Myocardial function improved by electromagnetic field induction of stress protein hsp70
8
Myocardial Function Improved by Electromagnetic Field Induction of Stress Protein hsp70 ISAAC GEORGE, 1 MATTHEW S. GEDDIS, 2 ZACHARY LILL, 1 HANA LIN, 3 TEODORO GOMEZ, 2 MARTIN BLANK, 4 MEHMET C. OZ, 1 AND REBA GOODMAN 3 * 1 Department of Surgery, Division of Cardiothoracic Surgery, Columbia University College of Physicians and Surgeons, New York, New York 2 Department of Surgery, Division of Surgical Sciences, Columbia University College of Physicians and Surgeons, New York, New York 3 Department of Anatomy and Pathology, Columbia University College of Physicians and Surgeons, New York, New York 4 Department of Physiology and Cellular Biophysics, Columbia University College of Physicians and Surgeons, New York, New York Studies on myocardial function have shown that hsp70, stimulated by an increase in temperature, leads to improved survival following ischemia-reperfusion (I-R). Low frequency electromagnetic fields (EMFs) also induce the stress protein hsp70, but without elevating temperature. We have examined the hemodynamic changes in concert with EMF pre-conditioning and the induction of hsp70 to determine whether improved myocardial function occurs following I-R injury in Sprague–Dawley rats. Animals were exposed to EMF (60 Hz, 8 mT) for 30 min prior to I-R. Ischemia was then induced by ligation of left anterior descending coronary artery (LAD) for 30 min, followed by 30 min of reperfusion. Blood and heart tissue levels for hsp70 were determined by Western blot and RNA transcription by rtPCR. Significant upregulation of the HSP70 gene and increased hsp70 levels were measured in response to EMF pre-exposures. Invasive hemodynamics, as measured using a volume conductance catheter, demonstrated significant recovery of systolic contractile function after 30 min of reperfusion following EMF exposure. Additionally, isovolemic relaxation, a measure of ventricular diastolic function, was markedly improved in EMF-treated animals. In conclusion, non-invasive EMF induction of hsp70 preserved myocardial function and has the potential to improve tolerance to ischemic injury. J. Cell. Physiol. 216: 816–823, 2008. ß 2008 Wiley-Liss, Inc. Cardiovascular disease is the leading cause of morbidity and mortality in the United States, accounting for 70–80% of deaths in men and women over the age of 65. Furthermore, congestive heart failure is the most common cause of hospitalization of the elderly, and its incidence continues to increase (Schneider, 1999). In open and percutaneous revascularization procedures (coronary artery bypass surgery and percutaneous coronary interventions (PCIs)) and in the treatment of myocardial infarction and heart failure, it is essential to protect cardiomyocytes from the effects of hypoxia and ischemia (Bolli et al., 2004). Currently, myocardial protection can be accomplished by induction of the stress protein hsp70 through the use of elevated temperature (heat shock) (Currie et al., 1993; Udelsman et al., 1993; Nitta et al., 1994; Plumier and Currie, 1996). Induction of stress proteins by heat to prevent stroke and myocardial infarction during reperfusion has been shown to partially protect the myocardium under ischemic stress in a variety of models (Heads et al., 1995; Mestril et al., 1996; Plumier and Currie, 1996; Benjamin and McMillan, 1998; Chong et al., 1998; Cornelussen et al., 1998). The use of heat stress pre-treatment leads to moderate increases in hsp70 levels, but does not improve ischemia tolerance in isolated hearts (Cornelussen et al., 1998). Moreover, heat stress pre-treatment (hyperthermia) is of limited clinical utility since it requires a temperature elevation to 428C, a level impractical for clinical use or to achieve sufficient hsp70 increases. We have shown previously that 60 Hz electromagnetic fields (EMFs) upregulate the heat shock gene, HSP70 and induce elevated levels of hsp70 protein in the absence of elevated temperature (Goodman et al., 1994; Goodman and Blank, 1998; Han et al., 1998; Lin et al., 1998, 1999, 2001; Carmody et al., 2000). Of particular relevance, we previously elevated hsp70 levels in cultured rodent cardiomyocytes using EMF pre-treatment (Goodman and Blank, 2002). Additionally, studies from Di Carlo et al. (1999) and Shallom et al. (2002) confirmed that cardiomyocytes were protected from anoxic damage in EMF exposed chick embryos. The induction of increased levels of hsp70 protein by low frequency EMF exposures offers multiple clinical advantages over thermal, chemical or gene-transfer methods of induction for both patient and clinician. EMF stimulation of cytoprotective proteins is a non-invasive procedure easily administered to the patient. EMF-induced hsp70 does not turn off baseline protein synthesis, in contrast to elevated temperature (Goodman et al., 1989). A significant increase in hsp70 stress protein is induced within 5 min at 14 orders of magnitude lower energy input than thermal stress. Additionally, unlike thermal stress, the induced protection can be restimulated even after the stress is already present, and restimulation with even higher hsp70 levels can be induced by a different field strength, higher (800 mG) or lower (8 mG; Blank et al., 1994; Lin et al., 1997). Abbreviations: LVSP, left ventricular systolic pressure; LV dP/dt max , left ventricular; P-A Max Power, pre-load adjusted maximum power; G, Gauss; rtPCR, reverse-transcriptase polymerase chain reaction.Contract grant sponsor: Robert I. Goodman Fund. Contract grant sponsor: National Institute of Health; Contract grant number: T32-HL07854. *Correspondence to: Reba Goodman, Department of Pathology, Columbia University College of Physicians and Surgeons, PH, Rm. 15-90, 630 W. 168th St., New York, NY 10032. E-mail: [email protected] Received 5 February 2008; Accepted 4 March 2008 DOI: 10.1002/jcp.21461 ORIGINAL ARTICLE 816 Journal of Journal of Cellular Physiology Cellular Physiology ß 2008 WILEY-LISS, INC.
-
Upload
isaac-george -
Category
Documents
-
view
212 -
download
0
Transcript of Myocardial function improved by electromagnetic field induction of stress protein hsp70

ORIGINAL ARTICLE 816J o u r n a l o fJ o u r n a l o f
CellularPhysiologyCellularPhysiology
Myocardial Function Improved byElectromagnetic Field Inductionof Stress Protein hsp70
ISAAC GEORGE,1 MATTHEW S. GEDDIS,2 ZACHARY LILL,1 HANA LIN,3 TEODORO GOMEZ,2MARTIN BLANK,4 MEHMET C. OZ,1 AND REBA GOODMAN3*1Department of Surgery, Division of Cardiothoracic Surgery, Columbia University College of Physicians and Surgeons, New York,
New York2Department of Surgery, Division of Surgical Sciences, Columbia University College of Physicians and Surgeons, New York, New York3Department of Anatomy and Pathology, Columbia University College of Physicians and Surgeons, New York, New York4Department of Physiology and Cellular Biophysics, Columbia University College of Physicians and Surgeons, New York, New York
Studies on myocardial function have shown that hsp70, stimulated by an increase in temperature, leads to improved survival followingischemia-reperfusion (I-R). Low frequency electromagnetic fields (EMFs) also induce the stress protein hsp70, but without elevatingtemperature. We have examined the hemodynamic changes in concert with EMF pre-conditioning and the induction of hsp70 todetermine whether improved myocardial function occurs following I-R injury in Sprague–Dawley rats. Animals were exposed to EMF(60 Hz, 8 mT) for 30 min prior to I-R. Ischemia was then induced by ligation of left anterior descending coronary artery (LAD) for 30 min,followed by 30 min of reperfusion. Blood and heart tissue levels for hsp70 were determined by Western blot and RNA transcription byrtPCR. Significant upregulation of the HSP70 gene and increased hsp70 levels were measured in response to EMF pre-exposures. Invasivehemodynamics, as measured using a volume conductance catheter, demonstrated significant recovery of systolic contractile function after30 min of reperfusion following EMF exposure. Additionally, isovolemic relaxation, a measure of ventricular diastolic function, wasmarkedly improved in EMF-treated animals. In conclusion, non-invasive EMF induction of hsp70 preserved myocardial function and has thepotential to improve tolerance to ischemic injury.
