
Nitrogen-responsiveRegulationofGATAProteinFamily ... · Intracellular Gln3 and Gat1...
17
Nitrogen-responsive Regulation of GATA Protein Family Activators Gln3 and Gat1 Occurs by Two Distinct Pathways, One Inhibited by Rapamycin and the Other by Methionine Sulfoximine * Received for publication, August 8, 2011, and in revised form, October 17, 2011 Published, JBC Papers in Press, October 28, 2011, DOI 10.1074/jbc.M111.290577 Isabelle Georis ‡ , Jennifer J. Tate § , Terrance G. Cooper §1 , and Evelyne Dubois ‡ From the ‡ Institut de Recherches Microbiologiques J. M. Wiame, Laboratoire de Microbiologie Université Libre de Bruxelles, B1070 Brussels Belgium and the § Department of Microbiology, Immunology, and Biochemistry, University of Tennessee Health Science Center, Memphis, Tennessee 38163 Background: Nitrogen availability and TorC1 regulate the localization and function of transcription factors Gln3 and Gat1. Results: Gln3 and Gat1 responses to rapamycin and methionine sulfoximine differ markedly and exhibit different phosphatase requirements. Conclusion: Gln3 and Gat1 localization and function are regulated by two distinct pathways, one inhibited by rapamycin and the other by methionine sulfoximine. Significance: Glutamine starvation regulates Gln3 via a TorC1-independent pathway. Nitrogen availability regulates the transcription of genes required to degrade non-preferentially utilized nitrogen sources by governing the localization and function of transcription acti- vators, Gln3 and Gat1. TorC1 inhibitor, rapamycin (Rap), and glutamine synthetase inhibitor, methionine sulfoximine (Msx), elicit responses grossly similar to those of limiting nitrogen, implicating both glutamine synthesis and TorC1 in the regula- tion of Gln3 and Gat1. To better understand this regulation, we compared Msx- versus Rap-elicited Gln3 and Gat1 localization, their DNA binding, nitrogen catabolite repression-sensitive gene expression, and the TorC1 pathway phosphatase require- ments for these responses. Using this information we queried whether Rap and Msx inhibit sequential steps in a single, linear cascade connecting glutamine availability to Gln3 and Gat1 con- trol as currently accepted or alternatively inhibit steps in two distinct parallel pathways. We find that Rap most strongly elicits nuclear Gat1 localization and expression of genes whose tran- scription is most Gat1-dependent. Msx, on the other hand, elic- its nuclear Gln3 but not Gat1 localization and expression of genes that are most Gln3-dependent. Importantly, Rap-elicited nuclear Gln3 localization is absolutely Sit4-dependent, but that elicited by Msx is not. PP2A, although not always required for nuclear GATA factor localization, is highly required for GATA factor binding to nitrogen-responsive promoters and subse- quent transcription irrespective of the gene GATA factor spec- ificities. Collectively, our data support the existence of two dif- ferent nitrogen-responsive regulatory pathways, one inhibited by Msx and the other by rapamycin. Saccharomyces cerevisiae cells live in a boom and bust nutri- tional environment. At times nitrogen sources are available in abundance, whereas at others they are not, and scavenging for whatever non-preferentially utilized/poor nitrogen sources can be found is imperative to survive (1). The finely tuned responses of yeast to these contrasting environments are controlled by the nitrogen-responsive localizations and functions of the GATA family transcriptional activators Gln3 and Gat1 (2–10). Initial characterization (before 1999) of nitrogen-responsive transcriptional regulation, designated nitrogen catabolite repression (NCR) 2 (1, 11) (Fig. 1, Pathway 1), demonstrated that Gln3 and Gat1 bind to UAS GATA elements in the promot- ers of NCR-sensitive genes. These two GATA factors are the activators responsible for NCR-sensitive transcription when cells are provided with a poor nitrogen source, e.g. proline, with a limiting amount of a preferentially utilized/good nitrogen source or when nitrogen assimilation pathways are impaired, e.g. in gdh1 or gln1 mutants (1, 12–15). Repression of Gln3- and Gat1-dependent, NCR-sensitive transcription depends upon the Ure2 ability to negatively regulate them in nitrogen-rich conditions (16 –19). The isolation of a Gln3-Ure2 complex pro- vided a more concrete biochemical function for Ure2 in this regulation and the beginnings of a molecular basis for NCR (7, 8, 20 –24). This rather primitive view of GATA factor control dramati- cally expanded when several correlations strongly implicated Tor complex 1 kinase (TorC1), Sit4, and glutamine synthetase as primary components of nitrogen-responsive transcription factor localization and transcriptional regulation (Refs. 7–10; see Ref. 5 for a comprehensive review of Tor). Treating cells with the TorC1-specific inhibitor, rapamycin, or the glutamine synthetase inhibitor, methionine sulfoximine (Msx), mimicked * This work was supported, in whole or in part, by National Institutes of Health Grant GM-35642 (to J. J. T. and T. G. C.). This work was also supported by Commission Communautaire Française (to E. D. and I. G.). 1 To whom correspondence should be addressed. Tel.: 901-448-6179; Fax: 901-448-7360; E-mail: [email protected]. 2 The abbreviations used are: NCR, nitrogen catabolite repression; TorC1, Tor complex 1 kinase; Msx, methionine sulfoximine; IP, immunoprecipitation; IN, input; Rap, rapamycin; YNB, yeast nitrogen base; Am, ammonia. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 286, NO. 52, pp. 44897–44912, December 30, 2011 © 2011 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. DECEMBER 30, 2011 • VOLUME 286 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 44897 by guest on September 9, 2019 http://www.jbc.org/ Downloaded from
Transcript of Nitrogen-responsiveRegulationofGATAProteinFamily ... · Intracellular Gln3 and Gat1...

Nitrogen-responsive Regulation of GATA Protein FamilyActivators Gln3 and Gat1 Occurs by Two Distinct Pathways,One Inhibited by Rapamycin and the Other by MethionineSulfoximine*
Received for publication, August 8, 2011, and in revised form, October 17, 2011 Published, JBC Papers in Press, October 28, 2011, DOI 10.1074/jbc.M111.290577
Isabelle Georis‡, Jennifer J. Tate§, Terrance G. Cooper§1, and Evelyne Dubois‡
From the ‡Institut de Recherches Microbiologiques J. M. Wiame, Laboratoire de Microbiologie Université Libre de Bruxelles, B1070Brussels Belgium and the §Department of Microbiology, Immunology, and Biochemistry, University of Tennessee Health ScienceCenter, Memphis, Tennessee 38163
Background:Nitrogen availability and TorC1 regulate the localization and function of transcription factors Gln3 andGat1.Results:Gln3 and Gat1 responses to rapamycin and methionine sulfoximine differ markedly and exhibit different phosphataserequirements.Conclusion: Gln3 and Gat1 localization and function are regulated by two distinct pathways, one inhibited by rapamycin andthe other by methionine sulfoximine.Significance: Glutamine starvation regulates Gln3 via a TorC1-independent pathway.
Nitrogen availability regulates the transcription of genesrequired todegradenon-preferentially utilizednitrogen sourcesby governing the localization and function of transcription acti-vators, Gln3 and Gat1. TorC1 inhibitor, rapamycin (Rap), andglutamine synthetase inhibitor, methionine sulfoximine (Msx),elicit responses grossly similar to those of limiting nitrogen,implicating both glutamine synthesis and TorC1 in the regula-tion of Gln3 and Gat1. To better understand this regulation, wecompared Msx- versus Rap-elicited Gln3 and Gat1 localization,their DNA binding, nitrogen catabolite repression-sensitivegene expression, and the TorC1 pathway phosphatase require-ments for these responses. Using this information we queriedwhether Rap andMsx inhibit sequential steps in a single, linearcascade connecting glutamine availability toGln3 andGat1 con-trol as currently accepted or alternatively inhibit steps in twodistinct parallel pathways.We find thatRapmost strongly elicitsnuclear Gat1 localization and expression of genes whose tran-scription is most Gat1-dependent. Msx, on the other hand, elic-its nuclear Gln3 but not Gat1 localization and expression ofgenes that are most Gln3-dependent. Importantly, Rap-elicitednuclear Gln3 localization is absolutely Sit4-dependent, but thatelicited by Msx is not. PP2A, although not always required fornuclear GATA factor localization, is highly required for GATAfactor binding to nitrogen-responsive promoters and subse-quent transcription irrespective of the gene GATA factor spec-ificities. Collectively, our data support the existence of two dif-ferent nitrogen-responsive regulatory pathways, one inhibitedby Msx and the other by rapamycin.
Saccharomyces cerevisiae cells live in a boom and bust nutri-tional environment. At times nitrogen sources are available inabundance, whereas at others they are not, and scavenging forwhatever non-preferentially utilized/poor nitrogen sources canbe found is imperative to survive (1). The finely tuned responsesof yeast to these contrasting environments are controlled by thenitrogen-responsive localizations and functions of the GATAfamily transcriptional activators Gln3 and Gat1 (2–10).Initial characterization (before 1999) of nitrogen-responsive
transcriptional regulation, designated nitrogen cataboliterepression (NCR)2 (1, 11) (Fig. 1, Pathway 1), demonstratedthat Gln3 and Gat1 bind to UASGATA elements in the promot-ers of NCR-sensitive genes. These two GATA factors are theactivators responsible for NCR-sensitive transcription whencells are provided with a poor nitrogen source, e.g. proline, witha limiting amount of a preferentially utilized/good nitrogensource or when nitrogen assimilation pathways are impaired,e.g. in gdh1 or gln1mutants (1, 12–15). Repression of Gln3- andGat1-dependent, NCR-sensitive transcription depends uponthe Ure2 ability to negatively regulate them in nitrogen-richconditions (16–19). The isolation of a Gln3-Ure2 complex pro-vided a more concrete biochemical function for Ure2 in thisregulation and the beginnings of a molecular basis for NCR (7,8, 20–24).This rather primitive view of GATA factor control dramati-
cally expanded when several correlations strongly implicatedTor complex 1 kinase (TorC1), Sit4, and glutamine synthetaseas primary components of nitrogen-responsive transcriptionfactor localization and transcriptional regulation (Refs. 7–10;see Ref. 5 for a comprehensive review of Tor). Treating cellswith the TorC1-specific inhibitor, rapamycin, or the glutaminesynthetase inhibitor, methionine sulfoximine (Msx), mimicked
* This work was supported, in whole or in part, by National Institutes of HealthGrant GM-35642 (to J. J. T. and T. G. C.). This work was also supported byCommission Communautaire Française (to E. D. and I. G.).
