
Occurrence of terminal a2 —> 8-linked disialylated - Glycobiology
11
Glycobiology vol. 7 no. 2 195-205, 1997 Occurrence of terminal a2 —> 8-linked disialylated poly-N-acetyllactosamine chains with Le x and I antigenic glycotopes in tetraantennary arms of an AMinked glycoprotein isolated from rainbow trout ovarian fluid Yoko Funakoshi, Tomohiko Taguchi, Chihiro Sato, Ken Kitajima 4 , Sadako Inoue 1 , Howard R.Morris 2 , Anne Dell and Yasuo Inoue lr3 Department of Biophysics and Biochemistry, Graduate School of Science, University of Tokyo, Tokyo 113, Japan, 'Institute of Biological Chemistry, Acaderrua Sinica, Nan Kang, Taipei, Taiwan, and 2 Department of Biochemistry, Imperial College of Science, Technology and Medicine, London SW7 2AZ, United Kingdom 'To whom correspondence should be addressed 4 Present address: Department of Applied Biological Sciences, School of Agricultural Sciences, Nagoya University, Chikusa, Nagoya 464-01, Japan The Pronase digestion of a 54K glycoprotein present in ovarian fluid of rainbow trout yielded a major glycopep- tide. Carbohydrate compositional analysis revealed that' this glycopeptide was likely to possess a single large N- glycan chain having low molecular weight oligomers of N- acetylneuraminic acid (oligoNeu5Ac). Structural studies of this glycopeptide revealed novel ct2 —> 8-linked disialylated poIy-A'-acetyllactosamine chains with Le x and I antigenic determinants on the A'-linked tetraantennary core glycan. In our recent studies (Kitazume,S., Kitajima,K., Inoue^S., Inoue,Y. and Troy,F.A. (1994) / . Biol Chem. 269, 10330- 10340) we presented evidence that synthesis of a2 — > 8-linked polysialic acid (polySia) chains is a two-step pro- cess in which chain initiation is catalyzed by an ct2 —• 8-sialyltransferase (at2 -» 8-ST; initiase) that catalyzes syn- thesis of the first Sia ot2 -* 8-linkage, forming the disialic acid (diSia) unit, Siaa2 -» 8-Siaa2 —» 6-Gal-.Chain poly- merization is then postulated to be catalyzed by a second enzyme, an a2 —» 8-polyST ("polymerase") that converts the diSia units to polySia chains. The present structural studies leading to the discovery of a2 —> 8-linked disi- alylated units that terminate poIy-A'-acetyllactosamine chains in an A'-linked glycoprotein is further evidence in support of our hypothesis that more than one sialyltrans- ferase activity is required for polySia chain synthesis and polymerization. Key words: disialosyl group/I antigen/Le x antigen/poly-A'- acetyllactosamineAetraantennary A'-glycan Introduction Mature eggs of rainbow trout are surrounded by and ovulated with ovarian fluid. Suggestions for the physiological function of ovarian fluid must take into account its location and that the fluid has a homeostatic role on unfertilized eggs immediately before ovulation. Other as yet undefined roles at fertilization have also been postulated (Kanamori et al., 1989). Previously, we identified in the ovarian fluid a unique highly acidic gly- coprotein (designated KDN-gp-OF), in which a large number of 0-linked ot2 —» 8-oligo/polyKDN-containing glycan chains were attached to the polypeptide core (Kanamori et al., 1989; Kitazume et al., 1992). A KDN-rich glycoprotein, which is similar if not identical, was also found as an egg cell surface glycoprotein, and denoted as KDN-gp-VE (Inoue et al., 1988; Kanamori et al., 1990). The first example of the occurrence of ot2 —> 8-linked oligo/polysialic acid chains in eukaryotes was made almost two decades ago by Inoue and Iwasaki (1978), who discovered an egg glycoprotein rich in sialic acid. There is now considerable evidence that the a2 —> 8-linked polySia glycotopes occur in glycoproteins from a variety of species ranging from bacteria to human (Inoue and Iwasaki, 1978; James and Agnew, 1987; Roth et al., 1992; Troy, 1992). Our finding of a2 —» 8-polySia chains in rainbow trout eggs stimu- lated our interest in the biosynthesis of such polySia chains. Recently, we undertook studies to elucidate the biosynthetic mechanism and developmental expression of 0-linked polySia chains attached a2 — ¥ 6 to the proximal GalNAc residue during oogenesis of rainbow trout eggs (Kitazume et al., 1994). As the results of these studies, the general outline of the biosynthetic pathway emerged, and was shown to involve at least three distinct sialyltransferases (ST), as follows: (l)an a2 —» 8-ST that catalyze the reaction -> 3Gaipi -» 3GaINAcal -» Ser/ Thr + CMP-Sia -> Siaa2 -> 6( -> 3Gal£ -> 3)GalNAcal -> Ser/Thr + CMP; (2) an a2 -> 8-ST (initiase) that is respon- sible for catalyzing the reaction, Siaa2 —» 6( —> 3Gal(31 -> 3)GalNAcal -> Ser/Thr + CMP-Sia -> Siaa2 -> 8Siaa2 -> 6( -> 3Gaip -» 3)GalNAcal -> Ser/Thr + CMP; and (3) an a2 — > 8-polyST (polymerase) catalyzing Siaa2 -> 8Siaa2 — ¥ 6( -> 3Galpl -> 3)GalNAcal ^ Ser/Thr + nCMP-Sia -^ [Siaa2 -> 8] n Siaa2 -» 8Siaa2 -^ 6( -> 3Galpl -* 3)- GalNAca 1 -» Ser^Tir + nCMP. However, definitive evidence as to how polySia chain synthesis starts and how the chains are elongated are still open questions. The impetus for the present study came from the finding of a unique sialoglycoprotein in rainbow trout ovarian fluid (OF) whose carbohydrate composition indicated the possibility that a glycopeptide derived from this glycoprotein possessed lower oligoSia residues on oligo-A'-acetyllactosamine units. Struc- tural analysis of the A'-linked glycan chain(s) present in this ovarian fluid sialoglycoprotein was of considerable interest for two reasons. First, the study has provided important structural information relevant to the mechanism of biosynthesis of poly- Sia chains. Second, since oligosialyl groups have been reported in only a limited number of glycoproteins (Finne et al., 1977; Fukuda et al., 1984a, 1985, 1987), despite being common con- stituents of gangliosides, a detailed knowledge of oligoSia- containing glycan structures is a prerequisite to understanding not only their biosynthesis but also their reactivity with anti- bodies. We report in this article the occurrence and identification of © Oxford University Press 195 Downloaded from https://academic.oup.com/glycob/article-abstract/7/2/195/598111 by guest on 03 April 2019
Transcript of Occurrence of terminal a2 —> 8-linked disialylated - Glycobiology

Glycobiology vol. 7 no. 2 195-205, 1997
Occurrence of terminal a2 —> 8-linked disialylated poly-N-acetyllactosamine chainswith Lex and I antigenic glycotopes in tetraantennary arms of an AMinkedglycoprotein isolated from rainbow trout ovarian fluid
Yoko Funakoshi, Tomohiko Taguchi, Chihiro Sato, KenKitajima4, Sadako Inoue1, Howard R.Morris2, AnneDell and Yasuo Inouelr3
Department of Biophysics and Biochemistry, Graduate School of Science,University of Tokyo, Tokyo 113, Japan, 'Institute of Biological Chemistry,Acaderrua Sinica, Nan Kang, Taipei, Taiwan, and 2Department ofBiochemistry, Imperial College of Science, Technology and Medicine,London SW7 2AZ, United Kingdom
'To whom correspondence should be addressed4Present address: Department of Applied Biological Sciences, School ofAgricultural Sciences, Nagoya University, Chikusa, Nagoya 464-01, Japan
The Pronase digestion of a 54K glycoprotein present inovarian fluid of rainbow trout yielded a major glycopep-tide. Carbohydrate compositional analysis revealed that'this glycopeptide was likely to possess a single large N-glycan chain having low molecular weight oligomers of N-acetylneuraminic acid (oligoNeu5Ac). Structural studies ofthis glycopeptide revealed novel ct2 —> 8-linked disialylatedpoIy-A'-acetyllactosamine chains with Lex and I antigenicdeterminants on the A'-linked tetraantennary core glycan.In our recent studies (Kitazume,S., Kitajima,K., Inoue^S.,Inoue,Y. and Troy,F.A. (1994) / . Biol Chem. 269, 10330-10340) we presented evidence that synthesis of a2 —>8-linked polysialic acid (polySia) chains is a two-step pro-cess in which chain initiation is catalyzed by an ct2 —•8-sialyltransferase (at2 -» 8-ST; initiase) that catalyzes syn-thesis of the first Sia ot2 -* 8-linkage, forming the disialicacid (diSia) unit, Siaa2 -» 8-Siaa2 —» 6-Gal-.Chain poly-merization is then postulated to be catalyzed by a secondenzyme, an a2 —» 8-polyST ("polymerase") that convertsthe diSia units to polySia chains. The present structuralstudies leading to the discovery of a2 —> 8-linked disi-alylated units that terminate poIy-A'-acetyllactosaminechains in an A'-linked glycoprotein is further evidence insupport of our hypothesis that more than one sialyltrans-ferase activity is required for polySia chain synthesis andpolymerization.