J. Cell. Physiol. 216: 816–823, 2008. � 2008 Wiley-Liss, Inc.
Abbreviations: LVSP, left ventricular systolic pressure; LV dP/dtmax,left ventricular; P-A Max Power, pre-load adjusted maximumpower; G, Gauss; rtPCR, reverse-transcriptase polymerase chainreaction.Contract grant sponsor: Robert I. Goodman Fund.Contract grant sponsor: National Institute of Health;Contract grant number: T32-HL07854.
*Correspondence to: Reba Goodman, Department of Pathology,Columbia University College of Physicians and Surgeons, PH, Rm.15-90, 630 W. 168th St., New York, NY 10032.E-mail: [email protected]
Received 5 February 2008; Accepted 4 March 2008
DOI: 10.1002/jcp.21461
Cardiovascular disease is the leading cause of morbidity andmortality in the United States, accounting for 70–80% of deathsin men and women over the age of 65. Furthermore, congestiveheart failure is the most common cause of hospitalization of theelderly, and its incidence continues to increase (Schneider,1999). In open and percutaneous revascularization procedures(coronary artery bypass surgery and percutaneous coronaryinterventions (PCIs)) and in the treatment of myocardialinfarction and heart failure, it is essential to protectcardiomyocytes from the effects of hypoxia and ischemia (Bolliet al., 2004). Currently, myocardial protection can beaccomplished by induction of the stress protein hsp70 throughthe use of elevated temperature (heat shock) (Currie et al.,1993; Udelsman et al., 1993; Nitta et al., 1994; Plumier andCurrie, 1996). Induction of stress proteins by heat to preventstroke and myocardial infarction during reperfusion has beenshown to partially protect the myocardium under ischemicstress in a variety of models (Heads et al., 1995; Mestril et al.,1996; Plumier and Currie, 1996; Benjamin and McMillan, 1998;Chong et al., 1998; Cornelussen et al., 1998). The use of heatstress pre-treatment leads to moderate increases in hsp70levels, but does not improve ischemia tolerance in isolatedhearts (Cornelussen et al., 1998). Moreover, heat stresspre-treatment (hyperthermia) is of limited clinical utility sinceit requires a temperature elevation to 428C, a level impracticalfor clinical use or to achieve sufficient hsp70 increases.
We have shown previously that 60 Hz electromagnetic fields(EMFs) upregulate the heat shock gene, HSP70 and induceelevated levels of hsp70 protein in the absence of elevatedtemperature (Goodman et al., 1994; Goodman and Blank, 1998;Han et al., 1998; Lin et al., 1998, 1999, 2001; Carmody et al.,2000). Of particular relevance, we previously elevated hsp70levels in cultured rodent cardiomyocytes using EMFpre-treatment (Goodman and Blank, 2002). Additionally,
� 2 0 0 8 W I L E Y - L I S S , I N C .
studies from Di Carlo et al. (1999) and Shallom et al. (2002)confirmed that cardiomyocytes were protected from anoxicdamage in EMF exposed chick embryos.
The induction of increased levels of hsp70 protein by lowfrequency EMF exposures offers multiple clinical advantagesover thermal, chemical or gene-transfer methods of inductionfor both patient and clinician. EMF stimulation of cytoprotectiveproteins is a non-invasive procedure easily administered to thepatient. EMF-induced hsp70 does not turn off baseline proteinsynthesis, in contrast to elevated temperature (Goodman et al.,1989). A significant increase in hsp70 stress protein is inducedwithin 5 min at 14 orders of magnitude lower energy input thanthermal stress. Additionally, unlike thermal stress, the inducedprotection can be restimulated even after the stress is alreadypresent, and restimulation with even higher hsp70 levels can beinduced by a different field strength, higher (800 mG) or lower(8 mG; Blank et al., 1994; Lin et al., 1997).

Fig. 1. EMF exposure system. Animals were exposed to 60 Hz/8 mTEMFs by Helmholtz coils (19 G copper wire, 164 turns, 1.5 inches thickcovered with electrical tape; part A) that was contained within aplastic exposure cage (part B). The EMF field was perpendicular to theexposure device.
M Y O C A R D I A L P R O T E C T I O N A N D E M F 817
Extensive reports in the literature have shown that elevatedlevels of hsp70 improve cardiac function after hypoxic stressand ischemia-reperfusion (I-R; Suzuki et al., 1997, 2000, 2002).In this set of experiments, we evaluated whether pre-treatmentwith EMF-induced levels of hsp70 can preserve myocardialfunction after ischemia-reperfusion.
MethodsAnimal care
Male Sprague–Dawley rats (250–400 g) were selected as theexperimental species. To assure their health, the animals wereexamined by the animal facility veterinarian upon arrival. Animalswere allowed to acclimate for at least 2 days and adjust to beinghandled before randomization into the study. Animals were housedin cages in an environmentally controlled room within the animalcare facility at Columbia University. Care and management ofrats was conducted according to facility standard operatingprocedures. At the conclusion of the acclimation period animalsjudged to be suitable for testing were assigned sequentially toeither treatment or control. This investigation conforms with theGuide for the Care and Use of Laboratory Animals published by the USNational Institutes of Health (NIH Publication No. 85-23, revised1996). This study was approved by the Institutional Animal Careand Use Committee of Columbia University.
