
Discuss nursing interventions that prevent complications of immobility

1521-0103/358/2/271–281$25.00 http://dx.doi.org/10.1124/jpet.116.233338THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS J Pharmacol Exp Ther 358:271–281, August 2016Copyright ª 2016 by The American Society for Pharmacology and Experimental Therapeutics
Ontogeny of SERT Expression and Antidepressant-like Responseto Escitalopram in Wild-Type and SERT Mutant Mice
Nathan C. Mitchell, Georgianna G. Gould, Wouter Koek, and Lynette C. DawsDepartments of Physiology (N.C.M., G.G.G., L.C.D.), Psychiatry (W.K.), and Pharmacology (L.C.D., W.K.), University of TexasHealth Science Center, San Antonio, Texas
Received March 4, 2016; accepted June 8, 2016
ABSTRACTDepression is a disabling affective disorder for which the majorityof patients are not effectively treated. This problem is exacerbatedin children and adolescents for whom only two antidepressantsare approved, both of which are selective serotonin reuptakeinhibitor (SSRIs). Unfortunately SSRIs are often less effectivein juveniles than in adults; however, the mechanism(s) underly-ing age-dependent responses to SSRIs is unknown. To this end,we compared the antidepressant-like response to the SSRIescitalopram using the tail suspension test and saturation bindingof [3H]citalopram to the serotonin transporter (SERT), the primarytarget of SSRIs, in juvenile [postnatal day (P)21], adolescent (P28),and adult (P90) wild-type (SERT1/1) mice. In addition, to modelindividuals carrying low-expressing SERT variants, we studiedmice with reduced SERT expression (SERT1/2) or lacking SERT
(SERT2/2). Maximal antidepressant-like effects were less inP21 mice relative to P90 mice. This was especially apparent inSERT1/2 mice. However, the potency for escitalopram to pro-duce antidepressant-like effects in SERT1/1 and SERT1/2micewas greater in P21 and P28 mice than in adults. SERT expressionincreased with age in terminal regions and decreased with age incell body regions. Binding affinity values did not change as afunction of age or genotype. As expected, in SERT2/2 miceescitalopram produced no behavioral effects, and there was nospecific [3H]citalopram binding. These data reveal age- andgenotype-dependent shifts in the dose-response for escitalopramto produce antidepressant-like effects, which vary with SERTexpression, and may contribute to the limited therapeutic re-sponse to SSRIs in juveniles and adolescents.
IntroductionDepression is a disabling affective disorder for which the
majority of patients are not effectively treated. This problemis exacerbated in juveniles and adolescents by limited thera-peutic options (Bylund and Reed, 2007). Only two selectiveserotonin (5-HT) reuptake inhibitor (SSRI) antidepressants,escitalopram and fluoxetine, have U.S. Food and DrugAdministration approval as treatments for pediatric depres-sion. SSRIs act on the serotonergic system by blocking the5-HT transporter (SERT) (SLC6A4), the high affinity clear-ance mechanism for extracellular 5-HT. The resultant in-crease in extracellular 5-HT is thought to trigger therapeuticdownstream effects. However, escitalopramand fluoxetine areoften less effective in treating symptoms of depression injuveniles and adolescents than in adults (Tsapakis et al., 2008;Hetrick et al., 2007, 2010). Furthermore, in adolescentscertain SERT gene variants are associated with increasedrisk of developing SSRI-resistant depression after stressfullife events, triggering a lifelong struggle with the disorder
(Serretti et al., 2007; Petersen et al., 2012). Given the highincidence of depression in adolescents, which comprises 4%–
8% of the U.S. population, and the fact that suicide is the thirdmost likely cause of death in this age group, there is a distinctneed to uncover mechanism(s) limiting the therapeutic bene-fits of SSRIs in young patients (Kessler et al., 2001; Bujoreanuet al., 2011). A better understanding of the age dependency ofSSRI efficacy is therefore an important first step towardimproving therapeutics for pediatric depression.Rodents are indispensable tools for studying the behavioral
and neurochemical effects of antidepressants. However, moststudies to date have focused on antidepressant-like behaviorin adult rodents and have largely ignored the use of juvenilesand adolescents. One study reported that SSRIs reducedimmobility time in the forced swim test (FST), an index ofantidepressant-like action, in juvenile [postnatal day (P)21]and adolescent (P28–P35) rats (Reed et al., 2008). Recently, wediscovered differences in behavioral responses to SSRIs injuvenile mice. We used escitalopram at a dose of 10 mg/kg,which is known to produce maximal antidepressant-likeeffects in adult mice in the tail suspension test (TST), anassay commonly used to assess antidepressant-like activity inmice (Cryan et al., 2005; O’Leary et al., 2007). At this dose theantidepressant-like effect of escitalopram was less in juvenilemice than in adult mice (Mitchell et al., 2013), a findingconsistent with clinical reports. The present study expandsupon this initial finding by examining the dose dependency of
This work was supported by the National Institutes of Health NationalInstitute of Mental Health [Grants MH106978, MH093320, and MH086708]and the Congressionally Directed Medical Research Programs [AwardAR110109].
The research was conducted in the absence of any commercial or financialrelationships that could be construed as a potential conflict of interest.
dx.doi.org/10.1124/jpet.116.233338.
ABBREVIATIONS: ANOVA, analysis of variance; Bmax, maximal binding; CA, cornu ammonis; Emax, maximal effect; FST, forced swim test; 5-HT,serotonin; Kd, affinity; P, postnatal day; SERT, serotonin transporter; SSRI, selective serotonin reuptake inhibitor; TST, tail suspension test.
271
at ASPE
T Journals on A
ugust 28, 2019jpet.aspetjournals.org
Dow
nloaded from

escitalopram’s antidepressant-like effects in juvenile (P21), ado-lescent (P28), and adult (P90) mice. In addition, because individ-uals carrying low-expressing variants of SERT are often reportedto benefit less from treatment with SSRIs than individuals whodo not harbor such gene variants, we studied mice with reducedSERT expression (SERT1/2) or lacking SERT (SERT2/2) andcompared them to wild-type mice (SERT1/1).In murine models, serotonergic neuronal innervation, den-
dritic connections, and extracellular 5-HT concentration havebeen shown to be similar to adults by P21 (Loizou and Salt,1970; Wallace and Lauder, 1983; Miranda-Contreras et al.,1998). These studies suggest that some aspects of the centralserotonergic system are functionally developed in juveniles;however, a developmental lag in SERT expression or functioncould limit the therapeutic benefit of SSRIs. In rats, SERTexpression reaches adult levels between birth and P21 withexpression patterns varying in different brain regions (Mollet al., 2000; Galineau et al., 2004; Bylund and Reed, 2007;Slotkin et al., 2008). However, these studies did not evaluatepossible developmental changes in SERT affinity (Kd). Re-cently, we discovered that SERT binding properties in juve-nile (P21) and adolescent (P28) mice may differ from adults.While maximal binding (Bmax) of the SERT-selective ligand,[3H]citalopram, in hippocampal membranes revealed no dif-ferences among ages, variance in the Kd value for P21 micewas significantly greater than in older mice (Mitchell et al.,2013), suggesting that the functionality of SERT may be at adevelopmentally dynamic transitional stage. To better char-acterize age- and brain region–dependent changes in SERTfunction, the present study used quantitative autoradiography,an anatomicallymore sensitive assay, to examine [3H]citaloprambinding properties and their relation to behavior in the TST.