Key words: disialosyl group/I antigen/Lex antigen/poly-A'-acetyllactosamineAetraantennary A'-glycan
Introduction
Mature eggs of rainbow trout are surrounded by and ovulatedwith ovarian fluid. Suggestions for the physiological functionof ovarian fluid must take into account its location and that thefluid has a homeostatic role on unfertilized eggs immediatelybefore ovulation. Other as yet undefined roles at fertilizationhave also been postulated (Kanamori et al., 1989). Previously,we identified in the ovarian fluid a unique highly acidic gly-
coprotein (designated KDN-gp-OF), in which a large numberof 0-linked ot2 —» 8-oligo/polyKDN-containing glycan chainswere attached to the polypeptide core (Kanamori et al., 1989;Kitazume et al., 1992). A KDN-rich glycoprotein, which issimilar if not identical, was also found as an egg cell surfaceglycoprotein, and denoted as KDN-gp-VE (Inoue et al., 1988;Kanamori et al., 1990). The first example of the occurrence ofot2 —> 8-linked oligo/polysialic acid chains in eukaryotes wasmade almost two decades ago by Inoue and Iwasaki (1978),who discovered an egg glycoprotein rich in sialic acid. There isnow considerable evidence that the a2 —> 8-linked polySiaglycotopes occur in glycoproteins from a variety of speciesranging from bacteria to human (Inoue and Iwasaki, 1978;James and Agnew, 1987; Roth et al., 1992; Troy, 1992). Ourfinding of a2 —» 8-polySia chains in rainbow trout eggs stimu-lated our interest in the biosynthesis of such polySia chains.Recently, we undertook studies to elucidate the biosyntheticmechanism and developmental expression of 0-linked polySiachains attached a2 —¥ 6 to the proximal GalNAc residue duringoogenesis of rainbow trout eggs (Kitazume et al., 1994). As theresults of these studies, the general outline of the biosyntheticpathway emerged, and was shown to involve at least threedistinct sialyltransferases (ST), as follows: (l)an a2 —» 8-STthat catalyze the reaction -> 3Gaipi -» 3GaINAcal -» Ser/Thr + CMP-Sia -> Siaa2 -> 6( -> 3Gal£ -> 3)GalNAcal-> Ser/Thr + CMP; (2) an a2 -> 8-ST (initiase) that is respon-sible for catalyzing the reaction, Siaa2 —» 6( —> 3Gal(31-> 3)GalNAcal -> Ser/Thr + CMP-Sia -> Siaa2 -> 8Siaa2-> 6( -> 3Gaip -» 3)GalNAcal -> Ser/Thr + CMP; and (3) ana2 —> 8-polyST (polymerase) catalyzing Siaa2 -> 8Siaa2 —¥6( -> 3Galpl -> 3)GalNAcal ^ Ser/Thr + nCMP-Sia-^ [Siaa2 -> 8]nSiaa2 -» 8Siaa2 -^ 6( -> 3Galpl -* 3)-GalNAca 1 -» Ser^Tir + nCMP. However, definitive evidenceas to how polySia chain synthesis starts and how the chains areelongated are still open questions.
The impetus for the present study came from the finding ofa unique sialoglycoprotein in rainbow trout ovarian fluid (OF)whose carbohydrate composition indicated the possibility thata glycopeptide derived from this glycoprotein possessed loweroligoSia residues on oligo-A'-acetyllactosamine units. Struc-tural analysis of the A'-linked glycan chain(s) present in thisovarian fluid sialoglycoprotein was of considerable interest fortwo reasons. First, the study has provided important structuralinformation relevant to the mechanism of biosynthesis of poly-Sia chains. Second, since oligosialyl groups have been reportedin only a limited number of glycoproteins (Finne et al., 1977;Fukuda et al., 1984a, 1985, 1987), despite being common con-stituents of gangliosides, a detailed knowledge of oligoSia-containing glycan structures is a prerequisite to understandingnot only their biosynthesis but also their reactivity with anti-bodies.
We report in this article the occurrence and identification of
© Oxford University Press 195
Dow
nloaded from https://academ
ic.oup.com/glycob/article-abstract/7/2/195/598111 by guest on 03 April 2019

Y.Funakoshl et aL
a2 —» 8-linked disialylated units that cap poly-A/-acetyllacto-samine chains that are attached to tetraantennary //-linked coreglycans.
Results
Isolation and purification of glycopeptide OF-gp fromrainbow trout ovarian fluid
As described under Materials and methods, the delipidatedphenol extracted ovarian fluid (39 mg as Neu5Ac) was di-gested with Pronase and chromatographed on DEAE-Toyopearl 650M (Figure la). The fractions containing sialicacid and neutral sugars (17 mg) were eluted at 0.07-0.21 MNaCl, pooled, and subjected to Sephacryl S-200 chromatogra-phy (Figure lb) to eliminate KDN-containing glycopeptides.The KDN-free sialoglycopeptide(s) was further chromato-graphed on a Sephadex G-50 column (results not shown), anda symmetrical peak at K,,v 0.33 was obtained and labeled OF-gp. The molecular mass of OF-gp was estimated to be about 11kDa based on the Kav value of 0.47 on Sephacryl S-200 chro-matography. The yield of OF-gp was 14 mg from 3.5 1 of theovarian fluid.
Chemical analysis of the glycopeptide, OF-gp
The carbohydrate composition of OF-gp is given in Table 1.Sialic acid was found to be exclusively Neu5Ac. The aminoacid composition of OF-gp was Asn/Thr/Ser/Ala = 1.0:1.2:0.98:0.53. OF-gp contained 1.0 Asn when Man was set equalto 3.0, suggesting that OF-gp had only one N-linked glycanchain.
The carbohydrate composition remained unchanged after
hydrazinolysis except for GalNAc, indicating that all of thesugar components besides GalNAc were constituents of an//-linked carbohydrate chain originally present in OF-gp. Thecalculated molecular weight of the glycan chain on the basis ofcomposition analysis was about 9600, which was in reasonablygood agreement with the molecular weight of 11 K estimatedby Sephacryl S-200 gel filtration (see above). Based on theseresults we concluded that OF-gp contains a single glycanchain.
Methylation analysis of glycopeptides
The results of methylation analysis of OF-gp, asialo-OF-gp,and dF-asialo-OF-gp are shown in Table H. The substitutionpatterns of the Man residues were identical within this series ofglycopeptides. In favor of a tetraantennary glycan core com-prising 2,4-Man/2,6-Man residues otl —» 3- and otl —» 6-linkedto the |3-Man residue, 1 mol each of three different di-0-substituted Man residues was identified and neither of mono-0-substituted or tri-<9-substituted Man was detected. On re-moval of Neu5Ac residues from OF-gp, the proportion of theterminal Gal increased at the expense of 3-0-substituted Gal,indicating Neu5Ac was originally attached to O-3 of the Galresidues. On removal of two-thirds of Fuc residues from asialo-OF-gp the proportion of 4-O-substituted GlcNAc increased atthe expense of parts of the 3,4-di-O-substituted GlcNAc and allthe 4,6-di-O-substituted GlcNAc residues, thus indicating thepresence of Fuel —» 3( —> 4GlcNAc) and Fuel —» 6( —> 4GlcNAc) linkages. The presence of these two types of struc-tures was also substantiated by the ^ -NMR measurements(see below). Detection of 0.61 mol of 4,6-di-O-substitutedGlcNAc residue in asialo-OF-gp suggested that 60% of the
(a) (b)
50 100 150Fraction number
200
2 •
1 -
1 /
.y *
vt
i
L200 400 600
Elution volume (ml)
Fig. 1. Preparation of OF-gp from the Pronase digests of rainbow trout ovarian fluid, (a) DEAE-Toyopearl 65OM chromatography. The elution profile wasmonitored by the absorbance at 230 run (open circle) and by the resorcinol method (A580, closed circle) for //-acylneuraminic acid. The broken linerepresents a gradient of NaCl. Fractions indicated by the bar were pooled and subjected to a gel filtration on Sephacryl S-200. (b) Sephacryl S-200 gelfiltration. The elution as described in (a) was examined by the absorbance at 230 nm (open circle) and for N-acylneuraminic acid by the TBA method aftermild acid hydrolysis (A549, solid circle). Fractions indicated by the bar were pooled, passed through a Sephadex G-50 column, and designated OF-gp.
196
Dow
nloaded from https://academ
ic.oup.com/glycob/article-abstract/7/2/195/598111 by guest on 03 April 2019

A novel disialylated tetraantennary A'-linked glvcan chains
Table L Carbohydrate composition of OF-gp, asialo-OF-gp, anddF-asialo-OF-gp
OF-gp asialo-OF-gp dF-asialo-OF-gp OF-GPf
FucManGalGalNAcb
GlcNAcNeu5Ac
6.33.0
121.0
167.4
6.33.0
12n.d.c
15d
2.3*3.0
11n.d.14—
1 63.09.20.8
145.4
Values are molar ratios relative to Man set equal to 3.0 mol.Two-thirds of the Fuc residues were removed from the glycan chain upontreatment with 0.05 N trifluoroacetic acid at 80°C for 2 h.•"GalNAc was quantitated by the ammo acid analysis as described inMaterials and methods.cn.d. not determined.d—, Not detected.
4-0-substituted nonreducing terminal GlcNAc residues werefucosylated at the O-6 position.