1 To whom correspondence should be addressed. Tel.: 901-448-6179; Fax:901-448-7360; E-mail: [email protected].
2 The abbreviations used are: NCR, nitrogen catabolite repression; TorC1, Torcomplex 1 kinase; Msx, methionine sulfoximine; IP, immunoprecipitation;IN, input; Rap, rapamycin; YNB, yeast nitrogen base; Am, ammonia.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 286, NO. 52, pp. 44897–44912, December 30, 2011© 2011 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
DECEMBER 30, 2011 • VOLUME 286 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 44897
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

the effects of nitrogen limitation, i.e. nuclear Gln3 localizationand high level NCR-sensitive gene expression (7–10, 25). Iden-tification and assembly of these correlations by multiple labo-ratories led to the following molecular model.In repressive, nitrogen-rich conditions, glutamine (or a glu-
taminemetabolite) is thought to activate the TorC1 kinase (Fig.1, Pathway 2) (25). TorC1, in turn, phosphorylates Tap42,which facilitates complex formation betweenTap42, the Sit4 orPP2A phosphatases, and TorC1 (26–30). When in ternarycomplexes with TorC1, Tap42-Sit4 and Tap42-PP2A binarycomplexes are inactive (29). Thus inhibited, these phosphatasesare unable to dephosphorylate Gln3 and thereby facilitate itsdissociation from the Ure2-Gln3 complex, the latter beingrequired for its nuclear localization (8). As a result, the UAS-GATA elements ofNCR-sensitive genes remain unoccupied, andNCR-sensitive transcription is repressed (31, 32).On the other hand, during nitrogen starvation, limitation, or
inhibition of glutamine production by Msx, TorC1 activation,needed for Sit4 and PP2A inactivation, does not occur. Thissituation can also be mimicked by rapamycin treatment, whichcauses release of Tap42-Sit4 or Tap42-PP2A binary complexesfrom TorC1, concomitantly activating Sit4 and PP2A (29).Activated Sit4-Tap42 complexes dephosphorylate Gln3, per-mitting it to dissociate from Ure2, enter the nucleus, bind toUASGATA elements, and activate transcription. The activeTap42-Sit4 and Tap42-PP2A complexes subsequently dissoci-ate, yielding inactive Sit4 or PP2A (29). Although analyzed inmuch less detail, Gat1 was reported to be regulated similarly toGln3 (8).Despite the elegance of the TorC1 regulation model
described above (Fig. 1, Pathway 2), accumulating evidencesuggests the characteristics of nitrogen-responsive GATA fac-tor regulation are not always congruent with the expectations itpredicts (26–30, 32–45). Amajor unanswered question criticalto our understanding of the mechanisms of nitrogen-respon-sive GATA factor regulation is whether glutamine and TorC1control Gln3 and Gat1 localization and/or function via a singlelinear pathway (Pathway 2) or two distinct pathways acting inparallel (Pathways 1 and 2). This is tantamount to asking (i)
whether Msx functions at the top of the TorC1 regulatory cas-cade (Pathway 2 in Fig. 1) or alternatively functions indepen-dently of TorC1 (Pathway 1 in Fig. 1) or (ii) whether there is oneinput signal directing Gln3 and Gat1 to relocate into thenucleus or two input signals.Two observations raised the possibility that rapamycin and
Msx may not function in the same cascade; Msx elicits nuclearGln3 but not Gat1 localization (42), and Msx addition leads toincreasedGln3 phosphorylation, whereas rapamycin treatmentdecreases it (40). Whether additional differences pertinent tothis question exist between Msx- and rapamycin-elicitedresponses is unknown because, in contrast with rapamycin,Msx-elicited responses remain largely uncharacterized.The purpose of this work has been to investigate the require-
ments of Msx-elicited GATA factor regulation at the levels oflocalization, DNA binding, and transcription using Gln3- ver-sus Gat1-specific genes and to compare them with those elic-ited by rapamycin treatment. Because one versus two pathwayGATA factor regulation predicts distinct experimental out-comes, the data we obtained specifically address this question.
MATERIALS AND METHODS
Strains and Culture Conditions—S. cerevisiae strains used inthis work are listed in Table 1. Deletion strains involving inser-tion of kanMX or natMX cassettes were constructed using thelong flanking homology strategy of Wach (46) using primersdescribed previously (43). Growth conditions were identical tothose described in Tate et al. (40, 41). Cultures (50 ml) weregrown to mid-log phase (A600 nm � 0.5–0.55) in YNB minimalammonia (ammonia, 0.1% final concentration) medium.Appropriate supplements (120 �g/ml leucine, 20 �g/ml uracil,histidine, tryptophan, and arginine) were added to the mediumas necessary to cover auxotrophic requirements. For live cellreal time fluorescence microscopy, strains were transformedwith CEN-based plasmids pRS416-Gat1-GFP and pRS416-Gln3-GFP whose construction and detailed validation for nor-mal regulation have appeared (44, 47, 48). Only freshly pre-pared transformants were assayed. Where indicated, cells weretreated with 200 ng/ml rapamycin or 2mMmethionine sulfoxi-
TABLE 1S. cerevisiae strains used in this work
Strain Pertinent genotype Parent Complete genotype Reference
TB50 W.T. MATa, leu2-3, 112, ura3-52, trp1, his3, rme1, HMLa 8TB123 W.T. Gln3-Myc13 MATa, leu2-3, 112, ura3-52, trp1, his4, rme1, HMLa, GLN3-MYC13[KanMX] 8TB136–2a sit4� Gln3-Myc13 MATa, leu2-3, 112, ura3-52, trp1, his4, rme1, HMLa, GLN3-MYC13[KanMX], sit4::kanMX 803644c pph22� Gln3-Myc13 MAT�, leu2-3, 112, ura3-52, trp1, his4, rme1, HMLa, GLN3-MYC13[KanMX],
pph22::kanMX43
03705d pph21�pph22� Gln3-Myc13 MAT�, leu2-3, 112, ura3-52, trp1, his3, rme1, HML�, GLN3-MYC13 [KanMX],pph21::kanMX, pph22::kanMX
43
03879c pph21�pph22� Gat1-Myc13 MAT�, leu2-3,112, ura3-52, trp1, his3, rme1, HML�, GAT1-MYC13[HIS3], pph21::kanMX,pph22::kanMX
44
FV005 gln3� MATa, leu2-3,112, ura3-52, trp1, his3, rme1, HMLa, gln3::kanMX 32FV006 gat1� MATa, leu2-3,112, ura3-52, trp1, his3, rme1, HMLa, gat1::kanMX 32FV014 pph22� TB50 MATa, leu2-3, 112, ura3-52, trp1, his3, rme1, HMLa, pph22::kanMX This workFV018 gat1� Gln3-Myc13 MATa, leu2-3, 112, ura3-52, trp1, his4, rme1, HMLa, GLN3- MYC13 [KanMX],
gat1::natMX32
FV029 sit4� MATa, leu2-3,112, ura3-52, trp1, his3, rme1, HMLa, sit4::natMX 32FV063 W.T. Gat1-Myc13 MATa, leu2-3, 112, ura3-52, trp1, his3, rme1, HMLa, GAT1- MYC13[HIS3] 32FV064 gln3� Gat1-Myc13 MATa, leu2-3, 112, ura3-52, trp1, his3, rme1, HMLa, GAT1- MYC13 [HIS3], gln3::kanMX 32FV066 sit4� Gat1-Myc13 MATa, leu2-3, 112, ura3-52, trp1, his3, rme1, HMLa, GAT1- MYC13 [HIS3], sit4::kanMX 32FV221 pph21�pph22� Gln3-Myc13 03644c MAT�, leu2-3, 112, ura3-52, trp1, his4, rme1, HMLa, GLN3- MYC13 [KanMX],
pph21::natMX, pph22::kanMXThis work
FV239 pph21�pph22� FV014 MATa, leu2-3, 112, ura3-52, trp1, his3, rme1, HMLa, pph21::natMX, pph22::kanMX This work
Yeast GATA Factor Regulation by Rapamycin and Msx
44898 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 52 • DECEMBER 30, 2011
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

mine (final concentrations). Fixed treatment times were 20minfor rapamycin and 30 min for Msx.Quantitative RT-PCR—RNA isolation and cDNA synthesis
were conducted as described in Georis et al. (49). Quantifica-tion of specific cDNA targets was measured by real time PCRperformed on a StepOnePlus device (Applied Biosystems)using primers GDH2-O1 (5�-CCCGACGATTTGATC-GAACA-3�) and GDH2-O2 (5�-TAGCCTTGGCTGAATTC-CGT-3�) or primers that have been described previously (DAL532;GAT1,DAL80, 49). The values reported represent the aver-ages of at least two experiments from independent cultures;error bars indicate S.E.
Chromatin Immunoprecipitation—Cell extracts and immu-noprecipitations were conducted as described in Georis et al.(32, 49). Concentrations of specific DNA targets in immuno-precipitation (IP) and input (IN) samplesweremeasured by realtime PCR performed on a StepOnePlus device (Applied Biosys-tems) using primers GDH2-P1 (5�-GGCCATATGCTC-CAGCTTT-3�), GDH2-P2 (5�-TTCAAGCGGACTTGGTT-TTC-3�), or primers described in Georis et al. (32). The valuesreported represent the averages of two immunoprecipitationsperformed in at least two experiments from independent cul-tures; error bars indicate S.E.
FIGURE 1. Simplified diagrammatic representation, based on data in theliterature, of the two possible pathways responsible for nitrogen-re-sponsive GATA factor regulation. This poses the question, Where does Msxfunction as an inhibitor, at the top of the TorC1 pathway (Pathway 2) or in aseparate pathway (Pathway 1)? Green colors in arrows or molecule designa-tions are used to indicate that a molecule is active or a reaction is facilitated.Red colors, on the other hand, indicate inactive molecules or inhibition of areaction. Phosphorylation of GATA factors and Ure2 are those associated withPathway 2. No specific molecular models related to Msx-elicited phosphoryl-ation have been proposed.
FIGURE 2. Transcription of NCR-sensitive Genes GDH2, GAT1, DAL80, andDAL5 in response to Msx and rapamycin. Total RNA was isolated from wildtype (TB50), gln3� (FV005), and gat1� (FV006) cells grown in YNB-ammoniamedium and treated with rapamycin (�Rap) or methionine sulfoximine(�Msx). Control cells were similarly grown but untreated (Am.). mRNA levelswere quantified by quantitative RT-PCR as described under “Materials andMethods.” GDH2 (panel A), GAT1 (panel B), DAL80 (panel C), and DAL5 (panel D)values were normalized with TBP1.