Materials and MethodsAnimals
Maleand femaleSERTwild-type (SERT1/1), heterozygote (SERT1/2),or homozygote knockout (SERT2 /2) mice (backcrossed toC57BL/6J for.10 generations) were used for all experiments. Colonyfounders were provided by Dr. Dennis Murphy (National Institute ofMental Health, Bethesda, MD), and were bred and identified aspreviously described (Bengel et al., 1998). P21 (juvenile), P28 (adoles-cent), and P90–P100 (adult) mice (Spear, 2000) were used. Mice ofeach genotype were generated by crossing male and female SERT1/2mice. Animals were housed in a temperature-controlled (24°C)vivariummaintained on a 12/12-hour light/dark cycle (lights on at 7:00AM) in plastic cages (29 � 18 � 13 cm) containing rodent bedding(Sani-chips; Harlan Teklad, Madison, WI) with free access to food(irradiated rodent sterilizable diet; Harlan Teklad) and water. Afterweaning on P28, mice were housed in groups of five with same-sexpeers. TST experiments were conducted prior to weaning inmice agedP21 and P28. All procedures were conducted in accordance with theNational Institutes of Health Guide for the Care and Use of LaboratoryAnimals (Institute of Laboratory Animal Resources, Commission on LifeSciences, National Research Council, https://grants.nih.gov/grants/olaw/Guide-for-the-Care-and-use-of-laboratory-animals.pdf), and with the ap-proval of the Institutional Animal Care and Use Committee at theUniversity of Texas Health Science Center.
Tail Suspension Test
The TST was conducted as originally described by Steru et al. (1985)(for a review, see Castagné et al., 2011). Naivemice weremoved from thecolony room to the testing room and allowed a 1- to 2-hour acclimation
period. All experiments were conducted between 12:00 and 5:00 PM. Allmice received saline vehicle i.p. 1 hourbefore testing, followed30minuteslater by s.c. injection of either escitalopram (0.1, 0.32, 1.0, 3.2, or10.0 mg/kg) or saline vehicle (control condition). This drug administra-tion protocol was selected to be consistent with previously publishedprocedures (Baganz et al., 2008;Horton et al., 2013;Mitchell et al., 2015).In addition, administration of antidepressant 30 minutes before testingis standard (Steru et al., 1985; Holmes et al., 2002; Ripoll et al., 2003).Immediately before testing, the distal portion of the tail was fastened to aflat aluminum bar (2� 0.3� 10 cm) using adhesive tape placed at a 90°angle to the longitudinal axis of the mouse tail, with 3 to 4 cm betweenthe base of the mouse tail and the aluminum bar. A hole opposite thetaped endof thebarwasused to secure the bar to ahook on the ceiling of avisually isolatedwhite box (40� 40� 40 cm).Eachmousewas suspendedby its tail for 6 minutes, allowing the ventral surface and the front andhind limbs to be recorded using a digital video camera facing the testingbox. Total immobility time was measured (in seconds) during the6-minute time period. Immobility was defined as the absence of initiatedmovements, and included passive swaying of the body. A mouse wasexcluded from the study if it climbed and held its tail or the aluminumbar for a period of 3 seconds or longer. Immobility was scored twice fromthe videos by observers who were blind to the treatments. Each mousewas tested only once, and allmicewere randomlyassigned to treatments.
Drugs
Escitalopram [s-citalopram or (S)-1[3-(Dimethylamino)propyl]-1-(4-fluorophenyl)-1,3-dihydroisobenzofuran-5-carbonitrile] oxalate(Sigma-Aldrich, St. Louis,MO) was dissolved in physiologic saline andinjected s.c. at doses expressed as base weight per kilogram bodyweight. The injection volume was 10 ml/kg. Drug doses were chosenthat ranged from subthreshold (0.32 mg/kg) to maximally effective(10 mg/kg) in adult SERT1/1 mice.
Autoradiography
SERT saturation binding was assessed in mouse brain by quantita-tive autoradiography using the SERT-selective ligand [3H]citalopramand methods adapted from D’Amato et al., (1987). Naive mice werekilled by decapitation and brains rapidly removed and frozen onpowdered dry ice before being stored at 280°C until sectioned forquantitative autoradiography. Brains were brought to 220°C in acryostat (Leica CM 1850,Meyer Instruments, Houston, TX) and coronalsections (20 mm) were collected at the level of plate 12 (prefrontalcortex), plate 47 [hippocampal subregions, cornu ammonis (CA) 1–3 anddentate gyrus; parietal cortex; amygdala; and ventromedial hypothal-amus], and plate 64 (raphe nuclei) according to the Paxinos andFranklin (1997) mouse brain atlas. Sections were thaw mounted ontogelatin-coated microscope slides, vacuum desiccated for 18–24 hours at4°C and stored at280°C until use. In preparation for binding, sectionswere thawed and then preincubated for 1 hour in a 50 mM Tris-HCl,120 mM NaCl, and 5 mM KCl pH 7.4 at room temperature (∼24°C).Incubation was carried out in slide mailers (VWR International,Radnor, PA) filled with 10 ml of the same buffer containing[3H]citalopram at concentrations of 0.32, 0.56, 1.0, 3.2, or 5.6 nM for1 hour at room temperature. Nonspecific binding was defined by20 mM sertraline (Pfizer, Groton, CT) and was ∼11%–23% totalbinding in low-binding regions (i.e., parietal cortex) and ∼5%–9% totalbinding in high-binding regions (i.e., dorsal raphe). The incubationwas terminated by two 10-minute washes in 50mMTris-HCl, 120mMNaCl, 5 mMKCl pH 7.4 at 4°C, followed by a 5-second dip in deionizedwater at 4°C. Slides were dried on a slide warmer for 20 minutes.[3H]citalopram-labeled sections were exposed to Carestream BiomaxMR film for 6 weeks, along with tritium standards (American Radio-labeled Chemicals, St Louis, MO). Films were developed in a film pro-cessor (AFP imaging, Elmsford, NY). Autoradiogram images werecaptured on a digital image system: 12-digital bit camera (CFW-1612M,Scion Corp., Frederick, MD), Nikon Lens, Northern Lights Illuminator,and Kaiser RS1 copy stand (all from InterFocus Imaging Ltd., Linton,
272 Mitchell et al.
at ASPE
T Journals on A
ugust 28, 2019jpet.aspetjournals.org
Dow
nloaded from

United Kingdom), and were calibrated and measured with the NationalInstitutes of Health Image J public access shareware https://imagej.nih.gov/ij/download.html on a MacBook (OS 10).
Data Analysis
Statistical analyses were performed using Prism 6.0 (GraphPad,San Diego, CA).
TST. Under vehicle conditions, Mitchell et al. (2013, 2015) pre-viously found that time spent immobile varies by age and SERTgenotype. Because similar findings were obtained here (Fig. 1A), dose-
response datawere expressed as percentage fromvehicle control (Fig. 1,B–D) and analyzed by two-factor analysis of variance (ANOVA), i.e., ageand drug dose, followed by Tukey’s andDunnett’smultiple comparisonstests. Within each genotype and age group there were no significantdifferences in drug-induced immobility between males and females(main effect of sex:P$ 0.11; interaction between sex and dose,P$ 0.10;data not shown); thus, data for both sexes were pooled. Maximal effect(Emax) and half-maximally effective dose (ED50) values, derived fromdata shown in Fig. 1, B and C are shown in Fig. 1, E and F. All data areexpressed as mean 6 S.E.M., except the ED50 values, which areexpressed as themean.P, 0.05was considered statistically significant.