Electromagnetic field exposures
All EMF exposures were performed prior to induction of ischemia.The exposures described here are pre-treatment exposures. Inprevious studies, we tested a variety of field strengths andfrequencies (Goodman et al., 1989; Wei et al., 1990; Jin et al., 1997),and eventually determined that a 60 Hz frequency and a fieldstrength of 8 mT consistently induced the highest level oftranscriptional activation of the HSP70 gene and the highest hsp70protein levels (reviewed in Goodman and Blank, 2002). In thestudies reported here, animals were exposed to 60 Hz/8 mT EMFsin a plastic exposure cage (16 cm� 24 cm) surrounded byHelmholtz coils (19 gauge copper wire, 164 turns, 1.5 inches thickcovered with electrical tape). The system was designed andcalibrated (R. Cangialosi, Electro-Biology, Inc., Fairfield, NJ) to ourspecifications for comfortably holding a large rodent; the cageholding the rats was suspended in a plastic enclosure (Fig. 1).EMF conditions were set using a function generator (BK Precision4011A 5 MHz, Yorba Linda, CA) and digital multimeter(BK Precision 2706A). Rectal temperature was continuouslymonitored with a thermocouple probe (�0.18C resolution;PhysiTemp., Cliffside Park, NJ). The digital multimeter was used tomeasure the field intensity and verify the systems operation. Fieldparameters were monitored with a Hitachi V-1065 100 MHzoscilloscope and a calibrated inductive search coil. Exposureconditions were monitored with a Sypris triaxial magnetic fieldmeter (Model 4080, Bell Laboratories, Orlando, FL). Experimentswere carried out at room temperature (approx. 258C).
Anesthesia
Rats were anesthetized with 2% isoflurane and mechanicallyventilated via a tracheostomy (Harvard rodent ventilator, model683, South Natick, MA) throughout the duration of theexperiment.
Surgical ProceduresBlood collection for hsp70 determination
The left femoral vein was cannulated with polyethylene tubing(OD 0.965 mm, Becton Dickinson, Franklin Lakes, NJ) forrepeated blood draws. Rats were then randomized to EMFexposure (60 Hz 8 mT) for 30 min (n¼ 6) or Control(no EMF exposure, n¼ 6). Blood was collected at baseline
JOURNAL OF CELLULAR PHYSIOLOGY
(pre-exposure) and after EMF exposure every 30 min up to120 min. Serum was spun down at 1,200 rpm for 10 min; packedred blood cells (RBCs) were flash-frozen for hsp70 proteinanalyses, and stored at �808C.
Terminal tissue for HSP70 RNA determination
Heart tissue (left ventricle; LV) was flash-frozen for analysisof HSP70 RNA by reverse-transcriptase polymerase chainreaction (rtPCR). GAPDH confirmed loading concentrations.
Ischemia-reperfusion protocol
The hemodynamic effects of EMF exposure on myocardialfunction after I-R (EMF n¼ 10, Control n¼ 10) were measuredusing the following protocol. Pre-anesthetized animals wererandomly assigned to EMF pre-treatment and underwent EMFexposure (60 Hz, 8 mT) for 30 min. During this time period,each individual animal was allowed to rest comfortably in the

818 G E O R G E E T A L .
exposure device under quiet conditions. No medications wereadministered prior or during this time. Animals designated ascontrols received no exposure and their cages were maintainedin an EMF-free area and monitored with a Sypris triaxialgaussmeter. At the conclusion of 30 min, all rats (both exposedand control) were anesthetized with 2% isoflurane andmechanically ventilated on 2% isoflurane after a tracheostomy.A 2 Fr volume conductance catheter (Millar Instruments,Houston, TX) was inserted into the left ventricle (LV) forcontinuous pressure-volume (PV) tracings immediatelyfollowing right carotid artery cannulation. After leftthoracotomy, coronary ischemia was created by ligating theleft anterior descending (LAD) coronary artery with a5-0 polypropylene suture for a total of 30 min before release ofthe suture. Reperfusion was monitored for an additional 30 min.Hemodynamics (Chart for Windows v5, ADI Instruments,Colorado Spring, CO) were recorded at baseline(pre-ischemia), 30 min of ischemia, and at 1, 10, 20, and 30 minafter reperfusion. Upon completion of the experiment, theheart was excised, and the LV was sectioned for histology.Ischemic and non-ischemic portions of the LV were flash-frozenfor extraction of protein for Western blot analysis of hsp70levels and RNA extraction for reverse-transcriptasepolymerase chain reaction (rtPCR) to determine upregulationof the HSP70 gene.
Hemodynamics and pressure-volume analysis
Hemodynamic determinations were made on all ratsundergoing I-R (n¼ 20). LV end-systolic pressure (LVSP), LVend-diastolic pressure (LVEDP), and LV volume were measuredusing the Millar conductance catheter placed into the LV acrossthe aortic valve. End-systolic volume (ESV) and end-diastolicvolume (EDV) were measured using standard techniques (Itoand Bassett, 1983). Cardiac output (CO), arterial elastance (Ea),and pre-load adjusted maximum power (P-A Max Power) werecomputed using a pressure-volume analysis program (PVAN v.3.2, Millar Instruments). The time constant of LV isovolumetricpressure relaxation, t, was calculated using the logarithmicmethod described by Raff and Glantz (1981). In experimentswhere animals had undergoneischemia-reperfusion, the hemodynamic and pressure-volumeanalyses indicated no adverse events or mortality associatedwith EMF exposure.
Protein sample preparation. Protein was extracted frommyocardial tissue and packed RBCs using methods previouslydescribed (Lin et al., 1998; Carmody et al., 2000). Proteinconcentrations were determined by Bradford assay (Bio-RadLaboratories, Hercules, CA).
Western blot. Equivalent (30 mg) amounts of protein wereseparated by gel electrophoresis on 10% polyacrylamide gelsusing appropriate molecular weight markers and transferred toPVDF membrane for immunoblotting. Blots were probedwith anti-hsp70 antibody (1:10,000; kindly provided byDr. Richard Morimoto, Northwestern University). The blotswere then stripped and reprobed with anti-b actin (1:1,000,Sigma-Aldrich, St. Louis, MO) to confirm equivalent loading.Visualization was by the ECL detection system as previouslydescribed (Lin et al., 1998).
Reverse-transcriptase polymerase chain reaction(rtPCR). Total RNA was extracted from non-ischemic andischemic left ventricular (LV) heart tissue using Trizol reagent(Life Technologies, Inc., Rockville, MD). Total RNA (0.1mg) wasprocessed directly to cDNA synthesis using the TaqMan1
Reverse Transcriptase Reagents kit (Applied Biosystems,Foster City, CA). All PCR primers and TaqMan probes weredesigned using PrimerExpress software (Applied Biosystems)and published sequence data from the NCBI database (Lin et al.,2001).
JOURNAL OF CELLULAR PHYSIOLOGY
Quantification of bands on films from Western blots andrtPCR. The films from Western blots and rtPCR analyseswere scanned into a computer. The density of the bands wasmeasured using image analysis software (ImageJ v1.38, NIH).
Statistical analysis. Continuous variables are expressedas mean� standard error and compared using two-tailedindependent t-testing with Levene’s Test for Equality ofvariances. Categorical variables were compared by x2 tests.Paired t-testing was used to evaluate significance within groupsat multiple time points. For all analyses, a P-value of less than0.05 was considered statistically significant. All analysis wasperformed using SPSS software (v. 11.5, Chicago, IL).