The finding of 3,6-di-(9-substituted Gal indicated that OF-gpcontained a branching structure. Either GlcNAc or Gal waspossibly attached to the branched Gal residues, but substitutionby GlcNAc was most likely because Gal-Gal sequence was notdetected on analysis of hydrazinolysis—nitrous acid deamina-tion products of dF-asialo-OF-gp (see the following section).Thus, the I antigenic determinant, Gal-GlcNAc —> 6(Gal-GlcNAc —» 3) Gal —> , was indicated to exist in OF-gp.
Hydrazinolysis-nitrous acid deamination of dF-asialo-OF-gp
The product formed by hydrazinolysis-nitrous acid deamina-tion of dF-asialo-OF-gp followed by NaBH4 reduction wasapplied to a column of Bio-Gel P-2. Neutral sugar analysis ofthe column effluent by the phenol-H2SO4 method gave twomajor peaks, HN-1 (lower-molecular-weight material) andHN-2 (higher-molecular-weight material eluted at the void vol-ume of the column) in the proportion of 5:7 on the basis ofhexose contenr(data not shown). When examined by TLC inl-propanol/NH4OH/water (6:1:2.5, v/v) solvent, peak HN-1gave a single spot with Rf = 1.8 relative to Gal, and it wasfound to contain Gal and anhydro Man-ol in a 1:1 ratio asestimated by GLC analysis. Methylation analysis showed thepresence of 1 mol each of unsubstituted terminal Gal and 4-O-substituted 2,5-anhydro Man-ol in HN-1. Peak HN-2 showed adiffuse band on TLC, and carbohydrate analysis indicated that
Table IL Methylation analysisdF-asialo-OF-gp
Terminal FucTerminal Gal3-O-Substituted Gal3,6-di-O-Substituted Gal2,4-di-O-SubstitutedMan2,6-di-O-Substituted Man3,6-di-O-Substituted ManTerminal GlcNAc4-0-Substituted GlcNAc3,4-di-O-Substituted GlcNAc4,6-di-O-Substituted GlcNAc
of OF-gp,
OF-gp
6.31.67.73.10.871.01.01.39.25.10.45
asialo-OF-gp, and
asiao-OF-gp
6.34.93.92.70.780.811.01.67.75.30.61
dF-asialo-OF-gp
2.34.83.23.11.00.911.01.8120.73—
Values are molar ratios relative to 3,6-di-O-substituted Man that set equal to1.0 mol.
HN-2 was derived from the trimannosidic core moiety of dF-asialo-OF-gp. However, owing to shortage of material no fur-ther structural studies were carried out on HN-2. The aboveresults showed, however, that OF-gp contained A'-acetyl-lacosamine units.
Endo-$-galactosidase digestion of asialo-OF-gpand dF-asialo-OF-gp
Since the presence of multiple N-acetyllactosamine units in theperipheral portion of the glycan chain was suggested, asialo-OF-gp and dF-asialo-OF-gp (20 \ig each as hexose) were eachsubjected to endo-3-galactosidase digestion (1.25 mU, 25°C,26h). Asialo-OF-gp was resistant to the enzyme, while dF-asialo-OF-gp gave a small amount (yield, <10% in hexoseamount base) of a product having the same migration rate asGlcNAcpi -> 3Gal on TLC when treated with the enzyme.These results were compatible with a structure in which N-acetyllactosamine units may constitute an I antigenic branchedstructure and/or a repeated linear Lex structure, both of whichare known to be poorly susceptible to E.freundii endo-(B-galactosidase (Fukuda et al., 1984b, c).
Structural elucidation of the sialyl moiety in OF-gp
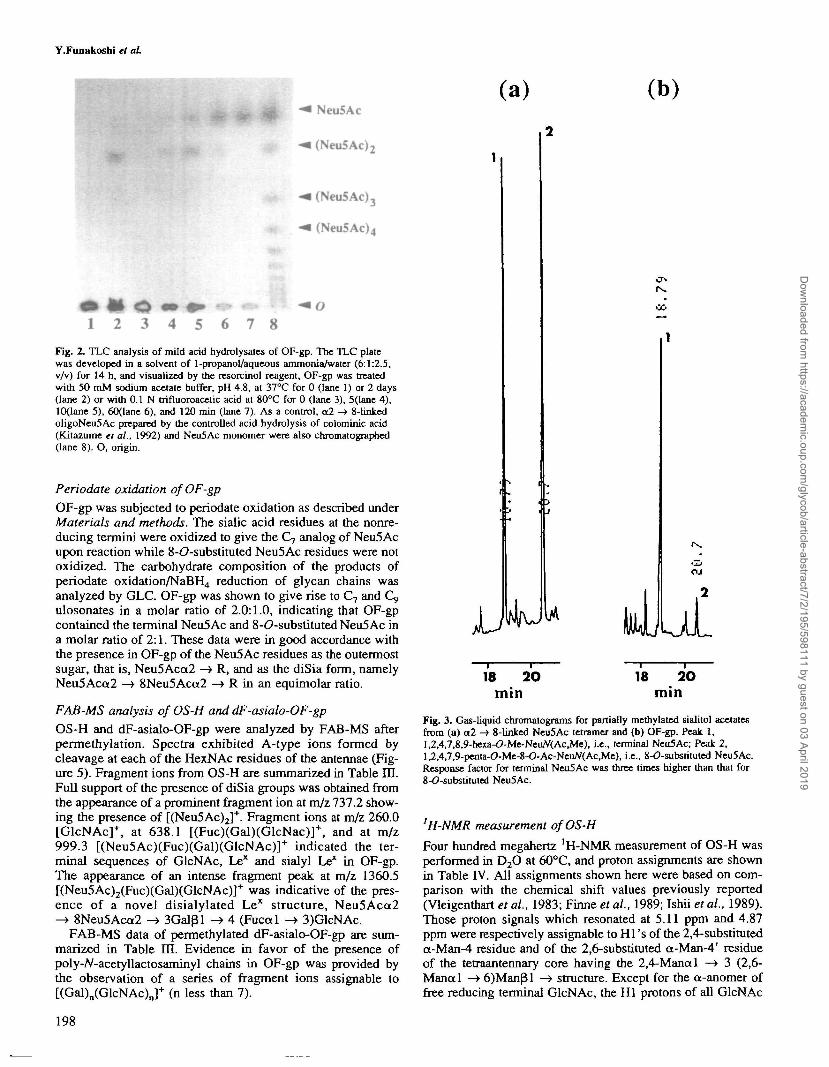
TLC analysis of mild acid hydrolysate of OF-gp
To investigate the possible presence of oligoSia groups in OF-gp, OF-gp and OS-H (free glycan prepared from OF-gp byhydrazinolysis as described under Materials and methods)were subjected to controlled hydrolysis in 50 mM sodium ac-etate buffer (pH 4.8) at 37°C for 2 days (Kitazume et al.,1992).The hydrolysates were examined by TLC (Figure 2, lane 2),showing two bands migrating identically to Neu5Ac monomerand Neu5Ac dimer. The same TLC pattern was obtained whenOF-gp was treated with 0.1 M trifluoroacetic acid at 80°C for10 min (Kitazume et al., 1992). No higher oligomer other thandimer was detected in hydrolysates of short incubations underthe same conditions (Figure 2, lanes 4-7). These data show thepresence of either disialyl groups (Neu5Acot2 —> 8Neu5Ac) ormono- and disialyl groups on the peripheral regions of theglycan chain in OF-gp.
Methylation analysis of the sialyl residues of OF-gp
Partially methylated sialitol from the glycopeptide OF-gp wasobtained as described under Materials and methods. On GLCanalysis l,2,4,7,8,9-hexa-O-Me-Neu7V(Ac,Me) and 1,2,4,7,9-penta-0-Me-8-0-Ac-Neu/V(Ac,Me) were identified (Figure 3),confirming the presence of the diSia structure, Neu5Aca2 —»8Neu5Ac, in OF-gp.
Chromatographic separation of diSia and its'H-NMR spectroscopy
To provide direct experimental evidence for the occurrence ofNeu5ca2 —> 8Neu5Ac, Neu5Ac monomer and dimer wereseparately obtained by mild acid hydrolysis of OF-gp followedby chromatography on a DEAE-Sephadex A-25 column. The400 MHz ' H-NMR spectra obtained with both the Neu5Acdimer thus derived from OF-gp and authentic Neu5Acct2 —»8Neu5Ac were completely superimposable (Figure 4).
197
Dow
nloaded from https://academ
ic.oup.com/glycob/article-abstract/7/2/195/598111 by guest on 03 April 2019
Y.Funakoshi et aL
(a) (b)
1 2 3 4 5 6 7 8
I Neu5Ac
I (Neu5Ac)2
i (Neu5Ac)3
i (NeuSAc)4
O
Fig. 2. TLC analysis of mild acid hydrolysates of OF-gp. The TLC platewas developed in a solvent of 1-propanol/aqueous ammonia/water (6:1:2.5,v/v) for 14 h, and visualized by the resorcinol reagent, OF-gp was treatedwith 50 mM sodium acetate buffer, pH 4.8, at 37°C for 0 (lane 1) or 2 days(lane 2) or with 0.1 N trifluoroaceric acid at 80°C for 0 (lane 3), 5(lane 4),10(lane 5), 60(lane 6), and 120 min (lane 7). As a control, a2 -> 8-linkedoligoNeu5Ac prepared by the controlled acid hydrolysis of colominic acid(Kitazume et a!., 1992) and Neu5Ac monomer were also chromatographed(lane 8). O, origin.
