Organization Oocyte Zebrafish
16
RESEARCH ARTICLE Reorganization of Cytoplasm in the Zebrafish Oocyte and Egg During Early Steps of Ooplasmic Segregation Juan Ferna ´ ndez, * Macarena Valladares, Ricardo Fuentes, and Andrea Ubilla The aim of this work is to determine when and how ooplasmic segregation is initiated in the zebrafish egg. To this end, the organization of the ooplasm and vitelloplasm were examined in oocytes and eggs shortly after activation. Ooplasmic segregation, initiated in the stage V oocyte, led to the formation of ooplasmic domains rich in organelles, and ribonucleoproteins. A linear array of closely arranged peripheral yolk globules separated an outer domain of ectoplasm from an inner domain of interconnected endoplasmic lacunae. The structure of this yolk array and the distribution of microinjected labeled tracers suggests that it may provide a barrier limiting ooplasm transit. Loosely arranged yolk globules at the animal hemisphere allow wide connections between the endoplasm and a preblastodisc domain. Activation caused further segregation of ooplasm, reorganization of endoplasmic lacunae, and blastodisc growth. The presence of an endoplasmic cytoskeleton suggests that these changes may be driven by microtubules and microfilaments. Developmental Dynamics 235:656 – 671, 2006. © 2006 Wiley-Liss, Inc. Key words: zebrafish; oocyte; egg; ooplasmic segregation Accepted 28 November 2005 INTRODUCTION Distinctive features of egg cytoplasm are the presence of numerous or- ganelles, yolk inclusions, maternal mRNAs, proteins, and sometimes lipid droplets and pigments. These cy- toplasmic components are at first in- termingled and preparation of the egg for development is often accompanied by separation of the organelles and maternal factors from the yolk, which in turn leads to establishment of oop- lasmic domains. Orderly partition and funneling of these domains into differ- ent blastomeres during cleavage is necessary for normal embryogenesis to take place. The process is called “ooplasmic segregation.” Its initiation, usually, is triggered by egg activation. Activation may be elicited by sperm penetration or by chemical or mechan- ical stimulation of the egg. The eggs of many invertebrate and some verte- brate phyla have been used as models for the study of ooplasmic segregation as for example the eggs of nematodes (Strome and Wood, 1983; Bowerman, 1999), ascidians (Reverberi, 1971; Jef- fery and Meier, 1983; Sardet et al., 1989), annelids (Shimizu, 1982; Fer- na ´ ndez and Olea, 1982; Ferna ´ndez et al., 1998), mollusks (Hess, 1971), am- phibians (Elinson and Rowning, 1988; Danilchik and Denegre, 1991), and fish (reviewed by Hart and Fluck, 1995). Although every cell model has advantages and limitations, the te- leost fish egg has particular advan- tages for studying ooplasmic segrega- tion. First, the egg is translucent and poleward transportation of ooplasm can be traced in live eggs using regu- lar light microscopy (Roosen-Runge, 1938; Devillers, 1961; Ivanenkov et al., 1990; Abraham et al., 1993; Kim- mel et al., 1995; Leung et al., 2000). Second, eggs are amenable for experi- mental manipulation such as microin- jection, ablation of parts, development within wide range of temperatures, ul- traviolet irradiation, drug treatment, and so on (Oppenheimer, 1936; Tung et al., 1945; Jesuthasan and Stra ¨ hle, 1996). In addition, information is available about the location and man- ner of transportation of maternal fac- tors during ooplasmic segregation (re- viewed by Pelegri, 2003). Another interesting feature of the fish egg is that yolk may be sequestered within a Department of Biology, Faculty of Sciences, University of Chile, Santiago, Chile Grant sponsor: FONDECYT; Grant number: 1030879. *Correspondence to: Juan Ferna ´ ndez, Department of Biology, Faculty of Sciences, University of Chile, Casilla 653, Santiago, Chile. E-mail: [email protected] DOI 10.1002/dvdy.20682 Published online 19 January 2006 in Wiley InterScience (www.interscience.wiley.com). DEVELOPMENTAL DYNAMICS 235:656 – 671, 2006 © 2006 Wiley-Liss, Inc.
Transcript of Organization Oocyte Zebrafish

RESEARCH ARTICLE
Reorganization of Cytoplasm in the ZebrafishOocyte and Egg During Early Steps ofOoplasmic SegregationJuan Fernandez,* Macarena Valladares, Ricardo Fuentes, and Andrea Ubilla
The aim of this work is to determine when and how ooplasmic segregation is initiated in the zebrafish egg.To this end, the organization of the ooplasm and vitelloplasm were examined in oocytes and eggs shortlyafter activation. Ooplasmic segregation, initiated in the stage V oocyte, led to the formation of ooplasmicdomains rich in organelles, and ribonucleoproteins. A linear array of closely arranged peripheral yolkglobules separated an outer domain of ectoplasm from an inner domain of interconnected endoplasmiclacunae. The structure of this yolk array and the distribution of microinjected labeled tracers suggests thatit may provide a barrier limiting ooplasm transit. Loosely arranged yolk globules at the animal hemisphereallow wide connections between the endoplasm and a preblastodisc domain. Activation caused furthersegregation of ooplasm, reorganization of endoplasmic lacunae, and blastodisc growth. The presence of anendoplasmic cytoskeleton suggests that these changes may be driven by microtubules and microfilaments.Developmental Dynamics 235:656–671, 2006. © 2006 Wiley-Liss, Inc.
Key words: zebrafish; oocyte; egg; ooplasmic segregation
Accepted 28 November 2005
INTRODUCTION
Distinctive features of egg cytoplasmare the presence of numerous or-ganelles, yolk inclusions, maternalmRNAs, proteins, and sometimeslipid droplets and pigments. These cy-toplasmic components are at first in-termingled and preparation of the eggfor development is often accompaniedby separation of the organelles andmaternal factors from the yolk, whichin turn leads to establishment of oop-lasmic domains. Orderly partition andfunneling of these domains into differ-ent blastomeres during cleavage isnecessary for normal embryogenesisto take place. The process is called“ooplasmic segregation.” Its initiation,usually, is triggered by egg activation.Activation may be elicited by sperm
penetration or by chemical or mechan-ical stimulation of the egg. The eggs ofmany invertebrate and some verte-brate phyla have been used as modelsfor the study of ooplasmic segregationas for example the eggs of nematodes(Strome and Wood, 1983; Bowerman,1999), ascidians (Reverberi, 1971; Jef-fery and Meier, 1983; Sardet et al.,1989), annelids (Shimizu, 1982; Fer-nandez and Olea, 1982; Fernandez etal., 1998), mollusks (Hess, 1971), am-phibians (Elinson and Rowning, 1988;Danilchik and Denegre, 1991), andfish (reviewed by Hart and Fluck,1995). Although every cell model hasadvantages and limitations, the te-leost fish egg has particular advan-tages for studying ooplasmic segrega-tion. First, the egg is translucent and
poleward transportation of ooplasmcan be traced in live eggs using regu-lar light microscopy (Roosen-Runge,1938; Devillers, 1961; Ivanenkov etal., 1990; Abraham et al., 1993; Kim-mel et al., 1995; Leung et al., 2000).Second, eggs are amenable for experi-mental manipulation such as microin-jection, ablation of parts, developmentwithin wide range of temperatures, ul-traviolet irradiation, drug treatment,and so on (Oppenheimer, 1936; Tunget al., 1945; Jesuthasan and Strahle,1996). In addition, information isavailable about the location and man-ner of transportation of maternal fac-tors during ooplasmic segregation (re-viewed by Pelegri, 2003). Anotherinteresting feature of the fish egg isthat yolk may be sequestered within a
Department of Biology, Faculty of Sciences, University of Chile, Santiago, ChileGrant sponsor: FONDECYT; Grant number: 1030879.*Correspondence to: Juan Fernandez, Department of Biology, Faculty of Sciences, University of Chile, Casilla 653, Santiago,Chile. E-mail: [email protected]
DOI 10.1002/dvdy.20682Published online 19 January 2006 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 235:656–671, 2006
© 2006 Wiley-Liss, Inc.

membrane-bound compartment at thecenter of the egg, as in Medaka, ormay be intermingled with ooplasm, asin zebrafish (reviewed by Hart andFluck, 1995). This situation deter-mines important differences on howooplasmic segregation is achieved.
The zebrafish egg has been used ex-tensively to understand the mecha-nisms of ooplasmic segregation. Earlystudies by Roosen-Runge (1938), Lewisand Roosen-Runge (1943), Hisaoka andFirlit (1960), and Beams et al. (1985)were descriptive, whereas that of Ka-tow (1983), using cytochalasin B, aimedto understand the role of actin in oop-lasmic segregation. Leung et al. (1998,2000) have focused on the role of cal-cium and cortical actin filaments inooplasmic segregation, whereas Je-suthasan and Strahle (1996) and Goreand Sampath (2002) have shown thatcortical microtubules transport foreignmicroinjected particles as well as cyto-plasmic determinants. Information isnow available from in situ hybridizationand pharmacological approaches aboutlocation and transportation of maternalmRNAs (reviewed by Pelegri, 2003).
Despite the importance of the oop-lasm in the construction of the zebrafishblastodisc, and consequently of the em-bryo (Lewis and Roosen-Runge, 1943;Leung et al., 2000), little is known onthe fine structure of its components,particularly of the endoplasm. Katow(1983) briefly described the structure ofendoplasm streamers in zebrafish earlyfertilized eggs using transmission elec-tron microscopy. He found mitochon-dria and 30-nm granules but no cy-toskeletal elements. A few years later,Beams et al. (1985) described the possi-ble presence of microtubules and micro-filaments across the endoplasm usingscanning electron microscopy. Numer-ous studies of the fine structure of theectoplasm have been made, particularlyof the cortical actin-rich region and itsparticipation in fertilization (Wolenskiand Hart, 1987; Hart et al., 1992), cor-tical reaction (Hart et al., 1977; Hartand Yu, 1980), and formation of theblastodisc (Katow, 1983).
In this study, we show that ooplasmicsegregation is initiated in the stage Voocyte by separation of yolk from oop-lasm. This process leads to the estab-lishment of 3 ooplasmic domains: ecto-plasm, endoplasm, and preblastodisc.The ooplasm encloses numerous or-
ganelles scattered across a granularmatrix of ribonucleoproteins. It is pre-sumed that ooplasmic domains areformed as result of concerted clusteringof yolk globules across the oocyte cyto-plasm. At most of the oocyte periphery,this process generated a palisade ofclosely arranged yolk globules that sep-arated ecto- and endoplasm, while atthe center of the oocyte yolk clusteringgave rise to irregularly shaped endo-plasmic lacunae. Narrow pathways, orcanaliculi, between peripheral globulesmay impose restrictions to the transit ofooplasm between ecto- and endoplasm.Loosely arranged peripheral yolk glob-ules at the top of the animal hemi-sphere, on the contrary, left wide gaps,allowing endoplasm flow into the pre-blastodisc. Activation caused furthernarrowing of the canaliculi between pe-ripheral yolk globules, a situation thatprobably imposed further restrictions tothe transit of organelles and ribonucleo-proteins between ecto- and endoplasm.This conclusion is supported by resultsdealing with the distribution of labeleddextrans. Narrowing of canaliculi be-tween central yolk globules provokedenlargement of endoplasmic lacunaedue to translocation of granular oop-lasm from between the yolk globules.These processes as well as the gather-ing of endoplasmic lacunae along theanimal/vegetal axis of the egg, the rear-rangement of organelles and ribonucle-oprotein granules in the endoplasm ofmany lacunae, and the blastodiscgrowth are likely to be driven by anubiquitous internal cytoskeleton of mi-crotubules and microfilaments.
