
Nutritional Value Harpacticoid Copepods Live Foodnlc-bnc.ca/obj/s4/f2/dsk2/ftp04/mq24887.pdf ·...
157
Nutritional Value -of Marine Harpacticoid Copepods as Live Food for Marine Fish Larvae. BY Dominic Andre Nanton Submitted in partial fulfillment of the requirements for the degree of Master of Science Dalhousie University Halifax. Nova Scotia CANADA May, 1997 O Copyright by Dominic Andre Nanton, 1997
Transcript of Nutritional Value Harpacticoid Copepods Live Foodnlc-bnc.ca/obj/s4/f2/dsk2/ftp04/mq24887.pdf ·...

Nutritional Value -of Marine Harpacticoid Copepods as Live Food for Marine Fish Larvae.
BY
Dominic Andre Nanton
Submitted in partial fulfillment of the requirements for the degree of Master of Science
Dalhousie University
Halifax. Nova Scotia
CANADA
May, 1997
O Copyright by Dominic Andre Nanton, 1997

Nationai L i i BiMimthèque nationale du Canada
Acquisitions and Acquisitions et Bibliograp hic Services services bibliographiques 395 Wellington Street 395, nre Wellington ûüawaON K1AON4 OttawaON K1AON4 Canada Canada
The author has granted a non- exclusive licence aJlowing the National Library of Canada to reproduce, loan, distnie or sell copies of this thesis in rnicroform, paper or electronic formats.
The author retains ownership of the copyright in this thesis. Neither the thesis nor substantial extracts fkom it may be printed or otherwise reproduced without the author's permission.
L'auteur a accordé une licence non exclusive permettant à la Bibliothèque nationale du Canada de reproduire, prêter, distn'buer ou vendre des copies de cette thèse sous la forme de microfiche/nlm, de reproduction sur papier ou sur format électronique.
L'auteur conserve la propriété du droit d'auteur qui protège cette thése. Ni la thése ni des extraits substantiels de celle-ci ne doivent être imprimés ou autrement reproduits sans son autorisation.

TABLE OF CONTENTS
TABLE OF CONTENTS .............................................. ...... ..... .. ..... i v LIST OF TABLES, FIGURES AND PLATES v i
0 0 0 ABSTRACT ............................................... ...... ............ . . . .... VI II
ACKNOWLEDGEMENTS ............................................ . ............ .. . ...... i x
Chapter 1 Fatty Acid Composition of Haipacticoid Copepods
and their Nutritional Value for Marine Flsh Larvae
O F m O N COPEpOD UpiDs IMnODUCTION . . .. . . . . . .. .. . . ... . .. . ... . . .. .. . .... .. . .. . . .. . .. . . .. . . .. . . . . . .. . . . . . . . . . . . . 1 5 MATERIALS AND M E M O D S . . ~ . . . . . . . . . . . . . . . . . . ~ ~ m . . . . . . . . . m a l 6
COPEPODCULTURE ...................................................... . ......1 6 ALGAL C U L T U R E . . . . . . . . . . . m . . . . . . a . . ~ . . . . . . o . . . . m 8 LlPlD ANALYSIS .................................... . ................... ........1 9 STATISTICAL ANALYSIS ..................... ........ .. ..... .. .. . . . . 2 3
RESULTS . ............... ..... ........ ............... . ......... .... ...... ......... ....... 2 3 DISCUSSION . . . .. . . .. . . . . . . . . .. . . . . . .. . . . . .. . . . . . . . . .. . . . . . .. . . . . . . . . . . . . . . . . . . ... . . . . . . . . . . O
EFFECT OF DlET ON COPEPOD UPIDS. ............ ... . ... .. . . .. . . ... . .. . ..A O NUTRITIONAL IMPLICATIONS FOR MARINE FISH LARVAE . . . . . ... 4 7
IMRODUCllON . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . .. . m l MATERIALS AND MRHODS ................................... ...... . . . . ...... . 3 RESULTS ............................................. . ............................. ........... 5 4 DISCUSSION . e . . . . . . . . . . . . . . . . . . . . . . m . . . . . . . m . . . . . . . 6 O
EFFECT OF TEMPERATURE ON COPEPOO UPIûS.. .. . . .. . . . ... . .. . . . .. .6 O NUTRITIONAL IMPLICATIONS FOR MARINE FISH LARVAE . . . . . . . .6 4
ID COMPOSITION OF qllVlNAtlVE -eFC(ES INTRODUCTION . . . . . .. . . .. . .. . . .. . . . . . .. . .. . . . . . .. . . .. . . . . . .. . . . .. . . . . . . . . . . . .. . . .. . .. . . . . . .6 6 MATERIALS AND METHODS ............. ........... . . .. . . . . . . . . . . . . . . . . . . . . . . . . . .. 6 6

. . . . . ....... ............................................................ . .
DISCUSSION ........................................ 5 LlPlD COMPOSITION OF ALTERNATNE UVE FOOD SPECIES ..... 75 NUTRITIONAL IMPLICATIONS FOR MARINE FlSH LARVAE ........ 77
Chapter 2 Free Amino Acid Composition of Harpacticoid Copepods
and their Nutritional Value for Marine Fish Lawae
.................... ............... ............................... INTRODUCTiûN .. ..i 2 FREE AMIN0 AClD M E T ' L I S M IN COPEPODS ...................... 82 FREE AMIN0 AClD METABOLISM IN MARINE FlSH LARVAE ....... 84 EXPERIMENTAL OBJECTIVES . . . ~ . m . ~ . . . . ~ . . . . m m ~ . ~ . m . . . ~ . . a m 8 6
MATERIALS AND METHODS .............................. ... .......................a... 0 8 RESULTS ....................................... 1 DISCUSSION ....................................... 9 5
Chapter 3 Mars Culture of a Harpacticoid Copepod Tisbe sp .
..... ............................................. INTROOUCTION ................... .. .. 0 1 ............................................................. MATERIALS AND MtlTnODS 102 RESULTS AND DISCUSSION .am.e.mmmmmm.mm..m~~ . . m m . . . m ~ ~ m m m m m m l 0 5
Chapter 4 Preliminary f rials Using a Harpacticoid
Copepod. Thbe sp., as a Dlet for Marine Fish Lawae
.................. ......................... JMRODUCTlON ...................... . . 1 1 ...... ................ MATERIALS AND MEMODS .. .............................. 1 1 4 RESULTS AND DISCUSSION .................... ... .. .... .....................m... 119
GENERAL CONCLUSIONS ........................................................ 2 7 APPENDICES
A . LlPlD ANALYSIS TECHNWES ........................... ... .................... 130 B . ASTkWüWHIN MEASUREMENTS ................... ... .... .... ...... ....A 32 C . FREE AMIN0 AClD IXITWCTION ............................. ..... ............ 135 ....................................... BIBLIOGRAPHY ........................... ... 1 37


UST OF FIGURES
Fig . l a . Fig . 1b . Fig . 2 . Fig . 3 . Fig . 4 . Fig . 5 . Fig . A1 . Fig . A2 . Fig . B I . Fig . 62 .
The n-3 and n-6 families of fatty acids ......................... 4 Alternative pathway to A 4 desaturase ......................... 5 Photographs of the harpacticoid copepods .................... 17 Tisbe sp . and Amonardia sp . Diagram of the nsbe mass culture systern ................ 103 Population counts from the nsbe masa .................... 106 culture system . Photographs of haddock l a ~ a e fed Tisbe .................. ..l 25 Bligh and Dyer lipid extraction method ...................... 130 Methyl ester formation method .................................. 131 ..... Visible absorption spectrum for wild zooplankton 133 astaxanthin at different wavelengths . Astaxanthin standard plot and regression .................. 134 analysis .
vii

ABSTRACT
Live food organisms containing relatively high concentrations of 13-3 highly unsaturated fatty acids (HUFA) are required for the first-feeding of cold-water marine fish lantae. In particular, long- chain fatty acids such as 20:5n-3 and 22:6n-3 are essential. Two species of harpacticoid copepods isolated from the Atlantic Oœan near Halifax, Nova Scotia, Tîsbe sp. and Amonardia sp., demonstrated the ability to elongate and desaturate 18:3n-3 to 20:5n-3 and 22:6n- 3 when fed a diet deficient in these essential fatty acids (EFA). Decreasing th8 culture temperature from 20 to 6OC increased the proportion of long-chain EFA (20:5n-3, 22:6n-3 and 20:4n-6) in the copepods. Both harpacticoid copepod species maintained a 22:6n-3 to 20:5n-3 ratio of greater than two for al1 dietary (the algae Chaetocenrs calcitrans, DunalieMa te rtiolecta , lsochrysis galbana and baker's yeast) and temperature (6, 15 and 20°C) regimens, suggesting their usefulness as a live food for marine fish larvae. For corn parison. trochophores of the common mussel Mytilus edulis and wild zooplankton obtained from St. Andrew's N.B. were examined for use as potential live food and were also found to have a high percentage of long-chah EFAs.
The relative amounts of free amino acids (FAA) in Tisbe rernained f airly consistent despite large differences in the amino acid composition of the diet. The propoition of essential free amino acids was nearly doubled in Tisbe fed 1. galbana (clone T-iso) compared with Tisbe fed yeast.
The harpacticoid copepod Tisbe was mass cultured in a system which produced nearly 10' individuals in a relatively small tank volume of 32 L.
P reliminary feeding trials comparing growth and su wival O f marine fish larvae (American plaice and haddock) fed rotifers or Tisbe were conducted. The plaice and haddock larvae were observed feeding on the harpacticoid copepods. Tisbe, when compared w i th the rotifers, gave superior growth but poorer survival of the haddock larvae.

I would like ta extend thanks and appreciation to al1 the
membem of my supervisory cornmittee, D~s. JmD. Castell, I.A.
McLaren, J.S. Craigie, SBJB Iverson, R a Ackman and my external
examiner, Dr. C.C. Parrish, for their advice and comments regarding
the research.
Special thanks to Dr. J.D. Castell foi his daily guidance and
encouragement, to Dr. I.A. McLaren for organizing the finances, and to
Dr. J.S. Craigie for the amino acid analysis of the copepod samples.
1 would like to thank L. Boston foi showing me the l ip id
analysis techniques and for her helpful advice. Statistical
assistance was kindly provided by DrB R Rodgers. Appreciation goes
to Dr. S. Johnson and Dr. R. Huys for harpacticoid copepod
identification. Thanks must also be extended to Dr. C. Monisson and
J. Martel1 for the sectioning and preparation of the lawal haddock
slides. I am grateful to the Aquarium staff, F. Rahey, P. Fraser and B.
Bassett, for their help in the setting-up and maintenance of the
culture systerns.
I would also like to thank my family for their support and
encouragement.
This work was funded in part by a research grant from Atlantic
Fisheries Adjustment Program (Dept. Fisheries and Oceans, Canada), the New Finfish Aquaculture Species Prograrn (CanadiadNew
Brunswick Cooperative Agreement on Economic Diversification and
Aquaculture Development for Nontraditional Species), and the
Patrick Lett Fund (Dalhousie University).

Chapter 1
Fatty Acld Comporit ion of Harpact icoid
Copepods and Implicatlonr on their
Nutritional Value for Marine Firh Larvae

GENERAL INTRODUCTION
OVEMON
Aquacuîture is the most significant growth component of the
Canadian fisheries industry. Salmon culture in the Maritimes now
represents an estimated 100 plus million dollar industry (McGeachy
et al., 1996). The world wide growth of salmonid culture has
resulted in a depression of th8 market value, and consequently
interest in the culture of other valuable marine fish species such as
halibut Uippoglossus hippoglossus, haddoock Melanogrammus
aeglefinus, flounder Pseudopleuronectes americanus and cod Gad-
morhua. lnterest in marine fish culture has dramatically increased
with the recent failures in the commercial fishery for northem cod
and other important fish stocks. There hm been some success in
rearing hali but, cod, sea bream Spanrs aurata, turbot Scophthalmus
maximus and other fish species in Noway, France, the United
Kingdom, Spain, Japan, Canada and other countries eround the world.
The main bottleneck for fry production in the species
mentioned above and other cold-water marine fish is associated
with larval first feeding. First feeding occum when the endogenous
energy resewes in the lama's yolk sac are depleted and the larva

must begin to feed exogenously. Lawal mortalities are generally
greatest at this stage. The high mortality is, in part, due to the lack
of nutritionall y adequate live food organisms. Fish l a ~ a e req u i re
live food organisms which have relatively high concentrations of the
long-chain, n-3 highly unsaturated (HUFA; M double bonds) or
essential fatty acids (EFA) such as 20:5n-3 (EPA; eicosapentaenoic
acid) and 22:6n-3 (DM; docosahexaenoic acid). These are essen tial
because the lawae do not have th8 necessary A-6, A95 and putative
6-4 desaturases to synthesize these fatty acids (FA) from shorter
chain n-3 fatty acids (Fig. 1). The bnne shrimp Artemia sp. and the
rotifer Brachionus plicatifis, which are widely used as live food f O r
cfustacean and fish lanral culture, have limited value in the culture
of cold-water marine fish larvae because of their low long-chain
EFA levels. Marine copepods, which are the principal cornponents of
the natural diet of many marine fish lawae, have higher amounts of
the long-chain EFA which makes them an attractive alternative l i ve
food source for the first feeding of cold-water manne fish larvae
under hatchery conditions.
My first objective was to evaluate the nutritional value of
indigenous marine copepods as live food for use in the commercial

Essential in Diet
Fig. la. The n-3 and n-6 families of fatty acids. The slanted and vertical arrows represent elongation and desaturation reactions, respectively. An alternative pathway to the putative 6-4 desaturase is depicted in Fig. 1 b.
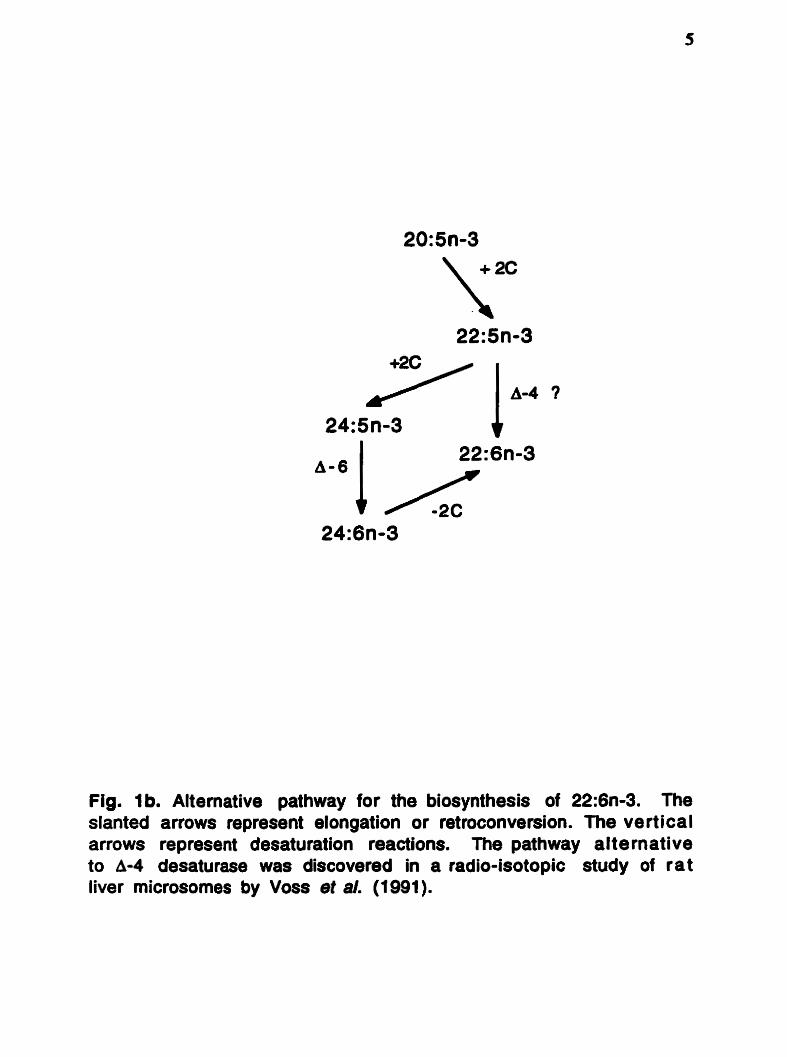
Fig. Ib. Alternative pathway for the biosynthesis of 22:6n-3. The slanted arrows represent elongation or retroconversion. The vertical arrows represent desaturation reactions. The pathway al t e rnative to A-4 desaturase was discovered in a radio-isotopic study of rat liver microsornes by Voss et al. (1 991).

culture of locally important marine fish larvae such as halibut, cod.
flounder, and haddock. This was to be achieved by establishing
cultures of the native harpacticoid copepods, Tisbe sp. and
Amonardia sp. The effects of algae and yeast in diets, as well as
those of temperature on the essential fatty acid composition of the
copepod populations were then to be detemiined.
FArrY AClD REQUIREMENTS OF MARINE flSH L A M
Most marine fish larvae have an essential requirement for the
n-3 long chain HUFAs, 20:5n-3 and 22:6n-3 (reviews: Greene and
Selivonchick, 1987; Watanabe, 1982). Cowey et al. (1976) were
among the first to examine the fatty acid requirements of marine
fish. Turbot Scophthalmus maximus juveniles were fed isocal O ric
diets containing differing amounts of the n-3 series of fatty acids.
They demonstrated that weight gain and protein efficiency fat i O
were highest in the turbot when polyunsaturated fatty acids (PUFA)
of the n-3 family were present in the diet. Scott and Middleton
(1979) examined the EFA requirements of turbot larvae, and
observed that the larvae had better sunrival and growth, when fed
rotif e ts which contained elevated concentrations of 20:Sn-3 and
22:6n-3 in their lipids. The ratio of DHA to €PA also significantly

affects the suwival of marine fish. lncreasing the ratio of DHA t o
EPA from 0.1 to 0.5 in the diet of turbot markedly decreased the
mortalities (Bell et al., 1985). There is other evidence for the need
of a high DHA to €PA ratio in the diet of marine fish lawae. For
example, the yolk of wild marine fish eggs contains a DHA to EPA
ratio of about 2.0 (Parrish et al., 1999, and this high ratio has been
observed in the polar lipids of copepods, the natural prey of marine
fish lawae (Kattner et al., 1981).
Tocher et al. (1 989) incorporated various radio-labelled PüFA
into cultured turbot fin cells to detemine which specif ic
desaturation and elongation reactions were taking place. The ce1 ls
exhibited A-6 desaturase activity converting 18:Zn-6 to 18:3n-6 and
18:3n-3 to 18:4n-3, however, they could not convert the supplied
18:3n-3 into 2031-3 indicating either a lack of 6 5 desaturase or of
the C-18 to C-20 elongase enzymes. Tocher (1993) found that the
A-5 desaturase was also lacking in the turbot brain astroglial cells.
The enzyme activity required to biosynthesize 22:6n-3 was also low.
When 20:5n-3 was the sole PUFA incorporated into turbot astroglial
cells, the elongation reaction of 2Mn-3 to 22:5n-3 predominated
over the reaction converting 22:5n-3 to 22:6n-3. This may explain

why turbot and other cold-water marine fish have sudi a large
dietary requirement for 22:6n-3 (Bell et al., 1985).
Arachidonic acid (AA; 20:4n-6) is another fatty acid which may
be considered essential fo i marine fish. Castell et al. (1 994)
discovered that, when turbot S. maximus juveniles which had
previously ben fed a diet with high proportions of 22:6n-3 but
deficient in 20:4n-6, were fed a diet containing 20:4n-6 as the sole
HUFA, they expenenced higher growth and suwival than those
supplied 22:6n-3 as the sole HUFA Arachidonic acid is the major
fatty acid precursor of several important eicosanoids including
prostaglandins, leukotrienes and hydroxytrienoic acids. Bell et al.
(1 995) discovered that increasing the amounts of 20~411-6 i ncreased
the levels of prostaglandin (POE and PGF) in the tissue homogenates
of the juvenile turbot.
The live food organisms traditionally used for warm-water
fish culture, the brine shrimp ARemia salina and rotifer Brachionus
plicatilis, are not as nutritionally valuable as copepods for col d-
water marine fish larvae. Brine shrimp cannot synthesize or
incorporate significant arnounts of the 22:6n-3 fatty acid which i s
crucial for the survival of most marine fish larvae (Watanabe et al.,

1978). Davis and Olla (1 992) fed lamal walleye pollock Theragra
chalcogramma a diet of Artemia sp. only, and found reduced g rowth
and suMval. However, when wild-caught copepods were added t O
the diet, growth and suwival of the lawae improved. Rotifers can
synthesize highly unsaturated fatty acids (HUFA) to a lirnited extent,
but to supply sufficient amounts of HUFA to marine fish lanrae, the
rotifers must be fed EFA-rich food (Lubzens et al., 1985). Kitajima
et al. (1980 a,b) demonstrated that ayu Plecoglossus altlvelis and
red sea brearn Pagnrs major lawae cultured with rotifers which
were fed bakers' yeast supplemented with cuttlefish liver oïl (high
in EFA) had superior growth than those fed a diet of unenriched
rotifers.
The value of the trocophores of the mussel MytiIus edulis, as
an alternate live feed for marine fish larvae, was evaluated by
Howell (1973). They did not support growth in lawal plaice
Pleuronectes platessa and sole Solea solea. However, if a diet O f
trocophore lanrae is followed by rotifers, the diet can support the
development of lemon sole to the point when newly-hatched Artemia
nauplii c m be used by the lawae for further growth t o
metamorphosis (Howell, lQ7l).

The future of marine fish lawal culture may lie wi th
fomiulated micro-particulate feeds. Micro-particulate diets WOU Id
eliminate the need to culture algae for consumption by live food
organisms thus simplifying larval culture. They could also eliminate
the need for live food if microcapsules could be fed directly to the
marine fish Ianrae. Up to the present, such diets have been
unsuccessful as food for manne fish lanrae. This is thought to be
due primarily to the deficiency of appropriate digestive enzymes i n
the incomplete digestive system of the earliest larval stages
(Walford, 1991). Munilla-Moran (1 990) proposed that exogenous
enzymes supplied by live food organisrns such as copepods play an
essential role in their own digestion in the gut of the marine f ish
larvae.
COPEPOD UPlD METABOUSM
Watanabe et al., (1 978) suggested that it is the copepods' high
HUFA levels that make them nutiitionally valuable for marine fish.
It is well established that manne calanoid copepods, parti CU l arl y
those from northem latitudes. are generally rich in lipid which i s
made up prirnarily of wax esters. In calanoid copepods the lipids can
mach 80% of the dry weight with the wax esters making up to 90%

of the total lipid (Sargent and Henderson. 1986). The marine wax
esters are contained in oil sacs which nm parallel to the gut and
consist primarily of long-chain fatty alcohols (usuaily saturated O r
monounsaturated) esterif ied to fatty acids (often pol yunsaturated) . Copepods accumulate wax esters after phytoplankton blooms (spring,
early fall) and we them as fuel resewes during prolonged periods
(usually winter) of food shortage (Sargent and Henderson, 1 986).
The long chain EFAs for cold-water marine fish, 20:5n-3 and 22:6n-
3, are prominent in the lipids of these copepods (Ackrnan et al..
1974; Sargent et al., 1977; Kattner et al., 1981; Nonbin et al.,
1990). For calanoid copepods the large amounts of n-3 HUFA are
probably incorporated directly from their phytoplankton diet. Prahl
et al. (1 984) demonstrated the selective absorption of n-3 HUFA i n
calanoids by feeding the green alga, Dunaiieiia primolecta, t O
Calanus helgolandicus and observing a drop in the n-3 HUFA
composition of the food as it passed through the gut to f o n faecal
pellets. Stattnip and Jensen (1 990) demonstrated that, because O f
the inability of Acartia tonsa to elongate and desaturate 18:3n-3 t O
longer chain HUFA, this calanoid shows reduced growth and egg
production when fed Dunaliella tertiolecta, an afgae deficient i n

20511-3 and 22:6n-3. These findings were confirmed with a series
of studies looking specifically at the fatty acid content of the diet
and relating it to egg production in the calanoids Acartia
(Jonasdottir, 1994; Jonasdottir and Kiarboe, 1996) and Temora
longicomis (Jonasdottir et al., 1995). The fatty acid requirements
were for high n-3 to n-6 and high 22:6n-3 to 20:5n-3 ratios. The
fatty acid 22:6n-3 was also positively correlated with egg
production for both species.
Harpacticoid copepods, which are benthic with some
exceptions, live in an environment with a more stable food supply
than calanoid species. They are known to feed opportunistically ,
efficiently utilizing various food sources such as vegetables (Kahan,
1 979), polychaete tissues (Guidi, l984), bacteria (Rieper, 1 Q78),
unicellular algae, detritus (Ustach, 1 QW), dried mussel powder,
yeast and macroalgae (Miliou and Moraitou-Apostolopoulou, 1 99 1 b) . Therefore, they do not need to accumulate wax esters to the same
extent as calanoids, and their lipid composition is typicalty around
10% of their dry weight (Miliou et al., 1992). Norsker and Stprttrup
(1 994) discovered that a harpacticoid copepod Tisbe holothuriae
does have the ability to elongate and desaturate the shorter chain n -

3 HUFAs from Duneliella tertiolecta to produce relatively large
amounts of the essential far( adds 20:5n-3 and 22:6n-3. When fed
D. tertiolecta, egg production of the harpacticoid was not
significantly decreased by the lack of long-chah €FA in the diet
when compared with Rhodomonas baltica, an alga with a relatively
high content of long-chain EFAs.
Poikilothermic animals, including copepods, tend to increase
their unsaturated to saturated fatty acid ratios as tem peratu res
decrease. The increase in this ratio is an adaptation thought t o
allow membrane fluidity to be maintained at lower temperatures
(Hazel and Williams, 1990). Farkas (1979) noted a large increase i n
the PUFA in the phospholipids of freshwater copepods as the
temperature decreased from approximately 20 to 1O0C. The largest
proportion of this increase in PUFA consisted of the EFAs, 22:6n-3
and 20:Sn-3. The levels of total PUFA including 20:5n-3 and 22:6n-3
were also obsewed to increase in the lipids of various species of
marine calanoid copepods in the North Sea during the colder winte r
months (Kattner et al., 1981).