Yeast GATA Factor Regulation by Rapamycin and Msx
DECEMBER 30, 2011 • VOLUME 286 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 44899
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Yeast GATA Factor Regulation by Rapamycin and Msx
44900 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 52 • DECEMBER 30, 2011
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Intracellular Gln3 and Gat1 Localization—Two methodswere used to determine Gln3 and Gat1 intracellular localiza-tion, indirect immunofluorescence of Gln3-Myc13 and Gat1-Myc13, and real time Gln3-GFP andGat1-GFP fluorescence forthe kinetics of intracellularGln3 andGat1movement. Compre-hensive descriptions of both procedures appear in Tate et al.(41, 43, 44).Images (subsequently designated, primary images) were col-
lected at room temperature using a Zeiss Axioplan 2 micro-scope equippedwith a 100�/1.40 (Myc13 fluorescence) or 63�/1.40 (live cell GFP assay) Plan-Aprochromat oil objective, aZeiss Axio camera, and Zeiss Axiovision 4.1 software. Only pri-mary, unaltered .zvi file images were used for scoring intracel-lular Gln-Myc13, Gat1-Myc13, Gln3-GFP, andGat1-GFP distri-butions as described below. For the microscopic imagespresented, primary .zvi files were converted to .tif files and pro-cessed for publication using Adobe Photoshop and Illustratorsoftware. Gamma settings of microscopic images presented inthis work were unaltered or only minimally altered where nec-essary to avoid loss of detail relative to what was observed in themicroscope. Changes were applied uniformly to the image pre-sented and as similarly as possible from one image to another.Scoring of Intracellular GATA Factor Localization—We
manually scored Gln3-Myc13 and Gat1-Myc13 localization in200 or more cells from multiple, randomly chosen fields fromwhich each immunofluorescence image was taken. All of thecells in the two images collected from each Gln3-GFP or Gat1-GFP culture sample (average number of cells scored per samplewas 91). In all cases, scoring was performed using primary,unaltered .zvi file images and Zeiss AxioVision 4.6.3 software.Cells were classified into one of three categories: cytoplasmic
(cytoplasmic fluorescence only), nuclear-cytoplasmic (fluores-cence in the cytoplasm and the nucleus), or nuclear (fluores-cence exclusively restricted to the nucleus). These categoriesare color-coded as red, yellow, and green bars, respectively, inthe histograms. Criteria applied in the determination of GATAfactor localization, including panels of standard images demon-strating the differences between the categories for both immu-nofluorescence and GFP fluorescence measurements appear inFig. 2 of Tate et al. (43) and Fig. 1 of Tate et al. (44), as hasdetailed evaluations of the precision of the scoring procedures(41, 44). Precision for both methods ranges from �4% whenGATA factors in �80% of the cells are situated in a single cel-lular compartment to �10% when GATA factors are more orless equally distributed in two or three cellular compartments.Further evaluation of scoring precision can also be obtained bycomparing the similar experiments within and between thisand previous publications from our laboratory (32, 41, 43, 44,50, 51). All experiments were performed two or more timeswith similar results.
As a result of the low concentration of Gln3 in the cell andhigh background fluorescence, it is not possible to confidentlydistinguish cells in which Gln3-GFP is nuclear from thosewhere it is nuclear-cytoplasmic (51). Therefore, only two-cate-gory scoring was performed for Gln3-GFP, i.e. it was scoredonly as completely cytoplasmic or nuclear-cytoplasmic (the lat-ter being the sum of nuclear plus nuclear-cytoplasmic valueswhen data are compared with those obtained using indirectimmunofluorescence of Gln3-Myc13).
RESULTS
The experiments described below were designed to addresstwo specific questions. (i) Do Msx and rapamycin inhibitsequential steps in a single linear cascade controlling Gln3 andGat1 (depicted as Pathway 2; Fig. 1) or, alternatively, individualsteps in two distinct parallel pathways (depicted as Pathways 1� 2 with Msx inhibiting only Pathway 1, Fig. 1)? (ii) Do Msx-and rapamycin-inhibited reactions impact equally on bothGln3 and Gat1 regulation, or do the inhibitors exhibit GATAfactor specificity? Alternative answers to each question gener-ate clearly testable predictions. If Msx and rapamycin inhibitsequential steps of a single, TorC1-dependent regulatory path-way (Pathway 2), then we would expect them to elicit uniformeffects on downstream processes, i.e.GATA factor localization,NCR-sensitive transcription, GATA factor DNA binding, andpossess common Tor pathway phosphatase requirements forthe effects. On the other hand, if Msx and rapamycin are situ-ated in distinct pathways, one inhibited by rapamycin but notMsx (Pathway 2) and the other inhibited by Msx but not rapa-mycin (Pathway 1), one might expect differing inhibitor-spe-cific responses on these downstream processes and their phos-phatase requirements. Stating the question in another way,does Msx function in TorC1-dependent Pathway 2 or TorC1-independent Pathway 1?To address these questions, we systematically compared
Msx- and rapamycin-elicited responses and GATA factorrequirements for well established NCR-sensitive catabolicgenes: GDH2, encoding the catabolic NAD-glutamate dehy-drogenase responsible for glutamate degradation and ammoniaproduction (37, 52, 53, 55); GAT1, encoding a GATA-familytranscriptional activator (56–58); DAL80, encoding a GATA-family repressor that down-regulates expression of some butnot allNCR-sensitive genes (49, 59–62);DAL5, encoding allan-toate permease (63, 64). The rationale for choosing these par-ticular genes was that the respective Gln3 and Gat1 contribu-tions to their overall transcriptional activation are known todiffer.NCR-sensitive Genes, GDH2, GAT1, DAL80, and DAL5
Respond Differently to Msx and Rapamycin—We initiated ourinvestigation by assessing the transcription profiles of GDH2,
FIGURE 3. Localization of Gln3-Myc13 and Gat1-Myc13 in response to Msx and rapamycin. Wild type (TB123), pph21�pph22� (03705d), or sit4� (TB136-2a)strains, containing Gln3-Myc13 (panels A and B) or wild type (FV063), pph21�pph22� (03879c), or sit4� (FV066) strains containing Gat1-Myc13 (panels C and D)were grown in YNB-ammonia medium. The cultures were sampled for indirect immunofluorescence assay of Gln3-Myc13 or Gat1-Myc13 localization (Am.).Rapamycin (�Rap) or methionine sulfoximine (�Msx) was then added, and the cultures were sampled again 20 and 30 min later, respectively. Indirectimmunofluorescence assays were performed and imaged as described under “Materials and Methods.” For each histogram, cells were scored for intracellularGln3-Myc13 or Gat1-Myc13 localization (cytoplasmic, red bars, nuclear-cytoplasmic, yellow bars; nuclear, green bars) using criteria described under “Materials andMethods.” Multiple examples of images in each category may be found in Fig. 2 of Tate et al. (43). Images from which the histograms were derived are shownabove each histogram. The upper member of each pair (panels A and C) depicts red Gln3-Myc13- or Gat1-Myc13-derived fluorescence, and the lower one showsDAPI-positive material fluorescence. The precision of the IF assays we employ has been analyzed in detail and is described under “Materials and Methods.”
Yeast GATA Factor Regulation by Rapamycin and Msx
DECEMBER 30, 2011 • VOLUME 286 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 44901
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
GAT1,DAL80, andDAL5 in ammonia-grown cells treated witheitherMsx or rapamycin.Msx, but not rapamycin, elicited highlevel GDH2 transcription. This transcription was dependentonly on Gln3 as shown by the fact that Msx-induced transcrip-tion was eliminated in a gln3� but unaffected in a gat1� (Fig.2A). A similar result was obtained for GAT1 transcription; itwas highly induced by Msx in a Gln3-dependent manner, withno transcription being elicited by rapamycin addition (Fig. 2B).In contrast with these two genes, which exhibited a response toonly one inhibitor and one GATA family activator, DAL80transcription increased after the addition of eitherMsx or rapa-mycin, stimulation being 5-fold greater with rapamycin thanwith Msx (Fig. 2C). However, the two inhibitors exhibitedGATA factor specificities. Msx-induced DAL80 transcriptionwas completely Gat1- and Gln3-dependent, whereas rapamy-cin-induced DAL80 transcription was absolutely Gat1-depen-dent but only partially required Gln3. Finally, DAL5 transcrip-tion responded about equally to both Msx or rapamycinaddition, and both transcription factors were required, but thespecificity was less obvious (Fig. 2D). The Msx response wasabsolutely Gln3-dependent and strongly Gat1-dependent(therewas a small response toMsx in the gat1�). Gat1 andGln3were equally required for rapamycin-induced DAL5 transcrip-tion. Overall, the GDH2, GAT1, DAL80, and DAL5 transcrip-tional responses elicited by Msx and rapamycin treatmentswere not uniform. They were gene-, GATA family activator-,and inhibitor-specific.Response of Intracellular Gln3 and Gat1 Localization to Msx
and Rapamycin Addition—The above data demonstrated cleardifferences in GATA factor-specific transcriptional responsestoMsx and rapamycin.Our next objectivewas to determine theextent to which these observations correlated with Msx- and
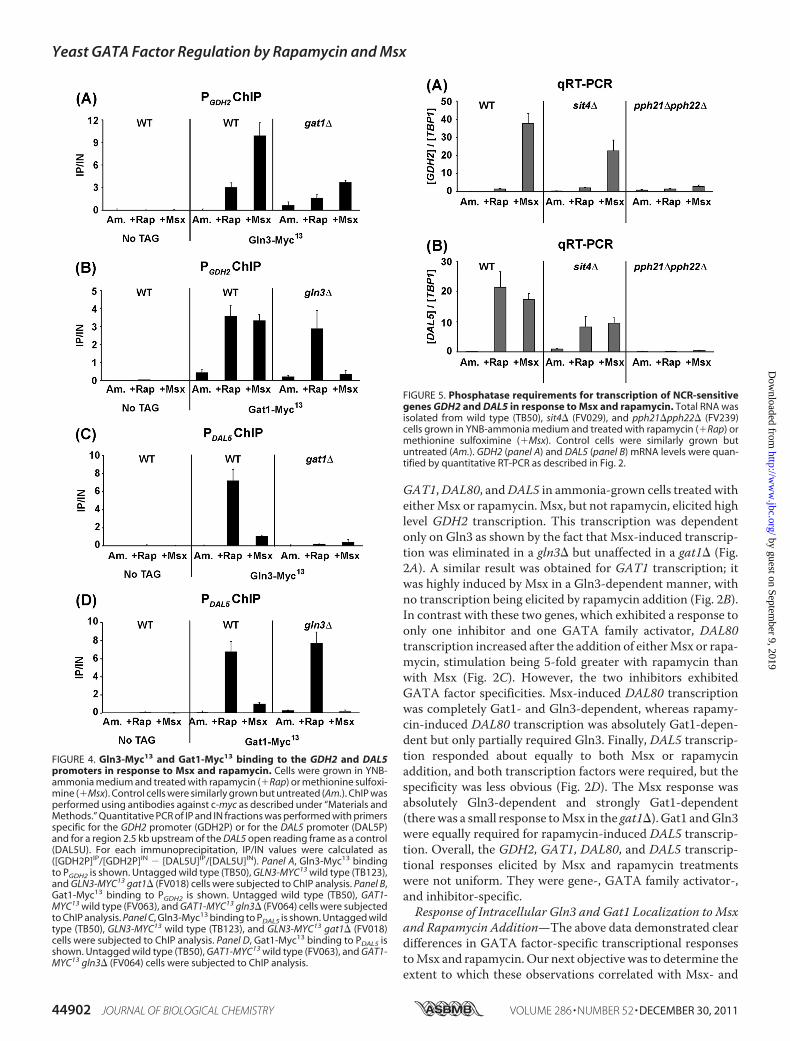
FIGURE 4. Gln3-Myc13 and Gat1-Myc13 binding to the GDH2 and DAL5promoters in response to Msx and rapamycin. Cells were grown in YNB-ammonia medium and treated with rapamycin (�Rap) or methionine sulfoxi-mine (�Msx). Control cells were similarly grown but untreated (Am.). ChIP wasperformed using antibodies against c-myc as described under “Materials andMethods.” Quantitative PCR of IP and IN fractions was performed with primersspecific for the GDH2 promoter (GDH2P) or for the DAL5 promoter (DAL5P)and for a region 2.5 kb upstream of the DAL5 open reading frame as a control(DAL5U). For each immunoprecipitation, IP/IN values were calculated as([GDH2P]IP/[GDH2P]IN � [DAL5U]IP/[DAL5U]IN). Panel A, Gln3-Myc13 bindingto PGDH2 is shown. Untagged wild type (TB50), GLN3-MYC13 wild type (TB123),and GLN3-MYC13 gat1� (FV018) cells were subjected to ChIP analysis. Panel B,Gat1-Myc13 binding to PGDH2 is shown. Untagged wild type (TB50), GAT1-MYC13 wild type (FV063), and GAT1-MYC13 gln3� (FV064) cells were subjectedto ChIP analysis. Panel C, Gln3-Myc13 binding to PDAL5 is shown. Untagged wildtype (TB50), GLN3-MYC13 wild type (TB123), and GLN3-MYC13 gat1� (FV018)cells were subjected to ChIP analysis. Panel D, Gat1-Myc13 binding to PDAL5 isshown. Untagged wild type (TB50), GAT1-MYC13 wild type (FV063), and GAT1-MYC13 gln3� (FV064) cells were subjected to ChIP analysis.
