Human Papilloma Virus and Epstein-Barr Virus in Sinonasal ...

[CAN@KRRESEARCH @,15g6-1@4,Aug@M19681
SUMMARY
Different isolates of bovine papilloma virus (BPV) originating from different cases induced well-differentiated subcutanoons fibrosarcomas in 67 to 100% of inoculated hamsters afteran induction period ranging from 25 to 56 weeks. Age at timeof inoculation had little if any effect upon the incidence orsubsequent development of the BPV-induced tumors. Disseminated tumor nodules occurred in about 10% of the animals. Several tumors were transplanted to other hamsters. Areview of reports on other viruses oncogenic for the hamsterrevealed that the incidence of tumors decreased when olderanimals were inoculated.
INTRODUCTION
Bovine papilloma virus (BPV) can induce tumors in bothepithelial and connective tissues of the bovine as well as connective tissue tumors in other species of animals. The lesioninduced by BPV in the bovine skin may properly be called afibropapilloma since it is a fibroma underlying a benign neoplastic epithelium. This feature was first described by Creechin 1929 (10) and has been observed by others (4, 9). Bovinepapulioma virus can induce fibroblastic tumors independent ofepithelial hyperplasia in the urinary bladder (28) and genitalmucosa of cattle (26), as well as in the dermis of the horse(27) and C3H/eB mice (6). The hamster is susceptible to theoncogenic action of BPV and develops connective tissue tumorsin the dermis (6, 8, 25, 36) and brain (8, 25). Various factorsinfluencing tumor induction by BPV in the hamster and othercharacteristics of the tumors were examined in this study.These factors included the use of different BPV isolates, dose,different sites of subcutaneous inoculation, and variations ofthe age of the hamster at time of inoculation. In addition,several of the BPV-induced tumors were transplanted to otherhamsters.
MATERIALS AND METHODS
Cutaneous bovine fibropapillomas from several differentanimals were used. Isolate 247 had been carried in 3 successivebovine animal passages. Tissue from Isolate 247 was storedfor several years at 4°C in 50% glycerin and saline prior to its
1 Published with the approval of the Director of the Wisconsin
Agricultural Experiment Station. Supported in part with the aidof RG-C4627 and USPHS Training Grant 817, NIH.
Received December 4, 1967; accepted April 15, 1968.
use in this experiment. Tissue from field Isolates 305, 306, and310 had been stored at —70°C.The fibropapilloma tissue wasground in a mortar and pestle with sterile sand and saline tomake a 10% tissue suspension. This suspension was sonifiedat a setting of 3, with 3 amp for 2 minutes in an ice water bath(Model LS-75, Branson Inst. Co., Danbury, Conn.). The suepension was centrifuged at 3500 x g for 10 minutes and thesupernatant fluid used as the inoculum. A suspension of normalbovine skin was prepared in the same manner and injectedinto control hamsters.
Hamsters were obtained from Con Olson Co. Inc., Madison,Wisconsin. One-tenth ml of supernatant fluid of the appropriateinoculum was injected subcutaneously over the dorsal midlinein the thoracic and/or lumbar area (Table 1) . Isolates 305,306, and 310 were inoculated into the pinna of the ear as well.The hamsters were kept in individual cages to avoid canniballam and observed at 2-week intervals for the duration of theirexperimental life.
The induction period (interval from inoculation until the firstappearance of tumors) was determined by palpation of therestrained animal. Tumors were measured in two diametersand size was expressed as the average diameter. Several of thetumors were removed surgically, and small pieces of tumorwere transplanted subcutaneously into young (less than 14day old) as well as one-year-old normal hamsters.
Representative tissues were selected at necropsy for histologic examination. These were fixed in formalin, processed byconventional paraffin embedding, and sections were stained bythe hematoxylin and eosin, van Gieson, Giemsa, and Kingsleytechnics.
RESULTS
Death from non-neoplastic disease occurred in less than 10%of the hamsters before tumors developed, and these animalswere not considered in the results of the respective experiments.
Tumors developed in 86% (37/43) of the hamsters inoculatedat ages ranging from 2 to 52 weeks. The induction period averaged 39 weeks and ranged from 25 to 50 weeks (Table 1, Experiment1).