OBJECTIVES
The nutritional or EFA value of indigenous mafine harpact i coid
copepod species as lhre food for the commercial culture of
important marine fish larvae (Le. halibut, floundei, cod, and
haddock) was evafuated by:
a) establishing cultures of native harpacticoid copepods;
b) determining the effects of diet (algae and yeast) on the fatty acid
composition of two species of cultured harpacticoid copepods (Tlsbe
and Amonardia) ;
c) detemining the effects of temperature on the fatty acid
composition of the cultured harpacticoid copepods (Tisbe and
Amonardia) fed the same algal diet;
d) comparing th8 fatty acid compositions of the two harpacticoid
copepod species (Tisbe and Amonardia);
e) cornparing the essential fatty acid composition of the
harpacticoid copepods with other potential native live food
organisrns for marine fish larvae. Wild zooplankton (made up almost
entirel y of calanoid copepods) and 24-h-old f ertilized eggs and
trochophores of M ' u s edulis were the other live foods evaluated.

Effect of Diet on Copepod Lipids
INlRUMlCTlON
One objective of this study was to determine if the
harpacticoid copepods are able to elongate and desaturate shorter
chain n-6 and n-3 fatty acids such as 182n-6 18:3n-3 into the long-
chain EFAs 20:4n-6. 20:5n-3 and 22:6n-3. This ability can be
detennined by feeding the copepods diets which have only trace
amounts of these EFAs (Le., the alga, Dunalieilla tertiolecta. and
bakers' yeast) and then detemining the copepods' fatty acid
compositions. Norsker and Stattrup (1 994) discovered that the
harpacticoid copepod. Tisbe holothuriae. does have the ability t O
elongate and desaturate the shorter chain n-3 HUFAs in Dunaliella
tertiolecta to produce relatively large amounts of the essential
fatty acids 20:5n-3 and 22:6n-3. lt was, therefore, expected that
our indigenous harpacticoid copepods (Tisbe sp. and Amonardia sp.)
would also posess the sarne desaturase and elongase enzymes needed
to produce 20:5n-3 and 22:6n-3 from the shorter chah n-3 f at ty
acids such as 18:3n-3.
The other main objective was to examine which diets gave
higher levels of the n-3 and n-6 essential fatty acids and high DHA

to EPA ratios in the copepod lipids. These parameters are crucial
for the survival and growth of cold-water marine fish larvae
(Watanabe, 1982). The copepod diets were chosen on the basis o f
their large differences in long-chain EFA composition. lt was
expected that, because of the harpacticoid copepods' elongating and
desaturating abillties, they would maintain relatively high levels O f
20:4n-6, 205n-3 and 22:6n-3, and a high DHA to EPA ratio even when
fed long-chain €FA deficient diets.
MATERlALS AND MElHWS
Copepod Culture
The harpacticoid copepods, Tisbe sp. and Amonardia sp. (Fig.
2), were captured and isolated from the plankton in Halifax Harbor,
Nova Scotia, Canada in Febniary 1994. They were cultured for over
10 generations on a rnixed algal diet at 20°C (generation time was
approx. 2 weeks). The calanoid species, Acartia hudsonica and
Eurytemora herdmani, were also captured in the same tow, but due
to low production rates and poor survival the numbers necessary for
lipid analysis were not obtained. The harpacticoid copepods we re
raised in 6-L cylindrical plexiglass (McDonald) jars containing
seawater (32 ppt salinity) which had been filtered through 10 prn
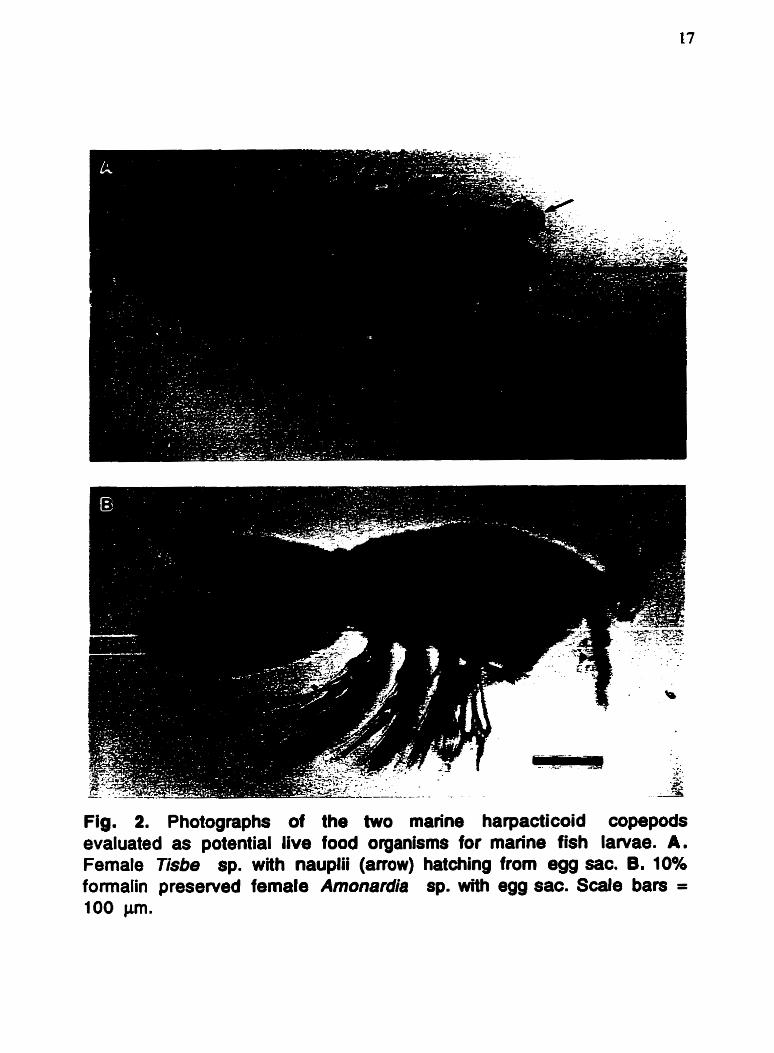
Fig. 2. Photographs of the two marine harpacticoid copepods evaluated as potential live food organisms for marine fish larvae. A. Female Tisbe sp. with nauplii (anow) hatching from egg sac. B. 10% formalin preserved femele Amonsrdie sp. with egg sac. Scale bars = 100 Pm.

pore size cartridges and UV treated. Oxygen saturation and wate r
circulation were maintained with air flowing through airstones at
the bottom of each cylinder. Temperatures were maintained a t
2WI0C. The seawater was changed approximately twice per week
by collecting the copepods on a 40 pm mesh and transferring them to
jars of fresh, filtered seawater. Three replicate cylinders of
copepods were used in the feeding trials and the algae and yeast
were fed to the copepods in excess (ca. 1 mg dry weight/L of sea
water) each time the water was replaced. The copepods were
cultured in this manner for approximately 4 weeks pn'or to l ipid
analysis.
Algal Culture
Pure strains of the algae lsochrpis galbana (UK isolate),
Chaetoceros calcitrans and DunaIieIIa tertioiecta were purchased
frorn the Provasoli-Guillard Center for Culture of Marine
Phytoplankton (West Boothbay Harbor, ME). ErJenmeyer flasks (2 5 0-
m l ) containing 150 mL of autoclaved sea water and f/2 medium
(Fritz Algae Food, Leland, MS) developed by Guillaid and Ryther
(1 962) were then aseptically inoculated with the pure strain of
algae and stoppered with cotton wool to allow for gas exchange.

The cultures were maintained at 18OC for 3-5 d before being
transferred to 1 1-L carboys. The 1 1 -L carboys contained 10 L O f
autoclaved seawater. 2 mL of Fritz Algae Food f/2 Solution A and 2
mL of Solution B were then added. The carboys were maintained a t
18°C and bubbled vigorously with air. Carbon dioxide was added at a
flow rate of 0.5 Umin. The algae were hawested while in the log
phase at 6-8 d (approx. 5 million cells/mL) for feeding to the
copepods.
Lipid Analysis
Samples of the algae used in the feeding trials were separated
from the culture medium by centrifugation. The algae were
harvested in the log phase and equal volumes were poured into 2 5 0 -
mL plastic flasks. These flasks were placed in a 5.75 in. maximum
radius rotor and spun at 5000 rpm (g force = 334) for 20 min using a
Serval RC28 centrifuge. The seawater was decanted. The algal
pellet was then scraped out of the plastic flask and analyzed f O r
lipids and dry weight.
Adult copepods were collected on 200 pm mesh nylon screens
and transferred to jars containing fresh 10 Pm filtered, UV-treated
sea water. They were then stanred for 24 h to allow for the

clearance of dietary algae present in the gut and rescreened t o
remove any accumulated waste products. More than 200 animals per
replicate were isolated in this manner for iipid analysis.
Subsequently, the copepods were suction-filtered on Whatman no. 1
filter pape? and rinsed with distilied water. The copepods plus 1 0
mL of chlorofonn:methanol (2:l v/v) were then homogenized in a 20-
mL glass culture tube using a polytron (8n'nkman Instruments).
Total lipids for the diets and copepods were extracted using a
modified method based on that of Bligh and Dyer (1959). For details
of method see Appendix A; Fig. Al. If, after removal of the
chloroform, the total amount of lipid was too small (4 mg) for an
accurate weight measurernent, a known amount of an interna1
standard fatty acid (23:O) was incorporated in the sample t o
estimate lipid weijht. Methyl ester derivatives of the total f ip id
were prepared by adding 2 mL 7% BF3 in methanol to the lipid sample
in 15-mL glass tubes with teflon-lined screw caps and heating t O
1 OO°C. After cooling, the methyl esters were recovered by adding 5
mL anhydrous sodium sulfate and extracting twice with 4 mL
hexane. The hexane was evaporated and the methyl esters were

purifieci by thin layer chromatography. For details of method see
Appendix A; Fig. A2.
The fatty acid methyl esters were t k n quantified using a
Varian 3400 mode1 gas liquid chromatograph (GU) equipped with a
hydrogen Rame ionization detector (FID) and a polar column
(Omegawax 320 flexible fused Mica capillary column; 20 m i n
length x 0.32 mm ID). The injection temperature in the GU= went
from 100 to 250°C in 0.51 min and was held at 250°C for 1 min so
that the sample was fully vaporized. The det8~tor temperature was
300°C, and the oven temperature was programmed as follows: the
initial colurnn temperature was 160°C increasing to 240°C at 3.S°C/
min for 22.85 min and then held for an additional 12 min at 240°C t o
clear any residual material from the column. Total nin time was
34.85 min. The pressure of the helium carrier gas was set at 82
kPa; gas flow through the column was 3.0 mumin. An inboard data
handling system detemined retention times and integrated the
areas under each methyl ester peak. Individual peaks were
tentatively identified by compaiing their retention times w i t h
known standard fatty acid methyl esters, typical of marine oils,

which were previously identified in an Ornegawax Column Test Mix
(Supelco, Bellefonte. PA).
Identification of some peakl not present in the reference
sample was f acilitated through the use of hydrogenation. The peak
that iepresented an unsaturated fatty acid methyl ester w i l l
disappear after hydrogenation and the equivalent chain lengt h
saturated fatty acid methyl ester will increase by an amount equal
to the previous unsaturated fatty acid methyl ester. Thus, we can
determine the carbon chain-length for the unknown unsaturated
fatty acid methyl ester. The methyl ester sample to be hydrogenated
was placed in a 250-mL Erlenmeyer flask with 20 mL methanol and
approximately 0.5 mg of platinum oxide catalyst. The flask was
flushed with hydrogen gas two or three times stoppered and left t O
stir (magnetic stirrer) for 1 h. The plaünum dioxide was removed
from the sample by suction-filtering the methanol through Whatman
no. 1 filter paper and rinsing with with 20 mL hexane. The sample
was then evaporated to dryness under nitrogen and the same volume
of hexane used for the unhydrogenated sample was added for dilution
prior to injection onto the GLC (Christie, 1982).

Statistical Analysis
Signif icant differences in the lipid compositions of the
materials examined were evaluated by analysis of variance (ANOVA)
using Systat 5.1 for personai cornputers (Wilkinson, 1990). Tukey's
HSD (Highly Significant Difference) test which is a post-hoc
paiwise comparison test was used at the pS0.05 level to compare
diff erences between means. The fatty acid compositions we re
calculated as a percent of total lipid. Data represented as
percentages are not normally distributed (the tails of percentage
data distributions are shortened due to the finite O and 100% l i m i ts)
which violates the assumptions for ANOVA. The data were therefore
normalized or transfomied using the arcsine of the square root o f
the proportion (arcsine X ln) before ANOVA was applied.
RESULTS
Analyses for the ash weight and lipid percent of dry weight of
the three algae and one yeast diet are presented in Table 1. The
diatorn Chaetoceros calcitrans had the highest ash weight due to the
presence of its siliceous shell. The ash content of the flagellates,
Dunaliella tertiole cta and lsochrysis galbana, were similar a t
around 10% dry weight. lsochrysis galbana had the highest lipid

Table 1. Ash weight and total llpid content of copepod diets.
Dunafiella tertiolecta 8.95 * 2.15 25.67 f 3.09
Chaetoceros caIcitrans 24.30 * 2.33 21 -67 f 4.24
Isochrysis galbana 10.65 1 .O0 28.41 f 0.99
Ba kers' yeast 5.12 î 0.85 2.19 i 0.14
Values are mean (n-3) f standard deviation on a dry matter basis.
The algae was washed with ammonium formate pikr to ashing.

content, followed in decreasing order by D. tertiolecta and C
calcitrans. Both the ash and lipid contents of the bakers' yeast
were very low.
The three species of aigae in this study were chosen because
of the large differences in their EFA compositions (Table 2). Lipids
of I. galbana had significantly more 22:6n-3 than the other diets
(about 1/4 of the total FA) and a small amount of 20:5n3.
Conversely, Cm calcitrans contained significantly more 20: 5n-3
(about 1/3 of the total FA) compared to the other diets and only a
small amount of 22:6n-3. 0. tertiolecta had only trace amounts of
both 20:5n-3 and 22:6n-3. However, Dm tertiolecta lipid had
significantl y higher amounts of the 18-carbon fatty acids, 1 8:2n-6
(almost 10%) and 18:3n-3 (almost 50%). The content of 20:4n-6 was
present at low levelç in al1 diets (<0.3%), but was significantly
higher in C. calcitrans than in any of the othei diets. These extreme
differences in the EFA levels allowed a cornparison of the effects of
dietary €FA on the fatty acid composition of the harpacticoid
copepods.
The bakers' yeast diet (Table 2) had small percentage of both
20:Sn-3 (not significantly different from the levels in I. galbana)

Table 2. Fatty acid compositions of the algae (C cal= Chetoceros calcitrans, D tert- Dunaliella tertiolecta and I gal= lsochrysis galbana) cultured at 18OC with f/2 media and of bakers' yeast used in the copepod feeding trials. Diffeient letters indicate sig ni f ican t differences at pc0.05 level.
Fatty acid C d 0 tert lad Yeast

mm 7.7 f l.7r 9.4 f 6.h 10.0 f 4.8r 23.7 f 0.66 33.8 f 2.3r 11 .O i 2.5b 23.5 & 3.lc 89.8 I 1.2d
C PUfA 57.1 f 1-68 76.1 f 5-36 65.1 & 6.6. 5.8 I 0.7~ C ri-3 37.4 f 3-98 57.8 * 3-86 51.6 & 6.5b 4.6 f 0 . 7 ~ C n-6 5.3 f 0.4. 15.1 * 5-76 11.8 f 0.9b 0.8 f 0 . 1 ~ DHAEEPA 0.1 f 0.0. 2.0 I 1-46 12.2 f 3 . 1 ~ 1.1 I 0.4a C UNK 0.6 f 0.4 1.6 f 1.3 0.2 f 0.1 t r
Results are expressed as 45 of total fatty acids and represent means and standard deviations of 3 repficates. Unknowns el% not included in table. Abbreviations; nd= not detected, tr= trace (<0.05%), 7 Me= 7 methyf, SA'= saturates, MûNQ monounsaturates. PUFk polyunsaturates, UNK= unknowns, DHAt docosahaaenoic acid, E P h eicosapentaenoic acid.

and 22:6n-3 (not significantly different from the levels in C
calcitrans). The fatty acid composition of bakers' yeast d i f f e red
markedly when compared with the algae. The levels of saturated and
monounsaturated fatty acids were at least double that of the dosest
algae. The fatty acids, l6:l n-7 (20.4%), 18:l n-7 (27.3%) and 1 8: 1 n-9
(17.9%) were the major contributors to the high levels of
monounsaturates.
The fatty acid composition of the harpacticoid copepod, Tisbe
sp., fed these four organisms are presented in Table 3a. The yeast
was fed to the copepod at a different time of year than the algae
(Jan., 95 W. Aug., 94, respectively), so that other factors, such as
season, may have affected the fatty acid composition of the
copepods when compan'ng the algaf and yeast diets.
In Tisbe, the relative amounts of the 18 carbon fatty acid,
1 8:3n-3, incorporated by the copepod corresponded with the amount
in the algal diet. 18:3n-3 was highest in D. tertiolecta (45.0%) and
the copepods fed with this diet. It was much lower in copepods fed
with C. calcitrans and I. galbana. This was not the case with 1 8:2n-
6. Tisbe fed with I. galbana incorporated slightly higher amounts
of 18:2n-6 than Tisbe fed with D. tertiolecta, evee though

Table 3a. Fatty acid composition of Tisbe sp. cultured at 20°C and fed algal (C cal= Chaetocem calcitrans, I gai= Isochrysî" galbana and D tert= Dunaliella tertiolecta ) and bakers' yeast diets. Differences in lettering indicate signîficant differences at the pc0.05 level.
l4:O l 4 : l n-7 l 4 : l n-5 l5:O DMA16 l6:O l6: l n-7 7-Me16 l 6 : l n-5 16:2n-6 16:2n-4 l7:O 16:3n-3 16:4n13 16:4n-1 DMA 18 l8:O 18:ln-9 l 8 : l n-7 l 8 : l n-5 18:2n-9 18:2n-6 18:Zn-4 18:3ri-6 18:3n-4 18:3n-3 18:4n-3 20:o 20:1 n-11 20:1 n-9 20:1 n-7 UNK 20:2n-6 20:3n-6 20:4n-6 20:3n-3 20:4n-3 20:Sn-3

22:o 6.9 * 2.9. 1.9 0.460 3.8 * O h 1.6 * 0.56~ 22:ln-11 0.3 * 0.1 0.3 * 0.1 0.2 * 0.1 0.5 * 0.4 22:In-9 0.4 I 0.1 0.3 * 0.1 0.3 * 0.1 0.2 * 0.0 22:ln-7 0.1 f 0.2 0.3 * 0.1 0.2 * 0.1 r d 21:Sn-3 0.2 f 0.2 0.2 * 0.1 0.6 * 0.2 nd 22:4n-6 0.1 f 0.1 0.7 * 0.7 0.4 * 0.5 nd 22:Sn-6 0.5 * 0.5 1.2 k 0.2 1.4 * 0.7 0.9 * 0.3 22:4n-3 0.2 t 0.3 0.1 î 0.0 0.3 * 0.4 0.2 * 0.1 22:5n-3 1.0 î 0.4 0.4 0.2 0.5 * 0.1 1.1 * 0.5 24:O 3.8 * 0.6. 1.5 0.2b 2.5 * 0.60 rid 22:6n-3 21.4 * 1.9.b 12.4 2.98 22.9 * 9.8.b 28.4 * 6.b 24:ln-11 6.6 * 4.1. 0.2 * 0.lbc 1.6 1.2.0 1.2 * 0.4bc 24:1 n-9 1.3 * 0.1. 0.6 0.lb 0.6 * 0 . lb nd 26 :O 4.0 * 2.3 2.7 * 1.1 3.7 * 0.5 0.9 * 0.3 C a . 40.3 * 3.3.6 43.7 6.8. 33.0 * 5.6.b 28.0 2 3.26 PW 18.0 3.2 18.3 * 1.2 13.8 * 2.7 20.7 * 3.7 z WFA 36.2 * 1.4.6 35.4 * 5.4. 45.7 * 9.- 50.1 * 3-06 C n-3 31.6 * 2.0.b 25.9 * 5.5. 32.1 * 11.0.6 43.3 3-76 2 n-6 4.1 * 1-38 8.3 & 0.7d 11.7 * 3.46 6.3 1.6.0 D M P A 2.6 * 0.3 2.0 * 0.1 3.3 * 0.5 3.1 1.3 C UNK 5.5 * 1.0. 2.7 1.h~ 5.4 * 2-24 1.2 * 0.46~
Results are expressed as the 96 of total fatty soi& and r q c r e n t means and standard deviations of 3 replicates. Unhomu ~ 1 % not inciuded in taôle. Abbraviations: n& not detected, tr= trace (<0.05%). 7 Me= 7 methyl, D M k dimethyl anteisu, SAT= saturates. M C N b monounsaturates, PUFA.: polyunratuiates. UNlG unknowns, DCUI doco98her~.noic acid, EPAr eicosapentaenoic acid.

Table 3b. Percent change in the fatty acid composition of Tisbe sp. cultured at 20°C as compared with the fatty acid composition of the algal (C cab Chaetoceros calcitrans, O tert= Dunaliella tertiolecta and I gal= I s o & ~ s galbana) and bakenr' yeast diets. The minus (-) symbol indicates the fatty acid is found in the algae but not detected in Tisbe sp. The plus (+) symbol indicates the fatty acid i s found in the ~0p8p0d but is not deteded in the algae.
Fatty acid C d D tert l ml veast

Results are expressed as 96 change in total fatty ad& from the diet to the q m p d and represent the change in the mean of 3 repficates. Abbreviations; 7 Me= 7 methyl, SAT= saturates, MON- rnonounsrrturatm, PUFk polyunsaturatcw.

Dm tertiolecta contained a higher proportion of this fatty acid. The
lipid from copepods fed al1 three algal spedes contained
substantially lower percentages of 18:3n-3 than was present in the
lipids from those algal species. The largest reduction in 18:3n-3
from the diet (ca. 45%) to the copepod (ce. 4%) was seen in the
copepods fed D. tertiolecta. This represented a 91% decrease of
18:3n-3 (Table 3b).
The propodon of 22:6n-3 was higher in the lipids of Tisbe fed
1. galbana and calcitrans but not significantly different (pS0.05)
from the levels in the copepods fed 0. tertiolecta. There were
sig nif icantl y higher concentrations of 22:6n-3 in these two algae,
especially 1. galbana, than in D. tertiolecta which produced SI i g h t l y
lower levels in the copepods. The amount of 20:5n-3 was
significantly higher in the copepods fed with C. calcitrans than in
those fed with D. tertiolecta. The level of 20:Sn-3 in the copepods
fed with 1. galbana was not significantly different from the other
two algal diets. The DHA to €PA ratio was much higher (2.6) in the
copepod fed with C. calcitrans than in C. calcitrans itself, wheie
this ratio was extremely low (0.05). This was due. to both a large
reduction in the proportion of 20:5n-3 (about a 75% decrease; Table

3b) in copepod lipid compared with that in the Cm calcitrans diet, and
a corresponding 10-fold increase (Table 3b) in th8 proportion of
22:6n-3 in the copepod. The lipids of D. tertiolecta-fed copepods
contained relatively large amounts of 20:Sn-3 and 22:6n-3, even
though only trace amounts of these EFAs were present in that alga,
and this accounted for a 45-fold and a 40-fold increase,
respectively (Table ab). In copepods fed with lm galbana, there was a
2-fold increase in the proportion of 20:5n-3, but a slight (10%)
decrease in the proportion of 22:6n-3. Tisbe fed the bakers' yeast
had extremely high amounts of both 20:5n-3 and especially 22:6n-3
in their lipids (Table 3a).
It was apparent from differences in the EFA content between
each of the diets and the copepod, that Tisbe managed to maintain a
consistently high DHA to €PA ratio despite the extreme diff erences
in the arnounts of these fatty acids in the diet. This ratio ranged
between 2.0 and 3.3 and there were no significant differences
(ps0.05) in this ratio between the ~0p8pods fed any of the diets.
The levels of 20:4n-6 were relatively high (~1.0%) for Tisbe
fed each of the diets. The copepods fed with Dm tertiolecta

contained 1.5% of 20:4n-6 in thelr lipids, even though this fatty acid
was below detectable levels in this alga.
Two of the algae (D. tertiolecta and I. galbana) were also f ed
to another species of harpacticoid copepod, Amonardia sp. (Table
4a). This copepud had low levels of 20:5n-3 (4%). when fed w i t h
either D. tertiolecta or 1. galbana. The levels of 22:6n-3 (>13%)
were much higher than those of 20:Sn-3 (Table 4a). Amonardia fed
with 1. galbana increased the percentage of 20:5n-3 in their l ipids
slightly, and those fed with D. tertiolecta dramatically increased
(>4-fold) the percentage of 20:Sn-3 in their Iipids compared to the
diet (Table 4b). The ~0p8pods fed with 0. tertiolecta also had a ve ry
high, almost 300-fold, increase in 22:6n-3 compared to the diet. As
was the case in Tisbe, the percentage of 22:6n-3 in the copepods fed
with 1. galbana was slightly reduced compared to the diet (Table
4b).
Unlike Tisbe sp.. Amonardia sp. did not maintain a constant
DHA to EPA ratio and there was a significant difference in th is
ratio between the copepods fed the two algal diets: 24.0 for those
fed 0. tertiolecta compared with 6.6 for those fed 1. galbana. This
was due prirnarily to the low level of 20:Sn-3 in the copepods fed O.

