith limited or even no phenotypic changesin defense in response to alteration in the car-bon:nutrient ratio (i.e., species with completegenetic determination of defenses and there-fore limited or no plasticity in “constitutive”defense levels) (Bryant et al. 1983; Tuomi etal. 1988, 1991). Importantly, the hypothesisallows for carbon surplus to be allocated tostorage and/or defense.
A meta-analysis of 147 studies suggestedthat the concentration of pooled nonnitro-gen secondary compounds and carbohy-drates responded to nutrients, shade, and car-bon dioxide enrichment as predicted by theCNB hypothesis, but that the concentrationof particular compounds, such as hydrolyza-ble tannins and terpenoids, did not (Kori-cheva et al. 1998). However, such meta-anal-ysis is only conclusive if the individual studiesare free of methodological problems, whichis not the case.
For instance, most studies only examinedthe changes in a few of the nonnitrogenousand nitrogenous phytochemicals. But a mea-sured secondary metabolite may not be cor-related with total secondary metabolite pro-duction (Zangerl and Berenbaum 1987).That is, it may be that the levels of some sec-ondary metabolites are “fixed” genetically,whereas the levels of others are affected byenvironmental conditions and so reflect thecarbon:nutrient ratio (Tuomi et al. 1991).The defensive patterns in the studies thatmade a strong case for having evaluated mostof the major defenses concluded that therewas support both for and against the CNBhypothesis; however, the predictions were interms only of genotypes with phenotypic plas-ticity (Folgarait and Davidson 1994, 1995).Furthermore, none of the studies clearly esti-mated total defense in terms of the car-bon:nutrient ratio of the plant and relative tothe baseline genetic defense, which is what isrequired for a full evaluation of the CNB
hypothesis. There is also the problem of erro-neous conclusions due to measuring defen-sive concentrations (which are a function ofplant biomass) when the issue is allocationpatterns of molecules per plant (Koricheva1999). Then there is the difficulty of measur-ing production of secondary metabolites thatare dynamic or constantly turned over. It hasbeen suggested that concentration of “static”defenses, such as condensed tannins, shouldfit the CNB hypothesis, whereas concentra-tion of more “dynamic” defenses, such as ter-penes, may not (Reichardt et al. 1991). Onlyabout half of the studies with data applicableto the CNB hypothesis (e.g., as cited in Kori-cheva et al. 1998) support the idea that con-centration of static defenses fits the CNBhypothesis. But even so, the CNB hypothesisis about a plant’s carbon surplus; it does not(nor was it formulated to) predict levels ofparticular types of defense (Bryant et al. 1983;Tuomi et al. 1988).
It is difficult to evaluate change in the car-bon to nitrogen resources within a plant. Ina study that tried to examine such changes,with “carbon” being equivalent to starch plusthe nonnitrogen-containing allelochemicalsthat were measured, enhanced CO2 increasedthe “carbon”:nitrogen ratio for aspen, oak,and maple (Kinney et al. 1997). High nitratedecreased the “carbon”:nitrogen ratio foraspen and maple but not for oak. But eventhis approach leaves the question unresolved.
Some of the explanations for the equivocalresults to the CNB hypothesis have beenattributed to the mechanisms of secondarymetabolite production. When an increase indefense requires an increase in both carbonand nitrogen, an increase in defense will notoccur with a change in just carbon or nitro-gen. For example, sufficient enzyme activa-tion is also necessary to convert an accumu-lation of substrate to secondary metabolites(Gershenzon 1994a). No change in the con-centration of a secondary metabolite inresponse to a change in the carbon:nutrientratio may occur anytime there are multinu-trient-requiring structures necessary to syn-thesize, transport, and sequester defenses(Gershenzon 1994b). Furthermore, alloca-tion tradeoffs for carbon and nitrogen prob-ably occur among metabolic pathways and

38 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
even within pathways (Folgarait and Davidson1995; Jones and Hartley 1999). This wouldaffect the amount and type of defenses pro-duced (Muzika 1993). For example, it hasbeen suggested that limited nitrogen affectsphenolic production more negatively thanterpene production (Muzika and Pregitzer1992; Lambers 1993). Also, some chemicaldefenses require a delivery or canal system,which may be made from cells such as latici-fers (Dussourd 1993), and production ofthese would require carbon and nitrogen.
Typically what is left out of the discussionis that phenotypic plasticity in defense due tothe effects of the carbon:nutrient ratio is onlypredicted by the CNB hypothesis for geno-types programmed to put carbon surplus intodefense. The hypothesis allows that some spe-cies will have primarily fixed allocation. Thelevel of secondary metabolite productiondoes seem to be genetically fixed for somespecies (Holopainen et al. 1995). Specifically,in the case of terpenes, it appears that someplant species or populations of plants exhibitmuch less variation in terpene production inresponse to environmental conditions thanothers (Muzika et al. 1989).
Consequently, to test the CNB hypothesis,it is first necessary to establish the baselinegenetic defense. That is, what is the level andarray of defenses at the optimal nutrition andmaximal growth rate for the plant species? Nostudies used to evaluate the CNB hypothesishave established this. To achieve optimalnutrition and maximal growth rate, and thenexamine effects of limiting resources,requires conducting “steady-state” nutritionexperiments (Ingestad 1982, 1991; Ingestadand Lund 1986).
Rapid induction of specific defenses inresponse (within hours followed by relaxationover days or weeks) to herbivory cannot beexplained by the nutrient stress view (car-bon:nutrient balance) (Tuomi et al. 1991),but does fit the defense view (OD hypothe-sis). For example, large amounts of monoter-penes are produced after wounding of grandfir, which are different than the constitutivemonoterpenes; furthermore, the enzymaticmachinery for these induced monoterpenes,which are effective against fungi and herbi-vores, is only apparent several days after
wounding (Gershenzon and Croteau 1991;Gijzen et al. 1992). Rapid induction of spe-cific defenses is clearly not simply chemicaloutcomes from a shift in the carbon:nutrientbalance due to loss of acquired resources, butthen the CNB hypothesis does not predictthat it is.
Thus, studies suggest that neither thenutrient stress view (CNB hypothesis) northe defense view (OD hypothesis) aloneadequately account for patterns of inducedresistance (Haukioja and Neuvonen 1985;Hammerschmidt and Schultz 1995). Thesehypotheses are difficult to test because someinduced chemicals may represent passiveresponse to damage (carbon:nutrient bal-ance), some may reflect active response(selection for defense), some may be involvedin both passive and active responses, and thenof course there may be no induction (whichcan be explained by either the nutrient stressor defense views). Therefore, the emphasisshould be on describing and understandingthe passive, active, and interaction of passive-active responses in plant species chosen forcontrasting patterns. Again, it is particularlyimportant to establish the baseline defense(under optimal nutrition and maximalgrowth conditions) first because without thisframe of reference, it will be difficult (if notimpossible) to evaluate plant responses interms of the CNB hypothesis.
In sum, there is much research purportingto test or provide insight on the CNB hypoth-esis, and there is also some derision of thehypothesis, much of which is flawed by mis-understanding. Although evidence relating tothe CNB hypothesis suggests that the relation-ship between available resources and plantdefense is more complicated than the originalCNB hypothesis predicts, the tests are onsubhypotheses and none have first establisheddefense (fixed and flexible) allocation pat-terns at optimal nutrition and maximalgrowth. The CNB hypothesis brought atten-tion to how resources influence both consti-tutive and induced plant defenses, however,and it contributed to the next hypothesis.
the growth rate hypothesisThe Growth Rate (GR) Hypothesis addresses
genotypic variation in plant defenses, via

March 2003 39PLANT DEFENSE HYPOTHESES
resources shaping inherent growth rate and,subsequently, the constraints on defenseagainst herbivores (Coley et al. 1985). GrowthRate Hypothesis is the term used by Coley(1987a,b); others have referred to it as theResource Availability Hypothesis, but thisterm does not distinguish it from the GDBhypothesis or the OD Allocation Cost ofDefense view (Subhypothesis 2d). The GRhypothesis is that among plant species, asmaximal growth rate (at optimal resources)decreases, the constitutive level of defenseincreases (Figure 1). Note that there is aproblem with the x-axis of Figure 1, in thatbiologically 100% of leaf mass cannot beinvested in defense. Either the curves canonly extend partway across the x-axis (from0 up to some maximal investment that allowsother leaf functions), or the x-axis shouldbe renamed “Defense investment (g actualdefense/g potential possible defense per g leaf ).”Either way, it does not change the basichypothesis. Importantly, plant growth rate isthe balance between a reduction in growthdue to defense costs and an increase ingrowth due to protection from herbivores(Coley et al. 1985).
Assumptions
The Growth Rate (GR) Hypothesis has sev-eral assumptions. It assumes that maximalrelative growth rate is determined by resourceavailability, and so the inherent growth rateof plants matches the resource limitation inthe preferred habitat. Data support this(Coley et al. 1985). The GR hypothesisassumes that herbivory is an important selec-tive force for production of secondary metab-olites and that defense has a cost. Data againsupport this (discussed in an earlier section).It assumes that herbivores consume plantmass as a function of herbivore mass(referred to as a “fixed amount”) rather thanas a fixed percentage of plant productivity,which is a reasonable assumption. It alsoassumes that slow turnover of plant parts isadvantageous in low-nutrient environmentsand fast turnover of plant parts is advanta-geous in high-nutrient environments. Datasupport this (Coley et al. 1985). The idea isthat fast-growing species have short-livedleaves because energy acquisition in nutrient-
rich environments is maximized by rapid leafturnover, whereas slow-growing species havelong-lived leaves because slow turnover isadvantageous in a nutrient-poor environmentwhere (re)growth is constrained.
