
Neuropeptide and catecholamine effects on tenocytes in ...619908/FULLTEXT01.pdf · Neuropeptide and...
90
Neuropeptide and catecholamine effects on tenocytes in tendinosis development – studies on two model systems with focus on proliferation and apoptosis Ludvig J. Backman Umeå, 2013 Dept of Integrative Medical Biology, Anatomy, and Dept of Surgical and Perioperative Sciences, Sports Medicine Umeå University, Umeå, Sweden in collaboration with Dept of Physical Therapy The University of British Columbia, Vancouver, BC, Canada
Transcript of Neuropeptide and catecholamine effects on tenocytes in ...619908/FULLTEXT01.pdf · Neuropeptide and...

Neuropeptide and catecholamine effects
on tenocytes in tendinosis development
– studies on two model systems with focus on proliferation and apoptosis
Ludvig J. Backman
Umeå, 2013
Dept of Integrative Medical Biology, Anatomy, and
Dept of Surgical and Perioperative Sciences, Sports Medicine
Umeå University, Umeå, Sweden
in collaboration with
Dept of Physical Therapy
The University of British Columbia, Vancouver, BC, Canada

ii
© Ludvig Backman
Responsible publisher under Swedish law: The Dean of the Faculty of Medicine
This work is protected by Swedish Copyright Legislation (Act 1960:729).
The original articles were reproduced with permission from the publishers.
ISBN for printed version: 978-91-7459-633-5
ISBN for e-publication version: 978-91-7459-634-2
ISSN: 0346-6612
New Series No: 1572
Electronic version available at http://umu.diva-portal.org/
Printed by: Print and Media, Umeå University
Umeå, Sweden, May 2013
Figures 1-9, 11, 14-21, and 23-24 are illustrated by Gustav Andersson.
Copyright with artist.
Figures 1-2 are reprinted from “Influences of paratendinous innervation and non-
neuronal substance P in tendinopathy – studies on human tendon tissue and an
experimental model of Achilles tendinopathy” by Gustav Andersson, Umeå, 2010.
Figure 2 is a drawing partly based on Fig. 2.6 in Jozsa, 1997.

iii

Ludvig J. Backman, 2013
iv
TABLE OF CONTENTS
TABLE OF CONTENTS ................................................................................. iv ABSTRACT .................................................................................................... vi ABBREVATIONS ......................................................................................... viii LIST OF ORIGINAL PAPERS ....................................................................... ix 1. BACKGROUND .................................................................. 1
1.1 The human Achilles tendon .......................................................... 1 1.1.1 Anatomy .............................................................................................. 1 1.1.2 Tendon structure ................................................................................ 2 1.1.3 Tendon extracellular matrix .............................................................. 3 1.1.4 Tendon cells ........................................................................................ 4 1.1.5 Biomechanics ..................................................................................... 4
1.2 Chronic tendon pathology ............................................................ 5 1.2.1 Definition of tendinopathy and tendinosis ....................................... 5 1.2.2 Characteristics of tendinosis .............................................................. 6 1.2.3 Theories of aetiology .......................................................................... 6 1.2.4 Achilles tendinopathy – epidemiology, symptoms,
and treatments ................................................................................... 6 1.3 Tendinosis models ........................................................................ 8
1.3.1 In vivo models .................................................................................... 8 1.3.2 In vitro models ................................................................................... 9
1.4 Neuropeptides and neurotransmitters of relevance for tendons ................................................................... 9
1.4.1 Neuropeptides .................................................................................... 9 1.4.2 Neurotransmitters ............................................................................. 12
1.5 Non-neuronal production of signal substances by tenocytes .....14 1.5.1 Introduction ...................................................................................... 14 1.5.2 SP and NK-1 R ................................................................................... 15 1.5.3 Catecholamines and α2-adrenoreceptors ......................................... 16
1.6 Mechanisms of basic cell functions .............................................16 1.6.1 Cell cycle and proliferation ............................................................... 16 1.6.2 Apoptosis .......................................................................................... 18
2. HYPOTHESES AND AIMS ............................................... 22 2.1 Hypotheses ................................................................................. 22 2.2 Aims ............................................................................................ 22
3. MATERIAL AND METHODS ........ FEL! BOKMÄRKET ÄR INTE DEFINIERAT.
3.1 Animal model of Achilles tendinosis .......................................... 23 3.1.1 Rabbit material ................................................................................. 23 3.1.2 Ethical considerations ...................................................................... 24 3.1.3 Experimental design ........................................................................ 24 3.1.4 Animal care and anaesthesia ........................................................... 26 3.1.5 Rabbit biopsies ................................................................................. 26
3.2 Primary human tendon cell culture model ................................ 27 3.2.1 Human material ............................................................................... 27 3.2.2 Ethical considerations ...................................................................... 28 3.2.3 Isolation of human Achilles tendon derived
tenocytes and cell culturing ............................................................. 28 3.2.4 Experimental set-up ......................................................................... 28
3.3 Methods of analysis ..................................................................... 31

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
v
3.3.1 General morphology ......................................................................... 31 3.3.2 Immunostaining ................................................................................ 31 3.3.3 In situ hybridization ......................................................................... 34 3.3.4 EIA .................................................................................................... 36 3.3.5 Vascularity assessment .................................................................... 38 3.3.6 Inflammation assessment ................................................................ 38 3.3.7 Analysis of cell number, viability, proliferation
and metabolic activity ...................................................................... 38 3.3.8 Analysis of cell death and apoptosis ................................................ 40 3.3.9 Western blot ...................................................................................... 41 3.3.10 Real-time quantitative polymerase chain reaction (RT-qPCR) ..... 44 3.3.11 Cell density used for different experiments in vitro ....................... 45
3.4 Statistics ...................................................................................... 46 3.4.1 In vivo studies .................................................................................. 46 3.4.2 In vitro studies ................................................................................. 47
4. RESULTS......................................................................... 48 4.1 Phenotyping of cells studied in vitro (tenocytes) ...................... 48
4.1.1 Complementary results: Additional phenotyping of cells .............. 49 4.2 Presence of signal substances and their receptors .................... 49
4.2.1 In vivo ............................................................................................... 49 4.2.2 In vitro .............................................................................................. 50
4.3 Changes in SP and NK-1 R expression in response to load ........ 51 4.3.1 In vivo ................................................................................................ 51 4.3.2 In vitro .............................................................................................. 52
4.4 Effects of exogenously added clonidine ..................................... 52 4.4.1 In vivo ............................................................................................... 52 4.4.2 In vitro .............................................................................................. 53
4.5 Effects of exogenously added SP ................................................ 53 4.5.1 In vivo ............................................................................................... 53 4.5.2 In vitro .............................................................................................. 54
5. DISCUSSION ................................................................... 57 5.1 Introductory remarks ................................................................. 57 5.2 Methodological aspects of the experimental models ................. 57
5.2.1 Animal model of Achilles tendinosis ............................................... 57 5.2.2 Primary human tendon cell culture model ..................................... 60
5.3 The possible importance of SP and catecholamines in tendons and tendinosis .......................................................... 62
5.3.1 Aspects of the biochemical theory in the onset/development of tendinosis .................................................... 62
5.3.2 Aspects of the biochemical theory in the chronic stages of tendinosis ............................................................. 64
5.4 Summarizing reflections ............................................................ 65 5.4.1 The role of SP/catecholamines and the
hypercellularity in tendinosis .......................................................... 65 5.4.2 The role of SP and apoptosis in tendinosis ..................................... 66 5.4.3 Possible clinical implications ........................................................... 66
6. CONCLUSIONS ............................................................... 67 ACKNOWLEDGEMENTS ............................................................................ 68 FUNDING ..................................................................................................... 70 POPULÄRVETENSKAPLIG SAMMANFATTNING PÅ SVENSKA .............. 71 REFERENCES .............................................................................................. 72

Ludvig J. Backman, 2013
vi
ABSTRACT
Background: Achilles tendinopathy is a common clinical syndrome of chronic Achilles tendon pain combined with thickening of the tendon and impaired tendon function. Tendinopathy is often, but not always, induced by mechanical overload, and is frequently accompanied by abnormalities at the tissue level, such as hypercellularity and angiogenesis, in which case the condition is called tendinosis. In tendinosis, there are no signs of intratendinous inflammation, but occasionally increased apoptosis is observed. Tendinosis is often hard to treat and its pathogenesis is still not clear. Recently, a new hypothesis has gained support, suggesting a biochemical model based on the presence of a non-neuronal production of classically neuronal signal substances by the primary tendon cells (tenocytes) in tendinosis. The possible functional importance of these signal substances in tendons is unknown and needs to be studied. In particular, the neuropeptide substance P (SP) and catecholamines are of interest in this regard, since these substances have been found to be up-regulated in tendinosis. As both SP and catecholamines are known to exert effects in other tissues resulting in changes similar to those characteristic of tendinosis, it is possible that they have a role in tendinosis development. It is furthermore unknown what elicits the increased intratendinous neuropeptide production in tendinosis, but given that tendon overload is a prominent risk factor, it is possible that mechanical stimuli are involved.
The hypothesis of this thesis work was that intratendinous production of SP is up-regulated in response to load of Achilles tendons/tenocytes, and that stimulation of the preferred SP receptor, the neurokinin-1 receptor (NK-1 R), as well as stimulation of the catecholamine α2 adrenoreceptors, contribute to the hypercellularity seen in tendinosis, via increased proliferation and/or decreased apoptosis, and that SP stimulates tendon angiogenesis. The purpose of the studies was to test this hypothesis. To achieve this, two model systems were used: One in vivo (rabbit Achilles tendon overload model of tendinosis) and one in vitro (human primary Achilles tendon cell culture model). Results: In the rabbit Achilles tendon tissue, SP and NK-1 R expression was extensive in the blood vessel walls, but also to some extent seen in the tenocytes. Quantification of endogenously produced SP in vivo confirmed intratendinous production of the peptide. The production of SP by human tendon cells in vitro was furthermore demonstrated. The catecholamine synthesizing enzyme tyrosine hydroxylase (TH), as well as the α2A adrenoreceptor (α2A AR), were detected in the tenocytes, both in vivo in the rabbit tissue and in vitro in the human tendon cells. As a response to mechanical loading in the in vivo model, the intratendinous levels of SP increased, and this elevation was found to precede distinct tendinosis changes. The in vitro model demonstrated the same response to load, i.e. an increased SP expression, but in this case also a decrease in the NK-1 R expression. In the in vivo model, exogenously administered SP, as well as clonidine (an α2 AR agonist), accelerated tenocyte hypercellularity, an effect that was not seen when administrating a specific α2A AR antagonist. Exogenous administration of SP also resulted in intratendinous angiogenesis

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
vii
and paratendinous inflammation. In the in vitro model, both SP and clonidine had proliferative effects on the human tenocytes, specifically mediated via NK-1 R and α2A AR, respectively; both of which in turn involved activation/phosphorylation of the extracellular signal-regulated kinases 1 and 2 (ERK1/2). Exogenously administered SP, in Anti-Fas induced apoptosis of the tenocytes in vitro, confirmed SP to have an anti-apoptotic effect on these cells. This effect was specifically mediated via NK-1 R and the known anti-apoptotic Akt pathway. Conclusions: In summary, this thesis concludes that stimulation of NK-1 R and α2A AR on tenocytes, both in vitro and in vivo, mediates significant cell signalling effects leading to processes known to occur in tendinosis, including hypercellularity. The pathological role of the hypercellularity in tendinosis is still unclear, but it is likely to affect collagen metabolism/turnover and arrangement, and thereby indirectly tendon biomechanical function. Additional evidence is here provided showing that SP not only causes tenocyte proliferation, but also contributes to anti-apoptotic events. Furthermore, it was concluded that SP may be involved in the development of tendinosis, since its production is increased in response to load, preceding tendinosis, and since SP accelerates tendinosis changes, through some mechanistic pathways here delineated. These findings suggest that inhibition of SP, and possibly also catecholamines, could be beneficial in the reconstitution/normalization of tendon structure in tendinosis.

Ludvig J. Backman, 2013
viii
ABBREVATIONS
AR Adrenoreceptor AP Alkaline phosphatase BSA Bovine serum albumin cDNA Complementary DNA DED Death effector domain DEPC Diethylpyrocarbonate DIG Digoxigenin DISC Death-inducing signalling complex ERK1/2 Extracellular signal-regulated kinases 1 and 2 EPI Epinephrine/adrenaline FADD Fas-associated protein with death domain FBS Foetal bovine serum GAG Glycosaminoglycans GPCR G-protein coupled receptors HRP Horseradish peroxidase IL-8 Interleukin-8 IQR Inter-quartile range kDa kilo Dalton L-DOPA L-dihydroxyphenylalanine LDH Lactate dehydrogenase LDV Large dense vesicle MAP Mitogen-activated protein MOA Monoamine oxidase mRNA Messenger ribonucleic acid MTJ Musculotendinous junction NE Norepinephrine/noradrenaline NEP Neutral endopeptidase NF-κB Nuclear factor kappa-light-chain-enhancer of activated B cells NK-1 R Neurokinin-1 receptor NK-2 R Neurokinin-2 receptor NK-3 R Neurokinin-3 receptor OTJ Osteotendinous junction P-Akt Phophorylated Akt PARP Poly (ADP-ribose) polymerase PBS Phosphate buffered saline PFA Paraformaldehyde PI3K Phosphoinositide 3-OH kinase PKB Protein kinase B PPT-A Pre-protachykinin-A rcf Relative centrifugal force SSV Small synaptic vesicle SP Substance P TBS-T Tris buffered saline with tween 20 TH Tyrosine hydroxylase VGluT2 Vesicular glutamate transporter 2

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
ix
LIST OF ORIGINAL PAPERS
I. Endogenous substance P production in the Achilles tendon
increases with loading in an in vivo model of tendinopathy – peptidergic elevation preceding tendinosis-like tissue changes Backman, L.J., Andersson, G., Wennstig, G., Forsgren, S., and Danielson, P. Journal of Musculoskeletal and Neuronal Interactions, 2011;11(2):133-140.
II. Substance P accelerates hypercellularity and angiogenesis
in tendon tissue and enhances paratendinitis in response to Achilles tendon overuse in a tendinopathy model Andersson, G., Backman, L.J., Scott, A., Lorentzon, R., Forsgren, S., and Danielson, P. British Journal of Sports Medicine, 2011;45(13):1017-1022.
III. Substance P is a mechanoresponsive, autocrine regulator of
human tenocyte proliferation Backman, L.J., Fong, G., Andersson, G., Scott, A., and Danielson, P. PLoS One, 2011;6(11):e27209.
IV. Alpha-2 adrenergic stimulation triggers Achilles tenocyte
hypercellularity: Comparison between two model systems Backman, L.J., Andersson, G., Fong, G., Alfredson, H., Scott, A., and Danielson, P. Scandinavian Journal of Medicine and Science in Sports; Published online ahead of print January 31, 2012. doi: 10.1111/j.1600-0838.2011.01442.x
V. Akt-mediated anti-apoptotic effects of substance P in
Anti-Fas induced apoptosis of human tenocytes Backman, L.J., and Danielson, P. Journal of Cellular and Molecular Medicine; Published online ahead of print April 11, 2013. doi: 10.1111/jcmm.12059
In the thesis, the original papers will be referred to by their Roman numerals and the figures in the original papers will be referred to by the Roman numeral of the paper followed by the number of the figure in that paper (e.g., Fig. III:3a = Fig. 3a in paper III). The original papers were reprinted with permission of the respective publisher.


Neuropeptide and catecholamine effects on tenocytes in tendinosis development
1
1. BACKGROUND
1.1 The human Achilles tendon
1.1.1 Anatomy
The cross sectional area of the Achilles tendon is 0.8-1.4 cm2, making this the thickest and, based on its architecture, strongest tendon in the body (O'Brien, 1992).
The Achilles tendon connects the triceps surae muscle – which consists of the medial and lateral gastrocnemius muscles and the soleus muscle – to the calcaneal bone and this tendon-muscle complex spans over two joints. Proximally, the tendon originates from the gastrocnemius and soleus muscles at the musculotendinous junction (MTJ). The gastrocnemius portion of the proximal tendon begins as a broad aponeurosis and the soleus portion of the tendon begins as a band 8-15 cm distal to that of the gastrocnemius portion (Dalla Costa et al., 2010). As the tendon descends it can spiral up to 90° laterally, so that fibres that were initially located to the posterior become lateral, lateral fibres become anterior, anterior fibres become medial, and medial fibres become posterior, at the site of insertion (Jozsa, 1997). The degree of rotation of the tendon has a great variance depending on the amount of fibres originating from the gastrocnemius and soleus muscles, respectively, and also on the level at which fusion takes place (Cummins et al., 1946; Jozsa, 1997). The lateral rotation of the tendon as it descends, assists in supination of the ankle, but is also suggested to result in increased internal stress on the tendon in the region 2-6 cm above the calcaneal insertion (Morimoto and Ogata, 1968; Jozsa, 1997; Teitz et al., 1997).
The tendon becomes narrower and more rounded distally before attaching to the dorsal and superior surface of the tuber calcanei by a stiff fibrocartilagenous expansion; the osteotendinous junction (OTJ) (Jozsa, 1997). The OTJ progressively transitions from tendon to bone and consists of different zones; the tendon, fibrocartilage, mineralized fibrocartilage and bone (Astrom and Rausing, 1995; Jozsa, 1997). In the distal region of the tendon, two bursae are present, the retrocalcaneal and the subcutaneous, with the main function to decrease the friction between the tendon and surrounding tissue (Reinherz et al., 1991). See figure 1 for an overview of the Achilles tendon anatomy.
Fig. 1 Drawing illustrating the anatomy of the Achilles tendon and the triceps surae muscle.

Ludvig J. Backman, 2013
2
The fascia that envelopes the triceps surae muscle continuously surrounds
the Achilles tendon and merges distally with the crural fascia at about 4 cm from the calcaneal insertion. This tendon-surrounding tissue is called the paratenon (Williams, 1986; Jozsa, 1997; Carmont et al., 2011). The paratenon functions as an elastic sleeve that permits free movement of the tendon against surrounding tissues (Jozsa, 1997). The paratenon is described to consist of thin gliding membranes on the dorsal, lateral and medial sides whereas the ventral side is unclearly described in the literature. Some argue that the ventral aspect is a fatty areolar tissue with rich vascularisation, called paratendinous loose connective tissue (Kvist et al., 1987; Schepsis et al., 1994), and others imply that the paratenon is comprised of a membrane also ventrally, as visualized with ultrasound, and that it is not continuous with the tissue mass consisting of adipocytes ventral to the Achilles tendon called Kager’s fat pad (Pierre-Jerome et al., 2010).
1.1.2 Tendon structure
Tendons are dense connective tissues mainly composed of closely packed collagen fibres in parallel arrangements that enable the matrix to resist stretching forces. The dry weight of the tendon is about 30% of the total mass; the remaining 70% accounts for water. Of the dry weight 75-90% consists of various collagen molecules, 0.2-5% proteoglycans and 1-2% elastin (Jozsa, 1997). Type I collagen is the main component of tendon tissue, but there is also type III and type V collagen. The collagen, which contributes to the structural integrity and resists the tensile force applied to the tendon, is arranged in hierarchical levels of increasing complexity. The elastin is suggested to contribute to the recovery of the wavy configuration in the collagen fibres following tendinous stretch (Butler et al., 1978).
The smallest structure of the tendon is tropocollagen, a triple-helix polypeptide chain, which unites into fibrils, the basic tendon unit. Fibrils bond and create collagen fibres and the collagen fibres create fibre bundles (primary bundles), fascicles (secondary bundles), and finally tertiary bundles that form the tendon itself (Jozsa, 1997). A sheath of connective tissue, called endotenon, enfolds each collagen bundle and binds the bundles together. The tendon itself is covered by the epitenon. The vascular, lymphatic, and nerve supply of the tendon follows the endotenon into the tendon tissue proper. Superficially, the epitenon is surrounded by the paratenon (Jozsa, 1997; O'Brien, 1997; Kannus, 2000). It is also discussed that there is an outermost epithelium layer. Recent studies have shown that flexor digitorum profundus tendon in mouse, as well as mouse and rat tail, have an epithelium layer (Taylor et al., 2011). If this is the case also for the human Achilles tendon, it is not fully described. See figure 2 for an overview of the tendon hierarchy.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
3
Fig. 2 The structural hierarchy of a tendon.
1.1.3 Tendon extracellular matrix
Extracellular matrix consists of a variety of substances, of which collagen fibrils and proteoglycans are the most highly expressed (Curwin et al., 1988).
The ground substance that surrounds the collagen in the tendon consists of proteoglycans, glycosaminoglycans (GAG), structural glycoproteins and a wide variety of other small molecules. Proteoglycans – heavily glycosylated glycoproteins – are composed of a protein core to which one or more GAG is covalently attached. The collagen in the tendon is held together by these proteoglycans; a component that also includes both decorin and, in compressed regions of the tendon (such as in the OTJ), aggrecan. The macromolecules (proteoglycans and GAGs) that are entrapped between collagen fibrils and fibres are negatively charged hydrophilic molecules and have a high water binding capacity through osmosis as they attract cations. This enables buffering of water up to 50 times their weight, which improves the biomechanical properties of withstanding compressive forces. The proteoglycans also enable rapid diffusion of water-soluble molecules and migration of cells. (O'Brien, 1997)
Less than 0.2% of the tendon dry mass accounts for inorganic components, which are known to be intimately involved in growth, development, and normal metabolism. The anorganic component found in the highest concentration is calcium, which has a key role in the development of the OTJ. Other anorganic components present are magnesium, manganese, copper etc. (Jozsa, 1997)

Ludvig J. Backman, 2013
4
1.1.4 Tendon cells
Tendons consist of numerous cell types. The primary collagen-synthesizing tendon cells, tenoblasts and tenocytes, constitute about 90-95% of the cellular elements within the extracellular matrix, and both these cell types belong to a subpopulation of fibroblasts (Riley, 2008). Tenoblasts are considered to be immature tenocytes. They appear rounded and have a high metabolic activity. As they mature, they become more elongated and spindle-shaped, and transform/differentiate into tenocytes, which have lower metabolic activity. The tenocytes synthesize collagen and all components of the extracellular matrix seen in tendons (Jozsa et al., 1979; Kvist et al., 1987; O'Brien, 1997).
The remaining 5-10% of the cellular element consists of chondrocytes at the OTJ, synovial cells in the tendon sheath, and vascular cells including capillary endothelial cells and smooth muscle cells of arterioles (Jozsa, 1997).
1.1.4.1 Phenotyping tenocytes
There is not one marker that is truly specific to identify tenocytes. However, there are markers that are abundantly expressed in developing as well as in mature tendons. The transcription factor scleraxis and the type II transmembrane glycoprotein tenomodulin are both candidate markers for tenocytes. It is described that scleraxis positively regulates the expression of tenomodulin, and that tenomodulin is necessary for the proliferation and maturation of a tendon (Docheva et al., 2005; Shukunami et al., 2006).
As there is no perfect marker for tenocytes, markers for products of the tenocytes are also used to characterize these cells. Collagen, predominantly collagen type I, constitutes approximately 90% of the total protein in a tendon and considering the high abundance it is often used as an indirect marker of tenocytes (Jozsa, 1997). It is also known that scleraxis regulates the transcription of the gene for collagen type I, which reinforces the relevance of both collagen type I and scleraxis as markers for tenocytes (Lejard et al., 2007). Also decorin is commonly used, as it is a protein that binds to collagen type I and plays an important role in the matrix assembly (Yoon and Halper, 2005).
Additionally, cytoskeleton markers for tenocyte confirmation can be used (Bjur et al., 2008b). Vimentin, an intermediate filament protein, is expressed in mesenchymal derived cells such as osteoblasts, chondrocytes, adipocytes and also tenocytes (Rufai et al., 1992).
1.1.5 Biomechanics
The Achilles tendon transmits force from muscle to bone and acts as a buffer by absorbing external forces to limit muscle damage. The structure of the Achilles tendon, the lateral rotation and its internal architecture, make elongation and elastic recoil possible, and the stored energy can be released during locomotion (Alexander and Bennet-Clark, 1977). It is suggested that more than 30% of the required energy during jumping activities can be stored within the Achilles tendon itself (Fukashiro et al., 1995).