ResultsEMF-induced hsp70 levels
As described above, rats were exposed to EMF (60 Hz, 8 mT)for 30 min, blood samples were collected from the left femoralvein from both EMF-pre-treated and unexposed (control) ratsjust before (baseline) and immediately after EMF exposure(time 0) and at 30, 60, 90, and 120 min. Blood samples wereprepared for protein extraction and subsequent Western blotanalyses of hsp70. As seen in Figure 2A, the levels of hsp70protein were significantly elevated following 30 min of EMFpre-exposure, and the increase in protein level was sustainedfor 120 min post-exposure (P< 0.05 vs. Control, n¼ 6 pergroup, Fig. 2A); peak levels were reached at 30 min. Lowbaseline (constitutive) levels of hsp70 protein exist undernon-stressed conditions. After EMF exposure, these levelswere significantly elevated (>40%).
EMF-induced transcript levels for HSP70 RNA
Terminal myocardial tissue was collected for determination ofHSP70 RNA by rtPCR. Transcript levels of HSP70 RNA weresignificantly increased in terminal myocardial tissue extractsin response to EMF pre-exposure as compared to controls(P< 0.05 vs. Control, n¼ 6 per group), demonstratingupregulation of the HSP70 gene (Fig. 2B). There was no increasein HSP70 transcript levels in Control animals during the sametime period. These results confirm previous literature that EMFinduces upregulation of the HSP70 gene and increases hsp70protein levels (Lin et al., 1997, 1998, 1999, 2001).
Ischemia-reperfusion: hsp70 levels
Levels of hsp70 were increased in both non-ischemic (N) andischemic (I) left ventricle (LV) tissue after the termination ofI-R in EMF-exposed samples as compared to Control animals(P< 0.05 vs. Control, n¼ 10 per group; Fig. 3A). rtPCR analysisof terminal LV tissue from ischemic (I) and non-ischemic (N),showed increased RNA transcript levels in EMF pre-exposedtissue as compared to controls (P¼ 0.055 vs. Control, n¼ 10per group; Fig. 3B). The current results confirm that, in thisischemia-reperfusion model, EMF exposure upregulates theHSP70 gene and significantly increases hsp70 levels.
Ischemia-reperfusion: Hemodynamics
No adverse events or mortality were associated with EMFexposure. The mean temperatures before and after 30 minof EMF exposure were 36.1� 0.28C and 36.0� 0.28C,respectively, versus 36.0� 1.18C in Control rats (EMF vs.Control, P¼NS). The hemodynamics at baseline, after 30 minof coronary ischemia and after 10–30 min of reperfusion, aresummarized in Table 1. LVSP was significantly improvedbeginning at 10 min of reperfusion until 30 min in EMF-treatedanimals versus Control (Fig. 4A). No significant differenceswere seen in LVEDP, ESV, and EDV. Coronary ischemiaproduced a significant reduction in LV dP/dtmax (a measure ofventricular contractility) after 30 min. Recovery of LV dP/dtmax

Fig. 3. Ischemia-reperfusion: hsp70 protein levels and HSP70 RNAtranscript levels were increased significantly following EMF exposure.A: Levels of hsp70 were increased in both non-ischemic (N) andischemic (I) blood samples after ischemia-reperfusion. However,the hsp70 increase in ischemia samples was significant as comparedwith non-ischemic and control samples (MP < 0.05 vs. Control).B: A significant increase in HSP70 transcript levels was seen in theEMF pre-exposed terminal myocardium tissue from ischemia (I) leftventricular tissue as compared with non-ischemic and controlsamples (MP U 0.055 vs. Control).
Fig. 2. hsp70 protein levels and HSP70 RNA transcript levels areprogressively elevated after a 30-min EMF exposure. A: In response to30 min of EMF exposure (60 Hz/8 mT), hsp70 levels in blood weresignificantly elevated at 30 min and sustained for 120 min as comparedto unexposed control samples (MP < 0.05 vs. Control). B: HSP70 RNAtranscript levels were significantly increased in terminal myocardialtissue extracts in response to EMF exposure as compared tounexposed control samples (MP < 0.05 vs. Control).
M Y O C A R D I A L P R O T E C T I O N A N D E M F 819
was only seen in EMF-treated groups compared to Controlfrom 10 until 30 min after reperfusion (Fig. 4B). Pre-loadadjusted maximum power, a load and heart rate independentindex of systolic contractile function (the ability of the heart toeject blood), was similarly improved in EMF-treated rats afterreperfusion, as shown in Figure 4C. These results, interpretedcollectively, show that overall organ perfusion was increasedduring reperfusion (manifested by higher LVSP). The ability toincrease ventricular pressure at the time of reperfusion despiteischemic injury, without changes in blood volume, ventriculardimensions, or exogenous pharmacologic agents, implicates adirect mechanism to improve cardiac myocytes, and myocardialsystolic contractility. In addition, the lack of ventriculardilatation and absence of end-diastolic pressure elevationimplies normal end-systolic and end-diastolic pressure volumerelationships, in contrast to acutely failing hearts, whichundergo pressure-volume overload, enlargement of ventriculardimensions, and elevated end-diastolic pressures.
The contribution of diastolic function was also evaluated bycalculating the time constant of LV isovolumetric relaxation. Aslight, but statistically significant decrease in t (a measure ofdiastolic function) was observed throughout the course ofreperfusion after EMF treatment (Fig. 5), indicating improved
JOURNAL OF CELLULAR PHYSIOLOGY
ventricular wall compliance, allowing faster and greaterdiastolic filling. Greater diastolic filling and relaxation of theventricular wall improves cardiac output, lessens myocardialoxygen consumption, and increases the overall efficiency of theheart. Although no significant changes in arterial elastance wereseen, a strong recovery trend of cardiac output to 71% ofbaseline with EMF versus only 43% in Control was observed.Adverse changes in arterial elastance would potentially affectlong-term remodeling of central arteries, leading to multi-organdysfunction (most notably, renal failure).