RESULTS
Comparison of EarlyDevelopment BetweenFertilized and UnfertilizedEggs at DifferentTemperatures
Tests were made to determine whetherthere were differences on how ooplas-mic segregation progressed in fertilizedand unfertilized eggs. For this purpose,we considered two developmental pa-rameters: (1) rate of growth of the blas-todisc, as a result of ooplasm accumula-tion; and (2) time at which the animalactin ring underwent the first contrac-tion, as shown by the appearance of aconstriction ring at the outer border of
the blastodisc. Adequate visualizationof the growing blastodisc and contrac-tion ring was achieved by following in-dividual properly oriented chorionatedeggs. At the normal developing temper-ature of 28°C, the blastodisc of the fer-tilized and unfertilized eggs grew at thesame rate and the first contraction ofthe actin ring occurred at approxi-mately the same time: 26–29 min ofdevelopment (data for 16 unfertilizedand 15 fertilized eggs).
At 28°C, ooplasmic segregation in theegg and zygote occur too rapidlyto be followed in detail. For this reason,it was important to determine whetherearly development at lower tempera-tures could alter the subsequent steps.In fertilized and unfertilized eggs at20°C, we found that the blastodisc grewsimilarly and the first contraction of theactin ring also occurred at approxi-mately the same time: 45–49 min ofdevelopment (data for 18 unfertilizedand 12 fertilized eggs). Comparison offertilized and unfertilized eggs develop-ing at either 28°C or 20°C indicatedthat the lower temperature slowed thedevelopmental rate approximately 85%,yet under these conditions, ooplasmicsegregation appeared to progress nor-mally. Gross abnormalities were not de-tected in fertilized eggs developed at20°C until cleavage had produced sev-eral blastomeres. Results confirmed thework of Schirone and Gross (1968) inthat at 20°C zebrafish eggs developed ata reduced rate and normally until the32-cell stage embryo.
Ooplasm and Yolk AreIntermingled in the StageIII Oocyte
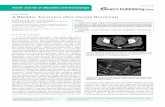
Formation of ooplasmic domains as re-sult of ooplasm and yolk separationmay be conveniently studied in acid-fixed cells. Unfortunately, this tech-nique failed to show the distribution ofooplasm in a nontranslucent cell likethe stage III oocyte. However, examina-tion of stained thick sections of resin-embedded medium and large sizedstage III oocytes showed that ooplasmand yolk globules were distributedevenly across the oocyte cytoplasm (Fig.1). Therefore, separation of ooplasmfrom yolk has not yet started in thestage III oocyte. For reasons explainedin the Experimental Procedures sec-
EARLY STEPS OF OOPLASMIC SEGREGATION 657
tion, it was not possible to examinestage IV oocytes.
Separation of Ooplasm FromYolk and Establishment ofOoplasmic Domains in theStage V Oocyte Constitutean Early Step in OoplasmicSegregation
Examination of whole-mountedacid-fixed stage V oocytes byvideo light microscopy.
Microscopic examination of nonacti-vated acid-fixed stage V oocytes showedthat their ooplasm was segregated from
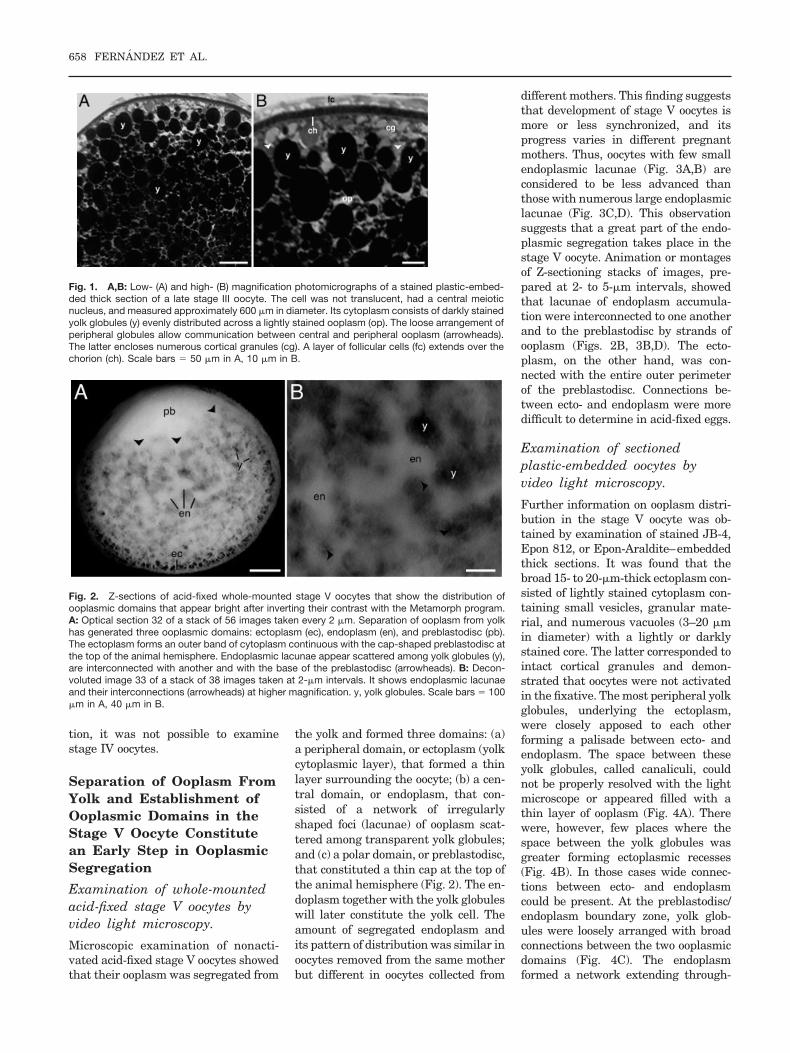
the yolk and formed three domains: (a)a peripheral domain, or ectoplasm (yolkcytoplasmic layer), that formed a thinlayer surrounding the oocyte; (b) a cen-tral domain, or endoplasm, that con-sisted of a network of irregularlyshaped foci (lacunae) of ooplasm scat-tered among transparent yolk globules;and (c) a polar domain, or preblastodisc,that constituted a thin cap at the top ofthe animal hemisphere (Fig. 2). The en-doplasm together with the yolk globuleswill later constitute the yolk cell. Theamount of segregated endoplasm andits pattern of distribution was similar inoocytes removed from the same motherbut different in oocytes collected from
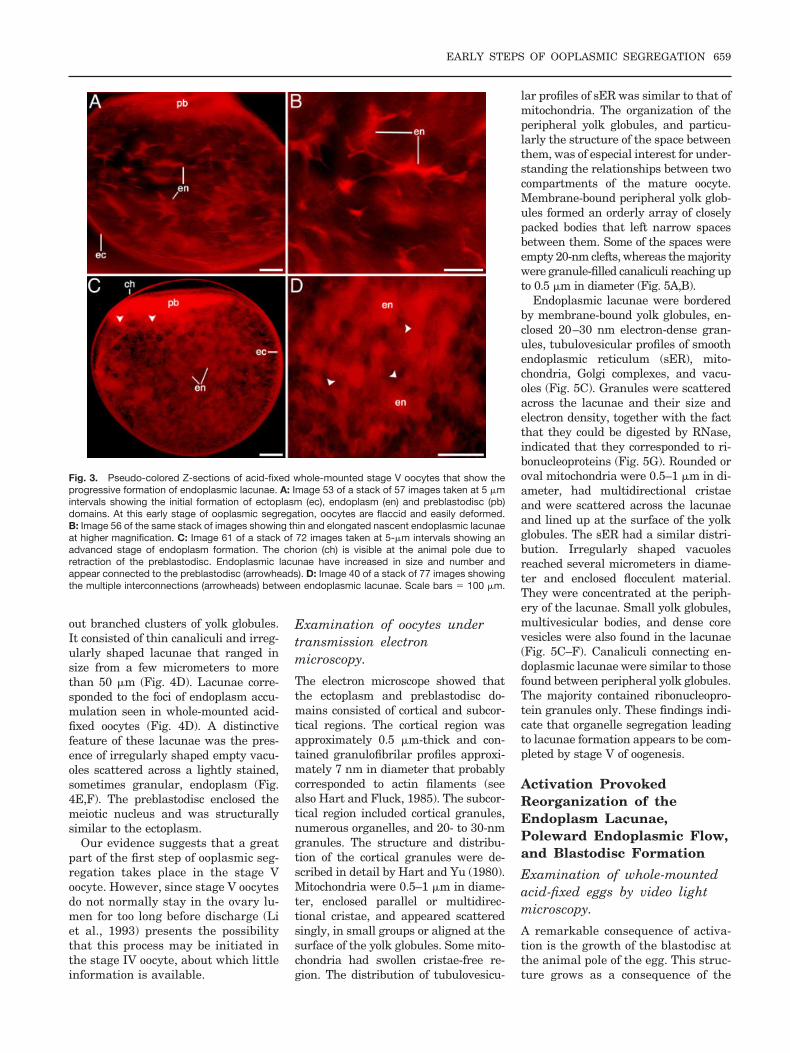
different mothers. This finding suggeststhat development of stage V oocytes ismore or less synchronized, and itsprogress varies in different pregnantmothers. Thus, oocytes with few smallendoplasmic lacunae (Fig. 3A,B) areconsidered to be less advanced thanthose with numerous large endoplasmiclacunae (Fig. 3C,D). This observationsuggests that a great part of the endo-plasmic segregation takes place in thestage V oocyte. Animation or montagesof Z-sectioning stacks of images, pre-pared at 2- to 5-�m intervals, showedthat lacunae of endoplasm accumula-tion were interconnected to one anotherand to the preblastodisc by strands ofooplasm (Figs. 2B, 3B,D). The ecto-plasm, on the other hand, was con-nected with the entire outer perimeterof the preblastodisc. Connections be-tween ecto- and endoplasm were moredifficult to determine in acid-fixed eggs.
Examination of sectionedplastic-embedded oocytes byvideo light microscopy.