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
5
The collagen fibrils at rest have a wavy configuration and part of the collagen fibres of the epitenon are in an approximately 60º angulation with the fibres of the tendon. This configuration disappears when the tendon is stretched to 2% elongation of its length and the angle decreases from 60º to approximately 30º. Up to 4% elongation, the tendon is deformed in a linear fashion, and the fibres become more parallel. The configuration and original length will be resumed at relaxation. If strain exceeds approximately 4%, the tendon fibres illustrate microscopic failure, and the tendon will rupture when the strain reaches 8-10%. (Jozsa, 1997)
The in vivo tensile force has been registered in a number of activities with an implanted transducer in the tendon. Running and also jumping has registered forces up to approximately 9,000 N, corresponding to about 12 times the body weight (Komi et al., 1992; Kader et al., 2002). In results from both static and dynamic testing of the tendon it is shown that it is the dorsal (superficial) part of the tendon that is the most loaded part and that the ventral part is less loaded (Almekinders et al., 2002).
1.2 Chronic tendon pathology
1.2.1 Definition of tendinopathy and
tendinosis
The definitions of tendinopathy and tendinosis have been somewhat confusing in the past. Different terms have been used to describe the same condition, and the definitions can still differ between research groups.
The most accepted definition of tendinopathy, and the one used in this thesis, is that the term describes a clinical condition of chronic tendon pain (>3 months), swelling, and impaired function of the tendon (Khan et al., 1999). If subjects with tendon pain, swelling and impaired function also demonstrate structural tissue changes in the tendon, as shown by ultrasound (US), magnetic resonance imaging (MRI), or histopathological examination of biopsies, it is generally (and in this thesis) defined as tendinosis (Alfredson and Ohberg, 2005). However, it has been suggested that tendinosis should only be regarded as a histopathological diagnosis, rather than radiological, since neither US nor MRI can confirm all the characteristics of tendinosis (see chapter 1.2.2) (Maffulli et al., 1998). The term tendinosis, as a histological description of degenerative pathology was first described by Puddu and collaborators in 1976 (Puddu et al., 1976). It should be pointed out that some authors, who define tendinosis as a mere description of tendon tissue changes as seen by histological (and/or radiological) examination, thus do not consider clinical symptoms a requirement for the diagnosis (Maffulli et al., 1998). Hence, in this meaning, an individual can have tendinosis without tendinopathy, if the latter is defined as a clinical symptomatic syndrome (as in this thesis). Finally, tendinopathy is by some interpreted in the literal meaning of the word (‘tendon pathology’), which

Ludvig J. Backman, 2013
6
then would include all tendon pathology and not a specific clinical syndrome (Riley, 2004).
1.2.2 Characteristics of tendinosis
Tissue changes seen in tendinosis are hypercellularity, increased vascularity, degeneration and disordered arrangement of collagen fibres (Khan et al., 1999). In a study by Åström and co-workers it was reported that 90% of biopsies of symptomatic tendons illustrates the mentioned tissue changes (Astrom and Rausing, 1995).
The increased amount of cells is also combined with an increased number of cells with rounded nuclei and less spindle-shaped appearance (Khan et al., 1999). These cells have both a higher proliferation and apoptosis index, indicating a cell population of high cell turnover (Chuen et al., 2004).
There are no signs of classic inflammation in chronic tendinosis. Hence, there is a lack of inflammatory cells in the tendon tissue (Khan et al., 1999) and no elevated intratendinous prostaglandin levels (Alfredson et al., 1999).
1.2.3 Theories of aetiology
The aetiology of Achilles tendinopathy is still unclear. However, it is likely that intrinsic and extrinsic factors interact (Khan and Maffulli, 1998). Some factors of intrinsic characteristics are age, gender, body weight, height, tendon vascularity, and lateral ankle instability (Maffulli et al., 2004). Intrinsic factors also include the genetic variants and it has been shown that variations in, for example, the COL5A1 and GDF-5 genes are associated with Achilles tendinopathy (September et al., 2009; Posthumus et al., 2010; Abrahams et al., 2013). Examples of extrinsic factors are changes in training pattern, poor technique, previous injuries, footwear, and training surface (Kvist, 1991).
Despite the fact that Achilles tendinopathy occurs among sedentary individuals (Rolf and Movin, 1997), the main pathological stimulus for developing tendinopathy is regarded to be excessive loading of the tendon (Jozsa, 1997; Kader et al., 2002). There are also studies linking obesity to tendinopathy (Gaida et al., 2009), which partly might explain why also inactive individuals are prone to develop the condition.
1.2.4 Achilles tendinopathy – epidemiology,
symptoms, and treatments
In recreational runners Achilles tendinopathy makes out about 6-18% of all injuries (Clement et al., 1984; Jozsa, 1997). In elite runners the 1-year incidence has been reported to be 7-9% (Lysholm and Wiklander, 1987). There are data showing that in individuals initially having unilateral Achilles tendinopathy, 41% also develop symptoms on the contralateral side (Paavola et al., 2000). The most common age group that develops Achilles tendinopathy is the one of 30-60 years of age; it is rarely seen in individuals younger than 30 years (Kvist,

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
7
1991). Tendinopathy in the Achilles tendon can be divided into mid-portion and distal Achilles tendinopathy, and the distal accounts for approximately 20% of all Achilles tendinopathies, meaning that the remaining 80% is mid-portion Achilles tendinopathy (Astrom and Rausing, 1995). There is a tendency that those diagnosed with distal Achilles tendinopathy are, in general, older, non-active, and overweight (Myerson and McGarvey, 1999). Considering that it has been shown that 90% of patients diagnosed with Achilles tendinopathy also illustrate structural changes, verified with histopathological examinations (i.e. tendinosis), the epidemiology for tendinopathy and that of tendinosis are closely related (Astrom and Rausing, 1995).
Individuals who develop tendinopathy most commonly have a gradual onset of pain, not associated with any trauma accompanied with acute pain. Initially, the pain is easy to neglect as the feeling of slight discomfort, as well as the pain and stiffness, disappear during activity. In the later stages, morning stiffness is common and the activity related symptoms start to increase. It is also common that a visible tendon swelling is observed in the region from which the symptoms derive. (Cook et al., 2002; Kader et al., 2002)
The first breakthrough in the treatment methods for chronic mid-portion Achilles tendinopathy was when the non-invasive eccentric training protocol was introduced in the late 1990s (Alfredson et al., 1998), based on ideas from Stanish and Curwin (Stanish et al., 1986). The eccentric training protocol has been reported to have a success rate of approximately 80% (Mafi et al., 2001). The effect of the treatment has been repeated by different research groups, however with varying results regarding the success rate in different activity level groups (Magnussen et al., 2009; Matheson, 2010). Thus, recent studies report lesser values in success rate, e.g. 30% at 12 weeks and 63% at 24 weeks of patient satisfaction (de Vos et al., 2010). It has furthermore been shown that other training regimes, such as stretching, can also be as effective as eccentric training (Norregaard et al., 2007). In addition, there are other non-invasive treatment methods such as shockwave, ultrasound, deep friction, rest and laser therapy reporting various results, alone or in combination with eccentric training, in the treatment of chronic mid-portion Achilles tendinopathy (Magnussen et al., 2009; Matheson, 2010; Rowe et al., 2012; Sussmilch-Leitch et al., 2012). In a recent systematic review of different conservative treatments it was concluded that there is strong evidence for the effectiveness of eccentric training and shockwave therapy, moderate evidence for the effectiveness for low-level laser therapy, and limited evidence for orthoses (Rowe et al., 2012). Also invasive treatment options have been used, such as longitudinal tenotomy (Maffulli et al., 1997) as well as injections of Polidocanol (sclerosing agent) (Alfredson and Ohberg, 2005), corticosteroids (Andres and Murrell, 2008) or platelet-rich plasma (de Vos et al., 2010). However, the most recently developed invasive method is the mini-surgery, or needle scraping method, that has obtained, in the short-term follow-up, results of approximately 90% satisfaction and most patients returning to full tendon loading activities after only 4-6 weeks (Alfredson, 2011).
One important thing that has to be stressed in the management of tendinopathy patients, is that it is a complex diagnosis and that the treatment regime chosen must be adapted to each individual; it is “no longer a ’one size fits

Ludvig J. Backman, 2013
8
all‘ diagnosis”, as expressed by well-renowned tendon researcher Jill Cook (Cook, 2011).
1.3 Tendinosis models
1.3.1 In vivo models
1.3.1.1 Assessment criteria of tendinosis animal
models
The assessment criteria, based on histopathology, of tendinosis animal models used, differ slightly between research groups. To facilitate the assessment, Lui and collaborators recently recommended that the histopathology parameters hypercellularity, hypervascularity, loss of matrix organization or collagen fragmentation, and rounding of cell nuclei, should be used to define tendinosis in all animal models (Lui et al., 2011).
1.3.1.2 Existing tendinosis animal models in general
There are two categories of in vivo models to induce tendinosis changes. The first category, which is the most frequently reported, consists of the mechanical loading models. The second category involves injections of chemical substances.
Mechanical loading is, as mentioned, the most popular model and it also best mimics the repetitive loading seen in humans that is considered the main reason for tendinosis development. Three different mechanical loading models are reported; forced treadmill running, tendon loading via artificial muscle stimulation, and finally direct repetitive tendon stretching via an external loading device (Lui et al., 2011). These three models can be subdivided into active participation models, which include the treadmill running, and passive participation models, which are the artificial muscle stimulation and stretching via the external loading device (Lake et al., 2008; Lui et al., 2011).
The second category of models is intratendinous injections of chemical substances. This is an alternative to inducing changes characteristic of tendinosis by the use of mechanical loading. However, injections cannot mimic the entire process. It only allows research into the later stages of pathology and not the preceding factors. Chemical substances used in animal models include collagenase, cytokines, PGE1, PGE2, and fluoroquinolone (Lake et al., 2008; Lui et al., 2011).
1.3.1.3 The experimental rabbit model used in this thesis work
The experimental rabbit model used in this thesis work is a passive participation mechanical loading model (see section 3.1.3.1) (Backman et al., 1990). This model, which uses New Zealand white rabbits, fulfils the assessment criteria

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
9
(hypercellularity, hypervascularity, rounding of cell nuclei) of tendinosis after 3 weeks of exercise according to the training protocol (Andersson et al., 2011).
1.3.2 In vitro models
In vitro models are used to study cellular responses hypothesised to be involved in the pathogenesis of tendinosis. Like the in vivo models, the in vitro models can be divided into two categories: mechanical models and models based on the introduction of exogenous substances.
Mechanical loading can be performed either in 2D or 3D cultures. In the 2D models, cells are basically cultured on an elastic membrane, most commonly a membrane that is pre-treated with collagen I, and stretched either uniaxially, i.e. stretched along one axis, or equibiaxial, i.e. stretched in multiple axes (Wall et al., 2007). In the 3D models, cells are embedded in a matrix gel, most commonly collagen I, and stretched uniaxially (Scott et al., 2011).
Models based on introduction of exogenous substances can also be used to study cellular responses (Qiu et al., 2012). By exogenously adding substances, it is possible to isolate the impact on the cells, by a certain biological agent, via using only one substance at the time.
1.4 Neuropeptides and
neurotransmitters of relevance
for tendons
1.4.1 Neuropeptides
Neuropeptides are small peptides and as the name implies, they help neurons communicate with each other. The process of forming neuropeptides includes pro-peptides in the endoplasmatic reticulum, which need to be spliced by enzymes in order to become the active neuropeptides. The splicing of the pro-peptides occurs during axonal transport to the large dense vesicles (LDVs) (Hokfelt et al., 2000; Hokfelt et al., 2001). LDVs, in which the active neuropeptides are stored, are sensitive to variations in local intracellular calcium, ten times more sensitive than small synaptic vesicles (SSVs), and a change of the intracellular calcium above a threshold results in a release mechanism, which in general is gradual (De Camilli, 1991). The released neuropeptides then enter the synaptic cleft and modulate or mediate communication by acting on cell surface receptors, which are often G-protein coupled receptors (GPCRs). The duration of neurotransmission by neuropeptides is counted in minutes, which is due to the time needed to remove the neuropeptide. For instance, the neuropeptide substance P (SP), is inactivated by neutral endopeptidase (NEP), angiotensin-converting-enzyme (ACE), or aminopeptidases, but sometimes a second enzymatic degradation step is also required to fully inactivate SP (Michael-Titus et al., 2002).

Ludvig J. Backman, 2013
10
In humans, around 70 genes coding for neuropeptides have been shown to give rise to approximately a 100 neuropeptides. All neuropeptides bind with preference to specific receptors, but there can also be cross-reactivity with receptors of similar characteristics (Li and Kim, 2008).
1.4.1.1 Substance P (SP)
Substance P (SP) belongs to the tachykinin neuropeptide family, which also includes, for instance, neurokinin A, neuropeptide K, neuropeptide gamma, and neurokinin B. All these tachykinins have similar primary structure and biological functions (Severini et al., 2002). SP consists of 11 amino acids and has a molecular weight of 1,348 g/mol (Harrison and Geppetti, 2001). It is encoded by the pre-protachykinin-A (PPT-A) gene, also called TAC1. When the PPT-A transcript is spliced it gives four distinct isoforms of messenger ribonucleic acid (mRNA): α-PPT-A, β-PPT-A, γ-PPT-A and δ-PPT-A (Harrison and Geppetti, 2001; Almeida et al., 2004; Burbach, 2010). Both α-PPT-A and δ-PPT-A encode for SP, whereas β-PPT-A encodes for SP, neurokinin A and neuropeptide K, and γ-PPT-A encodes for SP, neurokinin A and neuropeptide gamma (Carter and Krause, 1990; Lai et al., 1998b; Almeida et al., 2004; Burbach, 2010). For an overview see figure 3. There are only a few studies concerning the expression of the different PPT-A mRNA isoforms in different tissues, but it seems that the most abundantly expressed isoforms are the γ-PPT-A and β-PPT-A, i.e. the two mRNA encoding both SP and neurokinin A (Krause et al., 1987; Carter and Krause, 1990; Lai et al., 1998a; Pintado et al., 2003). When SP is secreted from the SP-producing cell it either binds to a receptor, preferably the neurokinin-1 receptor (NK-1 R) (cf. section 1.4.1.2) or is degraded by proteases.
1.4.1.1.1 Neuronal SP
Historically, SP has been considered to only exist in the nervous system, both in the central and peripheral. The production of SP in neurons takes place in the ribosomes (Harmar et al., 1980; Harmar and Keen, 1982; Keen et al., 1982) and SP is thereafter transported in LDVs to terminal endings of axons (Merighi et al., 1988), most commonly in Aδ- and C-fibres, where its final enzymatic processing is performed (Brimijoin et al., 1980). Following nociceptive stimuli, the stored SP is released from its fibres into the synaptic cleft, where it performs its effect as a signalling substance (McCarthy and Lawson, 1989). One of the well-known signalling effects of SP is its involvement in pain transmission (Harrison and Geppetti, 2001).
1.4.1.1.2 Non-neuronal SP
In contrary to the original belief, SP has been shown to also exist in multiple non-neuronal cell systems, which furthermore express SP’s preferred receptor, NK-1 R. Examples of such systems are human immune cells (Lai et al., 1998a; Douglas and Leeman, 2011), bone marrow stem cells (An et al., 2011), endothelial cells (Milner et al., 1990), fibroblasts (Bae et al., 2002), and tenocytes (Andersson et al., 2008).

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
11
Fig. 3 The biosynthesis of substance P and related peptides. The illustration is based on information from Almeida et al., 2004.

Ludvig J. Backman, 2013
12
1.4.1.2 The neurokinin-1 receptor (NK-1 R)
There are three known tachykinin (also called neurokinin) receptors, named neurokinin-1 receptor (NK-1 R), neurokinin-2 receptor (NK-2 R), and neurokinin-3 receptor (NK-3 R) (Harrison and Geppetti, 2001). NK-1 R is the preferred receptor for SP, but SP is also known to bind to NK-2 R and NK-3 R, which otherwise are the preferred receptors for neurokinin A and neurokinin B, respectively (Regoli et al., 1994). Although SP is the preferred ligand for NK-1 R, it is also shown that neurokinin A can bind to NK-1 R with high enough affinity to elicit a biological response (Sagan et al., 1996); it depends on the availability of the receptor and concentration of the peptide (Regoli et al., 1994). All three receptor types consist of seven hydrophobic transmembrane domains, that are connected by both extra- and intracellular loops, and are coupled to G-proteins (Harrison and Geppetti, 2001).
The human NK-1 R gene produces two spliced isoforms of mRNA: One that encodes for a 407 amino acid (full-length) receptor, and a shorter one encoding for a 311 amino acid (truncated) receptor (Lai et al., 2008; Tuluc et al., 2009; Douglas and Leeman, 2011). SP’s binding affinity to the truncated form of NK-1 R, is 10 times lower and results in a reduced calcium response as compared to when SP binds to the full-length NK-1 R (Douglas and Leeman, 2011). Other differences in functional properties are that the full-length NK-1 R induces expression of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and interleukin-8 (IL-8) mRNA, and also that it more rapidly activates extracellular signal-regulated kinases 1 and 2 (ERK1/2), as compared to the truncated NK-1 R (Lai et al., 2008).
It is furthermore known that NK-1 R has two N-linked glycosylation sites on the extracellular amino terminus, residues 14 and 18 (Tansky et al., 2007). The glycosylation of the NK-1 R do not affect the binding affinity for SP. However, the glycosylation may stabilize the receptor in the plasma membrane (Tansky et al., 2007).
When SP has bound to NK-1 R, it forms a SP/NK-1 R complex, which is internalized via endosomes, i.e. membrane-bound compartments inside the cells (Grady et al., 1995). In the early internalization of SP via endocytosis, SP is intact, but eventually it slowly degrades in the perinuclear vesicles (Grady et al., 1995). The NK-1 R is recycled in situations of ordinary stimulation, but in cases of chronic stimulation, the NK-1 R has been observed to be down-regulated (Grady et al., 1996; McConalogue et al., 1999; Cottrell et al., 2006).
1.4.2 Neurotransmitters
Neurotransmitters are small substances that transmit nerve impulses across a synapse. They are synthesised by enzymes from available amino acids in the body and therefore the biosynthesis can take place at the site of release, thus avoiding the time consuming steps that are seen in the synthesis of neuropeptides (cf. above) (Sulzer et al., 2005). The neurotransmitters are stored in SSVs and exocytosis is a fact when the calcium threshold is reached. The SSVs are mainly clustered at synaptic junctions and upon stimuli the neurotransmitters are released to the synaptic cleft and modulate or mediate

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
13
communication by acting on cell surface receptors. The duration of neurotransmission is only milliseconds due to the fact that the enzymes responsible for the degradation of neurotransmitters are abundantly present in synaptic clefts (Bortolato et al., 2008).
1.4.2.1 Catecholamines
Catecholamines are a group of neurotransmitters, which in humans consists of epinephrine /adrenaline (EPI), norepinephrine/noradrenaline (NE), and dopamine (DA), all of which are biosynthesised in a process starting with a first step catalyzed by an enzyme called tyrosine hydroxylase (TH). The name ‘catecholamine’ derives from the fact that these transmitters contain a catechol group and an amine group, both organic compounds. TH converts the amino acid tyrosine, the precursor of all catecholamines, to a component called L-dihydroxyphenylalanine (L-DOPA). This process is the first rate-limiting step of the catecholamine biosynthesis. L-DOPA is in turn converted to DA, the first catecholamine in the biosynthetic cascade, by an enzyme called aromatic L-amino acid decarboxylase (or DOPA decarboxylase). DA is converted to NE by dopamine-β-hydroxylase and NE is converted to EPI by phenylethanolamine-N-methyl transferase (Molinoff and Axelrod, 1971; Nagatsu, 1991). See figure 4 for an overview of the catecholamine synthesis.
Essentially, the degradation of NE and EPI is achieved by monoamine oxidase (MOA), which is a deaminase enzyme, meaning that it removes an amine group (Eisenhofer et al., 2004). The metabolism of DA occurs by two enzymatic pathways, the MAO and catechol-o-methyl transferase (Eisenhofer et al., 2004).
1.4.2.1.1 Neuronal catecholamines
Catecholamines are used for communication between neurons in both the central and peripheral nervous systems, and they belong to the sympathetic nervous system. The main source of the catecholamines NE and EPI in the body
Fig. 4 The catecholamine synthesis.

Ludvig J. Backman, 2013
14
is the neuroendocrine cells in the medulla of the adrenal glands, and the main sources of DA are neurons of the substantia niagra and the ventral tegmental area in the midbrain. The neurons using catecholamines as communicating neurotransmitters are called catecholaminergic (dopaminergic, adrenergic, noradrenergic).
1.4.2.1.2 Non-neuronal catecholamines
Apart from in the nervous system (including the adrenal medulla), the rate-limiting synthesising enzyme of catecholamines, TH, has been shown to be expressed also at non-neuronal sites, such as in mouse hepatic stellate cells (Oben et al., 2004) and human tenocytes (Danielson et al., 2007b, a; Bjur et al., 2008a). The release of NE has also been confirmed in mouse hepatic stellate cells (Oben and Diehl, 2004). Both hepatic stellate cells and tenocytes also express the adrenergic receptors (Oben and Diehl, 2004; Danielson et al., 2007b; Bjur et al., 2008a).
1.4.2.2 Adrenergic receptors
The adrenergic receptors (or adrenoreceptors [ARs]) can be classified as α- or β-subtypes, and are responsible for binding EPI and NE. The α ARs have a higher ligand specificity to NE, while β ARs mainly interact with EPI (Levitzki, 1978). However, in general, the catecholamines exert their effect via interaction with both α/β ARs.
The α ARs include six subtypes (α1A, α1B, α1C, α2A, α2B, α2C) and there are three subtypes of β ARs (β1, β2, β3), all receptors being GPCRs.
Dopamine exerts its effect via dopamine D1- and D2-like receptors (Missale et al., 1998).
1.5 Non-neuronal production of signal
substances by tenocytes
1.5.1 Introduction
The hypotheses for the pathophysiology of tendinopathy have been many over the years. The two previous main theories were inflammation and separation of collagen fibres. However, Khan and collaborators presented a new view, first by showing contradicting arguments for the previous hypotheses, and then by raising a novel hypothesis of a biochemical explanation for tendinopathy (Khan et al., 2000). They also furthermore speculated that if this biochemical hypothesis proves to have some validity, it might result in new future treatments of tendinopathy focusing on modifying the biochemical mediator response (Khan et al., 2000).
After this hypothesis was raised in 2000 (Khan et al., 2000), studies have been published that support the biochemical theory; studies showing evidence of a local production of an array of biochemical mediators in the tendons,

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
15
particularly in tendinopathy (Danielson, 2009). It has been shown, via microdialysis of tendinopathy tendons, that higher levels of glutamate are seen in these tendons as compared to in normal tendons (Alfredson et al., 1999); a suggested source of production being the tenocytes based on immunohistological expression of vesicular glutamate transporter 2 (VGluT2) in these cells (Scott et al., 2008). In addition, evidence favouring local productions of acetylcholine (Danielson et al., 2006; Bjur et al., 2008b), tumour necrosis factor alpha (TNF-α) (Gaida et al., 2012), catecholamines (Danielson et al., 2007a, b; Bjur et al., 2008a), and SP (Andersson et al., 2008) in human tenocytes have been demonstrated in immunohistological studies. Furthermore, the hypothesis of possible autocrine (stimulation on the same cell type) or paracrine (stimulation on neighbouring cells; e.g., on blood vessels and nerves) loops of these signalling systems in tendons, has been suggested based on evidence that the tenocytes also express the preferred receptors of all these substances (Danielson et al., 2006; Danielson et al., 2007b, c; Andersson et al., 2008; Bjur et al., 2008a, b). The evidence of endogenous production of classically neuronal signal substances in non-neuronal tendon cells and the importance of neurogenic mechanisms (Hart et al., 1998; Hart et al., 1999) have in combination resulted in much focus on neuropeptides and neurotransmitters in tendinopathy research of recent years.
1.5.2 SP and NK-1 R
SP has been observed in the paratendinous loose connective tissue ventral to the Achilles tendon in both normal and tendinosis tissue, confined to blood vessel walls and nerve fascicles (Bjur et al., 2005; Andersson et al., 2007). It should be mentioned that the tissue in the ventral part of the tendon is most affected in tendinosis, in terms of ultrasound changes and colour Doppler activity (Ohberg et al., 2001). Regarding the tenocytes themselves, it has been shown that they express SP mRNA, indicating endogenous production of SP (Andersson et al., 2008). However, the presence of SP on the level of protein in vivo has been difficult to prove, which is speculated to be because of the immunohistochemical method not being sensitive enough for the detection of the low SP concentration thought to be produced by the tenocytes (Andersson et al., 2008). However, recently a study by Schizas and co-workers (Schizas et al., 2012) could confirm the presence of the SP peptide in the tenocytes and also concludes that it was expressed to a higher extent in tendinosis tissue as compared to in normal controls.
The NK-1 R has been shown to be present at the level of both mRNA and protein in tenocytes, and it is also shown that it is expressed to a higher degree in tendinosis tendon tissue as compared to in control tendons (Andersson et al., 2008).

Ludvig J. Backman, 2013
16
1.5.3 Catecholamines and α2-adrenoreceptors
The presence of TH, indicating production of catecholamines, has been observed in the paratendinous tissue of tendons, and most abundantly so in large nerve fascicles but also in single nerve fibres in the proximities of blood vessels (Andersson et al., 2007; Danielson et al., 2007b; Bjur et al., 2008a). Receptors for catecholamines, α1 AR, α2A AR and β1 AR, have also been shown to be present in the paratendinous tissue of tendons (Andersson et al., 2007; Danielson et al., 2007b; Bjur et al., 2008a).
Regarding tenocytes in the tendon tissue proper, they also display reactions for both TH and ARs, most prominently in areas where hypercellularity and cells with a changed morphology are seen (Danielson et al., 2007b, a; Bjur et al., 2008a). The AR with the clearest reactions in the tenocytes was α2A AR, which also was seen to a higher extent in tendinosis tissue as compared to in normal tissue (Danielson et al., 2007b).
1.6 Mechanisms of basic cell functions
1.6.1 Cell cycle and proliferation
The cell cycle in eukaryote cells (such as tenocytes), basically consists of two phases, one in which the cell prepares for mitosis (cell division), the S-phase, and the other phase in which the mitosis occurs, the M-phase. There are also two checkpoint phases, one before the cell gets committed to DNA-synthesis (the S-phase), called the G1-phase, and the other before cells go into mitosis (the M-phase), called the G2-phase. One additional phase is also described, called the G0 in which the cell enters a long-term quiescent (non-dividing) state. However, this is a state from which the cell can be reactivated by growth
Fig. 5 Schematic drawing of the phases of the cell cycle.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
17
factors to re-enter the cycle. It is during the G1-phase the GPCRs, such as NK-1 R and α2A AR, are able to affect either mitogenic or growth inhibitory signal pathways, resulting in the cell either entering, pausing or exiting the cell cycle (New and Wong, 2007). See figure 5 for an overview of the cell cycle.
ERK1/2 are members of the mitogen-activated protein (MAP) kinase family that has integrative effects on the fate of the cell cycle, i.e. they are either involved in cell death or proliferative signalling (New and Wong, 2007).
The mechanisms of ERK1/2’s involvement in mediating cell death and suppression of survival pathways are in general unknown (Mebratu and Tesfaigzi, 2009). However, it is shown that ERK1/2 activation, which occurs when ERK1/2 are phosphorylated, causes cell death in certain conditions and also that it can decrease the well known cell survival signalling pathway called the Akt-pathway (Mebratu and Tesfaigzi, 2009) (see section 1.6.2.1).
The more well known effect of ERK1/2 is their role in activation of cell cycle regulatory proteins, such as cyclin-dependent kinases, resulting in mitogenic signalling, i.e. initiation of the cell cycle (Johnson and Walker, 1999; Meloche and Pouyssegur, 2007; New and Wong, 2007; Mebratu and Tesfaigzi, 2009). ERK1/2 can be phosphorylated as a result of a ligand binding to a GPCR, which subsequently results in downstream targets modulating proliferation, if the stimulus is sustained throughout the G1 phase (Meloche and Pouyssegur, 2007). It is also suggested that ERK1/2 activation is required for G0 arrested fibroblasts to re-enter into the cell cycle (Pages et al., 1993).
1.6.1.1 Substance P and proliferation
Stimulation with SP has been shown to result in increased number of cells in in vivo Achilles tendon rupture rat models (Burssens et al., 2005a; Carlsson et al., 2011) and also to be associated with increased proliferation and decreased apoptosis in a wound healing rat model (Jing et al., 2010). The studies on the Achilles tendon rupture models also confirmed SP to stimulate angiogenesis and nerve ingrowth, and to facilitate collagen organisation. Furthermore, it has been shown that locally produced SP increases the number of cells in the rat thymus, a conclusion based on the results from treatment only blocking the SP-degrading enzyme, neutral endopeptidase (NEP) (Amantini et al., 2008).
In vitro models also confirm SP to have a proliferative effect in many different cell types such as glioblastoma cells (Yamaguchi et al., 2005b), astrocytoma cells (Yamaguchi et al., 2005a), colonocytes (Koon et al., 2004), chondrocytes (Opolka et al., 2012), mesenteric preadipocytes (Gross et al., 2009), and skin fibroblasts (Chen et al., 2006). In the case of SP stimulation in glioblastoma cells (Yamaguchi et al., 2005b), astrocytoma cells (Yamaguchi et al., 2005a), and colonocytes (Koon et al., 2004), it was shown that the proliferative effect involved phosphorylation of ERK1/2, and in the colonocytes, that the SP induced increase in cell viability was blocked when the phosphorylation of ERK1/2 was inhibited, evidence confirming the important effect of phosphorylated ERK1/2 in SP induced increased cell viability.

