BACTERIAL MORPHOLOGY: PROCARYOTIC AND EUCARYOTIC CELLS Chapter 4

Jaan MännikSaturday Morning Physics Lecture Mar. 11, 2017
VBm
VBmmmmTkF sc
Btot�
��
�� ln
Physics and Nanotechnology to Study Bacterial Cells

Impact of physics on studies of living systems
2. Application of physics based theories to explain lifeprocesses.
1. Application of physics based methods and techniquesto experiments with living systems

Bacteria – the unseen majority• Majority of Earth’s biomass are bacteria and archea. They
are one of the main determinants how biospherefunctions
• There are estimated 10 times more bacterial cells inhuman body than our own cells
• Recognizing bacteria and other microorganisms asdisease agents have lead human life span to increasetwofold over past 100 years. This gain may not bepermanent.

Standard microbiology toolbox
The standard microbiology tools are not suitable to follow:
An agar plate Cell culture tubes
2. Molecular process in individual cells in real time.1. How bacteria behave in complex environments

Lab-on-a-chip based tools
• Well controlled environment for the cells which physical andchemical characteristics can be controlled and manipulated
• Compatible with high resolution microscopy of cells(including super-resolution imaging, SEM)

Advantages of lab-on-a-chip platform
2 Pm
• In situ biochemical analysis of cells
• Automation
F. K. Balagadde et al Science 309 (2005) 137.
J. W. Hong et al Nat. Biotech. 21 (2003) 1179
Device to extract bacterial DNA

Applications of lab-on-a-chip in studies of cells
S. Park et al. Science 301 (2003) 188
N. Q. Balaban et al. Science 305 (2004) 1622
Molecules Cells PopulationsOrganisms
Y. Mercy et al. PNAS 104 (2007) 11889
Sequencing Antibiotic resistancestudies Ecology,
evolution
Bacteria and tissue interactions
D.Huh et al. Science 328 (2010) 1622

Understanding how bacterial cells move in small pores using bio-mimetic lab-on-a-chip devices

Bacterial movement in channels and pores
• Most bacteria in different environments live in pores 10 Pmand smaller

Can’t do it in patient mouth …

Experimental setting
• Monitor bacterial movement through channels (constrictions) usingfluorescent microscopy
100 Pm
channel
chamber
• Fabricate on the single chip, large number of differently sized andshaped channels.

Microfabrication cleanroom

Microchip fabrication
Use electron beam or photolithography to write pattern of channels
Develop resist
Reactive ion etch Si wafer
Lift-off resist
Close channels
Repeat the process for different channel heights
1
2
3
4
PMMA
Si
5Si
PDMS coated glass coverslip
electron beam
channels
Drill access holes

Channels
1 µm
• On single chip, channel width is made to vary between ≈300 nm to 5 Pm to using a RIE cryoetch process
1 µm 1 µm
D
Side view Bacterium

Fluorescence
Most materials/objects do not fluoresce when excited with blue light (many materials fluoresce when exited with UV light)
Exception – jellyfishAequorea victoria
UV excitation
� Excitation with blue light allows to selectively observe engineered molecules such as GFP and have very little background from other molecules.