All 13 animals inoculated with dilutions of 10@ to 108from Isolate 247 developed tumors. Only 1 of 4 hamsters developed a tumor from the 10—4dilution, while the 10@ dilution failed to induce tumors in 5 hamsters. The induction periodaveraged 43 weeks and ranged from 36 to 56 weeks (Table 1,Experiment 2) . There was no appreciable difference in the
induction period with respect to the dilution of the viralsuspension.
1596 CANCER RESEARCH VOL.28
Oncogenic Action of Bovine Papilloma Virus in Hamsters'
Martin 0. RobI and Carl Olson
Department of Veterinary Science, University of Wisconsin, Madison, WiscOnsin 58706
Research. on February 14, 2019. © 1968 American Association for Cancercancerres.aacrjournals.org Downloaded from
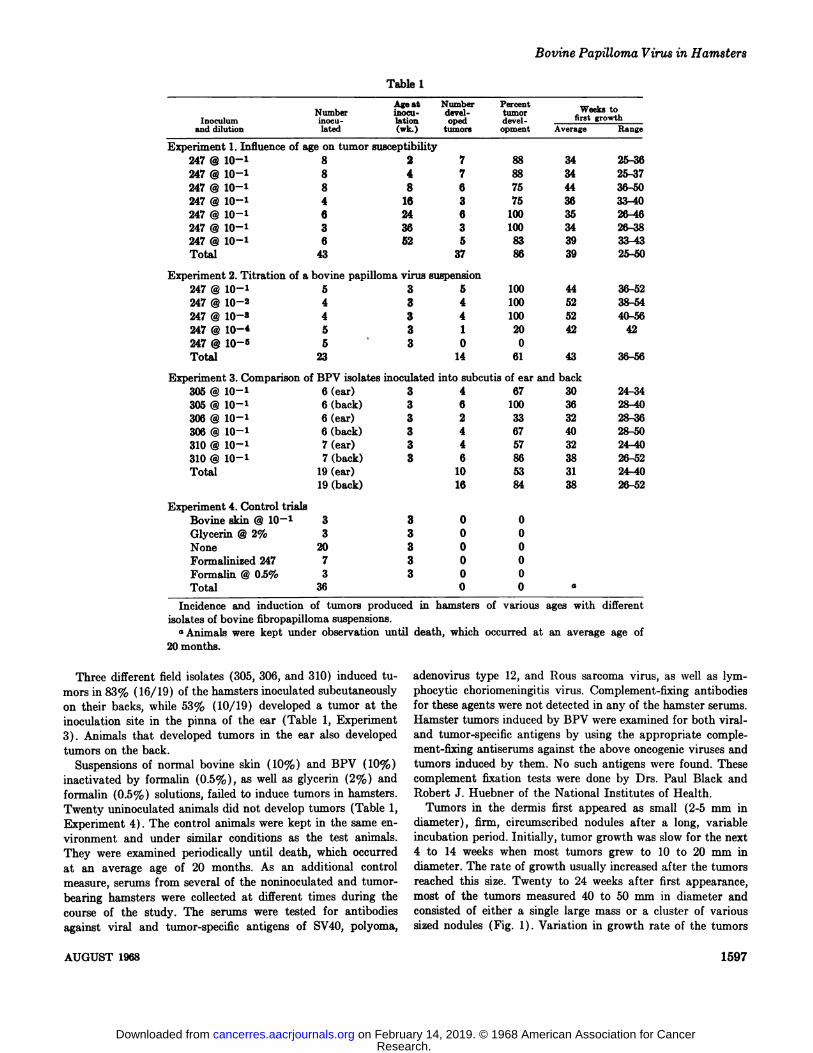
Age at Number PercentNumber rnoc@i- devel- tumor WO@k8to
Inoculum inocu- lation oped devel- fI@ gToWthand dilution lated (wk.) tumors opment AverageRangeExperiment
1. Influence of age on tumorsusceptibility247@10—'8 2 7 88 3425-36247@10—1
8 4 7 88 3425-37247@10—'8 8 6 75 4436-50247@10—1
4 16 3 75 3633—40247@10—'6 24 6 100 3526-46247@10—'3 36 3 100 3426—38247@10—16 52 5 83 3933-43Total
43 37 86 3925—SOExperiment
2. Titration of a bovine papilloma virussuspension247@1O—15 3 5 100 4436—52247@10—24 3 4 100 5238—54247@10—34 3 4 100 5240-56247@1O—@5 3 1 20 4242247@10—5
5@ 3 00Total23 14 61 4336-56Experiment
3. Comparison of BPV isolates inoculated into subcutis of ear andback305@1O—16(ear) 3 4 67 3024—34305
@ 10—1 6 (back) 3 6 100 3628-40306@10—16(ear) 3 2 33 3228—36306
@ 10—1 6 (back) 3 4 67 4028-50310@1O—'7(ear) 3 4 57 3224—40310@1O—'7(back) 3 6 86 3826-52Total
19(ear) 10 53 3124-4019(back) 16 84 3826-52Experiment
4. ControltrialsBovinesldn@10'3 3 00Glycerin@2%3 3 00None
20 3 00Formalinized247 7 3 00Formalin@OS%
3 3 00Total36 0 0 a
of various ages with different
Bovine Papilloma Virus in Hamsters
Table 1
Incidence and induction of tumors produced in hamstersisolates of bovine fibropapilloma suspensions.
a Animals were kept under observation until death, which
20 months.occurred at an average age of
Three different field isolates (305, 306, and 310) induced tumors in 83% (16/19) of the hamsters inoculated subcutaneouslyon their backs, while 53% (10/19) developed a tumor at theinoculation site in the pinna of the ear (Table 1, Experiment3) . Animals that developed tumors in the ear also developedtumors on the back.
Suspensions of normal bovine skin (10%) and BPV (10%)inactivated by formalin (0.5%), as well as glycerin (2%) andformalin (0.5%) solutions, failed to induce tumors in hamsters.Twenty uninoculated animals did not develop tumors (Table 1,Experiment 4) . The control animals were kept in the same environment and under similar conditions as the test animals.They were examined periodically until death, which occurredat an average age of 20 months. As an additional controlmeasure, serums from several of the noninoculated and tumorbearing hamsters were collected at different times during thecourse of the study. The serums were tested for antibodiesagainst viral and tumor-specific antigens of SV4O, polyoma,
adenovirus type 12, and Rous sarcoma virus, as well as lymphocytic choriomeningitis virus. Complement-fixing antibodiesfor these agents were not detected in any of the hamster serums.Hamster tumors induced by BPV were examined for both viraland tumor-specific antigens by using the appropriate complement-fixing antiserums against the above oncogenic viruses andtumors induced by them. No such antigens were found. Thesecomplement fixation tests were done by Drs. Paul Black andRobert J. Huebner of the National Institutes of Health.
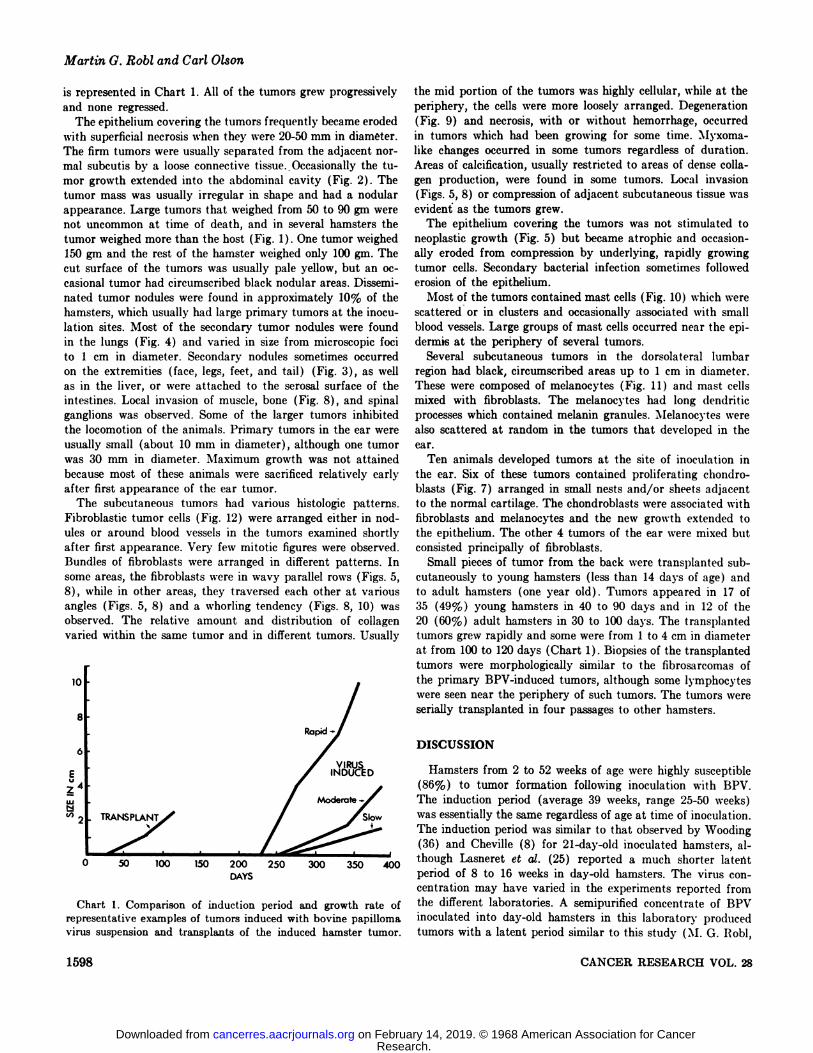
Tumors in the dermis first appeared as small (2-5 mm indiameter) , firm, circumscribed nodules after a long, variableincubation period. Initially, tumor growth was slow for the next4 to 14 weeks when most tumors grew to 10 to 20 mm indiameter. The rate of growth usually increased after the tumorsreached this size. Twenty to 24 weeks after first appearance,most of the tumors measured 40 to 50 mm in diameter andconsisted of either a single large mass or a cluster of varioussized nodules (Fig. 1) . Variation in growth rate of the tumors
AUGUST 1968 1597
Research. on February 14, 2019. © 1968 American Association for Cancercancerres.aacrjournals.org Downloaded from
Martin G. Robi and Carl Olson
is represented in Chart 1. All of the tumors grew progressivelyand none regressed.