Ludvig J. Backman, 2013
18
1.6.1.2 Catecholamines and proliferation
Catecholamine production and adrenergic receptor expression have been confirmed to be present in Achilles tendon tissue (Danielson, 2009). So far, the effects of catecholamines on tendons in vivo have not been studied in an experimental Achilles tendinosis model. However, the effects of catecholamines have been explored in a liver fibrosis mice model, in which continuous administration of NE markedly stimulated proliferation of the fibroblastic cells (Oben et al., 2004). It has also been shown that NE deficient mice expressed a markedly reduced collagen level in the liver fibrosis model, suggesting NE to play an important role in the collagen production (Oben and Diehl, 2004).
The proliferative effect of catecholamines has been studied in vitro in a mouse tumour cell line (Bruzzone et al., 2008; Bruzzone et al., 2011), in proximal tubule cells (Cussac et al., 2002), in mouse embryonic stem cells (Kim et al., 2008), in human intestinal epithelial cells (Schaak et al., 2000), and in other cell lines (Karkoulias et al., 2006). The proliferative effect of catecholamines in the mouse tumour cell line was specifically mediated via α2A AR (Bruzzone et al., 2008; Bruzzone et al., 2011). The other mentioned studies show that α2 AR stimulation, via ERK1/2, has a proliferative effect.
1.6.2 Apoptosis
The balance of the number of cells present in a tissue is not only regulated by proliferation, i.e. increasing the amount of cells, it is also controlled by decreasing the number of cells through cell death, which can be categorized into two types, either necrosis or apoptosis.
Necrosis is a more chaotic type of cell death than apoptosis. In many cases necrosis occurs after external injury to the cell. The process of necrosis is that the cell swells and ultimately bursts, resulting in increased membrane permeability and release of intracellular components leading to inflammation (Kroemer et al., 2009). In contrast, apoptosis is the controlled type of cell death that occurs during development but also following particular cell stimuli. Kerr and collaborators, who mounted the term apoptosis, first described the mechanisms of apoptosis and that it, in contrast to necrosis, is a molecular process in which inflammation is avoided through cell shrinkage and the formation of small membrane-enclosed vesicles, called apoptotic bodies, which are readily phagocytised and digested by macrophages or by neighbouring cells (Kerr et al., 1972). It is not only the cytoplasm but also the nucleus that is condensed and later fragmented (Kroemer et al., 2009). For an overview of apoptosis and necrosis, see figure 6.
Following apoptotic cell stimuli, an intracellular signalling cascade is initiated in which caspases play a central role. The caspases can be divided into initiating, effector (executioner), and inflammatory caspases. The group of initiator caspases consists of caspase-2, -8, -9, and -10, the group of effector caspases consists of caspase-3, -6, and -7, and the group of inflammatory caspases consists of caspase-1, -4, and -5 (Wolf and Green, 1999; Elmore, 2007). Death receptor or mitochondrion dependent apoptosis is initiated by the recruitment and activation of initiator caspases. One such caspase activation

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
19
(via so-called Anti-Fas stimulation) is when the Fas ligand (FasL) binds to its receptor (Fas or Fas R), resulting in Fas-associated protein with death domain (FADD) recruitment, which is an adapter protein. FADD then recruits procaspase-8 via dimerization of the death effector domain (DED) and forms a complex, death-inducing signalling complex (DISC), resulting in cleavage/activation of caspase-8 (Kischkel et al., 1995; Salvesen and Dixit, 1999). Activated caspase-8 and the other initiator caspases cleave and activate the downstream effector caspases, and among the effector caspases, activation/cleavage of caspase-3 is perhaps the most studied and a common detection method for apoptosis, since it appears to play a crucial role in many apoptotic models (Porter and Janicke, 1999).
Fig. 6 In a, necrosis is illustrated, showing a cell swelling and finally losing its membrane integrity which subsequently causes inflammation. In b, apoptosis is illustrated, showing a cell shrinking and formation of apoptotic bodies that are phagocyted.
Another apoptotic characteristic, which often correlates to caspase activity, is
DNA fragmentation (Wolf et al., 1999). During apoptosis, DNA fragmentation occurs when caspases activate DNase (Liu et al., 1998) and/or cleave poly (ADP-ribose) polymerase (PARP) (Oliver et al., 1998). PARP is involved in DNA repair, but the repairing function is inactivated by its cleavage, which thereby contributes to apoptosis (Alvarez-Gonzalez et al., 1999), and cleaved PARP (c-

Ludvig J. Backman, 2013
20
PARP) is thus an established marker of cells undergoing apoptosis (Oliver et al., 1998). PARP is cleaved by activated (i.e. cleaved) caspase-3 (Tewari et al., 1995).
In addition, activated caspases, such as caspase-8, can also disturb the balance of the pro- and anti-apoptotic proteins belonging to the BCL-2 family. These proteins can affect the pores in the mitochondria, which might result in release, or prevention of release, of the apoptotic inducing cytochrome C (Adams and Cory, 1998; Li et al., 1998). If the pro-apoptotic proteins (e.g., BID and BAX) have an advantage, cytochrome C will be released and this subsequently leads to the ultimate state of apoptosis (Adams and Cory, 1998; Li et al., 1998). For an overview see figure 7.
1.6.2.1 Anti-apoptotic mechanisms
Activation of phosphoinositide 3-OH kinase (PI3K) has been shown to provide cells with a survival signal that allows cells to withstand apoptotic insult (Yao and Cooper, 1995). It has also been shown that the major component of the survival signalling by the PI3K is the protein kinase Akt, also called protein kinase B (PKB), which is phosphorylated, and thereby activated, by PI3K into phosphorylated Akt (P-Akt) (Downward, 2004). There are many different anti-apoptotic pathways, not only involving regulators controlling the activity of Akt (Scheid and Woodgett, 2003). However, the focus here will be on the Akt subfamily, which consists of three isoforms, Akt1, Akt2, and Akt3, also called PKBα, PKBβ and PKBγ, respectively, which are all produced from distinct genes (New et al., 2007). Akt1 and Akt2 are generally expressed in the body (Hanada et al., 2004), whereas Akt3 is found predominantly in the brain, kidney, and heart (Masure et al., 1999). One of the first discovered targets of Akt, in its anti-apoptotic effect, was inactivation of BAD, which is a pro-apoptotic protein that exerts its effect by blocking anti-apoptotic proteins, like BCL-2 and BCL-XL and their effects in the mitochondria (Datta et al., 1997; Datta et al., 2002). In a similar way, Akt also blocks BAX, thus preventing its pro-apoptotic effects in the mitochondria (Maddika et al., 2007). Akt can also have an anti-apoptotic effect, indirectly, through increased expression of the anti-apoptotic protein BCL-2 (New et al., 2007). Other anti-apoptotic effects of the Akt pathway are achieved by inactivation of promoters of apoptosis such as Forkhead box (FOX) (Tzivion et al., 2011), proteins which are a family of transcription factors that transcribe mRNA coding for proteins that are apoptosis inducers, such as Fas ligand (Brunet et al., 1999). Finally, Akt can also target and inactivate the initiator caspases, and thereby reduce caspase-mediated apoptosis (New et al., 2007). For an overview see figure 7.
1.6.2.1.1 SP and anti-apoptotic mechanisms
In vivo rat tendon rupture models have shown that SP injections in the area of the rupture result in hypercellularity (Burssens et al., 2005a; Carlsson et al., 2011). In the case of tendon rupture models, which are known to express increased apoptosis (Lundgreen et al., 2011), the hypercellularity seen after SP stimulation is likely to also involve anti-apoptotic effects, not only proliferation, to explain the increased number of cells seen. Actually, in rat thymocytes, it has been shown that blocking of the SP-degrading enzyme, leading to higher SP

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
21
concentration, also results in a reduction of apoptosis, accompanied with an activation of Akt, suggesting endogenous SP to have an anti-apoptotic effect via the Akt-pathway (Amantini et al., 2008).
In vitro models confirm SP to have anti-apoptotic effects in human mesenteric preadipocytes (Gross et al., 2009), fibroblasts (Jing et al., 2010), bone marrow cells (An et al., 2011), and colonocytes (Koon et al., 2007). In the colonocytes, it was also shown that this anti-apoptotic effect of SP was mediated via the Akt-pathway.
Fig. 7 Simplified, schematic illustration of the apoptotic pathway after Anti-Fas induced apoptosis. The picture also illustrates a simplified, indirect pathway of Akt mediated anti-apoptotic effects.

Ludvig J. Backman, 2013
22
2. HYPOTHESES AND AIMS
2.1 Hypotheses
The overall hypothesis of our studies is that the non-neuronal production of classically neuronal signal substances in tenocytes is increased in response to load and is a possible contributing factor in the development and maintenance of tendinosis.
More specifically, in this thesis work it is hypothesized that intratendinous production of the neuropeptide substance P (SP) is up-regulated in response to load of tendons/tenocytes, and that SP, as well as adrenergic α2A AR stimulation, contribute to the hypercellularity seen in tendinosis, via increased proliferation and/or decreased apoptosis, and that SP stimulates tendon angiogenesis.
2.2 Aims
The specific aims of this thesis were:
1) To study the expression, in the Achilles tendon tissue and cells, of SP and its preferred receptor NK-1 R, in both an in vivo [I] and an in vitro [III] model, and also to study the presence of catecholamines, as evaluated by the expression of TH, and the presence of the α2A AR, in the in vivo and in vitro model [IV].
2) To study if SP is up-regulated in response to loading of tendons/tendon
cells in vivo [I] and in vitro [III], and if an up-regulation of SP precedes the tissue changes characteristic of tendinosis [I].
3) To study the possible effects of exogenous SP and an α2 AR agonist
(clonidine) on tenocyte hypercellularity/proliferation both in vivo [II, IV] and in vitro [III, IV, V], and if this involves the ERK1/2 pathway, as well as their possible effects on intratendinous vascularity in vivo [II, IV].
4) To study if SP, through NK-1 R stimulation, has an anti-apoptotic effect
via Akt-phosphorylation in vitro, in an apoptosis model for tenocytes [V].

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
23
3. MATERIAL AND METHODS
3.1 Animal model of Achilles tendinosis
3.1.1 Rabbit material
In total 47 female New Zealand white rabbits were used in the experimental studies, all obtained from a licensed laboratory animal breeder.
All rabbits used were female rabbits, which has to be taken into consideration when interpreting the results. Female rabbits do not have a true oestrous cycle with spontaneous ovulation. Ovulation only takes place after mating, as a result of luteinizing hormone secretion from the pituitary gland (Zutphen et al., 2001).
Another important factor to take into account in interpreting the results is whether the rabbits are skeletally mature, i.e. have closed epiphyses, when used for experiments. The closing of epiphyses for New Zealand white rabbits is described to take place at an age of 6-7 months (Woo et al., 1986), however some have reported this to occur at an older age (Hart et al., 1995). The rabbits used were delivered to the laboratory when they had reached a weight of approximately 4kg, which results in the rabbits being in the age span of 6-9 months.
All rabbits, except for the untrained controls, were subjected to an experimental protocol to induce tendinosis-like changes (see section 3.1.3). It was not clearly obvious that the animals showed amended movements or changed behaviours inbetween the experimental sessions. However, it was noticed that the experimental rabbits developed a palpable thickening of the tendon as the training went on.
3.1.1.1 Rabbit groups
Group N Mean weight at start (kg)
Papers
Untrained controls 6 4.5 I, II, IV
1 week training 6 4.1 I, II, IV
3 weeks training 6 4.1 I
6 weeks training 6 4.4 I
1 week training + SP injections
6 4.6 II
1 week training + Clonidine injections
6 4.2 IV
1 week training + BRL injections
6 4.6 IV
1 week training + NaCl-injections
5 4.2 II, IV

Ludvig J. Backman, 2013
24
3.1.2 Ethical considerations
The experiments were approved by the local ethical committee for research on animals in Umeå, Sweden (project numbers A34/07 and A95/07) and the animals were bred by a licensed breeder for the purpose of being used only as experimental animals. The Board of Agriculture approved the housing of the rabbits.
3.1.3 Experimental design
The rabbits included were divided into different groups with 6 rabbits in each group at the start of the experiments. The basic set-up consisted of groups that exercised one week, three weeks or six weeks according to the protocol (see section 3.1.3.1). One group did not train at all and served as an untrained control group. Four other groups received injections in conjunction with one week of training (cf. 3.1.3.2). See table in section 3.1.1.1 for an overview of the rabbit groups. All exercise was performed on the animals during general anaesthesia (see section 3.1.4).
Although all groups originally had 6 rabbits, the NaCl-injected group had 5 rabbits completing the protocol due to one rabbit being lost to complications of anaesthesia.
3.1.3.1 ”Exercise” protocol
A kicking machine that was originally designed by Backman and collaborators (Backman et al., 1990) was re-established and slightly modified (Andersson et al., 2011).
The kicking machine generates a passive dorsiflexion and plantarflexion of the right ankle using a pneumatic piston. The right foot of the rabbit was tightly attached to the piston, which moved 9.5 cm resulting in 20-25° dorsiflexion and 30-35° of plantarflexion in the ankle joint. To isolate the movement to the ankle joint, the pelvis and knee were strapped with bands to hold the pelvis fixed and to keep the knee joint in 90° angle, respectively. For an illustration of rabbits positioned in the kicking machine, see figure 8.
To mimic the situation of running in human the rabbit was stimulated with electrodes (pediatric electrodes 40 426A; Hewlett Packard, Andover, Massachusetts, USA) 2 cm apart on the triceps surae muscle to achieve a concentric contraction of the muscle at the plantarflexion movement. A microswitch was synchronized with a stimulation unit to achieve a muscle contraction in the plantarflexion movement 85 ms into the plantarflexion with a single impulse of 0.2
Fig. 8 Rabbits positioned in the kicking machine.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
25
ms. The total plantarflexion was 200 ms and the total time for the dorsiflexion was 197 ms. The amplitude of the stimulation varied in the range of 35-50 V depending on the contact of the electrodes, the amplitude of the stimulation was manually adjusted to make sure a muscle contraction occurred. The frequency of the movement (plantar and dorsiflexion) of the ankle joint was 2.5Hz, i.e. 150 movements per minute. The total training time was 2h for each session and the training sessions were repeated every second day. See figure 9 for an overview of the settings of the kicking machine.
The non-exercised, contralateral, leg was kept at total rest throughout the training sessions.
Fig. 9 Illustration of the settings in the kicking machine.
3.1.3.2 Exogenous injections
The New Zealand white rabbits that were subjected to injections were given their injections immediately after the exercise protocol, which was performed every second day for one week, i.e. a total of four injections. The injected groups were either injected with SP, clonidine (α2 agonist), BRL 44408 maleate (α2A antagonist) or NaCl. All injections were given with a volume of 1 mL distributed ventrally, along the entire length of the tendon.
3.1.3.2.1 Substances for in vivo experimental use
Substance Code Source Type Concentration
SP s6883 Sigma, St Louis, MO, USA
Tachykinin receptor agonist
10 -8 μmol/mL
BRL 44408 maleate
1133 Tocris Bioscience, Bristol, UK
α2A antagonist 400 μg/mL
Clonidine c7897 Sigma, Saint Louis, MO, USA
α2 agonist 400 μg/mL

Ludvig J. Backman, 2013
26
3.1.4 Animal care and anaesthesia
All animals were kept in ordinary individual cages, allowing free movement and free access to food and water. The laboratory kept a 12:12 hour light-dark cycle.
General anaesthesia was induced by intramuscular injection of fentyl-fluanisone (0.2-0.3 mM/kg) and diazepam (5 mg/mL, 0.2 mL/kg). The anaesthesia was kept with injections of fentyl-fluanisone (0.1 mL/kg) every 30-45 minutes. Subcutaneous injection of buprenorphine (0.01-0.05 mg/kg) was given after each training session for analgesia.
3.1.5 Rabbit biopsies
The rabbits were sacrificed with an overdose of pentobarbital one day after the last training session. The rabbit limbs were dissected and the entire Achilles tendon was taken to the laboratory immediately to achieve best preservation of the biopsies. Each tendon biopsy was divided into three parts, the distal part close to the OTJ (A), mid-portion part (B), and the proximal part, MTJ, (C). All samples had a cross sectional area of approximately 5x5 mm2 and the samples collected for ELISA had a weight of around 0.03 g.
In all groups but the one injected with NaCl, biopsies were taken from the Achilles tendon from both legs, i.e. the ipsilateral/exercised and contralateral leg.
3.1.5.1 Tissue preparation
Biopsies were either directly frozen or chemically fixed before freezing. The unfixed biopsies that were used for sectioning, immunohistochemistry and in situ hybridisation, were mounted on thin cardboard in OCT embedding medium (Miles Laboratories, Naperville, IL, USA) and frozen at -80°C in liquid nitrogen-chilled propane.
The biopsies to be chemically fixed were immediately placed in a 4% solution of formaldehyde diluted in 0.1 phosphate buffer (PBS), pH 7.0 at 4ºC overnight. The fixation is performed to preserve the tissue from decay, thereby preventing the self-digesting action by the tissues own enzyme, called autolysis. After carefully washed in 10% sucrose-containing Tyrode’s solution the biopsies were mounted on thin cardboard in OCT embedding medium (Miles Laboratories) and frozen at -80°C in liquid nitrogen-chilled propane.
Biopsies that later were homogenised and used for ELISA, were snap frozen in liquid nitrogen before stored in -80ºC.
3.1.5.2 Sectioning
Biopsies were sectioned at a thickness of 7 μm in a transverse orientation using a cryostat (Microsystem CM 3000, Heidelberg, Germany) set at a temperature of -24°C. The sections were mounted on chromealun gelatin pre-coated slides, dried and subsequently stained either with immunohistochemistry or hematoxyline-eosin. The sections that were not immediately stained were stored at -24°C until stained.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
27
For in situ hybridization the cryostat knife was washed in 70% EtOH in DEPC-H2O in order to prevent presence of any RNAse:s. The sections were cut at a thickness of 10 μm and mounted on Super Frost Plus slides (nr. 041300, Menzel-Gläser). Sections were done the same day as the start of the staining procedure.
3.2 Primary human tendon cell culture
model
3.2.1 Human material
The human material consisted of biopsies taken from a total of nine healthy individuals who volunteered to give a biopsy. Healthy controls were defined as individuals without history of Achilles tendon pain and with no abnormalities such as disorganized collagen structure or high intratendinous blood flow displayed at colour Doppler ultrasound examination. All examinations were performed by the same experienced ultrasound user (Prof. Alfredson).
The biopsies were harvested in strict sterile conditions. Under local anaesthesia the biopsies were taken from the ventral side of the mid-portion of the Achilles tendon (the most affected site in tendinosis patients) through a small skin incision lateral to the Achilles tendon (2-6 cm from its calcaneal insertion). The ventral side of the tendon was exposed and a small biopsy of 1-2 x 3-5 x 1-2 mm3 (w x h x d) was taken with a scalpel and directly placed in a sterile tube containing saline. The biopsies were immediately taken to the laboratory and prepared for cell culture. The small skin incision was sutured with 2 stitches and the patient was given crutches to use the first day and recommended to unload the tendon for the first 12h after the surgery. The regime for the first week was to leave the crutches after 1 day and gradually increase walking, then for the second week to progressively return to normal activity. Demanding sport activities were avoided the first two weeks to limit the risks of complications.
None of the donors reported any complications.
3.2.1.1 Human subjects
Code Sex Age Papers
P1 Male 25 III
P2 Male 31 III, IV
P3 Female 39 III, IV
P4 Male 23 III, IV
P5 Female 29 V
P6 Male 26 V
P7 Female 59 V
P8 Male 21 V
P9 Male 28 V

Ludvig J. Backman, 2013
28
3.2.2 Ethical considerations
The Regional Ethical Review Board in Umeå, Sweden, approved the harvesting and culture of human Achilles tendon tissue/cells (project number 04-157M). Everything was performed according to the principles of the Declaration of Helsinki and written informed consent was received from all participants.
3.2.3 Isolation of human Achilles tendon
derived tenocytes and cell culturing
The biopsies were thoroughly washed under sterile conditions in Hanks’ Balanced Salt Solution (HBSS; Invitrogen, Grand Island, NY, USA; code: 14170) and rinsed from any visible tissue that was not the tendon tissue proper, such as fat tissue and vessels, to reduce contamination of different cell types in the cell cultures. Subsequently, the biopsies were divided into equally small pieces and enzymatically digested. The digestion was done in 37°C at constant agitation in collagenase (Clostridopeptidase A, Sigma; code: C-0130) diluted in D-MEM (Invitrogen; code: 11960) to obtain a concentration of 2 mg/ml. The digestion product was centrifuged at 800 relative centrifugal force (rcf) for 5 minutes to obtain a cell pellet. The supernatant was discarded and the cell pellet was washed and finally dissolved in D-MEM supplemented with 10% foetal bovine serum (FBS; Invitrogen; code: 16000), 1% pen-strep (Invitrogen; code: 15410) and 0,2% L-Glutamine (Invitrogen; code: 25030) and cultured in a humidified atmosphere of 5% CO2 in air and 37°C. After 72 h the medium containing non-adherent cells was removed and fresh medium was added. At confluence the cells were enzymatically detached using 0.05% trypsin with EDTA (Invitrogen; code: 15400), removed and split into 1:3 ratio. Medium was changed every third day.
3.2.4 Experimental set-up
3.2.4.1 Cell passage and serum-starvation
For all experiments, cells from passage 3-6 were used to minimize the possible impact of phenotype drift in cultures. It has been shown that tendon cells in culture do exhibit phenotype drift in form of a decreased decorin expression, decreased tenomodulin expression, increased collagen type III, and a decreased collagen I expression, with increased number of passages, in this case when cells were compared from passage 1-8 (Yao et al., 2006; Mazzocca et al., 2012). We did not study the phenotype drift in our cells, however the morphology of the cultured cells did not appear to differ in the light microscope for the passages used. We did also confirm that almost all cells, approximately >95%, exhibited markers of tenocytes, such as tenomodulin, scleraxis, vimentin and collagen I, in all the passages used for the experiments (see chapter 4.1).
All cells used for experiments were serum-starved, i.e. incubated with medium only containing 1% FBS one day prior to and throughout the entire

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
29
experiments (if not otherwise stated), to reduce the metabolic activity seen in cells cultured in 10% FBS and to limit the unwanted effects of the serum. Macroscopically the cells had a normal appearance in 1% FBS and the metabolic activity in the cell cultures steadily increased over a four day period as shown with the metabolic assay MTS (for detailed information about the MTS assay, see chapter 3.3.7.4). However, there was clearly less metabolic activity as compared with cells cultured in 10% FBS, see figure 10.
Fig. 10 Metabolic activity of human tendon cells in culture, as measured with MTS assay, in different culture conditions: 0.1%, 1% or 10% FBS. The metabolic activity was measured over 4 days of incubation. Absorbance at 490 nm on the Y-axis.
3.2.4.2 Substances for in vitro experimental use
Substance Code Source Type Concentration Paper
SP 05-23-0600
Calbiochem, San Diego, USA
Tachykinin receptor agonist
10-7 – 10-9 M III, V
NK-1R antagonist
S3144 Sigma, St Louis, MO, USA
NK-1 R antagonist 10-7 M III
BRL 44408 maleate
1133 Tocris Bioscience, Bristol, UK
α2A antagonist 10-5 M IV
Clonidine C7897 Sigma, Saint Louis, MO, USA
α2 agonist 10-6 M IV
L-733,060 hydrochloride
1145 Tocris NK-1 R antagonist 10-6 – 10-8 M V
Akt Inhibitor V, Triciribine
124012 Calbiochem Akt inhibitor 40 μM V
Z-VAD-FMK G7231 Promega, Madison, WI, USA
Pan-Caspase inhibitor
10 μM V

Ludvig J. Backman, 2013
30
3.2.4.3 2D strain
Strain on cell cultures was achieved via vacuum deformation of a collagen I pre-coated elastic membrane (Bioflex BF-3001C). The vacuum deformed the membrane downwards at the outside edge of a 25 mm diameter cylindrical loading post, causing the membrane to stretch across the loading post face (figure 11). The cylindrical loading post results in an equibiaxial strain. Equibiaxial strain applied on cells is considered similar to that of a uniaxial loading for each individual cell. The strain on each individual cell depends on the orientation along the axis, however an average strain is 37 +/-8 % of the applied equibiaxial substrate strain (Wall et al., 2007).
A frequency of 1 Hz, operating for a total of 120 minutes with 10% strain, was applied in our study. Cells were strained once a day for 3 days before samples were collected. The protocol was based on an earlier study by Scott and collaborators (Scott et al., 2011). FlexCell strain unit FX-4000 tension plus biotechnology for cellular biomechanics (FlexCell International Corporation) was used to apply strain.
All samples were run at 6 wells/group.
Fig. 11 Schematic drawing of the FlexCell system showing the vacuum induced traction on the elastic membrane from the under-surface edge of the cylindrical loading post. As the membrane is strained downwards it results in an equibiaxial tension of the cells in culture.
Reprinted from the article Backman LJ, Fong G, Andersson G, Scott A, Danielson P: Substance P is a mechanoresponsive, autocrine regulator of human tenocyte proliferation. Plos One 2011;6(11):e27209 [III]