Table 4a. Fatty add composition of Amonardia sp. culhired at 20°C and fed the algae lsochrysis galbana (1 gaî) and Dunaliella tertiolecta (O tert). The asterisk indicates significant diff erences at the p<0.05 level.
Fat& acid l cral 0 tert ug LipW copepod 0.51 f 0.12 0.31 f 0.04 14:O 1.4 f 0.4 0.3 i 0.3 14:1 n-9 0.4 f 0.2 0.2 f 0.2 14:ln-7' 1.3 f 0.8 0.1 f 0.2 15:O' 0.4 f 0.1 0.9 i 0.2 16:O 32.9 f 3.8 47.9 f 6.5 16:l n-7' 3.3 f 0.4 0.4 f 0.1 7-Me 16 0.6 f 0.2 nd 16:1 n-5 0.9 f 0.5 0.2 f 0.2 16:2n-6 0.4 f 0.2 nd 16:2n-4' 0.9 f 0.2 1.7 i 0.3 17:O 2.0 f 0.4 2.5 f 0.6 16:3n-4 0.4 f 0.4 0.3 f 0.3 16:3n-3 0.2 f 0.1 0.2 f 0.2 l6:4n-3' 1.8 f 0.7 0.7 f 0.1 16:4n-1' 0.7 f 0.1 1.8 f 0.2 18:O' 7.5 f 1.2 3.9 i 0.6 18:1 n-9' 4.0 k 0.3 2.6 i 0.1 18:l n-7' 2.3 f 0.1 1.5 f 0.0 18:1 n-5 0.4 f 0.2 0.2 î 0.2 18:2n-9 0.4 f 0.3 nd 18:2n-6 1.0 f 0.1 0.3 î 0.2 18:2n-4' 0.3 f: 0.0 1.2 f 0.4 18:3n-6 1.1 f 0.7 0.4 f 0.2 18:3n-4 0.8 f 1.0 nd 18:3n-3' 1.1 f 0.1 2.3 f 0.7 t8:4n-3 0.8 f 0. 7 nd 18:4n-1 0.7 f 0.8 1.6 f 0.6 20:o 1.1 * 0.1 1.1 i 0.1 20:1 n-9 0.7 k 0.5 nd 20: 1 n-7 0.1 k 0.0 nd 20: 1 n-5 nd 2.7 * 0.4 20:2n-6' 0.1 f 0.0 1.4 f 0.4 20:3n-6' 1.2 f 0.3 0.3 I 0.1 20:4n-6 0.3 f 0.1 0.1 f 0.1 20:3tt-3 t r 0.2 I 0.3 20:4n-3' 0.6 I 0.2 1.2 i 0.3 20:5n-3' 2.7 i 0.8 0.8 f 0.1 UNK 0.3 * 0.6 0.4 k 0.0 22:O 2.0 f 0.3 1.8 f 0.3 22: 1 n-1 1 0.5 f 0.1 0.1 f 0.2 22: 1 n-9 0.2 f 0.1 nd

26:09 0.7 f 0.2 1.7 f: 0.8 C mT 52.1 f 5.8 60.1 f 6.2 C MONO' 14.5 f 1.4 9.4 f 0.6 C PüFA 34. 6 f 4.6 30.0 f 6.0 C n-3 26.0 4.1 19-9 + 4.7 C n-6 4-4 f 0.5 3.6 f 0.8 DHA/EPA* 6.6 * 3.0 24.0 * 3.8 C UNK" 1.4 f 0.8 0.6 f 0.2
Results are erqrressed as 96 of total futty a& and repnisent means and standard deviations of 3 replicates. Unknowns 4% not iriduded in taMe. Abbreviations; n& not detected, tr= trace (<O.OS%), ?-Me= 7-methyl, SATI saturates, MONOI monounsaturates, PUFR polyunsaturates, UNK= unknowns, DHAt docosahexaenoic acid, €PA= eicosapentaenoic acid.

Table 4b. Percent difference in the fatty add composition of Amonardia sp. cultured at 20% as compared with the fa€ty acid composRion of the algai (D tert- DunaIfella tertiolecta and I gai= lsochrysis galbana) diets. The minus ( 0 ) symbol indicates the fat t y acid is found in the algae but not detected in Amonardia sp. The plus (+) symbol indiates the fatty acid is found in the copepod but is not detected in the algae.
Fatty acid I ad D tert 14:O - 1 2 371 14:1 n-9 229 - 6 1 14:1 n-7 889 - 4 6 14:l n-5 - - 15:O 807 1 4 0 4 16:O 352 4 7 3 16:ln-9 - + 16:l n-7 4 8 -03 7-Met 6 - 7 0
16:l n-5 4967 - 8 7 16:Zn-6 1941 0
16:2n-4 - 8 1928 17:O 3283 525 16:3n-4 496 548 16:3n-3 262 - 8 9 16:4n-3 25827 - 9 4 16:4n-1 1 IO7 13321 18:O 1185 1 089 18:ln-9 -63 16 18:ln-7 7 6 2 f 8 : l n-5 685 + 18:2n-9 + - 18:2n-6 - 7 9 - 9 7 18:2n-4 + 6 3 1 3 18:3n-6 1126 - 9 0 18:3n-4 1873 + 18:3n-3 - 9 1 - 9 5 18:4n-3 - 9 2 - 18:4n-1 + + 20:o + + 20:1 n-11 - - 20:1 n-9 2757 - 20:1 n-7 + - 20: 1 n-5 + + 20:2n-6 - 7 8 2237 20:3n-6 592 21 8 7 20:4n-6 - 1 0 + 20:3n-3 - 298 20:4n-3 188 4456
Results are expressed es 96 change in total fatty adde from the diet to the cupfmd and represent the change in the mean of 3 replicates. Abbreviatkns; 7-Me= 7-methyl, SAfr saturates, MON- monounsaturates, PUFA= polyunsaturates.

tertiolecta, as the difference in the percentage of 22:6n-3 in the
copepod lipid fed the two diets was not significant.
The levels of 20:4n-6 were very low in Amonardia, 4).3% and
not significantly different (pS0.05) for those fed either alga.
DISCUSSION
Effects of Diet on Copepod Lipids
Dunaliella tertiolecta (CMorophyceae) contained only t race
arnounts (<O.S%) of the EFAs 20:Sn-3, 22:6n-3 and 20:4n-6. This
suggests that this alga contains low levels of active A-6 and A -5
desaturases, which are necessary to convert 1 8:3n-3 into 20: 5n-3
and 22:6n-3 and I8:2n6 into 20:4n-6. Chaetocmos calcitrans
(Bacillario ph yceae) contains relative1 y large amounts of the €FA
20:5n-3 and little 22:6n-3, which suggests that this algae contains
low levels of active enzymes necessary for 22:6n-3 synthesis.
lsochrysis galbana (Prymnesiophyceae) contained large amounts of
22:6n-3 and very little 20:5n-3, which is consistent with the
presence of either an active A 4 desaturase or the Schoenberg shunt,
which catalyzes the transfer of 20:Sn-3 on through to the 22:6n-3
end-product (Fig. 1 b). These fatty acid compositions are typical f O r
these classes of algae (Volkman et al., 1989). The bakers' yeast has

a large amount of A-9 desaturase activity as is evidenced by the
high level of 18:ln-9, 16:ln-7 and 18:ln-7 (a 16:ln-7 elongation
produd). These three A-9-desaturated fatty acids make up 65.5% O f
the lipid. The yeast contains low levels of th8 other desaturase
enzymes as is evidenced by its low amount of total PUFA (5.8%).
These species were chosen as diets because of their large
differences in €FA composition. The diets Mgh in 20:Sn-3 (C.
calcitrans) and 22:6n-3 (1. galbana) allowed us to examine the level
of incorporation and modification of these EFAs from the diet to the
copepod. The diets deficient in long-chain EFA (D. tertiolecta and
yeast) allow us to evaluate the desaturating and chain-elongating
abilities of the harpacticoid copepod.
Most crustaceans thus far used as live food organisms in cold-
water marine fish culture have no or only a limited ability t o
bioconvert shorter chain n-3 PUFA into the longer chain EFA, 20:Sn-3
and 22:6n-3 (Lubzens et al., 1985; Howell and Tzoumas, 1991). Both
harpacticoid copepods, Tisbe sp. and Amonarclia sp., that we
studied are able to synthesize significant amounts of 20:5n-3 and
22:6n-3, when fed Dunaliella tertiolecta, which contains only trace
amounts (<0.5% of lipid) of these EFAs. This confirms the work done

by Norsker and Stettrup (1994) on a European strain of Tisbe
holothuriae. They fed 0. tertiolecta to this harpacticoid and found
that il also could convert n-3 PUFAs from the algae into 20:5n-3 and
22:6n-3. This was further evidenced in their egg production study
with 1. holothuriae. There was no significant drop in the copepods'
egg production, when fed the €FA-poor Dm tertiolecte versus the
relatively €FA-rich aigae Rhodomonas baltica.
The harpacticoids, when fed D. tertiolecta, converted mainly
18:3n-3 (45.0% of lipid) into 20:Sn-3 and 22:6n9. This was
evidenced by a large decrease in 18:3n-3 from the amounts present
in the alga relative to the amount in both Tisbe and Amonardia
when fed this species. Along with this decrease, there was a big
increase for both copepods in the amounts of 20:5n-3 and 22:6n-3
relative to that in 0. tertlolecta. These copepods have the ability t O
convert 18:3n-3 into substantial quantities of 20:5n-3 and 22:6n-3.
Therefore, to be capable of this conversion the copepod must contain
suff icient amounts of the C 18-to-C20 and C20-to-C22 elongase, as
well as A-6, A-5 and putative A 4 desaturases (Fig. 1).
Tisbe, when fed C. calcitrans which is rich in 20:5n-3,
reduced its proportion by about 75% in its lipids. There was also a

concurrent 10-fold increase in aie percentage of 22:6n-3 in Tisbe
fed with calcitrans. This is consistent with the hypothesis t hat
the copepod uses C20-to-C22 elongase to convert 20:5n-3 to 22:5n-
3, and either A-4 desaturase or the Schoenberg shunt to convert
22:5n-3 to 22:6n-3. The disappearance of 20:Sn-3 may also be due,
in part, to oxidative catabolism. However, 20:Sn-3 constitutes over
90% of the total n-3 fatty adds in C. calcltram, and an n -3
precursor is needed to produce the levels of 22:8n-3 seen in the
copepods fed with C. calcitmns.
The copepods (Tisbe and Amonardia), fed an alga rich in 22: 6n-
3 (1. galbana) decreased the proportion of this EFA in its lipids when
compared with that in the alga. There was a slight decrease in Tisbe
and a somewhat larger decrease in Amonardia. As previously
mentioned, Tisbe when fed a diet rich in 20:Sn-3 (C. calcitrans)
reduced the proportion of this EFA in its lipid when compared with
the alga. When the copepods were f8d with the EFA-pooi a
tertiolecta and yeast, the proportions of 20:5n-3 were increased
dramatically in its lipid, relative to that in the diet (ca. 45-fold for
D. tertiolecta fed to Tisbe: ca. 5-fold for D. tertiolecta fed t o
Amonardia; ca. 6-fold for yeast fed to Tisbe). A similar increase was

obsewed for 22:6n-3 (40-fold for 0. tertioIecta fed to Tisbe; 4 5 - fold for Dm tertiolecta fed to Amonardia; almost 15-fold for yeast
fed to Tisbe). Because of these changes, the DHA to EPA ratio
remained high ( ~ 2 ) for boai harpacticoid copepod species for a l l
diets, despite the huge variations in the EFA composition of their
diets. In Tisbe, this ratio also remained remaikably constant.
There was also no significant difference in the DHA to EPA ratio
arnong the copepods fed any of the four diets. If the diet contained
quantities greater than a certain threshold for either 20:5n-3 or
22:6n-3, the harpacticoid copepods reduced the percentage of these
in its lipids. If the amount was below this threshold then the levels
were increased so that a consistently high DHA to EPA ratio was
maintained (Fig. 1 ).
The lipids of Tisbe fed with yeast contained th8 highest levels
of long-chah EFAs of any treatment group, despite the very l ow
levels of EFAs in the yeast itself (496 of both 20:5n-3 and 22:6n-3).
This can be partially explained by th8 copepods' low total I ipid
content. Tisbe fed with yeast had the lowest average lipid content
(0.20 pg/copepod) of any treatment group. Therefore, the i r
proportion of membrane phospholipids would necessarily be greater

and their storage lipids (triglycerides) less. The membrane lipids
are generally high in the EFAs, and their proportionate increase
would partially account for the high €FA levels of Tisbe fed wit h
yeast. The overall low lipid content of the copepod could be
accounted for by the low lipid content of the yeast (2.2% of the dry
W.). However, the yeast feeding trials were performed at a
different tirne of year from the algal feeding trials, so that seasonal
effects may also have played a role in lipid composition differences.
The other long-chain €FA, 20:4n6, was present at highei
levels in the harpacticoid Tisbe (A% for al1 the diets with no
significant differences among them) and at very low quantities in
Amonardia (<0.3% for those fed either diet). More evidence for A - 6
and A-5 desaturases in the harpacticoids can be seen in Tisbe fed
with D. tertiolecta. No 20:4n-6 was detected in D. tertiolecta, yet i t
was found in Tisbe fed this alga. For both harpacticoid species fed
with D. tertiolecta, there was a decrease in the levels of th8
ptecursor fatty acid 18:2n-6 (by 65% for Tisbe and by 97% f O r
Amonardia) and for Tisbe a concurrent increase in 20:4n-6 to 1.5%.
In Amonardie fed with O. tertiolecta the levels of 20:4n-6 were
very low (~0.5%). Most of the 18:2n-6 in these copepods was

elongated to 20:2n6 (>20-fold increase) and not desaturated to
18:3n-6 (the next step in the conversion to 20:4n6; Fig. l), which
was actually decreased in Amonadia by 90%.
Some general trends were seen in Tisbe fed al1 of the diets.
The levels of 18:O were increased, compared with the diet, for the
copepods fed al1 of the diets. 77368 may preferentially synthesize
this fatty acid de novo, as oppoged to 16:O. Th8 monounsaturate,
16:i n-7, decreased in the copepod lipid compared with the levels i n
the diet for the copepode fed al1 diets. There was a concurrent
increase in I8 : lnJ for the copepods fed al1 algal diets. suggesting
that the copepod rnay be elongating 16:l n-7 in the diet to 18:1 n-7.
Both 20:l n-7 and 22:l n-7 were also increased in Tisbe fed al1 diets
which indicating that 18:ln-7 may be further elongated to these
fatty acids in the copepod. However, 20:ln-7 and 22:l n-7 were
present in much lower quantities in the copepod lipid, compared
with 18:ln-7. In yeast. 18:ln-7 was present at extrernely high
levels, therefore, there was decrease in its level in the copepods fed
with yeast.
Amonardia, increased the total arnount of saturated f a t ty
acids, including 16:O and 18:0, relative to that in the dietary lipid.

Unlike Tisbe, Amonardia wntained a large amount of 16:0, which
was greater than the amount of 18:O. This suggests that there i s
less elongation of 16:O to 18:0, or that there is preferential de novo
synthesis of l6:O in Amonardia. As was the case in Tisbe, 16:O was
present at a higher level in the aie copepod fed with Dm tertiolecta
(48%) than in those fed with I. galbana (33%). However, th is
difference was not significant. The fatty acids 18:ln-9, 16:ln-7 and
18: 1 n-7 (a l6:l n-7 elongation produd) were al1 significantly higher
in Amonardia fed with I. galbana, indicating A-9 desaturase activity
was greater in this copepod. Those copepods fed Dm tertiolecta may
be channeling their energy into making the desaturases needed t o
convert 18:3n-3 into the EFAs (20:5n-3 and 22:6n-3), while l i rn iti ng
the synthesis or activity of the A-9 desaturase.
Nutritionai Implications for Marine Fish Larvae
In mafine fish larvae, 22:6n-3 has more €FA value than 20:5n-
3. This has been demonstrated in several species of manne fis h
larvae in feeding studies using the brine shrimp, which cannot
synthesize or incorporate high levels of 22:6n-3. Howell and
Tszournas (1991) fed brin8 shrimp from Brazil and Utah to larval
sole (Solea solea). The Brazilian strain was low in 22:6n-3, but had

significantly more than the Utah strain. The larvae had significantly
higher growth rates when fed the Brazillan strain, but there was not
enough of the 22:6n-3 to promote good survival. The ratio of EPA t o
DHA was also show to significantly affect the survival of marine
fish. Changing the ratio of €PA to DHA from 13.8 to 2.2 in the diet of
turbot markedly decreased mortalities (Bell et al., 1985). The
increased importance of 22:6n-3 compared . with 205n-3 as an EFA
for marine fish larvae was reviewed by Watanabe, 1993 f o r
yellowtail Serjola quinqueradiata. striped jack Pseudocaranx dentex.
striped knifejaw Oplegnathus fasciatus, red sea bream Pagms ma jar
and f lounder Paralichthys olivaceus.
Tisbe, because of its high desaturase activity, is able t o
produce large amounts of 22:6n-3 and maintain a consistently hig h
DHA to EPA ratio (2.0 to 3.4), despite the long-chah EFA
composition of the diet. Amonardie also had a very high DHA to EPA
ratio (23.4) when fed a long-chain EFA deficient diet. This
extrernely high ratio was due, in part, to very low levels of 20:5n-3.
These harpacticoids have a great potential as live food, because they
do not need to be fed an EFA-rich diet, but maintain high DHA to EPA

ratios within their lipids even when cultured on low cost and
maintenance foods such as yeast.
For both harpacticoids, higher 22:6n-3 values were achieved
when they were fed a long-chain €FA-rich diet. The exception was
Tisbe fed on yeast, which had lower total lipid levels. The long-
chain, EFA-rich algae, lsochrysis galbana and Chaetoceros
calcitrans, gave higher levels of 22:6n-3 in Tisbe, when compared
with Dunaliella tertlolecta ('Table 3a). Arnonardia also contained
higher levels of 22:6n-3 when fed with 1. galbena as oppoged to LX
tertiolecta (Table 4a). Therefore, feeding the harpacticoids w i t h
diets n'cher in long-chah €FA increases the amounts of 22:6n-3 in
the harpacticoid lipids, which can be made available to marine f i s h
larvae.
Arachidonic acid (20:4n-6) is also essential for marine fish.
Linares and Henderson (1 99 1 ) discovered that radiolabelled 20:4n-6
is incorporated into turôot phosphatidylinositol at a very specific
and high level. They suggested that, because of this specific
incorporation of 20:4n-6 into phosphatidylinositol, and because O f
its role as a precursor for prostaglandin synthesis, 20:4n-6 i s
probably essential for marine fish. Marine fish have very low levels

of A95 desaturase activity. Therefore, they cannot convert 18:2n-6
into sufficient amounts of 20:4n-6 and so require preforrned 20:4n-6
in their diet (Mourente and Tocher, 1993). Although marine f ish
require 20:4n-6, the demand is not as great as that for 22:6n-3 as
indicated by the higher whole-body retention of dietary 22:6n-3
compared to 20:4n6 in turbot juveniles (castell et al., 1994).
The harpacticoid Tisbe sp. contained greater than 1% 20:4n-6
with al! diets tested, including the alga, D. tertiolecta, which
contained only trace amounts of this EFA (Table 3a). Amonardla
produced lower levels of 20:4n-6 (4.3%) than Tisbe, producing
instead, increased proportions of 20:2n-6 and 20:3n-6, when fed the
algal diets (Table 4a). Tisbe, in terms of its arachidonic acid
composition, thus appears to be best suited for manne fish iarval
culture, when grown on various diets.

EFFECT OF TEMPERATURE ON COPEPOD LlPlDS
I M O N
Acute changes in environmental temperatures may have
deleterious effects on the structure and function of cellular
membranes of crustaceans as well as other poikilothermic animals.
Changes in temperature impact both the permeability of cel l
membranes and the activity of integral membrane proteins,
presumably via changes in the physical characteristics of the l ip i d
bilayer. It is generally assumed that, for a given set of membrane
constituents, there is an optimal range of ternperatures wi t h in
which suitable molecular interactions (either lipid-lipid or l i p i d - protein) occur for proper membrane structure and function. and that
this range is above the critical phase transition (gel to liquid-
crystalline) temperature of the bilayer (Pniitt, 1990).
This maintenance by poikilothemic animals of their membrane
lipids at a standard fluidity above the phase transition temperature
is known as "homeoviscous adaptation' and was first described f o r
Escherichia coli by Sinensky (1974). Poikilotherms tend t O
increase the ratio of unsaturated to saturated fatty acids of thei r
phospholipids in cell and organelle membranes as tempe rature

decreases (Chapelle, 1 078; Farkas, 1 979). Lower tem peratu res
increase the viscosity of the cell membrane by reducing the thermal
energy and causing greater cohesiveness of the macromolecules
which comprise the membrane. lncreased desaturation of membrane
phospholipids at lower temperatures increases the disorder of the
macromolecules in the membrane so that a standard fluidity i s
maintained.
Rainbow trout (Salmo gairdneri) hepatocytes had higher levels
of n-3 PUFA, in particular 22:6n-3 and 20:5n-3 as well as higher
levels of 20:4n-6 when acclirnated at 5OC rather than 20°C (Sellner
and Hazel, 1982). lncreases in 20:4n=6, 20:Sn-3, 22:6n-3 and to ta1
n-3 PUFA at lower temperatures have also been obsewed i n
crustaceans (Harrison, 199 1 ). Among copepods, large increases i n
2Mn-3 and 22:6n-3 fatty acids were also obsewed in freshwater
species as tempe rature decreased (Farkas, 1 979).
The objective of rny present study was to detemine i f
decreasing the rearing temperature will significantly increase the
level of EFAs (20:4n-6, 20:Sn-3 and 22:6n-3) in the harpacticoid
copepods, Tisbe and Amonardia, thereby making them more valuable
as a live food for marine fish larvae. In the copepod feeding t r ia ls

with various diets, a large potential for desaturation of fatty acids
was observed. In particular, harpacticoids had a great ability t o
convert 18:3n-3 into 20:5n-3 and 22:6n-3 and 18:2n-6 into 20:4n-6.
One would expect that, as an adaptation to the lower temperatures,
the copepods would further increase their already large amounts of
EFAs to maintain a standard membrane fluidity.
MATERIALS AND MEiHODG
The harpacticoid cupepods nsbe sp. and Amonardia sp. were
raised in 6-L cylindrical plexiglass jais containing seawater (32 ppt
salinity) which had been filtered through 10 prn pore site cartridges
and UV treated. Oxyg-en saturation and water circulation were
maintained with air flowing through eirstones at the bottom of each
cylinder. They were maintained for approximately 4 weeks at 3
temperatures 20, 15 and 6OC (i 1 OC) while being fed the same alga,
lsochrysis galbana. The medium was changed approximately t w i c e
per week by collecting the copepods on a 40 pm mesh and
transferring them to jars of fresh, filtered sea water. Three
replicate cylinders of copepoâs were used in the study and the alga
was fed to the copepods in excess (ca. 1 mg dry weight IL of sea
water) each time the water was replaced.