Predictions
The basic prediction is that among plantspecies, as maximal growth rate (i.e., intrinsicgrowth rate at optimal resources) increases,the level of constitutive defense shoulddecrease. A second prediction is that fast-growing species exhibit more sharply definedpeaks in the defense investment versus real-ized growth-rate curves, which means that“deviations from the optimal defense levelshave a larger negative impact on realizedgrowth rate than they would for slow-growingspecies” (Coley et al. 1985:897; Figure 1).Optimal defense level would refer to theexpression of defenses at the maximal growthrate with optimal nutrition for each plant spe-cies. In practice, the predictions have beenreduced to: 1) in high resource environments(e.g., agroecosystems, old-field habitats),competition favors fast-growing plant species,and so to grow fast means allocating little todefense; and 2) in low resource environments(e.g., dry or shaded habitats), resource limi-tation favors slow-growing plant species,which should allocate more to defense (Coleyet al. 1985). This latter group cannot com-pensate easily for herbivory because of theirslow growth, so replacement of resources lostto herbivores is more costly for plants in lowresource environments.
Coley et al. (1985) make some additionalpredictions. One prediction is that, via moreinvested in defense, the actual level of herbi-vory experienced by slow-growing species isless than that of fast-growing species. If fast-growing species are subject to more herbivorybecause the constitutive defensive level is rela-tively low, then another prediction is that fast-growing species have greater defensive plas-ticity than slow-growing species. Fast-growingspecies are predicted to have defenses thatinclude more secondary metabolites that aremobile, have high turnover rates, and thushave a reversible commitment to defense. Incontrast, slow-growing species are predictedto have immobile defenses and, thus, a fixed
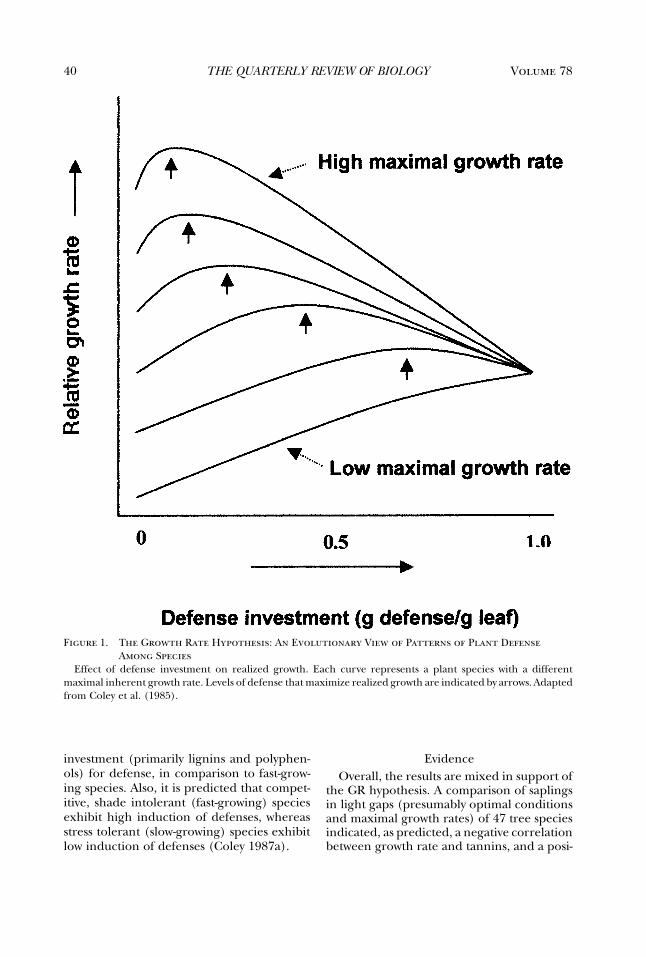
40 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
Figure 1. The Growth Rate Hypothesis: An Evolutionary View of Patterns of Plant DefenseAmong Species
Effect of defense investment on realized growth. Each curve represents a plant species with a differentmaximal inherent growth rate. Levels of defense that maximize realized growth are indicated by arrows. Adaptedfrom Coley et al. (1985).
investment (primarily lignins and polyphen-ols) for defense, in comparison to fast-grow-ing species. Also, it is predicted that compet-itive, shade intolerant (fast-growing) speciesexhibit high induction of defenses, whereasstress tolerant (slow-growing) species exhibitlow induction of defenses (Coley 1987a).
EvidenceOverall, the results are mixed in support of
the GR hypothesis. A comparison of saplingsin light gaps (presumably optimal conditionsand maximal growth rates) of 47 tree speciesindicated, as predicted, a negative correlationbetween growth rate and tannins, and a posi-

March 2003 41PLANT DEFENSE HYPOTHESES
tive correlation between growth rate and rateof herbivore damage to leaves (Coley 1987a).Growth rate was estimated in three ways:annual increase in height, annual leaf pro-duction, and highest individual tree growthrate per species (presumably an estimate ofmaximal growth rate). As predicted, specieswith long-lived leaves (i.e., slow-growing spe-cies) had higher concentrations of tanninsthan species with short-lived leaves (i.e., fast-growing species). But there was no negativecorrelation between rate of herbivore dam-age and tannin concentration. A comparisonof pioneer versus gap tree species provideddata both in support of and against the GRhypothesis (Folgarait and Davidson 1994,1995). Tests within species that comparedmale and female plants, which have differentgrowth rates, supported the GR hypothesis( Jing and Coley 1990; Herms and Mattson1992).
Other examples clearly do not fit the GRhypothesis. For instance, fast-growing earlysuccessional conifer species (pines) havehigh constitutive levels of defense with rela-tively low induction, whereas slower-growinglate successional conifer species (firs) lack themassive constitutive defenses of pine and relyon fast-acting massive increases in defense atthe wound site (Cates 1996). Furthermore, ithas become clear that secondary metabolitescannot simply be classified in terms of type orperceived function. Terpenes, for example,have been thought of as mobile defenses and,therefore, as appropriate defense for short-lived leaves, when actually they have traitsappropriate for both short- and long-livedleaves. Although relatively expensive to pro-duce, terpenes are cheap to maintain andmobile prior to senescence (Gershenzon1994a), and concentrations can be highenough to produce dosage or “quantitative”effects (Langenheim 1994).
Furthermore, the predictions about theshape of the defense investment versus real-ized growth-rate curves, degree of defensiveplasticity, and degree of induction have notbeen examined. In particular, while somesubhypotheses have been tested, data havenot been gathered to examine the hypothesismore directly. The curves of defense invest-ment versus realized growth rate are specified
mathematically (Figure 1), so in principlesuch a figure could be generated by compar-ison of plant species. But there are no studiesof the GR hypothesis taking that approach.To do this would require estimating the max-imal growth rate (at optimal resources foreach species) and the corresponding consti-tutive defense level, plus the consequences todefense level with deviations from the maxi-mal growth rate. Without such an assessment,it will be difficult to evaluate the GR hypoth-esis fully.
In sum, the research related to the GRhypothesis makes a compelling case that evo-lutionarily the effects of resource availabilityon growth are orders of magnitude morethan the effects of defense cost (Coley1987b). Furthermore, it indicates that intrin-sic growth rate and correlated characteristics,such as leaf life span, are so strongly shapedby abiotic conditions that these determinebasic defensive profiles.
the growth-differentiationbalance hypothesis
The Growth-Differentiation Balance (GDB)Hypothesis provides a framework for predict-ing how plants will balance allocation betweendifferentiation-related processes and growth-related processes over a range of environ-mental conditions (Loomis 1932, 1953).Growth refers to the production of roots,stems, and leaves, or any process that requiressubstantial cell division and elongation. Dif-ferentiation is everything else, so it refers toenhancement of the structure or function ofexisting cells (i.e., maturation and speciali-zation). Some differentiation traits can limitherbivory. Secondary metabolism, trichomeproduction, and thickening of leaf cuticle areexamples of those kinds of differentiation-related processes (Herms and Mattson 1992).Allocation to differentiation includes processand product, so it includes cost of enzymes,transport, and storage structures involved indefense. Research shows that growth pro-cesses and secondary metabolism can com-pete for available photosynthate (Wadleigh etal. 1946; Veihmeyer and Hendrickson 1961;Mooney and Chu 1974) and, thus, full carbonallocation to all functions cannot be metsimultaneously (Lorio 1986).

42 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
The GDB hypothesis states that any envi-ronmental factor that slows growth more thanit slows photosynthesis can increase theresource pool available for allocation to dif-ferentiation-related products (Loomis 1932,1953). For instance, growth is slowed substan-tially by shortages of nutrients and water,whereas photosynthesis is less sensitive tosuch limitations (reviewed by Herms andMattson 1992). In these situations, carbohy-drates accumulate in excess of growthdemands and, consequently, may be con-verted to secondary metabolites, with low costto plant fitness.
Although it was Loomis (1932, 1953) whooriginally outlined the Growth-Differentia-tion Balance Hypothesis for plants, it wasHerms and Mattson (1992) who used thehypothesis to explain how the physiologicaltradeoff between growth and differentiationprocesses interacts with the selective forces ofcompetition and herbivory to shape plantlife-history strategies. They proposed agrowth-differentiation continuum reflectingthe allocation of resources to these processes.Competition in resource-rich environmentsselects for a growth-dominated strategy,whereas stress of resource-poor environmentsselects for differentiation-dominated strategy.
Assumptions
The expanded GDB hypothesis assumesthat: 1) resource limitation (except for light)has a more negative effect on growth than onphotosynthesis; 2) maximal relative growthrate is determined by resource availabilityand, consequently, the inherent growth rateof plant species reflects the resource limita-tion of the preferred environment; 3) thereis a tradeoff between growth and differentia-tion due to competition for photosynthate;4) herbivory is a major selective force for pro-duction of secondary metabolites; 5) defensesreduce herbivory; and 6) defenses can have acost because they can divert resources fromgrowth.