Fluorescent proteins
• Insert genetic code for GFP molecule into bacterial genome or in plasmid (short circular DNA that many bacteria carry). Bacteria will synthesize the protein.
GFP (Green Fluoresccent Protein)
• Shine blue light on bacteria that express GFP. They will shine green light back. There is little background at green wavelength region.
Blue light
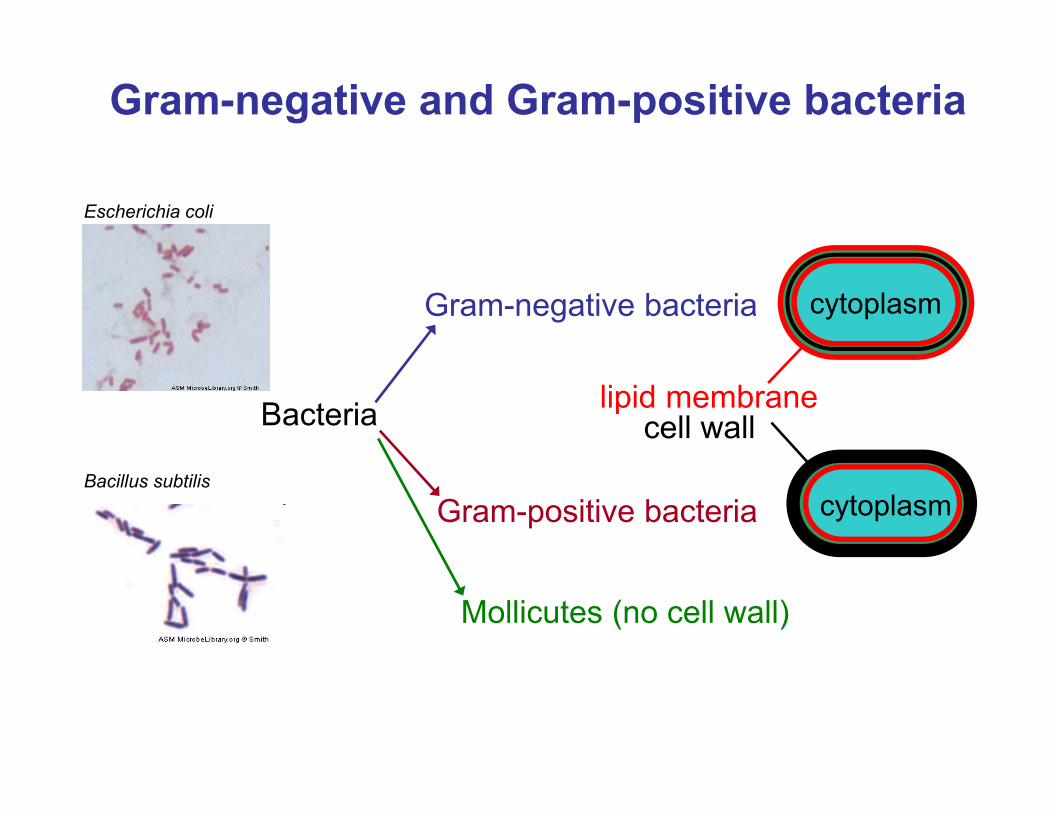
Gram-negative and Gram-positive bacteria
Bacteria
Gram-negative bacteria
Gram-positive bacteria
cell walllipid membrane
cytoplasm
cytoplasm
Mollicutes (no cell wall)
Escherichia coli
Bacillus subtilis