Periodate oxidation of OF-gp
OF-gp was subjected to periodate oxidation as described underMaterials and methods. The sialic acid residues at the nonre-ducing termini were oxidized to give the Oj analog of Neu5Acupon reaction while 8-O-substituted Neu5Ac residues were notoxidized. The carbohydrate composition of the products ofperiodate oxidation/NaBH4 reduction of glycan chains wasanalyzed by GLC. OF-gp was shown to give rise to Cq and C?ulosonates in a molar ratio of 2.0:1.0, indicating that OF-gpcontained the terminal Neu5Ac and 8-(9-substituted Neu5Ac ina molar ratio of 2:1. These data were in good accordance withthe presence in OF-gp of the Neu5 Ac residues as the outermostsugar, that is, Neu5Aca2 —» R, and as the diSia form, namelyNeu5Aca2 —> 8Neu5Aca2 —̂ R in an equimolar ratio.
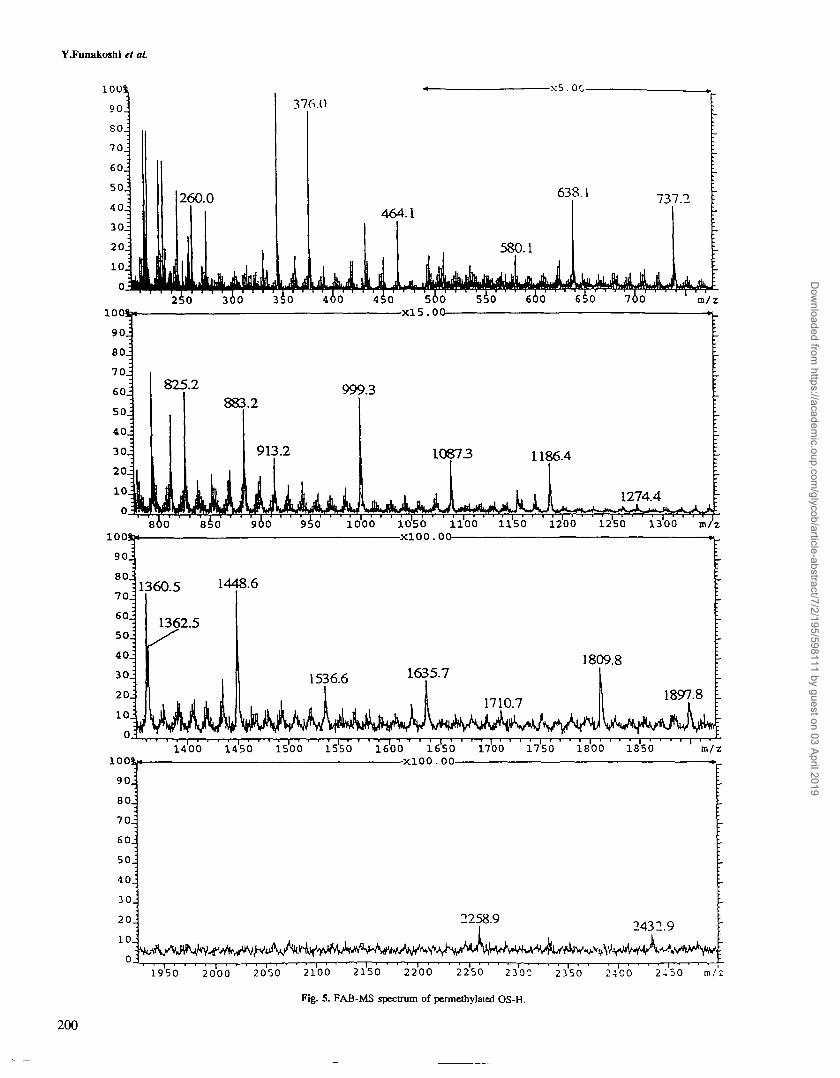
FAB-MS analysis of OS-H and dF-asialo-OF-gp
OS-H and dF-asialo-OF-gp were analyzed by FAB-MS afterpermethylation. Spectra exhibited A-type ions formed bycleavage at each of the HexNAc residues of the antennae (Fig-ure 5). Fragment ions from OS-H are summarized in Table HI.Full support of the presence of diSia groups was obtained fromthe appearance of a prominent fragment ion at m/z 737.2 show-ing the presence of [(Neu5Ac)2]+. Fragment ions at m/z 260.0[GlcNAc]+, at 638.1 [(Fuc)(Gal)(GlcNac)]+, and at m/z999.3 [(Neu5Ac)(Fuc)(Gal)(GlcNAc)]+ indicated the ter-minal sequences of GlcNAc, Lex and sialyl Lex in OF-gp.The appearance of an intense fragment peak at m/z 1360.5[(Neu5Ac)2(Fuc)(Gal)(GlcNAc)]+ was indicative of the pres-ence of a novel disialylated Lex structure, Neu5Acot2-> 8Neu5Aca2 -> 3Ga^ l -> 4 (Fucal -> 3)GlcNAc.
FAB-MS data of permethylated dF-asialo-OF-gp are sum-marized in Table EQ. Evidence in favor of the presence ofpoly-A'-acetyllactosaminyl chains in OF-gp was provided bythe observation of a series of fragment ions assignable to[(Gal)n(GlcNAc)n]
+ (n less than 7).
18 20min
18 20min
Fig. 3. Gas-liquid chromatograms for partially methylated sialitol acetatesfrom (a) a2 -» 8-linlced Neu5Ac tetramer and (b) OF-gp. Peak 1,l,2,4,7,8,9-hexa-O-Me-NeuAf(AcJvle), i.e., terminal Neu5Ac; Peak 2,1,2,4,7,9-penta-O-Me-8-O-Ac-NeiW(Ac,Me), i.e., 8-O-substituted Neu5Ac.Response factor for terminal Neu5Ac was three times higher than that for8-<?-substituted Neu5Ac.
'H-NMR measurement of OS-H
Four hundred megahertz : H-NMR measurement of OS-H wasperformed in D2O at 60°C, and proton assignments are shownin Table IV. All assignments shown here were based on com-parison with the chemical shift values previously reported(Vleigenthart etal, 1983; Finne et aL, 1989; Ishii etal, 1989).Those proton signals which resonated at 5.11 ppm and 4.87ppm were respectively assignable to HI's of the 2,4-substitutedot-Man-4 residue and of the 2,6-substituted ot-Man-4' residueof the tetraantennary core having the 2,4-Manotl —> 3 (2,6-Manal —» 6)Man^l —> structure. Except for the a-anomer offree reducing terminal GlcNAc, the HI protons of all GlcNAc
198
Dow
nloaded from https://academ
ic.oup.com/glycob/article-abstract/7/2/195/598111 by guest on 03 April 2019

A novel disialylated tetraantennary A'-linked glycan chains
(a)
A.ppm
2
(b)
Jkjppm
Fig. 4. 'H-NMR spectra of (a) Neu5Ac dimer derived from OF-gp and (b) the authentic Neu5Aca2 -> 8Neu5Ac. The authentic Neu5Aca2 -> 8Neu5Ac wasprepared by DEAE-Sephadex A-25 chromatography of mild acid hydrolysate of colominic acid as described previously (Kitazume et al, 1992). A large peakal 3.75 ppm in the control sample was due to contaminating Tris base used in Sephadex A-25 chromatography.
and Gal residues resonated between 4.4-4.8 ppm, indicatingthat their anomeric configurations were p. The proton signalswhich resonated at 5.11 ppm, 4.84 ppm, and 1.17 ppmwere respectively assignable to HI, H5, and methyl protonsof Fuc residues in sialyl Lex and/or Lex terminal struc-tures, whose presence was suggested from the FAB-MSand methylation analyses of OF-gp. The existence of theFucal -> 6( -> GlcNAcpi -» 4)GlcNAc structure, which wasalso suggested by methylation analysis (Table FT), was sup-ported by those resonances at 4.80 ppm assignable to the HIproton and at 1.21 ppm for methyl protons (Finne et al., 1989).H3 proton signals of Neu5Ac residues of three different link-age types were observed: (HS^, 2.74 ppm; H3M> 1.77 ppm) forNeu5Aca2 -» 3Gal, (H3~, 2.76 ppm; IB^, 1.74 ppm) forNeu5Aca2 -> 8Neu5Aca2 -> 3Gal, and (TO^,, 2.67 ppm;H3M , 1.73 ppm) for Neu5Aca2 -> 8Neu5Aca2 -» 3Gal(Michon et al., 1987; Ishii et al., 1989). Based on the peakarea ratio, molar proportion of Neu5Acot2 —> 3Gal andNeu5Aca2 -> 8Neu5Aca2 -> 3Gal was determined as 1.2:1.0,
which was compatible with the results of periodate oxidationexperiment (see above).
Isolation and identification of a glycoprotein containingdisialyl groups fi-om rainbow trout ovarian fluidA series of fractionation steps including chromatography onDEAE-Toyopearl 650M (Figure 6a) and repeated gel chroma-tography on Sephacryl S-200 (Figure 6b,c) allowed us to iso-late and identify a glycoprotein fraction, OF-GP, containingNeu5Aca2 -> 8Neu5Ac groups. OF-GP gave a band whosemobility was identical to that of the authentic Neu5Aca2 —»8Neu5Ac on TLC when subjected to mild acid hydrolysis (pH4.8, 37°C, 73 h) (data not shown). The approximate molecularweight of OF-GP was estimated to be 53 K by gel filtrationusing Sephacryl S-200. The SDS-PAGE gave a single majorband of 54 K with a trace of three contaminating faint bands(65 K, 67 K, and 69 K; data not shown).