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
31
3.3 Methods of analysis
3.3.1 General morphology
3.3.1.1 Hematoxylin and eosin staining
3.3.1.1.1 In vivo model
Pre-fixated sections were air-dried and afterwards stained in freshly filtered hematoxylin solution (Harris HTX Histolab; Gothenburg, Sweden; code: 1312002) for 5 minutes. The hematoxylin staining was followed by a single wash in MilliQ water before staining in Eosin Y (Sigma, St Louis, MO, USA; code: E6003) diluted in ethanol for 5 minutes. Subsequently, slides were placed in 0.1% acetic acid for 15 seconds followed by 4 minutes in warm water. The acetic acid is performed to make the Eosin Y staining more intense and the warm water neutralize the acid and form an insoluable blue hematin complex. Before mounting in xylene mounting solution cells were dehydrated in 95% alcohol 3 times for 3 minutes each.
The structures stained with hematoxylin (blue colour), are the basophilic structures such as cell nucleus and ribosomes. The cytoplasmic content and extracellular proteins, structures that are eosinophilic, are stained by Eosin Y (pink or red colour).
3.3.1.1.2 In vitro model
Cell cultures were fixed in 20% paraformaldehyde (PFA) diluted in 0.1 M phosphate buffer (PBS) (pH 7.4) for 5 minutes. The reason for a strong fixation was to try to fix the cells undergoing apoptosis to the culture slides, otherwise the apoptotic cells tend to detach from the culture surface or the neighbouring cells and be missed in the analysis. After the fixation 2 washes for 2 minutes each in PBS was performed before the staining procedure, which was the same as for the biopsies from the in vivo model (cf. above).
3.3.2 Immunostaining
Immunostaining was performed on both the rabbit Achilles tendon tissue sections (immunohistochemistry) and human primary tendon cell cultures in vitro (immunocytochemistry).
3.3.2.1 Immunohistochemistry
Sections stained for TH, and α2A AR were pre-treated with potassium permanganate for 2 minutes. This procedure is shown to enhance specific immunoreactions (Hansson and Forsgren, 1995). Afterwards sections were washed in PBS 3 times for 5 minutes each. Sections stained for SP, TH, α2A AR,
NK-1 R, and CD31 were incubated in 1% triton solution for 20 minutes to achieve membrane permeabilization. Subsequently, after a series of washing,

Ludvig J. Backman, 2013
32
incubation with normal serum at a concentration of 5% for 15 minutes was done to block any potential unspecific binding. The normal serum chosen is from the same species as that in which the secondary antibody was raised. The serum was discarded and the primary antibody was incubated 60 minutes at 37ºC, or overnight at 4ºC, in a humidified incubator. After washing, the blocking in normal serum was performed again before incubation with secondary antibody for 30 minutes at 37ºC. The secondary antibody is raised against the species that the primary antibody was raised in. Finally, after washing, the stainings were mounted in Vectashield H-1000 mounting medium (Vector Laboratories, Burlingame, California, USA).
In cases of double staining, incubation with a second primary antibody was performed after the incubation with the first secondary antibody. The same protocol as described was then repeated. In a double staining the primary antibodies must contain different Fab-fragments (i.e. be raised in different species) to make possible for the secondary antibodies to bind specifically to only one primary antibody each.
As for control stainings, the primary antibody was either replaced with PBS or pre-absorbed with the peptide to saturate the primary antibody. A Zeiss Axioskop 2 plus microscope equipped with epifluorescence and an Olympus DP70 digital camera was used for analysis.
3.3.2.1.1 Primary antibodies used for immunohistochemistry
Antigen Code Source Type Dilution Tissue Method Paper
SP SC-14104 Santa Cruz, CA, USA
Goat 1:50 Fixed Alexa I
NK-1 R SC-5220 Santa Cruz, CA, USA
Goat 1:50 Fixed Alexa I, II
CD31 M0823 Dako, Glostrup, Denmark
Mouse 1:100 Unfixed TRITC II, IV
TH P40101 Pel-Freez, Rogers, AR USA
Rabbit 1:100 Fixed FITC IV
α2A AR PC161 Oncogene, Boston, MA, USA
Rabbit 1:50 Fixed TRITC IV
3.3.2.1.2 Pre-absorption antigens for immunohistochemistry
Pre-absorption antigen
Code Source Used with antibody
Dilution Paper
SP s6883 Sigma, St Louis, MO, USA
SP; SC-14104 50 μg/mL I
NK-1 R SC-5220P Santa Cruz, CA, USA
NK-1 R; SC-5220
50 μg/mL I, II
3.3.2.2 Immunocytochemistry (in vitro)
Cells were plated and left to adhere before being stained for SP, vimentin, tenomodulin, TH, α2A AR, Cleaved-PARP, and FAS-receptor.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
33
The procedure started with fixation in 2% paraformaldehyde (PFA) diluted in
0.1 M PBS (pH 7.4) for 5 minutes followed by washing, i.e. cells were not pre-treated with potassium permanganate or triton as in the immunohistochemistry of tissue sections. After fixation cells were blocked in 5% serum for 15 minutes and afterwards incubated with the primary antibody for 60 minutes at 37ºC in a humidified incubator. The same incubation condition was used for all primary antibodies except the antibody for detection of Fas-receptor, which was incubated 120 minutes at room temperature. This was followed by washing and incubation with the secondary antibody for 30 minutes at 37ºC in a humidified incubator before cells were mounted. The mounting media was supplemented with 4',6-diamidino-2-phenylindole (DAPI; Vector Laboratories; code: H-1500). DAPI is a fluorescent stain that binds strongly to A-T rich regions in the DNA, i.e. giving the nuclei of the cells a blue colour.
As for control staining, the primary antibody was replaced with PBS. A Zeiss Axioscop 2 plus microscope equipped with epifluorescence and an Olympus DP70 digital camera was used for analysis.
Complementary staining was performed using antibodies against PGP 9.5 at a concentration of 1:50, SSEA-4 at a concentration of 1:200 and CD31 at a concentration of 1:100 (results not reported in any of the papers). The procedures of the staining were performed as described above.
3.3.2.2.1 Primary antibodies used for immunocytochemistry
Antigen Code Source Type Dilution Fixation Method Paper
NK-1R SC-5220 Santa Cruz, CA, USA
Goat 1:50 Fixed FITC III
Vimentin M0725 Dako, Glostrup, Denmark
Goat 1:100 Fixed TRITC III
Tenomodulin SC-49325
Santa Cruz, CA, USA
Goat 1:100 Fixed FITC III
SP 8450-0004
Serotec, Oxford, UK
Rabbit 1:100 Fixed TRITC III
TH P40101 Pel-Freez, Rogers, AR USA
Rabbit 1:100 Fixed FITC IV
α2A AR PC161 Oncogene, Boston, MA, USA
Rabbit 1:50 Fixed TRITC IV
c-PARP 9541 Cell Signal, Danvers, MA, USA
Rabbit 1:100 Fixed TRITC V
FasR 05-201 Millipore, Billerica, MA, USA
Mouse 5 μg/mL Fixed FITC V
PGP 9.5 7863-0504
Biogenesis, Poole, UK
Rabbit 1:50 Fixed TRITC *
CD31 M0823 Dako, Glostrup, Denmark
Mouse 1:100 Fixed TRITC *
SSEA-4 41-4000 Invitrogen Mouse 1:200 Fixed TRITC *
* complementary experiments, not reported in any of the original papers

Ludvig J. Backman, 2013
34
3.3.2.3 Normal sera
Serum Code Source Dilution Paper
Donkey 017-000-121 Jackson I.R. West Grove, PA, USA
1:20 I, II, III
Rabbit X0902 Dako, Copenhagen, Denmark
1:20 II, III, IV
Swine 014-000-121 Jackson I.R. West Grove, PA, USA
1:20 III, IV, V
3.3.2.4 Secondary antibodies used
Secondary antibody
Code Source Used against primary antibody
Dilution Paper
Alexa Fluor 488, donkey anti-goat
A-11055 Invitrogen, CA, USA
Goat 1:300 I, II, III
TRITC-conjugated rabbit anti-mouse
R0270 Dako, Copenhagen, Denmark
Mouse 1:40 II, III, IV
TRITC-conjugated swine anti-rabbit
R0156/Z0259 Dako, Copenhagen, Denmark
Rabbit 1:40
III, IV, V
FITC-conjugated donkey anti-goat
705-095-147 Jackson immunoresearch. West Groove, PA, USA
Goat 1:40 III
FITC-conjugated swine anti-rabbit
F0205 Dako, Copenhagen, Denmark
Rabbit 1:40 IV
FITC-conjugated goat anti-mouse
F9259 Sigma, St Louis, MO, USA
Mouse 1:200 V
3.3.3 In situ hybridization
In situ hybridization was used to detect mRNA in the rabbit tissue. The procedure is well established and used earlier in our laboratory (Panoskaltsis-Mortari and Bucy, 1995; Danielson et al., 2007a). Details of all solutions mentioned below in the explanation of the procedure are described in table 3.3.3.1.
Freshly made 10 m thick cryosections either fixed or unfixed, were air-dried at room temperature for 30 minutes and afterwards post-fixed for 60 minutes with 4% paraformaldehyde diluted in 0.1 M sterile filtered PBS. After rinsing two times in 2xSSC for 10 minutes, each section was incubated in 0.2 M HCI at room temperature for 8 minutes to inhibit the endogenous alkaline phosphatase (AP)-activity (inhibition of unspecific reactions). Sections were acetylated at room temperature, to reduce background-staining, for 15 minutes in a mixture

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
35
of 195 mL diethylpyrocarbonate (DEPC)-water; 2.7 mL triethanolamine, 0.355 mL concentrated HCI and 0.5 mL acetic anhydride. After another wash in 2xSSC the denaturated ssDNA probes were added and the slides were covered with cover slips, sealed with nail polish, and incubated at 56ºC overnight. Denaturation of the probes at 80ºC for 5 minutes in 15 uL hybridisation solution was performed to avoid any hairpin formation. The probes were cooled on ice before administered on sections.
To eliminate excess of probes, sections were washed 2 times in 2xSSC for 10 minutes each and incubation in STE-buffer for 5 minutes, all done in room temperature. Sections were thereafter incubated with 100 μL of RNase A solution at 37ºC for 30 minutes. This was followed by a series of washes; first in 2xSSC supplemented with 50% formamide and incubated at 56ºC for 20 minutes, 2 washes in 1xSSC for 5 minutes each at room temperature and finally 2 washes in 0.5xSSC for 5 minutes each. Sections were after this incubated in buffer 1 for 5 minutes before incubation of the same buffer supplemented with 4% horse normal serum and incubated 60 minutes in a humid chamber at room temperature. Thereafter the sections were incubated in the alkaline phosphatase (AP)-labelled anti-digoxigenin (DIG) antibody diluted 1:500 with buffer 1 supplemented with 5% horse serum, added at a volume of 100 μL to each section and incubated for another 60 minutes in the humid chamber at room temperature. Additional washes was performed, this time 2 washes 10 minutes each in buffer 1 followed by 2 washes 5 minutes each in buffer 2. Finally, freshly sterile-filtered (22 μm filtered) substrate solution was added and incubated overnight at 4ºC. During the overnight incubation the substrate solution reacted with the enzyme alkaline phosphatase, which resulted in a detectable reaction. The incubation was performed upside-down to avoid deposits in the tissue.
To stop the colour reaction the sections were placed in buffer 3. Finally, before sections were mounted using Pertex, a counterstaining with methyl green was performed. The sections were washed in a series of steps; 30 seconds in 75% ethanol, followed by 30 seconds in 95% ethanol, 4-5 seconds in 0.5%methyl green and finally dipped in pure ethanol three times.
The entire procedure was performed under sterile conditions to avoid RNA degradation.
3.3.3.1 In situ hybridization solutions
Solution Ingredients
Diethylpyrocarbonate (DEPC) 0.1% vol/vol, autoclaved
4% paraformaldehyde 2 g of PFA dissolved in RNase-free H2O (25 mL) with added phosphate-buffer saline (PBS) (25 mL)
0.2 N HCI Diluted in DEPC-H2O
0.25% Acetic anhydride/0.1M triethanolamine- HCI buffer
0.93 g triethanolamine-HCI powder dissolved in 45mL DEPC-H2O with NaOH added (pH8). Diluted to 50 mL with DEPC-H2O.
STE-buffer 500 mM NaCl, 1 mM EDTA and 20 mM Tris-HCI, pH 7.5.
RNase A solution 40 ug RNase A/mL STE
20x SSC 3 M NaCl and 0.3 M sodium citrate, pH7.0 - autoclaved
2xSSC, 50% formamide 25 mL formamide (Sigma Chemical, St.Louis, MO, USA), 5 mL 20xSSC. Diluted to 50 mL with double distilled H2O (ddH2O)

Ludvig J. Backman, 2013
36
Buffer 1 100 mM Tris-HCI, pH7.5 and 150 mM NaCl. Diluted to 1 L with ddH2O. Filtered.
Buffer 2 100 mM Tris-HCI, pH9.5, 100 mM NaCl and 50 mMMgCl2. Diluted to 1 L with ddH2O. Filtered.
Substrate solution 1 mL buffer 2, 20 μL NBT/BCIP (Roche, Mannheim, Germany), 10 μL levamisole solution (1 mM) (Vector Labs, Burlingame, CA, USA)
Buffer 3 (stop buffer) 10 mM Tris-HCI, pH 8.0 and 1 mM EDTA, diluted to 1 L with ddH2O. Filtered.
Hybridization solution 500 μL formamide (Sigma, St.Louis, MO, USA), 200 μL 20xSSC, 50 uL of 20x Denhardt´s solution (Sigma), 50 μL of heat-denaturated herring sperm DNA (10mg/ml), 25 μL baker’s yeast RNA (10mg/ml) and 175 uL of dextran sulphate 50%
3.3.3.2 ssDNA anti-sense probes
Probe Code Source Dilution Tissue Paper
SP mRNA probe1
GD1001-CS (custom made)
GeneDetect, Auckland, New Zealand
50-100 ng in 15 μL hybridization solution
Fixed I
TACR1 mRNA probe cocktail2
GD1001-DS (custom made)
GeneDetect, Auckland, New Zealand
50 ng in 15 μL hybridization solution
Fixed/ unfixed
I, II
TH
mRNA probe3
GD1811-OP GeneDetect, Auckland, New Zealand
50 ng in 15 μL hybridization solution
Unfixed IV
1 Sequence: CGTTTGCCCATTAATCCAAAGAACTGCTGAGGCTTGGGTCTCCG 2 Detecting NK-1 R mRNA. Sequence #1: GGCTGCACGAACTGGTTAGACTCAGAGGTGTTGGTGGAGATGTTGGGG Sequence #2: TGGAGCTTTCTGTCATGGTCTTGGAGTTGCTGCGAGAGGAGCCGTTGG Sequence #3: TGACCACCTTGCGCTTGGCAGAGACTTGCTCGTGGTAGCGGTCAGAGG 3 Sequence: AACCGCGGGGACATGATGGCCT
3.3.3.3 Positive controls, ssDNA probes
1 Sequence: CGTTTGCCCATTAATCCAAAGAACTGCTGAGGCTTGGGTCTCCG
3.3.4 EIA
3.3.4.1 Rabbit tissue homogenization
Rabbit tendon tissue samples that previously had been weighed were mechanically homogenized in a lysis buffer (100 Mm Tris-HCI buffer pH 7.0 containing 1 M NaCl, 2% Bovine Serum Albumine [BSA], 4 mM EDTA, 0.2% Triton X-100, 0.02% sodium azide buffer) supplemented with protease
Probe Code Source Dilution Tissue Paper
β-actin1 (all species)
GD5000-OP Gene detection New Zealand
50 ng in 15 μL hybridization solution
Fixed/ unfixed
I, II, IV
Poly(dT) probe
GD4000-OP Gene detection New Zealand
50 ng in 15 μL hybridization solution
Unfixed I, II, IV

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
37
inhibitors pepstatin A (P4265, 0.1 g/mL), aprotinin (A1153, 5 g/mL), antipain
(A6191, 0.5 g/mL), benzamidin (B6506, 167 g/mL) and PMSF (P7626, 5.2
g/mL), all from Sigma. The weighed tissue samples and the lysis buffer were mixed in a 1:20 ratio before mechanically homogenized. The homogenization was performed using 2 periods with 1 minute duration for each period at 5,000 revolutions per minute (rpm), repeated twice with a Precellys 24 (Bertil technologies) homogenizer. The supernatant was moved to a new Eppendorf tube that was centrifuged 13,000 rcf at 4ºC for 15 minutes to eliminate cell debris. The supernatant was collected and stored at -80ºC until analysed. The entire procedure was carried out on ice to minimize autolysis.
3.3.4.2 Cell culture lysis
Cells were mechanically detached and centrifuged at 500 rcf for 3 minutes to get a cell pellet. The cell pellet was lysed in a lysis buffer (100 Mm Tris-HCI buffer pH 7.0 containing 1 M NaCl, 2% Bovine Serum Albumine (BSA), 4 mM EDTA, 0.2% Triton X-100, 0.02% sodium azide buffer) supplemented with protease
inhibitors pepstatin A (P4265, 0.1 g/mL), aprotinin (A1153, 5 g/mL), antipain
(A6191, 0.5 g/mL), benzamidin (B6506, 167 g/mL) and PMSF (P7626, 5.2 g/mL) at a volume of 150 μL lysis buffer per 1 million cells. After dissolving the cell pellet in the lysis buffer the lysate was incubated on ice for 30 minutes. Finally, the lysates were centrifuged at 13,000 rcf at 4ºC for 15 minutes to eliminate cell debris. The supernatant was collected and stored at -80ºC until analysed. The entire procedure was carried out on ice to minimize autolysis.
3.3.4.3 Experimental procedure
The concentration of SP was analysed using a commercially available enzyme immunoassay kit with a detection range (linear range) of 0.07-2.24 ng/mL (EK-061-05 Phoenix Pharmaceuticals, CA, USA).
All samples were diluted in a 1:5 ratio with the 1x assay buffer before added into each well in a 96-well plate. All wells were, by the manufacturer, pre-coated with the secondary antibody and the non-specific bindings sites were blocked. The standard, ranging from 0-25 ng/mL, was also added into assigned wells. The rehydrated primary antibody and the rehydrated biotinylated peptide were added before incubation at room temperature for 120 minutes with constant agitation. This was followed with washing, using 350 μL/well of the 1x assay buffer. The washing was repeated 4 times. After washes, SA-HRP (horseradish peroxidase), an enzyme found in horseradish (plant) that oxidizes the chromogenic substrate TMB into a coloured molecule, was added and incubated 60 minutes at room temperature with constant agitation. The washing procedure was repeated and followed by incubation with the TMB substrate solution in dark conditions for 60 minutes with constant agitation. Finally, adding 2 N HCI into each well stopped the reaction. The plate was read at 450 nm using a spectrophotometer.
All samples were read in duplicates and the mean was calculated. Values with a CV% >15 were excluded. Statistics were based on triplicate samples.

Ludvig J. Backman, 2013
38
3.3.5 Vascularity assessment
To evaluate the vascular structure in the rabbit tendon tissues, the sections were stained using the antibody against CD31, which stain endothelial cells positive.
The semi-quantitative grading of each section was done according to a modified Bonar scale in a blinded manner. The original Bonar scale (Cook et al., 2004) assessed vascularity, tenocyte appearance, ground substance and collagen, but for the modified Bonar scale used here, only vascularity was assessed using a o-3 grading, the lower grading means less vascularity and the higher grading means greater vascularity (Andersson et al., 2011). Each section was thoroughly examined and the intratendinous vascularity was assessed and graded.
Each part of the rabbit Achilles tendon sections (proximal portion, mid-portion, and distal portion) were graded and summarized i.e. each tendon was given a total grade with the range 0-9.
3.3.6 Inflammation assessment
Evaluation of inflammation, in the paratendinous tissue, was performed on the rabbit tissue sections that were stained with hematoxylin and eosin.
The grading of the paratendinous inflammation was done with a score of grades 0-3, 0 being no visible inflammatory cells, and 3 being marked inflammatory infiltration of the tissue. The grading was done of each part of the tendon tissue (proximal portion, mid-portion, and distal portion) and analysed separately.
3.3.7 Analysis of cell number, viability,
proliferation and metabolic activity
3.3.7.1 Counting method
The quantification of the number of tenocytes on the rabbit tissue sections was done on hematoxylin and eosin stained sections.
Two researchers (G.A, L.J.B) independently took three digital micrographs from each part of the tendon tissue proper (proximal portion, mid-portion, and distal portion) from each section using the 40x/0.75 microscope objective size resulting in an image of 283 x 213 μm2. A square grid was laid over the pictures to facilitate the tenocyte counting. A mean of the three pictures from the same section was calculated from both examiners and finally a mean from both examiners was calculated. The same was calculated for all three parts of the rabbit Achilles tendon (Andersson et al., 2011). Since Friedman statistical test, which take into account that the values calculated come from the same individual, showed no significant difference in the number of cells between the three parts of the tendon, a mean for the whole tendon was calculated and used in the comparison between animals and/or tendons.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
39
3.3.7.2 Bromodeoxyuridine (BrdU)
BrdU is a synthetic nucleotide that substitutes thymidine with uridine in the DNA at the phase of the cell cycle where DNA replication takes place, the s-phase. An antibody specific for BrdU can then be used to detect the incorporated BrdU. For these experiments all cells labelled with BrdU were counted.
Cells were plated, left to adhere and serum-starved in 1% serum overnight before they were treated.
BrdU labelling reagent, 10 μmol, was added to all chamber wells followed by a 120 minute re-incubation in the humidified incubator at 37°C in 5% CO2 in air for the BrdU to incorporate into dividing cells. The cells were then fixed in a fixative solution (50 mM glycine, 100 mL EtOH, pH2.0) for 20 minutes at -20ºC. After washing three times in the washing buffer the anti-BrdU monoclonal antibody was incubated for 30 minutes at 37ºC. Afterwards, cells were washed three times and thereafter incubated with the anti-mouse-Ig-AP for 30 minutes at 37ºC. After further washing, the substrate solution (4.3 mM NBT, 3.3 mM BCIP) diluted in substrate solution buffer (100 mM Tris HCI, 100 mM NaCl, 50 mM MgCl2, ph 9.5) was added and incubated for 20 minutes at room temperature in darkness for the detectable colour reaction to occur. Finally, before cells were mounted using Vectashield hard set with DAPI (Vector Laboratories; code: H-1500) one more wash was performed.
Proliferative cells are expressed as a ratio (percentage) of BrdU positive cells and all cells (DAPI staining). For each chamber well, three randomized pictures with 10x objective was taken and the mean ratio of BrdU and DAPI positive cells was used for statistic calculations.
Statistics were based on triplicate samples.
3.3.7.3 Crystal violet
Crystal violet is a triarylmethane dye used for a general protein stain. The basis of the assay is that cells on average will have the same total amount of protein and that the reading of a certain number of cells can be translated into a relative number of cells per well. It is important to keep in mind that many conditions can increase or decrease total proteins in cells.
Cells were plated, left to adhere overnight and thereafter serum-starved in 1% serum overnight. At the end of the experiment, cells were carefully washed in PBS two times to remove non-adherent cells and thereafter fixed in 1% glutaraldehyde for 30 minutes at room temperature. After three washes in PBS cells were either immediately stained or if a time-course was performed, cells were left in PBS at 4°C until all time-courses were collected. When all samples were collected they were stained in 0.1% crystal violet (sigma; code: C3886) diluted in MilliQ water for 30 minutes at room temperature in darkness. This was followed by three washes in an abundance of water to remove any excess of staining. Cells were then left to air-dry before an acidic solution of 30% methanol and 10% acetic acid, diluted in MilliQ water, was added to each well. The samples were left on a shaker for 10 minutes to ensure complete resolution

Ludvig J. Backman, 2013
40
before the dye was removed to a 96-well plate and the absorbance was read at 590 nm.
Each sample was read in duplicates and the mean was used. Statistics were based on triplicate samples.
The results of cell viability measured with crystal violet [III] are only presented at one time-point, 24h. However, several time points between 4-48h were tested, and all time points from 12h had the same trend but the clearest results were seen at 24h.
3.3.7.4 MTS assay
The colorimetric MTS assay was used to evaluate cell viability. This assay is composed of solutions of a tetrazolium compound [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt; MTS] and an electron coupling reagent (phenazine methosulfate) PMS which is bioreduced by cells into formazan. This reduction takes place only when reductase, an enzyme that catalyzes a reduction reaction, is active, i.e., it measures cell viability. It is important to keep in mind that many conditions can increase or decrease a cells metabolic activity.
Cells were left to adhere and were serum-starved in 1% serum overnight. Cells were treated and 120 minutes before the end of the experiment, MTS was added into each well and incubated in a humidified incubator at 37°C and 5% CO2 in air. The resulting formazan was measured at 490 nm using a spectrophotometer.
The viability is expressed as a value of absorbance in cells and medium minus that of just medium. Statistics were based on triplicate samples.
The results of cell viability measured with MTS [III] are only presented at one time-point, 48h. However, also several time points between 2-72h were tested with varying results, but the clearest results for the experimental design used in paper III were seen at 48h.
3.3.8 Analysis of cell death and apoptosis
3.3.8.1 Lactate dehydrogenase (LDH) assay
Lactate dehydrogenase (LDH) is an enzyme that exists in four distinct enzyme classes in the cells; two of which are cytochrome C dependent enzymes and two of which are NAD(P)-dependent. As a result of apoptosis or necrosis, when the cells loses its membrane integrity, the enzyme LDH is released into the culture medium. The basics of analyzing the released LDH is that it catalyzes the reduction of NAD+ to NADH and H+ and that diaphorase uses the newly-formed NADH and H+ to catalyzes the reduction of a tetrazolium salt (INT) to formazan which easily can be measured. The reaction cocktail, containing NAD+, diaphorase and INT, is added in the analysing and contributed by the manufacturer.
Cells were plated, left to adhere and serum-starved in 1% serum overnight before they were treated. At the end of the experiment, the medium (supernatant), possibly containing LDH, was collected. The collected