Two bacterial species studied
E. coli RP437
1 µm 1 µm
D
Escherichia coli Gram-negative bacterium
Bacillus subtilisGram-positive bacterium
1 µm
D
• Superfically looking the two bacterial species are similar but onmolecular level they are different than humans are from roundworms.
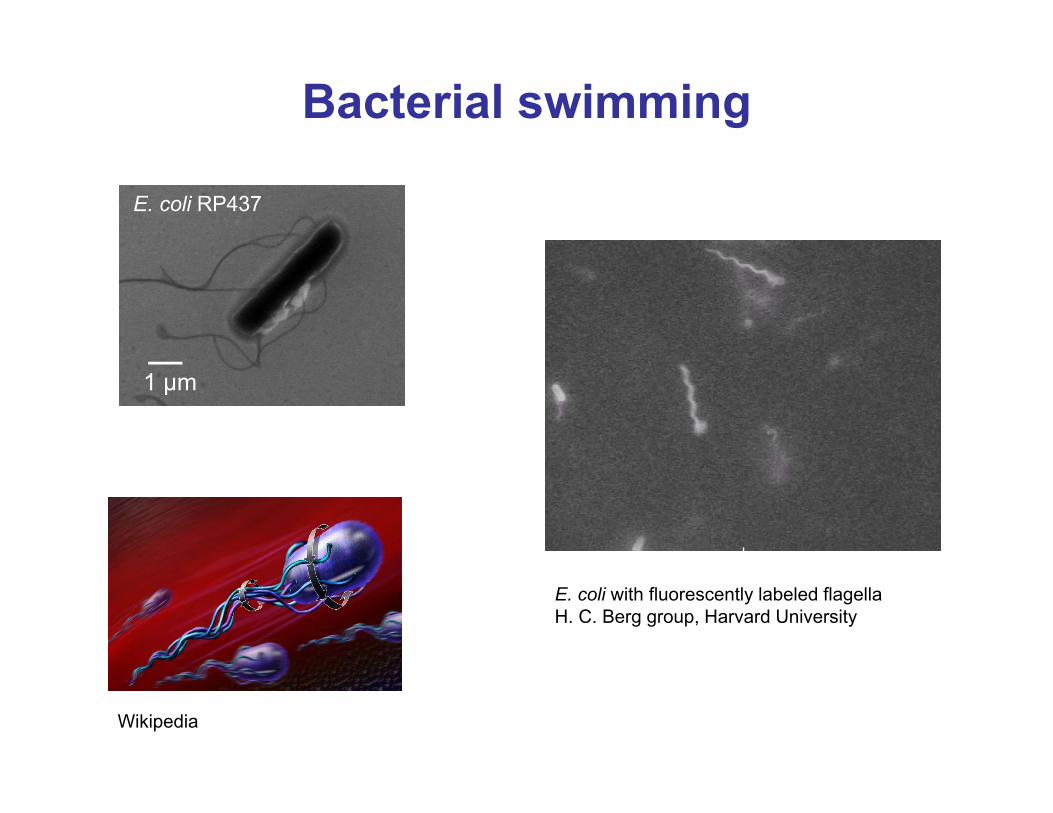
Bacterial swimming
E. coli with fluorescently labeled flagellaH. C. Berg group, Harvard University
Wikipedia
E. coli RP437
1 µm

Introduction: Bacterial motility
• Bacterial flagellar motor is a rotary motor superficially similar to DC electrical motor• Rotation speed controlled membrane potential and pH gradient across inner membrane• Bacterial sensory system (chemotaxis receptors through signaling cascade) control direction of rotations

Bacterial motility in channels
channel chamberchamber
W = 1.2 PmE. coli RP437
0.5 1.0 1.5 2.0 2.5
0
10
20
30
<v>
[Pm
/s]
W [Pm]
• E. coli and B. subtilis bacteria aremotile in channels which are only30-40% wider than their diameter
• In smaller channels bacteria losetheir ability to swim but ..

Growth and division is bacterial solution to penetrate narrower
channelsE. coli
channel chamberchamber
B. subtilis
W = 0.6 Pm

Different layout of channels
• More details of cells visible in the microscope
• Channel ceiling soft; Bacteria can deform it

Bacterial movement in sub-micron channels
• B. subtilis grows to filaments that buckle and finally divide
B. subtilis
E. coli
• Growth in confinement alter drastically E.coli shapes

Re-emergence of regularly shaped bacteria
5 hrs 49 hrs0 hrs
10 µm
• Over period 1-2 days regular rod-shaped bacteria replace initialpopulation of aberrantly shaped bacteria

Modes of penetration for different channel widths
• E. coli but not B. subtilis bacteria are able to grow through channelswhich widths are smaller than their diameters