Sialooligosaccharides, designated OF-GPf, were obtainedby DEAE-Sephadex A-25 chromatography of the products of
199
Dow
nloaded from https://academ
ic.oup.com/glycob/article-abstract/7/2/195/598111 by guest on 03 April 2019
Y.Funakoshl et aL
X5.00-
800 850 900 950 1000 1050 1100 1150 1200 1250 1300 m/z
1360.5 1448.6
100%
9 0J
80_i
TO.
6 0j
50j
40J
3 0j
20j
IOJ
0
1400 1450 1500 1550 1600 1650 1700 1750 1800 1850 m/zX100.00
2258.9 2432.9I - IHJ- .V
1950 2 0 0 0 2050 2100 2 1 5 0 2 2 0 0 2250 230C 2350 2400 2 4 5 0 m/
Fig. 5. FAB-MS spectrum of permethylaled OS-H.
200
Dow
nloaded from https://academ
ic.oup.com/glycob/article-abstract/7/2/195/598111 by guest on 03 April 2019

A novel disialylated tetraantennary /V-linked glycan chains
Table EQ.
OS-H
Fragmentionsobserved(m/z)
260.0376.0464.1580.1638.1737.2825.2883.2913.2999.3
1087.31186.41274.41360.51362.51448.61536.61635.71710.71809.81897.82258.92432.9
Summary of FAB-MS of OS
Assignment
HexNAc*NeuAc+
HexHexNAc+
NeuAcHex*FucHexHexNAc+
NeuAc+*NeuAcHexHexNAc+
FucHexHexNAc/HeXjHexNAc/NeuAcFucHexHexNAc*FucHeXjHexNAcj*NeuACjHexHexNAc+
NeuAcHex2HexNAc2+
NeuACjFucHexHexNAc*Hex3HexNAc3*NeuAcFucHexjHexNAc/FucHex3HexNAc3*NeuAcHexjHexNAc^FuCjHeXjHexNAc/NeuACjFucHeXjHexNA^NeuAcFucHex3HexNAc3
+
NeuAc2FucHex3HexNAc3+
NeuACjFuCjHexjHexNAcj*
-H and dF-asialo-OF-gp
dF-asialo-OF-GP
Fragmentionsobserved(m/z)
260.2464.3638.5709.5913.6
1087.81158.91362.91537.21608.21812.41987.22058.62262.12436.02711.73160.1
Assignment
HexNAc*HexHexNAc+
FucHexHexNAc*HexHexHAc/HeXjHexNAc^FucHex2HexNAc/Hex2HexNAc3
+
Hex3HexNAc3+
FucHex3HexNAc3+
Hex3HexNAc<+
Hex«HexNAc4+
FucHex4HexNAc/Hex4HexNAc,+
Hex5HexNAc5+
FuxHex5HexNAc,+
HeXsHexNAc/Hex7HexNAc/
the hydrazinolysis/A/-acetylation of 54 K OF-GP. Carbohydratecomposition of OF-GPf is shown in Table I, indicating thatOF-GPf had almost the same composition as OF-gp except forthe significantly low Fuc content and the presence of a traceamount of GalNAc. The Q/C^NeuSAc analysis showed thatOF-GPf contained disialosyl groups and monosialosyl groupsin a 1.2:1.0 molar proportion, exactly the same proportion asthat of OF-gp. These results strongly suggest that 54 K OF-GP
Table IV. Chemical shift values of protons in OS-H in D2O at 60°C
Residue Proton Chemical shift (ppm)
a-GlcNAc-1Man-4Man^*'Fuc(al -> 6GlcNAc)
Fuc(al -> 3GlcNAc)
Neu5Aca2 -> 3 Gal
Neu5Aco2 -> 8Neu5Aca2 -> 3C-al
Neu5Aca2 -> 8Neu5Aco2 -> 3 Gal
HIHIHICH3
HIHIH5H5CH3
H 3 «H3,,,H3H3-.
5.175.114.874.891.215.114.844.841.171.772.741.742.761.732.67
Residue numberings of the mannose-core are shown in the structure. Foroverall structure of OS-H, see Structure I.
4GlcNAcpi -> 4GlcNAc-GlcNAc^l^^
is a carrier protein of the OF-gp glycan that was determined bythe detailed structure analysis described above. The differencein content of Fuc residues may likely be ascribed to a differentbatch of fish because in trout polysialoglycoproteins the con-tent of Fuc has been shown to be quite different among troutfish of differing origins (Kitajima et al., 1986).
Discussion
In animal glycoproteins the sialic acids are most often found asthe outermost sugar residues except in the case of the polysialyllinkage (Inoue and Iwasaki, 1978; Finne, 1982; Roth et al.,1992; Troy, 1992). Although oligoSia groups are common con-stituents of gangliosides, they are rare in glycoproteins, havingbeen reported in only a limited number of glycoproteins (Finneet al., 1977; Fukuda et al., 1985, 1987). The presence of a2 ->8-linked diNeu5Ac group in glycoproteins was first suggestedby Finne and co-workers as early as in 1977. Subsequently, theprecise structures of ot2 —> 9- and a2 -» 8 linked diNeu5Acgroups were identified in collaborating work by Dell andFukuda in PA1 human embryonal carcinoma cells and humanerythrocyte glycophorins, respectively (Fukuda et al., 1984a,1985, 1987). The composition of a glycopeptide (OF-gp) de-rived from ovarian fluid glycoprotein of rainbow trout sug-gested the possible presence of lower oligoSia residues (TableI). When this was considered in the context of our currentunderstanding of biosynthesis of polySia chains (Kitazume etal., 1994), it was of special interest to determine if OF-gpmight contain a biosynthetically intermediate form of polySiagroups.
Our proposed biosynthetic pathway summarized in the In-troduction suggests how polysialylation of 0-linked glycanchains in rainbow trout PSGP occurs. This pathway is based onour findings that PSGP isolated at an early stage of develop-ment from the immature oocytes contained a high proportion ofnonelongated, mostly a2 —» 8-linked diSia residues that weresubsequently converted to polySia-glycan chain in a develop-mentally regulated manner (Kitazume et al., 1994). The factthat diSia appears to be a precursor of <9-linked polySia chains,and the finding of the identical diSia epitope in the presentstudies leads us to postulate the following pathway for synthe-sis of the polySia chains on AMinked glycans.[1] a2 —» 3-ST-catalyzed reaction:
4GlcNAc31 -+ + CMP-Sia ->> 3Gaipi -> 4GlcNAcpi -> + CMPSiaa2
[2] a2 -» 8-ST (initiase)-catalyzed reaction:
Siaa2 -> 3Gaipi -> 4GlcNAc31 -> + CMP-Sia ->Siaa2 -> 8Siaa2 -> 3Gaipi -» 4GlcNAc£l -> + CMP
[3] a2 —> 8-polyST (polvmerase)-catalyzed reaction:
Siaa2 -> 8Siaa2 -> 3Ga^l -> 4GlcNAcpinCMP-Sia -> (Siaot2 -^ 8)nSiaa2 -> 8Siaa2-^ 4GlcNAcpi -> + nCMP.
3Gaipi
jManaM
By analogy with synthesis of O-linked polySia chains, forma-tion of the AMinked polySia chains, if they occur in rainbowtrout, would also likely require at least three different sialyl-transferases, viz. an ot2 —> 3-ST, an a2 —> 8-ST, and an a2 —»8-polyST. Recently, Kojima et al. (1995a) reported the cloningand characterization of a novel sialyltransferase in mouse
201
Dow
nloaded from https://academ
ic.oup.com/glycob/article-abstract/7/2/195/598111 by guest on 03 April 2019

Y.Funakoshi el al
1M IS*
Fraction number200
2M 4M t«0
Elution volume (ml)2M 400 WHJ
Elution volume (ml)
Fig. 6. Isolation of diSia-containing glycoprotein OF-GP from rainbow trout ovarian fluid, (a) DEAE-Toyopearl 650M chromatogram of the ovarian fluid and(b) First Sephacryl S-200 chromatogram of the pooled fraction under the bar shown in (a). Fractions under the bar in (b) were shown to contain the diSiastructure, and were pooled, and subjected to repeated chromatography on S-200. (c) Third chromatogram of the diSia-containing fraction on Sephacryl S-200.Elution profiles were monitored by the absorbance at 280 nm (open circle) and for Neu5Ac by the resorcinol method (A580, solid circle). The broken line in(a) represents the gradient of NaCl.
which is involved in the transfer of Sia from CMP-Sia to the al—» 3-linked sialoside residues but not to nonreducing terminalGal residues. This enzyme would be analogous to the "initi-ase" in the above biosynthetic pathway for poly Sia chain syn-thesis. A possible candidate for the ST "initiase" was purified2900-fold from human neuroblastoma CHP-134 cells(Stoykova and Glick, 1995). This enzyme differs in substratespecificity from the cloned and expressed polySTs reported(Eckhardt et al., 1995; Kojima et al., 1995a,b; Yoshida et al,1995; Nakayama and Fukuda, 1996).