Predictions
Ecologically, the GBD hypothesis predicts:1) Plants experiencing low levels of resourcesshould be limited in both growth and pho-
tosynthetic capability. Limited resourcesshould be shunted preferentially to growthprocesses over differentiation processes (War-ing and Pitman 1985). The limitation ofgrowth processes would yield low growth rateand moderate concentrations of secondarymetabolites (Herms and Mattson 1992; seeFigure 2, point A). 2) Plants experiencingintermediate resource availability shouldhave high concentrations of secondarymetabolites but an intermediate accumula-tion of biomass (Loomis 1932, 1953). At anintermediate resource level, growth (throughcell division and enlargement) is limited bythe resource level, whereas photosynthesis isless affected (Chapin 1980; Korner 1991;Luxmoore 1991). Therefore, secondarymetabolites will tend to accumulate, due tothe excess pool of photosynthate (or assimi-lates), and the defenses will be produced rela-tively inexpensively (Figure 2, point B). 3)Plants experiencing high resource availabilityshould not be limited by photosynthesis orgrowth and, thus, should allocate a greaterproportion of the photosynthate (or assimi-lates) to growth rather than to differentiationtraits (Loomis 1932; Herms and Mattson1992) (Figure 2, point C). Overall, the pat-tern of allocation to secondary metabolitesshould be curvilinear across a resource gra-dient, with a peak at intermediate resourcelevels. This predicted pattern contrasts withthat of the OD Allocation Cost of PhenotypicDefense (Hypothesis 2d), which is that allo-cation to defense increases across a gradientof increasing resources. It is also similar to thepredicted pattern of the CNB hypothesis, butwith a different explanation. 4) The excep-tion to the GDB prediction is the effect oflight, which influences photosynthesis morethan growth. In this case, with increasinglight, secondary metabolites will increase pro-portionally with growth.
It might be useful to recast the ecological-side GDB graph to establish the real issues(Figure 3). Growth is measured as mass (oras a correlate of mass, such as height), butmass is composed of undifferentiated and dif-ferentiated products. Some of the differenti-ated items functionally belong in the“defense” category (e.g., glandular tri-chomes, secretory glands, resin ducts), others
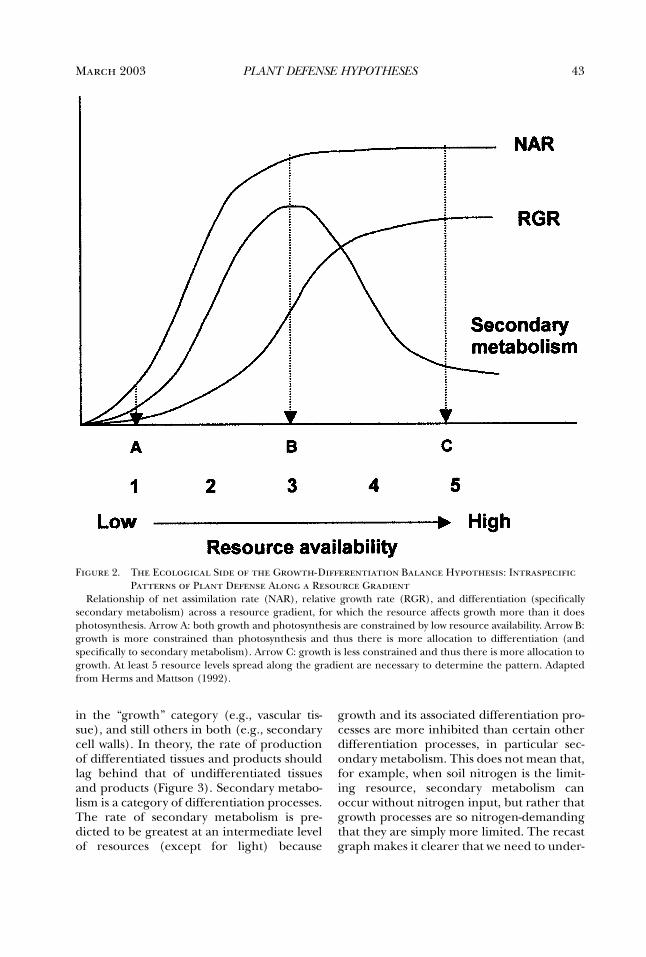
March 2003 43PLANT DEFENSE HYPOTHESES
Figure 2. The Ecological Side of the Growth-Differentiation Balance Hypothesis: IntraspecificPatterns of Plant Defense Along a Resource Gradient
Relationship of net assimilation rate (NAR), relative growth rate (RGR), and differentiation (specificallysecondary metabolism) across a resource gradient, for which the resource affects growth more than it doesphotosynthesis. Arrow A: both growth and photosynthesis are constrained by low resource availability. Arrow B:growth is more constrained than photosynthesis and thus there is more allocation to differentiation (andspecifically to secondary metabolism). Arrow C: growth is less constrained and thus there is more allocation togrowth. At least 5 resource levels spread along the gradient are necessary to determine the pattern. Adaptedfrom Herms and Mattson (1992).
in the “growth” category (e.g., vascular tis-sue), and still others in both (e.g., secondarycell walls). In theory, the rate of productionof differentiated tissues and products shouldlag behind that of undifferentiated tissuesand products (Figure 3). Secondary metabo-lism is a category of differentiation processes.The rate of secondary metabolism is pre-dicted to be greatest at an intermediate levelof resources (except for light) because
growth and its associated differentiation pro-cesses are more inhibited than certain otherdifferentiation processes, in particular sec-ondary metabolism. This does not mean that,for example, when soil nitrogen is the limit-ing resource, secondary metabolism canoccur without nitrogen input, but rather thatgrowth processes are so nitrogen-demandingthat they are simply more limited. The recastgraph makes it clearer that we need to under-
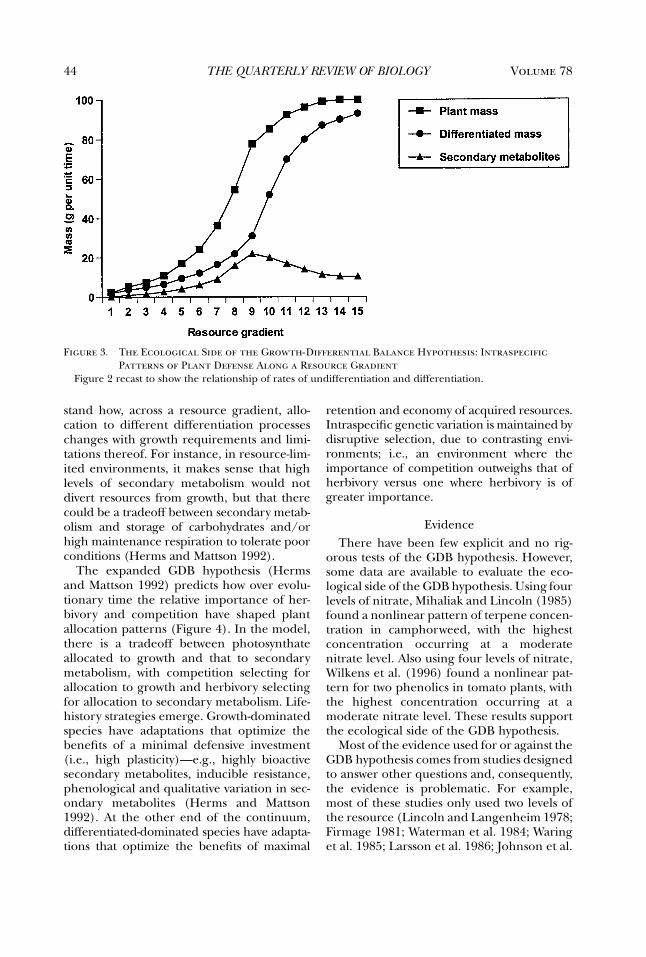
44 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
Figure 3. The Ecological Side of the Growth-Differential Balance Hypothesis: IntraspecificPatterns of Plant Defense Along a Resource Gradient
Figure 2 recast to show the relationship of rates of undifferentiation and differentiation.
stand how, across a resource gradient, allo-cation to different differentiation processeschanges with growth requirements and limi-tations thereof. For instance, in resource-lim-ited environments, it makes sense that highlevels of secondary metabolism would notdivert resources from growth, but that therecould be a tradeoff between secondary metab-olism and storage of carbohydrates and/orhigh maintenance respiration to tolerate poorconditions (Herms and Mattson 1992).
The expanded GDB hypothesis (Hermsand Mattson 1992) predicts how over evolu-tionary time the relative importance of her-bivory and competition have shaped plantallocation patterns (Figure 4). In the model,there is a tradeoff between photosynthateallocated to growth and that to secondarymetabolism, with competition selecting forallocation to growth and herbivory selectingfor allocation to secondary metabolism. Life-history strategies emerge. Growth-dominatedspecies have adaptations that optimize thebenefits of a minimal defensive investment(i.e., high plasticity)—e.g., highly bioactivesecondary metabolites, inducible resistance,phenological and qualitative variation in sec-ondary metabolites (Herms and Mattson1992). At the other end of the continuum,differentiated-dominated species have adapta-tions that optimize the benefits of maximal
retention and economy of acquired resources.Intraspecific genetic variation is maintained bydisruptive selection, due to contrasting envi-ronments; i.e., an environment where theimportance of competition outweighs that ofherbivory versus one where herbivory is ofgreater importance.