FIGURE 5. Phosphatase requirements for transcription of NCR-sensitivegenes GDH2 and DAL5 in response to Msx and rapamycin. Total RNA wasisolated from wild type (TB50), sit4� (FV029), and pph21�pph22� (FV239)cells grown in YNB-ammonia medium and treated with rapamycin (�Rap) ormethionine sulfoximine (�Msx). Control cells were similarly grown butuntreated (Am.). GDH2 (panel A) and DAL5 (panel B) mRNA levels were quan-tified by quantitative RT-PCR as described in Fig. 2.
Yeast GATA Factor Regulation by Rapamycin and Msx
44902 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 52 • DECEMBER 30, 2011
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
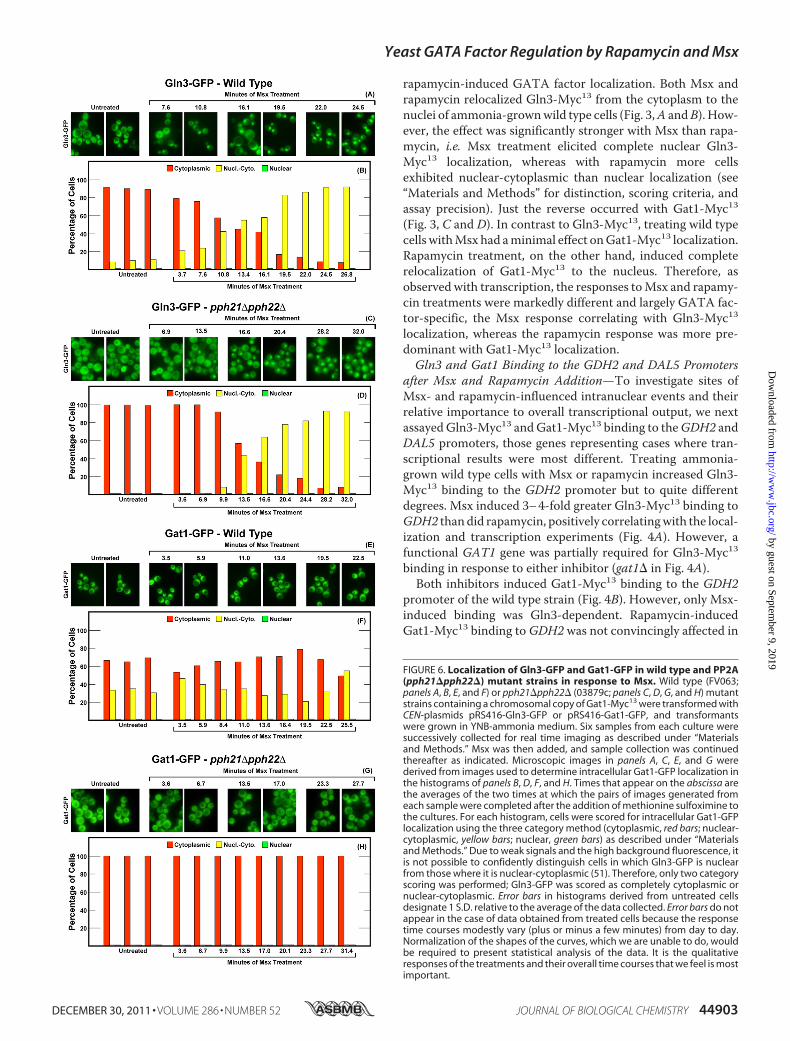
rapamycin-induced GATA factor localization. Both Msx andrapamycin relocalized Gln3-Myc13 from the cytoplasm to thenuclei of ammonia-grownwild type cells (Fig. 3,A andB). How-ever, the effect was significantly stronger with Msx than rapa-mycin, i.e. Msx treatment elicited complete nuclear Gln3-Myc13 localization, whereas with rapamycin more cellsexhibited nuclear-cytoplasmic than nuclear localization (see“Materials and Methods” for distinction, scoring criteria, andassay precision). Just the reverse occurred with Gat1-Myc13(Fig. 3, C and D). In contrast to Gln3-Myc13, treating wild typecells withMsx had aminimal effect onGat1-Myc13 localization.Rapamycin treatment, on the other hand, induced completerelocalization of Gat1-Myc13 to the nucleus. Therefore, asobserved with transcription, the responses toMsx and rapamy-cin treatments were markedly different and largely GATA fac-tor-specific, the Msx response correlating with Gln3-Myc13localization, whereas the rapamycin response was more pre-dominant with Gat1-Myc13 localization.Gln3 and Gat1 Binding to the GDH2 and DAL5 Promoters
after Msx and Rapamycin Addition—To investigate sites ofMsx- and rapamycin-influenced intranuclear events and theirrelative importance to overall transcriptional output, we nextassayedGln3-Myc13 andGat1-Myc13 binding to theGDH2 andDAL5 promoters, those genes representing cases where tran-scriptional results were most different. Treating ammonia-grown wild type cells with Msx or rapamycin increased Gln3-Myc13 binding to the GDH2 promoter but to quite differentdegrees. Msx induced 3–4-fold greater Gln3-Myc13 binding toGDH2 thandid rapamycin, positively correlatingwith the local-ization and transcription experiments (Fig. 4A). However, afunctional GAT1 gene was partially required for Gln3-Myc13binding in response to either inhibitor (gat1� in Fig. 4A).
Both inhibitors induced Gat1-Myc13 binding to the GDH2promoter of the wild type strain (Fig. 4B). However, only Msx-induced binding was Gln3-dependent. Rapamycin-inducedGat1-Myc13 binding toGDH2 was not convincingly affected in
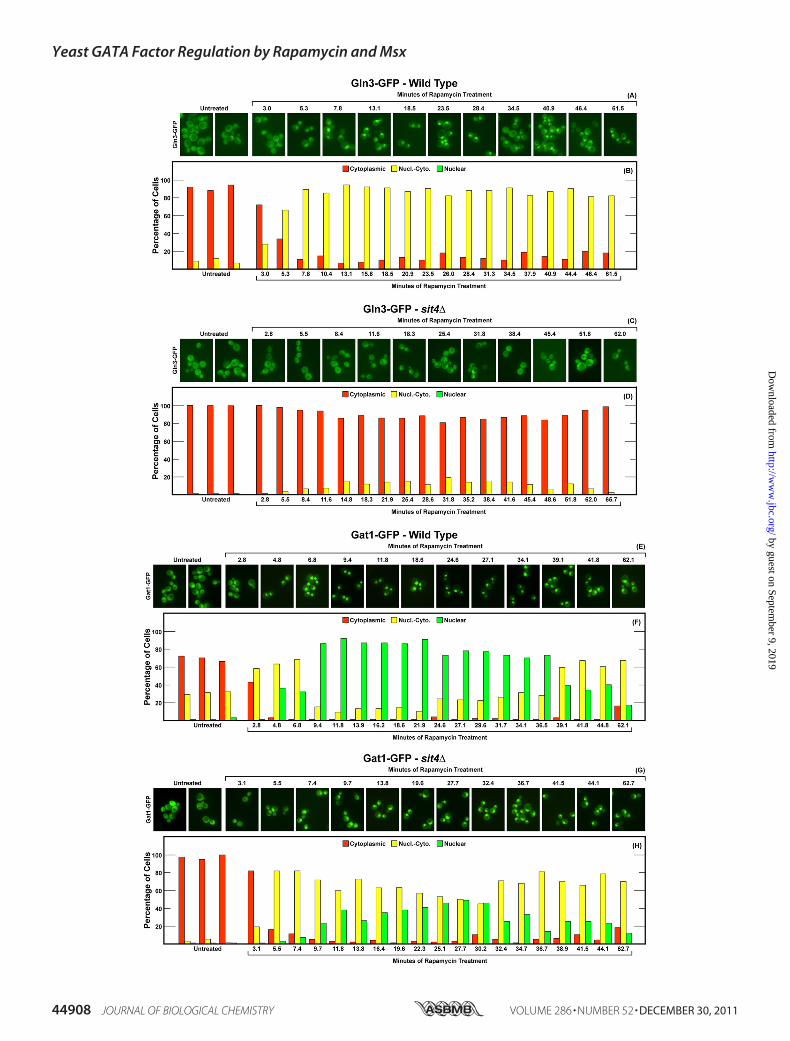
FIGURE 6. Localization of Gln3-GFP and Gat1-GFP in wild type and PP2A(pph21�pph22�) mutant strains in response to Msx. Wild type (FV063;panels A, B, E, and F) or pph21�pph22� (03879c; panels C, D, G, and H) mutantstrains containing a chromosomal copy of Gat1-Myc13 were transformed withCEN-plasmids pRS416-Gln3-GFP or pRS416-Gat1-GFP, and transformantswere grown in YNB-ammonia medium. Six samples from each culture weresuccessively collected for real time imaging as described under “Materialsand Methods.” Msx was then added, and sample collection was continuedthereafter as indicated. Microscopic images in panels A, C, E, and G werederived from images used to determine intracellular Gat1-GFP localization inthe histograms of panels B, D, F, and H. Times that appear on the abscissa arethe averages of the two times at which the pairs of images generated fromeach sample were completed after the addition of methionine sulfoximine tothe cultures. For each histogram, cells were scored for intracellular Gat1-GFPlocalization using the three category method (cytoplasmic, red bars; nuclear-cytoplasmic, yellow bars; nuclear, green bars) as described under “Materialsand Methods.” Due to weak signals and the high background fluorescence, itis not possible to confidently distinguish cells in which Gln3-GFP is nuclearfrom those where it is nuclear-cytoplasmic (51). Therefore, only two categoryscoring was performed; Gln3-GFP was scored as completely cytoplasmic ornuclear-cytoplasmic. Error bars in histograms derived from untreated cellsdesignate 1 S.D. relative to the average of the data collected. Error bars do notappear in the case of data obtained from treated cells because the responsetime courses modestly vary (plus or minus a few minutes) from day to day.Normalization of the shapes of the curves, which we are unable to do, wouldbe required to present statistical analysis of the data. It is the qualitativeresponses of the treatments and their overall time courses that we feel is mostimportant.