Structure 1 shows the most probable glycan structure for OF-gp, based on the experimental results from this structural study.While the a2 —» 8-linked diSia moiety is a definite chemicalentity of OF-gp, heterogeneity in the size and in the variety ofcarbohydrate chains exist, including: (1) monoSia and diSia butno higher oligoSia chains; (2) different numbers of N-acetyl-lactosamine units; and (3) Le* and I antigenic determinants in theperipheral region of the glycan chain although the nature of thetetraantennary structure is uniform. We have ascertained that OF-gp is formed from its original glycoprotein, OF-GP, by Pronasedigestion. No higher oligo- or polysialic acid chains were found toexist in the 54K glycoprotein isolated from the ovarian fluid whilepolymerase activity that is involved in biosynthesis of oligo/polyNeu5Gc chains on 0-linked glycan chains of polysialogly-coproteins was detected within the ovaries.
It will be important now to produce monoclonal antibodiesto the diSia glycotope for our future structural and functionalstudies of disialylated glycoproteins. Such antibodies will alsobe useful to search for the presence of an intracellular pool ofdiNeu5Ac-gps that may resemble glycan chains of mature N-CAM that we have recently found in brains of adult chickenand pigs (unpublished results). Thus, disialylglycan chain-containing glycoproteins may not be confined to a limitednumber of glycoproteins, but rather may be found more com-monly than presently recognized in animal cells and tissues aswas first suggested by Fukuda et al. (1987).
Materials and methodsMaterials
The ovarian fluid of rainbow trout was supplied by courtesy of the GunmaPrefectural Fisheries Station at Kawaba and the Okutama Fish Farm, Depart-
ment of Fishery, Tokyo Metropolitan Government, and stored at -30°C untiluse. Actinase E (Pronase) was obtained from JCaken Co. (Japan). Arthrobacterweafaciens exosialidase and Escherichia frcundii cndo-(3-galactosidase werepurchased from Nacalai Co. and Seikagaku Kogyo (Japan), respectively.
Chemical analysis
Sialic acid was quantitated or monitored by the TBA method (Aminoff, 1962;Uchida et al., 1977) and the resorcinol method (Svennerholm, 1963). Hexosewas quantitated by the phenol-sulfuric acid method (Dubois et al., 1956). Themolar ratios of carbohydrate were determined by GLC for oligosaccharidesand glycopeptides (Nomoto et al., 1982). Analysis of amino acids, GlcNAc,and GalNAc was carried out after hydrolysis in 6 M HC1 at 110°C for 24 hunder N2 and precolumn derivatization with phenylisothiocyanate (Taguchi etal., 1993). Permethylarion analysis for oligosaccharides and glycopeptides wascarried out as described previously (Anumula and Taylor, 1992; Taguchi et al.,1993). Linkage analysis of mono- and oligosialyl groups was carried outaccording to the method of Inoue et al. (1982). The partially methylatedsialitols were quantitated by GLC on a capillary column (CBJ5, 0.32 mm x 30m, Shimadzu) at 190-260°C at 4°C/min.
Isolation of Pronase-gtycopeptide having N-linked glycan
Ovarian fluid (3.5 1) was concentrated by evaporation under reduced pressureand dialyzed against distilled water at 4°C for 2 days. The dialysate waslyophilized and delipidated as described previously (Nishino et al., 1996). Thelyophilized powder (32 g) was delipidated by stirring first with 20 volumes(v/w) of chloroform/methanol (2:1, v/v) at room temperature for 2 h, filtered,and then delipidated with the same volume of chloroform/methanol (1:2, v/v)followed by filtration. The delipidated ovarian fluid (25 g) thus obtained wasdigested with 0.5 g of Pronase in 0.2 M Tris-HCl (pH 8.0) and 0.01 M CaCl2
at 60°C for 24 h (Tezuka et al., 1994). The mixture was centrifuged andaqueous phase was extracted with 90% phenol at room temperature for 1 h.The mixture was centrifuged at 5000 rpm (3500 x g) for 15 min and separatedinto three fractions, upper aqueous layer, middle cream layer, and lower layer.To the middle layer was added 100 ml of 0.01 M Tris-HCl (pH 8.0), stirred atroom temperature for 1 h, and centrifuged at 5000 rpm for 15 min. Theaqueous layer was collected and added to the upper layer. The combinedaqueous layer was dialyzed against distilled water at 4°C for 2 days. Thedialysate was applied to a column (2.8 x 67 cm) of DEAE-Toyopearl 650M(CT form; equilibrated with 0.01 M Tris-HCl, pH 8.0). The column was elutedby a linear gradient of NaCl (0-0.4 M, 4 1) in the same buffer, and 20 mlfractions were collected. The elution profile was monitored by the absorbanceat 230 nm, the thiobarbituric acid method (Aminoff, 1961; Uchida et al., 1977;Kitajima et al., 1992) and the resorcinol method (Svennerholm, 1963). Thefraction, which was sialic acid-positive, was collected (see Figure la) and thenapplied to a Sephacryl S-200 column (2.2 x 164 cm; equilibrated and elutedwith 0.1 M NaCl in 5 mM Tris-HQ, pH 8.0). The elution profile was moni-tored by the absorbance at 230 nm and the thiobarbituric acid method. The
202
Dow
nloaded from https://academ
ic.oup.com/glycob/article-abstract/7/2/195/598111 by guest on 03 April 2019

A novel disialylated tetraantennary A'-linked glycan chains
R
tFuctxl
GlcNAcpi'
R
VR
*Fucccl-/
tFucal-*'
R
GlcNAcp
4jGlcNAcpl.
jGalpl-—-
±Fucal -^ J
R7>^4
±Fucal -^ 3
4GlcNAcpi
4GlcNAcpi
4GlcNAcpi
4GlcNAcpi
±Fucal
\
R3, R4, Rs, R6, R7 = Neu5Aca2 8Neu5Aca2
Neu5Aca2
-*-3Galpl-
-^3Galpl-
Galpl-
H-
31%
31%
21%
17%
Structure 1. Proposed structure for glycan chain of OF-gp. The distinct features of the glycan chain are: (1) exclusive occurrence of a tetraantennarystructure; (2) the presence of oligo/poly-W-acetyllactosamine units up to seven repeats on the antennae; (3) occurrence of on average three branched Galresidues (3.1 residues of 3,4-substituted Gal/glycan chain, Table H), suggestive of the presence of I antigen; (4) exclusive occurrence of the disialyl structureand absence of oligoSia higher than dimer; (5) termination of on average seven branch arms present in the OF-gp glycan chains by GlcNAc (1.2 mol),-* 4GlcNAc and/or Le* (2.2 mol), or disialylated Lex (2.2 mol).
sialic acid-rich fraction (see Figure lb) was collected and applied to a Seph-adex G-50 column (1.6 x 100 cm; equilibrated with 0.1 M NaCl). The glyco-peptide fraction thus obtained was desalted on a column of Sephadex G-25 (1.3x 100 cm; equilibrated with 5% ethanol). This fraction was designated asOF-gp.
Preparation of asialo-OF-gp and partially defucosylated OF-gp
OF-gp (27 mg as Neu5Ac) was digested with 250 mu^ml of Arthrobacterureafaciens exosialidase in 1.5 ml of 50 mM sodium acetate buffer, pH 4.8, at37°C. After 24 h incubation, an additional 75 pJ of 2.5 (i/ml exosialidase wasadded three times at 6 h intervals and incubated further at 37°C. The resultingasialo-glycopeptide was separated by passage of the enzyme digests through aSephadex G-50 column (1.6 x 100 cm; equilibrated with 0.1 M NaCl), desaltedby Sephadex G-25 column, and designated as asialo-OF-gp.
To remove a majority of the Fuc residues selectively, asialo-OF-gp wastreated with 0.05 M trifluoroacetic acid at 80°C for 2 h, and the defucosylatedasialo-OF-gp was separated from Fuc by Sephadex G-50 column chromatog-raphy followed by desalting on a Sephadex G-25 column. The material thusobtained was designated dF-asialo-OF-gp.
Isolation of intact proteinfs) with disialyl groups
Ovarian fluid (1000 ml) was dialyzed against water at 4°C foT 16 h. Thedialysate was applied to a column of DEAE-Toyopearl 650M and eluted by3000 ml of 10 mM Tris-HCl buffer (pH 8.0) with a linear gradient of NaCl(0-0.4 M). Fifteen milliliter fractions were collected and monitored by theabsorbance at 280 and 230 run, by the resorcinol method for Neu5Ac (Sven-nerholm, 1963), by the TBA method for KDN (Kitajima et al., 1992), and by
the phenol-sulfuric acid method for neutral hexose (Dubois et al., 1956). Sialicacid-containing fractions were tested for the presence of disialic acid by themild acid hydrolysis, as described below. The sialic acid-positive fractionswere pooled, concentrated by limited filtration on YM10 (Amicon), and ap-plied to a Sephacryl S-200 column (2.2 x 164 cm; equilibrated with 0.1 MNaCl in 0.01 M Tris-HCl, pH 8.0). The peak fractions which gave disialic acidon mild acid hydrolysis were pooled and rechromatographed on the sameSephacryl S-200 column three times until a peak of the glycoprotein wassymmetrical. Each fraction was also tested for purity by SDS-PAGE (10%acrylamide; Laemmli, 1970). The purified glycoprotein fraction was desig-nated as OF-GP.
Preparation of N-linked glycan chain from OF-gp and OF-GP
OF-gp (1 mg as hexose) was treated with anhydrous hydrazine in a commercialhydrazinolysis apparatus, Hydraclub (HONEN Co, Japan), at 110°C for 2 h.The reaction mixture was W-acetylated with saturated NaHCOj/acetic anhy-dride as previously described (Tezuka et al., 1994) and then applied to aSephadex G-50 column (1.6 x 100 cm; equilibrated with 0.1 M NaCl). Freeglycan was collected and designated OS-H.