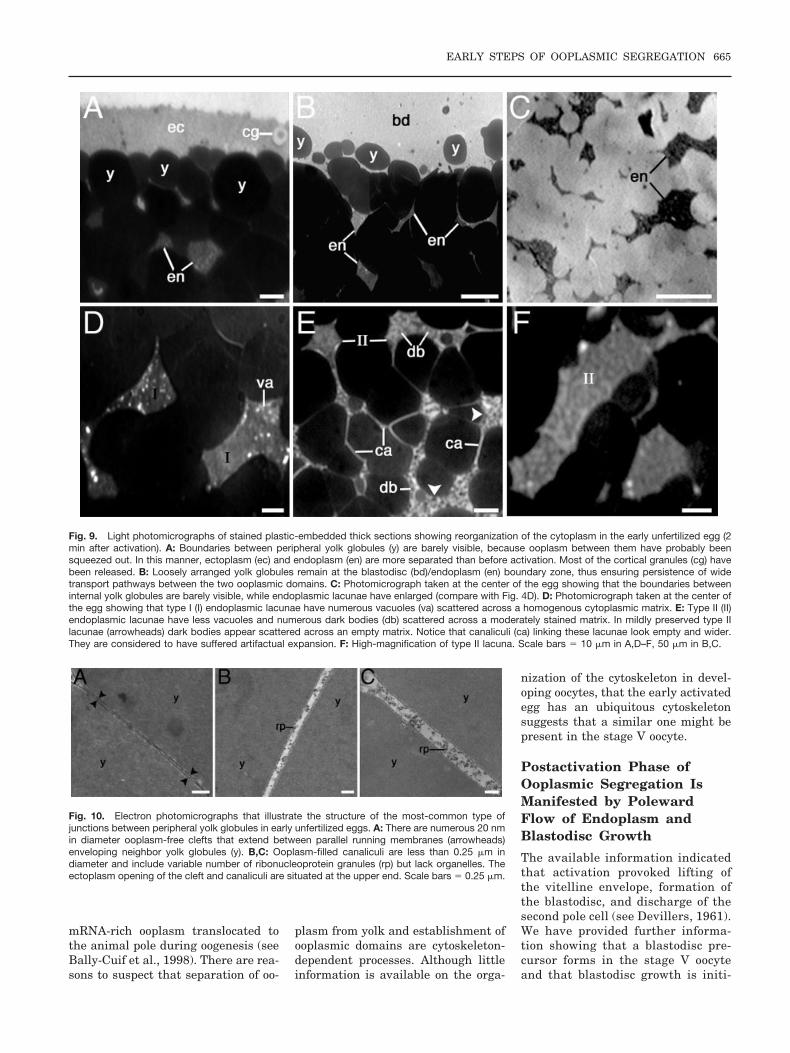
Further information on ooplasm distri-bution in the stage V oocyte was ob-tained by examination of stained JB-4,Epon 812, or Epon-Araldite–embeddedthick sections. It was found that thebroad 15- to 20-�m-thick ectoplasm con-sisted of lightly stained cytoplasm con-taining small vesicles, granular mate-rial, and numerous vacuoles (3–20 �min diameter) with a lightly or darklystained core. The latter corresponded tointact cortical granules and demon-strated that oocytes were not activatedin the fixative. The most peripheral yolkglobules, underlying the ectoplasm,were closely apposed to each otherforming a palisade between ecto- andendoplasm. The space between theseyolk globules, called canaliculi, couldnot be properly resolved with the lightmicroscope or appeared filled with athin layer of ooplasm (Fig. 4A). Therewere, however, few places where thespace between the yolk globules wasgreater forming ectoplasmic recesses(Fig. 4B). In those cases wide connec-tions between ecto- and endoplasmcould be present. At the preblastodisc/endoplasm boundary zone, yolk glob-ules were loosely arranged with broadconnections between the two ooplasmicdomains (Fig. 4C). The endoplasmformed a network extending through-
Fig. 1. A,B: Low- (A) and high- (B) magnification photomicrographs of a stained plastic-embed-ded thick section of a late stage III oocyte. The cell was not translucent, had a central meioticnucleus, and measured approximately 600 �m in diameter. Its cytoplasm consists of darkly stainedyolk globules (y) evenly distributed across a lightly stained ooplasm (op). The loose arrangement ofperipheral globules allow communication between central and peripheral ooplasm (arrowheads).The latter encloses numerous cortical granules (cg). A layer of follicular cells (fc) extends over thechorion (ch). Scale bars � 50 �m in A, 10 �m in B.
Fig. 2. Z-sections of acid-fixed whole-mounted stage V oocytes that show the distribution ofooplasmic domains that appear bright after inverting their contrast with the Metamorph program.A: Optical section 32 of a stack of 56 images taken every 2 �m. Separation of ooplasm from yolkhas generated three ooplasmic domains: ectoplasm (ec), endoplasm (en), and preblastodisc (pb).The ectoplasm forms an outer band of cytoplasm continuous with the cap-shaped preblastodisc atthe top of the animal hemisphere. Endoplasmic lacunae appear scattered among yolk globules (y),are interconnected with another and with the base of the preblastodisc (arrowheads). B: Decon-voluted image 33 of a stack of 38 images taken at 2-�m intervals. It shows endoplasmic lacunaeand their interconnections (arrowheads) at higher magnification. y, yolk globules. Scale bars � 100�m in A, 40 �m in B.
658 FERNANDEZ ET AL.
out branched clusters of yolk globules.It consisted of thin canaliculi and irreg-ularly shaped lacunae that ranged insize from a few micrometers to morethan 50 �m (Fig. 4D). Lacunae corre-sponded to the foci of endoplasm accu-mulation seen in whole-mounted acid-fixed oocytes (Fig. 4D). A distinctivefeature of these lacunae was the pres-ence of irregularly shaped empty vacu-oles scattered across a lightly stained,sometimes granular, endoplasm (Fig.4E,F). The preblastodisc enclosed themeiotic nucleus and was structurallysimilar to the ectoplasm.
Our evidence suggests that a greatpart of the first step of ooplasmic seg-regation takes place in the stage Voocyte. However, since stage V oocytesdo not normally stay in the ovary lu-men for too long before discharge (Liet al., 1993) presents the possibilitythat this process may be initiated inthe stage IV oocyte, about which littleinformation is available.
Examination of oocytes undertransmission electronmicroscopy.
The electron microscope showed thatthe ectoplasm and preblastodisc do-mains consisted of cortical and subcor-tical regions. The cortical region wasapproximately 0.5 �m-thick and con-tained granulofibrilar profiles approxi-mately 7 nm in diameter that probablycorresponded to actin filaments (seealso Hart and Fluck, 1985). The subcor-tical region included cortical granules,numerous organelles, and 20- to 30-nmgranules. The structure and distribu-tion of the cortical granules were de-scribed in detail by Hart and Yu (1980).Mitochondria were 0.5–1 �m in diame-ter, enclosed parallel or multidirec-tional cristae, and appeared scatteredsingly, in small groups or aligned at thesurface of the yolk globules. Some mito-chondria had swollen cristae-free re-gion. The distribution of tubulovesicu-
lar profiles of sER was similar to that ofmitochondria. The organization of theperipheral yolk globules, and particu-larly the structure of the space betweenthem, was of especial interest for under-standing the relationships between twocompartments of the mature oocyte.Membrane-bound peripheral yolk glob-ules formed an orderly array of closelypacked bodies that left narrow spacesbetween them. Some of the spaces wereempty 20-nm clefts, whereas the majoritywere granule-filled canaliculi reaching upto 0.5 �m in diameter (Fig. 5A,B).
Endoplasmic lacunae were borderedby membrane-bound yolk globules, en-closed 20–30 nm electron-dense gran-ules, tubulovesicular profiles of smoothendoplasmic reticulum (sER), mito-chondria, Golgi complexes, and vacu-oles (Fig. 5C). Granules were scatteredacross the lacunae and their size andelectron density, together with the factthat they could be digested by RNase,indicated that they corresponded to ri-bonucleoproteins (Fig. 5G). Rounded oroval mitochondria were 0.5–1 �m in di-ameter, had multidirectional cristaeand were scattered across the lacunaeand lined up at the surface of the yolkglobules. The sER had a similar distri-bution. Irregularly shaped vacuolesreached several micrometers in diame-ter and enclosed flocculent material.They were concentrated at the periph-ery of the lacunae. Small yolk globules,multivesicular bodies, and dense corevesicles were also found in the lacunae(Fig. 5C–F). Canaliculi connecting en-doplasmic lacunae were similar to thosefound between peripheral yolk globules.The majority contained ribonucleopro-tein granules only. These findings indi-cate that organelle segregation leadingto lacunae formation appears to be com-pleted by stage V of oogenesis.
Activation ProvokedReorganization of theEndoplasm Lacunae,Poleward Endoplasmic Flow,and Blastodisc Formation
Examination of whole-mountedacid-fixed eggs by video lightmicroscopy.
A remarkable consequence of activa-tion is the growth of the blastodisc atthe animal pole of the egg. This struc-ture grows as a consequence of the
Fig. 3. Pseudo-colored Z-sections of acid-fixed whole-mounted stage V oocytes that show theprogressive formation of endoplasmic lacunae. A: Image 53 of a stack of 57 images taken at 5 �mintervals showing the initial formation of ectoplasm (ec), endoplasm (en) and preblastodisc (pb)domains. At this early stage of ooplasmic segregation, oocytes are flaccid and easily deformed.B: Image 56 of the same stack of images showing thin and elongated nascent endoplasmic lacunaeat higher magnification. C: Image 61 of a stack of 72 images taken at 5-�m intervals showing anadvanced stage of endoplasm formation. The chorion (ch) is visible at the animal pole due toretraction of the preblastodisc. Endoplasmic lacunae have increased in size and number andappear connected to the preblastodisc (arrowheads). D: Image 40 of a stack of 77 images showingthe multiple interconnections (arrowheads) between endoplasmic lacunae. Scale bars � 100 �m.
EARLY STEPS OF OOPLASMIC SEGREGATION 659

accumulation of endoplasm from theyolk cell region of the activated egg.This process is particularly wellshown when acid-fixed stage V oo-cytes are compared with activatedacid-fixed eggs (Fig. 6). In the stageV oocyte, the preblastodisc is thinand transparent, when viewed fromthe animal pole, showing the profilesof underneath yolk globules. Thepreblastodisc is 50- to 90-�m-thickand 250 –350 �m in diameter. Threeminutes after activation, the blasto-disc has lost its transparency becom-ing thicker (approximately 130 �m)and wider (approximately 450 �m).Five minutes after activation, theblastodisc became highly opaquereaching 180 �m in thickness and upto 550–650 �m in diameter. There-fore, activation was followed byquick accumulation of ooplasm atthe animal pole. Evidence presentedbelow demonstrates that polewardflow of endoplasm is also accompa-
nied by reorganization of the endo-plasmic lacunae generated in thestage V oocyte.