EvidenceThere have been few explicit and no rig-
orous tests of the GDB hypothesis. However,some data are available to evaluate the eco-logical side of the GDB hypothesis. Using fourlevels of nitrate, Mihaliak and Lincoln (1985)found a nonlinear pattern of terpene concen-tration in camphorweed, with the highestconcentration occurring at a moderatenitrate level. Also using four levels of nitrate,Wilkens et al. (1996) found a nonlinear pat-tern for two phenolics in tomato plants, withthe highest concentration occurring at amoderate nitrate level. These results supportthe ecological side of the GDB hypothesis.
Most of the evidence used for or against theGDB hypothesis comes from studies designedto answer other questions and, consequently,the evidence is problematic. For example,most of these studies only used two levels ofthe resource (Lincoln and Langenheim 1978;Firmage 1981; Waterman et al. 1984; Waringet al. 1985; Larsson et al. 1986; Johnson et al.
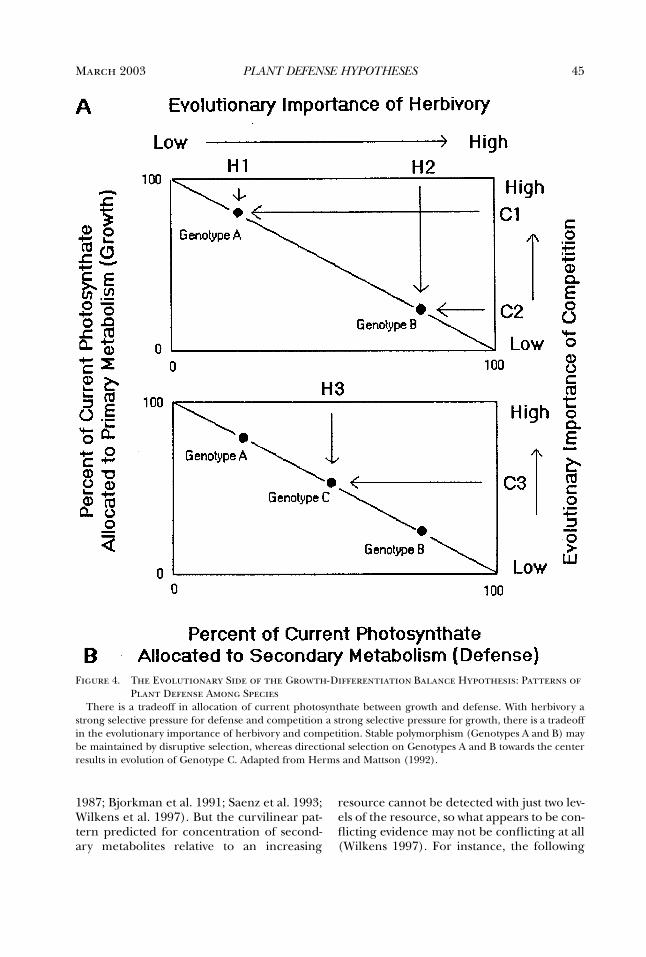
March 2003 45PLANT DEFENSE HYPOTHESES
Figure 4. The Evolutionary Side of the Growth-Differentiation Balance Hypothesis: Patterns ofPlant Defense Among Species
There is a tradeoff in allocation of current photosynthate between growth and defense. With herbivory astrong selective pressure for defense and competition a strong selective pressure for growth, there is a tradeoffin the evolutionary importance of herbivory and competition. Stable polymorphism (Genotypes A and B) maybe maintained by disruptive selection, whereas directional selection on Genotypes A and B towards the centerresults in evolution of Genotype C. Adapted from Herms and Mattson (1992).
1987; Bjorkman et al. 1991; Saenz et al. 1993;Wilkens et al. 1997). But the curvilinear pat-tern predicted for concentration of second-ary metabolites relative to an increasing
resource cannot be detected with just two lev-els of the resource, so what appears to be con-flicting evidence may not be conflicting at all(Wilkens 1997). For instance, the following

46 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
results would all be supportive of the GDBhypothesis: 1) a positive correlation betweenconcentration of secondary metabolites andlow versus intermediate resource levels (Fig-ure 2, points A and B); 2) a neutral patternacross low versus high resource levels (Figure2, points A and C); and 3) a negative corre-lation across intermediate versus highresource levels (Figure 2, points B and C).Even results from studies with three levels ofa resource can be difficult to interpret. Forexample, Muzika et al. (1989) found terpeneconcentration for a fir species was notaffected by soil nitrogen, but it is not clearwhere the three nitrogen levels would fallalong the ideal range for testing the GDBhypothesis (i.e., relative to the plant speciesand the experimental conditions). To detecta potential curvilinear pattern for plantdefense, Wilkens (1997) recommends a min-imum of five levels spread along an appro-priate gradient of the resource.
The GDB hypothesis has been applied towithin-plant patterns of defense allocation. Itwas suggested that terpene biosynthesisoccurring primarily in young rapidly growingleaves contradicts the GDB hypothesis (Ler-dau et al. 1994). But consideration of cellulargrowth and differentiation processes inyoung leaves indicates that it is not a contra-diction. The epidermal cells, which mature(i.e., differentiate) relatively early in leaves,often contain high levels of secondary metab-olites and most likely account for the high ter-pene levels (Herms and Mattson 1992).
The only analysis of the evolutionary sideof the GDB hypothesis is the discussion byHerms and Mattson (1992). No tests havebeen reported.
Relationship of These HypothesesThese hypotheses are not mutually exclu-
sive (Scriber and Ayres 1988; Jing and Coley1990; Herms and Mattson 1992; Tuomi 1992;Berenbaum 1995), and it is appropriate tointegrate them (Price 1991; Herms and Matt-son 1992; Tuomi 1992; Mole 1994).
comparison and integration of the odand gr hypotheses
In the GR hypothesis, herbivory is comple-mentary to the selective pressure of resources
(Coley et al. 1985) rather than being the driv-ing force as in the OD view (Rhoades 1979).Furthermore, in the OD framework the totalselective pressure from herbivores of differ-ent feeding specializations can vary for appar-ent and unapparent plants, and thereforedetermine the amount and type of defenseexhibited by a plant (Feeny 1975, 1976;Rhoades and Cates 1976; Rhoades 1979). Incontrast, in the GR framework, the interac-tion of resource availability and total herbi-vory, regardless of plant apparency or herbi-vore specialization, determines the amountand type of defense (Coley et al. 1985).
Recognizing that unapparency as an escapestrategy probably only applies to ruderal spe-cies, Coley (1987a) integrated the OD andGR hypotheses by creating the Habitat Tem-plate-Plant Defense (HT-PD) Hypothesis. TheHT-PD hypothesis is based on a conceptualmodel of plant strategies shaped by the levelof disturbance and stress (Grime 1977, 1979;Southwood 1977) and relates defense tointrinsic growth rate (Coley 1987a). The HT-PD hypothesis is certainly intriguing, but untilquite recently (Hodgson et al. 1999) it wasdifficult to know (and still is) where to assignparticular species relative to the disturbanceand stress axes.
The HT-PD integration did not resolve adiscrepancy in predictions about types ofdefense. Based on the OD hypothesis, it waspredicted that quantitative defenses (such astannins) are more costly than qualitativedefenses (such as alkaloids and terpenes),reflecting the differences in molecular sizeand concentration in a plant (Feeny 1975,1976; Rhoades and Cates 1976). In contrast,based on the GR hypothesis, it was predictedthat qualitative defenses are more costly thanquantitative defenses because qualitativedefenses are continually produced andmetabolized, whereas quantitative defensesare produced and then kept intact (Coley etal. 1985). Effort to resolve this is probablypointless, however, because it is clear that costof a defensive chemical is not as simple asthese predictions imply.
comparison and integration of cnband gr
The difference between the CNB and theGR hypotheses is that the GR hypothesis spe-

March 2003 47PLANT DEFENSE HYPOTHESES
cifically incorporates the effects of herbivoryand competition to select for optimal defense,whereas the CNB hypothesis addresses phe-notypic expression of the genetic potential fordefense. Thus, the CNB hypothesis needs theGR hypothesis to explain genotypic patterns.The GR hypothesis relies on the CNB hypo-thesis for predictions about defenses pheno-typically (e.g., Folgarait and Davidson 1994,1995).
Together the GR and CNB hypothesesexplain what otherwise would seem like acontradiction. Among tree species, shade-tolerant species have higher concentrations ofnonnitrogen-containing defenses than gap orpioneer species, whereas within species,shaded individuals have lower concentrationsof nonnitrogen-containing defenses thanthose in full sunlight (Bryant et al. 1983: sec-tions 2 & 6; Coley 1987a). Similarly, it takesthese two hypotheses to explain the differ-ence in inter- and intraspecific defensive pat-terns relative to nitrogen availability. That is,interspecific variation in defense reflects adap-tations to herbivory and absolute resource lev-els, whereas intraspecific variation in defensereflects relative resource levels or the car-bon:nutrient balance (Coley 1987a).
comparison of cnb-gr versus gdbAt this point, it may seem that the integra-
tion of the CNB and GR hypotheses yields theGDB hypothesis. That is not the case, however.
First, the GDB hypothesis predicts that anyenvironmental factor that slows growth morethan it slows photosynthesis can increase theresource pool available for allocation to sec-ondary metabolism (Loomis 1932, 1953).Thus, the GBD hypothesis goes beyond theCNB hypothesis (Herms and Mattson 1992),which focuses on the effects of shade and fer-tilization on allocation to secondary metabo-lism versus growth (Bryant et al. 1983).