The epithelium covering the tumors frequently became erodedwith superficial necrosis when they were 20-50 mm in diameter.The firm tumors were usually separated from the adjacent normal subcutis by a loose connective tissue. .Occasionally the tumor growth extended into the abdominal cavity (Fig. 2) . Thetumor mass was usually irregular in shape and had a nodularappearance. Large tumors that weighed from 50 to 90 gm werenot uncommon at time of death, and in several hamsters thetumor weighed more than the host (Fig. 1) . One tumor weighed150 gm and the rest of the hamster weighed only 100 gm. Thecut surface of the tumors was usually pale yellow, but an cccasional tumor had circumscribed black nodular areas. Disseminated tumor nodules were found in approximately 10% of thehamsters, which usually had large primary tumors at the inoculation sites. Most of the secondary tumor nodules were foundin the lungs (Fig. 4) and varied in size from microscopic focito 1 cm in diameter. Secondary nodules sometimes occurredon the extremities (face, legs, feet, and tail) (Fig. 3), as wellas in the liver, or were attached to the serosal surface of theintestines. Local invasion of muscle, bone (Fig. 8), and spinalganglions was observed. Some of the larger tumors inhibitedthe locomotion of the animals. Primary tumors in the ear wereusually small (about 10 mm in diameter) , although one tumorwas 30 mm in diameter. Maximum growth was not attainedbecause most of these animals were sacrificed relatively earlyafter first appearance of the ear tumor.
The subcutaneous tumors had various histologic patterns.Fibroblastic tumor cells (Fig. 12) were arranged either in nodules or around blood vessels in the tumors examined shortlyafter first appearance. Very few mitotic figures were observed.Bundles of fibroblasts were arranged in different patterns. Insome areas, the fibroblasts were in wavy parallel rows (Figs. 5,8) , while in other areas, they traversed each other at variousangles (Figs. 5, 8) and a whorling tendency (Figs. 8, 10) wasobserved. The relative amount and distribution of collagenvaried within the same tumor and in different tumors. Usually
Chart 1. Comparison of induction period and growth rate ofrepresentative examples of tumors induced with bovine papillomavirus suspension and transplants of the induced hamster tumor.
the mid portion of the tumors was highly cellular, while at theperiphery, the cells were more loosely arranged. Degeneration(Fig. 9) and necrosis, with or without hemorrhage, occurredin tumors which had been growing for some time. Myxomalike changes occurred in some tumors regardless of duration.Areas of calcification, usually restricted to areas of dense collagen production, were found in some tumors. Local invasion(Figs. 5, 8) or compression of adjacent subcutaneous tissue wasevident as the tumors grew.
The epithelium covering the tumors was not stimulated toneoplastic growth (Fig. 5) but became atrophic and occasionally eroded from compression by underlying, rapidly growingtumor cells. Secondary bacterial infection sometimes followederosion of the epithelium.
Most of the tumors contained mast cells (Fig. 10) which werescattered or in clusters and occasionally associated with smallblood vessels. Large groups of mast cells occurred near the epidermis at the periphery of several tumors.
Several subcutaneous tumors in the dorsolateral lumbarregion had black, circumscribed areas up to 1 cm in diameter.These were composed of melanocytes (Fig. 11) and mast cellsmixed with fibroblasts. The melanocytes had long dendriticprocesses which contained melanin granules. 1'@1elanocytes werealso scattered at random in the tumors that developed in theear.
Ten animals developed tumors at the site of inoculation inthe ear. Six of these tumors contained proliferating chondroblasts (Fig. 7) arranged in small nests and/or sheets adjacentto the normal cartilage. The chondroblasts were associated withfibroblasts and melanocytes and the new growth extended tothe epithelium. The other 4 tumors of the ear were mixed butconsisted principally of fibroblasts.
Small pieces of tumor from the back were transplanted subcutaneously to young hamsters (less than 14 days of age) andto adult hamsters (one year old) . Tumors appeared in 17 of35 (49%) young hamsters in 40 to 90 days and in 12 of the20 (60%) adult hamsters in 30 to 100 days. The transplantedtumors grew rapidly and some were from 1 to 4 cm in diameterat from 100 to 120 days (Chart 1). Biopsies of the transplantedtumors were morphologically similar to the fibrosarcomas ofthe primary BPV-induced tumors, although some lymphocyteswere seen near the periphery of such tumors. The tumors wereserially transplanted in four passages to other hamsters.
DISCUSSION
Hamsters from 2 to 52 weeks of age were highly susceptible(86%) to tumor formation following inoculation with BPV.The induction period (average 39 weeks, range 25-50 weeks)was essentially the same regardless of age at time of inoculation.The induction period was similar to that observed by Wooding(36) and Cheville (8) for 21-day-old inoculated hamsters, although Lasneret et a!. (25) reported a much shorter latentperiod of 8 to 16 weeks in day-old hamsters. The virus concentration may have varied in the experiments reported fromthe different laboratories. A semipurified concentrate of BPVinoculated into day-old hamsters in this laboratory producedtumors with a latent period similar to this study (M. G. Ilobl,
10
Ra@
0 100 150 200 250 300 350 400DAYS
1598 CANCER RESEARCH VOL.28
Research. on February 14, 2019. © 1968 American Association for Cancercancerres.aacrjournals.org Downloaded from

-Bovine Papilloma Virus in Hamsters
M. Tajima, and C. Olson, unpublished observations). The sueceptibility of hamsters from different sources may also haveinfluenced the results in the different laboratories.