Yeast GATA Factor Regulation by Rapamycin and Msx
DECEMBER 30, 2011 • VOLUME 286 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 44903
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Yeast GATA Factor Regulation by Rapamycin and Msx
44904 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 52 • DECEMBER 30, 2011
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

a gln3� mutant. The fact that Msx induced as much Gat1-Myc13 binding to GDH2 as did rapamycin was somewhat sur-prising given the nearly complete lack of effect ofMsx onGat1-Myc13 localization (Fig. 3, C and D). A possible explanation ofthis observation is that there is a substantial amount of nuclearGat1-Myc13 in untreated, ammonia-grown cells. If Gln3 facili-tated Gat1 binding, as Gat1 does for Gln3 binding (32), thenMsx addition, which greatly increases nuclear Gln3-Myc13localization, would generate enhanced binding of the limitedamount of Gat1-Myc13 already in the nucleus. Consistent withthis explanation is the fact thatMsx-inducedGat1-Myc13 bind-ing to GDH2 was Gln3-dependent, whereas rapamycin-in-duced Gat1-Myc13 binding was not.
The overall pattern of inhibitor-induced Gln3-Myc13 bind-ing to the DAL5 promoter was reversed relative to binding toGDH2. Gat1-dependent Gln3-Myc13 binding at theDAL5 pro-moter was induced by bothMsx and rapamycin, but rapamycinwas 7-fold more effective than Msx (Fig. 4C) despite the factthat Gln3-Myc13 was less nuclear in rapamycin-treated than inMsx-treated cells.Msx- and rapamycin-induced Gat1-Myc13 binding to the
DAL5 promoter also differed dramatically. Rapamycin inducedalmost a full order of magnitude more Gat1-Myc13 binding towild type ammonia-grown cells than did Msx (Fig. 4D). Theseeffects positively correlated with the effect of rapamycin andMsx on Gat1-Myc13 localization (Fig. 3, C and D). Again, thesmall amount of Msx-induced Gat1-Myc13 binding was fullyGln3-dependent, whereas rapamycin-induced Gat1-Myc13binding was unaffected in the gln3�. Overall, we observedgreater positive correlation between the effects of Msx withGln3 function than that of Gat1 (localization, binding, andGln3-dependent transcription) and, conversely, amore positivecorrelation with the effects of rapamycin and Gat1 function.Pph21/22 and Sit4 Requirements for Msx- and Rapamycin-
induced GDH2 and DAL5 Transcription—To assess whethercorrelations between the inhibitor responses and the GATAfactors required for them could be extended further, we deter-mined the phosphatase requirements forMsx- and rapamycin-induced transcription. Pph21/22was required forMsx-inducedGDH2 transcription in that the levels decreased almost 10-foldin a pph21�pph22� mutant relative to wild type (Fig. 5A). Incontrast, Sit4 was only partially required for this transcription,GDH2 transcription decreasing by less than half in the sit4�(Fig. 5A). Inasmuch as rapamycin-inducedGDH2 transcriptionwas minimal (Fig. 5A), its requirements were considered moot.In the case of DAL5, both inhibitors triggered transcription,
thus providing an opportunity to determine their respectivephosphatase requirements. Pph21/22 was required for bothMsx- and rapamycin-induced DAL5 transcription; only basallevels were observed in the pph21�pph22�mutant (Fig. 5B). Incontrast, Sit4 was only partially required for both Msx- andrapamycin-inducedDAL5 transcription (Fig. 5B). Overall, irre-spective of the inhibitor added, Pph21/22was strongly required
to induceGDH2 andDAL5 transcription, whereas Sit4was onlypartially required.Effects of Eliminating Pph21/22 and Sit4 Activities on Msx-
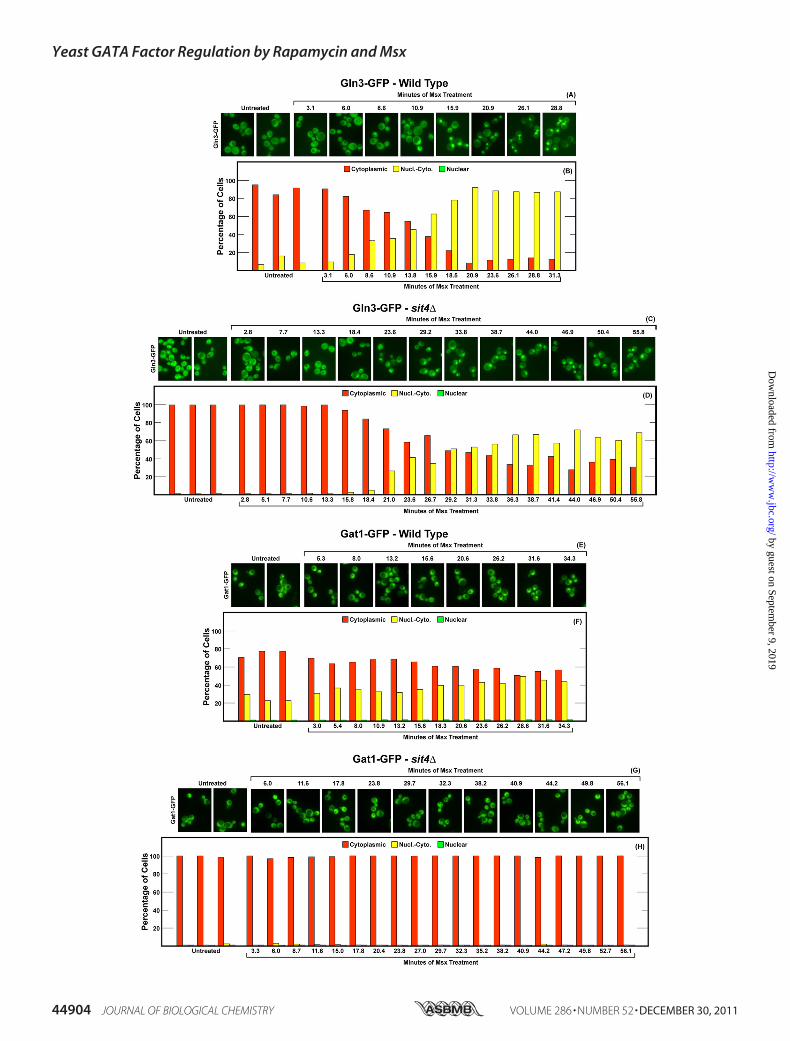
influenced Gln3-GFP and Gat1-GFP Localization—The abovetranscription prompted us to analyze the phosphatase require-ments of Msx-induced GATA factor localizations and thekinetics of their movements. Gln3-GFP was cytoplasmic in�90% of ammonia-grown wild type cells and promptly beganrelocating to the nucleus after Msx addition (Fig. 6, A and B).Even though high background fluorescence did not permitthree-category scoring of Gln3-GFP localization (see explana-tion in “Materials and Methods”), the effect of Msx on Gln3-GFP localization was clear. The fraction of cells exhibitingexclusively cytoplasmic Gln3-GFP fell from �90% to around10% in24–27min. Therewas no further change out to 60min(data not shown). These results correlated well with thoseobserved with Gln3-Myc13, which is amenable to three cate-gory scoring (Fig. 3, A and B).
The response of Gln3-GFP localization to Msx addition inthe pph21�pph22� mutant was indistinguishable from that inthe wild type except that 9–10 additional min elapsed betweenMsx addition and Gln3-GFP movement (Fig. 6, C and D). Athigher scoring resolution, using the Myc13-based indirect fluo-rescence assay, a measurable Pph21/22 requirement could bedetected (Fig. 3, A and B). Irrespective of the assay, however,Msx relocalized a major portion of Gln3 to the nucleus in alargely Pph21/22-independent manner.Gat1-GFP behaved quite differently from Gln3-GFP. First,
Gat1-GFP was more nuclear than Gln3-GFP in untreated,ammonia-grown, wild type cells, a reflection of its lower sensi-tivity to NCR regulation (Fig. 6, E and F). Foremost, however,Msx had only a small effect on Gat1-GFP localization (Fig. 6, Eand F). In an untreated pph21�pph22� mutant, nuclear-cyto-plasmic Gat1-GFP localization completely disappeared andwas not affected by the addition of Msx (Fig. 6, G and H). Incontrast, using the Myc13-based indirect fluorescence assay,Gat1-Myc13 was nuclear-cytoplasmic in 20% Msx-treatedpph21�pph22� cells (Fig. 3, C and D).The loss of Sit4 elicited a somewhat greater effect on Msx-
induced Gln3-GFP localization than did that of Pph21/22 (Fig.7, C and D). Nuclear-cytoplasmic plus nuclear Gln3-GFPdecreased from around 90% that of wild type cells to about60–70% that in the sit4�, indicating that Msx-induced nuclearGln3-GFP localization was only slightly Sit4-dependent. Thismodest Sit4 requirement was less obvious using the Gln3-Myc13 indirect fluorescent assay (Fig. 3, A and B) in that Msx-treated sit4� cells with nuclear Gln3-Myc13 localizationdecreased by only about 10–15%. The greatest differenceoccurred in the kinetics of the response. Loss of Pph21/22resulted in9–10min lag before themutant cells responded toMsx (compare Fig. 6, B with D). In contrast, the loss of Sit4doubled the lag to 19–20 min. The transition of Gln3-GFP
FIGURE 7. Localization of Gln3-GFP and Gat1-GFP in wild type and sit4� mutant strains in response to Msx. The experimental format and data presen-tation were the same as those in Fig. 7 except for the strains used as transformation recipients. In this experiment they were wild type (TB50; panels A, B, E, andF) and sit4� (FV029; panels C, D, G, and H) containing an untagged chromosomal copy of GAT1.
Yeast GATA Factor Regulation by Rapamycin and Msx
DECEMBER 30, 2011 • VOLUME 286 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 44905
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Yeast GATA Factor Regulation by Rapamycin and Msx
44906 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 52 • DECEMBER 30, 2011
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

from the cytoplasm in wild type cells was nearly completebefore it ever started in the sit4� (Compare Fig. 7, B and D).