Second, the GDB hypothesis explicitlyacknowledges that in plant developmentthere is a constant tradeoff between growthand differentiation requirements. Differentia-tion continually diverts resources from plantgrowth (e.g., away from production of new leafarea). Growth continually diverts resourcesfrom new differentiation (e.g., away frommaturation and specialization of tissue). For
instance, secondary metabolism and struc-tural reinforcement are physiologically con-strained in dividing and enlarging cells, andresin ducts and other compartmentalizationof defenses depend on cell growth. There-fore, the phenotypic expression of a plant’sgenetic potential for defense will reflect thiscontinual tug-of-war between growth and dif-ferentiation processes. When environmentalconditions are optimal for a plant species(maximal growth rate), vegetative growth(plus differentiation to support that) isexpected to receive priority for resources oversecondary metabolism and storage. Whenenvironmental conditions are unfavorablefor growth, the carbon pool available for allo-cation to secondary metabolism increasessuch that there is less or even no tradeoff withgrowth.
Third, the mathematical model of the GRhypothesis (Coley et al. 1985) assumes that allresource allocation that does not go intodefense goes to growth, but that is unrealisticbiologically (Moles 1994). The mathematicalmodel of the GDB hypothesis (Herms andMattson 1992) recognizes that phenotypic(“realized”) growth rate is a function ofintrinsic defense investment, competition,herbivory, and environment, and associatedtradeoffs including compensatory regrowth,and thus incorporates third party tradeoffs(Mole 1994). Therefore, both the CNB andGR hypotheses need the GDB hypothesis toaccount for the tradeoff complexitiesbetween growth and differentiation, and inparticular to include the role of tolerance.
Fourth, because plants have so many routesof entry and egress of limiting resources, eval-uation of resource-based tradeoff models willrequire physiological and molecular genetictechniques to isolate and follow processes(Mole 1994). For instance, it appears thatdownregulation of growth rate, beyond thatneeded due to immediate resource require-ments of activated defense, allows plants toreallocate resources, for example, toregrowth processes, which would increase tol-erance to herbivory (Baldwin and Hamilton2000). The GDB hypothesis provides a frame-work for identifying cellular and tissuegrowth-differentiation and third-party trade-offs for study of plant defense issues.
48 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
There is limited utility in comparing thegraphical models of the CNB, GR, and GDBhypotheses, however, even though all threehave growth rate on the y-axis. The CNBgraph illustrates the extreme situation, whereinall carbon surplus is shunted to defense, anddefense is determined completely by that “phe-notypic plasticity” (Figure 5). The ecological-side GDB graph is too simplistic (Figure 2).The exact form of the tradeoff betweengrowth and secondary metabolism has notbeen established for any species (Herms andMattson 1992). Furthermore, the graph failsto capture the intimacy of growth and differ-entiation processes, and the implications ofthat for withstanding herbivory, which is thekey contribution of the GDB hypothesis. TheGR graph illustrates “defense investment” onthe x-axis, but it does not suggest the degreeto which that investment comes from fixedand/or flexible allocations (Figure 1). It isalso difficult to incorporate tolerance into“defense investment” because tolerancerefers to more than storage reserves; it also
refers to processes, such as regulation of pho-tosynthetic rate and nutrient uptake.
comparsion of the od and expandedgdb hypotheses
As discussed by Herms and Mattson (1992),the expanded GDB hypothesis also addressesthe evolutionary aspects of the OD and GRhypotheses. The expanded GDB hypothesisincludes a model of the evolution of plantallocation trajectories that takes into accountthe selective pressures of herbivory and com-petition (Figure 4). In contrast to the ODview, the GDB explanation resides in recog-nition of the physiological tradeoffs betweengrowth and differentiation (and whichincludes third party tradeoffs) at the cellularand tissue levels relative to the selective pres-sures of resource availability. Predictions canbe made and tested. Therefore, by subsumingthe other hypotheses and providing explicitand logically compatible predictions, theexpanded GDB is the most theoreticallymature of the hypotheses of plant defense.
Figure 5. The Carbon:Nutrient Balance Hypothesis: An Ecological View of Intraspecific Patternsof Total Plant Defense Along a Resource Gradient
Model of plant carbon:nutrient balance in which secondary metabolite production is supported by carbonsurplus. Ct � total available carbon; Cg � carbon diverted to growth; Ce � excess carbon; and Cd � level of“carbon-based” allelochemicals. In this extreme situation, all carbon surplus is shunted to defense, and defenseis determined completely by that “phenotypic plasticity.” Adapted from Tuomi et al. (1988).

March 2003 49PLANT DEFENSE HYPOTHESES
Conclusions About the Theory ofPlant Defense
None of the plant defense hypotheses haveever been firmly rejected (Herms and Matt-son 1992; Berenbaum 1995). Consideringhow theory develops, this is not surprising.The theory of plant defense has the charac-teristics of immature theory. To expect thatwe would have a mature theory at this pointis unreasonable.
So where do we go from here? The currenttheory of plant defense addresses the diverseand ubiquitous ways that plants cope withenemies, in particular why some plant speciesare so well defended and others not so, but itis in danger of stagnation (and so remainingin a state of confusion) if hypotheses are notrigorously tested. Each of the hypotheses hascontributed to our current understandingand could contribute more, but we need torecognize that each has its limitations andeach has different kinds of contributions.However, while the basic OD hypothesesserved a useful purpose at the outset, they areno longer appropriate as the focal frameworkfor investigation because the tendency is that,whatever the results of a test, they can be (and
have been) interpreted as supporting the ODview. In contrast, the subhypotheses of theOD view are more testable, but they shouldbe referred to with specific names that distin-guish them from the two basic OD hypothe-ses. In particular, focus on cost per se will beless useful than that on mechanisms of cost,such as for ecological costs. Any use of theCNB and GR hypotheses should clearly andfairly address the crux of those hypotheses (asoutlined herein). Further research on thesubhypotheses of the CNB and GR hypothe-ses is unlikely to be productive, however. Theexpanded GDB hypothesis, which subsumesthe other plant defense hypotheses (Hermsand Mattson 1992) and, furthermore, is themost mature of the hypotheses, should betested carefully so that we can determine thedegree to which it is a useful hypothesis.
acknowledgmentsThanks to Deane Bowers, Daniel Herms, Rick Karban,Peter Lorio, Tod Osier, John Titus, Rich Wilkens, andan anonymous reviewer for comments on one versionor another of this evolved manuscript. The author’sresearch was supported by USA National ScienceFoundation grant DEB 9726222 and USDA grant NRI98–35302–6878.
REFERENCES
Abrams P A. 1986. Adaptive responses of predators toprey and prey to predators: the failure of the arms-race analogy. Evolution 40:1229–1247.
Adler F R, Karban R. 1994. Defended fortresses ormoving targets?: another model of inducibledefenses inspired by military metaphors. AmericanNaturalist 144:813–832.
Agrawal A A. 1998. Induced responses to herbivory andincreased plant performance. Science 279:1201–1202.
Agrawal A A. 2000. Specificity of induced resistance inwild radish: causes and consequences for two spe-cialist and two generalist caterpillars. Oikos 89:493–500.
Agrawal A A, Karban R. 1999. Why induced defensesmay be favored over constitutive strategies inplants. Pages 45–61 in The Ecology and Evolution ofInducible Defenses, edited by R Tollrian and C D Har-vell. Princeton (NJ): Princeton University Press.
Agren J, Schemske D W. 1993. The cost of defenseagainst herbivores: an experimental study of tri-chome production in Brassica rapa. American Nat-uralist 141:338–350.
Baas W J. 1989. Secondary plant compounds, their
ecological significance and consequences for thecarbon budget. Pages 313–340 in Causes and Con-sequences of Variation in Growth Rate and Productivityof Higher Plants, edited by H Lambers et al. TheHague (The Netherlands): SPB Academic Publish-ing.
Baldwin I T. 1998. Jasmonate-induced responses arecostly but benefit plants under attack in native pop-ulations. Proceedings of the National Academy of Sci-ences of the United States of America 95:8113–8118.
Baldwin I T, Hamilton W, III. 2000. Jasmonate-inducedresponses of Nicotiana sylvestris results in fitnesscosts due to impaired competitive ability for nitro-gen. Journal of Chemical Ecology 26:915–952.
Baldwin I T, Karb M J. 1995. Plasticity in allocation ofnicotine to reproductive parts in Nicotiana attenu-ata. Journal of Chemical Ecology 21:897–909.
Baldwin I T, Ohnmeiss T E. 1994. Swords into plow-shares?: Nicotiana sylvestris does not use nicotine asa nitrogen source under nitrogen-limited growth.Oecologia 98:385–392.
Baldwin I T, Schmelz E A. 1994. Constraints on aninduced defense: the role of leaf area. Oecologia97:424–430.

50 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
Baldwin I T, Sims C L, Kean S E. 1990. The reproduc-tive consequences associated with inducible alka-loidal responses in wild tobacco. Ecology 71:252–262.
Berenbaum M. 1980. Adaptive significance of midgutpH in larval lepidoptera. American Naturalist115:138–146.
Berenbaum M. 1981. Patterns of furanocoumarin dis-tribution and insect herbivory in the umbelliferae:plant chemistry and community structure. Ecology62:1254–1266.
Berenbaum M. 1983. Coumarins and caterpillars: acase for coevolution. Evolution 37:163–179.
Berenbaum M R. 1995. The chemistry of defense: the-ory and practice. Proceedings of the National Academyof Sciences of the United States of America 92:2–8.
Berenbaum M R, Zangerl A R. 1998. Chemical phe-notype matching between a plant and its insectherbivore. Proceedings of the National Academy of Sci-ences of the United States of America 95:13743–13748.
Bergelson J. 1994. The effects of genotype and theenvironment on costs of resistance in lettuce. Amer-ican Naturalist 143:349–359.
Bergelson J, Purrington C B. 1996. Surveying patternsin the cost of resistance in plants. American Natu-ralist 148:536–558.
Bergelson J, Purrington C B, Palm C J, Lopez-Gutier-rez J-C. 1996. Costs of resistance: a test using trans-genic Arabidopsis thaliana. Proceedings of the RoyalSociety of London Series B 263:1659–1663.