Lipid and statistical analyses were perfoned using the same
methods descilbed for the feeding trials (pp. 19-29).
RESULTS
There was a direct relationship between the percentage of
saturated fatty acids and temperature. As the temperature of the
environment decreased from 20 to 15 and e0C, the copepod,
Amonardia sp., significantly decreased the levels of saturated f a t t y
acids from about 52% to 32% in its lipid (Table 5). This difference
in the levels of saturated fatty acids was due principally to the
lower content of 16:0, which decreased by about half as the
temperature decreased from 20 to 15 and 6OC. The percentage o f
total PUFA increased at 6OC compared to 15 and 20°C. The PUFA
were at their lowest levels at 15OC. None of these differences i n
the PUFA at the various temperatures were significant (pS0.05).
The EFA 20:5n-3 in Amonardia increased significantly at 6°C
compared with 20°C, but there was a decrease in this PUFA at W C .
The pattern was similar for 22:6n-3, which showed a slight increase
at 6°C compared with 20°C and a significant decrease at 15OC
compared with 6 and 20°C. There were no significant differences in
the DHA to EPA ratio for the three ternperatures. As expected, the
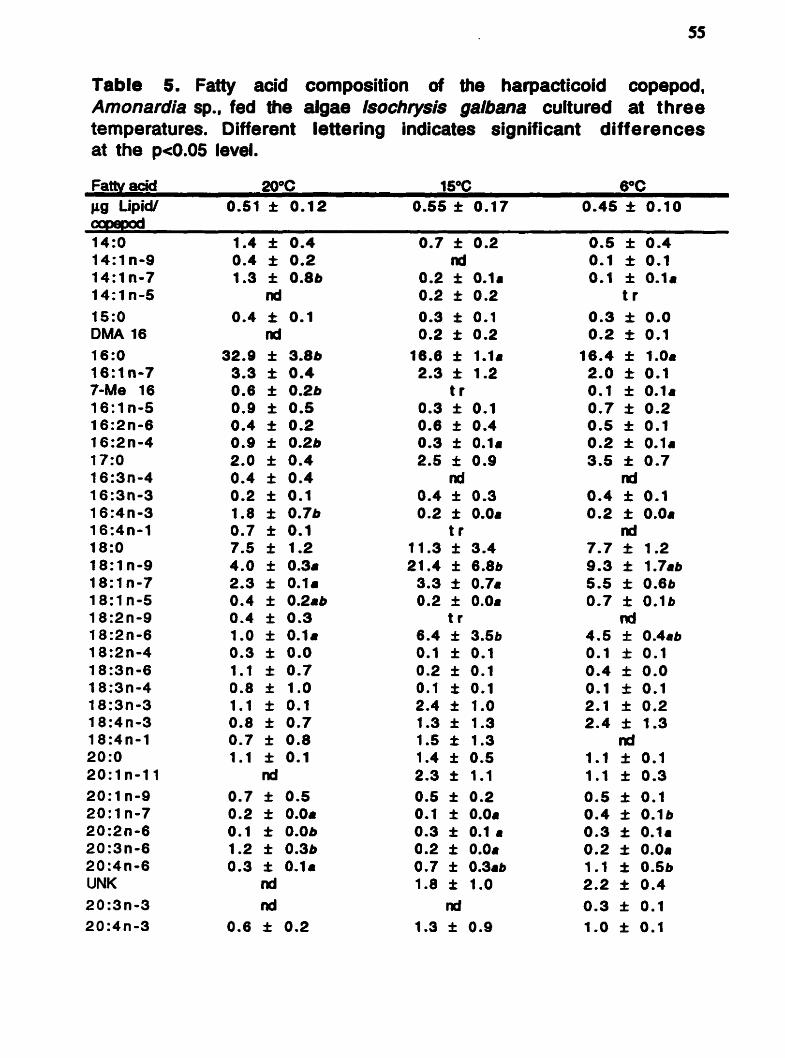
Table 5. Fatty acid composition of the harpacticoid copepod, Amonardia sp., fed the aîgae Isochvsis galbana cultured at three temperatures. Different lettering indicates significant differences at the pe0.05 level.
l4:O 14:ln-9 14:ln-7 l 4 : l n-5 l5:O DMA 16 l6:O 16:ln-7 7-Mû 16 16:ln-5 16:2n-6 16:2n-4 17:O 16:3n-4 16:3n-3 16:4n-3 16:4n-1 l8:O t8: ln-9 l 8 : l n-7 l 8 : l n-5 18:2n-9 18:2n-6 18:2n-4 18:3n-6 t8:3n-4 18:3n-3 18:4n-3 18:4n-1 20:o 20: 1 n-11 20:1 n-9 20:1 n-7 20:Zn-6 20:3n-6 20:4n-6 UNK 20:3n-3 20:4n-3

CMoNo 14.5 k 1.4b 32.0 i 9.9r 22.9 I 1.Oab C PUFA 34.6 f 4.6 29.6 f 8 A 41.8 f 3.9 x n-3 26.0 î 4.0ab 18.0 i 5.78 31.0 I 4.3b C n-6 4.4 * 0.56 11.6 * 4.58 10.5 I 1.0a D HA/ E PA 6.6 f 3.0 7.2 f 0-4 3.4 f 0.6 C UNK 1.4 * 0.8 2.7 i 0.6 3.1 f 0.7
Results are expressed as % of total fatty ecids and represent means and standard deviations of 3 replicates. Unhomu, 4% not induded in taMe. Abbreviations; nd- not detected, tr= trace (4.05%). 7-Met 7- rnethyl, D M k dimethyl anteiso, SAT= satutates, MCWQ rnonounsatumt8~, PUFA= poîyunsaturates, ü h k unknowns, DHA= docogahexaenoic acid, €PA+ eicosapentaenoic acid.

content of 20:4n-6 in the lipid increased as the temperature
decreased. The diierence between 20 and 6OC was significant.
The effects of temperature on the fatty acid composition of
Tisbe sp. were evaluated (Table 6). One replicate sample out of
three for Tisbe sp. cultured et 6OC was contaminated, so that
statistical cornparisons were only made between 20 and lS°C. The
amounts of PUFA increased only slightly at 6OC compared with 20°C,
but there was a significant decrease in the PUFA at 15OC compared
with 20°C. Th8 latter was a similar temperature response to t hat
shown by Amonardia. The saturated fatty acids rernained nearly
constant at about 30% for the three temperatures. The levels of
20:5n-3 were about 9% at 20°C and decreased substantially to 4%
at 15OC and 6OC (significantly so at 15°C). The amounts of 22:6n-3
increased at 6OC compared with 20°C (no significance att r i b uted) . However, there was a slight drop in the level of 22:6n-3 at lS°C,
which is anothei temperature response similar to that of Amonardia.
There was a steady increase in the DHA to EPA ratio as the
temperature decreased. As in Amonardia sp., 20:4n-6 reached i t s
highest levels in Tisbe at the lowest temperature tested.

Table 6. Fatty acid composition of the harpacticoid copepod, Tlsbe sp., fed the algae lsochryss galbana cultured at three temperatures. The asterisk symbol indicates significant differences at the pe0.05 level for 20 and lS°C.
Çaîty Acid 20°C 15OC 6°C 14:O 0.7 I 0.3 nd 0.1 14:ln-9 0.2 * 0.3 nd t r 14:l n-7 0.3 f 0.3 nd t r 14:1 n-5 0.1 f 0.2 nd O. 1 1 5:0 0.5 f 0.2 nd 0.2 DMA 16 0.1 f 0.1 nd 0 -2 DMA 16 0.1 f 0.2 0.1 16:O' 19.5 f 5.4 9.9 * 2.6 14.1 16:1 n-7 7.0 f 6.5 0.9 f 1.2 0.2 16:ln-5 0.4 0.2 0.3 î 0.1 0.3 16:2n-6 0.2 f 0.1 nd nd 16:2n-4 0.4 f 0.2 0.9 f 0.5 0.1 17:O 2.8 i 0.9 1.6 f 0.5 1 .O 16:3n-4 0.5 i 0.2 nd 0.3 16:ln-3 0.5 I 0.5 0.2 f 0.2 0.4 16:4n-1 1.1 f 1.1 1.7 f 1.0 0.6 18:O 8.1 f 1.8 11.0 1.9 7.4 18:ln-9 3.9 f 0.8 10.3 f 5.1 1.8 18:ln-7' 2.8 f 0.7 5.5 î 1 .3 1.5 18:1 n-5 0.4 i 0.3 0.6 f 0.4 0.1 18:2n-6 1.7 f 0.2 1.2 f 0.8 0.5 18:Zn-4" 0.3 i 0.2 2.5 f 1.3 0-3 18:3n-6 0.1 I 0.1 0.2 f 0.1 O. 1 18:3n-3 0.8 f 0.1 0.6 f 0.2 0.4 18:4n-3 0.6 f 0.2 0.4 f 0.1 0.3 18:4n-1' 0.2 i 0.2 2.7 f 1.7 0.4 20:0* 0.4 f 0.2 1.1 f 0.2 2.1 20:1 n-11 0.4 f 0.1 nd nd 20: 1 n-9 0.9 0.7 1.4 f 0.4 0.3 20:l n-7* 0.3 f 0.1 0-8 î 0.2 0 .5 20:l n-5 0.1 f 0.2 0.5 f 0.9 nd 20:2n-9 0.3 f 0.4 0.4 f 0.4 0.6 20:2n-6' 0.2 f 0.0 2.6 f 1.6 0.3 20:3n-6 0.8 I: 0.4 0.9 f 0.3 3.0 20:4n-6 0.7 î 0.1 0.5 f 0.3 2.1 20:3n-3 0.1 f 0.0 nd 0 -4 20:4n-3 0.5 f 0.2 0.3 f 0.1 0 .6 UNK nd 2.5 f 1.5 nd 20:Sn-3' 9.3 f 0.5 2.2 * 0.5 2.5 22:O' 1.2 f 0.5 3.6 f 1.4 5.0 22:1 n-11 0.1 f 0.1 0.3 f 0.1 t r 22:1 n-9 0.2 f 0.3 0.2 i 0.2 0.4

22:Sn-6 1.3 f 0.5 1.6 0.3 1 .1 22:ln-3' 0.3 i 0.5 2.4 f 1.3 1.8 22:Sn-3 0.3 f 0.3 nd 0.6 24: 1 n-11 0.6 f 0.4 1.5 f 0.6 2.6 22:6n-3 26.9 f 9.4 20.5 f 1.6 38.4 24: 1 n-9 0.4 f 0.3 nd 1.3 26:O' 0.4 f 0.2 1.4 f 0.3 1.7 C SAT' 33.7 f 2.2 28.6 f 1.5 31.8 C m 18.1 f 7.9 22.4 f 7.8 9.5 C PüFA 47.2 f 9.8 42.0 f 7.1 56.0 C n-3 39.5 f 10.5 26.5 f 1.9 45.5 C n-6' 5.0 i 0.7 7.3 f 1.2 8.3 D HNEPA* 2.9 f 1.0 9.9 I 3.4 15.7 C UNK* t r 5.5 f 2.9 0 -2
Results are qressed as % of total fatty adds and represent means and standard deviations of 3 replicates except for 6OC which is the mean of 2 replicattw. Unknomis ~ 1 % not included in table. Abbreviations; nd= not detected, tr= trace (<O.OS%), SAT= saturates, MONOc monounsatutates, PUFA= polyunsaturates, UNlG unlaiomis, D W dooosahexaenoic acid, €PA= eicosapentaenoic acid.

0lSCUSSl0N
Effects of Temperature on Copepod Lipids
As the temperature decreased from 20 to 6OC, the percentage
of saturated fatty acids in Amonardia decreased from about one half
to one third. In Tisbe, the saturated fatty A acids were slightly, but
significantly higher at 20°C than et lS°C. but slightly higher again
at 6OC. This suggests that there was either more desaturase
activity, or less de norio synthesis of the saturated fatty acids a t
the lower ternperatures, particularly in Amonardia. There were
slightly higher levels of PUFA in Amonardia at 6OC than at 20°C, and
in Tisbe at 6OC than at 20°C (significance not attributed). At the
intermediate temperature of lS°C, the PUFAs were at their lowest
level in Arnonardia and in Tisbe. The monounsaturated fatty acids
were at their highest levels at 15OC in both Amonardia and Tisbe.
perhaps making up for the lower levels of PUFAs at th is
temperature. Thus, the total amount of PUFAs were highest in both
copepods at 6OC relative to 15 and 20°C. In Amonardia at 6OC, th is
increase in the level of PUFAs was at the expense of the saturated
fatty acids. Thus, these copepods seem to be undergoing

homeoviscous adaptation at 6OC to maintain a constant membrane
fluidity at this lower temperature.
The EFAs also exhibited distinct temperature responses i n
Amonanlia and Tisbe. The EFAs, 20:5n-3 and 22:6n-3, showed the
sarne general trend. In both harpactlcoids, except for 20:5n-3 in
Tisbe, the EFA occurred at higher levels at 20°C, decreased at 15°C
and then increased at 6OC. From 15 to B°C, which aie within the
range that the copepod would naturally encounter in the collection
site at Halifax Harbor, the levels of 22:6n-3 and 20:5n-3
dramatically increased with decreasing temperature for both
harpacticoids. This increase in 20:5n-3 and 22:6n-3 was also
observed for f reshwater copepods at lower temperatures (Farkas,
1979). There are some possible explanations for the increase in n-3
EFA a? 20°C compared to 15OC. The most probable explanation f O r
the increase in n-3 EFAs at 20°C, is that the phospholipid levels
increased over the neutral lipids as a proportion of the total lipid.
The stress of the abnormally high temperature of 20°C cornbined
with a higher metabolic rate could release depot lipid reserves
(Miliou and Moraitou-Apostopoulou, 1991 c; Pollero et al., 1 99 1 ).
There were no significant differences in the total lipid per copepod

for Amonardia at the three temperatures. At this highest
temperature, a higher proportion of females undergoing I i pi d
analysis may contain egg sacs, so that the total lipid per individual
copepod remains fairly constant, but more of it may be phospholipid.
There may also be selective catabolism of the monounsaturates,
such as 18:ln-9, to meet the higher energy demands at the higher
tem peratures. In Amonardia 1 8: 1 n-9 decreased 5-fold and in Tisbe
almost 3-fold as the temperature increased from 15 to 20°C (both of
these decreases are significant at pSO.10). This large decrease in
the monounsaturates, especially in Amonardia, would help to make
the n-3 EFAs more prominent in the lipid. Some or al1 of these
factors might explain the increase in n-3 EFAs from 15 to 20°C.
The EFA, 20:4n-6, generally followed the expected trend f O r
PUFAs, increasing steadily as the temperature decreased from 2 0
to 15 to 6OC in Amonardia and, in Tisbe, remaining nearly constant
at 20 and 15°C while increasing dramatically at 6OC. The increase
in this PUFA at the lower temperatures may be another example of
homeoviscous adaptation in the copepods.
For both harpacticoid copepods, the saturated fatty acid, 1 6:O,
was twice as high at 20°C compared with 15OC. although it remained

neai constant between 15 and 6°C. The high levels of this saturated
fatty acid may help maintain cell membranes at the proper liquid-
crystalline state at 20°C. The level of 16:O at 20°C is probably
achieved for the most part by de novo synthesis. The drop i n
monounsaturates frorn 15 to 20°C might indicate that they are the
primary energy source at 20°C, freeing up 16:O to be incorporated
into the phospholipid bilayer.
The main explanation given above for the increased levels of n-
3 PUFA at the highest temperature of 20°C was that the neutral
lipids were being bumed off du8 to increased metabolic rate or
stress, thereby decreasing the levels of saturated fatty acids and
increasing the phospholipid charader of the copepods' lipids. The
explanation for the increased levels of n-3 PUFA at the lowest
temperature of 6OC was that these decreased the viscosity o f
membranes by increasing the level of unsaturated fatty acids in the
phospholipid bilayer. To test these hypotheses, fatty acid analyses
would have to be performed on the neutral and phospholipid fractions
of the copepod lipids to detemine if the changes with respect t o
temperature in fatty acid composition occur in the membrane or the
depot lipids. If the total phospholipid was further divided into

classes. an increase in the levels of phosphatidylethanolamine a t
lower temperatures might be predicted. Phosphatidylethanolamine
in poikilothermic animals tends to be the most unsaturated of the
lipid classes, and increases in this phospholipid have been observed
in cold-acclimated f ish (Hazel, 1 979) and crustaceans (Chapelle,
1986). Evaluating the fatty acid composition of the various l ipid
classes in th8 copepods is n0t essential for the purpose of marine
fish larval nutrition. Suwival and growth in first-feeding marine
fish lantae c m be related to the EFA composition of the total l i p i d
in the live food (Watanabe, 1993).
Nutritional Implications for Marine Fish Larvae
The lowest temperature (6OC) gave the highest amounts o f
22:6n-3 for both harpacticoids. However, there is a trade-off in
decreasing the culture temperature. Loweiing the temperatu re
lowers the biomass of copepods available from a mass culture
system by decreasing egg production and growth rate and by
increasing generation times (Miliou and Moraitou-Apostolopoulou,
1991a). The nutritional value of the individual copepods for the
marine fish larvae will be increased, but there will be fewer
copepods for feeding to th8 larvae. The copepods had very high

levels of 22:6n-3 and high DHA to EPA ratios at 2Q°C, so that the
EFA value of the food would not be limiting for marine fish larvae
fed copepodg cultured at high temperatures. Therefore, it i s
piobably not necessary to lowei temperature to increase the
harpacticoid copepods' nutritional value.
As the temperature decreased from 20 to 6OC, the amounts of
arachidonic acid (20:4n-6) increased in both harpacticoids over 4 - fold for Amonardia and 3-fold for Tisbe (Tables 5 and 6). The
biggest advantage in ternis of increasing the EFA composition of the
live food with lowered temperature, lies with increasing the
amounts of arachidonic acid. However, requirement of marine fis h
lawae for 20:4n-6 is relatively low when compared to that for the
n-3 fatty acids 20:5n-3 and 22:6n-3, as is evidenced by the l ow
total amount of 20:4n-6 in the laivae (Castell et al., 1994).

LlPlD COMPOSITION OF ALTERNATlVE LlVE FOOD SPECIES
I r n W O N
The lipid compositions of some other native potential live food
organisms were exarnined in addition to those of the harpacticoid
copepods, Tisbe and Amonardia. Wild zooplankton (consisti ng
primarily of calanoid copepods), useâ in lanral marine fish feeding
trials at the St. Andrews Biologicai Station, and the trochophores of
a local mussel species. M'lus edulis, were the alternative l ive
food species evaluated. Cornparisons were then made with the
harpacticoid copepods, to determine which live foods had the most
appropriate EFA compositions for marine fish lanrae. It was
expected that th8 harpacticoid copepods would have the highest
levels of 22:6n-3 and the highest DHA to €PA ratios because of their
extrernely high desaturase activity, thus meking them the most
attractive live food for marine fish larvae, in ternis of fatty acid
composition. The wild zooplankton tend to inhabit environments rich
in n-3 EFAs (20:5n-3 and 22:6n-3) and incorporate high levels of
these EFAs in their lipid. The DHA to EPA ratio in the lipids of most
north Atlantic calanoid copepod species is usually around 1. although
this number cm Vary widely with respect to species and

environmental parameters (Kattner et al., 1981). Shellfish tend t O
have higher levels of 20:5n-3 and DHA to €PA ratios of less than 1
(Kluytmans et al., 1985). The trochophores of M. eduM may not have
as high an €FA value as the copepods as Iive food for marine f is h
larvae, because of this low DHA to €PA ratio.
MATERlALS AND METHODS
Freshly collected local (Nova Scotian) mussels were held in
buckets at ambient seawater temperature (6OC). They were then
isolated in trays containing filtered (10 pm), UV-treated sea water
and brought up to room temperature (lQ°C) to trigger a spawning.
The spetm and eggs of an individual male and fernale mussel were
cornbined in a 20-L bucket and after 24 h the fertilized eggs and
trochophores were captured on 40 jm screen and suction f i l te red ,
using a Buchner funnel, on Whatman no. 1 filter paper. Fatty acid
compositions were detemined for three replicates in the same
manner as for the copepods (p. 19).
The lipid composition of wild zooplankton taken f rom
Passamaquoddy Bay, Na B., in a tow (mesh site = 84 pm) on 27 J uly ,
1995 was also analyzed. The sample consisted of freeze-dried
calanoid copepods (Eurytemora sp. and Acarlia sp.) which were used

as live food for marine fish larvae at St. Andrews Biological Station.
Three replicate samples were weighed and anaiyzed for lipids using
the methods described for copepods (p. 19).
Differences in the relative amount of EFAs in the harpacticoid
copepods, zooplankton and mussel trochophore larvae were compared
using Tukey's HSD pairwise cornparison test.
RESULTS
The fatty acid composition of 24 h-old fertilized eggs and
trochophores of the mussel Mytilus edulis were determined (Table
7). The trochophores contained relatively large amounts of the €FA
20:5n-3 and 22:6n-3, but had a low DHA to €PA ratio (0.37). They
also contained 1.3% of the €FA 20:4n-6. The 16 carbon fatty acids
16:0 (18.5%) and 16:ln-7 (10.6%) comptised the greatest portions of
the saturated and monunsaturated lipid compositions, respectively.
The total PUFA content of the trochophores was extremely high
(alrnost half of the total FA), and approximately equal to the sum of
the saturates and monounsaturates combined. It was predominantly
made up of the n-3 family of fatty acids.
The lipid composition of wild zooplanMon captured i n
Passamaquoddy Bay is shown in Table 8. The percentage of total

Table 7. The fatty acid composition of 24 hour old fertilized eggs and trochophores of the blue mussel, Mytilus eduk
FaW acÏd Percentaae 14:O 1 .O I 0.9 15:O 0.1 f 0.1 16:O 18.5 f 1.0 16:ln-7 10.6 f 0.5 .
7-Me 16 0.1 f 0.0 16:l n-5 0.5 f 0.0 16:Zn-6 0.4 f 0.1 16:2n-4 0.5 f 0.1 17:O 0.3 f 0.0 16:3n-4 0.2 f 0.2 16311-3 0.4 f 0.1 16:4n-3 0.1 f 0.0 16:4n-1 4.8 f 0.6 18:O 2.7 f 0.4 1811 n-9 1.5 f 2.0 18:l n-7 3-1 f 0.6 18:1 n-5 0.2 f 0.0 18:Zn-6 1.2 f 0.2 18:2n-4 0.7 f 0.2 18:3n-6 0-1 f 0.0 18:3n-4 0.3 f 0.0 18:3n-3 1 -6 f 0.3 18:4n-3 2-7 f 0.5 18:4n-1 0.4 f 0.1 20 :O o. 1 f 0.0 20:1 n-11 0-7 f 0.6 20:l n-9 1.9 f 0.6 20:1 n-7 1.6 f 0.3 UNK 1.4 f 0.1 20:2n-6 0-6 f 0.0 20:3n-6 O. 1 f 0.0 20:4n-6 1.3 f O -8 20:3n-3 0. 1 f 0.0 20:4n-3 0.5 I 0.0 20:5n-3 24.0 f 3-5 22:o 0.2 * 0.0 22:1 11-31 O. 1 f 0.0 22:1 n-9 0.3 k 0-2 22:1 n-7 0.4 f 0.3 21 :5n-3 0.9 I O. 1 22:4n-6 0.2 f 0.0 22:5n-6 O. 1 f 0.0 22:4n-3 O. 1 f 0.0 22:5n-3 1.3 f 0.1 24:O O. 1 f 0.0

22:6n-3 8.8 f 1 ,O 24: 1 n-1 1 0.2 f O. 1 C m 23.5 I 1.2 CMoNo 20.6 f 2.2 C PUFA 46.7 f 4.1 C n-3 40.6 f 5.1 C n-6 4.0 f 0.7 D W A 0.4 i 0.0 C UNK 3.8 f 1.8
Resulfs are expraned as 96 of total fuüy adds and represent means and standard deviations of 3 mplicates. Unknowns 4% not induded in table, Abbreviations; ncb not detected, tr= trace (~0.05%). ?-Me= 7-methyl, DMA= dimeîhyl anteiw, SAT= saturates, monounsaturates, PUFA= polyunsaturates, UNlb unknowns, D H k docosahexaenoic acid, €PA= eicosapentaenoic acid.
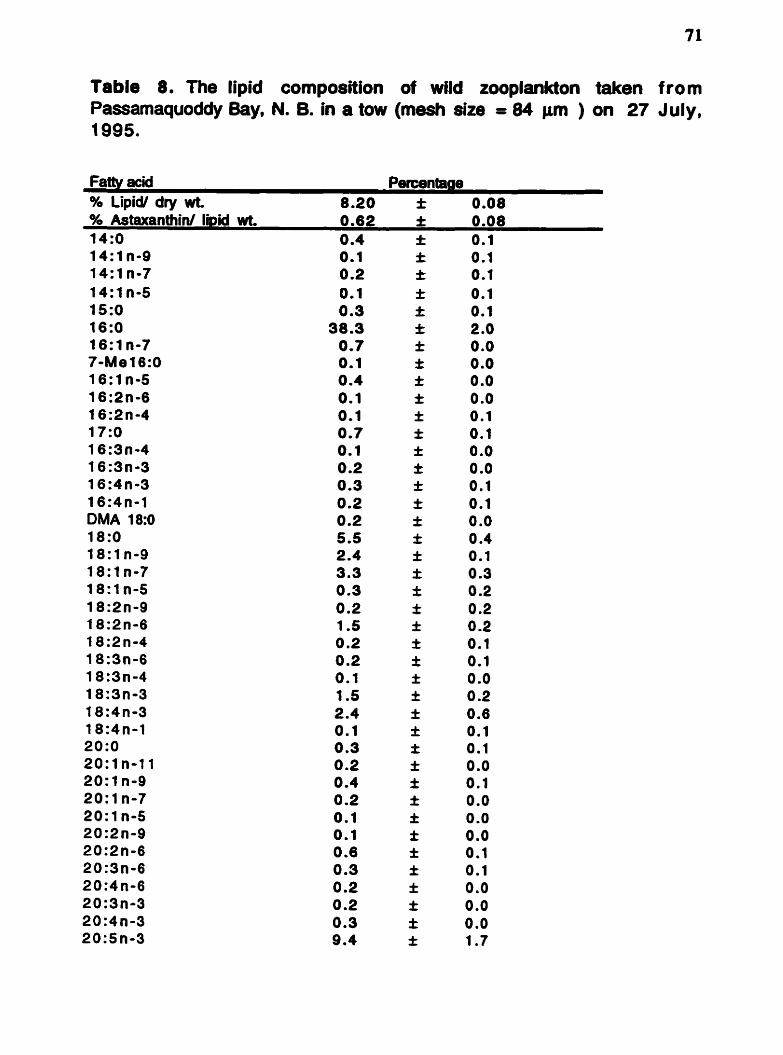
Table 8. The lipid composition of wild zooplankton taken f rom Passamaquoddy Bay, N. B. in a tow (mesh size = 84 pm ) on 27 July, 1995.
Fatty acid Percentage % Lipidl dry wt 8 -20 * 0.08 % Astaxanthinl lbid wt, 0.62 f 0.08 14:O 0.4 l 4 : l n-9 14: 1 n-7 l 4 : l n-5 l5:O l6:O l 6 : l n-7 7-Mû16:O l 6 : l n-5 16:2n-6 16:2n-4 17:O 16:3n-4 16:3n-3 16:4n-3 16:4n-1 DMA 18:O 18:O 18:ln-9 18:ln-7 l 8 : l n-5 18:2n-9 18:2n-6 18:2n-4 18:3n-6 18:3n-4 18:3n-3 18:4n-3 18:4n-1 20:o 20: 1 n-11 20: 1 n-9 20:1 n-7 20:1 n-5 20:2n-9 20:Zn-6 20:3n-6 20:4n-6 20:3n-3 20:4n-3 20:5n-3