Fig. 1. Escitalopram reduces immobility time in the TST in juvenile, adolescent, and adult mice as a function of SERT genotype. (A) Basal immobility time(saline-injected controls) in seconds across age and SERT genotype. Solid symbols represent a significant difference from P21 with Dunnett’s post hocmultiple comparisons test after two-factor ANOVA (age, SERT genotype) *P , 0.05 represents the difference from age-matched SERT+/+ and #P , 0.05represents the difference from age-matched SERT+/2with Tukey’s post hoc multiple comparisons test. (B–D) Escitalopram dose-effect curves across age inSERT+/+, SERT+/2, and SERT2/2mice. Immobility expressed as a percentage of saline-injected controls. Solid symbols represent a significant differencefrom vehicle treatment with Dunnett’s post hoc multiple comparisons test after two-factor ANOVA (age, drug dose) performed separately for each genotype.*P , 0.05, **P , 0.01 represent a significant difference from SERT gene–matched P90; Tukey’s post hoc multiple comparisons test. (E) Half-maximallyeffective dose (ED50); *P, 0.05, **P, 0.01 represent a significant difference fromSERT gene–matched P90; † P, 0.05 P90 SERT+/+ versus P90 SERT+/2.(F) Maximal effect (Emax, expressed as a percent from control) of dose-effect curves (B and C); *P , 0.05, **P , 0.01 represent a significant difference fromSERT gene–matched P90 with Tukey’s post hoc multiple comparisons test after two-factor ANOVA (age, SERT genotype). There were no statisticallysignificant main effects of sex or sex� dose interactions in any age group or SERT genotype; therefore, data for both genders were pooled. Data are mean6S.E.M., except for ED50 values, which are expressed asmean. SERT+/+, n = 17–31 (males n = 4–16 and females n = 9–21 pooled); SERT+/2, n = 14–20 (malesn = 8–10 and females n = 6–11 pooled); and SERT2/2, n = 18–26 (males n = 9–14 and females 8–12 pooled) per data point.
Ontogeny of SERT Expression and Antidepressant-like Response 273
at ASPE
T Journals on A
ugust 28, 2019jpet.aspetjournals.org
Dow
nloaded from

The Emax value was defined as the greatest observed percentagechange in immobility from the saline control condition. To calculatetheED50 values, time spent immobilewas expressed as the percentageof the vehicle control, and the linear portion of the dose-responsecurves was analyzed by log-linear regression of data from individualsubjects with the following equation: effect 5 slope � log(dose) 1intercept, using methods detailed elsewhere (Koek et al., 2009).Maximal effects were analyzed using two-factor ANOVA (age andgenotype) followed by Tukey’s multiple comparisons test.
Quantitative Autoradiography. Nonspecific binding was sub-jected to unweighted linear regression and subtracted from total[3H]citalopram binding to give specific binding. Specific [3H]citaloprambinding data were submitted to unweighted nonlinear regression andthe saturation binding isotherm fitted according to a one-site model:Y5Bmax * X/(Kd1 X). MeanBmax andKd values were analyzed usingtwo-factor (age and genotype) ANOVA (Fig. 4), with Tukey’s posthoc test for multiple comparisons. Within genotype-, age-, andbrain region–matched groups there were no statistically significantdifferences in Bmax and Kd values for [3H]citalopram bindingbetween males and females; thus, data from both sexes were pooled(P$ 0.051) (data not shown). All data are expressed as mean6 S.E.M.P , 0.05 was considered statistically significant.
Correlations. As shown in Fig. 4, A–F, Pearson’s correlation wasused to examine the relationship between the Emax value forescitalopram to reduce immobility time in the TST values taken fromFig. 1F, and maximal specific [3H]citalopram binding values takenfrom Fig. 4, A–F. All data are expressed as mean 6 S.E.M.
ResultsMaximal Effect and Potency of Escitalopram to ReduceImmobility Time in the TST Is Age- and SERT-GenotypeDependent
Under control (saline-injected) conditions (Fig. 1A), basalimmobility varied in an age- and genotype-dependent manner[age versus SERT-genotype interaction: F(4,169) 5 7.35, P ,0.0001]. Baseline immobility increased in age SERT1/2mice,but not in SERT1/1 or SERT2/2 mice (Fig. 1A). At P21,differences among the genotypes were not statistically signif-icant. At P28 and P90, SERT2/2mice showed less immobilitythan SERT1/1 mice. At P90, SERT2/2 mice showed lessimmobility than at P21; in contrast, SERT1/2 mice showedmore immobility at P90 than at P21.In SERT1/1mice, escitalopram reduced immobility time in
the TST in all age groups [F(4,299) 5 56.3, P , 0.0001] (Fig.1B). Overall, P21 and P28 mice spent less time immobile thanP90mice following escitalopram [F(2,299)5 7.86,P5 0.0005].In addition, escitalopram dose interacted with age [F(8,299)53.39, P 5 0.001]: at low doses, P21 and P28 mice showed lessimmobility thanmice aged P90. P21 SERT1/1mice spent lesstime immobile than P90 mice following 1.0 and 3.2 mg/kgdoses of escitalopram (P5 0.013 and 0.012, respectively), andP28 SERT1/1 mice spent significantly less time immobilethan P90 mice following 0.32, 1.0, and 3.2 mg/kg doses ofescitalopram (P , 0.0001 and P 5 0.013 and 0.005, respec-tively). There was no significant difference in time spentimmobile among ages following 10 mg/kg escitalopram,although there was a trend for P21 mice to spend more timeimmobile than P90 mice (P 5 0.11). The lowest effective dosefor P28 was 0.32 mg/kg escitalopram (P 5 0.0004), while1.0 mg/kg was the lowest for P21 (P , 0.0001) and P90 mice(P 5 0.008).As for SERT1/1 mice, escitalopram reduced immobility in
all age groups of SERT1/2mice [F(4,250)5 37.4, P, 0.0001]
and its effects interacted with age [F(8,250) 5 2.60, P 50.0095] (Fig. 1C). P21 SERT1/2 mice spent significantly lesstime immobile than P90 SERT1/2 mice following 0.32 mg/kgescitalopram (P5 0.01), but significantly more time immobilefollowing 10 mg/kg escitalopram (P 5 0.006). The lowesteffective dose for P21 and P28 SERT1/2mice was 0.32 mg/kg(P , 0.001 and P 5 0.014, respectively), while thelowest effective dose for P90 SERT1/2 mice was 1.0 mg/kg(P , 0.001).Because previous studies have shown that SSRIs do not
produce antidepressant-like effects in SERT2/2 mice(Holmes et al., 2002), we evaluated only the 10 mg/kg dose ofescitalopram, which is maximally effective in P90 SERT1/1and SERT1/2 mice. As expected, there was no effect of drug[F(1,115)5 0.86, P5 0.42], age [F(2,115)5 0.86, P5 0.42], orinteraction [F(2,115)5 0.86,P5 0.42] (Fig. 1D). Therewere nosignificant differences in immobility among ages and betweenvehicle and 10 mg/kg escitalopram.The ED50 and Emax values for escitalopram to reduce
immobility time in SERT1/1 and SERT1/2 mice are sum-marized in Fig. 1, E and F (SERT2/2 mice were not includedin the analysis because escitalopram did not alter immobilitytime in these mice). The ED50 value of escitalopram to inhibitimmobility varied as a function of age [SERT1/1: F(2,191) 519.45, P , 0.001; SERT1/2: F(2,156) 5 4.18, P 5 0.017];escitalopram more potently inhibited immobility in P21 andP28 mice than in P90 mice for both genotypes [SERT1/1:F(1,133) 5 15.7, P , 0.001, and F(1,137) 5 34.7, P , 0.001;SERT1/2: F(1,104)5 6.24, P5 0.014, and F(1,110)5 5.7, P50.019] (Fig. 1E). Within-age comparisons showed no potencydifference of escitalopram to reduce immobility in SERT1/1and SERT1/2 mice aged P21 and P28. However, at P90 theED50 value was significantly lower in SERT1/2 mice than inSERT1/1 mice [F(1,138) 5 8.35, P 5 0.005].In SERT1/1 and SERT1/2 mice, the Emax values for
escitalopram were smaller in young mice than in adult (P90)mice [F(2,103) 5 7.59, P , 0.001] (Fig. 1F). Genotype had noeffect on the Emax values [F(1,103) 5 0.55, P 5 0.46], nor wasthere an age-by-genotype interaction [F(2,103) 5 7.59, P 50.86]. The Emax values in SERT1/1 and SERT1/2mice agedP21 were smaller than those for genotype-matched P90 mice(P 5 0.047 and 0.008, respectively). No significant differencewas found between P28 and P90Emax values in SERT1/1 andSERT1/2 mice (P 5 0.12 and 0.10, respectively).In summary, for SERT1/1 and SERT1/2 mice the ED50
value for escitalopram to decrease immobility time was lowerin P21 and P28 mice than in P90 mice, suggesting thatescitalopram is more potent in juvenile and adolescent micethan in adult mice. In contrast, the Emax values were smallerin young mice than in P90 mice, suggesting that escitalopramis less efficacious in juvenile and adolescentmice than in adultmice.