Tumor development occurred only when suspensions of bovine papilloma virus were inoculated. The different isolates ofBPV induced a similar incidence of tumors of similar histologictypes. Normal bovine skin, formalin-inactivated bovine papilloma virus suspensions, formalin, and glycerin failed to inducetumors in hamsters. Antibodies to other viruses oncogenic forthe hamster (SV4O, polyoma, adenoviruses, and Rous sarcomavirus) were not detected by the complement fixation test.
Secondary tumor nodules were probably derived from metastatic tumor cells ; however, the possibility of their being causedby a viremia cannot be ruled out. Histologically, the BPVinduced tumors did not appear anaplastic, and mitotic figureswere rather infrequent ; however, the fact that some of themdid metastasize and were successfully transplanted to otherhamsters is evidence that they should be considered malignant.
The tumors probably were derived from young fibroblastsor undifferentiated mesenchymal cells which are present bothin young and adult connective tissues ( 19, 35) . In addition tothe fibromas and fibrosarcomas, chondromas, and possibly melanofibromas and mast cell neoplasms were observed in thehamster. The reasons for such various histologic reactions canbe only a matter of conjecture. The type of cellular responsewas influenced to some extent by the site of inoculation, sincethe chondromas developed only in the ear and the melanofibromas developed in areas with an abundance of melanocytes.Explanation for the large numbers of mast cells, also observedby Wooding (36) and Allison (2), in BPV-induced tumors isnot known. They could be transformed fibroblasts or they couldbe a manifestation of an abortive attempt by the hamster toreject the tumor (31, 32).
The development of tumors 30 to 90 days after transplantation was much shorter than the induction period for initialtumor development after the bovine papilloma virus suspensionswere inoculated. This is interpreted as growth of the graftedcells, thus confirming the observations of Lasneret et al. (25),who noted growth 6 months after grafting small pieces of tumor, and a statement by Allison (2) that grafts were successful.
Bovine papilloma virus particles and associated viral antigenshave been demonstrated only in the superficial epithelial cellsof the cutaneous neoplasm in the bovine (7, 9, 15, 33, 34) . Ourattempts to demonstrate virus in the hamster tumors by immunofluorescence, by electron microscopy of thin sections andsuspensions, as well as by inoculation of cell-free preparationsinto other hamsters have been negative (M. G. Robi, M.Tajima, and C. Olson, unpublished observations) . The relationship of the BPV to the fibromatous response in the variousspecies appears to be a manifestation of a highly integratedvirus-cell complex.
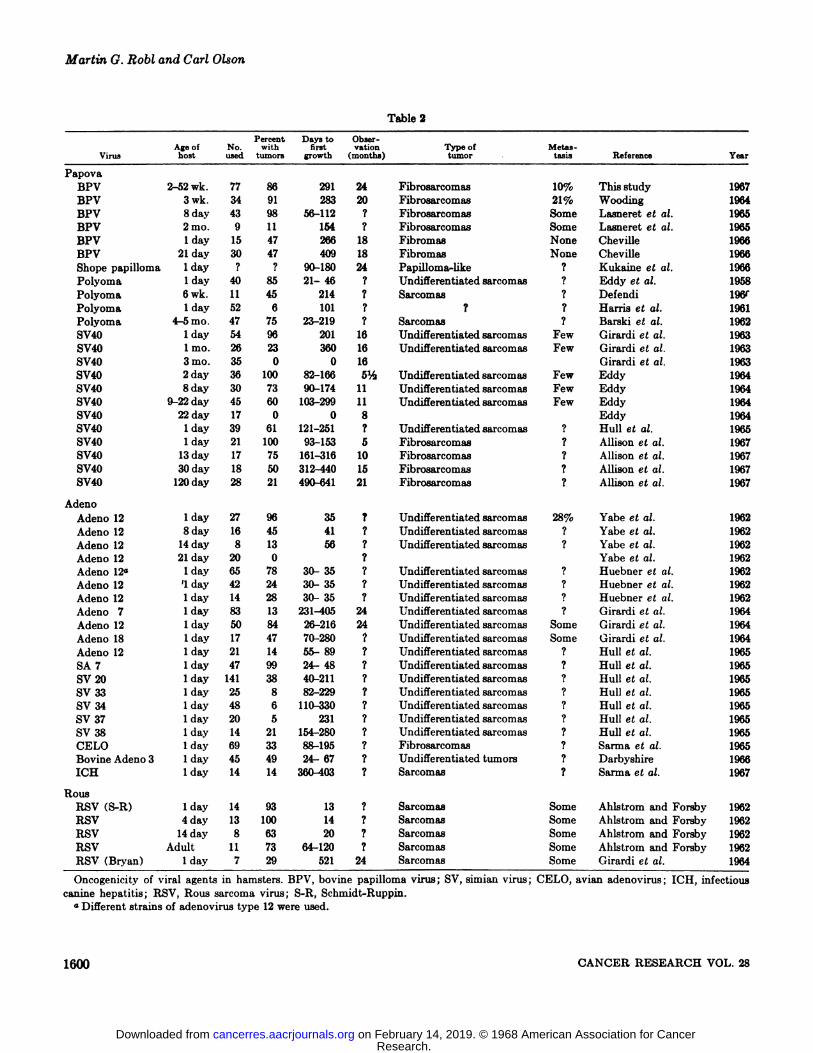
Data accumulated on viruses oncogenic for the hamster arecompiled in Table 2 and provide comparative information. Theinduction period varies with the same virus, with differentviruses, with the age of the hamster, and with the individualresearcher. Demonstration of viral carcinogenesis is usually believed to require that a large quantity of virus be given to ayoung, immunologically incompetent animal (23). The percent
age of tumor development usually decreases as the age of thehost at time of @inoculation increases. The induction period isusually lengthened and the percentage of tumor developmentdecreased when the quantity of virus was decreased.
The influence of age upon subsequent development of tumorsinduced by the polyoma virus in the hamster is difficult toevaluate. Eddy et at. ( 14) reported that the neonate hamsterwas highly susceptible (85%), and tumors developed in a relatively short time (21—46)days. Harris et at. (20) found a lowincidence of tumor development in neonate hamsters (6%) witha latent period of about 101 days. Barski et al. (5) reportedthat 75% of 4- to 5-month-old hamsters developed tumors withthe induction period ranging from 23 to 219 days.