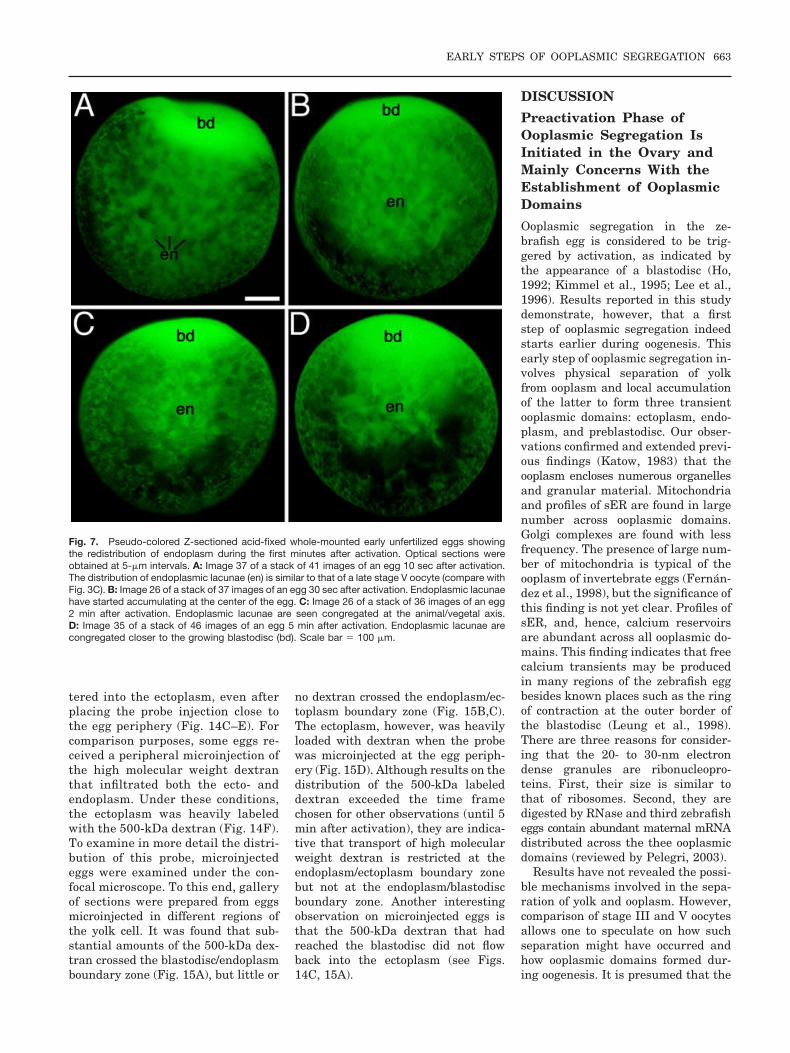
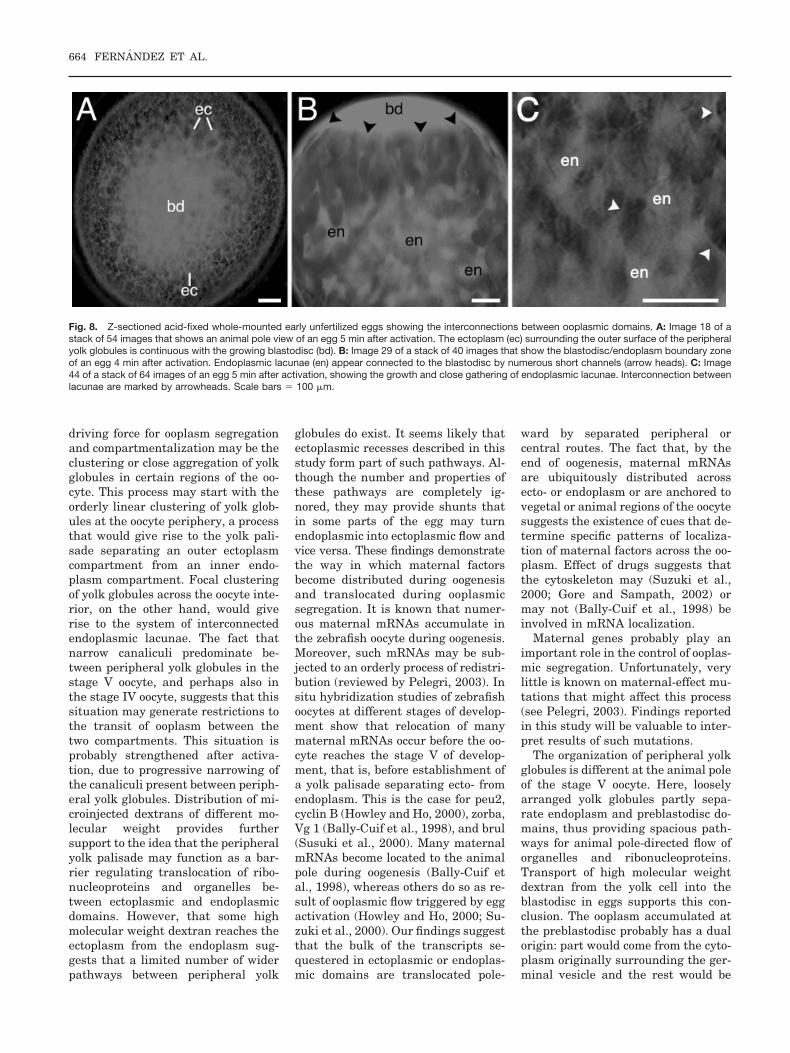
The redistribution of the endoplas-mic lacunae was also studied in acti-vated whole-mounted acid-fixed eggsprepared every 10 sec during the firstminute of development and every 30sec during the following 4 min. Stacksof Z-sectioned eggs allowed examina-tion of digitally processed optical sec-tions and preparation of animationsor reconstructions. Activation pro-duced changes in the distribution ofendoplasmic lacunae (Fig. 7) as a re-sult of endoplasm flowing into theblastodisc. Because the blastodisc andectoplasm have multiple connections(Fig. 8A) ectoplasm may also flow intothe embryonic disc. In many eggs, thenetwork of endoplasmic lacunae accu-mulated at the animal/vegetal axis ofthe egg, in close proximity to the grow-ing blastodisc. Here, lacunae wereseen to extend into the blastodisc
forming short vertical channels remi-niscent of the axial streamers formedin the zygote (Fig. 8B). Higher resolu-tion microscopic observation of acid-fixed eggs showed that the enlargedendoplasmic lacunae were intercon-nected forming a complex network(Fig. 8C).
Examination of sectionedplastic-embedded eggs by videolight microscopy.
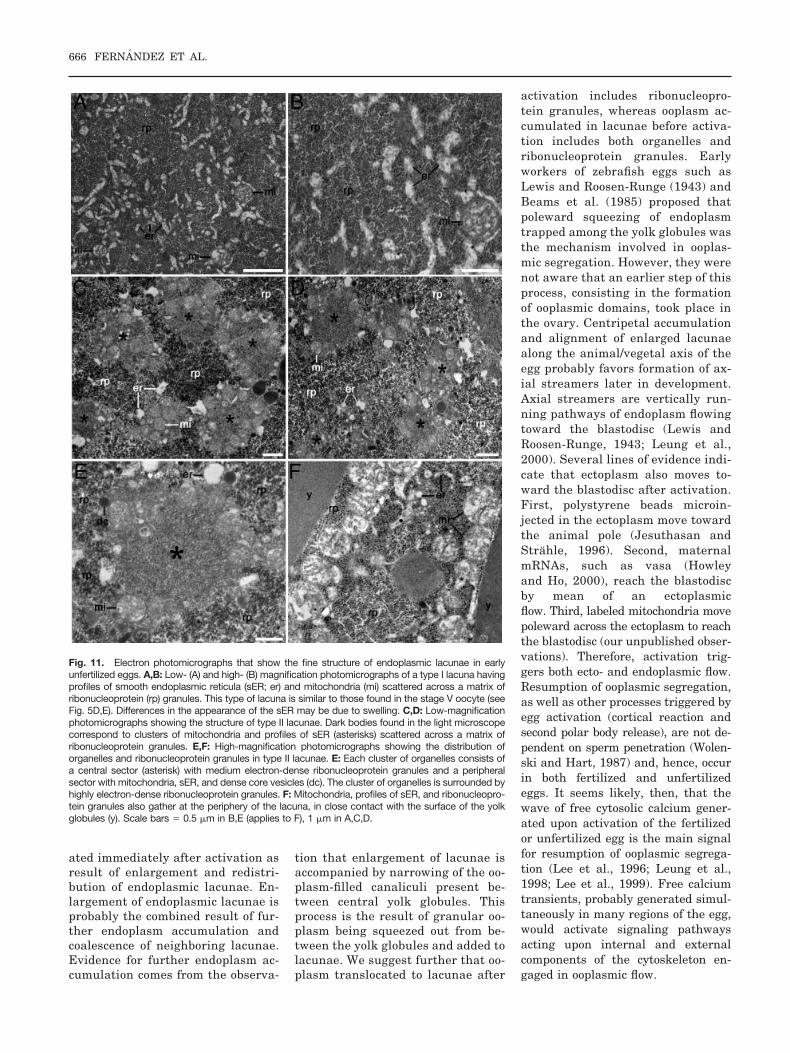
Plastic sections of fertilized and unfer-tilized eggs examined by video micros-copy during the first 5 min after acti-vation confirmed results obtained inacid-fixed eggs. First, it was difficultto visualize the lateral border betweenadjacent peripheral yolk globules (Fig.9A). This observation suggested thatcanaliculi between yolk globules nar-rowed. Second, loosely arranged yolkglobules persisted at the blastodisc/endoplasm boundary zone (Fig. 9B),
Fig. 4. Light photomicrographs of stained plastic-embedded thick sections of stage V oocytes showing the structure of ooplasmic domains (comparewith Fig. 1). A: Peripheral yolk globules (y) are closely apposed to one another forming a palisade separating a continuous outer ectoplasm (ec) froma system of inner endoplasmic lacunae (en). The ectoplasm encloses intact cortical granule (cg). Notice that ooplasm-filled canaliculi (ca) are presentbetween peripheral and central yolk globules. B: Ectoplasmic recess (arrowheads) formed between peripheral yolk globules may sometimes beconnected to endoplasmic lacunae. The ectoplasm encloses several layers of cortical granules. C: Loosely arranged yolk globules at the animal poleregion of the oocyte allow continuity between the endoplasm and the preblastodisc (pb). D: Low-magnification photomicrograph showing thedistribution of endoplasmic lacunae and canaliculi connecting them. E,F: High-magnification photomicrographs showing that endoplasmic lacunaeenclose empty vacuoles (va) and a lightly stained, sometimes granular, matrix. Canaliculi between yolk globules interconnect endoplasmic lacunae andare filled with lightly stained ooplasm. Scale bars� 10 �m in B,E,F, 30 �m in A,C,D.
660 FERNANDEZ ET AL.

maintaining spacious pathways forpoleward endoplasmic flow. Third, en-doplasmic lacunae enlarged and theoutlines of the central yolk globuleswere in general difficult to detect (Fig.9C). Canaliculi had narrowed, pre-sumably as result of ooplasm flowinginto the lacunae. Detailed microscopy
of activated eggs revealed that someendoplasmic lacunae had undergonestructural changes. Thus, two types ofendoplasm lacunae could be detectedin the activated egg. Type I lacunaewere similar to those seen in stage Voocytes (Fig. 9D), whereas type II la-cunae presented a different distribu-
tion of their endoplasmic components(Fig. 9E,F). Vacuoles diminished innumber, and darkly stained bodies,2–7 �m in diameter, appeared scat-tered across the lacunae. Type I and IIlacunae may be interconnected, andsome displayed a mixed structure. Insome eggs, type II lacunae were abun-dant in the blastodisc neighborhood.
Examination of eggs bytransmission electronmicroscopy.