In summary, a clear effect on systolic contractile function inEMF-treated animals was found after reperfusion, as shown byincreases in global indices of contractile function (LV dP/dtmax
and pre-load adjusted maximum power). These changesoccurred without evidence of concurrent LV hypertrophy or at
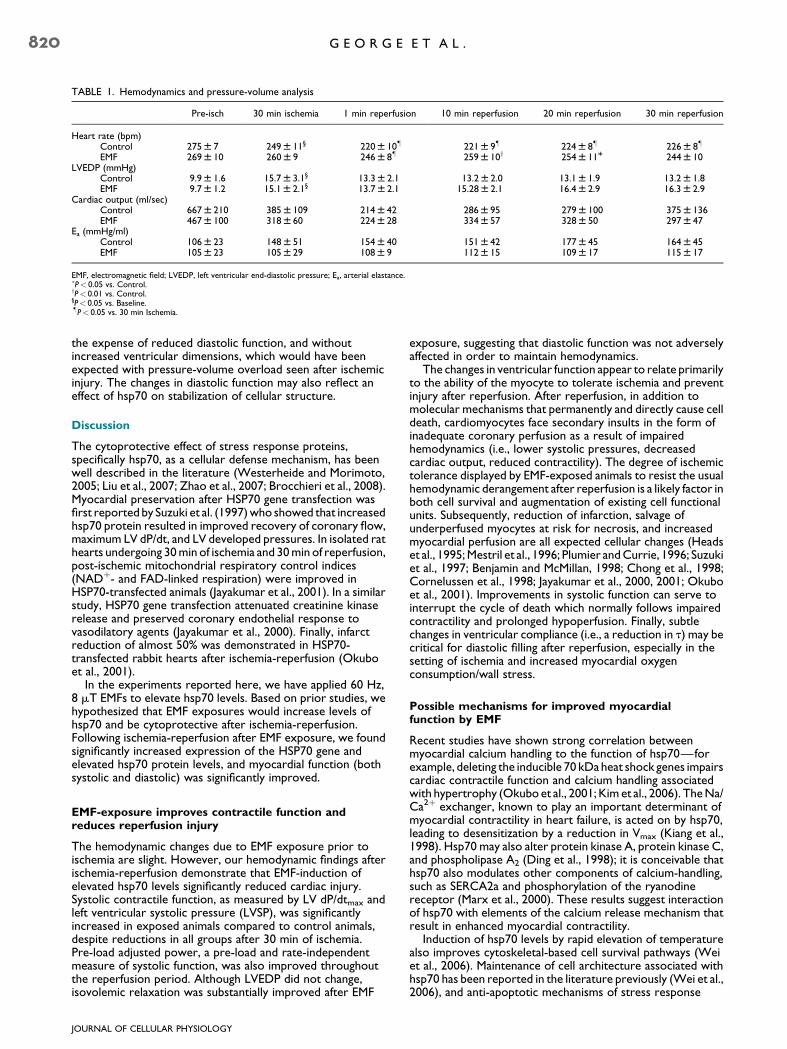
TABLE 1. Hemodynamics and pressure-volume analysis
Pre-isch 30 min ischemia 1 min reperfusion 10 min reperfusion 20 min reperfusion 30 min reperfusion
Heart rate (bpm)Control 275 W 7 249 W 11§ 220 W 10� 221 W 9� 224 W 8� 226 W 8�
EMF 269 W 10 260 W 9 246 W 8� 259 W 10y 254 W 11M 244 W 10LVEDP (mmHg)
Control 9.9 W 1.6 15.7 W 3.1§ 13.3 W 2.1 13.2 W 2.0 13.1 W 1.9 13.2 W 1.8EMF 9.7 W 1.2 15.1 W 2.1§ 13.7 W 2.1 15.28 W 2.1 16.4 W 2.9 16.3 W 2.9
Cardiac output (ml/sec)Control 667 W 210 385 W 109 214 W 42 286 W 95 279 W 100 375 W 136EMF 467 W 100 318 W 60 224 W 28 334 W 57 328 W 50 297 W 47
Ea (mmHg/ml)Control 106 W 23 148 W 51 154 W 40 151 W 42 177 W 45 164 W 45EMF 105 W 23 105 W 29 108 W 9 112 W 15 109 W 17 115 W 17
EMF, electromagnetic field; LVEDP, left ventricular end-diastolic pressure; Ea, arterial elastance.�P< 0.05 vs. Control.yP< 0.01 vs. Control.§P< 0.05 vs. Baseline.�P< 0.05 vs. 30 min Ischemia.
820 G E O R G E E T A L .
the expense of reduced diastolic function, and withoutincreased ventricular dimensions, which would have beenexpected with pressure-volume overload seen after ischemicinjury. The changes in diastolic function may also reflect aneffect of hsp70 on stabilization of cellular structure.
Discussion
The cytoprotective effect of stress response proteins,specifically hsp70, as a cellular defense mechanism, has beenwell described in the literature (Westerheide and Morimoto,2005; Liu et al., 2007; Zhao et al., 2007; Brocchieri et al., 2008).Myocardial preservation after HSP70 gene transfection wasfirst reported by Suzuki et al. (1997) who showed that increasedhsp70 protein resulted in improved recovery of coronary flow,maximum LV dP/dt, and LV developed pressures. In isolated rathearts undergoing 30 min of ischemia and 30 min of reperfusion,post-ischemic mitochondrial respiratory control indices(NADþ- and FAD-linked respiration) were improved inHSP70-transfected animals (Jayakumar et al., 2001). In a similarstudy, HSP70 gene transfection attenuated creatinine kinaserelease and preserved coronary endothelial response tovasodilatory agents (Jayakumar et al., 2000). Finally, infarctreduction of almost 50% was demonstrated in HSP70-transfected rabbit hearts after ischemia-reperfusion (Okuboet al., 2001).
In the experiments reported here, we have applied 60 Hz,8 mT EMFs to elevate hsp70 levels. Based on prior studies, wehypothesized that EMF exposures would increase levels ofhsp70 and be cytoprotective after ischemia-reperfusion.Following ischemia-reperfusion after EMF exposure, we foundsignificantly increased expression of the HSP70 gene andelevated hsp70 protein levels, and myocardial function (bothsystolic and diastolic) was significantly improved.
EMF-exposure improves contractile function andreduces reperfusion injury
The hemodynamic changes due to EMF exposure prior toischemia are slight. However, our hemodynamic findings afterischemia-reperfusion demonstrate that EMF-induction ofelevated hsp70 levels significantly reduced cardiac injury.Systolic contractile function, as measured by LV dP/dtmax andleft ventricular systolic pressure (LVSP), was significantlyincreased in exposed animals compared to control animals,despite reductions in all groups after 30 min of ischemia.Pre-load adjusted power, a pre-load and rate-independentmeasure of systolic function, was also improved throughoutthe reperfusion period. Although LVEDP did not change,isovolemic relaxation was substantially improved after EMF
JOURNAL OF CELLULAR PHYSIOLOGY
exposure, suggesting that diastolic function was not adverselyaffected in order to maintain hemodynamics.
The changes in ventricular function appear to relate primarilyto the ability of the myocyte to tolerate ischemia and preventinjury after reperfusion. After reperfusion, in addition tomolecular mechanisms that permanently and directly cause celldeath, cardiomyocytes face secondary insults in the form ofinadequate coronary perfusion as a result of impairedhemodynamics (i.e., lower systolic pressures, decreasedcardiac output, reduced contractility). The degree of ischemictolerance displayed by EMF-exposed animals to resist the usualhemodynamic derangement after reperfusion is a likely factor inboth cell survival and augmentation of existing cell functionalunits. Subsequently, reduction of infarction, salvage ofunderperfused myocytes at risk for necrosis, and increasedmyocardial perfusion are all expected cellular changes (Headset al., 1995; Mestril et al., 1996; Plumier and Currie, 1996; Suzukiet al., 1997; Benjamin and McMillan, 1998; Chong et al., 1998;Cornelussen et al., 1998; Jayakumar et al., 2000, 2001; Okuboet al., 2001). Improvements in systolic function can serve tointerrupt the cycle of death which normally follows impairedcontractility and prolonged hypoperfusion. Finally, subtlechanges in ventricular compliance (i.e., a reduction in t) may becritical for diastolic filling after reperfusion, especially in thesetting of ischemia and increased myocardial oxygenconsumption/wall stress.