Adenoviruses appear to be oncogenic only in young hamsters.The oncogenic effect and induction period for tumor development has varied with the different types of adenovirus, as wellas with different isolates of adenovirus type 12. For example,Girardi et al. (17) reported tumors in only 13% of neonatehamsters with an induction period ranging from 231 to 405days, whereas Yabe et al. (37) and Huebner et al. (21) obtamed an incidence of 96 and 78% respectively with an induction period of less than 35 days.
Neonate hamsters developed tumors from SV4O after a relatively long induction period. Girardi et at. (17) observed aninduction period of 201 days, Eddy (13) observed an inductionperiod ranging from 82 to 166 days, and Allison et al. (3) observed an induction period of 139 days. Inoculation of olderhamsters produced fewer tumors and the induction period wasprolonged. For example, Allison et al. (3) found tumors in only21% of hamsters inoculated at the age of 120 days, and theinduction period averaged 575 days.
Ahlstrom and Forsby (1) found that young hamsters werequite susceptible (93-100%) to Rous sarcoma virus (RSV,Schmidt-Ruppin strain) and developed tumors in 14 to 20 days.Adult hamsters were also quite susceptible (63%) , but the induction period was from 64 to 120 days. Girardi et al. (16)noted a somewhat lower incidence of tumors (29% ) in dayold hamsters inoculated with the Bryan strain of RSV.
BPV, polyoma, and SV4O of the papova virus group areoncogenic in the Syrian hamster (Mesocricatus auratus) (Table2). Possibly the Shope rabbit papilloma virus, also a memberof the papova virus group, can be included as oncogenic in theSyrian hamster, although the published abstract (24) gives onlybrief details. In the adenovirus group, at least 8 human, 6simian, 1 bovine, 1 avian, and 1 canine type are oncogenic(Table 2) . Two strains of the Rous sarcoma virus (RSV) arealso capable of inducing tumors in the hamster (Table 2). Onlytwo (BPV and RSV) and possibly a third (polyoma) of theviruses oncogenic in hamsters are known to induce tumors intheir species of origin. The Shope rabbit papilloma virus induces tumors in its natural host, but the tumor-like protuberances described by Kukaine and Nagayeva (24) in the hamsterhave not been reported by other investigators. With the cxception of several adenoviruses that produce an acute infectiousdisease in their natural host, almost all of the other virusesoncogenic for the hamster produce only a latent or inapparentinfection in their natural host.
The quantity of BPV required to induce a tumor in the ham
AUGUST 1968 1599
Research. on February 14, 2019. © 1968 American Association for Cancercancerres.aacrjournals.org Downloaded from
Age ofVirus hostNo. usedPercent
withtumorsDays
tofirst
growthObservation
(months)Typeof
tumor .Metes teals Reference@ Year
Martin G. Robl and Carl Olson
Table 2
PapovaBPV2-52wk.778629124Fibrosarcomas10%Thisstudy1967BPV3
wk.349128320Fibrosarcomas21%Wooding1964BPV8day439856-112?FibrosarcomasSomeLasneret etat.1965BPV2mo.911154?FibrosarcomasSomeLasneret etal.1965BPV1
day154726618FibromasNoneCheville1966BPV21day304740918FibromasNoneCheville1966Shope
papilloma1 day??90-18024Papilloma-like?Kukaine etal.1966Polyoma1day408521- 46?Undifferentiated sarcomas?Eddy etal.1958Polyoma6
wk.1145214?Sarcomas?Defendi19&Polyoma1day526101??Harris etal.1961Polyoma4—5mo.477523—219?Sarcomas?Barski etat.1962SV4O1day549620116Undifferentiated sarcomasFewGirardi etat.1963SV4O1mo.262336016Undifferentiated sarcomasFewGirardi etal.1963SV403mo.350016Girardietal.1963SV4O2
day3610082-1665@UndifferentiatedsarcomasFewEddy1964SV4O8day307390-17411UndifferentiatedsarcomasFewEddy1964SV4O9-22day4560103-29911UndifferentiatedsarcomasFewEddy1964SV4O22
day17008Eddy1964SV4O1day3961121—251?Undifferentiated sarcomas?Hull etat.1965SV4O1day2110093—1535Fibrosarcomas?Allison etal.1967SV4O13day1775161—31610Fibrosarcomas?Allison etal.1967SV4O30day1850312-44015Fibrosarcomas?Allison etal.1967SV4O120day2821490-64121Fibrosarcomas?Allison etal.1967AdenoAdeno
121 day279635?Undifferentiated sarcomas28%Yabe etal.1962Adeno128 day164541?Undifferentiated sarcomas?Yabe etal.1962Adeno1214 day81356?Undifferentiated sarcomas?Yabe etat.1962Adeno1221 day200?Yabe etat.1962Adeno12@'1 day657830- 35?Undifferentiated sarcomas?Huebner etat.1962Adeno12‘1 day422430- 35?Undifferentiated sarcomas?Huebner etat.1962Adeno121 day142830- 35?Undifferentiated sarcomas?Huebner etat.1962Adeno71 day8313231—40524Undifferentiated sarcomas?Girardi etat.1964Adeno
121 day508426-21624Undifferentiated sarcomasSomeGirardi etat.1964Adeno181 day174770—280?Undifferentiated sarcomasSomeGirardi etat.1964Adeno121 day211455- 89?Undifferentiated sarcomas?Hull etat.1965SA
71 day479924— 48?Undifferentiated sarcomas?Hull etat.1965SV201 day1413840-211?Undifferentiated sarcomas?Hull etat.1965SV331 day25882—229?Undifferentiated sarcomas?Hull etat.1965SV341 day486110-330?Undifferentiated sarcomas?Hull etat.1965SV371 day205231?Undifferentiated sarcomas?Hull etat.1965SV381 day1421154—280?Undifferentiated sarcomas?Hull etat.1965CELO1
day693388—195?Fibrosarcomas?Sarma etat.1965BovineAdeno 31 day454924— 67?Undifferentiatedtumors?Darbyshire1966ICH1
day1414360-403?Sarcomas?Sarma etat.1967RousRSV
(S-R)1 day149313?SarcomasSomeAhlstrom andForsby1962RSV4day1310014?SarcomasSomeAhlstrom andForsby1962RSV14day86320?SarcomasSomeAhlstrom andForsby1962RSVAdult117364—120?SarcomasSomeAhlstrom
andForsby1962RSV(Bryan)1 day72952124SarcomasSomeGirardi et at.1964
Oncogenicity of viral agents in hamsters. BPV, bovine papilloma virus; SV, simian virus; CELO, avian adenovirus ; ICH, infectiouscanine hepatitis ; ItSV, Rosa sarcoma virus ; S-R, Schmidt-Ruppin.
a Different strains of adenovirus type 12 were used.