Mechanical properties and propagation in narrow channels
• Cell-wall has high Young modulus but is easily bendable (think of inflated balloon)
• The thicker the cell wall the higher the Young modulus and the higher osmotic pressure can bacterium maintain in its interior
DWchannel wall
channel wall
E. coli B. subtilis
Posm=2-3 atmA. Boulbitch et al PRL 85 (2000) 5246
Posm=26 atmV. R. F. Matias et al Mol. Microbiol 56 (2005) 240
cell wall |3 nm cell wall 30-40 nm
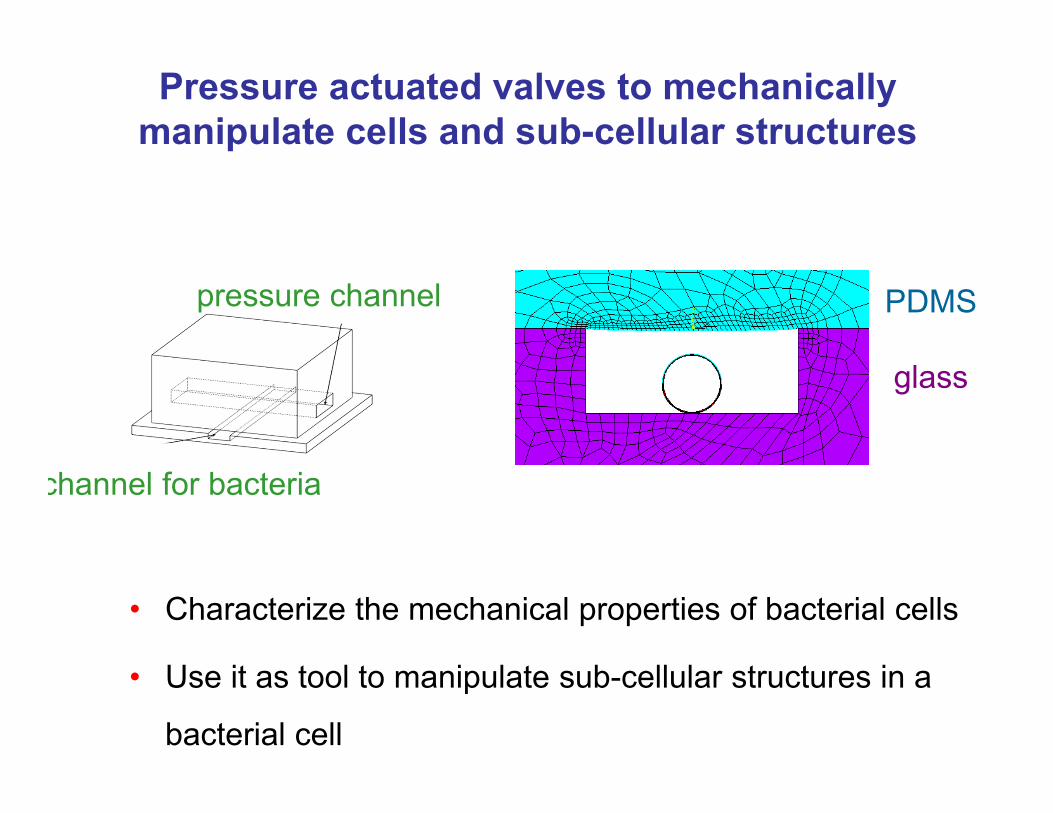
Pressure actuated valves to mechanically manipulate cells and sub-cellular structures
• Characterize the mechanical properties of bacterial cells
glass
PDMSpressure channel
channel for bacteria
• Use it as tool to manipulate sub-cellular structures in a
bacterial cell

Mechanical limits of bacterial deformations
• Bacteria break open at isolated weak spots at cell poles
Step #
pmax = 2 atm
pmax0
pmax = 3 atm
• Bacteria can be stretched up 25% elastically

Short vs long time response of E. coli
1
2
3
• Elastic deformation after entry in shallow channel• Slow cellular response after prolonged stay in channel
1
2
3

Summary on lab-on-a-chip and squeezing measurements
• Lab-on-a-chip platform allows to create biomimetic environments where cell behavior can be studied using high resolution optical microscope and bioanalytical tools.
• E. coli and B. subtilis are well adapted to swim in small channels despite their long flagella. Both species retain ability to swim in channels which only 30% exceed their body diameter
• Surprisingly, bacteria can get through even narrower channels! Forthat they use growth and division.
• Bacterial growth is robust despite drastic changes in their shapes
• Mechanical properties of bacteria determine how small channelsthey can penetrate.