22:o 0.2 f O -2 22:1 n-11 0.1 f 0.0 22:1 n-9 0.5 f 0.0 22:1 n-7 0.1 f 0.0 21 :Sn-3 0.2 f 0.1 22:4n-6 0.1 f 0.1 22:s n-6 0.2 f 0.0 22:4n-3 0.2 f 0.1 22:5n-3 0.5 f 0.2 24:l n-11 0.2 f 0 .2 22:6n-3 16.6 f 2.9 24:1 n-9 4.1 f 0.7 26:O 0.4 f 0.2 CmT 46.2 f 1.6 CMaw 12.8 f O -7 C PUFA 35.3 f 4.7 C n-3 31.5 f 4.7 C ri-6 2.9 f 0 .2 D M P A 1.8 f 0 .2 t UNK 5.7 f 5-7
Results are expressed as % of total fatty ad& and reprewnt means and standard deviations of 3 replicates. Unknowns t1% not induded in taMe, Abbreviations; nd= not detecteâ, tr= trace (<O.OS%), 74- 7-methyl, D M k dimethyl anteisa, SAT= saturates, monounsaturates, PUFA= poiyunsatutates, UNb unknowns, DHAt docosahexaenaic acid, EPAs eicosapentaenoic acid.

lipid was low for calanoid copepods at 8.2% of the dry weight. The
zooplankton sample was freeze-dried for storage which may cause a
lower lipid yield when extracted (Dunstan et al., 1093). The
carotenoid pigment astaxanthin made up 0.6% of the lipid weight
(Appendix 6). The EFAs, 20:5n-3 and 22:6n-3, were present in
relatively high arnounts. The DHA to €PA ratio was also hlgh at 1.8.
The €FA, 20:4n-6, was present in very low quantities. The
zooplankton contained large amounts of saturates (almost half O f
the total FA), somewhat less polyunsaturates and much less
monounsaturates. Most of the saturated fatty acid was 16:O.
The fatty acid compositions of the four live food species (the
two harpacticoids, the wild zooplankton, and the 24 h old M. edulis
trochophores) are compared in Table 9. The 18-carbon fatty acids
18:2n-6 (precursor for 20:4n-6) and 18:3n-3 (precursor for 20:Sn-3
and 22:6n-3) were relatively highest in Amonardia. The percentage
of 20:4n-6 was highest in the trochophores and lowest in the w i I d
zooplankton. Significantly larger quantities of 20:Sn-3 occured i n
mussel larvae, followed by the wild zooplanldon, with the
harpacticoids containing comparatively the lowest percentages.
Tisbe, however, contained the highest percentage of 22:6n-3, not
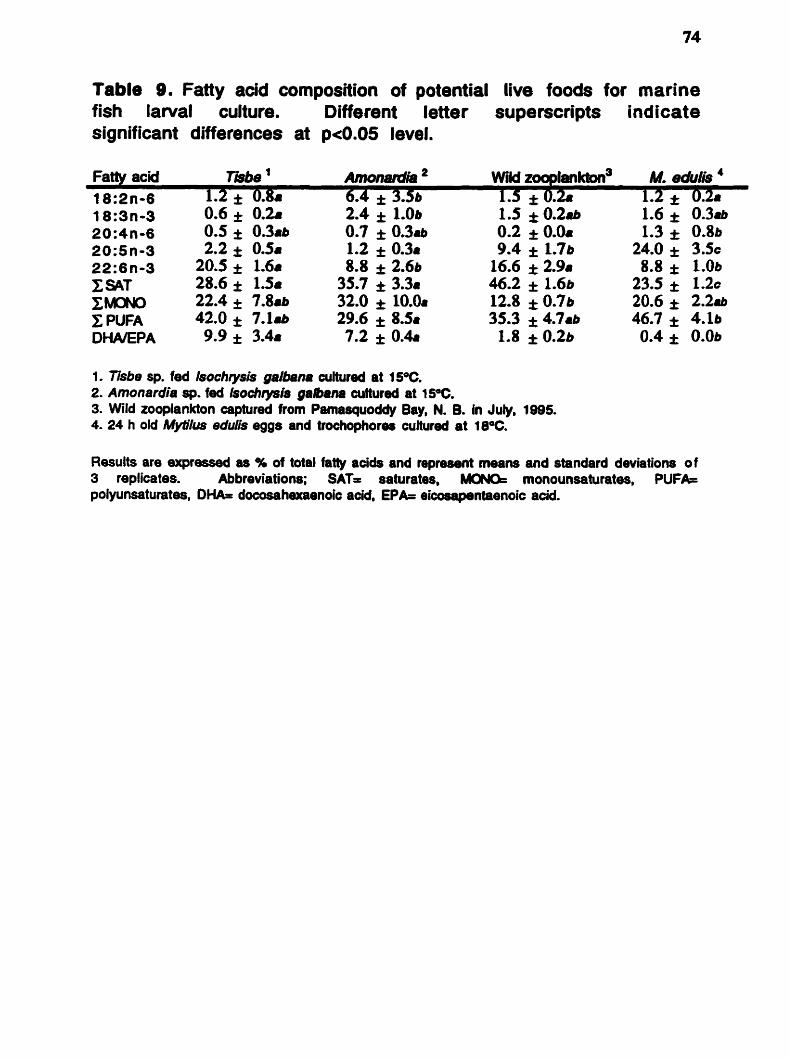
Table 9. Fatty add composition of potential live foods for marine fish larval culture. Different letter superscripts indicate significant differences at pe0.05 level.
Fatty acid riabe ' Amondia wild wopl-kton3 M. 18:2n-6 1.2 * 0.84 6.4 * 3.56 1-5 i 0.û 1.2 * 0.2.
1. Tisbe sp. fed lsochrysis galbana cultured at 1 SOC. 2. Amonardia sp. fed Isochrysis flaibana culturd at 15%. 3. Wild zooplankton capturd fiom Pamcuguoddy Bay, N. B. in July. 1895. 4. 24 h old Mytilus eduiis eggs and trochophores cultured at 18OC.
Results are wressed as % of total fatly acids and represent means and standard deviations of 3 replicates. Abbreviations; SAT= saturates, MONOc monounsaturates, PUFh polyunsaturates, D b docosahexaenoic acid. EPA= ricosapentaenoic acid.

significantly greater than the wild zooplankton, but significantly
more than was present in Amonardla and M. edulis. Not suprisingly,
the harpacticoids had significantly higher DHA to EPA ratios when
compared wïth the other live food specïes. The M. edulis larvae had
a lower DHA to EPA ratio than the wild zooplankton (0.37 vs. 1.77).
DlSCUSSiûN
Lipid Composition of Alternative Live Food Species
The larvae of M edulis had a significantly higher level of
20:Sn-3 when compared to the copepod species. The lower levels of
20:5n-3 and higher levels of 22:6n-3 in the copepods could be an
adaptation to their more motile lifestyle when compared to that of
M. edulis (suggestion by Dr. R. G. Ackman). The fatty acid 22:6n-3 i s
present at high levels in the nervous systems of motile animals. The
functioning of the nervous systern is based on the activity of ion
channels and of receptors situated within the lipid bilayer. The
proper functioning of these proteins depends on a bilayer wit h
certain physical properties (i.e., fluidity and permeability) which the
n-3 PUFA confer, in particular 22:6n-3 (Dumont et al., 1992). The
mussel trochophore larvae were the only potential live food species
we studied which had a DHA to EPA ratio of less than 1. This fa tt y

acid composition is typical of other marine bivalve larvae (Marty e t
al., 1992). Unlike the copepods. M edulis larvae do not use
desaturase and elongase enzymes to convert most of their 20:5n-3
into 22:6n-3.
Both 77sh and Amonardia had significantly highei DHA to EPA
ratios compared to the wild zooplankton. The latter (consist ing
almost entirely of Acartia sp. and Eurytemora sp.) captured i n
Passamaquoddy Bay during the month of July, contained very low
lipid levels (about 896). These copepods do not contain an oil sac,
thus they do not accumulate the large lipid reserves of some other
calanoid species. The phyîoplankton concentrations in July also tend
to be low, so that the wild zooplankton will not have a high energy
intake, or b8 able to accumulate high levels of lipid (Sargent and
Henderson, 1 986).
The lipid compositions of the two harpacticoid copepods
differed considerably. Tlsbe seems to have a greater desaturating
capacity than Amonardia. When both species were fed I. galbana a t
1 5OC, Tisbe produced almost twice as much of the n-3 EFAs, 20:5n-
3 and 22:6n-3. Amonardia, however, contained signif icantl y more
18C precursor fatty acid 18:3nB (Table 9). This suggests that Tisbe

contains more of the desaturases necesssary for this conversion
( g . 1 ) . Amonardia also had a signif icantly higher level of 1 8:2n-6,
but the levels of 20:4n-6 were not significantly different from
those of Tisbe, a further indication of less active A-6 and A - 5
desaturase. Also, the overall levels of PUFA were much higher i n
Tisbe, compared to Amonardia, whereas, Amonanfia contained much
higher levels of the saturates and monounsaturates.
Nutritional Implications for Manne Fish Lawae
The nutritional value of the wild zooplankton for marine f ish
larvae will Vary considerably with season. Our zooplankton sample
taken in midsummer was lipid poor (about 8% dry wt.). Amounts of
lipid reach their lowest levels in July for North Atlantic calanoid
copepods. Also, the fatty acid composition (particularly of the
neutral lipids) would be quite different, immediately after or during
a phytoplankton bloom (Kattner et al., 1981 ; Norrbin et al., 1 990).
The wild zooplankton have a much higher caloric and nutritional
value for mafine fish larvae dun'ng or directly after the spring or
fall ph ytoplankton bfoorns, when some calanoid species accumulate
wax esters, which can make up to 70% of their dry weight (Sargent
and Henderson, 1986). The advantage of using cultured animals

versus wild moplankton is that the cultured animals can be
manipulated to give a consistently high lipid and EFA composition.
The large arnounts of both long-drain EFAs, 20:5n-3 and 22:6n-
3 and their high D M to EPA ratio indicate that these calanoid
copepoûs have high EFA value for marine fish lawae. This high EFA
value is one reason why field-captured zooplankton aie widely and
successfully used for the culture of cold-water manne fish larvae
(aga Kuhlman et al., 1981 ; Boxsspen et al., 1990; Shields, 1992).
The value of trochophores of the mussel, Mytilus eduiis, as an
alternate [ive feed for marine fish lawae was evaluated by Howell
(1973) and was found not to support growth in lawal plaice,
Pleuronectes platessa, and lemon sole, Solea solea, when used as
the only food. However, when the trochophores were offered in
combination with rotifers to the lemon sole l a ~ a e at the early
first-feeding stages, a survival rate of 32% and an average growth
increase from 5.8 to 8.1 mm length for the first 30 days post-hatch
was obtained. Ch days 12 and 20 post-hatch, al1 the first-feeding
larvae contained only trochophores in their guts. This demonstrates
their preference for trochophores over rotifers during the early
first-feeding stage (Howell, 1971).

The advantage of using mussel lawae as live food is the ease
of culture. Simply raise the culture temperatures of adult male and
fernale mussels and they will spawn. Combine the spem and eggs of
the mussels and within 24 h trochophore larvae are produced.
Trochophores may not have as high an €FA value as copepods
for manne fish larvae because of their lower DHA to €PA ratio.
However, the trochophores do contain much higher levels of the
long-chain EFAs, 20:5n-3 and 22:8n-3, than do the traditional l ive
food organisms, bfine shrimp and rotifers. The rnwsel trochophores
also contain a higher proportion of the EFA 20:4n-6 than do the other
alternative live food organisms analyzed.
Live foods, which contain either the most 22:6n-3 or the
higher ratio of 22:6n-3 to 20:Sn-3 tend to have the highest EFA value
for cold-water, marine fish larvae (Bell et al., 1985; Watanabe,
1993). The two harpacticoid copepod species studied had the
highest ratios of DHA to EPA, when cotnpared with wild zooplankton
or mussel lawae. Tisbe hed the highest percentage of 22:6n-3 of al1
the live foods that we examined, and it contained much more of th is
EFA than the traditional live food organisms such as the brine
shrimp and rotifers. Tisbe is able to produce large amounts o f

22:6n-3 and maintain consistently high DHA to EPA ratios (2.0 t O
3.4) despite the €FA composition of its diet, due to its high levels O f
desaturase activity . This 22:6n-3 producing capability toget h er
with its ability to be mass produceci at high densities make the
harpacticoid Tlsbe, and to a lesser extent Amonardia, very
attractive potential live food organisms for marine fish l arval
culture.

Chapter 2
Free Amino Acid Composition of Harpacticoid
Copepodr and lmplicationr on their
Nutritional Value for Marine Fish Larvae

Introduction
Free Amino Acid Metabolism in Copepods
Marine invertebrates, crustaceans especially, accumulate large
reserves of free amim acids (concentrations around 300-500 mM
are typical) for use as osmolytes to help maintain their intracellular
osmotic stasis relative to that of their seawater medium (Pierce,
1981; Fyhn et al., 1993). Theiefore, changing the salinity of the
seawater medium will effect a diange in the free amino acid pool.
With exposure to reduced salinities, the free amino acid levels are
reduced in both calanoid (Farrner and Reeve, 1978; Fyhn et al., 1 993)
and harpacticoid (Goolish and Burton, 1 989) copepods. The f re s h l y-
caught calanoid copepod AcaHia tonsa starved during sa l i n i t y
acclimation did not increase its free amino acid pool at salinities
above 100% seawater, perhaps du8 to a lack of nutritional reserves
needed for the extra synthesis of amino acids at the higher
salinities (Farmer and Reeve, 1 978). The calanoid Calanus
finnmarchicus, however, did increase its free amino acids as
salinity increased from 15 to 45 ppt (Fyhn et al., 1993). Jeffries
and Alzara (1969) also obsewed an increase in the total amount o f
free arnino acids as salinity increased, when comparing calanoid

copepod species which live in f reshwater environments wit h t hose
from estuarine and marine environments. The harpacticoid copepod,
Tigriopus, inhabits splash-pools of the intertidal zone and
experiences a wide range of salinities, being active in water from O
to 80 ppt salinity. Tisbe sp. occupies the sublittoral zone or t ide
pools that are regularly inundated by seawater and so experîences a
narrower range of salinity in its environment. It is only active f rom
30 to 45 ppt salinity (Finney, 1979), therefore, one might expect a
larger free amino acid pool in the euryhaline Timpus when
compared to Tisbe, particularly when acclimated to higher
salinities. Tigriopus californicus was the subject of a number o f
studies on free amino acids versus salinity (Burton, 1986; Burton
and Feldman, 1982; Goolish and Burton, 1989). The non-essential
amino acids proline, glycine and alanine, in decreasing order of
concentration are the dominant contributors to the free amino acid
pool when Tm californicus is put under hyperosmotic stress. These
high free amino acid levels drop rapidly (within 5 h proline dropped
40%), when the copepods are transfened from water of 68 to 34 ppt
salinity (Burton and Feldman, 1982). Therefore, the copepods must
be ingested within a short period of time after exposure to the

hyperosmotic medium. if the marine fish larvae are to benefit
maximally from the increased level of non-essential free amino
acids of the copepods.
The dominant contributor to the free amino acid pool differs
between the harpacticoids and calanoids which have been acclimated
to 100% seawater. Glycine dominates the free amino acid pool in the
marine calanoids (Fyhn et al., 1993; Famer and Reeve, 1978),
whereas proline dominates the free amino acid pool in the
harpacticoid copepod T 'opus (Burton and Feldman, 1 982).
Crustaceans, when injected with radiolabelled acetate, were
shown to lack the enzymes needed to synthesize the same 10 amino
acids which are essential for marine fish larvae (Cowey and Forster,
1971). Therefore, any increase in these 10 essential amino acids as
a response to either salinity or diet is due to increased
incorporation from the diet and not de novo synthesis.
Free Amino Acid Metabolism in Manne Fish Larvae
Manne fish larvae also require certain amino acids. Halver
(1 957a, b) discovered that chinook salenon Oncorhynchus fsha wytscha
required 10 essential amino acids (phenylalanine, tryptophan,
histidine, arginine, lysine, valine, isoleucine, leucine, threonine and

methionine). These amino acids were omitted from test diets and
growth responses were compared with those of complete diets. No
significant growth was obtained on diets devoid of any one of these
amino acids. The marine f latf ish plaice Pleuronectes platessa end
sole Solea solea were ako unable to synthesize these amino acids
from radiolabelled glucose. Thus, these amino acids are also thought
to be essential for marine fish (Cowey et al., 1970). nierefore. a
higher level of essential free amino adds will increase the
nutritional value of the live food for manne fish larvae.
Free amino acids have a special role in the eggs and early
larvae of marine fish. Marine fish eggs and yolk-sac l ame contain
much higher amounts of free amino acids compared to the adolt
tissues. The large pool of free amino acids is contained almost
exclusively in the yolk-sac cornpartment of the fish lawae. The free
amino acid pool is depleted during development and reaches l ow
levels at first feeding (Rsnnestad and Fyhn, 1993). As this free
amino acid level decreases with development, there is a concomitant
increase in ammonia excietion and oxygen consurnption, suggesting
that the free amino acids are being aerobically catabolized as a
source of energy (Fyhn and Sergistad, 1987; Finn and Fyhn, 1993).

First-feeding mafine fish lanrae have a relatively undifferentiat ed
digestive system and low proteolytic capability (Hjelmeland et al..
1993; Pedersen et al., 1987). Dietary proteins will, for the most
part, be unuseable as an energy source by the larvae at this stage.
Also their supply of free amino acids from th8 yolk sac will have
been largely depleted. Thus, at the initiation of exogenous feeding,
the marine fish lawae may require large amounts of free amino
acids. As noted above, marine zooplankton typical l y accum ulate
large amounts of free amino acids which play a dominant role in
rnaintaining the osmotic balance of their intracellular fluids. Fyhn
(1989) suggested that, in choosing a live food organism, the free
amino acid pool of the prey should match the free amino acid pool
consumed by the fish embryo during endogenous feeding.
Experimental Objectives
The purpose of this study was to determine if the diet fed t O
the harpacticoid copepod, Tisbe sp., affects the size and
composition of the free amino acid pool in the copepod. The t wo
diets used in the copepod feeding trial, the alga lsochrpis galbana
(clone T-iso) and bakersg yeast, were chosen because of their large
differences in amino acid composition (Chau et al., 1967; Ren et al.,

1993). It is expected that the proportions of nonessential free
amino acids in the copepod fed the two diets should remain f ai rl y
uniform with nonessential free amino acids such as glycine, tau rine,
proline and alanine dominating. However, the increased amount of
essential amino acids available in the alga 1. galbana (T-iso)
compared with yeast, should increase the level of essential free
amino acids in the copepods fed T h D An increase in the size of the
free amino acid pool might increase growth in fish lawae R the free
arnino acids remain an important source of fuel and precursors f O r
protein synthesis after yolk sac absorption (Fyhn et al., 1993). Large
variations in the total free amino acid pool, foi the copepods fed the
two diets should be expected, considering the dynamic nature of
these metabolites (Stnrttrup et al., 1995). A higher level of
essential free arnino acids in the copepods will also increase the
nutritional value of th8 live food (Cowey et al., 1970).
The free amino acid composition of wild zooplankton (calanoid
copepods) from a tow in Passamaquoddy Bay, Ne B. (27 July, 1995)
was also compared with that of the harpacticoid copepod Tisbe. The
free amino acid compositions should be similar for the two copepod

88
classes with the non-essential amino adds glycine, proline and
alanine dominating (Farrner and Reeve, 1978).
The salinity tolerance of Tisbe sp. was also tested. It i s
expected that this copepod will be stenohaline as it lives in an
environment with a low variability in salinity (Finney, 1979).
Because of this stenohalinity Tisbe will not have the same potential
as Tigropius for increasing its free amino acid pool, and therefore
energy value, for marine fish lawae when subjected to h yperosmotic
stress (Burton and Feldman. 1982).
Materials and Methods
The free amino acid composition of an alga, lsochrysis galbana
(clone T-iso), a bakers' yeast diet, and of Ti- sp. fed these diets,
was evaluated. The alga, T-iso, was cultured and centiifuged using
the sarne methods described earlier for lipid analyses of aigae (p.
19). T-iso, was grown at 18°C on f/2 medium (Guillard and Ryther,
1962) and harvested by centrifugation while in th8 log phase at 8 d.
The copepods were cultured for over one generation on the yeest and
algal diets using the same culture setup as described for the lipid
study (p. 16). The copepods were screened through a 200 pm mesh
and subsequently transferred to jars containing fresh 1 0-prn

filtered, UV-treated sea water. They were then stanred for 24 h t o
allow for the clearance of dietary algae or yeast present in the gut
and rescreened to remove any accumulated waste products. The
copepods were then counted, suction filtered through Whatman no. 1
filter paper and rinsed with seawater.
Free amino acids were extracted from the copepods and t hei r
diets using boiling 80% ethanol. Two replicate samples of the
copepods containing approximately 500 individuals per sample we re
analyted. Three replicate samples of the diets consisting of an
average of 84.0 mg dry weight of bakers' yeast and 86.1 mg dry
weight of T-iso were used for free amino acid analysis. The dietary
and copepod samples were immersed in the boiling ethanol and
homogenized in 20-mL glass culture tubes using a polytron
(Brinkman Instruments). The homogenized samples were t hen
filtered through Whatman no. 1 filter paper and rinsed with excess
boiling ethanol. The ethanol extract from each sample was then used
for free amino acid analysis.
An emount of standard norleucine was added to the extract so
that area percent could be converted to pmoles of free amino acid
units. The standard norleucine peak area was approximately 10930%

of the largest peak area. The amount of norleudne to be added for a
given amount of sample was detemined by either the estimated dry
weight of the sample. or by preliminary analysis with various
dilutions of the sample. The extract was then evaporated to dryness
in a rotary flash evaporator (Haakebuchler Instruments). Samples
were redissolved in a citrate buffer (pH 2.2) and injected on the
automatic amino acid analyzer (Bedunan 119 CL), which was based
on the system developed by Moore et al. (1958). The presence of
glutamine in the yeast samples interfered with quantification of the
threonine and serine peaks. Therefore half of the yeast sample was
made to 2 M with trifluoroacetic acid and hydrolyzed at 1 OO°C f O i
1 5 min (sufficient to quantitatively destroy glutamine), after w h i ch
it was evaporated to dryness and dissolved in citrate buffer (pH 2.2)
for injection on the amino acid analyzer.
A cornparison between two free amino acid extraction
rnethods- the aqueous phase of the Bligh and Dyei (1 959) versus the
boiling 80% ethanolic extraction method- was conducted using a
hornogenized wild zooplankton sample (Appendix C). The 80% ethanol
method gave significantly higher extractions for 75% of the amino
acids detected in the zooplankton. Thus, the 80% ethanol method

was chosen over that of Bligh and Dyet (1959), for the extraction o f
free amino acid from the copepod samples.
The salinity tolerance of Tisbe was also tested using
different amounts of lnstant* O-' (Aquarium Systems) dissolved
in distilled water. Approxirnately 20 adult copepods were e~posed
to different salinities (20, 30, 40 and 54 ppt.) for 5 hours. The
copepods were then examined under a dissecting microscope (20X
magnificat ion) to determine percent survival . Results
The free amino acid composition of bakers' yeast and
Isoch~sis galbana (clone 1-iso) differed considerably (Table 1 O) .
The alga had a total free amino acid concentration greater than 3.5
times that of the yeast. The dominant free amino acids in the T-iso
sample were gamma aminobutyric acid (ca. 39 pmol/g dry W.) and
alanine (ca. 19 pmollg). Proline, taurine and arginine were the other
major free amino acids in T-iso (ca. 11 prnoVg). The dominant free
amino acid in the yeast was glutamic acid (ca. 25 pmoVg diy wt.).
Alanine (ca. 5 pmollg), aspartic acid (ca. 3 pmol/g) and proline (ca.
3 pmol/g) were the next largest contributors to the free amino acid
pool in the yeast.