Observations with the transmissionelectron microscope confirmed that ac-tivation led to a reduced width of thecanaliculi running between peripheralyolk globules. Figure 10A–C showsjunctions between peripheral yolkglobules of activated eggs. Junctionswere 20-nm-wide granule-free cleftsor under 0.25 �m-wide ooplasm-filledcanalicules. In this manner, activa-tion imposed further restriction in thesize of the pathways connecting ecto-and endoplasm. Although preserva-tion of the endoplasm was difficult,the two type of lacunae were easilyrecognized across the yolk cell. Type Ilacunae appeared similar to thosefound in the stage V oocyte. Their en-doplasm consisted of mitochondriaand profiles of sER scattered across amatrix of electron-dense granules(Fig. 11A,B). The electron microscopealso showed that dark bodies in type IIlacunae were clusters of medium sizedelectron-dense granules surroundedby mitochondria, profiles of sER, andoften dense core vesicles. These clus-ters were dispersed across a matrix ofregularly spaced highly electron-dense granules (Fig. 11C–E). As ithappened in type I lacunae, numerousmitochondria and profiles of sER linedup at the surface of the yolk globulesfacing the lacunae (Fig. 11F). RNasedigestion demonstrated that electron-dense granules in type II lacunae werealso ribonucleoproteins (Fig. 12).
The cytoskeleton of the zebrafishegg was poorly preserved after regularfixation in the Karnovsky solution.However, prefixation treatment of theeggs with cytoskeleton stabilizers suchas Taxol and phalloidin allowed visu-alization of microtubules and actinfilaments throughout the ecto- andendoplasm as well as blastodisc. We
Fig. 5. Electron photomicrographs that illustrate the fine structure of the stage V oocyte. A,B:These photomicrographs show the most common type of canaliculi present between peripheralyolk globules (y). Small canaliculi (A) only include electron-dense granules (rp), whereas largecanaliculi (B) may also have organelles such as mitochondria (mi). G: Granules are digested byRNase A and, hence, are considered to be ribonucleoproteins. The ectoplasm opening of thecanaliculi is at its upper end. C–F: These photomicrographs illustrate the structure of the endo-plasmic lacunae. C: Low-magnification lacuna showing that the endoplasm includes numerousmitochondria that are scattered across a matrix of ribonucleoprotein granules or line up at thesurface of the yolk globules (y). D,E: Higher-magnification photomicrographs of endoplasmiclacunae showing the abundance of smooth endoplasmic reticulum (er), mitochondria and small yolkglobules scattered across a granular matrix. F: Closely apposed mitochondria lining up the surfaceof a yolk globule. G: A substantial number of electron-dense granules disappeared from theendoplasm after RNase A digestion. This finding caused the appearance of empty spaces sur-rounded by organelles. va, vacuole. Scale bars � 0.25 �m in A,B,D–G, 1 �m in C.
EARLY STEPS OF OOPLASMIC SEGREGATION 661

put particular attention to the endo-plasmic cytoskeleton, because its exis-tence has been controversial. Loosebundles of microtubules were seenalong the endoplasmic lacunae inproximity to organelles. Bundles ofmicrofilaments, approximately 7 nmin diameter, were also seen scatteredthroughout the endoplasmic lacunaenear microtubules (Fig. 13A,B). Thesebundles of microfilaments closely re-sembled those present at the egg cor-tex (Fig. 13C), suggesting that theyalso consisted of actin filaments. De-finitive identification of actin fila-ments in endoplasmic bundles awaitsdecoration with heavy meromyosin.The structure of the cytoskeletoncould not be investigated in the stageV oocyte, because these flaccid cellswere difficult to microinject with cy-toskeleton stabilizers. However, itwas possible to explore in the eggwhether changes in the structure anddistribution of endoplasmic lacunae,as result of activation, were accompa-nied by changes in the structure oftheir cytoskeleton. Observations withthe electron microscope have not yetprovided sufficient information on thismatter. However, is clear that duringearly remodeling of the endoplasmiclacunae microtubule and microfila-ment bundles remain scatteredacross the endoplasm. It is expected
that immunocytochemical stainingof the cytoskeleton during ooplasmicsegregation will provide the neededinformation.
Distribution of MicroinjectedLabeled Tracers SuggestsThat a Transport Barrier IsPresent at the Ectoplasm/Endoplasm Boundary Zone
To determine whether there was a re-lationship between the organization ofperipheral yolk globules and trans-port between different ooplasmic do-mains, labeled proteins and dextranswere microinjected into the yolk cell.Transport of fluorescent tracers wasmonitored by time-lapse fluorescencevideo and confocal microscopy ofwhole-mounted eggs microinjectedshortly after activation. Distributionof the tracer, on the other hand, wasanalyzed in video Z-series of similareggs microinjected in different regionsof the yolk cell.
Distribution of labeled proteins.
Labeled proteins, ranging between21.5- and 66-kDa, diffused throughoutthe endoplasmic lacunae and enteredboth into the blastodisc and ectoplasmwithin the first minutes after microin-jection. Of interest, the layer of pe-ripheral ectoplasm became more fluo-
rescent than the endoplasmic lacunaeand blastodisc (not shown). These re-sults indicated that mid molecularweight proteins microinjected in theendoplasm crossed the closely ap-posed layer of peripheral yolk globulesat the endoplasm/ectoplasm boundaryzone. Brighter fluorescence of the ec-toplasm may be indicative that micro-injected labeled proteins were moreconcentrated in the ectoplasm than inother ooplasmic domains.
Distribution of labeled dextrans.
Distribution of 10- to 70-kDa dex-trans was similar to that of labeledproteins, but their rate of diffusionacross the ooplasm was slower. Ittook no less than 5–15 min for theseprobes, microinjected at the center ofthe yolk cell, to become distributedacross the endoplasmic lacunae,blastodisc and ectoplasm (Fig.14A,B). In the case of microinjected500-kDa dextrans, the time of diffu-sion of the probe across the endo-plasm and blastodisc was evenslower, sometimes extending up to20 –25 min. Substantial amount ofthe probe reached the blastodisc in-dicating that the 500-kDa dextrancrossed the blastodisc/yolk cellboundary zone. However, very littlehigh molecular weight dextran en-
Fig. 6. A–F: Lateral (A,C,E) and animal pole (B,D,F) views of whole-mounted acid-fixed cells showing development of the blastodisc from the earlystage V oocyte (A,B) to the egg shortly after activation (C–F). The early stage V oocyte (A and B) is a flaccid cell exhibiting numerous folds (f). Itspreblastodisc (pb) is thin and remains largely translucent after fixation. At 3 (C,D) and 5 (E,F) min after activation, the blastodisc (bd) has enlarged andthickened considerably. Arrowheads mark the outer border of the preblastodisc and blastodisc. Scale bar � 100 �m.
662 FERNANDEZ ET AL.
tered into the ectoplasm, even afterplacing the probe injection close tothe egg periphery (Fig. 14C–E). Forcomparison purposes, some eggs re-ceived a peripheral microinjection ofthe high molecular weight dextranthat infiltrated both the ecto- andendoplasm. Under these conditions,the ectoplasm was heavily labeledwith the 500-kDa dextran (Fig. 14F).To examine in more detail the distri-bution of this probe, microinjectedeggs were examined under the con-focal microscope. To this end, galleryof sections were prepared from eggsmicroinjected in different regions ofthe yolk cell. It was found that sub-stantial amounts of the 500-kDa dex-tran crossed the blastodisc/endoplasmboundary zone (Fig. 15A), but little or
no dextran crossed the endoplasm/ec-toplasm boundary zone (Fig. 15B,C).The ectoplasm, however, was heavilyloaded with dextran when the probewas microinjected at the egg periph-ery (Fig. 15D). Although results on thedistribution of the 500-kDa labeleddextran exceeded the time framechosen for other observations (until 5min after activation), they are indica-tive that transport of high molecularweight dextran is restricted at theendoplasm/ectoplasm boundary zonebut not at the endoplasm/blastodiscboundary zone. Another interestingobservation on microinjected eggs isthat the 500-kDa dextran that hadreached the blastodisc did not flowback into the ectoplasm (see Figs.14C, 15A).
DISCUSSION
Preactivation Phase ofOoplasmic Segregation IsInitiated in the Ovary andMainly Concerns With theEstablishment of OoplasmicDomains
Ooplasmic segregation in the ze-brafish egg is considered to be trig-gered by activation, as indicated bythe appearance of a blastodisc (Ho,1992; Kimmel et al., 1995; Lee et al.,1996). Results reported in this studydemonstrate, however, that a firststep of ooplasmic segregation indeedstarts earlier during oogenesis. Thisearly step of ooplasmic segregation in-volves physical separation of yolkfrom ooplasm and local accumulationof the latter to form three transientooplasmic domains: ectoplasm, endo-plasm, and preblastodisc. Our obser-vations confirmed and extended previ-ous findings (Katow, 1983) that theooplasm encloses numerous organellesand granular material. Mitochondriaand profiles of sER are found in largenumber across ooplasmic domains.Golgi complexes are found with lessfrequency. The presence of large num-ber of mitochondria is typical of theooplasm of invertebrate eggs (Fernan-dez et al., 1998), but the significance ofthis finding is not yet clear. Profiles ofsER, and, hence, calcium reservoirsare abundant across all ooplasmic do-mains. This finding indicates that freecalcium transients may be producedin many regions of the zebrafish eggbesides known places such as the ringof contraction at the outer border ofthe blastodisc (Leung et al., 1998).There are three reasons for consider-ing that the 20- to 30-nm electrondense granules are ribonucleopro-teins. First, their size is similar tothat of ribosomes. Second, they aredigested by RNase and third zebrafisheggs contain abundant maternal mRNAdistributed across the thee ooplasmicdomains (reviewed by Pelegri, 2003).
Results have not revealed the possi-ble mechanisms involved in the sepa-ration of yolk and ooplasm. However,comparison of stage III and V oocytesallows one to speculate on how suchseparation might have occurred andhow ooplasmic domains formed dur-ing oogenesis. It is presumed that the
Fig. 7. Pseudo-colored Z-sectioned acid-fixed whole-mounted early unfertilized eggs showingthe redistribution of endoplasm during the first minutes after activation. Optical sections wereobtained at 5-�m intervals. A: Image 37 of a stack of 41 images of an egg 10 sec after activation.The distribution of endoplasmic lacunae (en) is similar to that of a late stage V oocyte (compare withFig. 3C). B: Image 26 of a stack of 37 images of an egg 30 sec after activation. Endoplasmic lacunaehave started accumulating at the center of the egg. C: Image 26 of a stack of 36 images of an egg2 min after activation. Endoplasmic lacunae are seen congregated at the animal/vegetal axis.D: Image 35 of a stack of 46 images of an egg 5 min after activation. Endoplasmic lacunae arecongregated closer to the growing blastodisc (bd). Scale bar � 100 �m.