For reasons that we cannot explain, two opposite results havebeen reported for characterizations of the effects of Msx onGln3-Myc13 localization. In the initial report describing Msx-elicited nuclear Gln-Myc13 localization, Sit4 was required forthe effect (Fig. 2A of Ref. 25). As seen in this work (Fig. 3,A andB), however, we did not observe a Sit4 requirement for Msx-elicited nuclear Gln3-Myc13 localization. Although the samestrains were assayed, there wereminor differences in themediaused in the present and earlier experiments. Therefore, werepeated the Gln3-Myc13 localization experiments using con-ditions identical to those initially reported but were still unableto obtain a demonstrable Sit4 requirement (data not shown).We did observe a modest effect of Sit4 loss when the kinetics ofMsx-elicited Gln3-GFP relocalization were followed (compareFig. 7, A and Bwith C andD), but it was minor when comparedwith that reported earlier. A possible explanation for the differ-ing observations might be that Gln3-Myc13 localization wasquantitatively determined in the present work, whereas only asingle image was reported earlier. However, this reasoningwould not explain that other equally important differenceswere observed between the initial and subsequent reports ofMsx-elicited Gln3-Myc13 phosphorylation (25, 40).In untreated andMsx-treated sit4� cells (Fig. 7,G andH), as
in pph21/22� cells (Fig. 6,G andH), nuclear-cytoplasmicGat1-GFP localizationwas eliminated. In sit4�mutant cells,Msx hadno effect whatever on Gat1-GFP localization (Fig. 7, G and H),unlike the small amount of Msx elicited relocalization in thewild type (Fig. 7, E and F). However, as in pph21/22� cells,nuclear-cytoplasmic Gat1-GFP localization was eliminated inuntreated and Msx-treated sit4� cells. In contrast with theGat1-GFP assay, the loss of Sit4 had little effect on nuclear-cytoplasmic or nuclear Gat1-Myc13 localization that occurredin20–30% of untreated orMsx-treated cells (Fig. 3,C andD).Potential explanations for this differencewill be discussed later.Overall, the effects of Msx on GATA factor localization in
ammonia-grown cells were almost exclusively restricted toGln3.Msx effects onGat1 localization wereminimal. Althoughthere was detectable deterioration in the extent of nuclear Gln3localization when Pph21/22 (Fig. 3, A and B) or Sit4 (Fig. 7, Cand D) activities were eliminated, the effects were minimal tomodest at best, i.e. nuclear Gln3 localization in response toMsxaddition was largely Pph21/22- and Sit4-independent.Effects of Eliminating Pph21/22 and Sit4 Activities on Rapa-
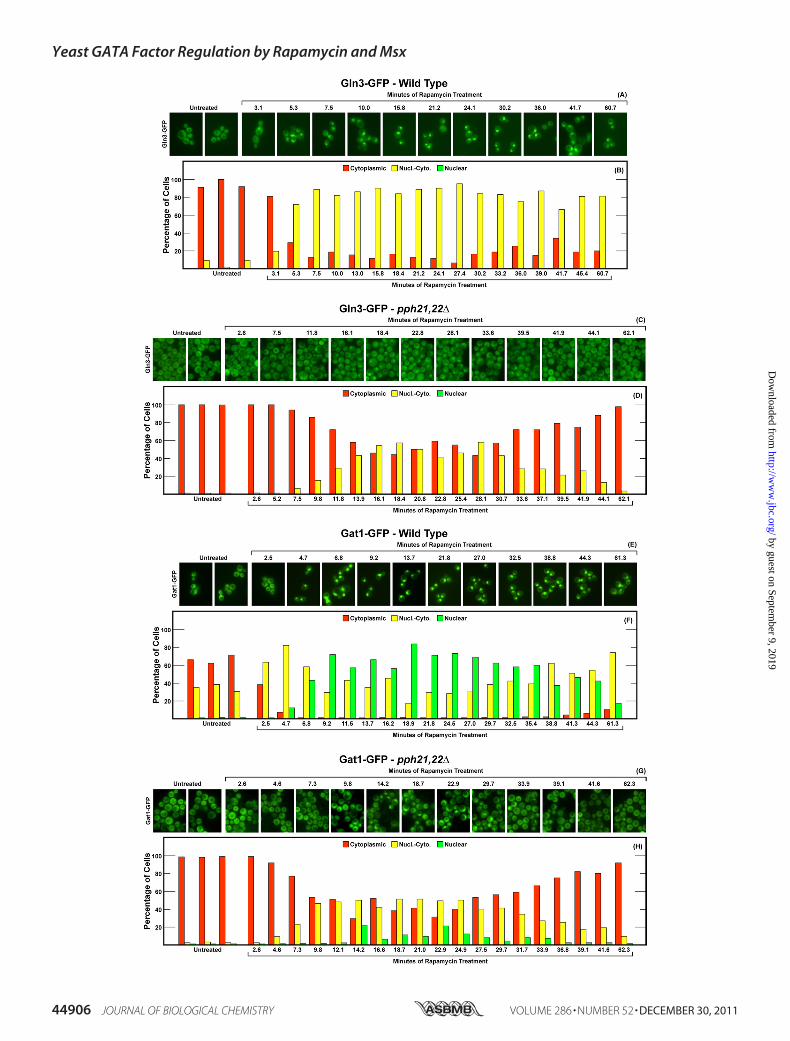
mycin-influenced Gln3-GFP and Gat1-GFP Localization—Thekinetics of rapamycin-induced Gln3-GFP movement in wildtype cells differed from those of Msx. The response to rapamy-cin was faster, being largely complete in 5–7 min, whereas bythis time after Msx addition, Gln3-GFP movement into thenucleus was just getting well under way (compare Fig. 8, A andB, with Fig. 6, A and B). Furthermore, the loss of Pph21/22,which minimally affected Msx-induced Gln3-GFP movement,resulted in a moderate decrease in the degree to which Gln3-
GFP moved from the cytoplasm into the nuclear-cytoplasmicand nuclear scoring categories (Fig. 8, C and D). Additionally,the response in the pph21�pph22� became transitory withGln3-GFP remaining maximally nuclear for only 10–12min.In contrast with the lack of a response to Msx, Gat1-GFP
rapidly relocated to the nucleus after rapamycin addition (Fig.8, E and F). Although there was some evidence that Gat1-GFPbegan to leave the nuclei of rapamycin-treated cells, the effectwas relatively modest (Fig. 8, E and F). Gat1-GFP relocalizationfrom the cytoplasm to the nuclei of pph21�pph22� mutantcells, on the other hand, was markedly muted and clearly tran-sient (Fig. 8, G and H). There was also noticeable similarity inthe kinetics of Gat1-GFP and Gln3-GFP responses to rapamy-cin in the pph21�pph22�mutant (compare Fig. 8,C andDwithG and H).The effect of deleting SIT4 on rapamycin-elicited nuclear
localization of Gln3-GFP was far stronger than that ofPph21/22 (compare Fig. 9,C andD, with Fig. 8,C andD) or thatfor Msx responses in either pph21�pph22� or sit4� (compareFig. 9, C and D, with Fig. 7, C and D, and Fig. 8, C and D). TheSit4 requirement for rapamycin-elicited nuclear localization ofGat1-GFP was more modest than was that of Pph21/22 (com-pare Fig. 9, G and H with Fig. 8, G and H) or that of Sit4 forGln3-GFP (Fig. 9, G and H with Fig. 8, C and D).
One additional parenthetic result emanates from Figs. 6 and7. Occasions arise when double-tagged strains are required,raising the question of whether the dual presence of Gat1-Myc13 and Gln3-GFP or Gat1-GFP influences the outcomesobserved. To comprehensively assess this possibility, all of theexperiments in Figs. 6–9 were performed both in strains thatdid or did not contain Gat1-Myc13. The presence of Gat1-Myc13 had no convincing effect on either wild type or mutantoutcomes. This conclusion can be directly assessed by compar-ing Gln3-GFP and Gat1-GFP results observed in the wild typestrains in Figs. 6 and 7.Effects of Eliminating Pph21/22 and Sit4 Activities on Msx
and Rapamycin-influenced Gln3-Myc13 and Gat1-Myc13 Bind-ing to the GDH2 and DAL5 Promoters—To ascertain thePph21/22 and Sit4 requirements for Msx- and rapamycin-in-duced GATA factor binding to the GDH2 and DAL5 promot-ers, we performed ChIP assays in wild type and mutant strains.Msx induced high levels Gln3-Myc13 binding to theGDH2 pro-moter that was substantially Sit4- and Pph21/22-independent(Fig. 10A). These observations correlated well with Gln3 local-ization as in all three instances there was substantial nuclearGln3. Surprisingly, despite its reduced nuclear localization inthe phosphatase mutants, rapamycin-induced Gln3-Myc13binding to GDH2 was largely Sit4- and partially Pph21/22-in-dependent (Fig. 10A).Msx-induced Gat1-Myc13 binding was partially Sit4-inde-
pendent, whereas binding elicited by rapamycin treatment wasfully Sit4-independent (Fig. 10B). In the pph21�pph22�, Gat1-Myc13 binding toGDH2, whether triggered byMsx or rapamy-cin, was minimally over background levels, demonstrating a
FIGURE 8. Localization of Gln3-GFP and Gat1-GFP in wild type and pph21�pph22� mutant strains in response to rapamycin. The experimental formatand data presentation were the same as those in Fig. 7 with two exceptions. First, the strains used as transformation recipients were wild type (TB50; panels A,B, E, and F) and pph21�pph22� (FV239; panels C, D, G, and H). Second, rapamycin replaced methionine sulfoximine as the inhibitor added to the cultures.
Yeast GATA Factor Regulation by Rapamycin and Msx
DECEMBER 30, 2011 • VOLUME 286 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 44907
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Yeast GATA Factor Regulation by Rapamycin and Msx
44908 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 52 • DECEMBER 30, 2011
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Pph21/22 requirement correlating with the requirement ofPP2A for Gat1 localization.When analogous experiments were performed with the
DAL5 promoter, Gln3-Myc13 binding to the DAL5 promoterwas both Sit4- and Pph21/22-dependent (Fig. 10C). In contrastwith Gln3, Gat1-Myc13 binding to DAL5 responded asexpected, being greatly stimulated by rapamycin in a Sit4-inde-
pendent manner (Fig. 10D). The Pph21/22 requirement forGat1-Myc13 binding toDAL5 paralleled its subcellular localiza-tion and the transcriptional results (Fig. 5B). Overall, in mostcases Sit4 was never strongly required for GATA factor bindingon the two promoters tested. Pph21/22 wasmostly required forGat1 binding and also, probably as a consequence, for Gln3binding at the DAL5 promoter.
DISCUSSION
The evidence presented above suggests that Msx and rapa-mycin inhibit distinct nitrogen-responsive regulatory pathwaysrather than sequential steps in a single linear cascade. Thus, theMsx-inhibited glutamine synthetase reaction is not situated atthe top of the TorC1 pathway (Pathway 2, Fig. 1) as currentlyaccepted (25), but is situated in a separate pathway (Location 1,in Fig. 1). This conclusion implies the existence of at least twodistinct nitrogen-responsive inputs regulating Gln3 and Gat1.In addition to answering the question of one pathway or two,the data presented demonstrate that Gln3 and Gat1 eachexhibit remarkably strong sensitivity to regulation by one of thetwo pathways relative to the other. These conclusions are basedon GATA factor responses to Msx and rapamycin treatmentmeasured at three different levels: GATA factor localization,binding to nitrogen-responsive promoters, and GATA factor-mediated transcription.At the level of GATA factor localization inwild type cells, the
responses were both GATA factor- and inhibitor-specific. Msxrelocalized Gln3 from the cytoplasm to the nuclei of essentiallyall ammonia-grown cells but minimally if at all affected Gat1localization. In contrast, rapamycin relocalized Gat1 to thenuclei of essentially all cells but had only a moderate compara-tive effect onGln3 localization, i.e.Gln3was completely nuclearin less than half the cells.Transcriptional responses as well as the GATA factor
requirements for these responses were inhibitor-specific. Inaddition, the responses and GATA factor requirements werealso gene-specific, reflecting the contribution of promoterstructure to the overall end response. GDH2 transcription wasstrongly induced byMsx treatment in a highly Gln3-dependentandGat1-independentmanner. Rapamycin, on the other hand,was strikingly ineffective; no response whatever was observed.GAT1 transcription behaved similarly. DAL80 transcriptionwas intermediate in the spectrumof responses. Rapamycin elic-ited amuch greater response than didMsx, that response beingGat1-dependent but also partially Gln3-dependent. Not sur-prisingly given Gln3 participation, Msx also elicited moderateDAL80 transcription that was absolutely Gln3-dependent. Atthe other end of the spectrum, Msx and rapamycin elicitedequal levels of DAL5 transcription that was largely dependenton both GATA factors. Thus at transcription, as with localiza-tion,Msx and rapamycin elicited responses correlated bestwithGln3 and Gat1, respectively. Importantly, these results empha-size the inadequacy of using nitrogen catabolic gene expressionas a reporter of nitrogen-responsive regulation in the absence of
FIGURE 9. Localization of Gln3-GFP and Gat1-GFP in wild type and sit4� mutant strains in response to rapamycin. The experimental format and datapresentation were the same as those in Fig. 7 with two exceptions. First, the strains used as transformation recipients were wild type (TB50; panels A, B, E, andF) and sit4� (FV029; panels C, D, G, and H). Second, rapamycin replaced methionine sulfoximine as the inhibitor added to the cultures.