SERT Expression Increases with Age in Select 5-HTTerminal Regions and Decreases with Age in Cell BodyRegions
Maximal specific [3H]citalopram binding (Bmax) values andKd values derived from curve fitting were analyzed separatelyfor each brain region with two-factor ANOVA (age and geno-type) (Figs. 2–4; Table 1). As expected, [3H]citalopram bindingin SERT1/2 mice was less than that in SERT1/1 mice and
274 Mitchell et al.
at ASPE
T Journals on A
ugust 28, 2019jpet.aspetjournals.org
Dow
nloaded from

there was no [3H]citalopram binding detected in SERT2/2mice (Figs. 2 and 3).Terminal Regions. The Bmax value for [3H]citalopram
binding increased with age in CA1, CA2, and dentate gyrus[F(2,44)5 9.84,P, 0.001;F(2, 44)5 9.42,P, 0.001;F(2,44)57.17, P 5 0.002]; however, age did not significantly influencethe Bmax value for [3H]citalopram binding in CA3 [F(2,44) 52.80, P5 0.071] (Fig. 4, A–D). As expected, the Bmax values for[3H]citalopram binding in SERT1/2 mice were lower thanthose in SERT1/1 mice [for CA1, CA2, CA3, and dentategyrus, respectively: F(1,44) 5 182, P , 0.001; F(1,44) 5 206,P , 0.001; F(1,44) 5 240, P , 0.001; and F(1,44) 5 132, P ,0.001]. No interactions were found between age and SERTgenotype in the CA1, CA2, and CA3 regions of hippocampus ordentate gyrus [F(2,44) 5 0.24, P 5 0.79; F(2,44) 5 0.66, P 50.52; F(2,44) 5 0.13, P 5 0.88; F(2,44) 5 0.56, P 5 0.58].Among-age comparisons showed theBmax values to be smallerin P21 SERT1/1 and P28 SERT mice than in P90 SERT1/1mice in CA2 and dentate gyrus (P , 0.01 and 0.05 for P21and P28 mice, respectively). In CA1, the Bmax values for[3H]citalopram binding were smaller in P21 SERT1/1 andP21 SERT1/2mice than in the SERT genotype–matched P90group (P , 0.01 and 0.05 for SERT1/1 and SERT1/2 mice,respectively).The effects of age on the Bmax value for [3H]citalopram
binding in either the prefrontal cortex, parietal cortex, orventromedial hypothalamus were not statistically significant[F(2,44) 5 0.68, P 5 0.51; F(2,44) 5 2.858, P 5 0.07; and F(2,44) 5 2.16, P 5 0.13, respectively] (Table 1), except thatthe Bmax value in the parietal cortex of SERT1/1 micewas lower in P21 than in P90 mice. The Bmax values for[3H]citalopram binding in SERT1/1 mice were higher thanthose for SERT1/2mice in the prefrontal cortex, parietal cortex,and ventromedial hypothalamus [F(1,44) 5 154, P , 0.01;F(1,44) 5 64.9, P , 0.01; and F(1,44) 5 90.27, P , 0.01,respectively], and no interaction was found between age andSERT genotype [F(2,44) 5 0.32, P 5 0.73; F(2,44) 5 0.47, P 50.63; and F(2,44) 5 0.61, P 5 0.55, respectively].The Bmax value for [
3H]citalopram binding increased withage in the amygdala [F(2,44) 5 5.17, P , 0.01] (Table 1). Asexpected, lower [3H]citalopram binding was found in amyg-dala of SERT1/2 mice compared with SERT1/1 mice[F(1,44)5 112.7, P, 0.01]. There was no interaction betweenage and SERT genotype [F(2,44)5 0.97, P5 0.39]. Among-agecomparisons showed the Bmax value for [3H]citalopram bind-ing in amygdala to be lower in P28 SERT1/2mice than in P90SERT1/2 mice (P 5 0.046) (Table 1). There was no effect ofage or genotype on theKd values for [
3H]citalopram binding inany terminal region (data not shown, P$ 0.06). Values rangedfrom 0.6 to 1.4 nM [3H]citalopram.Cell Body Regions. The Bmax value for [3H]citalopram
binding significantly decreased with age in the median raphenucleus [F(2,44) 5 10.3, P , 0.01] (Fig. 4F). In the dorsalraphe, a statistically nonsignificant trend showed lower Bmax
values in younger mice than in older mice [F(2,44)5 2.46, P50.098] (Fig. 4E). In both cell body regions, higher Bmax valueswere observed in SERT1/1 mice than in SERT1/2 mice[F(1,44)5 86.7, P, 0.01, and F(1,44)5 78.7, P, 0.01, for themedian and dorsal raphe, respectively]. For both median ordorsal raphe, no interactions between SERT genotype and agewere observed [F(2,44) 5 0.1, P 5 0.91; F(2,44) 5 0.3, P 50.74]. Among-age comparisons in themedian and dorsal raphe
of SERT1/1 mice showed greater Bmax values in P21 than inP90 mice (P , 0.01, P 5 0.04). Similarly, greater Bmax valueswere found in themedian raphe of P21 SERT1/2mice than inP90 SERT1/2 mice (P , 0.01). There were no statisticallysignificant effects of age or SERT genotype on Kd values for[3H]citalopram binding (data not shown, P $ 0.051). Valuesranged from 0.54 to 0.85 nM [3H]citalopram.