Understanding how cells are built from molecules up using
quantitative high resolution microscopy and modelling

Escherichia coli as living “hydrogen atom”
Escherichia coli { E. coli
• Bacteria present the simplest systems to understand howthe cellular processes unfold using basic physicsprinciples
Jacque Monod, “What is Valid for E. coli is also valid for the elephant”1 Pm

What is known
1 Pm
FtsZ MatP tetramer SlmA tetramer
2 nm
• Functions of about 70% genes to some degree
• 50% of protein structures (most based on homology)

What is not known: from genes to cell
How nanometer-scale proteins, DNA and lipids come together and form the micro-size cell?
1 Pm
DNAlipid
polysaccharideprotein
?

Cellular organization in bacteria
• Bacterial cells are highly organized despite their apparent simplicity
Bag with soup of molecules Assembly of molecular machines
Wikipedia

envelope
• How are chromosomes and cell division apparatus organized in a bacterial cell?
• How cell division proteins position relative to nucleoids?

Chromosome organization

Genetic information is tightly packed in the cell
Escherichia colicircular chromosome
4.6�106
basepairs
1.6 mm
1 μ
• Thousand fold compaction of DNA in the cell
HupA-mCherry labelled chromosome

Nucleoid
1 μ
• DNA spreads just to a fraction of cell volume (~50%) – the nucleoid (not to be confused with nucleus)
HupA-mCherry labelled chromosome
nucleoid
• There is no membrane surrounding the nucleoid
cytosol

What compacts DNA?

Depletion Forces
• Molecules move so that to minimize excluded volume. This leads to appearance of an average force (depletion force) that pushes larger molecules together.
D
Volume gained (Vgained) by small molecules if two large molecules come together
• Alternative view: smaller molecules exert (osmotic) pressure to big ones which pushes them together.
abundant small molecules (crowders)
few largemolecules

Alignment and compaction of DNA strands as result of depletion
forces
• DNA strands are pushed together and aligned

Depletion forces compact DNA and lead to phase separation
1 μ
Nucleoid contains DNA and occludes most proteins and ribosomes
Cytosol phase is protein and ribosome rich
• Proteins and ribosomes are crowders. They push and compact DNA. In doing so DNA and proteins phase separate from each other (like water and ice do)

Dream of a bacterium is to make from one cell two cells (Jacque Monod)
• To divide bacterium needs to duplicate its DNA
Bacterium living up its dream

DNA within nucleoid is dynamic
• Some chromosomal regions occupy well-defined locations in the cell
• How exactly DNA is folded in the nucleoid not yet understood.

Modelling replicating chromosome in a cylindrical confinement
• Model DNA as self-avoiding chain of 100 nm blobs thatmoves under thermal force (Langevin thermostat)
Ter
oriC
oriC
100 nm
blob
Partially replicatedchromosome

Dynamics of replication terminus region
• Replication terminus region positions itself in the cellcenter as a result of entropic force
Ter
oriC
oriC
Ter
starting configuration equilibrium configuration