FIGURE 10. Phosphatase requirements for Gln3-Myc13 and Gat1-Myc13
binding to the GDH2 and DAL5 promoters promoter in response to Msxand rapamycin. Cells were grown in YNB-ammonia medium and treatedwith rapamycin (�Rap) or methionine sulfoximine (�Msx). Control cells weresimilarly grown but untreated (Am.). ChIP was performed as described in Fig.4. Panel A, Gln3-Myc13 binding to PGDH2 is shown. GLN3-MYC13 wild type(TB123), GLN3-MYC13 sit4� (TB136-2a), and GLN3-MYC13 pph21�pph22�(FV221) cells were subjected to ChIP analysis. Panel B, Gat1-Myc13 binding toPGDH2 is shown. GAT1-MYC13 wild type (FV063), GAT1-MYC13 sit4� (FV066), andGAT1-MYC13 pph21�pph22� (03879c) cells were subjected to ChIP analysis.Panel C, Gln3-Myc13 binding to PDAL5 is shown. GLN3-MYC13 wild type (TB123),GLN3-MYC13 sit4� (TB136-2a), and GLN3-MYC13 pph21�pph22� (FV221) cellswere subjected to ChIP analysis. Panel D, Gat1-Myc13 binding to PDAL5 isshown. GAT1-MYC13 wild type (FV063), GAT1-MYC13 sit4� (FV066), and GAT1-MYC13 pph21�pph22� (03879c) cells were subjected to ChIP analysis.
Yeast GATA Factor Regulation by Rapamycin and Msx
DECEMBER 30, 2011 • VOLUME 286 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 44909
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

detailed information concerning the specific GATA factorrequirements of that transcription under the growth conditionsbeing used (54). If this detailed information is ignored, the con-clusions obtained can easily be dictated by the gene assayed orgrowth conditions employed thus making broad generaliza-tions risky.GATA factor binding was perhaps the most difficult to
straightforwardly correlate because not only did it displayinhibitor and gene specificity, but one also sees the influencethat binding of one GATA factor has on the other. The Gat1requirement for Gln3 binding was gene- but not inhibitor-spe-cific, whereas the Gln3 requirement for Gat1 binding wasinhibitor- but not gene-specific.Keeping in mind that overall GDH2 transcription relies on
Gln3, that DAL80 is Gat1-dependent, and that DAL5 expres-sion requires bothGln3 andGat1, we can draw reasonable con-nections between the inhibitor added to the culture and thefinal outcome, gene transcription. For example, Msx treatmentleads to nuclear Gln3 localization, which correlates well withincreased Gln3 binding to the GDH2 promoter and transcrip-tion. In contrast, Gat1 is largely if not completely immune toMsx treatment, resulting in an absence of Gat1-dependentGDH2 expression. Rapamycin treatment, on the other hand,elicits low nuclearGln3 localization correlatingwith lowGDH2binding and transcription. Rapamycin treatment also results inhigh nuclear Gat1 localization correlating with elevated Gat1binding to theDAL5 promoter and with high, Gat1-dependentDAL5 and DAL80 transcription. Inhibitor-induced DAL5 acti-vation requires both Gat1 and Gln3. Lower Gln3 binding suf-fices to support some transcriptional activation in the absenceof Gat1. In contrast, even elevated Gat1 binding is not able toactivate DAL5 to wild type levels in the absence of Gln3 (rapa-mycin-treated gln3� cells), suggesting that, in ammonia-growncells, Gln3 is more “competent” than Gat1 to support DAL5transcriptional activation when the GATA factors are assayedseparately.Although compelling positive correlations exist among the
three different levels of inhibitor responses (localization, bind-ing, expression), there are also negative correlations, especiallywith respect to GATA factor binding, some of which can beexplained and others that at present cannot. Such negative cor-relations emphasize the limits thatmust be placed on our inter-pretations, conceding that much still remains to be learnedabout these regulatory pathways. For example, Gln3 binding tothe DAL5 promoter in response to Msx and rapamycin treat-ment does not correlate with Gln3 localization but may beexplained by the Gat1 requirement for Gln3 binding irrespec-tive of the inhibitor used. Among the negative correlations thatcannot be explained are the following three examples. (i) Rapa-mycin failed to elicit GDH2 transcription despite the fact thatrapamycin elicited nuclear localization of both GATA factorsas well as binding of the GATA factors in wild type, gln3�, andgat1� strains. At face value, this observation raises the specu-lative possibility that a rapamycin-inhibited event plays a nec-essary intranuclear rolewith respect toGDH2 transcription. (ii)Msx-elicited GDH2 transcription did not decrease in a gat1�mutant despite reduced Gln3-Myc13 binding to the promoter.(iii) Equivalent levels of Msx- and rapamycin-elicited DAL5
transcription occur despite much lower binding of both GATAfactors in theMsx-treated cells. These yet unexplained negativecorrelations suggest that more than just GATA factor localiza-tion and binding account for the regulated transcriptionaloutputs.DifferingMsx- andRapamycin-elicitedResponses in the Phos-
phatase Mutants—Comparing the phosphatase requirementsfor the Msx and rapamycin responses provided an importantinsight into the question ofwhether one or two regulatory path-ways are operative. If Msx and rapamycin inhibit sequentialsteps in a linear pathway, then the phosphatase requirementsfor the most downstream inhibitor, rapamycin, should beobserved for both Msx and rapamycin. Msx-elicited responsesmight be expected to exhibit additional requirements notrequired for a rapamycin response, but the requirements for arapamycin response should always be present whether cells aretreatedwith rapamycin orMsx. The reasoning here is similar tothat of the epistasis relationships expected of two loss of func-tion mutations situated sequentially in a linear cascade. Onlythe Gln3 response can be evaluated in this manner becauseGat1 localization does not respond sufficiently toMsx addition.Msx-elicited Gln3 localization is nearly Sit4-independent,whereas the rapamycin-elicited response is absolutely Sit4-de-pendent. In other words, the rapamycin and Msx responsesexhibit opposite Sit4 requirements. Although more complexschemes can be envisioned, the Sit4 independence ofMsx-elic-ited nuclear Gln3 localization is the expected result if Msx andrapamycin are inhibiting steps in two different regulatory path-ways or a branched pathway. They are not consistent with theinhibitors acting on sequential steps of a single linear cascade.Taken together, the data presented in this work also demon-strate theGATA factor and inhibitor specificity of the phospha-tase requirements.At transcription, the phosphatase requirements were more
uniform, PP2A (Pph21/22) being absolutely required irrespec-tive of the gene and the inhibitor added, whereas the Sit4requirement was only partial. The uniformity mainly reflectsthe intranuclear influence of one GATA factor effects on thebinding of the other as noted below.At promoter binding, the phosphatase requirements became
gene- and inhibitor-specific, reflecting the interdependence ofone GATA factor binding on the other. Msx- and rapamycin-induced Gln3 binding to the GDH2 promoter was partiallyPP2A- and Sit4-dependent, whereas binding to the DAL5 pro-moter exhibited absolute requirements of both phosphatases.Rapamycin- and Msx-elicited Gat1 binding to the GDH2 andDAL5 promoters exhibited a much greater PP2A than Sit4requirement.There were both positive and negative correlations with
respect to phosphatase requirements. In rapamycin-treatedammonia grown sit4� cells, lowGln3 binding to theDAL5 pro-moter correlated with minimal nuclear Gln3 localization. Onthe other hand, rapamycin elicited substantial nuclear Gat1localization in the sit4�, which again correlated with binding toDAL5 and GDH2. Furthermore, rapamycin-induced DAL5transcription was reduced in the sit4�, correlating with wildtypeGat1 binding but lowGln3 binding. In a rapamycin-treatedpph21�pph22�, nuclear Gln3 and Gat1 localizations are
Yeast GATA Factor Regulation by Rapamycin and Msx
44910 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 52 • DECEMBER 30, 2011
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

reduced, positively correlating with reduced binding at GDH2and DAL5 and abolished transcription in DAL5.Unexplained negative correlations, however, suggested that
phosphatases could be responsible for regulated GATA factorfunctions beyond localization and DNA binding. For example,there is no Msx-elicited GDH2 transcription in apph21�pph22� despite nearly wild type levels of Gln3 bindingto the GDH2 promoter.Kinetics of GATA Factor Movement Are Inhibitor-specific
—The responses of GATA factor localizations to the inhibitorsfollowed quite different time courses. InMsx-treated wild type,pph21�pph22�, and sit4� cells, Gln3 relocated to the nucleusand remained there, although the onset of relocation was mod-estly and more dramatically delayed in the pph21�pph22� andsit4� strains, respectively. In the case of rapamycin, transientresponses were the norm. For Gln3, a transient response wasonly observed in the pph21�pph22� mutant, whereas for Gat1all of the responses were transient, those in the mutants beingmore marked than in the wild type.One other observation merits noting. This is the first time
that Myc13 and GFP-tagged GATA factor localization could becompared in phosphatasemutants.With one significant excep-tion, the results obtained with Myc13- and GFP-tagged GATAfactorswere consistentwith one anotherwhen considering thatit was possible to employ three category scoring of Gln3-Myc13but only two category scoring of Gln3-GFP (comparisonrequires combining nuclear cytoplasmic and nuclear values ofthe Myc13 results). The exception occurred with the responsesof Gat1-Myc13 and Gat1-GFP localizations in untreated,ammonia-grown sit4� mutant cells and in Msx-treatedpph21�pph22� cells (compare Fig. 3,C andDwith Fig. 6,G andH, and Fig. 7, G and H). Using the Myc13 tag, Gat1 is nuclear-cytoplasmic in about 15–20% of the cells, whereas with theGFPtag it is absolutely cytoplasmic. Although the two assays differboth in the tag and the use of formalin fixation when theMyc13tag is used (51), we cannot explain with confidence the differingresults. It is, however, important to keep in mind that suchdifferences can occur.We endwherewe began, with the overall goal of thework, i.e.
to decide whether rapamycin and Msx inhibited two steps in alinear cascade linking nitrogen availability with GATA factorresponses or the inhibitors participated in separate regulatorypathways. The preponderance of data supports the conclusionthat Msx and rapamycin are not inhibiting tandem steps of alinear pathway. The differences in the inhibitor responses andrequirements at all levels are just too great. On the other hand,one cannot rigorously conclude that the pathways are totallyseparate unless we ignore the negative correlations.If, as the data suggest, there are two regulatory pathways, it
prompts the question of whether or not they are redundant.Gene expression data in Fig. 1 argue that the twoGATA factorsand twoputative regulatory pathways are notmerely redundantas clear differential effects of both the inhibitors and GATAfactors (gln3� and gat1�) are observed. The question that pres-ent data will not answer is whether both regulatory systems arerequired for growth on a poorly utilized nitrogen source, e.g.proline, or a single functional pathway would suffice. Althougha comprehensive explanation of GATA factor regulation will
undoubtedly require integrating the responses to individualinputs and outputs of both pathways, specific GATA factormutations eliminating responses to one or the other of the reg-ulatory pathways, should effectively help to solve that issue.Furthermore, additional factors and regulatory pathwayrequirements involved in localization, DNA binding, and tran-scription will almost certainly emerge from such studies. It istoward this end that future work will be directed.