Free glycan(s) which was obtained from OF-GP (60 u,g as NeuSAc) wasalso prepared by the hydrazinolysis/W-acetylation, as described above. ThediSia-containing free glycan was purified by DEAE-Sephadex A-25 columnchromatography: C\~, 1 x 1 cm; eluted with 0.2 M NaCl after washing withwater. The 0.2 M fraction was desalted by Sephadex G-25 column and des-ignated OF-GPf.
Hydrazinolysis-nitrous acid deamination
A freeze-dried sample of dF-asialo-OF-gp (0.5 mg as Gal) was reacted with 2ml of anhydrous hydrazine in the presence of hydrazine sulfate in a commer-
203
Dow
nloaded from https://academ
ic.oup.com/glycob/article-abstract/7/2/195/598111 by guest on 03 April 2019

Y.Funakoshi et aL
cial apparatus, Hydraclub (HONEN Co., Japan), at 110°C for 24 h (Taguchi etal., 1993). After cooling to room temperature, the reaction mixture was driedfor 16 h in a vacuum desiccator over concentrated HjSO,,. Twenty milligramsof sodium nitrite and 2 ml of 0.5 M acetic acid were then added. After reactionat room temperature for 2 h, an additional 10 mg of NaNO2 was added, anddeamination was continued for 16 h. Ethylamine was then added to destroyexcess nitrite, and the solution was treated with 10 mg of NaBH4 at roomtemperature for 2 h. The pH was adjusted to 5-6 with 2 M acetic acid, and thereaction products were then applied to a Bio-Gel P-2 column (1.3 x 73 cm;equilibrated and eluted with 50 mM acetic acid). The effluent was monitoredfor neutral hexose, and the phenol-sulfuric acid-positive fractions were col-lected and analyzed.
Thin-layer chromatographic (TLC) analysis of mild acid hydrolysales ofsialoglycopeptides and glycoproteins
OligoSia-containing glycopeptides or glycoproleins were hydrolyzed in 50mM sodium acetate buffer, pH 4.8, at 37°C for 2 or 3 days, or in 0.1 Mtnfluoroacetic acid at 80°C for 0, 5, 10, 20, 60, and 120 min, and analyzed byTLC (Kitazume et al., 1992). TLC was carried out on Silica Gel 60 plate(0.2-mm-thick silica gel coated plastic plate; Merck) in l-propanol/NH4OH/water (6:1:2.5, v/v), developed for 11-14 h at room temperature. Neutral sugarbands were visualized after spraying with 1% orcinol/50% sulfuric acid andheating at 120°C for 20 min. Neu5Ac-containing bands were visualized byspraying with the resorcinol reagent, followed by hearing at 80°C for 30 min(Kitazume et al., 1992).
As a routine assay for diSia in fractions of the column chromatographydescribed above, an aliquot of each sample was partially hydrolyzed in 0.1 Mtrifluoroacetic acid at 80°C for 10 min followed by the TLC analysis.
Isolation of diSia released from OF-gp and its characterization by'H-NMR measurement
The mild acid hydrolysate of OF-gp obtained after treatment with 50 mMsodium acetate buffer, pH 4.8, at 37°C for 3 days was subjected to DEAE-Sephadex A-25 chromatography to obtain mono- and oligosialic acids as de-scribed by Kitazume et al. (1992). The diSia-containing fraction was charac-terized by ' H-NMR (see below) after desalting on a Sephadex G-25 column.Authentic Neu5Ac dimer was prepared as previously described (Nomoto et al.,1982; Kitazume et al., 1992), and also characterized by NMR spectroscopy.
Periodate oxidation of OF-gp
To 40 jil of OF-gp (40 p.g as Neu5Ac) in water was added 10 jil of 0.2 Msodium acetate buffer, pH 5.5, and 4 uJ of 0.25 M NaI04. The sample wasincubated at 0°C for 3 h in the dark. Under these conditions, nonreducingterminal Neu5Ac residues were oxidized to the C, analogue of Neu5Ac, whilethe internal al —» 8-linked Neu5Ac (C,) residues remained intact (McLean etal., 1971). Water (736 nJ) and 200 uJ of 0.5 M sodium borate buffer (pH8.O)/D.5 M NaBH4 were then added and the mixture was incubated at 0°C for12 h. The reaction mixture was neutralized with 1 M acetic acid and appliedto a Sephadex G-25 column for desalting. The periodate-oxidized sugars werethen analyzed by GLC (Nomoto et al., 1982) for the ratio of C7/G, Neu5Acderivatives. As the standard to quantify the C, analog of Neu5Ac and intactNeu5Ac (C,) residue formed in oligosaccharides after the oxidation/reduction,L-hyosophorin from Oryiias latipes (Taguchi et al., 1994) was analyzed byGLC before and after the oxidation/reduction. The Neu5Ac residues present inthe L-hyosophorin were converted quantitatively to C7 analog of Neu5Ac.
'H-NMR spectroscopy
'H-NMR measurements of oligosaccharides and glycopeptides were carriedout in D2O on a 400 MHz 'H-NMR instrument as described previously (Ishiiet al., 1989). Sample solutions were exchanged by D2O as described previ-ously (Ishii et al., 1989).
FAB-MS spectrometry
OS-H and dF-asialo-OF-gp were permethylated (see above) and analyzed byFAB-MS as described previously (Dell et al., 1994).
AcknowledgmentsThis research was supported in part by Grants-in-Aids for International Sci-entific Research Program: Joint Research 04044055 (to YJ.) and for GeneralScientific Research 06680576 (to Y.I.) and 07680658 (to S.L) from the Min-
istry of Education, Science, and Culture of Japan when Y.I. and S.I. were at theUniversity of Tokyo and Showa University, respectively. This work was alsosupported by an MRC Programme grant, a BBSRC grant, and a WellcomeTrust Grant 030826 (to H.R.M. and A.D.). We thank Ms. Mild Nagasao,Showa University, for obtaining the 400 MHz 'H-NMR spectra.
Abbreviationsasialo-OF-gp, a glycopeptide fraction obtained by digestion of OF-gp withexosialidase; CMP-Sia, cytidine 5'-sialic acid phosphate; diSia, al -» 8-linkeddisialic acid(Neu5Ac); dF-asialo-OF-gp, a partially defucosylated glycopep-tide obtained by treatment of asialo-OF-gp with 0.05 M trifluoroacetic acid at80°C for 2 h; FAB-MS, fast atom bombardment-mass spectrometry; GLC,gas-liquid chromatography; KDN, 2-keto-3-deoxy-D-g/ycero-D-ga/ac»o-nononic acid; N-CAM, neural cell adhesion molecule; NeuSAc, N-acetylneuraminic acid; NMR, nuclear magnetic resonance; OF-gp, a diSia-containing sialoglycopeptide fraction that was prepared and purified from Pro-nase digest of the ovarian fluid of rainbow trout; OF-GP, a diSia-containingsialoglycoprotein isolated from the ovarian fluid of rainbow trout; oligoSia,oligosialic acid; OS-H, free glycan chain released from OF-gp by hydrazinoly-sis/re-N-acetylation; polySia, polysialic acid; TBA, thiobarbituric acid; TLC,thin-layer chromatography.
References
Aminoff.D. (1961) Methods for the quantitative estimation of N-acetylneuraminic acid and their application to hydrolysatcs of sialomucoids.Biochem J., 81, 384-392.
AnumulajCR. and Taylor,P.B. (1992) A comprehensive procedure for prepa-ration of partially methylated alditol acetates from glycoprotein carbohy-drates. Anal. Biochem., 203, 101-108.
Dell,A., Reason^., Khoojt-H., Panico.M., McDowell,R.A. and Morris,H.R.(1994) Mass spectrometry of carbohydrate-containing biopolymers. Meth-ods Enzyml., 230, 108-132.
Dubois,M., GillesJCA., HamiltonJK., Rebers,P.A. and Smithy. (1956) Col-orimetric method for determination of sugars and related substances. Anal.Chem., 28, 350-356.
Eckhardt,M., Waibeljl., Hartmann.U., Mulenhoff,M., Bethe,A., Frosch,M.and Gerardy-Schahn,R. (1995) Biosynthesis of polysialic acid in mammalsand the regulation of its expression. Glycoconjugate J , 12, 508.
FinneJ. (1982) Occurrence of unique polysialosyl carbohydrate units in gly-coproteins of developing brain. / . Biol. Chem., 257, 11966-11970.
FinneJ., Krusius.T., Rauvala.H. and HemminkiJC. (1977) The disialosyl groupof glycoproteins. Occurrence in different tissues and cellular membranes.Eur. J. Biochem., TJ, 319-323.
FinneJ., Breimer,M.R, Hansson.G.C, Karlsson,K.-A., Leffler.H., Vliegent-hartJ.F.G. and van Halbeekji. (1989) Novel polyfucosylated N-linked gly-copeptides with blood group A, H, X, and Y determinants from human smallintestinal epithelial cells. J. Biol. Chem , 264, 5720-5735.
Fukuda,M., Dell,A. and Fukuda,M.N. (1984a) Structure of fetal lactos-aminoglycan. The carbohydrate moiety of Band 3 isolated from humanumbilical cord erythrocytes. J. Biol. Chem., 259, 4782-4791.
Fukuda,M., Dell A , OatesJ.E. and FukudaJvI.N. (1984b) Structure of branchedlactosaminoglycan, the carbohydrate moiety of Band 3 isolated from adulthuman erythrocytes. / . Biol. Chem., 259, 8260-8273.