EARLY STEPS OF OOPLASMIC SEGREGATION 663
driving force for ooplasm segregationand compartmentalization may be theclustering or close aggregation of yolkglobules in certain regions of the oo-cyte. This process may start with theorderly linear clustering of yolk glob-ules at the oocyte periphery, a processthat would give rise to the yolk pali-sade separating an outer ectoplasmcompartment from an inner endo-plasm compartment. Focal clusteringof yolk globules across the oocyte inte-rior, on the other hand, would giverise to the system of interconnectedendoplasmic lacunae. The fact thatnarrow canaliculi predominate be-tween peripheral yolk globules in thestage V oocyte, and perhaps also inthe stage IV oocyte, suggests that thissituation may generate restrictions tothe transit of ooplasm between thetwo compartments. This situation isprobably strengthened after activa-tion, due to progressive narrowing ofthe canaliculi present between periph-eral yolk globules. Distribution of mi-croinjected dextrans of different mo-lecular weight provides furthersupport to the idea that the peripheralyolk palisade may function as a bar-rier regulating translocation of ribo-nucleoproteins and organelles be-tween ectoplasmic and endoplasmicdomains. However, that some highmolecular weight dextran reaches theectoplasm from the endoplasm sug-gests that a limited number of widerpathways between peripheral yolk
globules do exist. It seems likely thatectoplasmic recesses described in thisstudy form part of such pathways. Al-though the number and properties ofthese pathways are completely ig-nored, they may provide shunts thatin some parts of the egg may turnendoplasmic into ectoplasmic flow andvice versa. These findings demonstratethe way in which maternal factorsbecome distributed during oogenesisand translocated during ooplasmicsegregation. It is known that numer-ous maternal mRNAs accumulate inthe zebrafish oocyte during oogenesis.Moreover, such mRNAs may be sub-jected to an orderly process of redistri-bution (reviewed by Pelegri, 2003). Insitu hybridization studies of zebrafishoocytes at different stages of develop-ment show that relocation of manymaternal mRNAs occur before the oo-cyte reaches the stage V of develop-ment, that is, before establishment ofa yolk palisade separating ecto- fromendoplasm. This is the case for peu2,cyclin B (Howley and Ho, 2000), zorba,Vg 1 (Bally-Cuif et al., 1998), and brul(Susuki et al., 2000). Many maternalmRNAs become located to the animalpole during oogenesis (Bally-Cuif etal., 1998), whereas others do so as re-sult of ooplasmic flow triggered by eggactivation (Howley and Ho, 2000; Su-zuki et al., 2000). Our findings suggestthat the bulk of the transcripts se-questered in ectoplasmic or endoplas-mic domains are translocated pole-
ward by separated peripheral orcentral routes. The fact that, by theend of oogenesis, maternal mRNAsare ubiquitously distributed acrossecto- or endoplasm or are anchored tovegetal or animal regions of the oocytesuggests the existence of cues that de-termine specific patterns of localiza-tion of maternal factors across the oo-plasm. Effect of drugs suggests thatthe cytoskeleton may (Suzuki et al.,2000; Gore and Sampath, 2002) ormay not (Bally-Cuif et al., 1998) beinvolved in mRNA localization.
Maternal genes probably play animportant role in the control of ooplas-mic segregation. Unfortunately, verylittle is known on maternal-effect mu-tations that might affect this process(see Pelegri, 2003). Findings reportedin this study will be valuable to inter-pret results of such mutations.
The organization of peripheral yolkglobules is different at the animal poleof the stage V oocyte. Here, looselyarranged yolk globules partly sepa-rate endoplasm and preblastodisc do-mains, thus providing spacious path-ways for animal pole-directed flow oforganelles and ribonucleoproteins.Transport of high molecular weightdextran from the yolk cell into theblastodisc in eggs supports this con-clusion. The ooplasm accumulated atthe preblastodisc probably has a dualorigin: part would come from the cyto-plasm originally surrounding the ger-minal vesicle and the rest would be
Fig. 8. Z-sectioned acid-fixed whole-mounted early unfertilized eggs showing the interconnections between ooplasmic domains. A: Image 18 of astack of 54 images that shows an animal pole view of an egg 5 min after activation. The ectoplasm (ec) surrounding the outer surface of the peripheralyolk globules is continuous with the growing blastodisc (bd). B: Image 29 of a stack of 40 images that show the blastodisc/endoplasm boundary zoneof an egg 4 min after activation. Endoplasmic lacunae (en) appear connected to the blastodisc by numerous short channels (arrow heads). C: Image44 of a stack of 64 images of an egg 5 min after activation, showing the growth and close gathering of endoplasmic lacunae. Interconnection betweenlacunae are marked by arrowheads. Scale bars � 100 �m.
664 FERNANDEZ ET AL.
mRNA-rich ooplasm translocated tothe animal pole during oogenesis (seeBally-Cuif et al., 1998). There are rea-sons to suspect that separation of oo-
plasm from yolk and establishment ofooplasmic domains are cytoskeleton-dependent processes. Although littleinformation is available on the orga-
nization of the cytoskeleton in devel-oping oocytes, that the early activatedegg has an ubiquitous cytoskeletonsuggests that a similar one might bepresent in the stage V oocyte.
Postactivation Phase ofOoplasmic Segregation IsManifested by PolewardFlow of Endoplasm andBlastodisc Growth
The available information indicatedthat activation provoked lifting ofthe vitelline envelope, formation ofthe blastodisc, and discharge of thesecond pole cell (see Devillers, 1961).We have provided further informa-tion showing that a blastodisc pre-cursor forms in the stage V oocyteand that blastodisc growth is initi-
Fig. 10. Electron photomicrographs that illustrate the structure of the most-common type ofjunctions between peripheral yolk globules in early unfertilized eggs. A: There are numerous 20 nmin diameter ooplasm-free clefts that extend between parallel running membranes (arrowheads)enveloping neighbor yolk globules (y). B,C: Ooplasm-filled canaliculi are less than 0.25 �m indiameter and include variable number of ribonucleoprotein granules (rp) but lack organelles. Theectoplasm opening of the cleft and canaliculi are situated at the upper end. Scale bars � 0.25 �m.
Fig. 9. Light photomicrographs of stained plastic-embedded thick sections showing reorganization of the cytoplasm in the early unfertilized egg (2min after activation). A: Boundaries between peripheral yolk globules (y) are barely visible, because ooplasm between them have probably beensqueezed out. In this manner, ectoplasm (ec) and endoplasm (en) are more separated than before activation. Most of the cortical granules (cg) havebeen released. B: Loosely arranged yolk globules remain at the blastodisc (bd)/endoplasm (en) boundary zone, thus ensuring persistence of widetransport pathways between the two ooplasmic domains. C: Photomicrograph taken at the center of the egg showing that the boundaries betweeninternal yolk globules are barely visible, while endoplasmic lacunae have enlarged (compare with Fig. 4D). D: Photomicrograph taken at the center ofthe egg showing that type I (I) endoplasmic lacunae have numerous vacuoles (va) scattered across a homogenous cytoplasmic matrix. E: Type II (II)endoplasmic lacunae have less vacuoles and numerous dark bodies (db) scattered across a moderately stained matrix. In mildly preserved type IIlacunae (arrowheads) dark bodies appear scattered across an empty matrix. Notice that canaliculi (ca) linking these lacunae look empty and wider.They are considered to have suffered artifactual expansion. F: High-magnification of type II lacuna. Scale bars � 10 �m in A,D–F, 50 �m in B,C.
EARLY STEPS OF OOPLASMIC SEGREGATION 665
ated immediately after activation asresult of enlargement and redistri-bution of endoplasmic lacunae. En-largement of endoplasmic lacunae isprobably the combined result of fur-ther endoplasm accumulation andcoalescence of neighboring lacunae.Evidence for further endoplasm ac-cumulation comes from the observa-
tion that enlargement of lacunae isaccompanied by narrowing of the oo-plasm-filled canaliculi present be-tween central yolk globules. Thisprocess is the result of granular oo-plasm being squeezed out from be-tween the yolk globules and added tolacunae. We suggest further that oo-plasm translocated to lacunae after
activation includes ribonucleopro-tein granules, whereas ooplasm ac-cumulated in lacunae before activa-tion includes both organelles andribonucleoprotein granules. Earlyworkers of zebrafish eggs such asLewis and Roosen-Runge (1943) andBeams et al. (1985) proposed thatpoleward squeezing of endoplasmtrapped among the yolk globules wasthe mechanism involved in ooplas-mic segregation. However, they werenot aware that an earlier step of thisprocess, consisting in the formationof ooplasmic domains, took place inthe ovary. Centripetal accumulationand alignment of enlarged lacunaealong the animal/vegetal axis of theegg probably favors formation of ax-ial streamers later in development.Axial streamers are vertically run-ning pathways of endoplasm flowingtoward the blastodisc (Lewis andRoosen-Runge, 1943; Leung et al.,2000). Several lines of evidence indi-cate that ectoplasm also moves to-ward the blastodisc after activation.First, polystyrene beads microin-jected in the ectoplasm move towardthe animal pole (Jesuthasan andStrahle, 1996). Second, maternalmRNAs, such as vasa (Howleyand Ho, 2000), reach the blastodiscby mean of an ectoplasmicflow. Third, labeled mitochondria movepoleward across the ectoplasm to reachthe blastodisc (our unpublished obser-vations). Therefore, activation trig-gers both ecto- and endoplasmic flow.Resumption of ooplasmic segregation,as well as other processes triggered byegg activation (cortical reaction andsecond polar body release), are not de-pendent on sperm penetration (Wolen-ski and Hart, 1987) and, hence, occurin both fertilized and unfertilizedeggs. It seems likely, then, that thewave of free cytosolic calcium gener-ated upon activation of the fertilizedor unfertilized egg is the main signalfor resumption of ooplasmic segrega-tion (Lee et al., 1996; Leung et al.,1998; Lee et al., 1999). Free calciumtransients, probably generated simul-taneously in many regions of the egg,would activate signaling pathwaysacting upon internal and externalcomponents of the cytoskeleton en-gaged in ooplasmic flow.
Fig. 11. Electron photomicrographs that show the fine structure of endoplasmic lacunae in earlyunfertilized eggs. A,B: Low- (A) and high- (B) magnification photomicrographs of a type I lacuna havingprofiles of smooth endoplasmic reticula (sER; er) and mitochondria (mi) scattered across a matrix ofribonucleoprotein (rp) granules. This type of lacuna is similar to those found in the stage V oocyte (seeFig. 5D,E). Differences in the appearance of the sER may be due to swelling. C,D: Low-magnificationphotomicrographs showing the structure of type II lacunae. Dark bodies found in the light microscopecorrespond to clusters of mitochondria and profiles of sER (asterisks) scattered across a matrix ofribonucleoprotein granules. E,F: High-magnification photomicrographs showing the distribution oforganelles and ribonucleoprotein granules in type II lacunae. E: Each cluster of organelles consists ofa central sector (asterisk) with medium electron-dense ribonucleoprotein granules and a peripheralsector with mitochondria, sER, and dense core vesicles (dc). The cluster of organelles is surrounded byhighly electron-dense ribonucleoprotein granules. F: Mitochondria, profiles of sER, and ribonucleopro-tein granules also gather at the periphery of the lacuna, in close contact with the surface of the yolkglobules (y). Scale bars � 0.5 �m in B,E (applies to F), 1 �m in A,C,D.