1600 CANCER RESEARCH VOL.28
Research. on February 14, 2019. © 1968 American Association for Cancercancerres.aacrjournals.org Downloaded from

Bovine Papilloma Virus in Hamsters
ster is not known. However, it is probably relatively small incomparison to quantities reported for tumor induction by polyoma, SV4O, and the adenoviruses. Habel (18) stated that certam strains of polyoma virus, even in large doses, failed toinduce tumors in a high percentage of hamsters. BPV can bediluted to the point where the suspension no longer producestumors in the hamster (Table 1, Experiment 2).
REFERENCES
1. Ahlstrom, C. G., and Forsby, N. Sarcomas in Hamsters afterInjection with Rosa Chicken Tumor Material. J. Exptl. Med.,115: 839—852,1962.
2. Allison, A. C. Viruses Inducing Skin Tumors in Animals. In:A. J. Rook and G. S. Walton, (eds.), Comparative Physiologyand Pathology of the Skin, pp. 665-684. Oxford : BlackwellScientific Publications, 1965.
3. Allison, A. C., Chesterman, F. C., and Baron, S. Induction ofTumors in Adult Hamsters with Simian Virus 40. J. Natl.CancerInst.,38:567—572,1967.
4. Bagdonas, V., and Olson, C. Observations on the Epizootiologyof Cutaneous Papillomatosis (Warts) of Cattle. J. Am. Vet.Med. Assoc., 140: 50-52, 1953.
5. Barski, G., Chlap, Z., Gotlieb-Stemateky, T., and Charlier, M.Neoplasms and Haemorrhagic Disease in Adult Hamsters withPolyoma Virus. Nature, 195: 298-299, 1962.
6. Boiron, M., Levy, J. P., Thomas, M., Friedmann, J. C., andBernard, J. Some Properties of Bovine Papilloma Virus.Nature, Wi: 423—424,1964.
7. Brobst, D., and Hinsman, E. J. Electron Microscopy of theBovine Cutaneous Papilloma. Pathol. Vet. (Basel), 8: 193-207, 1966.
8. Cheville, N. F. Studies on Connective Tissue Tumors in theHamster Produced by Bovine Papilloma Virus. Cancer Rca.,@6:2334—2339,1966.
9. Cheville, N. F., and Olson, C. Epithelial and FibroblasticProliferation in Bovine Cutaneous Papillomatosis. Pathol.Vet. (Basel), 1: 248—257,1964.
10. Creech, G. T. Experimental Studies of the Etiology of Common Warts in Cattle. J. Agr. Res., @9:723—737,1929.
11. Darbyshire, J. H. Oncogenicity of Bovine Adenovirus Type 3
in Hamsters. Nature, p11: 102, 1966.12. Defendi, V. Induction of Tumors by Polyoma in Adult Ham
stem. Nature, 188: 508—509,1960.13. Eddy, B. E. Simian Virus 40 (SV4O) : An Oncogenic Virus.
Progr. Exptl. Tumor Res., 4: 1—26,1964.14. Eddy, B. E., Stewart, S. A., Young, R., and Mider, G. B. Neo
plasms in Hamsters Induced by Mouse Tumor Agent Passedin Tissue Culture. J. Natl. Cancer Inst., 20: 747—762,1958.
15. Fujimoto, Y., and Olson, C. The Fine Structure of the Bovine
Wart. Pathol. Vet. (Basal), 3: 659-684, 1966.16. Girardi, A. J., Hilleman, M. R., and Zwickey, R. E. Tests in
Hamsters for Oncogenic Quality of Ordinary Viruses IncludingAdenovirus Type 7. Proc. Soc. Exptl. Bid. Med., 115: 1141—1150, 1964.
17. Girardi, A. J., Sweet, B. H., and Hilleman, M. R. Factors
Influencing Tumor Production in Hamsters by VacuolatingVirus SV-40. Proc. Soc. Exptl. Biol. Med., 11@: 662-667, 1963.
18. Habel, K. Malignant Transformation by Polyoma Virus. Ann.Rev. Microbiol., 17: 167—178,1963.
19. Ham, A. W. Histology. The Integumentary System, Ch. 23,pp. 613-644. Philadelphia : J. P. Lippincott Co., 1965.
20. Harris, J. C., Chesterman, F. C., and Negroni, G. Inductionof Tumors in Newborn Ferrets with Mill Hill Polyoma Virus.Lancet, 1: 788—791,1961.
21. Huebner, R. J., Rowe, W. P., and Lane, W. T. OncogenicEffects of Human Adenovirus Types 12 and 18. Proc. Nat!.Acad. Sci., 48: 2051—2058,1962.
22. Hull, R. N., Johnson, T. S., Culbertson, C. G., Reimer, C. B.,and Wright, H. F. Oncogenicity of the Simian Adenoviruses.Science, 150: 1044—1046,1965.
23. Koprowski, H. Viral-induced Tumors and Leukemias. Am. J.Med.,38: 716—725,1965.
24. Kukaine, R. A., and Nagayeva, L. I. Further Studies of SpecificSelection Inherent to Shope Papilloma Virus. Abstracts 9thIntern. Cong. Microbiol., Moscow, p. 519, 1966.
25. Lasneret, J., Cherat, J. C., Levy, J. P., and Boiron, M. EtudeDes Tumeurs ProvoquéesChez Le Hamster Par Le Virus DcLa Papillomatose Bovine. Pathol. Biol., 13: 1174-1179, 1965.
26. McEntee, K. Transmissible Fibropapillomas of the ExternalGenitalia of Cattle. Rept. N. Y. Vet. Coll., p. 28, CornellUniversity, Ithaca, New York, 1952.
27. Olson, C., and Cook, R. H. Cutaneous Sarcoma-like Lesionsof the Horse Caused by the Agent of Bovine Papilloma. Proc.Soc. Exptl. Biol. Med., 77: 281—284,1951.
28. Olson, C., Pamukcu, A. M., Brobst, D. F., Kowalczyk, T.,Satter, E. J., and Price, J. M. A Urinary Bladder Tumor Induced by a Bovine Cutaneous Papilloma Agent. Cancer Res.,19: 779—782,1959.
29. Sarma, P. S., Huebner, R. J., and Lane, W. T. Induction ofTumors in Hamsters with an Avian Adenovirus (CELO).Science, 149: 1108, 1965.