Cell division and organization cell division
apparatus
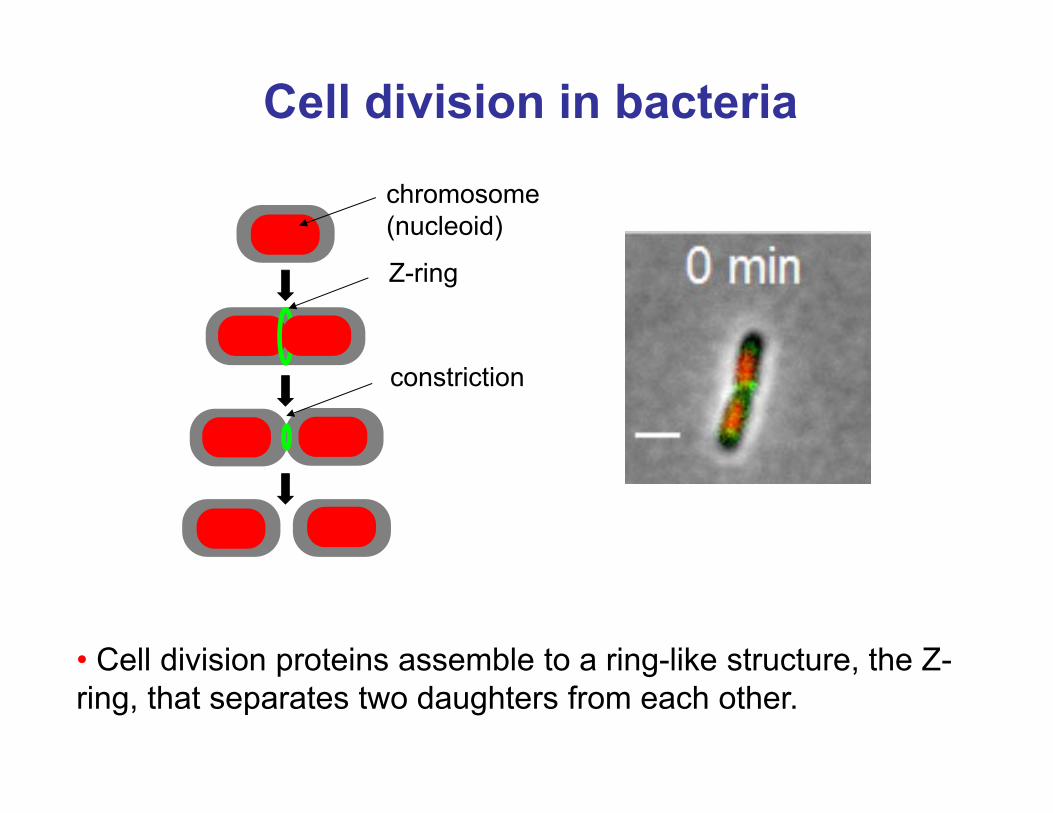
Cell division in bacteriachromosome (nucleoid)
Z-ring
constriction
• Cell division proteins assemble to a ring-like structure, the Z-ring, that separates two daughters from each other.

Z-ring �{ FtsZ ring
FtsZ monomer
FtsZ filamentH. P. Erickson et al., Microbiol. Mol. Biol. Rev. 74 (2010) 504
• FtsZ filaments act as a scaffold. This scaffold recruitsabout 30 other proteins to the ring (the fellowship of thering).
• Assembly of FtsZ filaments is the first step in cell division
D. W. Adams, J. Errington, Nat. Rev. Micro. 7 (2009) 642
Z-ring

Assembly of the Z-ring
Green, Red – FtsZ pointing up and down
Patchy band modelContinuous cohesive ring
Patchy band
S. Holden et al PNAS 2014 (111) 4566
• How filaments are exactly organized in the Z-ring is notknown

Positioning of cell division proteins
• Fellowship of the ring locates within 2% accuracy in the center of the cell

Go to the right place !!!
PDB: 1FSz
First view
• There is a little control freak inside every cell who knows where things need to go and it takes care of it.

• Self-assembly of molecules to macromolecularstructures driven by diffusion, intermolecularbinding, conformational entropy, excludedvolume effect etc.
Second view
PDB: 1FSz
Let’s get together at the
middle

Two known molecular systems in E. coli to localize its Z-ring:
• The Min system• Nucleoid occlusion mechanism

-2 -1 0 1 2
-2
-1
0
1
2
x [Pm]
y [P
m]
Min operon: minC, minD, minE
Min oscillations in E. coli cells
• MinD, MinE and MinC proteins oscillate between two poles of rod-shaped E. coli.
MinD::GFP
• Their time-averaged concentration is high at the cell poles. This defines inhibitory zone for Z-ring localization.