Possible mechanisms for improved myocardialfunction by EMF
Recent studies have shown strong correlation betweenmyocardial calcium handling to the function of hsp70—forexample, deleting the inducible 70 kDa heat shock genes impairscardiac contractile function and calcium handling associatedwith hypertrophy (Okubo et al., 2001; Kim et al., 2006). The Na/Ca2þ exchanger, known to play an important determinant ofmyocardial contractility in heart failure, is acted on by hsp70,leading to desensitization by a reduction in Vmax (Kiang et al.,1998). Hsp70 may also alter protein kinase A, protein kinase C,and phospholipase A2 (Ding et al., 1998); it is conceivable thathsp70 also modulates other components of calcium-handling,such as SERCA2a and phosphorylation of the ryanodinereceptor (Marx et al., 2000). These results suggest interactionof hsp70 with elements of the calcium release mechanism thatresult in enhanced myocardial contractility.
Induction of hsp70 levels by rapid elevation of temperaturealso improves cytoskeletal-based cell survival pathways (Weiet al., 2006). Maintenance of cell architecture associated withhsp70 has been reported in the literature previously (Wei et al.,2006), and anti-apoptotic mechanisms of stress response

Fig. 4. Ischemia-reperfusion: Contractile function was significantlyimproved in response to EMF exposure. Contractile (systolic)function was significantly increased after ischemia-reperfusion inresponse to 30 min of EMF exposure, as measured by ventricularsystolic pressure (LVSP; A), left ventricular dP/dt (LV dP/dtmax; B),and pre-load adjusted maximum power (P-A Max Power; C;yyP < 0.05 vs. Baseline, §P < 0.05 vs. Ischemia 30, MP < 0.05 vs. Control,zzP < 0.01 vs. Control, ��P < 0.005 vs. Control).
Fig. 5. Ischemia-reperfusion: diastolic function was significantlyimproved in response to EMF exposure. Diastolic function, orisovolumetric ventricular relaxation as measured by Tau (t), wassignificantly improved throughout Reperfusion after 30 minof EMF exposure (yyP < 0.05 vs. Baseline, MP < 0.05 vs. Control).
M Y O C A R D I A L P R O T E C T I O N A N D E M F 821
proteins, such as attenuation of the ASK-JNK/p38 signalingcascades (Fan et al., 2005), has been documented. There is alsoevidence that hsp70 has a direct effect on apoptosis bypreventing caspase-3 activation, PARP cleavage, DNAladdering, and cell death in vitro, but its actions appear to bedownstream of cytochrome C release (Westerheide andMorimoto, 2005). The ATP-binding domain of HSP70suppresses SAPK/JNK activation, either through suppression of
JOURNAL OF CELLULAR PHYSIOLOGY
phosphorylation and activation of the upstream kinase SEK,or by inhibition of stress-induced suppression of JNKdephosphorylation (Meriin et al., 1999; Yaglom et al., 1999; Parket al., 2001). A secondary effect of HSP70 gene upregulation isincreased manganese superoxide dismutase activity, whichserves to limit mitochondrial apoptosis in ischemia-reperfusion(Suzuki et al., 2002). Interestingly, MDA and LDH levels,markers of cardiac injury, were unchanged in our experiments(data not shown); this suggests that EMF does not diminish theextent of cardiac injury but rather enhances the contractilefunction of remaining myocytes.
EMF and biological interactions
EMF interaction with cells and tissues has been extensivelystudied in vivo and in vitro (reviewed in Goodman and Blank,2002; Blank and Goodman, 2004, 2007). EMF is known toinduce elevated DNA transcript levels of several genesincluding HSP70 as well as elevated levels of hsp70 protein in theabsence of increased temperature (reviewed in Goodman andBlank, 1998). The interaction mechanism of EMF with DNA incells and tissues to stimulate protein synthesis remainsunknown. Several theoretical approaches to EMF mechanismhave been proposed including cyclotron resonance (Liboff,1985; Lednev, 1991) and forced vibration of ions (Panagopouloset al., 2002). There is evidence from biochemical reactions thatEMFs can accelerate electron transfer and move within DNA(Blank and Soo, 2001; Goodman and Blank, 2002; Blank andGoodman, 2004, 2007).
An important clue to EMF stimulation of biosynthesis comesfrom identification of a specific EMF-sensitive DNA sequenceon both the c-myc and the HSP70 gene promoters (Lin et al.,1999, 2001). The HSP70 promoter has three nCTCTnrecognition motifs/response elements (�158 to �203 relativeto the transcription initiation site) that are EMF-sensitive (Linet al., 1999). The heat shock element (HSE), lying between�180 and �203, is required for induction of HSP70 geneexpression by EMFs (Lin et al., 1999, 2001). Mutation ofthe nCTCTn sequences (EMRE, electromagnetic responseelements) eliminates the EMF sensitivity of the HSP70promoter (Lin et al., 2001). The EMF domain and the heat shockdomain function independently. This is an HSF-1 dependentprocess (Lin et al., 1998, 1999; Lin et al., 2001). Furthermore,

822 G E O R G E E T A L .
the HSE in the heat shock domain is not interchangeable withthe HSE in the EMF domain (Lin et al., 2001). nCTCTnsequences, placed upstream of CAT or luciferase reporterconstructs (that were otherwise unresponsive to EMFs) weretransfected into HeLa cells and exposed to EMFs. Proteinextracts from EMF-exposed transfectants had significantincreases in both CAT and luciferase activity, as compared withidentical transfectants that were sham exposed (Lin et al.,2001).
Interaction with electrons could account for activation ofDNA by both low and high frequency EMFs. An EMF sensitiveDNA sequence suggests that EMFs may interact both directlyand indirectly with DNA. The initial interaction could involvethe displacement of electrons in the H-bonds that hold DNAtogether, thereby causing chain separation and initiatingtranscription and translation. Blank and Goodman (2007), usingexperimentally observed processes as links in a causal chain,have proposed that DNA activation of transcription is based onEMF’s displacement of electrons in DNA by the EMF and thatthis causes transient charging of small groups of base pairs (e.g.,nCTCTn). At the charged sites disaggregation forces overcomethe H-bonds, and this disaggregation of the two chains at thosesites permits transcription.
Clinical use of EMF technology
An abundance of hsp70 is clearly important to limit myocardialinjury, following coronary occlusion by reducing infarct size andby increasing contractile function. Modulation of hsp70 levels inthe heart, using heat stimulation, is currently problematic froma temporal standpoint. It is known that a twofold induction ofhsp70 improves heart muscle cell resistance to oxidation,ischemia and hypoxia (Heads et al., 1995; Mestril et al., 1996;Chong et al., 1998). Endogenous hsp70 doubles after only 1 hfollowing coronary artery occlusion (Loncar et al., 1998). It maytake up to 24 h for hsp70 levels to reach a four- to fivefold level.The four- to fivefold level has historically been required, at aminimum, to provide improved ischemic tolerance. This isoutside the ‘‘golden window of opportunity’’ to protect themyocardium, which is at the greatest risk in the first 6 h afterartery occlusion. Lower levels of hsp70 have not providedsufficient ischemic tolerance to prevent permanent myocardialdamage. In our experiments, augmented hsp70 levels occur asearly as 30 min after exposure, and last up to 3 h. This iswell-within an average door-to-intervention period for an acutecoronary syndrome. The importance of this specificity isthat EMF exposure produces hsp70 without the untowarddownstream effects of ubiquitous stress response activation.