Fukuda,M., Spooncer£., OatesJ.E., Dell,A. and KlockJ.C. (1984c) Structureof sialylated fucosyl lactosaminoglycan isolated from human granulocyte. J.Biol. Chem., 259, 10925-10935.
FukudaJvi.N., Dell,A., OatesJ.E. and Fukudajvl. (1985) Embryonal lactos-aminoglycan. The structure of branched lactosaminoglycans with novel di-sialosyl (sialyla 2 -» 9sialyl) terminals isolated from PA1 human embryonalcarcinoma cells. / . Biol. Chem., 260, 6623-6631.
Fukuda.M., Lauffenburger,M., Sasaki,H., Rogers,M.E. and Dell,A. (1987)Structures of novel sialylated O-linked oligosaccharides isolated from hu-man erythrocyte glycoproteins. J. Biol. Chem., 262, 11952-11957.
InoueJS. and Iwasaki,M. (1978) Isolation of a novel glycoprotein from the eggsof rainbow trout: Occurrence of disialosyl groups on all carbohydrate chains.Biochem. Biophys. Res. Commun., 83, 1018-1023.
Inoue.S., Matsumura,G. and Inoue,Y. (1982) Stability of N-acyl groups inmethylated a2 - • 8-linked oligosialosyl chains toward methanolysis. Analy-sis by chemical ionizanon-mass spectrometry. Anal. Biochem., 125, 118-124.
lnoue.S., Kanamori^A., KitajimaJC. and Inoue.Y. (1988) KDN-glycoprotcin: anovel deaminated neuraminic acid-rich glycoprotein isolated from vitelline
204
Dow
nloaded from https://academ
ic.oup.com/glycob/article-abstract/7/2/195/598111 by guest on 03 April 2019

A novel disialylated tetraantennary JV-linked glycan chains
envelope of rainbow trout eggs. Biochem. Biophys. Res. Commun., 153,172-176.
IshiiJC, Iwasaki,M., Inoue.S., Kenny,P.T.M., Komurajt and Inoue,Y. (1989)Free sialooligosaccharides found in the unfertilized eggs of a freshwatertrout, Plecoglossus alrivelis. A large storage pool of complex-type bi-, tri-,and tetraantennary sialooligosaccharides. / . Biol. Chem., 264, 1623-1630.
James.W.M. and Agnew.W.S. (1987) Multiple oligosaccharide chains in thevoltage-sensitive Na channel from Electrophorus electricus. Evidence foro-2,8-linked polysialic acid. Biochem. Biophys. Res. Commun., 148, 817-826.
Kanamori,A., KitajimaJC., Inoue.S. and Inoue.Y. (1989) Isolation and charac-terization of deaminated neuramiriic acid-rich glycoprotein (KDN-gp-OF) uithe ovarian fluid of rainbow trout {Salmo gairdneri). Biochem. Biophys Res.Commun., 164, 744-749.
KanamorirA., Inoue.S., Iwasakijvl., KitajimaJC, Kawai.G., Yokoyama,S. andInoue.Y. (1990) Deaminated neuraminic acid-rich glycoprotein of rainbowtrout egg vitelline envelope. J Biol. Chem., 265, 21811-21819.
KitajimaJC, Inoue.Y. and Inoue.S. (1986) Polysialoglycoproteins of Salmom-dae fish eggs. Complete structure of 200-kDa polysialoglycoprotein fromthe unfertilized eggs of rainbow trout (Salmo gairdneri) J. Biol. Chem., 261,5256-5269.
KitajimaJC., Inoue.S., Kitazume.S. and Inoue.Y. (1992) Analytical methodsfor identifying and quantitating deaminated sialic acid (2-keto-3-deoxy-D-glycero-D-galacto-nononic acid) and a2 —> 8-Unked poly(oligo)nonulos-onate residues in glycoconjugates. Anal. Biochem., 205, 244—250.
Kitazume.S., KitajimaJC., Inoue.S. and Inoue.Y. (1992) Detection, isolation,and characterization of oligo/poly(sialic acid) and oligo/poly(deami-noneuraminic acid) units in glycoconjugates. Anal. Biochem., 202, 25-34.
Kitazume.S., KitajimaJC., Inoue.S., Inoue.Y. and TroyJ'.A,, U (1994) Devel-opmental expression of trout egg polysialoglycoproteins and the prerequisitea2,6-, and a2,8-sialyl and o2,8-polysialyltransferase activities required fortheir synthesis during oogenesis. / . Biol Chem., 269, 10330-10340.
KojimaJ^., Yoshida,Y., Kurosawa^., Lee,Y.-C. and Tsuji.S. (1995a) Enzy-matic activity of a developmentally regulated member of the sialyltransfer-ase family (STX): evidence for ot2,8-sialyltransferase activity toward N-linked oligosaccharides. FEBS Lett., 360, 1-4.
Kojima>J., Yoshida,Y. and Tsuji,S. (1995b) A developmentally regulatedmember of the sialyltransferase family (ST8Sia n, STX) is a polysialic acidsynthetase. FEBS Lett., 373, 119-122.
Laemmli.U.K. (1970) Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature, 227, 680-685.
McLeanJ^.L., Suttajitjd., BeidlerJ. and WinzlerJU. (1971) W-Acetylneur-aminic acid analogues. I. Preparation of the 8-carbon and 7 carbon com-pounds. / . Biol. Chem., 246, 803-809.
MichonJ3., BrissonJ.-R. and JenningsJU. (1987) Conformational differencesbetween linear (ot2 —> 8)-linked homooligosaccharides and the epitope ofthe group B meningococcal polysaccharide. Biochemistry, 26, 8399-8405.
Nakayamaj. and FukudaJVL (1996) A human polysialyltransferase directs invitro synthesis of polysialic acid. J. Biol. Chem., 271, 1829-1832.
Nishino.S., Kuroyanagiji., Terada,T., Inoue.S., Inoue.Y., TroyJ^.A., D andKitajima, K. (1996) Induction, localization, and purification of a novel si-alidase, deaminoneuraminidase (KDNase), from Sphingobacterium multivo-rum. J. Biol. Chem., 271, 2909-2913.
Nomoto.H., Iwasaki.M., Endo.T., Inoue.S., Inoue.Y. and Matsumura,G. (1982)Structures of carbohydrate units isolated from trout egg polysialoglycopro-teins: short-cored units with oligosialosyl groups. Arch. Biochem Biophys.,218, 335-341.
RothJ., Kempf,A., Reuter.G., Schauerjt. and Gehring,WJ. (1992) Occurrenceof sialic acids in Drosophila melanogaster. Science, 256, 673—675.
StoykovaJ^.1., and Glickjvl. (1995) Purification of an a-2,8-sialyltransferase,a potential initiating enzyme for the biosynthesis of polysialic acid in humanneuroblastoma cells. Biochem. Biophys. Res. Commun., 217, 777-783.
SvennerholmJL. (1963) Sialic acids and derivatives: estimation by the ion-exchange method. Methods Enzymol., 6, 459—462.
Taguchi.T., Seko,A., KitajimaJC, Inoue.S., Iwamatsu.T., KhooJC.-H., Mor-risJi.R., DellA and Inoue.Y. (1993) Structural studies of a novel type oftetraantennary sialoglycan unit in a carbohydrate-rich glycopeptide isolatedfrom the fertilized eggs of Indian medaka fish, Oryzias melastigma. J. Biol.Chem., 268, 2353-2362.
Taguchi.T., Seko.A., KitajimaJC., Muto.Y., Inoue.S., Khoo,K.-H., Mor-ris.H.R., DeIl,A. and Inoue.Y. (1994) Structural studies of a novel type ofpentaantennary large glycan unit in the fertilization-associated carbohy-drate-rich glycopeptide isolated from the fertilized eggs of Oryzias latipes.J. Biol. Chem., 269, 8762-8771.
Tezuka,T., Taguchi.T., KanamorirA., Muto.Y., KitajimaJC, Inoue.Y. and In-oue.S. (1994) Identification and structural determination of the KDN-
containing N-linked glycan chains consisting of bi-, triantennary complex-type units of KDN-glycoprotein previously isolated from rainbow trout vi-telline envelopes. Biochemistry, 33, 6495-6502.
TroyJ.A., D (1992) Polysialylation: from bacteria to brains. Gtycobiolgy, 2,5-23.
Uchida,Y., Tsukada,Y. and Sugimori.T. (1977) Distribution of neuraminidasein Arthrobacter and its purification by affinity chromatography / . Biochem(Tokyo), 82, 1425-1433.
VliegentharU-F.G., DorlandJ- and van HalbeekJI. (1983) High-resolution,'H-nuclear magnetic resonance spectroscopy as a tool in the structuralanalysis of carbohydrates related to glycoproteins. Adv. Carbhydr. Chem.Biochem., 41, 209-374.
Yoshida,Y., KojimaJ")., KurosawaJM., Hamamoto.T. and Tsuji,S. (1995) Mo-lecular cloning of Sia 2,3Gaipi,4GlcNAc a2,8-sialytransferase from mousebrain. / . Biol. Chem., 270, 14628-14633.
Received May 20, 1996; revised on August 6, 1996; accepted on August 11,1996.
205
Dow
nloaded from https://academ
ic.oup.com/glycob/article-abstract/7/2/195/598111 by guest on 03 April 2019


