The mol % free amino acid composition of the copepods was
very similar (Table 1 1). among diets (T-iso W. yeast) and copepods
(calanoid vs. harpacticoid). The wild zooplanîcton sample was made
up almost entirely of the calanoid spedes, Eurytemora sp. and
Acartia sp. Glycine was the dominant free amino acid in al1
copepod samples. It was highest in th8 wild zooplankton (43.1%),
followed by nsbe f8d yeast (38.0%) and T-iso (33.5%). In the w i l d
zooplankton, glycine was foiîowed in decreasing order, by taurine,
alanine, proline and arginine. In nsbe fed with yeast, glycine was
followed in decreasing order by alanine, taurine. arginine and
proline. In Tisbe fed with T-iso, glycine was followed by arginine,
taurine and alanine. Proline was present at a comparatively l o w
level in Tisbe fed with T-iso. Thus, the three copepod samples had
fairly similar patterns with respect to mol % free amino acid
composition. The nonessential free amino acid glycine dominated,
followed in lesser arnounts by alanine, taurine and proline (Tisbe
fed T-iso excepted). The essential free amino acids were present i n
much smaller amounts in the copepods, when compared with the
nonessential free amino acids. Arginine was the only essentiel free
amino acid which exceeded 5% of the total free amino acids. Tisbe

Table 1 O. Free amino acid composition of the alga Isochrysis gaibana (clone T-iso) and bakers' yeast used in copepod feeding trials. Results are expressed as pmol/ g dry wt. and represent means and standard deviations of three replicates.
Amino acid Yeast T-iso taurine nd 11.18 f 1.85 aspartic acid threonine* serine glutamic acid proline glycine alanine cysteine valine methionine* leucine* isoleucine* tyrosine phenylalanine* GABA histidine* lysine* arginine* Total EFAA % Total NEFAA % Total FAA
Abbreviations: nt#= mt deticted, GA&= gamma amino butyric acid. FAA= free amino acids, EFAA= essential free amim acids. NEFM= nonessential free mino acids. denotes individual essential free amino acids.
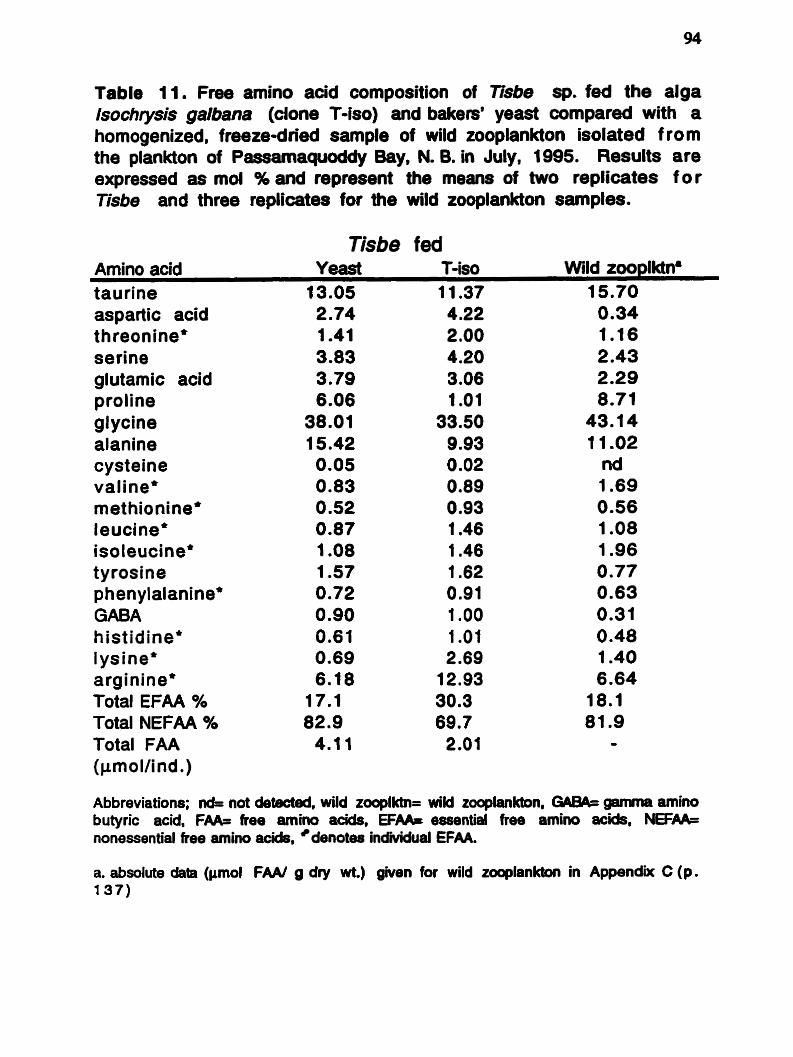
Table 11. Free amino acid composition of Tisbe sp. fed the alga Isochrysis galbana (clone T-iso) and bakers' yeast compared with a homogenized, freeze-dried sample of wild zooplankton isolated f ro m the plankton of Passamaquoddy Bay, N. B. in July, 1995. Results are expressed as moi % and represent th8 means of two replicates f O r Tisbe and three replicates for the wild zooplankton samples.
Tisbe fed Amino acid Yeast T-iso Wild zooplktn8 taurine 13.05 11.37 15.70 aspartic acid threonine* serine glutamic acid proline glycine alanine cysteine valine* methionine* leucine* isoleucine* tyrosine phenylalanine* GABA histidine' lysinet arginine* Total EFAA % Total NEFAA % Total FAA (pmol/ind.)
Abbreviations; mi= not detected. wild zooplktn= m'id zoaplankton, gamma amino butyric acid, FAA= free amino acida, €FM= essentiai free amim acids. NBAA- nonessentiai free amino acids, denotes individual €FM.
a. absolute data (pmol F M g dry W.) given for wild zocplankbn in Appendk C (p. 137)

fed T-iso contained th8 largest fraction of essential free amino
acids, with levels of the basic amino acids arginine, lysine and
histidine approximately double those in the other copepod samples.
The free arnino acid level per copepod was approximately
twice as high in Tisbe fed yeast as in Tisbe fed T-iso. The level of
total free amino acids in the homogenized, freeze-dried wi ld
zooplankton was expressed as pmoV g dry wt. and was not d i rectl y
corn pared.
The salinity tolerance of Tisbe tested using Instant Oceana
(Aquarium Systems) and was found to be fairly stenohaline. The
survival was 1ûô% at 30 and 40 ppt. after 5 h exposure. At 20 ppt,
survival was 7ï%, but the copepods were quite inactive and at 50
ppt, survival was only 29% after 5 h.
Discussion
The large differences in the free amino acid composition
between the yeast and alga used for th8 copepod feeding t r ia ls
should be expected considering that the orgmisms are classified i n
different kingdoms and inhabit diff erent environments. T-iso has a
much higher total free amino acid content than the yeast. The role
of free amino acids as osmotic effectors in marine algae was f i r s t

demonstrated for the red aiga Porptyn'dium purpureum (Gilles and
Pequeux, 1968). The total free amino acids increased 4-fold when
this alga was moved from fresh to sea water culture. Presumably,
T-iso also uses free amino acids to help maintain an osmotic
balance in its marine environment.
The high level of glutamic acid, which malces up approximately
50% of the total free amino acids, is typical for yeast (Ren et al.,
1 993). The alga T-iso (Isochrysis galbana ) has a large percentage
of free gamma eminobutyric acid (about 20%). There appears to be
no published data on the free amino acid composition of T-iso.
Gamma aminobutyric acid does not play a role in the structural
proteins (Enright et aL, 1986) but is the dominant free amino acid i n
T-iso. This indicates that its major role in the algae could be as an
osmotic effector.
In general, the harpacticoid copepod Tisbe demonstrated a
similar free amino acid pattem when fed the yeast and T-iso diets,
despite the very different free amino acid compositions of the diet.
The wild zooplankton also demonstrated a very similar pattern t O
Tisbe. The total amount of free amino acids per individual Tisbe
was about twice as great in those fed on yeast as aiose fed on T-iso.

Such variations in total free amino acids were also seen in the
rotifer Brachionus plicatilis fed various algal diets (Stmrttrup et al.,
1995). and might be expected considering the transitory nature of
the free amino acid pool.
Glycine was the dominant free amino acid in the copepod
samples including those from the wild zooplankton. The free glycine
levels of Tisbe more closely resembled those of the calanoid
copepods (wild zooplankton sample), than those of the i n t e rt idal
harpacticoid copepod Tigropius califomicus examined by Burton and
Feldman (1982). Th8 proline concentration was 5 times higher than
glycine in T. californ~cus, and was the primary free amino acid used
by this euryhaline copepod to osmoregulate in extremely variable
salinities. It appears that the non-essential free amino acid
composition of marine copepods is more dependent on their f u nct ion
in osmoregulation than on their classification (harpacticoid vs.
calanoid). Free amino acids of euryhaline copepods are dominated by
proline. whereas stenohaline copepods store large amounts of free
glycine. The stenohaline copepods with dominant glycine in the free
amino acid pool include calanoids such as Acartia tonsa (Famer and

Reeve, 1978) and Calanus finmarchicus (Cowey and Corner. 1 963),
as well as our harpacticoid copepod Tisbe . The percentage of essential free amino acids in 77sbe fed the
alga T-iso was almost twice that of nsbe fed yeast. This may
partially reflect the higher total amount of free amino acids
available in T-iso, which also has a much higher percentage of f ree
essential amino acids approximately double that in the yeast.
Copepods have high enzyme levels and contain large amounts of
protease (Munilla-Moran et el., 1990; Mayzaud et al., 1992).
Therefore, the proteins in the yeast and T-iso can also contribute to
the free amino acid pool in Tisbe . Like the copepods, marine flatfish l a ~ a e aie unable to
synthesize the 10 essential amino acids and must, theref ore,
incorporate them from their diet (Cowey et al., 1970). Tisbe fed
with the alga T-iso had a higher percentage of essential free amino
acids than either the Tisbe fed with yeast, or the wild zooplanidon,
and thus might provide better nutrition fo i the fish lawae. Marine
fish larvae tend to have simple digestive systerns and l irnited
protein catabolizing ability. This is demonstrated in the marine fish
larvae Chrysophrys major, Plecoglossus aitiveiis and L ates

calcarifer, which can ingest protein-membrane microcapsules, but
are unable to digest the walls of these capsules (Kanazawa et al.,
1982; Walford et al., 1991). As noted above, the proteins in l ive
food may be partially digestible because the fish lanrae use the
exogenous enzymes of the live food foi digestion. Walford et al.
(1991) demonstrated that adding rotifers to a microencapsulated
diet caused increased digestion of the protein membranes of the
microcapsules, suggesting that the fish larvae are using the
exogenous proteases of the rotifers to aid in digestion of the protein
membrane of the microcapsules.
Amino acid composition of proteins in crustaceans fed
different diets tends to remain fairly constant (Frolov et al., 1991;
Tamani et al., 1993). Therefore, assuming a constant amino acid
composition foi the proteins, Tisbe fed with T-iso rather than yeast
would appeai to be the better food for marine fish larvae because its
free essential amino acid content is double that of Tisbe fed w i t h
yeast.

Chaptei 3
Mars Culture of the Harpacticoid Copepod, T l s b sp.

Introduction
Harpadicoid copepods have been mass cultured in vaiious
laboratories for use as an alternative live food source. The marine
harpacticoid copepods, Tigropius jatponicus (Fukusho, 1 98 O),
Amphiascoides atopus (Sun and Fleeger, 1995) and Tisbe sp. (Kahan
et el., 1982; Uhlig, 1984) are among those that have been cultuied on
a large scale.
The harpacticoid copepod genus Tisbe is epibenthic and l ives
in the subtidai zone. Tisbe is therefore more suited to production
with a more constant supply of fresh seawater. A funning water
system shows decisive advantages over a closed system. Zhang and
Uhlig (1993) found that lawal mortality is about 20% lower in
running water systems, the time of larval development is about two
days shorter and thete is a high rate of naopliar production. Uhlig
(1981) also demonstrated that an increase in the production of Tisbe
was correlated more to the available substrate or base area in the
container, than to seawater volume.
U hlig (1 084) gave five reasons why harpacticoid copepods,
such as Tisbe, are more suitable than calanoids in mass culture as
live food for marine fish larvae. Of particular importance are: 1 )

their tolerance of a wide range of environmental conditions, 2) their
ability to utilize many different food sources, 3) their high
reproductive capacity. 4) their relatively short life cycles, and 5)
their ability to produce high population densities in appropriate
culture systems.
The purpose of my study was to create an appropriate mass
culture system for Tisbe, and evaluate the copepod population
present in the system over time. The copepod populations in the
mass culture system were evaluated for two diets: a commercial
larval diet, Microfeast L-1 O@, and the alga lsochrysis galbana. Given
its ability to thrive on a van'ety of food sources (Miliou and
Moraitou-Apostopoulou, 1991 b; U hlig, 1 Q8l), high densities of Tisbe
were expected in this system for both diets.
Materials and Methods
The mass culture system used in this study (Fig. 3). contained
32 L of seawater (20°C) in a plastic culture tank. The water was
successively filtered through 5 pm and 0.25 pm pore size cartridge
f Mers (model #150071; Atlantic Purification Systems Ltd.,
Dartmouth, N. S.) end approximately 15-20 L was added every 2-3 d.
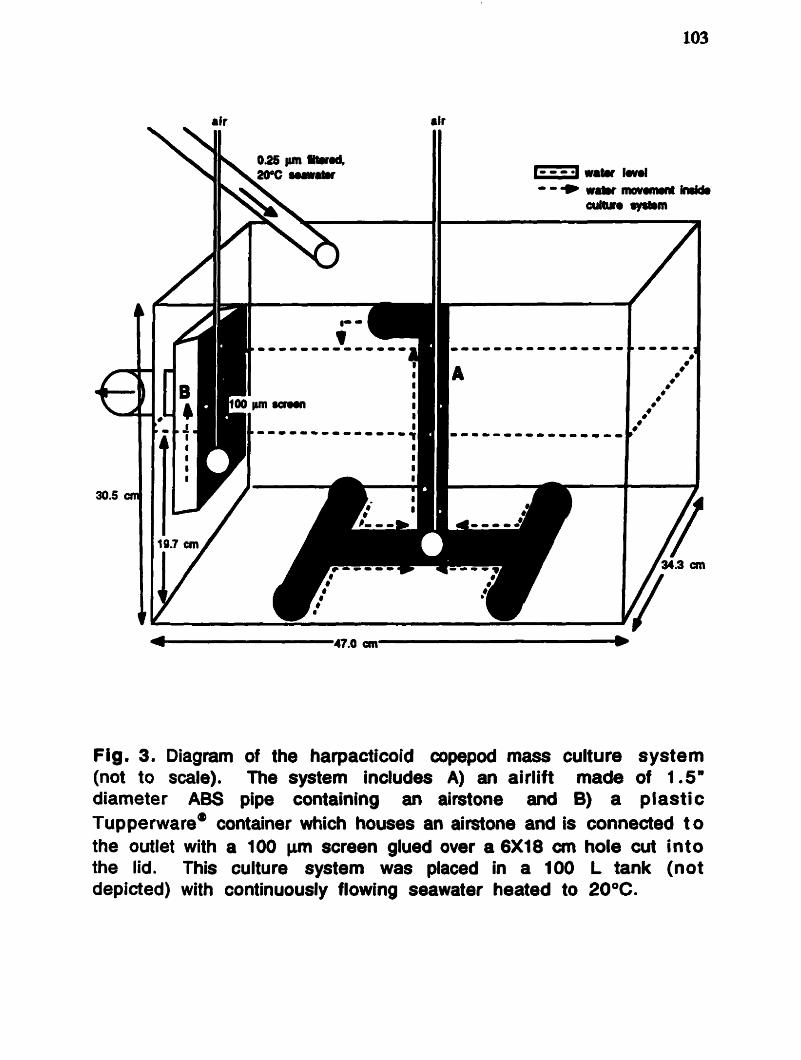
Fig. 3. Diagram of the harpacticoid copepod mas$ culture system (not to scale). The system indudes A) an airlift made of 1 .5" diameter A M pipe containing an airstone and B) a plastic ~upperware~ container which houses an airstone and is connected to the outlet with a 100 pm screen glued over a 6x18 an hole cut in t O
the lid. This culture system was placed in a 100 L tank (not depicted) with continuously flowing seawater heated to 20°C.

A constant flow rate was not used, because the 100 pm screened
outlet became obstnided in the tank containing the oily ~ i c r o f e a s t ~
diet, causing the tank to overflow within 12 hm An airlift pump kept
the water in the tanks well aerated, and provided a means of
bringing copepods from the bottom of the tank to the surface f O r
collection. A plastic ~upperware@ container (24x12 cm) was affixed
to the outlet. The ~upperware* lid had an 18x6 cm hole cut out and a
100 pm screen glued over the opening. The 100 pm screen was large
enough to permit good water flow through the outlet pipe, but it did
allow th8 escape of some of the copepod nauplii, although the adults
and copepodites were ietained. An airstone was used to maintain
water circulation in the ~upperware' container, thereby keeping the
mesh free from obstruction. The culture system was placed in a 100
L tank, with a continuous flow of 20°C seawater to maintain a
constant water temperature.
The harpacticoid copepod Tisbe which we had been culturing in
srnall, 6 L cylindrical jars for biochemical analyses, was introduced
to our mass culture system. Two replicate culture systems were
used for each of the two dietary treatments. The copepods were fed

twice per week with either 0.5 g of Microfeast L-IO* lawal diet or
3 L (appiox. 5 million cells/mL) of log phase lsochrysis galbana
(clone T-iso).
Enurneration of copepod populations in the mass culture tanks
were perfomed at various time intewals. The copepods were
transferred from the mass culture tanks into 20 L buckets using
0.25 pm filtered sea water. They were then collected on a 40 pm
screen to decrease the water volume and transferred to a 2 L
graduated cylinder. They were then thoroughly mixed and a known
volume was removed.. The majority of adult copepods were
separated from the copepodites and nauplii, by seiving through a 200
pm screen. After a number of sirnilar dilutions, copepods of the two
size fractions were evenly mixed in a 20 mL graduated cylinder, and
0.5 mL drops were micropipetted onto a petri dish for counting using
a dissecting microscope (20X magnification). Five drops were
counted to determine an average. Dilution factors were used to
calculate the number of copepods in the original culture.
Rerults and Diacurrion
The copepods fed the Microfeast L-IO@ larval diet were first
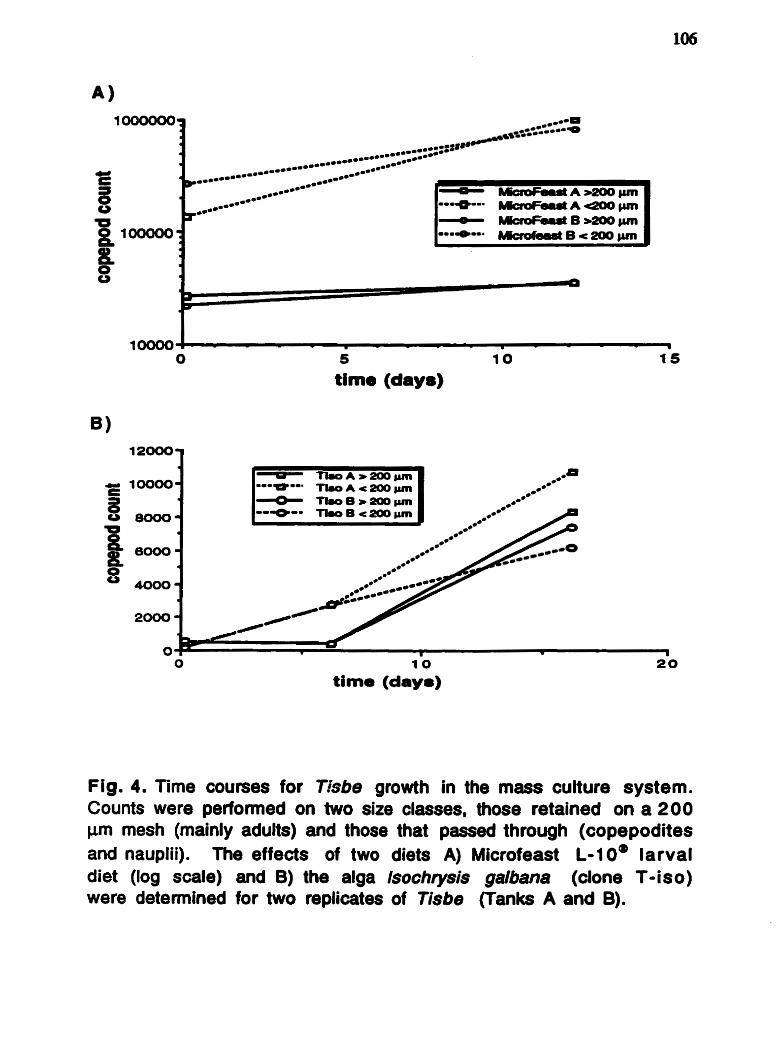
- . . O 5 10 15
time (days)
m
2000 - 10 20
timr (dayr)
Fig. 4. Time courses for Tisbe growth in the mass culture system. Counts were performed on two sire classes, those retained on a 200 ym mesh (mainly adults) and those that passed through (copepodites and nauplii). The effects of two diets A) Microfeast L-1 O' larval diet (log scale) and 6) the alga lsochrysis galbana (clone T-iso) were determined for two ieplicates of Tisbe (Tanks A end B).

counted after approximately 3 weeks of culture (Fig. 4). Over a
period of 12 days, the adults (~200 pm size class) increased on
average from about 22,000 to 32,000 indviduals. During this same
time period, the juveniles and nauplii (QOO pm size class)
increased on average from about l89,OOO to 829,000 individuals.
The copepods in the mass culture tanks fed T-iso were counted f ro m
the first day, when 384 adult copepods wefe introduced to each of
the two tanks (Fig. 4). Growth of the populations were very si mil ar
in the replicate tanks fed 1-iso. Over the first 6 d, the adult size
ciass decreased from 384 to an average of about 225 individuals
(41.4% mortality). Meanwhile, the juveniles and nauplii increased
from O to an average of about 2,500 individuals. The adult
populations thus produced an average of about 1.4 nauplii par
individual per day in this mass culture system over the first 6 d.
After 16 d, the adult population (>ZOO pm) reached about 7,000 as a
result of the maturation of the first generation of nauplii. There
was a large difference in the population growth of the 9 0 0 pm size
class between tanks from 6 to 16 d. In Tank A, the population
increased from 2,500 to alrnost 11,000 and in Tank B. from 2,500 to
6,000 individuals.

The expefiment was terminated du8 to contamination by
other harpacticoid copepod species in the tanks. Thus. we were not
able to compare the carrying capacities of the mass culture tanks
containing Tisbe fed the two diets. To prevent contamination i n
future studies, it is recommended that the copepods be cultured i n
an isolated area to prevent untreated se% water from getting into
the tanks. The copepod populations fed T-iso had not reached the
levels of those fed Microfeast. The mass culture of Tisbe fed w i t h
T-iso demonstrated the short time span needed to obtain a f a i rl y
large population of copepods. After 16 d the total copepod
population (both site classes) had reached a level of alrnost 20,000
copepods per tank. The terminal population of the Microf east-fed
animais in Tank A approached 1 million of the d l 0 pm class and
32.000 adults in a relatively small tank volume of 32 L. This
indicates that Tisbe has the capacity necessary to produce the
numbers needed for larval fish culture. The original purpose of the
experirnent was to detemine the carrying capacity of this system
for Tisbe fed both diets. Unforhinately, the copepod population was
still growing in al1 tanks at the time of contamination. Once the
canying capacity of the system is determined, a study of harvesting

rates for the copepod population could be carried out. This study
should simulate the exploitation levels needed for the culture of
marine fish larvae. Different proportions of the population would be
taken out at various time intervals to determine the ability of the
population to retum to previous levels (Ohno et al., 1990).
The copepod population could be further increased if the
available substrate area in the tank was increased. Uhlig (1 98 1 )
dernonstiated that the mass production of Tisbe is essentially
related to the available substrate area, and less to the available
water volume. A series of black removable plates placed in the tank
might be an advantageous addition to the m a s culture system.
Harpacticoid copepods are negatively phototactic and will be
attracted to the surface of the black plates. These plates could then
be rernoved and the copepods washed off to be fed to the fish larvae.

Chapter 4
Preliminary Trials Uring a Harpacticoid
Copepod, Tlrbe rp., as a Diet for Marine Fish Larvae

Introduction
Calanoid (Last, 1978; McLaren and Avendano, 1995)
harpacticoid (Jindasa et al., 1 991 ; McCall, 1 992) copepodites
nauplii constitute the principal food of many mafine fish lawae.
culture of most cold-water marine fish species requires
and
and
The
the
provision of live prey for a variable period .from the onset of larval
feeding. When the marine Rsh larvae are 0ffer8d C0p8pOd~ and other
zooplankton @.gag rotifers Brachionus plicatilis) in their diets, the
fish larvae tend to select the copepods (Kuhlmann, Quantz, and Witt,
1981; van der Meeren, 1991). Meng and Orsi (1991) demonstrated
that larval striped bass in the San Joaquin Estuary select native
over introduced species of copepods.
Growth of marine fish larvae fed on a copepod diet alone has
been demonstrated for both calanoids and harpacticoids. for
example, the harpacticoid Tisbe sp. has been used in preliminary
studies as a live food for laival saury Scomberesox saurus
(Brownell, 1983) and sea bream Spanrs aurata (Kahan et al., 1 98 1 ) .
Euterpina a cutiforons was fed to larval mahimahi Cotyphaena
hippurus (Kraul et al., 1991). Tigropius japonicus was fed to the

112
mud dab Limanda ydrohamae (Fukusho et el., 1979) and the black
sea bream Mylio macrocephaius (Lee et al., 1981).
The calanoid copepoâs have ais0 been used as live food for
manne fish larvae. Euryiemora affinis has been wed as the sole
food foi striped b a s larvae Morone saxtiIis (Chesney, 1989; Tsai Ce,
1991), and for turbot larvae Scophthalmus maximus (Kuhlmann e t
al., 1981 ; Witt et al., 1984). Wild zooplankton, presumed to be made
up almost entirely of calanoid copepods, have been used as food fo r
hali but larvae Hippoglossus hnippglossus (Boxaspen et al., 1 990 ;
Shields, 1992).
The main advantages in using harpacticoid rather than calanoid
copepods are the ability of the former to be mass cultured and thei r
high €FA (20:Sn-3 and 22:6n-3) composition which is independent o f
the long-chah €FA composition of the diet. A further advantage o f
using harpacticoids in marine fish culture is that those which are
not eaten are able to find nourishment in the fish tanks by feeding on
detritus or the rapidly developing biofilm, including live bacteria on
the tank surfaces. Thus, they maintain their nutritional value while
helping to keep the tank ctean, both important factors in the
successful rean'ng of marine fish larvae (Stnrttrup, 1 993).