It is clear that significant potential exists for this technology.It is most applicable to the coronary revascularizationpopulation, consisting of patients undergoing coronary arterybypass grafting and PCIs. With open-heart surgeries and PCIbeing performed for expanding indications of revascularization,the need for protective strategies is even more pressing. Boththerapies aim to restore coronary flow to underperfusedmyocardium after periods of critical or sub-acutehypoperfusion. Any portion of myocardium that faceshypoperfusion followed by reperfusion faces the hazards ofreperfusion injury (i.e., hemodynamic dysfunction, infarctexpansion, or arrhythmias).
In the controlled setting of cardiopulmonary bypass in theoperating room or in the angiography suite, EMF exposure caneasily be incorporated into the clinical care protocols. Exposuretimes of 30 min prior to intervention can easily be coordinatedwith ischemia and timed reperfusion. An EMF device for use inthe operating room or the emergency room would beuncomplicated. For example, in current EMF devices used forbone non-union and wound healing, coils are within lightweightbinding strap that are placed on the patient before, during and/
JOURNAL OF CELLULAR PHYSIOLOGY
or following surgery. The exposure technology is entirely non-invasive. This device is portable, weighs about one pound, andcan be easily applied by any technician.
The use of EMFs for the induction of hsp70 for post-ischemia-reperfusion treatment has clear advantages over the invasiveelevated temperature treatment efforts tested to date.Non-ionizing EMF induction of hsp70 is safe, efficient andpractical. These methods can be administered andreadministered prior to and during coronary interventions.Furthermore, hsp70 levels can be increased repeatedly withEMF versus the limited single use of the thermal method.
Summary and Conclusion
In these experiments, we report a novel non-invasive techniqueto increase hsp70 levels using exposure to low energy, lowfrequency EMF. While stress proteins in cells and tissues havebeen previously utilized as diagnostic markers and prognosticindicators, a safe, non-invasive method of augmentingendogenous defense mechanisms as a therapeutic tool, such asEMF exposure, has significant clinical potential. Our dataindicate that pre-exposure with EMF prior to ischemia andreperfusion, in a mammalian model, induces upregulation of theHSP70 gene, subsequently increased levels of hsp70 protein,and, most importantly, improved ventricular function afterischemia-reperfusion.
Acknowledgments
This work was partially supported by National Institute ofHealth Grant T32-HL07854 (I.G.) and the Robert I. GoodmanFund (R.G.). We would also like to acknowledge Eve Vagg forher invaluable assistance with the figures.
Literature Cited
Benjamin IJ, McMillan DR. 1998. Stress (heat shock) proteins: Molecular chaperones incardiovascular biology and disease. Circ Res 83:117–132.
Blank M, Khorkova O, Goodman R. 1994. Changes in polypeptide distribution stimulated bydifferent levels of electromagnetic and thermal stress. Bioelectrochem Bioenerg 33:109–114.
Blank M, Goodman R. 2004. Initial interaction in electromagnetic field-induced biosynthesis.J Cell Physiol 199:359–363.
Blank M, Goodman R. 2007. A mechanism for stimulation of biosynthesis by electromagneticfields: Charge transfers in DNA and base pair separation. J Cell Physiol 214:20–26.
Blank M, Soo L. 2001. Electromagnetic acceleration of electron transfer reactions. J CellBiochem 81:278–283.
Bolli R, Becker L, Gross G, Mentzer R, Balshaw D, Lathrop D. 2004. Myocardial protection ata crossroads: The need for translation into clinical therapy. Circ Res 95:125–134.
Brocchieri L, Conway de Marcario E, Marcario AJ. 2008. hsp70 genes in the human genome:Conservation and differentiation patterns predict a wide array of overlapping andspecialized functions. BMC Evol Biol 8:19.
Carmody S, Wu XL, Lin H, Blank M, Goodman R. 2000. Cytoprotection by electromagneticfield-induced hsp70: A model for clinical application. J Cell Biochem 79:453–459.
Chong KY, Lai CC, Lille S, Chang C, Su C. 1998. Stable overexpression of the constitutiveform of heat shock protein 70 confers oxidative protection. J Mol Cell Cardiol 30:599–608.
Cornelussen RN, Garnier AV, van der Vusse GJ, Reneman RS, Snoeckx LHEH. 1998. Biphasiceffect of heat stress pretreatment on ischemic tolerance of isolated rat hearts. J Mol CellCardiol 30:365–372.
Currie RW, Tanguay R, Klingma JG. 1993. Heat-shock response and limitation of tissuenecrosis during occlusion/reperfusion in rabbit hearts. Circulation 87:863–871.
Di Carlo AL, Farrell JM, Litovitz TA. 1999. Mycardial protection conferred byelectromagnetic fields. Circulation 99:813–816.
Ding XZ, Tsokos GC, Kiang JG. 1998. Overexpression of HSP-70 inhibits thephosphorylation of HSF1 by activating protein phosphotase and inhibiting protein kinase Cactivity. FASEB J 12:451–459.
Fan GC, Ren X, Qian J, Yuan Q, Nicolaou P, Wang Y, Jones WK, Chu G, Kranias EG. 2005.Novel cardioprotective role of a small heat-shock protein, Hsp20, against ischemia/reperfusion injury. Circulation 111:1792–1799.
Goodman R, Wei L-X, Xu J-C, Henderson A. 1989. Exposure of human cells to low-frequencyelectromagnetic fields results in quantitative changes in transcripts. Biochim Biophys Acta1009:216–220.
Goodman R, Blank M, Lin H, Khorkova O, Soo L, Weisbrot D, Henderson A. 1994. Increasedlevels of hsp transcripts are induced when cells are exposed to low frequencyelectromagnetic fields. Bioelectrochem Bioenerg 33:115–120.
Goodman R, Blank M. 1998. Magnetic field stress induces expression of hsp7. Cell StressChaperones 3:79–88.
Goodman R, Blank M. 2002. Insights into electromagnetic interaction mechanisms. J CellPhysiol 192:16–22.
Han L, Lin H, Jin M, Blank M, Goodman R. 1998. Application of magnetic field-induced heatshock protein 70 for presurgical cytoprotection. J Cell Biochem 71:577–583.

M Y O C A R D I A L P R O T E C T I O N A N D E M F 823
Heads RJ, Yellon DM, Latchman DS. 1995. Differential cytoprotection against heat stress orhypoxia following expression of specific stress protein genes in myogenic cells. J Mol CellCardiol 27:1669–1678.
Ito H, Bassett CAL. 1983. Effect of weak pulsing electromagnetic fields on neural regenerationin the rat. Clin Orthop 181:283–290.
Jayakumar J, Suzuki K, Khan M, Smolenski RT, Farrell A, Latif N, Raisky O, Abunasra H,Sammut IA, Murtuza B, Amrani M, Yacoub M. 2000. Gene therapy for myocardialprotection: Transfection of donor hearts with heat shock protein 70 gene protects cardiacfunction against ischemia-reperfusion injury. Circulation 102:302–306.