Relationship between Antidepressant-like Effects ofEscitalopram in the TST and [3H]Citalopram Binding inHippocampus and Raphe Nuclei
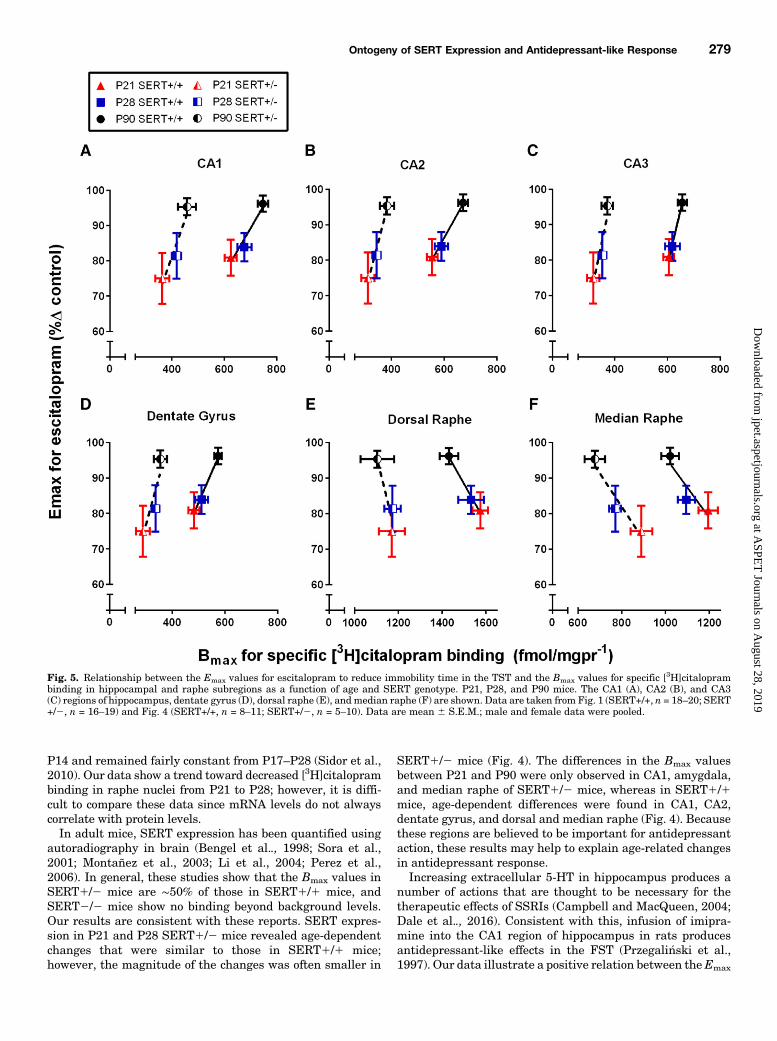
Terminal Regions. Figure 5, A–Dshows a positive relationbetween the Emax values (i.e., antidepressant-like response) forescitalopram and the Bmax values for [
3H]citalopram binding inhippocampal subregions as a function of age and SERT geno-type. In both SERT1/1 and SERT1/2 mice this positiverelation can be adequately described by a straight line in each ofthe hippocampal regions (r $ 0.80–0.99, Pearson’s correlation)(Fig. 5, A–D).Cell Body Regions. Figure 5, E and F shows a negative
relation between theEmax values (i.e.,maximal antidepressant-like response) and the Bmax values for [3H]citalopram bindingin dorsal and median raphe nuclei as a function of age andSERT genotype. In both SERT1/1 and SERT1/2 mice thisnegative relation can be adequately described by a straightline in both the median and dorsal raphe (r $ 0.83–0.99,Pearson’s correlation) (Fig. 5, E and F).
DiscussionTo our knowledge, we are the first to investigate the dose
dependency of escitalopram’s antidepressant-like effect andits relation to SERT expression and affinity in juvenile andadolescent SERT1/1, SERT1/2, and SERT2/2 mice. TheEmax value for escitalopram to reduce immobility time in theTST was lower in juvenile (P21) and adolescent (P28) micethan in adults (P90) (Fig. 1F), while the potency of escitalo-pram was greater in younger mice than in adults (Fig. 1 E).These age-related variations in the Emax values and potencywere apparent in SERT1/1 and SERT1/2 mice. SERTexpression, quantified by [3H]citalopram binding using auto-radiography, increased with age in CA1 and CA2 regions ofhippocampus, dentate gyrus, and amygdala (Fig. 4, A–D;Table 1), and decreased with age in dorsal and median raphenuclei (Fig. 4, E and F). Age-related increases in the Emax
value for escitalopram to reduce immobility time in the TSTwere positively related to age-related increases in the Bmax
value for [3H]citalopram binding (SERT expression) in hippo-campus (Fig. 5, A–D), and negatively related to SERTexpression in raphe nuclei (Fig. 5, E and F). Our data furthersupport the feasibility of using juvenile and adolescent mice tostudy antidepressant activity (Mitchell et al., 2013), andsuggest a link between antidepressant-like activity of escita-lopram and relative expression of its target, SERT.Adolescent (P28–P35)mice respond to SSRIs in the TST and
FST (Bourin et al., 1998; Mason et al., 2009) and juveniles(P21) respond to SSRIs in the TST; however, the magnitude ofthe effect in P21 mice was less than in adult (P90) mice(Mitchell et al., 2013). Our data in SERT1/1 mice areconsistent with these findings (Fig. 1B), as well as withexperiments with rats that have shown escitalopram to reduce
Ontogeny of SERT Expression and Antidepressant-like Response 275
at ASPE
T Journals on A
ugust 28, 2019jpet.aspetjournals.org
Dow
nloaded from

Fig. 2. Specific binding of [3H]citalopram to SERT in hippocampal subregions as a function of age and SERT genotype. Brain sections from P21, P28,and P90 mice were incubated with increasing concentrations of [3H]citalopram. Nonspecific binding was defined by sertraline (20mM). (A)Representative coronal sections showing [3H]citalopram binding at the level of plate 47 (Paxinos and Franklin, 1997) in P21, P28, and P90, SERT+/+,SERT+/2, and SERT2/2 mice. Saturation binding isotherms in CA1 (B), CA2 (C), and CA3 (D) regions of hippocampus, and dentate gyrus (E) of P21,P28, and P90, SERT+/+, SERT+/2, and SERT2/2mice. The Bmax values for each curve are summarized in Fig. 4. There were no significant differencesin Kd values among ages or between SERT+/+ and SERT+/2 mice. There was no significant difference between males and females for any age or SERTgenotype; therefore, data for both genders were pooled. SERT+/+, n = 8–11 (males n = 5 and females n = 3–6 pooled); SERT+/2, n = 5–10 (males n = 3–5and females 2–5 pooled); and SERT2/2, n = 2–4 (males n = 1 to 2 and females n = 1 to 2 pooled) mice per group. Note that because binding in SERT2/2mice was not different from background the sample size was not further increased.
276 Mitchell et al.
at ASPE
T Journals on A
ugust 28, 2019jpet.aspetjournals.org
Dow
nloaded from

immobility in the FST in juveniles (P21) and adolescents (P28)(Reed et al., 2008). Results from these studies parallel clinicalliterature suggesting SSRIs yield limited therapeutic effectsin many children (Tsapakis et al., 2008; Hetrick et al., 2007,2010).Another population that is reported to show a limited
therapeutic response to SSRIs is comprised of individualscarrying low-expressing SERT gene variants (Serretti et al.,2007). SERT1/2 mice provide a model to study these genevariants (Fox et al., 2007; Homberg and Lesch, 2011).Escitalopram reduced immobility in P21, P28, and P90SERT1/1 and SERT1/2 mice (Fig. 1C), a finding consistentwith previous work in adults showing that imipramine
(a SERT and norepinephrine transporter blocker) and fluox-etine (an SSRI) reduced immobility time in the TST equallyin SERT1/1 and SERT1/2 mice (Holmes et al., 2002).SERT2/2 mice did not respond to escitalopram at 10 mg/kg,a dose that is maximally effective in wild-type mice (Fig. 1D),underscoring the selective nature of escitalopram’s actions atSERT.The ED50 value for escitalopram to reduce immobility was
lower in P21 and P28 SERT1/1 and SERT1/2 mice than ingenotype-matched P90 mice (Fig. 1E). This observation isconsistent with the finding that adolescent mice (P35) weremore sensitive to the anti-immobility effects of low SSRI dosesthan adults in the FST (David et al., 2001). Pharmacokinetic
Fig. 3. Specific binding of [3H]citalopram to SERT in dorsal and median raphe nuclei as a function of age and SERT genotype. Brain sections from P21,P28, and P90 mice were incubated with increasing concentrations of [3H]citalopram. Nonspecific binding was defined by sertraline (20mM). (A)Representative coronal sections showing [3H]citalopram binding at the level of plate 64 (Paxinos and Franklin, 1997) in P21, P28, and P90, SERT+/+,SERT+/2, and SERT2/2 mice. Saturation binding isotherms in dorsal raphe (B) and median raphe (C) of P21, P28, and P90, SERT+/+, SERT+/2, andSERT2/2 mice. The Bmax values for each curve are summarized in Fig. 4. There were no significant differences in Kd values among ages or betweenSERT+/+ and SERT+/2 mice. There was no significant difference between males and females for any age or SERT genotype; therefore, data for bothgenders were pooled. SERT+/+, n = 8–11 (males n = 5 and females n = 3–6 pooled); SERT+/2, n = 5–10 (males n = 3–5 and females 2–5 pooled); andSERT2/2, n = 2–4 (males n = 1 to 2 and females n = 1 to 2 pooled) mice per group. Note that because binding in SERT2/2 mice was not different frombackground the sample size was not further increased.