A disadvantage to using harpacticoids for marine fish culture
is their inability to remain suspended in the water column.
However, Kahan et al. (1 981) devised a floating tray culture system
for Tisbe, in which a basket with a 80-100 pin mesh bottom f loats
on the surface of a larval marine fish rearing tank. This allowed
newly hatched nauplii to fall through the mesh, thereby making the
copepods more available in the water column as a live food for the
fish lawae.
A preliminary, small-scale study was perfoned w i t h
American plaice Hippoglossoides platessoides and haddock
Melanogrammus aeglefinus larvae feeding on a sole diet of either
rotifers Brachionus plicatilis or the harpacticoid copepod Tisbe sp.
The main objective was to detemine if the marine fish larvae w i l l
grow and survive, on a diet of Tisbe alone. The lanrae should show
increased growth and suwival, due to the high EFA value of Tisbe
cornpared with rotifers. However, this benefit might not be realized
by the possibility that Tisbe may be less available than the rot i f ers
to the fish lanrae in the water column. As an aside. a f i rst-f eeding
haddock larvae fed with Tisbe was longditudinally thin-sectioned,
to determine if there were copepods in its gut.

Materials and Yethods
Eggs f rom one naturally ripe American plaice Hippoglossoides
platessoides were stfipped on March 29, 1996, and placed in dry
plastic containers floated on ambient temperature sea water (Z°C).
The sperm from an adult male was then added to the container and
mixed with the eggs. Fertilization occured over a perlod of 5 min.
Filtered (5 pm), ambient temperature (2OC) sea water was then
added to the fertilized eggs and they were tiansferred to floating
trays with a 200 pm mesh bottom in a flow-through ambient sea
water (2OC) tank. The trays were checked at regular intervals and
unfertilized or undeveloped doudy eggs were removed to prevent
disease or anaerobic conditions. Hatching took place approximatel y
14 d after fertilization. Initial lengths of the larvae were measured
using slide calipers before they were placed in larval culture
containers. The containers consisted of 15 cm (6 in.) diameter
white PVC pipe 15 cm in height with a 40 pm mesh screen inserted
5 cm from the bottom (total volume 1.9 L). These were placed i n
flow-through ambient seawater tanks at 2.0°C (range 1.2-2.6°C).
The l a ~ a e were cultured in green water with approximately 250 mL
(a. 5 million cells/ml) of the alga, lsochtysis galbana (clone T -

iso), added daily. To allow for optimal aeration, a stream of air was
directed over the surface. Two arches (2 X 3 cm) were cut out of the
bottom of each container to allow for better water flow and
exchange through the mesh bottom. Only one container with 20
plaice larvae was used for feeding with the copepod (Tisbe) diet and
four containers with 50 plaice larvae each were used for feeding
with the rotifer Brachionus plicatilis. This irnbalance was due t O
the fact that our mass copepod culture system was just being
established and there were insuffident copepods for feeding a large
number of fish larvae. The main objective was to determine whether
the l a ~ a e would grow and survive fed on a sole diet of Tisbe. The
rotifers were offered at a concentration of approximately 10
animals/mL. The concentration of Tisbe was approximatel y 0.9
copepodlmL in the >200 pm size fraction (mainly adults) and 3.5
copepods/mL in the 4 0 0 pm size fraction (copepodites and nauplii).
Tisbe was cultured in 6 4 cylindrical jars and fed bakers'
yeast using the procedures described for the copepods produced f O r
lipid analysis (p. 16).
The rotifers were fed 3 g of Microfeast L-1 O" lawal diet
daily. The rotifers were cultured at 20°C in 100 L of filtered (5

pm), 20
with an
replaced
ppt seawater contained in a 500-L plastic cylindrical
airstone. Approximateiy two thirds of the seawater
116
tank
was
each week. The rotifer population averaged about 30,000
individuals per L.
A second group of plaice larvae (hatched 6 June, 1996) was
cultured in 27 an (11 in.) diameter Mack buckets (total volume 7.8
L) with two 10 X 10 cm square holes cut in the sides. These holes
were covered with 40 mesh to allow water flow through the
container yet retain the live food. These buckets were placed in the
flow-through ambient seawater tanks (at average 6.1 O C ; range 5.4-
6.8OC). The larvae were cultured in green water with approximately
250 mL (ca. 5 million cells/mL) of the alga, I. galbana (clone T-iso),
being added daily. Thirty plaice larvae were cultured in each of three
buckets and fed rotifers at approximateiy 10 rotifers/mL. Nine days
after hatching, DHA-enriched brine shnmp (1-2 d post hatch) were
also fed in excess to the larvae. Unfortunately, most of these larvae
died about 20 d post hatch due to water-flow problems. However,
sixty larve8 of this same batch were cultured in floating trays w i t h
a 40 pm mesh bottom and were unaffected by these water-flow
problems. These plaice larvae were fed in the same manner (rotifers

and brine shrimp) as those cultured in the black buckets. The tim ing
of metamorphosis and the dimensions of these plaice larvae at
metamorphosis were recorded.
The bn'ne shrimp wem cultured using the following procedure.
Ten mL of brine shiimp cysts (8iomarine Inc., Hawthorne, CA) were
added to a 6-1 cylindrical jar (with a bottom drain), containing 4 L
of 20°C, filtered (5 pm) seawater. Most of the brin8 shn'mp hatched
after 48 h. The newly-hatched brine shrimp were then sepaiated
from the dead cysts which rose to the top of the container. This was
done in a dark room, by holding a light near the bottom of the
container. The newly-hatched brine shrimp were attracted to the
light as the water was drained from the bottom, and caught on a 40
pm screen. lhey were then transferred to another 6 L cylindrical
jar, containing an airstone and approximately 0.25 g of DHA-rich
~e lco ' (Artemia Systems N.V., Ghent, Belgium) emulsified in 4 L of
sea water. The brine shrimp were enfiched for 24 h, and rescreened
through a 40 pm mesh to remove excess oil prior to feeding to the
fish larvae.
Fertilized eggs of haddock Melanogrammus âeglefinus, obtained
from Dr. Ken Waiwood (St. Andrews Biological Research Station, N

B.) were placed in floating trays and treated similarly to those o f
the American plaice. The eggs were fertilized April 5, 1996 and
hatched approximately 10 d later. The newly-hatched larvae were
measured and separated into two culture containers (average 2.0°C;
range 1.2-2.6OC) with the same setup as used for the plaice lafvae.
Thirty larvae were placed in each container and fed either the
rotifers or copepods (Tisbe). The concentration of Tïsbe was
approximately 0.7 copepod/mL in the a200 pm size fraction (mainly
adults) and 2.7 copepodlmL in the 4 0 0 Fm size fraction
(copepodites and nauplii). The rotifers were fed at a concent ration
of approximately 10 animals/ml. The rotifers were cultured on
Microfeast L-10" larval diet while Tisbe was grown on bakers'
yeast . A first-feeding haddock larva (17 d post hatch) was taken from
each dietary treatment (Tisbe and rotifer) and fixed in 1%
glutaraldehyde and 4% fonnaldehyde and thin-sectioned, so that the
gut content could be microscopically examined. The fixed specimens
were dehydrated in methanol and embedded in historesin blocks
which were sectioned et 2 pn and stained with toluidine blue. After
staining , the sections were dried with compressed

chlorodifluoromethane (MG. Chemicals, Surrey, B. C.) instead O f
solvents. Sections were photographed with a Zeiss photomicroscope.
Reaults and Discussion
The plaice larvae fed with copepods (Tisbe) had significantly
higher growth (psO.05) compared to those fed with rotifers, when
assessed after 6 d (Table 12). This may have b e n due more to the
lower density of copepod-fed larvae present in their containers than
to diet. Suwivai of the copepod-fed plaice larvae (65%) was very
similar to that of the rotifer-fed plaice l a m e with the highest
survival (68%) 12 days after hatching. Their was a large variation in
the suwival of plaice larvae fed rotifers (Table 12). Suwival of
plaice larvae declined precipitously 2-3 weeks after hatching. This
mortality could have been due to a number of factors: 1 )
deteriorating water quality, particulaily in the tanks with Tisbe as
food. because we were unable to completely separate the benthic
copepods frorn the detritus in the copepod culture container before
feeding them to the fish larvae; 2) inappropriate fish lawae culture
containers (containers with dark walls and conical bottoms would be
preferable); and 3) rotifers may have been too small as live food f O r
the two week old l a~ae . Even so, the lame were obsewed to be

Table 12. Survival and growth measurements of American plaice (HippogIossoides platessoides) lame ieared at 2OC and f ed harpacticoid oopepods (Tisbe sp.) or rotifers (Brachionus plicatilis). The copepod was fed bakenr' yeast and the rotifer was fed Microf easte L-10 Larval Diet. The asterisk symbol indicates significant ciifferences at the pe0.05 level.
Copepod-feda Rotifer-fedb Survivat% Survival% (n=5û)
Time (days) Length (mm) (W02 Length (mm) Median Range O 4.21 f 0.31 1 O0 4.35 f 0.38 1 O0 100-1 00 6 6.32 f 0.43' - 5.56 f 0.58 - 12 O 65 - 24 68-1 O 15 9 25 9 12 48-6 23 O O 7.61 f 0.72 4 14-0
a Survival in phice lanrae fed with appods foi 1 container containing an initial population of 20 larvae.
b. Survival in plaice larvae fed with rotifen is the median aid range for 4 containers containing an initial population of 50 lawae each.

feeding on Tisbe and we were able to obtain relatively good growth
and sundval of the lanrae for two weeks after hatching. considering
that the culture conditions were less than optimal.
The plaice larvae, which were cultured in the second
experiment in three black buckets, had extremely good growth and
suwival during the first three weeks after hatching. The results
were as follows: day 1 post hatch (suwival 1 W%, length 5.37i0.21
mm), day 6 (suwival 1 00%. length 6.1 3I0.38 mm), day 13 (survival
87.8fl.9%, length 7.5if0.10) and day 20 (survival 78.9I5.1%, length
8.4810.38). These length measurements were taken f rom a sample
of five larvae. All larvae in the black buckets died shortly after 2 0
d due to water-flow problems. The increased early growth and
survival of these plaice larvae raised in the black buckets, compared
to those in the white PVC pipe could have been due to a number of
factors: 1) a higher percentage of the eggs fiom this batch were
transparent floaters and the hatching success was higher; 2) the
density of the larvae in the black buckets was much lower; 3) the
black buckets provide better contrast for the larvae to see and
capture their live food prey; and 4) DHA-enriched brin8 shrimp were
added to the diet 9 d post hatch.

Approximately 60 plaice lame of the same batch as those in
the black buckets, were reared on the sarne rotifer and brine shrirnp
diets in floating trays with a 40 pm mesh bottom. These lawae
were unaffected by the water flow problems. 01 these larvae, 2 0
were raised through metamorphosis. Metamorphosis was defined by
complete eye migration, fin ray and scale development. At this
point, they started to feed on aie pelleted marine fish lawal diet
developed by Dr. J. D. Castell. Metamorphosis occured approxirn atel y
85-90 days post hatch. The newly metamorphosed larvae were
25.59I2.74 mm long and 11.39I0.41 mm wide (n=5). The
temperature rose from 6 to g0C over this time period. To my
knowledge, there are no published studies of Amen'can plaice larvae
being raised through metamorphosis.
Haddock l a ~ a e were also cültured with Tisbe and rotifers
(Table 13). The haddock lawae demonstrated better growth but
poorer survival when fed the copepods compared to the rotif ers.
Unlike the plaice larvae, the containers with haddock larvae fed the
two diets had similar numbers of lawae suggesting that the better
growth was due more to diet than stocking density. Feeding behavior
of the haddock on the copepod lanrae was also obsented. The biggest

Table 13. Sunrival and growth measurements of haddock (Melanogremmus aeglefinus) larvae reared at 2OC and fed harpacticoid copepods ( Tisbe sp.) or rotifers (Brachionus plica tilis) . The copepod was fed bakers' yeast and the rotifer was fed Microfeaste L-IO Lawal Oiet.
copepod-fed rotifer-fed s u n r ~ v a ~ sunrival96
time (days) length (mm) (ndo) lerigth (mm) b 3 0 ) O 3-17 * 0.41 1 O0 3.47 * 0.41 100 8.5 5.36 f 0.78. 57 4.57 f 0.15 93
18.5 5.89 f 0.61" 23 5.00 * 0.41 57 21.5 - 1 O - 10 25.5 - 3 - 7
Significant differences at P O . 1 O.
" Significant differences at p40.05.

drop in suivival was durlng the first two weeks post hatch for the
copepod-fed larvae and after approximately 2.5 weeks for the
rotifer-fed larvae. The reasons are probably similar to those
proposed for the plaice larvae.
Proof that the first-feeding haddock larvae ingested Tisbe can
be seen in photographs of a thin-sectioned lama (Fig. 5). This
haddock larva contained a partially digested adult or l ate r-stage
copepodite (over 200 pm ir; length) in the stomach lumen, a smaller
early copepodite (100-200 pm in length) in the intestinal lumen and
possibly a third welkdigested copepod in the most posterior section
of the intestinal lumen. A haddock larva taken from the tank with
rotifers as food showed nothing in the gut lumen.
These preliminary larval feeding trials demonstrated that the
haddock and plaice lawae will ingest the harpacticoid copepod Tisbe
and will grow on this diet over the first two weeks after hatching.
The culture conditions were not optimal and work needs to be done
on a larger scale with better culture parameters to determine
whether rotifers or Tisbe is superior as a live food diet. Tisbe
spent most of ils time on the walls and bottom of the culture
container, and so was not as accessible to the larvae as were the

Fig . 5. Thin section of a first-feeding haddock (Melanogrammus aeglefinus) larva (17 d post hatch) fed the harpacticoid copepod (Tisbe sp.) and reared at 2°C. A. Whole rnount of the larva with arrows depicting the gut lumen containing the partial1 y-digested copepod (bar= 500 pm). B. Magnified view of the gut content of the lama (bar= 1 00 pm).

rotifers which swim freely in th8 water column. This too, may be a
reason for the drop in suwival of the larvae approximately two
weeks after hatching.
In summary, the increased growth of the marine fish larvae
fed with Tisbe compared with rotifers, could be due to both the
increased EFA vdue of th8 copepod. and the decreased densities O f
l a ~ a e in the tanks that were fed Tisbe. The decreased suwival o f
the fish larvae in the copepod-fed haddock larvae could be due to the
inaccessbility of the lhre food to the lawae in the water column. To
make Tisbe more accessible as a live food foi the fish larvae, a
floating tray system similar to the one designed by Kahan et al.
(1981) might be used. Because of their detritivorous, benthic
nature, Tisbe may better serve as a tank cleaner and EFA-rich, live
food supplement to rotifers and bn'ne shn'rnp, rather than as the sole
live food diet (Stattrup et al., 1995). Further studies comparing
growth and suwlval of fish larvae on a diet of Tisbe, rotifers and
brine shrimp, versus rotifers and brine shrimp alone, should be
performed to determine the value of Tisbe as a live food
supplement.

GENERAL CONCLUSIONS
The greatest obstacle in the culture of cold-water marine fis h
is the provision of a nutritionally suitable live food for the larvae a t
f irst-feeding . Harpadcoid copepods ( Tlsbe sp. in particular) are
prornising candidates as potential live food foi marine fish larvae.
They are easy to culture, can be grown at high densities and have the
necessary nutritional composition, in ternis of EFA, for marine fish.
Pemaps the rnost important requirement for marine fish larvae is a
diet with a high DHA to EPA ratio. The harpadcoids maintained this
high ratio despite a diet low in EFA or a high culture temperature.
Tisbe also contained >1% of the €FA arachidonic acid in its lipid at
20°C, irrespective of the diet's EFA composition.
Unlike fatty acid composition, the effects of free amino acid
composition of the live food on growth and survivai in manne fis h
larvae is largely unknown. Free amino acids are used primarily as an
energy source in the developing larvae. The harpacticoid copepods'
free amino acid pattern is fairly consenrative for the major
components, irrespective of the free amino acid composition of the
diet, with glycine contributing over 30% of the free arnino acids
followed by alanine, taurine and arginine. The essential free arnino

128
acids in 7ïsbe fed T-iso (30%) were airnoet double that of Tisbe fed
yeast (17%). The levels of total free amino adds per copepod was
highly variable.
The harpacticoid copepod Tnsbe demonstrated the necessafy
production parameters for mass culture, and could give the numbers
needed for marine fish lanral culture. ln a 32 L container nearly 1 O6
copepods were produced. This system could easily be modified t o
increase this production significantly by increasing the surface area
available to the copepod.
Survival of the marine fish larvae was poor when fed only on
the harpacticoid copepod Tisbe. This could have been due to several
factors including inappropriate culture containers and the
unavailability of the copepod in the watei column. Tsbe may better
serve as a tank cleaner and EFA-rich live food supplement to
rotifers and brine shrimp, rather than as the sole live food (Stettrup
et al., 1995). Further studies should be performed to confimi this.
Presently there is much local interest in using the
harpacticoid copepod Tisbe sp. as a live food organism for marine
fish culture. Research on the nutritional composition of Tisbe and
related studies are critical if marine finfish culture is to progress

in the Atlantic provinces. To date, replacement of live food has not
k e n accomplished for any of the marine finfish species considered
important for cultivation (Holt, 1993). Thus EFA-rich, live food
organisms such as Tisbe are, at this point the only viable
alternative for getting the larvae past first-feeding stage en route
to metamorphosis.

Appendlx A - Lipid Analysis Techniques
comôine sample with 10 mL methanol + 5 m l chloroform
homogenize for 1 min in polytron
suction filter through Whatman no. 1 filter paper rinse sample tube and filtrate with 5 mL chlorofom
add 9 mC distilled water to give 0:10:10 solvent ratio
dry chlorofom and lipid discard top aqueous with anhydrous sodium sulfate methanol layer*
filter through glass wool in a Pasteur pipette to test tube
evaporate chloroform under nitrogen
transfer to preweighed via1 and obtain total lipid weight
Fig. Al . Diagram of the Bligh and Dyer (1 959) method for total l ip id extraction. The asterisk symbol indicates where the sample w as extracted for amino acids.

Obtain total lipid using Bligh and Dyer (1959) method. Add 2 mL 7% boron trifluoride in methanol and 0.5 mL of toluene per 1 mg of lipid. Boil for 30 min at 100°C or ovemight at 50°C in pressure-tested 15 mL teflon-lined screw cap culture tubes. Cool tubes and add 10 mL of distilled water. Extract the top hexane layer mice with 2 to 3 mL of hexane. Discard the water layer. Dry the hexane with anhydrous sodium sulfate. Filter through Pasteur pipette containing gless wool. Evaporate hexane under nitrogen till dry. Add 30pL of chloroform to 1 mg of sample. Apply a 1 cm streak 2.5 cm from bottom of a precoated silica gel thin layer chromatographie (TLC) plate. 3 or 4 sarnples can be added to a 20 X 20 cm TLC plate.
10)Also add 15 pL of a reference methyl ester sample on the side of the plate 2.5 cm from the bottom.
1 1) Develop the plate in hexane: diethyl ether: acetic acid (90: 10:l V/V/V) for 45 min.
12)Allow the solvent to evaporate and spray the section of the plate containing the reference rnethyl esters with the visualking agent 0.1 % 2',7'-dichlorofluorescein in methanol while covenng the experimental sarnples.
13)Under ultraviolet light the reference methyl esters appear as two averlapping spots or bands. The upper band containing the more saturated and the lower containing the more unsaturated methyl esters. Mark the portion of the plate where your methyl esters would be and scrape the silica off the plate into a test tube.
14)Add ca. 7 mL of chloroform to methyl esters and silica in the test tube. Filter through a Pasteur pipette containing glass wool to remove the silica.
1 5) Evaporate to dryness under nitrogen and iedissolve in hexane prïor to analysis of fatty acid methyl esters on the GLC.
Fig. A2. Method for obtaining methyl esters from total lipids for analysis on GLC.

Appendix B - Astuanthin Analysis
The wild zooplankton captured in a plankîon tow (84 pm mesh)
on 27 July, 1995 was analyzed for both lipid (8.2% of dry wt.) and a
carotenoid pigment tentatively identified as astaxanthin (0.6% o f
total lipid). The chlorofom layer of the Bligh and Dyer (1959) lipid
extraction contained al1 of the astaxanthin pigment. The peak
absorbance of the pigment found in the zooplankton occuned at
approximately 460 nm wavelength (Fig. BI), typical of the esterified
astaxanthin pigments dissolved in hexane (467 nm; Czeczuga. 1971).
The absorbance of the pigment at 467 nm was related to the we i g h t
of astaxanthin through the use of known amounts of standard
astaxanthin sample. The positive linear relationship is shown i n
Fig. 82.

300 400 500 600
wavelength (nm)
Fig. BI . Absorbante spectnim of wild zooplankton astaxanthin measured in hexane.
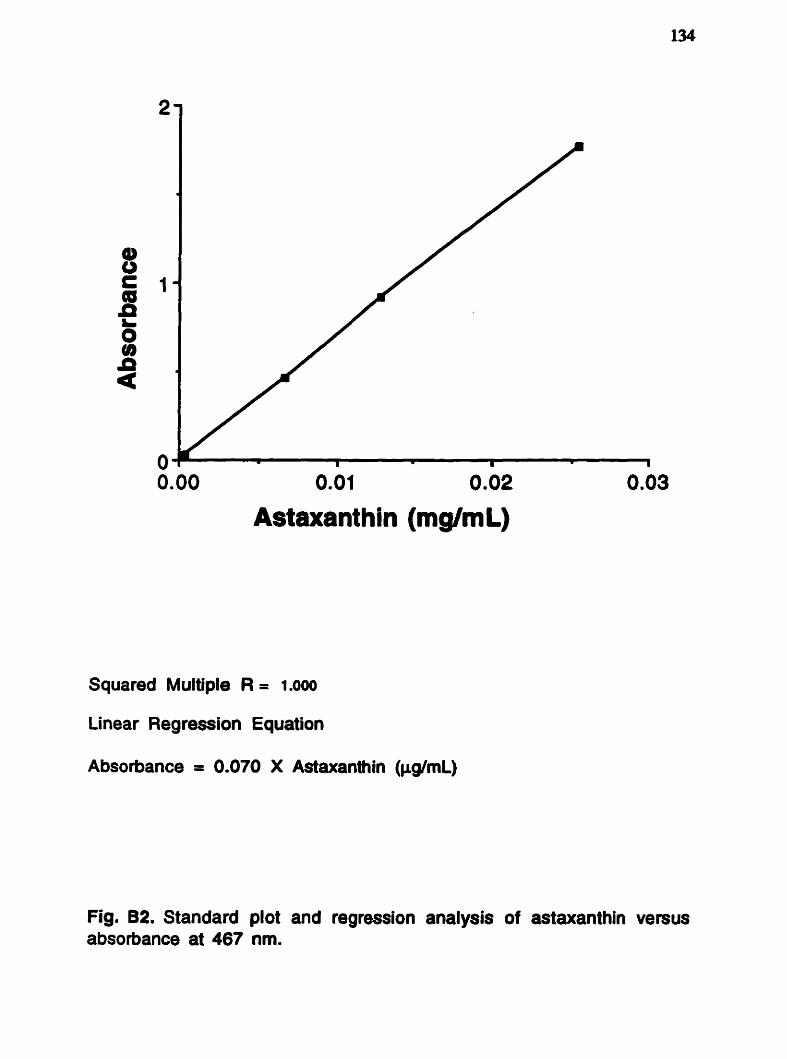
.O0 0.01 0.02 0.03
Astaxanthin (mg/mL)
Squared Multiple R = i .ow
Linear Regression Equation
Absorbance = 0.070 X Astaxanthin (pg/ml)
Fig. B2. Standard plot and iegression analysis of astaxanthin versus absorbante at 467 nm.

Appendix C - Free Amino Acid Extraction
A freeze-dried sample of wild zooplankton was obtained f rom
the plankton of Passamaquoddy Bay, N. B. in July, 1995, using an 8 4
pm mesh plankton net. There appeared to be some leaching of f ree
amino acids in this sample, pehaps due to thawing in the f reeze-
drying process. This was evidenced by large variances in the
zooplankton f ree amino add compositions. This zooplankton sam ple
was then homogenized, so that two extraction techniques could be
compared; the water phase of th8 Bligh and Dyer (1959) method
versus the boiling 80% ethanol method. The boiling 80% ethanol
method gave significantly higher extractions (p<O.OS) for 75% of the
free amino acids detected in the zooplankton (Table Cl). Migration
of some of the free amino acids from the water phase into the
chloroforrn layer in the Bligh and Dyer method might explain the
lower free amino acid extraction values obtained with this method.
Thus, the Bligh and Dyer method, aMhough advantageous in ternis o f
allowing one to analyze both lipid and free amino acid compositions
with a single sample, significantly underestimates the free amino
acid content. Therefore, it should not be used for the purpose of
free amino acid extraction.

Table Cl . A cornparison of two amino acid extraction techniques, the Bligh and Dyer aqueous phase extraction and the boiling 80% ethanol methods. Amino acid composition of a homogenized, freeze- dried sample of wild zooplankton isolated from the plankton of Passamaquoddy Bay, N. B. in July, 1995. The asterisk symbol indicates significant differences at the p<O.OS level.
Amino acid Bliah and mer h m d g diy wt.) ûô% Ethaml (umdlg dry wt.) taurine* 66.67 * 4-06 82.10 i 2.39 aspartic acide 1.43 f 0.08 3.67 f 0.73 threonine* 4.91 f 0.20 5-78 i 0.12 serine' glutamic acid* prolinee glycine* alanine* valine methionine leucine* isoleucine* tyrosine* phenylalanine histidine lysine' ammonia 3.83 I 0.99 arginine* 28.20 f 0.63
See Appendix A (p. 130) for more information on extraction techniques.