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
41
supernatant was centrifuged at 400 rcf for 5 minutes to remove any cell debris and cells and the new supernatant was subsequently removed, and, if not immediately used, frozen in -80°C until used.
To analyse the concentration of LDH in the supernatant, 100 μL of freshly mixed reaction mixture, containing NAD+, diaphorase and INT, were mixed with 50 μL of supernatant in a 96-well plate and incubated at room temperature on constant agitation for 30 minutes protected from light. The absorbance was read at 490 nm using a spectrophotometer.
As positive control, cells were lysed in 1% triton to achieve membrane breakage and release of LDH. This was done to confirm successful methodology and not presented in the results.
Each sample was read in duplicates and the mean was used. Statistics were based on triplicate samples.
3.3.8.2 TUNEL-assay
A cell undergoing apoptosis is characterized by discontinuity in the double stranded DNA, i.e. the bond between the adjacent nucleotides in the double stranded DNA is damaged. The basics of the TUNEL-assay are that the terminal deoxynucleotidyl transferase, recombinant enzyme (rTdT), can detect the damage in the double-stranded DNA and incorporate fluorescein-12-dUTP, which can be visualized.
Cells were plated, left to adhere and serum-starve in 1% serum overnight before they were treated. At the end of the experiment the cells were fixed for 5 minutes in 2% paraformaldehyde in 0.1 M phosphate buffer (pH7.4) before washing in PBS twice for 5 minutes each. To accomplish membrane permeabilization the cells were incubated 5 minutes in 0.2% triton x-100 diluted in PBS. After washing, the cells were incubated in equilibration buffer for 10 minutes at room temperature and thereafter incubated for 60 minutes at 37°C with the nucleotide and rTdT mix. To stop the reaction the 2xSSC solution was added and incubated for additional 15 minutes. Finally, cells were washed and mounted in Vectashield Hard Set Medium with DAPI (Vector Laboratories; code: H-1500).
As negative control, the rTdT was left out, and as positive control, cells were treated with DNase before incubation with the equilibration buffer. This was done to confirm successful methodology and was not presented in the results.
The results of apoptosis measured with TUNEL assay [V] are only presented at one time point, 24h. However, also the time point 12h was tested, showing the same trend, but the clearest results were seen at 24h.
3.3.9 Western blot
3.3.9.1 Cell lysis
Cells were collected and lysed in RIPA lysis buffer (150 mM Sodium chloride, 1% triton, 0.5% Sodium deoxycholate, 0.1% Sodium Dodecyl Sulphate [SDS], 50 mM Tris, pH 8.0) supplemented with protease inhibitor at a concentration of 1:200, all from Sigma. Cells were mixed well with the lysis buffer to ensure

Ludvig J. Backman, 2013
42
complete dissolving of the cell pellet and thereafter incubated on ice for 30 minutes with mixing on regular basis of each sample. To abolish cell debris, the lysate was centrifuged at 11,300 rcf for 5 minutes. The supernatant was collected and a small volume was used to determine total protein concentration using Protein Assay Dye Reagent Concentrate (Bio-Rad, Hercules, CA, USA; code: 500-0006) with Bovine Serum Albumin (BSA, Sigma; code: A9647) as a standard. The remaining supernatant was immediately diluted in Laemmli Sample Buffer (Bio-Rad; code: 161-0737), supplemented with beta-mercaptoethanol and heat-denatured at 95°C for 5 minutes. The function of beta-mercaptoethanol is mainly to reduce the disulphide bonds and the heat-denaturation to disrupt both the secondary and tertiary structures (not strong enough to break peptide bonds between the amino acid subunits and therefore the primary structure will remain). After the samples had been heat-denatured they were mixed well and either immediately used for Western blot or frozen for later use. The frozen samples were thawed at 37°C for 10 minutes and mixed before used.
3.3.9.2 Procedure
Samples of similar total protein concentration, confirmed with the protein assay, were loaded onto sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) separating gel to separate proteins after their molecular weight, in kilo Dalton (kDa). The pre-made TGX gels used were run at 160V for 45 minutes in the running buffer. Details of all solutions mentioned in the explanation of the procedure are described in table 3.3.9.6. The concentration of the TGX-gel used was based on the analysed proteins expected kDa. As a rule of thumb, the smaller size of the protein of interest, the higher percentage of the gel, and the reversed for bigger kDa proteins. This was followed by transferring the proteins from the gel to a Polyvinylidene Fluoride Transfer Membrane (PVDF, Santa Cruz; code: sc-3723), which was done at 100V for 60 minutes in the transfer buffer. After the transfer was done the membrane was stained with Poncea red (0.1% poncea red, 1% acetic acid and MilliQ water), a reversible protein stain to ensure successful transfer. To remove the Poncea red staining washing in MilliQ water and/or tris buffered saline with Tween 20 (TBS-T) was done before the membrane was blocked in TBS-T supplemented with 5% non-fat milk at room temperature for 60 minutes at constant agitation. Subsequently, the primary antibody, diluted in either TBS-T supplemented with 5% BSA or 5% non-fat milk, was incubated overnight at 4°C on constant agitation.
The second day the membrane was washed with TBS-T for 6x5 minutes before the secondary antibody, an antibody conjugated to the horseradish peroxidase (HRP) diluted in TBS-T supplemented with 5% non-fat milk, was incubated at room temperature at constant agitation for 6o minutes. After additional washing 6x5 minutes in TBS-T the membrane was exposed to a chemiluminescence substrate for 5 minutes. The chemiluminescence consist of peroxidase and the substrate which, when oxidased by HRP using perioxidase as the oxidising agent, produces light which can be detected on high performance chemiluminescence film (GE healthcare; code: 28-9068-38).

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
43
To confirm equal loading the membrane was stripped using Western blot stripping buffer (Thermo Scientific 21059). This was done at 37°C for 25 minutes under constant agitation. Following the stripping procedure the membrane was thoroughly washed in TBS-T 6x5 minutes and then blocked before re-stained with the house keeping antibody, β-actin.
The results of NK-1 R expression decrease after submission of SP [III] are only presented at one time point, 4h. However, several time points between 2-48h were tested and the clearest difference was seen at 4h. The difference steadily decreased and became more similar to untreated cells with time. Also, the results of c-PARP and c-caspase-3 [V] are only presented at one time-point, 12h. However, both 6h and 24h were also used and all time-points showed the same trend. The results of P-Akt [V] and P-ERK1/2 [III, IV] were presented in a time dependent manner.
3.3.9.3 Primary antibodies used for Western blot
Antigen Code Source Type Dilution Paper
P-p44/42 MAPK (ERK1/2)
4370 Cell signal, Danvers, MA, USA
Rabbit 1:2000 III, IV
NK-1 R sc-5220 Santa Cruz, CA, USA. Goat 1:1000 III
β-actin 4967 Cell signal, Danvers, MA, USA
Rabbit 1:2000 III, IV, V
TH P40101 Pel-Freez, Rogers, AR, USA
Rabbit 1:2000 IV
c-PARP 9541 Cell signal, Danvers, MA, USA
Rabbit 1:2000 V
c-Caspase-3 9664 Cell signal, Danvers, MA, USA
Rabbit 1:1000 V
P-Akt 9271 Cell signal, Danvers, MA, USA
Rabbit 1:2000 V
3.3.9.4 Negative and positive controls used for Western blot
Extract Treated Code Source Paper
Jurkat Apoptosis Cell Lysate
Untreated (negative control) Etoposide treated (positive control)
2043 Cell signal, Danvers, MA, USA V
Akt Control Cell Extract
LY294002 treated (negative control) Calyculin A treated (positive control)
9273 Cell signal, Danvers, MA, USA V

Ludvig J. Backman, 2013
44
3.3.9.5 Secondary antibodies used for Western blot
Secondary ab Code Source Dilution Paper
HRP-conjugated donkey anti-goat
sc-2020 Santa Cruz, CA, USA 1:5000 III
HRP-conjugated goat anti-rabbit
7074 Cell signal, Danvers, MA, USA
1:2000 III, IV, V
3.3.9.6 Western blot solutions
Solution Ingredients
TBS-T (1.5 L) 1.82 g Tris-Base. 8.76 g NaCl. 1,5ml Tween-20 pH 7.4
Running buffer (1.0 L) 3.03 g Tris-base. 14.4 g glycine. 1 g SDS pH 8.3
Transfer buffer (1.0 L) 3.03 g Tris-base. 14.4 g glycine. 200 mL Methanol
3.3.10 Real-time quantitative polymerase chain
reaction (RT-qPCR)
3.3.10.1 RNA extraction
RNA was extracted using the RNeasy Mini Kit (Qiagen, Hilden, Germany; code: 74106) and the manufacturer’s instructions were followed.
Cell cultures were washed and lysed in the RLT buffer supplemented with 1% β-mercaptoethanol in the recommended volume of buffer. In the lysate, 1 volume of 70% ethanol was added and thoroughly mixed before transferred to the RNeasy spin columns and centrifuged 15 seconds at 8,000 rcf. In some cases a QIAshredder was used to ensure a good lysis in the RLT buffer before adding samples into RNeasy spin columns. The first washing buffer, called RW1 by the manufacturer, was added followed by a centrifugation of 15 seconds at 8,000 rcf. An additional washing in the second washing buffer, called RPE, was added and again centrifuged for 15 seconds at 8,000 rcf. Finally, one last wash in the RPE buffer was done followed by centrifugation at 8,000 rcf for 2 minutes and immediately additional centrifuged at 13,400 rcf for 1 minute. To collect the RNA, RNase free water was added and collected in a new tube by centrifugation at 13,400 rcf for 1 minute.
3.3.10.2 Reverse transcription
Reverse transcription was performed using the High-capacity cDNA Reverse Transcription kit (Applied Biosystems [ABI], Warrington, Cheshire, UK; code: 4368813). RNA was diluted with 2 μL of the RT buffer (keeps the enzyme stable), 2 μL of the RT random primers (bind to mRNA in order for reverse transcription to occur), 1 μL of the multiscribe Reverse transcriptase (RNA dependent-DNA polymerase), 0.8 μL of the dNTP mix (used for cDNA synthesis) and finally nuclease free water to get the sufficient volume of 20 μL for each reaction. The cycler conditions consisted of an initial step of 10 minutes at 25°C followed by 120 minutes at 37°C, 5 minutes at 85°C, and finally

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
45
refrigerated to 4°C. The complementary DNA (cDNA) was either used immediately or stored at -20°C until used.
The optimal mRNA concentration for reversed transcriptase in cultured human tenocytes was tested for and proven to be 1,000 ng/reaction tube.
3.3.10.3 RT-qPCR
Samples of equal cDNA concentration were diluted with the 2x TaqMan Fast Advanced master mix, the 20x TaqMan MGB probe and nuclease free water. All samples were amplified at the following settings; 95°C for 20 seconds in the first cycle, then followed by 2 step cycles with the denaturation at 95°C for 1 second (first step), and annealing, extension at 60°C for 20 seconds (second step). During the extension the reporter is separated from the quencher on the probe thus resulting in fluorescent light proportional to the product.
3.3.10.4 Probes used for RT-qPCR
Probe name Code Source Paper
TAC11 Hs00243225 Applied Biosystems, Warrington, Cheshire, UK
III
TACR12 Hs00185530 Applied Biosystems, Warrington, Cheshire, UK
III
18s (house-keeping gene)
4308329 Applied Biosystems, Warrington, Cheshire, UK
III
1 Detects mRNA coding for SP 2 Detects mRNA coding for NK-1 R
3.3.11 Cell density used for different
experiments in vitro
Plate formate
Code Source Density (cells/well)
Experiment Paper
96-well plate
3300 Corning, Tewksbury, MA, USA
5,000 MTS III
Pre-coated collagen I 6-well plates
BF-3001C Bioflex, Hillsborough, NC, USA
175,000 2D strain III
8-well plate
354118 BD Bioscience /BD Falcon, Bedford, MA, USA
15,000 Immunocytochemistry, TUNEL-assay, BrdU
III, IV, V
6-well plate
83.1839 Sarstedt, Newton, NC, USA
150,000 Crystal violet, LDH, Western blot
III, IV, V
4-well plate
354104 BD Bioscience /BD Falcon, Bedford, MA, USA
35,000 Hemotoxylin and Eosin
V
12-well plate
734-2324 VWR, Stockholm, Sweden
75,000 LDH V

Ludvig J. Backman, 2013
46
3.4 Statistics
All statistical calculations were done using computer software (PASW statistics 18.0.0; SPSS Inc., Chicago, IL, USA). Statistical significance was predetermined at p<0.05.
3.4.1 In vivo studies
For the in vivo model, it was not assumed that the data was normally distributed due to the low number of rabbits used in each experimental group, and therefore non-parametric methods, i.e. methods using ranks to replace original data, were used in all statistical calculations.
The results and distribution of data in paper I, II and IV (the in vivo results) are presented in medians and error bars showing the inter-quartile range (IQR), which is a measure of variability based on dividing a data set into quartiles. IQR is the appropriate measure of the variability in non-parametric data (McCluskey and Lalkhen, 2007). The bars in the tables illustrate the distance between quartile 1 (Q1) and quartile 3 (Q3) which represents the middle 50% of the observations, thus resulting in 25% represented below Q1 and the remaining 25% above Q3 (McCluskey and Lalkhen, 2007). The median is Q2.
3.4.1.1 Friedman test
This is a method to detect differences across multiple test attempts, similar to the parametric repeated measure ANOVA. This method takes into account that the values calculated come from the same individual and therefore this test was applied in paper II, IV when the different parts (distal part close to the OTJ [A], mid-portion part [B], and the proximal part, MTJ, [C]) of the Achilles tendon were compared in the same animal.
3.4.1.2 Kruskal-Wallis test and Mann-Whitney U test
The Kruskal-Wallis test was used for comparing three or more independent samples. The test does, however, not identify between which groups the differences occur. For that purpose, pair-wise Mann-Whitney U test was used, which analyses the specific sample pairs for statistically significant differences. The Mann-Whitney U test had to be complemented with Bonferroni correction, which counteracts the problem of multiple comparisons.
These statistical methods were applied in paper I, II and IV.
3.4.1.3 Wilcoxon signed rank test
This test was used when comparing two related or dependent samples, i.e. it differs from Mann-Whitney U test, which tests independent sample pairs.
This statistical method was applied in paper I, II and IV to calculate if there was a statistically significant difference between the two legs of the same rabbit.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
47
3.4.2 In vitro studies
For the in vitro model, normal distribution of the data was assumed and therefore parametric methods, i.e. using original data, were used in all statistical tests.
The results of the in vitro experiments are presented as means and the distribution of data is indicated with either standard error of the mean, which is an estimate of how close sample means are to the population mean, used in paper III (Fig. III:7), or standard deviation, which is the degree to which data within the sample differ from the sample mean, used in paper III, IV and V.
3.4.2.1 Independent samples t-test
This test was applied to determine if there was a statistically significant difference between the mean of two independent samples. This test was applied in paper III when only two samples were compared.
3.4.2.2 One-way ANOVA with Bonferroni post hoc test
The one-way ANOVA test was used to compare the mean of three or more samples, and was complemented with a Bonferroni post hoc test to identify differences between sample pairs as well as adjust for multiple comparisons. These tests were applied in paper III, IV, and V.

Ludvig J. Backman, 2013
48
4. RESULTS
4.1 Phenotyping of cells studied in vitro (tenocytes)
The vast majority, approximately >95%, of primary human tendon cells in culture displayed an elongated appearance and immunopositive reactions for tenomodulin and vimentin (Fig. III:3a, and Fig. III:3b, respectively). Additional control studies by us on these cells confirmed that they also express scleraxis mRNA and display immunoreactions for scleraxis (figure 12c; results reported by us in Fong et al., 2013, although the controls were done continuously on the model). Furthermore, Western blot for scleraxis showed an immunopositive band at the expected kDA (Fong et al., 2013) (inset in figure 12c). Finally, the cells also expressed a high level of collagen I in relation to collagen III (figure 12a-b) and showed no positive reactions for alpha-smooth muscle actin (Fong et al., 2013) (figure 12d). Based on these results, confirming the presence of tenocyte phenotype, we called the tendon derived cells in culture tenocytes.
Fig. 12 Immunoflourescence for collagen I (a) shows that this protein is abundantly expressed in the primary human tendon cell cultures, whereas staining for collagen III (b) shows very scarce reactions. Almost all cells were immunopositive for scleraxis (c). Western blot for scleraxis (SCX) shows a band at 22 kDa, corresponding to the known molecular weight of SCX (inset in c). All cells were immunonegative for alpha-smooth muscle actin (d). Cell nuclei counter-stained in blue using DAPI.
Rearranged original photos reprinted from the article Fong G, Backman LJ, Andersson G, Scott A, Danielson P: Human tenocytes are stimulated to proliferate by acetylcholine through an EGFR signalling pathway. Cell and Tissue Research 2013;351(3):465-475.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
49
4.1.1 Complementary results: Additional
phenotyping of cells
In the results of paper III it was mentioned that a few cells occasionally showed a more plump appearance and also that some cells sporadically exhibited extended dendritic-like processes. Additional stainings, in an attempt to confirm the cell type of these particular cells, have later shown that the cells with a plump appearance (approximately <5% of the cells) are immunopositive for SSEA-4, which is a tendon stem cell marker (Zhang and Wang, 2010) (figure 13a). The rare cells with extended dendritic-like processes have been found to be immunopositive for PGP9.5, which is a marker for neurons (figure 13b). No cells had positive immunoreactions for CD31, a marker for endothelial cells.
Fig. 13 A cell from the primary human tendon cell cultures immunopositive for the stem cell marker SSEA-4 is seen in a. In b, a cell with extended dendritic-like processes is seen to be immunopositive for the nerve cell marker PGP9.5, whereas other cells are seen to be immunonegative in this regard (nuclei counter-stained in blue using DAPI).
4.2 Presence of signal substances and
their receptors
4.2.1 In vivo
SP mRNA was extensively detected in several of the blood vessels located in the paratendinous tissue. These reactions were mainly seen in the endothelial cells of the blood vessels (Fig. I:3a). Occasional reactions for SP mRNA were detected in the tenocytes in the tendon tissue proper. On the level of protein, reactions for SP were also extensively seen in the blood vessels located in the paratendinous tissue, however in this case both in the endothelial and smooth muscle cells of the blood vessels (Fig. I:2). Some fine nerve fascicles in the paratendinous tissue also exhibited immunoreactions for SP (Fig. I:2 inset). Quantification of the SP peptide, using EIA assay, showed a concentration of 175 pg/mg in the unexercised control group (Fig. I:1).
NK-1 R mRNA was detected in the walls of the blood vessels, both in the paratendinous tissue and the tendon tissue proper, predominantly in the endothelium but also in the smooth muscle cells (Fig. I:4c, and Fig. II:1b). The

Ludvig J. Backman, 2013
50
tenocytes in the tendon tissue proper also expressed NK-1 R mRNA reactions. On the protein level, reactions for NK-1 R were seen at the same locations as the expression of NK-1 R mRNA. The nerves in the paratendinous tissue showed positive immunoreactions for NK-1 R, which was not the case for NK-1 R mRNA.
TH mRNA reactions were found in the tenocytes in the tendon tissue proper (Fig. IV:1b, inset). On the level of protein, TH immunoreactions were seen in the tenocytes in the tendon tissue proper, but also in the nerves seen in the paratendinous tissue (Fig. IV:1a). Immunoreactions for the receptor α2A AR were displayed in the tenocytes in the tendon tissue proper (Fig. IV:1b).
For an overview of the immunoreactions, see figure 14.
Fig. 14 Both SP and NK-1 R were expressed in blood vessels, nerves and tenocytes of the rabbit Achilles tendons. Expression of both TH and α2A AR were seen in the tenocytes, but only TH immunoreactions were seen in the nerves.
4.2.2 In vitro
The cultured tenocytes expressed SP mRNA (Fig. III:7a), and on the level of protein, frequent immunoreactions were observed (Fig. III: 5a). Quantification of SP, using EIA assay, showed a concentration of 170 pg/2x106 tenocytes.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
51
NK-1 R mRNA was expressed by the cultured tenocytes (Fig. III:7b), and on the level of protein immunoreactions were frequently observed (Fig. III:5b). The detection of NK-1 R protein was also confirmed using Western blot, which clearly showed two bands, suggested to be the full-length glycosylated and truncated unglycosylated receptor, at molecular weights of 80 kDa and 37 kDa, respectively (Fig. III:6).
Clear immunoreactions for TH were displayed in the tenocytes (Fig. IV:4a), which was also confirmed using Western blot, with detection of TH at the expected molecular weight of 60 kDA (Fig. IV:4c). The tenocytes also expressed clear immunoreactions for the α2A AR (Fig. IV:4b).
For an overview, see figure 15.
4.3 Changes in SP and NK-1 R
expression in response to load
4.3.1 In vivo
Biopsies from the mid-portion of the rabbit Achilles tendon showed that 1 week of exercise resulted in a significant increase of endogenously produced SP as compared to the control group (i.e. rabbits not having exercised). The significant increase of SP production that was seen after 1 week of exercise was also seen in the 3 weeks and 6 weeks exercise groups, in comparison to the control group (Fig. I:1) (figure 16). Unexpectedly, the level of SP did not differ
Fig. 15 The tenocytes in culture expressed SP, NK-1 R, TH, and α2A AR.
Fig. 16 Endogenously produced SP in the Achilles tendon was increased in response to mechanical loading.

Ludvig J. Backman, 2013
52
significantly when comparing the exercised and unexercised leg of the same rabbit in any of the groups. The expression of NK-1 R was not quantified. However, subjective evaluation of immunoreactions concluded no obvious difference in response to loading.
Interestingly, the rabbit model has previously been shown to result in tendinosis-like changes, i.e. hypercellularity and angiogenesis, only after a minimum of 3 weeks of exercise (Andersson et al., 2011), implying that the increased production of intratendinous SP, seen here after only 1 week of exercise, precedes the onset of tendinosis-like changes.
4.3.2 In vitro
In the cultured tenocytes, a significant increase of SP mRNA was seen as a response to the 2D loading protocol as compared to the unloaded controls (Fig. III:7a), and the reverse was seen for NK-1 R mRNA in response to load, i.e. a significant decrease of mRNA expression as compared to in the unloaded controls (Fig. III:7b). See figure 17 for an overview.
4.4 Effects of exogenously added
clonidine
4.4.1 In vivo
Paratendinous injections of clonidine (an α2 AR agonist) in the connective tissue ventral to the rabbit Achilles tendon, in combination with 1 week of exercise, resulted in a significant increase in the number of tenocytes in the tendon tissue proper when comparing cell density to that of the exercise control group, i.e. the group undergoing only 1 week of exercise without injections (figure 18). Exercise in combination with BRL-injections (an α2A AR antagonist) resulted in a significant reduction of the number of tenocytes as compared to after clonidine injections. Injections of NaCl in combination with exercise (NaCl-group) also resulted in a significant increase in the number of tenocytes as compared to the exercise control group. However, when comparing the NaCl-group with the BRL-injection group no significant difference was seen (Fig. IV: 2).
Fig. 17 Loading of tenocytes in culture resulted in increased SP expression and decreased NK-1 R expression.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
53
4.4.2 In vitro
Clonidine stimulation of tenocytes resulted in a significant increase of cell viability as compared to untreated cells, and the effect of clonidine stimulation was abolished when the cells were pre-incubated with BRL (Fig. IV:5). Results from the proliferative assay, BrdU, showed the same result as the cell viability assay, i.e. clonidine stimulation had a proliferative effect through an α2A AR specific pathway (Fig. IV:6). See figure 19.
Western blot analysis showed that clonidine stimulation of tenocytes resulted in ERK1/2 phosphorylation, peaking after 10 minutes of stimulation. Pre-incubation with BRL successfully blocked the effect of clonidine on phosphorylation of ERK1/2 (Fig. IV:7).
4.5 Effects of exogenously added SP
4.5.1 In vivo
SP-injections in combination with 1 week of exercise resulted in a significant increase in the number of tenocytes as compared to the unexercised control group and the 1 week exercise control group (none of these control groups receiving injections) (figure 20). The injection exercise control group (the NaCl-group) also showed a significant increase in the number of cells as compared to the unexercised control group (Fig. II:3). The number of tenocytes did not differ significantly when comparing the exercise SP-injected Achilles tendon with the unexercised SP-injected Achilles tendon of the same rabbit (Fig. II:4).
Fig. 19 Clonidine stimulated proliferation of tenocytes in culture.
Fig. 18 Paratendinous injections of clonidine, in combination with 1 week of exercise, resulted in hypercellularity.