Ontogeny of SERT Expression and Antidepressant-like Response 277
at ASPE
T Journals on A
ugust 28, 2019jpet.aspetjournals.org
Dow
nloaded from

factors could explain differences in antidepressant drugpotency (Bylund and Reed, 2007); however, the eliminationrate of escitalopramhas yet to be determined in youngmice. Incontrast to the effects of age on the potency of escitalopram,the Emax values were lower in P21 SERT1/1 and SERT1/2mice than in SERT genotype–matched P90 mice (Fig. 1F),which is consistent with and expands upon previous studies inyoung SERT1/1 mice (Mitchell et al., 2013).During postnatal development, 5-HT serves as a mitogen to
regulate neuronal growth, dendritic pruning, and additionalaspects of neurodevelopment (Migliarini et al., 2013). Expres-sion of SERT, as a function of age, may vary by brain region toregulate extracellular 5-HT and direct, for example, neuronalgrowth, which is known to vary by brain region (Daval et al.,1987; Krogsrud et al., 2014; Sussman et al., 2016). However,there is a paucity of studies investigating antidepressant
mechanisms in juvenile and adolescent mice. In an effort toaccount for age-related changes in antidepressant response,we quantified SERT expression using autoradiography (Figs.2 and 3). SERT density increased with age in many seroto-nergic terminal regions, including the CA1 and CA2 regions ofhippocampus, dentate gyrus, and amygdala, but did not varywith age in the CA3 region of hippocampus, cortex, orhypothalamus (Fig. 4; Table 1). These data partially divergefrom a previous study, where [3H]citalopram saturationbinding was used in hippocampal membrane preparationsand SERT expression in P21mice was found to be the same asthat in adults (Mitchell et al., 2013). This is likely becauseanalysis of whole hippocampal homogenates does not affordthe anatomic resolution of quantitative autoradiographywhere hippocampal subregions can be readily discerned. Theaffinity values for [3H]citalopram binding in the current study(Kd: 0.54–1.4 nM) were similar to those in the previous study(Kd: 1.3–2.3 nM) (Mitchell et al., 2013).Experiments describing SERT ontogeny in terminal regions
of rat brain report peak SERT expression to occur duringadolescence (∼ P35), with expression decreasing to plateau inadulthood (P70–P100) (Galineau et al., 2004; Slotkin et al.,2008; Daws and Gould, 2011). In contrast, we found either nodifference in SERT expression between adolescent (P28) andadult (P90) mice (CA1, CA3, amygdala, hypothalamus, andprefrontal cortex), or lower SERT expression in adolescentmice compared with adults (CA2, dentate gyrus, and parietalcortex) (Fig. 4; Table 1). The brain regions studied possiblyexplain this disparity; for example, no study in rats hascompared hippocampal expression of SERT across these agegroups. In rats, SERT density in serotonergic cell bodiesdecreased from the early postnatal period to adulthood (Mollet al., 2000; Galineau et al., 2004). Our findings in dorsal andmedian raphe of SERT1/1 mice (Fig. 3; Fig. 4, E and F) areconsistent with those in rats. One study compared mouseSERT mRNA levels in dorsal raphe and found they peaked at
Fig. 4. Bmax values for [3H]citalopram binding toSERT in SERT+/+ and SERT+/2mice aged P21, P28,and P90. The Bmax values were determined from one-site curve fits to data plotted in Fig. 2, B–D and Fig.3, B and C from P21 (red), P28 (blue), and P90 (black)SERT+/+ and SERT+/2mice. Data aremean6S.E.M.pooled from male and female mice. *P , 0.05,**P , 0.01 represent a significant difference fromSERT gene–matched P90, Tukey’s post hoc multiplecomparisons test after repeated-measure two-factorANOVA (age, brain region). SERT+/+, n = 8–11(males n = 5 and females n = 3–6 pooled); and SERT+/2, n = 5–10 (males n = 3–5 and females 2–5 pooled).
TABLE 1Summary of Bmax values for specific [3H]citalopram binding in SERT+/+and SERT+/2 miceData are mean 6 S.E.M. fmol/mg protein. There was no significant differencebetween males and females for any age or SERT genotype; therefore, data for bothgenders were pooled. Tukey’s multiple comparisons test after two-factor ANOVA, n =8–11.
Genotype P21 P28 P90
Prefrontal cortexSERT+/+ 568 6 25 588 6 25 583 6 18SERT+/2 322 6 32 331 6 15 363 6 28
Parietal cortexSERT+/+ 342 6 24* 382 6 21 431 6 24SERT+/2 186 6 37 214 6 24 225 6 22
AmygdalaSERT+/+ 970 6 31 1032 6 29 1058 6 29SERT+/2 669 6 58 678 6 21a 801 6 48
Ventromedial hypothalamusSERT+/+ 999 6 38 933 6 37 938 6 30SERT+/2 696 6 57 677 6 29 597 6 37
aDifference from SERT genotype–matched P90 group, *P , 0.05.
278 Mitchell et al.
at ASPE
T Journals on A
ugust 28, 2019jpet.aspetjournals.org
Dow
nloaded from
P14 and remained fairly constant from P17–P28 (Sidor et al.,2010). Our data show a trend toward decreased [3H]citaloprambinding in raphe nuclei from P21 to P28; however, it is diffi-cult to compare these data since mRNA levels do not alwayscorrelate with protein levels.In adult mice, SERT expression has been quantified using
autoradiography in brain (Bengel et al.., 1998; Sora et al.,2001; Montañez et al., 2003; Li et al., 2004; Perez et al.,2006). In general, these studies show that the Bmax values inSERT1/2 mice are ∼50% of those in SERT1/1 mice, andSERT2/2 mice show no binding beyond background levels.Our results are consistent with these reports. SERT expres-sion in P21 and P28 SERT1/2 mice revealed age-dependentchanges that were similar to those in SERT1/1 mice;however, the magnitude of the changes was often smaller in
SERT1/2 mice (Fig. 4). The differences in the Bmax valuesbetween P21 and P90 were only observed in CA1, amygdala,and median raphe of SERT1/2 mice, whereas in SERT1/1mice, age-dependent differences were found in CA1, CA2,dentate gyrus, and dorsal and median raphe (Fig. 4). Becausethese regions are believed to be important for antidepressantaction, these results may help to explain age-related changesin antidepressant response.Increasing extracellular 5-HT in hippocampus produces a
number of actions that are thought to be necessary for thetherapeutic effects of SSRIs (Campbell and MacQueen, 2004;Dale et al.., 2016). Consistent with this, infusion of imipra-mine into the CA1 region of hippocampus in rats producesantidepressant-like effects in the FST (Przegali�nski et al.,1997). Our data illustrate a positive relation between theEmax
Fig. 5. Relationship between the Emax values for escitalopram to reduce immobility time in the TST and the Bmax values for specific [3H]citaloprambinding in hippocampal and raphe subregions as a function of age and SERT genotype. P21, P28, and P90 mice. The CA1 (A), CA2 (B), and CA3(C) regions of hippocampus, dentate gyrus (D), dorsal raphe (E), andmedian raphe (F) are shown. Data are taken from Fig. 1 (SERT+/+, n = 18–20; SERT+/2, n = 16–19) and Fig. 4 (SERT+/+, n = 8–11; SERT+/2, n = 5–10). Data are mean 6 S.E.M.; male and female data were pooled.