30. Sarma, P. S., Vass, W., Huebner, R. J., Igel, H., Lane, W. T.,and Turner, H. C. Induction of Tumors in Hamsters withInfectious Canine Hepatitis Virus. Nature, 215 : 293—294,1967.
31. Selye, H. The Mast Cells. Baltimore : Waverly Press, Inc.,
1965.32. Simpson, W. L. Distribution of Mast Cells as a Function of
Age and Exposure to Carcinogenic Agents. Ann. N. Y. Acad.Sci., 103: 4—19,1963.
33. Smithies, L. K., and Olson, C. Antigen of Bovine CutaneousPapilloma Detected by Fluorescent Antibodies. Cancer Res.,51: 1557—1559,1961.
34. Smithies, L. K., and Olson, C. Correlation of Immunofluorescence and Infectivity in the Developing Bovine CutaneousPapilloma. Cancer Res., 24: 674-681, 1964.
35. Willis, R. A. Pathology of Tumors, p. 647, Washington, D. C.:Butterworth Inc., 1960.
36. Wooding, W. Susceptibility of Various Animals to BovineCutaneous Papilloma Material. M. S. Thesis, University ofWisconsin, Madison, 1964.
37. Yabe, Y., Tretin, J. J., and Taylor, T. Cancer Induction inHamsters by Human Type 12 Adenovirus. Effect of Age andof Virus Dose. Proc. Soc. Exptl. Biol. Med., 11: 343—344,1962.
1601AUGUST 1968
Research. on February 14, 2019. © 1968 American Association for Cancercancerres.aacrjournals.org Downloaded from

Martin 0. RobI and Carl Olson
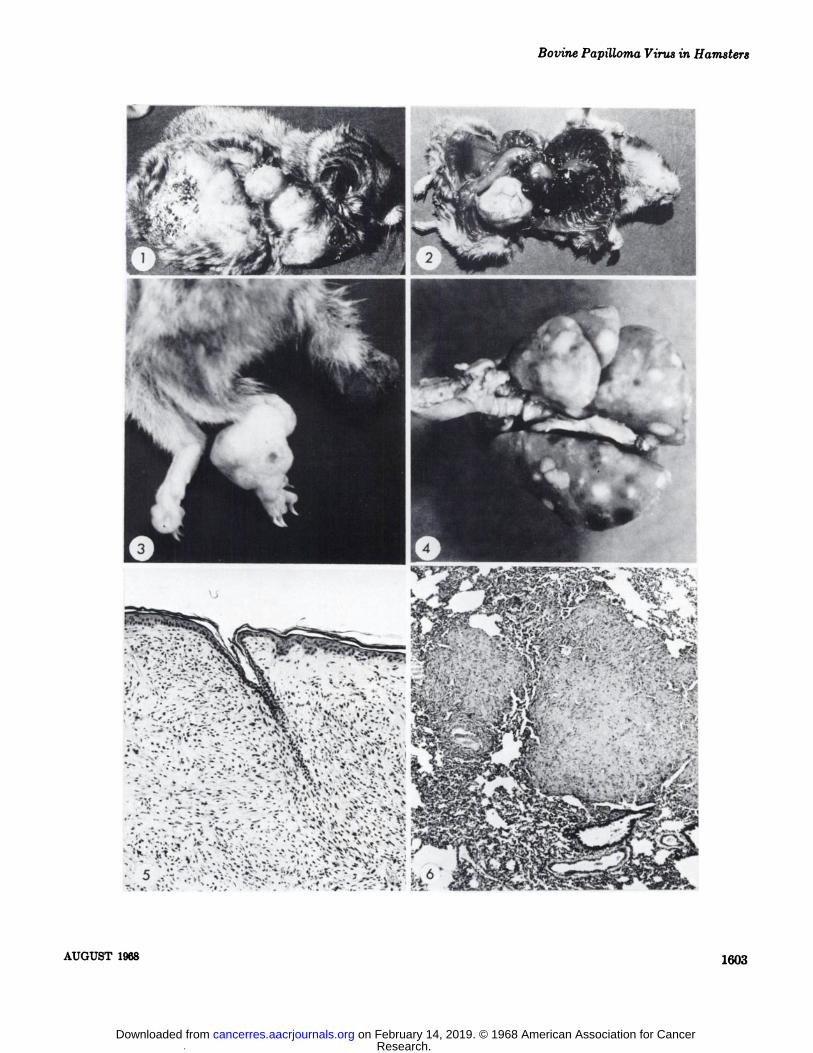
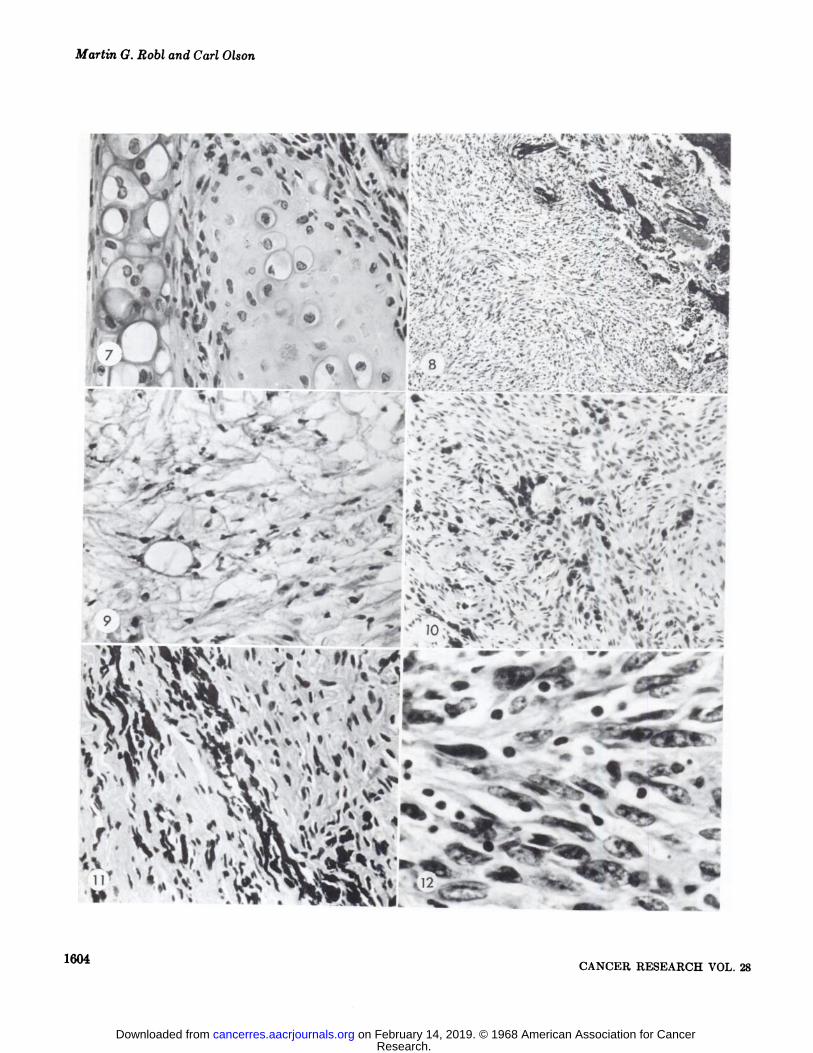
Fig. 1. Large tumor of a hamster one year after subcutaneous injection of bovine papilloma virus suspension. The tumor was firstobserved 28 weeks after inoculation.