666 FERNANDEZ ET AL.

Activation ProvokesRedistribution of Organellesand RibonucleoproteinGranules in EndoplasmicLacunae
A surprising effect of activation wasthe appearance, across the yolk cell, ofa different type of lacuna called type IIlacuna. Type I and type II lacunaehave similar structures but differ inthe way that their components are ar-ranged. In type I lacunae, ribonucleo-protein granules and most organellesare evenly distributed, whereas intype II lacunae, granules and or-ganelles form conspicuous clusters.Type II lacunae could be newly formed
structures or modified type I lacunae.The last alternative is probably cor-rect, because by the time of activation,most organelles have already been ac-cumulated in type I lacunae and theooplasm remaining among the yolkglobules is essentially granular. In ad-dition, there are transitional lacunaehaving a mixed arrangement of theirorganelles. It is not known how acti-vation provokes such redistributionof organelles and ribonucleoproteingranules. We suspect, however, that[Ca2�] transients accompanying acti-vation might stimulate a cascade ofinternal signals affecting the functionof an internal system of motors andcytoskeleton components. The latter
have been described in this study and,hence, microtubules and microfila-ments may be involved in the rear-rangement of organelles and granulesacross endoplasmic lacunae. Thattype II lacunae are sometimes concen-trated in the animal sector of the yolkcell may be taken as indicative thatthese lacunae may be more activelyinvolved in ooplasmic flow than type Ilacunae. If this presumption is cor-rect, aggregation of organelles and ri-bonucleoprotein granules may expe-dite endoplasmic flow. Assembly ofmitochondria and granular materialin clusters along transport pathwaysis also seen in the leech zygote at thepeak of ooplasmic segregation (Fer-nandez et al., 1998).
Ubiquitous Distribution ofMicrotubules andMicrofilaments Across theEgg Suggests ThatOoplasmic Segregation IsDriven by Both Peripheraland Central Cytoskeletons
Findings reported in this study showthat microtubules and microfila-ments are present throughout thethree ooplasmic domains. These re-sults confirm the work of others inthat actin filaments (Hart et al.,1977, 1992; Hart and Yu, 1980; Ka-tow, 1983; Wolensky and Hart, 1987)and microtubules (Jesuthasan andStrahle, 1996; Gore and Sampath,2002) are present across the ecto-
Fig. 12. A,B: Low- (A) and high- (B) magnification photomicrographs showing type II lacunae ofegg fragments digested with RNase A. A great number of electron-dense granules have beenremoved by the enzyme causing formation of numerous empty spaces. Mitochondria (mi), smoothendoplasmic reticula (sER; er), and some granules (rp) persist at the periphery of the empty spaces.Scale bars � 0.5 �m.
Fig. 13. A,B: Low- (A) and high- (B) magnification electron photomicrographs that show presence of microtubules (mt) and microfilaments (mf) inendoplasmic lacunae. Microtubules are scattered singly or in small loose bundles running into different directions near profiles of sER (er) andribonucleoprotein particles (rp). Microfilaments are seen forming bundles adjacent to microtubules. C: Microfilament bundles in the endoplasm aresimilar to those seen in the cortical ectoplasm. Individual microfilaments are indicated by arrowheads. pm, plasma membrane. Scale bars � 0.2 �m.
EARLY STEPS OF OOPLASMIC SEGREGATION 667

plasm. However, there is only onereport (Beams et al., 1995) based onthe observation of zebrafish eggs un-der the scanning electron micro-scope, in which the authors inter-preted lineal elements as belongingto microtubules or microfilaments.For reasons explained in the text, itwas not possible to compare the cy-toskeleton of stage V oocytes withthat of early developing eggs. Forthat reason, we could not tracechanges in the cytoskeleton relatedto the first steps of ooplasmic segre-gation. However, that the cytoskele-ton of developing eggs remains em-bedded across the ooplasm indicatesthat sliding of microtubules and ac-tin filaments may provide the forcefor ooplasmic flow across both ecto-and endoplasm. This alternativedoes not exclude the possibility thatmicrotubules and actin filamentsmay also function as tracks alongwhich organelles and ribonucleoproteinswould move thanks to the activity ofmolecular motors.
Fig. 14.
Fig. 15.
668 FERNANDEZ ET AL.

EXPERIMENTALPROCEDURES
Collection and Handling ofOocytes and Eggs
Adult zebrafish (Danio rerio) weremaintained at 28°C in aeratedaquaria subjected to a 14-hr light/10-hr dark photoperiod. Males and fe-males were separated a day beforeand crossed the next day. Nontranslu-cent medium and large sized stage IIIoocytes (400–600 �m in diameter),and translucent stage V oocytes (ap-proximately 750 �m in diameter),were surgically removed from the ova-ries. Selected stage III oocytes had acentrally located meiotic nucleus (seeSelman et al., 1993). Stage V oocyteswere also procured by gently pressingthe abdomen of pregnant mothersanesthetized with tricaine (Sigma).Stage IV oocytes are difficult to iden-tify, because they overlap in size withlarge stage III oocytes and are rapidlyconverted in stage V oocytes (Selmanet al., 1993; Li et al., 1993). For thisreason, they were not included in thiswork. Early activated stage V oocytes(unfertilized eggs) were obtained bybriefly soaking the cells in filteredtank water. A small fraction of thestage V oocytes removed from theovary failed to be activated. Fertilizedeggs were obtained by natural or arti-
ficial procedures. Fertilized and unfer-tilized eggs developing in conditionedtank water at 20°C were removed formicroscopic examination at differenttime points during the first 5 min af-ter activation. To determine the effectof temperature and fertilization on oo-plasmic segregation, development offertilized and unfertilized eggs werecompared at 20°C and 28°C.
Light Microscopy
Preparation of whole-mountedacid-fixed oocytes and eggs.
Stage V oocytes and eggs are translu-cent, but their ooplasm may be ren-dered opaque by fixing the cells in tis-sue culture dishes (35 � 10 mm,Sarstedt) using freshly prepared neu-tralized 10% formaldehyde, to which afew droplets of glacial acetic acid (2–3droplets in 4–5 ml of formaldehyde)were added at the beginning of thefixation (Fernandez, 1980). The cho-rion did not interfere with the acidfixation, but it could be removed tofacilitate positioning of the oocyte dur-ing microscopic examination. Formore detailed observation, acid-fixedcells may be further cleared by trans-ferring them to a 1:1 mixture of glyc-erol and formaldehyde/acetic acid.Study of the displayed images wereimproved by converting opacity into
brightness, using the reverse contrastfacility of the Metamorph program.
Loading cells with labeledproteins and dextrans.
To test the permeability of the ecto-plasm/endoplasm yolk barrier thetransport of proteins and dextranswere studied in eggs microinjectedshortly after activation. The probeswere injected with micropipettes pre-pared from borosilicate glass capillar-ies (Harvard Apparatus Ltd.) using aNarishige IM 300 microinjector. A to-tal of 4 to 8 nl of the probes weremicroinjected into different regions ofthe yolk cell: close to the blastodisc,close to the ectoplasm or at the eggcenter. The amount of injected tracerrepresented less than 5% of the eggvolume (approximately 200 nl).Labeled proteins were column-purifiedand its purity tested by electrophoresisand Western blot. The following labeledproteins were used: rhodamine-labeleddimeric tubulin (approximately 45-kDa,see Fernandez et al., 2002, 2004), Alexafluor 594–labeled bovine serum albu-men (approximately 66-kDa, MolecularProbes). These probes were dissolved inglutamate buffer (Hyman et al., 1991)to a final concentration at the pipette of12–90 �M. Dextrans were also dis-solved in glutamate injection buffer to afinal concentration at the pipette of25–50 mM. Fluorescein-labeled dex-trans (Molecular Probes) of the follow-ing molecular weight were tested: 10kDa, 70 kDa, and 500 kDa. All microin-jected proteins and dextrans.
Preparation of stained sectionsof resin-embedded oocytes andeggs.
Cells were fixed and postfixed as forelectron microscopy (see below) andembedded in JB-4 (Polysciences), asoft mixture of Epon–Araldite or hardEpon 812 (Pelco) resins. In the firstcase, 1- to 2-�m-thick sections werestained with methylene blue and basicfuchsine and in the second case, with1% toluidine blue in borate buffer.Sections obtained with a MT 5000 Sor-vall ultramicrotome were floated anddried in an aqueous diluted ammoniasolution.
Video microscopy and image pro-cessing. Live, microinjected and acid-fixed cells were examined in a Zeiss
Fig. 14. Low-magnification micrographs of whole-mounted live zygotes showing the distribution offluorescein-labeled dextrans microinjected shortly after activation. The site of injection is marked byan asterisk. A: Zygote microinjected with 10-kDa dextran close to the cell center. The probediffused across endoplasmic lacunae (en) and reached the ectoplasm (ec) and blastodisc (bd).B: Zygote microinjected with 70-kDa dextran close to the cell center. The probe also diffusedacross the endoplasmic lacunae and reached the ectoplasm that became labeled. C: Egg micro-injected with 500-kDa dextran under the blastodisc. The probe diffused across the endoplasm andblastodisc but the ectoplasm remained unlabeled. For this reason, a fluorescent rim is absent at thezygote periphery. D,E: Zygotes microinjected at the cell center with 500-kDa dextran. In bothcases, the probe diffused across the endoplasm but did not accumulate in the ectoplasm. F: Eggswere microinjected with 500-kDa dextran at the periphery of the cell, so that the probe infiltratedboth the ecto- and endoplasm. Under these conditions, the heavy molecular weight dextrandiffused across the endoplasm and ectoplasm, where it formed a fluorescent rim. Scale bar � 100�m.
Fig. 15. Combined phase contrast and confocal images of whole-mounted live zygotes showingthe distribution of 500-kDa fluorescein-labeled dextran microinjected shortly after activation. Thesite of injection is marked with an asterisk. A: The probe was microinjected close to the cell centerand entered the blastodisc (bd). Notice that dextran accumulated in the blastodisc did not diffuseinto the ectoplasm (ec). B,C: Optical section 20 (B) and 35 (C) of a stack of 37 images taken every3.3 �m The probe was microinjected close to the zygote center from where it diffused acrossendoplasmic lacunae and entered the blastodisc. Very little of the probe gained direct access to theectoplasm (arrow in the inset). D: Zygote microinjected at the periphery of the cell so that the probeinfiltrated both the ecto- and endoplasm. Notice transportation of abundant labeled dextran acrossboth the endoplasm and ectoplasm. White spots in the images are not empty spaces but phasecontrast effects. Scale bar � 100 �m.
EARLY STEPS OF OOPLASMIC SEGREGATION 669

135 inverted fluorescence microscopeequipped with a Z motor (Prior) and aHamamatsu chilled CCD camera(Model C5985). For lower magnifica-tion, Achroplan objectives (�10–40)were used. For higher magnification, aPlan Neofluar objective (�100, NA1.4) and the Optovar (�1.6–2.5) al-lowed the projection of images ontothe camera chip at 160–250 magnifi-cation. Image grabbing and analysiswere performed on a PC computer us-ing the Metamorph (6.1 version) pro-gram. This strategy allowed prepara-tion of stacks of images for time lapseand Z-sectioning analysis, elaborationof superficial topographic maps, bestfocus, and deconvolution. Assignmentof pseudo coloring was possible in 24bit-converted images. Further pro-cessing of the images was performedwith a 7.0 version of Adobe Photoshop.For observation, cells were placed inFalcon 3072 tissue culture wells or be-tween two coverslips separated byplasticine stoppers. The coverslipswere mounted on the window of a me-tallic slide and the edges sealed withVaseline. This type of cell mountingallowed better resolution of the im-ages and inspection of the two sides ofthe cells by turning the preparation180 degrees.
Confocal Microscopy
Whole-mounted eggs microinjectedwith labeled proteins or dextrans weremounted as for video microscopy andexamined in a Zeiss LSM 510 METAlaser scanning microscope, equippedwith argon and He-Ne lasers. Time-lapse series of microinjected eggs weretaken every 5–10 min, and Z-sectionswere performed every 3–5 �m. Galleryof images allowed preparation of ani-mations or reconstructions.