Ontogeny of SERT Expression and Antidepressant-like Response 279
at ASPE
T Journals on A
ugust 28, 2019jpet.aspetjournals.org
Dow
nloaded from

value for escitalopram to produce antidepressant-like effectsand the Bmax value for [3H]citalopram binding in hippocam-pus (Fig. 5, A–D). Expression does not always indicatefunction; however, it is tempting to speculate that a devel-opmental lag in hippocampal SERT expression could limitthe antidepressant-like response to SSRIs.In contrast, there was a negative relation between the Emax
value for antidepressant-like response and the Bmax value for[3H]citalopram binding in raphe nuclei (Fig. 5, E and F).Greater SERT expression in the raphe of juvenile andadolescent mice than in adult mice could produce age-dependent differences in brain region–specific extracellular5-HT concentrations after SSRI administration; however, itis unclear exactly how region-specific differences in 5-HTmay limit the therapeutic effects of SSRIs. Of course, age-dependent shifts in expression and/or function of 5-HTreceptors may also contribute to the limited antidepressant-like response of escitalopram in young mice and should not beoverlooked (Carr and Lucki, 2011). Future studies are neces-sary to better understand the mechanism(s) contributing tothe limited antidepressant-like response of escitalopram inyoung mice. Moreover, because SSRIs differ in their affinityfor SERT as well as other targets (including receptors andtransporters), studies examining a broader range of SSRIsmay provide valuable insight into identifying antidepressantsthat may be more effective for the treatment of depression injuveniles.To our knowledge, this is the first study to evaluate the
ontogeny of SERT expression and antidepressant-like re-sponse in P21, P28, and P90 SERT1/1 and SERT1/2 mice.Its findings help build the foundation needed to discernmechanisms underlying the limited therapeutic benefit ofSSRIs in juveniles and adolescents.
Acknowledgments
The authors gratefully acknowledge the technical assistance ofMelissa Vitela and Myrna Herrera-Rosales.
Authorship Contributions
Participated in research design: Mitchell, Gould, Koek, Daws.Conducted experiments: Mitchell, Gould.Performed data analysis: Mitchell.Wrote or contributed to the writing of the manuscript: Mitchell,
Gould, Koek, Daws.
References
Baganz NL, Horton RE, Calderon AS, Owens WA, Munn JL, Watts LT, Koldzic-Zivanovic N, Jeske NA, Koek W, and Toney GM, et al. (2008) Organic cationtransporter 3: Keeping the brake on extracellular serotonin in serotonin-transporter-deficient mice. Proc Natl Acad Sci USA 105:18976–18981.
Bengel D, Murphy DL, Andrews AM, Wichems CH, Feltner D, Heils A, Mössner R,Westphal H, and Lesch KP (1998) Altered brain serotonin homeostasis and loco-motor insensitivity to 3, 4-methylenedioxymethamphetamine (“Ecstasy”) in sero-tonin transporter-deficient mice. Mol Pharmacol 53:649–655.
Bourin M, Colombel MC, Redrobe JP, Nizard J, Hascoët M, and Baker GB (1998)Evaluation of efficacies of different classes of antidepressants in the forced swim-ming test in mice at different ages. Prog Neuropsychopharmacol Biol Psychiatry22:343–351.
Bujoreanu S, Benhayon D, and Szigethy E (2011) Treatment of depression in childrenand adolescents. Pediatr Ann 40:548–555.
Bylund DB and Reed AL (2007) Childhood and adolescent depression: why do chil-dren and adults respond differently to antidepressant drugs? Neurochem Int 51:246–253.
Campbell S and MacQueen G (2004) The role of the hippocampus in the patho-physiology of major depression. J Psychiatry Neurosci 29:417–426.
Carr GV and Lucki I (2011) The role of serotonin receptor subtypes intreating depression: a review of animal studies. Psychopharmacology (Berl) 213:265–287.
Castagné V, Moser P, Roux S, and Porsolt RD (2011) Rodent models of depression:forced swim and tail suspension behavioral despair tests in rats and mice, in CurrProtoc Neurosci 55:8.10A.1–8.10A.14, John Wiley and Sons, Hoboken.
Cryan JF, Mombereau C, and Vassout A (2005) The tail suspension test as a modelfor assessing antidepressant activity: review of pharmacological and geneticstudies in mice. Neurosci Biobehav Rev 29:571–625.
Dale E, Pehrson AL, Jeyarajah T, Li Y, Leiser SC, Smagin G, Olsen CK, and SanchezC (2016) Effects of serotonin in the hippocampus: how SSRIs and multimodalantidepressants might regulate pyramidal cell function. CNS Spectr 21:143–161.
D’Amato RJ, Largent BL, Snowman AM, and Snyder SH (1987) Selective labeling ofserotonin uptake sites in rat brain by [3H]citalopram contrasted to labeling ofmultiple sites by [3H]imipramine. J Pharmacol Exp Ther 242:364–371.
Daval G, Vergé D, Becerril A, Gozlan H, Spampinato U, and Hamon M (1987)Transient expression of 5-HT1A receptor binding sites in some areas of the ratCNS during postnatal development. Int J Dev Neurosci 5:171–180.
David DJ, Bourin M, Hascoët M, Colombel MC, Baker GB, and Jolliet P (2001)Comparison of antidepressant activity in 4- and 40-week-old male mice in theforced swimming test: involvement of 5-HT1A and 5-HT1B receptors in old mice.Psychopharmacology (Berl) 153:443–449.
Daws LC and Gould GG (2011) Ontogeny and regulation of the serotonin transporter:providing insights into human disorders. Pharmacol Ther 131:61–79.
Fox MA, Andrews AM, Wendland JR, Lesch KP, Holmes A, and Murphy DL (2007) Apharmacological analysis of mice with a targeted disruption of the serotonintransporter. Psychopharmacology (Berl) 195:147–166.
Galineau L, Kodas E, Guilloteau D, Vilar MP, and Chalon S (2004) Ontogeny of thedopamine and serotonin transporters in the rat brain: an autoradiographic study.Neurosci Lett 363:266–271.
Hetrick SE, McKenzie JE, and Merry SN (2010) The use of SSRIs in children andadolescents. Curr Opin Psychiatry 23:53–57.
Hetrick SE, Merry SN, McKenzie J, Sindahl P, and Proctor M (2007) Selectiveserotonin reuptake inhibitors (SSRIs) depressive disorders in children and ado-lescents. Cochrane Database Syst Rev, Issue 3: CD004851. 1–91.
Holmes A, Yang RJ, Murphy DL, and Crawley JN (2002) Evaluation ofantidepressant-related behavioral responses in mice lacking the serotonin trans-porter. Neuropsychopharmacology 27:914–923.
Homberg JR and Lesch KP (2011) Looking on the bright side of serotonin transportergene variation. Biol Psychiatry 69:513–519.
Horton RE, Apple DM, Owens WA, Baganz NL, Cano S, Mitchell NC, Vitela M, GouldGG, Koek W, and Daws LC (2013) Decynium-22 enhances SSRI-inducedantidepressant-like effects in mice: uncovering novel targets to treat depression.J Neurosci 33:10534–10543.