Ackman R.G., Linke B.A. and Hingley J., 1974. Some details of fatty acids and alcohols in the lipids of north Atlantic copepods. J. Fish. Res. Board Can. 31(11): 1812-1817.
Bell M.V., Henderson RA, Pirie B.J.S. and Sargent J.R., 1985. Effects of dietary polyunsaturated fetty acid deficiencies on mortality, growîh and gill structure in the turbot, Scophthalmus maximus. J. Fish Biol. 26: 181-191.
Bell J.G., Castell J.O., Tocher D.R., MacDonald F., and Sargent J.R., 1995. Effects of dietary arachidonic acid: docosahexaenoic acid ratios on phospholipid fatty acid compositions and prostaglandin production in juvenile turbot (Scophthalmus maximus). Fish Physiol. Biochem., l4(2): 139-1 51.
Bligh E.C. and Dyer W.J., 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37: 911- 927.
Boxaspen K., Harbe Tm and Skjolddal L.H., 1990. A pilot study of halibut larvae (Hippoglossus hippoglossus L.) reared f rom start feeding to metamorphosis on diets of wild zooplankton and Artemia. ICES, F:52, 8 pp.
Brownell CL., 1983. Early growth rate and feeding of a small group of laboratory-reared saury, Scomberesox saunrs Scombroides (Pisces: Scomberesocidae). S. Afr. J. Mar. Sci. 1: 245-248.
Burton R.S., 1986. Incorporation of C-14 bicarbonate into the free amino acid pool during hyperosmotic stress in an intertidal copepod. J. Exp. Zool. 238: 55-61.
Burton R.S. and Feldman M.W., 1982. Changes in free amino acid concentrations during osmotic stress in the intertidal copepod Tigriopus califomicus. Comp. Biochem. Ph ysiol. 73A(3): 441 -445.
Castell J.D., Bell J.O., Tocher D.R. and Sargent J.R., 1994. Effects of purified diets containing different combinations of arachidonic

acid and docosahexaenoic add on survival, growth and fatty acid compostion of juvenile turbot (Scophthalmus maximus). Aquaculture 128: 81 5-333.
Chapelle S., 1878. The influence of acdimation temperature on the fatty acid composition of an aquatic crustacean (Carcinus maenas). J. Exp. Zool. 204: 337-346.
Chapelle S., 1986. Aspects of phospholipid metaboiism in crustaceans as related to changes in environmental temperatutes and salinities. Comp. Biochem. Physiol. 84B(4): 423439.
Chau Y.K., Chuecas L and Riley J.P.. 1967. The component amino acids of some marine phytoplanktonic species. J. Mar. Biol. Assoc. U.K. 47: 543-554,
Chesney E.J. Jr., 1989. Estimating the food requirements of striped bass larvae Morone saxitilis : Effects of light, turbidity and turbulence. Mar. Ecol. Prog. Ser. 53( 2): 191-200.
Christie, W .M., 1 982. Lipid Analysis. Isolation, Separation, Identification and Structural Analysis of Lipids, 2nd Ed. New York, Pergarnon Press, 207 p.
Cowey C.B., Adron J. and Blair A., 1970. Studies on the nutrition of manne flatfish: the essential amino acid requirements of plaice and sole. J. Mar. Biol. Assoc. U.K. 50: 87-95.
Cowey C.B., Adron J. W., Owen J.M., and Roberts R.J., 1976. The ef f e cts of different dietary oils on tissue fatty acids and tissue pathology in turbot Scophthalmus maximus. Comp. Biochem. Physiol. 538: 399-403.
Cowey C.B. and Corner E.D.S., 1963. Amino acids and some other nitrogenous compounds in Calanus finmarchicus. J. Mar. Biol. Assoc. U.K. 43: 485493.
Cowey C.B. and Forster J.R.M., 1971. The essential amino acid requitements of the prawn Palaemon serratus. The growth of

prawns on diets containing proteins of different amino acid compositions. Mar. Biol. 10: 77-81.
Czeczuga B., 1971. Studies on the carotenoids in Alternia salina L. eggs. Comp. Biochem. Physiol. 408: 47-52.
Davis M.W., and Olla B.L., 1902. Cornparison of growth, behavior and lipid concentrations of walleye pollock Theragra chalcogramma larvae fed lipid enriched, lipid deficient and field collected prey. Mar. Ecol. Prog. Ser., 90(1): 23-30.
Dumont E., Chichery M.P., Chichery R. and Nouvelot A., 1992. Analysis of the fatty acids in the centrai nervous system of Sepia officinalis . Comp. Biochem. Physiol. 1 01 B(3): 341 -344.
Dunstan G.A., Volkman J.K. and Barrett S.M., 1993. The effect of lyphilization on the solvent extraction of lipid classes, fatty acids and sterols from the oyster Crassostrea gigas. Lipids 28(1 O): 937-944.
Enright C.T.. Newkirk G.F., Craigie J.S. and Castell J.O., 1986. Evaluation of phytoplanicton as diets for juvenile Ostrea edulis L. J . Exp. Mar. Biol. 96: 1-1 3.
Farkas T., 1979. Adaptation of fatty acid compositions to temperature- a study on planktonic crustaceans. Comp. Biochem. Physiol. 648: 71 -76.
Famer L. and Reeve M.R., 1978. Role of the free arnino acid pool of the copepod Acartia tonsa in adjustment to salinity change. Mar. Biol. 48: 311-316.
Finn RN. and Fyhn H.J., 1993. Energy substrates in the metabolism of developing marine fish embryos. Lp; Reinertsen H., Oahle LA., Jsrgensen L., and Tvinnereim K. (eds.). Proceedings of the First International Conference on Fish Farming Technology in Trondheim, Norway, 9-1 2 August 1 993. A. A. Balkema, Rotterdam. pp. 117-119.

Finney C.M., 1979. Salinity stress in harpacticoid copepods. Estuaries 2(2): 1 32-1 85.
Frolov A.V., Pankov S.L., Geradze K m N o l Pankova S.A. a d Spektorova L.V., 1991. Influence of the biochemical composition of food on the biochemical composition of the rotifer Brachionus plicatilis. Aquaculture 97: 1 81 -202.
Fukusho K., 1980. Mass production of a copepod, Tigriops japonicus in combination with a rotifer Brachionus plicatilis fed omega- yeast as a food source. Bull. Jpn. Soc. Sci. Fish. 46(5): 625-629.
Fukusho K., Arakawa T., and Watanabe T., 1980. Food value of a copepod, Tigriopus japonicus , cultured with omega-yeast for larvae and juveniles of mud dab Limanda yokohamae. Bull. Jpn. Soc. Sci. Fish. Nissuishi., 46(4): 499-503.
Fyhn H.J.. 1989. First feeding of marine fish larvae: Are free amino acids the source of energy? Aquaculture 80: 1 1 1-120.
Fyhn H.J., Finn R.N., Helland S., Rennestad l., and Lsmsland E.R., 1993. Nutritional value of phyto- and zooplankton as live food for marine fish larvae. Proceedings of the First International Conference on Fish Farming Technology in Trondheim, Norway, 9- 12 August 1993. A.A. Balkema, Rotterdam. pp. 121-126.
Fyhn H.J. and Sergistad B., 1987. Free amino acids as energy substrate in developing eggs and larvae of the cod Gadus mothua. Mar. Biol. 96: 335-341.
Gilles R. and Pequeux A., 1968. Effect of salinity on the free amino acid pool of the red alga Povhyridium purpureum (= P. cruentum). Comp. Biochem. Physiol. 57A: 183-1 85.
Goolish E.M. and Burton R.S., 1989. Energetics of ogmoregulation in an intertidal copepod: effects of anoxia and lipid reserves on the pattern of free amino acid accumulation. Funct. Ecol. 3: 81-89.
Greene D.H.S. and Selivonchick O.P., 1987. Lipid metabolism in fish. Prog. Lipid Res.. 26: 53-85.

Guidi L.D., 1984. The effect of food composition on ingestion. development, and suwival of a harpacticoid copepod, nsbe cucumariae. J. Exp. Mar. Biol. Ecd., 84: 101-1 10.
Guillard R.R.L. and J.H. Ryther. 1962. Studies on manne planktonic diatoms. 1. CycIoteIIa nana Hustedt and Detonula confervacea Cleve. Cm. J. Microbiol. 8: 229-239.
Halver J.E., 1957a. Nutrition of salmonoid fishes: IV. An amino acid test diet for chinook salmon. J. Nutr. 62: 245-254.
Halver J.E., DeLong D.C., and Mertz E.T., 1957b. Nutrition of salmonoid fishes: V. Classification of essential amino acids for chinook salmon. J. Nutr. 63: 95-105.
Harrison K. E., 1 990. Essential fatty acid metabolism and requirements of the iabster, Homarus ametïcanus. Ph. 0. Dissertation, Dalhousie University, Halifax N. S.. 358 p.
Hazel J.R.. 1979. Influence of thermal acclimation on membrane l ipid composition of rainbow trout liver. Am. J. Physiol. 236(1): R91- R101.
Hazel J.R. and Williams E.E., 1990. The role of alterations in membrane lipid compositions in enabling physiological adaptation of organisms to their physical environment. Prog. Lipid Res. 29: 167 -227.
Hjelmeland K., Ugelstad I., Homme J.M., Lein 1. and Pedersen Tm, 1993. ln; Reinertsen H., Dahle L.A., Jsrgensen L., and Tvinnereim K. (eds.). Proceedings of the First International Conference on Fish Farming Technology in Trondheim, Norway, 9-12 August 1993. A.A. Balkema, Rotterdam. p. 127.
Holt G.J., 1993. Feeding larval r8d drum on microparüculate diets in a closed recirculating watei system. J. Woild Aquacult. Soc. 24(2): 225-230.

Howell B.R., 1971. Rearing the larvae of the lemon sole, Microstomus kitt, Walbaum, on cultured foods. CES, E:B6. 4 PP*
Howell BR., 1973. Marine fish culture in Britain VIII. A marine rotifer Brachhus plicatilis Muller, and the lawae of the mussel, M ' u s edulis L. as foods foi l a ~ a l flatfish. J. Cons. I nt. Explor. Mer., 35: 1 -6.
Howell BR. and Tzoumas T.S., 1991. The nutritional value of Artemia nauplii for larval sole, Solea solea (L.) with respect to their (n- 3) HUFA content. Europ. Aquacult. Soc. Spec. Publ. 15: 63-65.
Jeffries H.P. and Alrare L., 1969. Dominance diversity relationships of the free amino acids in coastal zooplankton. Comp. Biochem. Physiol. 37: 21 5-223.
Jindasa J., Howell B.R. and Baynes S.M., 1991. Preliminary experiments on the development of extensive culture methods for juvenile sole, Solea solea (1.). Aquacult. Fish. Manag., 22: 25-36.
Jonasdottir S.H., 1994. Effects of food quality on the reproductive success of Acartia tonsa and Acartia hudsonica : laboratory observations. Mar. Biol. 121 : 67-81.
Jonasdottir S.H., Fields 0. and Pantoja S., 1995. Copepod egg production in Long Island Sound, USA, as a function of the chernical composition of seston. Mar. Ecol. Prog. Ser. 1 1 9: 87-98.
Jonasdottir S.H. and Kirarboe T., 1996. Copepod iecruitment and food composition: Do diatoms affect hatching success? Mar. Biol. 125: 743-750.
Kahan D., 1979. Vegetables as food for marine harpacticoid copepods. Aquaculture, 16: 345950.
Kahan D., Uhlig O., Schwenzer De and Horowitz L., 1982. A simple rnethod for cultivating harpacticoid copepods and offering them to fish larvae. Aquaculture 26: 303-310.

Kanazawa A., Teshima S., lnamori S., Sumida S. and lwashita T., 1982. Rearing of larval red sea bream and ayu with artificial diets. Mem. Fac. Fish. Kagoshima Univ. 31: 185-192.
Kattner O., Krause M. and Trahms JO, 1981. Lipid composition of some typical North Sea copepods. Mar. Ecol. Prog. Ser. 4(1): 69- 74.
Kitajima Cm, Arakawa Tm, Oowa Fm, Fujita S., lmada O., Watanabe T o and Yone, Y., 1980. Dietary value for red sea . bream larvae of rotifer Brachionus plicatilis cultured with a new type of yeast. Bull. Jpn. Soc. Sci. Fish. 46: 43-46.
Kitajima Cm, Yoshida M. and Watanabe T., 1980. Dietary value for Ayu. Plecoglossus altivelis, of rotifer, Brachionus plicatilis, cultured with bakers' yeast, Saccharomyces cerevisiae, supplemented with cuttlefish liver oil. Bull. Jpn. Soc. Sci. Fish. 46: 47-50.
Kluytmans J.H., Boot J.H., Oudejans R.C.H.M. and Zandee Dolet 1985. Fatty acid synthesis in relation to gametogenesis in the mussel Mytius edulis L. Comp. Biochern. Physiol. 818: 959-963.
Kraul S., Ako Ho. BiSttain K., Ogasawara A., Cantrell R. and Nagao Tm, 1991. Cornparison of copepods and enriched Alternia as feeds for laml Mahi mahi (Coryphaena hippuns). in: Lavens P., Sorgeloos P., Jaspers E., and Olevier Fm (eds), LaM' 91-Fish and Crustacean Lawiculture Symposium. pp. 4547. European Aquaculture Soc., Ghent, Belgiurn.
Kuhlmann Dm, Quantz 0. and Witt U., 1981. Rearing of turbot larvae (Scophthalmus maximus Lm) on cultured food organisms and postmetamorphosis growth on natural and artificial food. Aquaculture 23: 183-1 96.
Last C.M., 1978. The food of three species of gadoid larvae in Eastern English Channel and Southem North Sea. Mar. Biol. 48: 377-386.

Lee CS., Hu F. and Hirano R., 1981. Organisms suitable as food for larvae of Black Sea bream. Progr. Fish Cult. 43(3): 121-125.
Linares F. and Henderson RJ., 1991. Incorporation of C-14 labelled polyunsaturated fatty acids by juvenile turbot, Scophfhalmus maximus (L.) in vivo. J. Fish 6id. 38: 335-347.
Lubzens E., Marko A. and Tietz A., 1985 De novo synthesis of fatty acids in the rotifer, Brachionus plicitais Aquaculture 47: 27- 37.
Marty Y., Delaunay F., Moal J. and Samain J.-F., 1992. Changes in the fatty acid composition of Pecten maximus (L.) during l a ~ a l development. J. Exp. Mar. Biol. Ecol. 163: 221-234.
Mayzaud P., Roche-Mayzaud 0. and Razouls S., 1992. Medium terni time acclimation and digestive enzyme activity in marine copepods: inRuence of food concentration and copepod species. Mar. Ecol. Prog. Ser. 89: 197-212.
McCall J.N., 1992. Source of harpacticoid copepods in the diet of juvenile starry flounder. Mar. Ecol. Prog. Ser. 86: 41-50.
McGeachy S.A., OsFlynn F.M., Benfey T.J. and Friars G.W., 1996. Seawater performance of triploid Atlantic salmon in New Brunswick aquaculture. Bull. Aquacult. Assoc. Canada 96(2): 24- 28.
McLaren I.A. and Avendano P., 1995. Prey field and diet of lawal cod on Western Bank, Scotian Shen. Cm. J. Fish. Aquatic. Sci. 52: 448- 463.
Miliou H. and Moraitou-Apostolopoulou M., 1991 a. Combined effects of temperature and salinity on the population dynamics of Tisbe holothuriae Humes (Copepoda: Harpacticoida) . Archo H ydrobiol. 121(4): 431-448.
Miliou H. and Moraitou-Apostolopoulou M., 1991b. Effects of seven diets on the population dynamics of laboratory cultured Tisbe

holothuriae Humes (Copepoda: Harpactiooida) . Helgol. Meeresunt.
Miliou H. and Moraitou-Apostolopoulou M., 1 991 c. Variations of respiratory rate of Tsbe holothuriae Humes (Copepoda: Harpacticoida) in relation to temperature, salinity and food type. Corn p. Biochem. Physiol. 1 OOA(1): 169-1 74.
Miliou H., MoraitouApostopoulou M. and Argyridou M., 1992. Biochemical composition of fisbe holothut#ae (Copepoda: Harpacticoida) and its differentiation in relation to developmental stages. Ophelia 36(2): 159-1 68.
Moore S., Spackman D.H. and Stein W.H., 1958. Chrornatography of amino acids on sulfonated polystyrene resins: An improved system. Anal. Chem. 30(7): 1 1 85-1 190.
Mourente Go and Tocher D., 1993. Incorporation and metabolisrn of C- 14 labelled polyunsaturated fatty acids in juvenile gilthead sea bream Spams aurata (L.) in vivo. Fish Physiol. Biodiern. 1 O(6): 443-453.
Munilla Moran R.. Stark J.R. and Barbour A., 1990. The role of exogenous enzymes in digestion in cultured turbot l a ~ a e (Scopthalamus maximus L.). Aquaculture 88: 337-50.
Nonbin M.F., Olsen R.-E. and Tande Kasml 1990. Seasonal variation in lipid class and fatty acid composition of two small copepods in Balsfjorden, northem Norway. Mar. Biol. 105: 205-211.
Nonker N. H. and S t ~ t t ~ p J., 1994. The importance of dietary HUFAs for fecundity and HUFA content in the harpacticoid Tisbe holothuriae Humes. Aquaculture 1%: 155-1 66.
Ohno Aml Takahashi T. and Taki Y., 1990. Dynamics of exploited populations of the calanoid copepod Acarlia tsuensis. Aquaculture 84: 27-39.
Pamsh C.C., Castell J.D., Brown J.A., Boston L., Strickland J.S. and Somerton D.C., 1994. Fatty acid compostition of Atlantic halibut

eggs in relation to fetülization. Bull. Aquacult. Assoc. Can. 94(2): 36-38.
Pedersen B.H., Nilsen E.M. and Hjelmeland K., 1987. Variations in the content of trypsin and trypsinogen in larval herring (Clupea harengus) digesüng copepod nauplii. Mar. Biol. 94: 1 71 -1 81 .
Pierce S.K., 1981. Introduction: Symposium on cell volume regulation. J. Exp. 2001. 21 5: 235-236.
Pollero R.J., Gonzalez M.R and l w u C.E.. 1991. Lipid ciasses' consumption related to fasting and themial stress in the shrimp Macrobrachium borellîi. Cornp. Biochem. Physiol. QQB(1): 233-238.
Prahl F.G.8 Eglington G., Corner E.D.S., OnHam S.C.M. and Forsberg T.E.V., 1984. Changes in plant lipids dun'ng passage thiough the gut of Calanus. J. Mar. Biol. Assoc. U.K. 64: 31 7-334.
Pruitt N.L., 1990. Adaptations to temperature in the cellular membranes of cn#itacea: membrane structure and metabolism. J. Them. Biol. 15(1): 1-8.
Ren Ha, Hayashi T., Endo H. and Watanabe E., 1993. Quality evaluation of seasoning extracts on the basis of their beta-buffer capacity and amino acid composition. Nippon Suisan Gakkashai 59(1): 177- 182.
Rieper M.L., 1978. Bacteria as food for marine harpacticoid copepods. Mar. Biol. 45: 337-3445
Rsnnestad 1. and Fyhn H.J., 1993. Metabolic aspects of free amino acids in developing marine fish eggs and lame. Rev. Fish. Sci. l(3): 239-259.
Sargent J.R., Gatten R.R. and Mclntosh R., 1977. Wax esters in the marine environment, their occurence, formation, transformation and ultimate fates. Mar. Chem. 5: 573-584.

Sargent J.R. and Henderson R. J., 19û6. Upids. la; E.D.S. Corner and S.C.M. O'Hara (eds.). The Biological Chemistry of Marine Copepods. Clarendon Press. Oxford, England. pp. 59-1 08.
Scott A.P. and Middleton C., 1979. Unicellular algae as a food for turbot (Scophthalmus maximus L.) lame. The importance of dietary long chain polyunsaturated fatty - acids. Aquaculture 18: 227.240.
Sellner P.A. and Hazd J.R., 1982. Desaturation and elongation of unsaturated fatty acids in hepatocytes of thermally acclimated rainbow trout. Arch. Biochem. Biophys. 21 3(l): 58-66.
Shields R.J., 1992. First feeding of lawal halibut: A review of Norwegian studies. Sea Fish Authority lntemal Report, 11 pp. Ardtoe, Norway.
Sinensky H., 1974. Homeoviscous adaptation-a homeostatic process that regulates the viscosity of membrane lipids in Escherlchia coli. Proc. NatBl. Acad. Sci. U.S.A. 71: 522-525.
Stnrttrup J.G., 1993. First feeding in marine fish lantae: nutritional and environmental aspects. In; B.T. Walther and H.J. Fhyn (eds.). Physiological and Biochemical Aspects of Fish Development. University of Bergen, Norway. pp. 123-131.
Strattnip J.G., Gravningen K. and Norsker N.H., 1995. The role of different algae on th8 growth and suwival of turbot. Scophthlamug maximus larvae in intensive rearing systems. ICES Mar. Sci. Symp. 201 : 173-186.
SWttnip J.O. and Jensen J., 1990. Influence of algal diet on feeding and egg production of the calanoid copepod Acaltia tonsa. Dana. J. Exp. Mar. Biol. 141: 87-105.
Sun B. and Fleeger J.W., 1995. Sustained mass culture of Amphiascoides atopus, a marine harpacticoid copepod in a recirculating systern. Aquaculture 1 36: 31 3-32 1.

Tamani C.S., Murashige R., Lee C.S., Ako Hm and Sato V., 1993. Rotifers fed various diets of bakers' yeast andlor NannochIoropsis oculata and their effect on the growth and survival of striped mullet (Mugi1 cephalus ) and milkfish (Chanos chanos ) larvae. Aquaculture 1 1 0: 361 -372.
Tocher D.R., 1 993. Elongation predominates over deaturation in the metabolism of 18:3n-3 and 20511-3 in turbot (Scophthalmus maximus) brain astroglial cells in pfirnary culture. Lipids 28: 267-272.
Tocher D.R., Carr J. and Sargent J.R., 1989. Polyunsaturated fatty acid metabolism in fish cells: Differential metabolism of (n-3) and (ne) series fatty acids by wltured cells originating from a freshwatei teleost fish and from a marine teleost fish. Comp. Biochem. Physiol. Q4B(2): 367-374.
Tsai C., 1991. Prey density requirements of the striped bass Morone saxitilis lanrae. Estuaries 1 4(2): 207-21 7.
Uhlig G., 1984. Progress in mass cultivation of harpacticoid copepods for mariculture purposes. Spec. Publ. Europ. Maricult. SOC. 8: 261-273.
Ustach J.F., 1982. Algae, bacteria and detritus as food foi the harpacticoid copepod, HetempsyNus pseudonunni- J. Exp. Mar. Biol. €COI. 64: 203-214.
van der Meeren T., 1991. Selective feeding and prediction of food consumption in turbot tanrae (Scophthalmus maximus Lm) reared on the rotifer Brachionus plicatilis and natural zooplankton. Aquaculture 93: 35-55.
Volkman J.K., Jeffiey S.W., Nichols P.D., Rogers G.I. and Garland C.D.,1 989. Fatty acid and lipid composition of 10 species of microalgae used in mariculture. J. Exp. Mar. Biol. 128: 219-240.
Voss A., Reinhart M., Sankarappa S. and Sprecher H., 1991. The metabolism of 7,10,13,16,19-docosapentaenoic acid to 4,7,10,13,16,1Q-docosahexaenoic acid in rat liver is

independent of a 4-desaturase. J. Biol. Chem. 266(30): 19995- 20000.
Walford J., Lim T.M. and Lam T.J., 1991. Replacing live foods with microencapsulated diets in the rearing of seabass (Lates calcarifer) larvae: Do the larvae ingest and digest protein membrane microcapsules? Aquaculture 92: 225-235.
Watanabe T., 1982. Upid nutrition in fish. Comp. Biochem. Physiol. 738(1): 3-1 5.
Watanabe T., 1993. Impoctance of docosahexaenoic acid in marine lawal fish. J. Wodd Aquacult. Soc. 24(2): 152-1 6 1.
Watanabe T., Arakawa T., Kiiima C., Fukusho K and Fujita S., 1978. Nutritional quality of living feed from the viewpoint of essential fatty acids fo i fish. Bull. Jpn. Soc. Sci. Fish. 44(11): 1223- 1227.
Watanabe T., Oowa F., Kitajima C. and Fujita S., 1978. Nutritional quality of brine shrimp Artetnia salina as a living feed from the viewpoint of essential fatty acids. Bull Jpn. Soc. Sci. Fish. 44(lO): 1 1 15-1 121.
White A.W., 1981. Manne zooplankton can accumulate and retain dinoflagellate toxins and cause fish kills. Umnol. Oceanogr. 26(1): 103-1 09.
Wilkinson L., 1990. Systat: The System for Statistics. Systat Inc., Evanston, IL., pp. 485-489.
Witt U., Quantz G., Kuhlmann O. and Kaattner G., 1984. Survival and growth of turbot lawae Scophthalmus maximus reared on different food organisms with special regard to long chah PUFA. Aquacult. Eng. 3: 177-1 90.
Zhang Q. and Uhlig G., 1993. Effect of density on larval development and female productivity of Tisbe holothuriae (Copepoda, Harpacticoida) under laboratory conditions. Helgol. Meersunters 47: 229-241.


