Ludvig J. Backman, 2013
54
Regarding vascularity, the SP-injected group resulted in a significantly higher grade of vascularity as compared to the unexercised control (Fig. II:5) (figure 20).
Finally, the proximal part of the tendon, i.e. myotendinous junction, displayed a significantly higher grade of paratendinous inflammation in the SP-injected group as compared to the NaCl-injected group (Fig. II:6).
4.5.2 In vitro
4.5.2.1 Proliferation
SP stimulation of tenocytes resulted in a significant increase in metabolic activity and cell viability as compared to controls (Fig. III:9, and III:10). The effect of SP regarding cell viability was effectively blocked when cells were pre-incubated with the NK-1 R inhibitor (Fig. III:10). Results from the proliferative assay, BrdU, showed the same result as the cell viability assay did, i.e. SP stimulation having a proliferative effect through a NK-1 R specific pathway (Fig. III:12). See figure 21.
Western blot analysis showed that SP stimulation of tenocytes resulted in
ERK1/2 phosphorylation in a biphasic manner with peaks at 5 minutes and 15 minutes after stimulation. Pre-incubation with the NK-1 R blocker effectively abolished the effect of SP on phosphorylation of ERK1/2 (Fig. III:13).
4.5.2.1.1 Complementary results: Proliferation
The result of paper III showed that SP, at a concentration of 10-7 M, had a significant proliferative effect on tenocytes. This effect of SP has been further studied, showing, by the use of SP at concentrations of 10-7, 10-8, and 10-9 M, that the effect is dos-dependent (see figure 22 for detailed results).
Fig. 20 Paratendinous injections of SP, in combination with 1 week of exercise, resulted in hypercellularity as well as angiogenesis.
Fig. 21 SP stimulated proliferation of tenocytes in culture.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
55
Fig. 22 As compared to the control, SP at a concentration of 10-7 M had a significant proliferative effect on human tendon cells. It was also shown that the effect of 10-7 M of SP had significantly superior effect as compared to 10-8 M SP and 10-9 M SP. Y-axis showing fraction of proliferating (BrdU positive) cells. Error bars show standard deviations. *p< 0.05, **p < 0.01. (One-way ANOVA; Bonferroni post hoc test).
4.5.2.2 Anti-apoptosis
Exposing cells to Anti-Fas resulted in a statistically significant dose-dependent response in cell death (as measured with the LDH assay), and there was also a clear increase over time in the response to Anti-Fas (Fig V:3), confirming Anti-Fas treatment to be a good model to analyse cell death in tenocytes. The induced cell death with Anti-Fas was dose-dependently reduced when cells were pre-incubated with SP at different concentrations as confirmed with both cell viability and cell death assays (Fig. V:4, and V:5). Furthermore, it was shown that this effect of SP was specifically blocked, in a dose-dependent manner, with the NK-1 R inhibitor (Fig. V:6, and V:7). Neither cell death nor cell viability distinguish between necrosis and apoptosis, therefore a TUNEL-assay, which is an apoptosis specific assay, was performed and the same results were seen as for the cell death and cell viability assays, i.e. Anti-Fas induced apoptosis which was reduced with SP in a NK-1 R specific manner (Fig. V:8). Also apoptotic specific markers, such as caspase-3 and PARP, confirmed SP to have an anti-apoptotic effect through a NK-1 R specific pathway after Anti-Fas induced cell death (Fig V:9).
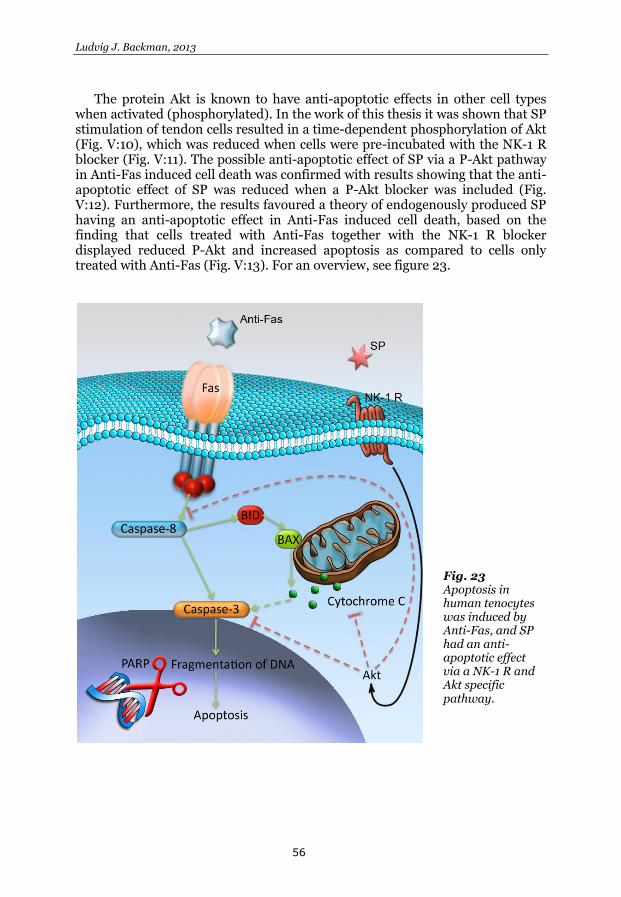
Ludvig J. Backman, 2013
56
The protein Akt is known to have anti-apoptotic effects in other cell types when activated (phosphorylated). In the work of this thesis it was shown that SP stimulation of tendon cells resulted in a time-dependent phosphorylation of Akt (Fig. V:10), which was reduced when cells were pre-incubated with the NK-1 R blocker (Fig. V:11). The possible anti-apoptotic effect of SP via a P-Akt pathway in Anti-Fas induced cell death was confirmed with results showing that the anti-apoptotic effect of SP was reduced when a P-Akt blocker was included (Fig. V:12). Furthermore, the results favoured a theory of endogenously produced SP having an anti-apoptotic effect in Anti-Fas induced cell death, based on the finding that cells treated with Anti-Fas together with the NK-1 R blocker displayed reduced P-Akt and increased apoptosis as compared to cells only treated with Anti-Fas (Fig. V:13). For an overview, see figure 23.
Fig. 23 Apoptosis in human tenocytes was induced by Anti-Fas, and SP had an anti-apoptotic effect via a NK-1 R and Akt specific pathway.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
57
5. DISCUSSION
5.1 Introductory remarks
There is a lack of knowledge regarding the mechanisms explaining hypercellularity and angiogenesis in tendinosis tissue. However, lately a biochemical explanation model, i.e. a model suggesting that biochemical mediators produced by tenocytes are involved in the pathogenesis, has been thoroughly discussed (Danielson, 2009). Due to the current lack of experimental data supporting this biochemical theory, the overall aim of this thesis was to experimentally study the importance of NK-1 R and α2A AR stimulation of tenocytes, and its potential role in tendinosis development. In the following chapter the results from the studies will be discussed and integrated in a wider context.
5.2 Methodological aspects of the experimental models
5.2.1 Animal model of Achilles tendinosis
As previously introduced, there are different types of experimental in vivo models to study tendinosis, all with different advantages and disadvantages. The in vivo model used in this thesis has some aspects that have to be considered when interpreting the results.
5.2.1.1 Electrical stimulation
In the original experimental model, upon which our refined model is based, an impulse train of electrical stimulation was applied to the rabbits calf muscle in the concentric movement of the ankle joint to facilitate the muscle contraction and mirror the tension in the tendon during running in man (Backman et al., 1990). In our re-established model the impulse train was not performed, instead only one twitch of stimulation was applied (Andersson et al., 2011). The relevance of this is unknown, but it is likely that the muscle recruitment is reduced and therefore likewise the tension in the tendon, making this re-established model in that sense perhaps less physiological than the original model. On the other hand, the set-up in the re-established model results in tendinosis, which was the wanted outcome, and since less electrical stimulation was given its possible negative effects on the rabbit and on the results were reduced (cf. next paragraph).
Electrical stimulation is known to affect nerves to cause a neurogenic response, including efferent release of SP and other factors (White and Helme, 1985; Burssens et al., 2005b). This might affect the analysed outcome in our

Ludvig J. Backman, 2013
58
model, and possibly explain the fact that hypercellularity and angiogenesis was observed as a response to the electrical stimulation protocol (Andersson et al., 2011), as known effects of SP are hypercellularity and angiogenesis (Burssens et al., 2005a; Carlsson et al., 2011). However, the fact that the contralateral side did not differ in either of the morphological changes or in the level of SP as compared to the exercised side contradicts that the electrical stimulation alone, via release of SP, would explain the morphological changes. Nevertheless, one could speculate on central mechanisms causing the elevated levels of SP in the contralateral leg and subsequently resulting in the morphological changes. All the same, should endogenous SP be the reason for observed effects, this would only strengthen our hypothesis further.
5.2.1.2 Terminology – exercised and non-exercised leg
The terminology, “exercised” and “non-exercised leg”, used in the studies on the rabbit model is perhaps not the most appropriate terminology, since we cannot ensure that the contralateral leg is truly “non-exercised”. It might have been more accurate to simply use the terminology ipsilateral, for the experimental leg, and contralateral, for the non-experimental leg. The first reason why this should be considered is that the rabbits might have had a slight amended movement as a result of the exercise protocol (although no obvious limping was observed), which in turn could result in overloading of the contralateral leg. The second reason is the fact that EMG activity can be recorded from a non-exercised limb during exercise of the other leg (Devine et al., 1981). It is possible that this also occurred in the experimental model of our studies, i.e. the contralateral leg also becoming activated, although this was not evaluated with EMG in these studies.
5.2.1.3 Anaesthesia and analgesia
Since the effect of the anaesthesia and the analgesia of the rabbits are not controlled for, it cannot be ruled out that these factors caused a general effect on the tendons in the rabbit. One possible explanation for such a hypothesis is the usage of the opioid agonist buprenorphine for analgesia. Since opioids are known to inhibit SP release from peripheral afferent neurons (Brodin et al., 1983) it is possible that buprenorphine results in a systemic effect in the rabbit. The possible impact of both the anaesthesia and analgesia could be controlled for in the future by collecting and analysing other tendons, for instance from the upper extremities. However, again, should endogenous SP be of importance for observed effects, this would only goes to further strengthen our overall hypothesis. It is possible that without the buprenorphine, the changes in SP expression and effects would have been even more distinct.
5.2.1.4 Injections
A puzzling result in the injection experiments was that the NaCl-injections, which served as an injection control, gave the same results, regarding hypercellularity, as SP and clonidine did. This was an unexpected result that has

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
59
to be considered in the interpretation. One could speculate if the increased pressure following the injections might explain the hypercellularity. However, the results of the BRL-injections (α2A AR inhibitor) directly contradict this possible explanation, since it resulted in significantly less cell counts than after injections with clonidine and no difference in cell number compared to uninjected controls. In other words, only the increased pressure following injections cannot alone explain the effects of hypercellularity seen after injections, since the same volume was injected in all cases.
The fact that it is known that NaCl-injection can result in elevated levels of endogenous SP (Chen et al., 2007), makes it tempting to explain the results of hypercellularity after NaCl-injections with an increase in the level of endogenously produced SP.
Since it is known that the tendon tissue has an endogenous production of SP and catecholamines, it might be that further increasing the concentrations of the substances in question by injections will not increase their specific effect. Thus, unspecific effects, for instance an effect of increased pressure (cf. above), might be what is predominantly seen. On the other hand, by blocking the receptor the effect is abolished, as evident by BRL-injection, indicating that a down-regulation of a biochemical system could be the approach to use to investigate its effects in vivo in future studies.
5.2.1.5 Anatomical differences – rabbit vs. human
There are some differences regarding the anatomy of the triceps surae muscle (gastrocnemius and soleus) in the rabbit as compared to in the human. The soleus muscle and the contribution to the Achilles tendon of this muscle is proportionally smaller in rabbits, the medial and lateral gastrocnemius tendon fibres fuse together more distally in rabbits, and finally rabbits have their flexor digitorum superficial tendon just dorsal to the Achilles tendon and within the Achilles tendon paratenon (Cummins et al., 1946; Doherty et al., 2006). The relevance of these anatomical differences in the use of rabbits as a model system is not obvious, but it is an advantage that both rabbit and human Achilles tendon-/triceps surae muscle-complex spans over two joints and that the rabbit has a fairly large Achilles tendon.
Regarding the source of SP production, as evident by the expression of SP mRNA, it differs between rabbit and human. In humans, an important source of locally produced SP in the tendon seems to be the tenocytes (Andersson et al., 2008) [III], whereas in rabbits it seems like the source mainly is the blood vessels, more specifically the endothelium [I].
5.2.1.6 Suggested future complementary
experiments
For future experiments using the experimental rabbit model, more controls are needed to interpret the results. For example controls for the electrical stimulation, having rabbits stimulated without being in the kicking apparatus, and controls for the anaesthesia (i.e. anesthetized but not exercised animals) would have been desirable. Furthermore, the possible impact of

Ludvig J. Backman, 2013
60
anaesthesia/analgesia could also be controlled for by collecting and analysing other tendons, for instance from the upper extremities. In addition, to be able to draw more solid conclusions from the effects of SP, a NK-1 R blocker injection group should be included, both alone and in combination with SP injections.
5.2.2 Primary human tendon cell culture model
Culturing isolated cells in an artificial environment has advantages as it makes it possible to analyse the effect of specific substances under controlled circumstances on the specific cell type studied. However, the generalization of the results from such environments has to be made carefully. Important structures in the tendon, such as collagen, blood flow, pressure, nerves, and other surrounding factors that are present in vivo, are ruled out in the in vitro model. To mirror the in vivo environment, some in vitro experiments in the studies of this thesis were performed in a loading environment. Also, a great advantage by culturing human tendon cells is that the results from the experimental rabbit model can be controlled for on human tissue, without being done in human patients. The use of a two-model system, both the experimental rabbit model and the human cell cultures, is a strength of the studies.
5.2.2.1 What cells are we culturing?
There is currently no single established specific tenocyte marker only detecting tenocytes (Riley, 2008). However, tenomodulin is considered a fairly specific marker, which is a type II transmembrane glycoprotein, necessary for tenocyte proliferation and tendon maturation (Docheva et al., 2005; Jelinsky et al., 2010). Scleraxis, which is a transcription factor that has been shown to positively regulate the expression of tenomodulin and also collagen type I, is also used as a marker for tenocytes (Shukunami et al., 2006; Lejard et al., 2007; Jelinsky et al., 2010). Vimentin, a type III intermediate filament protein, present in mesenchymal derived cells such as fibroblasts, can also be used to confirm presence of tenocytes (Rufai et al., 1992; Bjur et al., 2008b), at least to differ them from cells of a non-fibroblastic origin. Results from our studies on the human tendon cells show that a vast majority of the cells in vitro, approximately >95%, is positive for tenomodulin, scleraxis and also vimentin, and that this phenotype is preserved through the passages used for our experiments.
However, these markers are not only present in tenocytes. For example, tenomodulin is also expressed in bone marrow, muscle, cartilage and fat (Jelinsky et al., 2010). Therefore, other cell types cannot be excluded to be present in the cell cultures in our in vitro model. In addition, we have noticed cells with characteristics of nerve cells, i.e. cells being PGP 9.5 positive and showing very long dendritic-like processes, which suggests that the cells have the ability to differentiate, since no nerve cell bodies are harvested with the biopsies due to them being located in the dorsal root ganglion or ventral horn. This means that, apart from the likeliness that other cells than tenocytes are cultured, also stem cells that have the ability to differentiate are included. Actually, SSEA-4, a marker for tendon stem/progenitor cells (Zhang and Wang,

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
61
2010), was present in a minority of our cells, those cells with a more plump appearance. It was however confirmed that the cell cultures did not contain any endothelial cells since no positive reactions were seen for CD31.
The knowledge that the cells in culture may not only consist of tenocytes, does not necessarily mean that it is a disadvantage, rather the opposite, i.e. that it might be more similar to the combination of cells as seen in vivo.
5.2.2.2 Interpretation of different forms of NK-1 R
One of the studies [III] raised the suggestion that the tenocytes express two different isoforms of NK-1 R, one glycosylated full-length and one unglycosylated truncated form. The basis of such suggestion was the results from Western blot, clearly indicating two different bands of 80 kDa and 37 kDa, respectively. It has previously been shown that migration of glycosylated full-length NK-1 R and glycosylated truncated NK-1 R results in bands corresponding to 80 kDa and 50 kDa, respectively, and that after deglycosylation the migration results in bands of 46 kDa (full-length unglycosylated) and 37 kDa (truncated unglycosylated) (Li et al., 1997). The fact that the SP induced phosphorylation of ERK1/2 was shown to be biphasic in this thesis work [III], further confirms the likeliness of the occurrence of two different forms of NK-1 R, as different time-response to stimulation is a known effect of different isoforms of NK-1 R (Lai et al., 2008).
5.2.2.3 Suggested future complementary
experiments
Further research is needed to establish the possible different forms of NK-1 R present in tenocytes (discussed in previous section), and an interesting aspect would be to investigate if SP has different effects depending on the form of NK-1 R expressed on the cell surface.
Additional experiments are also desirable to confirm some of the here obtained results, such as for instance analysing also the response in SP peptide levels following loading in the in vitro model, not just the mRNA levels as measured here.
It would also be interesting to compare tendon cells harvested from normal healthy individuals with cells from patients suffering from tendinosis. Actually, preliminary data of ours indicate that cells in culture harvested from healthy individuals have a lower SP production as compared to cells in culture from tendinosis patients, making it interesting to further study this.

Ludvig J. Backman, 2013
62
5.3 The possible importance of SP and
catecholamines in tendons and tendinosis
5.3.1 Aspects of the biochemical theory in the onset/development of tendinosis
Excessive training is the most accepted pathological stimulus leading to the development of tendinosis (Kader et al., 2002) (although also sedentary people are affected to a lesser degree (Rolf and Movin, 1997)). However, the underlying mechanisms that initiate tendinosis in response to load are unknown. There have been theories suggesting microtrauma to the collagen fibres, followed by insufficient time for repair, as part of the tendinosis pathogenesis, and also suggestions that overload causes an inflammation in the tendon tissue that triggers the development of tendinosis (Kader et al., 2002). In addition, it is not clear whether the initial event in the pathological cascade occurs in the collagen matrix or in the tenocytes (Jozsa, 1997). It may be that overloading the tendon results in collagen fibre disruption that either triggers mechanoreceptors or directly damage the tenocytes (Cook et al., 2004).
However, there are studies that provide data inconsistent with these mentioned theories. Firstly, the theory of microtrauma does not hold up considering the fact that the patients who have patellar tendon autograft for anterior cruciate ligament reconstruction, only develop minimal donor site knee pain even though a severe trauma is evident in the patellar tendon after harvesting of the graft (Adriani et al., 1995; Kiss et al., 1998). Also, the donors for Achilles tendon biopsies in our studies had a minor trauma to their tendon but no one developed tendinopathy. Secondly, the theory of intratendinous inflammation, as a cause of tendinosis is also opposed by contradicting data showing that patients with tendinosis do not display any inflammatory cells or increased levels of prostaglandin E2 (a classical marker of inflammatory processes) (Alfredson et al., 1999; Khan et al., 1999). It should here be mentioned that the results of paper II showed inflammation in the paratendinous tissue, i.e. outside the tendon tissue proper. Due to the inflammatory process being outside the tendon tissue it might be insufficient to have any impact in the tendon tissue proper. However, we raised the speculation that the paratendinous inflammation, caused by SP, is essential for the angiogenesis [II]. It has also been confirmed by others that inflammatory reactions are seen as an early response to overload in tendinosis models and that the inflammatory substances, such as prostaglandin E2, may lead to tendinosis changes in experimental models, which has raised the discussion of reviving the inflammatory hypothesis (Rees et al., 2013). Thus, it cannot be ruled out that a paratendinous inflammatory process is one of the components of importance in the early phase of tendinosis development. It should finally be pointed out, in the context of inflammation, that SP, among other substances, is said to be involved in ’neurogenic inflammation’ – an efferent effect of nervous

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
63
SP leading to vasodilatation, extravasation of plasma proteins, and leukocyte adhesion to the endothelium (Harrison and Geppetti, 2001).
The many opposing arguments for the theories of microtrauma and inflammation, prompted the alternative theory of a biochemical involvement (Khan et al., 2000), which has been supported by data of later studies (Danielson, 2009). However, it has so far not been possible to speculate on where in the process of tendinosis development the biochemical hypothesis has its role, since most data have only confirmed presence of biochemical substances (SP, catecholamines, acetylcholine, glutamate, TNF-α) in the chronic stage of tendinosis. This thesis confirms that a SP increase precedes tendinosis changes [I], and it can therefore be speculated that SP has a central role in the development of tendinosis. This speculation is furthermore strengthened by the fact that exogenously added SP accelerates tendinosis changes such as hypercellularity and angiogenesis [II]. It could therefore also be speculated that the initial event in the pathological cascade is activated by the tenocytes themselves – and not indirectly by tenocytes being activated by trauma in the extracellular matrix – since mechanical loading of tenocytes in vitro results in elevated levels of SP [III]. Cook and Purdam (Cook and Purdam, 2012) have also been speculating that the tendon cells are central components in tendinosis, not least in tendinosis development, a speculation that complements the biochemical theory discussed here. It is not farfetched to speculate, that the increased SP from tenocytes, due to load, via binding to NK-1 R on the same cell type (autocrine) or on neighbouring cells (paracrine; e.g., on blood vessels and nerves), is an important factor in the cell-central theory, especially considering the known effects of SP in other tissues, including stimulation of cell proliferation, angiogenesis, and pain. Frequently occurring, and cumulative, increases in SP levels, as a result of overload, might not leave enough time for recovery and tendinosis will be a fact. See figure 24 for a summarizing overview.
One aspect that needs to be considered is the fact that loading of tenocytes not only resulted in an SP mRNA increase, but also in a down-regulation of NK-1 R mRNA. The same result for NK-1 R, but on the level of protein, was also observed in cells exposed to exogenous SP, i.e. both internalized and membrane expressed NK-1 R was down-regulated. These latter findings indicate that new synthesising of the receptor is needed after SP exposure, not only recycling of the internalized NK-1 R, for complete recovery. The reduced expression of NK-1 R mRNA in response to load could thus be interpreted as a way of the cells to adjust to the mechanical stimuli, and thereby to the effect of increased SP, by decreasing synthesis of NK-1 R, resulting in less NK-1 Rs being expressed on the cell surface and being susceptible to SP (to minimize the SP effects). In view of this, the fact that NK-1 R is down-regulated in response to load, could possibly be seen as a confirmation that SP peptide production is increased (not only SP mRNA expression), since NK-1 R decrease is a known effect after SP stimulation (Grady et al., 1995; Grady et al., 1996) and as mentioned is also shown for the cells of the present studies [III]. Thus, this might confirm that SP truly binds to NK-1 R and initiate its effect, shown here to be both anti-apoptotic [V] and proliferative [III]. One should also bear in mind that the results of down-regulated NK-1 R mRNA in response to load are based on a specific time-point, and therefore it is not known when the recovery of the receptor expression

Ludvig J. Backman, 2013
64
occurs. In addition, the return of receptor responsiveness to SP (NK-1 R resensitisation) was not examined here, but is something that is explained to occur before the recovery of NK-1 R expression (Bennett et al., 2005). An important aspect is also the fact that we do not know how long the effect of SP is sustained after binding to NK-1 R, although data from this thesis indicate that stimulation with SP at least have sustained effects up to 48 h [e.g., Fig. V:4-5].
Fig. 24 This picture illustrates the speculation that endogenously produced SP by the tenocytes is central in the onset of tendinosis. Loading of the tenocytes/tendon results in increased SP production, with known proliferative and angiogenic effects, both of which are characteristics of tendinosis. Since loading results in elevated endogenous SP production, which precedes the presence of tendinosis changes, it is speculated that SP is involved in the onset of tendinosis (i.e. that SP is ‘pro-tendinotic’). There are other non-neuronal signal substances known to be up-regulated in tendinosis, albeit with so far unknown effects related to loading and the onset of tendinosis. Nevertheless, catecholamines have known proliferative effects on tenocytes and are likely to be involved in the onset of tendinosis since exogenous clonidine injections accelerate tendinosis changes.
5.3.2 Aspects of the biochemical theory in the
chronic stages of tendinosis
It has been shown that patients suffering from tendinosis have an up-regulated local production of SP and catecholamines in the chronic stage, and that the tenocytes in tendinosis also express the receptors NK-1 R and α2A AR to a higher extent than in healthy tendons (Danielson et al., 2007b; Andersson et al., 2008). Other signal substances have also been shown to follow the same pattern, i.e. up-regulation in tendinosis, such as acetylcholine (Danielson et al., 2007c), glutamate (Alfredson et al., 1999) and tumour necrosis factor alpha (TNF-α) (Gaida et al., 2012). The cells expressing evidence of increased production of signal substances and their preferred receptors are mainly the

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
65
cells with more rounded nuclei, most commonly in the areas of hypercellularity and referred to by some as tenoblasts (Chuen et al., 2004). The tenoblasts are demonstrating significantly higher proliferation as compared to the tenocytes (Chuen et al., 2004). This suggests that biochemical substances are not only involved in the development of tendinosis but also that they play a role in the chronic stage, i.e. maintaining the state of tendinosis. As has been repeatedly pointed out, the characteristics of tendinosis are, among others, hypercellularity and angiogenesis, and both NK-1 R and α2A AR stimulation are, in this thesis work, shown to result in tenocyte hypercellularity (both models), and NK-1 R stimulation also causes angiogenesis in vivo. These functions of the signal substances make it plausible that they play a central role in the maintenance of tendinosis, especially since SP in addition is a known pain signalling peptide (Helke et al., 1982). Worth mentioning is furthermore the fact that SP stimulates nerve ingrowth, as seen in tendon healing (Burssens et al., 2005a), which might serve as one possible pain-signalling pathway in tendinosis. These speculations on the roles of SP and catecholamines produced by tenocytes in chronic tendinosis, are based on the assumption that there are autocrine/paracrine effects, i.e. that the tenocytes producing catecholamines and SP influence the tenocytes themselves (autocrine), or neighbouring cells such as blood vessel cells or nerves (paracrine), by stimulating NK-1 R and α2A AR on the surface of these cells (all of which have been confirmed to be positive for these receptors in human tendon tissue in vivo (Danielson, 2009)).
Rolf and collaborators have shown that tendinosis cells are metabolically hyperactive and that their degenerative behaviour persists for many generations, even when removed from the tendinosis environment (Rolf et al., 2001). They concluded that once a normal spindle shaped tenocyte differentiates into a cell with more rounded nuclei seen in tendinosis, the process becomes difficult to reverse (Rolf et al., 2001). Speculatively, it is possible that continuous autocrine stimulation of the cells by SP, and maybe also catecholamines, may partly explain this phenomenon that the cells are difficult to reverse.
5.4 Summarizing reflections
5.4.1 The role of SP/catecholamines and the hypercellularity in tendinosis
One of the major characteristics of tendinosis is the hypercellularity seen in the tendon tissue proper. The area of hypercellularity mainly consists of the more metabolically active cells displaying abnormal characteristics of tendon cells, not least in their up-regulation of the SP and catecholamine systems, which in turn might contribute to the hypercellularity – an assumption corroborated by the results of this thesis. The existence of hypercellularity in tendinosis might initially be an attempt for the tendon to adjust in direct response to load, or to lack thereof, since it might be beneficial/physiological to increase cellularity and

Ludvig J. Backman, 2013
66
thereby collagen metabolism in early stages of recovery, but instead it ends up in a stage of ‘failed adaptation’, since the cells exhibit dysfunctional collagen synthesis, i.e. the tendon tissue does not normalise and the result is decreased tendon biomechanical function. Speculatively, hypercellularity might also occur indirectly as a response to load, as loading is here shown to be accompanied with increased SP production, with shown effects in stimulating proliferation (and anti-apoptosis), leading to hypercellularity.
5.4.2 The role of SP and apoptosis in
tendinosis
In tendinosis, not only hypercellularity is seen, but also excessive apoptosis. This might be an attempt of the tendon itself to regulate the number of cells, thereby limiting further presence of the metabolically hyperactive tendon cells that might cause unwanted effects, such as inadequate collagen arrangement (cf. previous section). Substances hindering this process of apoptosis are thus possibly also hindering an attempt of self-healing in tendinosis. SP, in addition to its proliferative effects, is here also shown to harbour an anti-apoptotic effect, implying that it hampers a possible self-regulating mechanism. Hence, it is speculated that SP, via binding to NK-1 R, has effects that partly explain why tendinosis is chronic, i.e. maintenance of the disease, since it prevents apoptosis of the tendinosis cells, in addition to stimulating their proliferation.
5.4.3 Possible clinical implications
In future clinical management of tendinosis, the aim of experimental treatment approaches could be to modify the biochemical microenvironment in the tendon, based on findings in this thesis and other previous and future studies. The validity of the biochemical hypothesis is here confirmed since the SP and catecholamine systems are found to be present and functional in the tendon tissue/cells. In addition, this thesis adds speculation to the biochemical hypothesis, that the signal substances produced by tendon cells, via their effect on hypercellularity, are responsible for the development and maintenance of tendinosis. Furthermore, the anti-apoptotic effect of SP is speculated to prevent self-healing of tendinosis. Based on these raised speculations, it might be worthwhile to attempt to block the NK-1 R and α2A AR receptors by administrating inhibitors, as a possible future treatment in the onset and/or the chronic stage of tendinosis, to prevent tendinosis and/or achieve reconstitution/normalization of tendon structure in later stages. However, further research is needed to confirm these hypotheses.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
67
6. CONCLUSIONS
This thesis is based on studies on two model systems: An in vivo rabbit Achilles tendinosis model and an in vitro human primary tendon cell culture model. In these studies it is concluded that:
1. Tenocytes express SP and NK- 1 R both in vitro and in vivo, and SP and
NK-1 R are also present in blood vessel walls, as well as on nerve fascicles, in the paratendinous tissue in vivo. In addition, there is an endogenous production of catecholamines in tenocytes, as evident by their expression of TH, as well as an expression of α2A AR on tenocytes, both in vivo and in vitro. TH-positive nerves are also present in the paratendinous tissue in vivo.
2. SP is up-regulated in response to loading both in vivo of the whole
Achilles tendon and in vitro of tenocytes, and the increased SP levels precedes the tendinosis changes in vivo. In addition, expression of NK-1 R is down-regulated in response to loading of tenocytes in vitro.
3. Stimulation with SP results in tenocyte hypercellularity both in vivo and
in vitro, as well as intratendinous angiogenesis and paratendinous inflammation in vivo. Furthermore, α2 AR stimulation with clonidine also results in tenocyte hypercellularity, both in vivo and in vitro. The effects of SP and clonidine on tenocytes in vitro are, at least in part, proliferative through activation/phosphorylation of ERK1/2, and mediated through stimulation of NK-1 R and α2A AR, respectively.
4. SP has an anti-apoptotic effect in Anti-Fas induced apoptosis of human
tenocytes, in a NK-1 R and Akt specific pathway.
The overall conclusion of the thesis is that:
Intratendinous SP is up-regulated in response to mechanical loading, preceding tendinosis, and stimulation of NK-1 R, as well as of α2A AR, contribute to tenocyte hypercellularity, through proliferative effects. In addition, SP also has anti-apoptotic and angiogenic effects in tendons/tendon cells, of possible importance in the development and maintenance of tendinosis.