K. C. Huang et al PNAS 100 (2003) 12724
Reaction-diffusion system:
Min system: Turing instability in nature
, , …

Nucleoid occlusion
• Nucleoid occlusion: presence of nucleoid producesinhibition for the Z-ring assembly
D. W. Adams, J. Errington, Nat. Rev. Micro. 7 (2009) 642

Nucleoid occlusion factor SlmA
• SlmA binds to specific sequences of DNA that are missing in replication terminus region.
N.K. Tonhat et al EMBO J. 30 (2011) 154
• SlmA is inhibitor for Z-ring formation.
Terminus
Origin
H. Cho et al PNAS 108 (2011) 3773
Terminus
• Replication terminus region positions itself in the center of the cell. o No inhibition of Z-ring formation at the cell center.
SlmA
DNA

What happens to cells that lack these positioning systems?
WT
'slmA 'minC
'minC
a minicell
'slmA
Evidence for divisome localization mechanisms independent of the Min system and SlmA in Escherichia coli, M. W. Bailey, P. Bisicchia, B. T. Warren, D. J. Sherratt and J. Männik, PLoSGenet. 10 (2014) e1004504.

How can E . coli cells without known positioning systems still
divide about its middle?

-2 0 20.0
0.5
1.0
Inte
nsity
(arb
. uni
ts)
Length along the long axes [Pm]
�'Xz
�'Xn phase
DAPI
GFP-0.4 -0.2 0.0 0.2 0.4
-0.4
-0.2
0.0
0.2
0.4
'Xz/L
'Xn/L
'slmA'min
Localization of Z-ring in cells w/o known positioning systems
• Z-ring localizes the center of the nucleoid rather than tonucleoid free regions.
-0.30 -0.15 0.00 0.15 0.300
20
40
60
coun
ts
Xz-Xn [Pm]
'slmA'min
V = 66 nm
'slmA 'min

Positive regulation
• A positive signal that guides localization of cell divisionproteins from the cell center to the nucleoid center.
time [min]Le
ngth
alo
ng lo
ng
axes
[Pm
]
Z-ring
Chromosome
Leng
th a
long
long
ax
es [P
m]
'slmA 'min
2 Pm
-4 -2 0 2 4 60
1
2
I [ar
b. u
nits
]
Length along long axes [Pm]
nucleoid
Z-ring

Realization of the positive signal
ΔslmA ΔminC ΔzapA ΔslmA ΔminC ΔzapB
MisplacedMisplaced
'slmA 'min 'matP
-1.0 -0.5 0.0 0.5 1.00
20
40
60
coun
tsXz-Xn [Pm]
N=123
-1.0 -0.5 0.0 0.5 1.00
20
40
60
80
coun
ts
Xz-Xn [Pm]
N=218
-1.0 -0.5 0.0 0.5 1.00
20
40
60
coun
ts
Xz-Xn [Pm]
N=145
V = 202 nm V = 150 nmV = 222 nm
• ZapA, ZapB and MatP proteins are required for thepositive signal

Summary on organization of bacterial cells
• Min system and nucleoid occlusion are two known systemsfor positioning of bacterial Z-ring via a negative regulation.
• ZapA, ZapB and MatP proteins are needed for the positioning.
• Replication terminus region can spontaneously position itselfin mid-cell as a result of entropic force. This spontaneousprocess appears ultimately to control the placement of celldivision proteins.
• We have found a positive regulatory mechanism thatpromotes Z-ring formation at the vicinity of the Ter region ofthe chromosome.

Thanks
Matthew Bailey, UTKGeorge Siopsis, UTKDan Castillo, UTKJaana Männik, UTKPaola Bisicchia, Oxford Univ.David Sherratt, Oxford Univ. Piet de Boer, Case Western Reserve Univ.Alex Dajkovic, Univ. of Paris 5Rodrigo Lamothe-Reyes, McGill Univ.Cees Dekker, Delft University of Tech. Conrad Woldringh, Univ. of AmsterdamArieh Zaritsky, Ben Gurion University NSF CAREER award
CNMS User Program: 2015-231UTK start-up funding

Thank you for your attention!
There is lab tour starting in 5 min.