Fig. 2. Tumor mass that extended into the abdominal cavity and displaced the viscera.Fig. 3. Secondary tumor nodules that developed on the feet and tail. The primary tumor at the inoculation site had developed to 3
cm in diameter before these tumors were first observed.Fig. 4. Disseminated tumor nodules in the lung.Fig. 5. Tumor growth originated in the subcutaneous tissue. Tumor growth replaced the adnexal structures of the skin. The epithe
hum is not involved in the neoplastic growth. H & E, x 115.Fig. 6. Microscopic tumor nodules in the lung. Nodules are in close proximity to blood vessels. There is no host response to these
tumor nodules. H & E, x 75.Fig. 7. Chondroblast proliferation adjacent to normal cartilage of ear. Chondroblast.s, fibroblasts, and melanocytes comprised the
tumor that developed at site of inoculation with bovine papilloma virus suspension in the ear. H & E, X 450.Fig. 8. Tumor cells that have invaded through the periosteum, endosteum, and into the bone marrow cavity of this vertebra. Note
the disorganized pattern of the fibroblasts. This hamster had disseminated tumor nodules in the lung, liver, and on the serosal surface of the intestines. H & E, x 115.
Fig. 9. An area of tumor undergoing degeneration. Note the small pyknotic nuclei and the loose fine fibrillar remnants of the degenerating fibroblasts. Calcium deposition was located adjacent to this area. H & E, X 500.
Fig. 10. An area of tumor with a large number of mast cells. Mast cells were confirmed by the Kingsley and Giemsa technics. Notethe unorganized pattern of the fibroblasts with a whorling tendency. H & E, x 325.
Fig. 11. Junction of pigmented nodule with an adjacent fibroblastic nodule. H & E, X 500.Fig. 12. High power of fibroblastic tumor cells. Note the variation in the size and shape of the nuclei and the absence of mitotic
figures. H & E, x 800.
1602 CANCER RESEARCH VOL.28
Research. on February 14, 2019. © 1968 American Association for Cancercancerres.aacrjournals.org Downloaded from
@4f@ .,. -.@...? @$
@, @‘@‘@:@
) .. ;@L ‘@. t
:@
‘..j. . . . - . .::@!J.@.@ - . .@ . .@ 4'. ;@.@ .,
1603
t@ri;@:@ &a.
@rs@-
3 7 .@!@A.
I
3
.-@@
.@ .- ..--@@@@ ‘
@ •:@@@ _•@@@ @.c.
:@.@ • ,@-.@ @.‘“@.@ . @,@..
@ — 4@@
@ .- ..‘,@@@ ‘. ..“
@ ‘\‘@—@ :-@ , @k \
;..c. .@ .: .@ @. _@@@@@
.@ .@ @: @:@@
@ ‘ S@ -@ @.i . i@ : @.@ .@ :@ @:#@@
@. ,,tt,l@ S':.@ ,@ —.; @@‘,:_•@@@. 5@@
Bovine Papilloma Virus in Hamsters
AUGUST 1968
Research. on February 14, 2019. © 1968 American Association for Cancercancerres.aacrjournals.org Downloaded from
Martin G. Robi and Cart Olson
.,@ I@
,@.,,,
@ 4 ... ‘ @—@ , .
1% • •@@ :.,,y@@@
@ .-‘.‘@‘ ,‘@/.a, i.,' : ;@ .@ . -@—
@ f4@1 .@.... —-—-@ .@
. ‘I, I@ •y@—@:@ _ •‘I, .‘ _ .@i -,.. •:@ •‘,@-:-td -‘.@ ,@V ,@, -@ I.
. 8 / €.,_, r@@@I@@ ,‘* - .-.@ ‘ .@.@
r- @-.@‘@H:qI@@@ ‘,:@ :: ,@ ;‘i@@lIP@―•@:1.:r@
@• , - . — @â€%_‘•_. :@ -4•@ ‘__@ .:. —-
@- $ e@ .‘@ s- ‘@ •@,@ ‘. .@@
. , .; .— ,.‘:.@@ - c@@
“ @. ,@. _, -•. - ••@. #l@p_
:@( 1@@L:y- -dl@'@ @#@ 5.d@ (@@
@ ‘@- # ‘ .,@..@ . . ,@ . -.@• %@@@ . I •@
@ C @. @\.,.._•. —@ , , •1r' .@ â€â€˜@‘.:@-•‘i-,;@_@:.@ #‘•s‘@.-@Ib:'@ 4, , . % @. @— F.@@ L., ‘@ @.@ ‘@@ ,@@ :â€-@ ‘
5' S@ db;@@.F@ :*t@@4 . @,
J_•@ .@ £@@@
@ ,-.@.@ ,@ @.‘ t •:@@ 4b.@@L* .@
,l1@ø@ - C
@ •@Ii ‘P •:@:;-@@@
a
{a'
,
. t@@ :@‘t@u@
.. .@
@ @‘,#:@@@
/4/
I-@
-,@A‘ 9(i@
‘@—@‘
‘ii
1604CANCER RESEARCH VOL. 28
4•@@; eL@‘Al@
@l‘@
C
Research. on February 14, 2019. © 1968 American Association for Cancercancerres.aacrjournals.org Downloaded from

1968;28:1596-1604. Cancer Res Martin G. Robl and Carl Olson Oncogenic Action of Bovine Papilloma Virus in Hamsters
Updated version
http://cancerres.aacrjournals.org/content/28/8/1596
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/28/8/1596To request permission to re-use all or part of this article, use this link
Research. on February 14, 2019. © 1968 American Association for Cancercancerres.aacrjournals.org Downloaded from