Ludvig J. Backman, 2013
68
ACKNOWLEDGEMENTS
The studies of this thesis were conducted in a collaborative project between Umeå University, Departments of Integrative Medical Biology, Anatomy, and Surgical and Perioperative Sciences, Sports Medicine, and the University of British Columbia (UBC), Department of Physical Therapy, and the Centre for Hip Health and Mobility (CHHM), Vancouver Coastal Health and Research Institute, Vancouver, Canada.
During the work of this thesis many people have contributed to my work and I am indebted to you all. I wish, in particular, to express my sincere appreciation and gratitude to: Patrik Danielson, my supervisor, for your tremendous help. You are the most intelligent person I have ever met, you can handle more than anyone can imagine and with complete structure, I am deeply impressed by your capability. I am greatly thankful for your never-ending patience, generosity and guidance throughout this work, I would not have achieved this without you. I could not have asked for a better and more perfect supervisor. Alex Scott, at UBC and CHHM, my assistant supervisor, for inviting me to your laboratory in Vancouver and for very productive and rewarding research visits there. You have been a very knowledgeable supervisor, always with valuable input throughout my work. Håkan Alfredson, my assistant supervisor, for your endless enthusiasm and encouragement. Discussions with you have been incredibly inspiring and given me lots of energy. Gustav Andersson, my assistant supervisor, for always being extremely supportive and helpful. You are not only a great researcher but also an amazing illustrator. Your career will be as fast as your car! Sture Forsgren, the perfect example of a true and humble researcher and professor, for always having your door open and being helpful. You are a man of great qualities. Arthur Sampaio, your expertise and willingness to help with scientific advice have been invaluable. Ulla Hedlund, for your help and support in the laboratory. Lotta Alfredson, for your help in coordinating donations of tissue material. Adrian Lamouroux, Fellon Robson-Long, Ronny Lorentzon and Jamie Gaida for technical service and co-operation in the work with the animal model.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
69
To co-author and fellow researcher Gloria Fong, for your patience and support when you introduced me to the laboratory, I am also grateful for your friendliness during my stay in Vancouver. To fellow researchers Sandrine Le Roux and Peter Boman, for technical advice and inspiring scientific discussions, I really enjoyed sharing office and laboratory space with both of you! I also want to extend my appreciation to Filip Löfgren for excellent technical service, to Gabriel Wennstig for collaboration on the animal project, and to Per-Arne Oldenborg, Daniel Öhlund, Andreas Hult, Lars Frängsmyr, and Paul Kingham for valuable technical advice. To Göran Dahlgren for technical support and house-restoration advice, and to Anna-Lena Tallander and Anita Dreyer-Perkiömäki for your administrative assistance. To all colleagues at both Anatomy and Sports Medicine, and in Vancouver, especially to: Johan Bagge, Christoph Spang, Yafeng Song, Lina Renström, Jens Christensen, Qingjun Zhou, Björn Lindell, Ali Janbaz, Vahid Mohaghegh Harandi, Anton Tjust, Catarina Conde, Farhan Khalid Shah, Anna-Karin Olofsson, Mona Lindström, Lena Carlsson, Eva Carlsson, Jingxia Liu, Fatima Pedrosa Domellöf, Lev Novikov, Ludmila Novikova, Gunvor Hällström, Gunnel Folkesson, Mallappa Kolar, Maria Brohlin, Siggan Kilter, Rebecka Wiberg, Mikael Wiberg, and Per Stål for great company at the department. To my loving family, for always being supportive and generous, I cannot thank you all enough! Thanks to my father Clas Backman (who is actually the person behind the original “Backman model”) and my mother Margareta Backman, for being who you are. Finally, to Linda for literally always supporting me.....

Ludvig J. Backman, 2013
70
FUNDING
The studies of this thesis were financially supported by:
the National Swedish Research Council (VR)
the Swedish Society of Medicine (SLS)
the Faculty of Medicine, Umeå University (through a ‘samfinansierad doktorandtjänst’ for L.J.B.) and the board of Umeå University (through a Young Researcher Award to P.D.)
the Tore Nilsson Foundation for Medical Research
the Magn. Bergvall Foundation
the Sven and Dagmar Salén Foundation
the Sigurd and Elsa Golje Memorial Foundation
the J.C. Kempe and Seth M. Kempe Memorial Foundations
the Swedish National Centre for Research in Sports
the Knut and Alice Wallenberg Foundation (Vice-chancelor’s fund at Umeå University)
the J.C. Kempe Memorial Foundation Academic Funds at Umeå University
the Fund-raising Foundation (‘Insamlingsstiftelsen’) of the Faculty of Medicine at Umeå University
the Canadian Institutes of Health Research
the Michael Smith Foundation for Health Research

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
71
POPULÄRVETENSKAPLIG SAMMANFATTNING PÅ SVENSKA
Kronisk smärta i en muskelsena, kombinerat med nedsatt funktion och förtjockning av senan kallas för tendinopati, och är ett kliniskt syndrom som ofta, men inte alltid, utlöses av överbelastning. Tendinopati är i regel också förknippat med strukturella vävnadsförändringar, såsom ökad mängd celler (hypercellularitet), nybildning av blodkärl (angiogenes) samt ibland ökad mängd programmerad celldöd (apoptos), och tillståndet kallas då för tendinos. Tendinos är relativt vanligt, angriper ofta hälsenan (Achillessenan) och drabbar såväl elit- och motionsidrottare som icke-aktiva, och anses ofta vara svårt att behandla. De bakomliggande sjukdomsmekanismerna är till stor del okända och det existerar en rad olika teorier om möjliga orsaker. På senare tid har det presenterats en ny, ”biokemisk” teori som bygger på att sencellerna (tenocyterna) producerar signalämnen som man tidigare trodde främst producerades av nervceller, som exempelvis substans P (SP) och katekolaminer (adrenalin och noradrenalin). Vid tendinos har det visats att tenocyterna, av okänd anledning, har en ökad produktion av både SP och katekolaminer jämfört med i en frisk sena. Dessa substanser har kända effekter som kan leda till några av de vävnadsförändringar som är karaktäristiska vid tendinos, t.ex. hypercellularitet och angiogenes.
Hypotesen i detta avhandlingsprojekt är att belastning av en sena/senceller leder till ökad produktion av SP, samt att stimulering med SP av dess receptor neurokinin-1 receptorn (NK-1 R), liksom stimulering av katekolaminreceptorn α2A AR, bidrar till vävnadsförändringarna vid tendinos. För att studera detta har två modellsystem använts, en in vivo, kaninmodell, samt en in vitro, cellodlingsmodell.
Resultaten visar att både SP och NK-1 R samt katekolaminproduktion och α2A AR kan påvisas i tenocyterna i båda modellerna. Efter belastning sågs en ökad produktion av SP och en minskad produktion av NK-1 R. Intressant var att ökningen av SP-produktionen i kaninmodellen kom innan man kunde se tendinosförändringarna. Stimulering av både NK-1 R (med SP) och α2A AR resulterade i hypercellularitet i båda modellerna. Man kunde även i kaninmodellen konstatera att SP-injektioner ledde till angiogenes inuti senan och inflammation i vävnaden runt senan. Vad gäller hypercellularitet, vilket både kan förklaras av en ökad delning av celler (proliferation) och/eller minskad mängd programmerad celldöd (apoptos), kunde man här se att stimulering av både NK-1 R (med SP) och α2A AR hade proliferativa effekter, samt att SP även hade anti-apoptotisk effekt.
Studierna i denna avhandling visar alltså att stimulering av NK-1 R och α2A AR på tenocyter leder till förändringar som ses vid tendinos, såsom hypercellularitet. Betydelsen av hypercellularitet i tendinos är inte klarlagd, men troligtvis innebär det en negativ påverkan på tenocyternas produktion av, och strukturen på, den viktigaste byggstenen i en sena, fiberproteinet kollagen, så att senans biomekaniska funktion försämras. Med tanke på att produktion av SP ökar vid belastning och att ökningen kommer innan tendinos, samt att SP leder till typiska tendinosförändringar, är det rimligt att anta att SP inte bara är involverat i det kroniska tillståndet, utan även i utvecklingen av tendinos. Resultaten i denna avhandling indikerar att blockering av SP, och möjligen katekolaminer, kan vara fördelaktigt i behandlingen av de strukturella vävnadsförändringar som ses vid tendinos.

Ludvig J. Backman, 2013
72
REFERENCES
Abrahams Y, Laguette MJ, Prince S, Collins M (2013) Polymorphisms within the COL5A1 3'-UTR That Alters mRNA Structure and the MIR608 Gene are Associated with Achilles Tendinopathy. Ann Hum Genet Published online first: Jan 24; doi: 10.1111/ahg.12013.
Adams JM, Cory S (1998) The Bcl-2 protein family: arbiters of cell survival. Science 281; 1322-1326. Adriani E, Mariani PP, Maresca G, Santori N (1995) Healing of the patellar tendon after harvesting
of its mid-third for anterior cruciate ligament reconstruction and evolution of the unclosed donor site defect. Knee Surg Sports Traumatol Arthrosc 3; 138-143.
Alexander RM, Bennet-Clark HC (1977) Storage of elastic strain energy in muscle and other tissues. Nature 265; 114-117.
Alfredson H, Pietila T, Jonsson P, Lorentzon R (1998) Heavy-load eccentric calf muscle training for the treatment of chronic Achilles tendinosis. Am J Sports Med 26; 360-366.
Alfredson H, Thorsen K, Lorentzon R (1999) In situ microdialysis in tendon tissue: high levels of glutamate, but not prostaglandin E2 in chronic Achilles tendon pain. Knee Surg Sports Traumatol Arthrosc 7; 378-381.
Alfredson H, Ohberg L (2005) Neovascularisation in chronic painful patellar tendinosis--promising results after sclerosing neovessels outside the tendon challenge the need for surgery. Knee Surg Sports Traumatol Arthrosc 13; 74-80.
Alfredson H (2011) Ultrasound and Doppler-guided mini-surgery to treat midportion Achilles tendinosis: results of a large material and a randomised study comparing two scraping techniques. Br J Sports Med 45; 407-410.
Almeida TA, Rojo J, Nieto PM, Pinto FM, Hernandez M, Martin JD, Candenas ML (2004) Tachykinins and tachykinin receptors: structure and activity relationships. Curr Med Chem 11; 2045-2081.
Almekinders LC, Vellema JH, Weinhold PS (2002) Strain patterns in the patellar tendon and the implications for patellar tendinopathy. Knee Surg Sports Traumatol Arthrosc 10; 2-5.
Alvarez-Gonzalez R, Spring H, Muller M, Burkle A (1999) Selective loss of poly(ADP-ribose) and the 85-kDa fragment of poly(ADP-ribose) polymerase in nucleoli during alkylation-induced apoptosis of HeLa cells. J Biol Chem 274; 32122-32126.
Amantini C, Mosca M, Lucciarini R, Perfumi MC, Santoni G (2008) Thiorphan-induced survival and proliferation of rat thymocytes by activation of Akt/survivin pathway and inhibition of caspase-3 activity. J Pharmacol Exp Ther 327; 215-225.
An YS, Lee E, Kang MH, Hong HS, Kim MR, Jang WS, Son Y, Yi JY (2011) Substance P stimulates the recovery of bone marrow after the irradiation. J Cell Physiol 226; 1204-1213.
Andersson G, Danielson P, Alfredson H, Forsgren S (2007) Nerve-related characteristics of ventral paratendinous tissue in chronic Achilles tendinosis. Knee Surg Sports Traumatol Arthrosc 15; 1272-1279.
Andersson G, Danielson P, Alfredson H, Forsgren S (2008) Presence of substance P and the neurokinin-1 receptor in tenocytes of the human Achilles tendon. Regul Pept 150; 81-87.
Andersson G, Forsgren S, Scott A, Gaida JE, Stjernfeldt JE, Lorentzon R, Alfredson H, Backman C, Danielson P (2011) Tenocyte hypercellularity and vascular proliferation in a rabbit model of tendinopathy: contralateral effects suggest the involvement of central neuronal mechanisms. Br J Sports Med 45; 399-406.
Andres BM, Murrell GA (2008) Treatment of tendinopathy: what works, what does not, and what is on the horizon. Clin Orthop Relat Res 466; 1539-1554.
Astrom M, Rausing A (1995) Chronic Achilles tendinopathy. A survey of surgical and histopathologic findings. Clin Orthop Relat Res 151-164.
Backman C, Boquist L, Friden J, Lorentzon R, Toolanen G (1990) Chronic achilles paratenonitis with tendinosis: an experimental model in the rabbit. J Orthop Res 8; 541-547.
Bae SJ, Matsunaga Y, Takenaka M, Tanaka Y, Hamazaki Y, Shimizu K, Katayama I (2002) Substance P induced preprotachykinin-a mRNA, neutral endopeptidase mRNA and substance P in cultured normal fibroblasts. Int Arch Allergy Immunol 127; 316-321.
Bennett VJ, Perrine SA, Simmons MA (2005) Neurokinin-1 receptor resensitization precedes receptor recycling. J Pharmacol Exp Ther 313; 1347-1354.
Bjur D, Alfredson H, Forsgren S (2005) The innervation pattern of the human Achilles tendon: studies of the normal and tendinosis tendon with markers for general and sensory innervation. Cell Tissue Res 320; 201-206.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
73
Bjur D, Danielson P, Alfredson H, Forsgren S (2008a) Immunohistochemical and in situ hybridization observations favor a local catecholamine production in the human Achilles tendon. Histol Histopathol 23; 197-208.
Bjur D, Danielson P, Alfredson H, Forsgren S (2008b) Presence of a non-neuronal cholinergic system and occurrence of up- and down-regulation in expression of M2 muscarinic acetylcholine receptors: new aspects of importance regarding Achilles tendon tendinosis (tendinopathy). Cell Tissue Res 331; 385-400.
Bortolato M, Chen K, Shih JC (2008) Monoamine oxidase inactivation: from pathophysiology to therapeutics. Adv Drug Deliv Rev 60; 1527-1533.
Brimijoin S, Lundberg JM, Brodin E, Hokfelt T, Nilsson G (1980) Axonal transport of substance P in the vagus and sciatic nerves of the guinea pig. Brain Res 191; 443-457.
Brodin E, Gazelius B, Panopoulos P, Olgart L (1983) Morphine inhibits substance P release from peripheral sensory nerve endings. Acta Physiol Scand 117; 567-570.
Brunet A, Bonni A, Zigmond MJ, Lin MZ, Juo P, Hu LS, Anderson MJ, Arden KC, Blenis J, Greenberg ME (1999) Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 96; 857-868.
Bruzzone A, Pinero CP, Castillo LF, Sarappa MG, Rojas P, Lanari C, Luthy IA (2008) Alpha2-adrenoceptor action on cell proliferation and mammary tumour growth in mice. Br J Pharmacol 155; 494-504.
Bruzzone A, Pinero CP, Rojas P, Romanato M, Gass H, Lanari C, Luthy IA (2011) alpha(2)-Adrenoceptors enhance cell proliferation and mammary tumor growth acting through both the stroma and the tumor cells. Curr Cancer Drug Targets 11; 763-774.
Burbach JP (2010) Neuropeptides from concept to online database www.neuropeptides.nl. Eur J Pharmacol 626; 27-48.
Burssens P, Steyaert A, Forsyth R, van Ovost EJ, Depaepe Y, Verdonk R (2005a) Exogenously administered substance P and neutral endopeptidase inhibitors stimulate fibroblast proliferation, angiogenesis and collagen organization during Achilles tendon healing. Foot Ankle Int 26; 832-839.
Burssens P, Forsyth R, Steyaert A, Van Ovost E, Praet M, Verdonk R (2005b) Influence of burst TENS stimulation on collagen formation after Achilles tendon suture in man. A histological evaluation with Movat's pentachrome stain. Acta Orthop Belg 71; 342-346.
Butler DL, Grood ES, Noyes FR, Zernicke RF (1978) Biomechanics of ligaments and tendons. Exerc Sport Sci Rev 6; 125-181.
Carlsson O, Schizas N, Li J, Ackermann PW (2011) Substance P injections enhance tissue proliferation and regulate sensory nerve ingrowth in rat tendon repair. Scand J Med Sci Sports 21; 562-569.
Carmont MR, Highland AM, Rochester JR, Paling EM, Davies MB (2011) An anatomical and radiological study of the fascia cruris and paratenon of the Achilles tendon. Foot Ankle Surg 17; 186-192.
Carter MS, Krause JE (1990) Structure, expression, and some regulatory mechanisms of the rat preprotachykinin gene encoding substance P, neurokinin A, neuropeptide K, and neuropeptide gamma. J Neurosci 10; 2203-2214.
Chen J, Wang JH, Zhuang HX (2006) [Influence of substance P on the proliferation and apoptosis of fibroblasts of pathological scars]. Zhonghua Shao Shang Za Zhi 22; 277-280.
Chen SC, Tsai MH, Wang HJ, Yu HS, Chang LW (2007) Involvement of substance P and neurogenic inflammation in arsenic-induced early vascular dysfunction. Toxicol Sci 95; 82-88.
Chuen FS, Chuk CY, Ping WY, Nar WW, Kim HL, Ming CK (2004) Immunohistochemical characterization of cells in adult human patellar tendons. J Histochem Cytochem 52; 1151-1157.
Clement DB, Taunton JE, Smart GW (1984) Achilles tendinitis and peritendinitis: etiology and treatment. Am J Sports Med 12; 179-184.
Cook J (2011) Tendinopathy: no longer a 'one size fi ts all' diagnosis. Br J Sports Med 45; 385. Cook JL, Khan KM, Purdam C (2002) Achilles tendinopathy. Man Ther 7; 121-130. Cook JL, Feller JA, Bonar SF, Khan KM (2004) Abnormal tenocyte morphology is more prevalent
than collagen disruption in asymptomatic athletes' patellar tendons. J Orthop Res 22; 334-338. Cook JL, Purdam C (2012) Is compressive load a factor in the development of tendinopathy? Br J
Sports Med 46; 163-168. Cottrell GS, Padilla B, Pikios S, Roosterman D, Steinhoff M, Gehringer D, Grady EF, Bunnett NW
(2006) Ubiquitin-dependent down-regulation of the neurokinin-1 receptor. J Biol Chem 281; 27773-27783.

Ludvig J. Backman, 2013
74
Cummins EJ, Anson BJ, et al. (1946) The structure of the calcaneal tendon (of Achilles) in relation to orthopedic surgery, with additional observations on the plantaris muscle. Surg Gynecol Obstet 83; 107-116.
Curwin SL, Vailas AC, Wood J (1988) Immature tendon adaptation to strenuous exercise. J Appl Physiol 65; 2297-2301.
Cussac D, Schaak S, Gales C, Flordellis C, Denis C, Paris H (2002) alpha(2B)-Adrenergic receptors activate MAPK and modulate proliferation of primary cultured proximal tubule cells. Am J Physiol Renal Physiol 282; F943-952.
Dalla Costa AP, Clemente CF, Carvalho HF, Carvalheira JB, Nadruz W, Jr., Franchini KG (2010) FAK mediates the activation of cardiac fibroblasts induced by mechanical stress through regulation of the mTOR complex. Cardiovasc Res 86; 421-431.
Danielson P, Alfredson H, Forsgren S (2006) Immunohistochemical and histochemical findings favoring the occurrence of autocrine/paracrine as well as nerve-related cholinergic effects in chronic painful patellar tendon tendinosis. Microsc Res Tech 69; 808-819.
Danielson P, Alfredson H, Forsgren S (2007a) In situ hybridization studies confirming recent findings of the existence of a local nonneuronal catecholamine production in human patellar tendinosis. Microsc Res Tech 70; 908-911.
Danielson P, Alfredson H, Forsgren S (2007b) Studies on the importance of sympathetic innervation, adrenergic receptors, and a possible local catecholamine production in the development of patellar tendinopathy (tendinosis) in man. Microsc Res Tech 70; 310-324.
Danielson P, Andersson G, Alfredson H, Forsgren S (2007c) Extensive expression of markers for acetylcholine synthesis and of M2 receptors in tenocytes in therapy-resistant chronic painful patellar tendon tendinosis - a pilot study. Life Sci 80; 2235-2238.
Danielson P (2009) Reviving the "biochemical" hypothesis for tendinopathy: new findings suggest the involvement of locally produced signal substances. Br J Sports Med 43; 265-268.
Datta SR, Dudek H, Tao X, Masters S, Fu H, Gotoh Y, Greenberg ME (1997) Akt phosphorylation of BAD couples survival signals to the cell-intrinsic death machinery. Cell 91; 231-241.
Datta SR, Ranger AM, Lin MZ, Sturgill JF, Ma YC, Cowan CW, Dikkes P, Korsmeyer SJ, Greenberg ME (2002) Survival factor-mediated BAD phosphorylation raises the mitochondrial threshold for apoptosis. Dev Cell 3; 631-643.
De Camilli P (1991) Co-secretion of multiple signal molecules from endocrine cells via distinct exocytotic pathways. Trends Pharmacol Sci 12; 446-448.
de Vos RJ, Weir A, van Schie HT, Bierma-Zeinstra SM, Verhaar JA, Weinans H, Tol JL (2010) Platelet-rich plasma injection for chronic Achilles tendinopathy: a randomized controlled trial. JAMA 303; 144-149.
Devine KL, LeVeau BF, Yack HJ (1981) Electromyographic activity recorded from an unexercised muscle during maximal isometric exercise of the contralateral agonists and antagonists. Phys Ther 61; 898-903.
Docheva D, Hunziker EB, Fassler R, Brandau O (2005) Tenomodulin is necessary for tenocyte proliferation and tendon maturation. Mol Cell Biol 25; 699-705.
Doherty GP, Koike Y, Uhthoff HK, Lecompte M, Trudel G (2006) Comparative anatomy of rabbit and human achilles tendons with magnetic resonance and ultrasound imaging. Comp Med 56; 68-74.
Douglas SD, Leeman SE (2011) Neurokinin-1 receptor: functional significance in the immune system in reference to selected infections and inflammation. Ann N Y Acad Sci 1217; 83-95.
Downward J (2004) PI 3-kinase, Akt and cell survival. Semin Cell Dev Biol 15; 177-182. Eisenhofer G, Kopin IJ, Goldstein DS (2004) Catecholamine metabolism: a contemporary view
with implications for physiology and medicine. Pharmacol Rev 56; 331-349. Elmore S (2007) Apoptosis: a review of programmed cell death. Toxicol Pathol 35; 495-516. Fong G, Backman LJ, Andersson G, Scott A, Danielson P (2013) Human tenocytes are stimulated to
proliferate by acetylcholine through an EGFR signalling pathway. Cell Tissue Res 351; 465-475. Fukashiro S, Komi PV, Jarvinen M, Miyashita M (1995) In vivo Achilles tendon loading during
jumping in humans. Eur J Appl Physiol Occup Physiol 71; 453-458. Gaida JE, Ashe MC, Bass SL, Cook JL (2009) Is adiposity an under-recognized risk factor for
tendinopathy? A systematic review. Arthritis Rheum 61; 840-849. Gaida JE, Bagge J, Purdam C, Cook J, Alfredson H, Forsgren S (2012) Evidence of the TNF-alpha
system in the human Achilles tendon: expression of TNF-alpha and TNF receptor at both protein and mRNA levels in the tenocytes. Cells Tissues Organs 196; 339-352.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
75
Grady EF, Garland AM, Gamp PD, Lovett M, Payan DG, Bunnett NW (1995) Delineation of the endocytic pathway of substance P and its seven-transmembrane domain NK1 receptor. Mol Biol Cell 6; 509-524.
Grady EF, Gamp PD, Jones E, Baluk P, McDonald DM, Payan DG, Bunnett NW (1996) Endocytosis and recycling of neurokinin 1 receptors in enteric neurons. Neuroscience 75; 1239-1254.
Gross K, Karagiannides I, Thomou T, Koon HW, Bowe C, Kim H, Giorgadze N, Tchkonia T, Pirtskhalava T, Kirkland JL, Pothoulakis C (2009) Substance P promotes expansion of human mesenteric preadipocytes through proliferative and antiapoptotic pathways. Am J Physiol Gastrointest Liver Physiol 296; G1012-1019.
Hanada M, Feng J, Hemmings BA (2004) Structure, regulation and function of PKB/AKT--a major therapeutic target. Biochim Biophys Acta 1697; 3-16.
Hansson M, Forsgren S (1995) Immunoreactive atrial and brain natriuretic peptides are co-localized in Purkinje fibres but not in the innervation of the bovine heart conduction system. Histochem J 27; 222-230.
Harmar A, Schofield JG, Keen P (1980) Cycloheximide-sensitive synthesis of substance P by isolated dorsal root ganglia. Nature 284; 267-269.
Harmar A, Keen P (1982) Synthesis, and central and peripheral axonal transport of substance P in a dorsal root ganglion-nerve preparation in vitro. Brain Res 231; 379-385.
Harrison S, Geppetti P (2001) Substance p. Int J Biochem Cell Biol 33; 555-576. Hart, CB F, RC B (1995) Inflammatory processes in repetitive motion and overuse syndromes:
Potential role of neurogenic mechanisms in tendons and ligaments. Repetitive motion disorders of the upper extremity 247-262.
Hart DA, Archambault JM, Kydd A, Reno C, Frank CB, Herzog W (1998) Gender and neurogenic variables in tendon biology and repetitive motion disorders. Clin Orthop Relat Res 44-56.
Hart DA, Kydd A, Reno C (1999) Gender and pregnancy affect neuropeptide responses of the rabbit Achilles tendon. Clin Orthop Relat Res 237-246.
Helke CJ, Neil JJ, Massari VJ, Loewy AD (1982) Substance P neurons project from the ventral medulla to the intermediolateral cell column and ventral horn in the rat. Brain Res 243; 147-152.
Hokfelt T, Broberger C, Xu ZQ, Sergeyev V, Ubink R, Diez M (2000) Neuropeptides--an overview. Neuropharmacology 39; 1337-1356.
Hokfelt T, Pernow B, Wahren J (2001) Substance P: a pioneer amongst neuropeptides. J Intern Med 249; 27-40.
Jelinsky SA, Archambault J, Li L, Seeherman H (2010) Tendon-selective genes identified from rat and human musculoskeletal tissues. J Orthop Res 28; 289-297.
Jing C, Jia-Han W, Hong-Xing Z (2010) Double-edged effects of neuropeptide substance P on repair of cutaneous trauma. Wound Repair Regen 18; 319-324.
Johnson DG, Walker CL (1999) Cyclins and cell cycle checkpoints. Annu Rev Pharmacol Toxicol 39; 295-312.
Jozsa L, Balint JB, Reffy A, Demel Z (1979) Histochemical and ultrastructural study of adult human tendon. Acta Histochem 65; 250-257.
Jozsa L (1997) Human Tendons. Anatomy, Physiology and pathology. Human Kinetics, Champaign, IL; USA.
Kader D, Saxena A, Movin T, Maffulli N (2002) Achilles tendinopathy: some aspects of basic science and clinical management. Br J Sports Med 36; 239-249.
Kannus P (2000) Structure of the tendon connective tissue. Scand J Med Sci Sports 10; 312-320. Karkoulias G, Mastrogianni O, Lymperopoulos A, Paris H, Flordellis C (2006) alpha(2)-Adrenergic
receptors activate MAPK and Akt through a pathway involving arachidonic acid metabolism by cytochrome P450-dependent epoxygenase, matrix metalloproteinase activation and subtype-specific transactivation of EGFR. Cell Signal 18; 729-739.
Keen P, Harmar AJ, Spears F, Winter E (1982) Biosynthesis, axonal transport and turnover of neuronal substance P. Ciba Found Symp 145-164.
Kerr JF, Wyllie AH, Currie AR (1972) Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. Br J Cancer 26; 239-257.
Khan KM, Maffulli N (1998) Tendinopathy: an Achilles' heel for athletes and clinicians. Clin J Sport Med 8; 151-154.
Khan KM, Cook JL, Bonar F, Harcourt P, Astrom M (1999) Histopathology of common tendinopathies. Update and implications for clinical management. Sports Med 27; 393-408.
Khan KM, Cook JL, Maffulli N, Kannus P (2000) Where is the pain coming from in tendinopathy? It may be biochemical, not only structural, in origin. Br J Sports Med 34; 81-83.