Bergey D R, Howe G A, Ryan C A. 1996. Polypeptidesignaling for plants defensive genes exhibits anal-ogies to defense signaling in animals. Proceedings ofthe National Academy of Sciences of the United States ofAmerica 93:12053–12058.
Bernays E A. 1981. Plant tannins and insect herbi-vores: an appraisal. Ecological Entomology 6:353–360.
Bernays E A, Chamberlain D, McCarthy P. 1980. Thedifferential effects of ingested tannic acid on dif-ferent species of acridoidea. Entomologia Experimen-talis et Applicata 28:158–166.
Bjorkman C, Larsson S, Gref R. 1991. Effects of nitro-gen fertilization on pine needle chemistry and saw-fly performance. Oecologia 86:202–209.
Bilbrough C J, Richards J H. 1993. Growth of sage-brush and bitterbrush following simulated winterbrowsing: mechanisms of tolerance. Ecology 74:481–492.
Briggs M A, Schultz J C. 1990. Chemical defense pro-duction in Lotus corniculatus L. II. Trade-offsamong growth, reproduction and defense. Oecolo-gia 83:32–37.
Broadway R M, Duffey S S, Pearce G, Ryan C A. 1986.Plant proteinase inhibitors: a defense against her-bivorous insects? Entomologia Experimentalis et Appli-cata 41:33–38.
Bryant J P, Chapin F S, III, Klein D R. 1983. Carbon/
nutrient balance of boreal plants in relation to ver-tebrate herbivory. Oikos 40:357–368.
Bryant J P, Chapin F S, III, Reichardt P B, Clausen TP. 1987. Response of winter chemical defense inAlaska paper birch and green alder to manipula-tion of plant carbon/nutrient balance. Oecologia72:510–514.
Cates R G. 1996. The role of mixtures and variationin the production of terpenoids in conifer-insect-pathogen interactions. Pages 179–216 in RecentAdvances in Phytochemistry, Volume 30, edited by JT Romeo et al. New York: Plenum Press.
Chapin F S, III. 1980. The mineral nutrition of wildplants. Pages 233–260 in Annual Review of Ecologyand Systematics, Volume 11, edited by R J Johnstonet al. Palo Alto (CA): Annual Reviews.
Chapin F S, III, McNaughton S J. 1989. Lack of com-pensatory growth under phosphorus deficiency ingrazing-adapted grasses from the Serengeti Plains.Oecologia 79:551–557.
Cipollini D. 1998. Induced defenses and phenotypicplasticity. Trends in Ecology & Evolution 13:200.
Coley P D. 1983. Herbivory and defensive character-istics of tree species in a lowland tropical forest.Ecological Monographs 53:209–233.
Coley P D. 1987a. Interspecific variation in plant anti-herbivore properties: the role of habitat qualityand rate of disturbance. New Phytologist 106 (Sup-plement):251–263.
Coley P D. 1987b. Effects of plant growth rate and leaflifetime on the amount and type of anti-herbivoredefense. Oecologia 74:531–536.
Coley P D, Bryant J P, Chapin F S, III. 1985. Resourceavailability and plant antiherbivore defense. Science230:895–899.
Dawkins R, Krebs J R. 1979. Arms races between andwithin species. Proceedings of the Royal Society of Lon-don Series B 205:489–511.
Dethier V G. 1954. Evolution of feeding preferencesin phytophagous insects. Evolution 8:33–54.
DeWitt T J, Sih A, Wilson D S. 1998. Costs and limitsof phenotypic plasticity. Trends in Ecology & Evolu-tion 13:77–81.
Dirzo R, Harper J L. 1982. Experimental studies onslug-plant interactions. III. Differences in theacceptability of individual plants of Trifolium repensto slugs and snails. Journal of Ecology 70:101–117.
Dussourd D E. 1993. Foraging with finesse: caterpillaradaptations for circumventing plant defenses.Pages 92–131 in Caterpillars: Ecological and Evolu-tionary Constraints on Foraging, edited by N E Stampand T M Casey. New York: Chapman & Hall.
Ehrlich P R, Raven P H. 1964. Butterflies and plants:a study in coevolution. Evolution 18:586–608.
Fagerstrom T, Larsson S, Tenow O. 1987. On optimaldefence in plants. Functional Ecology 1:73–81.
Farrell B D, Mitter C. 1993. Phylogenetic determi-

March 2003 51PLANT DEFENSE HYPOTHESES
nants of insect/plant community diversity. Pages253–266 in Species Diversity in Ecological Communities:Historical and Geographical Perspectives, edited by RE Ricklefs and D Schluter. Chicago (IL): Universityof Chicago Press.
Farrell B D, Mitter C. 1994. Adaptive radiation ininsects and plants: time and opportunity. AmericanZoologist 34:57–69.
Farrell B D, Mitter C. 1998. The timing of insect/plantdiversification: might Tetraopes (Coleoptera: Cer-ambycidae) and Asclepias (Asclepiadaceae) haveco-evolved? Biological Journal of the Linnean Society63:553–577.
Feeny P. 1975. Biochemical coevolution betweenplants and their insect herbivores. Pages 3–19 inCoevolution of Animals and Plants, edited by L E Gil-bert and P H Raven. Austin (TX): University ofTexas Press.
Feeny P. 1976. Plant apparency and chemical defense.Pages 1–40 in Recent Advances in Phytochemistry, Vol-ume 10, edited by J W Wallace and R L Mansell.New York: Plenum Press.
Feeny P. 1991. Theories of plant chemical defense: abrief historical survey. Pages 163–175 in Insects andPlants, edited by T Jermy and A Szentesi. TheHague (The Netherlands): SPB Academic Publish-ing.
Felton G W, Korth K L. 2000. Trade-offs between path-ogen and herbivore resistance. Current Opinion inPlant Biology 3:309–314.
Fineblum W L, Rausher M D. 1995. Tradeoff betweenresistance and tolerance to herbivore damage in amorning glory. Nature 377:517–520.
Firmage D H. 1981. Environmental influences on themonoterpene variation in Hedeoma drummondii.Biochemical Systematics and Ecology 9:53–58.
Folgarait P J, Davidson D W. 1994. Antiherbivoredefenses of myrmecophytic Cecropia under differ-ent light regimes. Oikos 71:305–320.
Folgarait P J, Davidson D W. 1995. MyrmecophyticCecropia: antiherbivore defenses under differentnutrient treatments. Oecologia 104:189–206.
Fowler S V, Lawton J H. 1985. Rapidly induceddefenses and talking trees: the devil’s advocateposition. American Naturalist 126:181–195.
Fox L R. 1981. Defense and dynamics in plant-herbi-vore systems. American Zoologist 21:853–864.
Fraenkel G S. 1959. The raison d’etre of secondaryplant substances. Science 129:1466–1470.
Futuyma D. 1976. Food plant specialization and envi-ronmental predictability in lepidoptera. AmericanNaturalist 110:285–292.
Futuyma D. 1983. Evolutionary interactions amongherbivorous insects and plants. Pages 207–231 inCoevolution, edited by D J Futuyma and M Slatkin.Sunderland (MA): Sinauer Associates.
Futuyma D, Keese M C. 1992. Evolution and coevolu-
tion of plants and phytophagous arthropods. Pages439–475 in Herbivores: Their Interactions with Second-ary Plant Metabolites, Second Edition, Volume II,edited by G A Rosenthal and M R Berenbaum. SanDiego (CA): Academic Press.
Futuyma D, Slatkin M. 1983. Introduction. Pages 1–13in Coevolution, edited by D J Futuyma and M Slat-kin. Sunderland (MA): Sinauer Associates.
Gershenzon J. 1984. Changes in the levels of plantsecondary metabolites under water and nutrientstress. Pages 273–320 in Recent Advances in Phyto-chemistry, Volume 18, edited by B N Timmermannet al. New York: Plenum Press.
Gershenzon J. 1994a. Metabolic costs of terpenoidaccumulation in higher plants. Journal of ChemicalEcology 20:1281–1328.
Gershenzon J. 1994b. The cost of plant chemicaldefense against herbivory: a biochemical perspec-tive. Pages 105–173 in Insect-Plant Interactions, Vol-ume 5, edited by E A Bernays. Boca Raton (FL):CRC Press.
Gershenzon J, Croteau R. 1991. Terpenoids. Pages165–219 in Herbivores: Their Interactions with Second-ary Plant Metabolites, Second Edition, Volume I,edited by G A Rosenthal and M R Berenbaum. SanDiego (CA): Academic Press.
Gijzen M, Lewinsohn E, Croteau R. 1992. Antigeniccross-reactivity among monoterpene cyclases fromgrand fir and induction of these enzymes uponstem wounding. Archives of Biochemistry and Biophys-ics 294:670–674.
Green T R, Ryan C A. 1972. Wound-induced protein-ase inhibitor in plant leaves: a possible defensemechanism against insects. Science 175:776–777.
Grime J P. 1977. Evidence for the existence of threeprimary strategies in plants and its relevance toecological and evolutionary theory. American Nat-uralist 111:1169–1194.
Grime J P. 1979. Plant Strategies and Vegetation Processes.New York: Wiley.
Hammerschmidt R, Schultz J C. 1995. Multipledefenses and signals in plant defense against path-ogens and herbivores. Pages 121–154 in RecentAdvances in Phytochemistry, Volume 30, edited by JT Romeo et al. New York: Plenum Press.
Hamilton J G, Zangerl A R, DeLucia E H, BerenbaumM R. 2001. The carbon-nutrient balance hypothe-sis: its rise and fall. Ecology Letters 4:86–95.
Haukioja E, Neuvonen S. 1985. Induced long-termresistance of birch foliage against defoliators:defensive or incidental? Ecology 66:1303–1308.