Acknowledgments—We thank Fabienne Vierendeels for excellenttechnical assistance, Tim Higgins and André Feller for preparing theartwork, and Rajendra Rai for suggestions to improve themanuscript.
REFERENCES1. Cooper, T. G. (1982) in Molecular Biology of the Yeast Saccharomyces:
Metabolism and Gene Expression (Strathern, J. N., Jones, E. W., andBroach, J. R., eds) pp. 39–99, Cold Spring Harbor Laboratory, Cold SpringHarbor, New York
2. Hofman-Bang, J. (1999)Mol. Biotechnol. 12, 35–733. Magasanik, B., and Kaiser, C. A. (2002) Gene 290, 1–184. Cooper, T. G. (2004) In Topics in Current Genetics (Winderickx, J., and
Taylor, P. M., eds) Vol. 7, Chapter 9, pp. 225–257, Springer-Verlag, Berlin5. Zaman, S., Lippman, S. I., Zhao, X., and Broach, J. R. (2008) Ann. Rev.
Genet. 42, 27–816. Rohde, J. R., and Cardenas, M. E. (2004) Curr. Top. Microbiol. Immunol.
279, 53–727. Bertram, P. G., Choi, J. H., Carvalho, J., Ai, W., Zeng, C., Chan, T. F., and
Zheng, X. F. (2000) J. Biol. Chem. 275, 35727–357338. Beck, T., and Hall, M. N. (1999) Nature 402, 689–6929. Cardenas,M. E., Cutler, N. S., Lorenz,M. C., Di Como, C. J., andHeitman,
J. (1999) Genes Dev. 13, 3271–327910. Hardwick, J. S., Kuruvilla, F. G., Tong, J. K., Shamji, A. F., and Schreiber,
S. L. (1999) Proc. Natl. Acad. Sci. U.S.A. 96, 14866–1487011. Wiame, J.M., Grenson,M., andArst, H.N., Jr. (1985)Adv.Microb. Physiol.
26, 1–8812. Grenson, M., and Hou, C. (1972) Biochem. Biophys. Res. Commun. 48,
749–75613. Dubois, E., Grenson, M., and Wiame, J. M. (1974) Eur. J. Biochem. 48,
603–61614. Dubois, E. L., and Grenson, M. (1974) Biochem. Biophys. Res. Commun.
60, 150–15715. Mitchell, A. P, and Magasanik, B. (1984)Mol. Cell. Biol. 4, 2758–276616. Drillien, R., and Lacroute, F. (1972) J. Bacteriol. 109, 203–20817. Drillien, R., Aigle,M., and Lacroute, F. (1973)Biochem. Biophys. Res. Com-
mun. 53, 367–37218. GrensonM, Dubois E, PiotrowskaM, Drillien R, and AigleM. (1974)Mol.
Gen. Genet. 128, 73–8519. Courchesne, W. E., and Magasanik, B. (1988) J. Bacteriol. 170, 708–71320. Blinder, D., Coschigano, P. W., andMagasanik, B. (1996) J. Bacteriol. 178,
4734–473621. Kulkarni, A. A., Abul-Hamd, A. T., Rai, R., El Berry, H., and Cooper, T. G.
(2001) J. Biol. Chem. 276, 32136–3214422. Carvalho, J., and Zheng, X. F. (2003) J. Biol. Chem. 278, 16878–1688623. Cunningham, T. S., Andhare, R., and Cooper, T. G. (2000) J. Biol. Chem.
275, 14408–1441424. Shamji, A. F., Kuruvilla, F. G., and Schreiber, S. L. (2000) Curr. Biol. 10,
1574–158125. Crespo, J. L., Powers, T., Fowler, B., and Hall, M. N. (2002) Proc. Natl.
Acad. Sci. U.S.A. 99, 6784–678926. Di Como, C. J., and Arndt, K. T. (1996) Genes Dev. 10, 1904–191627. Jiang, Y., and Broach, J. R. (1999) EMBO J. 18, 2782–279228. Wang, H., Wang, X., and Jiang, Y. (2003)Mol. Biol. Cell 14, 4342–435129. Yan, G., Shen, X., and Jiang, Y. (2006) EMBO J. 25, 3546–355530. Düvel, K., Santhanam, A., Garrett, S., Schneper, L., and Broach, J. R. (2003)
Mol. Cell 11, 1467–1478
Yeast GATA Factor Regulation by Rapamycin and Msx
DECEMBER 30, 2011 • VOLUME 286 • NUMBER 52 JOURNAL OF BIOLOGICAL CHEMISTRY 44911
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

31. Cox, K.H., Rai, R., Distler,M., Daugherty, J. R., Coffman, J. A., andCooper,T. G. (2000) J. Biol. Chem. 275, 17611–17618
32. Georis, I., Tate, J. J., Cooper, T.G., andDubois, E. (2008) J. Biol. Chem.283,8919–8929
33. Carvalho, J., Bertram, P. G., Wente, S. R., and Zheng, X. F. (2001) J. Biol.Chem. 276, 25359–25365
34. Cooper, T. G. (2002) FEMS Microbiol. Rev. 26, 223–23835. Tate, J. J., Cox, K. H., Rai, R., and Cooper, T. G. (2002) J. Biol. Chem. 277,
20477–2048236. Cox, K. H., Tate, J. J., and Cooper, T. G. (2002) J. Biol. Chem. 277,
37559–3756637. Tate, J. J., and Cooper, T. G. (2003) J. Biol. Chem. 278, 36924–3693338. Cox, K. H., Kulkarni, A., Tate, J. J., and Cooper, T. G. (2004) J. Biol. Chem.
279, 10270–1027839. Cox, K. H., Tate, J. J., and Cooper, T. G. (2004) J. Biol. Chem. 279,
19294–1930140. Tate, J. J., Rai, R., and Cooper, T. G. (2005) J. Biol. Chem. 280,
27195–2720141. Tate, J. J., Feller, A., Dubois, E., andCooper, T.G. (2006) J. Biol. Chem. 281,
37980–3799242. Kulkarni, A., Buford, T. D., Rai, R., and Cooper, T. G. (2006) FEMS Yeast
Res. 6, 218–22943. Tate, J. J., Georis, I., Feller, A., Dubois, E., and Cooper, TG. (2009) J. Biol.
Chem. 284, 2522–253444. Tate, J. J., Georis, I., Dubois, E., andCooper, T.G. (2010) J. Biol. Chem.285,
17880–1789545. Georis, I., Tate, J. J., Feller, A., Cooper, T. G., and Dubois, E. (2011) Mol.
Cell. Biol. 31, 92–10446. Wach, A. (1996) Yeast 12, 259–26547. Liu, Z., Sekito, T., Spírek, M., Thornton, J., and Butow, R. A. (2003) Mol.
Cell 12, 401–41148. Giannattasio, S., Liu, Z., Thornton, J., and Butow, R. A. (2005) J. Biol.
Chem. 280, 42528–4253549. Georis, I., Feller, A., Vierendeels, F., and Dubois, E. (2009)Mol. Cell. Biol.
29, 3803–381550. Tate, J. J., and Cooper, T. G. (2007) J. Biol. Chem. 282, 18467–1848051. Tate, J. J., and Cooper, T. G. (2008) FEMS Yeast Res. 8, 1223–123552. Roon, R. J., and Even, H. L. (1973) J. Bacteriol. 116, 367–37253. Daugherty, J. R., Rai, R., el Berry, H. M., and Cooper, T. G. (1993) J. Bac-
teriol. 175, 64–7354. Georis, I., Feller, A., Tate, J. J., Cooper, T. G., and Dubois, E. (2009) Ge-
netics 181, 861–87455. Scherens, B., Feller, A, Vierendeels, F., Messenguy, F., and Dubois, E.
(2006) FEMS Yeast Res. 6, 777–79156. Coffman, J. A., Rai, R., and Cooper, T. G. (1995) J. Bacteriol. 177,
6910–691857. Coffman, J. A., Rai, R., Cunningham, T., Svetlov, V., and Cooper, T. G.
(1996)Mol. Cell. Biol. 16, 847–85858. Stanbrough,M., Rowen, D.W., andMagasanik, B. (1995) Proc. Natl. Acad.
Sci. U.S.A. 92, 9450–945459. Chisholm, G., and Cooper, T. G. (1982)Mol. Cell. Biol. 2, 1088–109560. Cunningham, T. S., and Cooper, T. G. (1991) Mol. Cell. Biol. 11,
6205–621561. Coornaert, D., Vissers, S., André, B., and Grenson, M. (1992) Curr. Genet.
21, 301–30762. Coffman, J. A., Rai, R., Loprete, D. M., Cunningham, T., Svetlov, V., and
Cooper, T. G. (1997) J. Bacteriol. 179, 3416–342963. Turoscy, V., and Cooper, T. G. (1979) J. Bacteriol. 140, 971–97964. Rai, R., Genbauffe, F., Lea, H. Z., and Cooper, T. G. (1987) J. Bacteriol. 169,
3521–3524
Yeast GATA Factor Regulation by Rapamycin and Msx
44912 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 286 • NUMBER 52 • DECEMBER 30, 2011
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Isabelle Georis, Jennifer J. Tate, Terrance G. Cooper and Evelyne DuboisOther by Methionine Sulfoximine
Gat1 Occurs by Two Distinct Pathways, One Inhibited by Rapamycin and the Nitrogen-responsive Regulation of GATA Protein Family Activators Gln3 and
doi: 10.1074/jbc.M111.290577 originally published online October 28, 20112011, 286:44897-44912.J. Biol. Chem.
10.1074/jbc.M111.290577Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/286/52/44897.full.html#ref-list-1
This article cites 63 references, 40 of which can be accessed free at
by guest on September 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from


