Jayakumar J, Suzuki K, Sammut IA, Smolenski RT, Khan M, Najma L, Abusnasra H, Murtuza B,Amrani M, Yacoub M. 2001. Heat shock protein 70 gene transfection protectsmitochondrial and ventricular function against ischemia-reperfusion injury. Circulation104:I-303–I-307.
Jin M, Lin H, Han L, Opler M, Maurer S, Blank M, Goodman R. 1997. Biological and technicalvariables in myc expression in HL60 cells exposed to 60 Hz electromagnetic fields.Bioelectrochem Bioenerg 44:111–120.
Kiang JG, Ding XZ, McClain DE. 1998. Overexpression of HSP-70 attenuates increasesin Ca2þ and protects human epidermoid A-431 after chemical hypoxia. Toxicol ApplPharmacol 149:185–194.
Kim YK, Suarez J, Hu Y, McDonough PM, Boer C, Dix DJ, Dillmann WH. 2006. Deletion of theinducible 70-kDa heat shock protein genes in mice impairs cardiac contractile function andcalcium handling associated with hypertrophy. Circulation 113:2589–2597.
Lednev VV. 1991. Possible mechanism for influence of magnetic fields on biological systems.Bioelectromagnetics 12:71–75.
Liboff AR. 1985. Geomagnetic cyclotron resonance in membrane transport. J Biol Phys13:99–102.
Lin H, Opler M, Head M, Blank M, Goodman R. 1997. Electromagnetic field exposure inducesrapid transitory heat shock factor activation in human cells. J Cell Biochem 66:482–488.
Lin H, Head M, Blank M, Jin M, Goodman R. 1998. Myc-mediated transactivation of HSP70expression following exposure to magnetic fields. J Cell Biochem 69:181–188.
Lin H, Blank M, Goodman R. 1999. Magnetic field-responsive domain in human HSP70promoter. J Cell Biochem 75:170–176.
Lin H, Blank M, Goodman R. 2001. Regulating genes with electromagnetic response elements.J Cell Biochem 81:143–148.
Liu JC, He M, Wan L, Cheng XS. 2007. Heat shock protein gene transfection protects ratmyocardium cell against anoxia-reoxygenation injury. Chin Med J (Engl) 120:578–583.
Loncar R, Flesche CW, Deussen A. 1998. Regional myocardial heat-shock protein (HSP70)concentrations under different blood flow conditions. Pflugers Arch 437:98–103.
Marx SO, Reiken S, Hisamatsu T, Jayaraman T, Burkhoff D, Rosemblit N, Marks AR. 2000.PKA phosphorylation dissociates FKBP12.6 from the calcium release channel (ryanodinereceptor): Defective regulation in failing hearts. Cell 101:365–376.
Meriin AB, Yaglom JA, Gabai VL, Zon L, Ganiatsas S, Mosser DD, Zon L, Sherman MY. 1999.Protein-damaging stresses activate c-Jun terminal kinase via inhibition of itsdephosphorylation: A novel pathway controlled by HSP7. Mol Cell Biol 19:2547–2555.
JOURNAL OF CELLULAR PHYSIOLOGY
Mestril R, Giordano FJ, Conde AG, Dillmann WH. 1996. Adenovirus-mediated gene transferof a heat shock protein 70 (hsp) protects against stimulated ischemia. J Mol Cell Cardiol26:2351–2358.
Nitta Y, Abe K, Aoki M, Ohno I, Isoyama S. 1994. Diminished heat shock protein 70 mRNAinduction in aged rat hearts after ischemia. Am J Phyisol 267:H1795–H1803.
Okubo S, Wildner O, Shah MR, Chelliah JC, Hess ML, Kukreja RC. 2001. Gene transfer ofheat-shock protein 70 reduces infarct size in vivo after ischemia/reperfusion in the rabbitheart. Circulation 103:877–881.
Panagopoulos DJ, Karabarbounis A, Margaritis LH. 2002. Mechanism of action ofelectromagnetic fields on cells. Biochem Biophys Res Commun 298:95–102.
Park HS, Lee JS, Huh SH, Seo JS, Choi EJ. 2001. Hsp72 functions as a natural inhibitory proteinof c-Jun N-terminal kinase. EMBO J 20:446–456.
Plumier JCL, Currie RW. 1996. Heat shock-induced myocardial protection against ischemicinjury: A role for Hsp70? Cell Stress Chaperones 1:13–17.
Raff GL, Glantz SA. 1981. Volume loading slows left ventricular isovolemic relaxation rate.Evidence of load-dependent relaxation in the intact dog heart. Circ Res 48:813–824.
Schneider EL. 1999. Aging in the third millenium. Science 283:796–797.Shallom JM, DiCarlo AL, Ko D, Penafiel LM, Nakai A. 2002. Microwave exposure induces
hsp70 and confers protection against hypoxia in chick embryos. J Cell Biochem 86:490–496.
Suzuki K, Sawa Y, Kaneda Y, Ichikawa H, Shirakura R, Matsuda H. 1997. In vivo genetransfection with heat shock protein 70 enhances myocardial tolerance to ischemia-reperfusion injury in rat. J Clin Invest 99:1645–1650.
Suzuki K, Sawa Y, Kagisaki K, Taketani S, Ichikawa H, Kaneda Y, Matsuda H. 2000. Reductionin myocardial apoptosis associated with overexpression of heat shock protein 7. Basic ResCardiol 95:397–403.
Suzuki K, Murtuza B, Sammut IA, Latif N, Jayakumar J, Smolenkski RT, Kaneda Y, Sawa Y,Matsuda H, Yacoub MH. 2002. Heat shock protein 72 enhances manganese superoxidedismutase activity during myocardial ischemia-reperfusion injury, associated withmitochondrial protection and apoptosis reduction. Circulation 106:I-270–I-276.
Udelsman R, Blake MJ, Stagg CA, Li D-G, Putney D, Holbrook NJ. 1993. Vascular heat shockprotein expression in response to stress. J Clin Invest 91:465–473.
Wei H, Campbell W, Vander-Heide RS. 2006. Heat shock-induced cardioprotectionactivates cytoskeletal-based cell survival pathways. Am J Physiol Heart Circ Physiol291:H638–H647.
Wei L-X, Goodman R, Henderson A. 1990. Changes in levels of c-myc and histone H2Bfollowing exposure of cells to low frequency sinusoidal electromagnetic fields: Evidence fora window effect. Bioelectromagnetics 11:269–272.
Westerheide SD, Morimoto RI. 2005. Heat shock response modulators as therapeutic toolsfor diseases of protein conformation. J Biol Chem 280:33097–33100.
Yaglom JA, Gabai VL, Meriin AB, Mosser DD, Sherman MY. 1999. The function of HSP72 insuppression of c-Jun N-terminal kinase activation can be dissociated from its role inprevention of protein damage. J Biol Chem 274:20223–20228.
Zhao Y, Wang W, Qian L. 2007. Hsp70 may protect cardiomyocytes from stress-inducedFAS-mediated apoptosis. Cell Stress Chaperones 12:83–95.