Hay M E. 1991. Marine-terrestrial contrasts in the ecol-ogy of plant chemical defenses against herbivores.Trends in Ecology & Evolution 6:362–365.
Heil M, Bostock R M. 2002. Induced systemic resis-tance (ISR) against pathogens in the context of

52 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
induced plant defenses. Annals of Botany 89:503–512.
Herms D A, Mattson W J. 1992. The dilemma of plants:to grow or defend. Quarterly Review of Biology67:283–335.
Hodgson J G, Wilson P J, Hunt R, Grime J P, Thomp-son K. 1999. Allocating C-S-R plant functionaltypes: a soft approach to a hard problem. Oikos85:282–294.
Holopainen J K, Rikala R, Kainulainen P, Oksanen J.1995. Resource partitioning to growth, storage anddefence in nitrogen-fertilized Scots pine and sus-ceptibility of the seedlings to the tarnished plantbug Lygus rugulipennis. New Phytologist 131:521–532.
Howe G A, Lightner J, Browse J, Ryan C A. 1996. Anoctadecanoid pathway mutant ( JL5) of tomato iscompromised in signaling for defense againstinsect attack. Plant Cell 8:2067–2077.
Ingestad T. 1982. Relative addition rate and externalconcentration: driving variables used in plantnutrition research. Plant Cell and Environment5:443–453.
Ingestad T, Lund A. 1986. Theory and techniques forsteady state mineral nutrition and growth of plants.Scandinavian Journal of Forest Research 1:439–453.
Janzen D H. 1980. When is it coevolution? Evolution34:611–612.
Janzen D H. 1985. A host plant is more than its chem-istry. Illinois Natural History Survey Report 33:141–174.
Jing S W, Coley P D. 1990. Dioecy and herbivory: theeffect of growth rate on plant defense in Acernegundo. Oikos 58:369–377.
Johnson N D, Liu B, Bentley B L. 1987. The effects ofnitrogen fixation, soil nitrate, and defoliation onthe growth, alkaloids, and nitrogen levels of Lupi-nus succulentus (Fabaceae). Oecologia 74:425–431.
Jones C G, Hartley S E. 1999. A protein competitionmodel of phenolic allocation. Oikos 86:27–44.
Karban R. 1993. Costs and benefits of induced resis-tance and plant density for a native shrub, Gossy-pium thurberi. Ecology 74:9–19.
Karban R, Baldwin I T. 1997. Induced Responses to Her-bivory. Chicago (IL): University of Chicago Press.
Karban R, Myers J H. 1989. Induced plant responsesto herbivory. Pages 331–348 in Annual Review ofEcology and Systematics, Volume 20, edited by R FJohnston et al. Palo Alto (CA): Annual Reviews.
Karban R, Strauss S Y. 1993. Effects of herbivores ongrowth and reproduction of their perennial host,Erigeron glaucus. Ecology 74:39–46.
Kinney K K, Lindroth R L, Jung S M, Nordheim E V.1997. Effects of CO2 and NO3- availability on decid-uous trees: phytochemistry and insect perfor-mance. Ecology 78:215–230.
Koricheva J. 1999. Interpreting phenotypic variation
in plant allelochemistry: problems with the use ofconcentrations. Oecologia 119:467–473.
Koricheva J. 2002. Meta-analysis of sources of variationin fitness costs of plant antiherbivore defenses.Ecology 83:176–190.
Koricheva J, Larsson S, Haukioja E, Keinanen M. 1998.Regulation of woody plant secondary metabolismby resource availability: hypothesis testing bymeans of meta-analysis. Oikos 83:212–226.
Korner C H. 1991. Some often overlooked plant char-acteristics as determinants of plant growth: areconsideration. Functional Ecology 5:162–173.
Kunkel B N, Brooks D M. 2002. Cross talk betweensignaling pathways in pathogen defense. CurrentOpinion in Plant Biology 5:325–331.
Lambers H. 1993. Rising CO2, secondary plant metab-olism, plant-herbivore interactions and litterdecomposition—theoretical considerations. Vege-tatio 104:263–271.
Langenheim J H. 1994. Higher plant terpenoids: aphytocentric overview of their ecological roles.Journal of Chemical Ecology 20:1223–1280.
Larsson S, Wiren A, Lundgren L, Ericsson T. 1986.Effects of light and nutrient stress on leaf phenolicchemistry in Salix dasyclados and susceptibility ofGalerucella lineola (Coleoptera). Oikos 47:205–210.
Lincoln D E, Langenheim J H. 1978. Effect of lightand temperature on monoterpenoid yield andcomposition in Satureja douglasii. Biochemical System-atics and Ecology 6:21–32.
Loehle C. 1987. Hypothesis testing in ecology: psycho-logical aspects and the importance of theory mat-uration. Quarterly Review of Biology 62:397–409.
Loehle C. 1988. Philosophical tools: potential contri-butions to ecology. Oikos 51:97–104.
Loomis W E. 1932. Growth-differentiation balance vscarbohydrate-nitrogen ratio. Proceedings of the Amer-ican Society for Horticultural Science 29:240–245.
Loomis W E. 1953. Growth and differentiation—anintroduction and summary. Pages 1–17 in Growthand Differentiation in Plants, edited by W E Loomis.Ames (IA): Iowa State College Press.
Lorio P L. 1986. Growth-differentiation balance: abasis for understanding southern pine beetle-treeinteractions. Forest Ecology and Management 14:259–273.
Luxmoore R J. 1991. A source-sink framework for cou-pling water, carbon, and nutrient dynamics of veg-etation. Tree Physiology 9:267–280.
Marquis R J. 1984. Leaf herbivores decrease fitness ofa tropical plant. Science 226:537–539.
Martin J S, Martin M M, Bernays E A. 1987. Failure oftannic acid to inhibit digestion or reduce digesti-bility of plant protein in gut fluids of insect her-bivores: implications for theories of plant defense.Journal of Chemical Ecology 13:605–621.
Maschinski J, Whitham T G. 1989. The continuum of

March 2003 53PLANT DEFENSE HYPOTHESES
plant responses to herbivory: the influence ofplant association, nutrient availability, and timing.American Naturalist 134:1–19.
Mauricio R. 1998. Costs of resistance to natural ene-mies in field populations of the annual plant Ara-bidopsis thaliana. American Naturalist 151:20–28.
Mauricio R, Rausher M D. 1997. Experimental manip-ulation of putative selective agents provides evi-dence for the role of natural enemies in the evo-lution of plant defense. Evolution 51:1435–1444.
Mauricio R, Rausher M D, Burdick D S. 1997. Varia-tion in the defense strategies of plants: are resis-tance and tolerance mutually exclusive? Ecology78:1301–1311.
McKey D. 1974. Adaptive patterns in alkaloid physi-ology. American Naturalist 108:305–320.
McKey D. 1979. The distribution of secondary com-pounds within plants. Pages 55–133 in Herbivores:Their Interactions with Secondary Plant Metabolites,edited by G A Rosenthal and D H Janzen. NewYork: Academic Press.
McNaughton S J. 1983. Compensatory plant growth asa response to herbivory. Oikos 40:329–336.
Mihaliak C A, Lincoln D E. 1985. Growth pattern andcarbon allocation to volatile leaf terpenes undernitrogen-limiting conditions in Heterotheca subaxil-laris (Asteraceae). Oecologia 66:423–426.
Mitchell-Olds T, Siemens D, Pedersen D. 1996. Physi-ology and costs of resistance to herbivory and dis-ease in Brassica. Entomologia Experimentalis et Appli-cata 80:231–237.
Mitter C, Brooks D R. 1983. Phylogenetic aspects ofcoevolution. Pages 65–98 in Coevolution, edited byD J Futuyma and M Slatkin. Sunderland (MA): Sin-auer Associates.
Mole S. 1994. Trade-offs and constraints in plant-her-bivore defense theory: a life-history perspective.Oikos 71:3–12.
Mooney H A. 1972. Carbon balance of plants. Pages315–346 in Annual Review of Ecology and Systematics,Volume 3, edited by R F Johnston et al. Palo Alto(CA): Annual Reviews.
Mooney H A, Chu C. 1974. Seasonal carbon allocationin Heteromeles arbutifolia, a California evergreenshrub. Oecologia 14:295–306.
Mooney H A, Gulmon S L, Johnson N D. 1983. Phys-iological constraints on plant chemical defenses.Pages 21–36 in Plant Resistance to Insects, edited byP A Hedin. Washington (DC): American ChemicalSociety.
Muzika R M. 1993. Terpenes and phenolics inresponse to nitrogen fertilization: a test of the car-bon/nutrient balance hypothesis. Chemoecology4:3–7.
Muzika R M, Pregitzer K S. 1992. Effects of nitrogenfertilization on leaf phenolic production of grand
fir seedlings. Trees—Structure and Function 6:241–244.
Muzika R M, Pregitzer K S, Hanover J W. 1989.Changes in terpene production following nitrogenfertilization of grand fir (Abies grandis (Dougl.)Lindl.) seedlings. Oecologia 80:485–489.
Ohnmeiss T E, Baldwin I T. 1994. The allometry ofnitrogen allocation to growth and an inducibledefense under nitrogen-limited growth. Ecology75:995–1002.
Orozco-Cardenas M, McGurl B, Ryan C A. 1993.Expression of an antisense prosystemin gene intomato plants reduces resistance toward Manducasexta larvae. Proceedings of the National Academy ofSciences of the United States of America 90:8273–8276.
Paige K N, Whitham T G. 1987. Overcompensation inresponse to mammalian herbivory: the advantageof being eaten. American Naturalist 129:407–416.
Painter E L. 1987. Grazing and intraspecific variationin four North American grass species [Doctoraldissertation]. Fort Collins (CO): Colorado StateUniversity.