Kessler RC, Avenevoli S, and Ries Merikangas K (2001) Mood disorders in childrenand adolescents: an epidemiologic perspective. Biol Psychiatry 49:1002–1014.
Koek W, Mercer SL, Coop A, and France CP (2009) Behavioral effects of g-hydroxy-butyrate, its precursor g-butyrolactone, and GABAB receptor agonists: timecourse and differential antagonism by the GABAB receptor antagonist3-aminopropyl(diethoxymethyl)phosphinic acid (CGP35348). J Pharmacol ExpTher 330:876–883.
Krogsrud SK, Tamnes CK, Fjell AM, Amlien I, Grydeland H, Sulutvedt U, Due-Tønnessen P, Bjørnerud A, Sølsnes AE, and Håberg AK, et al. (2014) Developmentof hippocampal subfield volumes from 4 to 22 years. Hum Brain Mapp 35:5646–5657.
Li Q, Ma L, Innis RB, Seneca N, Ichise M, Huang H, Laruelle M, and Murphy DL(2004) Pharmacological and genetic characterization of two selective serotonin trans-porter ligands: 2-[2-(dimethylaminomethylphenylthio)]-5-fluoromethylphenylamine(AFM) and 3-amino-4-[2-(dimethylaminomethyl-phenylthio)]benzonitrile (DASB).J Pharmacol Exp Ther 308:481–486.
Loizou LA and Salt P (1970) Regional changes in monoamines of the rat brain duringpostnatal development. Brain Res 20:467–470.
Mason SS, Baker KB, Davis KW, Pogorelov VM, Malbari MM, Ritter R, Wray SP,Gerhardt B, Lanthorn TH, and Savelieva KV (2009) Differential sensitivity to SSRIand tricyclic antidepressants in juvenile and adult mice of three strains. Eur JPharmacol 602:306–315.
Migliarini S, Pacini G, Pelosi B, Lunardi G, and Pasqualetti M (2013) Lack of brainserotonin affects postnatal development and serotonergic neuronal circuitry for-mation. Mol Psychiatry 18:1106–1118.
Miranda-Contreras L, Mendoza-Briceño RV, and Palacios-Prü EL (1998) Levels ofmonoamine and amino acid neurotransmitters in the developing male mouse hy-pothalamus and in histotypic hypothalamic cultures. Int J Dev Neurosci 16:403–412.
Mitchell NC, Gould GG, Smolik CM, Koek W, and Daws LC (2013) Antidepressant-like drug effects in juvenile and adolescent mice in the tail suspension test: Re-lationship with hippocampal serotonin and norepinephrine transporter expressionand function. Front Pharmacol 4:131.
Mitchell NC, Koek W, and Daws LC (2015) Antidepressant-like effects and basalimmobility depend on age and serotonin transporter genotype. Genes Brain Behav14:543–549.
Moll GH, Mehnert C, Wicker M, Bock N, Rothenberger A, Rüther E, and Huether G(2000) Age-associated changes in the densities of presynaptic monoamine trans-porters in different regions of the rat brain from early juvenile life to late adult-hood. Brain Res Dev Brain Res 119:251–257.
Montañez S, Owens WA, Gould GG, Murphy DL, and Daws LC (2003) Exaggeratedeffect of fluvoxamine in heterozygote serotonin transporter knockout mice. JNeurochem 86:210–219.
O’Leary OF, Bechtholt AJ, Crowley JJ, Hill TE, Page ME, and Lucki I (2007) De-pletion of serotonin and catecholamines block the acute behavioral response todifferent classes of antidepressant drugs in the mouse tail suspension test. Psy-chopharmacology (Berl) 192:357–371.
Paxinos G and Franklin KBJ (1997) The Mouse Brain in Stereotaxic Coordinates,Academic Press, San Diego, CA.
Perez XA, Bianco LE, and Andrews AM (2006) Filtration disrupts synaptosomesduring radiochemical analysis of serotonin uptake: comparison with chro-noamperometry in SERT knockout mice. J Neurosci Methods 154:245–255.
280 Mitchell et al.
at ASPE
T Journals on A
ugust 28, 2019jpet.aspetjournals.org
Dow
nloaded from

Petersen IT, Bates JE, Goodnight JA, Dodge KA, Lansford JE, Pettit GS, LatendresseSJ, and Dick DM (2012) Interaction between serotonin transporter polymorphism(5-HTTLPR) and stressful life events in adolescents’ trajectories of anxious/depressedsymptoms. Dev Psychol 48:1463–1475.
Przegali�nski E, Tatarczy�nska E, Dere�n-Wesołek A, and Chojnacka-Wojcik E (1997)Antidepressant-like effects of a partial agonist at strychnine-insensitive glycinereceptors and a competitive NMDA receptor antagonist. Neuropharmacology 36:31–37.
Reed AL, Happe HK, Petty F, and Bylund DB (2008) Juvenile rats in the forced-swimtest model the human response to antidepressant treatment for pediatric de-pression. Psychopharmacology (Berl) 197:433–441.
Ripoll N, David DJ, Dailly E, Hascoët M, and Bourin M (2003) Antidepressant-like effectsin various mice strains in the tail suspension test. Behav Brain Res 143:193–200.
Serretti A, Kato M, De Ronchi D, and Kinoshita T (2007) Meta-analysis of serotonintransporter gene promoter polymorphism (5-HTTLPR) association with selective sero-tonin reuptake inhibitor efficacy in depressed patients. Mol Psychiatry 12:247–257.
Sidor MM, Amath A, MacQueen G, and Foster JA (2010) A developmental charac-terization of mesolimbocortical serotonergic gene expression changes followingearly immune challenge. Neuroscience 171:734–746.
Slotkin TA, Ryde IT, Levin ED, and Seidler FJ (2008) Developmental neurotoxicity oflow dose diazinon exposure of neonatal rats: effects on serotonin systems in ado-lescence and adulthood. Brain Res Bull 75:640–647.
Sora I, Hall FS, Andrews AM, Itokawa M, Li XF, Wei HB, Wichems C, Lesch KP,Murphy DL, and Uhl GR (2001) Molecular mechanisms of cocaine reward: com-bined dopamine and serotonin transporter knockouts eliminate cocaine placepreference. Proc Natl Acad Sci USA 98:5300–5305.
Spear LP (2000) The adolescent brain and age-related behavioral manifestations.Neurosci Biobehav Rev 24:417–463.
Steru L, Chermat R, Thierry B, and Simon P (1985) The tail suspension test: a newmethod for screening antidepressants in mice. Psychopharmacology (Berl) 85:367–370.
Sussman D, Leung RC, Chakravarty MM, Lerch JP, and Taylor MJ (2016) De-veloping human brain: age-related changes in cortical, subcortical, and cerebellaranatomy. Brain Behav 6:e00457.
Tsapakis EM, Soldani F, Tondo L, and Baldessarini RJ (2008) Efficacy of antide-pressants in juvenile depression: meta-analysis. Br J Psychiatry 193:10–17.
Wallace JA and Lauder JM (1983) Development of the serotonergic system in the ratembryo: an immunocytochemical study. Brain Res Bull 10:459–479.
Address correspondence to: Dr. Lynette C. Daws, Department of Physiology,University of Texas Health Science Center, 7703 Floyd Curl Drive, San Antonio,TX. E-mail: [email protected]
Ontogeny of SERT Expression and Antidepressant-like Response 281
at ASPE
T Journals on A
ugust 28, 2019jpet.aspetjournals.org
Dow
nloaded from


