Ludvig J. Backman, 2013
76
Kim MO, Na SI, Lee MY, Heo JS, Han HJ (2008) Epinephrine increases DNA synthesis via ERK1/2s through cAMP, Ca(2+)/PKC, and PI3K/Akt signaling pathways in mouse embryonic stem cells. J Cell Biochem 104; 1407-1420.
Kischkel FC, Hellbardt S, Behrmann I, Germer M, Pawlita M, Krammer PH, Peter ME (1995) Cytotoxicity-dependent APO-1 (Fas/CD95)-associated proteins form a death-inducing signaling complex (DISC) with the receptor. EMBO J 14; 5579-5588.
Kiss ZS, Kellaway DP, Cook JL, Khan KM (1998) Postoperative patellar tendon healing: an ultrasound study. VIS Tendon Study Group. Australas Radiol 42; 28-32.
Komi PV, Fukashiro S, Jarvinen M (1992) Biomechanical loading of Achilles tendon during normal locomotion. Clin Sports Med 11; 521-531.
Koon HW, Zhao D, Na X, Moyer MP, Pothoulakis C (2004) Metalloproteinases and transforming growth factor-alpha mediate substance P-induced mitogen-activated protein kinase activation and proliferation in human colonocytes. J Biol Chem 279; 45519-45527.
Koon HW, Zhao D, Zhan Y, Moyer MP, Pothoulakis C (2007) Substance P mediates antiapoptotic responses in human colonocytes by Akt activation. Proc Natl Acad Sci U S A 104; 2013-2018.
Krause JE, Chirgwin JM, Carter MS, Xu ZS, Hershey AD (1987) Three rat preprotachykinin mRNAs encode the neuropeptides substance P and neurokinin A. Proc Natl Acad Sci U S A 84; 881-885.
Kroemer G, Galluzzi L, Vandenabeele P, Abrams J, Alnemri ES, Baehrecke EH, Blagosklonny MV, El-Deiry WS, Golstein P, Green DR, Hengartner M, Knight RA, Kumar S, Lipton SA, Malorni W, Nunez G, Peter ME, Tschopp J, Yuan J, Piacentini M, Zhivotovsky B, Melino G (2009) Classification of cell death: recommendations of the Nomenclature Committee on Cell Death 2009. Cell Death Differ 16; 3-11.
Kvist M, Jozsa L, Jarvinen MJ, Kvist H (1987) Chronic Achilles paratenonitis in athletes: a histological and histochemical study. Pathology 19; 1-11.
Kvist M (1991) Achilles tendon injuries in athletes. Ann Chir Gynaecol 80; 188-201. Lai JP, Douglas SD, Ho WZ (1998a) Human lymphocytes express substance P and its receptor. J
Neuroimmunol 86; 80-86. Lai JP, Douglas SD, Rappaport E, Wu JM, Ho WZ (1998b) Identification of a delta isoform of
preprotachykinin mRNA in human mononuclear phagocytes and lymphocytes. J Neuroimmunol 91; 121-128.
Lai JP, Lai S, Tuluc F, Tansky MF, Kilpatrick LE, Leeman SE, Douglas SD (2008) Differences in the length of the carboxyl terminus mediate functional properties of neurokinin-1 receptor. Proc Natl Acad Sci U S A 105; 12605-12610.
Lake SP, Ansorge HL, Soslowsky LJ (2008) Animal models of tendinopathy. Disabil Rehabil 30; 1530-1541.
Lejard V, Brideau G, Blais F, Salingcarnboriboon R, Wagner G, Roehrl MH, Noda M, Duprez D, Houillier P, Rossert J (2007) Scleraxis and NFATc regulate the expression of the pro-alpha1(I) collagen gene in tendon fibroblasts. J Biol Chem 282; 17665-17675.
Levitzki A (1978) Catecholamine receptors. Rev Physiol Biochem Pharmacol 82; 1-26. Li C, Kim K (2008) Neuropeptides. WormBook, ed. The C. elegans Research Community,
WormBook, www.wormbook.org. doi/10.1895/wormbook.1891.1142.1891. Li H, Leeman SE, Slack BE, Hauser G, Saltsman WS, Krause JE, Blusztajn JK, Boyd ND (1997) A
substance P (neurokinin-1) receptor mutant carboxyl-terminally truncated to resemble a naturally occurring receptor isoform displays enhanced responsiveness and resistance to desensitization. Proc Natl Acad Sci U S A 94; 9475-9480.
Li H, Zhu H, Xu CJ, Yuan J (1998) Cleavage of BID by caspase 8 mediates the mitochondrial damage in the Fas pathway of apoptosis. Cell 94; 491-501.
Liu X, Li P, Widlak P, Zou H, Luo X, Garrard WT, Wang X (1998) The 40-kDa subunit of DNA fragmentation factor induces DNA fragmentation and chromatin condensation during apoptosis. Proc Natl Acad Sci U S A 95; 8461-8466.
Lui PP, Maffulli N, Rolf C, Smith RK (2011) What are the validated animal models for tendinopathy? Scand J Med Sci Sports 21; 3-17.
Lundgreen K, Lian OB, Engebretsen L, Scott A (2011) Tenocyte apoptosis in the torn rotator cuff: a primary or secondary pathological event? Br J Sports Med 45; 1035-1039.
Lysholm J, Wiklander J (1987) Injuries in runners. Am J Sports Med 15; 168-171. Maddika S, Ande SR, Panigrahi S, Paranjothy T, Weglarczyk K, Zuse A, Eshraghi M, Manda KD,
Wiechec E, Los M (2007) Cell survival, cell death and cell cycle pathways are interconnected: implications for cancer therapy. Drug Resist Updat 10; 13-29.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
77
Maffulli N, Testa V, Capasso G, Bifulco G, Binfield PM (1997) Results of percutaneous longitudinal tenotomy for Achilles tendinopathy in middle- and long-distance runners. Am J Sports Med 25; 835-840.
Maffulli N, Khan KM, Puddu G (1998) Overuse tendon conditions: time to change a confusing terminology. Arthroscopy 14; 840-843.
Maffulli N, Sharma P, Luscombe KL (2004) Achilles tendinopathy: aetiology and management. J R Soc Med 97; 472-476.
Mafi N, Lorentzon R, Alfredson H (2001) Superior short-term results with eccentric calf muscle training compared to concentric training in a randomized prospective multicenter study on patients with chronic Achilles tendinosis. Knee Surg Sports Traumatol Arthrosc 9; 42-47.
Magnussen RA, Dunn WR, Thomson AB (2009) Nonoperative treatment of midportion Achilles tendinopathy: a systematic review. Clin J Sport Med 19; 54-64.
Masure S, Haefner B, Wesselink JJ, Hoefnagel E, Mortier E, Verhasselt P, Tuytelaars A, Gordon R, Richardson A (1999) Molecular cloning, expression and characterization of the human serine/threonine kinase Akt-3. Eur J Biochem 265; 353-360.
Matheson GO (2010) Nonoperative treatment of midportion Achilles tendinopathy: a review. Clin J Sport Med 20; 72-73.
Mazzocca AD, Chowaniec D, McCarthy MB, Beitzel K, Cote MP, McKinnon W, Arciero R (2012) In vitro changes in human tenocyte cultures obtained from proximal biceps tendon: multiple passages result in changes in routine cell markers. Knee Surg Sports Traumatol Arthrosc 20; 1666-1672.
McCarthy PW, Lawson SN (1989) Cell type and conduction velocity of rat primary sensory neurons with substance P-like immunoreactivity. Neuroscience 28; 745-753.
McCluskey A, Lalkhen A (2007) Statistics II: Central tendency and spread of data. Continuing Education in Anaesthesia, Critical Care & Pain 7; 127-130.
McConalogue K, Dery O, Lovett M, Wong H, Walsh JH, Grady EF, Bunnett NW (1999) Substance P-induced trafficking of beta-arrestins. The role of beta-arrestins in endocytosis of the neurokinin-1 receptor. J Biol Chem 274; 16257-16268.
Mebratu Y, Tesfaigzi Y (2009) How ERK1/2 activation controls cell proliferation and cell death: Is subcellular localization the answer? Cell Cycle 8; 1168-1175.
Meloche S, Pouyssegur J (2007) The ERK1/2 mitogen-activated protein kinase pathway as a master regulator of the G1- to S-phase transition. Oncogene 26; 3227-3239.
Merighi A, Polak JM, Gibson SJ, Gulbenkian S, Valentino KL, Peirone SM (1988) Ultrastructural studies on calcitonin gene-related peptide-, tachykinins- and somatostatin-immunoreactive neurones in rat dorsal root ganglia: evidence for the colocalization of different peptides in single secretory granules. Cell Tissue Res 254; 101-109.
Michael-Titus AT, Fernandes K, Setty H, Whelpton R (2002) In vivo metabolism and clearance of substance P and co-expressed tachykinins in rat striatum. Neuroscience 110; 277-286.
Milner P, Kirkpatrick KA, Ralevic V, Toothill V, Pearson J, Burnstock G (1990) Endothelial cells cultured from human umbilical vein release ATP, substance P and acetylcholine in response to increased flow. Proc Biol Sci 241; 245-248.
Missale C, Nash SR, Robinson SW, Jaber M, Caron MG (1998) Dopamine receptors: from structure to function. Physiol Rev 78; 189-225.
Molinoff PB, Axelrod J (1971) Biochemistry of catecholamines. Annu Rev Biochem 40; 465-500. Morimoto I, Ogata T (1968) [Torsion of the achilles tendon in man and its anatomical and surgical
significance]. Kaibogaku Zasshi 43; 295-302. Myerson MS, McGarvey W (1999) Disorders of the Achilles tendon insertion and Achilles
tendinitis. Instr Course Lect 48; 211-218. Nagatsu T (1991) Genes for human catecholamine-synthesizing enzymes. Neurosci Res 12; 315-345. New DC, Wong YH (2007) Molecular mechanisms mediating the G protein-coupled receptor
regulation of cell cycle progression. J Mol Signal 2; 2. New DC, Wu K, Kwok AW, Wong YH (2007) G protein-coupled receptor-induced Akt activity in
cellular proliferation and apoptosis. FEBS J 274; 6025-6036. Norregaard J, Larsen CC, Bieler T, Langberg H (2007) Eccentric exercise in treatment of Achilles
tendinopathy. Scand J Med Sci Sports 17; 133-138. O'Brien M (1992) Functional anatomy and physiology of tendons. Clin Sports Med 11; 505-520. O'Brien M (1997) Structure and metabolism of tendons. Scand J Med Sci Sports 7; 55-61. Oben JA, Diehl AM (2004) Sympathetic nervous system regulation of liver repair. Anat Rec A
Discov Mol Cell Evol Biol 280; 874-883.

Ludvig J. Backman, 2013
78
Oben JA, Roskams T, Yang S, Lin H, Sinelli N, Torbenson M, Smedh U, Moran TH, Li Z, Huang J, Thomas SA, Diehl AM (2004) Hepatic fibrogenesis requires sympathetic neurotransmitters. Gut 53; 438-445.
Ohberg L, Lorentzon R, Alfredson H (2001) Neovascularisation in Achilles tendons with painful tendinosis but not in normal tendons: an ultrasonographic investigation. Knee Surg Sports Traumatol Arthrosc 9; 233-238.
Oliver FJ, de la Rubia G, Rolli V, Ruiz-Ruiz MC, de Murcia G, Murcia JM (1998) Importance of poly(ADP-ribose) polymerase and its cleavage in apoptosis. Lesson from an uncleavable mutant. J Biol Chem 273; 33533-33539.
Opolka A, Straub RH, Pasoldt A, Grifka J, Grassel S (2012) Substance P and norepinephrine modulate murine chondrocyte proliferation and apoptosis. Arthritis Rheum 64; 729-739.
Paavola M, Kannus P, Paakkala T, Pasanen M, Jarvinen M (2000) Long-term prognosis of patients with achilles tendinopathy. An observational 8-year follow-up study. Am J Sports Med 28; 634-642.
Pages G, Lenormand P, L'Allemain G, Chambard JC, Meloche S, Pouyssegur J (1993) Mitogen-activated protein kinases p42mapk and p44mapk are required for fibroblast proliferation. Proc Natl Acad Sci U S A 90; 8319-8323.
Panoskaltsis-Mortari A, Bucy RP (1995) In situ hybridization with digoxigenin-labeled RNA probes: facts and artifacts. Biotechniques 18; 300-307.
Pierre-Jerome C, Moncayo V, Terk MR (2010) MRI of the Achilles tendon: a comprehensive review of the anatomy, biomechanics, and imaging of overuse tendinopathies. Acta Radiol 51; 438-454.
Pintado CO, Pinto FM, Pennefather JN, Hidalgo A, Baamonde A, Sanchez T, Candenas ML (2003) A role for tachykinins in female mouse and rat reproductive function. Biol Reprod 69; 940-946.
Porter AG, Janicke RU (1999) Emerging roles of caspase-3 in apoptosis. Cell Death Differ 6; 99-104.
Posthumus M, Collins M, Cook J, Handley CJ, Ribbans WJ, Smith RK, Schwellnus MP, Raleigh SM (2010) Components of the transforming growth factor-beta family and the pathogenesis of human Achilles tendon pathology--a genetic association study. Rheumatology (Oxford) 49; 2090-2097.
Puddu G, Ippolito E, Postacchini F (1976) A classification of Achilles tendon disease. Am J Sports Med 4; 145-150.
Qiu YW, Zhu LW, Zhang X, Zhang P (2012) [The effects of insulin-like growth factor 1 and transforming growth factor beta-3 at various concentration on tenocyte survival and collagen formation]. Zhonghua Wai Ke Za Zhi 50; 744-747.
Rees JD, Stride M, Scott A (2013) Tendons - time to revisit inflammation. Br J Sports Med Published online first: March 9; doi: 10.1136/bjsports-2012-091957.
Regoli D, Nguyen QT, Jukic D (1994) Neurokinin receptor subtypes characterized by biological assays. Life Sci 54; 2035-2047.
Reinherz RP, Granoff SR, Westerfield M (1991) Pathologic afflictions of the Achilles tendon. J Foot Surg 30; 117-121.
Riley G (2004) The pathogenesis of tendinopathy. A molecular perspective. Rheumatology (Oxford) 43; 131-142.
Riley G (2008) Tendinopathy--from basic science to treatment. Nat Clin Pract Rheumatol 4; 82-89.
Rolf C, Movin T (1997) Etiology, histopathology, and outcome of surgery in achillodynia. Foot Ankle Int 18; 565-569.
Rolf CG, Fu BS, Pau A, Wang W, Chan B (2001) Increased cell proliferation and associated expression of PDGFRbeta causing hypercellularity in patellar tendinosis. Rheumatology (Oxford) 40; 256-261.
Rowe V, Hemmings S, Barton C, Malliaras P, Maffulli N, Morrissey D (2012) Conservative management of midportion Achilles tendinopathy: a mixed methods study, integrating systematic review and clinical reasoning. Sports Med 42; 941-967.
Rufai A, Benjamin M, Ralphs JR (1992) Development and ageing of phenotypically distinct fibrocartilages associated with the rat Achilles tendon. Anat Embryol (Berl) 186; 611-618.
Sagan S, Chassaing G, Pradier L, Lavielle S (1996) Tachykinin peptides affect differently the second messenger pathways after binding to CHO-expressed human NK-1 receptors. J Pharmacol Exp Ther 276; 1039-1048.
Salvesen GS, Dixit VM (1999) Caspase activation: the induced-proximity model. Proc Natl Acad Sci U S A 96; 10964-10967.

Neuropeptide and catecholamine effects on tenocytes in tendinosis development
79
Schaak S, Cussac D, Cayla C, Devedjian JC, Guyot R, Paris H, Denis C (2000) Alpha(2) adrenoceptors regulate proliferation of human intestinal epithelial cells. Gut 47; 242-250.
Scheid MP, Woodgett JR (2003) Unravelling the activation mechanisms of protein kinase B/Akt. FEBS Lett 546; 108-112.
Schepsis AA, Wagner C, Leach RE (1994) Surgical management of Achilles tendon overuse injuries. A long-term follow-up study. Am J Sports Med 22; 611-619.
Schizas N, Weiss R, Lian O, Frihagen F, Bahr R, Ackermann PW (2012) Glutamate receptors in tendinopathic patients. J Orthop Res 30; 1447-1452.
Scott A, Alfredson H, Forsgren S (2008) VGluT2 expression in painful Achilles and patellar tendinosis: evidence of local glutamate release by tenocytes. J Orthop Res 26; 685-692.
Scott A, Danielson P, Abraham T, Fong G, Sampaio AV, Underhill TM (2011) Mechanical force modulates scleraxis expression in bioartificial tendons. J Musculoskelet Neuronal Interact 11; 124-132.
September AV, Cook J, Handley CJ, van der Merwe L, Schwellnus MP, Collins M (2009) Variants within the COL5A1 gene are associated with Achilles tendinopathy in two populations. Br J Sports Med 43; 357-365.
Severini C, Improta G, Falconieri-Erspamer G, Salvadori S, Erspamer V (2002) The tachykinin peptide family. Pharmacol Rev 54; 285-322.
Shukunami C, Takimoto A, Oro M, Hiraki Y (2006) Scleraxis positively regulates the expression of tenomodulin, a differentiation marker of tenocytes. Dev Biol 298; 234-247.
Stanish WD, Rubinovich RM, Curwin S (1986) Eccentric exercise in chronic tendinitis. Clin Orthop Relat Res 65-68.
Sulzer D, Sonders MS, Poulsen NW, Galli A (2005) Mechanisms of neurotransmitter release by amphetamines: a review. Prog Neurobiol 75; 406-433.
Sussmilch-Leitch SP, Collins NJ, Bialocerkowski AE, Warden SJ, Crossley KM (2012) Physical therapies for Achilles tendinopathy: systematic review and meta-analysis. J Foot Ankle Res 5; 15.
Tansky MF, Pothoulakis C, Leeman SE (2007) Functional consequences of alteration of N-linked glycosylation sites on the neurokinin 1 receptor. Proc Natl Acad Sci U S A 104; 10691-10696.
Taylor SH, Al-Youha S, Van Agtmael T, Lu Y, Wong J, McGrouther DA, Kadler KE (2011) Tendon is covered by a basement membrane epithelium that is required for cell retention and the prevention of adhesion formation. PLoS One 6; e16337.
Teitz CC, Garrett WE, Jr., Miniaci A, Lee MH, Mann RA (1997) Tendon problems in athletic individuals. Instr Course Lect 46; 569-582.
Tewari M, Quan LT, O'Rourke K, Desnoyers S, Zeng Z, Beidler DR, Poirier GG, Salvesen GS, Dixit VM (1995) Yama/CPP32 beta, a mammalian homolog of CED-3, is a CrmA-inhibitable protease that cleaves the death substrate poly(ADP-ribose) polymerase. Cell 81; 801-809.
Tuluc F, Lai JP, Kilpatrick LE, Evans DL, Douglas SD (2009) Neurokinin 1 receptor isoforms and the control of innate immunity. Trends Immunol 30; 271-276.
Tzivion G, Dobson M, Ramakrishnan G (2011) FoxO transcription factors; Regulation by AKT and 14-3-3 proteins. Biochim Biophys Acta 1813; 1938-1945.
Wall ME, Weinhold PS, Siu T, Brown TD, Banes AJ (2007) Comparison of cellular strain with applied substrate strain in vitro. J Biomech 40; 173-181.
White DM, Helme RD (1985) Release of substance P from peripheral nerve terminals following electrical stimulation of the sciatic nerve. Brain Res 336; 27-31.
Williams JG (1986) Achilles tendon lesions in sport. Sports Med 3; 114-135. Wolf BB, Green DR (1999) Suicidal tendencies: apoptotic cell death by caspase family proteinases. J
Biol Chem 274; 20049-20052. Wolf BB, Schuler M, Echeverri F, Green DR (1999) Caspase-3 is the primary activator of apoptotic
DNA fragmentation via DNA fragmentation factor-45/inhibitor of caspase-activated DNase inactivation. J Biol Chem 274; 30651-30656.
Woo SL, Orlando CA, Gomez MA, Frank CB, Akeson WH (1986) Tensile properties of the medial collateral ligament as a function of age. J Orthop Res 4; 133-141.
Yamaguchi K, Kugimiya T, Miyazaki T (2005a) Substance P receptor in U373 MG human astrocytoma cells activates mitogen-activated protein kinases ERK1/2 through Src. Brain Tumor Pathol 22; 1-8.
Yamaguchi K, Richardson MD, Bigner DD, Kwatra MM (2005b) Signal transduction through substance P receptor in human glioblastoma cells: roles for Src and PKCdelta. Cancer Chemother Pharmacol 56; 585-593.

Ludvig J. Backman, 2013
80
Yao L, Bestwick CS, Bestwick LA, Maffulli N, Aspden RM (2006) Phenotypic drift in human tenocyte culture. Tissue Eng 12; 1843-1849.
Yao R, Cooper GM (1995) Requirement for phosphatidylinositol-3 kinase in the prevention of apoptosis by nerve growth factor. Science 267; 2003-2006.
Yoon JH, Halper J (2005) Tendon proteoglycans: biochemistry and function. J Musculoskelet Neuronal Interact 5; 22-34.
Zhang J, Wang JH (2010) Characterization of differential properties of rabbit tendon stem cells and tenocytes. BMC Musculoskelet Disord 11; 10.
Zutphen LEMv, Baumans V, Beynen AC (2001) Principles of laboratory animal science, Revised edition. Elsevier, USA; ISBN: 978-970-444-50612-50612.


