Price P W. 1991. The plant vigor hypothesis and her-bivore attack. Oikos 62:244–251.
Purrington C B. 2000. Costs of resistance. CurrentOpinion in Plant Biology 3:305–308.
Reichardt P B, Chapin F S, III, Bryant J P, Mattes B R,Clausen T P. 1991. Carbon/nutrient balance as apredictor of plant defense in Alaskan balsam pop-lar: potential importance of metabolite turnover.Oecologia 88:401–406.
Rhoades D F. 1979. Evolution of plant chemicaldefense against herbivores. Pages 1–55 in Herbi-vores: Their Interaction with Secondary Plant Metabo-lites, edited by G A Rosenthal and D H Janzen. NewYork: Academic Press.
Rhoades D F, Cates R G. 1976. Toward a general the-ory of plant antiherbivore chemistry. Pages 168–213 in Recent Advances in Phytochemistry, Volume 10,edited by J W Wallace and R L Mansell. New York:Plenum Press.
Rosenthal J P, Kotanen P M. 1994. Terrestrial planttolerance to herbivory. Trends in Ecology & Evolu-tion 9:145–148.
Saenz L, Santamarıa J M, Villanueva M A, Loyola-Var-gas V M, Oropeza C. 1993. Changes in the alkaloidcontent of plants of Catharanthus roseus L. (Don).as a result of water stress and treatment with abs-cisic acid. Journal of Plant Physiology 142:244–247.
Schultz J C, Lechowicz M J. 1986. Hostplant, larval age,and feeding behavior influence midgut pH in thegypsy moth (Lymantria dispar). Oecologia 71:133–137.
Scriber J M, Ayres M P. 1988. Leaf chemistry as adefense against insects. ISI Atlas of Science 1:117–123.

54 Volume 78THE QUARTERLY REVIEW OF BIOLOGY
Seger J. 1992. Evolution of exploiter-victim relation-ships. Pages 3–25 in Natural Enemies: The PopulationBiology of Predators, Parasites and Diseases, edited byM J Crawley. Oxford: Blackwell Scientific.
Simms E L. 1992. Costs of plant resistance to herbi-vory. Pages 392–425 in Plant Resistance to Herbivoresand Pathogens: Ecology, Evolution, and Genetics,edited by R S Fritz and E L Simms. Chicago (IL):University of Chicago Press.
Simms E L, Rausher M D. 1989. The evolution of resis-tance to herbivory in Ipomoea purpurea. II. Naturalselection by insects and costs of resistance. Evolu-tion 43:573–585.
Simms E L, Triplett J. 1994. Costs and benefits of plantresponses to disease: resistance and tolerance. Evo-lution 48:1973–1985.
Southwood T R E. 1977. Habitat, the templet for eco-logical strategies? Journal of Animal Ecology 46:337–365.
Steinly B A, Berenbaum M. 1985. Histopathologicaleffects of tannins on the midgut epithelium of Pap-ilio polyxenes and Papilio glaucus. Entomologia Exper-imentalis et Applicata 39:3–9.
Stowe K A, Marquis R J, Hochwender C G, Simms EL. 2000. The evolutionary ecology of tolerance toconsumer damage. Pages 565–595 in AnnualReview of Ecology and Systematics, Volume 31, editedby D G Fautin et al. Palo Alto (CA): AnnualReviews.
Strauss S Y. 1991. Direct, indirect, and cumulativeeffects of three native herbivores on a shared hostplant. Ecology 72:543–558.
Strauss S Y, Rudgers J A, Lau J A, Irwin R E. 2002.Direct and ecological costs of resistance to herbi-vory. Trends in Ecology & Evolution 17:278–285.
Thompson J N. 1989. Concepts of coevolution. Trendsin Ecology & Evolution 4:179–183.
Thompson J N. 1999. What we know and do not knowabout coevolution: insect herbivores and plants asa test case. Pages 7–30 in Herbivores: Between Plantsand Predators, edited by H Olff et al. London: Black-well Science.
Tuomi J. 1992. Toward integration of plant defencetheories. Trends in Ecology & Evolution 7:365–367.
Tuomi J, Fagerstrom T, Niemela P. 1991. Carbon allo-cation, phenotypic plasticity, and induceddefenses. Pages 85–104 in Phytochemical Induction byHerbivores, edited by D W Tallamy and M J Raupp.New York: Wiley.
Tuomi J, Niemela P, Chapin F S, III, Bryant J P, SirenS. 1988. Defensive responses of trees in relation totheir carbon/nutrient balance. Pages 57–72 inMechanisms of Woody Plant Defenses Against Insects:Search for Pattern, edited by W J Mattson et al. NewYork: Springer.
Tuomi J, Niemela P, Haukioja E, Siren S, Neuvonen S.1984. Nutrient stress: an explanation for plant anti-
herbivore responses to defoliation. Oecologia 61:208–210.
van Dam N M, van der Meijden E, Verpoorte R. 1993.Induced responses in three alkaloid-containingplant species. Oecologia 95:425–430.
van Dam N M, Vrieling K. 1994. Genetic variation inconstitutive and inducible pyrrolizidine alkaloid lev-els in Cynoglossum officinale L. Oecologia 99:374–378.
van Dam N M, Witte L, Theuring C, Hartmann T.1995. Distribution, biosynthesis and turnover ofpyrrolizidine alkaloids in Cynoglossum officinale. Phy-tochemistry 39:287–292.
van der Meijden E, Wijn M, Verkaar H J. 1988. Defenceand regrowth, alternative plant strategies in thestruggle against herbivores. Oikos 51:355–363.
Veihmeyer F J, Hendrickson A A. 1961. Responses ofa plant to soil-moisture changes as shown by gua-yule. Hilgardia 30(20):621–637.
Vrieling K, de Vos H, van Wijk C A M. 1993. Geneticanalysis of the concentrations of pyrrolizidine alka-loids in Senecio jacobaea. Phytochemistry 32:1141–1144.
Vrieling K, van Wijk C A M. 1994. Cost assessment ofthe production of pyrrolizidine alkaloids in rag-wort (Senecio jacobaea L.). Oecologia 97:541–546.
Wadleigh C H, Gauch H G, Magistad O C. 1946.Growth and rubber accumulation in guayule asconditioned by soil salinity and irrigation regime.USDA Technical Bulletin 925.
Wallace S K, Eigenbrode S D. 2002. Changes in theglucosinolate-myrosinase defense system in Bras-sica juncea cotyledons during seedling develop-ment. Journal of Chemical Ecology 28:243–256.
Waring R H, McDonald A J S, Larsson S, Ericsson T,Wiren A, Arwidsson E, Ericsson A, Lahammar T.1985. Differences in chemical composition ofplants grown at constant relative growth rates withstable mineral nutrition. Oecologia 66:157–160.
Waring R H, Pitman G B. 1985. Modifying lodgepolepine stands to change susceptibility to mountainpine beetle attack. Ecology 66:889–897.
Waterman P G, Mole S. 1989. Extrinsic factors influ-encing production of secondary metabolites inplants. Pages 107–134 in Insect-Plant Interactions,Volume 1, edited by E A Bernays. Boca Raton (FL):CRC Press.
Waterman P G, Ross J A M, McKey D B. 1984. Factorsaffecting levels of some phenolic compounds,digestibility, and nitrogen content of the matureleaves of Barteria fistulosa (Passifloraceae). Journalof Chemical Ecology 10:387–401.
Whittaker R H. 1975. Communities and Ecosystems. Sec-ond Edition. New York: Macmillan.
Whittaker R H, Feeny P P. 1971. Allelochemics: chem-ical interactions between species. Science 171:757–770.
Wilkens R T. 1997. Limitations of evaluating the

March 2003 55PLANT DEFENSE HYPOTHESES
growth-differentiation balance hypothesis with onlytwo levels of light and water. Ecoscience 4:319–326.
Wilkens R T, Ayres M P, Lorio P L, Hodges J D. 1997.Environmental effects on pine tree carbon budgetsand resistance to bark beetles. Pages 591–616 inThe Productivity and Sustainability of Southern ForestEcosystems in a Changing Environment, edited by RMickler and S Fox. New York: Springer.
Wilkens R T, Spoerke J M, Stamp N E. 1996. Differ-ential responses of growth and two soluble phe-nolics of tomato to resource availability. Ecology77:247–258.
Williams D H, Stone M J, Hauck P R, Rahman S K.1989. Why are secondary metabolites (naturalproducts) biosynthesized? Journal of Natural Prod-ucts 52:1189–1208.
Wisdom C S, Crawford C S, Aldon E F. 1989. Influenceof insect herbivory on photosynthetic area andreproduction in Gutierrezia species. Journal of Ecol-ogy 77:685–692.
Wittstock U, Gershenzon J. 2002. Constitutive planttoxins and their role in defense against herbivoresand pathogens. Current Opinions in Plant Biology5:300–307.
Zangerl A R, Bazzaz F A. 1992. Theory and pattern inplant defense allocation. Pages 363–391 in PlantResistance to Herbivores and Pathogens: Ecology, Evo-lution, and Genetics, edited by R S Fritz and E LSimms. Chicago (IL): University of Chicago Press.
Zangerl A R, Berenbaum M R. 1987. Furanocoumarinsin wild parsnip: effects of photosynthetically activeradiation, ultraviolet light, and nutrients. Ecology68:516–520.
Zangerl A R, Berenbaum M R. 1990. Furanocour-marin induction in wild parsnip: genetics and pop-ulational variation. Ecology 71:1933–1940.
Zangerl A R, Rutledge C E. 1996. The probability ofattack and patterns of constitutive and induceddefense: a test of optimal defense theory. AmericanNaturalist 147:599–608.


