Transmission ElectronMicroscopy
Preparation and observation of sam-ples. Use of several fixative solutionsand embedding media indicated thatthe fine structure of the zebrafish oo-cyte and egg was difficult to preserve.The best results were obtained withthe use of the Karnovsky fixative andthe application of a slow resin-embed-ding procedure. The fixative contained
0.1% tannic acid (Merck), 1 �g/ml ofphalloidin (Sigma), and 1 �g/ml ofTaxol (Molecular Probes). Whole oo-cytes and mechanically dechorionatedeggs were fixed for 2–2:30 hr at roomtemperature. During fixation, cellswere sliced to favor fixative and resinpenetration. After rinsing in cacody-late buffer (containing the same con-centration of phalloidin and Taxol) forthe same amount of time, cells werepostfixed for 1:30 hr in 1% OsO4 in thecacodylate buffer at room temperatureand darkness. After dehydration ingraded ethanol, cells were embeddedin Epon 812 under vacuum. For thispurpose, cells remained for approxi-mately 12 hr in each of the followingEpon/propylene oxide mixtures: 1:2,1:1, and 2:1. They finally remained for36 hr in pure Epon replaced every 12hr. Thin sections were double stainedwith uranyl acetate and lead citrateand examined with a Jeol 100 SX elec-tron microscope. Photographic nega-tives were scanned with an Epson Ex-pression 1600 Professional scannerand processed with Adobe Photoshopin a G3 McIntosh computer.
Preservation of microtubules andmicrofilaments was greatly improvedby pretreatment of the eggs with amixture of cytoskeleton stabilizers.For this purpose, chorionated acti-vated eggs were microinjected at theequatorial region with a mixture of 5mM Taxol (4 nl) and 0.8 �M phalloidin(10 nl) in dimethyl sulfoxide. After 10min, eggs were fixed, rinsed, dehy-drated, and embedded as outlinedabove.
RNase Digestion
Egg fragments fixed as for electronmicroscopy were rinsed for 2 hr in ca-codylate buffer and then incubated for45 min in RNase A (Sigma) with con-tinuous agitation. RNase was dilutedin cacodylate buffer pH 7.6 at a con-centration of 10 �g/ml. After bufferrinsing, digested egg fragments werepost-fixed and embedded in Eponresin.
ACKNOWLEDGMENTSWe thank Vıctor Guzman, Raul Fu-entes, and Lorena Saragoni for tech-nical assistance and Dr. John Nichollsand Duncan Stuart for critical readingof the manuscript.
REFERENCES
Abraham VC, Gupta S, Fluck RA. 1993.Ooplasmic segregation in the medakaOryzias latipes egg. Biol Bull 184:115–124.
Bally-Cuif L, Schatz WJ, Ho RK. 1998.Characterization of the zebrafish Orb/CPEB-related RNA-binding protein andlocalization of maternal components inthe zebrafish oocyte. Mech Dev 77:31–47.
Beams HW, Kessel RG, Shih CY, TungHN. 1985. Scanning electron microscopystudies on blastodisc formation in thezebrafish, Brachidanio rerio. J Morphol184:41–39.
Bowerman B. 1999. The maternal controlof polarity and patterning during embry-ogenesis in the nematode Caenorhabdi-tis elegans. In: Moody SA, editor. Celllineage and fate determination. San Di-ego: Academic Press. p 97–117.
Danilchik MV, Denegre JM. 1991. Deepcytoplasmic rearrangements duringearly development in Xenopus laevis. De-velopment 111:845–856.
Devillers CH. 1961. Structural and dy-namic aspects of the development of theteleostean egg. Adv Morphol 1:379–428.
Elinson RP, Rowning B. 1988. A transientarray of parallel microtubules in frogeggs: potential tracks for a cytoplasmicrotation that specifies the dorso-ventralaxis. Dev Biol 128:185–197.
Fernandez J. 1980. Embryonic develop-ment of the glossiphoniid leech Theromy-zon rude: characterization of develop-mental stages. Dev Biol 76:245–262.
Fernandez J, Olea N. 1982. Embryonic de-velopment of glossiphoniid leeches. In:Harrison FW, Cowden RR, editors. De-velopmental biology of freshwater inver-tebrates. New York: Alan R. Liss. p 317–366.
Fernandez J, Olea N, Ubilla A, CantillanaV. 1998. Formation of polar cytoplasmicdomains (teloplasms) in the leech egg isa three-step segregation process. Int JDev Biol 42:149–162.
Fernandez J, Cantillana V, Ubilla A. 2002.Reorganization and translocation of theectoplasmic cytoskeleton in the leech zy-gote by condensation of cytasters and in-teractions of dynamic microtubules andactin filaments. Cell Motil Cytoskeleton53:214–230.
Fernandez J, Toro J, Ubilla A. 2004. Con-focal and video imaging of cytoskeletondynamics in the leech zygote. Dev Biol271:59–74.
Gore AV, Sampath K. 2002. Localization oftranscripts of the zebrafish morphogensquint is dependent on egg activationand the microtubule cytoskeleton. MechDev 112:153–156.
Hart NH, Fluck RA. 1985. Cytoskeleton inteleost eggs and early embryos: contribu-tions to cytoarchitecture and motileevents. In: Capco ED, editor. Currenttopics in developmental biology. San Di-ego: Academic Press. p 343–381.
Hart NH, Yu S. 1980. Cortical granule exo-cytosis and cell surface reorganization in
670 FERNANDEZ ET AL.

eggs of Brachidanio rerio. J Exp Zool213:137–159.
Hart NH, Yu SF, Greenhut VA. 1977. Ob-servations on the cortical reaction ineggs of Brachidanio rerio as seen withthe scanning electron microscope. J ExpZool 201:325–331.
Hart NH, Becker KA, Wollenski JS. 1992.The sperm entry site during fertilizationof the zebrafish egg: localization of actin.Mol Reprod Dev 32:217–228.
Hess O. 1971. Freshwater gastropoda. In:Reverberi G, editor. Experimental em-bryology of fresh-water invertebrates.Amsterdam: New Holland-Publ Co. p215–247.
Hisaoka KK, Battle HI. 1958. The normaldevelopmental stages of the zebrafish,Brachidanio rerio (Hamilton-Buchanan). J Morphol 107:205–226.
Hisaoka KK, Firlit CF. 1960. Further stud-ies on the embryonic development of thezebrafish, Brachidanio rerio (Hamilton-Buchanan). J Morphol 107:205–226.
Ho RK. 1992. Axis formation in the embryoof the zebrafish, Brachidanio rerio. Se-min Dev Biol 3:53–64.
Howley C, Ho RK. 2000. mRNA localiza-tion patterns in zebrafish oocytes. MechDev 92:305–309.
Hyman E, Drechsel D, Kellog D, Salser S,Sawin K, Steffen P, Wordeman L,Mitchison T. 1991. Preparation of modi-fied tubulins. Methods Enzymol 196:478–485.
Ivanenkov VV, Meshcheryakov VN, Marty-nova LE. 1990. Surface polarization inloach eggs and two-cell embryos: correla-tions between surface relief, endocytosisand cortex contractility. Int J Dev Biol34:337–349.
Jeffery WR, Meier S. 1983. A yellow cres-cent cytoskeletal domain in ascidianeggs and its role in early development.Dev Biol 96:125–143.
Jesuthasan S, Strahle U. 1996. Dynamicmicrotubules and specification of the ze-brafish embryonic axis. Curr Biol 7:31–42.
Katow H. 1983. Obstruction of blastodiscformation by cytochalasin B in the ze-brafish, Brachidanio rerio. Dev GrowthDiffer 25:477–484.
Kimmel C, Ballard WW, Kimmel SR, Ull-mann B, Schilling TF. 1995. Stages ofembryonic development in the zebrafish.Dev Dyn 203:253–310.
Lee KW, Baker R, Galione A, Gilland EH,Hanlon RT, Miller AL. 1996. Ionophore-induced calcium waves activate unfertil-ized zebrafish (Danio rerio) eggs. BiolBull 191:265–267.
Lee KW, Webb SE, Miller AL. 1999. Awave of free cytosolic calcium traverseszebrafish eggs on activation. Dev Biol 214:168–180.
Leung CF, Webb SE, Miller AL. 1998. Cal-cium transients accompany ooplasmicsegregation in zebrafish embryos. Dev.Growth Differ 40:313–326.
Leung CF, Webb SE, Miller AL. 2000. Onthe mechanism of ooplasmic segregationin single-cell zebrafish embryos. DevGrowth Differ 42:29–40.
Lewis WH, Roosen-Runge EC. 1943. Theformation of the blastodisc in the egg ofthe zebrafish, Brachidanio rerio. AnatRec 85:326.
Li S, Mao Z, Han W, Sun Z, Yan W, ChenH, Yan S. 1993. In vitro oocyte matura-tion in the zebrafish, Brachidanio rerio,and the fertilization and development ofthe mature egg. Chin J Biotechnol 9:247–255.
Oppenheimer JM. 1936. The developmentof isolated blastoderms of Fundulus het-eroclitus. J Exp Zool 72:247–269.
Pelegri F. 2003. Maternal factors in ze-brafish. Dev Dyn 228:535–554.
Reverberi G. 1971. Ascidians. In: ReverberiG, editor. Experimental embryology ofmarine and fresh-water invertebrates.Amsterdam: North-Holland Publ Co. p507–549.
Roosen-Runge EC. 1938. On the early de-velopment-bipolar differentiation andcleavage of the zebrafish, Brachidaniorerio. Biol Bull 75:119–133.
Sardet C, Speksnijder J, Inoue S, Jaffe L.1989. Fertilization and ooplasmic move-ments in the ascidian egg. Development105:237–249.
Schirone RC, Gross L. 1968. Effect of tem-perature on early embryological develop-ment of the zebra fish, Brachidanio rerio.J Exp Zool 169:43–52.
Selman K, Wallace RA, Sarka A, XiaopingQI. 1993. Stages of development in thezebrafish, Brachidanio rerio. J Morphol218:203–224.
Shimizu T. 1982. Ooplasmic segregation inthe Tubifex egg: mode of pole plasm ac-cumulation and possible involvement ofmicrofilaments. Roux Arch Dev Biol 191:246–256.
Strome S, Wood WB. 1983. Generation ofasymmetry and segregation of germ-linegranules in early C. elegans embryos.Cell 35:15–25.
Susuki H, Maegawa S, Nishibu T, Sug-iyama T, Yasuda K, Inoue K. 2000. Veg-etal localization of the maternal mRNAencoding an EDEN-BP/Bruno-like pro-tein in zebrafish. Mech Dev 93:205–209.
Tung TC, Chang CY, Tung YFY. 1945. Ex-periments on the developmental poten-ties of blastoderms and fragments of te-leostean eggs separated latitudinally.Proc Zool Soc Lond 115:175–188.
Wolenski JS, Hart NH. 1987. Scanningelectron microscope studies of sperm in-corporation into the zebrafish (Brachy-danio) eggs. J Exp Zool 246:206–215.
EARLY STEPS OF OOPLASMIC SEGREGATION 671